a parthenogenetic life cycle in a population of canthocamptus staphylinus (copepoda, harpacticoida)
TRANSCRIPT

A PARTHENOGENETIC LIFE CYCLE IN A POPULATION OF CANTHOCAMPTUS STAPHYLINUS(COPEPODA, HARPACTICOIDA)
J . SARVALA
Lammi Biological Station, University of Helsinki, SF-i69oo Lammi, Finland
Received February 22, 1978
Keywords : Life cycle, parthenogenesis, encystment, Copepoda
Abstract
The harpacticoid copepod Canthocamptus staphylinus (Jurine)was shown to reproduce parthenogenetically in an oligotrophicFinnish lake . The population was univoltine with peaks of eggproduction in winter and early spring. Young from both peaksbecame adults in spring and aestivated as cysts . Laboratory ex-periments suggested that either high temperature (12°C or more)or long days will inhibit egg production, but that both factorstogether are needed to induce encystment . Embryonic survivalwas low, on the average only 60% . The population was limitedby low food levels in winter, and it is suggested that food shortageprevents the species from colonizing the profundal zone of oligo-trophic lakes .
Introduction
Although parthenogenesis is common in freshwaterCladocera and Ostracoda, bisexual reproduction is therule in Copepoda. Among the copepods, partheno-genetic reproduction has so far been verified only for twoharpacticoid species, Elaphoidella bidens (Schmeil) (Roy,1931) and Epactophanes richardi Mrazek (Lang, 1935),both from the family Canthocamptidae . In addition,parthenogenesis has been suspected on the basis of thescarcity of males in some other Elaphoidella species : E.leruthi Chappuis and E. elaphoides (Chappuis) in thenorthernmost part of their range (Chappuis, 1955), andE. grandidieri (Guerne & Richard) (Lang, 1948) .
In the present paper a parthenogenetic life cycle isdescribed in a population of the harpacticoid Cantho-
Dr. W. Junk b .v. Publishers - The Hague, The Netherlands
. Hydrobiologia vol . 62, 2 , pag. 113-129, 1979
camptus staphylinus (Jurine) . Both field and laboratoryresults are compared with earlier accounts of sexuallyreproducing populations .
Study area, methods and material
C. staphylinus populations were studied in the meso-humic, oligotrophic lake Paajarvi (61°04' N, 25°08' E) in1972-1974, in connexion with an investigation on theproduction of benthic copepods . The lake Paajarvi hasthe following characteristics : area 13 .4 km2; maximumdepth 87 m, mean depth 14 .4 m; water colour 40-6o mgPt 1-, ; conductivity 6o µS (20'Q ; low nutrient content ;oxygen concentration fairly high throughout the year inthe hypolimnion. Macroscopic vegetation extends to adepth of 1 .5 m, and the lower limit of microscopic algaeis at about 5 m . During summer, the daily mean temper-ature rises to about 20-24'C at the surface and to about5-7°C in the lower profundal zone ; winter temperaturesvary from below 1°C in the littoral zone to about 2-3°Cin the lower profundal zone (Fig . 1) . Ice cover forms inNovember-December and usually melts at the beginningof May. A more detailed description of the lake is givenby Ruuhijarvi (1974) .
To survey the bathymetric and horizontal distributionof benthic copepods, 3-1o replicate cores with singlecorers of different sizes (Hakala, 1971 and unpublished)were taken at 6o sites at depths between 0 .2 and 65 m indifferent parts of the lake . The corers were attached to theend of a pole when sampling depths less than 3 m .
Five sites of different depths were sampled regularlythrough all seasons, but C. staphylinus occurred only at
1 1 3

25-0 -0
1 1 4
10-
5-
LakePaajarvi 1972
Western bay
1973
1 m
1974
J
I
F
1
M
I
A
1
M
1
J
I
J
I
A
1
S
1
0
1
N
I
Fig . i . The annual course of the mean temperature of 5-day periods during three years at a depth of i min the westernbay of the lake Paajarvi, southern Finland . Shading indicates the range of daily mean temperatures in May-November
1973 .
the i and 2 m sites. At the 2 m site samples were takenbetween July 1972 and February 1974, at the i m site be-tween June 1973 and June 1974, with five occasionalsamplings between April 1972 and March 1973 . Thesampling interval was 2-3 weeks in summer and aboutone month in winter. Five replicate cores were taken oneach date. As the sampling scheme was aimed at pro-viding quantitative information on all benthic copepodsand as the methods were improved during the study,various core sizes combined with differing laboratorytreatment were used at different sites and in successiveyears. A detailed account of the methods is given else-where (Sarvala, in preparation) ; here it is sufficient tostate that core sizes of 15 .2 and 7 .9 cm 2 with 1 .9 cm 2 sub-cores for smaller nauplii were used at the i m and 2 msampling sites, and that after February 1973 all life stagesof C. staphylinus were quantitatively retained on thefinest sieve mesh used (0 .05 or 0.07 mm) .
In diagnostic characters C. staphylinus adults from thelake Paajarvi agreed with the descriptions of the speciesby Gurney (1932), Lang (1948) and Dussart (1967) . Thecopepodids were distinguished from those of C. micro-
staphylinus Wolf found at the i m site only in their greatersize (length of cephalothorax) at each stage . The naupliiwere identified to stage and species according to thedescriptions by Sarvala (1977) .C. staphylinus was reared in the laboratory in small
plastic vials with 2 or 1o ml of lake water which had beenfiltered through a 0.05 mm mesh to remove all copepod
larvae . Sieving residues of sediment from the lake withyeast extract (0.6 g I-) or pieces of decomposing Isoeteslacustris were offered as food . The rearing vials were keptin darkness, natural light or constant light at various con-stant temperatures between 1 .5 and 15° C, and checked1-2 times daily .
Results
Life cycle of C. staphylinus in Lake PadjdrviC. staphylinus was found only in the western part of thelake, in a narrow zone about i km long and 30-40 mbroad and between the depths of , and 2 m. The highestdensities were recorded in the region of the permanenti m sampling site, which was located in a sparse stand ofisoetids 10-15 m outside a dense Equisetum stand . Popu-lation densities recorded at 2 m were low, and the bestdata were obtained from I m (Figs . 2-4) . The spatial dis-tribution of the species at the i m site was usually con-tagious with a variance to mean ratio significantly greaterthan unity. Each sample comprised only five samplingunits, so that the figures presented for abundances are notvery precise, as indicated by the large confidence belts inFig. 2 . In spite of the low statistical precision of abun-dance estimates and relatively long sampling intervals, aclear pattern emerges from the present data . The basicfeatures of the life cycle were similar at both permanentstudy sites and in all study years, and only slight year-to-
D I

7 100E
v
vc
rnO
10-
150-
100-
50
200
150
50
30
20
10
20-
10
J
S
0
N
D
J
F
M
A
M
J1973
1974Fig . 2 . The seasonal abundance of cysts, adults, eggs, nauplii and copepodids of Canthocamptus staphylinus at thei m sampling site in the lake Paajarvi in 1973-1974. Vertical bars show the 95% confidence intervals based on Taylor'slaw, blocks denote the standard error of the mean, and each mean is indicated by a horizontal bar . Note the different
vertical scale for eggs and nauplii .
1 1 5
l-
Copepodids
Nauplii
504 700 287 200
287 000Eggs 247 300
Adults
Females:----- Mates
\ \ ~
-----~ -~
Cysts

year variation in the timing of reproduction was noted .The population seemed to reproduce mainly parthe-
nogenetically. Altogether 1082 adults of C. staphylinusfrom Paajarvi were examined, and only 3 males (0.28%)and 8 females with spermatophores (0.74%) were found .It is not known whether the eggs of the mated femaleswere actually fertilized . The population was univoltinewith peaks of egg-production in winter and early spring .Adult females spend the summer encysted in the bottommud. The first encysted females were found in early June,and the majority of the population apparently encystedduring late June, as free-swimming individuals wereabsent in July . The average maximum external diameterof the cysts was 0.622 mm (range 0.574-0.697 mm, n =16), and the internal hyaline layer was always surroundedby a layer of detritus .In autumn 1972, free-swimming individuals were not
captured before October . In 1973, a few free-swimmingfemales were found in August, and the main part of thepopulation had emerged from the cysts by the middle ofSeptember. In both years, emergence from the cysts oc-curred soon after the water temperature had fallen below12°C (Figs . i and 2) . Between September 1973 and June1974 no encysted individuals were found . Mortalityduring the cyst period could be estimated from the num-bers of cysts containing remnants of a dead individual .Dead cysts were not counted separately before August1973 . By comparing the mean density of dead cysts inAugust-September with this figure plus the mean densityof living cysts in late June-August, a mortality of about13% was indicated. This figure is likely to be an over-estimate, since empty and dead cysts seemed to be pre-served for a long time : the observed numbers of bothvaried little during the period from August 1973 to June1974 . From the mean density of dead cysts relative tothat of all cysts during this period, a mortality percentageof 4.6% was obtained. In any case, mortality during thecyst period appears low . In permanent water bodies sum-mer encystment is thus an efficient way of avoiding pre-dation .The maximum numbers of free-swimming adults
were recorded in September after which a rapid decreaseoccurred until December (Fig . 2) . From December to theend of March numbers of the old generation remainedconstant; a rapid drop occurred again during May andJune. These changes in adult mortality seemed to becorrelated with water temperature . During the period ofhigh mortality in September-December, the tempera-ture gradually fell from above i0 to about ,°C. During
1 1 6
the winter period temperature remained below i°C andlittle or no mortality was noticed . The higher mortality inspring and early summer was associated with increasingtemperatures. These changes in adult numbers thereforeprobably reflect seasonal variations in the predationpressure, although they may also involve seasonal migra-tions. Immigration to the i m site from shallower areasmight have occurred especially in late winter when thelower surface of ice lay at a depth of o.8 m owing to a lowwater level .
In 1973, the first females with large ova in the oviductswere observed in August, and their proportion in the totalnumber of females was greatest in October (Fig . 3) . Thefirst egg-sacs appeared in September, and the highestpercentage of females carrying egg-sacs and the mosteggs per m2 were both recorded in December . Thereafter,both the percentage of females with large ova in the ovi-ducts, the percentage of females with egg-sacs, and thenumber of eggs per m 2 decreased through the winter, allhaving their minimum in late March . The percentage offemales with large ova in oviducts increased again inApril, but then fell rapidly, so that in late May no femaleswith full oviducts were found . The percentage of femaleswith egg-sacs and the number of eggs per m' reachedtheir spring peak slightly later, in early May, and by earlyJune females with egg-sacs had disappeared .
The number of eggs per sac varied characteristicallyduring the reproductive period (Fig . 3) . The mean num-ber of eggs in a sac was relatively high (24 .4) in October,attained a maximum value (28 .2) in December, and thendecreased through winter and spring to a minimumvalue (18.4) in late May. This minimum value containedtwo abnormal sacs with only 3 and 9 eggs, but the valueof 21 .5 obtained after excluding these sacs would stillhave been the lowest during the whole reproductiveperiod .
Some first stage nauplii were caught in September1973, but greater numbers of nauplii were not seen untilDecember. At that time all naupliar stages were present(Fig . 4) . First and second stage copepodids were alsopresent in the December sample ; growth had been rapidin October-December when the temperatures were stillmoderate (Fig . 1) . In December temperature fell to thewinter level (0 .8-0 .9° C) and the rate of development of thejuveniles of C. staphylinus also slowed down, so that fifthstage copepodids were not observed until April 1974 .Most of the juveniles derived from the winter egg peakbecame adults during May and encysted by early June .Juveniles from the spring egg peak passed through the

tna
aE) 50w
(U
0
0
30
25
20
15
100
Eggs per sac
1
Females with large ova in oviducts
56
97
7
739
10
38
J J A S
D J
M
M J1973
1974Fig. 3 . The number of females with full oviducts and of ovigerous females as percentages of the total number of C .staphylinus females, and the number of eggs per m' at the i m site in 1973-1974 . Inserted figures show the numberof individuals on which each percentage or mean is based ; statistically significant differences between successive ob-servations are shown by asterisks (t-test or binomial t-test) . For the number of eggs per sac, vertical bars show the 957
confidence interval, blocks denote the standard error of the mean, and the mean is indicated by a horizontal bar .
1 1 7

1 1 8
E -5
C4
C3
C2
J
J
M1973
1974Fig. 4 . The seasonal development of the numbers of each developmental stage of C . staphylinus at i m in the lake
Paajarvi from the first nauplius (Ni) to the fifth copepodid stage (C5) .

early naupliar stages during May ; the first of thembecame adults during June, and the last were advancedcopepodids on the final sampling date in late June .
The year-to-year variations in the time of emergencefrom the cysts and the commencement of reproductionwere visible throughout the winter. Emergence was late(October) in autumn 1972 and early (September) in 1973 .In 1974 emergence was not studied but judging fromtemperature conditions it should have been early . InMarch 1973 the most advanced juveniles at i m weresecond stage copepodids, at the corresponding time in1974 fourth stage copepodids. In 1973 the first youngadults were caught in early June, in 1974 in May, whereasin 1975 they had already appeared in April . During earlysummer these differences disappeared so that in everyyear both free-swimming and encysted individuals werefound in late June and only encysted females in July .
Juvenile mortality could be roughly estimated byutilizing laboratory data on the developmental rates .Assuming isochronal development (see Miller et al.,
1977) each nauplius stage takes 1/6 of the duration of thewhole naupliar development, and each copepodid stage/s of the whole copepodid development . The duration ofeach life stage could thus be calculated from the develop-ment time-temperature regressions given by Sarvala(1978) for the eggs, nauplii and copepodids of C. staphy-linus .
The observed numbers of each stage on each date weredivided by the appropriate duration and the resultingcurve integrated over the winter and spring reproductiveperiods separately . These integrals represented the totalnumber of individuals which passed through each stage,and mortality could then be estimated from the decreaseof numbers from stage to stage . Deviations from the ex-pected duration of development and the low statisticalprecision of the abundance estimates caused considerablescatter in the results, but some general features seemevident. About 700 ooo eggs per m 2 were producedduring the winter reproductive period, and only 145 000
eggs per m z during the spring reproductive period . How-ever, only 6% of the winter eggs passed the sixth naupliarstage and 0 .3% became adult, while for the spring eggsthe respective figures were 20% and 6%. In absolutefigures, the spring reproduction therefore yielded at leastfour times as many adults as the winter reproduction. Inwinter, survival between the eggs and the first naupliarstage was estimated to be only 33%, but a smoothed sur-vival curve suggested that a more realistic estimate mightbe about 5o% . No estimate was possible for the spring
egg peak. In winter, the greatest losses seemed to occurbetween the sixth naupliar and the first copepodid stageas well as between the second and third copepodid stages,while in spring they occurred between the first and secondcopepodid stages .
The new generation which became adult in spring 1974was much less abundant than the old generation, so thatthe total numbers of the species in late June 1974 wereonly 22% of the corresponding figure in 1973 . In 1972 stilllower numbers had been recorded . The year-to-year vari-ations in the abundance of the population at the 1 msampling site were thus considerable . It should be noted,however, that the area of high density around the 1 m sitein 1973 was very small, and the densities observed in 1972and 1974 correspond more closely to those found in otherparts of the bay .
Laboratory experimentsThe capacity for parthenogenetic reproduction in thePaajarvi population of C. staphylinus was confirmedexperimentally . Altogether 15 females isolated as naupliior copepodids were kept individually in small culturingvials. All of these unmated females produced egg sacswhich yielded viable larvae . The sex ratio of the labora-tory population also approached that observed in thelake: among the 242 individuals reared from egg to adultin the laboratory, only 5 were males (2.1%) .
Twenty-four C. staphylinus females with large ova inoviducts or with an egg-sac were collected from Paajarviin late November 1973, and kept in the laboratory atseven constant temperatures between 1 .5 and 15'C untilearly June 1974. Three females had an attached sperma-tophore; the remaining 21 were probably unmated . Fournon-ovigerous females were full of sporozoan endopara-sites and did not produce any egg sacs, two produced 3,seven 4, two 5, six 6, one 7 and two 8 egg sacs during thisperiod. The number of sacs produced was dependent ontemperature, but light conditions also seemed influential .All six females kept at 6.o and 7 .5°C in natural daylightceased egg-production after 4-8 sacs in March or April(daylength was then increasing from about 12 h to 14 h)and became full of the coloured oil droplets that alwaysappear before encystment (see below). By contrast, six ofthe eight females kept at 1 .5 and 3 .5'C in darkness werestill continuing egg production after the same number ofsacs when the experiment was terminated in June ; thetwo others had died earlier . A female kept at 12°C indarkness stopped reproduction after 4 sacs in early March,and died in April . Five females transferred from darkness
1 1 9
at io°C to weak diffuse constant light at I5°C producedthere only one or two egg sacs ; four of these females en-cysted within 2-4 weeks after dropping the last egg sac .Young females reared at 15°C or transferred there fromlower temperatures did not develop eggs but became fullof orange oil droplets and usually encysted within twoweeks . Young females reared at 12'C in darkness or at6.o or 7.5°C in natural daylight (daylength 12-14 h) didnot develop eggs either, but they did not encyst, even
1 20
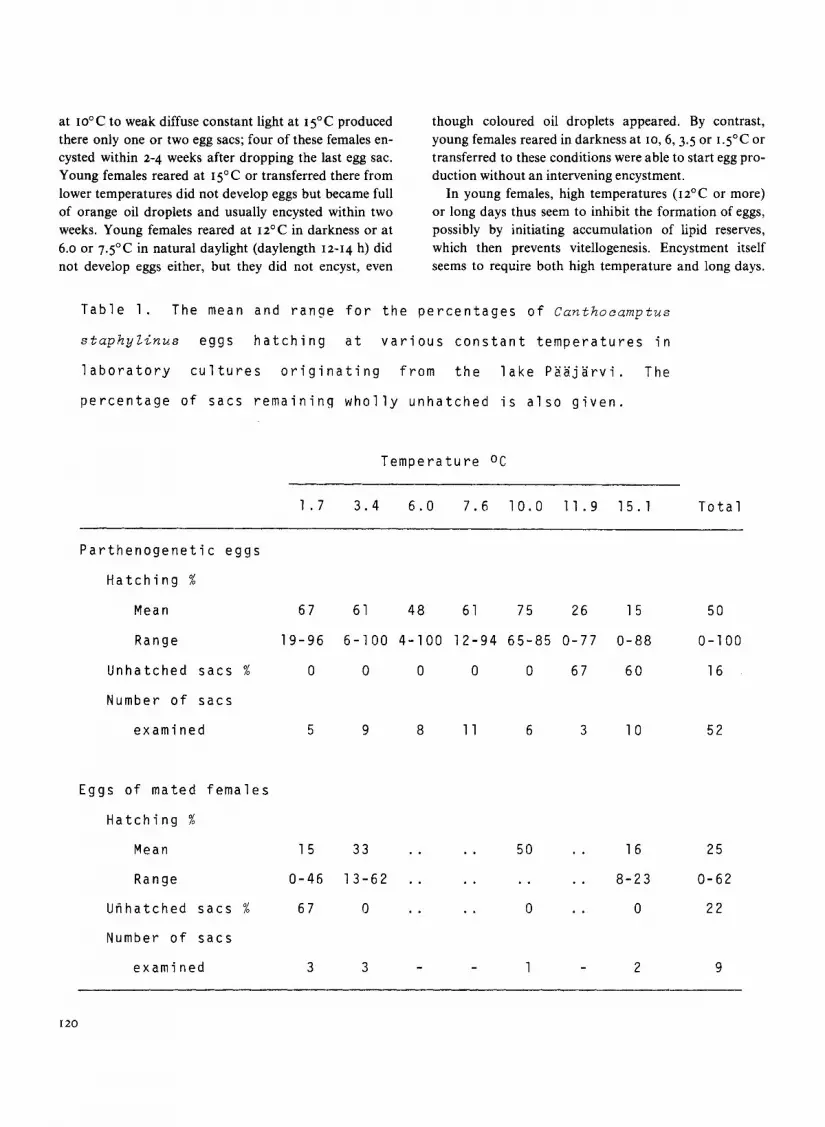
Table 1 . The mean and range for the percentages of Canthocamptus
though coloured oil droplets appeared . By contrast,young females reared in darkness at lo, 6, 3 .5 or 1 .5°C ortransferred to these conditions were able to start egg pro-duction without an intervening encystment .
In young females, high temperatures (i2°C or more)or long days thus seem to inhibit the formation of eggs,possibly by initiating accumulation of lipid reserves,which then prevents vitellogenesis . Encystment itselfseems to require both high temperature and long days .
staphyZinus eggs hatching at various constant temperatures in
laboratory
cultures
originating
from
the
lake Paa.jarvi .
The
percentage of sacs remaining wholly unhatched is also given .
Temperature oC
1 .7 3 .4 6 .0 7 .6 10 .0 11 .9 15 .1 Total
Parthenogenetic eggs
Hatching
Mean
Range
67
19-96
61
6-100
48
4-100
61
12-94
75
65-85
26
0-77
15
0-88
50
0-100
Unhatched sacs 0 0 0 0 0 67 60 16
Number of sacs
examined 5 9 8 11 6 3 10 52
Eggs of mated females
Hatching
Mean 15 33 50 16 25
Range 0-46 13-62 8-23 0-62
Unhatched sacs 67 0 0 0 22
Number of sacs
examined 3 3 - 1 - 2 9

The lipid reserves seem to be only partially consumedduring the cyst period, most being used for a rapid for-mation of eggs after emergence from the cyst . The cystsformed in cultures were usually like those collected fromthe lake, but sometimes an encysting female had crept tothe cover of the vial, and then the cyst was wholly trans-parent and without the usual detritus layer .
Thirty cysts collected from the lake on September 6and 8, 1972, were kept in the laboratory at 7-8° C in dark-ness. The first adult female emerged on September i i,the second one four days later, the third one on Septem-ber 24, and the remaining 27 females emerged by October2, or only slightly earlier than the majority of the popula-tion in the lake . Excystment thus seems to be controllednot only by environmental temperature but also byinternal factors .
In comparison with other harpacticoid copepods, theembryonic survival of C. staphylinus from Paajarvi waslow and highly variable . The percentage of eggs hatchingvaried according to temperature, the number of the eggsac, and possibly also the mode of reproduction . In spiteof the large between-sac variation, the average hatchingpercentages of the parthenogenetic eggs varied little be-tween temperatures of 1 .5 and io°C (Table i), with anoverall average of 60% within this temperature range . At12 and 15'C the average hatching percentage was verylow, and more than half of the parthenogenetic egg-sacsremained wholly unhatched . The data for sexually pro-duced egg sacs were very small, but show that the hatch-ing percentages were at least as low as for the partheno-genetic eggs, and at the lowest temperatures they seemedto be even lower .
Variation in the percentage hatch between the succes-sive egg sacs of the same female usually clearly exceededthe differences in the mean percentage hatch betweendifferent females . Generally the percentage hatch of eggsin the successive sacs of a parthenogenetic female washighest in the first sac and then seemed to decrease in thelater sacs (Table 2) . A similar trend at a lower level wasevident also in the sexually produced sacs . This effectmight have been due to adverse culture conditions, suchas defective food or excessive handling of animals . How-ever, ten other harpacticoid species, including the closelyrelated C. microstaphylinus, have always showed a highpercentage hatch of eggs in my cultures, so the presentresult may not be an artefact . Transferring a sac from oneconstant temperature to another during embryonicdevelopment seemed to have an adverse effect on thehatching success .
Table 2 . The mean and range of the percentages of
eggs hatching in successive broods laid by partheno-
genetic Canthocamptus staphylinus females in labora-
tory cultures originating from the lake Paajarvi .
The values come only from those temperatures (1 .5 -
10 .0 °C) at which a prolonged production of viable
eggs was possible (n = number of sacs examined) .
Discussion
Bathymetric distributionCharacteristic habitats of C . staphylinus are temporaryor permanent shallow ponds, slow-flowing rivers and thelittoral zone of lakes (see e .g. Lang, 1948 and Dussart,1967). In Finland, too, most finds have come from thelittoral of oligotrophic lakes (Sarvala, unpublished) . Insome mountain lakes in the Pyrenees and in the Alps thespecies is found also in the littoriprofundal and profundalzones (Monard, 1819 ; Pelosse, 1934; Smyly, 1964 ; Dus-sart, 1966 ; Rey & Dupin, 1973a) .Owing to the low temperature and weak illumination
which prevail all through the year, the profundal zone oflakes might provide a favorable environment for C. sta-phylinus. In most lakes, however, the species is confinedto the littoral areas . Of the commonest harpacticoidspecies of southern Finland, C. staphylinus has the high-est absolute rate of biomass growth during juveniledevelopment and egg production (Sarvala, 1978) . It istherefore likely to have relatively high food demands . Thelittoral and ponds habitats of C . staphylinus tend to beproductive or to have a high input of allochthonous or-ganic matter. The tendency of this species to prefereutrophic localities has been noted previously by e .g .Redeke & Vos (1933) in the Netherlands . In Lake Balaton,it lives only in regions covered by a thick layer of detritus(Ponyi et al., 1971). So far as is known, C. staphylinus
1 2 1
Mean Range n
1 . sac 76 19-100 11
2 . sac 55 6-88 10
3 . sac 64 24-94 5
4 . sac 39 7-65 5
5 . sac 33 4-85 4
6 . sac 90 80-100 2
Total 60 4-100 37

occurs at greater depths only in relatively productivelakes. In Sweden, Lilljeborg (1902) found the speciesdown to a depth of 36 m in eutrophic Lake Malaren, butelsewhere only in shallow water . In oligotrophic LakeWigry in Poland (no algal blooms, oxygen always abun-dant in hypolimnion) the species had its maximum in thelittoral at a depth of 1-3 m, whereas in the more produc-tive Lake Kiekrz (algal blooms, oxygen deficit in hypo-limnion during summer) it had its maximum in thelittoriprofundal zone at a depth of 8-io m (Rzoska,1936) . All the deep lakes at the foot of the Alps, whereC. staphylinus has been collected in the profundal (LakesNeuchatel, Geneva, Bourget, Annecy, Maggiore, Mergoz-zo), were at least mesotrophic at the time of the collec-tions (Dussart, 1963; Gerletti, 1972 ; Laurent, 1972) . InLake Port-Bielh, in the Pyrenees, the species occurs in thelittoriprofundal zone, in the same zone as where thebenthic alga Nitella flexilis has its peak abundance (Rey &Dupin, 1973a; Capblancq, 1973). Moreover, the organiccontent of the bottom sediments of this lake is of thesame order as in Polish mesotrophic lakes (Giani &Lucas, 1974), and an oxygen deficit develops in thedeepest basin during late winter (Capblancq & Laville,1968) . In Finland, the only find of C. staphylinus fromgreater depths (Kuopio, Maljalahti, 23 m; Sarvala, un-published) comes from a eutrophicated area . It thusseems probable that food shortage prevents C. staphyli-nus from colonizing the otherwise favorable profundalzone of oligotrophic lakes . The American Canthocamp-tus staphylinoides Pearse has a similar distributional pat-tern : in oligotrophic lakes it is present only in the littoralzone, but at all depths in more eutrophic lakes (Deevey,1941) . The reason for its exclusion from the profundal ofoligotrophic lakes may again be the poor food supply(Deevey, 1941) .
In contrast to C. staphylinoides, the cysts of whichendure total lack of oxygen (Deevey, 1941 ; Cole, 1953),C. staphylinus seems to be absent from the periodicallyanoxic hypolimnion of the most productive lakes . Forexample, in Lake Esrom in Denmark, C. staphylinusdoes not occur in the deepest parts where anoxic condi-tions develop during summer, although it is present in thelittoral zone and has been found down to a depth of 12 m(Berg, 1938). In the eutrophic Piburger See in Austriacomplete oxygen depletion occurred below a depth of 15m in 1973 and below 18-2o m in 1974-1975 ; C. staphylinuswas lacking below II m in 1973/74 and below 14 m in1974/75 (Thaler, 1977 ; Pechlaner, 1978) .
1 2 2
Life cycleMost earlier accounts of the biology of C. staphylinusconcern pond populations in central and western Europe(e .g. Wolf, 1905 ; Kruger, 1911 ; Dietrich, 1915 ; Donner,1928; Gurney, 1932; Roy, 1932 ; Lang, 1948; Smyly,1957; Roen, 1957 ; Tetart, 1974 ; Young, 1974) ; with the ex-ception of the work by Rey & Dupin (1973a, 1973b), thereare only short notes on lake populations (Thi6baud,19o8; Monard, 1919; Pelosse, 1934 ; Rzoska, 1936; Berg,1938; Thaler, 1977) . Although quantitative informationis almost lacking, especially of the naupliar stages, the lifecycle of C. staphylinus seems similar in most of the pondpopulations and also, so far as can be judged from thepublished notes, in the populations living in the littoralzone of lakes. Free-swimming individuals are scarce orabsent during summer, their nadir usually being observedin July . The number of free-swimming adults begins toincrease in August, but high numbers do not usually ap-pear until September-October . Even males are thenabundant and pairs in precopula are common through-out the autumn months. Egg production is maximal inlate autumn or early winter, usually in December, and thenumbers of the old generation as well as the reproductiveactivity decline through the winter. Males seem to dis-appear more rapidly than females . The new generationappears as first-stage copepodids in November-Decem-ber and it becomes adult in late March-May, causing anew peak in the abundance of adults . The species thendisappears from samples in late May-early June probablybecause of encystment .
The littoral population considered in the present in-vestigation differed from this pattern in its partheno-genetic mode of reproduction (see later) ; all populationsstudied previously have been bisexual. In addition, thenew winter generation in the lake Paajarvi seems toappear slightly later and develop more slowly than inother populations . The development times mentionedby Dietrich (1915), Smyly (1957) and Rouch (1961) foreggs and larval stages of this species are also shorter thanthose for the Paajarvi population (Sarvala, 1978) .
Furthermore, the increase in the number of egg-bearing females in April-May, which was so pronouncedin Paajarvi, has been noted in ponds only by Kruger(1911), Roen (1957) and Young (1974 ; in only one of threeyears) . Roen (1957) also recorded egg sacs through thewhole summer in a permanent Danish pond. He inter-preted the spring reproductive activity as due to the newgeneration, and he thus reported two generations peryear for his temporary ponds and three or more for a

permanent pond . According to other authors (Wolf,1905; Donner, 1928; Smyly, 1957), however, the genera-tion becoming adult in spring does not develop eggs be-fore the summer disappearance, and this was also thecase in the present study . Moreover, the present labora-tory experiments suggest that excessive illumination,such as exists in Denmark during ice-free conditions inApril-May, might inhibit ovogenesis in young females .It seems more likely that the Danish pond populationsof C. staphylinus were univoltine . Variations in the gener-al annual pattern seem to be possible, however. Accord-ing to Kruger (1911), numerous pairs in precopula andovigerous females had already appeared in a Germanpond in 1909 by late July. Reproduction continued inAugust, until September and early October only juvenileswere present, but adults appeared and reproductioncommenced again in late October . The summer 1909 wasexceptionally cold (Kruger, 1911), and the temperaturesmay have been low enough to allow emergence from thecysts and commencement of reproduction .
The deep-water population studied by Rey & Dupin(1973a, 1973b and personal communication) in LakePort-Bielh in the High Pyrenees was univoltine like theshallow-water populations but reproduced in summer .Ovigerous females were found during about one monthin July-early August just after the ice-break, which in thisalpine lake occurs in late June-early July . Juveniles fromthis egg peak evidently became adult in early October,,and they did not reproduce until the next summer . Onlypart of the population encysted and cysts were found bothin summer and in winter (see below) . The few other ob-servations on deep-water populations of the species alsosuggest an annual rhythm different from that in shallow-water populations . In the profundal zone of Lake Neu-chatel, Monard (1919) found ovigerous females fromMarch to November . Likewise, in Lake Bourget, Pelosse(1934) found ovigerous females mainly from Decemberto February in the littoral but also in June in the pro-fundal .
Donner (1928) and Roen (1957) counted the number ofeggs per egg sac, and although they, like Rouch (1968),found mean values clearly higher than those observed inPaajarvi, the seasonal variations were similar to those inthe present study : the largest egg sacs were produced inearly winter and towards spring the number of eggs persac decreased . The same pattern was thus observed inwidely differing habitats, and may be associated with theageing of females . Earlier laboratory work on other cope-pods supports this assumption : in Tisbe gracilis (T . Scott)
the number of eggs per egg sac begins to decrease after the
second sac (Battaglia, 1957), in Tigriopus brevicornis
(0. F. Muller) after the third sac (Comita & Comita,1966) ; in Acanthocyclops viridis (Jurine) the mean num-ber of eggs laid in successive broods tends to fall with ageindependently of food, although the overall mean perbrood varies with food (Smyly, 1970) .
It is generally agreed that the disappearance of C. sta-
phylinus during summer is due to encystment of theadults, but few authors have found these cysts in abun-
dance. Rz6ska (1936) found a maximum of 16 00o cystsper mz in the Polish Lake Wigry, and Smyly (1957) foundencysted adults in a British pond from late May until earlySeptember with a maximum in July; the observed num-bers of cysts were, however, only a fraction of the springand autumn population of free-swimming adults . This is
quite natural, since the cysts are difficult to find, particu-larly in unstained material . Even in the present study themaximum numbers of cysts in summer did not quiteequal those of free-swimming adults in the followingautumn (Fig . 2) .
The encysted individuals found by Berg (1938), Fryer &Smyly (1954), Rey & Dupin (1973b) and Young (1974)were enclosed only in a single, thin and transparent disc-formed case . In the cysts of C. staphylinus found by Kess-ler (1913) and Donner (1928) and in those ascribed to C .microstaphylinus by Lauterborn & Wolf (1909) this hya-line wall was surrounded by a layer of detritus cementedaround the cyst. All cysts found in Paajarvi were of thislatter type, but the laboratory experiments showed thatthe presence or absence of this outer detritus layer isdependent on the availability of suitable materials (cf .Fryer & Smyly, 1954) .
Although exact temperature data are usually lacking,all authors agree that in shallow-water habitats the massappearance of free-swimming adults always occurs aftera considerable cooling of water in the autumn . In 1902the first adults in a German pond were found on August19 at 21°C, but greater numbers appeared after a drop intemperature from 19 to 11 .5'C between August 27 andSeptember 7 (Wolf, 1905) . In the littoral of Lake Bourgetin France the species appeared in October, a little beforetemperature had fallen below 1o°C (Pelosse, 1934) . In aBritish pond, the main part of adult population emergedin 1968 and 1970 soon after temperature had fallen belowr1°C (Young, 1974) . In the present study the temperatureallowing emergence from cysts also seemed to be about10-12 °C. In temporary ponds, although the temperaturemay be low enough, emergence from the cysts after
12 3

drought is possible only after refilling of the pond, so thatthe appearance of C. staphylinus in such habitats is oftendelayed relative to that in permanent water-bodies or inyears with abundant water (e .g . Reen, 1957 ; Tetart, 1974 ;Young, 1974: the autumn 1969) .
Kessler (1913) showed experimentally that encysted C .staphylinus emerged from cysts when they were kept at atemperature of 9-io°C . Donner (1928) found that hightemperatures allowed encystment and, inversely, lowtemperatures caused encystment . Donner's experimentssuggested, however, that high temperature alone does notinduce encystment, since animals collected from a pondin May and kept over summer at about 16'C did notencyst . Likewise, Smyly (1957) failed to induce free-swimming adults to encyst at 18-2o°C . If the food con-ditions were poor, the failure to encyst at high tempera-tures might be due to inability to gather the energy re-serves necessary for encystment . Neither Donner norSmyly mentioned the light conditions, but Donner'sculture was kept in a cellar, and thus probably in darkness .If so, Donner's finding would agree with those presentedhere and suggest that light, as well as temperature, is im-portant in the control of reproduction and encystment inC. staphylinus, with long-day conditions inhibiting repro-duction and favoring encystment . In the control of theresting stage of several cyclopoid species, photoperiod isthe primary causal factor (e .g . Einsle, 1967 ; Elgmork &Nilssen, 1978), but whose effect may be modified bytemperature (Champeau, 1971 ; Watson & Smallman,1971). Even other environmental factors may intervene :Deevey (1941) showed that in Canthocamptus staphy-linoides a lack of oxygen at first inhibited emergencefrom the cysts ; however, after a longer period (5 months)excystment had occurred . This later observation indi-cates that the control of excystment may be partly endo-genous, as suggested also by the present experiments .
As mentioned above, the occurrence of cysts in thedeep-water population of Lake Port-Bielh (Rey & Dupin,1973b and personal communication) differed from thegeneral pattern found in shallow-water populations . InLake Port-Bielh cysts were found at all seasons, and theirproportion of the total adult population was low, exceptin September when about 50% of adults were encystedfemales. The numbers of C. staphylinus adults and es-pecially cysts were usually low and varied widely fromsample to sample . It seems clear, however, that the cystsformed in September were mainly old females which hadceased egg production, since 69% of these encystedfemales had a spermatophore, and, moreover, males of
1 24
the new generation did not appear until early Octoberwhen also the number of adult females without sperma-tophores suddenly increased . Some 50% of the old gener-ation thus encysted in September, probably as a responseto the relatively high temperatures (11-12°C which is theannual maximum) and the moderately long days . Al-though the majority of encysted individuals usually areunmated young adults., encysted females bearing an old,black spermatophore have been noticed by Kessler (1913)and in the present study (one individual in June), and,moreover, in my laboratory experiments, old femalescould be induced to encyst after a period of reproduction .Judging from the appearance of a few male cysts andfrom the more frequent appearance of encysted femaleswithout spermatophores in Lake Port-Bielh in winter,some individuals of the new generation may have encystedin October-November although temperature was lowand decreasing and illumination diminishing . Theappearance of some possibly new cysts in early summer(late July-early August)-if real-might be explained as aresponse to the suddenly increased illumination after theice-break . Thus, it seems that in the population of LakePort-Bielh tendency to encyst is weak, but that, on theother hand, encystment or at least postponement ofreproduction may occur even without the appropriateenvironmental cues .
Recent culture work (Sarvala, unpublished) suggeststhat there might be genetic differences in this respectbetween the shallow-water and the deep-water popula-tions. When reared in darkness at 8'C C. staphylinus in-dividuals from a pond in the High Pyrenees behaved likethose from Paajarvi and started reproduction immedi-ately after becoming adults without intervening encyst-ment. In the same conditions, individuals originatingfrom a depth of 55 m in Lake Geneva did not developeggs, but accumulated the lipid globules which alwaysprecede encystment .
Summer encystment in C. staphylinus may first haveevolved as a means of surviving in temporary pools whichdry up in summer . Increasing temperature and illumina-tion (long days?) have been fixed as environmental cuesinducing encystment, and excystment, on the other hand,has been linked with low temperature . In permanentponds and in the lake littoral summer encystment is nomore necessary as an assurance against drying up, but itis still advantageous for the species as a means of avoidingthe period of the most intense predation . As a relativelylarge harpacticoid C. staphylinus is subject to a con-siderable predation pressure at least in lakes (see e .g .

Lang, 1948 and Dussart, 1967), and as a univoltine winterbreeder it is not well adapted to endure intense preda-tion . Thus it is not surprising that in lake littoral practical-ly all adults encyst before summer .
Deeper in the lakes the reproductive success duringwinter may be low owing to the scarcity of food (seebelow). Then it might be advantageous for the species toencyst automatically for a given period after becomingadult, or to postpone reproduction until spring in someother way. As predation pressure in the low temperaturesof lake profundal is probably low in comparison withlake littoral, encystment may not give any decisive selec-tive advantage in the profundal. Indeed, as discussedabove, the present deep-water populations of the speciesshow a lowered percentage of encysting individuals and atendency to postpone reproduction in free-swimmingphase.
The basic univoltine life pattern of C. staphylinus thusshows a series of adaptive modifications in different typesof environments, from a unimodal cycle with a singlemid-winter period of egg production in most ponds,through a clearly two-peaked cycle with winter and springperiods of egg production in the littoral of Paajarvi, to aunimodal cycle with spring egg production only in thelittoriprofundal of Lake Port-Bielh .
In the littoral of Paajarvi, due to the scarcity of food inwinter, part of the reproductive capacity of the femalesremains unused until spring. In spring when food con-ditions improve temperature is still low, and therefore aspring peak of egg production develops . In ponds, on theother hand, rapid warming of water hinders reproduc-tion in spring and egg production is thus restricted towinter only . Deeper in the lakes, low temperature andillumination would allow reproduction all through theyear, but the food level is so low that the seasonal regimeof food production may impose restrictions . Microbialproduction, which is the main energy source for profun-dal animals, is dependent on temperature and sedimen-tation rate, both of which usually reach their maximumvalues during late summer . A tendency towards summerbreeding might thus be expected among profundal detri-tivores. Indeed, in Paajarvi some benthic copepods whichbreed in winter or in spring and autumn in the littoralzone tend to have a single summer peak of egg productionin the profundal (Sarvala, unpublished) . These con-siderations may not strictly apply to the population of C .staphylinus in Lake Port-Bielh, as that population isliving in a zone where the benthic alga Nitella flexilis isabundant . However, even there the food level in winter
may be too low for egg production and especially for thegrowth of young, and, on the other hand, temperaturesare so low throughout the year as to allow reproduction inspring or summer. Adaptation to different seasonalcycles of temperature and food availability may wellhave resulted in genetic differences between old deep-water and shallow-water populations of this species .
ParthenogenesisIn the present study, parthenogenetic reproduction wasconfirmed in a Finnish littoral population of C. staphy-linus. In addition, parthenogenesis is suspected in anoth-er population at a distance of 8o km from the first one. Onthe other hand, four certainly bisexual populations areknown from central and eastern Finland (Sarvala, un-published), and all earlier accounts of the life cycle of thisspecies from more southern areas (see references above)indicated a bisexual mode of reproduction .
The restricted distribution suggests that partheno-genesis in C. staphylinus may be of relatively recentorigin . This would also be in accordance with the highembryonic mortality . In insects high embryonic mor-tality is a characteristic feature of diploid parthenogenesiswhich is usually the first stage in the evolution of par-thenogenesis (see Suomalainen, 1969) .
Parthenogenesis in insects is often accompanied bypolyploidy (Suomalainen et al., 1976). Polyploid animalsare externally characterized by greater body size . C. sta-phylinus is one of the biggest freshwater harpacticoids,and about twice the weight of its close relative C. micro-staphylinus, but according to my own material from thePyrenees, the Alps, Denmark and Finland there is no sizedifference between the bisexual and parthenogeneticpopulations. The haploid chromosome number of thebisexual C. staphylinus is 12 (KrUger, 1911), which is thesame number as has been found in the marine harpacti-coid genera Tisbe and Tigriopus (see Lazzaretto-Colom-bera, 1976) . The caryology of the parthenogenetic C. sta-phylinus has not yet been studied. In Epactophanes ri-chardi cyclical parthenogenesis has been established byLang (1935), and males are also known in the Elaphoidel-la species, but, as in C. staphylinus, the cytological basisof their parthenogenesis is unknown .
The change from reproduction by fertilization to par-thenogenesis requires that (i) the female must be able tolay eggs without first mating, and (2) the unfertilized eggsmust be able to develop (see e .g. Suomalainen, 1969) . Incopepods, copulation and fertilization are not necessaryfor egg-laying. According to Lowndes (1933) and my own
1 2 5

observations (Sarvala, unpublished), isolated virginfemales of cyclopoids and harpacticoids often produceegg sacs which, however, do not develop further .
C. staphylinus females from the parthenogeneticpopulation of the lake Paajarvi seem to copulate if malesare present, but only few males are produced. Sex deter-mination in copepods as in many other crustaceans ap-pears to be complex and in many cases still obscure . Al-though sex chromosomes have been found in somespecies (Beermann, 1954; Riisch, 1960), a multifactorialcontrol of the sex ratio seems more likely (see Volk-mann-Rocco, 1972) . Whatever the underlying mecha-nisms, in many copepod species the sex ratio seems tovary according to external conditions, e .g. the proportionof females usually increases at low temperatures in bothnatural populations and in laboratory experiments (forreferences, see Battaglia, 1959 and Dussart, 1969) . C. sta-phylinus reproduces at low temperatures so that a pre-ponderance of females in its populations would be ex-pected .
There are few exact data on the sex ratio of the bisexualpopulations of C. staphylinus . Both Donner (1928) andSmyly (1957) noticed that the numbers of males de-creased in winter more rapidly than those of females, sothat the effective sex ratio could be observed only in thevery beginning of the reproductive period . Already Lillje-borg (1902) noted that females were more abundant thanmales in Sweden. In Germany, according to Wolf (1905),both sexes were about equally abundant from Septemberto February, whereas Donner (1928) stated that femaleswere usually more abundant, the numbers of malesapproaching those of females only in some samples inOctober. In Britain, according to Smyly (1957 and per-sonal communication), the proportion of males amongadults newly emerged from the cysts was about 25% in1953 and about 15% in 1954 . Further, Young (1974)reported a preponderance of females throughout the yearin a British pond, and, according to the tables of Bever-combe et al. (1973) the population in a British pool con-sisted almost exclusively of females and juveniles be-tween March and July . In the C . staphylinus populationof Lake Port-Bielh (Rey & Dupin, 1973b and personalcommunication) females clearly dominated : in October-November 18-23% of adults were males and 17% of cystsfound during winter contained a male . Even in the sexual-ly reproducing populations of C . staphylinus the sex ratiothus seems to be far from unity, females being morenumerous than males .
Parthenogenesis should give a species short-term
126
selective advantage in two days . First, it increases theprobability of successful dispersal because every singleindividual transported to a new locality is potentially ableto reproduce and found a new colony. This would be par-ticularly important for colonization of new areas or if theextinction rate of established colonies were high. Second,it reduces the amount of energy consumed per femaleoffspring produced . This would be particularly importantif food were limiting.
The potential advantages of parthenogenesis are con-siderably diminished in the Paajarvi population of C .staphylinus because of the reduced embryonic survival.
Part of the high mortality of eggs in the laboratory ex-periments could have been due to culture conditions, butthe field data suggested a survival percentage of the sameorder. A lowered embryonic survival has also been notedfor parthenogenetic eggs of Elaphoidella bidens (about56%, calculated from Roy, 1931) .
In a parthenogenetic population consisting only offemales, both the energy investments in juvenile develop-ment and the survival percentage of embryos required tomaintain a stable population with a given number ofmature females, would be only half of the correspondingfigures in a bisexually reproducing population with ai : i sex ratio, and 80% of those in a bisexual populationwith males representing 20% of adult numbers . This as-sumes equal reproductive effort per female and equalmortalities in both populations . The parthenogeneticpopulation might use the energy saved in juvenile growthto increase reproductive effort per female, which wouldthen allow still lower survival of embryos without de-creasing the reproduction potential . This would be pos-sible only if food is limiting the rate of egg production .Alternatively, the number of reproducing females mightbe increased, diminishing thus the probability of extinc-tion due to chance fluctuations, or competitive abilitymight be increased in some other way .
Thus it seems probable that due to its low embryonicsurvival, the parthenogenetic population now studied hasa reproductive economy only about equal to or worsethan a bisexual population with a ioo-% survival of em-bryos. The hatching percentage of eggs produced bymated females from the Paajarvi population seemed tobe even lower than that of parthenogenetic eggs, butearlier data from purely bisexual populations seem toimply a high hatch percentage (9o% or more). Hoppen-heit (1976) has shown that in laboratory-reared Tisbeholothuriae Humes the percentage hatch of eggs de-creased with increasing population density. Changes in

embryonic mortality might thus be a mechanism in theregulation of copepod densities . The low embryonic sur-vival in the population studied might thus be the result ofa relatively high density, independent of parthenogenesis,although this seems unlikely .
The suppression of egg production during winter inPaajarvi, and the simultaneous high mortality of juve-niles,. strongly suggest that this population was severelyfood-limited during winter . This assumption is supportedby the revival of reproductive activity in spring after foodconditions improved owing to a considerable growth ofbenthic algae under the ice during April. Further, duringearly summer the juvenile mortality was quite low . Com-parable quantitative material does not exist from themore southern localities, so that juvenile mortalitiescannot be compared . The mean number of eggs per egg-sac, however, provides indirect information on the nutri-tional status of the population (see e .g. Smyly, 1970 ; Elg-
mork & Halvorsen, 1976) . In Denmark, the number ofeggs per sac was higher in localities with higher pH andconductivity, which probably indicate a higher produc-tivity. Even in the poorest habitat with pH 4.5, the egg-sacs were slightly larger than in Paajarvi (Reen, 1957) .The numbers of eggs per brood given by Donner (1928)and Rouch (1968) were also higher than in Paajarvi. Itseems quite clear that the majority of the central andwestern European localities inhabited by C. staphylinusare more productive than the Finnish localities . The con-siderations presented above suggest that when food islimiting even a parthenogenetic population with a ratherlow embryonic survival is able to compete with a bi-sexual population which has a high survival of embryosand males representing about 20-30% of the total num-ber of adults . In Paajarvi, food seems to be a limitingfactor for C. staphylinus, and the parthenogenetic modeof reproduction should give selective advantage . Butwhere food is abundant, the bisexual population de-scribed would be able to produce more female offspringper time unit, and consequently outcompete a partheno-genetic strain with lowered embryonic survival . Low per-centage hatch of eggs might thus prevent the partheno-genetic strain of C. staphylinus from colonizing morefavorable areas .
Summary
The life cycle of Canthocamptus staphylinus (Jurine) inan oligotrophic Finnish lake is described . Partheno-
genetic reproduction was confirmed by the results fromboth field and laboratory studies .
Adult females aestivated as cysts . The main part of thepopulation emerged from the cysts in autumn after watertemperature had fallen below i2°C . The population wasunivoltine with peaks of egg production in winter andearly spring . The new generation became adult in springand encysted in June . Free-swimming individuals werescarce or absent in July and August .
Adult mortality was low in winter and during the cystperiod, but higher in autumn and spring . Egg productionwas suppressed and juvenile mortality was high duringmidwinter, probably due to food shortage, but reproduc-tive activity revived and juvenile survival increased inspring after the food conditions improved .
In the laboratory, prolonged egg production was pos-sible only at a temperature of io°C or less, and em-bryonic survival was low (mean 60%) and decreasing insuccessive sacs . The laboratory experiments suggestedthat either high temperature (12° C or more) or long dayswill inhibit egg production, but that both factors togetherare needed to induce encystment .
A survey of earlier literature showed that the species isnormally found in shallow-water habitats and deep-water populations occur only in mesotrophic lakes . Foodshortage seems to prevent the species from colonizingthe profundal zone of oligotrophic lakes, and lack ofoxygen from inhabiting the profundal of eutrophiclakes .
Acknowledgements
My best thanks are due to the personnel of the LammiBiological Station for providing me with good workingfacilities, to Jeanne Rey (Toulouse) and W. J . P. Smyly(Windermere) for kindly putting their original data atmy disposal, to Maija Sarvala for help and encourage-ment, to J . Lokki, K. Salonen, A. Saura, J. Tiainen andK. Vepsalainen for constructive criticism, and to SusanPowell and R . Jones for improving the English of themanuscript. This work was supported by the Universityof Helsinki and in part by the Academy of Finland .
References
Battaglia, B . 1957 . Ricerche sul ciclo biologico di Tisbe gracilis(T. Scott), (Copepoda, Harpacticoida), studiato in condizionidi laboratorio . Arch. Oceanogr. Limnol . 11 : 29-46 .
1 2 7

Battaglia, B . 1959 . Facteur thermique et differenciation saison-niere chez un Copepode Harpacticoide de la Lagune de Venice .Vie et Milieu to : 1-13 .
Beermann, W . 1954 . Weibliche Heterogametie bei Copepoden .Chromosoma 6: 381-396 .
Berg, K . 1938 . Studies on the bottom animals of Esrom Lake . K .danske vidensk . Selsk . Skrifter, Naturv . Math . Afd ., IX : 8 :1-255 .
Bevercombe, A . M., Cox, N ., Thomas, M . P. & Young, J . O .1973 . Studies on the invertebrate fauna of a wet slack in a sanddune system . Arch . Hydrobiol. 71 : 487-516 .
Capblancq, J . 1973 . Phytobenthos et productivity primaire d'unlac de haute montagne dans les Pyrenees Centrales . Ann .Limnol . 9 : 193-230.
Capblancq, J . & Laville, H . 1968. Etude morphometrique etphysico-chimique de neuf lacs du Massif de Neouvielle(Hautes-Pyren6es). Ann . Limnol . 4 :275 -324 .
Champeau, A . 197 t . Recherches sur ('adaptation a la vie latentedes Copepodes Cyclopoides et Harpacticoides des eaux tem-poraires Provengales . Bull . Soc . Ecol . 2: 151-167 .
Chappuis, P . A . 1955 . Notes sur les Copepodes . 18. NouveauxHarpacticoides des Pyrenees . 19 . Harpacticoides cavernicolesde Grece . 20 . Copepodes Harpacticoides des Iles du Pacifique .Notes Biospeol . to : 89-10, .
Cole, G. A . 1953 . Notes on copepod encystment. Ecology 34 :208-211 .
Comita, G. W . & Comita, J . J . 1966 . Egg production in Tigriopusbrevicornis. In: Some contemporary studies in marinescience (H . Barnes, ed .), pp . 171-185 . London : Allen & Unwin .
Deevey, E . S ., Jr . 1941 . Notes on the encystment of the harpacti-coid copepod Canthocamptus staphylinoides Pearse . Ecology22 : 1 97-200 .
Dietrich, W. 1915 . Die Metamorphose der freilebenden Suss-wasser-Copepoden . I . Die Nauplien and das erste Copepodid-stadium . Zeitschr. wiss . Zool . 113 : 252-323 .
Donner, F . 1928 . Die Harpacticiden der Leipziger Umgebungand der Schneeberger Erzbergwerke . Int . Rev. ges . Hydrobiol .Hydrogr . 20 : 221-353 .
Dussart, B . H . 1963 . Les grands lacs d'Europe occidentale. Ann .Biol . 2 : 499-572 .
Dussart, B. H . 1966 . Copepodes de la faune benthique du Leman .Vie et Milieu 17 (1-B) : 283-302 .
Dussart, B . 1967 . Les Copepodes des eaux continentales d'Europeoccidentale . Tome I : Calanoides et Harpacticoides . Paris :Boubee .
Dussart, B . 1969 . Les Copepodes des eaux continentales d'Europeoccidentale . Tome II : Cyclopoides et Biologie . Paris : Boubee .
Einsle, U . 1967 . Die ausseren Bedingungen der Diapause plank-tisch lebender Cyclops-Arten . Arch . Hydrobiol . 63 : 387-403 .
Elgmork, K . & Halvorsen, G . 1976 . Body size of free-livingcopepods . Oikos 27 : 27-33 .
Elgmork, K . & Nilssen, J . P . 1978 . Equivalence of copepod andinsect diapause . Verb . Int . Ver . Limnol . 20 .
Fryer, G . & Smyly, W . J . P . 1954 . Some remarks on the restingstages of some freshwater cyclopoid and harpacticoid cope-pods. Ann . Mag. nat . Hist . 12, 7 : 65-72.
Gerletti, M . 1972 . Comparative trends of primary productivityand some chemical parameters in Lake Maggiore on a plurian-nual basis . In : Productivity problems of freshwaters (Z. Kajak,A. Hillbricht-Ilkowska, eds .), pp . 709-713 . Warszawa-Krakow :Polish Scientific Publishers .
Giani, N . & Lucas, C . 1974 . Les sediments d'un lac de haute
1 28
montagne : structure, nature et peuplement . Ann . Limnol. lo :223-244 .
Gurney, R. 1932 . British Freshwater Copepoda. Vol . II . London :Ray Society .
Hakala, 1 . 1971 . A new model of the Kajak bottom sampler, andother improvements in the zoobenthos sampling technique .Ann. Zool . Fennici 8 : 422-426 .
Hoppenheit, M . 1976 . Zur Dynamik exploitierter Populationenvon Tisbe holothuriae (Copepoda, Harpacticoida) . III . Repro-duktion, Geschlechtsverhaltnis, Entwicklungsdauer and Ober-lebenszeit . Helgolander wiss . Meeresunters. 28 : 109-137 .
Kessler, E . 1913 . Ober eine Abart von Canthocamptus staphyli-nus: Canthocamptus staphylinus var . Thallwitzi nov . var.Arch . Hydrobiol. Plankt . 8 : 179-198 .
Kruger, P. 1911 . Beitrage zur Kenntnis der Oogenese bei Har-pacticiden, nebst biologischen Beobachtungen. Arch . Zell-forsch. 6 : 165-189 .
Lang, K . 1935 . Studien in der Gattung Epactophanes (CopepodaHarpacticoida) . Arkiv for Zoologi 28A (I 1) : 1-27 .
Lang, K . 1948 . Monographie der Harpacticiden . Vol . 1-11 . Lund .Laurent, P . J . 1972. Lac Leman : Effects of exploitation, eutro-
phication, and introductions on the salmonid community .J . Fish . Res . Bd . Canada 29 : 867-875 .
Lauterborn, R . & Wolf, E . 1909 . Cystenbildung bei Cantho-camptus microstaphylinus . Zool . Anz . 34 :130-136 .
Lazzaretto-Colombera, I . 1976. Chromosome number in tenspecies of the genus Tisbe (Copepoda : Harpacticoida) . Mar .Biol . 38 : 159-162 .
Lilljeborg, W. 1902 . Synopsis specierum huc usque in aquis dul-cibus Sueciae observatarum familiae Harpacticidarum . K .svenska Vetensk .-Akad . Handl . 36 (1) : 1-75 .
Lowndes, A . G . 1933 . Sexual reproduction in copepods . Nature131 : 240-241 .
Miller, C . B ., Johnson, J . K. & Heinle, D . R . 1977 . Growth rulesin the marine copepod genus Acartia . Limnol . Oceanogr . 22 :326-335 .
Monard, A. 1819 . La faune profonde du lac de Neuchatel . Bull .Soc . neuchatel . Sci . nat . 44 : 1-236 .
Pechlaner, R. 1978 . Response of a eutrophicated lake (PiburgerSee) to reduced nutrient load and selective water renewal . In :Jubilee symposium on lake metabolism and lake manage-ment . Uppsala: August 22-27, 1977 . Ergebn. Limnol .
Pelosse, J . 1934 . Etude sur la faune des Cladoceres et des Cope-podes de la region moyenne des Alpes frangaises . These, Lyon :Bosc & Riou .
Ponyi, J . E., Olah, J ., Biro, P. & Biro, K . 1971 . Comparativeinvestigations on the benthic fauna at two sewage inflows ofLake Balaton . Ann . Biol . Tihany 38 : 199-226 .
Redeke, H. C . & Vos, A. P . C. de 1933 . Beitrage zur Kenntnis derFauna niederlandischer oligotropher Gewasser . Int . Rev . ges .Hydrobiol. hydrogr. 28 : 1 -45 .
Rey, J . & Dupin, B . 1973a . Ecologie des Crustaces benthiques dulac de Port-Bielh (Pyrenees Centrales) : I . Repartition . Ann .Limnol. 9 :121-134 .
Rey, J . & Dupin, B . 1973b. Ecologie des Crustaces benthiques dulac de Port-Bielh (Pyrenees Centrales) : II . Cycles biologiquesAnn. Limnol . 9 :259-271 .
Rouch, R . 1961 . Le developpement et la croissance des Cope-podes Harpacticides cavernicoles (Crustaces) . C .r . hebd .Seanc . Acad . Sci . Paris IV, 252 : 4062-4064 .
Rouch, R . 1968 . Contribution a la connaissance des Harpactici-des hypoges (Crustaces-Copepodes) . Ann . Speleol. 23, 1 : 1-167 .

Roy, J . 1931 . Sur 1'existence de la parth6nogenese chez une especede Cop6podes (Elaphoidella bidens) . C. r. hebd . S6anc . Acad .Sci . Paris 192, 8 :507-509 .
Roy, J . 1932 . Cop6podes et Cladoceres de l'Ouest de la France .Recherches biologiques et faunistiques sur le plancton d'eaudouce des Vall6es du Loir et de la Sarthe . These, Gap : Jean .
Ruuhijarvi, R . 1974. A general description of the oligotrophiclake Paajarvi, southern Finland, and the ecological studies onit . Ann . Bot . Fennici 11 :95-104 .
Rusch, M .-E . 1960 . Untersuchungen caber Geschlechtsbestim-mungsmechanismen bei Copepoden . Chromosoma (Berl.)11 :419-432 .
Rzoska, J . 1936 . Uber die Okologie der Bodenfauna im Seen-litoral. Arch . Hydrobiol . Rybactwa 10 : 76-172 .
Roen, U. 1957 . Contributions to the biology of some Danishfree-living freshwater copepods . Biol . Skr. Dan . Vid . Selsk . 9,2 : 1-101 .
Sarvala, J . 1977 . The naupliar development of six species of fresh-water harpacticoid Copepoda . Ann . Zool. Fennici 14 : 135-161 .
Sarvala, J . 1978 . Effect of temperature on the duration of egg,nauplius and copepodid development in some freshwaterbenthic Copepoda. Unpublished manuscript .
Smyly, W . J, P . 1957 . Observations on the life-history of the har-pacticoid copepod, Canthocamptus staphylinus (Jurine) . Ann .Mag. nat . Hist. 12, 10: 509-512 .
Smyly, W. J, P . 1964 . An investigation of some benthic Entomo-straca of three lakes in northern Italy . Mem . Ist . Ital . Idrobiol .17 : 33-56 .
Smyly, W . J . P . 1970 . Observations on rate of development, lon-gevity and fecundity of Acanthocyclops viridis (Jurine) (Cope-poda, Cyclopoida) in relation to type of prey . Crustaceana 18 :21-36 .
Suomalainen, E . 1969 . Evolution in parthenogenetic Curcu-lionidae . In : Evolutionary Biology, Vol . 3 (T. Dobzhansky,M. K. Hecht, W. C . Steere, eds.), pp . 261-296. New York :Meredith Corporation .
Suomalainen, E ., Saura, A . & Lokki, J . 1976 . Evolution ofparthenogenetic insects. In: Evolutionary Biology, Vol . 9 (M .Hecht, W . C . Steere, B . Wallace, eds .), pp . 209-257. New York-London : Plenum Press .
Tetart, J . 1974 . Les Entomostrac6s des milieux peu profonds dela vall6e du Rhone . Essai d'6tude &cologique : composition desassociations et r6partition des especes . Trav . Lab . Hydrobiol .Piscicult . Grenoble 64-65 : 109 - 245 .
Thaler, B . 1977 . Die benthischen Crustaceen des Piburger Sees(Otztal, Tirol) . Diss . Abt. Limnol. Innsbruck 8 : 1 - 177 .
Thi&baud, M . 1908 . Contribution a la Biologie du lac de Saint-Blaise . Ann . Biol. Lacustre 3 : 54 - 140.
Volkmann-Rocco, B . 1972 . The effect of delayed fertilization insome species of the genus Tisbe (Copepoda, Harpacticoida) .Biol . Bull . (Woods Hole) 142 : 520-529 .
Watson, N . H . F. & Smallman, B . N . 1971 . The role of photo-period and temperature in the induction and termination ofan arrested development in two species of freshwater cyclopidcopepods. Can . J . Zoo1 . 49 : 855 - 862 .
Wolf, E . 1905 . Die Fortpflanzungsverhaltnisse unserer einheimi-schen Copepoden . Zool . Jahrb . Syst . 22 : 101-280 .
Young, J . O. 1974. Seasonal changes in the abundance of Micro-crustacea in a small pond . Hydrobiologia 45 : 373- 389 .
1 29