2 3 trees structure and function transpiration in a eucalypt plantation and a savanna in venezuela
TRANSCRIPT

1 23
TreesStructure and Function ISSN 0931-1890Volume 26Number 6 Trees (2012) 26:1759-1769DOI 10.1007/s00468-012-0745-0
Transpiration in a eucalypt plantation anda savanna in Venezuela
Ana Herrera, Rosa Urich, ElizabethRengifo, Caín Ballestrini, ArmandoGonzález & Williams León

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL PAPER
Transpiration in a eucalypt plantation and a savanna in Venezuela
Ana Herrera • Rosa Urich • Elizabeth Rengifo •
Caın Ballestrini • Armando Gonzalez •
Williams Leon
Received: 11 January 2012 / Revised: 4 June 2012 / Accepted: 6 June 2012 / Published online: 28 June 2012
� Springer-Verlag 2012
Abstract In Venezuela 30,000 km2 of land is covered by
savannas, of which 410,000 ha have been planted with
several species and hybrids of Eucalyptus for lumber and
pulp production. Popular concern about possible diminu-
tions in water availability of reservoirs near eucalypt
plantations prompted our interest in measuring water use
by these trees. Since these savannas are markedly seasonal,
the response of species to seasonal drought is important.
We aimed to compare the seasonal changes in single-leaf
and whole-plant transpiration in a seasonally dry savanna
with that of trees of E. urophylla in an experimental
plantation. We also examined the seasonal changes in
xylem water potential and stomatal response to air water-
vapour saturation deficit (D). Transpiration in eucalypts
and the dominant savanna species Trachypogon vestitus
and Curatella americana was evaluated using measure-
ments of leaf gas exchange in all three species, sap flux in
eucalypts, microclimatic variables and allometric and
photometric determinations of green area. In E. urophylla
and T. vestitus, but not in C. americana, stomatal
conductance (gs) proved sensitive to D. Integrated values of
daily courses of transpiration rate were scaled to one ha in a
preliminary approach to estimating ecosystem transpiration
(Eha). The Eha of the savanna (the sum of Eha of T. vestitus
and C. americana) was on average 2.4 times that of
eucalypts during the daytime; when nocturnal eucalypt
transpiration was included, the value was 1.9. The evapo-
transpiration calculated by the Penman–Montieth equation
(ETc) of eucalypts was lower than the savanna all year
round. The reference crop ET (ETo) varied little throughout
the seasons, the highest value occurring in March. The ratio
Eha/ETo for the savanna was on average near one during
the dry season and almost two during the rainy season; the
corresponding value for E. urophylla was 0.6 for both
seasons. The ratio Eha/ETc was on average 0.8 for the
species and the savanna. The cumulative Eha for the days of
measurements was higher in the savanna than in the eu-
calypts during the daytime (39.8 and 17.3 mm, respec-
tively), as was the cumulative ETc (37.5 vs. 20.3 mm).
Measured and calculated cumulative ET in eucalypts,
including nocturnal values, were 22.0 and 28.4 mm,
respectively. At the leaf level, both eucalypts and trees of
C. americana apparently may have accessed water from
deep horizons, since their values of W changed less sea-
sonally than in T. vestitus. At the ecosystem level, the
species that presented the largest changes in transpiration
was T. vestitus, which markedly increased savanna tran-
spiration during the rainy season. Our results suggest that,
for the days of this study, and considering the environ-
mental conditions of the ecosystems studied, the type of
measurements and the scaling procedures, stands of
E. urophylla transpire less water than the savanna.
Keywords Curatella � Eucalyptus urophylla �Trachypogon � Savanna � Transpiration
Communicated by M. Adams.
A. Herrera (&) � R. Urich � C. Ballestrini � A. Gonzalez
Instituto de Biologıa Experimental, Universidad Central
de Venezuela, Caracas 1020, Venezuela
e-mail: [email protected]
E. Rengifo
Centro de Ecologıa, Instituto Venezolano de Investigaciones
Cientıficas, Altos de Pipe, Caracas, Venezuela
W. Leon
Facultad de Ciencias Forestales, Universidad de Los Andes,
Merida, Venezuela
123
Trees (2012) 26:1759–1769
DOI 10.1007/s00468-012-0745-0
Author's personal copy

Introduction
Replacement of grasslands with eucalypt plantations may
result in a change in the water balance, mainly through
increased transpiration and interception (Calder 1986). An
analysis of 504 annual catchment observations showed that
afforestation strongly decreased stream flow within a few
years of planting (Jackson et al. 2005).
Concern about possible diminutions in water availability
of reservoirs near eucalypt plantations has motivated
interest in measuring transpiration and related physiologi-
cal and environmental variables (Whitehead and Beadle
2004). Eucalypts have a reputation for being great water
users, but this contention is not justified in all species or
environments (Calder 1986). In Australia, eucalypts have
been planted to lower the water table and thus help reduce
soil salinity (Calder 1986).
To give just one example of great water expenditure by
eucalypts, water use of trees of E. camaldulensis exceeded
rainfall by 62 % (Calder et al. 1997). In contrast, in
Northern Australia, total water loss of the canopy of a
eucalypt open-forest making part of the tropical savanna
mosaic was 870 mm, evapotranspiration of the understory
contributing 64 % of this flow (Hutley et al. 2000). During
the rainy season, this evapotranspiration was due mainly to
evaporation from the bare soil and transpiration by the C4
grass Sorghum spp., and the annual transpiration of the
trees was lower than in the grass. Indiscriminate specula-
tion concerning the water use of eucalypts may be mis-
leading, as wide variations across species can be expected
in, among other variables, transpirational losses in a given
environment (Calder 1986).
The different components of evapotranspiration are
difficult to measure in a natural forest, as is scaling up the
leaf level measurements of gas exchange with IRGAs to
stand-level data (Kallarackal and Somen 2008). This dif-
ficulty can be partly circumvented by measuring sap flux
density (V) and multiplying V by sapwood area of the
plantation. Other appropriate scalars are total leaf area
(difficult to determine) and stem diameter, which can be
easily measured with less error than sapwood or leaf area
(Hatton et al. 1995). Attempts to extrapolate values of
transpiration measured with porometers to canopy tran-
spiration have met with mixed success because of the
poorly quantified influence of boundary layers in limiting
whole-plant transpiration (Wullschleger et al. 1998).
In Venezuela, 30,000 km2 of land is covered by savan-
nas, of which 410,000 ha have been planted with several
species of Eucalyptus. Near the town of Mapire, Estado
Anzoategui, E. urophylla has been planted at a density of
1,111 trees ha-1 (Gonzalez et al. 2005) for lumber and pulp
production. Eucalypt plantations are usually within a few
km of morichales (palm swamps), which not only constitute
important ecosystems and have high recreational value, but
are also the natural water sources for the town’s domestic
use. Afforestation companies have found resistance on the
part of the population in Mapire to eucalypt growing, since
people fear that the plantations could dry out the moric-
hales. We have no evidence that this has happened since the
eucalypts were planted more than 15 years ago.
The vegetation in the understory of the eucalypt
plantations in Mapire is almost nonexistent; plantations
are surrounded by natural savannas composed mostly of
Trachypogon vestitus (syn. montufari, plumosus and spica-
tus; C4 grass) and Curatella americana (C3 evergreen tree),
these being the most important grass and arboreal compo-
nents of the ecosystem, respectively. In sparsely treed sav-
annas of Estado Anzoategui C. americana represents 3.9 %
and T. vestitus 41 % of the importance value index (Dezzeo
et al. 2008). In trees of C. americana, both dawn and midday
xylem water potential (W) remained relatively unchanged
throughout the year at C-0.5 and -1.0 to -1.5 MPa,
respectively (Goldstein et al. 1990; Medina and Francisco
1994). Transpiration rate was as high in the middle of the
dry season as during the rainy season, which, together with a
relative seasonal constancy in W and leaf renewal during the
dry season, has been attributed to root access to water in
deep soil layers (Goldstein et al. 1990).
In T. vestitus, marked daily changes in W and transpi-
ration rate have been reported (Goldstein et al. 1990).
Tussocks of T. vestitus are perennial; leaves dieback under
drought and re-sprout after the first rains. In the grass
stratum of a Venezuelan savanna composed mostly by
T. vestitus, LAI increased with rains from 0.21 to 7.33 (San
Jose and Medina 1975). A decrease in gs of 50 % with an
increase in D of nearly three times was found in plants of
T. vestitus collected in a Venezuelan savanna and subjected
to drought under controlled conditions (Baruch et al. 1985).
From our own observations for over 10 years, eucalypt
plantations in Mapire apparently maintain a constant leaf
area index (LAI) the year round.
Among the mechanisms of drought avoidance in several
species of eucalypt, deep rooting and a high sensitivity of
gs to D have been identified (Dye and Olbrich 1993;
Phillips et al. 2010; White et al. 2000); therefore, exami-
nation of seasonal changes in gs in response to W and D is
important from the viewpoint of the water status of
eucalypts.
Endeavouring to gather data of relatively easy acquisi-
tion to begin to elucidate whether plantations use more
water than the savanna, we aimed to compare, using
measurements of leaf gas exchange, V and microclimatic
variables, the transpiration of eucalypts in an experimental
plantation in Mapire with that in a savanna. Since these
savannas are markedly seasonal, the response of species to
seasonal drought is important. We predicted that (1) in
1760 Trees (2012) 26:1759–1769
123
Author's personal copy

T. vestitus and E. urophylla, transpiration would diminish
during the dry season due to water deficit in the former,
higher evaporative demand in the latter and a reduction in
LAI in T. vestitus, and (2) in C. americana, no seasonal
changes in transpiration would be found.
Materials and methods
Field site and plant material
This study was carried from January to October 2008.
Measurements in eucalypts were carried out in five trees of
Eucalyptus urophylla S.T. Blake, with an average height of
15 m and a mean diameter at breast height of 35 cm,
experimentally planted in an area of approximately
100 m2; trees were planted in a rough circle at an
approximate distance of 5 m from each other. The location
was the Ricardo Alfonso Rojas Experimental and Techni-
cal School, Mapire, Estado Anzoategui (Venezuela) at
78420N, 64�460W. Measurements in commercial planta-
tions were not feasible because leaves grew out of reach; in
addition, access to commercial plantations is heavily
restricted by owners, and plantations are not guarded
against vandalism or attacks to the equipment by cattle. In
a savanna approximately 2 km away from the experimental
plantation, measurements were done in tussocks of Trac-
hypogon vestitus Andersson (average height 0.5 m) and
trees of Curatella americana L. (average height 6 m). The
other important arboreal component of this savanna,
Byrsonima crassifolia, is present in very low numbers,
although an IVI of 22.5 has been reported for a sampling
area of 475 km2 (Dezzeo et al. 2008).
Soil water content
Soils are ultisols, of the kandistults group, very poor,
excessively drained and with a predominantly sandy or
sandy loam texture between 40 and 100 cm; they show
deposits of kaolinite clay, are deep, friable and much lix-
iviated (Gonzalez et al. 2005). Gravimetric soil water
content was determined as SWC = 100 (fresh weight-dry
weight)/dry weight in samples taken at depths of 10 and
20 cm (five samples per depth), maintained in sealed glass
vials at 10 �C and transported to the laboratory, where
weights were determined. In the trees at the experimental
site and in tussocks in the savanna, the fine-root density at
20 cm was high and almost nonexistent below; roots of
E. urophylla probably explore deeper horizons, but we
were not allowed to excavate the tree root system. We
found in a few individuals in the commercial plantations
with partly exposed roots a pivoting root that apparently
grew very deep into the soil. In a plantation of E. urophylla
in Brazil, 64 % of fine root biomass was found in the first
20 cm (Witschoreck et al. 2003).
Microclimatic variables
Photosynthetic photon flux density (PPFD) was measured
with a 190-S quantum sensor connected to a LI-185 meter
(LI-COR Inc., Lincoln, NE). In the experimental planta-
tion, air temperature and relative humidity were measured
using two HOBO Pro V2 loggers and data dumped with a
HOBO Waterproof Shuttle (Onset Computer Corporation,
Pocasset, MA). In the savanna, air temperature was mea-
sured with HOBOs or with YSI 405 thermistors connected
to a telethermometer (Yellow Springs Instruments, Yellow
Springs, OH) and relative humidity with HOBOs or a hair
strand hygrometer (Abbeon mod. AB167B, Santa Barbara,
CA). Net radiation, wind velocity, air temperature, relative
humidity and rainfall data for two weather stations near the
study site were obtained from the Instituto Nacional de
Meteorologıa e Hidrologıa (INAMEH, Venezuela).
Xylem sap flux velocity
Xylem sap flux velocity (V) was measured with thermal
dissipation probes, TDPs, designed after Granier (1987).
Two TDPs were inserted at breast height in each one of five
trees, thermally insulated and connected to a DL2 data
logger (Dynamax, Houston, TX). Constant voltage was set
up to yield a maximum temperature difference between the
reference and the heating elements of the probe (dTmax) of
approximately 8 �C. The data logger recorded data every
minute and averaged them every 30 min. Records were
taken for a minimum of 72 h; values shown are those
collected 48–72 h after set-up. In this study, records were
not taken for longer because of safety reasons and based on
the observation that in previous determinations of V in
Ficus obtusifolia, also a latex-containing species, constant
dTmax was obtained after 24 h (Ballestrini et al. 2011).
Values of V were converted to transpiration rate (E) by
multiplying V by sapwood area. Sapwood depth was
determined in oblique sections of the trunk.
Xylem water potential
Xylem water potential was measured at 0530–0630 and
1200–1300 h with a pressure chamber (PMS, Corvallis,
OR) in leaf-bearing branches of five different trees, and in
one leaf of five different tussocks.
Leaf area index and ground cover
Leaf area index of commercial plantation eucalypts and
trees of C. americana was measured in November using a
Trees (2012) 26:1759–1769 1761
123
Author's personal copy
LAI-2000 Plant Canopy Analyzer (LI-COR, Lincoln, NE).
For the determination of LAI in T. vestitus, the living mass
of five tussocks per season was harvested, and whole-plant
green leaf area obtained by multiplying the living mass by
the specific leaf area (SLA) determined in a subsample.
The LAI was calculated as LAI = green leaf area/area of
tussock. Ground cover by T. vestitus and C. americana was
estimated by counting the number of individuals in five
plots (5 9 5 m for T. vestitus and 9 9 9 m for C. ameri-
cana), estimating tussock or tree canopy area, multiplying
these values by the number of individuals in the plot and
extrapolating these values to 1 ha. Area of tussocks
was approximated to a circle; canopy area of trees of
C. americana, to polygons of variable shapes.
Leaf gas exchange
Stomatal conductance and instantaneous transpiration rate
(E) were measured with both a CIRAS 2 IRGA connected to
a PLC (B) chamber (PP Systems, Hoddesdon, UK) and an
LC4 IRGA connected to a PLC (B) chamber (Analytical
Development Co., Hitchin, UK). Previous measurements
determined that values obtained with both systems on the
same leaves at the same time were not significantly different.
Leaves of E. urophylla and C. americana used for mea-
surements were 1.5–3.5 m, and 2 m above ground, respec-
tively. Measurements were made under full sun exposure.
Individual and large-scale transpiration
For individuals of all three species, daily transpiration per unit
leaf area was calculated as the integral of the daily courses of
E. Since plant cover varies widely on a large scale, daily
integrated transpiration was calculated on a one hectare basis
(Eha), in a preliminary approach to estimate ecosystem tran-
spiration, by multiplying Ea by plant cover and LAI (savanna
species) and by LAI (eucalypt commercial plantations). Ref-
erence crop ET (ETo) and species evapotranspiration (ETc)
were calculated, with mean leaf resistances in the case of the
species, using the Penman–Monteith equation. Values of cli-
matic variables were obtained from INAMEH.
Statistics
Values at the individual level are presented as mean ± SE
where applicable, one- or two-way- ANOVA were performed
using the Statistica package.
Results
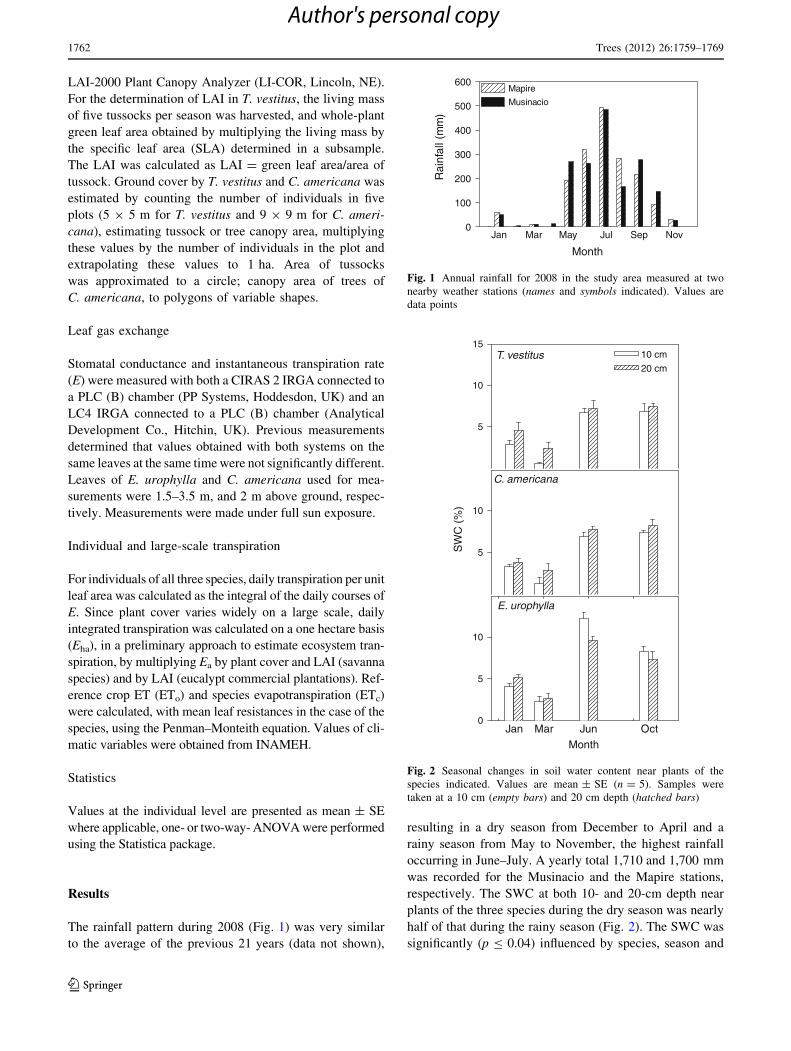
The rainfall pattern during 2008 (Fig. 1) was very similar
to the average of the previous 21 years (data not shown),
resulting in a dry season from December to April and a
rainy season from May to November, the highest rainfall
occurring in June–July. A yearly total 1,710 and 1,700 mm
was recorded for the Musinacio and the Mapire stations,
respectively. The SWC at both 10- and 20-cm depth near
plants of the three species during the dry season was nearly
half of that during the rainy season (Fig. 2). The SWC was
significantly (p B 0.04) influenced by species, season and
Month
Rai
nfal
l (m
m)
0
100
200
300
400
500
600
Jan Mar May Jul Sep Nov
Mapire
Musinacio
Fig. 1 Annual rainfall for 2008 in the study area measured at two
nearby weather stations (names and symbols indicated). Values are
data points
C. americana
SW
C (
%)
5
10
5
10
1510 cm
20 cmT. vestitus
E. urophylla
Month
0
5
10
OctJunMarJan
Fig. 2 Seasonal changes in soil water content near plants of the
species indicated. Values are mean ± SE (n = 5). Samples were
taken at a 10 cm (empty bars) and 20 cm depth (hatched bars)
1762 Trees (2012) 26:1759–1769
123
Author's personal copy
their interaction at both depths. Near the eucalypts, SWC
was twice as high in March and 50 % higher in June than
near the savanna species, differences disappearing in
October. These differences with the savanna could be due
to the plantation being closer to a morichal than the field
site in the savanna.
The SLA of T. vestitus was significantly (p \ 0.05)
higher in March and June (149 cm2 g-1) than in January
(110 cm2 g-1) and October (129 cm2 g-1). From March to
June, SLA in C. americana changed from 103 to
209 cm2 g-1 and in E. urophylla from 94 to 84 cm2 g-1,
changes indicating that leaves were younger (C. ameri-
cana) or older (eucalypts) in March than in June. Green
leaf area per tussock of T. vestitus increased significantly
with rains; consequently, LAI on a stand scale varied from
2.68 (January) to 1.62 (March), 5.08 (June) and 4.77
(October). The canopy remained apparently unchanged in
trees of E. urophylla and C. americana throughout the
period of study. The LAI was 2.77 ± 0.38 in E. urophylla
and 1.14 ± 0.15 in C. americana. Savanna live plant cover
was estimated to vary in T. vestitus from 26,048 (January)
to 15,754 (March), 49,316 (June) and 46,313 (October)
m2 leaf ha-1, and remained apparently unchanged in
C. americana at 5,862 m2 leaf ha-1.
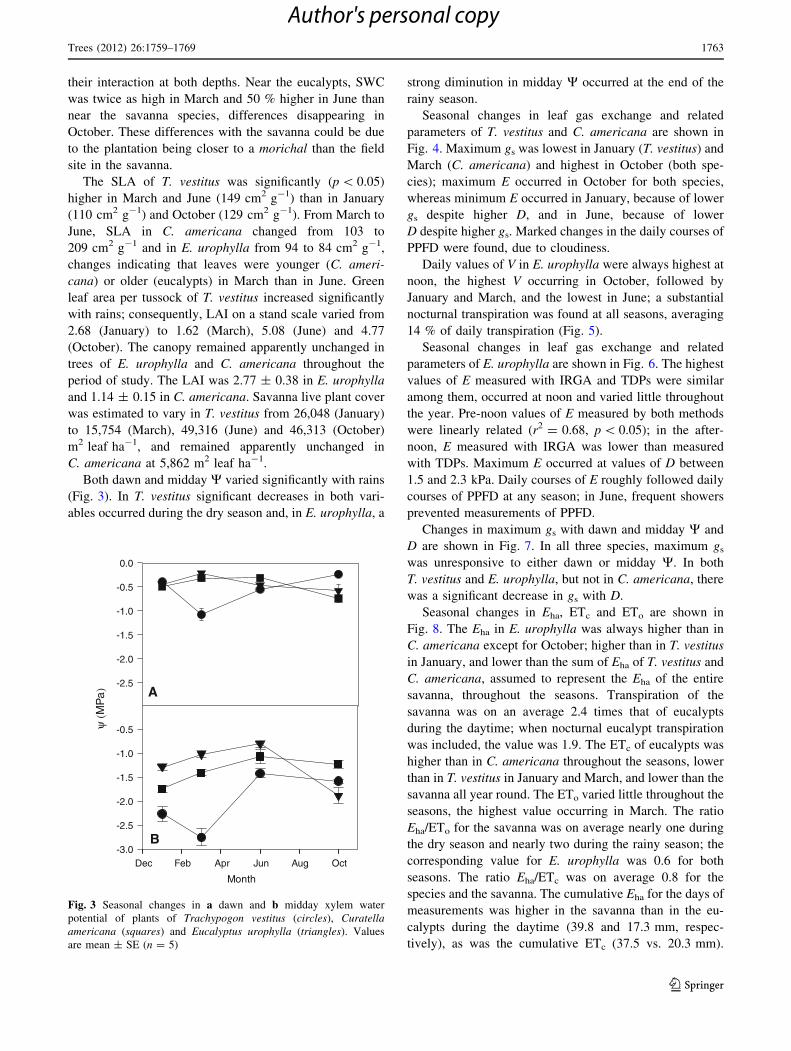
Both dawn and midday W varied significantly with rains
(Fig. 3). In T. vestitus significant decreases in both vari-
ables occurred during the dry season and, in E. urophylla, a
strong diminution in midday W occurred at the end of the
rainy season.
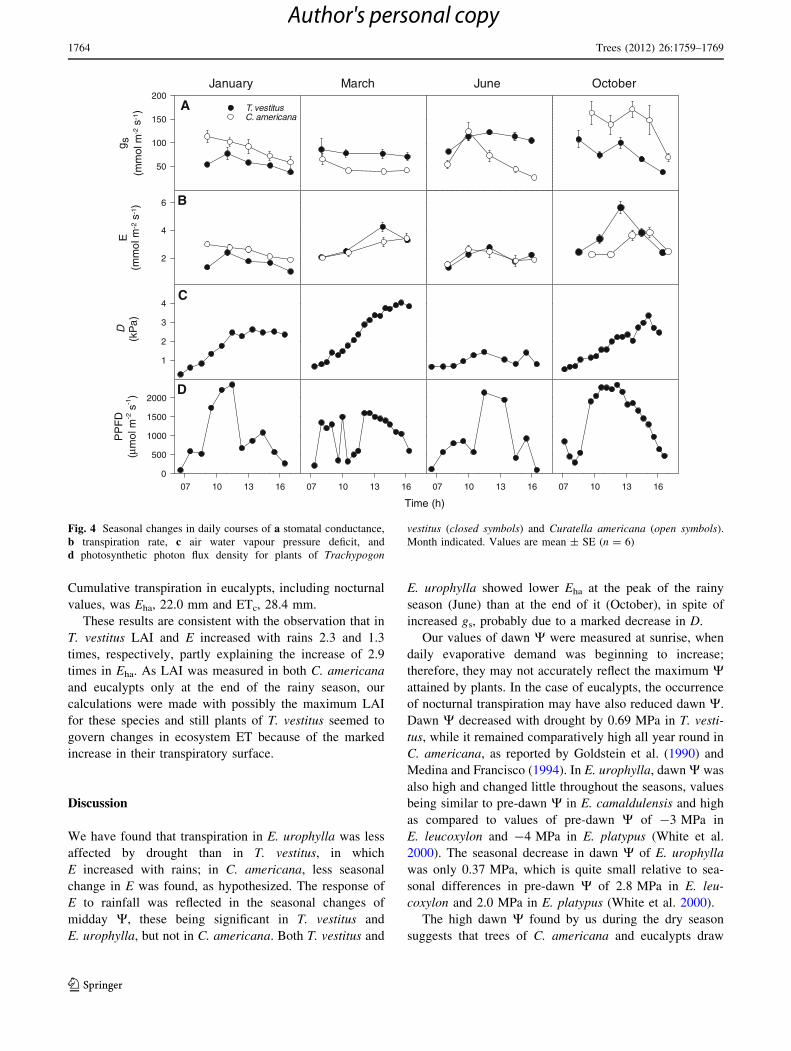
Seasonal changes in leaf gas exchange and related
parameters of T. vestitus and C. americana are shown in
Fig. 4. Maximum gs was lowest in January (T. vestitus) and
March (C. americana) and highest in October (both spe-
cies); maximum E occurred in October for both species,
whereas minimum E occurred in January, because of lower
gs despite higher D, and in June, because of lower
D despite higher gs. Marked changes in the daily courses of
PPFD were found, due to cloudiness.
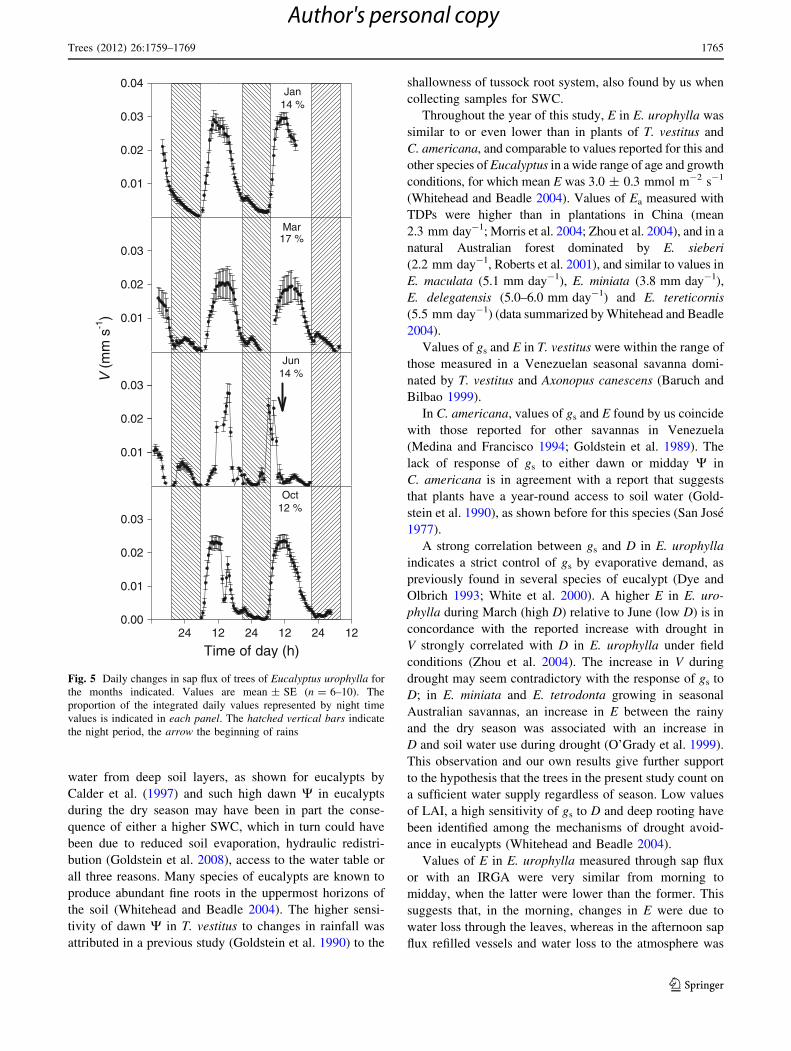
Daily values of V in E. urophylla were always highest at
noon, the highest V occurring in October, followed by
January and March, and the lowest in June; a substantial
nocturnal transpiration was found at all seasons, averaging
14 % of daily transpiration (Fig. 5).
Seasonal changes in leaf gas exchange and related
parameters of E. urophylla are shown in Fig. 6. The highest
values of E measured with IRGA and TDPs were similar
among them, occurred at noon and varied little throughout
the year. Pre-noon values of E measured by both methods
were linearly related (r2 = 0.68, p \ 0.05); in the after-
noon, E measured with IRGA was lower than measured
with TDPs. Maximum E occurred at values of D between
1.5 and 2.3 kPa. Daily courses of E roughly followed daily
courses of PPFD at any season; in June, frequent showers
prevented measurements of PPFD.
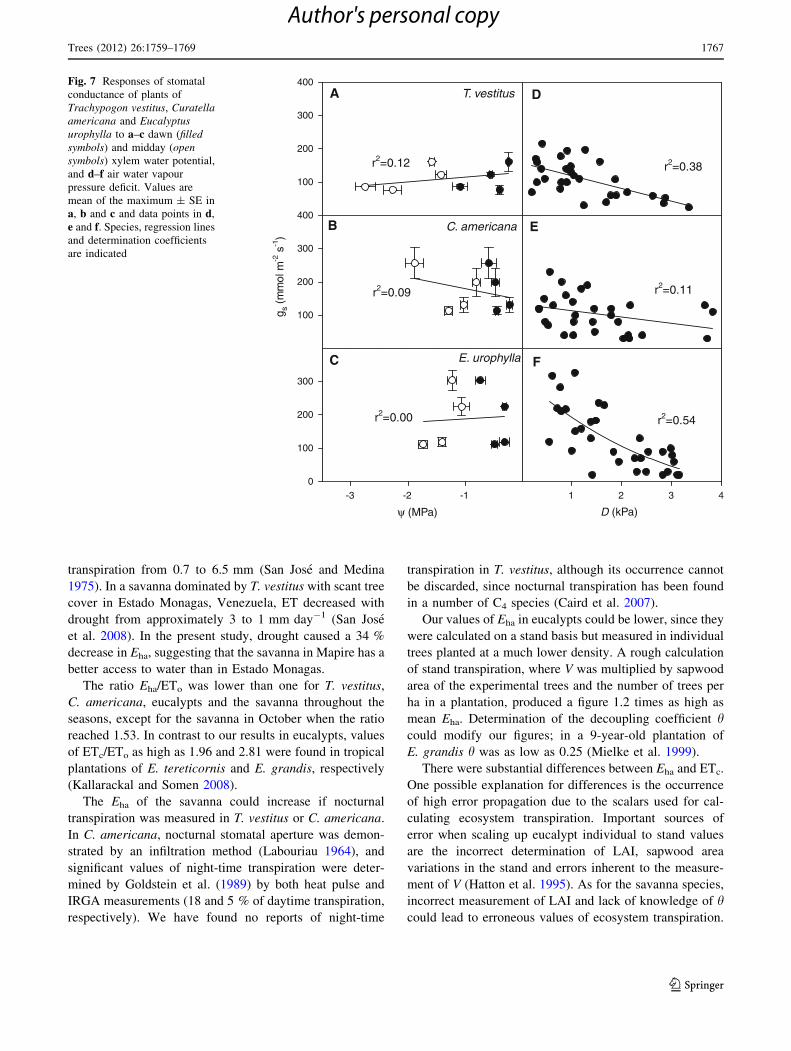
Changes in maximum gs with dawn and midday W and
D are shown in Fig. 7. In all three species, maximum gs
was unresponsive to either dawn or midday W. In both
T. vestitus and E. urophylla, but not in C. americana, there
was a significant decrease in gs with D.
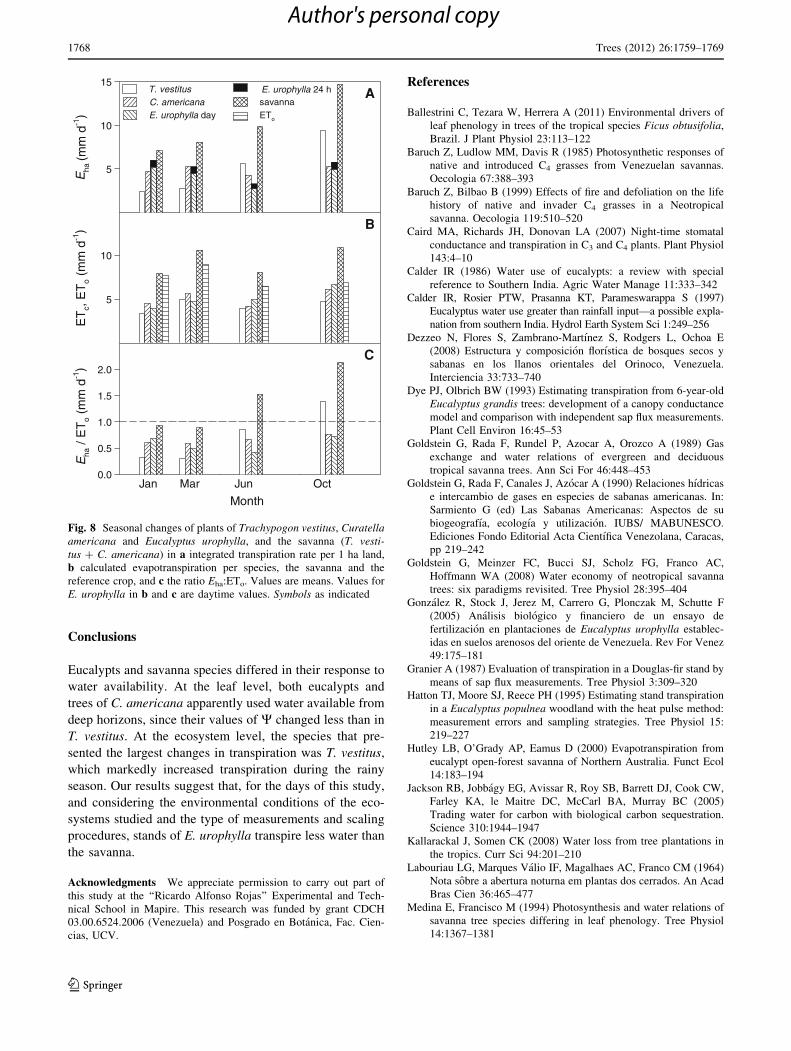
Seasonal changes in Eha, ETc and ETo are shown in
Fig. 8. The Eha in E. urophylla was always higher than in
C. americana except for October; higher than in T. vestitus
in January, and lower than the sum of Eha of T. vestitus and
C. americana, assumed to represent the Eha of the entire
savanna, throughout the seasons. Transpiration of the
savanna was on an average 2.4 times that of eucalypts
during the daytime; when nocturnal eucalypt transpiration
was included, the value was 1.9. The ETc of eucalypts was
higher than in C. americana throughout the seasons, lower
than in T. vestitus in January and March, and lower than the
savanna all year round. The ETo varied little throughout the
seasons, the highest value occurring in March. The ratio
Eha/ETo for the savanna was on average nearly one during
the dry season and nearly two during the rainy season; the
corresponding value for E. urophylla was 0.6 for both
seasons. The ratio Eha/ETc was on average 0.8 for the
species and the savanna. The cumulative Eha for the days of
measurements was higher in the savanna than in the eu-
calypts during the daytime (39.8 and 17.3 mm, respec-
tively), as was the cumulative ETc (37.5 vs. 20.3 mm).
A-2.5
-2.0
-1.5
-1.0
-0.5
0.0
Month
Dec Feb Apr Jun Aug Oct
ψ (M
Pa
)
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
B
Fig. 3 Seasonal changes in a dawn and b midday xylem water
potential of plants of Trachypogon vestitus (circles), Curatellaamericana (squares) and Eucalyptus urophylla (triangles). Values
are mean ± SE (n = 5)
Trees (2012) 26:1759–1769 1763
123
Author's personal copy
Cumulative transpiration in eucalypts, including nocturnal
values, was Eha, 22.0 mm and ETc, 28.4 mm.
These results are consistent with the observation that in
T. vestitus LAI and E increased with rains 2.3 and 1.3
times, respectively, partly explaining the increase of 2.9
times in Eha. As LAI was measured in both C. americana
and eucalypts only at the end of the rainy season, our
calculations were made with possibly the maximum LAI
for these species and still plants of T. vestitus seemed to
govern changes in ecosystem ET because of the marked
increase in their transpiratory surface.
Discussion
We have found that transpiration in E. urophylla was less
affected by drought than in T. vestitus, in which
E increased with rains; in C. americana, less seasonal
change in E was found, as hypothesized. The response of
E to rainfall was reflected in the seasonal changes of
midday W, these being significant in T. vestitus and
E. urophylla, but not in C. americana. Both T. vestitus and
E. urophylla showed lower Eha at the peak of the rainy
season (June) than at the end of it (October), in spite of
increased gs, probably due to a marked decrease in D.
Our values of dawn W were measured at sunrise, when
daily evaporative demand was beginning to increase;
therefore, they may not accurately reflect the maximum Wattained by plants. In the case of eucalypts, the occurrence
of nocturnal transpiration may have also reduced dawn W.
Dawn W decreased with drought by 0.69 MPa in T. vesti-
tus, while it remained comparatively high all year round in
C. americana, as reported by Goldstein et al. (1990) and
Medina and Francisco (1994). In E. urophylla, dawn W was
also high and changed little throughout the seasons, values
being similar to pre-dawn W in E. camaldulensis and high
as compared to values of pre-dawn W of -3 MPa in
E. leucoxylon and -4 MPa in E. platypus (White et al.
2000). The seasonal decrease in dawn W of E. urophylla
was only 0.37 MPa, which is quite small relative to sea-
sonal differences in pre-dawn W of 2.8 MPa in E. leu-
coxylon and 2.0 MPa in E. platypus (White et al. 2000).
The high dawn W found by us during the dry season
suggests that trees of C. americana and eucalypts draw
Ag s
(mm
ol m
-2 s
-1)
50
100
150
200T. vestitus
B
E(m
mol
m-2
s-1
)
2
4
6
C. americana
Hora del día
January March June October
C
D(k
Pa)
1
2
3
4
D
07 10 13 16
PP
FD
(μm
ol m
-2 s
-1)
0
500
1000
1500
2000
Time (h)
07 10 13 16 07 10 13 16 07 10 13 16
Fig. 4 Seasonal changes in daily courses of a stomatal conductance,
b transpiration rate, c air water vapour pressure deficit, and
d photosynthetic photon flux density for plants of Trachypogon
vestitus (closed symbols) and Curatella americana (open symbols).
Month indicated. Values are mean ± SE (n = 6)
1764 Trees (2012) 26:1759–1769
123
Author's personal copy
water from deep soil layers, as shown for eucalypts by
Calder et al. (1997) and such high dawn W in eucalypts
during the dry season may have been in part the conse-
quence of either a higher SWC, which in turn could have
been due to reduced soil evaporation, hydraulic redistri-
bution (Goldstein et al. 2008), access to the water table or
all three reasons. Many species of eucalypts are known to
produce abundant fine roots in the uppermost horizons of
the soil (Whitehead and Beadle 2004). The higher sensi-
tivity of dawn W in T. vestitus to changes in rainfall was
attributed in a previous study (Goldstein et al. 1990) to the
shallowness of tussock root system, also found by us when
collecting samples for SWC.
Throughout the year of this study, E in E. urophylla was
similar to or even lower than in plants of T. vestitus and
C. americana, and comparable to values reported for this and
other species of Eucalyptus in a wide range of age and growth
conditions, for which mean E was 3.0 ± 0.3 mmol m-2 s-1
(Whitehead and Beadle 2004). Values of Ea measured with
TDPs were higher than in plantations in China (mean
2.3 mm day-1; Morris et al. 2004; Zhou et al. 2004), and in a
natural Australian forest dominated by E. sieberi
(2.2 mm day-1, Roberts et al. 2001), and similar to values in
E. maculata (5.1 mm day-1), E. miniata (3.8 mm day-1),
E. delegatensis (5.0–6.0 mm day-1) and E. tereticornis
(5.5 mm day-1) (data summarized by Whitehead and Beadle
2004).
Values of gs and E in T. vestitus were within the range of
those measured in a Venezuelan seasonal savanna domi-
nated by T. vestitus and Axonopus canescens (Baruch and
Bilbao 1999).
In C. americana, values of gs and E found by us coincide
with those reported for other savannas in Venezuela
(Medina and Francisco 1994; Goldstein et al. 1989). The
lack of response of gs to either dawn or midday W in
C. americana is in agreement with a report that suggests
that plants have a year-round access to soil water (Gold-
stein et al. 1990), as shown before for this species (San Jose
1977).
A strong correlation between gs and D in E. urophylla
indicates a strict control of gs by evaporative demand, as
previously found in several species of eucalypt (Dye and
Olbrich 1993; White et al. 2000). A higher E in E. uro-
phylla during March (high D) relative to June (low D) is in
concordance with the reported increase with drought in
V strongly correlated with D in E. urophylla under field
conditions (Zhou et al. 2004). The increase in V during
drought may seem contradictory with the response of gs to
D; in E. miniata and E. tetrodonta growing in seasonal
Australian savannas, an increase in E between the rainy
and the dry season was associated with an increase in
D and soil water use during drought (O’Grady et al. 1999).
This observation and our own results give further support
to the hypothesis that the trees in the present study count on
a sufficient water supply regardless of season. Low values
of LAI, a high sensitivity of gs to D and deep rooting have
been identified among the mechanisms of drought avoid-
ance in eucalypts (Whitehead and Beadle 2004).
Values of E in E. urophylla measured through sap flux
or with an IRGA were very similar from morning to
midday, when the latter were lower than the former. This
suggests that, in the morning, changes in E were due to
water loss through the leaves, whereas in the afternoon sap
flux refilled vessels and water loss to the atmosphere was
Mar17 %
0.01
0.02
0.03
Oct12 %
Time of day (h)24 12 24 12 24 12
0.00
0.01
0.02
0.03
Jun14 %V
(m
m s
-1)
0.01
0.02
0.03
Jan14 %
0.01
0.02
0.03
0.04
Fig. 5 Daily changes in sap flux of trees of Eucalyptus urophylla for
the months indicated. Values are mean ± SE (n = 6–10). The
proportion of the integrated daily values represented by night time
values is indicated in each panel. The hatched vertical bars indicate
the night period, the arrow the beginning of rains
Trees (2012) 26:1759–1769 1765
123
Author's personal copy
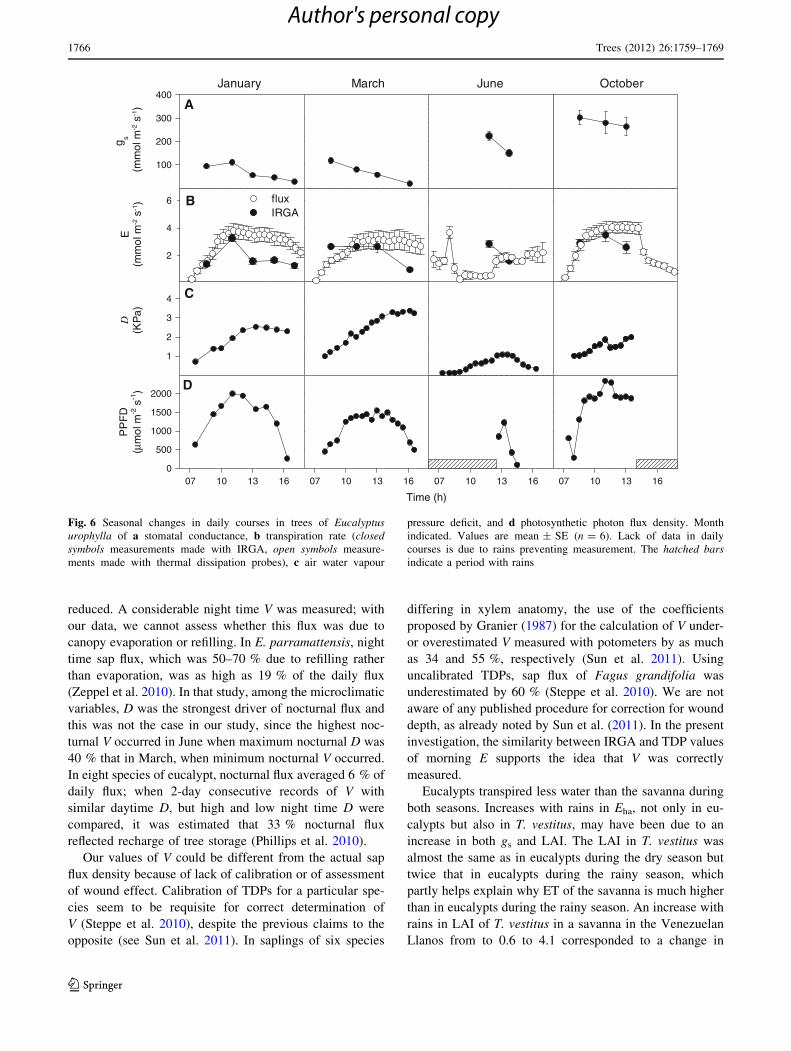
reduced. A considerable night time V was measured; with
our data, we cannot assess whether this flux was due to
canopy evaporation or refilling. In E. parramattensis, night
time sap flux, which was 50–70 % due to refilling rather
than evaporation, was as high as 19 % of the daily flux
(Zeppel et al. 2010). In that study, among the microclimatic
variables, D was the strongest driver of nocturnal flux and
this was not the case in our study, since the highest noc-
turnal V occurred in June when maximum nocturnal D was
40 % that in March, when minimum nocturnal V occurred.
In eight species of eucalypt, nocturnal flux averaged 6 % of
daily flux; when 2-day consecutive records of V with
similar daytime D, but high and low night time D were
compared, it was estimated that 33 % nocturnal flux
reflected recharge of tree storage (Phillips et al. 2010).
Our values of V could be different from the actual sap
flux density because of lack of calibration or of assessment
of wound effect. Calibration of TDPs for a particular spe-
cies seem to be requisite for correct determination of
V (Steppe et al. 2010), despite the previous claims to the
opposite (see Sun et al. 2011). In saplings of six species
differing in xylem anatomy, the use of the coefficients
proposed by Granier (1987) for the calculation of V under-
or overestimated V measured with potometers by as much
as 34 and 55 %, respectively (Sun et al. 2011). Using
uncalibrated TDPs, sap flux of Fagus grandifolia was
underestimated by 60 % (Steppe et al. 2010). We are not
aware of any published procedure for correction for wound
depth, as already noted by Sun et al. (2011). In the present
investigation, the similarity between IRGA and TDP values
of morning E supports the idea that V was correctly
measured.
Eucalypts transpired less water than the savanna during
both seasons. Increases with rains in Eha, not only in eu-
calypts but also in T. vestitus, may have been due to an
increase in both gs and LAI. The LAI in T. vestitus was
almost the same as in eucalypts during the dry season but
twice that in eucalypts during the rainy season, which
partly helps explain why ET of the savanna is much higher
than in eucalypts during the rainy season. An increase with
rains in LAI of T. vestitus in a savanna in the Venezuelan
Llanos from to 0.6 to 4.1 corresponded to a change in
C
D(K
Pa)
1
2
3
4
B
2
4
6 fluxIRGA
D
PP
FD
(μm
ol m
-2 s
-1)
0
500
1000
1500
2000
Ag s
(mm
ol m
-2 s
-1)
100
200
300
400January
March June
October
Time (h)
E(m
mol
m-2 s
-1)
07 10 13 1607 10 13 16 07 10 13 16 07 10 13 16
March June
Fig. 6 Seasonal changes in daily courses in trees of Eucalyptusurophylla of a stomatal conductance, b transpiration rate (closedsymbols measurements made with IRGA, open symbols measure-
ments made with thermal dissipation probes), c air water vapour
pressure deficit, and d photosynthetic photon flux density. Month
indicated. Values are mean ± SE (n = 6). Lack of data in daily
courses is due to rains preventing measurement. The hatched barsindicate a period with rains
1766 Trees (2012) 26:1759–1769
123
Author's personal copy
transpiration from 0.7 to 6.5 mm (San Jose and Medina
1975). In a savanna dominated by T. vestitus with scant tree
cover in Estado Monagas, Venezuela, ET decreased with
drought from approximately 3 to 1 mm day-1 (San Jose
et al. 2008). In the present study, drought caused a 34 %
decrease in Eha, suggesting that the savanna in Mapire has a
better access to water than in Estado Monagas.
The ratio Eha/ETo was lower than one for T. vestitus,
C. americana, eucalypts and the savanna throughout the
seasons, except for the savanna in October when the ratio
reached 1.53. In contrast to our results in eucalypts, values
of ETc/ETo as high as 1.96 and 2.81 were found in tropical
plantations of E. tereticornis and E. grandis, respectively
(Kallarackal and Somen 2008).
The Eha of the savanna could increase if nocturnal
transpiration was measured in T. vestitus or C. americana.
In C. americana, nocturnal stomatal aperture was demon-
strated by an infiltration method (Labouriau 1964), and
significant values of night-time transpiration were deter-
mined by Goldstein et al. (1989) by both heat pulse and
IRGA measurements (18 and 5 % of daytime transpiration,
respectively). We have found no reports of night-time
transpiration in T. vestitus, although its occurrence cannot
be discarded, since nocturnal transpiration has been found
in a number of C4 species (Caird et al. 2007).
Our values of Eha in eucalypts could be lower, since they
were calculated on a stand basis but measured in individual
trees planted at a much lower density. A rough calculation
of stand transpiration, where V was multiplied by sapwood
area of the experimental trees and the number of trees per
ha in a plantation, produced a figure 1.2 times as high as
mean Eha. Determination of the decoupling coefficient hcould modify our figures; in a 9-year-old plantation of
E. grandis h was as low as 0.25 (Mielke et al. 1999).
There were substantial differences between Eha and ETc.
One possible explanation for differences is the occurrence
of high error propagation due to the scalars used for cal-
culating ecosystem transpiration. Important sources of
error when scaling up eucalypt individual to stand values
are the incorrect determination of LAI, sapwood area
variations in the stand and errors inherent to the measure-
ment of V (Hatton et al. 1995). As for the savanna species,
incorrect measurement of LAI and lack of knowledge of hcould lead to erroneous values of ecosystem transpiration.
E. urophylla
ψ (MPa)
0
100
200
300
T. vestitus
100
200
300
400
C. americana
g s (
mm
ol m
-2 s
-1)
100
200
300
400
r2=0.12
r2=0.09
r2=0.00
r2=0.38
r2=0.11
D (kPa)
-3 -2 -1 1 2 3 4
r2=0.54
A
B
C
D
E
F
Fig. 7 Responses of stomatal
conductance of plants of
Trachypogon vestitus, Curatellaamericana and Eucalyptusurophylla to a–c dawn (filledsymbols) and midday (opensymbols) xylem water potential,
and d–f air water vapour
pressure deficit. Values are
mean of the maximum ± SE in
a, b and c and data points in d,
e and f. Species, regression lines
and determination coefficients
are indicated
Trees (2012) 26:1759–1769 1767
123
Author's personal copy
Conclusions
Eucalypts and savanna species differed in their response to
water availability. At the leaf level, both eucalypts and
trees of C. americana apparently used water available from
deep horizons, since their values of W changed less than in
T. vestitus. At the ecosystem level, the species that pre-
sented the largest changes in transpiration was T. vestitus,
which markedly increased transpiration during the rainy
season. Our results suggest that, for the days of this study,
and considering the environmental conditions of the eco-
systems studied and the type of measurements and scaling
procedures, stands of E. urophylla transpire less water than
the savanna.
Acknowledgments We appreciate permission to carry out part of
this study at the ‘‘Ricardo Alfonso Rojas’’ Experimental and Tech-
nical School in Mapire. This research was funded by grant CDCH
03.00.6524.2006 (Venezuela) and Posgrado en Botanica, Fac. Cien-
cias, UCV.
References
Ballestrini C, Tezara W, Herrera A (2011) Environmental drivers of
leaf phenology in trees of the tropical species Ficus obtusifolia,
Brazil. J Plant Physiol 23:113–122
Baruch Z, Ludlow MM, Davis R (1985) Photosynthetic responses of
native and introduced C4 grasses from Venezuelan savannas.
Oecologia 67:388–393
Baruch Z, Bilbao B (1999) Effects of fire and defoliation on the life
history of native and invader C4 grasses in a Neotropical
savanna. Oecologia 119:510–520
Caird MA, Richards JH, Donovan LA (2007) Night-time stomatal
conductance and transpiration in C3 and C4 plants. Plant Physiol
143:4–10
Calder IR (1986) Water use of eucalypts: a review with special
reference to Southern India. Agric Water Manage 11:333–342
Calder IR, Rosier PTW, Prasanna KT, Parameswarappa S (1997)
Eucalyptus water use greater than rainfall input—a possible expla-
nation from southern India. Hydrol Earth System Sci 1:249–256
Dezzeo N, Flores S, Zambrano-Martınez S, Rodgers L, Ochoa E
(2008) Estructura y composicion florıstica de bosques secos y
sabanas en los llanos orientales del Orinoco, Venezuela.
Interciencia 33:733–740
Dye PJ, Olbrich BW (1993) Estimating transpiration from 6-year-old
Eucalyptus grandis trees: development of a canopy conductance
model and comparison with independent sap flux measurements.
Plant Cell Environ 16:45–53
Goldstein G, Rada F, Rundel P, Azocar A, Orozco A (1989) Gas
exchange and water relations of evergreen and deciduous
tropical savanna trees. Ann Sci For 46:448–453
Goldstein G, Rada F, Canales J, Azocar A (1990) Relaciones hıdricas
e intercambio de gases en especies de sabanas americanas. In:
Sarmiento G (ed) Las Sabanas Americanas: Aspectos de su
biogeografıa, ecologıa y utilizacion. IUBS/ MABUNESCO.
Ediciones Fondo Editorial Acta Cientıfica Venezolana, Caracas,
pp 219–242
Goldstein G, Meinzer FC, Bucci SJ, Scholz FG, Franco AC,
Hoffmann WA (2008) Water economy of neotropical savanna
trees: six paradigms revisited. Tree Physiol 28:395–404
Gonzalez R, Stock J, Jerez M, Carrero G, Plonczak M, Schutte F
(2005) Analisis biologico y financiero de un ensayo de
fertilizacion en plantaciones de Eucalyptus urophylla establec-
idas en suelos arenosos del oriente de Venezuela. Rev For Venez
49:175–181
Granier A (1987) Evaluation of transpiration in a Douglas-fir stand by
means of sap flux measurements. Tree Physiol 3:309–320
Hatton TJ, Moore SJ, Reece PH (1995) Estimating stand transpiration
in a Eucalyptus populnea woodland with the heat pulse method:
measurement errors and sampling strategies. Tree Physiol 15:
219–227
Hutley LB, O’Grady AP, Eamus D (2000) Evapotranspiration from
eucalypt open-forest savanna of Northern Australia. Funct Ecol
14:183–194
Jackson RB, Jobbagy EG, Avissar R, Roy SB, Barrett DJ, Cook CW,
Farley KA, le Maitre DC, McCarl BA, Murray BC (2005)
Trading water for carbon with biological carbon sequestration.
Science 310:1944–1947
Kallarackal J, Somen CK (2008) Water loss from tree plantations in
the tropics. Curr Sci 94:201–210
Labouriau LG, Marques Valio IF, Magalhaes AC, Franco CM (1964)
Nota sobre a abertura noturna em plantas dos cerrados. An Acad
Bras Cien 36:465–477
Medina E, Francisco M (1994) Photosynthesis and water relations of
savanna tree species differing in leaf phenology. Tree Physiol
14:1367–1381
B
ET
c, E
To
(mm
d-1
)
5
10
C
Month
Eha
/ E
To
(mm
d-1
)
0.0
0.5
1.0
1.5
2.0
AE
ha(m
m d
-1)
5
10
15
Jan Mar Jun Oct
T. vestitusC. americanaE. urophylla day
savannaETo
E. urophylla 24 h
Fig. 8 Seasonal changes of plants of Trachypogon vestitus, Curatellaamericana and Eucalyptus urophylla, and the savanna (T. vesti-tus ? C. americana) in a integrated transpiration rate per 1 ha land,
b calculated evapotranspiration per species, the savanna and the
reference crop, and c the ratio Eha:ETo. Values are means. Values for
E. urophylla in b and c are daytime values. Symbols as indicated
1768 Trees (2012) 26:1759–1769
123
Author's personal copy

Mielke MS, Oliva MA, de Barros NF, Penchel RM, Martinez CA, de
Almeida AC (1999) Stomatal control of transpiration in the
canopy of a clonal Eucalyptus grandis plantation. Trees 13:152–
160
Morris J, Ningnan Z, Zenjiang Y, Collopy J, Daping X (2004) Water
use by fast-growing Eucalyptus urophylla plantations in southern
China. Tree Physiol 24:1035–1044
O’Grady AP, Eamus D, Hutley LB (1999) Transpiration increases
during the dry season: patterns of tree water use in eucalypt
open-forests of northern Australia. Tree Physiol 19:591–597
Phillips NG, Lewis JD, Logan BA, Tissue DT (2010) Inter- and intra-
specific variation in nocturnal water transport in Eucalyptus.
Tree Physiol 30:586–596
Roberts S, Vertessy R, Grayson R (2001) Transpiration from
Eucalyptus sieberi (L. Johnson) forests of different age. For
Ecol Manage 143 153:161
San Jose JJ, Medina E (1975) Effects of fire on organic matter
production and water balance on a tropical savanna. In: Golley
FB, Medina E (eds) Tropical ecological systems. Springer,
Berlin, pp 251–264
San Jose JJ (1977) Potencial hıdrico e intercambio gaseoso de
Curatella americana L. en la temporada seca de la sabana de
Trachypogon. Acta Cient Venez 28:373–379
San Jose JJ, Montes R, Grace J, Nikonova N, Osıo A (2008) Land-use
changes alter radiative energy and water vapor fluxes of a tall-
grass Andropogon field and a savanna–woodland continuum in
the Orinoco lowlands. Tree Physiol 28:425–435
Steppe K, De Pauw DJW, Doody TM, Teskey RO (2010) A
comparison of sap flux density using thermal dissipation, heat
pulse velocity and heat field deformation methods. Agric For
Meteorol 150:861–1158
Sun H, Aubrey DP, Teskey RO (2011) A simple calibration improved
the accuracy of the thermal dissipation technique for sap flow
measurements in juvenile trees of six species. Trees 26:631–640
White DA, Turner NC, Galbraith JH (2000) Leaf water relations and
stomatal behaviour of four allopatric Eucalyptus species planted in
Mediterranean southwestern Australia. Tree Physiol 20:1157–1165
Whitehead W, Beadle CL (2004) Physiological regulation of
productivity and water use in Eucalyptus: a review. For Ecol
Manage 193:113–140
Witschoreck R, Schumacher MV, Marcos Vinicius Winckler Caldeira
MV (2003) Estimating of biomass and length of fine roots in
Eucalyptus urophylla S.T. Blake in the county of Santa Maria,
RS. Rev Arvore 27:177–183
Wullschleger S, Meinzer F, Vertessy R (1998) A review of whole-
plant water use studies in trees. Tree Physiol 18:99–512
Zeppel M, Tissue D, Taylor D, Macinnis-Ng C, Eamus D (2010)
Rates of nocturnal transpiration in two evergreen temperate
woodland species with differing water-use strategies. Tree
Physiol 30:988–1000
Zhou G, Yin G, Morris J, Bai J, Chen S, Chu G, Zhang N (2004)
Measured sap flux and estimated evapotranspiration of tropical
Eucalyptus urophylla plantations in South China. Acta Bot Sin
46:202–210
Trees (2012) 26:1759–1769 1769
123
Author's personal copy