381 p 274 n° 130
TRANSCRIPT

ANALYTICAL LETTERS, 18(B13), 1593-1606 (1985) 381 P 274 n° 130
UNIVERSITAS BRUXtLLfcNSIS
DETERMINATION DE L'ACIDE L-ASCORBIQUE A L'AIDE
D'ELECTRODES BACTERIENNES, TISSULAIRES ET ENZYMATIQUES.
K£Y WORDS: bacterial, tissue and enzymic biosensors, L-ascorbic
acid in pharmaceucicals, sensor.
f
B.J. Vincké, M.J. Devleeschouwer°, G.J. Patriarche.
L-ascorbic acid membrane électrodes based upon the use of four
classes of biocatalysts immobilized at an oxygen électrode are eva-
luated and compared in terms of électrode properties and operating
requirements. Isolated ascorbate oxydase enzyme in soluble form and
in covalent binding matrices, peel of cucumber and living bacterial
cells of Enterobacter agglomerans strain, respectively, are employed
as biocatalytic layers. The physico-chemical factors, life times
and interférences are discussed in détails. The low stability of the
soluble enzyme sensor does not allow its analytical utilization,
but the immobilized enzyme, bacterial and tissue électrodes can be
used, even in multivitamin pharmaceutical formulations. The common -6 -4
linear range of those three biosensors are of 4.10 M to 7.10 M
with a précision and a reproducibility of + 3 %.
Institut de Pharmacie Université Libre de Bruxelles, Campus Plaine 205/6 et 205/2°
Boulevard du Triomphe, B-1050 Bruxelles, Belgium.
ABSTRACT
1593
Copyright © 1985 by Marcel Dekker, Inc. 0003-2719/8S/1813-1593$3.50/0

1594 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
INTRODUCTION
Les réactions enzymatiques, de par leur spécificité, ont été
couplées à de nombreux systèmes de détection tels que la fluori-1 2 3 . , . ,
métrie , 1'électrochimie ' , les mesures enthalpimetriques a 4
l'aide de thermistances , les mesures de fréquence par quartz
piézoélectriques ^ et plus récemment les fibres optiques
Cependant, ce sont les méthodes électrochimiques qui ont rencontré
le plus grand intérêt dans le domaine analytique et qui ont permis 3
la détermination de plus de 40 substrats . Parallèlement aux 7-9
électrodes à enzymes, des électrodes a membranes microbiennes ,
tissulaires '0"'' Q^ ^ organelles 1^>'2 sont développées.
L'importance de l'acide ascorbique (vitamine C) dans le domaine
pharmaceutique et agroalimentaire a donné lieu à de nombreuses 13
méthodes de dosage de celui-ci; citons les méthodes coulometriques
voltarame'triques impulsionnelles différentielles ainsi que l'uti
lisation d'électrodes enzymatiques .
Le but du présent travail est de présenter et de comparer
quatre électrodes biocatalytiques (à enzyme soluble ou immobilisée,
à mésocarpe de concombre ou à cellules bactériennes) riches en ascor-
bate oxydase et permettant de déterminer l'acide L-ascorbique dans
des formes pharmaceutiques. La réaction d'oxydation de la vitamine C
en présence d'ascorbate oxydase (EC 1.10.3.3) est schématisée
ci-dessous : . , , ascorbate
Acide L-ascorbique + 1/2 0^ > acide déhydroascorbique oxydase • ^
Elle peut être suivie stoechiométriquement par la mesure ide la
déplétion en oxygène grâce à une électrode à diffusion gazeuse d'
oxygène. Les caractéristiques et les performances analytiques de ces
quatre biosenseurs seront examinées en détails.

DETERMINATION DE L'ACIDE L-ASCORBIQUE
PARTIE EXPERIMENTALE
1595
Appareillage
Le dispositif de mesure comporte une électrode à diffusion ga
zeuse d'oxygène (0^ électrode Orion research model 97-08-00) dont
la membrane de dialyse est remplacée par une membrane de polypropy-
lène de + 50 im obtenue par pressage à chaud (190°C) sous pression 2 R
(160 kg/cm") pendant 1 minute de polypropylène Novolen BASF. La
différence d'intensité de courant résultant de la réduction de 1'
oxygène à la cathode est convertie linéairement en potentiel et lue
sur l'échelle pX d'un millivoltmetre Orion reseach digital ionanaly-
ser type 601 I. La courbe de variation de pX en fonction du temps
est tracée à l'aide d'un enregistreur E = f(t) (Goërtz Servogor 120)
relié en synchronisation avec le millivoltmètre.
Les mesures sont réalisées sous agitation constante dans une
cellule thermostatisée à 25,0 + 0,2°C.
Les coupes de tissus végétaux ont été effectuées à l'aide d'un
microtome à congélation (Leitz type 1310).
Réactifs_et milieux microbiologi2ues_de croissance
Toutes les solutions sont préparées à partir d'eau bidistillée.
Tous les réactifs utilisés sont de qualité P.A. ou biochimique.
L'ascorbate oxydase extraite de cucumis species est de prove
nance Boehringer (170 U/mg).
La souche d'Enterobacter agglomerans Indole (-) Saccharose (-)
(Erwinia herbicola) est une souche sauvage isolée à partir de thés
pharmaceutiques . Sa culture mère est maintenue à 4°C sur gélo
se inclinée H.I.A. (Difco).
Les milieux de croissance sont de deux types:
a) Milieu d'induction (0,2 % en substrat):
Extrait de levure 1g, Tryptone 1g, NH Cl 2g, KH„P0, Ag, Na,HPO, Ag
Fe (<FeSO^.7H,0) lOmg, Mg (<MgCl2) 28mg, Eau distillée IL.
Cette solution est autoclavée à 121°C pendant 15 minutes. Avant 1'
ensemencement, 0,2 g% d'acide L-ascorbique sont ajoutés extempora-
nément et la solution totale est stérilisée par filtration par
fraction de 100 ml.

1596 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
b) Milieu B.H.I. (Difco):
Il est utilisé avec ou sans inducteur (0,2 % d'acide L-ascorbique)
1) Electrode bactérienne:
La croissance d'Enterobacter agglomerans est effectuée dans le
milieu d'induction à 0,2 % en substrat pendant 24 heures et sous
agitation orbitale (100 tours min. ') à 27°C. Cent millilitres sont
filtrés sur filtre d'acétate de cellulose Millipore (HAWP 047 SO).
Le filtre saturé est lavé à l'aide de liquide physiologique. Une
partie adéquate du filtre est prélevée et fixée contre la membrane
de polypropylène de l'électrode par le biais d'un embout amovible
de P.V.C.
L'électrode est conservée soit à 4°C dans le tampon phosphate
0,1 M de pH 6 ,5 soit dans son milieu de culture à 27°C sous agitation
2) Electrode tissulaire:
Lors d'un travail se rapportant aux électrodes tissulaires, Smit
et Rechnitz avaient proposé Cucumis sativus comme support mem-
branaire lors du dosage de la cystéine.
Des membranes de 500 à 600 pm sont taillées dans le mésocarpe
de concombre. Celles-ci peuvent être conservées durant plus de deux
semaines dans un tampon phosphate 0,1 M de pH 6 , 5 .
La membrane tissulaire fixée à la surface de l'électrode par
un filet de nylon est maintenue au moyen d'un anneau de caoutchouc.
L'électrode est alors conservée à 4°C dans le tampon phosphate.
3) Electrodes enzymatiques:
0,30 mg d'enzyme (= 51 U) sont dissous dans 50 >J1 de tampon
phosphate 0,1 M et fixé directement à la surface de l'électrode par
l'intermédiaire d'une membrane de dialyse (cellophane 50 Jm) . L'élec
trode est conditionnée pendant 2 heures et stockée dans ce même tam
pon.

DETERMINATION DE L'ACIDE L-ASCORBIQUE 1597
à_£[!22SS_iE!i!2èili2ÎÊ_E§E_liâi^2G^ covalentes:
Deux méchodes d'immobilisation par liaison covalente ont été
réalisées: l'une par un greffage direct de l'enzyme sur un support
insoluble , l'autre par coréticulation avec une protéine inerte 1 9
selon Lubrano et Guilbault sur le même support insoluble d'acé
tate de cellulose. ..vS
- Type I: par greffage direct: Une membrane d'acétate de cellulo-2
se (membrane hydrophyle Tacussel) de 1 cm est imprégnée de 50 pl
de tampon phosphate 0,1 M de pH 6,50 contenant 0,30 mg d'ascorbate
ûxydase. La solution est évaporée sous vide à 4°C. La phase de ré-
ticulation est obtenue par adjonction d'une solution à 2,5 % de
glutaraldéhyde dans le même tampon. Après dessiccation sous vide,
la membrane est plongée dans une solution de glycine 0,1 M durant
3 heures à WC et lavée à l'eau distillée jusqu'à ce que l'eau de
ringage ne présente plus d'absorption spectrale à 280 nm.
- Type II: par coréticulation sur support insoluble: La membrane
d'acétate de cellulose est imprégnée de 14 JJI d'une solution tampon
contenant 50 mg d'albumine d'oeuf et 21,4 mg d'enzyme par ml et de
3 pl d'une solution à 2,5 % de glutaraldéhyde. La membrane est des
séchée à température ambiante durant 1 à 2 heures.
Les membranes enzymatiques sont conservées dans le tampon pen
dant une heure avant d'être fixées sur l'électrode au moyen d'un
filet de nylon. L'électrode est stockée à 4°C dans le tampon.
RESULTATS ET DISCUSSION
L'influence de l'activité enzymatique sur la pente et la linéa
rité de réponse de l'électrode a été réalisée pour les deux types d'
immobilisation enzymatique, sur le nombre de subcultures de la bacté
rie et sur l'épaisseur des coupes de mésocarpe de concombre.
Pour l'enzyme immobilisée, la pente maximale est obtenue à
partir de 10 U d'ascorbate oxydase dans la membrane, indépendament
du type d'immobilisation.Le temps de réponse de l'électrode est

I S W VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
indépendant de la concentration en enzyme dans la gamme de 10 à
170 U et du type d'immobilisation (type I ou II).
Pour l'électrode tissulaire, les activités enzymatiques sont
fort variables d'un concombre à l'autre, comme le montre la pente
maximale qui varie de 5,0 à 6,6ApX/mM, mais le domaine de linéarité
reste constant. Cependant, la pente maximale est constante au sein
d'un même concombre et est obtenue à partir d'une épaisseur de 400 jjm.
Le temps de réponse en phase stationnaire varie au maximum de 1,5 %
entre des épaisseurs de 400 à 1.450 ^m.
En ce qui concerne les électrodes bactériennes, l'ascorbate
oxydase a une activité très variable selon les souches d'Enterobacter
agglomerans testées, mais cette activité est très stable pour une
même souche et s'exprime dès le premier ensemencement en milieu d'
induction. Seule la souche présentant le maximum d'activité a été
utilisée. Lorsque la bactérie croît en milieu B.H.I. avec ou sans
inducteur, la pente de l'électrode diminue de plus de 60 % impli
quant la nécessité d'un milieu pauvre pour effectuer l'induction
enzymatique.
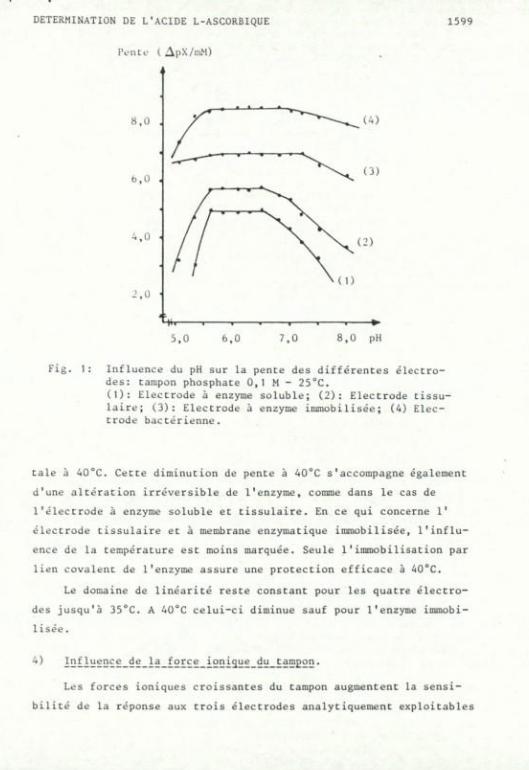
L'effet du pH a été étudié sur les 4 types d'électrodes. Pour
chacune le pH influence peu le domaine de linéarité. Cependant, comme
le montre la figure 1, l'étude de la pente de réponse en fonction du
pH fait apparaître deux types de courbes: celles de l'électrode à
enzyme soluble et à fragment tissulaire qui montrent une forte dé
pendance du pH, de part et d'autre du domaine de pH 5,60 à 6,60, et
celles correspondant aux électrodes bactériennes et à enzymes immobi
lisées mettent en évidence une plus large gamme de pH exploitables
(de pH 5,30 à 7,20). Pour les quatre électrodes, le même pH d'une
valeur de 6,50 peut être utilisé avec succès.
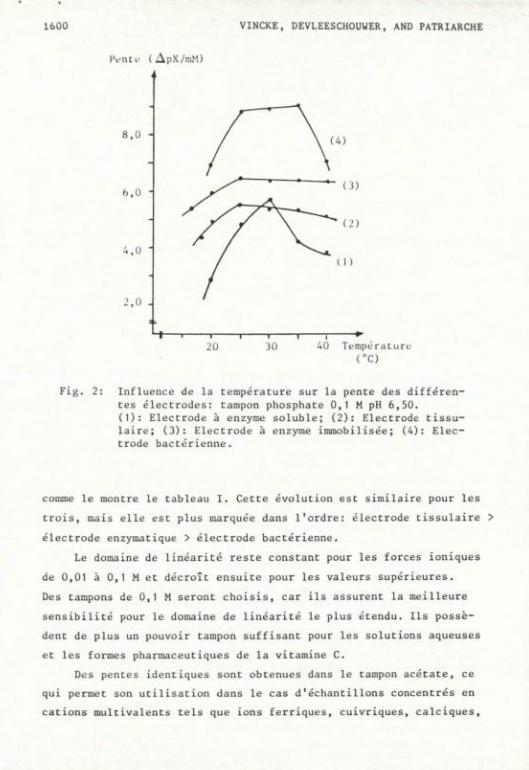
^) ISfiyS2£Ê de_la_tem2érature. '
La figure 2 démontre une influence marquée de la température sur
la réponse de l'enzyme soluble. Pour l'électrode bactérienne, l'on
observe une stabilité de pente entre 25 et 35°C avec une chute bru-
DETERMINATION DE L'A
l'eiUe (
8,0
6,0
H,0
2,0
5,0 6,0 7,0 8,0 pH
Fig. 1 : Influence du pH sur la pente des différentes électrodes: tampon phosphate 0,1 M - 25°C. (1): Electrode à enzyme soluble; (2): Electrode tissu-laire; (3): Electrode à enzyme immobilisée; (4) Electrode bactérienne. , ,
taie à 40°C. Cette diminution de pente à 40°C s'accompagne également
d'une altération irréversible de l'enzyme, comme dans le cas de
l'électrode à enzyme soluble et tissulaire. En ce qui concerne 1'
électrode tissulaire et à membrane enzymatique immobilisée, l'influ
ence de la température est moins marquée. Seule l'immobilisation par
lien covalent de l'enzyme assure une protection efficace à 40°C.
Le domaine de linéarité reste constant pour les quatre électro
des jusqu'à 35°C. A AO^C celui-ci diminue sauf pour l'enzyme immobi
lisée .
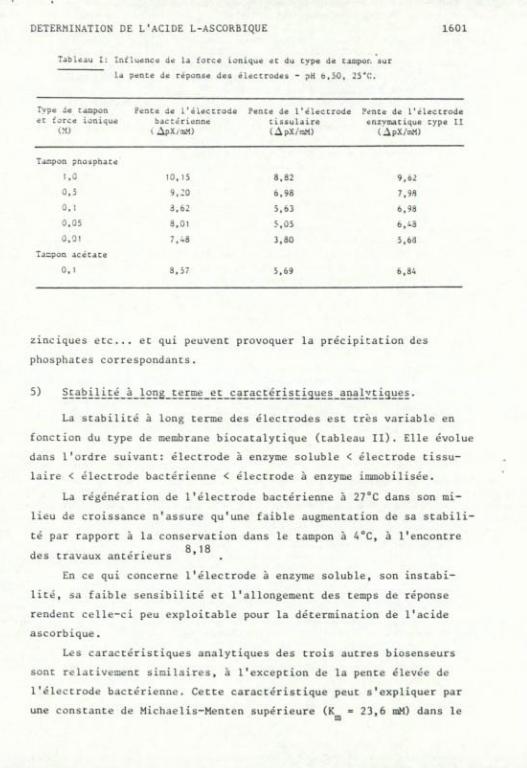
Les forces ioniques croissantes du tampon augmentent la sensi
bilité de la réponse aux trois électrodes analytiquement exploitables
1 6 0 0 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
Pi-nti" (ApX/mM)
. H 1 1 1 1 1 i ^ 20 30 40 Température
(°C)
Fig. 2: Influence de la température sur la pente des différentes électrodes: tampon phosphate 0,1 M pH 6,50. (1): Electrode à enzyme soluble; (2): Electrode tissu-laire; (3): Electrode à enzyme immobilisée; (A): Electrode bactérienne.
comme le montre le tableau I. Cette évolution est similaire pour les
trois, mais elle est plus marquée dans l'ordre: électrode tissulaire >
électrode enzymatique > électrode bactérienne.
Le domaine de linéarité reste constant pour les forces ioniques
de 0,01 à 0,1 M et décroît ensuite pour les valeurs supérieures.
Des tampons de 0,1 M seront choisis, car ils assurent la meilleure
sensibilité pour le domaine de linéarité le plus étendu. Ils possè
dent de plus un pouvoir tampon suffisant pour les solutions aqueuses
et les formes pharmaceutiques de la vitamine C.
Des pentes identiques sont obtenues dans le tampon acétate, ce
qui permet son utilisation dans le cas d'échantillons concentrés en
cations multivalents tels que ions ferriques, cuivriques, calciques.
DETERMINATION DE L'ACIDE L-ASCORBIQUE 1601
Tableau I; Influence de la torce ionique et du cype de tampor. sur
la pence de réponse des électrodes - pH o,5û, 25''C.
Type de cacipon ec force ionique
I.M)
Pence de L'éleccrode bactérienne l ApX/iM)
Pence de l'éleccrode cissulaire ( ApX/aM)
Pence de l'éleccrode enzymacique cype II
( ApX/mM)
Tampon pho:àphace
1 ,û 10, 15 8,82 9,62 0 ,5 9,:o 6,98 7,98
0 ,1 3,62 5,63 6.98
0 ,05 3,01 5,Û5 6 , i 3 û,Ol 7 , - 8 3,30 5,6a
Tacpon acécace
0, 1 3,57 5,69 6,84
zinciques etc.. et qui peuvent provoquer la précipitation des
phosphates correspondants.
La stabilité à long terme des électrodes est très variable en
fonction du type de membrane biocatalytique (tableau II). Elle évolue
dans l'ordre suivant: électrode à enzyme soluble < électrode tissu
laire < électrode bactérienne < électrode à enzyme immobilisée.
La régénération de l'électrode bactérienne à 27°C dans son rai-
lieu de croissance n'assure qu'une faible augmentation de sa stabili
té par rapport à la conservation dans le tampon à A°C, à l'encontre 8 18
des travaux antérieurs '
En ce qui concerne l'électrode à enzyme soluble, son instabi
lité, sa faible sensibilité et l'allongement des temps de réponse
rendent celle-ci peu exploitable pour la détermination de l'acide
ascorbique.
Les caractéristiques analytiques des trois autres biosenseurs
sont relativement similaires, à l'exception de la pente élevée de
l'électrode bactérienne. Cette caractéristique peut s'expliquer par
une constante de Michaelis-Menten supérieure (K = 23,6 mM) dans le

1 6 0 2 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
Tableau II: Caraccérisciques des électrodes dans le tampon phosphate 0,1M,
25'C. pH 6.50.
Type d'éleccrode Pente Linéarité Temps de réponse Stabilité en et nombre d' (ApX/mM) exprimée à l'état station- jours électrodes testées en moles/L naire exprimé en
minutes
Electrode 8,6 + 0,3 4 lû'*-7 10-* 2,5 a 3,0 10 à n
bactérienne (3)
8,6 (à 4'C)
15 à 16 (par régénération)
Electrode 5,8 0,8 4 10-^-1 .0-5 1,5 à 2,0 5 à 6
tissulaire (9)
Electrode à 0,4 4 ,0-^-5 4,0 à 5,0 2 a 3
enzyme soluble(7)
Electrode à enzy- + 0,2 4 t0"^-9 2,0 à 2,5 16 à 18
ise inmobilisée
type I et II (6)
cas de l'enzyme bactérienne démontré par Volk et Larsen pour
Enterobacter aerogenes par rapport à l'enzyme végétale (K^=1,1 mM)
6) Etude_des interférences. , .i-•
Etant donné que la vitamine C peut être oxydée de façon aspéci-
fique par certaines enzymes, telles les polyphénols oxydases et les 21
cytochromes oxydases en présence de leurs substrats , la présence
de celles -ci a été recherchée pour les trois biocatalyseurs. La
recherche des cytochromes oxydases a été effectuée sur disques R
réactifs d'oxydase test (Biomérieux) et les polyphénols oxydases en
utilisant la L-Dopa comme substrat. Ces réactions sont négatives pour
les différentes électrodes.
Parmi les autres substrats pouvant être oxydés par 1'ascorbate
oxydase tels que l'acide D-isoascorbique, l'acide D-glucoascorbique 22
et l'acide réductinique décrit par Dodds , seul l'acide D-isoas
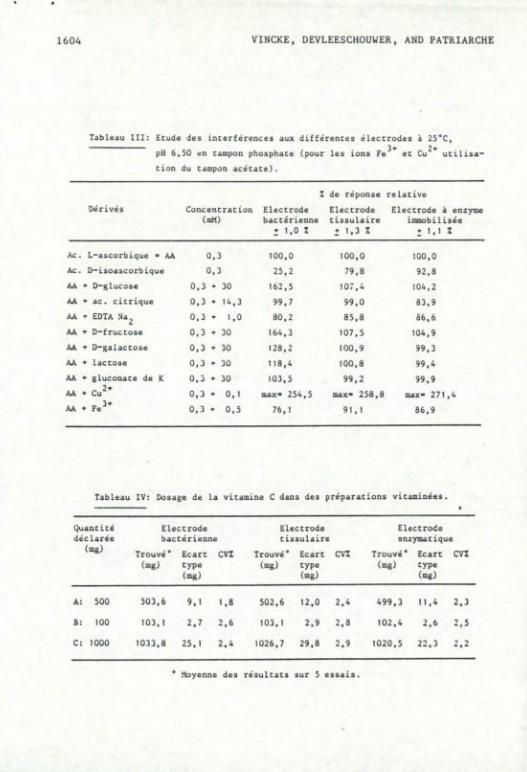
corbique a été étudié. Comme le montre le tableau III, sa réponse •

DETERMINATION DE L'ACIDE L-ASCORBIQUE 1603
varie en fonction de la biomembrane utilisée et est plus faible pour
l'électrode bactérienne.
Les ions cuivriques et ferriques, qui catalysent l'autooxyda-
tion de la vitamine C, interfèrent fortement aux électrodes. Les
ions ferriques diminuent la réponse par leur effet catalytique dans
1'autooxydation de la vitamine C en solution, par contre les ions
cuivriques augmentent la réponse par une oxydation beaucoup plus
rapide impliquant la formation de H.O, au lieu d'H„0 par la réaction
enzymatique ~ . O n obtient en effet un maximum rapide de réponse
qui ensuite décroît progressivement. Le peroxyde d'hydrogène formé
inhibe ensuite l'activité enzymatique de façon irréversible. Aussi
l'action de l'ascorbate oxydase (une cuproprotéine) peut être imitée 21
par des ions cuivriques liés aux protéines inertes . C'est ainsi
qu'une membrane d'albumine liée à 2.10 ^ M de CuSO^, réalisée dans
les mêmes conditions que la membrane de type II, peut oxyder l'acide
ascorbique avec une réponse relative de 30 % par rapport à celle
obtenue par la réaction enzymatique sur la membrane de type II conte
nant 51 U. La cinétique de réponse sera dans ce cas plus lente, d'une
durée de 5 à 6 minutes. 0 n peut distinguer cette oxydation de la
vitamine C par le cuivre de la réaction enzymatique par l'inhibition
de la réponse en présence d'EDTA Na2. En effet, si la réponse à 1' électrode enzymatique est réversiblement diminuée de façon constan-
-5 -3
te pour des concentrations de 1.10 à 1.10 M en EDTA Na2 (tableau
III), la réponse de l'électrode cuivre-protéine inerte est irréversi
blement inhibée de 80 % en présence de 1.10 ^ M en EDTA Na„. 23 •
Gerwin et al. ont montré que le citrate monoanion est un
inhibiteur compétitif de l'ascorbate. L'électrode enzymatique est la
plus sensible à la présence d'acide citrique, mais cette inhibition
est parfaitement réversible.
En ce qui concerne les sucres et alcools rentrant dans la syn
thèse de la vitamine C (D-glucose, L-sorbose, D-sorbitol) ainsi que
d'autres sucres et le gluconate fréquemnentutilisés comme excipients
des formes pharmaceutiques, seuls le glucose et le fructose interfè
rent légèrement et de manière semblable aux électrodes tissulaires
et enzymatiques. Par contre, un plus grand nombre de sucres interfè-
1 6 0 4 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
Tableau III: Etude des InLerférences aux différences éleccrodes à 25*C,
pH 6, 30 ân tampon phosphate (pour les ions Fe 3-» 2«-
et Cu utilisa-tion du tampon acétate).
Z de réponse relative
Dé rivés Concentration Electrode Electrode (mM) bactérienne tissulaire immobilisée
• 1,0 Z 1 1.3 Z î 1,1 I
Ac L-ascorbique - AA 0,3 100,0 100,0 100,0
Ac 0-isoascorbique 0,3 25,2 79,8 92,8
AA •*• EHglucose 0,3 » 30 162,5 107,4 104,2
AA + ac. citrique 0,3 • 14,3 99,7 99,0 83.9
AA * EDTA Na^ 0,3 *• 1,0 80,2 85,8 36,6
AA • D-truccose 0,3 + 30 164,3 107,5 104,9
AA • D-galactose 0,3 » 30 128,2 100,9 99,3
AA + lactose 0,3 •>• 30 118,4 100,8 99,4
AA * gluconate de K 0,3 * 30 103,5 99,2 99,9 -
AA * Cu^* 0,3 • 0,1 oiax- 254,5 max- 258,8 max- 271,4
AA * F e ^ * 0,3 • 0,5 76,1 91,1 86,9
Tableau XV: Dosage de la vicamine C dans des préparations vicamiaées.
Quantité Electrode Electrode Electrode déclarée bactérienne tissulaire enzymatique
(mg) Trouvé ° Ecart cvz Trouvé* Ecart cvt Trouvé* Ecart CVI (mg) type (mg) type (mg) type
(mg) (mg) (mg)
A: 500 503,6 9.1 1.8 502,6 12,0 2,4 499,3 11,4 2.3
B: 100 103,1 2.7 2,6 103,1 2.9 2,3 102,4 2.6 2.5
C: 1000 1033,8 25,1 2,4 1026,7 29,8 2,9 1020,5 22,3 2,2
* Moyenne des résultats sur S essais.

DETERMINATION DE L'ACIDE L-ASCORBIQUE 1^05
renc au niveau de l'électrode bactérienne en raison de leur métabo-
lisation.
Il est possible d'éliminer toutes ces interférences par la
méthode des additions standards à l'exception du cuivre.
7) àE2li£âli2D_âiî_Ë25^S^_'^^^ formes_£harmaceuti2ues.
Le dosage de l'acide L-ascorbique a été réalisé sur divers com
primés par la méthode des additions standards. Les résultats sont
rapportés dans le tableau IV.
Comme le montre le tableau, il existe une excellente corréla
tion entre les résultats obtenus sur les diverses électrodes.
CONCLUSIONS
L'étude comparative des quatre types de biocatalyseurs (enzyme
soluble, enzyme immobilisée, tissus végétaux et cellules bactériennes)
a permis de déterminer que les conditions opératoires optimales pour
chaque type de biocatalyseur sont fort semblables. Certaines diffé
rences ont été mises en évidence, en particulier en ce qui concerne
la stabilité à long terme et les interférences. Seules les électrodes
à enzymes immobilisées, bactériennes et tissulaires de par leur plus
longue stabilité ou leur faible coût sont exploitables analytiquement.
Les résultats obtenus avec celles-ci pour le dosage des formes
pharmaceutiques sont en parfaite corrélation.
REMERCIEMENTS
Nos remerciements vont au F.N.R.S. (Fonds National de la Recher
che Scientifique) pour l'aide apportée à l'un d'entre nous (G.J.P.).
Nos très sincères remerciements vont également au Prof. M. Van
Haelun pour l'aide apportée à l'élaboration des coupes au microtome.
BIBLIOGRAPHIE
1. G.G. GUILBAULT, In "Comprehensive Analytical Chemistry", WILSON and WILSON'S, chap. "Molecular Fluorescence Spectroscopy", vol 8, p. 71-205, G. SVEHLA, Elsevier, Amsterdam, (1977).

1 6 0 6 VINCKE, DEVLEESCHOUWER, AND PATRIARCHE
2. G.G. GUILBAULT, "Analytical uses of Iiranobilizèd Enzymes", chap.3: "Enzyme Eleccrode Probes", p. 112-224, Marcel Dekker Inc., New-York, (1984).
3. G.G. GUILBAULT, Enzyme Microb. Technol., 2, 258, (1980).
4. K. MOSBACH, B. DANIELSSÛN, Anal. Chem., 52, 83 A, (1981). .
5. G.G. GUILBAULT, Anal. Chem., 55, 1682, (1983).
6. M.A. ARNOLD, Anal. Chem., 57, 565, (1985).
7. B.J. VINCKE, M.J. DEVLEESCHOUWER, G.J. PATRIARCHE, Anal. LetCers, \b_, 673, (1983).
8. B.J. VINCKE, M.J. DEVLEESCHOUWER, J. DON-Ï, G.J. PATRIARCHE, Int. J. Pharm., 2_1_. 265, (1984) .
9. B.J. VINCKE, M.J. DEVLEESCHOmjER, G.J. PATRIARCHE, Anal. Letters,
sous presse, (1985).
10. M.A. ARNOLD, G.A. RECHNITZ, Anal. Chem., 5^, 1170, (1980).
11. M.A. ARNOLD, G.A. RECHNITZ, Anal. Chem., 53, 1837, (1981).
12. F. SCHUBERT, D. KIRSTEIN, F. SCHELLER, P. MOHR, Anal. Letters,
22, 1167, (1980). 13. M. CHATEAU-GOSSELIN, G.D. CHRISTIAN, G.J. PATRIARCHE, J. Pharm.
Belg., 22, 19, (1980).
14. A. LECHIEN, P. VALENTA, H.W. NURNBERG, G.J. PATRIARCHE, Fresenius
Z. Anal. Chem., 21i, 105, (1982).
15. K. MATSUMOTO, K. YAMADA, Y. ÛSAJIMA, Anal. Chem., 52, 1974, (1981).
16. M.J. DE\'LEESCHOUWER, J. DONY, J. Pharm. Belg., 2i. 260, (1979).
17. N. SMIT, G.A. RECHNITZ, Biotechnol. Letters, 6, 209, (1984), Chem. Abst. 101: 35283a.
18. C.R. LINDERS, B.J. VINCKE, M.J. DEVLEESCHOUWER, G.J. PATRIARCHE,
J. Pharm. Belg., 40, 19, (1985).
19. G.J. LUBR.-UiO, G.G. GUILBAULT, Anal. Chim. AcCa, 97., 229, ( 1978).
20. W.A. VOLK, J.L. LARSEN, Biochim. Biophys. Acta, 67, 576, (1963). 21. V.S. BUTT, In "The Biochemistry of Plants, a Comprehensive Trea-
cise", chap. "Direct Oxydases and Related Enzymes", vol 2: "Me-tabolism and Respiration", p. 81-123, D.D. DAVIES, Académie Press. New-York, (1980).
22. M.L. DODDS. Arch. Biochem., 28, 51, (1948).
23. B. GERWIN, S.R. BURSTEIN, J. WESTLEY, J. Biol. Chem., 249, 2005, (1974).
Received May 20, 1985 Accepted May 31, 1985