consejo nacional de ciencia y tecnologÍa …glifos.concyt.gob.gt/digital/fodecyt/fodecyt...
TRANSCRIPT

CONSEJO NACIONAL DE CIENCIA Y TECNOLOGÍA -CONCYT-
SECRETARIA NACIONAL DE CIENCIA Y TECNOLOGÍA -SENACYT-
FONDO NACIONAL DE CIENCIA Y TECNOLOGÍA -FONACYT-
UNIVERSIDAD DE SAN CARLOS DE GUATEMALA -USAC-
INFORME FINAL
EVALUACIÓN Y MONITOREO NACIONAL DE INFLUENZA, UTILIZANDO
LA PRUEBA DE INHIBICIÓN DE LA HEMOAGLUTINACIÓN (HI), PARA
DETECCIÓN DE CERDOS SEROPOSITIVOS EN LA REPÚBLICA DE
GUATEMALA
PROYECTO FODECYT No. 62-2009
LUCERO SERRANO ARRIAZA DE GAITÁN
Investigador Principal
GUATEMALA, AGOSTO 2011

1

i
AGRADECIMIENTOS
La realización de este trabajo, ha sido posible gracias al apoyo financiero dentro del
Fondo Nacional de Ciencia y Tecnología, -FONACYT- otorgado por la Secretaria
Nacional de Ciencia y Tecnología, -SENACYT- y al Consejo Nacional de Ciencia y
Tecnología -CONCYT-. Corresponde al proyecto del Fondo de Desarrollo Científico y
Tecnológico -FODECYT-.

ii
OTROS AGRADECIMIENTOS
Se agradece la participación del MV MSc Federico Villatoro, Coordinador de la Escuela
de Postgrado de la Facultad de Medicina Veterinaria y Zootecnia -FMVZ- de la
Universidad de San Carlos de Guatemala -USAC-, por su contribución en el
enriquecimiento del documento,
Se agradece al personal del Laboratorio de Ornitopatología y Avicultura -LOA-, de la
FMVZ/USAC por su ayuda en la preparación del material y en el procesamiento de las
muestras
La obtención de las muestras ha sido posible gracias la colaboración de los profesores y
estudiantes del Ejercicio Profesional Supervisado -EPS-, de la FMVZ/USAC.
Agradecemos la colaboración del personal administrativo, técnicos y médico veterinario
de los rastros municipales de Santa Catarina Pínula y de Quetzaltenango.

iii
TABLA DE CONTENIDO
RESUMEN 1
SUMMARY 2
PARTE I 3
I.1 INTRODUCCIÓN 3
I.2 PLANTEAMIENTO DEL PROBLEMA 6
I.2.1 Antecedentes 6
I.3 OBJETIVOS E HIPÓTESIS 8
I.3.1 Objetivos 8
I.3.1.1 General 8
I.3.1.2 Específicos 8
I.3.2 Hipótesis nula 8
I.4 Metodología 9
I.4.1 Variables 9
I.4.1.1 Variables Dependientes 9
I.4.1.2 Variables Independientes 9
I.4.2 Indicadores 9
I.4.3 Localización 9
I.4.4 Materiales y Métodos 9
I.4.4.1 Metodología Estadística 9
PARTE II 13
II.1 MARCO TEÓRICO 13
II.1.1 Etiología 13
II.1.2 Epidemiologia 14
II.1.3 Patogenia 16
II.1.4 Síntomas 17
II.1.5 Transmisión 18
II.1.6 Diagnóstico 19
II.1.7 Pandemias 22
II.1.8 Influenza Porcina, Implicaciones en Salud Pública 23

iv
PARTE III 31
III.1 resultados y discusión 31
PARTE IV 40
IV.1 CONCLUSIONES 40
IV.2 RECOMENDACIONES 42
IV.3 REFERENCIAS BIBLIOGRÁFICAS 43
IV.4 ANEXO 1: Protocolo de Recepción y Análisis de Muestras 47
IV.5 ANEXO 2: Protocolo de resultado del análisis de las muestras 48
IV.6 ANEXO 3: Registro fotográfico del proyecto FODECYT 62-2009 49
IV.7 ANEXO 4: Títulos de anticuerpos 56
IV.8 ANEXO 5: Currículum de Investigadores 58
IV.9 Informe Financiero 65
IV.10 Cronograma de la investigación 66

v
ÍNDICE DE FIGURAS
Figura 1: Número de muestras obtenidas por departamento de la República de Guatemala, 2010 10
Figura 2: Diagrama del virus de Influenza 14
Figura 3: Línea del tiempo de infección por Influenza Porcina 18
Figura 4: Fases de la Influenza Pandémica 23
Figura 5. Mutación del virus de Influenza 26
Figura 6: Tipo de Virus Respiratorios de casos positivos, en la semana de 1-16 del 2009 en Guatemala.
Análisis realizados por el Laboratorio Nacional de Salud. 27
Figura 7: Casos de Influenza sospechosa en humanos durante el año 2010 en la República de Guatemala. 28
Figura 8: Número de casos de Influenza sospechosos en humanos por regiones de la República de
Guatemala durante el año 2010. 28
Figura 9: Prevalencia de H1N1 en cerdos de explotaciones tecnificadas por áreas de la República de
Guatemala, 2010 33
Figura 10: Prevalencia de Influenza H1N1 en cerdos de explotaciones no tecnificadas por áreas en la
República de Guatemala, 2010 33
Figura 11: Prevalencia de Influenza H3N2 en cerdos de explotaciones tecnificadas, Guatemala, 2010 34
Figura 12: Prevalencia de Influenza H3N2 en cerdos de explotaciones no tecnificadas en la República de
Guatemala, 2010 34
Figura 13: Número de muestras que presentaron seropositividad a ambos serotipos de Influenza (H1N1 y
H3N2) por áreas en la República de Guatemala, 2010. 35
Figura 14: Prevalencia de Influenza H1N1 en cerdos machos de la República de Guatemala, 2010 36
Figura 15: Prevalencia de Influenza H1N1 en cerdas hembras de la República de Guatemala, 2010 36
Figura 16: Prevalencia de Influenza H3N2 en cerdos machos de la República de Guatemala, 2010 37
Figura 17: Prevalencia de Influenza H3N2 en cerdas hembras en la República de Guatemala, 2010 37
Figura 18: Prevalencia de Influenza H1N1 en cerdos jóvenes de la República de Guatemala. 2010 38
Figura 19: Prevalencia de Influenza H1N1 en cerdos adultos de la República de Guatemala, 2010 38
Figura 20: Prevalencia de Influenza H3N2 en cerdos jóvenes de la República de Guatemala, 2010 39
Figura 21: Prevalencia de Influenza H3N2 en cerdos adultos de la República de Guatemala, 2010 39

vi
ÍNDICE DE TABLAS
Tabla 1: Hemaglutinación del virus de influenza con diferentes especies de Glóbulos rojos. 21
Tabla 2: Virus de influenza H1N1 aislado en cerdos, por país y número de casos detectados a partir del
2009. 29
Tabla 3: Número de Muestras y Prevalencia por departamento, por área por tipo de explotación, H1N1 y
H3N2, Guatemala, 2010, Proyecto FODECYT 62-2009 32
Tabla 4: Títulos de Anticuerpos contra Influenza H1N1 de cerdos de explotaciones tecnificadas por áreas
de la República de Guatemala, 2010, Proyecto FODECYT 62-2009 56
Tabla 5: Títulos de anticuerpos contra influenza H1N1 de cerdos de explotaciones no tecnificadas por áreas
de la República de Guatemala, 2010, Proyecto FODECYT 62-2009 56
Tabla 6: Títulos de anticuerpos contar influenza H3N2 de cerdos de explotaciones tecnificadas de la
República de Guatemala, 2010, Proyecto FODECYT 62-2009 56
Tabla 7: Títulos de anticuerpos contra Influenza H3N2 de cerdos de explotaciones no tecnificadas en la
República de Guatemala, 2010, Proyecto FODECYT 62-2009 57

1
RESUMEN
Los virus clásicos de Influenza Porcina, son virus de influenza tipo A que infectan
cerdos, son comunes en América del Norte, América del Sur, Europa y países de Asia.
Los virus H1N1 de América y Asia pertenecen al linaje porcino clásico, son
genéticamente ligados al virus H1N1 humano responsable de la influenza española de
1918. En el año 2000 se presentan los primeros informes del virus Influenza en el campo
veterinario en Guatemala clasificado como virus de origen aviar H5N2 de baja
patogenicidad. La transmisión entre especies puede ocurrir bajo condiciones naturales por
cambios leves en los genes HA y NA. La circulación de virus de Influenza A de origen
porcino con Hemoaglutininas diferentes, debe ser una causa de alerta y preocupación
para las autoridades de salud pública en todo el mundo. La presencia de segmentos
genéticos de virus porcinos, los virus pandémicos es evidencia que existen deficiencias
en la vigilancia en las poblaciones porcinas.
Con el presente proyecto se realizó el primer monitoreo de circulación viral de
Influenza H1N1 y H3N2 en porcinos de la República de Guatemala. El monitoreo se
ejecutó durante el periodo de 10 meses, de febrero a noviembre de 2010, colectándose un
total de 2,055 sueros, de los cuales 1,017 provienen de cerdos de explotaciones
tecnificadas y 1,038 de cerdos de explotaciones no tecnificadas, de diferentes áreas
geográficas de la República de Guatemala. Las muestras fueron analizadas en el
Laboratorio de Referencia Regional de Sanidad Avícola (LARRSA) de la Facultad de
Medicina Veterinaria y Zootecnia -FMVZ-, empleando la técnica de Inhibición de la
Hemoaglutinación (HI) los antígenos H1N1 (A/SW/Iowa/H1N1) y el A/SW/TX/4199/98.
Se consideró positivo para ambos serotipos el suero cuyo título fue ≥ 1:80. La
prevalencia para H1N1 en explotaciones tecnificadas fue de 20.16% y 41.04% en
explotaciones no tecnificadas. Para H3N2 el 6.59% fueron positivas provenientes de
explotaciones tecnificadas, y 16.96% para no tecnificadas. Para ambos serotipos (H1N1 y
H3N2) la positividad fue 5.41% en explotaciones tecnificadas y 14.64% en explotaciones
no tecnificada, lo cual sugiere mayor riesgo de recombinación genética, y de mutación
viral en explotaciones no tecnificadas. Adicionalmente se analizaron 100 sueros de
porcinos provenientes de la República Mexicana a través del contrabando en un rastro
local. De los cuales 7% fueron positivos a H1N1, 6% a H3N2 y 2% a ambos serotipos.
Debido a que las pruebas serológicas no diferencian entre las cepas de H1N1 (cepa
porcina) y la pandémica H1N1/2009, se recomienda emplear pruebas virológicas y
moleculares para la vigilancia epidemiológica.

2
SUMMARY
The classical virus of Swine Influenza infecting pigs is common in North and South
America, Europe and Asia. The virus H1N1 from America and Asia belongs to the
classical swine linage; genetically bind to Human H1N1, responsible of the Spanish Flu
in 1918. The first report of Influenza in Guatemala of avian source, low pathogenicity
H5N2 was on the year 2000. The inter species transmission can occur under natural
conditions by slightly changes in the genes HA and NA. The viral circulation of an
Influenza virus type A of swine source with different hemagglutinins should be a warning
for Public Health authorities around the world. The presence of genetic segments of
swine virus, in the pandemic virus is evidence of the existence of differences in the
surveillance of swine population.
The first surveillance at Guatemala for viral circulation of Influenza H1N1 and
H3N2 in swine was completed over a period of 10 months, from February up to
November 2010 collecting a total of 2055 serums, from which 1038 were from basic
farms and 1017 from modern farms. The samples were analyzed in the Laboratorio de
Referencia Regional de Sanidad Avícola (LARRSA) at Faculty of Veterinary Medicine
and Animal Husbandry (FMVZ), using the Hemagglutination Inhibition technique (HI).
The serums, whose titles were ≥ 1:80, are considered positive for both serotypes.
The prevalence for H1N1 in modern farms was 20.16% and 41.04% in basic farms. For
H3N2 6.59% were positive in modern farms and 16.96% in basic farms. For both
serotypes (H1N1 and H3N2) positivity was 5.41% in modern farms and 14.64% in basic
farms. This suggests a higher risk of genetic recombination and viral mutation on basic
farms. A 100 serums where obtain, in a local slaughterhouse, from pigs smuggled from
Mexico; of which 7% were positive for H1N1, 6% for H3N2 and 2% to both serotypes.
This suggests a high-risk of genetic recombination and viral mutation in basic farms.
Since the serological tests do not distinguish between strains of H1N1 (swine) and
pandemic influenza H1N1 2009, it is recommended to use virological and molecular test
for epidemiological surveillance.

3
PARTE I
I.1 INTRODUCCIÓN
Los virus clásicos de Influenza Porcina, son virus de influenza tipo A, que infectan
cerdos, los subtipos más frecuentes son H1N1, H1N2 y H3N2, las principales cepas
involucradas en la gripe del cerdo son la H1N1 y la H3N2. Esporádicamente se han
aislado otras cepas como la H3N3 (Quebec) y H1N2 (Japón y Europa). (Taylor, 1999)
La Influenza Porcina es común en América del Norte, América del Sur, Europa y
países de Asia, los virus de Influenza H1N1 de Europa y América difieren en sus
componentes genómicos, los virus H1N1 de América y Asia pertenecen al linaje porcino
clásico, que es genéticamente ligado al virus H1N1 humano responsable de la influenza
española de 1918. En contraste los ocho segmentos del virus de Influenza H1N1 que
circula en Europa están filogenéticamente relacionados con el linaje aviar. (Scholtissek,
1990)
Los primeros informes en el campo veterinario del virus Influenza en Guatemala
datan del año 2000, cuando se realiza el primer aislamiento del virus en aves no
tecnificadas, en el Laboratorio de Ornitopatología y Avicultura de la Facultad de
Medicina Veterinaria y Zootecnia de la Universidad de San Carlos de Guatemala
(FMVZ), clasificado posteriormente por el USDA/APHIS como virus H5N2 de baja
patogenicidad (Serrano, Santizo, & Motta, 2002).
Ante la circulación del virus H5N2 en parvadas avícolas y a la costumbre de
producir simultáneamente aves y cerdos se decide en el 2,008 realizar un monitoreo en
cerdos utilizando la prueba de Reacción en cadena de la Polimerasa (PCR)
exclusivamente a nivel de explotaciones no tecnificadas, los resultados obtenidos
demostraron que las piaras en Guatemala, no han sido infectadas por el virus H5N2.
Estudios de Influenza en cerdos, determinando otro tipo de hemaglutininas (H1N1 y
H3N2), no han sido realizados anteriormente. (Marroquín, 2011)
La transmisión entre especies, específicamente de cepas humanas a porcinos puede
ocurrir bajo condiciones naturales por cambios leves en los genes HA y NA.
Los virus Influenza tipo A infectan animales de varias especies, siendo una
importante causa de enfermedad respiratoria en cerdos; las aves pueden infectarse de
forma subclínica, y estas son un reservorio de virus de todas las 16 Hemaglutininas (HA)
y de los nueve (9) subtipos de Neuraminidasa (NA) conocidos. Las infecciones de
Influenza en las poblaciones humanas son comunes y causa aproximadamente 20,000
muertes por año en Estados Unidos (Spickler, 2009).
Los cerdos poseen receptores en las células epiteliales que permiten la infección de
virus influenza, tanto de aves como de otros mamíferos. (Krause, 2006; Scholtissek,
Source for Influenza Pandemics, 1994; Scholtissek, Pigs as the “mixing vessel” for the
creation of new pandemic influenza A viruses, 1990).

4
La influenza pandémica de 1918 fue causada por un virus H1N1 que se transmitió
de aves a humanos y cerdos. Sin embargo, estudios de cepas pandémicas de Influenza
humana revela que algunos segmentos de los genes virales pueden transmitirse a virus
influenza de mamíferos a través de recombinación genética (Acuña L, 2004).
En noviembre de 1976, en una granja al sur de Wisconsin, se aisló el primer caso
de una infección zoonótica de Influenza Porcina en humanos. (Easterday & Hinshaw,
1992)
Generalmente los virus de la gripe H1N1 porcina no se contagian a los seres
humanos. Sin embargo se han registrado en forma esporádica; estos casos se presentan en
personas que tienen contacto directo con los cerdos. El contagio de la gripe H1N1 entre
humanos también es posible, se cree que esto sucede de la misma manera en que se
desarrolla una gripe de temporada en las personas, es decir, principalmente a través de la
tos y los estornudos de personas infectadas con el virus de la influenza (Bronze, 2010).
Olsen, Brammer, Easterday, Arden, Belay, & Cox, 2002, realizaron un estudio
para determinar la existencia de anticuerpos de Influenza Porcina en humanos asociados
ocupacionalmente con los mismos (propietarios, galponeros, veterinarios), sus resultados
apoyan la hipótesis que los humanos son infectados con virus de Influenza Porcina en
forma frecuente. (Olsen, Brammer, Easterday, Arden, Belay, & Cox, 2002)
Infecciones zoonóticas en humanos con virus de la Influenza Porcina, han sido
diagnosticadas en Estados Unidos, Europa, Nueva Zelanda y Asia, sin embargo el
número total de estas infecciones es muy pequeño comparado con el número de personas
directa e indirectamente involucradas con producción porcina. En muchos casos la
sintomatología en humanos por Influenza pandémica H1N1/2009 es indistinguible de las
infecciones por otros tipos de Influenza (Ferrari, y otros, 2009).
En abril del 2009 se reporta por primera vez en México la presencia de un virus de
Influenza pandémica H1N1 en humanos, el estudio genético del mismo reveló ser el
producto de la recombinación genética de cuatro cepas diferentes del virus de Influenza
quienes proporcionaron (i) un segmento genético de la influenza humana, (ii) segmentos
genéticos aviares de América del Norte, (iii) segmentos genéticos de la Influenza Porcina
de América del Norte y (iv) segmentos genéticos aviares similares a las porcinas de
Eurasia, la recombinación de los tres primeros segmentos se cree ha circulado desde hace
más de diez años entre los cerdos, causando enfermedades leves en humanos, sin
embargo la presencia del segmento de Eurasia no se había detectado anteriormente en
cerdos ni en humanos. Aunque la enfermedad producida por el nuevo virus se denominó
“gripe porcina” no se determinó ningún vínculo entre los casos porcinos y humanos, en
tanto que la transmisión de humano a humano fue evidente (Garcia, 1999; Ferrari, y
otros, 2009; Ferrari, y otros, 2009).
En octubre del 2009 se reporta en Japón el primer brote de gripe pandémica H1N1
en cerdos sin sintomatología previa, el diagnóstico se realizó durante la vigilancia
epidemiológica, mediante hisopados nasales, ninguno de los trabajadores de la granja
había presentado manifestaciones gripales hasta un mes antes al aislamiento, las pruebas
realizadas fueron la Prueba de Reacción en Cadena de la Polimerasa y la Inhibición de la
Hemoaglutinación, ambas positivas (Departamento de Salud y Servicios Humanos,
2009).

5
La Organización Mundial de la Sanidad Animal (OIE) define la vigilancia de las
enfermedades de animales como “las operaciones sistemáticas y continuas de
recolección, comparación y análisis de datos zoosanitarios y la difusión de información
en tiempo oportuno a quienes la necesiten para tomar medidas” (OIE, 2008).
En los últimos años, ha habido un incremento en las actividades de vigilancia en
cerdos y aves en Europa y América del Norte, sin embargo, y por innumerables razones
(desinformación, pocos recursos, insuficiente capacidad instaurada), muchos países no
cuentan con información sobre la existencia y evolución de la circulación del virus de
influenza en cerdos para implementar medidas apropiadas así como el asignarle la
preponderancia necesaria en la salud animal y en la salud pública.
Este estudio comprobó la presencia de la circulación de los subtipos H1N1 y H3N2
en cerdos de explotaciones tecnificadas y explotaciones no tecnificadas.

6
I.2 PLANTEAMIENTO DEL PROBLEMA
I.2.1 Antecedentes
En Guatemala el estudio de la Influenza se inicia a partir del año 1996, como parte
del programa de vigilancia epidemiológica consecuente al aparecimiento del brote de
Influenza Aviar de elevada patogenicidad de 1995 en México.
En marzo del año 2000, se realizó el primer aislamiento del virus de Influenza
Aviar en aves no tecnificada en el Laboratorio de Ornitopatología y Avicultura de la
Facultad de Medicina Veterinaria y Zootecnia de la Universidad de San Carlos de
Guatemala, siendo clasificado posteriormente por el USDA/APHIS como virus influenza
H5N2 de baja patogenicidad (Serrano, Santizo, & Motta, 2002).
Heinen (2003) reporta que a nivel mundial un 25% de los animales presentan
anticuerpos de Influenza Porcina, en Guatemala es el primer estudio de virus influenza
H1N1 (cepa porcina) y H3N2, es un estudio pionero cuyo objetivo es apoyar la vigilancia
epidemiológica de los virus Influenza a nivel nacional, determinar la existencia de virus
de Influenza H1N1 y H3N2 en poblaciones porcinas en Guatemala, determinar regiones
que presentan seropositividad a H1N1 y H3N2, existencia de riesgo de transmisión viral
cerdo humano y la diferencia entre la seroprevalencia de explotaciones tecnificadas y no
tecnificadas, entre edades y sexo de los cerdos. Así mismo se evaluó e implementó la
técnica de Inhibición de la Hemoaglutinación para el monitoreo rutinario de Influenza
Porcina.
La transmisión entre especies, puede ocurrir bajo condiciones naturales; los cerdos
poseen receptores en las células epiteliales que permiten la infección de virus influenza,
tanto de aves como de otros mamíferos (Scholtissek, Source for Influenza Pandemics,
1994).
El virus de la gripe porcina H1N1, generalmente no se contagia a los seres
humanos, si bien, se han registrado casos en forma esporádica; estos casos se presentan
en personas que tienen contacto directo con los cerdos. (Bronze, 2010)
Infecciones zoonóticas en humanos con virus de la Influenza Porcina, han sido
diagnosticadas en Estados Unidos, Europa, Nueva Zelanda y Asia, sin embargo el
número total de infecciones zoonóticas es muy pequeño comparado con el número de
personas directa e indirectamente involucradas con producción porcina (Heinen, 2003).
En abril del 2009 se reporta por primera vez en México la presencia de un virus de
Influenza pandémica H1N1, posteriormente en octubre del 2009 se reporta en Japón el
primer brote de gripe pandémica H1N1 en cerdos sin sintomatología previa (HHS, 2009).
Se necesitan, recursos y capacitación para aquellos países que no cuentan con la
información pertinente sobre la existencia y evolución de la circulación del virus de
Influenza Porcina, para que se puedan implementar medidas adecuadas tanto en salud
pública como en salud animal.

7
El presente proyecto se justifica, debido al reciente brote de Influenza pandémica
H1N1/2009; a la posible recombinación entre los serotipos de Influenza Porcina (H1N1,
H3N2) y la potencial recombinación con el virus de Influenza Aviar H5N2 de baja
patogenicidad que se ha presentado desde el año 2,000 en Guatemala, otro argumento
para realizar este estudio, es que los resultados obtenidos de positividad en cerdos,
pueden servir para la toma de decisiones de las autoridades de Salud pública, en este caso
los humanos que conviven con cerdos pueden servir de centinelas para la aparición de
posibles brotes.

8
I.3 OBJETIVOS E HIPÓTESIS
I.3.1 Objetivos
I.3.1.1 General
Evaluar y monitorear a nivel nacional la presencia de Influenza, utilizando la
prueba de Inhibición de la Hemoaglutinación (HI), para detección de cerdos seropositivos
en la República de Guatemala.
I.3.1.2 Específicos
Evaluar a nivel nacional la presencia de los virus de Influenza, utilizando la
prueba de Inhibición de la Hemoaglutinación (HI), para detección de cerdos
seropositivos en la república de Guatemala
Realizar monitoreo a nivel nacional de la presencia de los virus de Influenza
utilizando la prueba de Inhibición de la Hemaglutinación (HI), para la detección
de cerdos seropositivos en la República de Guatemala.
Evaluar los riesgos de transmisión viral cerdo humano.
Evaluar la seroprevalencia de las granjas y áreas a nivel nacional
Declarar áreas, zonas, compartimentos, granjas o el país libre de la infección
Evaluar técnicas de Inhibición de la Hemoaglutinación.
Evaluar e implementar la técnica de Inhibición de la Hemoaglutinación como
prueba rutinaria, para el monitoreo de Influenza Porcina H1N1.
Divulgar a las autoridades, actores sociales e instituciones en el campo de su
competencia la información obtenida de la investigación.
I.3.2 Hipótesis nula
Los cerdos en Guatemala son seronegativos a Influenza Porcina H1N1 y H3N2,
demostrado por la prueba de Inhibición de la Hemaglutinación.

9
I.4 METODOLOGÍA
I.4.1 Variables
Los indicadores a considerar para determinar la prevalencia, están representados
por la presencia de anticuerpos contra los serotipos de H1N1 y H3N2, expresado en
títulos de anticuerpos. Se comparó la prevalencia de la infección en cuanto a explotación
tecnificada y no tecnificada, sexo y edad de los cerdos muestreados y entre las regiones
de Guatemala de donde provienen las muestras.
I.4.1.1 Variables Dependientes
Seropositividad, Títulos de anticuerpos
I.4.1.2 Variables Independientes
Tipo de explotación, Sexo, edad y regiones.
I.4.2 Indicadores
Prevalencia y títulos de anticuerpos
I.4.3 Localización
Guatemala se encuentra en la área Centroamericana y limita al norte con México, al
oeste con Belice y al sur con Honduras y El Salvador, bordeando el Golfo de Honduras.
El relieve se caracteriza por ser montañoso y con mesetas de caliza. Está en la latitud 15º
30' N y la longitud 90º 15' W. Cuenta con 22 departamentos. El área total de Guatemala
es de 108,890 kilómetros cuadrados con una costa que se extiende hasta 400 kilómetros.
El clima de Guatemala es mayormente de tipo tropical pero cálido y húmedo en la tierra
baja y frio en tierras altas.
I.4.4 Materiales y Métodos
I.4.4.1 Metodología Estadística
Se utilizó la fórmula de Cannon 2001, para obtener la muestra de la población
animal (Cannon, 2001).

10
Para determinar la heterogeneidad en las regiones de Guatemala se utilizó la
fórmula de G replicada (Sokal & Rohlf, 1995):
Obtención de sueros
Para determinar la seropositividad de los cerdos a nivel nacional para los virus de
Influenza H1N1 y H3N2 se tomaron 2155 muestras estériles de sangre de la vena yugular
(5 ml) de porcinos procedentes de diferentes departamentos del país, en los rastros
Municipales de Santa Catarina Pínula y Quetzaltenango, en donde se obtuvo muestras
tanto de explotaciones tecnificadas como de no tecnificadas.
Las muestras fueron adquiridas por los investigadores, con apoyo de los
estudiantes de EPS directamente de explotaciones tecnificadas y no tecnificadas, de los
departamentos de Guatemala, Sacatepéquez, Chimaltenango, Alta Verapaz, Baja
Verapaz, Petén, Quiché, Zacapa, El Progreso, Jutiapa, Santa Rosa, Escuintla,
Quetzaltenango, San Marcos, Suchitepéquez, y Retalhuleu; para hacer un total de 1017
tecnificadas y 1038 no tecnificadas; así como 100 sueros procedentes del contrabando de
cerdos de México. Al momento del sangrado, se anotó por cada animal, departamento de
procedencia, sexo y edad de los animales muestreados. El suero se obtuvo mediante
centrifugación trasvasándose a viales estériles. El procesamiento de las muestras para el
estudio serológico fue llevado a cabo en el Laboratorio de Referencia Regional
(LARRSA) de la Facultad de Medicina Veterinaria y Zootecnia, El período de muestreo
así como el análisis serológico se realizó de febrero a Noviembre del año 2010.
Figura 1: Número de muestras obtenidas por departamento de la República de Guatemala, 2010
Fuente: Proyecto FODECYT 62-2009

11
Inactivación y Tratamiento de los sueros
Para eliminar las Hemaglutininas inespecíficas, los sueros fueron tratados con
(RDE) enzima destructora de receptores de Vibrio Cholera, este proceso se realizó en una
cabina de Bioseguridad; se añadió 50µl de suero a 200µl de RDE, se incubó durante toda
la noche (12-18 horas) en baño maría a 37°C, se añadió 150 µl de solución de Citrato de
sodio al 2.5% y se inactivó por calor durante 30 minutos a 56°C. Se mezcló 200 µl de
suero tratado con 25 µl de PBS, se añadió 50 µl de eritrocitos de pavo macho al 50%
para dar una dilución final del suero 1:10. Se agitó e incubó 30 minutos a temperatura
ambiente.
Titulación inicial del antígeno de Influenza (H1N1; H3N2)
Los antígenos inactivados y purificados utilizados en la prueba fueron el
A/SW/Iowa/H1N1 para H1N1 y el A/SW/TX/4199/98 para H3N2.
Colocando una placa de 96 pozos en forma vertical, se agregó 25 µl de PBS con
una pipeta multicanal a cada pozo de la misma, se añadió 25µl del antígeno puro en el
primer foso y se diluyó en forma seriada. Posteriormente, se adicionó 25 µl de glóbulos
rojos de pavo lavados al 0.5% y se colocó en un agitador de placas. A continuación se
incubó a temperatura ambiente 40 minutos para proceder a su lectura, el antígeno se tituló
en forma pareada.
Preparación y lavado de Glóbulos rojos de pavo
Los glóbulos rojos fueron obtenidos de un pavo macho de 8 semanas de edad, al cual se
le realizaron análisis previos y se comprobó que estaba libre de Influenza Aviar. Por
cada mililitro de sangre se adicionó un mililitro de solución de Alsevers, se homogenizó
la mezcla y se transfirió a un tubo de centrifuga, se adicionó PBS (solución salina
buferada pH7.2), hasta llegar a 10 mililitros, se centrifugó 5 minutos a 500 RPM, se
eliminó el sobrenadante; se repitió el procedimiento tres veces hasta eliminar la totalidad
de glóbulos blancos de la muestra, ya lavados los glóbulos rojos se diluyeron para
obtener las concentraciones a usar (50% para el tratamiento de los sueros y 0.5% para la
prueba de hemoaglutinación e inhibición de la hemoaglutinación).
Preparación de la solución de antígeno de trabajo.
Se diluyó el antígeno a manera de obtener 4 Dosis Hemaglutinantes –DHA-/25µl en PBS
pH 7.0
Prueba de inhibición de la hemoaglutinación:
Se adicionó 25 µl de PBS con una pipeta multicanal a todos los pozos de la microplaca,
con la placa en posición horizontal, se colocó 25µl de los diferentes sueros tratados en la
línea H de la línea 1 a la 12, se hicieron diluciones dobles seriadas de la “H a la “A”, se
adicionó 25 µl de antígeno previamente diluido en 4 DHA de la A a la H y de la línea 1 a
la 12, se agitó durante 1 minuto, se incubó a temperatura ambiente durante 30 minutos, se

12
agregó 25 µl de glóbulos rojos lavados al 0.5% se agitó nuevamente e incubó 30 minutos
hasta observar un botón de eritrocitos bien definido en el fondo del pozo, se procedió a
realizar la lectura.
Lectura e Interpretación de Resultados
La lectura de la placa se realizó de izquierda a derecha (pozo 1 al 12 diferentes sueros) y
de abajo hacia arriba (título del mismo suero en diluciones dobles), la positividad fue
considerada al observar la formación de un botón completo de glóbulos rojos, cuando
existió duda de la lectura se inclinó la placa en ángulo de 45°, se buscó la mayor dilución
de suero que mostró una completa inhibición de la hemoaglutinación (formación de
lágrima).
Se tomó como título de HI la dilución más alta de suero que causó una completa
inhibición de las 4 DHA. Para confirmar la validez de los resultados se realizó en cada
corrida la titulación de dos sueros de control de referencia H1N1 y H3N2 (USDA).

13
PARTE II
II.1 MARCO TEÓRICO
II.1.1 Etiología
El agente causal de la Influenza Porcina es el virus de Influenza Tipo A, que
pertenece a la familia Orthomyxoviridae. (Organización Mundial de la Salud, 2009)
La influenza es una enfermedad infecciosa de aves y mamíferos causada por un
virus ARN de la familia de los Orthomyxoviridae, existen tres tipos que constituyen hoy
día los géneros: A, B, C, y Thogotovirus (World Health Organization, 2005). La
Influenza Porcina (IP), es una enfermedad respiratoria común en cerdos, causada por un
virus de Influenza Tipo A. (OIE, 2008; Easterday & Hinshaw, 1992)
El virus es pleomórfico, de simetría helicoidal, envuelto, la membrana lipídica del
virión se deriva de la células del hospedero en las que el virus se replica, posee un
diámetro de 80 a 120 nm dotado de una cadena sencilla de 8 segmentos de RNA
(nucleoproteína) negativos polarmente. (Lamb & King, 2001)
De la superficie del virus se extienden dos glicoproteínas la Hemaglutinina y la
Neuraminidasa, existen en la naturaleza 17 subtipos de Hemaglutinina (HA) y nueve
subtipos de neuraminidasa que al reasociarse genes de distintos virus dan un amplio
espectro de combinaciones identificadas en las aves silvestres, una tercera proteína
transmembrana es la denominada M2 presente de 30-60 moléculas por virión y la
proteína de matriz M1 que forma una capa debajo de la envoltura y es la que le da la
estructura al virus y encapsula la ribonucleoproteina. (Lamb & King, 2001)
El complejo de ribonucleoproteina consiste en Ácido Ribonucleico (RNA) asociado
a nucleoproteínas (NP) así como las polimerasas PA, PB1, y PB2.
Los subtipos H1, H5 y H7 son importantes debido a la inestabilidad de su
Hemaglutinina que se traduce en cuadros respiratorios leves causados por virus de baja
patogenicidad y cuadros septicémicos agudos con elevada mortalidad por virus de alta
patogenicidad.
Dos proteínas no estructurales también están asociadas con el virus, la NS2
encontrada en el virión, y la NS1 encontrada únicamente en células infectadas. (Heinen,
2003)

14
Fuente: http://www.micro.magnet.fsu.edu
II.1.2 Epidemiologia
La Influenza Porcina (IP) ha evolucionado de ser una enfermedad estacional
causada por un genotipo estable a una enfermedad respiratoria endémica causada por
múltiples genotipos sometidos a continuos cambios.
Los cambios de los virus influenza ocurren a través de dos mecanismos el primero
por una alteración en una o ambas glicoproteínas shift (cambio, conversión); es una
transformación dramática en el virus que ocurre cuando dos virus afectan
simultáneamente al mismo individuo y estos determinan que los virus influenza se
dividan en subtipos.
Los subtipos conocidos hasta el momento de Hemaglutinina difieren en un 30%
de la secuencia aminoacídica de la HA1, no presentando reactividad cruzada
serológicamente. (OIRSA, 2009)
La otra forma de mutación denominada drift (tendencia) se refiere a mutaciones
puntuales que ocurren en los genes durante la replicación del genoma viral.
Todos los virus de influenza A han sido aislados de aves, mientras que una menor
variedad han sido aislados de cerdos, caballos y otros mamíferos. Del humano sólo se
han aislado virus con tres Hemaglutininas (H1, H2, H3) y dos Neuraminidasas (N1, N2).
(OIRSA, 2009)
Las principales cepas implicadas en la gripe del cerdo son H1N1 y H3N2,
esporádicamente se han aislado otras cepas como H3N3 (Quebec) y la H1N2 (Japón y
Europa) (Mandell, Bennett, & Dolin, 2006).
El virus de Influenza H1N1 y H3N2 es endémico en las poblaciones de cerdos; se
presenta en los meses de baja temperatura y a veces con el ingreso de nuevos cerdos. Los
estudios han demostrado que un 25% de los animales presentan anticuerpos a la
infección. (Heinen, 2003; Mandell, Bennett, & Dolin, 2006).
Los cerdos son susceptibles a la infección experimental con todos los subtipos de
Influenza Aviar, y recientemente el virus H9N2 ha sido aislado de cerdos en Hong Kong
(World Health Organization, 2005).
Durante 80 años el virus de Influenza que circuló en los cerdos de Norte América
como cepa endémica fue la cepa de influenza H1N1, hasta 1998 cuando emerge una
Figura 2: Diagrama del virus de Influenza

15
recombinación en la población porcina de H3N2, pudiendo ser una doble recombinación
de aves y humanos (A/Sw/NC/98) o tripe recombinación con humanos aves y cerdos
(A/Sw/TC/98).
En 1999 la Hemaglutinina de la preexistente influenza H1N1 se recombinó con un
virus H3N2 para crear una segunda recombinación de virus H1N2.
Otras recombinaciones de virus han sido reconocidas en porcinos incluyendo
H3N2, semejante al humano H1N1 y H1N2 (Mandell, Bennett, & Dolin, 2006).
Henein (2003) cita el mecanismo molecular propuesto para la susceptibilidad de
los cerdos a la infección por virus aviares; los porcinos poseen receptores de
carbohidratos en el epitelio respiratorio, es decir los receptores virales sialoligosacáridos
que están presentes en las células traqueales de los cerdos poseen: acido N-
acetilneuramínico-α 2,3 galactosa (NeuAcα2,3Gal) y Acido N-acetilneuramínico-α 2, 6
galactosa (NeuAcα2,6Gal).
Los cerdos entonces reconocen virus de la gripe porcina, humana y aviar. En
consecuencia, los cerdos se han considerado como “recipientes de mezcla” para el
desarrollo de nuevos virus de la gripe cuando los virus de la gripe porcina, aviar y/o
humana se recombinan en los cerdos.
Mastrovich M, Tusikov, Bovin, Klimov, Kastruchi, 2000, refieren que los virus
humanos tienen preferencia a unirse a los sitios NeuAcα2, 6Gal mientras que los virus
aviares se unen a los NeuAcα2,3Gal esto es muy importante porque el usar carbohidratos
como receptores resulta en un amplio rango de huéspedes. (Mastrosovich, y otros, 2000)
Una vez el virus se adhiere al receptor, la replicación viral ocurre en el tracto
respiratorio de los cerdos y una gran cantidad de virus es producido después de un corto
periodo de incubación de 12 a 24 horas.
Las células epiteliales respiratorias se tornan necróticas y se desprenden en el
lumen alveolar, bronquial o traqueal. Los signos clínicos de neumonía incluyendo
disnea, taquipnea, tos, descarga nasal, letargo y fiebre ocurren brevemente después de la
infección.
El virus es expelido por la nariz durante la tos, los cerdos pueden eliminar el virus
durante 5 a 7 días post infección.
Los síntomas clínicos y la excreción nasal pueden aparecer a las 24 horas de la
infección, es decir se caracteriza por un corto período de incubación (1-3 días) después
del cual los animales parecen anoréxicos, inactivos y tienen la tendencia a acurrucarse.
(Ferrari, y otros, 2009)
La inmunidad desarrollada es incompleta pero efectiva para reducir los virus a no
detectables o insuficientes para una re infección. Cuando la inmunidad se desarrolla, las
lesiones en pulmones y los signos clínicos se resuelven.
La gripe porcina es una enfermedad vírica muy contagiosa que en una piara
afectada puede tener un impacto importante.
Los subtipos de IP que se identifican con más frecuencia en cerdos son H1N1,
H1N2 y H3N2. Otros subtipos identificados en cerdos son H1N7, H3N1, H4N6 y H9N2.

16
Los virus H1N1, H1N2 y H3N2 de Europa son antigénica y genéticamente
distintos de los encontrados en EE.UU.
La circulación de un virus de Influenza A de origen porcino con Hemaglutininas
diferentes, debe ser una causa de alerta y preocupación para las autoridades de salud
pública en todo el mundo. (Garten RJ, 2009;) La presencia de segmentos genéticos de
virus porcinos, en los virus pandémicos es evidencia que existen deficiencias en la
vigilancia en las poblaciones porcinas.
En patos, la mayoría de cepas aviares del virus de influenza, se replica
principalmente en las células que recubren el tracto intestinal y también en los pulmones
y en el tracto respiratorio superior. Los virus toman acceso pasando a través del tracto
digestivo del pato, a causa del bajo pH de la molleja son eliminados en altas
concentración en las heces. Los signos clínicos asociados con infecciones por influenza
tipo A varían considerablemente con la cepa del virus. La infección en los patos con
muchas cepas del virus de influenza son asintomáticas, sin embargo algunas cepas
producen infección sistémica acompañada con sintomatología del sistema nervioso
central y muerte. (World Health Organization, 2005)
II.1.3 Patogenia
El periodo de incubación en la Influenza varía según la especie, vía de exposición y
dosis de virus, de tres horas a tres días, la patogenicidad varia de inaparente a 100% de
mortalidad (Swayne, 2008)
La infección inicia con la destrucción de células que cubren el tracto respiratorio,
incluidos tráquea y bronquios; el virión penetra principalmente por vía aerógena hasta la
nasofaringe y se disemina por el tracto respiratorio infectando células susceptibles. El
virus debe primero atravesar las secreciones respiratorias, que contienen una gran
cantidad de mucoproteinas, la cual es hidrolizada por la neuraminidasa viral. (Easterday
& Hinshaw, 1992; Swayne, 2008)
La adhesión del virus a la célula susceptible se produce por la interacción entre la
Hemaglutinina y los receptores de ácido sálico. La Hemaglutinina tiene un sitio
específico de reconocimiento en su cabeza globular a modo de una pequeña hendidura
donde alberga al ácido siálico de la glicoproteína receptora de la célula hospedadora. La
proteína NA también puede reconocer al receptor mediante un sitio análogo.
El ingreso del virus dentro de la célula es dado por endocitosis. En la vacuola
cenocítica se produce una disminución del pH (acidificación) que induce un cambio
conformacional de la HA, dando lugar a la fusión de la membrana del endosoma y la
envoltura vírica.
Dentro del endosoma, el canal iónico, formado por la proteína M2, expone al
interior del virus a un pH bajo, lo que favorece la disociación de la proteína M1 del
complejo ribonucleoproteina vírica (vRNPs), quedando libres en el citoplasma. Las
vRNPs son transportadas al núcleo de la célula por señales de localización nuclear,
contenidas en el complejo ribonucleoprotéico -RNP- (PB1, PB2 y NP), donde tiene lugar
la transcripción.

17
El primer paso del proceso de transcripción consiste en la ruptura, mediante una
endonucleasa codificada por el virus, de la región del extremo 5´ del ARN mensajero -
ARNm- donador de 10 a 13 nucleótidos corriente abajo del “capuchón”. Aunque las
ribonucleasas dejan generalmente en el extremo 3´ un grupo fosfato en sus productos de
digestión, la endonucleasa de los virus de influenza genera productos con un grupo
hidroxilo en dicho extremo. De esta forma, el oligonucleótido resultante puede actuar
directamente como iniciador o “primer” sin necesidad de desfosforilación.
La síntesis de los distintos ARNm se lleva a cabo por el complejo ARN polimerasa,
dependiente del ARN del virus. Cinco de los ocho segmentos de ARN vírico se
transcriben de forma monocistrónica y se traducen en las proteínas HA, NA, NP, PB2 y
PA, sin embargo, tres de los segmentos de ARN vírico se transcriben, cada uno de ellos,
en dos ARN mensajeros mediante un mecanismo de procesamiento de transcripto. Los
genes M, NS, PB1 son traducidos en diferentes marcos de lectura generando M1 y M2,
NS1 y NS2, y PB1 y PB1-F2, respectivamente. (Swayne, 2008)
La NP y la NS1 son proteínas tempranas que se sintetizan en los primeros
momentos de la infección (transcripción primaria). La síntesis de ARNm celular queda
bloqueada.
La NP y la NS1 migran al núcleo. Se cree que un incremento en la concentración
de la NP libre cambia la síntesis de ARNm víricos por ARNc y ARNv. Los ARN víricos
sintetizados de nuevo son encapsulados con la NP en el núcleo y sirven como molde para
la transcripción secundaria de los ARN mensajeros víricos. Los productos mayoritarios
de la traducción, en esta fase tardía de la infección, son M1, HA y NA.
El ARN del virión unido a las nucleoproteínas abandona el núcleo celular y se
dirige hacia la membrana junto con proteínas víricas transmembranales (Hemaglutinina,
neuraminidasa y proteína M) y una capa subyacente de proteína M1 que se acumulan en
determinadas regiones de la membrana celular y protruyen sobre la membrana en esas
zonas, liberando finalmente un nuevo virus completo, con envoltura, en el fluido
extracelular.
Durante la infección el epitelio ciliado de las vías respiratorias altas son las
primeras células infectadas. Poco después de la infección sobreviene la necrosis de estas.
En esta fase puede haber una descamación extendida del epitelio respiratorio con lo que
se presentan los primeros signos respiratorios de la enfermedad. El virus se disemina
fácilmente a otros órganos, pudiendo afectar incluso el sistema nervioso central.
El virus se elimina a través de cualquier secreción y de heces fecales, por lo tanto,
las formas probables de transmisión incluyen tanto contacto directo entre animales
infectados y susceptibles como contacto indirecto, con aerosoles y fómites.
II.1.4 Síntomas
Las infecciones por IP causan una enfermedad respiratoria caracterizada por tos,
estornudos, descargas nasales, temperaturas rectales elevadas, letargo, dificultad
respiratoria y apetito disminuido.
Se pueden observar respiraciones abdominales con la boca abierta y los
movimientos van acompañados de ataques fuertes de tos. Se puede presentar

18
conjuntivitis, secreciones nasales y estornudos. Cuando a los animales infectados por IP
se les obliga a moverse, la fatiga respiratoria se vuelve más evidente.
Figura 3: Línea del tiempo de infección por Influenza Porcina
Fuente: Marie Gramer, DVM, PHD, Universidad de Minnesota, Laboratorio de diagnóstico veterinario.
En las infecciones por IP las tasas de morbilidad pueden alcanzar el 100%, aunque
las tasas de mortalidad son en general bajas. El impacto económico más importante está
relacionado con la pérdida de peso, que se traduce en un mayor número de días para
alcanzar el peso adecuado para el mercado.
En el cerdo se dan dos formas de la enfermedad, la epidémica o la endémica. En la
forma epidémica el virus pasa por todas las fases con una recuperación rápida del cerdo si
no existen factores complicantes bacterianos. En la forma endémica los síntomas pueden
ser menos aparentes, y no todos los cerdos muestran los síntomas clínicos tradicionales
de la infección (Panamerican Health Organization, 2009)
Entre los agentes que pueden originar una enfermedad respiratoria similar en cerdos
están el virus del síndrome reproductivo y respiratorio porcino, el virus de la enfermedad
de Aujeszky (seudorabia), el coronavirus respiratorio porcino, Actinobacillus
pleuropneumoniae y Mycoplasma hypopneumoniae, aunque la mayoría de éstos tienen
otros síntomas que no son similares a los de la gripe porcina; sólo Actinobacillus
pleuropneumoniae, en infección aguda, presenta síntomas clínicos similares a la gripe
porcina, como disnea, taquipnea, respiración abdominal, tos, fiebre, depresión y anorexia.
II.1.5 Transmisión
La Influenza Porcina no es una enfermedad de declaración obligatoria. La
transmisión se realiza por contacto con secreciones, como los aerosoles que se originan
por la tos y el estornudo, y por descargas nasales. (OIE, 2008)
Pueden darse infecciones por IP en humanos y se han descrito algunas muertes.
Deben tomarse precauciones para impedir las infecciones humanas.

19
Anteriormente, el CDC recibía informes de aproximadamente un caso de contagio
humano con el virus de influenza H1N1 cada uno a dos años en los EE.UU., pero desde
diciembre de 2005 hasta febrero de 2009, se reportaron 12 casos de contagio humano con
influenza H1N1.
El contagio de los virus de influenza H1N1 no se produce a través de los alimentos.
No puede contagiarse influenza H1N1 por ingerir cerdo o productos derivados del cerdo.
La ingesta de cerdo cocido y manipulado adecuadamente es segura ya que la cocción de
la carne porcina a una temperatura interna de 160°F mata el virus de la gripe H1N1 como
lo hace con otros virus y bacterias.
II.1.6 Diagnóstico
Identificación del agente: La identificación del virus es más fácil cuando se
recogen muestran a las 24–48 horas de la aparición de los síntomas clínicos.
El cerdo de elección es un animal sin tratamiento, con manifestación aguda y
temperatura rectal elevada.
El virus puede detectarse fácilmente en el tejido pulmonar y en frotis nasales.
El aislamiento se puede realizar en huevos embrionados de pollo de 10 días de edad
incubados a 35°C. (FAO, 2009)
Para el aislamiento en cultivo celular, se requieren células Madin-Darby de riñón de
perro (MDCK), inoculando 200 µl de espécimen en cada botella, chequeando cada 24
horas para observar el efecto citopático.
La evolución en los virus de influenza puede resultar en cambios en la
susceptibilidad del huésped. Los virus de Influenza Aviares usualmente crecen mejor en
embriones de pollo. Los virus de influenza de mamíferos, especialmente de cerdos,
pueden crecer pobremente en embriones de pollo, por lo que deben de ser aislados en
células MDCK (World Health Organization, 2005).
La subtipificación de los virus aislados se hace por pruebas de inhibición de la
hemaglutinación (HI) y de inhibición de la Neuraminidasa. Es de gran importancia que
el material clínico obtenido de cerdos o aves sea procesado en laboratorios en donde no
se procesen muestras humanas (World Health Organization, 2005)
La detección directa por Inmunoensayo cromatográfico, es una forma rápida de
detección de influenza tipo A y B, ha existido en el mercado desde hace varios años.
Pueden llevarse a cabo en cualquier laboratorio ya que todos los reactivos y materiales
están incluidos en el kit. El resultado puede ser obtenido en 15 a 20 minutos. El mayor
beneficio en estudios de influenza animal es la detección de huéspedes infectados. Las
muestras para este ensayo son hisopados traqueales, nasales o de garganta. Las muestras
fecales no son de elección, ya que pueden dar falsos resultados. Es utilizado para
detección de influenza A en embriones o cultivos celulares inoculados con material
sospechoso. (World Health Organization, 2005)
Para la prueba Directigen Flu, el primer paso es tratar la muestra con un buffer de
extracción conteniendo detergente para lisar las células y moco. La muestra es
homogenizada y luego filtrada por una membrana, el antígeno viral se une a esta

20
membrana, luego el material no unido es lavado y las uniones no específicas de los
reactivos subsiguientes se previenen por el bloqueo de la membrana con IgG de conejo.
Anticuerpos monoclonales marcados con enzima se adicionan y se repite el lavado, la
actividad de la unión de la enzima es detectada por incubación subsecuente de substratos
cromógenos. En presencia de antígeno viral se observa un triángulo purpura. El antígeno
viral en una pequeña área en el centro de la membrana sirve como control, los controles
positivos y negativos están incluidos en el kit. (World Health Organization, 2005)
El FLU OIA, involucra la extracción y detección de la nucleoproteina A o B, el
inmunoensayo óptico es visible a través del cambio físico en el grosor de una tira delgada
molecular, lo cual altera el patrón de luz reflejado y se percibe con un cambio de color.
(World Health Organization, 2005)
Pruebas serológicas
Existen otras pruebas para el diagnóstico serológico de influenza como la fijación
de complemento, Inmunoensayo enzimático e inmunodifusión en agar gel, sin embargo
la prueba de HI es preferida. (World Health Organization, 2005)
La prueba serológica más importante para la detección de anticuerpos contra el IP
es la prueba HI realizada con pares de sueros siendo específica para los diferentes
subtipos. (OIE, 2008)
Se consideran como positivas las muestras cuyos Títulos son iguales o mayores a
1:80 en la técnica de inhibición de la hemoaglutinación (HI) (Rodríguez Torres, Ramírez
Mendoza, Carreón Nápoles, & Mercado García, 1996)
Para confirmar la enfermedad se recomienda analizar sueros recolectados en un
intervalo de 10-21 días; al observarse un aumento de los títulos de anticuerpos sugiere
una infección reciente con el virus de IP.
Otras pruebas serológicas adicionales para el diagnóstico de la infección, son la
prueba de inmunodifusión en gel de agar, la inmunofluorescencia indirecta, la
neutralización vírica y ELISA. (OIE, 2008).
Dentro de los reportes previos en América utilizando la prueba de Inhibición de la
Hemoaglutinación se citan los realizados en Colombia durante 1971 y 1972, cuando se
detectan anticuerpos específicos contra Influenza en porcinos en el departamento de
Antioquia, reportando 14% frente al serotipo H3N2 (Hanssen, Hincapié, & López T,
2003)
Rodríguez Torres et al 1994 realizando un muestreo a nivel de un rastro en la
República de México y utilizando la técnica de inhibición de la hemoaglutinación para
detectar anticuerpos contra Influenza Porcina H1N1, reporta una positividad 20.25%.
En Colombia se realizó un monitoreo serológico con sueros de porcinos colectados
en granjas de producción intensiva entre 1997 - 1999 y durante los años 2000 - 2001, con
el fin de determinar la presencia de reactores serológicos frente al virus de la Influenza
Porcina subtipos H1N1 y H3N2 a través de la técnica de Inhibición de la
Hemaglutinación. En el primer estudio se analizaron 621 sueros por el serotipo H1N1 y
472 muestras para el subtipo H3N2, en el segundo monitoreo se analizaron 300 sueros
porcinos para el subtipo H3N2. Se observó una reactividad serológica del 0.8% y del
41.3% para los serotipos H1N1 y H3N2 respectivamente, en los sueros colectados entre

21
1997 - 1999. En las muestras colectadas durante los años 2000 - 2001 la reactividad
serológica frente al subtipo H3N2 fue del 10%. El mayor porcentaje de reactores
serológicos se detectó en el grupo de cerdas multíparas. En este estudio se sugiere que
siendo la enfermedad exótica en Colombia los títulos positivos provienen de animales
importados con historia de vacunación (Mogollón, y otros, 2009).
Ramírez realizando un estudio de Influenza Porcina en el año 2005 en granjas
tecnificadas en la República de Venezuela reporta porcentajes muy parecidos para ambos
serotipos 7.9% para H1N1 y 8.2 % para H3N2.
Piñeryo et al en el año 2006 en Argentina, reporta una prevalencia para H1 de
84% y 73.8% para H3, no observando diferencias significativas en la prevalencia de
H1N1 y H3N2 a nivel de granja, sin embargo si observó diferencias significativas en la
evaluación individual de los cerdos (Piñeyro, Cappuccio, Machuca, Teodoroff,
Baumeister, & Perfumo, 2006).
Janke, 2000 realizando un estudios serológico de la enfermedad a través del tiempo
en los Estados Unidos, informa de variaciones tendientes a la disminución en la
prevalencia del subtipo H3N2 del 46% en el año 1999 a 35% en el año 2000 (Janke,
2000).
En el año 2000 Richard Webby y col., al estudiar la evolución de los virus de
influenza H1N1 y H3N2 en los Estados Unidos, al analizar 4,382 sueros sanguíneos
provenientes de 23 estados, con la técnica de inhibición de la hemoaglutinación, reporta
que 28,3% de los animales han sido expuestos al virus clásico H1N1, mientras que el
20.5% han sido expuestos a la triple recombinación H3N2.
Tabla 1: Hemaglutinación del virus de influenza con diferentes especies de Glóbulos rojos.
Pollo Pavo Cobayo Tipo O
humana
Concentración 0.5% 0.5% 0.75% 0.75%
Microplaca “V” “V” “U” “U”
Tiempo de
incubación a
25°C,
30 min 30 min 1 hora 1 hora
Apariencia de
glóbulos rojos
controles.
Botón* Botón* Halo Halo
______________________
* = Fluye cuando titula
Fuente: (World Health Organization, 2005)

22
Los eritrocitos o glóbulos rojos de aves y mamíferos hemoaglutinan virus de
influenza. Los eritrocitos de pollo son utilizados con más frecuencia para las pruebas
porque el tiempo de sedimentación es más corto y los patrones de inhibición son claros.
Se sabe que algunas cepas de influenza durante los pasajes iniciales pueden no
hemoaglutinar los eritrocitos de pollo. Los eritrocitos de cobayo han sido constantemente
más sensibles para el diagnóstico de virus de influenza humana. Para la selección de los
eritrocitos debe tomarse en cuenta el huésped de origen de los aislamientos de virus de
influenza o la cepa de referencia. Es importante el uso de sueros control, ya que estos
deben de dar un resultado constante. (World Health Organization, 2005).
Las moléculas no específicas presentes en el suero son capaces de unirse a la
Hemaglutinina, resultando en inhibiciones no específicas y produciendo interpretaciones
falsas. Este efecto ocurre a causa de que los componenentes del suero contienen residuos
de ácido sialico que imitan a los receptores de los glóbulos rojos y compiten con los
receptores de la Hemaglutinina de influenza. Tres tipos moleculares de inhibidores
designados como alfa, beta y gama han sido descritos en sueros de humanos y animales.
Los inhibidores muestran diferentes niveles de actividad contra la Hemaglutinina con
diferentes cepas de influenza. Existen diferentes métodos para inactivar las
hemaglutininas inespecíficas en el suero (World Health Organization, 2005).
La prueba de inhibición de la hemoaglutinación es útil como prueba rutinaria para
la detección y subtipificación de Virus influenza tipo A. Sin embargo, el uso de técnicas
moleculares para la detección directa de las muestras facilita la rápida identificación y
caracterización genética de los virus de influenza A. (World Health Organization, 2005).
II.1.7 Pandemias
Desde 1900, se han producido tres pandemias y varias "amenazas de pandemia" de
Influenza.
La pandemia española es la catástrofe que se toma como parámetro para evaluar al
resto de las pandemias modernas; se calcula que entre un 20 a un 40 por ciento de la
población mundial enfermó y más de 50 millones de personas fallecieron entre
septiembre de 1918 y abril de 1919; se registraron alrededor de 675,000 muertes
provocadas por la gripe únicamente en los Estados Unidos. (HHS, 2009)
Uno de los aspectos poco comunes de la gripe española era su capacidad de
provocar la muerte en adultos jóvenes. Aún se desconocen las causas de tal fenómeno.
(Departamento de Salud y Servicios Humanos, 2009)

23
Figura 4: Fases de la Influenza Pandémica
Fuente: Bronze, M.S, 2009
II.1.8 Influenza Porcina, Implicaciones en Salud Pública
La Influenza Porcina fue observada inicialmente durante la pandemia en 1918, el
virus fue clasificado hasta 1930, como virus IP H1N1; en los Estados Unidos este virus se
ha mantenido y conservado antigénicamente, mientras que en Europa, el virus concluida
la pandemia desapareció, reapareciendo nuevamente en el año 1976, en 1979 es
reemplazado por virus aviares H1N1 similares a los porcinos, pero antigénicamente
distinguibles de los originales H1N1 (Heinen, 2003)
En febrero de 1957, se presenta la pandemia de influenza asiática (H2N2) en el
Lejano Oriente. La inmunidad a esta cepa no era común en las personas menores de 65
años de edad; a modo de preparación, se comenzaron a generar vacunas a finales del mes
de mayo de 1957, las autoridades de salud incrementaron la vigilancia de los brotes de
gripe. (Beveridge, 1977)
A diferencia del virus que provocó la pandemia de 1918, el virus de la pandemia de
1957 se identificó rápidamente gracias a los avances en materia de tecnología científica.
La vacuna estuvo disponible en cantidades limitadas en agosto de 1957 (Beveridge, 1977;
Acuña L, 2004).
En enero y febrero de 1958, se registró una nueva ola de la enfermedad entre los
ancianos. Esto ejemplifica la posible "segunda ola" de infecciones que se puede
desarrollar durante una pandemia. La enfermedad infecta a un grupo de personas en
primer lugar, luego parece disminuir y se incrementa más tarde en un sector diferente de
la población. A pesar de que la pandemia de gripe asiática no fue tan severa como la de

24
gripe española, alrededor de 69,800 personas fallecieron en los EE. UU. (Departamento
de Salud y Servicios Humanos, 2009).
A comienzos de 1968, se produce una nueva pandemia ahora con un virus H3N2; la
influenza de Hong Kong. Los primeros casos de la enfermedad en EE.UU. se registraron
en septiembre del mismo año, pero la enfermedad no se generalizó en los EE.UU. hasta el
mes de diciembre. El número de muertes entre septiembre de 1968 y marzo de 1969
provocadas por esta pandemia ascendió a 33,800, convirtiéndola en la pandemia menos
severa del siglo 20. El mismo virus regresó en el año 1970 y 1972. El virus H3N2
humano se transmitió a los porcinos y circuló en la población porcina en Europa y Asia
durante algunos años causando esporádicamente casos clínicos; no fue sino hasta 1984
que incrementa su virulencia, probablemente por una recombinación con un virus porcino
H1N1, este nuevo virus presentaba Hemaglutininas y Neuraminidasas del virus humano
pero con proteínas internas de origen aviar, llegando a reemplazar al virus original H3N2,
recientemente este virus ha comenzado a circular en Estados Unidos, donde causa
enfermedad severa y patologías reproductivas en cerdas (Heinen, 2003; Departamento de
Salud y Servicios Humanos, 2009; Beveridge, 1977).
Un virus nuevo denominado la "gripe asesina" H1N1 se identificó por primera vez
en Fort Dix, en Nueva Jersey (1976), EE.UU. El virus despertó una gran preocupación
por parte de los expertos quienes pensaban que dicho virus podía estar relacionado con el
virus de la gripe española de 1918. La idea de que una pandemia pudiera azotar al
mundo, alentó una campaña de vacunación masiva en los Estados Unidos. De hecho, el
virus después conocido como "gripe porcina" nunca se extendió más allá del área de Fort
Dix. Las posteriores investigaciones sobre este virus revelaron que en caso de haberse
diseminado, el virus probablemente hubiese sido mucho menos mortal que la gripe
española. (Departamento de Salud y Servicios Humanos de EE. UU. (Gaydos, Top,
Hodder, & Russell, 2006)
En el mes de mayo de 1977, los virus A/H1N1 de influenza aislados en la región
norte de China se diseminaron con rapidez causando una enfermedad epidémica de
alcance mundial en los niños y los adultos jóvenes (< 23 años). El virus de 1977 era
similar a otros virus A/H1N1 que ya habían circulado antes de 1957. (En 1957, el virus
A/H1N1 fue reemplazado por los virus nuevos A/H2N2). Debido al momento de la
aparición de estos tipos de virus, las personas nacidas antes del año 1957 podrían haber
estado expuestas a los virus A/H1N1 y haber desarrollado así una inmunidad contra los
virus A/H1N1. Por lo tanto, cuando el virus A/H1N1 apareció nuevamente en 1977,
mucha gente mayor de 23 años de edad ya contaba con algún tipo de protección en contra
del virus y fueron las personas más jóvenes quienes contrajeron las infecciones causadas
por el virus A/H1N1. Antes de enero de 1978, el virus ya se había diseminado por el
mundo entero, incluso en los Estados Unidos, ya que la enfermedad afectó
principalmente a los niños, el episodio no fue considerado una verdadera pandemia. Las
vacunas que contenían este tipo de virus no se generaron a tiempo para la temporada
1977-78, pero el virus se incluyó finalmente en la vacuna de 1978-79 (Acuña L, 2004;
Beveridge, 1977).
Entre diciembre de 2005 y enero de 2009, se reporta al CDC un incremento en la
notificación de virus Influenza H1N1 porcina de 1 a 2 casos por año hasta 12 casos de
infección en humanos; cinco de esos casos ocurrieron en pacientes que tuvieron contacto

25
directo con los cerdos, seis pacientes que reportaron proximidad a cerdos, y en un caso el
contacto se desconoce. (Departamento de Salud y Servicios Humanos, 2009)
En Estados Unidos se presenta una nueva pandemia, en marzo del año 2009
implicando nuevamente a un virus Influenza Tipo A H1N1, el llamado “caso cero”, una
niña de nueve años de edad, presentó un cuadro de tos y fiebre superior a los 40 grados
centígrados, extendiéndose a nivel mundial. El 13 de abril de 2009, se notificó al CDC
de una enfermedad respiratoria en un niño de 10 años procedente de San Diego,
California. El niño presentaba tos, fiebre y vómitos, los cuales se informó habían
aparecido el 30 de marzo, el niño se recuperó sin problema, ese mismo día, en el Hospital
General de Oaxaca, había fallecido una paciente, a causa de una neumonía provocada por
lo que hasta entonces se pensaba que era un coronavirus. El 14 de abril de 2009 la CDC
confirma al agente etiológico como un virus influenza (H1N1), el cual se llamó
posteriormente como Influenza Tipo A/H1N1. (Departamento de Salud y Servicios
Humanos, 2009)
Posteriormente el CDC confirma que el origen de la infección es una variante de la
cepa H1N1, con material genético proveniente de una cepa aviaria, dos cepas porcinas y
una humana que sufrió una mutación dando un salto inter especies (o heterocontagio) de
los cerdos a los humanos, llegando a contagiarse de persona a persona (Panamerican
Health Organization, 2009; Radio France International, 2009; Departamento de Salud y
Servicios Humanos, 2009).
Los estudios en ecología de los virus de influenza han llevado a la hipótesis que
todos los virus de influenza mamífera derivan de un reservorio de Influenza Aviar.
Análisis filogenético del gen de la glicoproteína muestran que los virus de Influenza
Aviar han evolucionado en cinco linajes especifico de huéspedes, un linaje equino que no
se ha aislado en más de 15 años y uno recientemente aislado, un linaje de gaviotas, uno
de cerdos y un linaje de humanos (World Health Organization, 2005).

26
Figura 5. Mutación del virus de Influenza
Fuente: htt://www3.niaid.nih.gov/news/focuson/flu/illustration/antigenic/antigenicshift.htm
Este brote de Influenza Tipo A (H1N1) se expandió a nivel mundial. El ingreso de
la Influenza pandémica H1N1 a Guatemala tuvo serias repercusiones económicas en la
industria porcina, debida a la baja en el consumo de carne y subproductos (Panamerican
Health Organization, 2009).
Generalmente los virus de la gripe H1N1 no se contagian a los seres humanos. Sin
embargo, se han registrado casos en forma esporádica (HHS, 2009; Departamento de
Salud y Servicios Humanos, 2009).
Ayora Talavera, Cadavieco y Canul realizando un estudio en poblaciones mayas en
la península de Yucatán, con condiciones de producción de cerdos no tecnificados
similares a las guatemaltecas encontraron una positividad en humanos a influenza H1 del
26% superando la seropositividad a influenza H3 de un 80.8%, encontrando,
mayormente, estos casos se presentan en personas que tienen contacto directo con los
cerdos. El contagio de la gripe H1N1 entre humanos también es posible, se cree que esto
sucede de la misma manera en que se desarrolla una gripe de temporada en las personas,
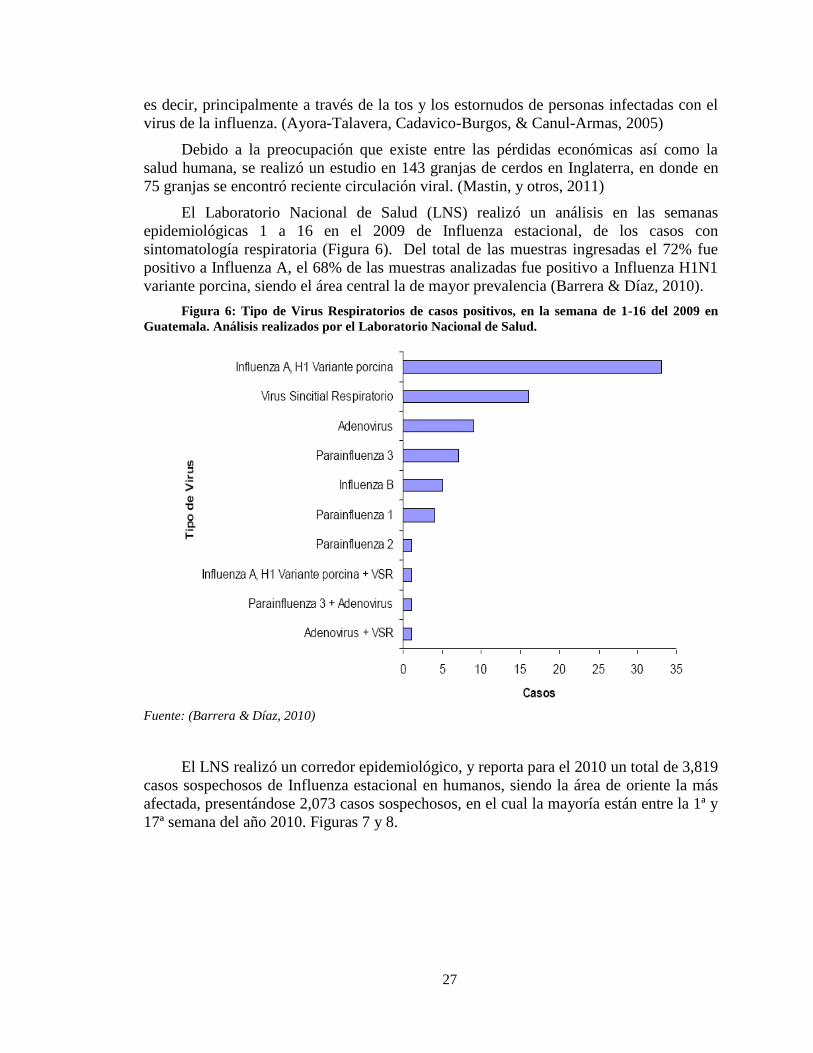
27
es decir, principalmente a través de la tos y los estornudos de personas infectadas con el
virus de la influenza. (Ayora-Talavera, Cadavico-Burgos, & Canul-Armas, 2005)
Debido a la preocupación que existe entre las pérdidas económicas así como la
salud humana, se realizó un estudio en 143 granjas de cerdos en Inglaterra, en donde en
75 granjas se encontró reciente circulación viral. (Mastin, y otros, 2011)
El Laboratorio Nacional de Salud (LNS) realizó un análisis en las semanas
epidemiológicas 1 a 16 en el 2009 de Influenza estacional, de los casos con
sintomatología respiratoria (Figura 6). Del total de las muestras ingresadas el 72% fue
positivo a Influenza A, el 68% de las muestras analizadas fue positivo a Influenza H1N1
variante porcina, siendo el área central la de mayor prevalencia (Barrera & Díaz, 2010).
Figura 6: Tipo de Virus Respiratorios de casos positivos, en la semana de 1-16 del 2009 en
Guatemala. Análisis realizados por el Laboratorio Nacional de Salud.
Fuente: (Barrera & Díaz, 2010)
El LNS realizó un corredor epidemiológico, y reporta para el 2010 un total de 3,819
casos sospechosos de Influenza estacional en humanos, siendo la área de oriente la más
afectada, presentándose 2,073 casos sospechosos, en el cual la mayoría están entre la 1ª y
17ª semana del año 2010. Figuras 7 y 8.

28
Figura 7: Casos de Influenza sospechosa en humanos durante el año 2010 en la República de
Guatemala.
Fuente: Ministerio de Salud Pública y Asistencia Social
Figura 8: Número de casos de Influenza sospechosos en humanos por regiones de la República de
Guatemala durante el año 2010.
Fuente: Ministerio de Salud Pública y Asistencia Social
En Guatemala la vigilancia de Influenza en cerdos es inadecuada porque la
enfermedad no es de notificación obligatoria (MAGA), así mismo las implicaciones
económicas que representa la mención simultanea de las palabras cerdo e influenza hace
que la mayoría de porcinocultores y veterinarios no hayan hecho de las pruebas
serológicas una actividad rutinaria.
0
20
40
60
80
100
120
1 5 9 13 17 21 25 29 33 37 41 45 49
Nú
me
ro d
e ca
sos
sosp
ech
oso
s
Semana
Central
Sur
Occidente
Oriente
Norte
0
500
1000
1500
2000
Central Sur Occidente Oriente Norte

29
Tabla 2: Virus de influenza H1N1 aislado en cerdos, por país y número de casos detectados a partir
del 2009.
Fuente: Gray and Baker, 2011)
AÑO FECHA DE INICIO DEL
EVENTO PAÍS
NÚMERO DE
CASO
2009
Abril 30 México 2
Agosto 18 Irlanda 1950
Agosto 27 Indonesia Sin reporte
Septiembre 2 Irlanda 4,500
Septiembre 22 Irlanda 4
Septiembre 25 Irlanda (2 focos) 105
Octubre 2 Japón 10
Octubre 9 Noruega 196
Octubre 22 EEUU (Indiana) Sin reporte
Octubre 23 Irlanda Sin reporte
Octubre 24 Islandia 10
Noviembre 6 Islandia 9
Noviembre 10 Noruega 83
Noviembre 10 Rusia 45
Noviembre 18 Finlandia 800
Noviembre 23 Italia 375
Noviembre 27 Irlanda Sin reporte
Diciembre 2 Inglaterra 58
Diciembre 3 Italia 9
Diciembre 4 Tailandia 34
Diciembre 14 Corea 334
Diciembre 22 Inglaterra 1124
2010
Enero 4 Dinamarca 1670
Enero 7 Hong Kong 1
Enero 12 Japón 164
Enero 13 Dinamarca Sin reporte
Enero 18 Serbia 20
Abril 12 Corea 36
Agosto 23 Finlandia 1500
Septiembre 2 Taipei (China) 4

30
La Organización Mundial de la Salud ha establecido una red mundial para la
vigilancia de la influenza humana. La meta principal de esta red es el detectar e
identificar nuevas variantes epidémicas emergentes, y contribuir a la selección apropiada
de cepas de vacunas. (World Health Organization, 2005)

31
PARTE III
III.1 RESULTADOS Y DISCUSIÓN
Es importante conocer la distribución e incidencia de las enfermedades,
especialmente de aquellas que poseen un potencial zoonótico. En este estudio se evalúo
la presencia de Influenza H1N1 y H3N2 en cerdos mediante la prueba de inhibición de la
hemoaglutinación (HI), se consideraron positivos los sueros con títulos ≥80, (Rodríguez
Torres, Ramírez Mendoza, Carreón Nápoles, & Mercado García, 1996).
La prevalencia serológica determinada en este estudio para H1N1 (cepa porcina) en
explotaciones tecnificadas es de 20.16% y de 41.04% para no tecnificadas. Existe
diferencia significativa de la prevalencia de Influenza H1N1 y H3N2 en explotación
tecnificada y no tecnificada (H1N1 Gh = 139.31 p<0.5 y H3N2 Gh= 81.87 p<0.5) ver
Tabla 3.
En el presente estudio el subtipo H1N1 se encuentra distribuido en todo el país
tanto en explotación tecnificada como en no tecnificada (Figuras 9 y 10).
Para el subtipo H3N2 la seropositividad establecida fue de 6.59% en explotaciones
tecnificadas y de 16.96 % en no tecnificadas; nuestros resultados son menores a los
reportados por Piñeyro en Argentina 2006 quien reporta 73.8%, pero son similares a los
reportados por Ramírez en Colombia quien reporta un 8.2% de positividad en
explotaciones tecnificadas para este serotipo
Ya que en Guatemala no se utilizan vacunas comerciales contra Influenza Porcina,
y a que el análisis serológico evidenció anticuerpos contra el virus de IP subtipos H1N1 y
H3N2, se puede inferir que la presencia de animales positivos es producto de infecciones
naturales.
El subtipo H3N2 en explotaciones no tecnificadas se encuentra distribuido en todo
el país, mientras que el mismo serotipo en explotación tecnificada se encuentra en el área
central, sur y oriente del país (Figuras 11 y 12).
Las explotaciones tecnificadas de las regiones Norte y Occidente fueron 100%
negativas Para el subtipo H3N2, mientras que en las regiones restantes la prevalencia
encontrada fue: Área Central = 10.72%, Área Sur = 3.41% y Área Oriente = 6.14%,
(Tabla 3). En contraste con nuestros resultados Darío Mogollón en Colombia reporta una
amplia dispersión de reactores para el subtipo H3N2, sin embargo reporta para el subtipo
H1N1 lo encuentra circunscrito en una área determinada.

32
Tabla 3: Número de Muestras y Prevalencia por departamento, por área por tipo de explotación, H1N1 y H3N2, Guatemala, 2010, Proyecto FODECYT 62-2009
Explotación Tecnificada Explotación No Tecnificada
Departamento H1N1 H3N2 H1N1 H3N2
Prevalencia por Departamento (+/n)
Prevalencia por área (+/n)
Prevalencia por Departamento (+/n)
Prevalencia por área (+/n)
Prevalencia por Departamento (+/n)
Prevalencia por área (+/n)
Prevalencia por Departamento (+/n)
Prevalencia por área (+/n)
Central Guatemala 32.78%(80/244) 24.46%(105/429) 14.75%(36/244) 10.72%(46/429) 0%(0/2) 49.41%(84/170) 0%(0/2) 20.59%(35/170)
Sacatepéquez 26.03%(19/73) 4.11%(3/73) 63.44%(59/93) 25.81%(24/93)
Chimaltenango 5.36%(6/112) 6.25%(7/112) 33.33%(25/75) 14.67%(11/75)
Norte Alta Verapaz 0%(0/0) 100%(1/1) 0(0/0) 0%(0/1) 64%(16/25) 51.02%(75/147) 24%(6/25) 27.21%(40/147)
Baja Verapaz 0%(0/0) 0(0/0) 94%(47/50) 64%(32/50)
Petén 100%(1/1) 0(0/1) 16.67%(12/72) 2.78%(2/72)
Occidente Totonicapán 0%(0/0) 16.13%(10/62) 0(0/0) 0%(0/62) 0%(0/0) 32.25%(188/583) 0%(0/0) 11.84%(69/583)
Sololá 0%(0/0) 0(0/0) 0%(0/0) 0%(0/0)
Quiché 0%(0/15) 0(0/15) 30.61%(15/49) 42.86%(21/49)
Quetzaltenango 30%(6/20) 0%(0/20) 31.11%(159/511) 8.22%(42/511)
San Marcos 14.81%(4/27) 0%(0/27) 60.87%(14/23) 26.09(6/23)
Huehuetenango 0%(0/0) 0(0/0) 0%(0/0) 0%(0/0)
Oriente Zacapa 0%(0/19) 21.93%(25/114) 5.26%(1/19) 6.14%(7/114) 0%(0/1) 66.67%(48/72) 0%(0/1) 29.17%(21/72)
Chiquimula 0%(0/0) 0%(0/0) 0%(0/0) 0%(0/0)
El Progreso 0%(0/1) 0%(0/1) 0%(0/0) 0%(0/0)
Izabal 0%(0/0) 0%(0/0) 0%(0/0) 0%(0/0)
Jutiapa 26.60%(25/94) 6.38%(6/94) 67.61%(48/71) 29.58%(21/71)
Jalapa 0%(0/0) 0(0/0) 0%(0/0) 0%(0/0)
Sur Santa Rosa 15.45%(17/110) 15.57%(64/411) 2.72%(3/110) 3.41%(14/411) 0%(0/8) 46.97%(31/66) 0%(0/8) 16.67%(11/66)
Escuintla 17.61%(31/176)
4.54%(8/176) 91.30%(21/23) 30.43%(7/23)
Suchitepéquez 7.14%(6/84) 3.57%(3/84) 37.04%(10/27) 14.81%(4/27)
Retalhuleu 0%(0/41) 0%(0/41) 0%(0/8) 0%(0/8)
Prevalencia 20.16%(205/1017) 6.59%(67/1017) 41.04%(426/1038) 16.96%(176/1038)
TOTAL DE MUESTRAS 1017 1038 2055

33
Figura 9: Prevalencia de H1N1 en cerdos de explotaciones tecnificadas por áreas de la República de
Guatemala, 2010
Fuente: FODECYT, 62-2009
Figura 10: Prevalencia de Influenza H1N1 en cerdos de explotaciones no tecnificadas por áreas en la
República de Guatemala, 2010
Fuente: FODECYT, 62-2009
23%12% 16% 1 22%
77%
88%
84%0
78%
0
100
200
300
400
500
600
Central Sur Occidente Norte Oriente
Nú
me
ro d
e m
ue
stra
s
Áreas
Negativos
Positivos
0
100
200
300
400
500
600
Central Sur Occidente Norte Oriente
47%53%
32%
51% 67%
53%
47%
68%
49%
33%
Nú
me
ro d
e M
ue
stra
s
Área

34
Figura 11: Prevalencia de Influenza H3N2 en cerdos de explotaciones tecnificadas por áreas, en la
República de Guatemala, 2010
Fuente: FODECYT, 62-2009
Figura 12: Prevalencia de Influenza H3N2 en cerdos de explotaciones no tecnificadas por área en la
República de Guatemala, 2010
Fuente: FODECYT, 62-2009
9% 4% 0% 0 6%
91%
96%
100%100%
94%
0
100
200
300
400
500
600
Central Sur Occidente Norte Oriente
Nú
me
ro d
e m
ue
stra
s
Área
Negativos
Positivos
0
100
200
300
400
500
600
Central Sur Occidente Norte Oriente
20% 19%12% 27% 29%
80%
81%
88%
73%
71%
Nú
me
ro d
e m
ue
stra
s
Área

35
Figura 13: Número de muestras que presentaron seropositividad a ambos serotipos de Influenza
(H1N1 y H3N2) por áreas en la República de Guatemala, 2010.
Fuente: FODECYT 62-2009
Existe una evidente diferencia entre la seropositividad obtenida en porcinocultura
tecnificada y no tecnificada para ambos serotipos, sugiriendo que las medidas de
bioseguridad utilizadas en las explotaciones tecnificadas juegan un importante papel para
la ausencia de infección. (H1N1 tecnificada Gh1= 15.16 p < 0.05, no tecnificada Gh= 43.72
p < 0.05, H3N2 tecnificada Gh= 15.03, p < 0.05 no tecnificada Gh= 17.64, p < 0.05).
El 5.7% de las muestras de explotaciones tecnificadas fueron positivos a ambos
serotipos (H1N1 y H3N2); mientras que en explotaciones no tecnificada este porcentaje
fue del 17%, sugiriendo que existe un mayor riesgo de recombinación genética, y de
mutación viral en explotaciones no tecnificadas, al respecto Brown (2000) afirma que los
intercambios genéticos cuando se presentan serotipos simultáneamente, pueden resultar en
la aparición de variantes virales con diferentes características genéticas y antigénicas.
En el año 2000 Richard Webby y col. Al estudiar la evolución de los virus de
influenza H1N1 y H3N2 en los Estados Unidos, y analizar 4,382 sueros sanguíneos
provenientes de 23 estados, con la técnica de inhibición de la hemoaglutinación, reporta
que 28,3% de los animales han sido expuestos al virus clásico H1N1, mientras que el
20.5% han sido expuestos a la triple recombinación H3N2.
Existe diferencia significativa entre la seropositividad a H1N1 entre sexo en
explotación tecnificada y no tecnificada (Sexo Gh= 4.44, p<0.5) Figuras No. 14 y 15, las
hembras tienen una mayor prevalencia. Sin embargo no existe diferencia significativa para
el serotipo H3N2 (Gh= 0.18, p<0.5) Figuras No. 16 y 17.
1 (Sokal & Rohlf, 1995)
0
10
20
30
40
50
60
70
80
Centro Norte Occidente Oriente Sur
Nú
me
ro d
e M
ue
stra
s
Áreas

36
Figura 14: Prevalencia de Influenza H1N1 en cerdos machos de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
Figura 15: Prevalencia de Influenza H1N1 en cerdas hembras de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
73%
27%
Negativos
Positivos
69%
31%
Negativos
Positivos

37
Figura 16: Prevalencia de Influenza H3N2 en cerdos machos de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
Figura 17: Prevalencia de Influenza H3N2 en cerdas hembras en la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
89%
11%
Negativos
Positivos
88%
12%
Negativos
Positivos

38
Existe diferencia significativa entre la seropositividad de H1N1 entre edades en
explotación tecnificada y no tecnificada (Gh= 10.37, p<05) ver Figuras 18 y 19, no existe
diferencia significativa para el serotipo H3N2 (Gh= 0.28, p<0.5 Figura No. 20 y 21.
Figura 18: Prevalencia de Influenza H1N1 en cerdos jóvenes de la República de Guatemala. 2010
Fuente: FODECYT, 62-2009
Figura 19: Prevalencia de Influenza H1N1 en cerdos adultos de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
73%
27%
Negativos
Positivos
67%
33%
Negativos
Positivos

39
Figura 20: Prevalencia de Influenza H3N2 en cerdos jóvenes de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
Figura 21: Prevalencia de Influenza H3N2 en cerdos adultos de la República de Guatemala, 2010
Fuente: FODECYT, 62-2009
88%
12%
Negativos
Positivos
89%
11%
Negativos
Positivos

40
PARTE IV
IV.1 CONCLUSIONES
1. Se monitorearon y evaluaron 2,055 sueros de cerdos procedentes de las
diferentes áreas geográficas de la República de Guatemala utilizando la
prueba de Inhibición de la Hemoaglutinación.
2. A nivel nacional se determinó la presencia del virus influenza H1N1
encontrándose una prevalencia de 24.46% en el área central, un 15.57% en el
área sur, 21.93% en oriente y 16.13% en occidente para explotaciones
tecnificadas y en explotaciones no tecnificadas se encontró una prevalencia de
46.97% en el área sur, 49.41% en área central, 51.02% en el área norte
32.25% en el occidente y 66.67% en oriente, por lo que se encuentra
distribuido en todo el país.
3. Se determinó la presencia del virus influenza H3N2, encontrándose una
prevalencia de 10.72% en el área central, 6.14% en oriente y un 3.41% en el
área sur en explotaciones tecnificadas y 20.59% en el área centro, 27.21 en el
área norte, 11.84% en el occidente, 29.17% en el oriente y 16.67% en el área
sur de explotaciones no tecnificadas. No existe diferencia significativa en
cuanto a edad (Gh= 0.28, p > 0.5) ni sexo (Gh= 0.18, p > 0.5).
4. Se demostró la presencia de Anticuerpos contra el virus de la Influenza
Porcina H1N1 y H3N2 en cerdos en diferentes áreas de la República de
Guatemala, así mismo se determinó la distribución de ambos subtipos (H1N1
y H3N2).
5. El 14.64% de las muestras de explotaciones no tecnificadas fueron positivas a
ambos serotipos (H1N1 y H3N2); La elevada prevalencia de ambos serotipos
en porcinocultura no tecnificada, sugiere un mayor riesgo de infección,
propagación, recombinación y/o mutación viral en este sector productivo,
principalmente por las técnicas de producción utilizadas en las que los cerdos
viven (se encuentran dentro de las viviendas), comen y comparten espacio con
humanos, elevando el riesgo zoonótico, al respecto Brown (2000) afirma que
los intercambios genéticos cuando se presentan serotipos simultáneamente y
en diversas especies, pueden resultar en la aparición de variantes virales con
diferentes características genéticas y antigénicas.
6. Se comprobó en este estudio que los cerdos poseen anticuerpos contra
Influenza subtipos H1N1 y H3N2 pudiendo actuar como reservorio para
ambos.
7. Las explotaciones tecnificadas en las áreas Norte y Occidente fueron 100%
negativas para el subtipo H3N2.

41
8. Los resultados de la prueba de inhibición de la hemoaglutinación son
altamente confiable, evidenciándose a los resultados obtenidos en los sueros
controles positivos de referencia H1N1 A/SW/Iowa/73 y H3N2
A/SW/TX/1/98 procedentes del Laboratorio Nacional de Servicios
Veterinarios del Departamento de Agricultura de los Estados Unidos
(NVSL/USDA).
9. Al evaluar la técnica de inhibición de la hemoaglutinación utilizando ambos
subtipos se pudo comprobar que el proceso de la inactivación y remoción de
hemaglutininas inespecíficas es largo y complejo, requiriendo de tiempo y
personal técnico capacitado.
10. Para realizar la prueba de HI para esta enfermedad se requiere de equipo
complejo presente únicamente en laboratorios especializados, ya que para la
inactivación y el tratamiento de los sueros se emplea Incubadora a 37°C, baño
María y campanas de flujo laminar para manipular sueros y enzima
destructora de receptores (RDE).
11. Se observó mayor prevalencia de subtipo H1N1 en cerdos hembras (31%
hembras y 27% machos) en la república de Guatemala.
12. Existe una mayor prevalencia del subtipo H1N1 en cerdos adultos (33%
adultos 27% jóvenes, Gh= 10.37, p<05) en la República de Guatemala.
13. Los resultados obtenidos al realizar la prueba de Inhibición de la
Hemoaglutinación serán implementados dentro del laboratorio para ofrecer
esta técnica como prueba diagnóstica rutinaria.
14. Para lograr difundir los resultados se publicará un artículo con los resultados
de la investigación en “La Revista” de la Facultad de Medicina Veterinaria y
Zootecnia, así como en otras revistas científicas.
15. Los investigadores informarán sobre los resultados obtenidos en esta
investigación por medio de participación en diferentes conferencias.
16. Los resultados del análisis serán transferidos al Ministerio de Agricultura y
Ganadería (MAGA) y al Ministerio de Salud y Asistencia Social (MSPAS),
presentándoles el proyecto para que se realicen monitoreos en granjas de
cerdos así como del personal de la granja, para explotaciones tecnificadas y a
los cerdos de explotaciones no tecnificadas, así como a los integrantes de la
comunidad.

42
IV.2 RECOMENDACIONES
1. Debido a que los virus influenza H1 circulan en menor proporción que los H3 y
que en este estudio se ha demostrado la presencia de circulación viral de ambos
serotipos, este estudio sugiere proporcionar mayor atención a este último
serotipo.
2. Realizar el aislamiento de los virus H1N1 y H3N2 utilizando cultivo celular
con células Madin-Darby de riñón de perro (MDCK); realizar pruebas
moleculares (PCR) para analizar la ocurrencia de variaciones antigénicas, así
como la secuenciación aminoacídica del material genético.
3. Implementar a nivel nacional monitoreos periódicos de Influenza pandémica
H1N1/2009 en explotaciones porcinas.
4. Promover la adaptación de esquemas de vigilancia de las enfermedades para
diferentes sistemas de producción, realizando monitoreos serológicos para el
análisis con la prueba de inhibición de la hemoaglutinación para serotipo
H1N1.
5. Los resultados obtenidos de positividad en cerdos, servirá para la toma de
decisiones de las autoridades de Salud pública, en este caso los humanos que
conviven con cerdos pueden servir de centinelas para la aparición de posibles
brotes.
6. Debido a que las pruebas serológicas no diferencian entre las cepas de H1N1
(porcina) y la pandémica H1N1/2009, para vigilancia epidemiológica de virus
pandémico en porcinos se recomienda emplear pruebas virológicas y
moleculares.
7. Organizar una red de laboratorios de diagnóstico (veterinarios y humanos) para
el monitoreo de virus respiratorios y otras enfermedades infecciosas, con
metodología estandarizada; el aislamiento de los virus es crucial para
determinar su tipo, subtipo y su siguiente clasificación.

43
IV.3 REFERENCIAS BIBLIOGRÁFICAS
1. Acuña L, G. (2004). Influenza: Historia y amenzas. Rev Chil Infect, 21(2),
162-164.
2. Ayora-Talavera, G., Cadavico-Burgos, J. M., & Canul-Armas, A. B.
(Enero de 2005). Serologic Evidence of Human and Swine
Influenza in Mayan Persons. CDC, 11(1), 158-160.
3. Barrera, M., & Díaz, S. (Septiembre de 24 de 2010). Centro Nacional de
Epidemiiología. Recuperado el 21 de Enero de 2011, de
Laboratorio Nacional de Salud:
http://epidemiologia.mspas.gob.gt/vigepi/2010/laboratorio/Virus%2
0Respiratorios%202009.pdf
4. Beveridge, W. I. (1977). Influenza: The last great plague. New York:
Heinemann Educational Publishers.
5. Bronze, M. S. (19 de Octubre de 2010). eMedicine. Recuperado el 20 de
Enero de 2011, de http://emedicine.medscape.com/article/1807048-
overview
6. Cannon. (2001). Sense and sensitivity –designing surveys based on an
imperfect test. 49: 141-163. Prev. Vet. Med,., 49, 141-163.
7. Departamento de Salud y Servicios Humanos. (6 de mayo de 2009).
PandemicFlu.gov. Recuperado el 20 de septiembre de 2009, de
Pandemias y amenazas de pandemia desde 1900:
http://espanol.pandemicflu.gov/pandemicflu/enes/24/_pandemicflu
_gov/general/historicaloverview.htm
8. Easterday, B. C., & Hinshaw, V. S. (1992). Diseases of swine. Ames:
Iowa State Press.
9. FAO. (2009). Guía de la FAO para la vigilancia del virus pandémico
H1N1/2009 y otros virus de influenza en poblaciones porcinas.

44
10. Ferrari, G., Pinto, J., Honhold, N., Zingeser, J., Dauphin, G., Kim, M., y
otros. (2009). Guía de la FAO para la vigilancia del virus
pandémico H1N1/2009 y otros virus de la influenza en poblaciones
porcinas. FAO. Departamento de Agricultura y Protección al
Consumidor.
11. Garcia, J. (1999). La Influenza Aviar en México. Simposio Internacional
sobre Influenza Aviar, (págs. 1-19). Maracay.
12. Garten RJ, D. C. (2009;). Antigenic and genetic characteristics of swine
origin 2009 A (H1N1) influenza circulating in humans. Science,
197-201.
13. Gaydos, J. C., Top, F. H., Hodder, R. A., & Russell, P. K. (Enero de
2006). Swine Influenza A Outbreake, Fort Dix, New Jersey, 1976.
Emergin Infectious Diseases, 12(1), 23-25.
14. Hanssen, H. V., Hincapié, O. N., & López T, J. H. (1 de Abril de 2003).
Recuperado el 5 de Diciembre de 2010, de Influenza en porcinos de
Antioquía, Colombia:
http://hist.library.paho.org/spanish/Bol/v82n1p35.pdf
15. Heinen, P. (15 de septiembre de 2003). Veterinary Science Tomorrow.
Recuperado el 20 de octubre de 2009, de Swine Influenza: a
zoonosis: http://www.vetscite.org/
16. HHS. (2009). U.S. Departament of Health & Human Services. Recuperado
el 21 de enero de 2011, de http://www.hhs.gov/pandemicflu/plan/
17. Janke, B. H. (Enero de 13 de 2000). American Association of Swine
Veterinarians . Recuperado el 12 de Enero de 2011, de
http://www.aasv.org/shap/issues/v8n2/v8n2p79.html
18. Krause, R. (2006). The Swine Flu Episode and the Fog of Epidemics.
Emergin Infectious Diseases, 12(1), 40-43.
19. Lamb, R. A., & King, R. M. (2001). Orthomixoviridae: The viruses and
their replication. En D. M. Knipe, P. M. Howley, D. E. Griffin, M.
A. Lamb, M. A. Martin, B. Roizman, y otros, Filds Virology (4th
ed., Vol. 2, págs. 1487-1532). Philadelphia, Pensilvania, Estados
Unidos: Lippincot-Raven.
20. Mandell, G. L., Bennett, J. E., & Dolin, R. (2006). Enfermedades
Infeeciosas. España: Elsevier.

45
21. Mastin, A., Alarcon, P., Pfeiffer, D., Wood, J., Williamson, S., Brown, I.,
y otros. (Febrero de 2011). Prevalence and risk factors for swine
influenza virus infection in the English pig population. PloS Curr.
22. Mastrosovich, M., Tuzikov, A., Bovin, N., Gambarayan, A., Klimov, A.,
Castrucci, M. R., y otros. (Septiembre de 2000). Early alterations of
the receptor-binding properties of H1, H2, and H3 avian influenza
virus hemagglutinins after their introduction into mammals. J Virol.,
74(18), 8502-12.
23. Mogollón, J. D., Rincón, M. A., Arbeláez, G., Sabogal, R., Ruíz, s., Orjuela, N., y
otros. (Septiembre de 18 de 2009). Mundo Veterinario. Recuperado el 20
de Septiembre de 2009, de Reactividad serologica frente al virus de la
influenza porcina en explotaciones intensivas de Colombia:
http://www.mundoveterinario.net/nueva/referencias/porcinos/influenza.htm
24. OIE. (16 de Abril de 2008). Organización Mundial de Sanidad Animal.
Recuperado el 23 de Enero de 2009, de Manual de las Pruebas de
Diagnóstico y de las Vacunas para los Animales Terrestres:
http://www.oie.int/esp/normes/mmanual/e_summry.htm
25. Olsen, C. W., Brammer, L., Easterday, B. C., Arden, N., Belay, E., & Cox, N. J.
(Agosto de 2002). Serologic Evidence of H1 Swine Influenza Virus
Infection in Swine Farm Residents and Employees. Emerg Infect Dis
[serial online] , 8(8), http://www.cdc.gov/ncidod/EID/vol8no8/01-
0474.htm.
26. Organización Mundial de la Salud. (4 de Mayo de 2009). OIE. Recuperado el 7
de Mayo de 2009, de http://www.oie.int
27. Panamerican Health Organization. (24 de abril de 2009). Panamerican Health
Organization. Recuperado el 22 de 10 de 2009, de Influenza-Like Illness
in Mexico and the United States :
http://new.paho.org/hq/index.php?option=com_content&task=view&id=1
259&Itemid=1
28. Piñeyro, P. E., Cappuccio, J. A., Machuca, M. A., Teodoroff, T. A., Baumeister,
E., & Perfumo, C. J. (2006). Prevalencia serológica del virus de Influenza
en Argentina
29. Radio France International. (27 de 4 de 2009). Radio France International.
Recuperado el 22 de 10 de 2009, de OMS: "Enfermedad con potencial
pandémico": http://www.rfi.fr/actues/articles/112/article_11682.asp

46
30. Ramírez, O. J., Boulanger, A., & Moscardi, A. (julio de 2005). Evidencia
Serológica de Infección por el virus de Influenza Porcina en Granjas de
Cerdos en Venezuela. Revista de la Facultad de Ciencias Veterinarias,
46(2).
31. Rodríguez Torres, J., Ramírez Mendoza, H., Carreón Nápoles, R., & Mercado
García, C. (1996). Muestreo serológico a nivel de rastro para detectarr
anticuerpos contra el virus de influenza porcina. Vet. Mex, 27(1), 17-21.
32. Scholtissek, C. (1990). Pigs as the “mixing vessel” for the creation of new
pandemic influenza A viruses. Med Principles Pract, 2, 65-71.
33. Scholtissek, C. (1994). Source for Influenza Pandemics. European Journal of
Epidemiology, 10, 455-458.
34. Serrano, L., Santizo, B., & Motta, L. (2002). Primer Aislamiento de Virus de
Influenza Aviar en Guatemala. La Revista Facultad de Medicina
Veterinaria y Zootecnia, 16(2), 18-20.
35. Sokal, R. R., & Rohlf, F. J. (1995). Biometry (3a ed.). Estados Unidos: W.H.
Freeman and Company.
36. Spickler, A. R. (29 de Diciembre de 2009). The Center for food security and
public health. Recuperado el 20 de Enero de 2011, de
http://www.cfsph.iastate.edu/Factsheets/pdfs/influenza.pdf
37. Swayne, D. E. (2008). Avian Influenza. Oxford, UK.: Blacwell Publishing.
38. Vetsweb. (4 de septiembre de 2009). Singapore detects H1N1 in pigs. Recuperado
el 22 de octubre de 2009, de Vetsweb:
http://www.vetsweb.com/news/singapore-detects-h1n1-in-pigs-413.html
39. World Health Organization. (2005). WHO Manual on Animal Influenza
Diagnosis and Surveillance. Switzerland: WHO.

47
IV.4 ANEXO 1: Protocolo de Recepción y Análisis de Muestras
Código: SENACYT 062-2009/______
CUESTIONARIO
EVALUACION Y MONITOREO DE INFLUENZA
El objetivo del presente cuestionario es identificar las muestras que serán analizadas para el monitoreo nacional, marque con una X la casilla correspondiente:
1. Fecha de recolección de muestra: _____________________________
2. Procedencia de la muestra:
Tecnificada No Tecnificada
3. Sexo del animal
Macho Hembra
4. Rango de edad visible:
Joven Adulto
5. Localización de la explotación, favor especifique departamento: _______ __________________________________________________________ -------------------------------------------------------------------------------------------------
PROCESAMIENTO DE LA MUESTRA
a. Fecha de Recepción: _______________________________
b. No. de Identificación de la muestra: ____________________
c. Temperatura de almacenamiento: _____________________
d. Fecha de tindalización: ______________________________
e. Fecha de tratamiento: _______________________________

48
IV.5 ANEXO 2: Protocolo de resultado del análisis de las muestras

49
IV.6 ANEXO 3: Registro fotográfico del proyecto FODECYT 62-2009
Fotografía 1: Preparación de Material Para la
toma de muestras
Fotografía 2: Entrada al Rastro de Municipal de
Santa Catarina Pinula
Fotografía 3: Toma de muestras en el Rastro
Municipal de Santa Catarina Pínula
Fotografía 4: Presentación ante estudiantes de EPS.
Fotografía 5: Presentación de teoría a los
estudiantes de EPS para la toma de muestras
en cerdos de explotaciones no tecnificadas
Fotografía 6: Estudiantes de EPS

50
Fotografía 7: Clase práctica para los
estudiantes de EPS
Fotografía 8: Toma de muestra en las vena ocular
Fotografía 9: Sujeción de cerdos grandes
Fotografía 10: Toma de muestra de la vena
Auricular
Fotografía 11: Sujeción de lechones
Fotografía 12: Indicación del lugar para realizar el
sangrado a nivel de vena yugular

51
Fotografía 13: Toma de muestra de la vena
yugular
Fotografía 14: Preparación del área de trabajo para
el procesamiento de los sueros. Se realizó en una
campana de flujo Laminar.
Fotografía 15: Trasvase de las muestras previo
a su tindalización.
Fotografía 16: Trasvase de las muestras a viales de
1.5 ml.
Fotografía 17: Trasvase de las muestras
Fotografía 18: Identificación de las muestras.

52
Fotografía 19: Desinfección del material
utilizado con Rayos Ultravioleta durante 8
horas antes de descartar el material
Fotografía 20: Desinfección del área de trabajo
Fotografía 21: Lavado del Material previo a la
esterilización
Fotografía 22: Colocación de los viales con los sueros
para su procesamiento
Fotografía 23: Tindalización de los sueros en
baño maría, a 56 ° C por 30 minutos
Fotografía 24: Tindalización de los sueros en baño
maría, a 56 ° C por 30 minutos

53
Fotografía 25: Almacenamiento de las muestras
a -20°C antes del tratamiento.
Fotografía 26: Uso del agitador magnético para la
preparación de buffers
Fotografía 27: Tratamiento de sueros con las
enzima RDE
Fotografía 28: Enzima RDE
Fotografía 29: Tratamiento de los sueros con
glóbulos rojos al 50%
Fotografía 30: Agitador de placas utilizado para el
tratamiento de los sueros con glóbulos rojos de pavo
al 50%.

54
Fotografía 31: Agitación de las muestras
tratadas con la Enzima Destructora de
Receptores.
Fotografía 32: Inactivación de la Enzima
Destructora de Receptores, después del tratamiento
Fotografía 33: Antígenos
Fotografía 34: Material a Utilizar
Fotografía 35: Micropipetas digitales utilizadas
para la prueba de Inhibición de la
Hemoaglutinación
Fotografía 36: Titulación de los Antígenos

55
Fotografía 37:Adición de glóbulos rojos de
pavo al 0.5%
Fotografía 38: Agitación de las placas después de la
adición de glóbulos rojos al 0.5%
Fotografía 39: Lectura de resultados de la
titulación del antígeno
Fotografía 40: Realización de la prueba de
Inhibición de la Hemoaglutinación.
Fotografía 41:Lectura de Resultados
Fotografía 42: Lectura de Resultados

56
IV.7 ANEXO 4: TÍTULOS DE ANTICUERPOS
Tabla 4: Títulos de Anticuerpos contra Influenza H1N1 de cerdos de explotaciones tecnificadas por
áreas de la República de Guatemala, 2010, Proyecto FODECYT 62-2009
ÁREA TÍTULOS DE ANTICUERPOS
0 20 40 80 160 320 640 1280 2560
Central 411 3 3 17 21 32 30 14 8
Sur 259 0 5 4 8 5 5 7 8
Occidente 52 0 0 1 4 2 3 0 0
Norte 0 0 0 0 0 1 0 0 0
Oriente 88 0 1 2 8 9 4 2 0 Fuente: FODECYT 62-2009
Tabla 5: Títulos de anticuerpos contra influenza H1N1 de cerdos de explotaciones no tecnificadas por
áreas de la República de Guatemala, 2010, Proyecto FODECYT 62-2009
ÁREA TÍTULOS DE ANTICUERPOS
0 20 40 80 160 320 640 1280 2560
Central 79 3 12 19 37 11 9 5 3
Sur 27 0 0 5 7 3 10 5 1
Occidente 385 6 4 43 46 56 29 12 2
Norte 64 2 6 26 22 15 4 3 5
Oriente 23 0 1 5 11 12 9 9 2
Fuente: FODECYT 62-2009
Tabla 6: Títulos de anticuerpos contar influenza H3N2 de cerdos de explotaciones tecnificadas de la
República de Guatemala, 2010, Proyecto FODECYT 62-2009
ÁREA TÍTULOS DE ANTICUERPOS
0 20 40 80 160 320 640 1280 2560
central 487 0 3 5 0 4 11 25 4
sur 289 0 1 1 0 1 6 3 0
occidente 62 0 0 0 0 0 0 0 0
norte 1 0 0 0 0 0 0 0 0
oriente 107 0 0 0 0 3 3 1 0
Fuente: FODECYT 62-2009

57
Tabla 7: Títulos de anticuerpos contra Influenza H3N2 de cerdos de explotaciones no tecnificadas en
la República de Guatemala, 2010, Proyecto FODECYT 62-2009
ÁREA TÍTULOS DE ANTICUERPOS
0 20 40 80 160 320 640 1280 2560
Central 138 1 4 5 12 5 8 4 1
Sur 47 0 0 1 1 6 1 1 1
Occidente 507 4 3 7 21 12 16 11 2
Norte 103 0 4 5 5 13 9 8 0
Oriente 50 1 0 3 2 3 4 3 6 Fuente: FODECYT 62-2009

58
IV.8 ANEXO 5: CURRÍCULUM DE INVESTIGADORES
Lucero Serrano Arriaza de Gaitán
Sexo: Femenino
Nacionalidad: guatemalteca
Estado civil: Casada
Profesión u oficio: Médico Veterinario
Puesto actual: Coordinadora del Laboratorio de referencia Regional de Sanidad avícola.
IDIOMAS:
Inglés: Escribe y lee
Alemán: Escribe lee y habla.
ESTUDIOS REALIZADOS:
UNIVERSITARIOS:
Universidad de San Carlos de Guatemala, Facultad de Medicina Veterinaria y Zootecnia, 1978-
1983, Obteniendo el grado de Licenciatura.
POST GRADO:
Maestría en Gerencia Educativa de la Universidad Panamericana 2007
Tierarztliche Hochshule Hannover. Alemania 1989 - 199; Facultad de Veterinaria de la
Universidad de Hannover, Diagnóstico de Enfermedades aviares
Munchen Universitaat. Universidad de Múnich, Alemania; Clínica de enfermedades Aviares;
Mayo a julio de 1998
Especialidad en Biología Molecular; Universidad de San Carlos de Guatemala
Facultad de Medicina; Septiembre 2,006 Octubre 2,007
Especialización en Formulación y Gestión de proyectos de Cooperación, Internacional. Agencia
Catalana de Cooperación. Fundación Universitaria; Iberoamericana. USAC. 26 de Febrero 2,007 al
25 de enero 2,008.
EXPERIENCIA LABORAL:
- Docente del Departamento de Avicultura de la Facultad de Medicina Veterinaria y Zootecnia,
cátedra obtenida por oposición curricular junio 1987
- Coordinador del Depto. de Ornitopatología y Avicultura.
- Coordinador del Área de Producción avícola de la Granja Experimental de la Facultad de
Medicina Veterinaria y Zootécnica.

59
- Directora de la Escuela de Medicina Veterinaria 2,005-2,007
- Consultor Programa regional de enfermedades aviares PREA/OIRSA 2,007
- Consultor de empresas Productoras avícolas, a la fecha.
- Directora de la Escuela de Medicina Veterinaria de la Facultad de Medicina Veterinaria y
Zootecnia (USAC) 2,005 -2,007.
- Coordinadora del Curso de Enfermedades Infecciosas. Facultad de Medicina Veterinaria y
Zootecnia. 2,006
- Coordinadora del Laboratorio de Referencia Regional de Sanidad Avícola LARRSA
- Asesor de Empresas Avícolas.
INVESTIGACIONES REALIZADAS Y PUBLICACIONES:
- Primer aislamiento de Influenza aviar de baja patogenicidad en Guatemala. Vol. 16 Guatemala,
mayo 2,002 Revista de la Facultad de Medicina Veterinaria y Zootecnia.
- Determinación de variantes antigénicas del virus de Influenza H5N2 en Guatemala. Congreso
Latinoamericano de avicultura. Cuba 2,003
- Evaluación de una vacuna recombinante de Influenza aviar Congreso de Médicos veterinarios
Guatemala 2,007
- Hipopteronosis quística estudio de caso. Guatemala 2,003
- Primer aislamiento de Ornithobacterium rinotraqueale.

60
YERY EDGARDO VELIZ PORRAS
NOMBRE: YERI VELIZ PORRAS
ESTADO CIVIL: CASADO
RELIGIÓN: CATÓLICO
RESIDENCIA: 0 CALLE “C” 3-16 ZONA 4 COLONIA EL VESUBIO, ESCUINTLA
TELÉFONO CASA: (502) 78898480
TELÉFONO CELULAR: (502) 55789663
CORREO ELECTRÓNICO: [email protected]
IDIOMAS: Español, Inglés, Alemán
PROFESIÓN: Médico Veterinario
Master en Administración Industrial y Empresas de Servicio
TRABAJO ACTUAL
Coordinador del Instituto de Reproducción Animal e Inseminación Artificial. Facultad de
Medicina Veterinaria y Zootecnia. Universidad de San Carlos de Guatemala. Zona 12
Ciudad.
Catedrático Titular IX de los cursos de Reproducción Animal e Inseminación Artificial.
Escuela de Veterinaria y Zootecnia.
Catedrático Titular IX de los cursos de Reproducción Animal e Inseminación Artificial,
Obstetricia y Patología de la Reproducción animal. Escuela de Veterinaria.
Catedrático titular IX de los cursos de Clínicas I y II de Porcinos. Escuela de Veterinaria
Consultor Topigs Dalland Porcina para Guatemala, El Salvador y Honduras.
Consultor de Granjas Porcinas en Guatemala, El Salvador y Honduras.
ESTUDIOS REALIZADOS.
Maestro de Educación Primaria urbana. Escuintla. 1,978.
Médico Veterinario. Facultad de Medicina Veterinaria y Zootecnia. USAC. 1,987
Especialidad en Clínica, Medicina Forense y Reproducción de cerdos, ovejas y cabras.
Hannover. Alemania. 1,991
Curso sobre Formulación y Evaluación de Proyectos. INAP. 1,997
Curso sobre Especialización y Actualización en Sanidad de Cerdos. Fac. Veterinaria;
APOGUA. GRETECEG. 1.,997
Curso Evaluación del Rendimiento Académico. Instituto de Investigaciones y Mejoramiento
Educativo. USAC. 1,997.

61
INVESTIGACIONES REALIZADAS
Evaluación del Destete parcial en cerdas Multíparas y su efecto en el desempeño
reproductivo, en una Granja Tecnificada, en el Municipio de Sumpango, Sacatepéquez.
Facultad de Medicina Veterinaria y Zootecnia. Agosto 2003
Evaluación del uso de Leche descremada Fluida UHT como Extensor de Semen Porcino,
Sobre la Fertilidad y número de Nacidos Totales en Cerdas Inseminadas. Facultad de
Medicina Veterinaria y Zootecnia. Mayo 2004
Evaluación del Propofol como anestésico general en Cerdos. Facultad de Medicina
Veterinaria y Zootecnia. Septiembre 2005
Efecto de la Utilización de Plasma seminal Sintético como presensibilizador, previo a la
inseminación Artificial en cerdas Multíparas. Facultad de Medicina Veterinaria y Zootecnia.
Septiembre 2005.
Utilización de Melaza en cerdas y su efecto sobre el aparecimiento del Celo post destete,
Porcentaje de Fertilidad y Nacidos Totales. Facultad de Medicina Veterinaria y Zootecnia.
USAC. Mayo 2006
Comparación de dos tipos de hierro para aplicación en lechones Recién Nacidos. Facultad de
Medicina Veterinaria y Zootecnia. USAC. Octubre 2006
Uso de Leche descremada en Polvo a diferentes concentraciones como Extensor de semen
Porcino. Facultad de Medicina Veterinaria y Zootecnia. USAC. Septiembre 2006.
Evaluación del Desempeño productivo de dos líneas genéticas Porcinas en una granja
Tecnificada a partir del destete hasta el peso al mercado. Facultad de Medicina Veterinaria y
Zootecnia. USAC. Noviembre, 2007
Evaluación Bioeconomica de dos tipos de comederos (seco/húmedo y Lineal) en cerdos de
la tercera a la onceava semana de edada post destete en una granja Tecnificada. Facultad de
Medicina Veterinaria y Zootecnia. USAC. Noviembre, 2007.
Determinación de la Presencia de Anticuerpos contra Leptospira interrogans en Cerdas de
Cinco granjas Tecnificadas de Guatemala, utilizando la prueba de Microaglutinación
(MAT). Facultad de Medicina Veterinaria y Zootecnia. Febrero 2008.
Comparación entre el uso de una dosis seminal en Inseminación Artificial de Cerdas Vrs. La
utilización de 3 dosis seminales. Facultad de Medicina Veterinaria y Zootecnia. USAC.
Octubre 2008
Estudio retrospectivo de las Causas más comunes de decomiso en Pulmones de cerdos en
Rastro CECARSA en el período 2006-2007. Facultad de Medicina Veterinaria y Zootecnia.
USAC. Octubre 2009
Efectividad de la prueba de Elisa de Captura para la Detección de Anticuerpos contra el
virus de la Peste Porcina Clásica en el Pecari de Collar (Tayassu tajaca). Facultad de
Medicina Veterinaria y Zootecnia. USAC. Octubre 2009
PUBLICACIONES
Coautor de Manual de Planes Profilácticos, Manejo y Sanidad de los Animales Domésticos.
Facultad Veterinaria. USAC. 1,994
Manejo Reproductivo del Verraco. Revista Facultad de Medicina veterinaria y Zootecnia.
Vol.13 No.2 1,996
Manual de Inseminación Artificial en Bovinos. 1,997
Sitios de Aplicación de Medicamentos en Porcinos. Revista Asociación Nacional de
Porcicultores. APOGUA. Febrero 2,000
Como ser un trabajador productivo en granjas porcinas. Revista Asociación Nacional de
Porcicultores. APOGUA. Mayo 2,000
Manual de Inseminación Artificial en Porcinos. 2,001

62
Necrosis de la orejas en Porcinos. Revista asociación Nacional de porcicultores. APOGUA.
Abril 2,002.
Erisipela Porcina, ataca de nuevo. Revista Asociación Nacional de Porcicultores. APOGUA.
Abril 2,003.
Manual de Inseminación Artificial en Porcinos. Vademecun Porcícola. Edifarm . 2003
Patología del Aparato Reproductor Masculino. Facultad de Medicina Veterinaria y
Zootecnia. USAC. Julio, 2003
Preparemos nuestras cerdas para la época de calor. Revista Asociación Nacional de
Porcicultores. APOGUA. Diciembre 2003.
Manual de Planes Profilácticos, manejo y sanidad de los Animales Domésticos y Silvestres.
Cuarta edición. Facultad de Medicina Veterinaria y Zootecnia. 2004
Rinitis Atrófica Porcina. ¿Reapareció?. Revista Asociación Nacional de Porcicultores.
APOGUA. Marzo, 2006

63
MAYRA LISSETTE MOTTA PADILLA DE HODGSON
KM. 16.5 Carretera a El Salvador
Residencial Los Diamantes, Sector "C", Lote 27
Santa Catarina Pinula, Guatemala
Teléfono: (502) 6637-4551
Fax: (502) 6637-4550
Móvil: (502) 53167799
INFORMACIÓN PERSONAL
Nacionalidad: Guatemalteca
Lugar del nacimiento: Guatemala
Fecha del nacimiento: 17 de enero de 1971
Edad: 40 años
Estado Civil: Casada
Cédula de Vecindad: Registro A-1, No. 823391
EMPLEOS
ASESOR PROFESIONAL: AÑOS DE TRABAJO (OCTUBRE – A LA FECHA)
Universidad de San Carlos, Laboratorio de Referencia Regional de Sanidad Avícola
Preparar la Planificación Estratégica, estudio e investigación de las normas ISO/IEC 17025:2005
para laboratorio, las normas ISO 9000 para la producción de biológicos, y las normas ISO
referentes a la protección del medio ambiente.
Investigación en el área de ciencias avícolas. Producción de antígenos de Influenza.
PROFESOR INTERINO: AÑOS DE TRABAJO (2002 – 2005)
Universidad de San Carlos, Departamento Ornitopatología y Avicultura. Guatemala, Ciudad
Impartir clases a alumnos del último año de zootecnia y veterinaria, investigación, práctica de la
ciencia Avícolas, pruebas serológicas y diagnóstico. Trabajo conjunto con el ministerio de la
agricultura de la erradicación de la Influenza Aviar en Guatemala.
AUXILIATURAS (2001 - 2002)
Universidad de San Carlos, Departamento Ornitopatología y Avicultura. Guatemala, Ciudad
Auxiliar de Cátedra de la clase de ciencias avícolas, preparación de clases y laboratorios. Trabajo
en diagnóstico y serología.

64
EDUCACIÓN
MAESTRÍA EN PRODUCTIVIDAD DE CIENCIAS AGRÍCOLAS (EX: 2004 - 2005)
Universidad Galileo, Ciudad
POSTGRADO EN ADMINISTRACIÓN DE EMPRESAS AGRÍCOLAS (EX: 2005)
Universidad Galileo, Ciudad
LICENCIATURA EN MEDICINA VETERINARIA (EX: 1990 - 2000)
Universidad de San Carlos de Guatemala, Ciudad
PUBLICACIONES
1. Quiste folicular en un loro (informe de caso), noviembre de 2002
2. Comparación de la prueba de la inhibición del hemoaglutinación (HI) y la prueba de
inmunoensayo de enzima asociada (ELISA) para la detección de los anticuerpos circulantes
contra le enfermedad de Newcastle. Guatemala, Octubre de 2002
3. Guatemala y su tecnología, XX Congreso Mundial de Avicultura, Nueva Delhi, India 1996
PROYECTOS
1. Monitoreo nacional de Influenza tipo H1N1, utilizando la prueba de inhibición de la
hemoaglutinación para detección de cerdos seropositivos en la república de Guatemala,
presentado a CONCYT y DIGI.
2. Evaluación de una vacuna recombinante de Influenza aviar, en pollitos desafiados con virus
H5N2 de baja patogenicidad, Revista FMVZ, 2011
3. Correlación entre las pruebas de ELISA, inmunodifusión e histopatología para el diagnóstico
de Laringotraqueítis infecciosa, presentada ante el MAGA.
HABILIDADES DEL LABORATORIO
Cultivo Celular
Prueba de ELISA indirecta.
Prueba de Inhibición de la Hemoaglutinación.
Prueba de Inmunodifusión en Agar Gel (AGP)
Aislamiento de Salmonella
Aislamiento Viral
Producción de antígeno de Influenza Aviar y Newcastle para la prueba de Inhibición de la
Hemoaglutinación -HI-
Producción de antígeno de Influenza Aviar para la prueba de Aglutinación en agar gel -
AGP-
Necropsias para diagnóstico en aves
Preparación de Medios de cultivo
Aislamiento Bacteriano

65
IV.9 INFORME FINANCIERO
FICHA DE EJECUCIÓN PRESUPUESTARIA
LÍNEA: FODECYT
Nombre del Proyecto: "Evaluación y monitoreo Nacional de Influenza, utilizando la prueba de inhibición de la
hemoaglutinación (HI), para detección de cerdos seropositivos en la República de
Guatemala"
Numero del Proyecto: 062-2009
Investigador Principal y/o Responsable del Proyecto:
DRA. LUCERO SERRANO ARRIAZA DE GAITÁN
Monto Autorizado: Q229,240.00
Orden de Inicio (y/o Fecha primer pago):
Plazo en meses 6 MESES
1a. PRÓRROGA AL 30/11/2010
Fecha de Inicio y Finalización: 01/02/2010 al 31/07/2010 2a. PRÓRROGA AL 31/01/2011
Grupo Renglón Nombre del Gasto Asignación
Presupuestaria
TRANSFERENCIA En Ejecución
Menos (-) Mas (+) Ejecutado Pendiente de
Ejecutar
1 Servicios no personales
181
Estudios, investigaciones y proyectos de
factibilidad Q 45,000.00 Q 45,000.00 Q -
181
Estudios, investigaciones y proyectos de
factibilidad (Evaluación Externa de Impacto) Q 8,000.00 Q 8,000.00
2 MATERIALES Y SUMINISTROS
261 Elementos y compuestos químicos Q 100,000.00 Q 99,520.29 Q 479.71
292 Útiles de limpieza y productos sanitarios Q 1,250.00 Q 1,249.50 Q 0.50
295
Útiles menores, médico-quirúrgicos y de
laboratorio Q 54,150.00 Q15,367.73 Q 37,721.86 Q 1,060.41
3
PROPIEDAD, PLANTA, EQUIPO E
INTANGIBLES
323 Equipo médico-sanitario y de laboratorio Q 5,367.73 Q 15,367.30 Q 0.43
GASTOS DE ADMÓN. (10%) Q 20,840.00 Q 20,840.00 Q -
Q 229,240.00 Q5,367.73
Q
15,367.73 Q 219,698.95 Q 9,541.05
MONTO AUTORIZADO Q 229,240.00
Disponibilidad Q 9,541.05
(-) EJECUTADO Q 219,698.95
SUBTOTAL Q 9,541.05
(-) CAJA CHICA
TOTAL POR EJECUTAR Q 9,541.05

66
IV.10 CRONOGRAMA DE LA INVESTIGACIÓN
ACTIVIDAD
2010 2011
FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT
Preparación de material
para muestreo
Toma de muestras
Envío y recepción de
muestras
Preparación de las
Muestras
Prueba de Laboratorio
de Inhibición de la
Hemoaglutinación
Análisis de Resultados
Elaboración de
cuadros, conclusiones y
recomendaciones
Presentación de
Informe Final
Presentación y
publicación en revista