determinants of spider species richness in coastal dunes along a gradient of mediterraneity
TRANSCRIPT

Determinants of spider species richness in coastaldunes along a gradient of mediterraneity
JOSE C. CARVALHO,1 PEDRO CARDOSO,2,3 LUIS C. CRESPO,3 SERGIOHENRIQUES,4 RUI CARVALHO4 and PEDRO GOMES1 1CBMA – Molecular and
Environmental Biology Centre, Department of Biology, University of Minho, Braga, Portugal, 2Smithsonian Institution,
National Museum of Natural History, Washington, DC, USA, 3Azorean Biodiversity Group–CITA-A, University of Azores,
Angra do Heroısmo, Portugal and 4Department of Biology, University of Evora, Evora, Portugal
Abstract. 1. The Iberian Peninsula is one of the most relevant areas in terms of spe-cies richness, rarity and endemism in the Mediterranean Basin. Using spiders as amodel, we studied the relative importance of environmental and non-environmentalspatial variation along a gradient of mediterraneity on the western coast of the Ibe-rian Peninsula.2. We performed a spatially explicit analysis to evaluate the contribution of pure
environmental and pure spatial effects and their shared influence on spider speciesrichness by variation partitioning and principal coordinates of neighbour matrices(PCNM). We dissect the spatial variation of species richness into additive scale-spe-cific models through PCNM analysis and estimate the relative importance of envi-ronmental variables for each model.3. Variation partitioning revealed that 72.8% of species richness variation could
be explained by the environmental (climate and dune stability) and spatial variables.Most of this variation (51.3%) corresponds to the environmental spatially structuredcomponent.4. The influence of environmental variables was scale-dependent. At the broadest
scales, the latitudinal-climatic gradient was the dominant factor that influenced spi-der species richness variation. At the finest scale, stability of the dune system was themost important component.5. In conclusion, spider species richness patterns were determined mostly by spa-
tially structured environmental variation. More importantly, this study shows thatdifferent environmental factors act at multiple scales. Hence, our results reinforce theimportance of the incorporation of both local and broad-scale factors when examin-ing species richness patterns.
Key words. Araneae, grey dunes, habitat stability, latitudinal gradient, scaledependence, spatial analysis.
Introduction
The Iberian Peninsula is among the most relevant regions interms of species richness, rarity and endemism in theMediterra-
nean Basin due to its high climatic and physiographic complex-ity and its role as refugia during the Pleistocene glacial period
(Gomez & Lunt, 2007). Understanding the determinants of spe-
cies richness is crucial for the conservation of biodiversity andecosystem management (Bestelmeyer et al., 2003). The anthro-pogenic pressure in the Mediterranean ecological systems, such
as coastal dunes, reinforces the urgency to acquire an under-standing of species richness patterns and their determinants, inorder to develop efficient conservation strategies (Carboni et al.,
2009).The distribution patterns of species are determined by a com-
bination of environmental and spatial processes (Borcard et al.,
1992). Therefore, to understand the determinants of species
Correspondence: Jose Carlos Carvalho, CBMA – Molecular
and Environmental Biology Centre, Department of Biology, Uni-
versity of Minho, Campus de Gualtar, 4710-057 Braga, Portugal.
E-mail: [email protected]
Insect Conservation and Diversity (2012) 5, 127–137 doi: 10.1111/j.1752-4598.2011.00139.x
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society 127

richness variation it is necessary to disentangle the effects of envi-ronmental and spatial variables. The explanatory capacity of
non-spatial structured environmental variation is a measure ofthe importance of local environmental factors. Spatial variationmay be decomposed into pure spatial variation and environmen-
tal spatially structured variation. The pure spatial componentmay be related to historic factors that left a spatial signature,such as dispersal ⁄colonisation events, or to unmeasured environ-
mental variables, while environmental spatially structured varia-tion is determined by environmental variables that arethemselves spatially structured (Borcard et al., 1992; Borcard &
Legendre, 1994).Moreover, patterns in species richness are the result of a com-
bination of several mechanisms acting at multiple scales (Rah-bek & Graves, 2001; Whittaker et al., 2001). Climate has been
cited frequently as a major correlate of species richness patternsat broad-scale geographic gradients (Hawkins et al., 2003). Atfiner scales, the structural components of the habitat, such as
heterogeneity, vegetation structure and substrate have beenrecognised as important local determinants of species richness(Tews et al., 2004). Hence, understanding species richness pat-
terns in a region depends on our ability to decompose its varia-tion into different processes acting at multiple scales (Borcardet al., 2004; Laliberte et al., 2009).The present study focuses on richness patterns of spiders in
interior fixed dunes, known as grey dunes according to theCORINE biotope classification (Natura 2000), along theAtlantic coast of Portugal. Mediterranean coastal dunes are one
of most threatened ecological systems by human activities inEurope, such as tourism and urbanisation (Meulen & Salman,1996). Hence, determining the factors that drive species richness
patterns in this environmentally sensitive region is of crucialimportance for the conservation of its biodiversity.We chose spiders as a model group because they are abun-
dant, species rich, easy to sample and because they are at the topof invertebrate food chains, hence, are likely to play an impor-tant role in shaping terrestrial arthropod communities (Wise,1993). Their occurrence is frequently related to environmental
factors such as vegetation structure and soil humidity (Bonteet al., 2002; Entling et al., 2007; Jimenez-Valverde & Lobo,2007; Finch et al., 2008). Spiders are known to respond sensi-
tively to environmental changes and anthropogenic impacts,which makes them suitable for bioindication studies (Marcet al., 1999; Cardoso et al., 2010). In coastal habitats, in particu-
lar, spiders respond to habitat productivity, heterogeneity andstability (Bonte et al., 2004a).The first aim of this study was to evaluate the relative effects
of environmental and spatial variables on local species richness
of spiders in coastal dunes. Because of the strong climatic gradi-ent (Eurosiberian–Mediterranean) at the Atlantic coast of Por-tugal, we hypothesise that spatially structured environmental
variables are potentially important determinants of spider spe-cies richness. To test this hypothesis, we performed a spatiallyexplicit analysis to evaluate the contribution of pure environ-
mental and pure spatial effects and their shared influence on spi-der species richness.The second aim of this investigation was to assess the
scale-specific importance of environmental variables on species
richness. We hypothesise that species richness is determined bydifferent processes acting at multiple scales: at larger scales, cli-
mate or climatic induced changes on vegetation structure shouldbe more important; at finer scales, local factors, such as the sta-bility of the dune system, which is dependent on the local eolian
dynamics, should play a major role. To test this hypothesis, wedissect the spatial variation of species richness into additivescale-specificmodels through principal coordinates of neighbour
matrices (PCNM) analysis and estimate the relative importanceof environmental variables for eachmodel.
Materials and methods
Study area
This study was carried out in 18 sites located in grey dunes(CORINE habitat code 2130) along a total distance of 445 km
of the Portuguese Atlantic coast (Fig. 1). This area comprisestwo distinct biogeographic regions, the Northern coast isincluded in the Eurosiberian region, while the Centre and South
are include in the Mediterranean region (Costa et al., 1998;Rivas-Martınez et al., 2002). We defined three bioclimatic zonesin order to stratify our sampling scheme along the gradient ofmediterraneity (see Rivas-Martınez &Rivas-Saenz, 2009): Ther-
motemperate, Mesomediterranean and Thermomediterranean(Table 1). The average distance among the three sets of samplingsites was 15.5 km (SD = 10.9), 46.0 km (SD = 28.8) and
23.4 km (SD = 17.8) for the Thermotemperate, Mesomediter-ranean and Thermomediterranean bioclimatic zones, respec-tively.
Spider sampling
A standardised and optimised sampling protocol forMediter-ranean spiders was developed by Cardoso et al. (2008a,b, 2009)and Cardoso (2009). This methodology proved to be rapid, rig-
orous and efficient. This protocol is based on semi-quantitativesampling, using different collecting methods (e.g. pitfall, sweep-ing, beating and hand collecting). A combination of different
methods is necessary to achieve a reliable inventory in habitatswith complex vegetation structure (Scharff et al., 2003; Jimenez-Valverde & Lobo, 2005; Cardoso et al., 2008a,b). However, due
to the poor structuring of vegetation of the sampled dunes in thisinvestigation, we decided to adapt this protocol and used onlypitfall trapping. This methodology is inherently biased to catchepigeal species. However, due to the vegetation characteristics of
the sampling locations, the epigeal spiders may allow a betterdistinction than web-building aerial species between differentareas. It should also be noted that this methodology was previ-
ously used in grey dunes and enabled a clear assessment of theregional and local variations of spider richness and assemblagecomposition (Bonte et al., 2003). By doing this, we assume that
our sampling target is the epigeal spider guild and not the entirecommunity of spiders.The pitfall traps (cups of 8 cm diameter · 12 cm depth) were
filledwith ethylene glycol diluted inwater (1 ⁄1), with a few drops
128 Jose C. Carvalho et al.
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

of detergent to break surface tension and were covered with asquare cork plate placed 2 cm above ground to prevent dryingout.At each site a square sampling plot of 100 · 100 m wide
(1 ha) was delimited. At each plot, we placed 12 groups of fourpitfall traps (arranged in a 5 m side square), totalizing 48 trapsper sampling station. The 12 groups were distributed in three
rows parallel to the sea. The distance between two successivegroups was 20 m. Each group of four pitfalls was considered asa sample for the purpose of estimating the richness and com-
pleteness of the inventory. Traps were left in the field for2 weeks. Then the biological material was collected in individ-ual bags and transported to the laboratory for species sortingand identification. The sampling took place in 2008 and 2009,
during May and June, when spider species richness and abun-dance are highest in Mediterranean ecosystems (Cardoso et al.,2007).
Three important sources of error are common to many eco-logical studies (see Longino et al., 2002; Scharff et al., 2003):spatial effects, phenological effects and methodological effects.
Fig. 1. Map of the study area with the
location of the sampling sites. Sampling
sites were divided into three sets according
to the thermoclimatic belts recognised for
the region, from North to South, Thermo-
temperate, Mesomediterranean and Ther-
momediterranean.
Table 1. Location, bioclimatic zone (Tt, Thermotemperate; Mm, Mesomediterranean; Tm, Thermomediterranean) and environmental
predictors of the studied sites, PCclimate corresponds to the site scores of the first component obtained by principal component analysis
(PCA) carried on a matrix of climatic variables; PC-1vegetation and PC-2vegetation were obtained by PCA on a vegetation structure variables
matrix; dune stability was obtained by a detrended correspondence analysis on a matrix of coverages of indicator plants.
Site Code
Bioclimatic
zone
Latitude
(N)
Longitude
(W) PCclimate PC-1vegetation PC-2vegetation
Dune
stability
Area
(ha)
Width
(m)
Amorosa AMO Tt 41�39¢24 8�49¢26 )0.949 0.22 )0.02 0.14 37 192
Carruagem CAR Tt 41�36¢14 8�48¢22 )0.905 0.46 0.07 )0.09 30 183
Belinho BEL Tt 41�35¢17 8�48¢14 )0.922 0.16 0.33 )0.15 13 158
Esposende ESP Tt 41�32¢57 8�47¢24 )0.854 0.39 0.26 0.18 8 160
Apulia APU Tt 41�28¢35 8�46¢28 )0.826 0.65 )0.02 )0.07 9 188
Mindelo MIN Tt 41�19¢09 8�44¢15 )0.783 )0.26 )0.38 0.22 25 300
Furadouro FUR Mm 40�51¢40 8�40¢38 )0.197 0.61 )0.19 0.16 29 304
Torreira TOR Mm 40�47¢12 8�42¢10 )0.172 0.35 )0.06 )0.37 159 343
Sao Jacinto SJA Mm 40�43¢45 8�43¢18 )0.294 0.47 )0.04 )0.21 195 767
Mira MIR Mm 40�26¢17 8�48¢24 )0.187 )0.09 )0.39 0.37 47 208
Quiaios QUI Mm 40�22¢31 8�49¢40 0.015 )0.11 )0.02 )0.26 84 129
Leirosa LEI Mm 40�03¢52 8�53¢08 0.310 )0.23 0.13 )0.32 79 216
Osso da Baleia OBA Mm 40�00¢04 8�54¢51 0.323 )0.44 0.11 )0.32 58 52
Santo Andre STA Tm 38�05¢37 8�48¢18 1.073 )0.51 )0.02 0.03 22 120
Areias Brancas ABR Tm 38�03¢33 8�49¢09 1.035 )0.43 0.02 0.03 12 87
Lagoa da Sancha SAN Tm 38�01¢36 8�50¢01 1.093 )0.50 )0.04 0.00 21 111
Sao Torpes STO Tm 37�54¢43 8�47¢51 1.110 )0.32 0.17 0.33 95 480
Almograve ALM Tm 37�38¢29 8�48¢15 1.130 )0.43 0.09 0.23 38 442
Determinants of spider species richness 129
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

Any of these may cause an inflation of the number of speciesinhabiting a given site. These may be vagrants from other habi-
tat types surrounding the studied site (spatial effects), character-istic of other seasons outside the sampled period (phenologicaleffects) or typical from microhabitats not targeted by the
employed methodology (methodological effects). We tried tolimit the first and second by concentrating a high trap density ina short period, probably avoiding the dispersal season of most
species (see Bonte et al., 2004b) and the third by discarding occa-sional catches (singletons and doubletons) of web-building aerialspiders (Linyphiidae, Theridiidae andAraneidae).
The inventory completeness was assessed by calculating theratio of observed richness to several richness estimators availablein the EstimateS 8.2 package (Colwell, 2009): Chao1, Chao2,Jack1, Jack2, Boot andMMMeans. Calculations were based on
1000 randomizations of the species · 12 samples datamatrix foreach site. The observed species richness values were obtained bypooling the 12 samples for each site. These values were used in
the subsequent analyses.
Local environmental variables
Several climatic variables were extracted from an Iberiandatabase with a 100 m resolution (Ninyerola et al., 2005): mean
annual precipitation, precipitation of the least rainy month, pre-cipitation of the most rainy month, mean annual temperature,mean temperature of the coldest month, mean temperature of
the hottest month and summer drought, i.e. the number ofmonths where the mean precipitation is less than twice the meantemperature. The values of these variables were extracted for the
18 cells representing sampling sites. Due to the high correlationamong the climatic variables we combined them into a singlefactor by principal component analysis (first eigenvalue =
7.839; second eigenvalue = 0.882), which explained almost89% of the variance (Table 1). This factor represents an increas-ing gradient of temperature and the period of summer droughtand a decreasing gradient of precipitation from North to South
(which we refer as PCclimate), thus representing a gradient ofmediterraneity. This climatic gradient was strongly correlatedwith latitude (r = 0.983,P < 0.001).
The vegetation structure and stability of the dune system areimportant factors known to influence spider assemblages (Bonteet al., 2003, 2004a). Therefore, for each sampling site we esti-
mated the following vegetation structure variables: % cover ofbare sand, % cover of litter, % cover of mosses and lichens, %cover of herbaceous vegetation (divided in three classes:<5 cm;5–20 cm; >20 cm) and % cover by shrubs. To estimate cover
we used a photoplot methodology (Elzinga et al., 2001). At eachsampling site, 24 squares of 1 m2 were delimited (two squaresper each group of four pitfall traps). Each square was photo-
graphed, in high resolution, with a digital camera at 1.5 m high.With the aid of a computer, a grid of 20 interceptions was super-imposed over each photograph and the number of hits on the
vegetation classes was noted. By dividing the number of hits foreach variable by the total number of interceptions, we obtainedthe percentage of coverage. Finally, the obtained values were
averaged for the entire sampling site.
A principal component analysis was carried out on thesquared-root transformed vegetation variables, in order to
avoid collinearity. The first two factors (PC-1vegetation and PC-2vegetation) accounted for 82.8% of the variance (eigen-values = 0.170 and 0.035, respectively) and were retained for
further analysis. The first component represents a gradient fromdunes with high sand coverage to dunes dominated by mossesand lichens. The second component contrasts dunes with higher
coverage of herbaceous plants with dunes with higher coverageby shrubs (Table 1).The stability of the dune system was inferred by estimating
the coverage of three plant functional types associated to dunedynamics, following Garcıa-Mora et al. (1999) and Martınezet al. (2006). Plants of Type I includemainly winter annuals withsoft and thin leaves, showing no presumed adaptations to the
dune environment. Plants of Type II consist mostly of perennialswith a below-ground spreading root network and leaves withpresumed adaptations to coastal environmental stress (leaf suc-
culence, pubescence or reinforced cuticles). Type III includesplants mostly capable of withstanding sand burial and of beingdispersed by seawater. Type II and Type III plants were found
to be more abundant in unstable dunes while Type I plants wereassociated with relatively more stable dunes. The relative coverof each plant functional type was determined by the samemethod used for the vegetation structure.
A detrended correspondence analysis was performed in orderto summarise this information into a single factor. We retainedthe first axis, that represents a gradient from dunes with higher
proportion of type III plants (negative scores) to dunes withhigher coverage of type I plants (positive scores) (Table 1). Thus,this factor corresponds to a gradient of stability and we refer to
it as dune stability further in this paper.The area of each interior dune fragment was measured in a
geographic information system using digitised orthophotomaps.
The width of the interior dune was also measured as the perpen-dicular distance from the crest of the frontal dune to pine planta-tions, agricultural fields or heathland depending on each site,passing through the centre of each sampling plot.
Finally, we constructed the final matrix of environmentalpredictors with the following variables: PCclimate, PC-1vegetation,PC-2vegetation, dune stability, dune area and dune width
(Table 1).
Spatial variables
Spatial dependence in the study areawasmodelled using prin-cipal coordinates of neighbour matrices (PCNM, Borcard &
Legendre, 2002; Borcard et al., 2004; Dray et al., 2006). Thismethod allows the detection of spatial trends across awide rangeof scales. First, a Euclidean distance matrix among sampling
locations was calculated based on the UTM coordinates of sam-pling sites. Then, a truncated connectivity matrix was built. Thetruncation value is usually the largest value in the minimum
spanning tree linking the sites (Legendre & Legendre, 1998). Inour case, we used themaximum distance between two successivesampling locations, since our study area is aligned with the axis
of latitude and, therefore, distances could be described in one
130 Jose C. Carvalho et al.
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

dimension only. In order to allow the PCNM analysis to detectfeatures smaller than the widest gap between neighbouring sites,
two supplementary objects were added to the coordinatesmatrix. These objects were removed after PCNM analysis (Bor-card&Legendre, 2002; Borcard et al., 2004).
Then a principal coordinate analysis (PCoA) was performedon the truncated connectivity matrix to extract the eigenvaluesand eigenvectors. Ten eigenvectors associated with positive
eigenvalues were obtained (PCNM variables). The eigenvectorswith large eigenvalues correspond to large-scale spatial struc-tures, whereas the ones with small eigenvalues describe local spa-
tial structures. All of the PCNM eigenvectors are orthogonaland therefore uncorrelated independent variables and were usedas spatial variables in regression analysis (Fig. 2a). PCNMeigenvectors were created using the package spacemakeR (Dray
et al., 2006) for the R language (R Development Core Team2009).
Statistical analysis
The relationship between individual environmental variablesand spider species richness was assessed by simple linear regres-sion, in order to show the individual explanatory capacity of
each predictor. Then, we ran forward selection, using multipleregression, on the environmental dataset to select those variables
with a significant (P < 0.05 after 999 random permutations)contribution to explaining variation in species richness (Blanchetet al., 2008). This procedure avoids inflated Type I error and the
overestimation of the amount of explained variance. The areaand the width variables were standardised to 0 mean and 1 SDto avoid scale effects, prior to the analysis.
The same forward selection procedurewas used to select thosePCNM eigenvectors with a significant contribution to explain-ing species richness (P < 0.05 after 999 random permutations).
This procedure identified three significant PCNM variables(PCNM 1, 3 and 10), corresponding to two spatial scales(broad-scale = PCNM1 and PCNM3 and fine-scale =PCNM10).
Both sets of selected variables (environmental and PCNMeigenvectors) were used in variation partitioning to quantify theproportion of the variation in species richness explained by
purely environmental, purely spatial and spatially structuredenvironmental effects (Borcard et al., 1992). Partitioning wasdone through a series of partial regression analyses (Fig. 2b).
The R2 values were adjusted to account for the number of sam-pling sites and explanatory variables, as unadjusted R2 valuesare biased (Peres-Neto et al., 2006).
(a)
(b)
(c)
Fig. 2. Diagram showing the statistical pro-
cedures step-by-step. dij, distance between
sites i and j; max, maximum distance
between two successive sites; PCoA, princi-
pal coordinate analysis. The PCNMs 1, 3
and 10 were plotted against geographic dis-
tance, thus revealing that PCNM 1 and 3
represent variation at a much broader scale
than PCNM10.
Determinants of spider species richness 131
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137
In order to access the scale-specific importance of environ-mental variables on species richness, we constructed two spatial
models (broad-scale = PCNM1 + PCNM3 and fine-sca-le = PCNM10) by fitting species richness values against eachforward selected PCNM eigenvector, using linear regression.
The fitted values were then used in a variation partitioning pro-cedure involving the forward selected environmental variablesto estimate their relative importance at each specific scale
(Fig. 2c). A detailed description of this procedure is given byLaliberte et al. (2009).The analyses were performed in the R statistical language
(R Development Core Team 2009) using the packages vegan(Oksanen et al., 2007) for variation partitioning and packfor(Dray, 2005) for the forward selection of explanatory variables.
Results
A total of 7379 adult spiders belonging to 101 species and 22families were collected in the survey. The mean observed speciesrichness was 19.4 per location (SD = 5.3), with a minimum of
eight species in Leirosa and a maximum of 28 in Mindelo.Abundance ranged from 32 individuals in Leirosa to 1469 indi-viduals in Torreira. The completeness values for each site variedfor each estimator, but were in general high (Table 2). More-
over, the different estimators were in general highly correlatedwith the observed species richness values (r-values: Chao1 =0.63; Chao2 = 0.69; Jack1 = 0.94; Jack2 = 0.81; Boot =
0.99; MMmeans = 0.74; all P-values <0.01). Therefore, we
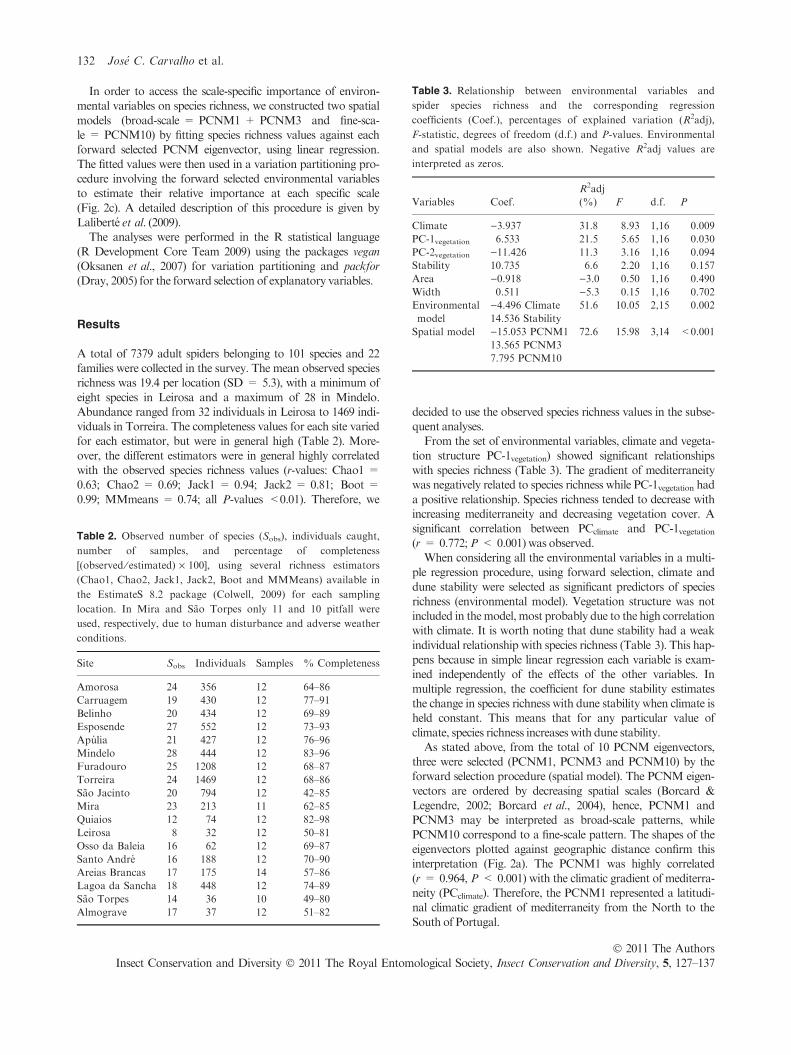
decided to use the observed species richness values in the subse-quent analyses.From the set of environmental variables, climate and vegeta-
tion structure PC-1vegetation) showed significant relationshipswith species richness (Table 3). The gradient of mediterraneitywas negatively related to species richness while PC-1vegetation had
a positive relationship. Species richness tended to decrease withincreasing mediterraneity and decreasing vegetation cover. Asignificant correlation between PCclimate and PC-1vegetation(r = 0.772;P < 0.001) was observed.When considering all the environmental variables in a multi-
ple regression procedure, using forward selection, climate and
dune stability were selected as significant predictors of speciesrichness (environmental model). Vegetation structure was notincluded in themodel, most probably due to the high correlationwith climate. It is worth noting that dune stability had a weak
individual relationship with species richness (Table 3). This hap-pens because in simple linear regression each variable is exam-ined independently of the effects of the other variables. In
multiple regression, the coefficient for dune stability estimatesthe change in species richness with dune stability when climate isheld constant. This means that for any particular value of
climate, species richness increases with dune stability.As stated above, from the total of 10 PCNM eigenvectors,
three were selected (PCNM1, PCNM3 and PCNM10) by theforward selection procedure (spatial model). The PCNM eigen-
vectors are ordered by decreasing spatial scales (Borcard &Legendre, 2002; Borcard et al., 2004), hence, PCNM1 andPCNM3 may be interpreted as broad-scale patterns, while
PCNM10 correspond to a fine-scale pattern. The shapes of theeigenvectors plotted against geographic distance confirm thisinterpretation (Fig. 2a). The PCNM1 was highly correlated
(r = 0.964, P < 0.001) with the climatic gradient of mediterra-neity (PCclimate). Therefore, the PCNM1 represented a latitudi-nal climatic gradient of mediterraneity from the North to the
South of Portugal.
Table 2. Observed number of species (Sobs), individuals caught,
number of samples, and percentage of completeness
[(observed ⁄ estimated) · 100], using several richness estimators
(Chao1, Chao2, Jack1, Jack2, Boot and MMMeans) available in
the EstimateS 8.2 package (Colwell, 2009) for each sampling
location. In Mira and Sao Torpes only 11 and 10 pitfall were
used, respectively, due to human disturbance and adverse weather
conditions.
Site Sobs Individuals Samples % Completeness
Amorosa 24 356 12 64–86
Carruagem 19 430 12 77–91
Belinho 20 434 12 69–89
Esposende 27 552 12 73–93
Apulia 21 427 12 76–96
Mindelo 28 444 12 83–96
Furadouro 25 1208 12 68–87
Torreira 24 1469 12 68–86
Sao Jacinto 20 794 12 42–85
Mira 23 213 11 62–85
Quiaios 12 74 12 82–98
Leirosa 8 32 12 50–81
Osso da Baleia 16 62 12 69–87
Santo Andre 16 188 12 70–90
Areias Brancas 17 175 14 57–86
Lagoa da Sancha 18 448 12 74–89
Sao Torpes 14 36 10 49–80
Almograve 17 37 12 51–82
Table 3. Relationship between environmental variables and
spider species richness and the corresponding regression
coefficients (Coef.), percentages of explained variation (R2adj),
F-statistic, degrees of freedom (d.f.) and P-values. Environmental
and spatial models are also shown. Negative R2adj values are
interpreted as zeros.
Variables Coef.
R2adj
(%) F d.f. P
Climate )3.937 31.8 8.93 1,16 0.009
PC-1vegetation 6.533 21.5 5.65 1,16 0.030
PC-2vegetation )11.426 11.3 3.16 1,16 0.094
Stability 10.735 6.6 2.20 1,16 0.157
Area )0.918 )3.0 0.50 1,16 0.490
Width 0.511 )5.3 0.15 1,16 0.702
Environmental
model
)4.496 Climate
14.536 Stability
51.6 10.05 2,15 0.002
Spatial model )15.053 PCNM1
13.565 PCNM3
7.795 PCNM10
72.6 15.98 3,14 <0.001
132 Jose C. Carvalho et al.
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

Variation partitioning revealed that 72.9% of the species rich-
ness variation could be explained by the environmental and thespatial variables recorded (Fig. 3). Most of this variation(51.3%) corresponded to the environmental spatially structured
component, while 21.3%of the variationwas due to pure spatialeffects and the pure environmental component was negligible.By further dissecting the variation explained by the environ-
mental component (51.6%), either alone or in conjunction with
spatial variation, into the unique contributions of each variable,we found that PCclimate was by far more important than dunestability (Fig. 3). It should be noted that negative effects between
the intersection of PCclimate with stability were observed. This isindicative of synergistic effects, i.e. the explanatory variablestogether explained the response variable better than the sum of
their individual effects (Legendre & Legendre, 1998). In thiscase, this meant that higher species richness could be observed instabilised dunes with a particular climate.
Dissecting the variation explained by spatial variables(72.6%), either alone or in conjunction with environment, intothe unique contributions of broad-scale and fine-scale eigenvec-tors, we found that the broad-scale latitudinal-climate gradient
of mediterraneity (PCNM1 + PCNM3 = 65.0%) was moreimportant than fine-scale processes (PCNM10 = 12.8%)(Fig. 3).
The scale-specific additive spatial models allowed dissectingthe total spatial variation into the environmental spatially struc-tured and pure spatial effects at the broad (PCNM1 +
PCNM3) and fine scales (PCNM10). At the broadest scale, envi-ronmental spatially structured variation and pure spatial varia-tion had similar importance. At the fine scale (PCNM10),spatial variation alone becomesmore important (Fig. 4).
The analysis conducted to assess the scale-specific importanceof environmental variables on the observed species richness,showed that different processes acted at multiple scales (Fig. 5).
At the broad-scale (PCNM1 and PCNM3), the PC-1climate vari-able was the only factor that influences spider species richness,revealing the strong influence of climate on the spider speciesrichness variation. At the fine-scale (PCNM10) only the stability
variable influenced the variation of species richness.
Discussion
Understanding the determinants of species richness, especially in
hyper-diverse ecoregions such as the Mediterranean Basin, is offundamental importance for the conservation and sustainableuse of biological diversity (Gaston, 2000). The Mediterranean
climate is characterised by a pattern with hot and dry summersand mild and humid winters. Dryness conditions become moreextreme towards the South of the Mediterranean Basin, wheresummer drought can last for several months (Blondel et al.,
2010). Therefore, the influence of Mediterranean climate on therichness and composition patterns of several arthropods groups
Fig. 4. Relative influences of spatially structured environmental
variation (Environment \ Space) and space on the spider species
richness at broad (PCNM1 and PCNM3) and fine scales
(PCNM10).
Fig. 3. Venn diagram showing the results of the variation parti-
tioning procedure carried out on the forward selected environ-
mental and spatial (PCNM eigenvectors) variables.
Fig. 5. Venn diagrams showing the results of the variation parti-
tioning carried out on the environmental variables at broad
(PCNM1 and PCNM3) and fine scales (PCNM10).
Determinants of spider species richness 133
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

is not surprising (butterflies: Hortal et al., 2004; dung beetles:Cabrero-Sanudo & Lobo, 2006; solitary wasps: Gonzalez et al.,
2009; leaf beetles: Baselga & Jimenez-Valverde, 2007). In thisstudy, we investigated the determinants of the variation of spiderspecies richness in coastal dunes along the Atlantic coast of Por-
tugal, characterised by a gradient of increasing mediterraneityfrom North to South, and the relative importance of differentfactors and scales. Our results showed that the climatic gradient
and the stability of the dune system were significant predictorsof spider species richness variation. Species richness decreasedwith increasing mediterraneity at a broad-scale and instability of
the dune system at a fine-scale.The decrease in species richness with increasing mediterrane-
ity contradicts the well known latitudinal diversity gradient thatpredicts an increase of species richness from North to South in
the Holarctic (Willig et al., 2003). As the use of pitfall traps asthe single collecting method may influence the lack of collectionof some species inhabiting within the vegetation, the observed
latitudinal gradient can be even higher. This inverse latitudinaldiversity gradient seems to be related with the gradient of medi-terraneity. Climate is one of themain factors determining species
richness patterns (Hawkins et al., 2003). This was verified alsofor spiders (Finch et al., 2008), which display substantial sensi-tivity to climatic factors, such as temperature and humidity(Wise, 1993; Bonte et al., 2002). The western coast of the Iberian
Peninsula is a biogeographic crossroad for spiders, encompass-ing elements from different biogeographic origins. The Eurosi-berian species were restricted to the Northern part of the
Atlantic coast, that represented the Southernmost limit of theirgeographic distribution, but Wide-range and Mediterraneanspecies were distributed along the entire coast (Carvalho et al.,
2011a). Therefore, the extreme temperatures and aridity in theSouthern locations may be acting as a filter for Eurosiberianspecies, being responsible for a decrease in species richness.
The climatic gradient may have influenced spider species rich-ness patterns indirectly by inducing changes in vegetation struc-ture, which has been reported as one of the most importantfactors determining spider species richness (e.g. Jimenez-Valver-
de & Lobo, 2007; Batary et al., 2008; Lambeets et al., 2008;Mas et al., 2009). Moreover, vegetation coverage and habitatopenness ⁄dryness have been reported as important factors to
explain spider species distribution in sand habitats (Bonte et al.,2004a; Buchholz, 2010). In the present case, Northern dunes arecharacterised by higher coverage of mosses and lichens, while
the Southern dunes are dominated by open sand, reflecting anincrease in aridity from North to South. We argue that mossesand lichens may act as keystone structures in dunes, whichfavour a higher richness of spiders, as they provide refuges
against micro-climatic variation, structures for attaching a weband ambush prey, but other indirect effects may be related, suchas, for example, prey availability or reduced cannibalism (Uetz,
1991;Marc et al., 1999).Our study indicates the importance of the stability of the dune
system as a driver of spider species richness patterns. This result
is consistent with earlier studies that showed that spider speciesrichness is negatively influenced by eolian dynamics and thelinked instability of the dune systems, by acting as disturbance
factors (Bonte et al., 2004a, 2006). Similar results were also
found in other dynamic habitats, such as gravel banks (Lambe-ets et al., 2008), thus habitat stability seems to be an important
driving factor of richness patterns of spiders.The variation partitioning procedure revealed that spider spe-
cies richness patterns were mainly determined by spatially struc-
tured environmental variation. The pure environmentalfraction was negligible due to the strong effects of the environ-mental spatially structured gradient of mediterraneity over spi-
der species richness patterns. In addition to the sharedenvironment–space component, the pure spatial componentaccounted for a significant part of the variability in species rich-
ness. The effect of historical unaccounted-for factors may havecontributed to generate a spatial gradient independently ofenvironmental conditions. The distance to the European sourceof some spider species may lead to a decrease in the ratio of
immigration ⁄ colonisation, from the base (mainland) to theWestof the Iberian Peninsula, generating a peninsular effect (Simp-son, 1964). In the Iberian Peninsula, similar patterns were also
found for butterflies (Martın & Gurrea, 1990) and forest birds(Tellerıa & Santos, 1993). However, this effect can only bereflected at the large spatial scale defined by the PCNM1 and 3.
At the local scale, defined by the PCNM10, the pure spatialvariation component may contain some residual environmentalvariation due to unmeasured local environmental variables(Borcard & Legendre, 1994; Legendre et al., 2005). Several
studies have shown the strong influence of the landscape heter-ogeneity on local richness of spiders (Clough et al., 2005;Schmidt et al., 2008; Mas et al., 2009) and this is also likely to
happen in grey dunes. Since, the landscape context surroundingeach dune patch was not assessed in this study, it is likely thatits influence on local spider species richness may have been
included in the pure spatial or in the unexplained variationcomponents.A small proportion of the variation (about 27%) in spider
species richness was undetermined. Undoubtedly, this is partlydue to so-called stochastic processes related to the strongdynamics of coastal environments (Hesp &Martınez, 2007), butit may also include deterministic variation caused by unmea-
sured environmental variables, such as landscape heterogeneitysurrounding dune patches, or spatial variation at finer scalesthan the ones encompassed by our sampling design.
By dissecting the spatial structure of species richness into addi-tive scale-specific models, we found that climate and stabilityacted at different scales determining spider species richness pat-
terns. Other studies have also shown that spider species richnesspatterns are influenced by a combination of different factors act-ing at multiple scales (Mas et al., 2009). In our case, climateacted at broader scales (PCNM1 and PCNM3) whilst stability
acted at finer scales (PCNM10). Climate has been repeatedlypointed as one of the major drivers of broad-scale geographicpatterns of species richness (Hawkins et al., 2003). However, on
finer scales, other factors such as stability become more impor-tant as was shown byBonte et al. (2004a) and in our study. Con-sequently, in order to understand the species richness gradients it
is necessary to assess the scale-specific importance of differentprocesses (Laliberte et al., 2009;Mas et al., 2009).In another contribution, we found that spider assemblage
structure, i.e. beta diversity, along the western Atlantic coast of
134 Jose C. Carvalho et al.
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

the Iberian Peninsula, was determined by a combination offactors (climate, vegetation structure and area of the dune patch)
operating at broader-scales. Moreover, the stability of the dunesystem had no effect on assemblage composition (Carvalhoet al., 2011b). This means that climate promotes two different
processes, richness differences among sites and the turnover ofspecies, while the stability of the dune system only induces rich-ness differences, probably determined by the loss of eurytopic
species in the more dynamic dune systems (Bonte et al., 2004a,2006). The corollary of these findings is that different mecha-nisms, operating at multiple scales, determine different biodiver-
sity attributes (alpha and beta diversity).In conclusion, our study showed that spider richness patterns
along the strong climatic gradient of mediterraneity characteris-tic of the Atlantic coast of Portugal are determined mostly by
spatially structured environmental variation. Dissecting the spa-tial structure of species richness at multiple scales allowed us tofind that climate was the major driver at the broad-scale, whilst
stability of the dune system acts at fine-scale. Hence, our resultsreinforce the importance of the incorporation of both local andbroad-scale factors when examining species richness patterns.
Acknowledgements
We acknowledge Jorge Lobo, Dries Bonte and an anonymousreviewer for suggestions on a previous version of the manuscriptand to Jose Antonio Barrientos, Carmen Urones and Stano
Pekar for their help with the taxonomic identification of somespider specimens.J.C.C. expresses his gratitude to the Luso-American Founda-
tion for its support (FLAD C & T Links 2010 grant). P.C. issupported by the Portuguese Foundation for Science andTechnology (SFRH ⁄BPD ⁄40688 ⁄2007).
References
Baselga, A. & Jimenez-Valverde, A. (2007) Environmental and
geographical determinants of beta diversity of leaf beetles
(Coleoptera: Chrysomelidae) in the Iberian Peninsula. Ecologi-
cal Entomology, 32, 312–318.
Batary, P., Baldi, A., Samu, F., Szuts, T. & Erdos, S. (2008) Are
spiders reacting to local or landscape scale effects in Hungarian
pastures? Biological Conservation, 141, 2062–2070.
Bestelmeyer, B.T., Miller, J.R. & Wiens, J.A. (2003) Applying
species diversity theory to land management. Ecological Appli-
cations, 13, 1750–1761.
Blanchet, F.G., Legendre, P. & Borcard, B. (2008) Forward selec-
tion of explanatory variables. Ecology, 89, 2623–2632.
Blondel, J., Aronson, J., Bodiou, J.-Y. & Boeuf, G. (2010) The
Mediterranean Region, Biological Diversity Through Time and
Space. Oxford University Press, Oxford, UK.
Bonte, D., Baert, L. & Maelfait, J.-P. (2002) Spider assemblage
structure and stability in a heterogeneous coastal dune system
(Belgium). Journal of Arachnology, 30, 331–343.
Bonte, D., Criel, P., Van Thournout, I. & Maelfait, J.-P. (2003)
Regional and local variation of spider assemblages (Araneae)
from coastal grey dunes along the North Sea. Journal of Bioge-
ography, 30, 901–911.
Bonte, D., Criel, P., Vanhoutte, L., Van Thournout, I. & Mael-
fait, J.-P. (2004a) The importance of habitat productivity, sta-
bility and heterogeneity for spider species richness in coastal
grey dunes along the North Sea and its implications for conser-
vation. Biodiversity and Conservation, 13, 2119–2134.
Bonte, D., Lens, L. & Maelfait, J.-P. (2006) Sand dynamics in
coastal dune landscapes constrain diversity and life-history
characteristics of spiders. Journal of Applied Ecology, 43, 735–
747.
Bonte, D., Maelfait, J.-P. & Baert, L. (2004b) Determining spider
species richness in fragmented coastal dune habitats by extrap-
olation and estimation (Araneae). European Arachnology 2002
(ed. by F. Samu and C. Szinetar), pp. 189–197. Akaprint kft,
Budapest, Hungary.
Borcard, D. & Legendre, P. (1994) Environmental control and
spatial structure in ecological communities, an example using
oribatid mites (Acari, Oribatei). Environmental and Ecological
Statistics, 1, 37–61.
Borcard, D. & Legendre, P. (2002) All-scale spatial analysis of
ecological data by means of principal coordinates of neighbour
matrices. Ecological Modelling, 153, 51–68.
Borcard, D., Legendre, P., Avois-Jacquet, C. & Tuomisto, H.
(2004) Dissecting the spatial structure of ecological data at
multiple scales. Ecology, 85, 1826–1832.
Borcard, D., Legendre, P. & Drapeau, P. (1992) Partialling out
the spatial component of ecological variation. Ecology, 73,
1045–1055.
Buchholz, S. (2010) Ground spider assemblages as indicators for
habitat structure in inland sand ecosystems. Biodiversity and
Conservation, 19, 2565–2595.
Cabrero-Sanudo, F.J. & Lobo, J.M. (2006) Determinant variables
of Iberian Peninsula Aphodiinae diversity (Coleoptera, Scarab-
aeoidea, Aphodiidae). Journal of Biogeography, 33, 1021–1043.
Carboni, M., Carranza, M.L. & Acosta, A. (2009) Assessing con-
servation status on coastal dunes, a multiscale approach. Land-
scape and Urban Planning, 91, 17–25.
Cardoso, P. (2009) Standardization and optimization of arthro-
pod inventories-the case of Iberian spiders. Biodiversity and
Conservation, 18, 3949–3962.
Cardoso, P., Arnedo, M.A., Triantis, K.A. & Borges, P.A.V.
(2010) Drivers of diversity in Macaronesian spiders and the
role of species extinctions. Journal of Biogeography, 37, 1034–
1046.
Cardoso, P., Gaspar, C., Pereira, L.C., Silva, I., Henriques, S.S.,
Silva, R.R. & Sousa, P. (2008a) Assessing spider species rich-
ness and composition in Mediterranean cork oak forests. Acta
Oecologica, 33, 114–127.
Cardoso, P., Henriques, S.S., Gaspar, C., Crespo, L.C., Carv-
alho, R., Schmidt, J.B., Sousa, P. & Szuts, T. (2009) Species
richness and composition assessment of spiders in a Mediterra-
nean scrubland. Journal of Insect Conservation, 13, 45–55.
Cardoso, P., Scharff, N., Gaspar, C., Henriques, S.S., Carvalho,
R., Castro, P.H., Schmidt, J.B., Silva, I., Szuts, T., Castro, A.
& Crespo, L.C. (2008b) Rapid biodiversity assessment of spi-
ders (Araneae) using semi-quantitative sampling, a case study
in a Mediterranean forest. Insect Conservation and Diversity, 1,
71–84.
Cardoso, P., Silva, I., Oliveira, N.G. & Serrano, A.R.M. (2007)
Seasonality of spiders (Araneae) in Mediterranean ecosystems
and its implications in the optimum sampling period. Ecologi-
cal Entomology, 32, 516–526.
Carvalho, J.C., Cardoso, P., Crespo, L.C., Henriques, S., Carv-
alho, R. & Pedro Gomes, P. (2011a) Biogeographic patterns of
Determinants of spider species richness 135
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

spiders in coastal dunes along a gradient of mediterraneity.
Biodiversity and Conservation, DOI: 10.1007/s10531-011-0001-8.
Carvalho, J.C., Cardoso, P., Crespo, L.C., Henriques, S., Carv-
alho, R. & Pedro Gomes, P. (2011b) Determinants of beta
diversity of spiders in coastal dunes along a gradient of medi-
terraneity. Diversity and Distributions, 17, 225–234.
Clough, Y., Kruess, A., Kleijn, D. & Tscharntke, T. (2005) Spider
diversity in cereal fields: comparing factors at local, landscape
and regional scales. Journal of Biogeography, 32, 2007–2014.
Colwell, R.K. 2009. EstimateS: Statistical Estimation of Species
Richness and Shared Species from Samples. Version 8.2.
<http://viceroy.eeb.uconn.edu/estimates.> Persistent URL:
<http://purl.oclc.org/estimates>1st May 2010.
Costa, J.C., Aguiar, C., Capelo, J.H., Lousa, M. & Neto, C.
(1998) Biogeografia de Portugal Continental. Quercetea, 0, 5–
56.
Dray, S. (2005) packfor: Forward selection with multivariate Y by
permutation under reduced model. Laboratoire Biometrie et
Biologie Evolutive, Lyon. <http://pbil.univ-lyon1.fr/members/
dray/software.php>1st May 2010.
Dray, S., Legendre, P. & Peres-Neto, P.R. (2006) Spatial model-
ling, a comprehensive framework for principal coordinate anal-
ysis of neighbour matrices (PCNM). Ecological Modelling, 196,
483–493.
Elzinga, C.L., Salzer, D.W., Willoughby, J.W. & Gibbs, J.P.
(2001) Monitoring Plant and Animal Populations. Blackwell Sci-
ence Inc., Malden, Massachusetts.
Entling, W., Schmidt, M.H., Bacher, S., Brandl, R. & Nentwig,
W. (2007) Niche properties of Central European spiders, shad-
ing, moisture, and the evolution of the habitat niche. Global
Ecology and Biogeography, 16, 440–448.
Finch, O.-D., Blick, T. & Schuldt, A. (2008) Macroecological pat-
terns of spider species richness across Europe. Biodiversity and
Conservation, 17, 2849–2868.
Garcıa-Mora, M., Rosario, M., Gallego-Fernandez, J.B. & Gar-
cıa-Novo, F. (1999) Plant functional types in coastal foredunes
in relation to environmental stress and disturbance. Journal of
Vegetation Science, 10, 27–34.
Gaston, K.J. (2000) Global patterns in biodiversity. Nature, 405,
220–227.
Gomez, A. & Lunt, D.H. (2007) Refugia within refugia, patterns
of phylogeographic concordance in the Iberian Peninsula. Phy-
logeography of Southern European Refugia (ed. by S. Weiss and
N. Ferrand), pp. 155–188. Springer, Dordrecht, The Nether-
lands.
Gonzalez, J.A., Gayubo, S.F., Asıs, J.D. & Tormos, J. (2009)
Diversity and biogeographical significance of solitary wasps
(Chrysididae, Eumeninae, and Spheciformes) at the Arribes del
Duero Natural Park, Spain, their importance for insect diver-
sity conservation in the Mediterranean region. Environmental
Entomology, 38, 608–626.
Hawkins, B.A., Field, R., Cornell, H.V., Currie, D.J., Guegan,
J.-F., Kaufman, D.M., Kerr, J.T., Mittelbach, G.G., Obe-
rdorff, T., O’brien, E.M., Porter, E.E. & Turner, J.R.G. (2003)
Energy, water, and broad-scale geographic patterns of species
richness. Ecology, 84, 3105–3117.
Hesp, P.A. & Martınez, M.L. (2007) Disturbance processes and
dynamics in coastal dunes. Plant Disturbance Ecology (ed. by
E.A. Johnson and K. Miyanishi), pp. 215–247. Academic Press,
Burlington, Massachusetts.
Hortal, J., Garcia-Pereira, P. & Garcıa-Barros, E. (2004) Butter-
fly species richness in mainland Portugal, predictive models of
geographic distribution patterns. Ecography, 27, 68–82.
Jimenez-Valverde, A. & Lobo, J.M. (2005) Determining a com-
bined sampling procedure for reliable estimation of Araneidae
and Thomisidae assemblages (Arachnida, Araneae). The Jour-
nal of Arachnology, 33, 33–42.
Jimenez-Valverde, A. & Lobo, J.M. (2007) Determinants of local
spider (Araneidae and Thomisidae) species richness on a regio-
nal scale, climate and altitude vs. habitat structure. Ecological
Entomology, 32, 113–122.
Laliberte, E., Paquette, A., Legendre, P. & Bouchard, A. (2009)
Assessing the scale-specific importance of niches and other spa-
tial processes on beta diversity, a case study from a temperate
forest. Oecologia, 159, 377–388.
Lambeets, K., Hendrickx, F., Vanacker, S., Van Looy, K., Mael-
fait, J.-P. & Bonte, D. (2008) Assemblage structure and conser-
vation value of spiders and carabid beetles from restored
lowland river banks. Biodiversity and Conservation, 17, 3133–
3148.
Legendre, P., Borcard, D. & Peres-Neto, P.R. (2005) Analyzing
beta diversity, partitioning the spatial variation of community
composition data. Ecological Monographs, 75, 435–450.
Legendre, P. & Legendre, L. (1998) Numerical Ecology. Elsevier,
Amsterdam, The Netherlands.
Longino, J.T., Coddington, J. & Colwell, R.K. (2002) The ant
fauna of a tropical rain forest, estimating species richness three
different ways. Ecology, 83, 689–702.
Marc, P., Canard, A. & Ysnel, F. (1999) Spiders (Araneae) useful
for pest limitation and bioindication. Agriculture, Ecosystems &
Environment, 74, 229–273.
Martın, J. & Gurrea, P. (1990) The peninsular effect in Iberian
butterflies (Lepidoptera, Papilionoidea and Hesperioidea). Jour-
nal of Biogeography, 17, 85–96.
Martınez, M.L., Gallego-Fernandez, J.B., Garcıa-Franco, J.G.,
Moctezuma, C. & Jimenez, C.D. (2006) Assessment of coastal
dune vulnerability to natural and anthropogenic disturbances
along the Gulf of Mexico. Environmental Conservation, 33,
109–117.
Mas, E.D., Chust, G., Pretus, J.L. & Ribera, C. (2009) Spatial
modelling of spider biodiversity, matters of scale. Biodiversity
and Conservation, 18, 1719–2000.
Meulen, F. & Salman, A.H.P.M. (1996) Management of Mediter-
ranean coastal dunes. Ocean and Coastal Management, 30,
177–195.
Natura (2000) Interpretation Manual of European Union habitats.
Eur 15 ⁄ 2, Nature Prediction, Coastal Zones and Tourism. Euro-
pean Commission and Environment, Brussels, Belgium.
Ninyerola, M., Pons, X. & Roure, J.M. (2005) Atlas Climatico
Digital de la Penınsula Iberica. Metodologıa y aplicaciones en
bioclimatologıa y Geobotanica. Universidad Autonoma de Bar-
celona, Barcelona. <http://www.opengis.uab.es/wms/iberia/
mms/index.htm> 1st May 2010.
Oksanen, J., Kindt, R., Legendre, P. & O’Hara, R.B. (2007)
vegan: community ecology package. R package version 1.9-25.
<http://cran.r-project.org>1st May 2010.
Peres-Neto, P.R., Legendre, P., Dray, S. & Borcard, D. (2006)
Variation partitioning of species data matrices, estimation and
comparison of fractions. Ecology, 87, 2614–2625.
R Development Core Team (2009) R: A Language and Environ-
ment for Statistical Computing. R Foundation for Statistical
Computing, Vienna, Austria. ISBN 3-900051-07-0. <http://
www.r-project.org>1st May 2010.
Rahbek, C. & Graves, G.R. (2001) Multiscale assessment of pat-
terns of avian species richness. Proceedings of the National Acad-
emy of Sciences of the United States of America, 98, 4534–4539.
136 Jose C. Carvalho et al.
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137

Rivas-Martınez, S., Dıaz, T.E., Fernandez-Gonzalez, F., Izco, J.,
Lousa, M. & Penas, A. (2002) Vascular Plant Communities of
Spain and Portugal. Addenda to the Syntaxonomical checklist
of 2001. Itinera Geobotanica, 15, 5–432.
Rivas-Martınez, S. & Rivas-Saenz, S. (2009) Worldwide Bio-
climatic Classification System. Phytosociological Research
Center, Spain. <http://www.globalbioclimatics.org>1st May
2010.
Scharff, N., Coddington, J.A., Griswold, C.E., Hormiga, G. &
Bjørn, P.P. (2003) When to quit? Estimating spider species rich-
ness in a Northern European deciduous forest Journal of
Arachnology, 31, 246–273.
Schmidt, M.H., Thies, C., Nentwig, W. & Tscharntke, T. (2008)
Contrasting responses of arable spiders to the landscape matrix
at different spatial scales. Journal of Biogeography, 35, 157–
166.
Simpson, G.G. (1964) Species density of North American recent
mammals. Systematica Zoologica, 12, 57–73.
Tellerıa, J.L. & Santos, T. (1993) Distributional pattern of insec-
tivorous passerines in the Iberian forests: does abundance
decrease near the border? Journal of Biogeography, 20, 235–
240.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann,
M.C., Schwager, M. & Jeltsch, F. (2004) Animal species diver-
sity driven by habitat heterogeneity ⁄ diversity, the importance
of keystone structures. Journal of Biogeography, 31, 79–92.
Uetz, G.W. (1991) Habitat structure and spider foraging. Habitat
Structure, the Physical Arrangement of Objects in Space (ed. by
S.S. Bell, E.D. McCoy and H.R. Mushinsky), pp. 325–348.
Chapman and Hall, London, UK.
Whittaker, R.J., Willis, K.J. & Field, R. (2001) Scale and species
richness, towards a general, hierarchical theory of species diver-
sity. Journal of Biogeography, 28, 453–470.
Willig, M.R., Kaufman, D.M. & Stevens, R.D. (2003) Latitudinal
gradients of biodiversity, pattern, process, scale and synthesis.
Annual Review of Ecology and Systematics, 34, 273–309.
Wise, D.H. (1993) Spiders in Ecological Webs. Cambridge Univer-
sity Press, New York.
Accepted 7 February 2011
First published online 7 March 2011
Editor: Calvin Dytham
Associate editor: Jorge Lobo
Determinants of spider species richness 137
� 2011 The AuthorsInsect Conservation and Diversity � 2011 The Royal Entomological Society, Insect Conservation and Diversity, 5, 127–137