développement folliculaire ovarien et ovulation chez les ... · vironnement nécessaire à sa...
TRANSCRIPT

HAL Id: hal-00793623https://hal.inria.fr/hal-00793623
Submitted on 22 Feb 2013
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Développement folliculaire ovarien et ovulation chez lesmammifères
Danielle Monniaux, Alain Caraty, Frederique Clement, Rozenn Dalbiès-Tran,J. Dupont, Stéphane Fabre, N. Gérard, Pascal Mermillod, Philippe Monget,
Svetlana Uzbekova
To cite this version:Danielle Monniaux, Alain Caraty, Frederique Clement, Rozenn Dalbiès-Tran, J. Dupont, et al..Développement folliculaire ovarien et ovulation chez les mammifères. Inra Prod.Anim, INRA, 2009,22 (6), pp.59-76. <hal-00793623>

La production de gamètes femellesfécondables, ou ovogenèse, est un pro-cessus de développement long et dis-continu. Chez les mammifères, la méiose qui aboutit à la formation degamètes haploïdes, ou ovules, débute austade fœtal, se bloque au stade de pro-phase de première division méiotiquelors de la formation des follicules ova-riens, puis reprend à l’ovulation pour sebloquer à nouveau au stade de méta-phase de deuxième division méiotique,stade auquel l’ovocyte mature est expulsé dans les voies génitales femelles.Ce dernier blocage ne sera levé qu’à lafécondation si elle a lieu, et l’ovule estdonc une cellule «fugace et rare».
L’ovogenèse est d’autant plus com-plexe qu’elle s’imbrique dans un autreprocessus de développement, la follicu-logenèse, avec laquelle elle entretientdes liens étroits. Ainsi, le folliculeapporte à l’ovocyte qu’il renferme l’en-vironnement nécessaire à sa croissanceet à l’acquisition de sa compétence à la fécondation et au développementembryonnaire. A l’inverse, la promotiondu développement folliculaire par l’ovo-cyte a été longtemps ignorée, mais estmaintenant bien établie (Matzuk et al2002, Knight et Glister 2006). L’en-semble des connaissances actuellesconfirme que l’ovogenèse est absolu-
ment indissociable de la folliculo-genèse.
Le développement des follicules ova-riens, ou folliculogenèse, commence audémarrage de croissance du folliculeprimordial et se termine à l’ovulation.L’ovulation, stade ultime de la follicu-logenèse, ne se produit que si les carac-téristiques endogènes (développemen-tales, hormonales, métaboliques) del’individu et son environnement (sai-son, nutrition, interactions sociales) lepermettent. L’influence des facteursenvironnementaux sur le développe-ment folliculaire et l’ovulation variefortement selon les espèces de mammi-fères considérées, comme l’illustre parexemple l’existence d’animaux à repro-duction saisonnée ou non, et d’animauxà ovulation spontanée ou déclenchéepar la saillie. Néanmoins, pour tous lesmammifères, la maturation folliculaireet l’ovulation sont sous le contrôledirect du système hypothalamo-hypo-physaire qui intègre les informationsdes facteurs endogènes et exogènes, etles retraduit par des modifications desécrétion des gonadotropines FSH etLH1. Le rôle crucial des gonadotropi-nes dans le contrôle de la folliculogenè-se et le déclenchement de l’ovulationest désormais bien établi. Cependant,ces processus sont aussi régulés par de
très nombreux autres facteurs, dontl’importance varie au cours de la folli-culogenèse. L’objectif de cet article derevue est de présenter l’état de nosconnaissances sur le développementfolliculaire ovarien et l’ovulation chezles mammifères, en intégrant le rôlerespectif de l’ovocyte et du folliculedans ces processus, et en tentant de hié-rarchiser l’importance des principauxfacteurs qui les régulent.
1 / Le développement folli-culaire : principaux chan-gements morphologiques etfonctionnels
1.1 / Caractéristiques morpho-logiques et dynamiques
Chez les mammifères, la colonisationdes crêtes génitales par les cellules ger-minales primordiales initie un pro-gramme de développement mâle (laspermatogenèse) ou femelle (l’ovoge-nèse) dans les gonades fœtales.L’orientation de ce programme estconditionnée par le caryotype (présen-ce ou absence d’un chromosome Y) descellules somatiques de la gonade et sondéroulement est le résultat d’interac-
Inra Productions Animales, 2009, numéro 2
Inra Prod. Anim., 2009,22 (2), 59-76
D. MONNIAUX1, 2, 3, 4, A. CARATY1, 2, 3, 4, F. CLÉMENT5, R. DALBIÈS-TRAN1, 2, 3, 4, J. DUPONT1, 2, 3, 4, S. FABRE,1, 2, 3, 4, N. GÉRARD1, 2, 3, 4, P. MERMILLOD1, 2, 3, 4, P. MONGET1, 2, 3, 4, S. UZBEKOVA1, 2, 3, 4
1 INRA, UMR85 Physiologie de la Reproduction et des Comportements, F-37380 Nouzilly, France2 CNRS, UMR6175 Physiologie de la Reproduction et des Comportements, F-37380 Nouzilly, France
3 Université François Rabelais de Tours, F-37041 Tours, France4 Haras Nationaux, F-37380 Nouzilly, France
5 INRIA, Centre de Recherche Paris-Rocquencourt, Domaine de Voluceau, Rocquencourt, F-78153 Le Chesnay, FranceCourriel : [email protected]
Développement folliculaire ovarienet ovulation chez les mammifères
Chez les mammifères, la fonction de l’ovaire est de produire à chaque ovulation un ouplusieurs ovocytes fécondables et de créer un environnement hormonal propice au démarrageéventuel d’une gestation. Au sein de l’ovaire, deux processus de développement étroitementimbriqués, l’ovogenèse et la folliculogenèse, déterminent le nombre et la qualité des ovocytesproduits. Ces processus, initiés pendant la vie fœtale, se poursuivent pendant toute la vie de lafemelle et sont étroitement contrôlés à chacune de leurs étapes par de nombreux facteurshormonaux et environnementaux.
1 Pour la définition des abréviations des protéines et des gènes cités dans l’article, cf. annexe.* La liste des co-auteurs, par ordre alphabétique, indique leur contribution identique à la réalisation de cet article.
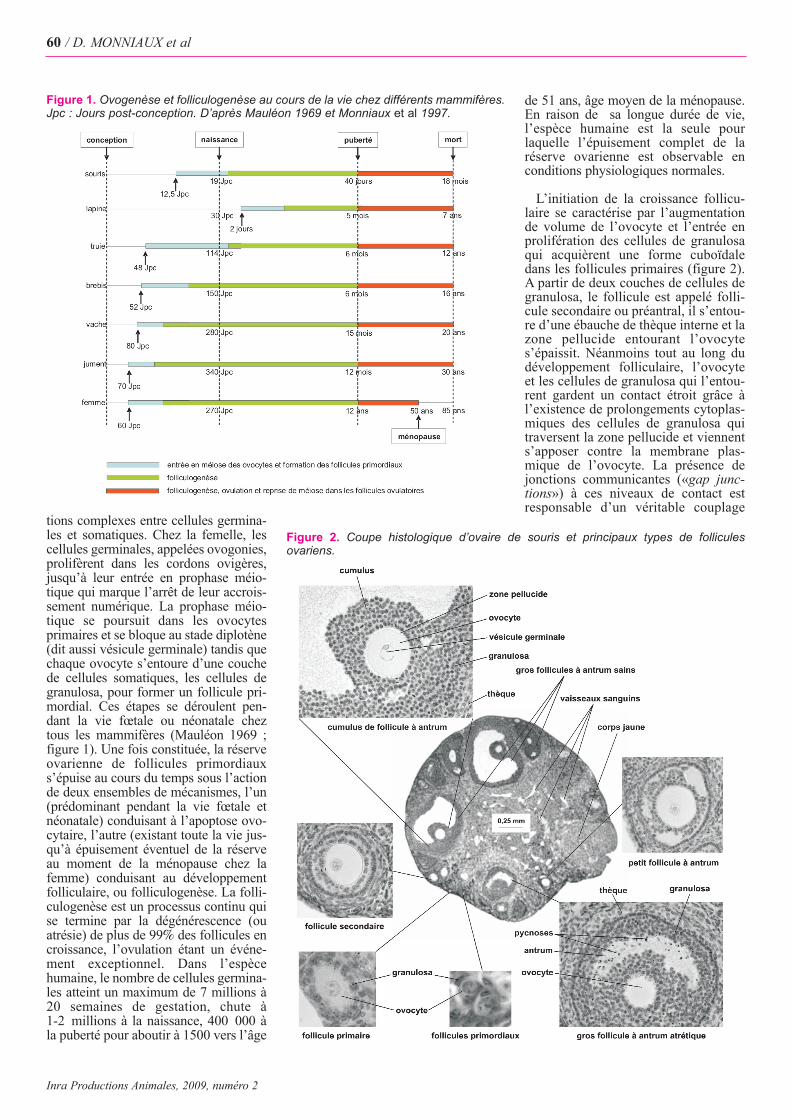
tions complexes entre cellules germina-les et somatiques. Chez la femelle, lescellules germinales, appelées ovogonies,prolifèrent dans les cordons ovigères,jusqu’à leur entrée en prophase méio-tique qui marque l’arrêt de leur accrois-sement numérique. La prophase méio-tique se poursuit dans les ovocytesprimaires et se bloque au stade diplotène(dit aussi vésicule germinale) tandis quechaque ovocyte s’entoure d’une couchede cellules somatiques, les cellules degranulosa, pour former un follicule pri-mordial. Ces étapes se déroulent pen-dant la vie fœtale ou néonatale chez tous les mammifères (Mauléon 1969 ; figure 1). Une fois constituée, la réserveovarienne de follicules primordiaux s’épuise au cours du temps sous l’actionde deux ensembles de mécanismes, l’un(prédominant pendant la vie fœtale etnéonatale) conduisant à l’apoptose ovo-cytaire, l’autre (existant toute la vie jus-qu’à épuisement éventuel de la réserveau moment de la ménopause chez lafemme) conduisant au développementfolliculaire, ou folliculogenèse. La folli-culogenèse est un processus continu quise termine par la dégénérescence (ouatrésie) de plus de 99% des follicules encroissance, l’ovulation étant un événe-ment exceptionnel. Dans l’espècehumaine, le nombre de cellules germina-les atteint un maximum de 7 millions à20 semaines de gestation, chute à 1-2 millions à la naissance, 400 000 àla puberté pour aboutir à 1500 vers l’âge
de 51 ans, âge moyen de la ménopause.En raison de sa longue durée de vie,l’espèce humaine est la seule pourlaquelle l’épuisement complet de laréserve ovarienne est observable enconditions physiologiques normales.
L’initiation de la croissance follicu-laire se caractérise par l’augmentationde volume de l’ovocyte et l’entrée enprolifération des cellules de granulosaqui acquièrent une forme cuboïdaledans les follicules primaires (figure 2).A partir de deux couches de cellules degranulosa, le follicule est appelé folli-cule secondaire ou préantral, il s’entou-re d’une ébauche de thèque interne et lazone pellucide entourant l’ovocytes’épaissit. Néanmoins tout au long dudéveloppement folliculaire, l’ovocyteet les cellules de granulosa qui l’entou-rent gardent un contact étroit grâce àl’existence de prolongements cytoplas-miques des cellules de granulosa quitraversent la zone pellucide et viennents’apposer contre la membrane plas-mique de l’ovocyte. La présence dejonctions communicantes («gap junc-tions») à ces niveaux de contact estresponsable d’un véritable couplage
60 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Figure 1. Ovogenèse et folliculogenèse au cours de la vie chez différents mammifères.Jpc : Jours post-conception. D’après Mauléon 1969 et Monniaux et al 1997.
Figure 2. Coupe histologique d’ovaire de souris et principaux types de folliculesovariens.

métabolique entre ces deux types cellu-laires, permettant des échanges d’ionset de petites molécules (PoidsMoléculaire < 1 kD). Quand le follicu-le atteint une taille voisine de 0,2 mmde diamètre, une cavité, appeléeantrum, se forme à l’intérieur de lagranulosa. C’est dans l’antrum ques’accumulent les produits de sécrétiondes cellules folliculaires (granulosa etthèque), ainsi que les substances plas-matiques diffusant librement à partirdes capillaires sanguins qui irriguent lathèque. A ce stade, le follicule àantrum, appelé aussi follicule tertiaire,comporte une thèque externe, unethèque interne séparée de la granulosapar la lame basale et un ovocyte entou-ré d’un massif de cellules appelé cumu-lus. Au cours du développement termi-nal du follicule à antrum, la taille del’ovocyte n’augmente plus guère, laprolifération des cellules de granulosadiminue progressivement, et la crois-sance folliculaire jusqu’au stade pré-ovulatoire s’effectue essentiellementpar accroissement du volume de l’an-trum. Cependant, l’immense majoritédes follicules n’atteint jamais le stadepréovulatoire. L’atrésie folliculairedébute par l’apoptose de l’ovocyte dansles follicules primaires et préantraux,et par l’apoptose des cellules de granu-losa dans les follicules à antrum. Dansces cellules, la phase finale de l’apopto-se est visible sous forme d’agré-gats d’ADN, les grains de pycnose(figure 2).
La durée totale du développementfolliculaire varie d’une vingtaine dejours chez les rongeurs à plusieurs mois
chez les mammifères de grande taille,dont la femme. Dans chaque espèce, ledéveloppement des follicules jusqu’àl’apparition de l’antrum est très lent etreprésente au moins 75% de la duréetotale du développement folliculaire(Monniaux et al 1997, McGee et Hsueh2000) (tableau 1). Le développementdes follicules à antrum, plus rapide, estcependant plus long que la durée d’uncycle sexuel pour toutes les espèces demammifères, sauf chez les rongeurs oùelle correspond exactement à la duréedu cycle.
1.2 / Les étapes-clés de la follicu-logenèse et du développementovocytaire
D’un point de vue fonctionnel, ledéveloppement folliculaire peut se sub-diviser en deux phases successives : lafolliculogenèse basale et la folliculoge-nèse terminale (figure 3).
La folliculogenèse basale se dérouleapparemment normalement en l’absen-ce de FSH, ce qui est observable expé-rimentalement après hypophysectomie(rat, brebis), désensibilisation hypo-physaire par traitement long avec unagoniste du GnRH (toutes espèces) ouinvalidation du gène (knock-out)codant la chaîne β de la FSH ou sonrécepteur chez la souris (Kumar et al1997). Un développement folliculairebasal existe aussi chez des femmes por-teuses de mutations affectant la fonc-tion de FSH ou de son récepteur. Danstous ces cas, le développement follicu-laire se poursuit jusqu’à une taille limi-
te, caractéristique de chaque espèce,variant de 0,2 mm chez les rongeurs à10 mm chez la jument (Monniaux et al1997) (tableau 2). La folliculogenèsebasale est contrôlée par de nombreuxfacteurs de croissance, d’origines ovo-cytaire et somatique, agissant essentiel-lement selon un mode paracrine derégulation (cf. § 2 ). C’est au cours decette phase que s’effectue l’essentiel dela croissance de l’ovocyte, à partir d’undiamètre initial de 20 à 30 microns, etque l’ovocyte acquiert la compétenceméiotique, c’est-à-dire la capacité àreprendre la méiose (bloquée au stadediplotène/diacinèse) quand il est extraitde son follicule.
La folliculogenèse terminale est stric-tement dépendante de la présence deFSH et, pour les stades terminaux dematuration du follicule préovulatoire,de la présence de LH. L’apparition derécepteurs de LH sur les cellules degranulosa est la «signature» d’unematurité complète du follicule, quidevient apte à ovuler. Cette maturitéfinale est atteinte pour des tailles folli-culaires variant entre 0,5 mm de dia-mètre chez les rongeurs et 15 mm dediamètre chez la jument (tableau 2). Lafolliculogenèse terminale est contrôléeessentiellement par FSH et LH, mais denombreux autres facteurs (facteurs decroissance, matrice extracellulaire, pro-téases, stéroïdes), d’origine locale ouendocrine, agissent en synergie avec lesgonadotropines pour réguler son dérou-lement (McGee et Hsueh 2000). C’estau cours de cette phase que s’effectuela sélection du ou des follicule(s) desti-né(s) à ovuler, grâce à un ensemble demécanismes dont la finalité biologiqueest de réguler le nombre d’ovulationscaractéristique de chaque espèce et dechaque race (cf. § 3). En début de folli-culogenèse terminale, l’ovocyte termi-ne sa croissance pour atteindre un dia-mètre de 80 à 125 microns selon lesespèces. Puis il subit des remaniementschromatiniens associés à l’acquisitionde la compétence au développement,c’est-à-dire la capacité à assurer undéveloppement embryonnaire normalaprès fécondation.
Développement folliculaire ovarien et ovulation chez les mammifères / 61
Inra Productions Animales, 2009, numéro 2
Tableau 1. Durée du développement folliculaire (en jours) chez différents mammifères.
ND : non déterminé.
Tableau 2. Diamètres folliculaires (en mm) aux principales étapes du développement folliculaire chez différents mammifères.

1.3 / Changements fonctionnelsdans les cellules somatiques etl’ovocyte au cours de la follicu-logenèse
De nombreux changements fonction-nels, aussi bien dans les cellules soma-tiques du follicule que dans l’ovocyte,accompagnent le développement folli-culaire (figure 4).
Lors du démarrage du développe-ment folliculaire, les cellules de granu-losa entrent en prolifération. Leur acti-vité de prolifération atteint unmaximum en fin de folliculogenèsebasale, stade pendant lequel près de100% des cellules sont proliférantes.Parallèlement, ces cellules de granulosasynthétisent de nombreux facteurs decroissance (EGF, FGF, AMH) et descytokines (KITLG). En particulier laproduction d’AMH, faible à indétecta-ble dans les follicules primaires, aug-mente dans les follicules préantrauxpour atteindre un maximum en fin defolliculogenèse basale (Visser etThemmen 2005). Les récepteurs deFSH apparaissent dans la granulosa desfollicules primaires, leur nombre aug-mente légèrement et progressivementdans les follicules préantraux, reste sta-ble dans les follicules à antrum et enfindiminue au stade préovulatoire. Les
cellules de thèque interne expriment,dès leur formation, des facteurs decroissance, des récepteurs de LH, ainsique des enzymes et des facteurs-clés de la stéroïdogenèse, permettant la syn-thèse de progestagènes (CYP11A1,STAR, HSD3B2) et d’androgènes(CYP17A1).
Au cours de la folliculogenèse termi-nale, les cellules de granulosa perdentprogressivement leur activité de proli-fération et de synthèse d’AMH, et sedifférencient en cellules stéroïdogènespar l’apparition et l’augmentation del’expression des gènes CYP11A1,STAR, HSD3B2, et surtout CYP19A1qui code l’enzyme aromatase, permet-tant la synthèse d’oestradiol à partir des androgènes d’origine thécale.Simultanément, ces cellules de granu-losa deviennent sensibles à LH (appari-tion et augmentation brutale des récep-teurs de LH, LHCGR). En fin de cycleovarien, le pic préovulatoire de LHinduit d’importants changements fonc-tionnels dans les cellules de granulosaet de thèque du follicule préovulatoire,conduisant à leur lutéinisation et à laformation du corps jaune (cf. § 4).
62 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Figure 3. Principales étapes du développement folliculaire et de la maturation ovocytaire.Dans le cas des rongeurs, la dépendance à FSH est acquise au moment de la forma-tion de l’antrum.
Figure 4. Changements fonctionnels dans les cellules de granulosa et l’ovocyte aucours du développement folliculaire. Dans le follicule après l’ovulation, l’expression de l’aromatase (CYP19A1) disparaît aumoment de la formation du corps jaune chez les ruminants, la truie et la jument, maispas chez les primates et les rongeurs (courbe orange en pointillé).

En parallèle avec le démarrage de sacroissance, l’ovocyte initie une intenseactivité transcriptionnelle. Une partiedes ARN supporte la synthèse pro-téique pour assurer le métabolisme del’ovocyte, tandis qu’une autre partieest stockée sous une forme stable ausein de particules ribonucléopro-téiques, à l’abri de la machinerie detraduction, pour un recrutement ulté-rieur. Certains gènes sont exprimésspécifiquement ou préférentiellementdans l’ovocyte, comme ceux codantles facteurs de croissance BMP15 etGDF9, les composants de la zone pel-lucide (protéines ZP), ainsi que desfacteurs dont le rôle est encoreinconnu (Zheng et Dean 2007).L’activité transcriptionnelle ralentitlorsque l’ovocyte approche de son dia-mètre maximal, jusqu’à devenir indé-tectable. Cette période de fin de crois-sance ovocytaire coïncide avecl’acquisition de propriétés fonction-nelles. L’ovocyte semble acquérir d’a-bord la compétence à reprendre laméiose (lorsqu’il atteint environ 80%de sa taille maximale) puis, à partird’un diamètre folliculaire caractéris-tique de chaque espèce, l’aptitude àsoutenir le développement précoce del’embryon (Mermillod et Marchal1999). Ce modèle repose sur des
observations statistiques, et rendcompte de la proportion d’ovocytescompétents dans une population defollicules à un stade donné, plutôt quede la dynamique d’un ovocyte particu-lier. Récemment, un autre modèle asuggéré que les ovocytes sont dansune certaine mesure prédestinés, etque les ovocytes incompétents sontéliminés progressivement au cours dela folliculogenèse via l’atrésie(Mermillod et al 2008). Quoi qu’il ensoit, la compétence de l’ovocyte sem-ble associée à l’évolution vers uneconfiguration chromatinienne conden-sée en un anneau périnucléolaire, quicoïncide avec l’inactivation des ARNpolymérases. En fin de folliculogenè-se, l’expression génique repose essen-tiellement sur le contrôle post-trans-criptionnel de la réserve d’ARN.
2 / Régulations du démar-rage de croissance follicu-laire et du développementfolliculaire basal
Les principales régulations du déve-loppement folliculaire basal sont sché-matisées dans la figure 5.
2.1 / Interactions cellules soma-tiques - ovocyte au cours de lamise en place de la réserve defollicules primordiaux et dudémarrage de croissance follicu-laire
L’effectif de la réserve de folliculesprimordiaux détermine la longévité dela fonction ovarienne. Il résulte vrai-semblablement de l’existence, dansl’ovaire fœtal, d’un dialogue molécu-laire étroit entre cellules somatiques etcellules germinales, mais les acteurs etles mécanismes en sont encore très malconnus. L’invalidation du gène codantle facteur de transcription ovocytaireFIGLA n’empêche pas la mise en placeet la multiplication des ovogonies dansl’ovaire fœtal, mais bloque la formationdes follicules primordiaux (Soyal et al2000). Par ailleurs, l’inactivation natu-relle ou expérimentale de nombreuxfacteurs impliqués dans le contrôle dudéclenchement (STRA8) et du bondéroulement de la méiose dans les cel-lules germinales (facteurs contrôlantles appariements ou les recombinaisonschromosomiques tels que CPEB1,DMC1, ATM et MSH5) conduit à uneperte massive des cellules germinales
Développement folliculaire ovarien et ovulation chez les mammifères / 63
Inra Productions Animales, 2009, numéro 2
Figure 5. Régulations du développement folliculaire basal.

par apoptose pendant la vie fœtale(Baltus et al 2006). En outre, des cyto-kines d’origine somatique telles queKITLG, LIF ou IL1A, les facteursmitochondriaux de la familleBCL2/BAX et les caspases (CASP2,CASP3) jouent un rôle essentiel dansla survie des cellules germinales et desfollicules primordiaux nouvellementformés ; leur inactivation, et à l’inver-se leur surexpression, modulent defaçon très importante le taux d’apop-tose des cellules germinales et par voie de conséquence la taille de laréserve ovarienne (Morita et Tilly1999).
Une fois les follicules primordiauxformés, certains vont démarrer leurcroissance immédiatement et d’autresvont attendre, pendant plusieurs moisou plusieurs années, un signal dedémarrage de nature encore inconnue.Nous ignorons aussi si ce signal pro-vient de l’ovocyte, des cellules de gra-nulosa qui l’entourent, du cortex ova-rien qui renferme les folliculesprimordiaux, ou s’il est endocrinien.
Plusieurs facteurs ovocytaires sem-blent jouer un rôle important dansl’initiation de croissance des folliculesprimordiaux. Chez la souris, l’invali-dation des gènes codant les facteurs detranscription ovocytaires NOBOX,LHX8, SOHLH1 ou SOHLH2 bloquece démarrage (Choi et Rajkovic 2006,Pangas et al 2006). Leur expressioncoordonnée contrôle l’expression d’ungrand nombre de gènes spécifiques del’ovocyte. En revanche, l’inactivationdans l’ovocyte des facteurs ubiquistesFOXO3 ou PTEN induit un démarragede croissance massif et un épuisementaccéléré de la réserve (Castrillon et al2003, Reddy et al 2008).
Des facteurs d’origine somatiqueseraient aussi impliqués dans ledémarrage de croissance folliculaire.Ainsi la cytokine KITLG, expriméepar les cellules de granulosa dès lestade de follicule primordial, est capa-ble de stimuler la croissance ovocytai-re in vitro, et la présence de mutationsinactivatrices dans ce gène bloque ledémarrage de croissance folliculaire invivo chez la souris, mais pas chez lafemme (Huang et al 1993, Klinger etDe Felici 2002). Le facteur de crois-sance BMP4, exprimé par les cellulesstromales du cortex ovarien, pourraitaussi réguler ce démarrage ; en effet,son administration à des rates ou dessouris stimule l’initiation de croissan-ce des follicules primordiaux (Nilssonet Skinner 2003).
2.2 / Interactions cellules soma-tiques - ovocyte au cours de lafolliculogenèse basale
Dès le stade de follicule primaire,l’ovocyte exprime et sécrète les fac-teurs BMP15 et GDF9 qui jouent unrôle crucial dans le déroulement de lafolliculogenèse basale. Ces facteurs sti-mulent la prolifération des cellules degranulosa et favorisent leur survie.L’invalidation du gène codant GDF9chez la souris conduit au blocage de lacroissance folliculaire au stade de folli-cule primaire et la présence, à l’étathomozygote, de mutations inactivatri-ces dans les gènes BMP15 ou GDF9 estassociée à un phénotype similaire chezla brebis (Juengel et McNatty 2005).En retour, les cellules de granulosa par-ticipent à la croissance ovocytaire parla production de KITLG, dont l’expres-sion est modulée par BMP15 et GDF9d’origine ovocytaire (Shimasaki et al2004).
L’existence d’un couplage métabo-lique entre ovocyte et granulosa consti-tue un autre élément déterminant dudialogue entre ces deux types cellulai-res. Par exemple, l’ovocyte contrôle letransport actif d’acides aminés dans lescellules de cumulus, et ces acides ami-nés peuvent être ensuite transférés àl’ovocyte par l’intermédiaire de jonc-tions communicantes (Eppig et al2005). La perte des jonctions commu-nicantes entre ovocyte et granulosa (parinvalidation du gène GJA4 codant laconnexine 37), a un effet délétère sur lacroissance et la survie de l’ovocyte(Carabatsos et al 2000).
Les cellules thécales contribuentaussi à la régulation du développementfolliculaire basal par la sécrétion d’an-drogènes (testostérone) et de nombreuxfacteurs de croissance (BMP4, BMP7,NGF, FGF7, EGF, TGF...) capables destimuler la prolifération des cellules degranulosa. L’activité des cellules dethèque est elle-même régulée par desfacteurs sécrétés par les cellules de gra-nulosa, en particulier le KITLG(Skinner 2005).
2.3 / Régulations endocrinesLe contrôle de la folliculogenèse
basale s’effectue essentiellement selonun mode paracrine de régulation.Néanmoins, le rôle modulateur dequelques facteurs endocriniens mérited’être signalé. En premier lieu, l’AMH,sécrétée par les cellules de granulosades follicules en croissance (mais pasdes follicules primordiaux), jouerait unrôle déterminant de «gardien» de la
réserve ovarienne puisque son invalida-tion chez la souris conduit à un démar-rage de croissance folliculaire massif etun épuisement accéléré de cette réserve(Durlinger et al 1999). D’autre part,même si le développement folliculairepeut se dérouler en absence de gonado-tropines comme nous l’avons dit, lescellules folliculaires n’y sont pas pourautant insensibles. Ainsi FSH et LHsont capables de stimuler le développe-ment folliculaire basal par leur actionsur les cellules de granulosa et dethèque, respectivement. En particulier,l’administration de FSH est capable destimuler la prolifération des cellules degranulosa des follicules préantraux etdes petits follicules à antrum. Au coursdes cycles sexuels, de modestes fluc-tuations de concentrations de FSH dansle sang régulent la transition vers ledéveloppement folliculaire terminal etle démarrage des vagues folliculaires.L’insuline et l’IGF1 stimulent cettetransition en augmentant la sensibilitéfolliculaire à FSH (Mazerbourg et al2003).
3 / Les événements et larégulation du développe-ment folliculaire terminal
3.1 / Principaux facteurs régula-teurs
Par définition, le développement fol-liculaire terminal est strictement dépen-dant de la présence de gonadotropines,et particulièrement de FSH qui en est lechef d’orchestre. Outre les gonadotro-pines, les principaux régulateurs de lacroissance folliculaire terminale sontl’IGF1 et l’insuline, qui sensibilisentles follicules à FSH. En particulier,l’action synergique de FSH et d’IGF1joue un rôle déterminant dans le débutde la phase terminale de croissance,permettant l’émergence du futur folli-cule préovulatoire. Ensuite, la domi-nance du follicule préovulatoire estassurée par la LH, hormone à laquellece follicule est progressivement devenuhypersensible et qui prend le relais dela FSH.
Les gonadotropines FSH et LH agis-sent sur leurs cellules-cibles via leursrécepteurs membranaires spécifiques,respectivement FSHR et LHCGR.L’activation de ces récepteurs par leursligands stimule la production d’AMPcpar l’enzyme adenylate cyclase ADCY,puis l’activation de la kinase PRKAC etde nombreuses autres kinases(MAPK1, MAPK14, PIK3). La
64 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2

phosphorylation de différents facteursde transcription (CREB1, NR5A1,SP1) par ces kinases va activer l’ex-pression d’un ensemble de gènes-ciblestels que CYP19A1, INHBB, FSHR etLHCGR dans la granulosa, CYP17A1dans la thèque, et CYP11A1, STAR etHSD3B2 dans ces 2 types cellulaires(Lécureuil et al 2007).
3.2 / Vagues folliculaires etmécanisme de sélection des folli-cules ovulatoires
Le développement folliculaire termi-nal se déroule sous forme de vaguesfolliculaires. Une vague folliculairecorrespond à la croissance synchroned’une cohorte de follicules, suivie de lasélection d’un ou plusieurs folliculesappelés dominants, et de leur évolutionvers l’ovulation ou de leur régressionquand les conditions endocriniennessont défavorables (phase lutéale ducycle ou gestation). Selon les espèces,une seule (femme, rongeurs, truie) ouplusieurs (brebis, vache, jument)
vagues de développement folliculaireterminal sont observables au cours ducycle sexuel (Driancourt et al 2001).Dans toutes les espèces, la vague dedéveloppement folliculaire terminal quiaboutit à l’ovulation démarre aumoment de la lutéolyse, l’ovulationayant lieu en fin de phase folliculairedu cycle sexuel. La jument présente laparticularité de pouvoir ovuler égale-ment pendant la phase lutéale du cycle.
Comme nous l’avons dit, la croissan-ce folliculaire terminale commence à0,2 mm chez les rongeurs, 1-2 mmchez la truie, 2 mm chez la brebis, 3-5mm chez la vache et la femme et 10mm chez la jument. C’est la taille cri-tique au-delà de laquelle la croissancedu follicule est strictement dépendantede niveaux minimaux de FSH. Dans lecas d’une espèce mono-ovulante, le fol-licule qui va ovuler se trouve parmi ungroupe de follicules qui atteignent «parhasard» cette taille au moment de lalutéolyse. Les teneurs sériques en FSHsont alors compatibles avec la poursui-
te de la croissance de ce groupe de fol-licules. Ainsi, lors de chaque vague fol-liculaire, le démarrage synchrone d’ungroupe de follicules en développementterminal est suivi d’une diminution desconcentrations circulantes de FSH, enréponse à la rétroaction négative del’oestradiol et de l’inhibine sécrétés parces follicules sur l’axe hypothalamo-hypophysaire (figure 6 et cf. § 5).
Or, le pool de follicules au seinduquel s’effectue la sélection n’est pashomogène. Bien que des follicules demême taille aient, a priori, les mêmespotentialités, ils ne sont pas fonction-nellement identiques en termes devitesse de prolifération des cellules fol-liculaires, de stéroïdogenèse et de syn-thèse de facteurs de croissance et/ou deprotéines de liaison de ces facteurs. Lesraisons de ces différences entre follicu-les apparemment tous au même stadede développement sont pour le momentinconnues. On peut penser que lescaractéristiques de chaque folliculerésultent à la fois de sa date d’entrée encroissance, de sa position dans l’ovaire,de l’importance de sa vascularisation,de son «historique» d’exposition auxgonadotropines, etc. Quoiqu’il en soit,chaque follicule est donc caractérisépar un équilibre local qui lui est propre,entre des facteurs paracrines stimulants(oestradiol, IGF1, activine...) et inhibi-teurs (androgènes, les IGFBP, protéinesde liaison des IGF, la follistatine qui estune protéine de liaison de l’activine...)de son développement. De fait, une hié-rarchie fonctionnelle s’établit entre fol-licules, pourtant a priori au même stadede développement.
Ainsi, au début de la phase folliculai-re du cycle sexuel, le dialogueovaire/axe hypothalamo-hypophysairemet en jeu plusieurs follicules.Progressivement, la petite «avance»dont pourrait bénéficier un follicule parrapport aux autres s’amplifie. Ce der-nier instaure alors une sorte de dialogueprivilégié en secrétant de plus en plusd’oestradiol, ce qui a pour conséquenceune diminution importante des teneursen FSH, qui provoque l’atrésie des fol-licules de la cohorte. Le follicule dit«dominant» ne souffre pas quant à luide cette baisse des teneurs en FSH,puisqu’il induit, par rétroaction positivecette fois, via l’oestradiol, une forteaugmentation de la fréquence des pul-ses de LH à laquelle il devient hyper-sensible, grâce à la très forte expressionde récepteurs de LH dans ses cellulesde granulosa. L’expression des deuxmarqueurs-clés de la différenciationfolliculaire, CYP19A1 et LHCGR, est
Développement folliculaire ovarien et ovulation chez les mammifères / 65
Inra Productions Animales, 2009, numéro 2
Figure 6. Régulations de la sélection et du développement terminal du follicule ovula-toire pendant la phase folliculaire du cycle ovarien chez une espèce mono-ovulante.La figure représente le dialogue endocrine existant entre les follicules ovariens de lavague ovulatoire et le système hypothalamo-hypophysaire, au début, au milieu et à lafin de la phase folliculaire. Au cours du temps, un seul des follicules de la vague devientprogressivement l’acteur essentiel de ce dialogue et du déclenchement de la déchargepréovulatoire de LH, et c’est ce follicule qui ovulera en réponse à cette décharge. E2 = oestradiol, INH = inhibine.

donc critique à l’établissement de lasélection du follicule préovulatoire(figure 6). Les follicules de la cohortedébutent un processus d’atrésie qui setraduit, dès ses premiers stades, par uneaugmentation de synthèse d’élémentsinhibiteurs (IGFBP, androgènes, TP53,BAX, caspases) et la perte de synthèsed’éléments stimulants (oestradiol,BCL2L1), changements qui ne font que précipiter leur dégénérescence(Monniaux et al 1999).
Ce mécanisme de sélection au coursdu développement folliculaire terminala fait l’objet d’une approche de modéli-sation dans l’espèce ovine (Echenim etal 2005). La figure 7 représente unexemple de compétition entre trois fol-licules en développement terminal, àpartir du pic de prolifération cellulaireau sein de la granulosa (follicules d'en-viron 1 mm de diamètre chez la bre-bis) jusqu'au déclenchement de ladécharge ovulatoire. Le taux de sélec-tion est déterminé conjointement par
les niveaux i) de sensibilité folliculaireà FSH, en termes de prolifération et dedifférenciation cellulaire, ii) de sensibi-lité hypophysaire vis-à-vis du rétro-contrôle négatif exercé par l'inhibine etl'oestradiol d'origine ovarienne et iii)de sensibilité hypothalamique au rétro-contrôle positif exercé par l'oestradiol.Ici, un seul follicule réussit à poursui-vre son développement jusqu'à l'ovula-tion. Il se distingue des deux autres parson effectif cellulaire final (de l'ordrede 7,5 millions de cellules), sa capaci-té de sécrétion en oestradiol (reflétéepar la maturité folliculaire) et une bio-disponibilité locale en FSH bien plusélevée, en liaison avec une vascularisa-tion plus développée.
Le follicule ovulatoire n’est pas pré-destiné, et sa sélection ne résulte pasnon plus d’un tirage au sort, mais du«choix» du follicule qui se trouve parhasard en parfaite harmonie avec lesniveaux circulants de gonadotropines,et ce pendant une fenêtre de temps bien
délimitée. Ainsi, entre une hypothèsedéterministe et une hypothèse stochas-tique de l’initiation de l’atrésie, c’estune hypothèse opportuniste qui semblese dégager. Le follicule qui ovule est lefollicule qui, par chance, était à labonne place au bon moment.
3.3 / La régulation du quotaovulatoire chez les mammifères
Le nombre d’ovulations par cyclesexuel, ou quota ovulatoire, est unecaractéristique de chaque espèce.Certaines espèces sont polyovulantes etdonc très prolifiques (rongeurs, lapine,truie), d’autres essentiellement mono-ovulantes (femme, vache, jument).L’espèce ovine a la particularité de ren-fermer à la fois des races polyovu-lantes (Romanov, Finnoise…) et desraces mono-ovulantes (Ile-de-France,Mérinos…). De plus, au sein de cesraces ovines, il existe des lignées(Booroola, Cambridge, Belclare…)présentant des caractéristiques excep-
66 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Figure 7. Modélisation du processus de sélection : compétition entre trois follicules en phase de développement terminal. Le graphique en haut à gauche illustre la chute de la concentration plasmatique de FSH en réponse à l'augmentation de la maturi-té ovarienne (représentant, de manière agrégée, l'inhibine et l'oestradiol). Les trois autres graphiques illustrent pour chaque follicu-le le niveau de FSH réellement disponible (biodisponibilité locale, graphique en bas à gauche), la maturité folliculaire (graphique enhaut à droite) et l'effectif cellulaire total de la granulosa (graphique en bas à droite). L'effectif et la maturité sont identiques entre fol-licules au début du développement terminal, mais leur sensibilité à FSH est légèrement différente. Seul un des follicules (courbe enbleu), poursuit son développement jusqu'à l'ovulation.

Développement folliculaire ovarien et ovulation chez les mammifères / 67
Inra Productions Animales, 2009, numéro 2
tionnelles en termes de quota ovulatoireet de prolificité. Dans ces lignées, l’ana-lyse du déterminisme génétique de lapoly-ovulation a permis d’identifier dif-férentes mutations dans les éléments dusystème BMP, responsables de change-ments du quota ovulatoire. Ainsi, desmutations conduisant à une perte defonction partielle de BMP15, GDF9, oudu récepteur BMPR1B ont toutes pourconséquence une augmentation duquota ovulatoire (Galloway et al 2000,Mulsant et al 2001, Hanrahan et al2004). Les éléments de cette famille defacteurs de croissance jouent en généralun rôle d’inhibiteurs de la différencia-tion cellulaire et de la lutéinisation.L’hypothèse la plus probable est que cesmutations induisent une avance à lamaturation folliculaire terminale, lesfollicules étant prêts à ovuler plus tôt, àdes tailles réduites pouvant aller de 3,5à 5 mm. Néanmoins, la capacité desynthèse d’oestradiol de chacun de cesfollicules étant inférieure à celle d’unfollicule préovulatoire d’une brebis nonporteuse d’une mutation de prolificité,chez une brebis porteuse c’est la sécré-tion simultanée de plusieurs folliculespréovulatoires qui permet d’atteindre
les niveaux sériques en oestradiolnécessaires au déclenchement du picpréovulatoire (figure 8, d’après Fabre etal 2006).
Comme nous l’avons dit précédem-ment (cf. § 2), la présence, à l’étathomozygote, de mutations conduisant àune perte de fonction totale des gènesBMP15 et GDF9 rendent les brebis sté-riles. Cela signifie que chez la brebis,une «dose» minimale de signalisationBMP est nécessaire à la folliculogenèseovarienne, et que de la valeur de cette«dose» dépend en partie le quota ovula-toire. Signalons enfin que chez la sou-ris, une perte partielle d’expression deBMP15 ou GDF9 (chez des individushétérozygotes pour le gène invalidé) n’aaucune conséquence sur le quota ovula-toire.
3.4 / Différenciation terminale del’ovocyte
Au cours du développement follicu-laire terminal, l’ovocyte subit des mofi-cations discrètes mais importantes d’unpoint de vue fonctionnel. En effet, laproportion d’ovocytes aptes à soutenir
le développement embryonnaire conti-nue à augmenter avec la taille folli-culaire, alors que le diamètre de l’ovocyte, lui, n’augmente quasimentplus. La période de transcription inten-se est achevée, et des vacuoles se déve-loppent dans le résidu nucléolaire(Hyttel et al 2001). La conservationglobale du niveau d’expression pendantcette période n’exclut cependant pas larégulation spécifique de certains trans-crits (Lequarre et al 2005, Mourot et al2006).
4 / L’ovulation et la matu-ration ovocytaire
4.1 / Déclenchement de l’ovula-tion et de la maturation ovocy-taire
L’ovulation est un processus com-plexe au cours duquel sont induits à lafois la reprise de méiose de l’ovocyte,l’expansion du cumulus, la rupture dupôle apical du follicule, et la restructu-ration tissulaire associée à la différen-ciation cellulaire nécessaire à la forma-tion du corps jaune. Au cours d’un cycle
Figure 8. Conséquences des mutations des gènes de prolificité sur la folliculogenèse et le nombre d’ovulations chez la brebis.L’activité réduite du système de signalisation des Bone Morphogenetic Proteins (BMP) chez les brebis porteuses de mutations dansles gènes de prolificité BMP15, GDF9 ou BMPR1B, conduit à diminuer l’action des BMPs, d’une part sur la prolifération et d’autrepart sur la sensibilité à FSH des cellules de la granulosa. Ainsi, les follicules antraux sont de plus petite taille avec moins de cellulesde la granulosa, mais présentent une sensibilité accrue à FSH. Ceci permet d’avancer la maturation des follicules. Ces follicules pluspetits produisent moins d’oestradiol et d’inhibine, mais ensemble, ils en produisent autant que le follicule préovulatoire des brebisnon-mutées. En conséquence, il s’établit le même dialogue endocrinien entre l’ovaire et l’axe hypothalamo-hypophysaire dans lesdeux génotypes. Finalement, cette régulation aboutit à la sélection et à l’ovulation de plusieurs follicules de plus petite taille chez lesbrebis porteuses des mutations dans les gènes de prolificité. E2 = oestradiol, INH = inhibine.
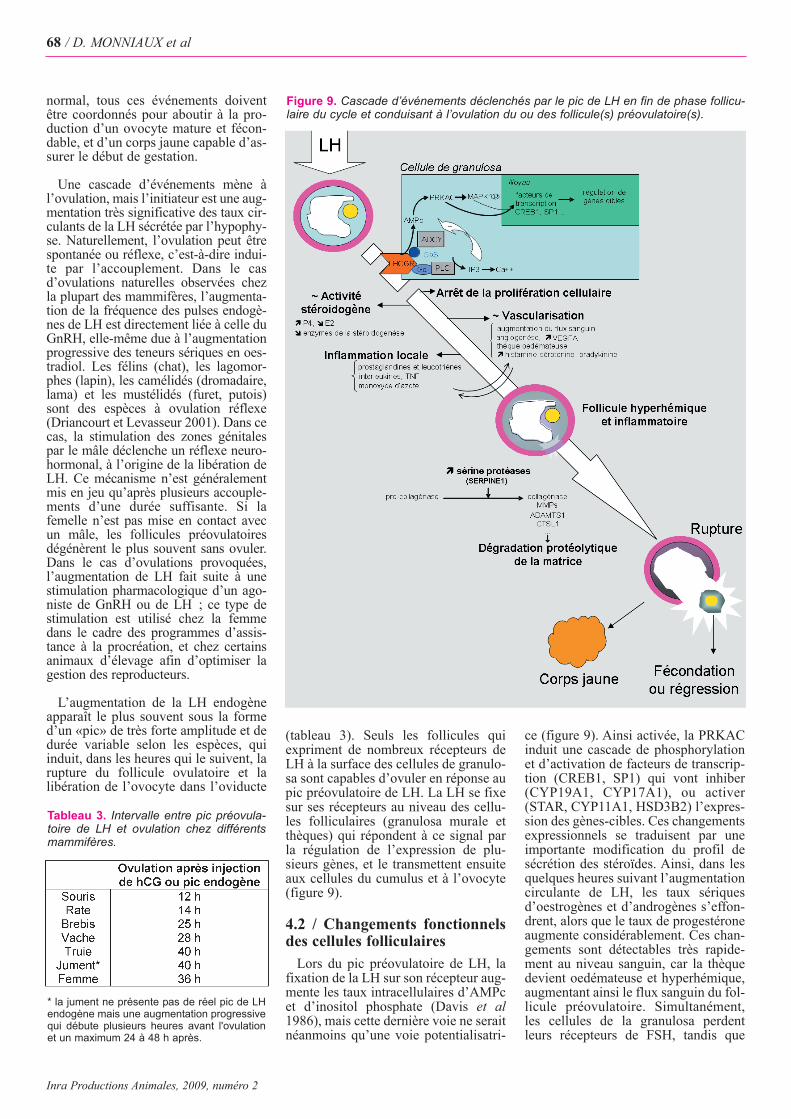
normal, tous ces événements doiventêtre coordonnés pour aboutir à la pro-duction d’un ovocyte mature et fécon-dable, et d’un corps jaune capable d’as-surer le début de gestation.
Une cascade d’événements mène àl’ovulation, mais l’initiateur est une aug-mentation très significative des taux cir-culants de la LH sécrétée par l’hypophy-se. Naturellement, l’ovulation peut êtrespontanée ou réflexe, c’est-à-dire indui-te par l’accouplement. Dans le cas d’ovulations naturelles observées chezla plupart des mammifères, l’augmenta-tion de la fréquence des pulses endogè-nes de LH est directement liée à celle duGnRH, elle-même due à l’augmentationprogressive des teneurs sériques en oes-tradiol. Les félins (chat), les lagomor-phes (lapin), les camélidés (dromadaire,lama) et les mustélidés (furet, putois)sont des espèces à ovulation réflexe(Driancourt et Levasseur 2001). Dans cecas, la stimulation des zones génitalespar le mâle déclenche un réflexe neuro-hormonal, à l’origine de la libération deLH. Ce mécanisme n’est généralementmis en jeu qu’après plusieurs accouple-ments d’une durée suffisante. Si lafemelle n’est pas mise en contact avecun mâle, les follicules préovulatoiresdégénèrent le plus souvent sans ovuler.Dans le cas d’ovulations provoquées,l’augmentation de LH fait suite à unestimulation pharmacologique d’un ago-niste de GnRH ou de LH ; ce type destimulation est utilisé chez la femmedans le cadre des programmes d’assis-tance à la procréation, et chez certainsanimaux d’élevage afin d’optimiser lagestion des reproducteurs.
L’augmentation de la LH endogèneapparaît le plus souvent sous la formed’un «pic» de très forte amplitude et dedurée variable selon les espèces, quiinduit, dans les heures qui le suivent, larupture du follicule ovulatoire et lalibération de l’ovocyte dans l’oviducte
(tableau 3). Seuls les follicules quiexpriment de nombreux récepteurs deLH à la surface des cellules de granulo-sa sont capables d’ovuler en réponse aupic préovulatoire de LH. La LH se fixesur ses récepteurs au niveau des cellu-les folliculaires (granulosa murale etthèques) qui répondent à ce signal parla régulation de l’expression de plu-sieurs gènes, et le transmettent ensuiteaux cellules du cumulus et à l’ovocyte(figure 9).
4.2 / Changements fonctionnelsdes cellules folliculaires
Lors du pic préovulatoire de LH, lafixation de la LH sur son récepteur aug-mente les taux intracellulaires d’AMPcet d’inositol phosphate (Davis et al1986), mais cette dernière voie ne seraitnéanmoins qu’une voie potentialisatri-
ce (figure 9). Ainsi activée, la PRKACinduit une cascade de phosphorylationet d’activation de facteurs de transcrip-tion (CREB1, SP1) qui vont inhiber(CYP19A1, CYP17A1), ou activer(STAR, CYP11A1, HSD3B2) l’expres-sion des gènes-cibles. Ces changementsexpressionnels se traduisent par uneimportante modification du profil desécrétion des stéroïdes. Ainsi, dans lesquelques heures suivant l’augmentationcirculante de LH, les taux sériquesd’oestrogènes et d’androgènes s’effon-drent, alors que le taux de progestéroneaugmente considérablement. Ces chan-gements sont détectables très rapide-ment au niveau sanguin, car la thèquedevient oedémateuse et hyperhémique,augmentant ainsi le flux sanguin du fol-licule préovulatoire. Simultanément,les cellules de la granulosa perdentleurs récepteurs de FSH, tandis que
68 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Tableau 3. Intervalle entre pic préovula-toire de LH et ovulation chez différentsmammifères.
* la jument ne présente pas de réel pic de LHendogène mais une augmentation progressivequi débute plusieurs heures avant l'ovulationet un maximum 24 à 48 h après.
Figure 9. Cascade d’événements déclenchés par le pic de LH en fin de phase follicu-laire du cycle et conduisant à l’ovulation du ou des follicule(s) préovulatoire(s).

l’expression des récepteurs de LHdiminue de façon transitoire pour aug-menter à nouveau par la suite (figu-re 4). Chez le rat, cette «reprogramma-tion» des cellules folliculaires dureenviron 7 h (Richards et Hedin 1988).
4.3 / La rupture de la paroi folli-culaire
Le processus d’ovulation est associéà une réaction de type inflammatoire(Espey 1980). La synthèse ovariennede cytokines inflammatoires (interleu-kines, TNF), de prostaglandines et decortisol (glucocorticoïde à action anti-inflammatoire) s’accentue dans le folli-cule préovulatoire au moment de l’ovu-lation. Le mécanisme d’action descytokines dans la maturation préovula-toire est probablement similaire à celuiobservé au cours d’une inflammation ;en particulier elles stimulent l’activitéd’enzymes protéolytiques et la produc-tion de prostaglandines et de monoxyded’azote (NO). De plus, elles sont capa-bles de moduler la stéroïdogenèse etinterviendraient aussi dans la matura-tion du complexe ovocyte-cumulus(Gérard et al 2004).
La rupture de la paroi folliculaire etl’expulsion de l’ovocyte dans l’oviduc-te nécessitent l’action d’enzymes pro-téolytiques dégradant la matrice extra-cellulaire. En réponse à la stimulationpar le pic préovulatoire de LH, la pro-duction locale d’activateur du plasmi-nogène, de plasmine et de collagénases(MMP1 et MMP2) augmente considé-rablement. L’activateur du plasminogè-ne transforme le plasminogène en plas-mine, qui elle-même activeraitcertaines collagénases (Beers et al1975). En parallèle, l’activité d’inhibi-teurs de sérines protéases (SERPINE1)et d’inhibiteurs des métalloprotéasesmatricielles (TIMP1 et TIMP2) aug-mente. L’expression de ces métallopro-téases et des TIMP est régulée par lesstéroïdes et les prostaglandines. Ainsi,l’expression concomitante de protéaseset d’anti-protéases dans le follicule pré-ovulatoire permet de moduler le site etle degré de dégradation du mur follicu-laire au moment de sa rupture (Curry etSmith 2006).
4.4 / La maturation du complexeovocyte-cumulus
En réponse au pic préovulatoire deLH, l’ovocyte entre en phase de matu-ration. La reprise de la méiose s’ac-compagne de modifications structura-les et biochimiques au sein ducytoplasme, et est associée à une diffé-
renciation des cellules du cumulus. Lasynchronisation de l’ensemble de cesévénements sous-tend le succès nonseulement de la maturation ovocytaireelle-même, mais aussi des étapes ulté-rieures de la fécondation et de l’initia-tion du développement embryonnaire.
Au cours de la folliculogenèse,depuis la formation du follicule primor-dial jusqu'à la phase de croissance fina-le du follicule dominant, l'ovocyte estbloqué en prophase de première divi-sion méiotique et ce blocage est main-tenu essentiellement par un niveauélevé d’AMPc intra-ovocytaire. Dansle follicule, ce facteur inhibiteur tran-site des cellules de granulosa muralesau cumulus et à l’ovocyte par des jonc-tions communicantes. In vitro, la repri-se de méiose intervient spontanémentquand le complexe ovocyte-cumulusest sorti du follicule et résulterait del’arrêt de l’apport de ce facteur inhibi-teur. In vivo, le pic préovulatoire de LHinduit la phosphorylation de laconnexine 43 (GJA1) qui constitue lesjonctions communicantes, bloquantainsi le passage d’AMPc par interrup-tion du couplage métabolique entre lescellules folliculaires et l’ovocyte(Norris et al 2008).
La rupture de la vésicule germinale(Germinal Vesicle BreakDown ouGVBD) est le premier signe visible dematuration. Elle se produit dans lesheures qui suivent le pic préovulatoirede LH et elle commence par un plisse-ment de l'enveloppe nucléaire. Lespores nucléaires disparaissent puis l'en-veloppe se fragmente avant d’êtredégradée (en 3 h chez la souris, 6 hchez les bovins). Le nucléole disparaîtrapidement au contact du cytoplasme.Les chromosomes se condensent durantet après la GVBD et les chiasmas mig-rent vers l’extrémité des bras chromo-somiques. Le fuseau se forme depuisles MTOCs (microtubule organizingcenter, équivalent du centrosome dansl’ovocyte) et ses tubules s’ancrent surles chromosomes qui se distribuent surla plaque métaphasique de la premièredivision méiotique (métaphase I).L’anaphase et la télophase sont rapides,les chromosomes homologues se sépa-rent et migrent aux pôles du fuseau,dont l’un provoque une boursouflure dela membrane qui deviendra le premierglobule polaire. Après la télophase, leschromosomes de l’ovocyte se répartis-sent rapidement sur une plaque méta-phasique alors que se forme le secondfuseau méiotique (métaphase II)(Brunet et Maro 2005, Terret etWassmann 2008). Le globule polaire
contenant la moitié du complémentchromatinien est expulsé dans l’espacepérivitellin. C’est à ce stade qu’inter-vient l’ovulation chez la plupart desmammifères, à l’exception des canidéspour lesquels la rupture de la vésiculegerminale a lieu lors du transit de l’ovocyte dans les voies génitales.L'ovocyte reste bloqué au stade demétaphase II par un facteur cytosta-tique (cytostatic factor ou CSF) jusqu’àsa fécondation ou son activation par-thénogénétique.
La reprise de méiose correspond à unpassage entre le stade G2 et le stade Mdu cycle cellulaire, et à ce titre reposesur l’activation du M-phase PromotingFactor ou MPF, hétérodimère composéd’une sous-unité catalytique, CDC2, etd’une sous-unité régulatrice, la cyclineCCNB1. La diversité des ciblesphosphorylées par le MPF (lamine B,histone H1 et certaines protéines asso-ciées aux microfilaments) peut expli-quer comment ce complexe coordonneles différents événements nécessaires àun bon déroulement de la maturation.Le niveau d’activité du MPF chute àl’anaphase I, la phosphorylation de sescibles étant alors maintenue par d’au-tres kinases (en particulier MAPK1).La progression de la méiose s’accom-pagne encore de l'activation de la kina-se AKT1 et de la déphosphorylation dela kinase PRKAC (Mermillod etMarchal 1999, Dekel 2005).
L’activité transcriptionnelle, déjàminimale depuis la fin de la croissanceovocytaire, s’arrête totalement dès lacondensation des chromosomes endébut de maturation. L’expressiongénique repose alors essentiellementsur la régulation post-transcriptionnelledes ARN présents dans le cytoplasme.La maturation s’accompagne d’unedégradation modérée des ARN, etd’une déadénylation sélective affectantles messagers à des degrés variables.Certains messagers, comportant dansleur région 3’ non traduite des signauxspécifiques, subissent au contraire unepolyadénylation cytoplasmique cou-plée à leur recrutement pour la traduc-tion.
Pendant cette période, l’élargisse-ment de l’espace périvitellin s’accom-pagne d’une réorganisation des compo-sants cytoplasmiques de l’ovocyte(Mermillod et Marchal 1999). Les gra-nules corticaux, qui avaient une locali-sation cytoplasmique sous-corticalediffuse dans l'ovocyte immature, mi-grent vers la zone corticale en s'asso-ciant au cytosquelette (filaments d'ac-
Développement folliculaire ovarien et ovulation chez les mammifères / 69
Inra Productions Animales, 2009, numéro 2

tine). Les mitochondries, qui consti-tuent l’organelle le plus abondant dansl’ovocyte, forment des agrégats avec leréticulum endoplasmique dans la régionpérinucléaire, ces déplacements étantsous la dépendance des microtubules. Laréserve en lipides et la concentration englutathion augmentent, indicateurs dechangement métaboliques. La reprise deméiose est corrélée à une augmentationde la concentration de calcium dans lecytosol. Le calcium est relargué destocks intracellulaires, et pourrait aussipénétrer par les canaux ioniques de lamembrane plasmique. Parallèlement,des changements de la machinerie designalisation calcique s’opèrent (Tosti2006).
Le pic préovulatoire de LH induitaussi l'expansion du cumulus quientoure l’ovocyte. Cette expansionrésulte de la sécrétion d’une matriceextracellulaire riche en acide hyaluro-nique, elle est stimulée par FSH et cer-tains facteurs de croissance (EGF,IGF1). Des facteurs ovocytaires inter-viennent également dans ce phénomè-ne, notamment GDF9. Le cumulusexpansé constitue un micro-environne-ment protecteur pour l'ovocyte et assu-re la captation du complexe par lepavillon de la trompe suite à l’ovula-tion. Il joue également un rôle dans lacapacitation des spermatozoïdes avantla fécondation (Tanghe et al 2002).
5 / Régulations par les fac-teurs externes
5.1 / Le système hypothalamo-hypophysaire : un intégrateurdes signaux endocriniens etenvironnementaux
La maturation folliculaire et l’ovula-tion sont contrôlées par le systèmehypothalamo-hypophysaire qui intègreles informations de différents facteursendogènes (signaux hormonaux etnutritionnels) et exogènes (photopério-de, température, stress…) (figure 10).
Le système hypothalamo-hypophy-saire est composé de l’hypothalamus,partie du cerveau située à la base dutroisième ventricule, et de l’hypophyse,placée sous l’hypothalamus et rattachéeà celui-ci par la tige pituitaire.L’hypophyse et plus précisément l’adé-nohypophyse exprime et sécréte lesgonadotropines, LH et FSH. La sécré-tion de ces hormones est sous le contrô-le du GnRH, une neurohormone hypo-thalamique secrétée directement dans
les capillaires sanguins de la tige pitui-taire. Le GnRH est un décapeptide,synthétisé par les neurones de l’hypo-thalamus médio-basal et de l’aireoptique, qui stimule la synthèse et lasécrétion de la LH et de la FSH en sefixant à des récepteurs situés à la surfa-ce des cellules gonadotropes. Les sté-roïdes, en particulier l’oestradiol d’ori-gine ovarienne, inhibent la sécrétionpulsatile de GnRH par un rétrocontrôlenégatif. Cependant, en fin de crois-sance folliculaire terminale et au-delà d’une concentration-seuil (5 à10 pg/mL chez la brebis) l’oestradiolexerce une action positive sur le systè-me hypothalamo-hypophysaire. Il pro-voque à la fois une augmentation de lafréquence des pulses de GnRH, puisune libération massive de ce dernier, etune augmentation importante de la sen-sibilité hypophysaire. L’association deces deux effets conduit à une déchargemassive de LH qui déclenchera l’ovu-lation des follicules préovulatoires pré-sents dans l’ovaire. Le peptide KISS1(ou kisspeptine), ligand d’un récepteurappelé KISS1R (ou GPR54), est unacteur majeur dans les mécanismes derétrocontrôle des stéroïdes sur la libéra-tion du GnRH et semble impliqué danstoutes les étapes de transition de la viereproductive (Caraty et Franceschini2008). De plus en plus de résultats indi-quent que cette molécule intervientégalement au niveau de l’hypophyse etdes gonades. Les premières manipula-
tions pharmacologiques de l’axe gona-dotrope par KISS1 sont très prometteu-ses. En injections intraveineuses, ellestimule fortement la sécrétion desgonadotropines chez l’animal commechez l’homme. Chez la brebis, elle per-met de synchroniser l’ovulation àl’heure près, en saison de reproduction,ou d’induire une cyclicité suivie d’uneovulation chez des animaux en état derepos sexuel (Caraty et al 2007).
Les différentes aires de l’hypothala-mus (ventromédian, paraventriculaire,noyau arqué) qui gouvernent le com-portement sexuel et la sécrétion duGnRH sont également au carrefour denombreux systèmes de contrôle del’homéostasie, tels que le contrôle dupoids corporel, du comportement ali-mentaire et de la thermogenèse. Ellessont donc capables d’intégrer toute per-turbation du bilan énergétique auniveau périphérique et de réagir enmodifiant un ensemble de fonctionsphysiologiques et de comportements.Chez la plupart des espèces étudiées,des perturbations de la ration alimentai-re entraînant un déséquilibre du bilanénergétique ont des conséquencesdirectes au niveau de l’axe hypothala-mo-hypophysaire. Par exemple, unesous-alimentation plus ou moinsimportante aboutit à une profonde déré-gulation du rétrocontrôle exercé parl’oestradiol sur la sécrétion de GnRH(Diskin et al 2003). Chez la vache et
70 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Figure 10. Implications des facteurs endocriniens, nutritionnels et saisonniers dans larégulation des fonctions ovariennes.

l’agnelle, cette dérégulation passe enpartie par un renforcement de larétroaction négative exercée par l’oes-tradiol sur la sécrétion de GnRH(Foster et Olster 1985). Cette dérégula-tion conduit à une diminution de lasécrétion de la LH et, à terme, à uneanovulation.
Chez les espèces à reproduction sai-sonnée (ovins, caprins, équins…), ladurée du jour (ou photopériode) est unfacteur supplémentaire qui modulel’activité de l’axe hypothalamo-hypo-physaire et est responsable de l’alter-nance entre une saison sexuelle et unesaison de repos sexuel, caractérisée parl’absence d’ovulation (Malpaux 2006).Cette action de la photopériode estrelayée par la mélatonine, une hormoneproduite exclusivement pendant la nuitpar la glande pinéale. La durée desécrétion de la mélatonine est propor-tionnelle à celle de la nuit et constitueun signal endocrinien permettant auxtissus cibles de «distinguer» les jourslongs d’été des jours courts d’hiver. Lamélatonine agit au niveau hypothala-mique pour contrôler la libération deGnRH, ce qui par voie de conséquencemodifie la secrétion des gonadotropi-nes. Cette action n’est toutefois pasdirecte sur les neurones à GnRH maiselle implique des interneurones multi-ples (dopamine, sérotonine, kisspepti-ne...) qui ne sont que partiellementconnus. L’action de la mélatonine passeégalement par une modulation de larétroaction négative de l’œstradiol avecun renforcement de cette dernière pen-dant les durées du jour inhibitrices(Lehman et al 2002). Enfin, d’autresfacteurs externes (alimentation, tempé-rature, facteurs sociaux, stress…)modulent les effets de la photopériodepour déterminer les caractéristiquesfines de la saison sexuelle. Ainsi, unerestriction d’apport alimentaire peutraccourcir la saison sexuelle sans per-turber la cyclicité pendant cette périodede l’année.
5.2 / Régulation nutritionnellede la fonction ovarienne
Des modifications quantitatives ouqualitatives de l’apport alimentaire,relayées par des changements designaux hormonaux (insuline, IGF, lep-tine) et par des variations importantesdes flux métaboliques (acides gras ouglucose), modulent l’activité de l’axehypothalamo-hypophyso-ovarien.
a) L’insuline et l’IGF1L’insuline, synthétisée exclusivement
par le pancréas, et l’IGF1, majoritaire-
ment produit par le foie en réponse àl’hormone de croissance (GH), jouentun rôle pivot dans la régulation gonado-trope et ovarienne. En effet, il existeune assez bonne corrélation in vivoentre les modifications des taux d’IGF1et d’insuline observées au cours de lacroissance, de la puberté ou lors demodifications du bilan énergétique, etla fréquence des pulses de LH. D’autrepart, les composants du système insuli-ne/IGF (ligands, récepteurs, IGFBP)sont retrouvés aux trois niveaux réglantla fonction ovarienne (hypothalamus,hypophyse, ovaires) où ils peuvent par-ticiper au contrôle de la réponse auxchangements des signaux nutritionnels(Monget et Martin 1997).
Au niveau central, l’IGF1 régule laneurosécrétion de GnRH et de LH. Eneffet, chez le rat, des injections d’IGF1par voie intra-cérébrale sont capablesde stimuler la sécrétion de LH et d’avancer l’âge à la puberté des femel-les. L’insuline modifie l’expressiongénique ou la libération de diversessubstances hypothalamiques interve-nant dans la régulation gonadotrope(IGF2, neuropeptide Y). Des études cli-niques suggèrent que l’hyperinsuliné-mie favorise l’amplitude des pulses desécrétion de LH chez les femmes obè-ses présentant un syndrome d’ovairespolykystiques.
Il existerait également des effetsdirects de l’insuline et/ou des IGF auniveau ovarien. Chez plusieurs espècesd’intérêt agronomique (brebis, truie etvache), une augmentation de la rationalimentaire pendant les derniers joursde la phase lutéale est capable d’aug-menter la vitesse de croissance, la tailleet le nombre de follicules dominantssans altérer les niveaux circulants deFSH et de LH (Downing et Scaramuzzi1991). Ces effets pourraient en partieêtre dus à l’insuline et/ou l’IGF1 quiaugmenteraient la sensibilité des folli-cules à l’action de la FSH au momentoù ces derniers rentrent dans leur phaseterminale de croissance (Mazerbourg etal 2003). Plusieurs mécanismes d’ac-tion de l’insuline/IGF1 au niveau ova-rien sont possibles, en particulier uneactivation directe de leurs récepteursrespectifs, ou une augmentation de labiodisponibilité de l’IGF1 par réduc-tion des taux intra-folliculairesd’IGFBP1.
b) La leptineLa leptine (LEP) est une hormone
principalement exprimée et sécrétéepar le tissu adipeux. L’injection de lep-tine à des souris femelles ob/ob, qui ont
un déficit congénital en leptine et sontinfertiles, augmente le taux de gonado-tropines circulantes, induit un dévelop-pement folliculaire ovarien normal etrestaure la fertilité (Chehab et al 1996).Chez plusieurs espèces animales, ilsemble que la leptine constitue unsignal nécessaire au système nerveuxcentral pour déclencher, en fonction del’état des réserves adipeuses, la pubertéet les premières ovulations chez les jeu-nes. In vivo, les teneurs en leptine dansle sérum augmentent jusqu’à l’âge de lapuberté (Zieba et al 2005).
Le mécanisme d’action de la leptinesur la fonction de reproduction n’estpas encore parfaitement connu. Lesrécepteurs de la leptine sont présentsdans de nombreux tissus y comprisdans l’hypothalamus, l’hypophyse etl’ovaire, indiquant que la leptine peutagir à tous les niveaux de l’axe hypo-thalamo-hypophyso-ovarien. In vitro,la leptine induit une libération de la LHà partir d’explants de complexes hypo-thalamo-infundibulaires et de culturesde cellules hypophysaires (Zieba et al2005), et exerce un effet direct sur lastéroïdogenèse des cellules de la granu-losa et de thèque (Spicer 2001).Comme la leptine, d’autres hormonesproduites par le tissu adipeux commel’adiponectine (ADIPOQ) ou la résisti-ne (RETN) pourraient jouer des rôlesimportants dans la régulation de l’axehypothalamo-hypophyso-ovarien(Mitchell et al 2005).
c) Le glucose et les acides grasLes nutriments, en particulier le glu-
cose et les acides gras, peuvent aussimoduler les fonctions ovariennes parune action directe et/ou indirecte.L’administration de 2-deoxy-D-gluco-se, un agent bloquant de la glycolyse,induit une chute brutale de la sécrétionde LH et bloque l’ovulation et la forma-tion du corps jaune (McClure et al1978, Funston et al 1995). De même,l’administration de méthyl-palmoxyra-te, un agent empêchant l’oxydation desacides gras, provoque une perturbationimportante de l’ovulation (Schneider etZhou 1999) et une lésion chirurgicalede l’area postrema, zone du cerveauinnervée par des afférences vagalesprovenant des viscères, lève totalementles effets inhibiteurs de la sous-alimen-tation et des agents bloquant le métabo-lisme du glucose et des acides gras, surla sécrétion de GnRH et sur le compor-tement sexuel (Wade et al 1996).
Les acides gras à longue chaîne pour-raient moduler la croissance folliculairepar une action directe au niveau ova-
Développement folliculaire ovarien et ovulation chez les mammifères / 71
Inra Productions Animales, 2009, numéro 2

rien. En effet, un récepteur de ces aci-des gras, le récepteur PPARG, est forte-ment exprimé dans les cellules de lagranulosa de brebis et de rongeurs, etun ligand synthétique de ce récepteurest capable de moduler in vitro la proli-fération et la stéroïdogenèse de ces cel-lules (Froment et al 2006). Ces résul-tats suggèrent qu’un métabolisme deslipides spécifique de l’ovaire pourraitjouer un rôle important dans les interfa-ces métabolisme/reproduction.
Conclusion
L’issue du développement folliculai-re, c’est-à-dire le déclenchement d’uneou plusieurs ovulations, est caractéris-tique de chaque espèce de mammifèreset de chaque race. Ce processus dedéveloppement est finement régulé àchacune de ses étapes, et la questionque l’on peut se poser est de savoircomment l’ensemble est orchestré.Chez la souris, il a été proposé que l’ovocyte pourrait piloter tout le déve-loppement folliculaire jusqu’à l’ovula-tion (Eppig et al 2002). Néanmoins, ilest difficile d’imaginer que l’ovocytepuisse orchestrer à lui seul l’ensembledu développement folliculaire chez desmammifères dont le diamètre du folli-cule préovulatoire dépasse plusieursmillimètres ou plusieurs centimètres.
Plus vraisemblablement, chaque étapedu développement folliculaire sembledépendre, pour son bon déroulement,de la présence de facteurs-clés. En l’état actuel des connaissances, on peutproposer que ces facteurs sont les BMP,le KITLG et l’AMH pour le démarragede croissance folliculaire, l’IGF et l’in-suline pour la transition entre dévelop-pement folliculaire basal et terminal, laFSH pour le début du développementfolliculaire terminal et la sélection dufollicule préovulatoire, et la LH pour ledéveloppement final de ce follicule, lamaturation ovocytaire et l’ovulation.Le bon pilote doit être présent à chaqueétape du développement folliculaire.Ainsi, pour un follicule ovarien qui abien démarré sa croissance mais quiarrive en début de son développementterminal juste après une ovulation chezla femme, le relais par FSH ne sera pasassuré. Ce passage de relais est égale-ment susceptible d’être modulé par lesfacteurs environnementaux, en particu-lier photopériodiques et nutritionnels. Iln’y a donc pas un seul, mais plusieurspilotes successifs et c’est le passage derelais de l’un à l’autre qui est détermi-nant pour le bon déroulement de la fol-liculogenèse.
Depuis la formation de la réserve defollicules primordiaux jusqu’à l’ovula-tion, l’ovaire est le siège d’un immensegaspillage de cellules germinales. Chez
les mammifères, ce gaspillage pourraitn’être qu’un résidu évolutif du méca-nisme de surproduction de gamètes quiexiste chez les poissons, et qui s’avèreindispensable pour la survie des espè-ces à fécondation externe. L’intérêt bio-logique éventuel de ce gaspillage pourles mammifères pourrait être in fine depermettre l’ovulation des seuls«meilleurs» ovocytes, aptes à êtrefécondés et à donner naissance à desdescendants viables. L’existence d’uneréserve folliculaire surdimensionnéeconfère surtout une grande plasticité auprocessus de développement folliculai-re, plasticité que l’on peut mettre enévidence expérimentalement. En effet,si l’on détruit par cautérisation les folli-cules à antrum présents sur un ovaire,ou si l’on enlève l’un des ovaires d’unanimal, dans les deux cas, le tissu ova-rien restant reconstitue rapidement unefolliculogenèse normale jusqu’à l’ovu-lation. De même, à partir d’un fragmentde cortex ovarien dépourvu de follicu-les à antrum, un démarrage de croissan-ce folliculaire s’initie «spontanément»et très rapidement in vitro, permettantde régénérer une folliculogenèse nor-male. Les mécanismes à l’origine deces phénomènes de compensation res-tent à élucider, et en particulier la natu-re et l’origine du signal de démarragede croissance des follicules primor-diaux sont actuellement inconnus.
72 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
Références
Baltus A.E., Menke D.B., Hu Y.C., GoodheartM.L., Carpenter A.E., de Rooij D.G., Page D.C.,2006. In germ cells of mouse embryonic ovaries,the decision to enter meiosis precedes premeio-tic DNA replication. Nat. Genet., 38, 1430-1434.
Beers W.H., Strickland S., Reich E., 1975.Ovarian plasminogen activator: relationship to ovu-lation and hormonal regulation. Cell, 6, 387-394.
Brunet S., Maro B., 2005. Cytoskeleton andcell cycle control during meiotic maturation ofthe mouse oocyte: integrating time and space.Reproduction, 130, 801-811.
Carabatsos M.J., Sellitto C., GoodenoughD.A., Albertini D.F., 2000. Oocyte-granulosacell heterologous gap junctions are required forthe coordination of nuclear and cytoplasmic mei-otic competence. Dev. Biol., 226, 167-179.
Caraty A., Franceschini I., 2008. Basic aspectsof the control of GnRH and LH secretions bykisspeptin: potential applications for better con-trol of fertility in females. Reprod. Domest.Anim., 43, Suppl 2, 172-178.
Caraty A., Smith J.T., Lomet D., Ben Said S.,Morrissey A., Cognie J., Doughton B., Baril G.,Briant C., Clarke I.J., 2007. Kisspeptin synchro-nizes preovulatory surges in cyclical ewes andcauses ovulation in seasonally acyclic ewes.Endocrinology, 148, 5258-5267.
Castrillon D.H., Miao L., Kollipara R., HornerJ.W., DePinho R.A., 2003. Suppression of ova-rian follicle activation in mice by the transcrip-tion factor Foxo3a. Science, 301, 215-218.
Chehab F.F., Lim M.E., Lu R., 1996.Correction of the sterility defect in homozygousobese female mice by treatment with the humanrecombinant leptin. Nat. Genet., 12, 318-320.
Choi Y., Rajkovic A., 2006. Genetics of earlymammalian folliculogenesis. Cell. Mol. LifeSci., 63, 579-590.
Curry T.E., Jr., Smith M.F., 2006. Impact ofextracellular matrix remodeling on ovulation andthe folliculo-luteal transition. Semin. Reprod.Med., 24, 228-241.
Davis J.S., Weakland L.L., West L.A., FareseR.V., 1986. Luteinizing hormone stimulates the for-mation of inositol trisphosphate and cyclic AMP inrat granulosa cells. Evidence for phospholipase Cgenerated second messengers in the action ofluteinizing hormone. Biochem. J., 238, 597-604.
Dekel N., 2005. Cellular, biochemical andmolecular mechanisms regulating oocyte matu-ration. Mol. Cell. Endocrinol., 234, 19-25.
Diskin M.G., Mackey D.R., Roche J.F.,Sreenan J.M., 2003. Effects of nutrition andmetabolic status on circulating hormones and
ovarian follicle development in cattle. Anim.Reprod. Sci., 78, 345-370.
Downing J.A., Scaramuzzi R.J., 1991.Nutrient effects on ovulation rate, ovarian func-tion and the secretion of gonadotrophic andmetabolic hormones in sheep. J. Reprod. Fertil.Suppl., 43, 209-227.
Driancourt M.A., Levasseur M.C., 2001.Cycles estriens et cycles menstruels. In: Lareproduction chez les mammifères et l'homme.Thibault C., Levasseur M.C., (Eds), Ellipses,Paris, France, 680-698.
Driancourt M.A., Gougeon A., Monniaux D.,Royere D., Thibault C., 2001. Folliculogenèse etovulation. In: La reproduction chez les mammi-fères et l'homme. Thibault C., Levasseur M.C.,(Eds), Ellipses, Paris, France, 316-347.
Durlinger A.L., Kramer P., Karels B., de JongF.H., Uilenbroek J.T., Grootegoed J.A.,Themmen A.P., 1999. Control of primordial fol-licle recruitment by anti-Mullerian hormone inthe mouse ovary. Endocrinology, 140, 5789-5796.
Echenim N., Monniaux D., Sorine M.,Clement F., 2005. Multi-scale modeling of thefollicle selection process in the ovary. Math.Biosci., 198, 57-79.

Eppig J.J., Wigglesworth K., Pendola F.L.,2002. The mammalian oocyte orchestrates therate of ovarian follicular development. Proc.Natl. Acad. Sci. U. S. A., 99, 2890-2894.
Eppig J.J., Pendola F.L., Wigglesworth K.,Pendola J.K., 2005. Mouse oocytes regulatemetabolic cooperativity between granulosa cellsand oocytes: amino acid transport. Biol. Reprod.,73, 351-357.
Espey L.L., 1980. Ovulation as an inflamma-tory reaction-a hypothesis. Biol. Reprod., 22, 73-106.
Fabre S., Pierre A., Mulsant P., Bodin L., DiPasquale E., Persani L., Monget P., MonniauxD., 2006. Regulation of ovulation rate in mam-mals: contribution of sheep genetic models.Reprod. Biol. Endocrinol., 4, 20.
Foster D.L., Olster D.H., 1985. Effect ofrestricted nutrition on puberty in the lamb: pat-terns of tonic luteinizing hormone (LH) secre-tion and competency of the LH surge system.Endocrinology, 116, 375-381.
Froment P., Gizard F., Defever D., Staels B.,Dupont J., Monget P., 2006. Peroxisome proli-ferator-activated receptors in reproductive tis-sues: from gametogenesis to parturition. J.Endocrinol., 189, 199-209.
Funston R.N., Roberts A.J., Hixon D.L.,Hallford D.M., Sanson D.W., Moss G.E., 1995.Effect of acute glucose antagonism on hypophy-seal hormones and concentrations of insulin-likegrowth factor (IGF)-I and IGF-binding proteinsin serum, anterior pituitary, and hypothalamus ofewes. Biol. Reprod., 52, 1179-1186.
Galloway S.M., McNatty K.P., CambridgeL.M., Laitinen M.P., Juengel J.L., Jokiranta T.S.,McLaren R.J., Luiro K., Dodds K.G.,Montgomery G.W., Beattie A.E., Davis G.H.,Ritvos O., 2000. Mutations in an oocyte-derivedgrowth factor gene (BMP15) cause increasedovulation rate and infertility in a dosage-sensiti-ve manner. Nat. Genet., 25, 279-283.
Gérard N., Caillaud M., Martoriati A., GoudetG., Lalmanach A.C., 2004. The interleukin-1 sys-tem and female reproduction. J. Endocrinol.,180, 203-212.
Hanrahan J.P., Gregan S.M., Mulsant P.,Mullen M., Davis G.H., Powell R., GallowayS.M., 2004. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 areassociated with both increased ovulation rate andsterility in Cambridge and Belclare sheep (Ovisaries). Biol. Reprod., 70, 900-909.
Huang E.J., Manova K., Packer A.I., SanchezS., Bachvarova R.F., Besmer P., 1993. Themurine steel panda mutation affects kit ligandexpression and growth of early ovarian follicles.Dev. Biol., 157, 100-109.
Hyttel P., Viuff D., Fair T., Laurincik J.,Thomsen P.D., Callesen H., Vos P.L., HendriksenP.J., Dieleman S.J., Schellander K., BesenfelderU., Greve T., 2001. Ribosomal RNA geneexpression and chromosome aberrations inbovine oocytes and preimplantation embryos.Reproduction, 122, 21-30.
Juengel J.L., McNatty K.P., 2005. The role ofproteins of the transforming growth factor-betasuperfamily in the intraovarian regulation of fol-licular development. Hum. Reprod. Update, 11,143-160.
Klinger F.G., De Felici M., 2002. In vitrodevelopment of growing oocytes from fetalmouse oocytes: stage-specific regulation by stemcell factor and granulosa cells. Dev. Biol., 244,85-95.
Knight P.G., Glister C., 2006. TGF-beta super-family members and ovarian follicle develop-ment. Reproduction, 132, 191-206.
Kumar T.R., Wang Y., Lu N., Matzuk M.M.,1997. Follicle stimulating hormone is requiredfor ovarian follicle maturation but not male ferti-lity. Nat. Genet., 15, 201-204.
Lécureuil C., Kara E., Guillou F., MonniauxD., Crépieux P., 2007. Does FSH signaling havea gender? Med. Sci. (Paris), 23, 75-80.
Lehman M.N., Coolen L.M., Goodman R.L.,Viguie C., Billings H.J., Karsch F.J., 2002.Seasonal plasticity in the brain: the use of largeanimal models for neuroanatomical research.Reproduction, Suppl., 59, 149-165.
Lequarre A.S., Vigneron C., Ribaucour F.,Holm P., Donnay I., Dalbies-Tran R., CallesenH., Mermillod P., 2005. Influence of antral folli-cle size on oocyte characteristics and embryodevelopment in the bovine. Theriogenology, 63,841-859.
Malpaux B., 2006. Seasonal regulation ofreproduction in mammals. In: Knobil and Neill'sphysiology of reproduction. Neill J.D., (Ed),Elsevier, UK, 2231-2281.
Matzuk M.M., Burns K.H., Viveiros M.M.,Eppig J.J., 2002. Intercellular communication inthe mammalian ovary: oocytes carry the conver-sation. Science, 296, 2178-2180.
Mauléon P., 1969. Oogenesis and folliculoge-nesis. In: Reprod. Domest. Anim., Cole H.H.,Cupps P.T., (Ed), Academic Press, New York,187-215.
Mazerbourg S., Bondy C.A., Zhou J., MongetP., 2003. The insulin-like growth factor system:a key determinant role in the growth and selec-tion of ovarian follicles? a comparative speciesstudy. Reprod. Domest. Anim., 38, 247-258.
McClure T.J., Nancarrow C.D., RadfordH.M., 1978. The effect of 2-deoxy-D-glucose onovarian function of cattle. Aust. J. Biol. Sci., 31,183-186.
McGee E.A., Hsueh A.J., 2000. Initial andcyclic recruitment of ovarian follicles. EndocrineRev., 21, 200-214.
Mermillod P., Marchal R., 1999. Mammalianoocyte maturation. Med. Sci. (Paris), 15, 148-156.
Mermillod P., Dalbiès-Tran R., Uzbekova S.,Thélie A., Traverso J.M., Perreau C., Papillier P.,Monget P., 2008. Factors affecting oocyte quali-ty: who is driving the follicle? Reprod. Domest.Anim., 43, Suppl 2, 393-400.
Mitchell M., Armstrong D.T., Robker R.L.,Norman R.J., 2005. Adipokines: implications forfemale fertility and obesity. Reproduction, 130,583-597.
Monget P., Martin G.B., 1997. Involvement ofinsulin-like growth factors in the interactionsbetween nutrition and reproduction in femalemammals. Hum. Reprod., 12, Suppl 1, 33-52.
Monniaux D., Huet C., Besnard N., ClémentF., Bosc M., Pisselet C., Monget P., Mariana J.C.,1997. Follicular growth and ovarian dynamics inmammals. J. Reprod. Fertil., Suppl., 51, 3-23.
Monniaux D., Mandon-Pepin B., Monget P.,1999. Follicular atresia, a programmed wastage.Med. Sci. (Paris), 15, 157-166.
Morita Y., Tilly J.L., 1999. Oocyte apoptosis:like sand through an hourglass. Dev. Biol., 213,1-17.
Mourot M., Dufort I., Gravel C., Algriany O.,Dieleman S., Sirard M.A., 2006. The influenceof follicle size, FSH-enriched maturation me-dium, and early cleavage on bovine oocytematernal mRNA levels. Mol. Reprod. Dev., 73,1367-1379.
Mulsant P., Lecerf F., Fabre S., Schibler L.,Monget P., Lanneluc I., Pisselet C., Riquet J.,Monniaux D., Callebaut I., Cribiu E., ThimonierJ., Teyssier J., Bodin L., Cognie Y., Chitour N.,Elsen J.M., 2001. Mutation in bone morpho-genetic protein receptor-IB is associated withincreased ovulation rate in Booroola Merinoewes. Proc. Natl. Acad. Sci. U.S.A., 98, 5104-5109.
Nilsson E.E., Skinner M.K., 2003. Bone mor-phogenetic protein-4 acts as an ovarian folliclesurvival factor and promotes primordial follicledevelopment. Biol. Reprod., 69, 1265-1272.
Norris R.P., Freudzon M., Mehlmann L.M.,Cowan A.E., Simon A.M., Paul D.L., LampeP.D., Jaffe L.A., 2008. Luteinizing hormonecauses MAP kinase-dependent phosphorylationand closure of connexin 43 gap junctions inmouse ovarian follicles: one of two paths to mei-otic resumption. Development, 135, 3229-3238.
Pangas S.A., Choi Y., Ballow D.J., Zhao Y.,Westphal H., Matzuk M.M., Rajkovic A., 2006.Oogenesis requires germ cell-specific transcrip-tional regulators Sohlh1 and Lhx8. Proc. Natl.Acad. Sci. U. S. A., 103, 8090-8095.
Reddy P., Liu L., Adhikari D., Jagarlamudi K.,Rajareddy S., Shen Y., Du C., Tang W.,Hamalainen T., Peng S.L., Lan Z.J., Cooney A.J.,Huhtaniemi I., Liu K., 2008. Oocyte-specific dele-tion of Pten causes premature activation of the pri-mordial follicle pool. Science, 319, 611-613.
Richards J.S., Hedin L., 1988. Molecularaspects of hormone action in ovarian folliculardevelopment, ovulation, and luteinization. Ann.Rev. Physiol., 50, 441-463.
Schneider J.E., Zhou D., 1999. Interactiveeffects of central leptin and peripheral fuel oxi-dation on estrous cyclicity. Am. J. Physiol., 277,R1020-1024.
Shimasaki S., Moore R.K., Otsuka F.,Erickson G.F., 2004. The bone morphogeneticprotein system in mammalian reproduction.Endocrine Reviews, 25, 72-101.
Skinner M.K., 2005. Regulation of primor-dial follicle assembly and development. Hum.Reprod. Update, 11, 461-471.
Soyal S.M., Amleh A., Dean J., 2000.FIGalpha, a germ cell-specific transcription fac-tor required for ovarian follicle formation.Development, 127, 4645-4654.
Spicer L.J., 2001. Leptin: a possible metabolicsignal affecting reproduction. Domest. Anim.Endocrinol., 21, 251-270.
Tanghe S., Van Soom A., Nauwynck H.,Coryn M., de Kruif A., 2002. Minireview:Functions of the cumulus oophorus duringoocyte maturation, ovulation, and fertilization.Mol. Reprod. Dev., 61, 414-424.
Terret M.E., Wassmann K., 2008. Meioticweakness: the first division. Med. Sci. (Paris),24, 197-203.
Tosti E., 2006. Calcium ion currents media-ting oocyte maturation events. Reprod. Biol.Endocrinol., 4, 26.
Visser J.A., Themmen A.P., 2005. Anti-Mullerian hormone and folliculogenesis. Mol.Cell. Endocrinol., 234, 81-86.
Développement folliculaire ovarien et ovulation chez les mammifères / 73
Inra Productions Animales, 2009, numéro 2

Wade G.N., Schneider J.E., Li H.Y., 1996.Control of fertility by metabolic cues. Am. J.Physiol., 270, E1-E19.
Zheng P., Dean J., 2007. Oocyte-specificgenes affect folliculogenesis, fertilization, andearly development. Semin. Reprod. Med., 25,243-251.
Zieba D.A., Amstalden M., Williams G.L.,2005. Regulatory roles of leptin in reproductionand metabolism: a comparative review. Domest.Anim. Endocrinol., 29, 166-185.
74 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
ANNEXES
Annexe. Abréviations utilisées pour les protéines et les gènes cités dans l’article.
Symbole Description Alias
ADAMTS1 ADAM metallopeptidase with thrombospondin type 1 motifADCY adenylate cyclaseADIPOQ adiponectin, C1Q and collagen domain containing adiponectinAKT1 v-akt murine thymoma viral oncogene homolog 1 AKT, PKB, PRKBAMH anti-Mullerian hormone MIS ATM ataxia telangiectasia mutated ATD BAX BCL2-associated X proteinBCL2 B-cell CLL/lymphoma 2 Bcl-2 BCL2L1 BCL2-like 1 BCLX BMP15 bone morphogenetic protein 15 GDF9B BMP4 bone morphogenetic protein 4 BMP2BBMP7 bone morphogenetic protein 7 OP1BMPR1B bone morphogenetic protein receptor, type IB ALK6CASP2 caspase 2, apoptosis-related cysteine peptidase NEDD2CASP3 caspase 3, apoptosis-related cysteine peptidase apopainCCNB1 cyclin B1CDC2 cell division cycle 2, G1 to S and G2 to M CDK1CPEB1 cytoplasmic polyadenylation element binding protein 1 CPEBCREB1 cAMP responsive element binding protein 1 CREB CTSL1 cathepsin L1 CATLCYP11A1 cytochrome P450, family 11, subfamily A, polypeptide 1 P450SCCCYP17A1 cytochrome P450, family 17, subfamily A, polypeptide 1 P450C17CYP19A1 cytochrome P450, family 19, subfamily A, polypeptide 1 aromatase, P450AroDMC1 DMC1 dosage suppressor of mck1 homolog, meiosis-specific
homologous recombination (yeast) LIM15 EGF epidermal growth factor urogastroneFGF fibroblast growth factorFGF7 fibroblast growth factor 7 KGF FIGLA folliculogenesis specific basic helix-loop-helix FIGalpha FOXO3 forkhead box O3 FKHRL1FSH follicle stimulating hormoneFSHR follicle stimulating hormone receptor FSH-RGDF9 growth differentiation factor 9GH1 growth hormone 1 GHGJA1 gap junction protein, alpha 1, 43kDa connexin 43GJA4 gap junction protein, alpha 4, 37kDa connexin 37GNRH gonadotropin-releasing hormone LHRHHSD3B2 hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 2 HSDB3, 3beta-HSD IGF1 insulin-like growth factor 1 Somatomedin-CIGF2 insulin-like growth factor 2 Somatomedin-AIGFBP1 insulin-like growth factor binding protein 1IL1A interleukin 1, alpha IL1-alphaINHBB inhibin, beta BINS insulin proinsulin KISS1 KISS-1 metastasis-suppressor kisspeptin KISS1R KISS1 receptor GPR54 KITLG KIT ligand SCF, Kitl, Steel factor

LEP leptin OBLH luteinizing hormoneLHCGR luteinizing hormone/choriogonadotropin receptor LHRLHX8 LIM homeobox 8LIF leukemia inhibitory factor CDF MAPK1 mitogen-activated protein kinase 1 ERK2MAPK14 mitogen-activated protein kinase 14 P38 MAPK, SAPK2AMMP1 matrix metallopeptidase 1 CLGNMMP2 matrix metallopeptidase 2 CLG4MSH5 mutS homolog 5 (E. coli)NGF nerve growth factor Beta-NGFNOBOX NOBOX oogenesis homeobox OG2NR5A1 nuclear receptor subfamily 5, group A, member 1 SF1, FTZF1PIK3 phosphoinositide-3-kinase PI3KPLC phospholipase CPPARG peroxisome proliferator-activated receptor gamma PPAR-gammaPRKAC protein kinase, cAMP-dependent PKAPTEN phosphatase and tensin homolog PTEN1 RETN resistin RSTN, FIZZ3SERPINE1 serpin peptidase inhibitor, clade E, member 1 PAI1SOHLH1 spermatogenesis and oogenesis specific basic helix-loop-helix 1 TEB2SOHLH2 spermatogenesis and oogenesis specific basic helix-loop-helix 2 TEB1 SP1 Sp1 transcription factor Gremlin-3 STAR steroidogenic acute regulatory proteinSTRA8 stimulated by retinoic acid gene 8 homolog (mouse)TGFA transforming growth factor, alpha TFGATGFB transforming growth factor, beta TGF-beta TIMP1 TIMP metallopeptidase inhibitor 1 TIMPTIMP2 TIMP metallopeptidase inhibitor 2TNF tumor necrosis factor TNF-alpha TP53 tumor protein p53 P53 VEGFA vascular endothelial growth factor A VEGF
Nomenclature suivant le «HUGO Gene Nomenclature Committee» http://www.genenames.org/
Développement folliculaire ovarien et ovulation chez les mammifères / 75
Inra Productions Animales, 2009, numéro 2
Cette revue présente l’état des connaissances sur la folliculogenèse et l’ovulation chez les mammifères. La folliculogenèse basale est unephase de croissance folliculaire lente, au cours de laquelle l’ovocyte acquiert la compétence méiotique. La folliculogenèse terminale est unephase de développement rapide, au cours de laquelle le follicule ovulatoire est sélectionné et termine sa maturation, tandis que l’ovocyteacquiert la compétence au développement. La revue décrit les différents changements fonctionnels qui s’opèrent dans le follicule et l’ovo-cyte au cours de ces deux phases, ainsi que les mécanismes qui les régulent et qui déterminent le quota ovulatoire caractéristique de chaqueespèce ou race. Les facteurs-clés identifiés de la folliculogenèse sont les BMP, le KITLG et l’AMH pour le démarrage de croissance folli-culaire, l’IGF et l’insuline pour la transition entre folliculogenèse basale et terminale, la FSH pour le début de la folliculogenèse termina-le et la sélection des follicules ovulatoires, et la LH pour le développement folliculaire final, la maturation ovocytaire et l’ovulation.L’ovulation est déclenchée par une décharge préovulatoire de LH qui induit la maturation du complexe cumulus-ovocyte dans le follicu-le ovulatoire, suivie de la rupture de la paroi folliculaire et de la formation du corps jaune. La folliculogenèse ovarienne est contrôlée pardes facteurs externes, tels que la photopériode et la nutrition, et c’est essentiellement le système hypothalamo-hypophysaire qui intègre lessignaux endocriniens et environnementaux. Des modifications de l’apport alimentaire, relayées par des variations de signaux hormonauxet métaboliques, modulent l’activité de l’axe hypothalamo-hypophyso-ovarien à ses différents étages.
Résumé
Ovarian follicular development and ovulation in mammals
This review is a state of the art on folliculogenesis and ovulation in mammals. Basal folliculogenesis is a phase of slow follicular growth,during which the oocyte acquires meiotic competence. Terminal folliculogenesis is a phase of rapid development, during which the ovula-tory follicle is selected and completes its maturation while the oocyte acquires developmental competence. This review describes the dif-ferent functional changes occurring within follicles and oocytes throughout these two phases, the underlying regulatory mechanisms and
Abstract

76 / D. MONNIAUX et al
Inra Productions Animales, 2009, numéro 2
the mechanisms that determine natural ovulation rate in different species and breeds. Known key factors for folliculogenesis are BMP,KITLG and AMH for the initiation of follicular growth, IGF and insulin for transition between basal and terminal folliculogenesis, FSHfor the outset of terminal folliculogenesis and the selection of ovulatory follicles, and LH for final follicular development, oocyte matura-tion and ovulation. Ovulation is triggered by a preovulatory LH surge that induces maturation of the cumulus oocyte complex within theovulatory follicle, rupture of the follicular wall and corpus luteum formation. Ovarian folliculogenesis is controlled by external factorssuch as photoperiod and nutrition, and the hypothalamo-pituitary system integrates endocrine and environmental signals. Nutritionalsupply, relayed by hormonal and metabolic signals, can modulate both the hypothalamo-pituitary and ovarian activity.
MONNIAUX D., CARATY A., CLÉMENT F., DALBIÈS-TRAN R., DUPONT J., FABRE S., GÉRARD N., MER-MILLOD P., MONGET P., UZBEKOVA S., 2009. Développement folliculaire ovarien et ovulation chez les mammifères. InraProd. Anim., 22, 59-76.