activation of t-cells by bryostatins: induction of the il2 receptor gene transcription and...
TRANSCRIPT

Int. J. Imraunopharmac., Vol. 12, No. 5, pp. 481-490, 1990. 0892-0561/90 $3.00 + .IX) Printed in Great Britain. International Society for immunopharmaeology.
ACTIVATION OF T-CELLS BY BRYOSTATINS: INDUCTION OF THE IL-2 RECEPTOR GENE TRANSCRIPTION AND DOWN-MODULATION
OF SURFACE RECEPTORS
AHMED H. ESA,** WILLIAM O. BOTO,* WILLIAM H. ADLER,* W. STRATFORD MAY *~ and ALLAN D. HESS *j
*The Johns Hopkins School of Medicine, Baltimore, Maryland, and the *Institute of Aging, the National Institutes of Health, Francis Scott Key Medical Center, Baltimore, Maryland, U.S.A.
(Received 4 December 1989 and in final form 25 January 1990)
Abstract - - Bryostatins are macrocyclic lactones isolated from the marine bryozoan Bugula neritina. They are currently evaluated for putative antineoplastic activity. Bryostatins bind and activate protein kinase C (PK-C), the cellular receptor for the phorbol esters, and elicit PK-C-dependent cellular functions. Such functions include the expression of the interleukin-2 receptor (IL-2R). Northern blot hybridization with a human IL-2R and an IL-2 eDNA showed that bryostatin I (bryo 1), like the phorbol ester, PMA, activates the IL-2R gene. Activation with bryo 1 or PMA in the presence of a calcium ionophore, tt23187, increased IL-2 message. These findings indicate that calcium mobilization is necessary for bryo 1 or PMA induced IL-2 gene expression. Unlike PMA, bryo 1 did not cause a vigorous proliferative response of T-lymphocytes unless A23187 was added to the cultures. A bryostatin congener, bryo 13, was inactive in the above assays. Short-term treatment of T-cells with bryo 1 and PMA resulted in an equivalent down-regulation of surface CD3 and CD4 receptors without affecting the CD8 receptor. Bryo 1 or PMA mediated expression of surface IL-2R and T-cell proliferation induced by bryo 1 or PMA were sensitive to inhibition by the PK-C antagonists staurosporine (Sts) and H-7. In contrast, CD4 and CD3 down-regulation were resistant to H-7, but could be blocked by Sts, although the Sts concentration required to block bryo 1 or PMA-indueed down- modulation was 2.5-fold higher than required to inhibit IL-2R expression and T-cell proliferation. These results indicate that bryostatins activate T-cells through PK-C.
Bryostatins constitute a group of macrocyclic lactones which are isolated from Bugula neritina (Pettit, Herald, Doubek & Herald, 1982; Petit, Herald, Kamano, Gust & Aoyagi, 1983). In initial screening tests, some bryostatin congeners were found to possess promising antineoplastic activity (Pettit, Eleet, Herald, Kamano, Boettner, Baczynskyj & Neiman, 1987). These novel agents are active against various tumors in vitro, and have also been demonstrated to be effective in in vivo tumor models (Dale & Geschen, 1989). In addition, bryostatins share certain properties with the granulocyte/macrophage colony stimulating factor (GM-CSF), such as the stimulation of normal bone marrow colony growth (May, Sharkis, Esa, Gebbia, Kraft, Pettit & Sensenbrenner, 1987).
Bryostatins activate human T lymphocytes to express the IL-2R, and when combined with a calcium ionophore, can effectively stimulate the production of IL-2 and proliferation of T-cells (Hess, Silanskis, Esa, Pettit & May, 1988). In vivo bryostatins appear to be synergistic with IL-4 in the generation of nonspecific cytotoxic T-cells (Trenn, Pettit, Takyama, Hu-Li & Sitkovsky, 1988). These properties make bryostatins promising candidates as antineoplastic biological response modifiers useful in imrnunotherapeutic interventions.
The precise mechanism of action of the bryostatins is still under investigation, but their ability to activate PK-C, a crucial intracellular enzyme important in lymphocyte activation, appears to be
tTo whom correspondence should be addressed at: The Johns Hopkins School of Medicine Oncology Center, RM 3-127, 600 N. Wolfe Street, Baltimore, MD 21205, U.S.A. ~WSM and ADH are Scholars of the Leukemia Society of America. Abbreviations used in this paper: PK-C, Ca2÷phospholipid dependent protein kinase; H-7, 5-(isoquinolinesulfonyl)-2- methylpiperazine dihydrochloride; Sts, staurosporine; PMA, phorbol myristate acetate; and DMSO, dimethylsulfoxide.
481

482
pivotal to their action. Structure- function relationship studies have now identified a site of some hryostatin congeners that fits the pharmaco- phore model of the phorbol esters (Wender, Cribbs, Koehler, Sharkey, Herald, Kamano, Pettit & Blumberg, 1988). Unlike phorbol esters however, byro 1 is poorly mitogenic for T-cells. The results described here extend our previous studies (Hess et al., 1988) on the effects of bryostatins on human T-lymphocytes and show that bryostatins induce the transciptional activation of the IL-2R gene, and down-modulation of specific T-ceU surface markers such as CD3 and CD4. The present data support the hypothesis that bryostatins, like phorbol esters, activate PK-C-dependent T-cell functions.
A. H. ESA et al.
resuspended at a density of 2 × 106/ml in culture medium (RPMI-1640 (Gibco, Grand Island, NY) supplemented with 10070 fetal bovine serum, 100/~g/ml streptomycin and 100 units/ml penicillin). Aliquots of the cells were pre-incubated with up to 10 nM staurosporine (Kyowa Hakko, Inc., NY) for 2 h in Falcon tissue culture flasks. The cells were stimulated with 50 nM bryo 1, bryo 13, PMA, alone or together with 0.3/aM calcium ionophore, A23187 (Sigma, St Louis, MO). Triplicate 200/al of these cells were incubated in microliter plates for proliferative response determination by 3H- thymidine ([3H]Tdr) incorporation assay. The cultures were incubated at 37°C, 5o70 CO2 and 95070 humidity for 5 h for northern blot analysis or 72 h for (3H)Tdr incorporation.
EXPERIMENTAL PROCEDURES
Preparation o f agents The two bryostatin congeners, bryostatin 1 and 13
used in this study were kind gifts from Dr G. R. Pettit, Arizona State University, and were prepared as described previously (Pettit et al., 1982, 1983, 1987). Stock preparations were stored at -200C in 100070 DMSO at 10 -3 M, and diluted in PBS before use. Stock phorbol myristate acetate (PMA) (Sigma, St Louis, MO) was stored in DMSO and diluted in RPMI when needed. Staurosporine (Sts, Kyogo, NY, NY) was diluted in PBS. H-7, 5-(isoquinolinsulfonyl)- 2-methylepiperazine dihydrochloride, (Seikagaku Kogyo Co., St Petersburg, FL) was dissolved in distilled water and diluted in PBS.
Detection of IL-2 and IL-2R mRNAs by northern blot analysis
Fifteen micrograms of total RNA isolated form pelleted cells (Chirgwin, Przybyla, MacDonald & Rutter, 1979; Glisin, Crkvenjakov & Byus, 1974), were denatured with glyoxal, fractionated by electrophoresis in 1 o70 agarose gels, and then blotted onto Hybond-N nylon membrane (Amersham, Arlington Heights, IL) or Genescreen Plus (NEN, Boston, MA). The blots were hybridized in 50070 formamide (LeMeur, Glanville, Mandel, Garlinger, Palmiter & Chambon, 1981; Kern, Reed, Daniele & Nowell, 1986) to (d-32P) dATP-labeled (Feinberg & Vogelstein, 1983) IL-2 or IL-2R cDNA's (Oncor, Gaithersburg, MD). The blots were exposed to X-ray films between intensifying screens for 12-48 h at -70°C.
Isolation and activation of T-cells T-lymphocytes were purified by passing peripheral
blood mononuclear cells of normal donors through nylon wool columns followed by E-rosetting as follows. Nonadherent cells were washed and then incubated with sheep erythrocytes which were pretreated for 15 rain with 0.15 M aminoethyl- isothioronium bromide (Sigma Chemical Co., St Louis, MO) at pH 8.0. Rosetting cells were separated from nonrosetting cells by centrifugation through lymphocyte separation medium (Litton Bionics, Kensington, MD). T-cells were recovered from the pelleted rosettes upon lysis of the erythroeytes with 5 ml of 58 mM Tris-ammonium chloride buffer (pH 7.2) for 5 - 8 rain. These cells were 97- 99~/0 CD2 ÷ as analysed by flow cytometry using OKTll monoclonal antibody. Purified T-cells were then
Flow cytometry for surface antigens The expression of IL-2R, CD3, CD4, and CD8 on
T-cells activated with bryo 1 or PMA was determined by flow cytometric analysis using respective monoclonal antibodies (anti-IL-2 R, CD4, CD8, CD3 [Beckton Dickinson]). The cells were incubated in RPMI-1640 + 10°70 FCS at 37°C and 5070 CO2 with or without 50 nM bryo 1 or PMA for 1 h. To evaluate the effects of inhibitors, Sts and H-7 were added to the cells for 20 rain prior to treatment with bryo 1 and PMA. The cells were then washed three times and stained with the respective FITC- conjugated mouse anti-human monoclonal antibodies. For analysis, a Coulter EPICS 752 flow cytometer was used to analyse 10,000 cells per sample. Percentage of positive cells were quantified as those with fluorescence intensities higher than
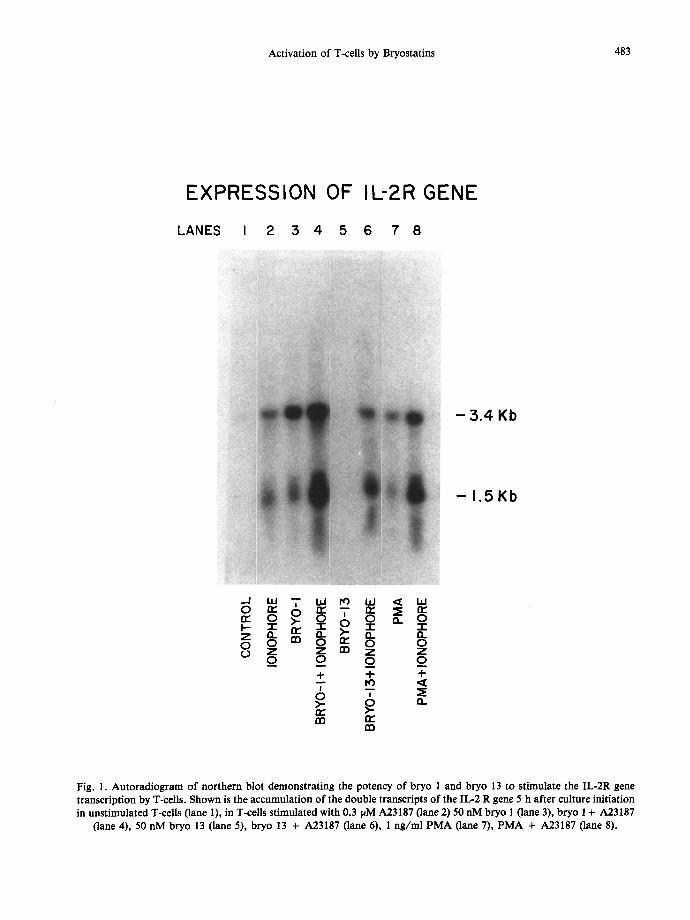
Activation of T-cells by Bryostatins 483
EXPRESSION OF IL-2R GENE
LANES I 2 3 4 5 6 7 8
- 3.4 K b
- 1 . 5 K b
7
+ "-I- -I- "7 rO < O "7 IE >. ~ o_
II1 I1~ m
Fig. 1. Autoradiogram of northern blot demonstrating the potency of bryo 1 and bryo 13 to stimulate the IL-2R gene transcription by T-cells. Shown is the accumulation of the double transcripts of the IL-2 R gene 5 h after culture initiation in unstimulated T-cells (lane 1), in T-cells stimulated with 0.3 pM A23187 (lane 2) 50 nM bryo 1 0ane 3), bryo 1 + A23187
(lane 4), 50 nM bryo 13 (lane 5), bryo 13 + A23187 (lane 6), 1 ng/ml PMA (lane 7), PMA + A23187 (lane 8).
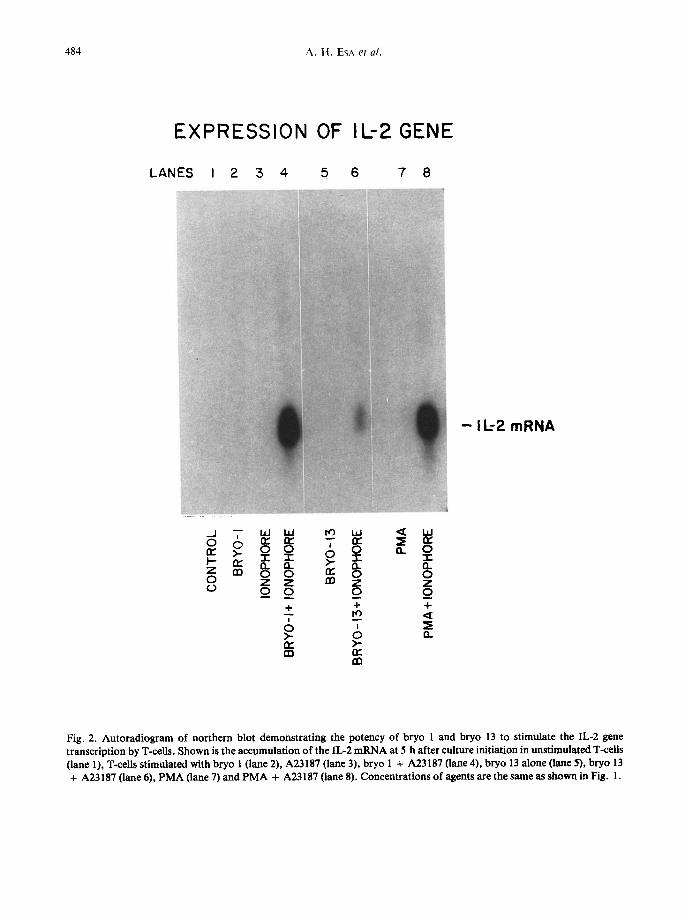
484 A . H . ESA et al.
EXPRESSION OF IL-2 GENE
L A N E S I 2 :5 4 5 6 7 8
- 11.=2 m R N A
o. a_ Z nn ~ 0 ry 0 0 0 Z Z r,n Z Z
_o o _o o + + -4-
m
Fig. 2. Autoradiogram of northern blot demonstrating the potency of bryo 1 and bryo 13 to stimulate the IL-2 gene transcription by T-cells. Shown is the accumulation of the IL-2 mRNA at 5 h after culture initiation in unstimulatexi T-cells (lane 1), T-cells stimulated with bryo 1 (lane 2), A23187 (lane 3), bryo 1 + A23187 (lane 4), bryo 13 alone (lane 5), bryo 13 + A23187 (lane 6), PMA (lane 7) and PMA + A23187 (lane 8). Concentrations of agents are the same as shown in Fig. 1.
Activation of T-cells by Bryostatins
Table 1. Proliferation and expression of the IL-2R by bryo 1- and PMA-aetivated T-cells
Treatment Per cent IL-2R positive cells Thymidine uptake
Experiment No. I Experiment No. 2
Medium 7.80 10.34 870 ± 118 H-7 8.90 ND ND Sts 9.70 ND ND
Bryo 1 only 61.80 68.10 12421 _+ 283 Bryo 1 + H-7 24.00 19.90 990 + 244 Bryo 1 + Sts 29.10 31.40 717 + 21
PMA 75.50 90.8 44634 __. 684 PMA + H-7 52.90 44.80 204 _+ 27 PMS + Sts 35.00 51.90 4081 + 269
T-cells were incubated with medium, 25/AVI H-7, or 4 nM Sts for 20 min, and then stimulated with 50 nM bryo 1 or PMA for 48 h, harvested and incubated with anti-IL-2R monoclonal antibody conjugated with FITC and analysed by flow cytometry for percent cells expressing the IL-2R. Thymidine uptake was determined by incubating triplicate aliquots in 0.1/aCi 3-H-thymidine for the last 4 h of the 48 h incubation period. The values show for thymidine uptake are means _+ 1 S.D. from a representative experiment.
485
99°70 of control cells stained with FITC-conjugated normal mouse Ig. Comparison between fluorescence intensities were made on the basis of arbitrary units using mean channel of fluorescence.
RESULTS
Stimulation o f the IL-2R gene by bryo 1, bryo 13 or PMA
We compared the relative effectiveness of bryo 1, bryo 13, A23187 and PMA for the regulation of the IL-2R gene function. The results in Fig. 1 show that both bryo 1 or PMA, as well as A23187 induce detectable levels of the IL-2R gene in T-cells. However, equimolar concentrations of bryo 13 did not induce detectable IL-2R gene. Bryo 1 and PMA, when combined with the Ca ~+ ionophore A23187, promoted maximal expression of the IL-2R gene. Although bryo 13 was ineffective when used alone, activity was noted when it was used in combination with A23187. As shown in Table 1, the expression of surface IL-2R induced by bryo 1 and PMA closely resembled the relative capacity of these agents to induce gene transcription. We further tested the role of PK-C activation in bryo 1 and PMA-induced IL-2R expression by examining the effects of two PK-C antagonists. The results shown in Table 1 indicate that bryo 1- or PMA-induced IL-2R expression was sensitive to 25/aM H-7 and 4 nM Sts.
In addition, the data in Table 1 show that the same concentration of bryo 1 was far less stimulatory for cellular proliferation than PMA, although these concentrations were comparably effective in the induction of IL-2R gene transcription. However, when A23187 was included in the culture, bryo 1 and PMA caused equivalent proliferation of T-cells (data not shown, similar data was previously reported by Hess et al., 1988).
Stimulation of the expression o f the IL.2 gene by bryo 1, bryo 13 or PMA
In contrast to their effectiveness in the stimulation of the IL-2R gene, bryo 1 and PMA were ineffective for transcriptional activation of the T-cell IL-2 gene (Fig. 2). Bryo 13 and A23187 alone were also inactive for the induction of the IL-2 gene transcrip- tion. However, sensitization of T-lymphocytes with bryo 1, or PMA, in combination with A23187 induced substantial accumulation of IL-2 message, as detected by northern blot hybridization. Bryo 13 also showed some activity when A23187 was used as a co-stimuhis, but this treatment was far less potent than when bryo 1 or PMA were used.
Regulation of cell surface markers by bryo 1
Given the rapid down-modulation of specific cell surface determinants induced by phorbol esters (Blue, Daley, Levine, Branton & Schlossman, 1989;
486
¢r" LU ¢n
Z .J ..J LU L.)
LU >
I-- < J I.,d n."
®
n~nt rol CD4
©
/ ~ ~ cryol
®
I PMA
' L ®
A. H. ESA et al.
®
Br yo 1
Control
D8
FLUORESCENCE INTENSITY
®
:o4 FLUORESCENCE INTENSITY
PMA
Control
CO8
FLUORESCENCE INTENSITY
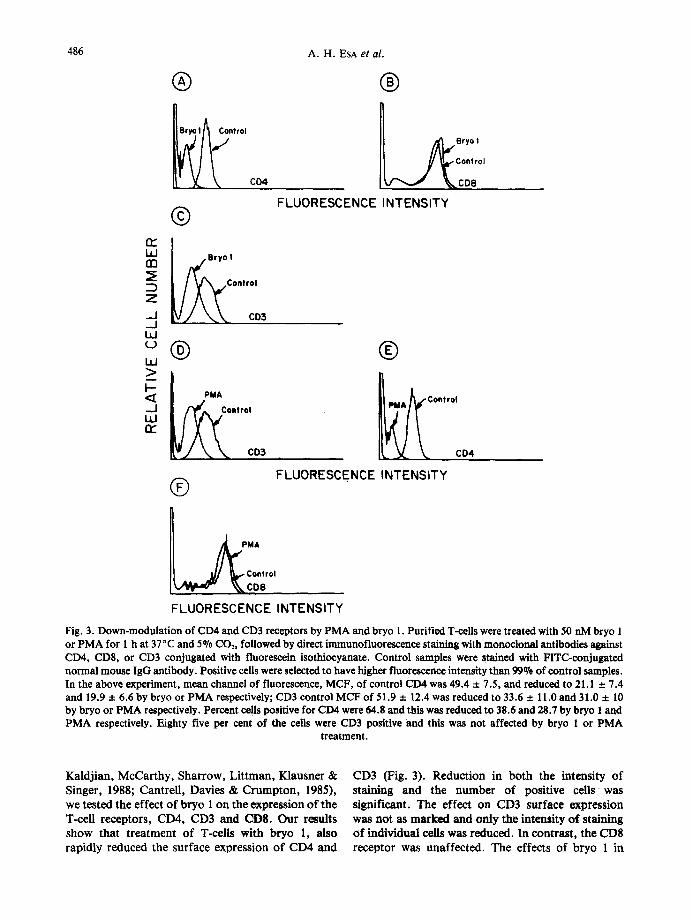
Fig. 3. Down-modulation of CD4 and CD3 receptors by PMA and bryo 1. Purified T-cells were treated with 50 nM bryo 1 or PMA for 1 h at 37°C and 5% CO2, followed by direct immunofluorescence staining with monoctonal antibodies against CD4, CDS, or CD3 conjugated with fluorescein isothiocyanate. Control samples were stained with FITC-conjugated normal mouse IgG antibody. Positive cells were selected to have higher fluorescence intensity than 99% of control samples. In the above experiment, mean channel of fluorescence, MCF, of control CD4 was 49.4 + 7.5, and reduced to 21.1 + 7.4 and 19.9 + 6.6 by bryo or PMA respectively; CD3 control MCF of 51.9 + 12.4 was reduced to 33.6 + 11.0 and 31.0 + 10 by bryo or PMA respectively. Percent cells positive for CD4 were 64.8 and this was reduced to 38.6 and 28.7 by bryo 1 and PMA respectively. Eighty five per cent of the cells were CD3 positive and this was not affected by bryo 1 or P M A
treatment.
Kaldj ian , McCar thy , Shar row, L i t tman , Klausner & Singer, 1988; Cantre l l , Davies & C r u m p t o n , 1985), we tested the effect o f b ryo 1 on the express ion o f the T-cell receptors , CD4, CD3 and CDS. O u r results show tha t t r ea tmen t o f T-cells wi th b ryo 1, also rapidly reduced the surface express ion o f CD4 and
CD3 (Fig. 3). Reduction in both the intensity of staining and the number of positive cells was significant. The effect on CD3 surface expression was not as marked and only the intensity of staining of indiv idual cells was reduced. In contras t , the CD8 receptor was unaffec ted . The effects of b ryo 1 in
Activation of T-cells by Bryostatins
® ® PMA
Control
LLJ ~3
- J ts ÷ PMA
LLJ
> F--
Ixl
I[ H-7 + PMA . ~ PMA
® Bryo I
i £ Control
®
t$ + 8 ryo 1
ryo 1
®
~ H-7 + Bryo ! Bryol
FLUORESCENCE INTENSITY (CD4)
487
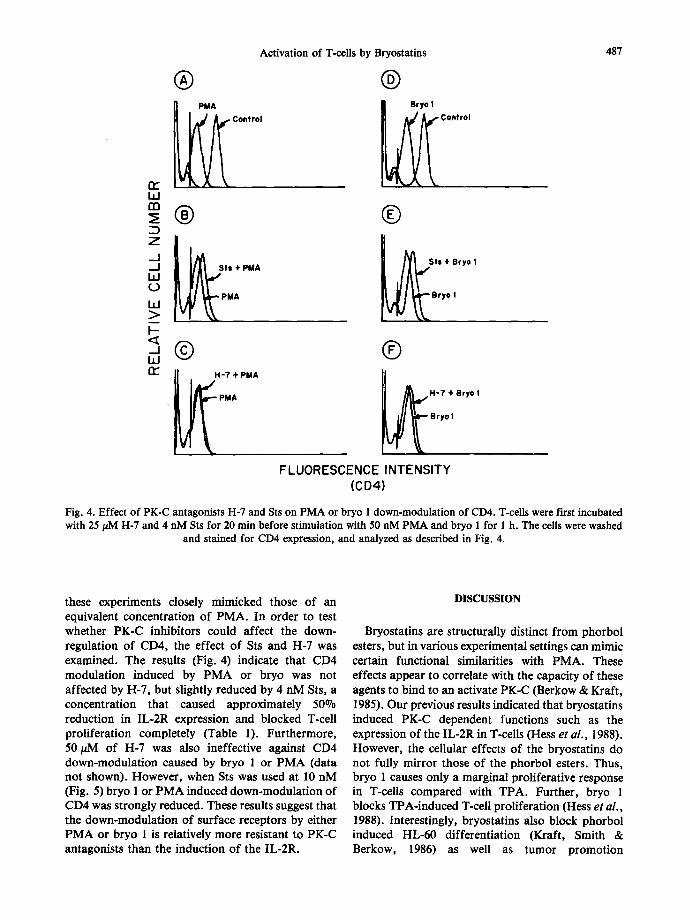
Fig. 4. Effect of PK-C antagonists H-7 and Sts on PMA or bryo 1 down-modulation of CD4. T-cells were first incubated with 25/~M H-7 and 4 nM Sts for 20 min before stimulation with 50 nM PMA and bryo 1 for 1 h. The cells were washed
and stained for CD4 expression, and analyzed as described in Fig. 4.
these experiments closely mimicked those of an equivalent concentration of PMA. In order to test whether PK-C inhibitors could affect the down- regulation of CD4, the effect of Sts and H-7 was examined. The results (Fig. 4) indicate that CD4 modulation induced by PMA or bryo was not affected by H-7, but slightly reduced by 4 nM Sts, a concentration that caused approximately 50°70 reduction in IL-2R expression and blocked T-cell proliferation completely (Table 1). Furthermore, 50/~M of H-7 was also ineffective against CD4 down-modulation caused by bryo 1 or PMA (data not shown). However, when Sts was used at 10 nM (Fig. 5) bryo 1 or PMA induced down-modulation of CD4 was strongly reduced. These results suggest that the down-modulation of surface receptors by either PMA or bryo 1 is relatively more resistant to PK-C antagonists than the induction of the IL-2R.
DISCUSSION
Bryostatins are structurally distinct from phorbol esters, but in various experimental settings can mimic certain functional similarities with PMA. These effects appear to correlate with the capacity of these agents to bind to an activate PK-C (Berkow & Kraft, 1985). Our previous results indicated that bryostatins induced PK-C dependent functions such as the expression of the IL-2R in T-cells (Hess et al., 1988). However, the cellular effects of the bryostatins do not fully mirror those of the phorbol esters. Thus, bryo 1 causes only a marginal proliferative response in T-cells compared with TPA. Further, bryo 1 blocks TPA-induced T-cell proliferation (Hess et ai., 1988). Interestingly, bryostatins also block phorbol induced HL-60 differentiation (Kraft, Smith & Berkow, 1986) as well as tumor promotion
488 A. H. ESA eta / .
30 ..... " ' ' ~ . . . . ~ . . . . ' . . . . ' . . . . ~ . . . . However, the potency of both bryo 1 and PMA was
25
20
15
10
Sts 10nM/Bryo
~ . L , , ' I I I I " • I . . . . 15 20 25 30
$ts 4nM/Bryo
B r y o o n l y ~
5 10
4-ControL
" ' ' 1 ' ' ' ' | ' ' ' ' I ' ' ' ' 1 . . . . I . . . .
5
E c :
3o
~ 25
2 0
1 5
3is I0 nM/PMA
5 I 0
ontroL
[ ~ t ' 1 , . , ,| [ a. . . . . 1 , , , ,
15 20 25 30
10
5i
Fluorescence intensity
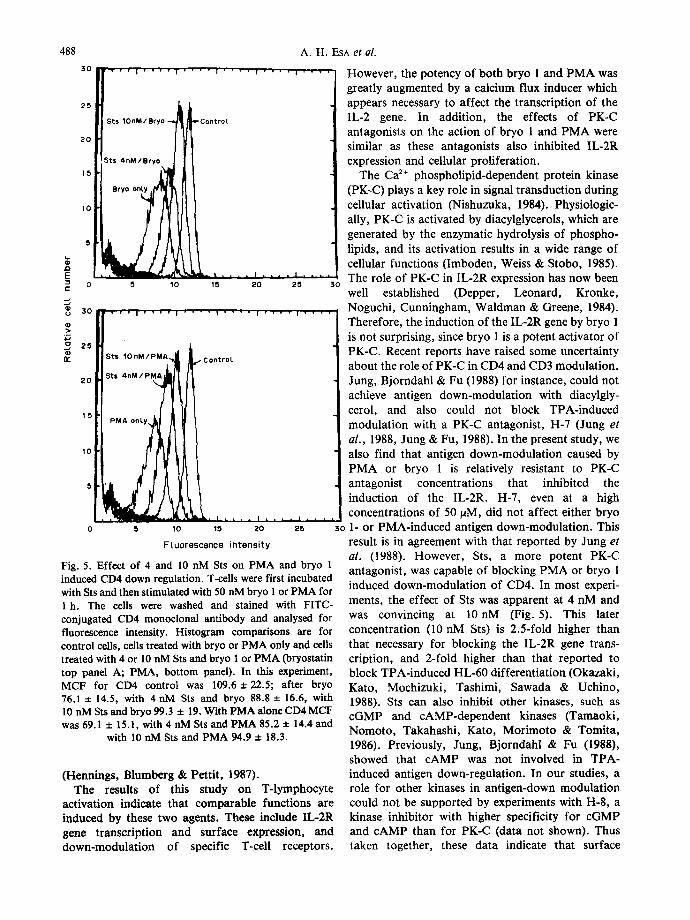
Fig. 5. Effect of 4 and 10 nM Sts on PMA and bryo 1 induced CD4 down regulation. T-cells were first incubated with Sts and then stimulated with 50 nM bryo 1 or PMA for 1 h. The cells were washed and stained with FITC- conjugated CD4 monoclonal antibody and analysed for fluorescence intensity. Histogram comparisons are for control cells, cells treated with bryo or PMA only and cells treated with 4 or 10 nM Sts and bryo 1 or PMA 0aryostatin top panel A; PMA, bottom panel). In this experiment, MCF for CD4 control was 109.6 + 22.5; after bryo 76.1 ± 14.5, with 4 nM Sts and bryo 88.8 ± 16.6, with 10 nM Sts and bryo 99.3 ± 19. With PMA alone CD4 MCF was 69.1 + 15.1, with 4 nM Sts and PMA 85.2 ± 14.4 and
with 10 nM Sts and PMA 94.9 ± 18.3.
(Hennings, Blumberg & Pettit, 1987). The results of this study on T-lymphocyte
activation indicate that comparable functions are induced by these two agents. These include IL-2R gene transcription and surface expression, and down-modulation of specific T-cell receptors.
greatly augmented by a calcium flux inducer which appears necessary to affect the transcription of the IL-2 gene. In addition, the effects of PK-C antagonists on the action of bryo 1 and PMA were similar as these antagonists also inhibited IL-2R expression and cellular proliferation.
The Ca 2÷ phospholipid-dependent protein kinase (PK-C) plays a key role in signal transduction during cellular activation (Nishuzuka, 1984). Physiologic- ally, PK-C is activated by diacylglycerols, which are generated by the enzymatic hydrolysis of phospho- lipids, and its activation results in a wide range of cellular functions (Imboden, Weiss & Stobo, 1985). The role of PK-C in IL-2R expression has now been well established (Depper, Leonard, Kronke, Noguchi, Cunningham, Waldman & Greene, 1984). Therefore, the induction of the IL-2R gene by bryo 1 is not surprising, since bryo 1 is a potent activator of PK-C. Recent reports have raised some uncertainty about the role of PK-C in CD4 and CD3 modulation. Jung, Bjorndahl & Fu (1988) for instance, could not achieve antigen down-modulation with diacylgly- cerol, and also could not block TPA-induced modulation with a PK-C antagonist, H-7 (Jung et al. , 1988, Jung & Fu, 1988). In the present study, we also find that antigen down-modulation caused by PMA or bryo 1 is relatively resistant to PK-C antagonist concentrations that inhibited the induction of the IL-2R. H-7, even at a high concentrations of 50/aM, did not affect either bryo 1- or PMA-induced antigen down-modulation. This result is in agreement with that reported by Jung et al. (1988). However, Sts, a more potent PK-C antagonist, was capable of blocking PMA or bryo 1 induced down-modulation of CD4. In most experi- ments, the effect of Sts was apparent at 4 nM and was convincing at 10nM (Fig. 5). This later concentration (10 nM Sts) is 2.5-fold higher than that necessary for blocking the IL-2R gene trans- cription, and 2-fold higher than that reported to block TPA-induced HL-60 differentiation (Okazaki, Kato, Mochizuki, Tashimi, Sawada & Uchino, 1988). Sts can also inhibit other kinases, such as cGMP and cAMP-dependent kinases (Tamaoki, Nomoto, Takahashi, Kato, Morimoto & Tomita, 1986). Previously, Jung, Bjorndahl & Fu (1988), showed that cAMP was not involved in TPA- induced antigen down-regulation. In our studies, a role for other kinases in antigen-down modulation could not be supported by experiments with H-8, a kinase inhibitor with higher specificity for cGMP and cAMP than for PK-C (data not shown). Thus taken together, these data indicate that surface

Activation of T-cells by Bryostatins
antigen down-modulation is more resistant to PK-C antagonists than the IL-2R expression, and our results with respect to H-7 are in agreement with those reported by Jung et al. (1988). The more potent PK-C antagonist, Sts, can however be used to block PMA or bryo 1 caused cell surface down-modula- tion. The possibility that different PK-C isoforms are responsible for the induction of the IL-2R and down-modulation of surface antigen receptors cannot be ruled out by the available data. This possibility would, however, mean (a) that the two diverse inhibitors used in this study (H-7 and Sts) have similar PK-C isozyme specificity, and (b) both bryo 1 and PMA activate the same isozymes. It is conceivable that the inhibitors used in this study may affect a step distal to PK-C mediated down-modu- lation of surface antigens but required for IL-2R
489
expression and cellular proliferation, and thus result in differential inhibition.
Despite the equivalent effects of bryo 1 and PMA for the T-cell properties found above, bryo 1 causes significantly less T-cell proliferation in comparison with equimolar concentrations of PMA. Thus, PK-C agonists such as PMA or teleocidin may engage a pathway(s) or a PK-C isoform(s) not affected by bryo 1.
Acknowledgements - - The authors wish to thank Ms Julie Mergerian for her technical studies and Mr Jim Flook for his expert help with the flow cytometric analysis. This work was supported by NCI Institutional Grant No. IN-11-28, NIH grant Nos. AI24682, CA15396, CA27903, CA47993 and ACS IM398.
REFERENCES
BERKOW, R. L. & KRAFT, A. S. (1985). Bryostatin, a non phorbol ester lactone, activates intact human polymorphonuclear leukocytes and binds to the phorbol ester receptor. Biochem. biophys. Res. Commun., 131, 1109-1116.
BLUE, M-L., DALLY, J. F., LEVINE, H., BRANTON, K. H. & SCHLOSSMAN, S. (1989). Regulation of CD4 and CD8 surface expression on human thymocyte subpopulations by triggering through CD2 and the CD3-T-cell receptor. J. Immun., 142, 374- 380.
CANTRELL, O. A., DAVIES, A.A. & CRUMPTON, M. J. (1985). Activators of protein kinase C down-regulate and phosphorylate the T3/T cell antigen receptor complex of human T lymphocytes. Proc. natn. Acad. Sci. U.S.A., 82, 8158-8162.
CHIRGWIN, J. M., PRZYBYLA, A.E. , MACDONALD, R. J. & ROTTER, W. J. (1979). Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry, 18, 5294-5298.
DALE, I. & GESCHEN, A. (1989). Effects of activators of protein kinase C, including bryostatin-I & 2, on the growth of A549 human lung carcinoma cells. Int. J. Cancer, 43, 158 - 163.
DEPPER, J. M., LEONARD, W. J., KRONKE, M., NOGUCHI, P. H., CUNNINGHAM, R. E., WALDMAN, T. A. & GREENE, W. C. (1984). Regulation of interleukin 2 receptor expression: effects of phorbol diester, phospholipase C, and exposure to lectin or antigen. J. Immun., 133, 3054-3061.
FEINBERG, A. & VOGELSTEIN, B. (1983). A technique for radiolabeled DNA restriction endonuclease fragments to high specific activity. Analyt. Biochem., 132, 6 - 13.
GLISlN, V., CRKVENJAKOV, R. & BYOS, C. (1974). Ribonucleic acid isolated by cesium chloride centrifugation. Biochemistry, 13, 2633 - 2637.
HENNINOS, H., BLUMBERG, P., PETTIT, G. R. et al. (1987). Bryostatin 1, an activator of PK-C, inhibits tumor promotion by phorbol esters in Sencar mouse skin. Carcinogenesis, 8, 1343- 1346.
HESS, A. D., SILANSKIS, M. K., ESA, A. H., PETTIT, G. R. & MAY, W. S. (1988). Activation of human T lymphocytes by bryostatin. J. Immun., 141, 3263- 3269.
IMBODEN, J. B., WEISS, A. & STOBO, J. D. (1985). Transmembrane signalling by the T cell antigen receptor: perturbation of the T3-antigen receptor complex generates inositol phosphates and release of calcium from intracellular stores. J. exp. Med., 161, 446- 456.
JUNG, L. K., BJORNDAHL, J. M. & FU, S. M. (1988). Dissociation of TPA-induced down-regulation of T-cell antigens from protein kinase C activation. Cell. Immun., 117, 352-359.
JUNG, L. K. L. & Fu, S. M. (1988). Effects of tumor promoter 12-O-tetradecanoylphorbol-13-acetate on CD7 expression by T lineage cells. Eur. J. lmmun., 18, 711-715.
KALDJIAN, E., MCCARTHY, S.A., SHARROW, S.O., LITTMAN, D.R., KLAUSNER, R.D. & SINGER, A. (1988). Nonequivalent effects of PKC activation by PMA on murine CD4 and CD8 cell-surface expression. FASEB J., 2, 2801 - 2806.
KERN, J. A., REED, J. C., DANIELE, R. P. & NOWELL, P. C. (1986). The role of the accessory cell in mitogen-stimulated human T cell gene expression. J. Immun., 137, 764-769.

490 A . H . ESA et al.
KRAFT, A. S., SMITH, J. B. & BERKOW, R. L. (1986). Bryostatin, an activator of the calcium phospholipid-dependent protein kinase, blocks phorbol ester induced differentiation of human promyclocytic leukemia cells HL-60. Proc. natn. Acad. Sci. U.S.A., 83, 1334- 1338.
LEMEUR, M., GLANVILLE, N., MANDEL, J. L., GARLINGER, P., PALMITER, R. & CHAMBON, P. (1981). The ovalbumin gene family: hormonal control of X and Y gene transcription and mRNA accumulation. Cell, 23, 561 - 571.
MAY, W. S., SHARKIS, S. J., ESA, A. H., GEaBIA, V., KRAFT, A. S., PETTIT, G. R. & SENSEr~aREr~,TER, L. L. (1987). Antineoplastic bryostatins are multipotential simulators of human progenitor cells. Proc. hath. Acad. Sci. U.S.A., 8 4 , 8 4 8 3 - 8487.
NISHUZUKA, Y. (1984). The role of protein kinase C in cell surface signal transduction and tumor promotion. Nature, Lond., 308, 693-698.
OKAZAKI, T., KATO, Y., MOCHIZUKI, T., TASHIMI, M. SAWADA, H. & Uchino, H. (1988). Staurosporine, a novel protein kinase inhibitor, enhances HL-60 differentiation by various compounds. Expl Hemat., 16, 42-48.
PETrIT, G. R., HERALD, C. L., DOUBEK, D. I. & HERALD, D. L. (1982). Isolation and structure of Bryostatinl. J. Am. Chem. Soc., 104, 6846- 6848.
PETTIT, G. R., HERALD, C. L., KAMANO, Y., GUST, D. & AOYAGI, R. (1983). The structure of bryostatin 2 from the marine bryozoan Bugula neritina. J. natn. Prod., 46, 528-531.
PETTIT, G. R., ELEET, J. E., HERALD, C. L., KAMANO, Y., BOETTNER, F. E., BACZYNSKYJ, L. & NEIMAN, R. A. (1987). Isolation and structure of bryostatins 12 and 13. J. org. Chem., 52, 2854-2860.
TAMAOKI, T., NOMOTO, H., TAKAHASHI, I., KATO, Y., MORIMOTO, M. & TOMITA, F. (1986). Staurosporine, a potent inhibitor of phospholipid/Ca 2+ dependent protein kinase. Biochem. biophys. Res. Commun., 135, 397- 402.
TRENN, G., PETTIT, G. R., TAKYAMA, H., HU.LI, J. & SITKOVSKY, M. V. (1988). Immunomodulating properties of a novel series of protein kinase C activators: the bryostatins. J. Immun,, 140, 433-439.
WENDER, P. A., CRIBBS, C. M., KOEHLER, K. F., SHARKEY, N. A., HERALD, C. L., KAMANO, Y., PETTIT, G. R. & BLUMBERG, P. M. (1988). Modeling of the bryostatins to the phorbol ester pharmacophore on protein kinase C. Proc. natn. Acad. Sci. U.S.A., 85, 7197-7201.