cyclic amp-adenosine pathway induces nitric oxide...
TRANSCRIPT

Cyclic AMP-Adenosine Pathway Induces Nitric Oxide Synthesis in Aortic Smooth Muscle Cells
Raghvendra K Dubey, Delbert G Gillespie, Edwin K Jackson
Abstract-The mam purpose of this mvestlgatlon was to evaluate whether the cychc AMP-adenosme pathway, le, the conversion of CAMP to AMP and, hence, to adenosme, 1s involved m the regulation of mtnc oxide (NO) synthesis by vascular smooth muscle cells (SMCs) Treatment of confluent monolayers of SMCs with adenosme, 2-chloroadenosme (stable analog of adenosme), and agents that elevate endogenous (SMC-derived) adenosme (EHNA and lodotubencldm) increased mtnte/mtrate (stable metabohtes of NO) levels m the medium and enhanced the conversion of “H-L-argmme to ‘H-L-cltrulhne by cytosohc extracts obtained from the pretreated SMCs The stlmulatory effects of adenosme were not mimicked by low (1 to 100 nmol/L) concentrations of CGS21680, an A,, receptor agomst, or CPA, a selective A, receptor agonist The stlmulatory effects of 2-chloroadenosme and EHNA plus lodotuberlcldm were slgmficantly inhibited by KF17837, a selective A, receptor antagomst, and by DPSPX, an AI/A, receptor antagonist, but not by DPCPX, a selective A, receptor antagonist DDA (adenylyl cyclase mhlbltor) and Rp-cyclic AMP (protein kmase A mhlbltor) did not block the effects of adenosme on NO synthesis Incubation of SMCs with exogenous cychc AMP, at concentrations previously shown to elevate levels of adenosme m the medium, also increased mtnte/mtrate levels and 3H-L-cltrulllne formation, and the effects of cychc AMP on NO synthesis were blocked by DPSPX and KF17837, but not by DPCPX These findings provide evidence that exogenous and SMC-derived adenosme mduce NO synthesis via A 2. receptors hnked to a pathway not mvolvmg adenylyl cyclase/protem kmase A Moreover, extracellular cychc AMP induces NO synthesis via conversion to adenosme and activation of A,, adenosme receptors The cychc AMP-adenosme pathway may be importantly involved m the vascular production of NO (Hypertension. 1998;31[part 2]:296-302.)
Key Words: cychc AMP n adenosme w nitric oxide w smooth muscle cells H A, adenosme receptors n vasculature
ascular endothehal as well as smooth muscle cells (SMCs) are known to synthesize mtrlc oxide (NO), a labile free
radical and intracellular messenger with a half-life of a few seconds I,* NO 1s synthesized by oxldatlon of the terminal guamdmo-nitrogen atoms of L-argmme by a family of en- zymes called NO synthase (NOS) ’ * Within the vessel wall, two distinct types of NOS have been identified The first 1~ a constltutlve Ca*+-dependent and calmodulm-dependent membrane-bound form that 1s present m endothehal cells and plays an important role m the dynamic control of vascular tone The second 1s an inducible Ca*+-independent form that IS mduced by endotoxm and some cytokmes and 1s present m vascular SMCs ’ * Endothehal cells contmuously generate low levels of NO, which maintain homeostasls m normal blood vessels I-? In contrast to endothehal cells, SMCs generate large amounts of NO m response to cytokmes and endotoxm
In SMCs, cyclic AMP and agents that increase cellular cyclic AMP levels (eg, forskolm and lsopreterenol) induce 1NOS expression and NO synthesis, as well as enhance cytokme- Induced NO synthesis m SMCs, by stablhzmg 1NOS mRNA 4mh Since adenosme stimulates cychc AMP levels m
vascular SMCs via A, receptors,’ we hypothesize that adeno- sine may induce NO synthesis m vascular SMCs and enhance cytokme induced NO synthesis (Fig 1)
Recent studies from our laboratory demonstrate that In vascular SMCs cychc AMP 1s an important determinant of adenosme production via a blochenucal mechanism we refer to as the cychc AMP-adenosme pathway (Fig 1) B This pathway mvolves the conversion of cychc AMP to AMP, and, hence, to adenosme by the enzymes phosphodlesterase (PDE) and S’- nucleotldase, respectively The cychc AMP-adenosme path- way may have both intracellular and extracellular arms, le. adenosme might be formed within the cell and transported to the extracellular space, or it might be formed directly m the extracellular space * We have recently shown that the mhlbl- tory effects of cychc AMP on SMC prollferatlon are blocked by A, adenoqme receptor antagomsts,8 thus lmplymg that the cychc AMP-adenosme pathway regulates SMC growth Since cychc AMP 1s a precursor for adenosmeX and adenosme may Induce NO synthesis, we hypothesize that the cychc AMP- adenosme pathway may be a potential pathway m regulating NO synthesis m vascular SMCs (Fig 1)
Recewed September 17, 1997, first dectsmn October 16, 1997, rews~on accepted October 29, 1997 From the Center for Pharmacology, Departments of Medmne (R K D , D G G ) and Pharmacology (E K J ), Umvenq of Plusburgh MedIcal Center,
Pmcburgh, Pa Correspondence to Dr Kaghvendra K Dubey, Center for Chmcal Pharmacology, Department of Medlcme, 623 Scalfe Hall, 200 Lothrop Street,
Unwerwy of Pittsburgh Medlcal Center, Ptttsburgh, PA 15213-2582 E-Mad dubey@novell2 dept-med putt edu 0 1998 American Heart Assoclatlon, Inc
296
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Dubey et al
Selected Abbreviations and Acronyms
BMC = basal medmm Eagle’s Fx:s = fetal calf Felum
DMEM = 11elbucco’s modified Eagle medmm HB\S -= Hanks balanced salt solutmn
1110 = lodotubencldm LPS = hpopolysacchatlde
MEM = mm,nnm~ essential medun NO = rutilc oxide
Accordingly, the aims of the present study were to deter-
mine whether: (1) exogenous adenosine induces NO synthesis
by SMCs; (2) specific adenosine receptor subtype(s) are in-
volved in inducing NO synthesis by SMCs; (3) SMC-derived
adenosine can effectively increase NO synthesis by SMCs; (4)
adenosine enhances LPS-induced NO synthesis by SMCs; and
(5) the cyclic AMFadenosine pathway induces NO synthesis
in SMCs.
Materials
Methods
Adenoune, 2-Lhlorondenoume (Cl-Ad), erythro-9-(2-hydroxy-3- nonyl) adenlne hydrochloride (EHNA), N”-mtro-L-algmlne methyl ecter (L-NAME), mtrate reductace (A~perglllus mger), hpopolysaccharlde (LPS), nlcotlnamlde-adenlne dlnucleotlde phosphate (NADI’H) and flawne ademne chnucleotlde (FAD) were purchased from Slgrna Chenucal Co. 2’,5’-Dldeoxyadenosme (DDA) was obtalned from Calblochem-Novabtochenl Corp. (R)- p-Adenoune 3’,.5-cychc phosphorothloate (Rp-CAMP), Nh- cyclopentyladenoslrl~ (CPA), 2-p-(2-carboxyethyl) phenethyl- nn~lllo-5’-N-ethylcnrboxamldo adenoslne hydrochloride (CGS21680), %cvclopentyl-1,3-dlpropylxanthme (DPCPX), lo- dotubexlcldm (IDO), and 1,3-dlpropyl-8-p-sulfophenylxanthlne (DPSPX) were pulchared from Research B~ochenucals lnterna- tlonal. Dulbecco’s mod&ied Eagle medium (DMEM), DMEM/F12 me&urn, nltrlte-nitrate free nunltnum etrentlal medium (MEM), Hanks balanced salt solutton (HBSS), pemcllhn, streptomycin,
Figure 1. Hypothesis: Cyclic AMP-adenosine pathway (ie, the conversion of cyclic AMP to AMP by phosphodiesterase [PDE] and hence to adenosine by 5’-nucleotidase [5’-NTJ) provides adenosine to the A2 receptor, which triggers the activation of inducible nitric oxide synthase via an adenylyl cyclase/protein kinase A dependent and/or independent pathway. Gs, stimulatoty G-protein; AC, adenylyl cyclase; ATP, adenosine triphosphate; CAMP, cyclic AMP; AMP, adenosine monophosphate; ADO, adenosine; Tr, nucleoside transporter; DPSPX, A,/A, receptor antagonist; KFl7837, a selec- tive A2 receptor antagonist; DDA, 2’,5’-dideoxyadenosine (adenylyl cyclase inhibitor); Rp-CAMP, protein kinase A inhibitor; INOS, inducible nitric oxide (NO) synthase.
0 25% trypsn-EDTA solwon, and all tissue culture wa*e were purchased from GIBCO Laboratorleq. Fetal calf Felum (FCS) wac obtamed from HyClone Lnbolatouet Inc. ‘H-L-argmme (cpeclfic activity 68 Cl/mmol) was purchased from Anxxsham Life Science 111~ KF17837 wa? genelou grft from Dr Funuo Suzuki, Kyowa Hakko Kogyo Co Ltd, Sunto, Shuzuoka, Japan. All other chenucalc used were of title culture 01 beqt grade avallable
Aortic Smooth Muscle Cell Culture Aortlc smooth muscle cells (SMCs) wele cultwed as explants from the awendmg thorauc aortx, obtained from ether-anesthetized male Sprague-Dawley rat5 (Charlet River, Wlhnmgton, Mass), after a nxdhne abdotnmal mclslon mcludmg the dlaphrngnl and as descllbed prewously by us.” SMC punlty \vas charactellzed by mm~m~ofluoles- cence stammg with smooth murcle specltic antlrmooth muxle alpha- actm monoclonal antlbodw and by molphologlc crlterla \peufic for wooth muscle a~ dewlbcd m detail plevloutly ” SMCF were pa\taged by trypwnzatlon, and cells between the 2nd and 31d pawge x\cre wed for NO blosynthew studies.
Treatment Protocols for NO Synthesis by SMCs SMCq were cultured m either 24-well tlqwe cnltule cl&es fool the rneawrement of mtrlte+mtrate production or m pert11 dxhes (100 tnrn dlarneter) for the cltrulllne astay When the cells reached confluence, the culture me&urn WRY arpnated and replaced with rerun-free MEM contalnmg 0.25% (wt/vol) of htty-acid free bovme serum albunun After overmght mcubatlon, the culture medun v ac replaced agam with flesh batal tnechun Eagles (BME) supplemented with albunnn and treated In the presence 01 absence of LPS (25 pg/mL) wth various combmatlont of other treatnxxx< as follows. (1) Cl-Ad, an analog of adenosme that 1~ not metnbohzed and mediates Its effects vta both A, and A, receptors,’ (2) adenosme; (3) CPA, an adenowe agomrt that mechates Its e&cts selectively wa A, leceptols,’ (4) CGS21680, an adenosme agomst that mediates Its effects seleuwely wa A,, receptors;’ (5) Cl-Ad plus DPCPX, a selective A, reLepto1 antagomst,’ (6) Cl-Ad plus DPSPX, an A,/A? adenowe receptor antagonIst,’ (7) Cl-Ad plw KFI 7837, a selective A, receptor antago- nest;” (8) EHNA, an mhlbltor of adenosme deamma\e,” (9) IDO, an mhlbltor of adenosme kmase, ,‘(’ (10) EHNA plus IDO; (1 1) DPSPX; (12) EHNA plus ID0 plu\ DPSPX, (13) DPCPX; (14) EHNA pluc ID0 plus DPCPX; (15) L-NAME, a NOS mhlbltol, (16) EHNA plus ID0 plus L-NAME; (17) KF17837; (18) EHNA plus ID0 plus KF17837; (19) cychc AMP, (20) cychc AMP plus DPSPX; (21) cychc AMP plus DPCPX; (22) cychc AMP plus KFl7837; (23) DDA, an mhlbltor of adenylyl cyclase;” (24) adenowle plur DDA; (25) EHNA plus ID0 plus DDA; (26) Rp-cAMP, a preferential mhlbltor of cychc AMP-dependent plotem kmase;” (27) adenosme plus Rp-CAMP, (28) EHNA plus IDO plus Rp-CAMP. For mtrlte/mtrate synthesis studies the cells were treated for 30 houl5, whereas, for cltrulhne assay f?x 20 hours.
Nitrite/Nitrate Analysis Nltrlte/Nltrate level% m the conditioned culture medun we1 r mea- sured ucmg the Grexs reagent as previously described.” Bllrfly, ahquotv (50 pL) of conditioned medun from confluent cells were collected and mcubated at loon, tetnpelature wth 25 PL of wbqtrate buffer (nmdazole 0 1 mol/L, NADPH 210 pnlol/L, flavme adenme dmucleot~de 3.8 /.unol/L) m the preteme of nitrate-reductase (14 mU enzyme/SO pL) from Aspergdlus nlger to convert NO, to NO, Total NO, was then analyzed by reactmg the wnplec with equal volume of Gelss reagenr (I’% sulfamlannde. 0 1% naphthalene-ethylene dlamme dlhydrochlonde m .5% H,PO,) for 4.5 nunutes at 27°C and nleatwmg absorbance at 540 nm spect~ophotometrlcall) Condltloned medium from untreated samples with no added Gelss reagent x\ere ured ac qalnple blanks. Atnounts of NO, III medium \vere e$tlmated from a standard curve of NaNO, obtamed by enzymatic convel$lon of NaNO, (O-32 pmol/L., >98’% convelqlon achieved; Merck) We did not attempt to differentiate between NO2 and NO, anloumtr, but rather enzymatuzally converted all NO, to NO, and therefore lepolt
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from
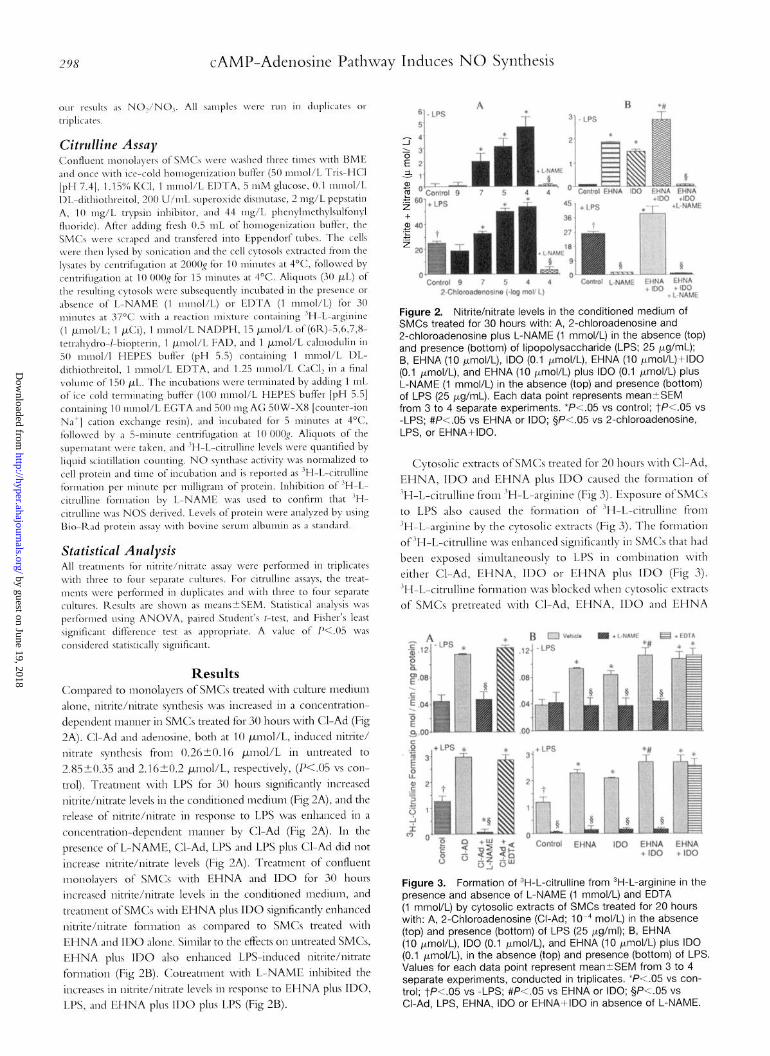
298 CAMP-Adenosine Pathway Induces NO Synthesis
our re&s as NOJNO,. All sampler were run in duplicates 01 triplicater.
Citrulline Assay Confluent monolayers of SMCt were wvhed three times with BME and once with ice-cold homogenization buffer (SO mmol/L Tris-HCI [pH 7.41, 1.15% K<:l, 1 mmol/L EDTA, 5 mM glucose, 0.1 mmol/L IIL-dithiothreitol, 200 U/mL superoxide dismutase, 2 mg/L pepstatin A, 10 mg/L trypsin inhibitor, and 44 mg/L phenyhnethylsulfonyl fluoride). After adding fresh 0.5 mL of homogenization buffu, the SMC? were scraped and transfered into Eppendorf tubes. The cells were then lysed by sonication and the cell cytosols extracted fi-om the lysates by centrifugation at 20002 for 10 minutes at 4’C, followed by centrif~lgation at 10 0002 for 15 minutes at 4°C. Aliquots (30 FL) of the rewlting cytosolq were subsequently incubated in the presence or absence of L-NAME (1 mmol/L) or EDTA (1 mmol/L) for 30 minutes at 37’C with a reaction mixture containing ‘H-L-arginine (1 ~~mol/L; I &i), 1 mmol/L NADPH, 15 pmol/L of ((iK)-5,6,7,X- tetrahydro-Cbiopterin, 1 p.mol/L FAD, and 1 +mol/L calmodulin in 50 mmol/l HEPES buffer (pH 5.5) containing 1 mmol/L DL- dithiothreitol, 1 mmol/L EDTA, and 1.25 mmol/L CaCl, iri a final volume of 150 PL. The incubations were terminated by adding 1 mL of ice cold terminating buffer (100 mmol/L HEI’ES buffer [pH 5.51 containjog 10 mmol/L EGTA and 500 mg AG SOW-XX [counter-ion Nail cation exchange resin), and incubated for 5 minutes at -1”C, followed by R 5-minute centrifugation at IO 000~~. Aliquon of the supernatant were taken, and ‘H-L-citrulline levels were quantified by liquid scintillation counting. NO synthase activity was normalized to cell pl-otein and time of incubation and is reported as ‘H-L-citrulline formation per minute per milligram of protein. Inhibition of ‘H-L- citrulline formation by L-NAME wns used to confirm that “H- citrulline was NOS derived. Levels of protein were analyzed by using Bio-Rad protein assay with bovine serum albumin as a standard.
Statistical Analysis All treatments for nitrite/nitrate assay were performed in triplicates with three to four separate cultures. For citrullinc assays, the treat-
ments \vere performed in duplicates and with three to four separate cultures. Results are shown as meansiSEM. Statistical analysis was performed using ANOVA, paired Student’s f-test, and Fisher’s least significant diff>rence teyt as appropriate. A value of I’<.05 was considered statistically significant.
Results Compared to monolayers of SMCs treated with culture lnedium
alone, nitrite/nitrate synthesis was increased in a concentration-
dependent manner in SMCs treated for 30 hours with C-Ad (Fig
2A). Cl-Ad and adenosine, both at 10 /~lol/L, induced nitrite/
nitrate synthesis from (3.26-+0.16 ~mol/L in untreated to
2.85kO.35 and 2.16+0.2 /~nol/L, respectively, (X.05 vs COW
trol). Treatment with LPS for 30 hours significantly increased
nitrite/nitrate levels in the conditioned medium (Fig 2A), and the
release of nitrite/nitrate in response to LPS was enhanced in a
concentratior-dependent manner by Cl-Ad (Fig 2A). In the
presence of L-NAME, Cl-Ad, LPS and LPS plus Cl-Ad did not
increase nitrite/nitrate levels (Fig 2A). Treatment of confluent
monolayers of SMCs with EHNA and ID0 for 30 hours
increased nitrite/nitrate levels in the conditioned medium, and
treatment of SMCs with EHNA plus ID0 significantly enhanced
nitrite/nitrate fornlation as compared to SMCs treated with
EHNA and ID0 alone, Similar to the effects on untreated SMCs,
EHNA plus ID0 also enhanced LB-induced nitrite/nitrate
formation (Fig 2B). Cotreatment with L-NAME inhibited the
increases in nitrite/nitrate levels in response to EHNA plus IDO,
LJ’S, and EHNA plus ID0 plus LPS (Fig 2B).
B
I.“.* EWJA 45 t *WI
+ LPS * rL t:nw
~- 4
Figure 2. Nitrite/nitrate levels In the conditioned medium of SMCs treated for 30 hours with: A, 2-chloroadenosine and 2-chloroadenosine plus L-NAME (1 mmol/L) in the absence (top) and presence (bottom) of lipopolysaccharide (LPS; 25 pg/mL); B, EHNA (10 kmol/L), ID0 (0.1 Fmol/L), EHNA (10 pmol/L)+IDO (0.1 pmol/L), and EHNA (10 wmol/L) plus ID0 (0.1 pmol/L) plus L-NAME (1 mmol/L) in the absence (top) and presence (bottom) of LPS (25 Kg/ml). Each data point represents meantSEM from 3 to 4 separate experiments. *P<.O5 vs control; tk.05 vs -LPS; #P<.O5 vs EHNA or IDO; §P<.O5 vs 2-chloroadenosine, LPS, or EHNA+IDO.
Cytosolic extracts of SMCs treated for 20 hours with Cl-Ad,
EHNA, ID0 and EHNA plus ID0 caused the formation of
“H-L-citrulline from ‘H-L-arginine (Fig 3). Exposure of SMCs
to LPS also caused the formation of ‘H-L-citrulline fi-om
“H-L-arginine by the cytosolic extracts (Fig 3). The formation
of “H-L-citrulline was enhanced significantly in SMCs that had
been exposed simultaneously to LPS in combination with
either Cl-Ad, EHNA, ID0 or EHNA plus ID0 (Fig 3).
“H-L-citrulline formation was blocked when cytosolic extracts
of SMCs pretreated with Cl-Ad, EHNA, ID0 and EHNA
Conlrol EHNA ID0 EHNA EHNA + ID0 + ID0
Figure 3. Formation of 3H-L-citrulline from 3H-L-arginine in the presence and absence of L-NAME (1 mmol/L) and EDTA (1 mmol/L) by cytosolic extracts of SMCs treated for 20 hours with: A, 2-Chloroadenosine (Cl-Ad; 1 Om4 mol/L) in the absence (top) and presence (bottom) of LPS (25 Kg/ml); B, EHNA (IO kmol/L), ID0 (0.1 WmollL), and EHNA (10 pmol/L) plus ID0 (0.1 PmollL), in the absence (top) and presence (bottom) of LPS. Values for each data point represent mean+-SEM from 3 to 4 separate experiments, conducted in triplicates. *P <.05 vs con- trol; tFY.05 vs -LPS; #P<.O5 vs EHNA or IDO; +3P<.O5 vs Cl-Ad, LPS, EHNA, ID0 or EHNA+IDO in absence of L-NAME.
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Dubey et al 299
” Control 9 7 5 ” Control 9 7 5
CGS 21680 (-log mol i L) CPA (-log mol / L)
Figure 4. Nitrite/nitrate levels in the conditioned medium of SMCs treated for 30 hours with adenosine agonists CGS21680 (A) and CPA (B), in the absence (top) and presence (bottom) of LPS (25 pg/mL). Each data point represent meantSEM from 3 to 4 separate experiments. *PC.05 vs control; tk.05 vs -LPS.
plus ID0 with and wvlthout LPS were incubated with ‘H-L-
arglnlne 1n the presence of L-NAME. In contrast, the addition
of 2 mM EDTA to cytosohc extracts did not influence
‘H-L-cltrulhne formatIon (Fig 3).
CGS21680 and CPA only increased nltrlte/nitrate levels at
extremely high concentrations, whether 111 the presence or not
of LPS (Fig 4). Sunmlarly, only extremely high concentratlonr
of CPA and CGS21680 (10 ’ mol/L) induced ‘H-L-citrulhne
formation in LPS treated and untreated SMCs (data not
shown).
KF17837 and DPSPX, but not DPCPX, cignlficantly attenu-
ated C-Ad induced nltnte/nltrate and ‘H-L-cltrulhne cynthesls
by SMCs (Fig 5). Snmlarly, the stnnulatoly effects of EHNA plug
ID0 on nltnte/nltrate and ‘H-L-citrulline fonnatlon by SMCs
were ugmficantly attenuated by KFI 7837 and DPSPX, but not by
DPCPX (Fig 5). Treatment of SMCs with exogenous cychc
AMP for 30 h ourc increased nltnte/rntrate levels in the condl-
Figure 5. Effects of DPCPX (10 nmol/L, selective A, receptor antagonist), DPSPX (10 nmol/L; AI/A* receptor antagonist) and KF 17837 (KF; 10 nmol/L; selective A2 receptor antagonist) on 2-chloroadenosine (Cl-Ad; 10 nmol/L; A) and EHNA (10 pmol/L) plus ID0 (0.1 pmol/L; B) induced nltnc oxide synthesis by SMCs (nitrite/nitrate levels In the conditioned medium of SMCs treated for 30 hours [top] and formation of 3H-L-cltrulline from 3H-L-ar- ginine by cytosolic extracts of SMCs treated for 20 hours [bot- tom]). Values for each data point represent meantSEM from 3 to 4 separate experiments. *P<.O5 YS control; Sk.05 vs Cl-Ad or EHNA+IDO.
CAMP (-log molt L)
Figure 6. A, (top): Nitrite/nitrate levels in the conditioned medium of SMCs treated for 30 hours with cyclic AMP (CAMP) or cyclic AMP plus L-NAME (1 mmol/L); A, (bottom): formation of 3H-L-cltrulline in the presence and absence L-NAME (1 mmol/L) or EDTA (1 mmol/L) by cytosolic extracts of SMCs treated for 20 hours with cyclic AMP (10 4 mol/L). B, Effects of DPCPX (10 nmol/L), DPSPX (10 nmol/L) and KF 17837 (KF; 10 nmol/L) on cyclic AMP (CAMP; 10 pmol/L) induced NO synthe- sis (nitrite/nitrate in the condltioned medium of SMCs treated for 30 hours [top] and formation of 3H-L-citrulline from WL-arginine by cytosolic extracts of SMCs treated for 20 hours [bottom]). Values for each data point represent meantSEM from 3 to 4 separate experiments. *PC.05 vs control; §P<.O5 vs cyclic AMP or 8-Br-CAMP.
tioned medium in a concentration-dependent manner (Fig 6). In
contrast to untreated SMCs, cytosolic extracts of SMCs treated for
20 l~ours with cyclic AMP convelted “H-L-arginine to ‘H-L-
citrulline, and these effects were abrogated by L-NAME, but not
EDTA (Fig 6). Similar to cyclic AMP, treatment of SMCs with
8-bromo-cyclic AMP induced nitrite/nitrate levels and ‘H-L-
citrulline formation (data not shown). The stimulatory effects of
cyclic AMP on nitrite/nitrate as well as “H-L-citrulhne fomiation
were significantly reduced in the presence of KF17837 and
DPSPX, but not DPCPX (Fig 6).
The effects of adenosine and EHNAt ID0 on nitrite/nitrate
and “H-L-citrulline fomlation were not blocked by the adenylate
cyclase inhibitor, DDA, or the cyclic AMP-dependent kinase
inhibitor, Rp-CAMP (Fig 7).
Discussion The results of the present study demonstrate that treatment of
rat aortlc SMCs with adenoslne, with a Ttable adenoslne analog
(Cl-Ad) and with agent? that elevate endogenouv adenoslne
(EHNA and IDO), induce NO syndiesls. The stnnulatory
effects ofadenorme are not mlnucked by low concentratlony of
CGS21680, a selective AZA receptor agomst, or CPA, a
selective A, receptor agonist. ’ Furthermore, the stimulatory
effects of C-Ad and EHNA plus ID0 are slgnlficantly
attenuated by KF17837, a relectlve Az receptor antagomrt,”
and by DPSPX, an AI/AZ receptor antagonist,’ but not by
DPCPX, a selective A, receptor antagoruct.’ Thur, the stlmu-
lately effects of adenoTlne are most hkely mediated via AzH
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

300 CAMP-Adenosine Pathway Induces NO Synthesis
Figure 7. Effects of 2’,5’-dideoxyadenosine (DDA; adenylyl cyclase inhibitor; 100 pmol/L), Rp-CAMP (10 pmol/L; inhibitor of cyclic AMP-dependent protein kinase) on adenosine (Ad; 10 PmollL) and EHNA (10 kmol/L) plus ID0 (0.1 pmol/L) induced NO synthesis by SMCs (nitrite/nitrate levels in the medium of SMCs treated for 30 hours [top] and formation of %L-citrulline from 3H-L-arginine by cytosolic extracts of SMCs treated for 20 hours [bottom]). Values for each data point repre- sent mean?SEM from 3 to 4 separate experiments. *PC.05 vs control.
receptors and not via A, or A?, receptors. These findings provide evidence that exogenous as well as SMC-derived adenosine induces NO synthesis in an autocrine/paracrine fashion and via the A2,1 receptor. The present study also establishes that treatment of SMCs with cyclic AMP, a precursor of adenosine, stimulates NO synthesis. Moreover, the stimulatory effects of cyclic AMP are blocked by KF17837 and DPSPX, but not by DPCPX. These findings suggest that cyclic AMPderived adenosine induces NO-synthesis via A,, receptors. Finally, our studie$ show that the stimulation of NO synthesis by exogenous and endogenous adenosine is not blocked by DDA, an adenylyl cyclase inhibitor,” or Rp- cAMP, a preferential inhibitor of cyclic AMP-dependent protein kinase. ’ ’ These findings suggest that adcnosine induces NO synthesis via a pathway that does not involve adenylyl cyclare/protein kinase A.
Local generation of NO within blood vessels plays a key role in the biology, physiology, and pathophysiology of the vascu- lature.‘-’ NO generated luminally prevents platelet aggregation and adhesion, as well as neutrophil-induced damage to endo- thelial cells.‘-” When generated abluminally, NO activates soluble guanylyl cyclase in SMCs, and thereby increases cGMP levels, decreases intracellular Ca’+ levels, induces vasodilation, and inhibits proliferation and migration of SMCs.‘-‘,‘I Under basal conditions cNOS within the endothelial cells is the main source for NO synthesis, whereas under pathological condi- tionc, such as sepsis, iNOS within the SMCs plays a key role in inducing hypotension.” In addition to NO, there are several other vasodilatory factors that regulate the vascular function in an autocrine/paracrine iashion.” Moreover, many of these Eactors are now known to induce their effects via generation of etidothelium-derived NO.‘-” In this regard, recent studies have shown that adenosine, which mimicks most of the anti-
vatoocclu<lve effects of N0,j,12 rtnnulates NO synthevlr by vascular endothehal cells via A, receptor$.‘3,‘4
Although the vascular effects of adenoyme may, m part, be mediated via endothehal cell-derived NO, adenoTme induces va?odilation both m the presence and absence of the endothelium.” However, the mechamcmc for the endothehum-Independent dilatory effects adenosme are not fully understood. Recent studies show that teveral endog- enous vasodllatory factors stmm!ate NO Fynthesis by SMCs. In this regard, cyclic AMP, as well as agents that elevate cychc AMP levels, Induce NO synthesis by SMCY and mesanglal cells via lNOS stmlulatlon.4-” ‘(, Because treat- ment of SMCs with adenosme mcrea<er mtracellular cychc AMP levels,’ we hypothe$lzed that adenosme mducer NO rynthesls m vascular SMCs. The findmgc that adenosme and its stable analog, Cl-Ad, stimulate mtrite/mtrate cynthe$ls and ‘H-L-citrullme formation by SMCc and that thic stlmulatlon 15 inhibited by L-NAME provides evidence that adenocme mduces NO syntheqs m SMCs. Moreover, the observation that EDTA does not inhibit ‘H-L-citrulhne formation suggests that adenosme induces NO synthesis via 1NOS. Conylstent with this conclusion, Ikeda et a1.17 very recently published a report that Cl-Ad mducey 1NOS mRNA m rat vascular SMCs, thur confirmmg our prevl- ouslx and present findings that, via 1NOS ytmiulatlon, adenosme induce? NO synthesl$ m SMCs.
Our observations that CPA (adenosme analog selective for A, receptors at pharmacologically low doses [<I nmol/L]‘) 1s unable to induce NO Tynthesis at low concentrations and DPCPX (selective A, adenosine receptor antagonist) does not block the $tnnulatoiy effects of Cl-Ad on NO gynthe?lr suggest that the stimulatory effect\ of adenosme are not mediated via A, receptors. Our findings that KF17837 and DPSPX, both A, receptor antagomstc, but not DPCPX, attenuate the stmiula- tory effects of Cl-Ad suggest that the stlmulatory effects of adenosine are Az receptor mediated. Moreover, our ob?erva- tion that only high, but not low, concentrations of CGS21680, an adenosme agonist which 1s highly selective for AZA receptors at concentrations of <IO mnol/L, stimulates NO synthecl?, provides evidence that the effects of adenosme are not medi- ated via AzA receptors. Rather, our data Tuggectu that the AZ” receptors may be mvolved m medlatmg the stmiulatory effects of adenosine.
The above findings provide clear evidence that exogenous adenosme stimulates NO syntheslc by SMCs; however, whether endogenous adeno$me has snnilar stnnulatory effects cannot be inferred from studies with agomrts. Elimmatlon of adenosine from the interstitial Tpace 1s mediated by faclhtated transport of adenosine mto cells followed by metabohrm of adenosme to mosine by adenosme deammate and to AMP by adenosme kinase.‘2 Inhibltlon of adenonne deaminase by EHNA and adenosme kmase by ID0 mcrease? endogenous levels of adenocme.‘” Moreover, we have shown that EHNA and ID0 increase adenosme levels in cultured SMCrS and cardiac fibroblasts.“’ Hence, these two compounds were used m the present study to assess the role of endogenous, le, SMC-derived, adenosine on NO rynthesls.
EHNA and ID0 mcrea$e NO synthesis m SMCY, and the stlmulatory effects of EHNA and ID0 are slgmficantly en-
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Dubey et al
hanced when SMCs are treated with a combmatlon of these agents Moreover, the stlmulatory effects of EHNA and ID0 on NO synthesis are blocked by L-NAME, but not by EDTA, suggesting that SMC NO 1s derived from lNOS The AZ receptor blockers, KF17837 and DPSPX, significantly attenu- ate the stlmulatory effects of EHNA plus IDO, supportmg our contention that the stlmulatory effects of these agents on NO synthesis m SMCs are mediated via generation of adenosme Moreover, the finding that DPCPX does not reverse the stlmulatory effects of EHNA plus ID0 on SMCs strongly suggests that the stlmulatory effects of endogenous adenosme are mediated via A2 receptors
Formation of adenosme wlthm and/or near the blood vessel wall occurs via three blochemlcal pathways (1) the mtracel- lular ATP pathway, which entails intracellular dephosphoryla- non of ATP to adenosme, (2) the extracellular ATP pathway, which 1s a source of extracellular adenosme, (3) the transmeth- ylatlon pathway, which mvolves the hydrolysis of S-adenosyl- L-homocysteme (SAH) to L-homocysteme and adeno- sme 8,12,1’1 Smce the intracellular and extracellular ATP pathways of adenosme production reqmre crlsls events and the transmethylatlon pathway 1s mostly constltutlve, the three tradltlonal routes of adenosme blosynthesls are not well suited for physlologlcal modulation More recently, we have dem- onstrated the presence of a fourth pathway, the cychc AMP- adenosme pathway, for adenosme production m vascular SMCs* that would be more amenable to physlologlcal modu- lation by hormones
The cychc AMP-adenosme pathway begms with the actl- vatlon of adenylyl cyclase and has both intracellular and extracellular sites of adenosme production * Wlthm the cell, cytosohc PDE and cytosohc 5’-nucleotldase metabolize cychc AMP to AMP and AMP to adenosme, respectively, and the adenosme formed reaches the extracellular space via faclhtated transport *,I* However, due to the competltlon of cytosohc 5’-nucleotldase and adenylate kmase for AMP and the com- petltlon of transport mechanisms with adenosme kmase for adenosme, the intracellular formation of adenosme may be dlmmlshed R l2 Therefore, the extracellular hmb of the cychc AMP-adenosme pathway, which mvolves the efficient metab- olism of cychc AMP to AMP and adenosme by ecto-PDE and ecto-5’-nucleotldase, respectively, may be quantltatlvely more important 8 I2
Regarding the extracellular cychc AMP-adenosme pathway, stimulation of adenylyl cyclase results m egress of cychc AMP,8,‘2 and relatively modest increases m cychc AMP production could give rise to slgmficant concentrations of adenosme at the cell surface, smce adenosme would be synthesized by a series of spatially linked enzymatic reac- tions * I2 We have recently shown that SMCS,~ mesangal cellQ” and cardiac fibroblasts*l express the extracellular hmb of the cychc AMP-adenosme pathway Moreover, m all these cell
types1 cychc AMP-derived adenosme mhlblts mltogen- induced cell growth. ?,** These studies led us to hypothesize that the cychc AMP-adenosme pathway may also induce NO synthecls m SMCs To test this hypothesis, we evaluated the effects of cychc AMP on NO synthesis m the presence and absence of the A, specific and nonspecific receptor antagonists,
KF17837 and DPSPX, respectively, and the A, adenosme receptor antagonist, DPCPX
Cychc AMP stmlulates NO production by SMCs, and the stlmulatory effect of cychc AMP on NO synthesis 1s sign& cantly attenuated by the adenosme receptor antagonists KF17837 and DPSPX, but not by DPCPX, quggestmg the mvolvement of A, receptor5 This conclusion 1s consistent with our observations that the stlmulatory effects of exogenous and endogenous adenosme on NO synthesis by SMCs are medl- ated via A, receptors and are blocked by KF17837 and DPSPX Taken together these findings strongly suggest that the cychc AMP-adenosme pathway may contribute lmpor- tantly to the regulation of vascular biology and more speclfi- caliy NO synthesis by SMCY
Although these findings provide evidence for the role of exogenous, endogenous, and cychc AMP-derived adenosme m mducmg NO synthesis by SMCr, the mechanisms involved m this effect are unclear Our data provides evidence that the effects of adenosme are mediated via AZ receptors Smce AZ receptors are posltlvely coupled to adenylyl cyclase and cychc AMP mducec NO qynthesls m SMCs by mcreasmg lNOS activity, It 1s feasible that the stlmulatory effects of adenosme on NO synthesis by SMCs 1s medlated largely via the second messenger cychc AMP However, neither DDA, an mhlbltor of adenylate cyclase that blocks cychc AMP generation, nor Rp-CAMP, an mhlbltor of cychc AMP-dependent kmase, block the stmmlatory effects of either adenocme or EHNA plus ID0 on NO synthesis These findings cuggest that the stlmu- latory effects of exogenous as well as SMC-denved adenosme on NO synthesis are mediated via A, adenosme receptor? linked to a pathway not mvolvmg adenylyl cyclase/protem kmase A
In conclusion, we provide evidence that both exogenous and vascular SMC-derived adenosme induces NO synthesis by vascular SMCs m an autocrme fashion As the effects of exogenous and endogenous adenosme on “H-L-cltrullme fol- matlon are not blocked by EDTA, the stlmulatory effects of adenosme are most likely mediated via lNOS Moreover, our findings provide evidence that adenosme generated from cychc AMP stimulates NO synthesis by SMCs m an autocrme/ paracrme fashion Since both adenosme and NO induce vasodllatlon, mhlblt platelet aggregation and adhesion, and prevent mltogen-induced prohferatlon and mlgratlon of SMCs,‘e3 ‘* processes importantly Involved m vaso-occlusive disorders m various vascular diseases, the current findmgq suggest that adenosme produced by SMCs may play a vital role as a local anti-vasooccluclve agent, which may induce Its effect? m part via generation of NO locally wlthm the vessel wall Moreover, decreased SMC synthesis of adenosme or increased catabolism of adenosme by adenosme deammase or adenosme kmase may importantly contribute to decreased qynthesls of NO This may m turn be associated with abnormal growth of SMCs and the vascular remodeling and vaso-occlu5lve dlsor- ders observed m hypertension, atherosclerosis, and restenoslq followmg balloon angloplasty 3 ‘* Agents that elevate endoge- nous adenosme could be chmcally important m preventing vaso-occlusive disorders associated with hypertension, athero- sclerosis, and restenosls by mducmg NO synthesis wlthm the vasculature, thus exerting beneficial effects on the vessel wall
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

302 CAMP-Adenosme Pathway Induces NO Synthesis
Finally, it IS possible that the cychc AMP-adenosme pathway may Importantly regulate vascular biology and physiology m VlVO
Acknowledgments This work was supported by grants from the NatIonal Institute of Health (HL 55314 and HL 35909)
1
2
3
4
5
6
References 14
Moncada S, Pahner RM, Hlggs EA NO Physiology. pathophyslology and
pharmacology Pharrnacol Rev 1991,43 109-142 Moms SM Jr, B~lhar TR New mslghts Into the regulation of mduable
mtrtc oxide synthws AmJ Phystol 1994,266 E82Y -E83Y Dubey RK, Jackson EK, Rupprecht HD, Sterzel RB Factors controlhng growth and matrx prodwon m vascular smooth muscle and glomerular nmangd celk Cwr Oyln Nephrvl Hyyertetu 1997,6 88-105 Sea HG, FUJII J, Asalu M, Okado A, Fu~‘wara N, Tamguclu N Roles of punne nucleot~des and adenosme m enhancmg NOSII gene expression m
lnterleukln-IP-stimulated rat vascular smooth muscle cells Free Rod Rex 1997,26 409-418
15
16
17
18 Ima~ T, Hlrata Y, Kanno K, Marumo F Inductmn of mtnc oxide synthase by CAMP ,n rat vascular smooth muscle cells J C/w Invesf 1994,93 543-549 19 Schml-Kerth VB, Flsslthaler B, Busse R CGRP enhances mductlon of NO synthase m vascular smooth muscle cells vu a cychc AMP-dependent mechamsm AnrJ Dhysrol 1994,267 H2483-H2490 D&xl HH, Westfall DP Receptors for adenme nucleoude~ and nucleo- rides subclawficauon, dlstnbutlon, and molecular characterlzatlon Pharmacol Rev lY94,46 449-466
20
Dubey KK, MI Z, CXleqxe DG, Jackcon EK Cychc AMP-Adenosme pathway ml&m vascular smooth muscle cell growth Hypewwston 1996, 28 765-771
21
Dubey RK, Gdlespuz DG, MI Z, Suzuki F, Jackson EK Smooth muscle cell-dewed adenosme mhlbas cell growth Hyperfe~~~~~~i 1996,27(suppl
II) 766-773 Dubey RK, Glllesple DG, Ml Z, Jackson EK Exogenous and endogenous adenosme mhlblts fetal calfserum-mduced growth of rat cardiac tibroblasts role of A2B receptors Grtulatton 1997,96 2656-2666
22
23
11
12
13
Dubey RK, Jackson EK, Luscher TF NltnL oxide mlublts ang~otensm
II-mduced tmgranon of rat aoruc smooth muscle cells Role of cychc
nucleotldes and anglotensm, receptors J Clln Znverr 1995,96 141-149. Jackson EK, Koehler M, Ml Z, Dubey RK, Tofowc SP, Carallo JA, Jones
GS Powble role of adenosme deammase m vase-occluswe chseages
/ Hypermr 1996.14 19-29
L.1 J-M, Fenton RA, Cutler BS, Dobson JG Jr Adenosme enhances mtnc
oxide production by vascular endothehal cells Aln J Pliyrlol 1995.269 C519-523
Vials A, Burnstock G A,-purmoceptor-mahated relaxauon 1n the
gumea-pig coronary vasadature R role for mtrlc oxide Br J Plramwwl 1993,109 424-42’)
Headrlck JP, Bane KM Endothehun-dependent and -Independent relax- anam to adenowe m gumea p’g aorta Ant J Physd 1990,259 H62-H67
Kunz D, Muhl H, Walker G, Pfellschlfter J Two chstmct slgnahng
pathways tugger the exprewon of mduable nmx oxide synthase m rat
mesang~al cells Proc Nor1 Acad Sn US A lYY4,91 5387-53Yl
Ikeda U, Kurosakl K, Ohya K, Shunada K Adenosme stnnulatrs mtnc oxide synthew m vascular smooth muscle cell Cnr&was Rcs 1997,35
168-174
Dubey RK, G~llespx DG, Jackson EK Adenosme mduces n,tr,c oxide
synthesis m rat aorac smooth muscle cells Hypermmn lYY6,28 552
(abstract P178) Saottl VM, Van-Wylrn DC Incrraws m mterstmal adenosme and cerebral
blood tlow wth mlubmon of ndenosmr kmase and adenosme deanunase
/ Cereb Blood Fh Melnb 1993,13 201-207 Dubey RK, G&nple DG, Ouka K, Suzuki F, Jackson EK Adenosme
mhlblts growth of rat aauc smooth muscle dells l’owble role of AZ,,
receptor Hypcrtc~~s~m 1996,27(suppl II) 786-793 Dubey RK, MI Z, Gdlesple DG, Jackson EK Cy& AMP-Admoanc
pathway mhlblts glomerular mesangul cell growth Hyperfewo~~ iYY7, 30 506 (abstract P13Y)
Dubey RK, Glllespx DG, MI Z, Jackson EK Cardiac fibroblasts metabohze
cych-AMP to generate adenosme Hypertemto~~ 1996,28 524 (abstract P7)
Dubey RK, Gdlesple DG, Jxkson EK Exogenous and ~nrdlac fibroblast cell-dewed adenosme mhlbas serum-mduced collagen and
protan synthesis III cardiac fibroblasts Hypertemtur~ 1996,28 541
(abstract PllO) by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Raghvendra K. Dubey, Delbert G. Gillespie and Edwin K. JacksonMuscle Cells
Cyclic AMP-Adenosine Pathway Induces Nitric Oxide Synthesis in Aortic Smooth
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 1998 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.31.1.296
1998;31:296-302Hypertension.
http://hyper.ahajournals.org/content/31/1/296the World Wide Web at:
The online version of this article, along with updated information and services, is located on
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer information about this process is available in the
is located, click Request Permissions in the middle column of the Web page under Services. Further requestedthe Editorial Office. Once the online version of the published article for which permission is being
can be obtained via RightsLink, a service of the Copyright Clearance Center, notHypertensionpublished in Requests for permissions to reproduce figures, tables, or portions of articles originallyPermissions:
by guest on June 19, 2018http://hyper.ahajournals.org/
Dow
nloaded from