development of thalamocortical projections in the south american gray short-tailed opossum...
TRANSCRIPT

Development of ThalamocorticalProjections in the South American
Gray Short-Tailed Opossum(Monodelphis domestica)
ZOLTAN MOLNAR,1* GRAHAM W. KNOTT,1,2 COLIN BLAKEMORE,1
AND NORMAN R. SAUNDERS2
1University Laboratory of Physiology, University of Oxford, Oxford OX1 3PT,United Kingdom
2Department of Anatomy and Physiology, University of Tasmania, Hobart, 7001,Tasmania, Australia
ABSTRACTWe determined the time-course and general pattern of thalamocortical development of
Monodelphis domestica by tracing projections with carbocyanine dye in fixed postnatal brainsbetween postnatal day 2 (P2) and P30. By P2, the first neurons have migrated to form thepreplate of the lateral cortex and have sent out axons into the intermediate zone. By P3, fibersfrom the preplate of more dorsal cortex have entered the intermediate zone, and, by P5, theyreach the primitive internal capsule. Crystal placements in the dorsal thalamus at P2–P3reveal thalamic axons extending down through the diencephalon and growing out through theinternal capsule among groups of back-labelled cells that already project into the thalamus.Thalamic axons arrive at the cortex after the arrival of cells of the true cortical plate has splitthe preplate into marginal zone and subplate. Axons from the ventral part of the dorsalthalamus reach the lateral cortex by P5: Dorsal thalamic fibers arrive at the extreme dorsalcortex by P9. The deeper layers of the cortex appear to mature relatively earlier inMonodelphis than in eutherian mammals, and the subplate becomes less distinct. Thalamicfibers and their side branches proceed into the cortex without an obvious period of waiting inthe subplate, but they do not penetrate the dense cortical plate itself. Monodelphis couldprovide an excellent model species, because the development of its thalamocortical connec-tions is entirely an extrauterine process: The period P0–P15 corresponds to that of E12-P0 inthe rat. J. Comp. Neurol. 398:491–514, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: cerebral cortex; internal capsule; thalamus; subplate; waiting period; marsupial
Marsupial and eutherian mammals are believed to havediverged from the same ancestor in the Cretaceous period,some 150 million years ago (Graves, 1991). The study ofmarsupial development can reveal which features areshared with eutherians and, thus, presumably are offundamental importance. We also wanted to determinewhether marsupials are sufficiently similar in the develop-ment of the cerebral cortex to justify their use as a modelspecies for further study, taking advantage of their accessi-bility for early extrauterine experimental manipulation(see, e.g., Saunders et al., 1989; Mark and Marotte, 1992;Saunders, 1997).
In eutherian mammals, axons of the first-generated cellsof the preplate (primordial plexiform layer), ganglioniceminence, and thalamic reticular nucleus establish earlyprojections. Subsequent ascending and descending connec-
tions may develop in the framework of these pioneeringnetworks (see, e.g., McConnell et al., 1989; Blakemore andMolnar, 1990; Shatz et al., 1990; Mitrofanis and Guillery,1993; Metin and Godement, 1996; Molnar et al., 1998).Indeed, it has been suggested that, in rodents, thalamocor-tical fibers grow through the intermediate zone within atopographic ‘‘scaffold’’ of descending preplate axons (see
Grant sponsor: United Kingdom Medical Research Council; Grant spon-sor: National Health and Medical Research Council, Australia; Grantsponsor: Australian Research Council.
*Correspondence to: Zoltan Molnar, University Laboratory of Physiology,University of Oxford, Parks Road, Oxford OX1 3PT, United Kingdom.E-mail: [email protected]
Received 25 November 1997; Revised 28 April 1998; Accepted 7 May 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 398:491–514 (1998)
r 1998 WILEY-LISS, INC.

Molnar and Blakemore, 1995; Molnar et al., 1998). Thecontribution of early-generated cells to forebrain develop-ment, however, is still controversial, and further investiga-tions are required to establish whether they play roles inaxon guidance in all mammalian species.
In carnivores (Shatz and Luskin, 1986; Ghosh andShatz, 1992) and primates (Rakic, 1977; Kostovic andRakic, 1990), as the cortical plate develops and thickens,thalamic fibers grow profusely into the subplate layer,where they accumulate, some of them probably formingsynaptic connections on subplate neurons, for a prolongedwaiting period before they invade the cortical plate. Inrodents, there is not such a distinct waiting period. Tha-lamic fibers do grow preferentially into and along thesubplate layer (Bicknese et al., 1994), where they accumu-late densely for 2–3 days (Lund and Mustari, 1977; Molnaret al., 1998). However, some thalamic axons and their sidebranches advance immediately, although slowly, into thedeepest, more mature part of the cortical plate (Catalanoet al., 1991, 1996; Kageyama and Robertson, 1993; Milleret al., 1993; Molnar et al., 1998). Thus, a distinct andprolonged period of waiting below the cortex may not be anessential part of the process of thalamocortical innervationin all mammalian species. The behavior of thalamic fibersmay simply reflect differences in the rate of maturation ofthe lower layers of the cortex and the onset of growth-permissive properties (Gotz et al., 1992; Molnar andBlakemore, 1995). Examination of marsupials could illumi-nate the question of how universal and important thewaiting period is.
The potential usefulness of marsupials for developmen-tal research has been recognized for many years (Renfreeand Tyndale-Biscoe, 1978; Reynolds et al., 1985; Saunderset al., 1989), but only recently have they begun to be usedfor detailed studies of neural development (see, e.g., Sand-erson and Weller, 1990; Mark and Marotte, 1992; Mark,1997; Saunders, 1997). Not all marsupials breed well incaptivity: Most are seasonal breeders, and many speciesproduce small numbers of offspring at each delivery (Saun-ders, 1997). The South American opossum (Monodelphisdomestica) is a small didelphid marsupial (adult bodyweight 80–120 g) that reproduces well in the laboratoryand is relatively disease free. It has a rather short gesta-tional period (usually 14 days, sometimes 15 days) andproduces large litters (6–12) as many as five times a year(Saunders et al., 1989).
Previous studies on marsupials indicate that the generalpattern of development of the neocortex is similar to thatin eutherians but that it occurs largely postnatally (forreferences, see Saunders, 1997). Just after birth, Monodel-phis neocortex is still at a two-layered ‘‘embryonic stage,’’and the cortical plate until about postnatal day 4 (P4)(Saunders et al., 1989). The equivalent stage of develop-ment occurs in utero in all eutherian mammals studied:around 31–35 days after conception (embryonic days 31–35; E31–E35) in the cat, with a total gestation of 65 days(Luskin and Shatz, 1985); approximately E45 in themonkey, with a term of E165 (Kostovic and Rakic, 1990);E15 in the rat (Valverde et al., 1989); and E14 in the mouse(Takahashi et al., 1995), with a term of E21–E22. At thisextremely early stage of cortical development, the youngMonodelphis is already ex utero, firmly attached to themother’s teats, but not enclosed in a pouch as in mostmarsupial species, including the North American opos-
sum, Didelphis. Thus, immature Monodelphis are particu-larly easily accessible (see Fig. 1).
Monodelphis, which has already been employed for avariety of in vitro investigations (see, e.g., Nicholls et al.,1990; Stewart et al., 1991; Møllgard et al., 1994; Nichollsand Saunders, 1996; Varga et al., 1996), could be aparticularly suitable species for experiments requiringearly manipulations of high precision (see, e.g., Saunders,1997). However, a prerequisite for its use as a modelspecies is the establishment of the time course, generalpattern, and peculiarities of its thalamocortical develop-ment. Comparative studies are also inherently importantfor understanding both the generalities and the idiosyncra-sies of forebrain development in various species. Forexample, marsupials have no corpus callosum and mighttherefore employ rather different strategies for the devel-opment of cortical connectivity. Although there have beennumerous studies of brain development in the tammarwallaby (Reynolds et al., 1985; Reynolds and Saunders,1988; Sanderson and Weller, 1990; Sheng et al., 1990,1991; Jones et al., 1991; Mark and Marotte, 1992; Marotteet al., 1997) and the related quokka (Harman et al., 1995),there has been no previous description of thalamocorticaldevelopment in Monodelphis. We have studied the entirecourse of development of both thalamocortical and earlycorticofugal projections in this species from shortly afterbirth to P30.
MATERIALS AND METHODS
Anesthesia and fixation
All experiments were carried out in accordance with theguidelines of the National Health and Medical ResearchCouncil (Australia) and with the approval of the Univer-sity of Tasmania Ethics (Animals) Committee. Monodel-phis pups of various postnatal ages were obtained from acolony in the Animal House at the University of Tasmania(for breeding and care, see Fadem et al., 1982; Saunders etal., 1989; Saunders, 1997). The first 24 hours after birthwere defined as P0. Young animals were killed with anoverdose of inhalation anesthetic (Methoxyflurane, Syd-ney, Australia) and immediately perfused transcardiallywith 0.1 M phosphate-buffered saline (PBS) at 4°C, pH 7.4.After exsanguination, this perfusate was replaced withfixative (see below). The perfused brain was then exposedand postfixed overnight in the same fixative.
Morphological studies
To study the general morphology of the Monodelphisbrain, young animals were perfused and postfixed witheither Bouin’s fixative or paraformaldehyde (4% in 0.1 MPBS, pH 7.4). The tissue was then dehydrated in a gradedalcohol series and embedded in paraffin wax (melting point54°C). Serial sections (3–5 µm thick) were cut in thecoronal plane and collected on gelatin-coated slides beforebeing stained with toluidine blue.
Placement of carbocyanine dye tracers
After perfusion and postfixation with 4% paraformalde-hyde in PBS, individual, small crystals (0.1 mm diameter)of the fluorescent carbocyanine dye 1,18-dioctadecyl 3,3,3838-tetramethylindocarbocyanine perchlorate (DiI; MolecularProbes, Inc. Eugene, OR) were inserted into different partsof the diencephalon or telencephalon (see Godement et al.,
492 Z. MOLNAR ET AL.

1987). For crystal placements in the dorsal or ventraldivisions of the dorsal thalamus, the brainstem was firsttransected coronally, rostral to the superior colliculus, toexpose the posterior thalamus. A carbocyanine crystal wasthen inserted into a small slit made in the tissue with thetip of a fine pair of forceps. In some animals older than P10,a number of dye crystals were inserted at points across thecortex, usually spaced about 0.75 mm apart, along either aparasagittal or a coronal row. For most of these experi-ments, we used alternating placements of DiI and anotherdye that is well transported, 4-(4-[dihexadecylamino]stryryl)-N-methylpyridinium iodide (DiA). These two dyesare clearly distinguishable under different wave lengths ofillumination.
After crystal insertion, the brains were stored at roomtemperature (22°C) or at 37°C in fixative or PBS contain-ing 0.1% sodium azide to prevent contamination. Afterincubation ranging from 3 days to 6 weeks, depending onage, the brains were embedded in 5% agar and, coronalsections were cut at 70–100 µm on a Vibratome (GeneralScientific, Redhill Surrey, UK). Table 1 summarizes thenumber of brains examined for each crystal placement siteat each age.
All sections were counterstained with bisbenzimide (10minutes; 2.5 µg/ml solution in PBS) to reveal majorboundaries and to confirm the presence of chromatin inback-labelled cells. The sections were coverslipped in PBSand then sealed with Paraseal (R.A. Lamb, London, UK)before being examined under epifluorescence illumination
and photographed. Camera lucida drawings were madefrom some of the serial sections.
RESULTS
Figure 2 demonstrates the size and overall shape ofMonodelphis brains at different stages from birth (P0)until P21. Their gross macroscopic appearance closelyresembles that of the developing rodent brain. However,there is a large difference in the relative maturity of thebrains at birth: The newborn Monodelphis brain is similarto that of a midterm embryonic rodent.
TABLE 1. Numbers of Monodelphis domestica Brains Used forCarbocyanine Dye Tracing and the Locations of the Crystal Placements1
Postnatal Age Ventral thalamus Dorsal thalamus Cortex
P2 2 2 3P3 2 3 4P5 2 3 4P7 2 2 2P8 1 2 1P9 1 1 1P10 2 2 3P11 1 1 1P13 2 2 2P15 1 — 1P17 2 2 2P21 1 — —P20 1 1 1P28 — — 1P30 1 — 1
1Age is indicated in postnatal days (first 24 hours after birth 5 P0).
Fig. 1. Photographs of Monodelphis domestica. A: Mother andpups 1 week after birth (postnatal day 7; P7). The pups, typically 6–12in number, attach themselves to the teats of the mother’s abdomen. Incontrast to many other marsupials, they are not enclosed in a pouch,
making them relatively easily accessible. B: A mature, 4-month-oldMonodelphis. C: P1 Monodelphis pup removed from the teat. Itscrown-rump length is 1 cm. Eye opening occurs during the fifth week,and the young are weaned 50 days after birth.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 493

Histological appearanceof Monodelphis forebrain
Development of cortical lamination. Toluidine blue-stained coronal sections through the cortex at the level ofthe anterior commissure reveal that, shortly after birth inMonodelphis, even the most mature lateral part of theneocortex is at a two-layered ‘‘embryonic’’ stage of develop-ment, consisting of a wide inner neuroepithelium (ventricu-lar and subventricular zones) and a narrow outer preplate(primordial plexiform layer) that consists of the firstpostmigratory neurons (Fig. 3P0). There is a clear rostro-lateral-to-mediocaudal gradient of maturation across thehemisphere (as in other mammals), such that, in coronalsections from the middle of the hemisphere, the extremelateral cortex is about 4 days more mature than the dorsalextremity of the cortex. The following description appliesto the lateral cortex unless specified otherwise.
Cells of the true cortical plate first appear between P2and P4, sandwiched within the preplate of the lateralcortex (Fig. 3). The splitting of the preplate is especiallyclear at P4, when the marginal zone and subplate of thelateral cortex are seen to be continuous with the stillunsplit preplate of the dorsal cortex. The similarity of thepresumed subplate in marsupials to that of mammals isfurther indicated by the fact that cells of the deepest partof the cortex in both Monodelphis and the tammar wallabyexpress the heavily glycosylated protein fetuin at thisearly stage, as do neurons of the true subplate and thedeepest cells of the cortical plate in eutherians (Adam etal., 1993).
At P8, the cortical plate of the lateral cortex consists oftwo distinct components (Fig. 3). Directly below the mar-ginal zone is a layer of densely packed, immature neurons,most of which have presumably just completed theirmigration. We use the term ‘‘dense cortical plate’’ todescribe this superficial band of immature cells. Immedi-ately beneath the dense cortical plate is a much looserlayer of clearly more mature neurons. At this stage, theoriginal, narrow subplate, with its somewhat denser popu-lation of neurons, can be distinguished as a separate layerbelow the plate. By P10, there has been a substantialincrease in the depth of the cortical wall, although thedense cortical plate, which contains the newly arrivedneurons, is similar in thickness to that at P6. This is in
contrast to eutherians, in which there is a progressive andsubstantial thickening of the dense cortical plate duringthe equivalent phase of cortical development.
In Monodelphis, the increase in overall cortical thick-ness at this early stage is due to a marked expansion of thelayers between the dense cortical plate and the ventricularzone (see P10 in Fig. 3). This region is filled with looselypacked cells of more mature appearance, often arranged inrather orderly, surface-parallel rows. A similar appearancehas been described in the tammar wallaby, in whichthymidine labelling studies (Reynolds et al., 1985) haveshown that the cells constituting these rows are bornrelatively early. At this stage, they presumably include thecells of true cortical layers VI and even V, which appear tomature more quickly than in eutherians.
By histological criteria alone, it is no longer possible todistinguish a separate subplate from P10 onward (Fig. 3).It is conceivable that the cells of the subplate are verytransient and that most have already died at this stage.Figure 3 illustrates that, by P20, the ventricular andsubventricular zones have reduced in size considerably,and all but the supragranular layers of the cortex appearto be quite mature.
General pattern of thalamic development. For thefirst few days after birth, the major diencephalic subdivi-sions in Monodelphis can be distinguished only by featuresof the diencephalic wall (Fig. 4). At P0 and P2, a ventricu-lar groove is seen on the anterior diencephalic wall,between the putative hypothalamus and the thalamus,and a protuberance marks an apparent boundary betweenthe future dorsal thalamus (containing the principal relaynuclei) and the ventral thalamus (containing the putativethalamic reticular nucleus). This stage corresponds to E12in the rat (Coggeshall, 1964). At P2, the wedge-shapedenlargement in the middle of the thalamus becomes moreapparent together with upper and lower furrows demarcat-ing the borders between thalamus and epithalamus, above,and hypothalamus, below. The central protrusion andlower groove become less distinct as the diencephalonmatures, and, by P5, the lower part of the ventricularspace has collapsed, leaving only a dorsal ventricularlumen. At P5, the beginnings of nuclear differentiation arevisible in the epithalamus and ventral thalamus in Mono-delphis but not yet in the dorsal thalamus, as observed
Fig. 2. Gross appearance of Monodelphis domestica brains viewedfrom above at various developmental stages (P0–P21). On the basis ofoverall macroscopic appearance, the development of the brain is very
similar to that of rat or mouse. However, there is a great difference inthe relative state of maturity at birth. Anterior is up, and posterior isdown. Scale bar 5 1 mm.
494 Z. MOLNAR ET AL.

previously at E16–E17 in the rat (Coggeshall, 1964). InMonodelphis, thalamic nuclear differentiation starts inthe posterolateral segment and proceeds anteriorly, medi-ally, and dorsally, as in other mammalian species (Rose,1942; Coggeshall, 1964; Jones, 1985). The dorsal lateralgeniculate nucleus begins to differentiate from the rest ofthe thalamus at around P5, a stage that corresponds toE16 in the rat (Coggeshall, 1964; Lund and Mustari, 1977).
Tracing of thalamocortical axons
Outgrowth of axons from the thalamus. At P2, axonshave started to grow out of all parts of the dorsal thalamus.By P3, DiI crystals placed either dorsally or ventrally inthe dorsal thalamus reveal anterogradely labelled tha-lamic fibers extending from the crystal placement sitethrough the basal diencephalon and into the nascentinternal capsule (Fig. 5). In addition, groups of retro-gradely labelled cell bodies are also seen, a few in theepithalamus, but most lying in the pathway of the growingthalamic axons (see below).
At P5, a crystal in either the ventral or the dorsal half ofthe dorsal thalamus reveals thalamocortical fibers thathave passed through the primitive internal capsule intothe intermediate zone of the basal telencephalon. From theventral segment, a few fibers are seen approaching the
most mature lateral part of the neocortex at this age (Fig.6). At each point in the hemisphere, thalamic fibersapproach the cortex a couple of days after the arrival of thefirst neurons of the dense cortical plate, when it is five toten cells thick. In accordance with this, cells in the moreposterior and ventral part of the dorsal thalamus first canbe back-labelled from crystal placements in the lateralcortex at P5, whereas more anterior and dorsal thalamicneurons can be labelled from dorsolateral cortex at P7 (seeFigs. 14 and 15 below).
Fiber ordering along the thalamocortical pathway.
In Monodelphis, as in the rat and mouse (see, e.g., Molnarand Blakemore, 1995), thalamic fibers show three distinctpatterns of organization along their pathway (see Fig. 6).Axons remain parallel within the diencephalon, and theyconverge to form a tight, homogeneous bundle as theyapproach the primitive internal capsule; however, onentering the corpus striatum, the bundle breaks up intoseparate fascicles. At the lateral edge of the striatum,thalamic fibers defasciculate and course toward the cortex,running as individual axons along parallel paths in theintermediate zone.
Progressive invasion of the cortex by thalamic axons.
Figures 6 and 7 illustrate the sequence of events as axons,which were labelled from a crystal placed in the dorsal
Fig. 3. Toluidine-blue-stained coronal sections of Monodelphisbrains illustrating the general appearance of the cerebral wall of theleft hemisphere from birth (P0) to P20. To facilitate comparison, thesections were all taken from the coronal level of the anterior commis-sure. Lateral is up, and dorsal is to the right. At birth and P2,Monodelphis neocortex is still at an extremely rudimentary, two-layered stage of development, with a wide inner ventricular zone (vz)merging with the subventricular zone (svz) and with a narrow outerpreplate (primordial plexiform layer; pp). Arrows (at P0, P2, and P8)indicate mitotic spindles in cells at the ventricular surface of theventricular zone (visualized under higher power). At P4–P6, the firstcells of the cortical plate (cp) arrive after migrating through the deepercells of the preplate from the proliferative layers below, hence,splitting the preplate into the marginal zone (mz) above and thesubplate (sp) below. Note that more dorsal cortex (to the right) is lessadvanced in its development compared with lateral cortex. At P8,
directly below the narrow marginal zone is a tightly packed layer ofnewly arrived, immature cells, constituting the dense cortical plate(dcp). Deep to this is a zone of more mature, widely spaced, postmigra-tory neurons, presumably the future layer VI and perhaps layer V.Cells of the original subplate are still distinguishable at the base of thecortex, although the boundary between layer VI and subplate isindistinct. At this age, numerous mitotic spindles are still visible (athigher power) in the ventricular zone (arrows indicate examples). AtP10 and P15, in contrast to the appearance in rodents, the densecortical plate has not increased significantly in thickness, but there isa substantial increase in the depth of the presumptive lower corticallayers. From P10 onward, it is impossible with Nissl staining todistinguish the original subplate. At P20, cortical neuronogenesisappears to be complete, and the adult layers (I–VI) can be discerned,although cells of the supragranular layers are still quite denselypacked. wm, White matter. Scale bar 5 100 µm.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 495

Figure 4
496 Z. MOLNAR ET AL.

thalamus, approach the dorsolateral segment of the cor-tex. At P5, the most advanced of these axons are ascendingas a parallel array through the intermediate zone, andthey arrive below this portion of the cortical plate at aboutP7 (Fig. 7, top). At this stage, the cortex appears relativelymore mature than the equivalent stage in the rat (E15–E16) or mouse (E14–E15), in which virtually the entirecortical plate consists of densely packed, less mature cellswith a relatively thin layer of more mature infragranularneurons underneath and a fairly clear residual subplatelayer below that (see Molnar and Blakemore, 1995). Whenthalamic axons arrive in Monodelphis, the dense corticalplate has not increased enormously in thickness, and asubstantial population of loosely packed, more maturepostmigratory neurons, which are presumably destined tobecome the future infragranular layers, has emergedbelow it (see Fig. 3). A separate subplate layer can nolonger be discerned.
By P9, some thalamic axons have turned to enter thecortical gray matter, running upward at a variety ofangles, but roughly radially. The difference between theregular parallel array in the intermediate zone and thedisorganized pattern of growth into the cortex is particu-larly clear in Figure 7 (middle).
By P11, most principal axons have entered and branchedwithin the lower parts of the cortex. Many extend upthrough the quite thick, more mature infragranular layersto the lower boundary of the dense cortical plate, althoughvery few actually penetrate the latter.
Early thalamofugal projections
Dye crystals placed either ventrally or dorsally in thedorsal thalamus, even as early as P2–P5, back labelnumerous cell groups along the growth pathway of thethalamocortical fibers, as described above. Many are seenin the ventral thalamus in a region presumed to beequivalent to the rodent thalamic reticular nucleus. An-other distinct group lies within the primitive internalcapsule, below the ganglionic eminence, in the region thatMitrofanis (1992) has called the ‘‘perireticular’’ cell groupin the rat (see Figs. 8–11). At these early ages, crystals
placed more ventrally within the thalamus also label cellsin the hypothalamus as well as in more dorsal thalamusand epithalamus (Fig. 11).
By P10, even dorsal thalamic crystal placements backlabel cells extensively throughout the diencephalon, includ-ing the epithalamus and the hypothalamus, as well as inthe perireticular group (Fig. 12). A greater density ofperireticular cells is labelled from the ventral division ofthe dorsal thalamus. In addition, by this age (but notbefore), a crystal placed in the ventral division labels cellsin a discrete region of the lateral cortex. These cells lie notonly at the boundary with the intermediate zone (possiblyequivalent to the original subplate) but also in the matureinfragranular cell layers, although not within the densecortical plate itself (Fig. 12). No cortical cells are labelledfrom extreme dorsal thalamus (using the same incubationprotocol), which implies that corticofugal projections haveinvaded the ventral diencephalon, but not the dorsaldiencephalon, by P10.
Labelling from the cortex
Early corticofugal projections. At P2–P3, about 3days before the arrival of the first thalamic fibers, a crystalplaced in the cortex labels axons leaving the most lateralsegment of the cortex (Fig. 13). Because this is still 1–2days before the appearance of the cortical plate, thesecorticofugal fibers must arise from preplate cells. Even asearly as P3, a crystal placed in the lateral cortex labels asubstantial bundle of fibers descending as a parallel arraythrough the intermediate zone; by P5, even corticofugalaxons from more dorsal cortex are approaching the primi-tive internal capsule (Fig. 13).
Up to P5, the fibers labelled from crystals placed in thedorsolateral convexity of the cortex and above end in largegrowth cones and are therefore almost certainly corticofu-gal. A few are seen to branch as they approach the anlageof the corpus striatum (Fig. 13E,F). No thalamic cells areback labelled from such crystal placements at this age.
Early projection into the anterior commissure.
Monodelphis, in common with other marsupials, does nothave a corpus callosum, and no medially directed axonswere seen even with the most dorsal cortical crystalplacements. However, axons directed toward the anteriorcommissure (the major forebrain commissure in marsupi-als) are seen diverging from the main population ofdescending corticofugal axons as early as P7 (Fig. 14). Atthis age, a single cortical crystal placement labels a bundle(consisting of corticofugal and thalamocortical fibers ex-tending all the way to the thalamus). At the boundarybetween the intermediate zone and the internal capsule, asmall filament of labelled fibers diverges from the mainbundle and runs ventrally and medially to form theanterior commissure.
Early corticopetal projections. Crystals placed inthe more mature lateral cortex first label cell bodiesretrogradely in the ventral part of the dorsal thalamus asearly as P5 (Figs. 14, 15). In addition, occasional axons canbe traced back to labelled cell bodies lying within theinternal capsule, in approximately the same region thatcan be back labelled from the thalamus at this age. Nosuch cells are labelled from more dorsal cortical crystalplacements. The serial camera lucida drawings in Figure15 demonstrate the distribution of cells in the internalcapsule and in the thalamus that were back labelled fromlateral cortex at P5.
Fig. 4. Toluidine-blue-stained coronal sections illustrating thedifferentiation of the subdivisions and nuclei of the thalamus from P0to adulthood. With respect to the telencephalon, the general pattern ofthalamic development is very similar in Monodelphis to that in rat andmouse, but with birth occurring at a much earlier stage. P0: Only themajor diencephalic subdivisions can be distinguished. A ventriculargroove appears on the anterior diencephalic wall between the hypo-thalamus below and the thalamus above (lower arrow). This stagecorresponds to embryonic day 12 (E12) in the rat (Coggeshall, 1964).The upper arrow indicates a protrusion defining a boundary betweenventral and dorsal thalamus. P2–P3: The wedge-shaped enlargementin the diencephalic wall (middle arrow) between the developing dorsalthalamus (DT) and ventral thalamus (VT) becomes more evident.Furrows (upper and lower arrows) demarcate the boundaries betweendorsal thalamus and epithalamus (ET) above and between centralthalamus and hypothalamus (HT) below. P5: The general appearanceof the diencephalon of Monodelphis is now similar to that of a fetal ratat E16–E17 (Coggeshall, 1964). The central protrusion and lowerfurrows have virtually disappeared, and the ventral part of theventricle has collapsed, leaving mainly a dorsal lumen. Nucleardifferentiation is commencing in the epithalamus and ventral thala-mus but has not yet begun in the dorsal thalamus at this coronal level.Adult: Coronal section through the middiencephalic level of the adultMonodelphis brain. Scale bars 5 200 µm for P5 (also applies to P0, P2,and P3), 1 mm for adult.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 497

Fig. 5. A–H: Anterograde labelling of thalamic axons and retro-grade labelling of cell groups after placement of a crystal of carbocya-nine dye (DiI) in the dorsal thalamus at P3. Series of coronal sectionsshown in ultraviolet illumination to show bisbenzimide counterstain(top row) and as fluorescence photomicrographs (bottom row) to revealDiI labelling. The sections proceed from posterior to anterior (left toright). A,E: At the level of the crystal placement itself (asterisk in E), amass of axons (both retrograde and anterograde labelling) are seenstreaming down from the dorsal thalamus (DT), through the ventralthalamus (VT), toward a group of back-labelled cells (arrow in E),which is presumed to be in the thalamic reticular nucleus. B,F andC,G: Farther rostral, the thalamic axons extend deeper through theventral diencephalon and have turned laterally toward the primitive
internal capsule (PIC), the entrance of which is indicated by thearrows in G. The group of back-labelled cells in the ventral thalamus,which are evident in F and G, extends rostral to the bundle of thalamicaxons, LV: lateral ventricle. D,H: At this more anterior plane, the frontof the advancing bundle of thalamic fibers (arrow) emerges from theprimitive internal capsule at the level of the middle of the ganglioniceminence (GE), heading toward the lateral cortex (CTX). The fibersdiverge slightly and form distinct fascicles as they pass through theanlage of the developing corpus striatum. In this particular specimen,very few back-labelled cells were visible in the region of the ‘‘perireticu-lar’’ cell group within the primitive internal capsule. ET, epithalamus.Scale bar 5 250 µm.
Fig. 6. A–I: Growth of thalamocortical fibers through the interme-diate zone. Each row shows a different age (P5, P9, and P11). A,D,G:Fluorescence photomicrographs at the level of the DiI crystal place-ments in the dorsal thalamus (arrowheads). B,E,H: More rostralsections at each age with major structural boundaries revealed bybisbenzimide counterstaining. C,F,I: Fluorescence photomicrographsof the same sections shown in B,E,H, respectively, showing the bundleof labelled thalamic fibers advancing through the intermediate zoneand then entering the more dorsal cortex. Arrows indicate the tips ofthe labelled thalamic fibers. At P5, the crystal in the lower part of thedorsal thalamus back labels cells farther dorsal in the thalamus aswell as some in the ventral diencephalon (in the presumed thalamicreticular nucleus; arrow in A). Labelled thalamic fibers have justpassed through the primitive internal capsule (PIC; delimited by
arrows in B) underneath the ganglionic eminence (GE). LV, lateralventricle; H, hippocampus. At P9, a similar crystal placement a littlemore dorsal within the dorsal thalamus reveals a denser cluster ofback-labelled cells in the presumed thalamic reticular nucleus (arrowin D). Thalamic fibers are arranged in fascicles as they traverse theanlage of the corpus striatum and then break up into a parallel arrayof individual axons as they enter the intermediate zone (IZ): Somehave already reached the lateral cortex. VZ, ventricular zone. By P11,labelled thalamic fibers have reached more dorsal parts of the cortexand are entering radially into the gray matter deep to the densecortical plate, presumed to consist of true layer VI and the residue ofthe subplate. Arrows in G demarcate the entrance of the internalcapsule. Scale bars 5 100 µm in A (also applies to D), 200 µm in C (alsoapplies to B,E,F,H,I), 500 µm in G.
498 Z. MOLNAR ET AL.
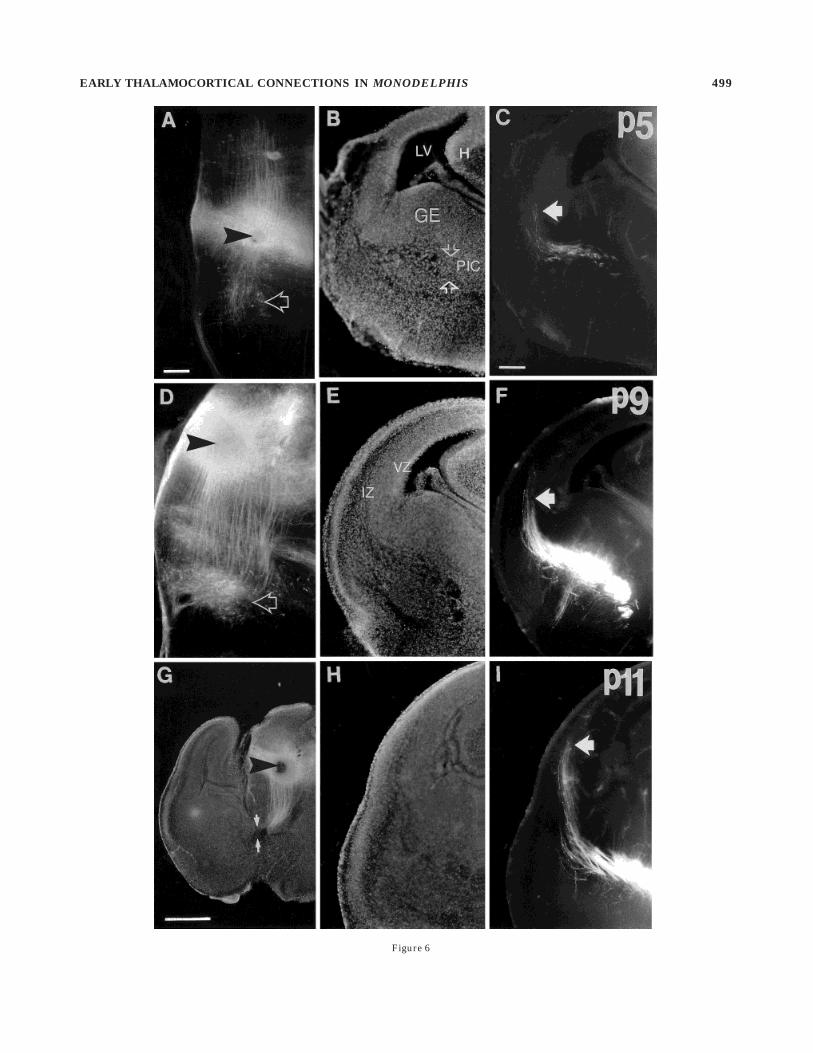
Figure 6
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 499

At progressively later ages, from P5 to P9, thalamic cellscan be back labelled from progressively more dorsal posi-tions in the cortex. These thalamic relay cells always lie in
the dorsal thalamus, i.e., within the region that containsthe principal projection nuclei. Cells in the ventral thala-mus (mainly consisting of the thalamic reticular nucleus)
Fig. 7. Bisbenzimide counterstaining (A,C,E) and fluorescencephotomicrographs (B,D,F) of individual coronal sections of the righthemisphere illustrating the development of lamination in the dorsolat-eral segment of the occipital cortex and the chronology of ingrowth ofaxons labelled by a DiI crystal placement in the dorsal thalamus.Lateral is up, and ventral is to the right. At P7 (A,B), thalamic axonsare still growing through the intermediate zone (IZ) and approachingthis more dorsal region of the occipital cortex (B). There is already arelatively thick layer of more mature postmigratory cells below thedense cortical plate (DCP) at this stage, and there is no clear boundarybetween these cells and the residual subplate. The cortex appearsrelatively more mature than at the equivalent stage in the rat (E16 for
occipital cortex), in which almost the entire true cortical plate consistsof densely packed immature cells. At P9 (C,D), most principal axonshave immediately invaded the more mature, lower layers of the cortex,where many have branched, although very few penetrate the densecortical plate. Note the way in which the regular parallel array ofthalamic axons in the intermediate zone becomes disordered as theaxons and their branches enter the cortex at a variety of angles. At P11(E,F), more thalamic fibers have arrived and proceeded into the deeplayers of the cortex. Although there is a high density of axons in theintermediate zone, they do not form a distinct plexus in the presumedsubplate layer. MZ, marginal zone; SVZ, subventricular zone; VZ,ventricular zone. Scale bar 5 100 µm.
500 Z. MOLNAR ET AL.

were never labelled from the cortex at any age studied.Indeed, the pattern of thalamic cell bodies and neuropilthat were back labelled from the cortex defines a rathersharp boundary between true projection thalamus (dorsalthalamus) and ventral thalamus (Figs. 14, 16).
Topography of the initial thalamocortical relation-
ship. After about P9–P10, a crystal placed at any point inthe hemisphere labels a group of cells in the dorsalthalamus (see Fig. 16). To examine the topographic relation-ship between thalamus and cortex at this early stage of
development, before the differentiation and separation ofthalamic nuclei, we placed a number of carbocyaninecrystals at different points in the same hemisphere. Whenthree dye crystals were placed approximately 750 µmapart in a parasagittal or coronal row along the convexityof the cortex, each crystal labelled a separate fiber bundleextending from cortex to thalamus and a discrete cluster ofthalamic cells. A coronal row of three crystals labels threegroups of thalamic cells that form a more-or-less continu-ous slab running obliquely through the dorsal thalamus
Fig. 8. In another P3 specimen (compare with Fig. 5), crystalplacements in the dorsal thalamus (on the left side of the brain) and inthe primitive internal capsule (right side) reveal the projections ofcells in the presumed perireticular cell group within the primitiveinternal capsule (the camera lucida drawings in Fig. 9 show the crystalplacement sites and the overall distribution of labelled cells in thisbrain). A,B: The left half of a single coronal section in which a DiIcrystal had been placed in the dorsal thalamus (see Fig. 9). A showsbisbenzimide counterstain, whereas the fluorescence photomicro-graph in B reveals a dense group of back-labelled cells amongpresumed thalamic axons within the primitive internal capsule (PIC).
Arrows in B indicate three examples of labelled cell bodies. C: Higherpower view of the group of labelled perireticular cells with the samethree neurons shown in B indicated by arrows: They have fusiform ormultiform morphology. D: Fluorescence photomicrograph of the basaltelencephalon of the right hemisphere at a coronal level close to thesite of a dye crystal in the primitive internal capsule. A group ofpresumed perireticular cells is back-labelled, lying in the primitiveinternal capsule (PIC) below the middle of the ganglionic eminence(GE). The arrow indicates a multiform cell (also shown at higherpower in the inset). CTX, lateral cortex. Scale bars 5 100 µm in A (alsoapplies to B), 100 µm in C,D, 50 µm in inset.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 501
when viewed in the horizontal plane. Movement fromlateral to dorsal along the surface of the cortex correlateswith a rostromedial-to-caudolateral sequence in the thala-mus. A parasagittal row of three cortical crystals leads tothe labelling of three distinct blocks of thalamic cells, eachsomewhat elongated orthogonal to the axis of their separa-tion. The most rostral cortical site labels the most caudome-dial slab of thalamic cells, whereas the most caudalposition in the cortex labels the most rostrolateral slab (seeFig. 17). Thus, the entire volume of the dorsal thalamus
appears to map continuously across the entire surface ofthe neocortex, in a pattern that implies that there is asingle rotation in the array of axons, probably occurring inthe basal diencephalon.
DISCUSSION
Monodelphis is so immature at birth that the entireprocess of formation and innervation of the neocortexoccurs postnatally. The first neurons start to arrive under
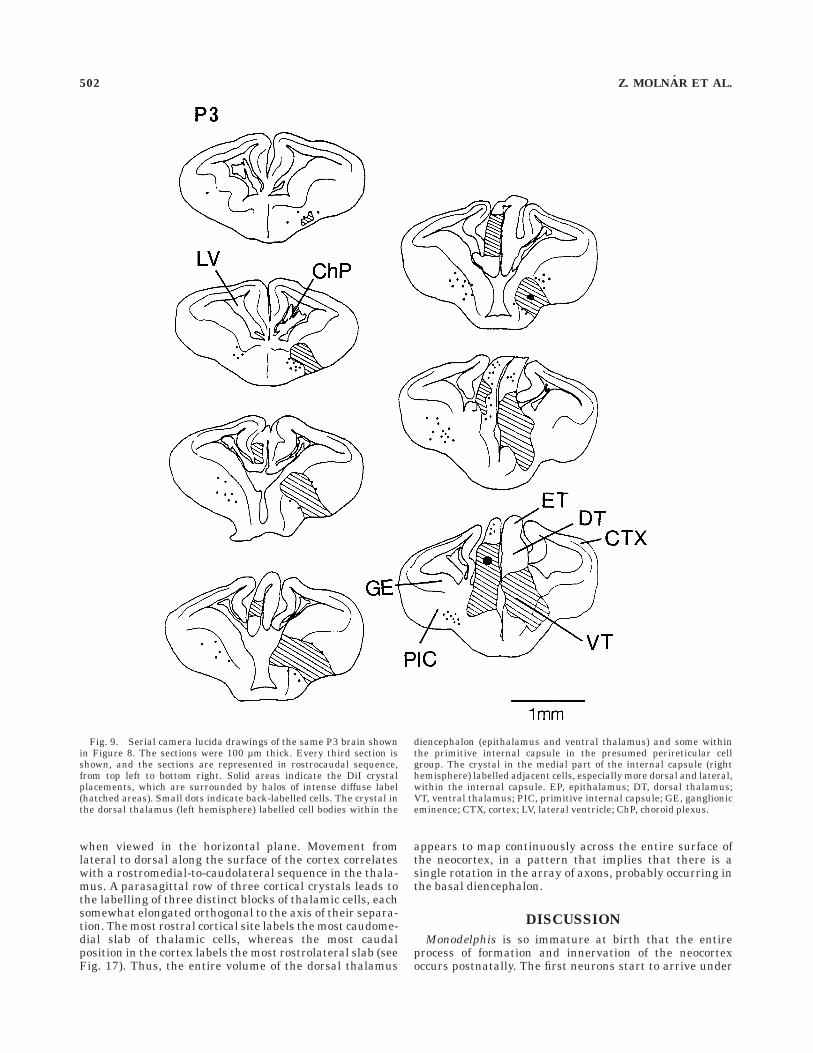
Fig. 9. Serial camera lucida drawings of the same P3 brain shownin Figure 8. The sections were 100 µm thick. Every third section isshown, and the sections are represented in rostrocaudal sequence,from top left to bottom right. Solid areas indicate the DiI crystalplacements, which are surrounded by halos of intense diffuse label(hatched areas). Small dots indicate back-labelled cells. The crystal inthe dorsal thalamus (left hemisphere) labelled cell bodies within the
diencephalon (epithalamus and ventral thalamus) and some withinthe primitive internal capsule in the presumed perireticular cellgroup. The crystal in the medial part of the internal capsule (righthemisphere) labelled adjacent cells, especially more dorsal and lateral,within the internal capsule. EP, epithalamus; DT, dorsal thalamus;VT, ventral thalamus; PIC, primitive internal capsule; GE, ganglioniceminence; CTX, cortex; LV, lateral ventricle; ChP, choroid plexus.
502 Z. MOLNAR ET AL.

the pial surface of the lateral cortex around the time ofbirth to form the preplate. There is a distinct gradient ofmaturation across the hemisphere, such that, in themiddle of the hemisphere, there is roughly a 4-day differ-
ence in the state of development of the most mature lateralsegment and the extreme dorsal convexity of the cortex.
Neurons of the preplate in Monodelphis, as described incarnivores (McConnell et al., 1989) and rodents (Molnar
Fig. 10. In this P5 specimen, a crystal placed at the boundarybetween dorsal thalamus (DT) and ventral thalamus (VT) of the righthemisphere reveals back-labelled perireticular cells within the primi-tive internal capsule as well as thalamic axons extending through theinternal capsule and into the intermediate zone, but not yet reachingthe lateral cortex. A and C show bisbenzimide counterstaining,whereas B and D are fluorescence photomicrographs of the samesections. A,B: At the coronal level of the crystal placement (asterisk inB), axons (presumably retrogradely labelled) as well as the processesof thalamic radial glia are seen running upward toward a group of
back-labelled cells in the dorsal thalamus. C,D: In this more rostralsection, thalamic axons are seen in the primitive internal capsule(PIC) within the anlage of the corpus striatum. Many back-labelledcell bodies were also seen among the thalamic axons in this region inthe presumed perireticular group. An arrow in D indicates one suchcell, and the enlarged, high-power inset shows its fusiform morphol-ogy, with dendrites extending perpendicular to the trajectory ofthalamocortical fibers. Scale bars 5 250 µm in A (also applies to B) andC (also applies to D), 50 µm in inset.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 503

Fig. 11. Serial camera lucida reconstructions of coronal sectionsfrom a P5 brain with crystal placements in the ventral segment of thedorsal thalamus (DT) of the left hemisphere and farther dorsal in theright hemisphere (for conventions, see Fig. 9). Dots represent indi-vidual back-labelled cells. Both ventral and dorsal thalamic crystals
label perireticular cells in the primitive internal capsule (PIC) be-neath the ganglionic eminence. The ventral thalamic (VT) crystalplacement labels larger numbers of hypothalamic cells in addition tothe cells in the dorsal thalamus and internal capsule. HT, hypothala-mus; GE, ganglionic eminence; CTX, cortex.
504 Z. MOLNAR ET AL.
and Blakemore, 1990; DeCarlos and O’Leary, 1992; Erzu-rumlu and Jhaveri, 1992), send out axons before thearrival of true plate cells. These pioneering fibers form an
ordered array, descending through the intermediate zone,the first of them reaching the lateral margin of the nascentinternal capsule at about P3. Meanwhile, axons from the
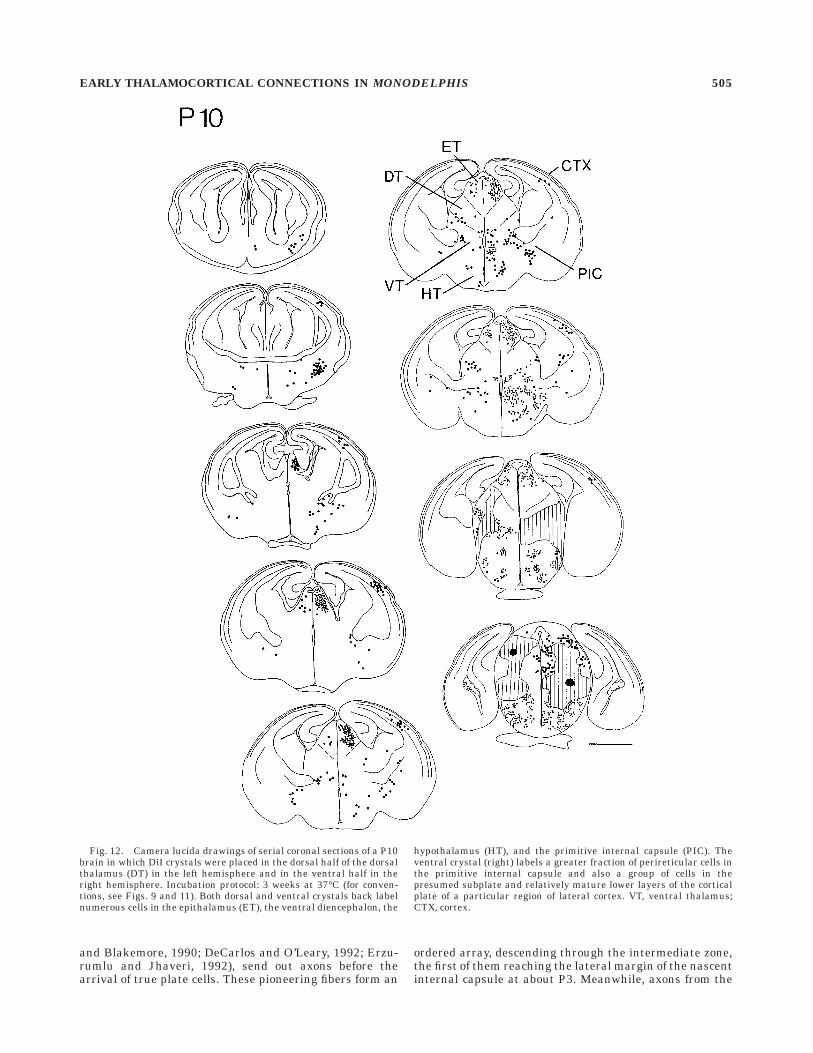
Fig. 12. Camera lucida drawings of serial coronal sections of a P10brain in which DiI crystals were placed in the dorsal half of the dorsalthalamus (DT) in the left hemisphere and in the ventral half in theright hemisphere. Incubation protocol: 3 weeks at 37°C (for conven-tions, see Figs. 9 and 11). Both dorsal and ventral crystals back labelnumerous cells in the epithalamus (ET), the ventral diencephalon, the
hypothalamus (HT), and the primitive internal capsule (PIC). Theventral crystal (right) labels a greater fraction of perireticular cells inthe primitive internal capsule and also a group of cells in thepresumed subplate and relatively mature lower layers of the corticalplate of a particular region of lateral cortex. VT, ventral thalamus;CTX, cortex.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 505

dorsal thalamus have grown through the primitive inter-nal capsule and are just emerging into the intermediatezone. In the rodent, there is evidence that early thalamocor-tical axons become intermingled with the scaffold of pre-plate axons, growing among them and perhaps using theirtopography to guide them toward the corresponding re-gions of the cortex (Molnar et al., 1998). The trajectoriesand distributions of the two sets of axons in Monodelphisalso suggest that they might meet and associate, butfurther work is needed to demonstrate such an interac-tion between descending and ascending fibers in marsupi-als.
Apart from the discrepancy in the general time scale ofcortical development, perhaps the most striking way inwhich Monodelphis differs from rodents is in the rapidmaturation of the early postmigratory neurons of itscortical plate. Just as in eutherians, cells of the truecortical plate migrate through the lower part of thepreplate, taking their place, in an inside-out sequence,between the marginal zone and the subplate. The mostnewly arrived of those neurons form the tightly packed,dense cortical plate below the marginal zone. In rodents,the dense cortical plate thickens progressively and consti-tutes much of the entire thickness of the cortical plate
Fig. 13. Early corticofugal projections revealed with crystals of DiIplaced in the dorsolateral occipital cortex at P3 (A–C) and P5 (D–F). Aand D show bisbenzimide counterstain, whereas B and E are fluores-cence photomicrographs of the same sections. C and F are higherpower views. At P3, the cerebral wall consists of only a loose preplate(pp) above the subventricular zone (svz) and ventricular zone (vz; seeFig. 3). The crystal, which was inserted at the site marked by thearrow in B, stained a large group of radial processes (presumed radialglia) that extend down to the ventricular surface. A bundle of labelledaxons is also visible within the region of the outline box (shownenlarged in C). The axons, which were presumably derived from
preplate cell bodies, are growing along curved trajectories through theintermediate zone toward the primitive internal capsule (PIC), thelateral entrance of which is demarcated by the arrows in C. Growthcones are visible at the tips of many axons. In another example, at P5,the crystal was placed a little more dorsal in the cortex, where thepreplate has still not been split by the arrival of true cortical platecells. Many of the corticofugal axons have growth cones at their tips,and some branch (arrows in E and F) as they enter the anlage of thecorpus striatum. Scale bars 5 250 µm in A (also applies to B), 100 µmin D (also applies to C,E), 50 µm in F.
506 Z. MOLNAR ET AL.

when thalamic axons arrive in the subplate layer below. Bycomparison, the dense cortical plate remains at much thesame thickness through most of the period of neuronogen-esis in Monodelphis. Cortical neurons appear to mature
more quickly after their migration and to drop out of thedense cortical plate to form a rapidly expanding layer ofloosely packed cells below. This layer of relatively maturecortical neurons, which presumably is destined to become
Fig. 14. Divergence of the corticofugal projection into the anteriorcommissure. The photomicrographs are from a P7 brain in which a DiIcrystal was placed in the dorsolateral convexity of the right hemi-sphere. A,B: Counterstaining (A) and axon staining (B) in a singlecoronal section containing the crystal site (large solid arrow). A singlebundle of descending axons emerges from the site: This consists notonly of early corticofugal axons but also of retrogradely stainedthalamic axons, which have already reached this region of the cortexat P7. At the boundary between the intermediate zone and theganglionic eminence (open arrow), the bundle splits into two (indi-cated by the small solid arrows). C: Fluorescence photomicrographtaken from an adjacent section in which the bundles have separated
farther. The thicker, medial group of fibers (top solid arrow) follows themain path through the internal capsule (IC), but the thinner, lateralbundle plunges ventrally and then runs horizontally (bottom solidarrow) into the nascent anterior commissure (AC). D,E: Higher powerviews of axon staining (D) and counterstain (E) of the region outlinedin C. F: Fluorescence photomicrograph of another section, which waslocated beyond the anterior commissure, showing the thalamic axonsof the main fiber bundle ending in a group of back-labelled neurons inthe lower part of the dorsal thalamus (DT). CTX, cortex; VT, ventralthalamus. Scale bars 5 250 µm in A (also applies to B,C), 100 µm in D(also applies to E,F).
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 507

the lower part of the infragranular layers, is already quitesubstantial at the time when thalamic axons arrive.
Similar evidence of early maturation of the lower layersof the cortex has been reported for other marsupials,
including the tammar wallaby (Reynolds et al., 1985;Marotte et al., 1997) and the Northern Native cat (Molnaret al., 1995). In these species, as in Monodelphis, cells ofthe lower layers not only mature much more rapidly but
Fig. 15. The origin of the first axons reaching the neocortex isshown in these camera lucida drawings of the distribution of back-labelled cells in serial coronal sections of a P5 brain in which a singleDiI crystal was placed into the lateral segment of the cortex on eachside near the middle of the rostrocaudal extent of the hemisphere (forconventions, see Fig. 9, except that every third 100-µm-thick sectionfrom the series is shown for the region around and immediately
posterior to the crystal placements). Even at this early age, a crystal inthe lateral cortex labels cells in the dorsal thalamus (DT), mainlyrestricted to its ventral part. A group of perireticular cells below theganglionic eminence is also labelled. It is not until P7 that a crystalinserted more dorsally in the cortex causes thalamic labelling. Cells inthe ganglionic eminence are not labelled from dorsal cortical sites evenat that age. CTX, cortex; PIC, primitive internal capsule.
508 Z. MOLNAR ET AL.

also have a degree of structural order that is not seen inthe eutherian species studied so far: Rows of neurons lineup in register, parallel to the surface of the brain (Reynoldset al., 1985). These differences have led to difficulties indefining the subplate in marsupials (Saunders, 1997).Unlike in eutheria, the histological boundary betweencortical plate and subplate (derived from the preplate)rapidly becomes indistinct. Moreover, in both the tammar(Reynolds et al., 1985) and in Monodelphis (Adam et al.,1993), immunoreactivity for fetuin and staining with thelectin Sambucus nigra (which binds to terminal carbohy-drate residues on fetuin) occur in the deepest cells of thecortex for only a few days immediately after their appear-ance and then rapidly disappear (Jones et al., 1991;unpublished results). On the other hand, subplate cells inthe sheep continue to stain for fetuin throughout most ofgestation (Saunders et al., 1992). This suggests either thatcells of the true subplate rapidly disappear in Monodelphis
(perhaps at the time of arrival of thalamic fibers) or thatfetuin expression is down-regulated early.
Is there a waiting period in Monodelphis?
Lund and Mustari (1977), who used degeneration meth-ods to study thalamocortical innervation in the rat, did notsee thalamic axons invading the cortical plate until about3 days after their arrival below it. This led to the concept ofthe waiting period, which is such a striking feature incarnivores (Shatz and Luskin, 1986; Ghosh and Shatz,1992) and primates (Rakic, 1977; Kostovic and Rakic,1990). However, Catalano et al. (1991), by using moresensitive axon-tracing techniques in the rat, showed notonly that thalamic axons arrive some 2–3 days beforeLund and Mustari (1977) estimated but also that someaxons and side branches immediately grow into the lowest,most mature part of the cortical plate. Much subsequentwork on rodents has confirmed that there is some early,
Fig. 16. A–G: Back labelling of cells in dorsal thalamus aftercortical DiI crystal placements at P10 (A,B,D,E) and P11 (C,F,G). Aand D show bisbenzimide counterstaining (A) and fluorescently la-belled axons (D; a mixture of corticofugal and thalamocortical) from acoronal section at the level of a crystal implantation in the dorsolateralcortex of the P10 animal (asterisk in D). B and E are a similar matchedpair of an adjacent section from the same specimen showing thelabelled fibers passing in densely packed fascicles through the internalcapsule (IC). The diencephalic portion of the bundle is also seen, withaxons extending through the ventral thalamus (VT) and ending in acluster of labelled cell bodies in the dorsal thalamus (DT). The dashedline marks the quite distinct boundary between dorsal and ventral
divisions of the thalamus. C: Similar example at P11 in which crystalswere implanted in the dorsolateral cortex on both sides of the brain.Both sides of the diencephalon are seen, with the two bundles of fibersrunning up to large groups of back-labelled cells in the dorsalthalamus. Note again the way in which the pattern of labelling of cellbodies and neuropil defines a sharp boundary between dorsal andventral thalamus (arrow and dashed line). F and G are matchedfluorescence (F) and counterstained images (G) of higher power viewsof the axons and back-labelled cell group in the left thalamus. Anarrow indicates the boundary of ventral and dorsal thalamus. Scalebars 5 250 µm in A (also applies to B–E), 100 µm in F (also appliesto G).
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 509

slow ingrowth (Kageyama and Robertson, 1993; Miller etal., 1993; Molnar and Blakemore, 1995; Catalano et al.,1996; Molnar et al., 1998). Nevertheless, there does seemto be a special relationship between thalamic axons andthe subplate, even in rodents. Axons grow preferentiallyinto and along the subplate layer (Bicknese et al., 1994),creating a dense plexus within it (see, e.g., Molnar et al.,1998), and they form functional synapses on subplate cells(Higashi et al., 1996). Indeed, in vitro experiments suggestthat the cortical plate does not express normal growth-permissive properties until some 3 days after thalamicfibers arrive (Gotz et al., 1992; Molnar and Blakemore,1995).
In Monodelphis, on the other hand, we see no evidence ofsubstantial accumulation of thalamic fibers in a presumedsubplate layer. The vast majority of axons appear to growquite quickly into and through the deep layers of the cortex(see Fig. 7), adopting the rather disorganized, roughlyradial pattern of ingrowth typical of the later stages ofinvasion in rats and mice (Kageyama and Robertson, 1993;Molnar and Blakemore, 1995; Catalano et al., 1996; Molnaret al., 1998), hamster (Naegele et al., 1988; Miller et al.,1993), and kitten (Ghosh and Shatz, 1992).
It seems likely that the wholesale, precocious invasion ofthe cortex in Monodelphis (as in the tammar wallaby;
Mark and Marrotte, 1992; Marotte et al., 1997) reflects therapid maturation of the infragranular layers and the lossor change in characteristics of the original subplate. If theexpression of fetuin in subplate neurons (Saunders et al.,1992) relates to their function as a temporary target forthalamic axons (see, e.g., Allendoerfer and Shatz, 1994),then its rapid disappearance in marsupials may explainwhy there is no obvious waiting period. Birth datingstudies are needed to determine whether the original,early-born preplate cells of the subplate actually die ormerely down-regulate fetuin in Monodelphis.
Figure 18 summarizes schematically the similaritiesand differences between rat and Monodelphis. The mostobvious difference is the age, relative to birth, at whichthalamic axons arrive. The other is the lack of a distinctsubplate by this stage in Monodelphis and the fact thatmost thalamic fibers extend quite quickly into the layer ofloosely packed, relatively mature cells below the thindense cortical plate.
Early projections into the thalamusin Monodelphis
In eutherian mammals, neurons of the thalamic reticu-lar nucleus (in the ventral thalamus) project into the
Fig. 17. Topographic relationship between the cortical surface andthe volume of the thalamus demonstrated with multiple crystalplacements in the cortex at P13. Three carbocyanine dye crystals (DiI,DiA, and DiI) were placed at roughly 750-µm intervals in a parasagit-tal row along the convexity of the right hemisphere. Another threecrystals (also DiI, DiA, and DiI) were implanted in a coronal row,running from lateral to dorsal convexities, roughly in the middle of theleft hemisphere. Five weeks of incubation at room temperatureallowed full anterograde and retrograde diffusion of dyes. Becausethalamic fibers have already arrived over the entire hemisphere byP13, the crystal placements reveal both corticofugal and thalamocorti-cal axons linking cortex and diencephalon. A: Fluorescence photomicro-graph of a 100-µm-thick horizontal section (rostral is up) that wasmade by triple exposure using rhodamine, fluorescein, and ultravioletfilters. The labelled axons from the parasagittal row of crystals in theright hemisphere form a broad horizontal fan in the telencephalon,running down toward the internal capsule: Those from the coronal rowin the left hemisphere form a somewhat narrower bundle in thishorizontal plane (although it is more extensive in the orthogonalplane). Those cells in the dorsal thalamus (within the box in A) thatare back labelled from the middle crystal of each row appear yellow-
green (DiA), whereas the flanking groups (DiI) appear more orange.With the quite close spacing of crystals used in this small brain and thelong incubation, there is considerable overlap of the labelled groups(as well as some ‘‘bleeding’’ between dye signals during triple expo-sure). However, the sequence of three colored groups can be discernedon both sides and is unequivocal in the original material. B: Enlargedview of the thalamic region within the outlined box in A. In the rightthalamus, the three crystals have labelled distinct, separate blocks ofthalamic cells (a–c, indicated by three arrows at right) that aredisplaced from one another along an oblique, rostrolateral-to-caudomedial axis. The most anterolateral group (c), which waslabelled from the most caudal cortical crystal, is quite elongated,forming a slab running orthogonal to the axis along which the threegroups are displaced. The most rostral cortical crystal labelled themost caudomedial block of thalamic cells (a). On the right side, thecoronal row of cortical crystals has labelled a more-or-less continuousslab of cells (axis indicated by arrow at left). Serial reconstruction ofthe entire array of axons showed that the most anterior cells in thisthalamic slab (a) were labelled from the most lateral cortical site,whereas the most posterior cells (c) were connected to the dorsalcortical site. Scale bars 5 300 µm in A, 50 µm in B.
510 Z. MOLNAR ET AL.

dorsal thalamus before thalamocortical axons have grownout of the diencephalon (Mitrofanis and Baker, 1993;Mitrofanis and Guillery, 1993; Adams and Baker, 1995;Metin and Godement, 1996). In addition, cells in theprimitive internal capsule, called the perireticular groupby Mitrofanis (1992), also project to the dorsal thalamus atearly stages (Metin and Godement, 1996; Cordery et al.,1997). The situation seems very similar in Monodelphis:The distribution of cells that are back labelled from thethalamus at P2–P5 is very similar to that seen in ratsaround E14 (Molnar and Blakemore, 1995; Cordery et al.,1996, 1997). At corresponding ages in the hamster, perire-ticular cells projecting into the thalamus lie exclusivelyunder the medial ganglionic eminence (Metin and Gode-ment, 1996). However, in Monodelphis, thalamopetalperireticular cells extend across the entire width of the
ganglionic eminence, as in rat and mouse (Molnar et al.,1995; Cordery et al., 1997).
The role of the early projections into the dorsal thalamusfrom the thalamic reticular nucleus and perireticular cellsremains to be clarified, but it is conceivable that they areinvolved in the guidance of thalamocortical axons in theproximal portion of the pathway through the ventraldiencephalon and nascent internal capsule. Monodelphiscould provide a useful model for their study.
Targets of early corticofugal projections
The exact patterns of growth and termination of corticofu-gal projections, both from the preplate and from neurons ofthe true infragranular layers, are rather uncertain. Re-cently, Clasca et al. (1995) have suggested that axons from
Fig. 18. Maturation of cortical lamination and in-growth of tha-lamic fibers summarized on schematic coronal sections of the leftoccipital cortex of rat (at E12, E16, E19, and birth, i.e., E21–E22) andof Monodelphis (at birth, P8, P10, and P15). At E12 in rat and at P0 inMonodelphis, the first postmitotic cells migrate to the outer edge of thecerebral wall to form the preplate, which is subsequently split intomarginal zone (the future layer I) and subplate by the arrival ofneurons of the true cortical plate. When thalamocortical fibers arrivein the rat (at E16), there is a distinct subplate, and the cortical plateconsists almost entirely of tightly packed, immature neurons (thedense cortical plate). At the equivalent stage in Monodelphis (P7–P8),
the subplate cannot be distinguished from the already mature,postmigratory cells of the lower layers. In the rat, thalamic fibersaccumulate within the subplate between about E16 and E19, althoughsome axons and side branches penetrate the lowest part of the corticalplate. In Monodelphis, most thalamic fibers pass immediately into thequite thick, mature, lower layers of the cortical plate, where many ofthem branch. In neither species is there significant axonal invasion ofthe dense cortical plate, which is always relatively thinner in Monodel-phis. PP, preplate; IZ, intermediate zone; VZ, ventricular zone; SVZ,subventricular zone; MZ, marginal zone; CP, cortical plate; DCP, densecortical plate; SP, subplate; V, layer V; VI, layer VI.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 511

the subplate and layer VI grow very slowly within thewhite matter of the telencephalon of the ferret. However,in the rat, Schreyer and Jones (1982) and Jones et al.(1982) were able to trace corticothalamic connections fromlayer VI to the ventral diencephalon by E19. At E16, dyeplacements in the rat ventral thalamus produce substan-tial back labelling in the cortical subplate and some inlayer VI; however, a small crystal a short distance away inthe dorsal thalamus does not back label cells in thecerebral wall at that stage (Molnar, 1994, 1998). Crandalland Caviness (1984) commented on the very sparse den-sity of corticofugal fibers within the thalamus after injec-tion of horseradish peroxidase (HRP) into the embryonicmouse cortex at E16. Thus, it seems likely that, in rodents,corticofugal projections do not reach the relay nuclei of thedorsal thalamus itself at early stages of development butmay pause in the ventral diencephalon, perhaps in theregion of the thalamic reticular nucleus (see, e.g., Shatzand Rakic, 1981; Miller et al., 1993; Molnar and Blake-more, 1995; Miller and Woolsey, 1996). In the ferret, thefirst corticofugal axons to penetrate the body of the dorsalthalamus (sometime after thalamic axons reach the cor-tex) arise from layer V: The usual corticothalamic projec-tion from layer VI does not enter until sometime later(Clasca et al., 1995).
A similar picture is seen in Monodelphis: At P10 andP13, after thalamic axons have reached the entire cortex,crystals placed ventrally in the diencephalon label cells inthe lower layers of the cortex, but placements high in thedorsal thalamus do not label cells in the cerebral wall. Inthe tammar wallaby, too, thalamic fibers appear to reachthe cortex long before the first cortical axons reach thethalamus (Sheng et al., 1991; Marotte et al., 1997). Thereis increasing evidence that corticofugal projections slowdown as they grow through the internal capsule andaccumulate in the ventral diencephalon. Subplate projec-tions might never enter the dorsal thalamus itself in manymammalian species.
A striking feature of the adult marsupial brain is theabsence of a corpus callosum, the anterior commissurebeing the equivalent forebrain commissure (for references,see Shang et al., 1997). Our results show that axonscontributing to the anterior commissure are evident at anearly stage, just a few days after the first corticofugaloutgrowth. They appear after about P5 as a small bundlethat diverges from the main array of descending axons inthe basal telencephalon and heads toward the putativeanterior commissure. Layer V is the main source of theearly projections into the anterior commissure. It is notuntil about P10 that the major brainstem projection(corticotectal, corticopontine, and corticospinal axons) isseen diverging in the region of the internal capsule to formthe cerebral peduncle. The mechanisms that are respon-sible for the separation of these axon populations areunknown.
Which axons reach the cortex first?
Recently, Metin and Godement (1996) described a popu-lation of cells in the lateral ganglionic eminence that couldbe back labelled from the cortex as early as E13.5 in themouse and E11.5 in the hamster—before the arrival ofthalamic axons. In the rat, however, corticopetal projec-tions from perireticular cells below the ganglionic emi-nence are not seen until about E17, after thalamic fibershave reached the cortex (Adams and Baker, 1995; Cordery
et al., 1996, 1997). In Monodelphis, we observed occasionalcells under the ganglionic eminence that were back la-belled from the cortex as early as P5 (corresponding toE14–E16 in the rat). However, in these cases, the dyeplacements were so lateral that they might have labelledprojections to the perirhinal cortex.
Early topography of thalamocorticalconnectivity
The basic topography of the interconnections betweenthe volume of the thalamus and the surface of the cortex,as determined with rows of separate carbocyanine dyes inthe cortex after the arrival of thalamic fibers, is verysimple and is remarkably similar to that in rodents (see,e.g., Blakemore and Molnar, 1990; Catalano et al., 1995;Molnar and Blakemore, 1995). The fiber bundles that arelabelled by each crystal remain discrete and orderedthroughout the pathway, terminating in groups of tha-lamic cells distributed in an ordered pattern throughoutthe volume of the dorsal thalamus, implying that the fiberarray undergoes a single rotation in the basal diencepha-lon. Thus, the entire thalamus initially projects continu-ously across the cortex, without regard to future nucleardivisions. This raises the possibility that rather simplemechanisms determine the initial topography of thalamicaxon targeting and that these mechanisms are shared bymammalian species as divergent as rodents and marsupi-als.
CONCLUSIONS
Our study shows that Monodelphis domestica has abasic pattern of cortical development very similar to thatof rodents, except for the relatively rapid maturation of theinfragranular layers and the wholesale early invasion bythalamic axons. The period equivalent to E13–P2 in therat occurs entirely postnatally in Monodelphis (roughlyP0–P11). The general similarity of the algorithm of thala-mocortical development to that in eutherian mammals,combined with its precocious birth, makes Monodelphis apotentially very valuable species for investigations of themechanisms of development.
ACKNOWLEDGMENTS
We are indebted to Laurence Waters, Lorraine Chapell,and William Hinkes for their help with photography. Theexcellent animal care of David Jacobs and Karen Lomas-ney is gratefully acknowledged. Z.M. was an MRC Train-ing Fellow.
LITERATURE CITED
Adam, E., K.M. Dziegielewska, N.R. Saunders, and U. Schumacher (1993)Neuraminic acid specific lectins as markers of early cortical plateneurons. Int. J. Dev. Neurosci. 11:451–460.
Adams, N.C. and G.E. Baker (1995) Cells of the perireticular nucleusproject to the developing neocortex of the rat. J. Comp. Neurol.359:613–626.
Allendoerfer, K.L. and C.J. Shatz (1994) The subplate, a transient neocorti-cal structure: Its role in the development of connections betweenthalamus and cortex. Annu. Rev. Neurosci. 17:185–218.
Bicknese, A.R., A.M. Sheppard, D.D.M. O’Leary, and A.L. Pearlman (1994)Thalamocortical axons extend along a chondroitin sulfate proteoglycan-enriched pathway coincident with the neocortical subplate and distinctfrom the efferent path. J. Neurosci. 14:3500–3510.
512 Z. MOLNAR ET AL.

Blakemore, C. and Z. Molnar (1990) Factors involved in the establishmentof specific interconnections between thalamus and cerebral cortex. ColdSpring Harbor Symp. Quant. Biol. 55:491–504.
Catalano, S., R.T. Robertson, and H.P. Killackey (1991) Early ingrowth ofthalamocortical afferents to the neocortex of the prenatal rat. Proc.Natl. Acad. Sci. USA 88:2999–3003.
Catalano, S.M., R.T. Robertson, and H.P. Killackey (1996) Individual axonmorphology and thalamocortical topography in developing rat somato-sensory cortex. J. Comp. Neurol. 367:36–53.
Clasca, F., A. Angelucci, and M. Sur (1995) Layer-specific programs ofdevelopment in neocortical projection neurons. Proc. Natl. Acad. Sci.USA 92:11145–11149.
Coggeshall, R.E. (1964) A study of diencephalic development in the albinorat. J. Comp. Neurol. 122:241–269.
Cordery, P., Z. Molnar, and C. Blakemore (1996) Origin of the firstprojections reaching the internal capsule and thalamus during embry-onic development. Brain Res. Assoc. Abstr. 13:52.
Cordery, P., C. Blakemore, and Z. Molnar (1997) Comparison of mammalianand reptilian pallium during development. Soc. Neurosci. Abstr. 23:75.
Crandall, J.E. and V.S. Caviness, Jr. (1984) Axon strata of the cerebral wallin the embryonic mice. Dev. Brain Res. 14:185–195.
DeCarlos, J.A. and D.D.M. O’Leary (1992) Growth and targeting ofsubplate axons and establishment of major cortical pathways. J.Neurosci. 12:1194–1211.
Erzurumlu, R.S. and S. Jhaveri (1992) Emergence of connectivity in theembryonic rat parietal cortex. Cerebral Cortex 2:336–352.
Fadem, B.H., G.L. Trupin, E. Maliniak, J.L. VandeBerg, and V. Hayssen(1982) Care and breeding of the gray, short-tailed opossum (Monodel-phis domestica). Lab. Animal Sci. 32:405–409.
Ghosh, A. and C.J. Shatz (1992) Pathfinding and target selection bydeveloping geniculocortical axons. J. Neurosci. 12:39–55.
Godement, P., J. Vanselow, S. Thanos, and F. Bonhoeffer (1987) A study indeveloping visual systems with a new method of staining neurones andtheir processes in fixed tissue. Development 101:697–713.
Gotz, M., N. Novak, M. Bastmeyer, and J. Bolz (1992) Membrane boundmolecules in the rat cerebral cortex regulate thalamic innervation.Development 116:507–519.
Graves, J.A. (1991) Mammalian genome evolution: new clues from compari-sons of eutherians, marsupials and monotremes. Comp. Biochem.Physiol. A 99:5–11.
Harman, A.M., N.J. Eastough, L.D. Beazley (1995) Development of thevisual cortex in a wallaby-phylogenetic implications. Brain Behav. Evol.45:138–152.
Higashi, S., Z. Molnar, T. Kurotani, H. Inokawa, and K. Toyama (1996)Functional thalamocortical connections develop during embryonic pe-riod in the rat: An optical recording study. Soc. Neurosci. Abstr. 22:1976.
Jones, E.G. (1985) The Thalamus. New York: Plenum Press.Jones, S.E., D.L. Christie, K.M Dziegielewska, L.A. Hinds, and N.R.
Saunders (1991) Developmental profile of a fetuin-like glycoprotein inneocortex, cerebrospinal fluid and plasma of post-natal tammar wallaby(Macropus eugenii). Anat. Embryol. Berlin 183:313–320.
Jones, E.G., D.J. Schreyer, and S.P. Wise (1982) Growth and maturation ofthe rat corticospinal tract. Prog. Brain Res. 57:361–379.
Kageyama, G.H. and R.T. Robertson (1993) Development of geniculocorticalprojections to visual cortex in rat; evidence for early ingrowth andsynaptogenesis. J. Comp. Neurol. 335:123–148.
Kostovic, I. and P. Rakic (1990) Developmental history of the transientsubplate zone in the visual and somatosensory cortex of the macaquemonkey and human brain. J. Comp. Neurol. 297:441–470.
Lund, R.D. and M.J. Mustari (1977) Development of the geniculocorticalpathway in rats. J. Comp. Neurol. 173:289–305.
Luskin, M.B. and C.J. Shatz (1985) Studies of the earliest-generated cells ofthe cat’s visual cortex: Cogeneration of subplate and marginal zones. J.Neurosci. 5:1062–1075.
Mark, R. (1997) Hugh Tyndale-Biscoe and the developmental neurobiologyof vision at the ANU. In N.R. Saunders and L. Hinds (eds): MarsupialBiology: Recent Research, New Perspectives. Sydney: University ofNew South Wales Press, pp. 312–326.
Mark, R.F. and L.R. Marotte (1992) Australian marsupials as models for thedeveloping mammalian visual system. Trends Neurosci. 15:51–57.
Marotte, L.R., C.A. Leamey, and P.M.E. Waite (1997) Timecourse ofdevelopment of the wallaby trigeminal pathway: III. Thalamocorticaland corticothalamic projections J. Comp. Neurol. 387:194–214.
McConnell, S.K., A. Ghosh, and C.J. Shatz (1989) Subplate neurons pioneerthe first axon pathway from the cerebral cortex. Science 245:978–982.
Metin, C. and P. Godement (1996) The ganglionic eminence may be anintermediate target for corticofugal and thalamocortical axons. J.Neurosci. 16:3219–3235.
Miller, B. and T.A. Woolsey (1996) The development of corticothalamicprojections in the mouse somatosensory system. Soc. Neurosci. Abstr.22:1012.
Miller, B., L. Chou, and B.L. Finlay (1993) The early development ofthalamocortical and corticothalamic projections. J. Comp. Neurol.335:16–41.
Mitrofanis, J. (1992) Patterns of antigenic expression in the thalamicreticular nucleus of developing rats. J. Comp. Neurol. 320:161–181.
Mitrofanis, J. and G.E. Baker (1993) Development of the thalamic reticularand perireticular nuclei in rats and their relationship to the course ofgrowing corticofugal and corticopetal axons. J. Comp. Neurol. 338:575–587.
Mitrofanis, J. and R.W. Guillery (1993) New views of the thalamic reticularnucleus in the adult and developing brain. Trends Neurosci. 16:240–245.
Møllgard, K., Y. Balslev, L.R. Christensen, T. Moos, O.B.F. Terkelsen, andN.R. Saunders (1994) Barrier systems and growth factors in thedeveloping brain. In H.C. Lou, G. Griesen, and J. Falck Larsen (eds):Brain Lesions in the Newborn: Alfred Benzon Symposium 37. Copenha-gen: Munksgaard, pp. 45–56.
Molnar, Z. (1994) Multiple mechanisms in the establishment of thalamocor-tical innervation (Ph.D. thesis). Oxford: University of Oxford.
Molnar, Z. (1998) Development of Thalamocortical Connections. Neurosci-ence Intelligence Unit Series. Austin, TX: R.G. Landes Company,Bioscience Publishers.
Molnar, Z. and C. Blakemore (1990) Relationship of corticofugal andcorticopetal projections in the prenatal establishment of projectionsfrom thalamic nuclei to the specific cortical areas of the rat. J. Physiol.430:104P.
Molnar, Z. and C. Blakemore (1995) How do thalamic axons find their wayto the cortex? Trends Neurosci. 18:389–397.
Molnar, Z., L. Krubitzer, J.E. Nelson, and C. Blakemore (1995) Develop-ment of thalamocortical innervation in the marsupial northern nativecat (Dasyurus hallucatus). Eur. J. Neurosci. 8(Suppl.):78.
Molnar, Z., R. Adams, C. Blakemore (1998) Mechanisms underlying theearly establishment of thalamocortical connections in the rat. J.Neurosci. (in press).
Naegele, J.R., S. Jhaveri, and G.E. Schneider (1988) Sharpening oftopographical projections and maturation of geniculocortical axonarbors in the hamster. J. Comp. Neurol. 277:593–607.
Nicholls, J.G. and N.R. Saunders (1996) Regeneration of immature mamma-lian spinal cord after injury. Trends Neurosci. 19:229–234.
Nicholls, J.G., R.R. Stewart, S.D. Erulkar, and N.R. Saunders (1990)Reflexes, fictive respiration and cell division in the brain and spinalcord of the newborn opossum, Monodelphis domestica, isolated andmaintained in vitro. J. Exp. Biol. 152:1–15.
Rakic, P. (1977) Prenatal development of the visual system in the rhesusmonkey. Phil. Trans. R. Soc. London B Biol. Sci. 278:245–260.
Renfree, M.B. and C.H. Tyndale-Biscoe (1978) Manipulation of marsupialembryos and pouch young. In J.C. Daniel (ed): Methods in MammalianReproduction. London: Academic Press, pp. 307–331.
Reynolds, M.L., and N.R. Saunders (1988) Differentiation of the neocortex.In The Developing Marsupial. Models for Biomedical Research. Eds.C.H. Tyndale-Biscoe and P. Janssens. Springer-Verlag, Berlin pp.101–116.
Reynolds, M.L., M.E. Cavanagh, K.M. Dziegielewska, L.A. Hinds, N.R.Saunders, and C.H. Tyndale-Biscoe (1985) Postnatal development ofthe telencephalon of the tammar wallaby (Macropus eugenii). Anaccessible model of neocortical differentiation. Anat. Embryol. 173:81–94.
Rose, J.E. (1942) The ontogenetic development of the rabbit’s diencephalon.J. Comp. Neurol. 77:61–129.
Sanderson, K.J. and W.L. Weller (1990) Gradients of neurogenesis inpossum neocortex. Dev. Brain Res. 55:269–274.
Saunders, N.R. (1997) Marsupials as models for studies of development andregeneration of the central nervous system. In N. Saunders and L.Hinds (eds): Marsupial Biology: Recent Research, New Perspectives.Sydney: University of New South Wales Press, pp. 382–410.
Saunders, N.R., E. Adams, M. Reader, and K. Møllgard (1989) Monodelphisdomestica (gray short-tailed opossum): An accessible model for studiesof early neocortical development. Anat. Embryol. 180:227–236.
EARLY THALAMOCORTICAL CONNECTIONS IN MONODELPHIS 513

Saunders, N.R., M.D. Habgood, R.A. Ward, and M.L. Reynolds (1992)Origin and fate of fetuin-containing neurons in the developing neocor-tex of the fetal sheep. Anat. Embryol. 186:477–486.
Schreyer, D.J. and E.G. Jones (1982) Growth and target finding by axons ofthe corticospinal tract in prenatal and postnatal rats. Neuroscience7:1837–1853.
Shang, F., K.W.S. Ashwell, L.R. Marotte, and P.M.E. Waite (1997) Develop-ment of commissural neurons in the wallaby (Macropus eugenii). J.Comp. Neurol. 387:507–523.
Shatz, C.J. and M.B. Luskin (1986) Relationship between the geniculocorti-cal afferents and their cortical target cells during development of thecat’s primary visual cortex. J. Neurosci. 6:3655–3668.
Shatz, C.J. and P. Rakic (1981) The genesis of efferent connections from thevisual cortex of the fetal rhesus monkey. J. Comp. Neurol. 196:287–307.
Shatz, C.J., A. Ghosh, S.K. McConnell, K.L. Allendoerfer, E. Friauf, and A.Antonini (1990) Pioneer neurons and target selection in cerebralcortical development. Cold Spring Harbor Symp. Quant. Biol. 55:469–480.
Sheng, X.-M., L.R. Marotte, and R.F. Mark (1990) Development of connec-
tions to and from the visual cortex in the wallaby (Macropus eugenii). J.Comp. Neurol. 300:196–210.
Sheng, X.-M., L.R. Marotte, and R.F. Mark (1991) Development of thelaminar distribution of thalamocortical axons and corticothalamic cellbodies in the visual cortex of the wallaby. J. Comp. Neurol. 307:17–38.
Stewart, R.R., D.-J. Zou, J.M. Treherne, K. Møllgard, N.R. Saunders, andJ.G. Nicholls (1991) The intact central nervous system of the newbornopossum in long term culture: Fine structure and GABA-mediatedinhibition of electrical activity. J. Exp. Biol. 161:25–41.
Takahashi, T., R.S. Nowakowski, and V.S. Caviness (1995) The cell cycle ofthe pseudostratified ventricular epithelium of the embryonic murinecerebral wall. J. Neurosci. 15:6046–6057.
Valverde, F., M.V. Facal-Valverde, M. Santacana, and M. Heredia (1989)Development and differentiation of early generated cells of sublayerVIb in the somatosensory cortex of the rat: A correlated Golgi andautoradiographic study. J. Comp. Neurol. 290:118–140.
Varga, Z.M., J. Fernandez, S. Blackshaw, K.J. Muller, W.B. Adams, and J.G.Nicholls (1996) Neurite outgrowth through lesions of neonatal opossumspinal cord in culture. J. Comp. Neurol. 366:1–13.
514 Z. MOLNAR ET AL.