accelerated features of age-related bone loss in zmpste24 metalloproteinase-deficient mice
TRANSCRIPT

Journal of Gerontology: BIOLOGICAL SCIENCES © The Author 2009. Published by Oxford University Press on behalf of The Gerontological Society of America.Cite journal as: J Gerontol A Biol Sci Med Sci All rights reserved. For permissions, please e-mail: [email protected]. 2009. Vol. 64A, No. 10, 1015–1024 Advance Access publication on July 8, 2009 doi:10.1093/gerona/glp089
1015
AGE-RELATED bone loss is the consequence of changes in hormones as well as cell number and func-
tion ( 1 ). Cellular changes include a decreasing number of bone forming osteoblasts at the expense of a higher number of adipocytes ( 2 , 3 ), thus affecting bone formation and bone quality ( 4 ). Although several studies have looked at the mo-lecular mechanisms of age-related bone loss, our knowl-edge of the factors involved in the switching from osteoblasts to adipocytes in aging bone remains limited.
The Hutchinson – Gilford progeria syndrome (HGPS) ( 5 ) is a particularly attractive example of the effect of aging on alterations in bone and fat. In addition to accelerated aging, these patients are characterized by short stature, microgna-tia, alopecia, sculptured nose, prominent scalp veins, loss of subcutaneous fat, prominent joints, skeletal hypoplasia and dysplasia, persistent open fontanelles, pathological frac-tures, hyperlipidemia, and early arteriosclerosis ( 5 , 6 ). The discovery that HGPS is caused by defective biogenesis of lamin A from prelamin A due to a mutation in the LMNA gene ( 7 ), as well as the concurrent description of similar phenotypes in mouse models of LMNA mutations ( 8 , 9 ), has increased the interest in the identifi cation of the potential role of lamins in bone biology ( 6 , 10 , 11 ).
Lamins form the lamina that keeps the nucleus shape and strength. They also play a role in a number of nuclear pro-cesses including DNA replication and transcription ( 12 ). The lamin gene family in mammals includes seven different proteins (A, A D 10, C, C2, B1, B2, and B3). Most adult mammalian somatic cells contain the three major lamins A, B1, and C. These various forms are grouped into two classes, A-type (A, A D 10, and C) and B-type (B1 and B2). Although B-type lamins are found in all nucleated somatic cells, the expression of A-type lamins is developmentally regulated
( 13 ). A-type lamins are absent from all preimplantation stage embryonic cells with their synthesis commencing at about Day 9 within the visceral endoderm and trophoblast. Subsequently, A-type lamins appear asynchronously in var-ious tissues ( 13 ).
The production of mature lamin A/C requires endoprote-olytic processing of prelamin A to mature lamin A ( 12 ). The main proteolytic enzyme involved in this process is known as Zmpste24 in mice and farnesylated proteins-converting enzyme 1 in humans ( 10 , 12 ). Lack of this enzyme would generate high levels of unprocessed prelamin A and its trun-cated form progerin/LA D 50, which are responsible for the progeria phenotype as well as the cellular defects seen in laminopathies ( 14 , 15 ). In humans, mutations in A-type lamins have been linked to several diseases including Emery-Dreifuss muscular dystrophy, partial lipodystrophy, cardiomyopathies, and HGPS ( 15 ). The common patterns observed in these pathologies, known as laminopathies, are alterations in mesenchymal stem cells (MSC) differentia-tion into adipocytes, myocytes, or osteoblasts ( 16 , 17 ).
We have recently tested the effect that absence of lamin A/C may have on the differentiation of MSC into osteo-blasts ( 18 ). Using a silencing RNA approach, we success-fully knocked down lamin A/C in a model of osteoblastogenesis. We found that lamin A/C inhibition af-fects osteoblastogenesis, whereas favors adipogenesis in vitro. In the present study, we further assessed the role of proteins of the nuclear envelope in MSC differentiation and bone quality in vivo using Zmpste24-null progeroid mice (Zmpste24 − / − ), which exhibit nuclear lamina defects and have high levels of unprocessed prelamin A ( 8 , 9 ). Al-though this model has been reported as showing low bone mineral density and spontaneous bone fractures ( 8 , 19 ), the
Accelerated Features of Age-Related Bone Loss in Zmpste24 Metalloproteinase-Defi cient Mice
Daniel Rivas , 1 Wei Li , 2 , 3 Rahima Akter , 1 Janet E. Henderson , 2 and Gustavo Duque 1 , 3
1 Lady Davis Institute and 2 JTN Wong Laboratories, McGill University, Montréal, Quebec, Canada . 3 Aging Bone Research Program, Nepean Clinical School, University of Sydney, Penrith, New South Wales, Australia .
Age-related bone loss is associated with changes in bone cellularity, which include marrow fat infi ltration and decreasing levels of osteoblastogenesis. The mechanisms that explain these changes remain unclear. Although nuclear lamina altera-tions occur in premature aging syndromes that include changes in body fat and severe osteoporosis, the role of proteins of the nuclear lamina in age-related bone loss remains unknown. Using the Zmpste24-null progeroid mice (Zmpste24 − / − ), which exhibit nuclear lamina defects and accumulate unprocessed prelamin A, we identifi ed several alterations in bone cellularity in vivo. We found that defective prelamin A processing induced accelerated features of age-related bone loss including lower osteoblast and osteocyte numbers and higher levels of marrow adipogenesis. In summary, processing of prelamin A could become a new approach to regulate osteoblastogenesis and bone turnover and thus for the prevention and treatment of senile osteoporosis.
Key Words : Lamin A/C — Osteoporosis — Mesenchymal stem cells — Osteoblastogenesis — Adipogenesis .
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

RIVAS ET AL. 1016
cellular mechanisms that explain this decrease in bone mass remain unknown. In addition, the elucidation of the poten-tial mechanisms that explain this bone loss in a model of progeria mice could provide new potential anabolic thera-peutic targets for senile osteoporosis.
Our results demonstrate that Zmpste24 − / − mice are osteopenic with low bone turnover and concomitant high levels of marrow fat, which are the hallmarks of age-related bone loss. Taken together, these data confi rm previous in vitro data on the role of proteins of the nuclear envelope in MSC differentiation and opens the path for the development of anabolic drugs that maintain the bone mass through the regulation of prelamin A processing in MSC.
Methods
Animals Zmpste24 +/ − mice were kindly provided by Dr. Carlos
Lopez-Otin from University of Oviedo ( 9 ). A colony of Zmpste24 − / − mice and their corresponding wild-type (WT +/+ ) controls was developed at the Lady Davis Institute for Medical Research (McGill University). Genotype was verifi ed by real-time polymerase chain reaction (RT-PCR). Male and female mice were housed in cages in a limited access room. Animal husbandry adhered to Canadian Coun-cil on Animal Care Standards. All protocols were approved by the McGill University Health Center Animal Care Utili-zation Committee. Mice (male and female) were killed at 3 months of age. Both side tibiae and femora were obtained for further analysis. All experiments and measurements were repeated at least three times.
Quantitative Radiological Imaging Micro-computed tomography (Micro-CT) was per-
formed, using a modifi cation of previously published meth-ods ( 20 ), on the left femur after removal of soft tissues and overnight fi xation in 4% paraformaldehyde. A Skyscan 1172 instrument (Skyscan, Antwerp, Belgium) equipped with a 1.3 Mp camera was used to capture two-dimensional (2D) serial cross-sections, which were used to reconstruct three-dimensional images for the quantifi cation of the vol-ume of bone in the distal metaphysis. Bone microarchitec-ture measurements were assessed using a set of calibrated phantoms purchased from Skyscan.
Histology and Histomorphometry For histomorphometric analyses, the left femur was fi xed
overnight in 4% paraformaldehyde, rinsed in three changes of phosphate-buffered saline (PBS), and embedded in poly-methyl methacrylate (MMA) or a mixture of 50% MMA and 50% glycol methacrylate (GMA). Serial 4- to 6- m m sections of MMA-embedded tissues were left unstained or stained with von Kossa, whereas 4- m m MMA-GMA sections were
stained for alkaline phosphatase (ALP; osteoblasts), toluidine blue (osteocytes), and tartrate resistance acid phosphatase (TRAP; osteoclasts) activity as described previously ( 20 ). Images were captured using a Leica DMR microscope (Leica Microsystems, Richmond Hill, Ontario, Canada) equipped with a Retiga 1300 camera (Qimaging, Burnaby, British Columbia, Canada) and the primary histomorphometric data obtained using Bioquant Nova Prime image analysis software (Bioquant Image Analysis Corp., Nashville, TN). Nomencla-ture and abbreviations conform to those recommended by the American Society for Bone and Mineral Research ( 21 ).
For marrow fat analysis, the right femur was cleaned of soft tissue, fi xed for 16 hours in 4% paraformaldehyde, rinsed thoroughly in PBS, decalcifi ed in 10% ethylenedi-aminetetraacetic acid (EDTA), and processed for paraffi n embedding. Serial 4- m m sections were cut on a modifi ed Leica RM 2155 rotary microtome (Leica Microsystems). Fat volume (FV) was quantifi ed using von Kossa-stained sections as previously described ( 22 ).
Serum Biochemistry and Bone Biomarkers Serum levels of calcium were determined at the Rodent Di-
agnostics Lab (McGill University, Montréal, Quebec, Canada) using routine automated techniques. Bone biomarkers, calcio-tropic, and sex hormones were analyzed at the Centre for Bone and Periodontal Research using commercial assays for serum C-telopeptides (C-Tx; Nordic Bioscience, Herlev, Denmark), 17 b -estradiol and testosterone (IBL Immuno-Biological, Hamburg, Germany), osteocalcin (OCN; ImmunoDiagnostic Systems Ltd, Fountain Hills, AZ), parathyroid hormone (PTH; Immunotopics Inc., San Clemente, CA), and 25(OH)-vitamin D (ImmunoDiagnostic Systems Ltd).
Ex vivo Cultures of Bone Marrow Cells To establish adherent bone marrow cultures, bone marrow
cells were obtained and induced to differentiate into osteo-blast as previously described ( 23 ). Briefl y, both tibiae from 3-month-old Zmpste24 − / − and WT +/+ ( n = 10 per group) were fl ushed using a 21-gauge needle attached to a 10-mL syringe fi lled with Dulbecco ’ s modifi ed Eagle ’ s Medium (DMEM; GIBCO BRL, Gaithersburg, MD). Cells from both tibiae were fi ltered through a cell strainer with 70- m nylon mesh (BD Bioscience, Bedford, MA) and then combined to pro-duce a volume of 2 mL containing approximately 10 7 cells/mL. Six -well plate cultures were then established in triplicate, with each well containing a 100- m L aliquot of cell suspension combined with 4 mL of fresh minimal essential medium. The cells were incubated in MSC growth media at 37°C with 5% humidifi ed CO 2 and isolated by their adherence to tissue culture plastic. Medium was aspirated and replaced with fresh medium to remove nonadherent cells every 2 – 3 days. The adherent MSC were grown to approximately 80% confl uence for about 7 days defi ned as MSC at passage 0, harvested with 0.25% trypsin and 1 mM EDTA for 5 minutes at 37°C,
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

ACCELERATED BONE LOSS IN PROGERIA MICE 1017
diluted 1:3 in MSC growth media, plated, and grown to confl uence for further expansion. After second and third passages, MSC were used for subsequent experiments.
To induce differentiation, a total of 10 4 cells were diluted in osteogenic medium (prepared with DMEM, 10% fetal calf serum [FCS], 0.2 mM dexamethasone, 10 mmol/L b -glycerol phosphate, and 50 m g/mL ascorbic acid) and plated in six-well plates. Media was aspirated and replaced with fresh osteogenic medium every 3 days. After 21 days in culture, cells were washed with PBS, ethanol fi xed, stained for ALP, and counterstained with hematoxylin (Sigma, Oakville, Ontario, Canada). The colonies with more than 10% of cells staining positive for ALP were con-sidered as colony-forming units – osteoblasts (CFU-OB).
For adipocyte differentiation, cells were obtained and plated as previously described. After passage 3, 10 4 MSC were diluted in adipogenic medium (prepared with DMEM, 4.5 g/L glucose, 1 m M dexamethasone, 0.2 mM indometha-cin, 1.7 m M insulin, 0.5 mM 3-isobutyl-1-methylxanthine, 10% FCS, 0.05 U/mL penicillin, and 0.05 m g/mL strepto-mycin) and plated in six-well plates for 3 days. To promote adipogenic phenotype, cells were then incubated for 3 days in adipogenesis maintenance medium (DMEM, 4.5 g/L glu-cose, 1.7 m M insulin, 10% fetal bovine serum, 0.05 U/mL penicillin, and 0.05 m g/mL streptomycin) and then switched to induction media again. After 3 cycles of induction or maintenance (Day 18), media was aspirated and cells were fi xed and adipocytes visualized by staining with Oil Red O and counterstaining with hematoxylin. Colonies with more than 10% of cells staining positive for Oil Red O were con-sidered as CFU – adipocytes (CFU-AD).
Semi-Quantitative RT-PCR Bone marrow cells were fl ushed and isolated as previ-
ously described. Total RNA was isolated from MSC using Trizol reagent (Invitrogen, Carlsbad, CA). Total RNA (1 m g) was reverse transcribed with either random hexamers or oligo-dT using Qiagen One Step RT-PCR Enzyme Mix (Qiagen, Valencia, CA). The resulting complementary DNA was amplifi ed by 35 PCR cycles with an annealing tempera-ture of 58°C. Primers for specifi c detection of messenger RNA (mRNA) expression included the following: Zmp-ste24 (F: 5 ′ -AGTCCGATCAGAGTGGGTTG-3 ′ and R: 5 ′ GGTGAAG GGGA CAGTGAAGA-3 ′ ), runt-related tran-scription factor 2 (Runx2; F: 5 ′ -ACAGGAAGTTGGG-ACTGTCG-3 ′ and R: 5 ′ -GAGGGACTA TGGCGTCAAAC-3 ′ ), osteopontin (OPN; F: 5 ′ -AGGTCCTCATCCGTGGCATC-3 ′ and R: 5 ′ -TGCAC CCAGATCCTATAGCC-3 ′ ); bone sialo-protein II (BSPII; F: 5 ′ -GTTCCTTCTGCACCTGCTTC-3 ′ and R: 5 ′ -GGGAGGC AGTGACTCTTCAG-3 ′ ); OCN (F: 5 ′ -ACTTGCAGGGCAGA GAGAGA-3 ′ and R: 5 ′ -CTGACCT-CACAGATGCCAAG-3 ′ ), CCAAT enhancer-binding pro-tein- a (C/EBP a ; F: 5 ′ -CAGTTTGG CAAGAATCAGAGCA-3 ′ and R: 5 ′ -GGGT GAGTTCATGGAGAATGG-3 ′ ), and per-
oxisome proliferator-activated receptor gamma 2 (PPAR g 2; F: 5 ′ -CCAACAGC TTCTCCTTCTCG-3 ′ and R: 5 ′ -CGAGT CTGT GGGGATA AAGC-3 ′ ). Amplifi cation of glyceraldehydes-3-phosphate dehydrogenase (GAPDH) was used as control. The expected sizes of the amplifi cation products were between 400 and 800 bp. Amplifi ed products were analyzed by 2% agarose gel electrophoresis. The signals were quantifi ed by densitometry (Bio-Rad laboratories, Her-cules, CA) and normalized according to GAPDH density.
Western Blot Analysis For Western blot analysis, 20 m g of total protein were
loaded on polyacrylamide gels and separated by standard sodium dodecyl sulfate – polyacrylamide gel electrophore-sis. To control for differences in gel migration, exposure time, antibody incubation, etc., samples were run on the same gels and transferred to the same polyvinylidene fl uo-ride membranes (Amersham, Little Chalfont, UK). Blots were blocked overnight in 2.5% nonfat dried milk and probed with antibodies directed against Zmpste24, Runx2, OPN, BSPII, OCN, C/EBP a , and PPAR g 2 (1:1000; Santa Cruz Biotechnology, Santa Cruz, CA). Positive controls were included in all experiments as provided by the manu-facturer (Santa Cruz Biotechnology), to confi rm antibody specifi city. Secondary antibodies conjugated to horseradish peroxidase were from Sigma (1:5000). Antigen – antibody complexes were detected by chemiluminescence using a kit of reagents from ECL (Amersham), and blots were exposed to high-performance chemiluminescence fi lm (Amersham). Films were scanned and the optical density of each specifi c band was analyzed using the ImageMaster program and expressed as OD/mm 2 /100 m g of total protein. Relative intensity of the samples was determined comparing the protein of interest in the mutant mice using the values of WT as controls (100%). Values are reported as the average of samples obtained from 8 to 10 mice.
Statistical Analysis Results are expressed as means ± SD and differences be-
tween groups of mice were determined by the Student ’ s t test and the Mann – Whitney test. A p value of ≤ .05 was considered statistically signifi cant.
Results
Accumulation of Unprocessed Prelamin A Affects Bone Mass and Reduces Bone Quality
Using a model of Zmpste24-null progeroid mice (Zmp-ste24 − / − ) ( 8 ), we analyzed whether accumulation of unpro cessed prelamin A has an effect on bone mass and mi-croarchitecture and whether gender differences are found. Figure 1 shows a qualitative (A – E) and quantitative (F – H) decline in bone density and architecture in both male and
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
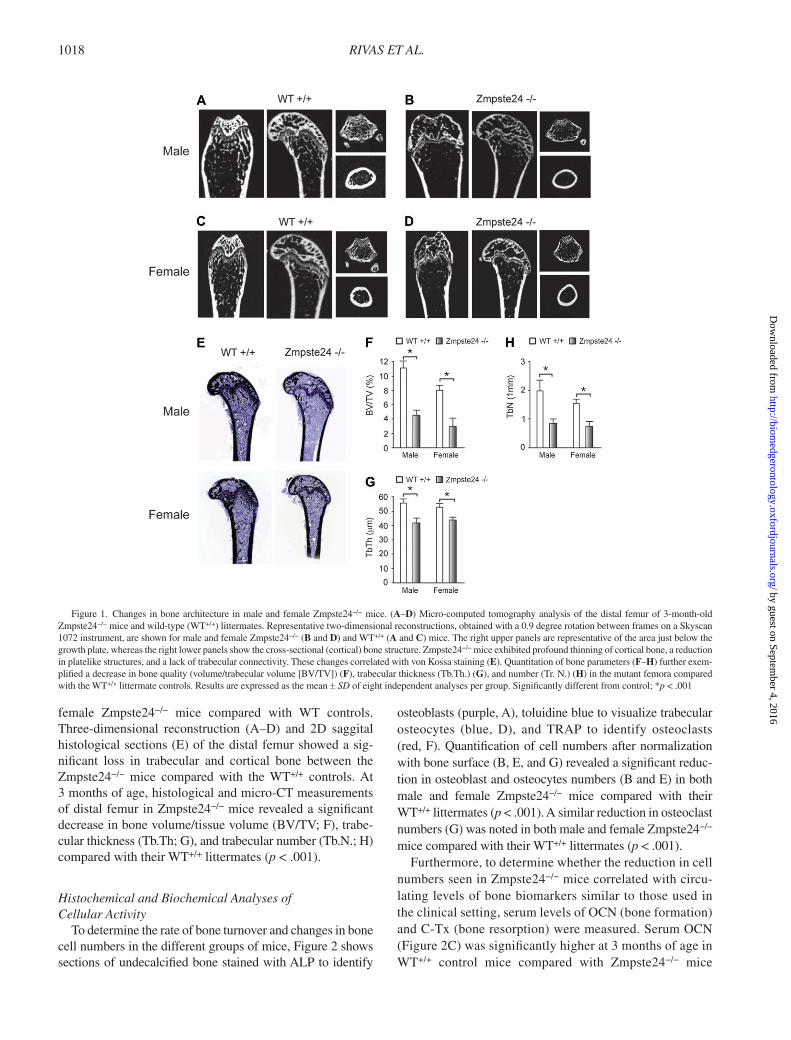
RIVAS ET AL. 1018
female Zmpste24 − / − mice compared with WT controls. Three-dimensional reconstruction (A – D) and 2D saggital histological sections (E) of the distal femur showed a sig-nifi cant loss in trabecular and cortical bone between the Zmpste24 − / − mice compared with the WT +/+ controls. At 3 months of age, histological and micro-CT measurements of distal femur in Zmpste24 − / − mice revealed a signifi cant decrease in bone volume/tissue volume (BV/TV; F), trabe-cular thickness (Tb.Th; G), and trabecular number (Tb.N.; H) compared with their WT +/+ littermates ( p < .001).
Histochemical and Biochemical Analyses of Cellular Activity
To determine the rate of bone turnover and changes in bone cell numbers in the different groups of mice, Figure 2 shows sections of undecalcifi ed bone stained with ALP to identify
osteoblasts (purple, A), toluidine blue to visualize trabecular osteocytes (blue, D), and TRAP to identify osteoclasts (red, F). Quantifi cation of cell numbers after normalization with bone surface (B, E, and G) revealed a signifi cant reduc-tion in osteoblast and osteocytes numbers (B and E) in both male and female Zmpste24 − / − mice compared with their WT +/+ littermates ( p < .001). A similar reduction in osteoclast numbers (G) was noted in both male and female Zmpste24 − / − mice compared with their WT +/+ littermates ( p < .001).
Furthermore, to determine whether the reduction in cell numbers seen in Zmpste24 − / − mice correlated with circu-lating levels of bone biomarkers similar to those used in the clinical setting, serum levels of OCN (bone formation) and C-Tx (bone resorption) were measured. Serum OCN ( Figure 2C ) was signifi cantly higher at 3 months of age in WT +/+ control mice compared with Zmpste24 − / − mice
Figure 1. Changes in bone architecture in male and female Zmpste24 − / − mice. ( A – D ) Micro-computed tomography analysis of the distal femur of 3-month-old Zmpste24 − / − mice and wild-type (WT +/+ ) littermates. Representative two-dimensional reconstructions, obtained with a 0.9 degree rotation between frames on a Skyscan 1072 instrument, are shown for male and female Zmpste24 − / − ( B and D ) and WT +/+ ( A and C ) mice. The right upper panels are representative of the area just below the growth plate, whereas the right lower panels show the cross-sectional (cortical) bone structure. Zmpste24 − / − mice exhibited profound thinning of cortical bone, a reduction in platelike structures, and a lack of trabecular connectivity. These changes correlated with von Kossa staining ( E ). Quantitation of bone parameters ( F – H ) further exem-plifi ed a decrease in bone quality (volume/trabecular volume [BV/TV]) ( F ), trabecular thickness (Tb.Th.) ( G ), and number (Tr. N.) ( H ) in the mutant femora compared with the WT +/+ littermate controls. Results are expressed as the mean ± SD of eight independent analyses per group. Signifi cantly different from control; * p < .001
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

ACCELERATED BONE LOSS IN PROGERIA MICE 1019
( p < .01), correlating with lower levels of osteoblast num-bers found in the histochemical analysis. In addition, Fig-ure 2H shows a signifi cant decrease in serum C-Tx in Zmpste24 − / − mice compared with their WT +/+ controls ( p < .01), also correlating with the lower levels of osteoclast numbers found in the histochemical analysis.
Circulating Concentrations of Calciotropic and Sex Steroid Hormones in Zmpste24 − / − Mice
To determine whether the changes in bone cellularity and microarchitecture were due to alterations in serum concentra-
tions of calciotropic and sex steroids, we measured serum lev-els of PTH, 25-dihydroxy(OH)-vitamin D, calcium, estradiol, and testosterone in both male and female Zmpste24 − / − and compared them with their WT +/+ controls. As shown in Table 1 , there were no differences in serum levels of PTH, 25(OH)-vitamin D, or calcium between Zmpste24 − / − and WT +/+ controls. As expected, levels of estradiol were higher in female than in male mice with no differences found between both groups. Similar results were found in testosterone levels where male have higher levels of testosterone without differ-ences found between Zmpste24 − / − and their WT +/+ controls.
Figure 2. Changes in bone cellularity of 3-month-old Zmpste24 − / − mice. Sections of plastic embedded tibiae from Zmpste24 − / − and wild-type (WT +/+ ) were stained sequentially for alkaline phosphatase (ALP) (osteoblasts, OB) ( A , arrows, magnifi cation ×20), toluidine blue (osteocytes, Ost) ( B , arrows, magnifi cation ×40), and tartrate resistance acid phosphatase (TRAP) (osteoclasts, OC) ( F , arrows, magnifi cation ×20). A signifi cant decrease in the number of ALP expressing OB ( B ) and OC showing TRAP enzyme activity ( E ) was seen in Zmpste24 − / − mice compared with WT +/+ mice. In addition, the number of Ost was signifi cantly reduced in Zmpste24 − / − mice compared with WT +/+ mice ( E ). Scale bar represents 50 m m for A and F and 25 m m for D. Micrographs are representative of those from fi ve differ-ent mice of each genotype. * p < .01 and ** p < .001. ( C and H ) Changes in bone cellularity correlated with changes in serum biochemical markers of bone formation ( C , OCN: osteocalcin) and bone resorption ( H , C-Tx: C-telopeptides) in Zmpste24 − / − mice compared with WT +/+ mice. * p < .01
Table 1. Circulating Concentrations of Calcium, Calciotropic (PTH and 25(OH)-vitamin D) and Sex Steroid Hormones in Zmpste24 − / − Mice Versus WT +/+ Controls
Assay
Male Female
WT +/+ Zmpste24 − / − p WT +/+ Zmpste24 − / − p
Ca 2+ (mmol/L) 2 ± 0.07 2.2 ± 0.08 NS 2.2 ± 0.1 2.3 ± 0.07 NS 25(OH)-vitamin D (nmol/L) 88 ± 13 107 ± 13 NS 107 ± 20 108 ± 21 NS PTH (pg/mL) 56 ± 22 36 ± 13 NS 101 ± 68 45 ± 27 NS Testosterone (ng/mL) 4.8 ± 0.2 5.2 ± 0.5 NS 0.3 ± 0.2 0.4 ± 0.1 NS Estradiol (pg/mL) 2.6 ± 0.4 1.8 ± 0.4 NS 9.8 ± 4 6.6 ± 3 NS
Note : NS = non signifi cant; PTH = parathyroid hormone; WT +/+ = wild-type.
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
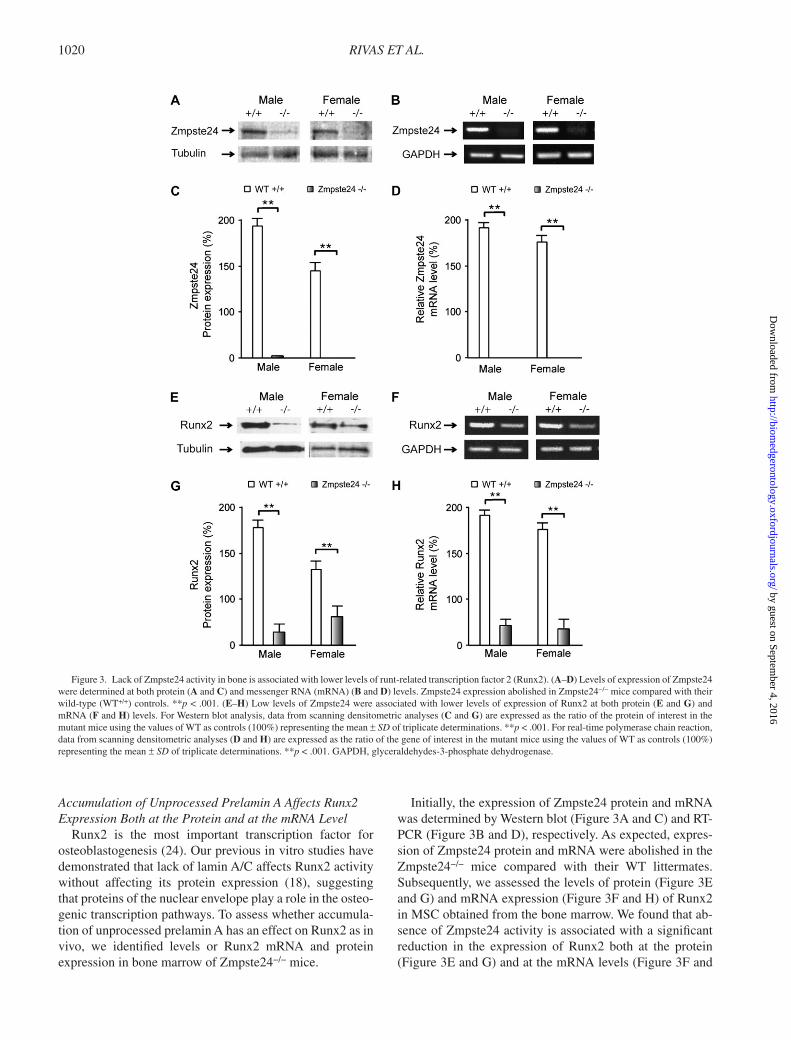
RIVAS ET AL. 1020
Accumulation of Unprocessed Prelamin A Affects Runx2 Expression Both at the Protein and at the mRNA Level
Runx2 is the most important transcription factor for osteoblastogenesis ( 24 ). Our previous in vitro studies have demonstrated that lack of lamin A/C affects Runx2 activity without affecting its protein expression ( 18 ), suggesting that proteins of the nuclear envelope play a role in the osteo-genic transcription pathways. To assess whether accumula-tion of unprocessed prelamin A has an effect on Runx2 as in vivo, we identifi ed levels or Runx2 mRNA and protein expression in bone marrow of Zmpste24 − / − mice.
Initially, the expression of Zmpste24 protein and mRNA was determined by Western blot ( Figure 3A and C ) and RT-PCR ( Figure 3B and D ), respectively. As expected, expres-sion of Zmpste24 protein and mRNA were abolished in the Zmpste24 − / − mice compared with their WT littermates. Subsequently, we assessed the levels of protein ( Figure 3E and G ) and mRNA expression ( Figure 3F and H ) of Runx2 in MSC obtained from the bone marrow. We found that ab-sence of Zmpste24 activity is associated with a signifi cant reduction in the expression of Runx2 both at the protein ( Figure 3E and G ) and at the mRNA levels ( Figure 3F and
Figure 3. Lack of Zmpste24 activity in bone is associated with lower levels of runt-related transcription factor 2 (Runx2). ( A – D ) Levels of expression of Zmpste24 were determined at both protein ( A and C ) and messenger RNA (mRNA) ( B and D ) levels. Zmpste24 expression abolished in Zmpste24 − / − mice compared with their wild-type (WT +/+ ) controls. ** p < .001. ( E – H ) Low levels of Zmpste24 were associated with lower levels of expression of Runx2 at both protein ( E and G ) and mRNA ( F and H ) levels. For Western blot analysis, data from scanning densitometric analyses ( C and G ) are expressed as the ratio of the protein of interest in the mutant mice using the values of WT as controls (100%) representing the mean ± SD of triplicate determinations. ** p < .001. For real-time polymerase chain reaction, data from scanning densitometric analyses ( D and H ) are expressed as the ratio of the gene of interest in the mutant mice using the values of WT as controls (100%) representing the mean ± SD of triplicate determinations. ** p < .001. GAPDH, glyceraldehydes-3-phosphate dehydrogenase.
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
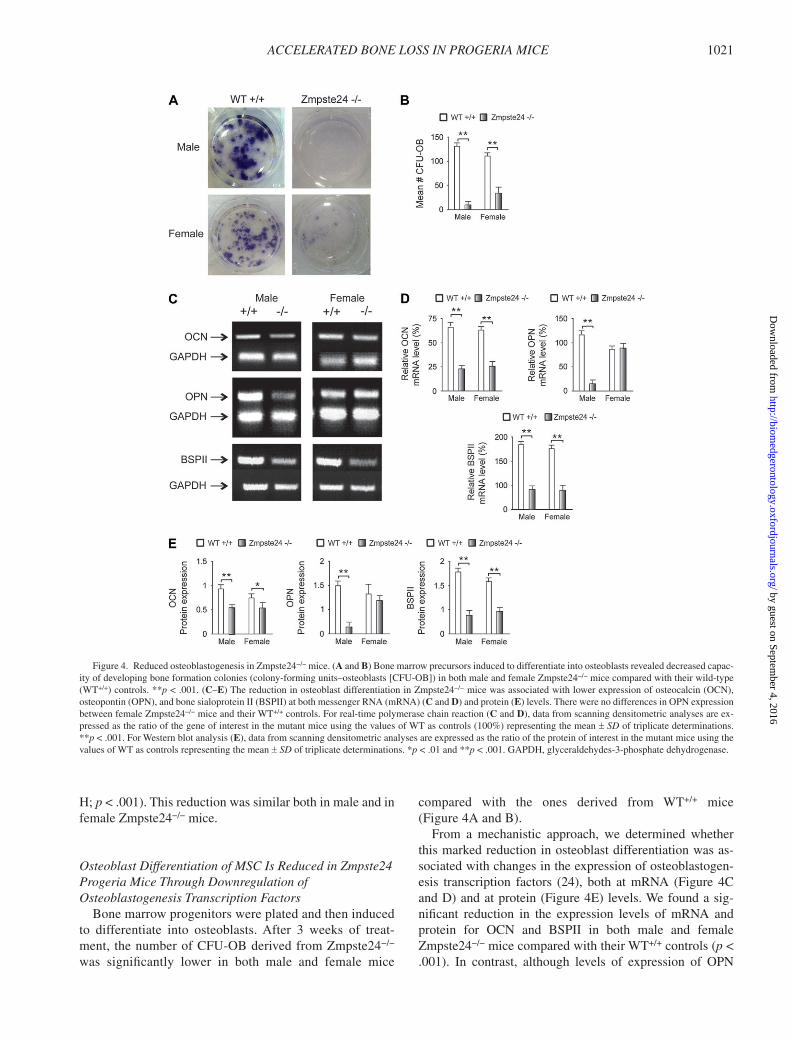
ACCELERATED BONE LOSS IN PROGERIA MICE 1021
H ; p < .001). This reduction was similar both in male and in female Zmpste24 − / − mice.
Osteoblast Differentiation of MSC Is Reduced in Zmpste24 Progeria Mice Through Downregulation of Osteoblastogenesis Transcription Factors
Bone marrow progenitors were plated and then induced to differentiate into osteoblasts. After 3 weeks of treat -ment, the number of CFU-OB derived from Zmpste24 − / − was signifi cantly lower in both male and female mice
compared with the ones derived from WT +/+ mice ( Figure 4A and B ).
From a mechanistic approach, we determined whether this marked reduction in osteoblast differentiation was as-sociated with changes in the expression of osteoblastogen-esis transcription factors ( 24 ), both at mRNA ( Figure 4C and D ) and at protein ( Figure 4E ) levels. We found a sig-nifi cant reduction in the expression levels of mRNA and protein for OCN and BSPII in both male and female Zmpste24 − / − mice compared with their WT +/+ controls ( p < .001). In contrast, although levels of expression of OPN
Figure 4. Reduced osteoblastogenesis in Zmpste24 − / − mice. ( A and B ) Bone marrow precursors induced to differentiate into osteoblasts revealed decreased capac-ity of developing bone formation colonies (colony-forming units – osteoblasts [CFU-OB]) in both male and female Zmpste24 − / − mice compared with their wild-type (WT +/+ ) controls. ** p < .001. ( C – E ) The reduction in osteoblast differentiation in Zmpste24 − / − mice was associated with lower expression of osteocalcin (OCN), osteopontin (OPN), and bone sialoprotein II (BSPII) at both messenger RNA (mRNA) ( C and D ) and protein ( E ) levels. There were no differences in OPN expression between female Zmpste24 − / − mice and their WT +/+ controls. For real-time polymerase chain reaction ( C and D ), data from scanning densitometric analyses are ex-pressed as the ratio of the gene of interest in the mutant mice using the values of WT as controls (100%) representing the mean ± SD of triplicate determinations. ** p < .001. For Western blot analysis ( E ), data from scanning densitometric analyses are expressed as the ratio of the protein of interest in the mutant mice using the values of WT as controls representing the mean ± SD of triplicate determinations. * p < .01 and ** p < .001. GAPDH, glyceraldehydes-3-phosphate dehydrogenase.
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
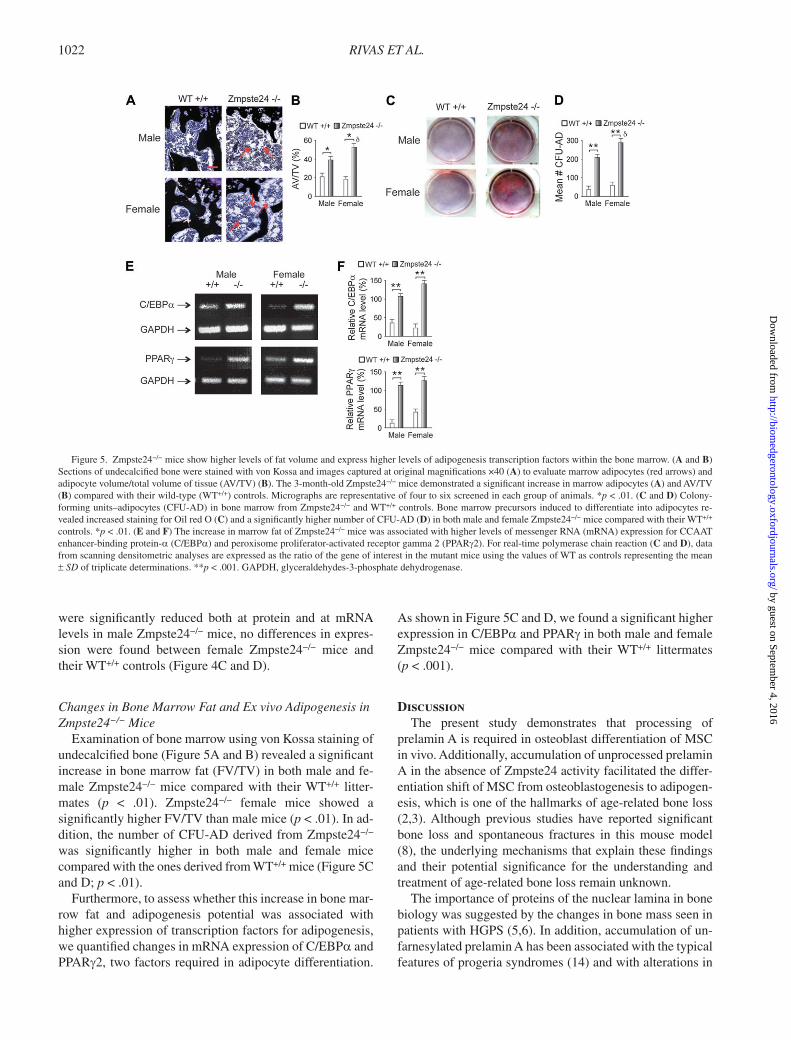
RIVAS ET AL. 1022
were signifi cantly reduced both at protein and at mRNA levels in male Zmpste24 − / − mice, no differences in expres-sion were found between female Zmpste24 − / − mice and their WT +/+ controls ( Figure 4C and D ).
Changes in Bone Marrow Fat and Ex vivo Adipogenesis in Zmpste24 − / − Mice
Examination of bone marrow using von Kossa staining of undecalcifi ed bone ( Figure 5A and B ) revealed a signifi cant increase in bone marrow fat (FV/TV) in both male and fe-male Zmpste24 − / − mice compared with their WT +/+ litter-mates ( p < .01). Zmpste24 − / − female mice showed a signifi cantly higher FV/TV than male mice ( p < .01). In ad-dition, the number of CFU-AD derived from Zmpste24 − / − was signifi cantly higher in both male and female mice compared with the ones derived from WT +/+ mice ( Figure 5C and D ; p < .01).
Furthermore, to assess whether this increase in bone mar-row fat and adipogenesis potential was associated with higher expression of transcription factors for adipogenesis, we quantifi ed changes in mRNA expression of C/EBP a and PPAR g 2, two factors required in adipocyte differentiation.
As shown in Figure 5C and D , we found a signifi cant higher expression in C/EBP a and PPAR g in both male and female Zmpste24 − / − mice compared with their WT +/+ littermates ( p < .001).
Discussion The present study demonstrates that processing of
prelamin A is required in osteoblast differentiation of MSC in vivo. Additionally, accumulation of unprocessed prelamin A in the absence of Zmpste24 activity facilitated the differ-entiation shift of MSC from osteoblastogenesis to adipogen-esis, which is one of the hallmarks of age-related bone loss ( 2 , 3 ). Although previous studies have reported signifi cant bone loss and spontaneous fractures in this mouse model ( 8 ), the underlying mechanisms that explain these fi ndings and their potential signifi cance for the understanding and treatment of age-related bone loss remain unknown.
The importance of proteins of the nuclear lamina in bone biology was suggested by the changes in bone mass seen in patients with HGPS ( 5 , 6 ). In addition, accumulation of un-farnesylated prelamin A has been associated with the typical features of progeria syndromes ( 14 ) and with alterations in
Figure 5. Zmpste24 − / − mice show higher levels of fat volume and express higher levels of adipogenesis transcription factors within the bone marrow. ( A and B ) Sections of undecalcifi ed bone were stained with von Kossa and images captured at original magnifi cations ×40 ( A ) to evaluate marrow adipocytes (red arrows) and adipocyte volume/total volume of tissue (AV/TV) ( B ). The 3-month-old Zmpste24 − / − mice demonstrated a signifi cant increase in marrow adipocytes ( A ) and AV/TV ( B ) compared with their wild-type (WT +/+ ) controls. Micrographs are representative of four to six screened in each group of animals. * p < .01. ( C and D ) Colony-forming units – adipocytes (CFU-AD) in bone marrow from Zmpste24 − / − and WT +/+ controls. Bone marrow precursors induced to differentiate into adipocytes re-vealed increased staining for Oil red O ( C ) and a signifi cantly higher number of CFU-AD ( D ) in both male and female Zmpste24 − / − mice compared with their WT +/+ controls. * p < .01. ( E and F ) The increase in marrow fat of Zmpste24 − / − mice was associated with higher levels of messenger RNA (mRNA) expression for CCAAT enhancer-binding protein- a (C/EBP a ) and peroxisome proliferator-activated receptor gamma 2 (PPAR g 2). For real-time polymerase chain reaction ( C and D ), data from scanning densitometric analyses are expressed as the ratio of the gene of interest in the mutant mice using the values of WT as controls representing the mean ± SD of triplicate determinations. ** p < .001. GAPDH, glyceraldehydes-3-phosphate dehydrogenase.
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

ACCELERATED BONE LOSS IN PROGERIA MICE 1023
the differentiation of MSC during the aging process ( 16 , 25 ). However, despite increasing evidence suggesting that either accumulation of unprocessed prelamin A or lower levels of lamin A/C may play a role in normal and pathological ag-ing, few studies have been pursued looking at their associa-tion with the cellular changes of aging bone.
A recent study has reported that levels of lamin A/C ex-pression are decreased during normal aging in osteoblasts ( 26 ). This reduction in lamin A/C expression by mature os-teoblasts could be associated with functional decline in osteo-blast function seen in normal aging bone. From the osteoblast differentiation perspective, the role of proteins of the nuclear lamina in osteoblast differentiation has been previously as-sessed in vitro ( 18 ). Lamin A/C knockdown decreased osteoblast differentiation, whereas it facilitated adipocyte differentiation of MSC. Interestingly, absence of lamin A/C affected the activation of the Runx2 nuclear binding complex without affecting Runx2 expression, subsequently affecting the expression of osteoblastogenesis transcription factors.
In this study, we assessed whether the low bone mass and spontaneous fractures found in the Zmpste24 − / − mice were due to the same mechanisms previously found in our ex-periments in vitro. In addition, we looked whether these abnormalities were similar in both sexes. Initially, we per-formed both micro-CT and histological analyses of bones of Zmpste24 − / − mice at the age of 3 months. This time point was selected due to the short life span of the homozygous mice and to previous reports that, after this age, Zmpste24 − / − mice start suffering spontaneous fractures ( 8 ).
Initially, von Kossa staining and micro-CT demonstrated that bone mass is signifi cantly lower in the Zmpste24 − / − mice compared with WT control mice, leading to an osteopenic phenotype. This bone loss is refl ected by a decrease in trabe-cular thickness and number, which were decreased to the same extent in both male and female Zmpste24 − / − mice. Fur-thermore, to analyze the cellular mechanisms of bone loss in this model, we assessed the changes in bone cellularity in Zmpste24 − / − mice compared with their WT littermates. We found that the number of osteoblasts and osteoclasts relative to the bone volume was signifi cantly reduced in Zmpste24 − / − mice compared with the WT controls. This reduction in cells number correlated with serum markers of osteoblastic (OCN) and osteoclastic (C-telopeptide) activities. No difference be-tween male and female was found. Overall, these changes in bone cellularity were not associated with changes in serum levels of either calcium or calciotropic and sex steroid hor-mones. Taken together, these results demonstrate that accu-mulation of unprocessed prelamin A induces bone loss through a signifi cant reduction in bone turnover. However, osteoblas-tic activity was more affected than osteoclastic activity, sug-gesting that processing of prelamin A may play a more important role in the regulation of osteoblast differentiation and function and that the reduction in osteoclast activity could be a consequence of lower number of osteoblast precursors which regulate osteoclast differentiation. Our fi ndings corre-
late with a recent study looking at the effect of high levels of prelamin A on osteoclast activity in vitro ( 27 ). The investiga-tors reported that the bone resorption activity of osteoclasts obtained in the presence of high prelamin A levels is lower with respect to control osteoclasts. Nevertheless, further in vivo studies looking at the role of proteins of the nuclear lam-ina in osteoclast differentiation and function will be required.
To further assess whether the reduction in osteoblast number was associated with alterations in Runx2 expression and activ-ity, we quantifi ed levels of Runx2 expression both at the protein and at the mRNA levels. We found that prelamin A accumula-tion induced a signifi cant reduction in Runx2 at both protein and mRNA levels. Considering that Runx2 is an essential fac-tor in osteoblastogenesis, we further assessed whether low levels of Runx2 in this model has an effect on osteoblast dif-ferentiation and on the expression of transcription factors downstream Runx2 activation ( 24 , 28 ). We found that MSC obtained from bone marrow in both male and female Zmp-ste24 − / − mice lost their capacity to differentiate into osteoblasts and expressed lower levels of the osteoblastogenesis transcrip-tion factors OCN and BSPII. Interestingly, whereas levels of OPN expression remained stable in female Zmpste24 − / − mice, male Zmpste24 − / − mice showed lower levels of OPN expres-sion suggesting that estrogens may play a protective effect to OPN expression in these mice. The mechanism involved in this gender-based difference should be assessed in further studies.
Furthermore, considering that osteocytes are the most abundant cell in bone, act as mechanosensors, and consti-tute the fi nal fate of a proportion of osteoblasts ( 29 ), we looked at the effect that lack of Zmpste24 activity may have on osteocytes number in this model. We found a signifi cant reduction in trabecular osteocyte number in Zmpste24 − / − mice compared with their WT +/+ controls. More studies looking at the role of proteins of the nuclear lamina in osteocyte function and mechanotransduction are required.
Finally, a particularly striking fi nding of this study was that the signifi cant bone loss in Zmpste24 − / − mice was associated with higher levels of fat infi ltration within the bone marrow. This evidence correlates with previous in vitro fi ndings show-ing that lack of lamin A/C favors the differentiation of MSC into adipocytes ( 18 ). In fact, lack of lamin A/C and accumula-tion of unfarnesylated prelamin A have been associated with lipodystrophy or redistribution of fat to nonadipose tissues ( 30 , 31 ). In this study, the abnormally increased amount of bone marrow fat correlated with higher levels of expression of proadipogenic transcription factors in both male and fe-male Zmpste24 − / − mice. Considering that age-related bone loss constitutes a type of lipodystrophy where fat is redistrib-uted into the bone marrow compartment ( 2 ), the regulation of the proteins of the lamina may play an important role in the prevention of this age-related phenomenon.
In summary, our fi ndings provide new insights into the role of prelamin A processing in bone quality and MSC differen-tiation. Our fi ndings allow us to postulate that prelamin A processing could be a determinant player in the pathogenesis
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

RIVAS ET AL. 1024
of age-related bone loss. Considering that levels of lamin A/C expression in bone progressively decrease with aging, it is tempting to propose that age-related changes in the process-ing of proteins of the nuclear lamina are associated with the key features of senile osteoporosis, namely decreasing osteo-blastogenesis, low bone turnover, and increasing adipogene-sis. In addition, recent studies have demonstrated that some of the pathological fi ndings associated with accumulation of unprocessed prelamin A could be reverted through the inhibi-tion of prenylation ( 32 , 33 ). Overall, we conclude that the pharmacological modulation of prelamin A and lamin A/C activities in bone could offer a new potential target to prevent bone loss thus maintaining appropriate levels of bone for-mation, whereas decreasing fat infi ltration within the bone marrow.
Funding
Grants from the Canadian Institutes of Health Research (2007031AP) and the Nepean Hospital Medical Research Foundation. G.D. holds a Fel-lowship from the University of Sydney Medical Research Foundation.
Acknowledgment
The authors would like to thank Mrs. Leigh Bambury for preparation of the manuscript.
Correspondence
Address correspondence to Gustavo Duque, MD, PhD, Aging Bone Research Program, Nepean Clinical School, University of Sydney, Level 5, South Block, Nepean Hospital, Penrith, NSW 2750, Australia. Email: [email protected]
References 1. Chan GK , Duque G . Age-related bone loss: old bone, new facts .
Gerontology . 2002 ; 48 : 62 – 71 . 2. Duque G , Troen BR . Understanding the mechanisms of senile osteo-
porosis: new facts for a major geriatric syndrome . J Am Geriatr Soc . 2008 ; 56 : 935 – 941 .
3. Gimble JM , Zvonic S , Floyd ZE , et al . Playing with bone and fat . J Cell Biochem . 2006 ; 98 : 251 – 266 .
4. Rosen CJ , Bouxsein ML . Mechanisms of disease: is osteoporosis the obesity of bone? Nat Clin Pract Rheumatol . 2006 ; 2 : 35 – 43 .
5. Merideth MA , Gordon LB , Clauss S , et al . Phenotype and course of Hutchinson-Gilford progeria syndrome . N Engl J Med . 2008 ; 358 : 592 – 604 .
6. de Paula Rodrigues GH , das Eiras Tâmega I , Duque G , et al . Severe bone changes in a case of Hutchinson-Gilford syndrome . Ann Genet . 2002 ; 45 : 151 – 155 .
7. De Sandre-Giovannoli A , Bernard R , Cau P , et al . Lamin a truncation in Hutchinson-Gilford progeria . Science . 2003 ; 27 : 2055 .
8. Navarro CL , De Sandre-Giovannoli A , Bernard R , et al . Lamin A and ZMPSTE24 (FACE-1) defects cause nuclear disorganization and identify restrictive dermopathy as a lethal neonatal laminopathy . Hum Mol Genet . 2004 ; 13 : 2493 – 2503 .
9. Pendás AM , Zhou Z , Cadiñanos J , et al . Defective prelamin A processing and muscular and adipocyte alterations in Zmpste24 metalloproteinase-defi cient mice . Nat Genet . 2002 ; 31 : 94 – 99 .
10. Bergo MO , Gavino B , Ross J , et al . Zmpste24 defi ciency in mice causes spontaneous bone fractures, muscle weakness, and a prelamin A pro-cessing defect . Proc Natl Acad Sci U S A . 2002 ; 99 : 13049 – 13054 .
11. Pajerowski JD , Dahl KN , Zhong FL , et al . Physical plasticity of the nucleus in stem cell differentiation . Proc Natl Acad Sci U S A . 2007 ; 104 : 15619 – 15624 .
12. Young SG , Fong LG , Michaelis S . Prelamin A, Zmpste24, misshapen cell nuclei, and progeria — new evidence suggesting that protein farnesylation could be important for disease pathogenesis . J Lipid Res . 2005 ; 46 : 2531 – 2558 .
13. Hutchison CJ , Worman HJ . A-type lamins: guardians of the soma? Nat Cell Biol . 2004 ; 6 : 1062 – 1067 .
14. Maraldi NM , Lattanzi G . Involvement of prelamin A in laminopa-thies . Crit Rev Eukaryot Gene Expr . 2007 ; 17 : 317 – 334 .
15. Worman HJ , Bonne G . “ Laminopathies ” : a wide spectrum of human diseases . Exp Cell Res . 2007 ; 313 : 2121 – 2133 .
16. Halaschek-Wiener J , Brooks-Wilson A . Progeria of stem cells: stem cell exhaustion in Hutchinson-Gilford progeria syndrome . J Gerontol A Biol Sci Med Sci . 2007 ; 62 : 3 – 8 .
17. Scaffi di P , Misteli T . Lamin A-dependent misregulation of adult stem cells associated with accelerated ageing . Nat Cell Biol . 2008 ; 10 : 452 – 459 .
18. Akter R , Rivas D , Drissi H , Duque G . Lamin A/C knockdown affects osteoblast differentiation of mesenchymal stem cells . J Bone Miner Res . 2009 ; 24 : 283 – 293 .
19. Jiang Y , Zhao J , Liao EY , et al . Application of micro-CT assessment of 3-D bone microstructure in preclinical and clinical studies . J Bone Miner Metab , 2005 ; 23 (suppl ) : 122 – 131 .
20. Valverde-Franco G , Liu H , Davidson D , et al . Defective bone miner-alization and osteopenia in young adult FGFR3-/- mice . Hum Mol Genet . 2004 ; 13 : 271 – 284 .
21. Parfi tt AM , Drezner MR , Glorieux FH , et al . Bone histomorphome-try: Standardization of nomenclature, symbols and units . J Bone Miner Res . 1987 ; 2 : 595 – 610 .
22. Richard S , Torabi N , Franco GV , et al . Ablation of the Sam68 RNA binding protein protects mice from age-related bone loss . PLoS Genet . 2005 ; 1 : e74 .
23. Duque G , Huang D , Macoritto M , et al . Autocrine Regulation of In-terferon g in mesenchymal stem cells plays a role in early osteoblas-togenesis . Stem Cells . 2008 ; 550 – 558 .
24. Marie PJ . Transcription factors controlling osteoblastogenesis . Arch Biochem Biophys . 2008 ; 473 : 98 – 105 .
25. Pekovic V , Hutchison CJ . Adult stem cell maintenance and tissue re-generation in the ageing context: the role for A-type lamins as intrin-sic modulators of ageing in adult stem cells and their niches . J Anat . 2008 ; 213 : 5 – 25 .
26. Duque G , Rivas D . Age-related changes in lamin A/C expression in the osteoarticular system: laminopathies as a potential new aging mechanism . Mech Ageing Dev . 2006 ; 127 : 378 – 383 .
27. Zini N , Avnet S , Ghisu S , et al . Effects of prelamin A processing in-hibitors on the differentiation and activity of human osteoclasts . J Cell Biochem . 2008 ; 105 : 34 – 40 .
28. Franceschi RT , Ge C , Xiao G , et al . Transcriptional regulation of osteoblasts . Ann N Y Acad Sci . 2007 ; 1116 : 196 – 207 .
29. Bonewald LF , Johnson ML . Osteocytes, mechanosensing and Wnt signaling . Bone . 2008 ; 42 : 606 – 615 .
30. Jacob KN , Garg A . Laminopathies: multisystem dystrophy syndromes . Mol Genet Metab . 2006 ; 87 : 289 – 302 .
31. Liu B , Zhou Z . Lamin A/C, laminopathies and premature ageing . Histol Histopathol . 2008 ; 23 : 747 – 763 .
32. Sagelius H , Rosengardten Y , Schmidt E , et al . Reversible phenotype in a mouse model of Hutchinson-Gilford progeria syndrome . J Med Genet . 2008 ; 794 – 801 .
33. Varela I , Pereira S , Ugalde AP , et al . Combined treatment with statins and aminobisphosphonates extends longevity in a mouse model of human premature aging . Nat Med . 2008 ; 14 : 767 – 772 .
Received September 9 , 2008 Accepted February 15 , 2009 Decision Editor: Huber R. Warner, PhD
by guest on September 4, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from