distribution of choline acetyl transferase mrna in … · tdepartment of neurobiology, ucla school...
TRANSCRIPT

Journal of Vestibular Research, Vol. 6, No. 3, pp. 203-212, 1996 Copyright © 1996 Elsevier Science Inc. Printed in the USA. All rights reserved
0957-4271196 $15 .00 + .00
ELSEVIER 0957-4271 (95)02041-1
Original Contribution
DISTRIBUTION OF CHOLINE ACETYL TRANSFERASE mRNA IN THE EFFERENT VESTIBULAR NEURONS OF THE CHINCHILLA
D. Bridgeman,* L. Hoffman,* P. A. Wackym, t P. E Micevych, :j: and P. Popper+
*Goodhill Ear Center, Division of Head and Neck Surgery, and tDepartment of Neurobiology, UCLA School of Medicine, Los Angeles, California, USA;
tDepartment of Otolaryngology, Mount Sinai School of Medicine, New York, New York, USA Reprint address: Dr. P. Popper, Division of Head and Neck Surgery, Box 951624,
UCLA School of Medicine, Los Angeles, CA 90095-1624 USA.
0 Abstract- The distribution of choline acetyltransferase messenger RNA (mRNA) among efferent vestibular neurons in the chinchilla was investigated. mRNA coding for choline acetyltransferase, the enzyme that synthesizes acetylcholine, was used as a marker for the cholinergic system. In order to retrogradely label the efferent vestibular neurons, Fluoro-gold was injected through the oval window into the inner ear of anesthetized young male chinchillas (6 to 12 months old). The animals were anesthetized and perfused through the heart 2 days post injection with 40Jo paraformaldehyde in phosphate buffer. Retrogradely labeled efferent vestibular neurons were mapped in brainstem sections prior to processing for in situ hybridization histochemistry using radiolabeled ribonucleic acid probes complementary to the 3' end of the choline acetyltransferase mRNA. At the levels of the ascending facial nerve and the genu of the facial nerve, we found that approximately 900Jo of the Fiuoro-gold labeled cells in group E] contained choline acetyhransferase mRNA. AH of the group E2 cells that were labeled with Fluoro-gold were found to be cholinergic (contain choline acetyltransferase mRNA). Finally, 600Jo of the Flu oro-gold-labeled cells in the caudal pontine reticular nucleus contained choline acetyltransferase mRNA.
0 Keywords- chinchilla; choline acetyltransferase; efferent vestibular neurons; Fluoro-gold; gene expression.
Introduction
Classical degeneration experiments, which demonstrated centrally originating fibers that terminate on peripheral vestibular end-organs, provided the first evidence that the central nervous system may be modulating input from the vestibular periphery (1 ,2). However, the mechanisms involved in the control of vestibular input by efferent vestibular neurons (EVN) have yet to be fully elucidated.
The location of efferent somata within the brainstems of the cat, gerbil, rat, and chinchilla (3-6) have been demonstrated with histological techniques. EVN are located in three primary groups within the brainstem medulla: Group E 1 and group E2, located lateral and medial, respectively, to the genu of the facial nerve, and a third group located medial to
caucia1 pont_mt reucula.t r1uc~eus ures , 1A2 ,
The neurochemical organization of EVN also have been investigated. Warr (3) utilized acetylcholinesterase (AChE) histochemistry to demonstrate that the efferent vestibular system of the cat is cholinergic. Recent studies have combined retrograde labeling, to identify the location of EVN, with choline acetyl trans-
RECEIVED 14 June 1994; AccEPTED 19 October 1995.
203
A .~/------.....'-, :r~c::
~. DC /. ) E.C ·u. ·. D Ve MVe I I '--- ~fH (
'"~' 1/Fi\ Sui /CPR
\d' "-.// • 8/ft mlf
spo C:') ·sc7 pd
-\ ~--- " Gi ,, / J ~ ... ( 7 . RMg
"". \_ G'A
~______;. ml RPa
~ PY
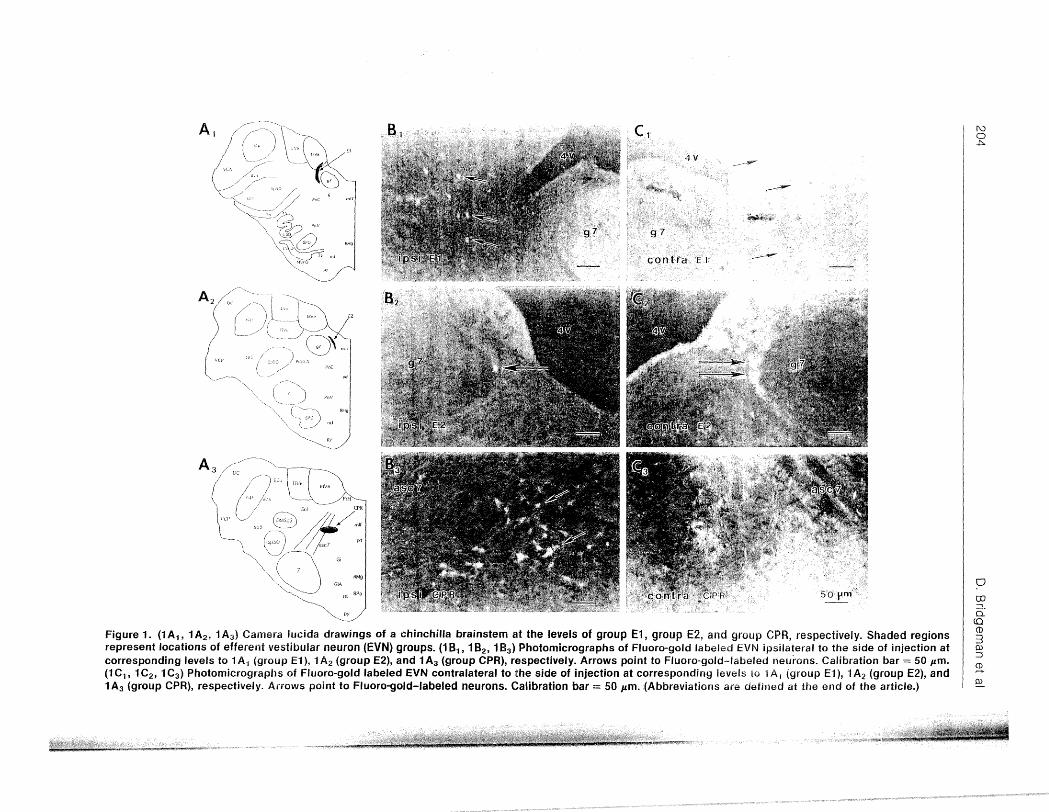
Figure 1. (1 A 1 , 1 A2 , 1 A3 ) Camera Iucida drawings of a chinchilla brain stem at the levels of group E1, group E2, and group CPR, respectively. Shaded regions represent locations of efferent vestibular neuron (EVN) groups. (1 8 1 , 182 , 1 8 3 ) Photomicrographs of Fluoro-gold labeled EVN ipsilateral to the side of injection at corresponding levels to 1 A 1 (group E1 ), 1 A2 (group E2), and 1 A3 (group CPR), respectively. Arrows point to Fluoro-gold--labeled neurons. Calibration bar = 50 pm. (1 C1 , 1 C2 , 1 C3 ) Photomicrographs of Fluoro-gold labeled EVN contralateral to the side of injection at corresponding levels to 1 A 1 (group E1 ), 1 A2 (group E2), and 1 A3 (group CPR), respectively. Arrows point to Fluoro-gold-labeled neurons. Calibration bar = 50 pm . . (Abbreviations are defined at the end of the article.)
(\..) 0 ~
0
co -. 0.: (Q CD 3 OJ ::J
~ OJ

ChAT mRNA in Vestibular Efferent Neurons
ferase (ChAT) immunohistochemistry to determine the distribution of cholinergic neurons in the efferent vestibular system (4,5). ChAT catalyzes the reaction that synthesizes acetylcholine (ACh) from choline and acetyl CoA in the nerve terminal, and it is a marker for cholinergic neurons. In rats, Schwarz and colleagues (5) demonstrated that 570Jo to 61 OJo of retrogradely labeled neurons in groups E 1 and E2 contain ChAT immunoreactivity, but none of the labeled cells in the CPR exhibited ChAT immunoreactivity. In gerbils, Perachio and Kevetter ( 4) showed that all of the efferent vestibular group El neurons were ChAT immunoreactive, however, these authors did not observe any labeled cells medial to the genu of the facial nerve. In addition, no retrogradely labeled neurons were observed in the CPR. This variability suggests that while a population of gerbil EVN are cholinergic, there may be a significant species difference where neurochemistry is concerned.
The chinchilla has frequently been used as a model to understand the anatomy and the physiology of the vestibular system. The bilateral and rostrocaudal extent of all efferent vestibular groups of the chinchilla has been published (6). Knowledge of the neurochemical anatomy of the chinchilla EVN will provide information with respect to the variety of neurotransmitters or neuromodulators among these cells, ultimately leading to a more comprehensive understanding of the function of the vestibular system. While it is generally accepted that in mammalian species certain EVN are cholinergic, it is not known whether all EVN in the chinchilla are cholinergic. In the present study in sjtu istry combined with retrograde tract tracing was used to study the distribution of ChAT mRNA among the cells of the EVN of the chinchilla. Portions of this work have been presented in abstract form (7).
Materials and Methods
Surgery
Chinchillas were prepared for perilymphatic administration of Fluoro-gold (FG;
205
Fluorochrome, Inc.) in the following manner. General anesthesia was achieved via intramuscular injection of ketamine (12 mg/kg) and xylazine (2.4 mg/kg). Atropine (0.2 cc) was delivered intramuscularly, followed by the subcutaneous administration of the antibiotic trimethoprim sulfa ( 40 mg/mL trimethoprim fraction) at a dosage of 5 mg/kg. Following surgical exposure of the middle ear, the head was positioned within a restraint device so that the round and oval windows were in clear view. Using an operating microscope, the stapes was gently removed while maintaining the integrity of the oval window, through which an FG-filled micropipette (50 p.m tip diameter) was inserted. Approximately 40 ~tL of the tracer was administered via pneumatic injection into the scala vestibuli. In a separate study, 3 chinchillas had FG injected on the stapedius muscle only. Upon completion of the injection, the oval window was sealed from the middle ear using a free fat graft, followed by a piece of cellulose foam. Animals were given trimethoprim sulfa antibiotic (0.1 cc) and fluids (sterile saline, 25 cc/kg) each day by subcutaneous administration during the 2 postoperative days, after which they were anesthetized and perfused intracardially with saiine followed by 500 mL of fixative (40Jo paraformaldehyde in 0.12 M phosphate buffer, pH 7 .6). The brainstems were dissected, freed of meningeal membranes, and cryoprotected overnight in a 250Jo sucrose/phosphatebuffered saline solution. Brainstems were frozen in dry ice, and serial sections (20 p.m) were collected using a cryostat microtome for
i.,lOh hist.ocnemistry processing.
lwapping of Fluoro-gold Cells
Initial trials of in situ hybridization histochemistry on FG-containing brainstem sections revealed that the FG signal was quenched from the tissue during processing. Therefore, it was necessary to map labeled cells in the brainstem prior to performing in situ hybridization histochemistry. Sections were collected onto Superfront Plus@ (Fisher, Tustin, CA) microscope

206
slides and dehydrated. FG-labeled cells were visualized using a wide-band ultraviolet (UV) excitation filter. Camera Iucida was used to map the position of the FG-labeled cells relative to brainstem landmarks under a 20x objective lens. After mapping, the slides were stored in desiccated boxes at -70°C until processed for in situ hybridization histochemistry.
A plasmid (pSPT 18, Pharmacia) containing a 2329 bp eDNA clone encoding rat ChAT was a gift from Dr. S. Berrard (Centre National de I a Recherche Scientifique, Gif sur Yvette, France). The plasmid was cut at an Hha1 restriction site within the eDNA insert, and a single-stranded 790-base long RNA probe, complementary to the 3'-end of the ChAT eDNA, was transcribed from the linearized template in a 20 ,uL reaction mixture containing 5 ,umol/L (100 pmol) 35S-UTP (1320 Ci/mmol, NEN), 0.7 ~L RNasin (40 U/,uL), 10 mmol/L DTT (d/-Dithiotheritol), 500 ,umol!L each of ATP, CTP, and GTP, 15 ,umol!L UTP, 4 ,uL of 5 x transcription buffer, 1 ,ug template eDNA, and 20 units of T7 RNA polymerase (all from Promega). The transcription was carried out for 2 h at 40°C and terminated by quenching on ice. The DNA template was digested with DNase RQ 1 for 10 min at 37°C. The transcript was labeled to a specific activity of 4.1 x 105 cpm/ng. The transcript was extracted with phenol/chloroform, precipitated with ethanol, reconstituted in 50 ,uL of 20 mM DTT, and stored at -20°C until used.
The in situ hybridization histochemistry protocol has been described previously (8-10), with the difference that the final formamide concentration was 400Jo and the hybridization was carried out at 60°C. Subsequently, tissue was washed in decreasing concentrations of sse (standard sodium citrate) containing 1 mmol/L DTT, with the final rinse done in o.5 x sse (1 x sse= o.1s M NaCl/0.04 M Na citrate, pH 7 .2) at room temperature for 10 min.
D. Bridgeman et al
Autoradiography
After washing, the slides were dipped in Kodak NTB-2 nuclear emulsion diluted 1:1 with distilled water and exposed at 4 oc for 5 to 7 d. After exposure, the slides were developed in Kodak D-19 developer for 4 min at l6°C, rinsed for 1 min in 0.1 mol/L PBS, and fixed for 5 min with Kodak Rapid Fix. The sections were then dehydrated and coverslipped with DPX mountant (Electron Microscopy Sciences, Ft. Washington, PA).
Analysis
Labeled cells were visualized with an Olympus Vanox-T microscope using dark field illumination and bright field illumination using Nomarski optics. Accumulation of reduced silver grains over cell bodies indicated ChAT cRNA hybridization to ChAT mRNA. The image of ChAT mRNA labeled cells was superimposed using camera-lucida onto the previously sketched FG maps and aligned with preexisting landmarks to reveal cells that are posirive for both FG and ChAT mRNA. A FG-labeled cell was considered to express ChAT mRNA if there was a cell-sized area of high density of silver grains, clearly different from the surrounding background. The experience in our laboratory is that this corresponds to 3 times the background level of silver grains (8).
Results
Anatomical Distribution of Efferent Vestibular 1Veurons
The injection of Fluoro-gold into the perilymph of 3 chinchillas resulted in the labeling of somata in the brainstem. Retrogradely labeled EVNs were concentrated approximately at the level of the genu of the seventh cranial nerve and at the level of the CPR. These labeled cells were located in regions similar to those described in a previous study using chinchillas, in which the anatomy of EVN was
ChAT mRNA in Vestibular Efferent Neurons
identified via administration of horseradish peroxidase (HRP) (6). As before, EVN were found distributed into 3 topologically distinct groups, identified as group El, group E2, and group CPR (Figure 1).
Group EJ. Group El consists of small ovoidshaped neurons with the long axis in the transverse plane oriented in the dorsal-ventral direction (Figures , 1 B 1 , 1 C 1 , and 2C). This group begins caudal to the rostral pole of the lateral vestibular nucleus and extends to the caudal pole of the genu of the facial nerve. The most ventral and rostral part of group El appears caudal to the rostral part of the medial vestibular nucleus (MVN) and caudal to the rostral part of the L VN. From rostral to caudal, group El appears lateral to
OS<: 7
207
the facial nerve genu and medial to the L VN. The nucleus extends lateral and ventral to the facial genu, surrounding the ventral parts of the MVN. Group El efferents were labeled both ipsilateral and contralateral to the injection site, with a larger number of neurons contralaterally than ipsilaterally. Group El contained the greatest number of vestibular efferents (Figure 1) (Table
E2 ~his is 2; group of cells sin1ilar in size and orientation to the cells of group El, but located between the medial part of the facial genu and the fourth ventricle (Figures 1A2 ,
1B2 , and 1C2 ). The rostrocaudal position of group E2 somata is caudal to the most rostral origin of the LVN. The rostral end of group E2 is more caudal than that of group E 1. Cells
4v
g7
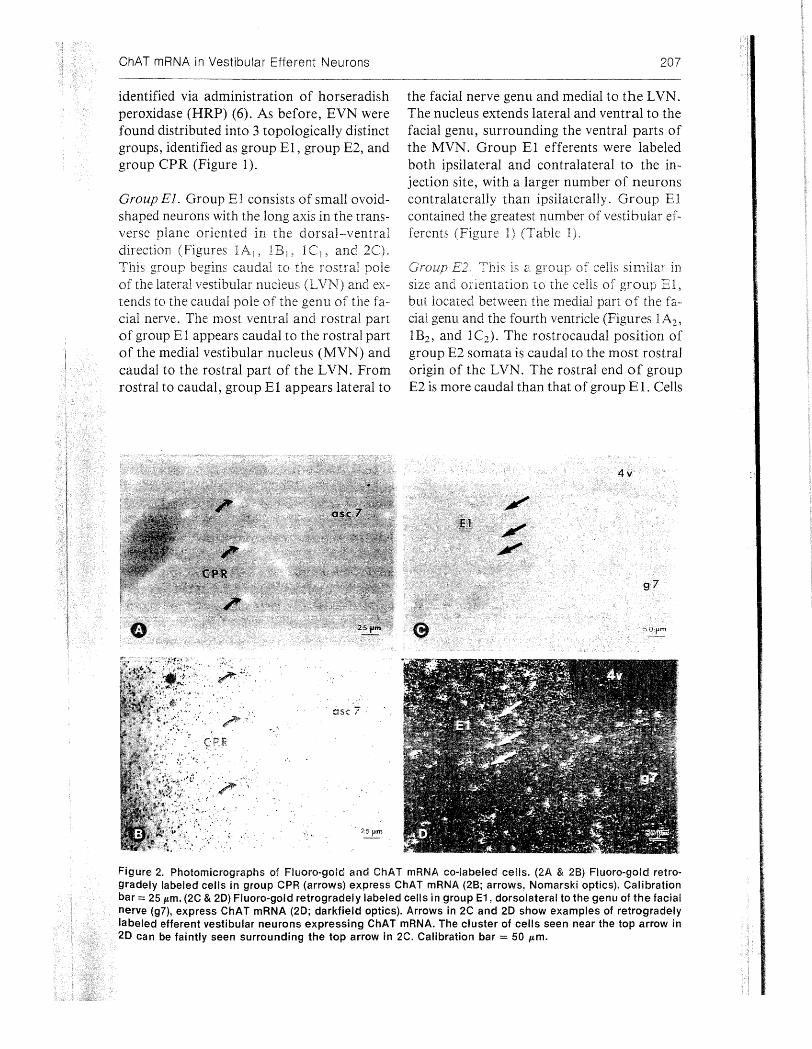
Figure 2. Photomicrographs of Fluoro-gold and ChAT mRNA co-labeled cells. (2A & 2B) Fluoro-gold retrogradely labeled cells in group CPR (arrows) express ChAT mRNA (2B; arrows, Nomarski optics). Calibration bar= 25 Jtffi. (2C & 20) Fluoro-gold retrogradely labeled cells in group E1, dorsolateral to the genu of the facial nerve (g7), express ChAT mRNA (20; darkfield optics). Arrows in 2C and 20 show examples of retrogradely labeled efferent vestibular neurons expressing ChAT mRNA. The cluster of cells seen near the top arrow in 20 can be faintly seen surrounding the top arrow in 2C. Calibration bar= 50 Jtffi.

208 D. Bridgeman et al
Table 1. Counts of Labeled Efferent Vestibular Somas (mean ± SD) for Each Group and Brainstem Side Following Unilateral Administration of Fluoro-gold*
n=3 Soma count Group E1 Group E2 Group CPR
Ipsilateral Contralateral
127 ± 35 (209) 1 07 ± 64 (224)
49 ± 22 (70) 99 ± 95 (209)
4 ± 3 (7) 8 ± 7 (15)
74 ±56 (132)
*Maximum number found in any one animal shown in parentheses.
of group E2 begin w be seen at the level of the were labeled with while no labeling was caudal pan group E . The orientation of observed in group E or CPR. the cells in this group follows the long axis of the narrow space between the genu and the fourth ventricle, cupping the genu at its most medial part. Again, neurons both ipsilateral and contralateral to the FG injection site were present, and a greater number of neurons projected contralaterally (Figure 1) and (Table 1). Of the 3 groups of cells identified in this study, group E2 contained the fewest vestibular efferents.
Group CPR. This group is formed by cells scattered among the fibers of the ascending limb of the facial nerve root in the caudal pontine reticular formation (Figures 1A3 , 1 B3 ,
1 C3 , and 2A). Rostrally, a very few cells of this group begin to appear at the level of the abducens nucleus, medial to the descending limb of the facial nerve root. The cells of the group CPR are ovoid in shape, are oriented along the fibers of the facial nerve, and are larger than the cells of group E1 or E2 (6). The rostral part was not present in all of the animals. When present, the rostral part contains neurons that project either ipsilaterally or contralaterally, while the caudal subgroup has only ipsilateral projections as described before (6). In addition, label was found in the superior olivary region, commonly associated with the cochlear efferent neurons. Since the retrograde tracer was placed into the vestibule, it was expected that both cochlear and vestibular efferents would be labeled.
Stapedius Injection Experiment. In these animals, the root of CN VII and fibers in the ascending and descending limb of the VII nerve
Localization of Choline Acetyltransferase (ChAT) mRNA in the Efferent Vestibular Neurons
The number and locations of cholinergic and FG-labeled EVN were assessed in 3 animals (Table 2). In the contralateral group El, 88.90Jo (±3.5) of FG-labeled neurons contained ChAT mRNA (Figure 2). In the ipsilateral group E1, 84.00Jo (± 7 .4) of FG-labeled neurons were cholinergic. In both ipsilateral and contralateral group E2, 1000Jo of the FGlaoeled neurons contained ChAT mRNA. In group CPR, 60.50Jo (±3.5) of the ipsilateral FGlabeled cells contained ChAT mRNA (Figure 2).
In the contralateral group E1, the percentage of FG-labeled cells that contained ChAT mRNA remained relatively constant over the entire rostrocaudal extent of this group. The percentage of ipsilateral group E 1 neurons that were FG-labeled and identified as cholinergic gradually increased through the rostral part of the group, but remained uniform
Table 2. Percentage of Fluoro-gold-Labeled Efferent Vestibular Somas (mean ± SO) Containing ChAT mRNA for Each Group and Brainstem Side*
Group E1 Group E2 Group CPR
Ipsilateral 84.0% ± 7.4 100% 60.5% ± 3.5% Contralateral 88.9% ± 3.5 100%
*Unilateral administration of Fluoro-gold. Fluorescent identification of retrograde-labeled efferent vestibular neurons and in situ hybridization histochemistry localization of choline acetyltransferase (ChAT) mRNA were performed in the same sections.

ChAT mRNA in Vestibular Efferent Neurons
through the caudal extent. Clusters of ChAT mRNA labeled cells shown in Figure 2D can be faintly seen in Figure 2C. The higher sensitivity of the in situ hybridization histochemistry and autoradiography shows the labeled somata better than the FG-containing, retrogradely labeled somata (Figure 2C). In the E2, throughout the entire rostrocaudal extent of both ipsilateral and contralateral groups, all of the FG-labeled neurons contained ChAT mRNA (Table 2). The percentage of FGlabeled cells that were cholinergic in group CPR remained relatively constant over the entire extent of this group.
Discussion
The main finding of this paper is that not all of the EVN in the chinchilla are cholinergic. Approximately 900Jo of the contralateral and 840Jo of the ipsilateral retrogradely labeled cells in group El contained ChAT mRNA. In addition, all of the labeled cells in group E2 were cholinergic. These results are at variance with the investigations of other mammalian species. Schwarz and colleagues (5) reported that in the rat approximately 40o/o of labeled cells in both groups El and E2 did not contain AChE. In addition, Perachio and Kevetter (4) observed in the gerbil that all labeled cells in group El were cholinergic, but none of the identified EVN in their ventral group displayed ChAT immunoreactivity or contained AChE. It appears that, in the rat and gerbil, groups El and E2 may consist of a heterogeneous population of cells that might function differently their different neuro-chemical properties. In the v>H .. U....,HHJ
all of the labeled cells in groups E 1 and E2 were cholinergic. In contrast to groups El and E2, approximately 40o/o of the cells labeled with Fluoro-gold did not express ChAT mRNA in group CPR (Table 2). This percentage was relatively consistent throughout the rostrocaudal extent of this group of neurons. Our results of EVN distribution in this group are also different from observations made in other species. Schwarz and colleagues (5) reported that while approximately half of the neurons ret-
209
rogradely labeled in the CPR of the rat were AChE positive, none were ChAT immunoreactive. Perachio and Kevetter (4) observed no labeled cells in the CPR. Again, it appears that the different results obtained in the rat and gerbil may be due to species differences or to the differences in the sensitivity of detecting ChAT mRNA and ChAT molecules. The use of in situ hybridization histochemistry permits a sensitive localization of cells expressing as few as 10 copies of the ChAT mRNA message, while the indirect visualization of ChAT antibody-antigen binding deperlds on issues related to transport out of the soma or other posttranscriptional and posttranslational events. For example, preprocholecystokinin mRNA has been demonstrated in a-motoneurons, while the immunohistochemical demonstration of cholecystokinin in motoneurons is controversial (11). Nonetheless, due to the large number of cells that did not contain ChAT mRNA, one may conclude that in the chinchilla, group CPR may consist of a heterogeneous population of cells.
The extent and location of EVN observed in this study are consistent with previous studies of the chinchilla that utilized retrograde tract tracing to identify efferent neurons (6). In groups El and E2, EVN were distributed bilaterally, with a greater proportion (2: 1) projecting contralateral to the side of injection. Two subgroups of EVN in the CPR were recognized: a rostral and a caudal subgroup. The rostral subgroup was not always labeled. When this subgroup was labeled, it was distributed bilaterally, while the caudal subgroup had only ipsilateral projections. In contrast, Schwarz and colleagues reported that
contra-laterally. However, Schwarz and colleagues (5) report observing only "a few labeled neurons in the caudal pontine reticular (CPR) nucleus" of the rat. Perachio and Kevetter (4) reported that they did not detect any labeled neurons in the CPR nucleus of the gerbil following injection of horseradish peroxidase into the inner ear. These variances may be due to species or technical differences.
Although the FG-labeled neurons could not be traced directly in the vestibular periphery,

210
they were interpreted as a putative source of vestibular efferents for the following reasons: 1) They were located in brainstem regions reported to contain vestibular efferent neurons in this and other mammalian species (4-6); 2) They were labeled consistently by injections of FG or HRP (6) into the 3) The locations of these neurons were different from those labeied in other mammalian species af-ter of label into the middle ear 4) the location of ;:he chinchilla EVl'J
monkey. In the latter bers from the squirrel monkey brainstem EVN nuclei were traced peripherally to the vestibular receptors, providing evidence that these efferent neurons were indeed efferents to the vestibular end-organs (12). Our study revealed similar patterns of labeled somata in comparison to the above-mentioned study.
Activation of efferent neurons has been shown to result in changes in discharge properties of vestibular primary afferent neurons, reflecting both excitatory and inhibitory influences (12-14). Though such influences have yet to be directly demonstrated for the chinchilla, they have been shown in experiments using bullfrogs (14), pigeons (13), and squirrel monkeys (12) and are therefore likely to be present in lower mammals as well. The results of the present study suggest that efferents originating in groups El and E2 are uniformly cholinergic and that therefore the aforementioned -excitatory and inhibitory influences (if indeed present in the chinchilla) upon primary afferent discharge mediated by these neurons are manifested through differential postsynaptic mechanisms. The :mme can be extended to the 6007o of group CPR efferents that are also shown to be choiinergic. Because a substantial fraction of CPR neurons were found to be noncholinergic, the possibility must be considered that some of these differential influences may be mediated through a transmitter mechanism other than acetylcholine.
These physiological findings are a reflection of the neurochemical characteristics of the ves-
D. Bridgeman eta!
tibular end-organs and the afferent and the efferent neurons. The effect of a neuron on a target cell is determined by the agent released by the neuron and by the type of receptor present on the target cell. For example, binding of ACh to neuronal nicotinic acetylcholine receptor (AChR) opens an ion channel resulting in a fast, short-lasting response. Muscarinic AChR are coupled to a second messenger system, and therefore binding of .ACh to these types of 1\ChR's slow,
afferent neurons. In the rat, a:4 and 132 nicotinic AChR subunit mRNAs are expressed by discrete populations of primary afferent neurons (9). Furthermore, mRNA coding for different muscarinic AChR subtypes have been shown to be selectively expressed in Scarpa's ganglia and the vestibular end-organs of the human and rat (15). Since cholinergic axo-somatic and axo-dendritic terminals exist in the human vestibular periphery (16), studies of the topographic distributions of nicotinic and muscarinic AChRs in the human vestibular end-organs will provide functional information regarding efferent function. · The physiological data (12-14) can be ex
plained by the putative neurochemistry of mammalian EVN. Fast responses may be due to the release of ACh on the vestibular afferents that express nicotinic AChR (9), whereas slow, long-lasting effects may be due to activation by ACh of muscarinic AChR (15,17) or to a different mechanism such as the release of neuropeptides by the vestibular efferents. Indeed, the electrical stimulation of the efferents required to obtain a slow response (typically prolonged, high frequency) were those that may release neuropeptides (18, l
In conclusion, we demonstrated that in the chinchilla nearly all of the group E efferents were cholinergic, while in the CPR only about half of the efferents were cholinergic. The complex physiologic response to efferent stimulation is mediated by the specific types of acetylcholine receptors expressed by primary afferent neurons and type II vestibular hair cells, as well as by noncholinergic neurotransmission by discrete populations of EVNs.

ChAT mRNA in Vestibular Efferent Neurons
Acknovvledgments- The authors thank Dr. Vicente Honrubia for critically reading the manuscript and for helpful comments and discussions. Supported (in part) by grants from the National Institutes of Deafness and Other Communication Disorders,
211
NIH/NIDCD P60DC01404-03, 1K08DC00062-04 (PAW), and IR29DC02971-01 (PAW); and the National Organization for Hearing Research (PP & PAW).
REFERENCES
1. Petroff E. An experimental investigation of the ori-gin of efferent fiber projections to the vestibular neuro-epithelium. Anat Rec. 1995; 121:352-60.
2. Rasmussen GL, Gacek R. Concerning the question of an efferent fiber component of the vestibular nerve of the cat. Anat Rec. 1958; 130:361-2.
3. Warr W. Olivocochlear and vestibular efferent neu-rons of the feline brain stem: their location, morpho!-ogy, and number determined by retrograde axonal transport and acetylcholinesterase histochemistry. J Comp Neurol. 1975;161:159-82.
4. Perachio AA, Kevetter GA. Identification of vestib-ular efferent neurons in the gerbil: immunohistochem-ical and retrograde labelling. Exp Brain Res. 1989; 64:315-26.
5. Schwarz DWF, Satoh K, Schwarz IE, Hu K, Fibiger HC. Cholinergic innervation of the rat's labyrinth. Exp Brain Res. 1986;64:19-26.
6. Marco J, Lee WS, Hoffman L, Honrubia V. Effer-ent vestibular neurons in the chinchilla. Adv Otorhi-nolaryngol. 1990;45:82-93.
7. Popper P, Wong Marco RA, Bridgeman D, Hoffman LF, Micevych PE~ Wackym PA. Coexistence of ace-tylcholine and calcitonin gene-related peptide in the vestibular efferent neurons of the chinchilla. Abstr Assoc Res Otolaryngol. 1993;16:23.
8. Popper P, Micevych PE. Steroid regulation of calcite-nin gene-related peptide mRNA expression in moto-neurons of the spinal nucleus of the bulbocavernosus. Brain Res Mol Brain Res. 1990;8: 159-66.
9. Wackym PA, Popper P, Lopez I, Ishiyama A, Micevych PE. Expression of a4 and {32 nicotinic ace-tylcholine receptor subunit mRNA localization of a-bungarotoxin binding proteins in the rat vestibu-lar periphery. Cell Bioi Int. 1995;19:291-300.
10. Wackym PA, Popper P, Micevych PE. Distribution
Abbreviations for Figures 1 and\ 2
4V 6 7n 8vn
= 4th ventricle = abducens nu = facial nerve = vestibular root of vestibula
cochlear nerve asc 7 g7
= ascending fibers of facial nerve = genu of facial nerve
of calcitonin gene-related peptide mRNA and immu-noreactivity in the rat central and peripheral vestib-ular system. Acta Otolaryngol (Stockh). 1993; 113: 601-8.
11. Abelson L, Micevych PE. Distribution of preprocho-lecystokinin mRNA in motoneurons of the rat brain-stem and spinal cord. Brain Res Mol Brain Res. 1991; 10:327-35.
12. Goldberg JM, Fernandez C. Efferent vestibular sys-tern in the squirrel monkey: anatomical location and influence on afferent activity. J Neurophysiol. 1980; 43:986-98.
13. Dickman JD, Correia MJ. Bilateral communication between vestibular labyrinths in pigeons. Neurosci-ence. 1993;57:1097-108.
14. Valli P, Costa J, Zucca G. Local mechanisms in ves-tibular receptor control. Acta Otolaryngol (Stockh). 1984;97:611-8.
15. Wackym PA, Chen CT, Ishiyama A, Pettis RM, Lopez I, Hoffman L. Muscarinic acetylcholine recep-tor subtype mRNAs in the human and rat vestibular periphery. Cell Bioi Int. 1996:20(In press).
16. lshiyama A, Lopez I, Wackym PA. Choline acetyl-transferase immunoreactivity in the human vestibu-lar end-organs. Cell Bioi Int. 1994;18:979-84.
17. Ogawa K, Schacht J. Receptor-mediated release of inositol phosphates in the cochlear and vestibular sen-sory epithelia of the rat. Hearing Res. 1994;69:207-14.
18. Sakaguchi M, lnaishi Y, Kashihara Y, Kuno M. Re-lease of calcitonin gene-related peptide from nerve ter-minals in rat skeletal muscle. J Physic!. 1991 ;434: 257-70.
19. Lundberg JM. Evidence for coexistence of vasoactive intestinal polypeptide (VIP) and acetylcholine neu-rons of cat exocrine glands. Acta Physiol (Scand). 1982;116(Suppl. 496):393-402.
contra CPR = caudal pontine reticular group contralateral to Fluoro-gold injection
contra El = efferent vestibular group lateral to g7, contralateral to Fluoro-gold injection
contra E2 = efferent vestibular group medial to g7, contralateral to Fluoro-gold injection
[·.

CPR
DC DMSp5
DVe El
E2
ECu FIX
GiA
lCp
ipsi CPR
ipsi El
ipsi E2
= efferent vestibular group in the caudal pontine reticular nu
= dorsal cochlear nu = spinal trigeminal nu/dorso
medial = dorsal vestibular nu = efferent vestibular group lat
eral to g7 = efferent vestibular group me
dial to g7 = external cuneate nu = F cell group vestibular /nu
~:!eu~ X reticular
= gigantocellular reticular nu/ alpha
= inferior cerebellar peduncle = caudal pontine reticular group
ipsilateral to Fluoro-gold injection
= El group ipsilateral to Fluorogold injection
= E2 group ipsilateral to Fluorogold injection
LSO LVe LVPO ml mlf MVe MVPO PCRtA
pd PnC PnV
py RlVIg RPa
rs Sol sp5 Sp50 Tz VCA VCP
0. Bridgeman et al
= lateral superior olive = lateral vestibular nu = lateroventral periolivary nu = medial lemniscus = medial longitudinal fasciculus = medial vestibular n u = medioventral periolivary nu = parvocellular reticular nu/
alpha = predorsal bundle = pontine reticular nu/caudal = caudal reticular nu/ventral = hypoglossal nu = pyramidal tract = raphe magnus nu = raphe pallidus (postpyramidal
raphe) nu = rubrospinal tract = nu solitary tract = spinal trigeminal tract = spinal trigeminal nu/oral = nu trapezoid body =ventral cochlear nu/anterior = ventral cochlear nu/posterior