2001 marino azzurro grouper reproduction jfb
TRANSCRIPT

Journal of Fish Biology (2001) 58, 909–927doi:10.1006/jfbi.2000.1502, available online at http://www.idealibrary.com on
Reproduction in the dusky grouper from the southernMediterranean
G. M*‡, E. A*, A. M†, M. G. F* A. M†
*ICRAM, Central Institute for Marine Research, Via Casalotti, 300, 00166 Rome, Italyand †Department of Experimental Biology, University of Genoa, Via le Benedetto XV 5,
16132 Genoa, Italy
(Received 1 April 2000, Accepted 27 October 2000)
Demographic data and gonad histology confirmed that the dusky grouper Epinephelusmarginatus is a protogynous hermaphrodite that follows a monandric pathway to sexualdevelopment. Females reached first sexual maturity at 36·7 cm LS and estimated mean lengthat first maturity (L50) was 43·8 cm LS for females and 81·3 cm LS for males. Adult sex ratiosduring the reproductive period were c. 3·5 : 1 females to males. Females exhibited group-synchronous ovarian development and multiple ovulation occurred over the spawning period.Gonads were ripe from early May and spawning occurred from June until early September.The size of ripe testes (0·6% W) indicated strong oligospermy and suggested a mating systemwith no sperm competition. Sexual transition was protogynous involving regression of ovariantissue and proliferation of testicular tissue in the gonads. Transitional individuals occurredfrom May through November and accounted for 9% of sampled adult population. Sex changeoccurred in fish 69–93 cm (LS) long and the size distributions of males and females overlappedover 27% of the LS range. Special zones were recognized as gathering areas for sexually maturedusky groupers during the reproductive period. � 2001 The Fisheries Society of the British Isles
Key words: reproduction; protogyny; gonad development; groupers; Epinephelus marginatus;Mediterranean Sea.
‡Author to whom correspondence should be addressed. Tel.: +39-06-61570495; fax: +39-06-61561906;email: [email protected]
INTRODUCTION
Groupers (Serranidae, sub-family Epinephelinae) are of considerable economicimportance in tropical, subtropical and temperate fisheries. Typically they aremonandric protogynous hermaphrodites (Smith, 1965; Shapiro, 1987), eventhough cases of gonochorism have been reported recently by Siau (1994) for theblue-spotted grouper Cephalopholis taeniops (Valenciennes) and by Sadovy &Colin (1995) for the Nassau grouper, Epinephelus striatus (Bloch). Despite theirsupposed uniformity of reproductive mode, they show a diverse array of matingsystems and reproductive dynamics, especially in the western Atlantic species.
The dusky grouper Epinephelus marginatus (Lowe), more widely known by itssynonym, Epinephelus guaza, is popular and recreationally important in theMediterranean Sea. Its protogynous hermaphroditism was suggested first bySparta (1935) and demonstrated later by Brusle & Brusle (1975, 1976). Firstsexual maturity in Tunisian waters occurs at age 5 as females (Chauvet, 1988).The sexual cycle and gametogenesis were reviewed by Bouain & Siau (1983) andBrusle (1985).
909
0022–1112/01/040909+19 $35.00/0 � 2001 The Fisheries Society of the British Isles

910 . .
E. marginatus is distributed in the East Atlantic Ocean, from the British Islesto the southern tip of Africa, along the coast of southern Brazil and throughoutthe Mediterranean Sea (Randall & Heemstra, 1993). Much of the southernMediterranean commercial catch is taken during summer aggregations, atspecific sites well known by local fishermen. Although specific fishery statisticsfor this species are not available, E. marginatus is believed to be overexploited inmost Mediterranean areas (Brusle & Brusle, 1976; Chauvet, 1991; Zabala et al.,1997a). It is the only serranid included in Annex 3 of the Berne Convention andin Annex 3 of the Protocol for specially protected areas and Mediterraneanbiodiversity of the Barcelona Convention (1995), as endangered species forwhich management measures should be implemented (Relini, 1999). In allgroupers, fishery management is complicated by sequential hermaphroditism(Bannerot et al., 1987; Shapiro, 1987). An understanding of their sexualsuccession, mating system and the potential for sperm limitation (Bannerot et al.,1987; Sadovy et al., 1994a) is necessary for an evaluation of the impact of fishingon wild populations, to develop appropriate management and conservationmeasures (Huntsman & Schaaf, 1994), to develop reproduction techniques incaptivity (Marino et al., 1998, 2000), and for producing quality seed forenhancing natural stocks (Art. 9·3·5, FAO, 1997).
The present study used histological analyses of gonads, population demo-graphic data and other reproductive parameters to investigate the reproductivebiology of E. marginatus. Reproduction of the dusky grouper in the southernMediterranean was characterized by addressing the following: (1) confirmationof the protogynous sexual pattern; (2) identification of gonad developmentalstages; (3) determination of size at first sexual maturation and size at sex change;(4) determination of the timing and duration of annual spawning activity; and (5)description of a reproductive gathering.
MATERIALS AND METHODS
COLLECTION OF SAMPLESThe study was conducted at the Pelagie Islands (Fig. 1), Mediterranean Sea (35–36� N;
10–14� E), between April 1994 and December 1997. Fishes (n=432) were collected mostlyduring May–August and occasionally during September–February (Table II).
Most specimens (n=380) were caught by professional hook-and-line in offshore waters,at 25–70 m depth and few (n=52) were collected by spearfishing in inshore shallowwaters. Individuals were analysed for: total length (LT) and standard length (LS),measured to the nearest 0·1 cm; body weight (W) and gonad weight (WG) measured to thenearest 0·1 g. Gonads were dissected immediately after capture and analysed macro-scopically on the basis of size, consistency, colour, vascularization, presence of lateralsperm sinuses, ovarian cavity, ovarian lamellae and identifiable oocytes. Apical, centraland caudal portions of gonads were fixed in Bouin’s solution, or in 2·5% glutaraldehydein 0·1 cacodylate buffer at pH 7·2.
GONAD HISTOLOGYGonads from 432 individuals were sexed histologically, but stage of maturity identified
for 405 specimens (Table I), because 27 gonads underwent post-mortem autolysis beforefixation. Gonads embedded in 5 �m paraffin were stained with haematoxylin-eosin andMann Dominici and gonads embedded in 2 �m resin were stained with toluidine blue.Gonads were sexed on the basis of tissue configuration and germ cell development into 13different developmental stages. Ovaries were staged according to the most advanced

911
oocytes (Johnson et al., 1998). Oocytes were classified according to their morphologyand to the presence and position of lipid droplets, yolk vesicles and granules (Mayeret al., 1988). Atresia, extent of vascularization and presence of both free and clusteredmacrophage centres were examined also.
According to Hastings (1981), transitional gonads were defined as those exhibitingregressing ovarian and developing testicular tissue. Three intersexual stages wereidentified by: (1) the proportion of male and female tissue; (2) the extent of degenerationof female germinal cells; (3) the presence of male cysts arranged in lobules; and (4) thedevelopment of sperm sinuses along the gonadal wall.
Those gonads in which a sperm duct was formed and testicular tissue well organized inlobules were classified as testes (Hastings, 1981) and staged on the basis of male germ celldevelopment and presence of sperm within the sperm sinuses.
SEX RATIO AND LENGTHFrequency distributions of standard length (LS) did not differ significantly from
normal (r=0·987) according to the Q-Q correlation test. Since the homogeneity ofvariances of LS was significant (�2
2=52·23, P<0·00, Bartlett’s test), LS values betweensexes were compared using the non-parametric test of Kruskal–Wallis and the Studentt-test for independent samples. Similarly, the Mann–Whitney test was used to comparegonad weights in males, females and transitionals, since the homogeneity of variances ofWG in the three sexual categories was significant (�2
2=61·257, P<0·001).Minimum size at sexual maturation was assessed for 59 males and 219 females, within
a size range of 36·7–105 cm LS. The smallest size (LS) class at which 50% of fish weresexually mature (L50) was determined according to Lowerre-Barbieri et al. (1996), byfitting the logistic function by nonlinear regression (Marquardt method) to the fraction ofmature females and males per 5 cm LS intervals using the SPSS 7·5 Advanced Software(Chicago, Illinois).
Adult sex ratios were calculated during the reproductive period, the non-reproductiveperiod and within a reproductive gathering and compared by applying the �2 test.
4 km
N
73
25 56
50
Lampedusa
300
314
6675
7264
F. 1. Map of study area showing the offshore capture locations for Epinephelus marginatus (�) and agathering area (�). Depths are given in metres.
PERIODICITY OF REPRODUCTION AND SEX CHANGEPeriodicity of reproduction was examined by monthly changes in relative gonad index
(IRG). The gonadosomatic index (de Vlaming et al., 1982) could not be used because the

912 . .
slopes of the regression relating gonad weight to body weight were not homogeneousamong different stages of ovary maturation (F=2·25, P<0·05). An alternative indexwas used, IRG=WG S�bj, proposed by Erickson et al. (1985) where bj is a parameter ofthe linear model. According to present data, IRG=WG S�0·6137 for females and IRG=WG S�1·02 for males. Temporal changes of IRG were compared by applying the �2 test.
Size at sexual transition was expressed as the size (LS) range of transitional individuals(T-range). According to Shapiro (1987) the following were calculated also: extent ofT-range: (T-range: maximum size); size overlap (range over which the LS distribution ofmales overlapped with that of females); range of overlap (overlap: maximum size).Timing of sex change was analyzed by plotting the occurrence of the three transitionalstages against the sampling period.
RESULTS
GONAD MORPHOLOGY
Ovaries and testesThe gonads of dusky grouper were composed of two unequal lobes, which fusein the medial-caudal portion. They were located in the posterior portion of thebody cavity, suspended by mesenteries and extended antero-dorsally along theventral wall of the swim bladder. The urinary bladder was placed postero-ventrally to the gonad and the mesonephric duct inserted anteriorly to theuro-genital papilla in the visceral cavity. In each lobe, ovaries, testes andtransitional gonads had a central lumen and lamellae of connective tissueprojecting into it. In the ovary, each lumen fused posteriorly into a commoncavity, which reduced gradually in size forming a small post-ovarian sinus. Ashort oviduct connected the sinus to the ovipore, which was posterior to the anusand anterior to the genital papilla. Most ovaries contained islets of testiculartissue, mainly represented by resting spermatogonia and a few cysts ofspermatocytes I and II. Testicular islets were intermingled rarely among theoocytes and distributed mostly along the border of the lamellae, in the medialportion of the ovaries.
Ovaries were categorized histologically into seven stages (Table I), whichembraced both ontogenetic changes in immature fish (J, immature females) andseasonal changes in adult females (F1-resting, F2-developing, F3-maturing,F4-mature, F5-partially running, F6-spent) [Fig. 2(a)–(f)].
Testes appeared as small soft grey gonads, sparsely vascularized. The gonadwall, usually thicker than in females, became thinner in spermiating males. Alltestes had a lamellar structure and a lumen, occluded posteriorly by connectivetissue. The duct system for sperm transport was comprised of multi-chamberedsinuses located within the muscular wall, enlarged in the dorsal part of thegonad. Spermatozoa drained peripherally and then collected in the sperm ductwhich extended, posteriorly to the post ovarian sinus through the urogenitalpapilla. Testicular organization was of the unrestricted spermatogonial testis-type (Grier, 1981). According to the degree of maturation of testicular tissue andthe presence of sperm into the sinuses, three maturity stages were identifiedhistologically (M1-maturing, M2-mature and M3-running ripe testes) [Table I;Fig. 3(d)].
Transitional gonadsIn early transition, gonads had asymmetric and compact lobes similar to an
inactive ovary, whereas in more advanced transitional stages these gonads
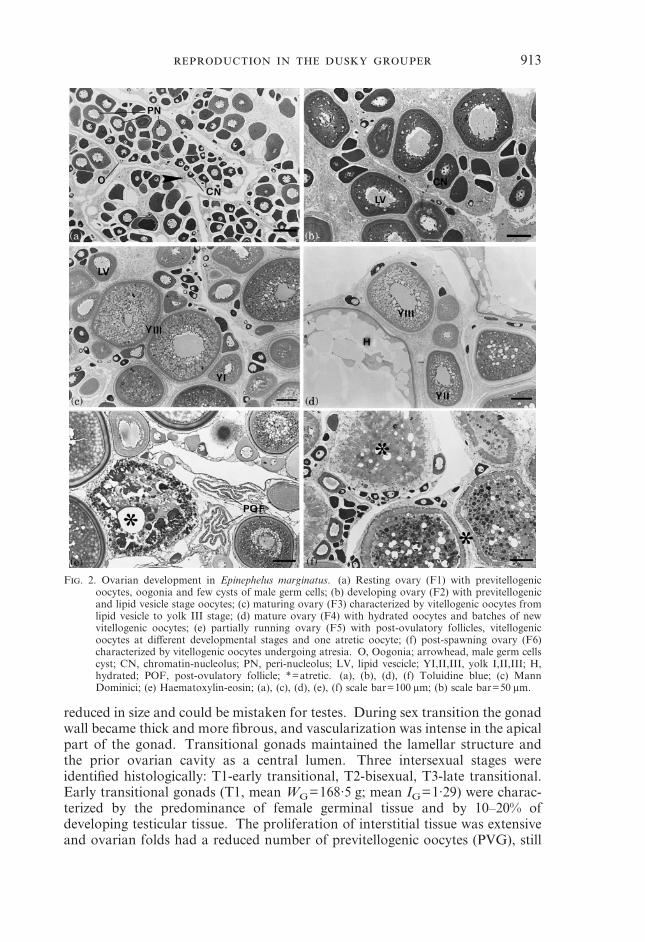
913
F. 2. Ovarian development in Epinephelus marginatus. (a) Resting ovary (F1) with previtellogenicoocytes, oogonia and few cysts of male germ cells; (b) developing ovary (F2) with previtellogenicand lipid vesicle stage oocytes; (c) maturing ovary (F3) characterized by vitellogenic oocytes fromlipid vesicle to yolk III stage; (d) mature ovary (F4) with hydrated oocytes and batches of newvitellogenic oocytes; (e) partially running ovary (F5) with post-ovulatory follicles, vitellogenicoocytes at different developmental stages and one atretic oocyte; (f) post-spawning ovary (F6)characterized by vitellogenic oocytes undergoing atresia. O, Oogonia; arrowhead, male germ cellscyst; CN, chromatin-nucleolus; PN, peri-nucleolus; LV, lipid vescicle; YI,II,III, yolk I,II,III; H,hydrated; POF, post-ovulatory follicle; *=atretic. (a), (b), (d), (f) Toluidine blue; (c) MannDominici; (e) Haematoxylin-eosin; (a), (c), (d), (e), (f) scale bar=100 �m; (b) scale bar=50 �m.
reduced in size and could be mistaken for testes. During sex transition the gonadwall became thick and more fibrous, and vascularization was intense in the apicalpart of the gonad. Transitional gonads maintained the lamellar structure andthe prior ovarian cavity as a central lumen. Three intersexual stages wereidentified histologically: T1-early transitional, T2-bisexual, T3-late transitional.Early transitional gonads (T1, mean WG=168·5 g; mean IG=1·29) were charac-terized by the predominance of female germinal tissue and by 10–20% ofdeveloping testicular tissue. The proliferation of interstitial tissue was extensiveand ovarian folds had a reduced number of previtellogenic oocytes (PVG), still

T
I.D
escr
ipti
onof
the
deve
lopm
enta
lst
ages
ofov
ary
and
test
isof
Epi
neph
elus
mar
gina
tus
inth
eP
elag
ieIs
land
s
Mac
rosc
opic
desc
ript
ion
Mic
rosc
opic
desc
ript
ion
J-Im
mat
ure
fem
ale
Smal
l,co
mpa
ctan
dtr
ansl
ucen
tbi
-lob
edgo
nad
adhe
rent
toth
esw
im-b
ladd
erw
all.
Ova
rian
cavi
tyan
dov
iger
ous
fold
sno
tdi
stin
guis
habl
e(m
ean
WG
=4·
1g;
mea
nI R
G=
0·26
)
Shor
tan
dcl
osel
ypa
cked
ovig
erou
sfo
lds,
proj
ecti
ngin
toth
eov
aria
nca
vity
.P
rim
ary
germ
cells
(PG
C),
oogo
nia
(O)
and
smal
lpr
evit
ello
geni
coo
cyte
s(P
VG
)bo
thal
ong
the
edge
and
the
axis
ofth
ela
mel
lae.
Few
mal
ege
rmce
llcy
sts
occa
sion
ally
obse
rved
.F
1-R
esti
ngfe
mal
eIn
acti
vefe
mal
es.
Ora
nge-
pink
ovar
y,la
mel
lae
not
dist
ingu
isha
ble
toth
ena
ked
eye
(mea
nW
G=
19·5
g;m
ean
I RG
=0·
39)
PG
C,
Oan
dsm
all
PV
Gin
chro
mat
innu
cleo
lus
stag
e(C
N)
and
inpe
rinu
cleo
lus
stag
e(P
N)
espe
cial
lylo
cate
dal
ong
the
peri
pher
yof
the
ovar
ian
fold
s,m
any
larg
eM
Mc
wer
eof
ten
pres
ent
wit
hin
the
stro
ma
alon
gth
eax
isof
the
fold
s.F
ewcy
sts
ofsp
erm
atog
onia
scat
tere
dam
ong
the
fem
ale
tiss
ue[F
ig.
2(a)
].F
2-D
evel
opin
gfe
mal
eO
vary
pink
inco
lour
wit
hnu
mer
ous
elon
gate
dov
iger
ous
fold
spr
ojec
ting
into
the
ovar
ian
cavi
ty(m
ean
WG
=54
·6g;
mea
nI R
G=
0·93
)
Ooc
ytes
inlip
idve
sicl
est
age
(LV
),an
doc
casi
onal
lyin
CN
stag
e,am
ongs
too
goni
a[F
ig.
2(b)
].B
oth
earl
yan
dla
teP
Noo
cyte
sus
ually
pres
ent.
Rar
ecy
sts
ofm
ale
germ
cells
obse
rved
.V
aria
ble
rate
ofL
Voo
cyte
atre
sia.
F3-
Mat
urin
gfe
mal
eSo
ftov
ary,
pale
-pin
kin
colo
ur,
wit
hop
aque
oocy
tes
clea
rly
visi
ble.
Wel
lde
velo
ped
ovig
erou
sfo
lds.
Dor
sal
enla
rged
bloo
dve
ssel
s(m
ean
WG
=36
5·9
g;m
ean
I RG
=4·
14)
Bat
ches
ofoo
cyte
sin
yolk
gran
ules
I(Y
I),
inyo
lkgr
anul
esII
(YII
)an
din
yolk
gran
ules
III
stag
e(Y
III)
,pr
edom
inan
tam
ongs
tP
VG
and
LV
oocy
tes
[Fig
.2(
c)].
Occ
asio
nal
atre
tic
oocy
tes
pres
ent.
Ver
yfe
wcy
sts
ofm
ale
germ
cells
,co
ntai
ning
sper
mat
ogon
ia,
sper
mat
ocyt
esI
and
IIan
dsp
erm
atid
s,de
tect
able
atth
epe
riph
ery
ofov
iger
ous
fold
s.F
4-M
atur
efe
mal
eSw
olle
nov
arie
s,st
rong
lyas
ymm
etri
cfil
ling
mos
tof
the
body
cavi
ty.
Tra
nslu
cent
hyal
ine
oocy
tes
scat
tere
dam
ongs
tm
any
opaq
ueoo
cyte
sw
ithi
nth
ehy
pert
roph
icla
mel
lae
(mea
nW
G=
730
g;m
ean
I RG
=7·
54)
One
batc
hof
hydr
ated
oocy
tes
(H)
scat
tere
dam
ongs
tgr
oups
ofvi
tello
geni
coo
cyte
sat
diff
eren
tde
velo
pmen
tal
stag
es[F
ig.
2(d)
].F
ewat
reti
cvi
tello
geni
coo
cyte
sde
tect
able
.

T
I.C
onti
nued
Mac
rosc
opic
desc
ript
ion
Mic
rosc
opic
desc
ript
ion
F5-
Par
tial
lyru
nnin
gfe
mal
eO
vige
rous
fold
sw
ith
opaq
ueoo
cyte
svi
sibl
eto
nake
dey
e(m
ean
WG
=42
5·3
g;m
ean
I RG
=5·
3)E
vide
nce
ofsp
awni
ngac
tivi
tyre
veal
edby
the
pres
ence
ofpo
st-o
vula
tory
folli
cles
(PO
F)
amon
gst
new
batc
hes
ofoo
cyte
sin
diff
eren
tst
ages
ofvi
tello
gene
sis,
incl
udin
gon
eba
tch
ofY
III
oocy
tes.
Occ
asio
nally
atre
tic
oocy
tes
[Fig
.2(
e)].
F6-
Pos
t-sp
awni
ngfe
mal
eF
lacc
idov
arie
s,re
ddis
hin
colo
ur,
wit
ha
thic
kene
dw
all.
Shor
tla
mel
lae,
wit
hop
aque
resi
dual
oocy
tes,
proj
ecti
ngin
toa
wid
eov
aria
nca
vity
(mea
nW
G=
59·3
g;m
ean
I RG
=0·
85)
Em
pty
ovig
erou
sfo
lds
wit
hfe
wre
sidu
alyo
lked
oocy
tes
amon
gst
num
erou
sat
reti
cvi
tello
geni
cfo
llicl
es.
Enl
arge
dbl
ood
vess
els
and
eosi
noph
ilic
gran
uloc
ytes
scat
tere
dal
ong
the
stro
ma
[Fig
.2(
f)].
M1-
Mat
urin
gm
ale
Smal
lsy
mm
etri
cgo
nads
delim
ited
bya
fibro
usw
all,
soft
inco
nsis
tenc
e,gr
eyin
colo
ur,
spar
sely
vasc
ular
ized
.C
entr
alca
vity
and
lam
ella
rst
ruct
ure
still
pres
ent.
Rec
ogni
zabl
esp
erm
sinu
ses,
wit
hout
sper
m,
unde
rth
ew
all
(mea
nW
G=
70·5
g;m
ean
I RG
=0·
43)
Lob
ules
cont
aini
ngcy
sts
ofSP
G(w
eakl
yba
soph
ilic
nucl
eian
dgr
anul
arcy
topl
asm
),SP
CI
and
SPC
II(s
tron
gly
baso
phili
cnu
clei
and
wea
kly
stai
ned
cyto
plas
m)
and
SPD
.SP
Zof
ten
clum
ped
insm
all
isle
tscl
ose
toth
elo
bule
wal
l.N
onsp
erm
wit
hin
the
sper
msi
nuse
s.M
2-M
atur
em
ale
Smal
lsy
mm
etri
cgo
nads
delim
ited
bya
thic
kw
all,
soft
inco
nsis
tenc
e,gr
ey-w
hiti
shin
colo
ur.
Cen
tral
cavi
tyan
dla
mel
lar
stru
ctur
est
illpr
esen
t.R
ecog
niza
ble
sper
msi
nuse
sun
der
the
wal
l(m
ean
WG
=78
·5g;
mea
nI R
G=
0·46
)
Enl
arge
dlo
bule
sw
ith
cyst
sof
mal
ege
rmce
llsin
all
deve
lopm
enta
lst
ages
and
wit
hSP
Zfr
eein
toth
elu
men
,st
arti
ngfr
omth
ece
ntra
lre
gion
sof
the
fold
san
dex
pand
ing
cent
ripe
tally
.L
ittl
eor
nosp
erm
wit
hin
the
sper
msi
nuse
s[F
ig.
3(d)
].M
3-R
ipe
Ivor
yan
dsy
mm
etri
cgo
nad
delim
ited
bya
thin
wal
l.C
entr
alca
vity
and
lam
ella
rst
ruct
ure
still
pres
ent.
Sper
mpr
esen
tin
toth
esp
erm
sinu
ses
(mea
nW
G=
82·3
g;m
ean
I RG
=0·
46)
Lob
ular
wal
lssu
rrou
ndin
gth
esp
erm
atog
enic
cyst
sof
ten
brok
endo
wn.
Num
erou
sfr
eeSP
Zin
the
larg
elu
men
ofth
elo
bule
san
din
the
sper
mat
icsi
nuse
s.
WG
=go
nad
wei
ght;
I RG
=re
lati
vego
nad
inde
x;M
Mc,
mel
ano-
mac
roph
age
cent
res;
SPC
,sp
erm
atoc
ytes
;SP
D,
sper
mat
ids;
SPG
,sp
erm
atog
onia
;SP
Z,
sper
mat
ozoa
.

916 . .
without any sign of degeneration. Vitellogenic oocytes underwent phase � ofatresia (Hunter & Macewicz, 1985). Blood vessels were hypertrophied, especiallyin correspondence with atretic oocytes and transported small melano-macrophages, which clustered around the atretic oocytes. Many eosinophilicgranulocytes were present in the middle of the folds amongst the connectivetissue. Primordial undifferentiated cells, often grouped in four to five units,increased in number. Cysts of spermatogonia (SPG), spermatocytes I and II(SPC I and II) and spermatids (SPD) were scattered irregularly among the femaletissue, along both the edge and the axis of ovarian lamellae. Small clusters ofspermatozoa were rarely present [Fig. 3(a)].
Bisexual gonads (T2, mean WG=66·6 g; mean IG=0·56 ) contained testiculartissue (from 30 to 70%) not yet organized in lobules. Cysts of spermatogeniccells (from SPG to spermatozoa (SPZ)) were intermingled irregularly withremnant previtellogenic oocytes within the interstitial tissue. Large melano-macrophage centres (MMc) were present typically in high numbers and partiallysubstituted the stroma [Fig. 3(b)].
Late transitional gonads (T3, mean WG=83·2 g; mean IG=0·51) were formedmostly by testicular tissue and by c. 10–20% of residual ovarian tissue, repre-sented by both healthy and atretic previtellogenic oocytes. Testicular tissue wasrearranged almost completely in lobules containing spermatogenic cysts withcells at different stages of development and free spermatozoa into the lumen.MMc were mostly present in the central part of the lamellae and werecharacterized by granular or acellular materials, PAS positive and rich inlipofuscins [Fig. 3(c)]. Within the gonad wall, muscle layers started to splitforming small and narrow spermatic sinuses, still not containing spermatozoa.Although very similar to testes, these gonads were categorized as transitionalssince the incomplete sperm duct formation prevented sperm release in thesespecimens.
REPRODUCTIVE CYCLESummer spawning was indicated by the presence of hydrated oocytes and
confirmed by the values of the relative gonad index (IRG) (Fig. 4). Femalesstarted early vitellogenesis in April and by May 55·1% showed yolked oocytes.Mature ovaries (F4) with hydrated oocytes and running ovaries (F5) withpost-ovulatory follicles were found from early June to early September (TableII). The frequency of stage F2 ovaries throughout the sampling period suggestedthe synchronous beginning of vitellogenic activity. In contrast, cessation ofspawning was asynchronous as indicated by some occasional regressing ovaries(F6) from July until early November.
In females, IRG ranged from 0·056 to 5·95. During the period September–April, IRG was low (0·1, range 0·056–0·184). Overall mean monthly IRG
calculated from the four-year survey, increased significantly (P<0·0007) in June(1·44) from May (0·78), and decreased significantly (P<0·0001) in September–October (0·22) from July. The highest mean values of IRG were observed in July(2·3 and 2·6 in 1995 and 1996, respectively), although these values did not differfrom June and August values in the same year.
In males, IRG values ranged from 0·42 to 1·83 and followed the same trend.Mature males (M2) were found from April to September and running ripe males
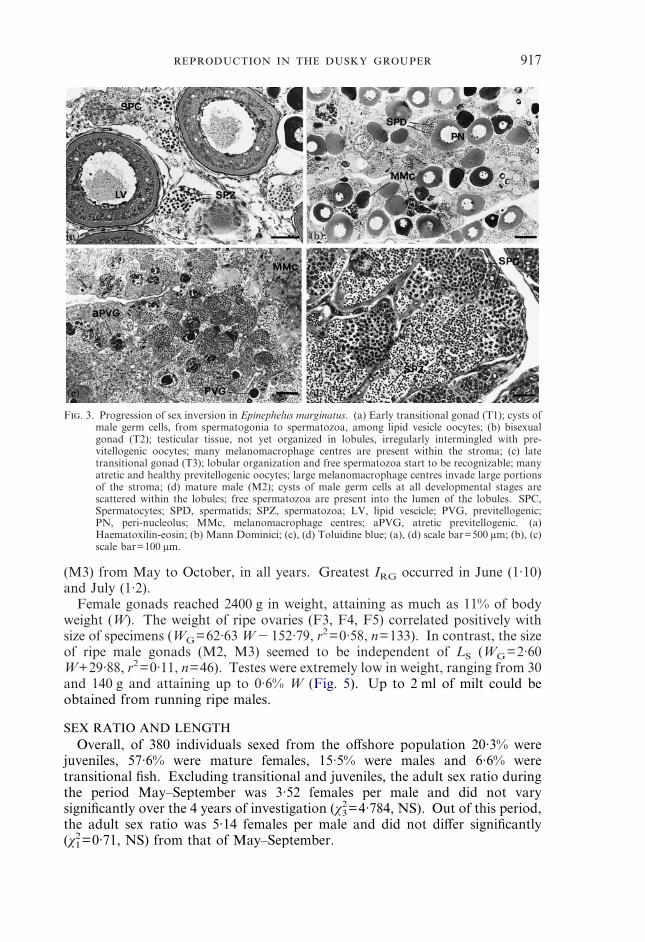
917
(M3) from May to October, in all years. Greatest IRG occurred in June (1·10)and July (1·2).
Female gonads reached 2400 g in weight, attaining as much as 11% of bodyweight (W). The weight of ripe ovaries (F3, F4, F5) correlated positively withsize of specimens (WG=62·63 W�152·79, r2=0·58, n=133). In contrast, the sizeof ripe male gonads (M2, M3) seemed to be independent of LS (WG=2·60W+29·88, r2=0·11, n=46). Testes were extremely low in weight, ranging from 30and 140 g and attaining up to 0·6% W (Fig. 5). Up to 2 ml of milt could beobtained from running ripe males.
F. 3. Progression of sex inversion in Epinephelus marginatus. (a) Early transitional gonad (T1); cysts ofmale germ cells, from spermatogonia to spermatozoa, among lipid vesicle oocytes; (b) bisexualgonad (T2); testicular tissue, not yet organized in lobules, irregularly intermingled with pre-vitellogenic oocytes; many melanomacrophage centres are present within the stroma; (c) latetransitional gonad (T3); lobular organization and free spermatozoa start to be recognizable; manyatretic and healthy previtellogenic oocytes; large melanomacrophage centres invade large portionsof the stroma; (d) mature male (M2); cysts of male germ cells at all developmental stages arescattered within the lobules; free spermatozoa are present into the lumen of the lobules. SPC,Spermatocytes; SPD, spermatids; SPZ, spermatozoa; LV, lipid vescicle; PVG, previtellogenic;PN, peri-nucleolus; MMc, melanomacrophage centres; aPVG, atretic previtellogenic. (a)Haematoxilin-eosin; (b) Mann Dominici; (c), (d) Toluidine blue; (a), (d) scale bar=500 �m; (b), (c)scale bar=100 �m.
SEX RATIO AND LENGTHOverall, of 380 individuals sexed from the offshore population 20·3% were
juveniles, 57·6% were mature females, 15·5% were males and 6·6% weretransitional fish. Excluding transitional and juveniles, the adult sex ratio duringthe period May–September was 3·52 females per male and did not varysignificantly over the 4 years of investigation (�2
3=4·784, NS). Out of this period,the adult sex ratio was 5·14 females per male and did not differ significantly(�2
1=0·71, NS) from that of May–September.
918 . .
The smallest juvenile analysed was 24·5 cm LS. Functional females rangedfrom 36·7 to 97·0 cm LS and males from 68·5 to 105·0 cm (Fig. 6). Estimatedmean length at first maturity (L50) was 43·8 cm LS for females and 81·3 cm LS formales (Fig. 7). The mean LS was different in the three different sexual categories(Table III), with females (67·8�12·7 cm) significantly smaller (P<0·0001) thantransitionals (84·0�6·4 cm) and transitionals significantly smaller (P<0·0001)than males (90·9�5·9 cm).
Out of 52 specimens sampled from inshore shallow waters, 41 were histologi-cally sexed as juveniles, 10 as females and 1 as male.
0
4
I RG
3
2
1
M J J A S O N
1994
M J J A S O N
Months1995
M J J A S O N
1996
D
F. 4. Relative gonadosomatic index and standard deviation for females (n=133) of Epinephelusmarginatus captured off the Pelagie Islands during May–November 1994–1996. Data is givenbiweekly.
SEXUAL SUCCESSIONDemographic features and gonadal analyses indicated a monandric
protogynous pattern of sexual development in the dusky grouper. No maleswere found in small size classes and all juveniles showed female gonads withprevitellogenic oocytes. Moreover, males smaller than the smallest functionalfemale were never observed.
Sex changing specimens represented 9% of adult population (25 of 278) andwere recorded in an LS range of 69–93 cm (T-range). This range can beconsidered representative of the size at which sex change usually occurs in thepopulation. The extent of T-range was 22·1%, the overlap between female andmale LS distributions of 68·5–97·0 cm and its extent was 27·1% (Table III). Ofsex changing individuals, 52% were in early transition (stage T1), 20% werebisexual (stage T2) and 28% were in late transition (stage T3). The frequencydistributions of the three transitional stages were not significantly different(�2
4=5·85, NS) for the different LS classes. All the transitional individuals werefound from May to July. The frequency distribution of the three transitionalstages in the different months were statistically non-homogeneous (�2
4=16·80;

T
II.
Mon
thly
freq
uenc
yof
mat
urit
yst
ages
offe
mal
e,tr
ansi
tion
alan
dm
ale
ofE
pine
phel
usm
argi
natu
sin
the
Pel
agie
Isla
nds
Mon
thF
emal
esT
rans
itio
nals
Mal
esT
otal
JF
1F
2F
3F
4F
5F
6T
1T
2T
3M
1M
2M
3
n1
1Ja
n.(%
)(1
00)
n1
11
3F
eb.
(%)
(33·
3)(3
3·3)
(33·
3)n
12
14
8A
pr.
(%)
(12·
5)(2
5)(1
2·5)
(50)
n6
1627
122
12
121
79M
ay(%
)(7
·6)
(20·
3)(3
4·2)
(15·
2)(2
·5)
(1·3
)(2
·5)
(15·
2)(1
·3)
n30
438
714
13
213
112
Jun.
(%)
(26·
8)(3
·6)
(33·
9)(6
·3)
(12·
5)(0
·9)
(2·7
)(1
·8)
(11·
6)n
132
467
81
22
116
210
0Ju
l.(%
)(1
3)(2
)(4
6)(7
)(8
)(1
)(2
)(2
)(1
)(1
6)(2
)n
35
11
31
115
Aug
.(%
)(2
0·0)
(33·
3)(6
·7)
(6·7
)(2
0)(6
·7)
(6·7
)n
91
21
13Se
p.(%
)(6
9·2)
(7·7
)(1
5·4)
(7·7
)n
3010
18
251
Oct
.(%
)(5
8·8)
(19·
6)(2
0)(1
5·7)
(3·9
)n
712
11
21N
ov.
(%)
(33·
3)(5
7·1)
(4·8
)(4
·8)
n2
2D
ec.
(%)
(100
)T
otal
n10
226
2211
715
2415
135
76
476
405
Dat
afr
om19
94–1
997
wer
epo
oled
bym
onth
.

920 . .
P<0·002). All the T1 stages were observed in April–May and stages T2 and T3in June and July, with the exception of one specimen with late transitionalgonads (T3) collected in November.
1500
0
3000
Gon
ad w
eigh
t (g
) 2500
1000
0 5 10 15 20 25 30 35
Body weight (kg)
2000
500
F. 5. Gonad weight of ripe males (�) (n=46) and ripe females (�) (n=133) against body weight inEpinephelus marginatus captured off the Pelagie Islands during May–November, 1994–1996.
6
0
12
Per
cen
t fr
equ
ency
10
4
35 40 50 55 60 70 80 90
Standard length (cm)
8
2
45 65 75 85 95 105100
F. 6. Size-frequency distribution for sexually mature females ( ), males ( ) and transitionals ( )(n=303) of Epinephelus marginatus, as determined histologically. Fish were captured off thePelagie Islands during April–December, 1994–1997.
SEASONAL GATHERINGOff the island of Lampedusa, from late April to the end of August, typically
local fishing boats catch large numbers of groupers in traditional locations,characterized as both cliffs and flat rocky platforms ranging from the surface to60–80 m. According to local fishermen’s reports, dusky grouper concentrationincreases in April–May and reaches maximum fish density in June–July. Somealong-shore sites of aggregation have been reported at depths of 20–30 m,although today most of the fishing areas are quite distant from the island(Fig. 1).
Between 1–3 July 1996, during the full moon, a gathering area was identified4 miles east of Lampedusa at 60–70 m depth. A single boat caught 46 individualscomprising 32 functional females, 3 transitionals, 8 functional males and

921
3 juveniles. The sex ratio, 1 : 4 males : females, was not significantly differentfrom the adult sex ratio calculated for fish captured during the reproductiveseasons (�2
1=0·078, NS). Gonad histology indicated the presence of fish at anadvanced spawning stage. Most of the aggregating females had maturing ovaries(F3), one had hydrated oocytes (F4) and four had gonads showing signs of arecent spawning (F5). Only one male had running ripe testes (M3), all the othershad mature testes (M2).
0
100
50
0 40 60 80 120
Standard length
10020
0
100
50
0 40 60 80 12010020
(a)
(b)
Per
cen
t m
atu
re
F. 7. Per cent mature female (a) and male (b) Epinephelus marginatus by 5 cm standard length intervals,fitted to a logistic function. Lines indicate median. (a) L50=43·8; n=219; r2=0·8; (b) L50=81·3;n=59, r2=0·9.
DISCUSSION
Protogynous hermaphroditism in the dusky grouper is indicated strongly bythree features (Sadovy & Shapiro, 1987): the occurrence of transitional individ-uals displaying inverting gonads, the presence of spermatic sinuses in the wall of
922 . .
T III. Size ranges (LS) of females, males, and transitionals of Epinephelus marginatusin the Pelagie Islands
Females Males Transitionals OverlapaExtent ofoverlapb
(%)
Extent ofT-rangec
(%)
Min. LS (cm) 36·7 68·5 69·0 68·5 27·1 22·9Max. LS (cm) 97·0 105·0 93·0 97·0Mean LS (cm) 67·8 90·9 84·0 82·2.. 0·85 0·76 1·28 0·60n 219 59 25 196
aOverlap of male and female length distributions; brange of overlap divided by maximum size; csizerange of transitional fish divided by maximum size.
male gonads and the presence, in all analysed testes, of a membrane lined centrallumen, not used for sperm transport. Also, sexual succession is supported by lessspecific features such as the lamellar organization of male gonads and a bimodalsize frequency distribution, with males significantly larger than females. Allthese findings confirm previous indications of protogyny for this species (Brusle& Brusle, 1975, 1976). Histological and demographic data also support amonandric pattern of development. Male gonads show no differences in weight,in general morphology or in the duct structures, suggesting the occurrence ofonly one type of testis, developed ontogenetically from a pre-existing ovarianstate. In several protogynous species and in some groupers sex change may takeplace before the female functions as an adult, in Cephalopsis taeniops, inPlectropomus maculatus (Bloch) and in Epinephelus morio (Valenciennes)(Siau, 1994; Ferreira, 1993; Moe, 1969), or directly in the juvenile phase, as inE. striatus (Sadovy & Colin, 1995). Unlike Brusle & Brusle (1976) no maleE. marginatus was found any smaller than the smallest functional female, andthus, alternative developmental sex sequences of males are likely to be rare, ifthey occur at all.
The configuration of germinal tissue of the gonads of E. marginatus falls intothe undelimited type 2 (Sadovy & Shapiro, 1987) in which male and femaletissues are not separated by connective tissue and are intermixed during sexreversal. The coexistence of both germinal tissues is not exclusive to the processof sexual transition, since small amounts of non functional male tissue are foundcommonly in the ovary of both immature and mature females [Fig. 2(a)]. Thispattern of germinal tissue configuration is not common in protogynousmonandric species, in which gonads of initial sex are generally purely ovarian,with no trace of testicular tissue (Sadovy & Shapiro, 1987). Female germinalcells have never been recorded in active testes, which fails to support thehypothesis that previtellogenic oocytes may differentiate for several years insidefunctional testes of E. marginatus (Brusle & Brusle, 1975).
First sexual maturity is reached at 36·7 cm LS in females and at 68·5 cm LS inmales and corresponds to an age of 5 and 12 years, respectively (data notpresented). These results agree with those reported by Bouain (1980) andChauvet (1988) for dusky groupers collected in the same continental shelf area.

923
The size at which sex change usually occurs can be derived from the size rangeof transitional individuals as well as from the extent of overlap of male andfemale sizes (Shapiro, 1987). Given the seasonal nature of sampling and therelatively small number of transitional individuals, the extent of overlap and theextent of T-range were used, to estimate the size range of occurrence of sexchange in E. marginatus. The extent of T-range was 22·9% and the extent ofoverlap 27·1%. Even if both indexes are far below the values of 58% and33–100%, respectively, calculated by Shapiro (1987) for other congeners, theyindicate that sex change in dusky grouper does not occur at any characteristicsize in the whole population. Moreover, in the specimens analysed sex changeoccurred in fish 69–93 cm LS long, a quite different size range from the68–110 cm (LT) range reported by Brusle & Brusle (1975). This difference in thesize and age at which females change sex supports the idea of a behaviouralinduction of sex change (Shapiro, 1987).
Some dusky grouper females approached the size of the largest males(95–100 cm LS) and over the size range of transitional individuals (the largestfemale was 97 cm LS, 20·6 kg W; the largest transitional was 93 cm LS, 16 kg W).The presence of large females (i.e. 20 kg W and 100 cm LT) was recorded also inTunisian coastal waters (Brusle & Brusle 1975, 1976), in Croatia (Skaramucaet al., 1989) and in the Medes Islands Marine Reserve (Zabala et al., 1997a). Theretaining of the initial sex in these large individuals could be due to lack ofenvironmental cues, either social or physical (Sadovy & Shapiro, 1987) or agenetic inability to undergo sex-change as suspected for E. marginatus (Brusle &Brusle, 1975) and for E. morio (Moe, 1969). However, since large dusky grouperfemales were never greater in size than the larger males (Brusle & Brusle, 1975,1976; Skaramuca et al., 1989; present study), their occurrence is not conclusiveevidence for considering E. marginatus as an incompletely metagonous species(Smith, 1965).
The reproductive season of dusky grouper lasts almost three months and mostactive spawning occurs between late June and early August. Multiple spawningduring this period is indicated by group-synchronous oocyte development and bythe occurrence of post-ovulatory follicles in running females. Batches ofvitellogenic oocytes also co-occur with hydrated oocytes in mature females. Theoccurrence of mature and running females between June and September, ofrunning ripe males from May until October, and of completely spent females inOctober, suggests a longer spawning season for dusky grouper than previouslyreported (Barnabe, 1974; Bouain, 1980; Brusle, 1985; Chauvet, 1988).
In most protogynous fish, sexual transition occurs outside the breeding season(Shapiro, 1987; Siau, 1994) and in many species of Serranidae it starts afterspawning (Moe, 1969; Sadovy & Shapiro, 1987; Ferreira, 1993; D. Y. Shapiro,unpubl. data). According to Shapiro (unpubl. data), sex change in groupers is arelatively rapid process (180 days), and in dusky grouper can be completedwithin one year, between two reproduction seasons (Brusle, 1985). The timingand duration of sex change in dusky grouper cannot be determined definitivelyfrom present data owing to the small number of individuals collected during thewinter. However, some indications emerge from the present study: (1) earlytransitional gonads have developing oocytes scattered among cysts of developingmale tissue, indicating that simultaneous development of both female and male

924 . .
germinal elements may occur in dusky grouper, as also reported for the redgrouper E. morio (Johnson et al., 1998); (2) sexual transition could occur infemales two months before the peak of gonad ripeness, as in E. morio (Johnsonet al., 1998); (3) the concomitant occurrence of all early transitionals in lateApril–May, and almost all bisexuals and late transitionals from June–July (TableII) suggests that at least part of the process of sex reversal may occur during thereproductive period. This supports the arguments of Shapiro (1987) that sexchange in groupers is not restricted to the immediate post-spawning period, butoccurs throughout much of the year.
In many teleosts, the size of ripe testes has been used to suggest the type ofmating system (Warner, 1984), extremely small testes suggesting the absence ofsperm competition (Stockley et al., 1997). In the present study, ripe testes wereextremely low in weight, attaining only 0·6% W. This finding is in accordancewith the observation that dusky groupers reproduce by pair-spawning insingle-male/multi-female social units (Zabala et al., 1997b), a mating system inwhich sperm competition is unlikely. A similar situation is known for the redhind Epinephelus guttatus (L.), in which males are also characterized by smalltestis size (<1% W) (Shapiro et al., 1993). In the case of species which spawn inmulti-male groups, with high sperm competition, ripe testes account for as muchas 10% of W (e.g. Nassau grouper) during the reproductive season (Sadovy &Colin, 1995).
Seasonal changes in the density of fish have been reported in the MarineNatural Reserve of the Medes Islands (Zabala et al., 1997a), where theabundance of dusky grouper increases five to eight fold during the summer,especially from mid April to late June, declining drastically during the winter.This poorly understood phenomenon has been associated with small-scalemigratory movements of fish, and is defined as a summer gathering (Zabalaet al., 1997a). Also in the Pelagie Islands the highest densities of dusky grouperoccur in the summer and then decline in the winter. Seasonal clumping of fishoccurs from May through August at traditional sites, often known and exploitedby fishermen. One indication of gathering behaviour in dusky grouper is a largenumber of fish (46) being caught in a single limited location in the space of a fewdays, which may be defined as an aggregation. Gonad analysis indicates that thisaggregation can be considered as reproductive, since some fish are ready tospawn and most are at an advanced stage of reproduction.
All intermediate and larger-sized grouper species aggregate to spawn (Sadovyet al., 1994a). In the dusky grouper, the aggregating behaviour described byZabala et al. (1997a, b) and that observed in the Pelagie Islands seems to besubstantially different from the reproductive aggregations of the tropical con-geners, characterized by massive movements, short duration and high densities(Smith, 1972; Colin, 1992; Sadovy et al., 1994b; Shapiro et al., 1994). Itresembles more closely the reproductive gathering described in E. guttatus andE. morio, which are distributed patchily over relatively large areas (Sadovy et al.,1994a). Like dusky grouper, both these species are protogynous and have smallsize testes, indicating the absence of sperm competition (Moe, 1969; Bannerotet al., 1987; Johnson et al., 1998).
Fish concentration renders grouper species prone to overexploitation (Sadovy& Eklund, 1999; Sadovy et al., 1994a,b) and nowadays gathering areas are

925
located easily by the increasing use of echo-sounders and on-board satelliteinstruments, and hence may be targeted by sports and artisan fishermen. Giventhe possible importance of aggregating behaviour for this species’ reproduction,additional details about the reproductive pattern and the impact of fishingactivity on aggregation units are needed to implement effective managementmeasures.
The authors thank P. di Marco for collecting part of these specimens and the fishermenof Lampedusa Island who made possible this study. We wish to thank two anonymousreferees for comments which considerably improved the manuscript. This work wassupported by a research grant from MiPA (low 41/82) to GM. Trials were performed inagreement with the Italian current laws.
References
Bannerot, S., Fox, W. W. Jr & Powers, J. E. (1987). Reproductive strategies and themanagement of snappers and groupers in the Gulf of Mexico and Caribbean. InTropical Snappers and Groupers: Biology and Fisheries Management (Polovina, J. J.& Ralston, S., eds), pp. 561–603. Boulder, Colorado: Westview Press.
Barnabe, G. (1974). La reproduction du merou Epinephelus gigas: observationspreliminaires de terrain. Aquaculture 4, 363–367.
Bouain, A. (1980). Sexualite et cycle sexuel des merous (Poissons, Teleosteens, Ser-ranides) des cotes du Sud tunisien. Bulletin Official National Peches Tunisie 4,215–229.
Bouain, A. & Siau, Y. (1983). Observations on the female reproduction cycle andfecundity of the species of groupers (Epinephelus) from the southeast Tunisianseashores. Marine Biology 73, 211–220.
Brusle, J. (1985). Expose synoptique des donnees biologiques sur les merous Epinephelusaeneus (Geoffroy Saint Hilaire, 1809) et Epinephelus guaza (Linnaeus, 1758) deOcean Atlantique et de la Mediterranee. FAO Synopsis sur les Peches 129, 64 pp.
Brusle, J. & Brusle, S. (1975). Ovarian and testicular intersexuality in protogynousMediterranean groupers, Epinephelus aeneus and Epinephelus guaza. In Inter-sexuality in the Animal Kingdom (Reinboth, R., ed.), pp. 222–227. New York:Springer Verlag.
Brusle, J. & Brusle, S. (1976). Contribution a l’etude de la reproduction de deux especesde Merous, Epinephelus aeneus et Epinephelus guaza des cotes de Tunisie. RevueTravail Institute Peches Maritime 39, 313–320.
Chauvet, C. (1988). Etude de la croissance du merou Epinephelus guaza des cotestunisiennes. Aquatic Living Resources 1, 277–288.
Chauvet, C. (1991). Statut d’ Epinephelus guaza (Linnaeus, 1758) et elements dedynamique des populations mediterranee et atlantique. In Les Especes Marinesa Proteger en Mediterranee (Boudouresque, F., Avon, M. & Graves, V., eds),pp. 255–275. Marseille: GIS Podisonie.
Colin, P. L. (1992). Reproduction of the Nassau grouper, Epinephelus striatus (Pisces:Serranidae) and its relationship to environmental conditions. EnvironmentalBiology of Fishes 34, 357–377.
De Vlaming, V., Grossman, G. & Chapman, F. (1982). On the use of the gonadosomaticindex. Comparative Biochemistry and Physiology 73A, 31–39.
Erickson, D. L., Hightower, J. E. & Grossman, D. (1985). The relative gonad index:an alternative index for quantification of reproductive condition. ComparativeBiochemistry and Physiology 81A, 117–120.
FAO Fisheries Department (1997). Aquaculture development. FAO Technical Guide-lines for Responsible Fisheries 5, 1–24. Rome: FAO.
Ferreira, B. P. (1993). Reproduction of the inshore coral trout Plectropomus maculatus(Perciformes: Serranidae) from the central Great Barrier Reef, Australia. Journalof Fish Biology 42, 831–844.

926 . .
Grier, G. J. (1981). Cellular organization of the testis and spermatogenesis in fishes.American Zoologist 21, 345–357.
Hastings, P. A. (1981). Gonad morphology and sex succession in the protogynoushermaphrodite Hemanthias vivianus (Jordan and Swain). Journal of Fish Biology18, 443–454.
Hunter, J. R. & Macewicz, B. J. (1985). Measurement of spawning frequency in multiplespawning fishes. In An Egg Production Method for Estimating Spawning Biomassof Pelagic Fish: Application to the Northern Anchovy, Engraulis mordax (Lasker,R., ed.), pp. 79–94. U.S. Department of Commerce: NOAA Technical ReportNMFS 36.
Huntsman, G. R. & Schaaf, W. E. (1994). Simulation of the impact of fishing onreproduction of a protogynous grouper, the graysby. North American Journal ofFisheries Management 14, 41–52.
Johnson, A. K., Thomas, P. & Wilson, R. R. Jr (1998). Seasonal cycles of gonaddevelopment and plasma sex steroid levels in Epinephelus morio, a protogynousgrouper in the eastern Gulf of Mexico. Journal of Fish Biology 52, 502–518.
Lowerre-Barbieri, S. K., Chittenden, M. E. Jr & Barbieri, L. R. (1996). The multiplespawning pattern of weakfish in the Chesapeake Bay and Middle Atlantic Bight.Journal of Fish Biology 48, 1139–1163.
Marino, G., Massari, A., Di Marco, P., Azzurro, E., Finoia, M. G. & Mandich, A.(1998). Reproduction of dusky grouper (Epinephelus marginatus, Lowe, 1834) inthe wild and under controlled conditions. Biologia Marina Mediterranea 5,1042–1051.
Marino, G., Azzurro, E., Finola, M. G., Messina, M. J., Massari, A. & Mandich, A.(2000). Recent Advances in Induced Breeding of the Dusky Grouper (Epinephelusmarginatus). Cahiers Options Mediterranees 47, 215–227.
Mayer, I., Shackley, S. E. & Ryland, J. S. (1988). Aspects of reproductive biology of thebass, Dicentrarchus labrax L. A histological and histochemical study of oocytedevelopment. Journal of Fish Biology 33, 609–622.
Moe, M. (1969). Biology of the red grouper Epinephelus morio from the eastern Gulf ofMexico. Professional Papers Series, Florida Department of Natural Resources.Marine Research Laboratory 10, 1–95.
Randall, J. E. & Heemstra, P. C. (1993). Groupers of the world (Family Serranidae,Subfamily Epinephelinae). FAO Fisheries Synopsis 125, 382 pp., Rome: FAO.
Relini, G. (1999). Italy and biodiversity conservation in the Mediterranean Sea. BiologiaMarina Mediterranea 6, 151–171.
Sadovy, Y. & Colin, P. L. (1995). Sexual development and sexuality in the Nassaugrouper. Journal of Fish Biology 46, 961–976.
Sadovy, Y. & Eklund, A. M. (1999). Synopsis of biological data on the Nassau grouper,Epinephelus striatus (Block, 1792), and the jewfish, Epinephelus itajara(Lichtenstein, 1822). FAO Fisheries Synopsis 157, 65 pp.
Sadovy, Y. & Shapiro, D. Y. (1987). Criteria for diagnosis of hermaphroditism in fishes.Copeia 1, 136–156.
Sadovy, Y., Rosario, A. & Roman, A. (1994a). Reproduction in an aggregating grouper,the red hind, Epinephelus guttatus. Environmental Biology of Fishes 41, 269–286.
Sadovy, Y., Colin, P. L. & Domeier, M. L. (1994b). Aggregation and spawning in thetiger grouper, Mycteroperca tigris (Pisces: Serranidae). Copeia 2, 511–516.
Shapiro, D. Y. (1987). Reproduction in groupers. In Tropical Snappers and Groupers,Biology and Fishery Management (Polovina, J. J., Ralston, S., eds), pp. 295–327.Boulder, Colorado: Westview Press.
Shapiro, D. Y., Sadovy, Y. & McGee, M. A. (1993). Periodicity of sex change andreproduction in the red hind, Epinephelus guttatus, a protogynous grouper.Bulletin of Marine Science 53, 1151–1162.
Shapiro, D. Y., Garcia-Moliner, G. & Sadovy, Y. (1994). Social system of an inshorestock of the red hind grouper, Epinephelus guttatus (Pisces: Serranidae). Environ-mental Biology of Fishes 41, 415–422.

927
Siau, Y. (1994). Population structure, reproduction and sex change in a tropical EastAtlantic grouper. Journal of Fish Biology 44, 205–221.
Skaramuca, B., Musin, D., Onofri, V. & Caric, M. (1989). A contribution to theknowledge on the spawning time of the dusky grouper (Epinephelus guaza L.).Ichtyologia 21, 79–85.
Smith, C. L. (1965). The pattern of sexuality and the classification of serranid fishes.American Museum Novitates 2207, 1–20.
Smith, C. L. (1972). A spawning aggregation of Nassau Grouper, Epinephelus striatus(Bloch). Transactions of the American Fisheries Society 2, 257–261.
Sparta, A. (1935). Contributo alla conoscenza dello sviluppo nei percidi di uova ovarichemature di Epinephelus guaza L. e stadi post-embrionali e larvali di Epinephelusalexandrinus. Cuv & Val. Comitato Talassografico Italiano Venezia, Memorie 224,1–13.
Stockley, P., Gage, M. J. G., Parker, G. A. & Møller, A. P. (1997). Sperm competitionin fishes: the evolution of testis size and ejaculate characteristics. AmericanNaturalist 149, 933–954.
Warner, R. R. (1984). Mating behaviour and hermaphroditism in coral reef fishes.American Scientist 72, 128–136.
Zabala, M., Louisy, P., Garcia-Rubies, A. & Gracia, V. (1997a). Socio-behavioralcontext of reproduction in the Mediterranean dusky grouper Epinephelusmarginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve(NW Mediterranean, Spain). Scientia Marina 61, 78–79.
Zabala, M., Garcia-Rubies, A., Louisy, P. & Sala, E. (1997b). Spawning behaviour ofthe Mediterranean dusky grouper Epinephelus marginatus (Lowe, 1834) (Pisces,Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean, Spain).Scientia Marina 61, 65–77.