crispr agbio workshop london...
TRANSCRIPT

Vladimir Nekrasov
CRISPR AgBio WorkshopLondon 2017


Nicotianabenthamiana
Tomato Barley
Maize Brassica
Wheat
Model plant: Crop plants:
Applying CRISPR-Cas9 technology in model and crop plants
Arabidopsisthaliana
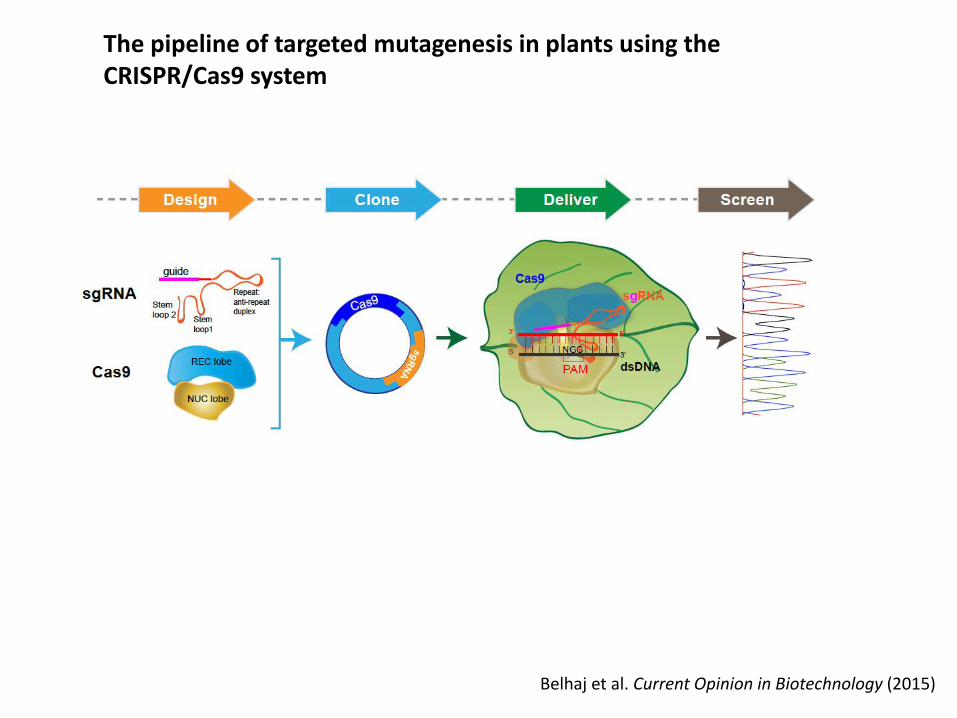
The pipeline of targeted mutagenesis in plants using theCRISPR/Cas9 system
Belhaj et al. Current Opinion in Biotechnology (2015)
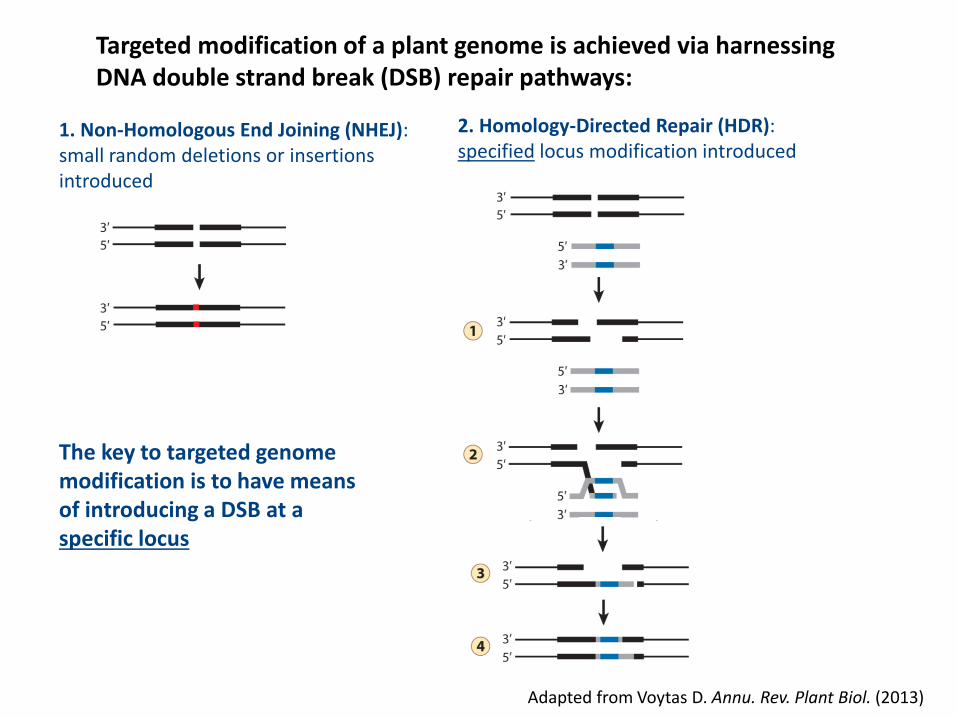
Targeted modification of a plant genome is achieved via harnessingDNA double strand break (DSB) repair pathways:
Adapted from Voytas D. Annu. Rev. Plant Biol. (2013)
1. Non-Homologous End Joining (NHEJ):small random deletions or insertions introduced
2. Homology-Directed Repair (HDR):specified locus modification introduced
The key to targeted genome modification is to have means of introducing a DSB at a specific locus

Single locus mutation
CRISPR/Cas9 applications in plants:targeted genome modification via DNA double strand break (DSB) repair by the non-homologous end joining (NHEJ)
5
Large scale chromosomal deletion
4 5 6
Cas9/sgRNA 1 Cas9/sgRNA 2
1
1 2 3
2 5 6
4 5 6
Cas9/sgRNA 1
1 2 3
5 61 2 3 4

http://www.rgenome.net
CRISPR RGEN Tools is a webtool for designing CRISPR/Cas targets and finding off-targets in genomes
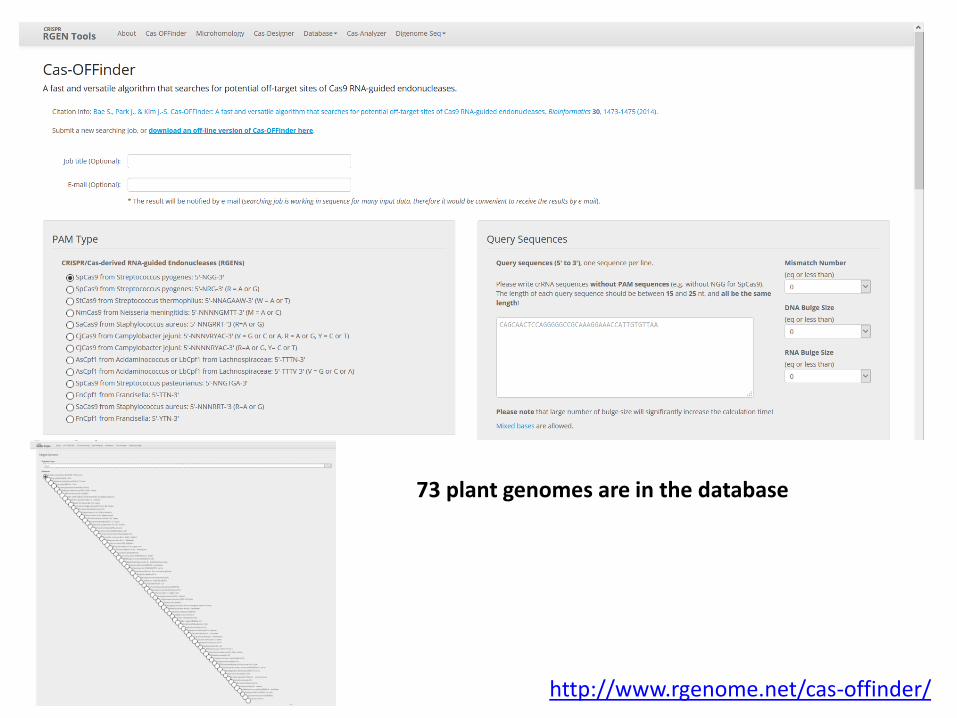
73 plant genomes are in the database
http://www.rgenome.net/cas-offinder/

PCR
Mutant (biallelic)
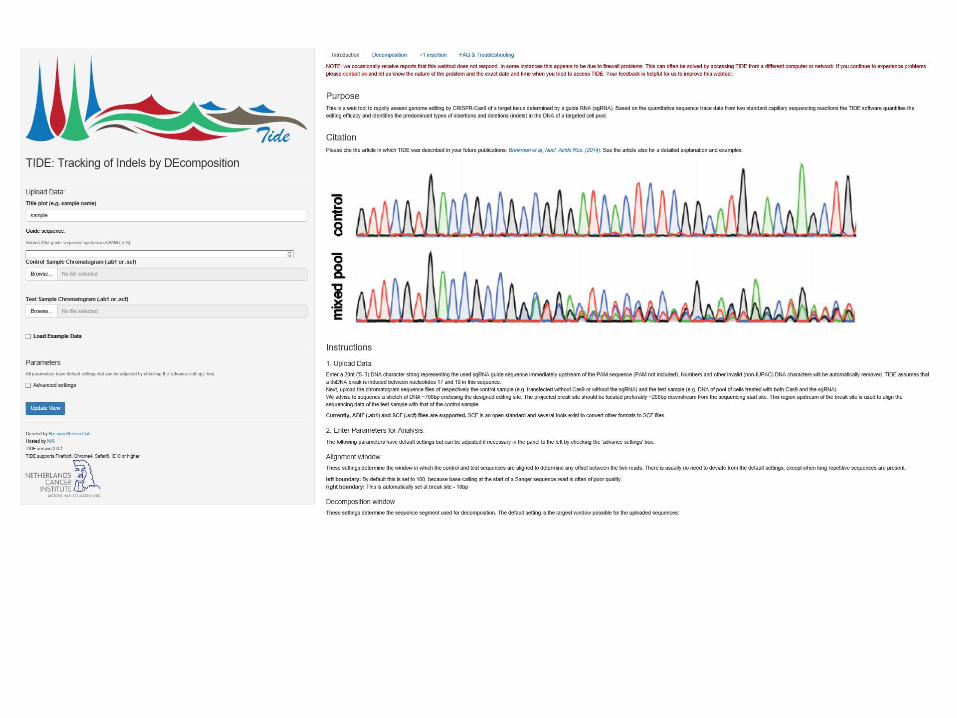
Decompositionusing TIDE webtool
Sanger sequencing
Methods for detecting CRISPR/Cas9-induced mutations
https://tide.nki.nl/


PCR with barcoded oligosMutant 1
Next GenerationSequencing (NGS)
Mutant 2
PCR
Mutant (biallelic)
Decompositionusing TIDE webtool
Sanger sequencing
Methods for detecting CRISPR/Cas9-induced mutations
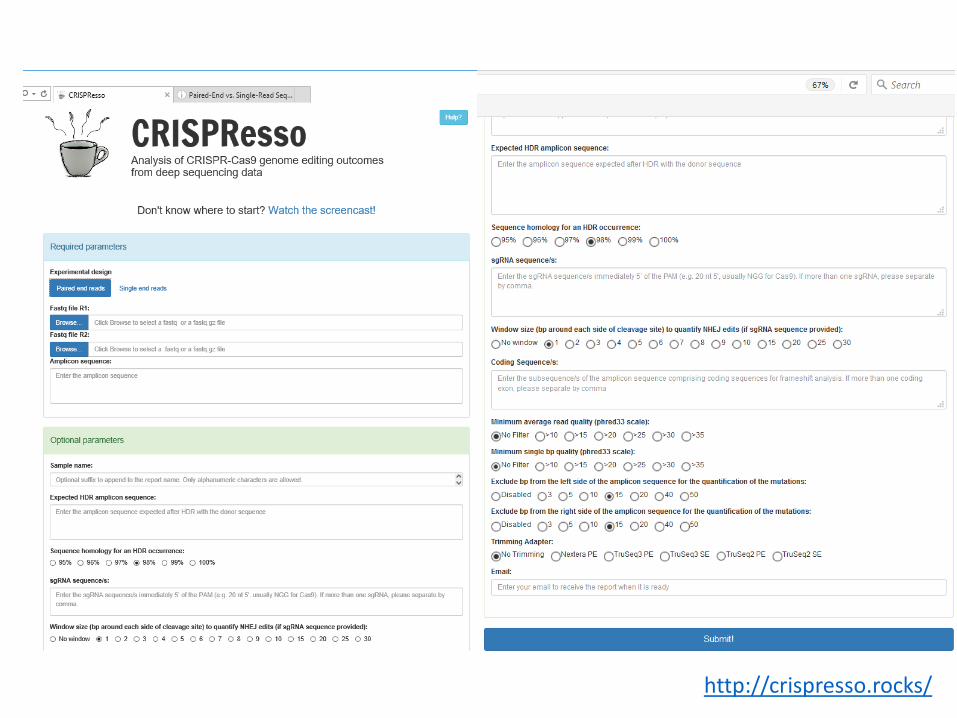


Fluorescent PCRCapillary electrophoresis
high-resolution fragment analysis (HRFA)
PCR with barcoded oligosMutant 1
Next GenerationSequencing (NGS)
Mutant 2
PCR
Mutant (biallelic)
Decompositionusing TIDE webtool
Sanger sequencing
Methods for detecting CRISPR/Cas9-induced mutations
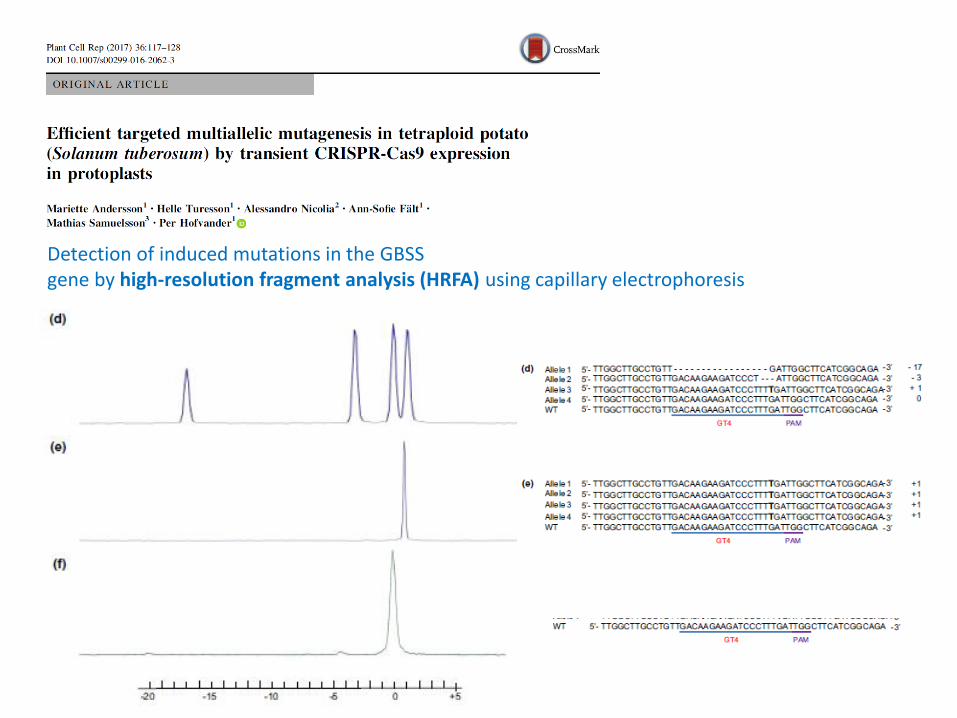
Detection of induced mutations in the GBSS gene by high-resolution fragment analysis (HRFA) using capillary electrophoresis

+
+
PCR, denaturation,annealing
PCR,denaturation
Surveyor assayCelI, T7
High ResolutionMelting analysis
Heteroduplex Mobility Assay (non-denaturing PAGE)
Single-strand conformationpolymorphism (SSCP)(non-denaturing PAGE)
Methods for detecting CRISPR/Cas9-induced mutations
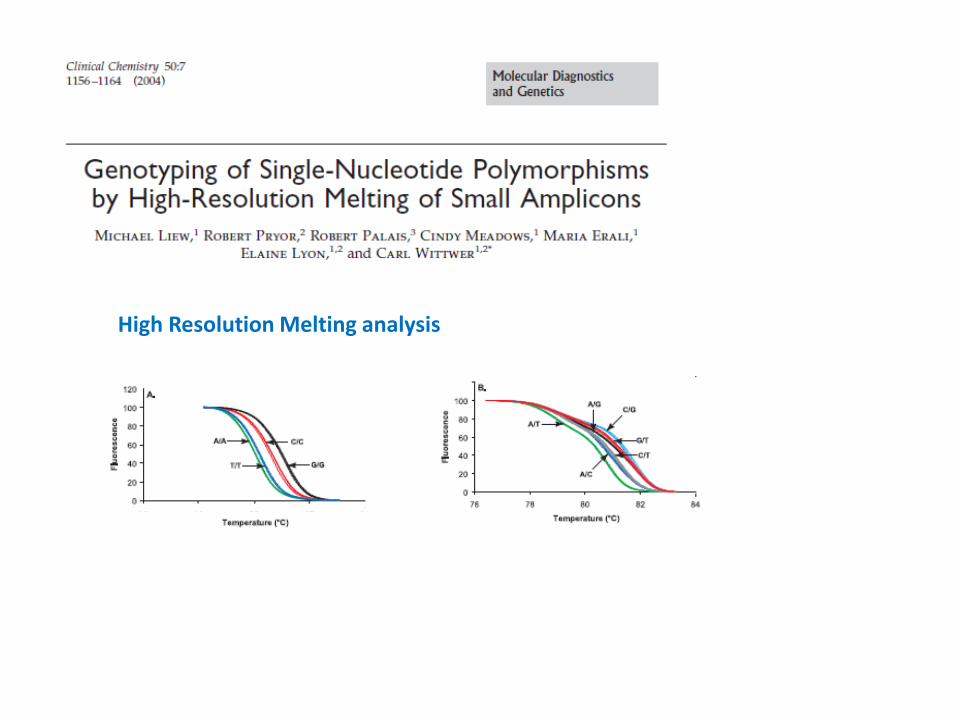
High Resolution Melting analysis
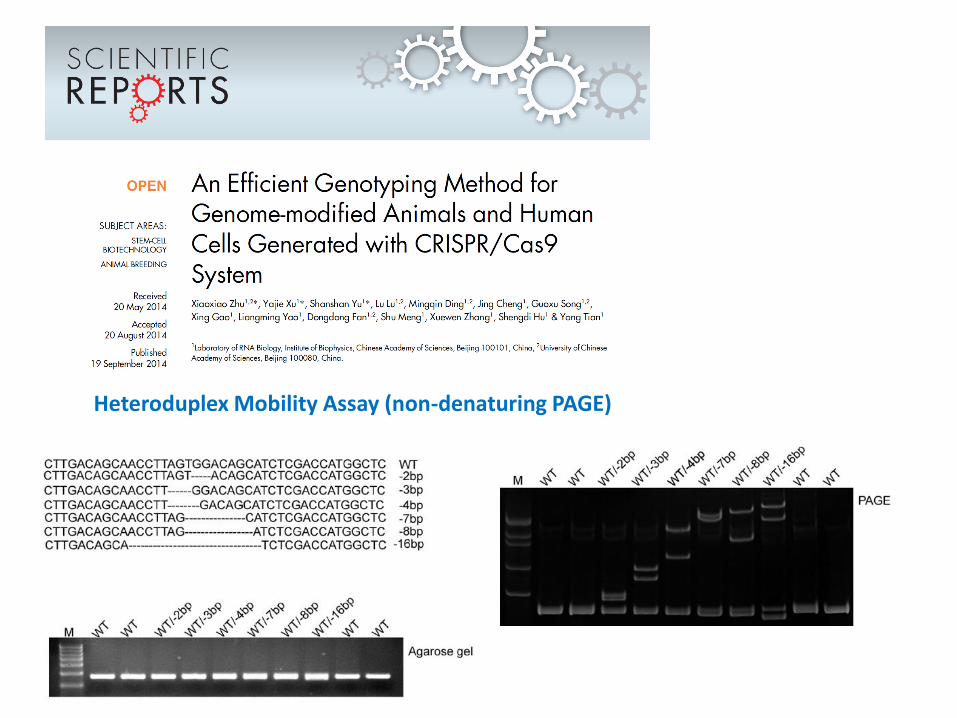
Heteroduplex Mobility Assay (non-denaturing PAGE)
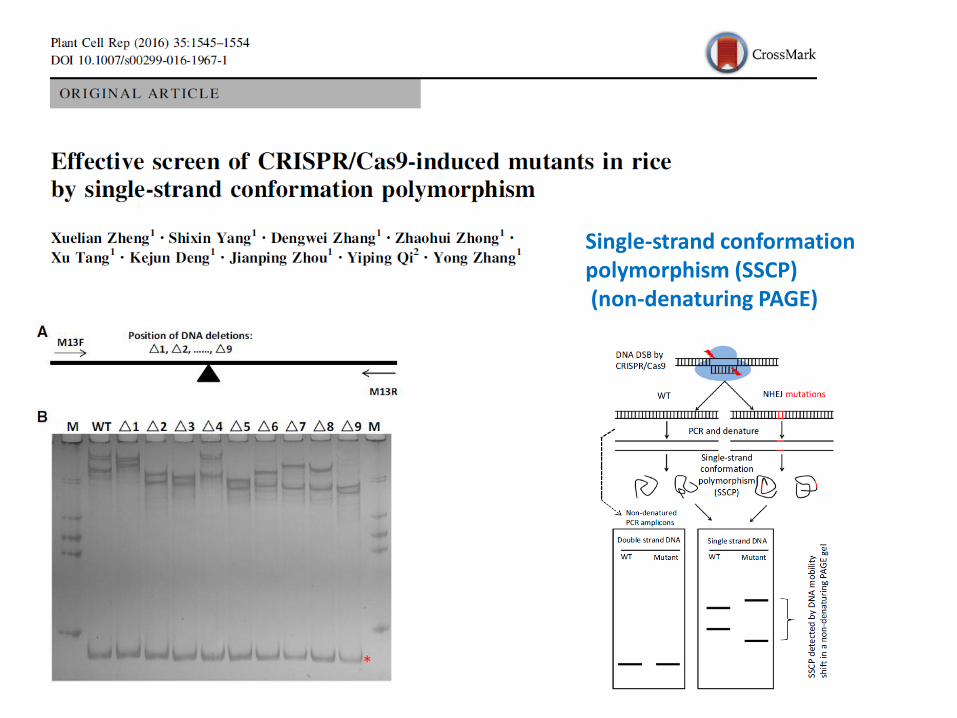
Single-strand conformationpolymorphism (SSCP)(non-denaturing PAGE)

PCR,digesting with a restriction enzyme
Restriction enzymesite loss assay
+
+
PCR, denaturation,annealing
PCR,denaturation
Surveyor assayCelI, T7
High ResolutionMelting analysis
Heteroduplex Mobility Assay (non-denaturing PAGE)
Single-strand conformationpolymorphism (SSCP)(non-denaturing PAGE)
Methods for detecting CRISPR/Cas9-induced mutations
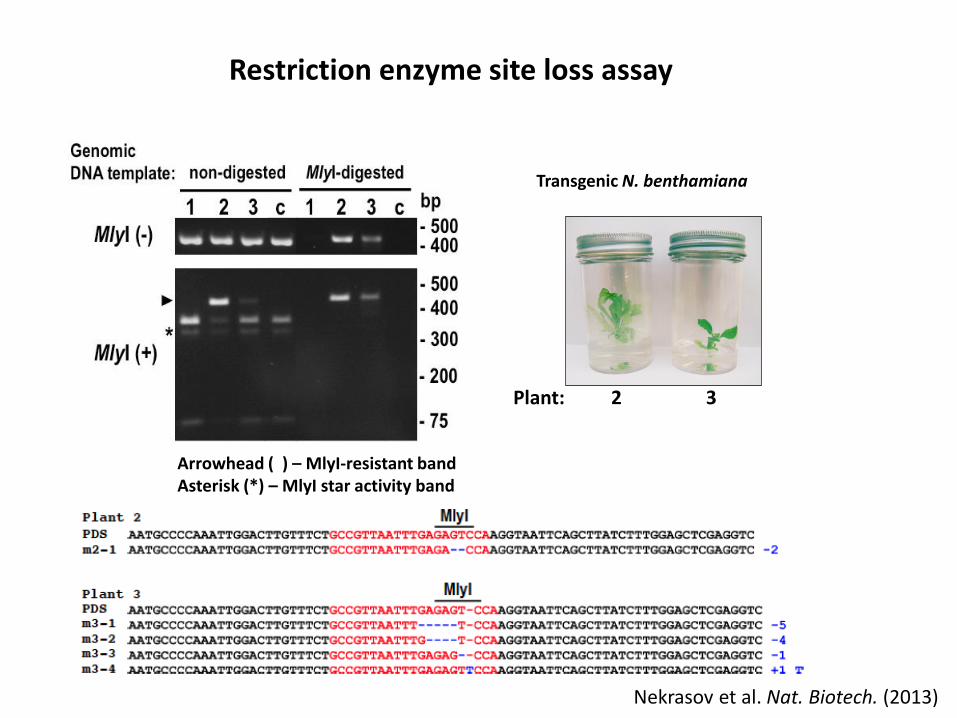
Transgenic N. benthamiana
Plant: 2 3
Arrowhead ( ) – MlyI-resistant bandAsterisk (*) – MlyI star activity band
Restriction enzyme site loss assay
Nekrasov et al. Nat. Biotech. (2013)

PCR,digesting with a restriction enzyme
Restriction enzymesite loss assay
+
+
PCR, denaturation,annealing
PCR,denaturation
Surveyor assayCelI, T7
High ResolutionMelting analysis
Heteroduplex Mobility Assay (non-denaturing PAGE)
Single-strand conformationpolymorphism (SSCP)(non-denaturing PAGE)
PCRAmplified fragmentlength polymorphism(AFLP)
Methods for detecting CRISPR/Cas9-induced mutations

LBP1 ORF1 T1 P2 ORF2 T2 P3 ORF3 T3 P4 ORF4 T4 P5 ORF5 T5
P6 ORF6 T6 P7 ORF7 T7 P8 ORF8 T8
P11 ORF11 T11
P9 ORF9 T9
P10
1 2 3 4 5
6 7 8
11
9
10 ORF10 T10
Level 2multigeneconstructs
35S ORF Ter
Level 1transcription
units
Level 0Choice ofmodules
P ORF Ter
Promoter 5' UTR ORF Ter
P1 Ter106
P2 Ter107
P3 Ter128
P4 Ter139
P5 Ter1410
Ter6
Ter7
Ter8
Ter
Ter9
Ter1
Ter2
Ter3
Ter4
Ter5
ORF1
ORF2
ORF3
ORF4
ORF5
ORF6
ORF7
ORF8
ORF9
ORF10
ORF11
ORF12
ORF13
ORF14
ORF15
P6
P7
P8
P9
P10
P11
P12
P13
P14
P15
1
2
3
4
5
SP1
SP2
SP3
SP4
SP5
SP6
SP7
SP8
SP9
SP10
SP11
SP12
SP13
SP14
SP15
Libraries of basicmodules
Promoters 5' UTR ORF TerminatorsSignal Peptides
RB
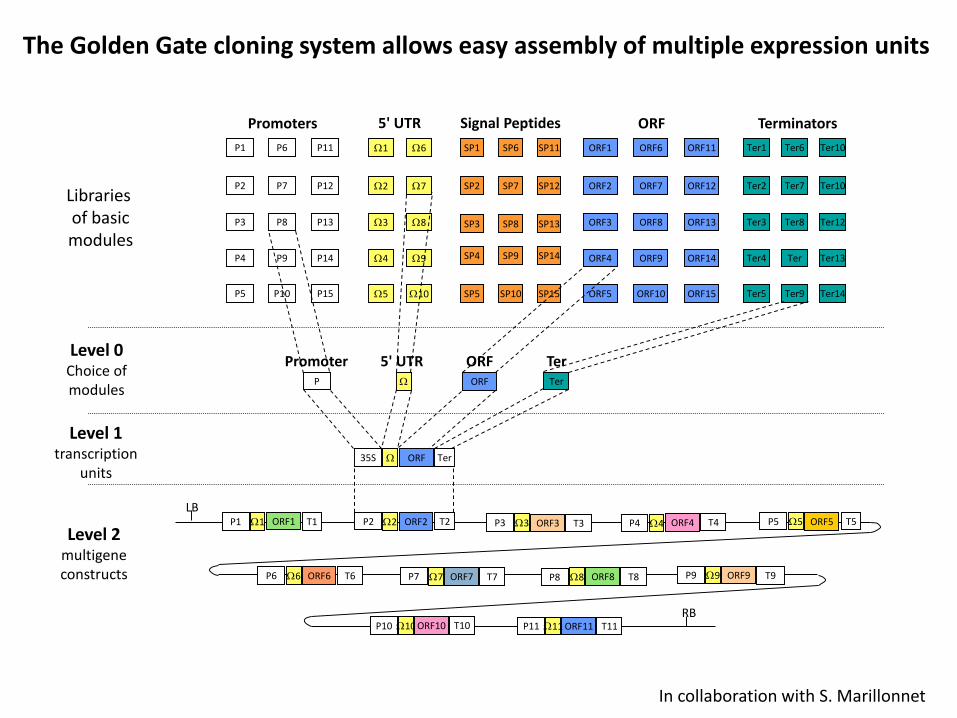
The Golden Gate cloning system allows easy assembly of multiple expression units
In collaboration with S. Marillonnet

GCAATTAC
ACTA
Kan
LB RB
+
Carb
NPTII
GGGA
Red
TGCC
TGCC
Bp
iI
GCAAB
piI
Bp
iI
Bp
iI
BpiI + ligase
+
Carb
Cas9Bp
iI
Bp
iI
ACTA
+
Carb
sgRNA1Bp
iI
Bp
iI
+
pICH47732::NOSp::NPTII-OCSTLevel 1
pICH47742::35Sp::Cas9-NOSTLevel 1
pICH47751AtU6p::sgRNA1Level 1
pAGM4723Level 2 vector
RBGGGA
Kan
LB
Cas9NPTII
ACTA
sgRNA1
TTACGCAATGCC
NPTII-Cas9-sgRNALevel 2
select white colonies
CAGA GGGA
Bp
iI
Bp
iI
Spec
L4E
pICH41780Linker
CAGATTAC
Carb
sgRNA2Bp
iI
Bp
iI
pICH47761AtU6p::sgRNA2Level 1
+
sgRNA2
CAGA
L4E
Using the Golden Gate cloning system to assemble CRISPR constructs
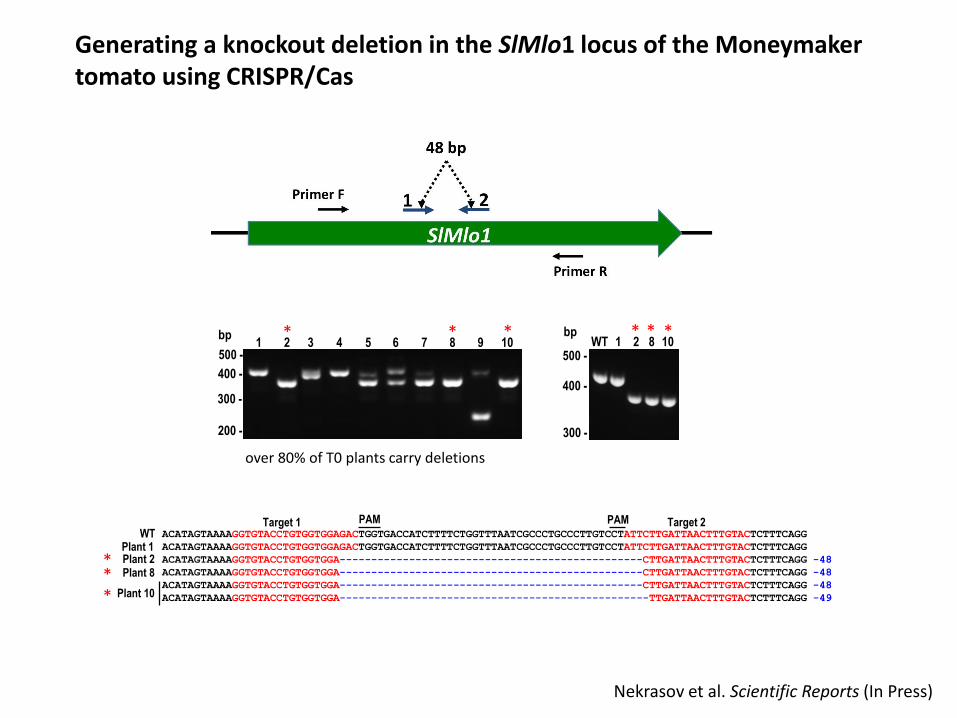
Generating a knockout deletion in the SlMlo1 locus of the Moneymaker tomato using CRISPR/Cas
500 -
400 -
300 -
bp
200 -
1 2 83 4 5 6 7 9 10* * *
500 -
400 -
300 -
bp1 2 8 10WT
* * *
ACATAGTAAAAGGTGTACCTGTGGTGGAGACTGGTGACCATCTTTTCTGGTTTAATCGCCCTGCCCTTGTCCTATTCTTGATTAACTTTGTACTCTTTCAGG
ACATAGTAAAAGGTGTACCTGTGGTGGAGACTGGTGACCATCTTTTCTGGTTTAATCGCCCTGCCCTTGTCCTATTCTTGATTAACTTTGTACTCTTTCAGG
ACATAGTAAAAGGTGTACCTGTGGTGGA------------------------------------------------CTTGATTAACTTTGTACTCTTTCAGG -48
ACATAGTAAAAGGTGTACCTGTGGTGGA------------------------------------------------CTTGATTAACTTTGTACTCTTTCAGG -48
ACATAGTAAAAGGTGTACCTGTGGTGGA------------------------------------------------CTTGATTAACTTTGTACTCTTTCAGG -48
ACATAGTAAAAGGTGTACCTGTGGTGGA-------------------------------------------------TTGATTAACTTTGTACTCTTTCAGG -49
WTPlant 1Plant 2Plant 8
Plant 10
PAM PAMTarget 1 Target 2
***
over 80% of T0 plants carry deletions
Nekrasov et al. Scientific Reports (In Press)

slmlo1
SlMLO1 (WT)
CRISPR/Cas-engineered slmlo1 tomato line is fully resistant to the powderymildew pathogen Oidium neolycopersici

T1 generation slmlo1 plants were screened for the absence of T-DNA
500 -400 -
300 -
8-1
500 -400 -
300 -
bp 8-2 8-3 8-4 8-5 WT
T-DNA
slmlo1
8-6
SlMlo1
T-DNA
slmlo1
WT
slmlo1 8-2
slmlo1 8-4
slmlo1 8-6 (T-DNA)
Illumina reads matching the T-DNA
T-DNA
LB RB
Nekrasov et al. Scientific Reports (In Press)

Illumina whole genome sequencing confirmed presence of a homozygous deletion in the SlMlo1 locus
WT
slmlo1 8-2
slmlo1 8-4
slmlo1 8-6 (T-DNA)
WT
slmlo1 8-2
slmlo1 8-4
slmlo1 8-6 (T-DNA)
Nekrasov et al. Scientific Reports (In Press)

Target sequence (PAM is in Green) Sites chromosome position off-target sequence Reference
base
Called base
(slmlo1 8-2)
Rate of the
called base
Mutation? Different
from wild
type?
Reference base Called base
(wild type)
Rate of the
called base
Mutation?
CCAATTCTTGATTAACTTTGTAC 1 chr0 15022112 CCAATTCTTtATTAttTTTGTAC C C 0.995 - - C C 0.992 -
2 chr0 15022113 CCAATTCTTtATTAttTTTGTAC C C 0.989 - - C C 0.985 -
3 chr0 15022114 CCAATTCTTtATTAttTTTGTAC A A 1 - - A A 1 -
4 chr0 15022115 CCAATTCTTtATTAttTTTGTAC A A 0.995 - - A A 1 -
5 chr0 15022116 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
6 chr0 15022117 CCAATTCTTtATTAttTTTGTAC T T 0.925 - - T T 0.947 -
7 chr0 15022118 CCAATTCTTtATTAttTTTGTAC C C 1 - - C C 0.985 -
8 chr0 15022119 CCAATTCTTtATTAttTTTGTAC T T 0.995 - - T T 0.993 -
9 chr0 15022120 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
10 chr0 15022121 CCAATTCTTtATTAttTTTGTAC t T 1 - - t T 0.993 -
11 chr0 15022122 CCAATTCTTtATTAttTTTGTAC A A 0.995 - - A A 0.993 -
12 chr0 15022123 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
13 chr0 15022124 CCAATTCTTtATTAttTTTGTAC T T 0.995 - - T T 1 -
14 chr0 15022125 CCAATTCTTtATTAttTTTGTAC A A 0.995 - - A A 1 -
15 chr0 15022126 CCAATTCTTtATTAttTTTGTAC t T 0.995 - - t T 0.986 -
16 chr0 15022127 CCAATTCTTtATTAttTTTGTAC t T 1 - - t T 0.98 -
17 chr0 15022128 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
18 chr0 15022129 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
19 chr0 15022130 CCAATTCTTtATTAttTTTGTAC T T 0.995 - - T T 1 -
20 chr0 15022131 CCAATTCTTtATTAttTTTGTAC G G 0.995 - - G G 0.988 -
21 chr0 15022132 CCAATTCTTtATTAttTTTGTAC T T 1 - - T T 1 -
22 chr0 15022133 CCAATTCTTtATTAttTTTGTAC A A 0.99 - - A A 0.987 -
23 chr0 15022134 CCAATTCTTtATTAttTTTGTAC C C 1 - - C C 0.994 -
Off-target analysis: 145 putative off-targets with up to 4 mismatches analysed
Result: no off-target mutations detected suggesting that CRISPR/Cas is a highly precise tool in tomato
Nekrasov et al. Scientific Reports (In Press)

Transform with Cas9/sgRNAs
Callustissue
T0 plantlets
T1 seeds slmlo1 T-DNA segregating
Screen forhomozygous slmlo1 T0 mutants
Screen T1 generationfor T-DNA-free plants
T2 seedsslmlo1T-DNA-free
0 3 6 9.5Time, months3 months 3 months 3.5 months
Homozygous T-DNA free slmlo1 tomato lines have been producedin less than 1 year
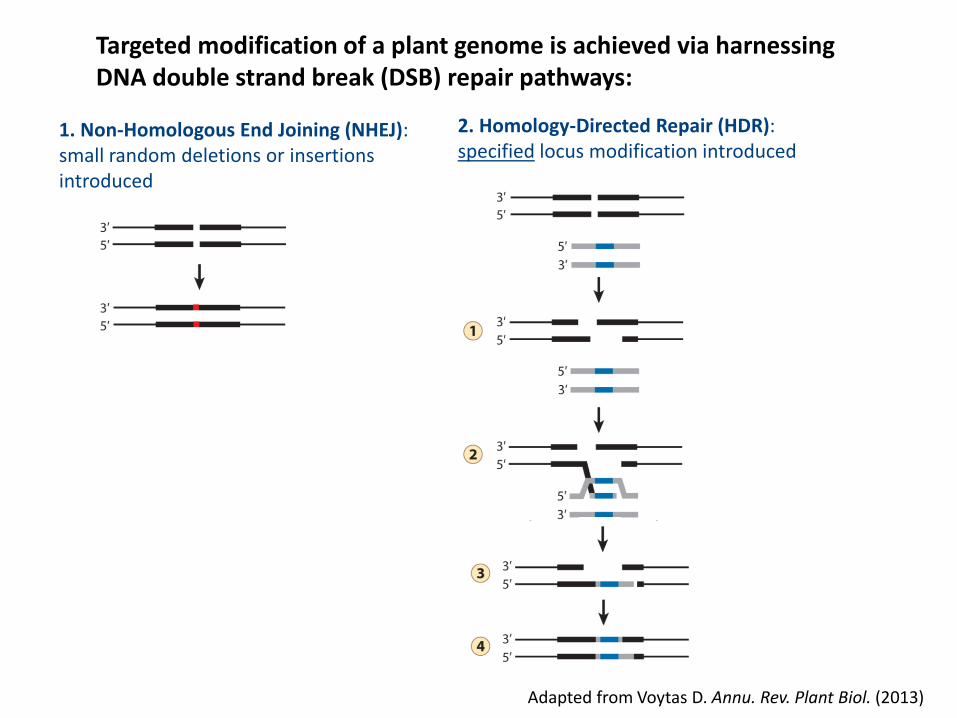
Targeted modification of a plant genome is achieved via harnessingDNA double strand break (DSB) repair pathways:
Adapted from Voytas D. Annu. Rev. Plant Biol. (2013)
1. Non-Homologous End Joining (NHEJ):small random deletions or insertions introduced
2. Homology-Directed Repair (HDR):specified locus modification introduced

CRISPR/Cas9 technology applications involving homologous recombination (HR)
• Targeted gene insertion (e.g. GFP tag)• Targeted gene replacement (introducing SNPs into a desired gene)
Targeted Gene Insertion
Gene 1
Gene 1
Repair template
Target locus
Targeted Gene Replacement
Cas9/sgRNA
Gene 1
Gene 1
Target locus
Repair templateCas9/sgRNA
GFP
GFP
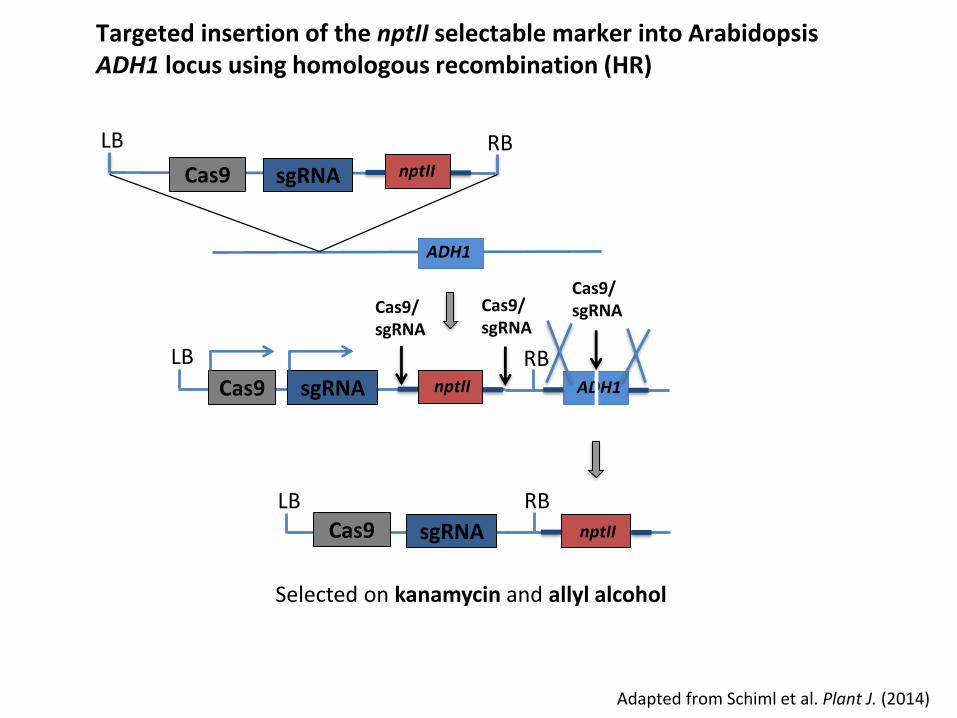
Cas9/sgRNACas9/
sgRNACas9/sgRNA
Adapted from Schiml et al. Plant J. (2014)
Targeted insertion of the nptII selectable marker into ArabidopsisADH1 locus using homologous recombination (HR)
LB
Cas9 sgRNA
ADH1
Cas9
LB RBnptIIsgRNA
nptII
RBADH1
Cas9LB RB
nptIIsgRNA
Selected on kanamycin and allyl alcohol

ZmALS2
ZmALS2
Cas9/sgRNA
Adapted from Svitashev et al. Plant Physiology (2015)
Cas9
ZmALS2
ZmALS2P165S+
DNA repair template
Plant cell nucleus
sgRNA
ZmALS2P165S
ZmALS2P165S
ZmALS2P165S
Maize resistant to chlorsulfuron
WTALS2P165S
Chlo
rsulfuro
n(1
00
µg
L−1)
HDR-mediated ZmALS2 allele replacement in maize using CRISPR/Cas9
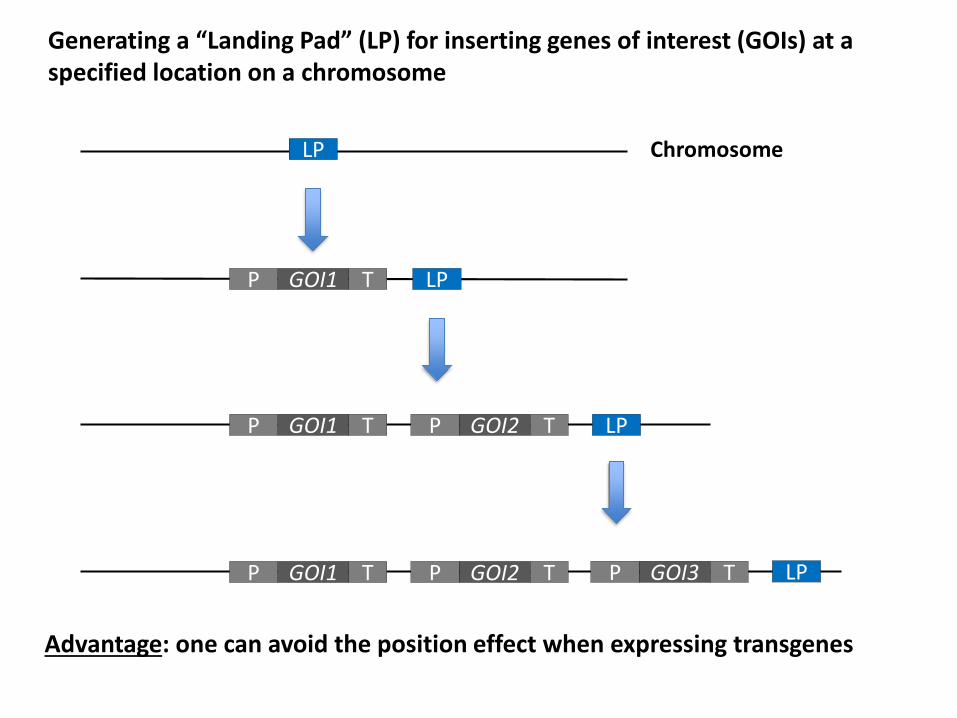
GOI1P T
Chromosome
GOI1P T GOI2P T
GOI1P T GOI2P T GOI3P T
LP
LP
LP
LP
Generating a “Landing Pad” (LP) for inserting genes of interest (GOIs) at aspecified location on a chromosome
Advantage: one can avoid the position effect when expressing transgenes

BARP T exon2 HPT Tintron Chromosome
Cas9P T
P intronexon1 HPT
sgRNAP
GOIP T
DNA repair template/ Cas9 + sgRNA vector
P exon1 HPTGOIP T exon2 HPT Tintron Chromosome
BAR – Basta selectable marker; HPT – Hygromycin selectable marker; GOI – gene of interest; P – promoter; T – terminator
Generating a “Landing Pad” for inserting genes of interest at a specifiedlocation on a chromosome

CRISPR/Cas system
Genome Editing As A Tool To Speed Up Breeding Of Crops With Improved Agronomic Traits

Advantages of genome editing as compared to traditional breeding:
• Speed: mutations can be introduced directly into an elite variety without lengthy crosses with e.g. a related wild species and then backcrosses to the elite line
• No linkage drag: unlike in conventional breeding, a mutation can be introducedinto an elite variety without closely linked genes that may confer an adversephenotype
• Precision: CRISPR practically does not generate off-target mutations, while e.g. EMS mutant lines have thousands of background mutations
• Multiplexing: ability to target a few genes at the same time (e.g. homologues or genesarranged in a gene cluster)

Challenges for CRISPR/Cas-assisted crop breeding:
• need to broadly connect genotype with phenotype - Gene function predictions based on model species e.g. Arabidopsis
- GWAS on accessions that show variation in the trait of interest- Forward genetic screen (e.g. using EMS-mutagenized population)
• aim to develop the technology to level when multiple precise genetic changes can be simultaneously introduced into an elite crop variety
Genetic locus fine-mapping


The Sainsbury Laboratory (Norwich)
Sophien Kamoun
Joe Win
and the rest of the Kamoun Lab
Jonathan Jones
Earlham Institute
Nicola Patron
Oleg Raitskin
Cold Spring Harbor Laboratories
Zachary Lippman
Chris Brooks
Boyce Thompson Institute
Joyce van Eck
MPI Tubingen
Detlef Weigel
Institute of Digital Agriculture,
Zhejiang Academy of Agriculture
Sciences
Congmao Wang
Acknowledgements
Achim DobermannMalcolm Hawkesford
Caroline SparksDamiano MartignagoLucy Hyde