college hospital - jb.asm.orgjb.asm.org/content/33/4/389.full.pdfandnewyorkmedical college andflower...
TRANSCRIPT

VIABILITY OF BACTERIA IN SEA WATER'
SELMAN A. WAKSMAN AND MARGARET HOTCHKISS
Woods Hole Oceanographic Institution, NewJersey Agricultural Experiment Station,and New York Medical College and Flower Hospital
Received for publication October 23, 1936
INTRODUCTORY
It has been repeatedly found that the bacterial population inthe sea is very limited. This is especially true of sea water in anatural state, although the sea floor, particularly the uppermostsurface layer, may contain large numbers of bacteria. Fre-quently, not more than ten or a dozen bacterial cells are found in1 cc. of sea water when it is plated out immediately upon asuitable agar or gelatin medium. Close to shore and at certainseasons of the year, this number may reach 500 to 1,000 cells,while far from land it may be as low as 1 or 2 cells per cubiccentimeter of sea water. This exceptionally small number ofviable cells is not due to a lack of food material in the sea water,since, when the latter is allowed to remain undisturbed in thelaboratory for 24 to 72 hours, the number of bacteria capable ofdeveloping into colonies on the plate may increase to 50,000 per1 cc. and even to 10 or 20 times that figure. An appreciableincrease may take place even within 2 to 4 hours, or during theperiod when the sample of sea water is being brought to thelaboratory.Among the explanations suggested for the very low numbers of
viable bacteria in natural waters, the following may be mentioned:(1) the presence in sea water of toxic substances which aredestructive to bacteria under natural conditions (ZoBell, 1936);(2) the presence of bacteriophage in the water (Arloing et al.,
1 Contribution No. 124 of the Woods Hole Oceanographic Institution; JournalSeries Paper of the New Jersey Agricultural Experiment Station, and contributionfrom New York Medical College and Flower Hospital.
389
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

390 SELMAN A. WAKSMAN AND MARGARET HOTCHKISS
1925); (3) the adsorption of the bacteria by the sea bottom andtheir sedimentation (Rubentschik et al., 1936); (4) the bactericidaleffect of sunlight (Gaarder and Spirck, 1931); (5) the consump-tion of the bacteria by protozoa and other small animal organisms(Waksman and Carey, 1935); (6) the possible presence in the seaof inactive bacteria which are capable of developing only undermore favorable conditions of temperature, aeration and foodsupply (Korinek, 1927); (7) the lack of sufficient nutriments inthe water; (8) the antagonistic relations of other microorganisms;(9) inadequacy of methods commonly used for counting marinebacteria.Although all of these suggestions may be partly justified, espe-
cially under controlled conditions, it is doubtful whether any oneof them can explain the phenomena observed in the sea. Mostof the investigations, upon which the above explanations rest,were carried out by introducing a pure bacterial culture or acomplex natural population into a medium to which it had notbeen previously accustomed; an example of this is the introduc-tion of typhoid bacteria into sewage or of sewage or river waterinto sea water.
It has been known since the work of Giaxa (1889) and Miquel(1891) that natural or raw water in which bacteria have grownbecomes unfit as a medium for the growth of bacteria, due to theformation of certain soluble and toxic products. Although Greig-Smith (1914) has found that the toxicity is increased by boiling,and ZoBell (1936) has reported that autoclaved and Berkefeld-filtered sea water are also bactericidal, other investigators (Giaxa,1889) have come to the conclusion that the toxic agents aredestroyed by heat.The following experiments were undertaken in order to obtain
further information concerning the survival of bacteria in seawater. Either the undisturbed natural population was used or aculture of a typical marine bacterium was added to the water andits survival determined. An agar-liquefying marine organism,kept in the Culture Collection of the Woods Hole OceanographicInstitution as No. 11, was selected for this purpose; it grewrapidly and the colonies on the plate could easily be distinguishedfrom the majority of other marine bacteria.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VIABILITY OF BACTERIA IN SEA WATER
EXPERIMENTAL
The first experiment deals with the survival of the marine agar-liquefying bacterium in fresh sea water, in heated sea water andin artificial salt water. It was found that heating of sea water at80'C., for 30 minutes, was sufficient for sterilization; highertemperatures and pressures were avoided to prevent formation ofany toxic substances. The salt water was prepared according tothe formula of Allen (1914) and sterilized as above. The seawater for this and subsequent experiments was obtained fromWoods Hole Harbor, one hundred or so meters from the dock,filtered through paper and used immediately.One hundred cubic centimeter quantities of sea water were
placed in sterile 250 cc. Erlenmeyer flasks and plugged withcotton. Bacterium No. 11 was grown for 24 hours in a liquidmedium containing 1 gram glucose, 1 gram peptone and 0.5 gramK2HPO4, in 1,000 cc. sea water. The major reason for using amedium so low in nutrients was to avoid the addition with theinoculum of any appreciable amounts of unused organic com-pounds to the sea water. The bacterial culture was added in 0.5and 2.5 cc. amounts to the various flasks of water. These wereincubated in the dark at room temperature (18 to 20'C.) andplated out, usually in two dilutions, on an agar medium containingthe above nutrients plus 1.5 per cent agar. The plates wereincubated for 48 hours and the colonies counted. This periodof incubation was found to be sufficient for the development ofthe agar-liquefying bacterium and for a number of the commonsea water bacteria.The results presented in table 1 show that there was active
bacterial multiplication in the fresh and heated sea water, aswell as in the prepared salt water. The increase in bacterialnumbers may have been due partly to their continued growthat the expense of the small amounts of nutrient material whichstill remained in the culture inoculum, and partly to a betterdispersion of the cells. In the fresh sea water, the bacterialpopulation of the water itself also began to grow rapidly. Themaximum numbers were attained in 24 to 48 hours. After thatperiod, the number of bacteria began to diminish rapidly in the
391
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

392 SELMAN A. WAKSMMAN AND MARGARET HOTCHKISS
fresh sea water, reaching a minimum in 13 to 29 days. Thisminimum was comparable to the number of bacteria found in theuninoculated fresh sea water at the same incubation period. Theplates showed few colonies of organism No. 11, but numerouscolonies of the usual sea water bacteria were present. In theheated water, however, Bacterium No. 11 remained in largenumbers up to 13 days and then began to decrease slowly, accord-ing to counts made on the 29th day of incubation. Between 29and 56 days the numbers decreased rapidly. The behavior of the
TABLE 1Survival of an agar-liquefying marine bacterium in fresh sea water, in sterile sea
water, and in prepared salt waterNumbers of bacteria in 1 cc., in thousands
TREATMExT OF WATER
PE9RIOD OFrINCIJ- Untreated fresh sea water Heatedseawater Salt waterNATION Amount of culture No. 11 added
None 0.5 cc. 2.5 cc. None 0.5 cc. 2.5 cc. None 0.5 cc. 2.5 cc.
Start 1.6 930 4,250 0 2,300 6,250 0 1,470 5,8001.5 hours 2.3 2,800 19,400 0 3 ,780 24,800 0 1,690 15, 8004.0 hours 2 4,950 35,600 0 4,600 31,000 0 3,930 24,30021.5 hours 29 15,100 70,000 0 5,420 23,600 0 3,260 27,400
2 days 120 9,000 56,000 0 9,500 34,000 0 2,700 29,5004 days 79 688 0 9, 150 32,500 0 2,450 27,7007 days 100 2,140 608 7,900 34,800 2,560 31,00013 days 52 33.5 25.5 5,300 21,200 2,980 21,50029 days 25 2.2 29.8 2,650 9,200 700 7,40056 days 21 39.8 63.4 88 215 105 205
bacteria added to the prepared salt water was very similar to thatof the bacteria in the heated sea water.The results of this experiment have thus brought out definitely
the fact that, in fresh sea water, a destructive effect was exertedupon the cells of a marine bacterium added to it. This destruc-tion of the bacteria was usually accompanied by the formationof a very fine coagulum throughout the water, which graduallysettled to the bottom of the container.
In order to determine the survival of bacteria in Berkefeld-filtered sea water, a quantity of fresh sea water was filtered
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
VIABILITY OF BACTERIA IN SEA WATER
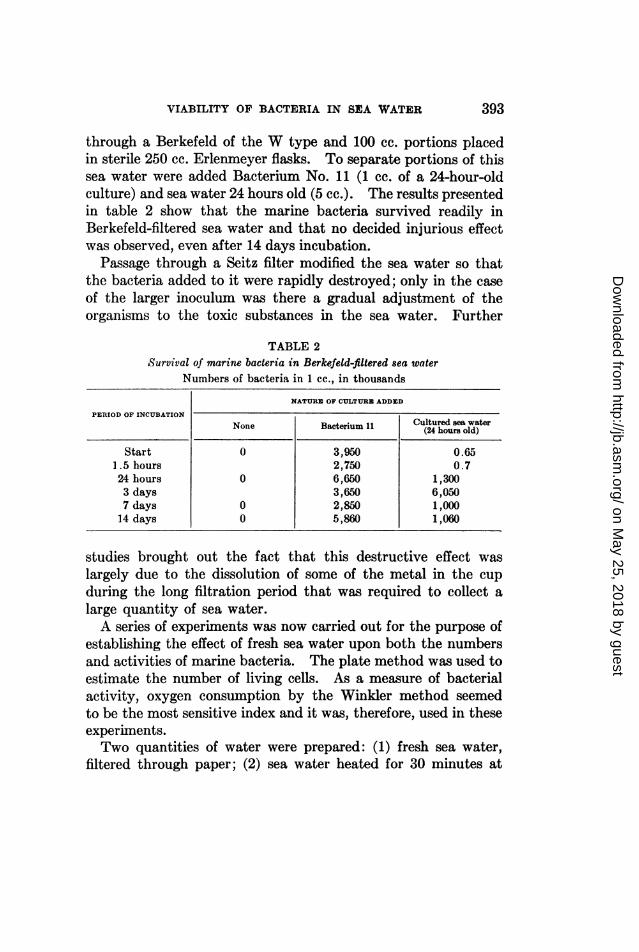
through a Berkefeld of the W type and 100 cc. portions placedin sterile 250 cc. Erlenmeyer flasks. To separate portions of thissea water were added Bacterium No. 11 (1 cc. of a 24-hour-oldculture) and sea water 24 hours old (5 cc.). The results presentedin table 2 show that the marine bacteria survived readily inBerkefeld-filtered sea water and that no decided injurious effectwas observed, even after 14 days incubation.
Passage through a Seitz filter modified the sea water so thatthe bacteria added to it were rapidly destroyed; only in the caseof the larger inoculum was there a gradual adjustment of theorganisms to the toxic substances in the sea water. Further
TABLE 2Survival of marine bacteria in Berkefeld-filtered sea water
Numbers of bacteria in 1 cc., in thousands
NATURE OF CULTURE ADDED
PERIOD OF INCUBATION
None ~~Bacterium 11 Cultured sea waterNone Bacterium 11 (24 hours old)
Start 0 3,950 0.651.5 hours 2,750 0.724 hours 0 6,650 1,3003 days 3,650 6,0507 days 0 2,850 1,00014 days 0 5,860 1,060
studies brought out the fact that this destructive effect waslargely due to the dissolution of some of the metal in the cupduring the long filtration period that was required to collect alarge quantity of sea water.A series of experiments was now carried out for the purpose of
establishing the effect of fresh sea water upon both the numbersand activities of marine bacteria. The plate method was used toestimate the number of living cells. As a measure of bacterialactivity, oxygen consumption by the Winkler method seemedto be the most sensitive index and it was, therefore, used in theseexperiments.Two quantities of water were prepared: (1) fresh sea water,
filtered through paper; (2) sea water heated for 30 minutes at
393
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SELMAN A. WAKSMAN AND MARGARET HOTCHKISS
80'C. In order to re-saturate the heated water with oxygen,the flasks containing this water were placed in a refrigeratorovernight; the water was then de-saturated by placing the flasks,for a few minutes, in warm water. The oxygen content of thefresh and heated sea water was 5.43 and 5.62 per liter, respec-tively. Both lots of water were distributed into a series ofcarefully cleaned and dried 200 cc. oxygen bottles. These wereinoculated with varying amounts of a 48-hour culture of Bac-terium No. 11. The bottles were incubated under water in thedark and removed at various intervals for the determination ofbacterial numbers and oxygen concentration.The destructive effect of the fresh sea water upon the added
bacteria was observed (table 3) within 23 hours incubation; thiseffect became especially striking after 3 days, when practicallyall the added bacteria had disappeared. In the heated seawater, however, the bacteria remained at a high level up to ninedays. Although the bacteria had been reduced to a minimumin the bottles containing the fresh sea water, there was greateroxygen consumption in these bottles than in those containingthe heated sea water, where the added bacteria survived in muchgreater numbers. The experiment was repeated, with similarresults. This could lead to but one conclusion, namely, that theagents responsible for the destruction of the bacterial cells inthe fresh sea water were not necessarily destructive to theprocesses concerned in the decomposition of organic matter inthe water, as measured by oxygen consumption. Either theseagents themselves consumed the oxygen in the water, or the fewernumbers of bacteria left possessed a much greater power ofoxygen consumption (and, therefore, of organic matter decom-position), or a large part of the oxygen consumption was broughtabout by bacteria which were unable to develop on the agar plate.
In the following experiment, an attempt was made to obtainfurther information in regard to the relationship between thesurvival of bacteria in sea water and oxygen consumption, tosearch for the presence of other organisms in sea water whichmight modify bacterial activities, and to determine the changestaking place in the water during the early periods after the addi-
394
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VIABILITY OF BACTERIA IN SEA WATER 395
tion of the bacteria. All the bottles were sterilized by dry heat,for 18 hours at 60 to 650C. Several check determinations weremade, to eliminate the variability of the individual bottles, whichhad been frequently obtained in the previous experiments. A
TABLE 3Survival of an agar-liquefying marine bacterium in fresh and in heated sea water
NATURE OF WATER FRESH HEATED
Incubation Inoculum Bacteria in Oxygen Bacteria in Oxygenadded 1 cc. consumed 1 cc. consumed
3.5 hours3.5 hours3.5 hours3.5 hours23 hours23 hours23 hours23 hours3 days3 days3 days3 days5 days5 days5 days5 days9 days9 days9 days9 days15 days15 days15 days15 days
cc.
00.10.31.000.10.31.000.10.31.000.10.31.000.10.31.000.10.31.0
thousands
1,6505,750
8.5685
3,250
26316.52.84.95.57.0
Few4.55.5
0.91.62.8
cc. per liter
0.060.090.390.120.66
2.16
0.751.664.380.390.871.745.16
2.405.43*
1.172.465.43*
thousands
9102,2508,150
01,2002,3355,150
1,4902,14010,680
50980
1,8506,390
4401,3505,350
1151,9301,850
cc. per liter
0.100.160.6100.160.250.82
0.400.432.740.010.400.702.89
0.520.883.58
0.581.183.88
* Oxygen all used up.
24-hour culture of No. 11 was employed. The results presentedin table 4 again confirmed those obtained previously.
In the case of the fresh sea water, the bacterial maximum wasobtained within 24 hours, followed by a drop in the number ofbacteria. Within 3 days, practically all the added bacteria had
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
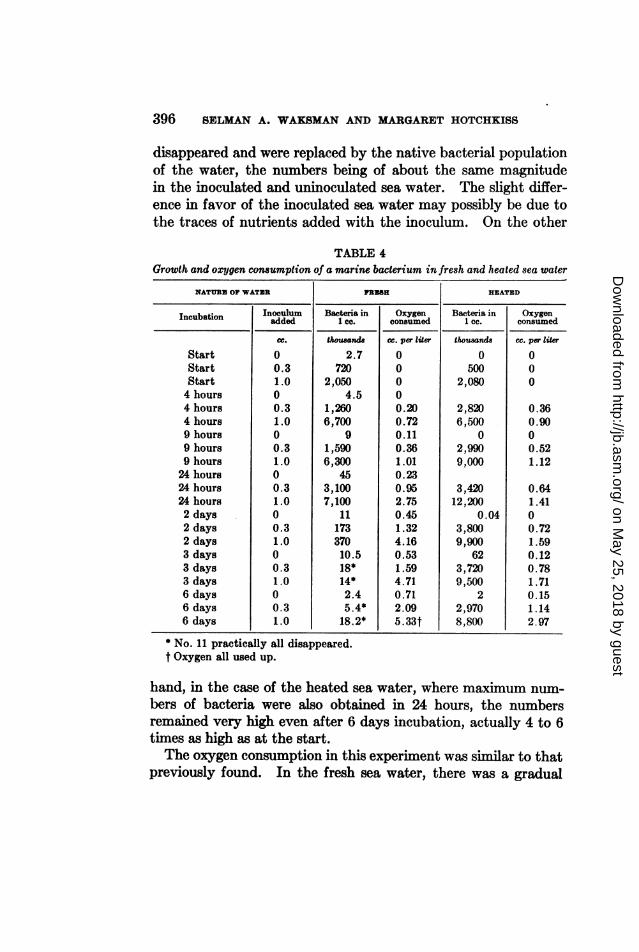
396 BELMAN A. WAKSMAN AND MARGARET HOTCHKISS
disappeared and were replaced by the native bacterial populationof the water, the numbers being of about the same magnitudein the inoculated and uninoculated sea water. The slight differ-ence in favor of the inoculated sea water may possibly be due tothe traces of nutrients added with the inoculum. On the other
TABLE 4Growth and oxygen consumption of a marine bacterium in fresh and heated sea water
NATUREl~ OF WATER FR19ES HEATEDInoculum~~ ~ ~BatraiOe atri n Oye
Incubation
StartStartStart4 hours4 hours4 hours9 hours9 hours9 hours24 hours24 hours24 hours2 days2 days2 days3 days3 days3 days6 days6 days6 days
Inoculumadded
cc.
00.31.000.31.000.31.000.31.000.31.000.31.000.31.0
Bacteria in1 cc.
thousand.2.7
7202,050
4.51,2606,700
91,5906,300
453,1007,100
1117337010.518*14*2.45.4*18.2*
* No. 11 practically all disappeared.t Oxygen all used up.
Oxygenconsumed
cc. per liter
00000.200.720.110.361.010.230.952.750.451.324.160.531.594.710.712.095.33t
Bacteria in1 cc.
thousand.
0500
2,080
2,8206,500
02,9909,000
3,42012,200
0.043,8009,900
623,7209,500
22,9708,800
Oxygenconsumed
cc. per liter
000
0.360.9000.521.12
0.641.4100.721.590.120.781.710.151.142.97
hand, in the case of the heated sea water, where maximum num-bers of bacteria were also obtained in 24 hours, the numbersremained very high even after 6 days incubation, actually 4 to 6times as high as at the start.The oxygen consumption in this experiment was similar to that
previously found. In the fresh sea water, there was a gradual
-I-IlI
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VIABILITY OF BACTERIA IN SEA WATER
disappearance of the oxygen, the rate of its consumption depend-ing on the amount of inoculum added. The greater consumptionof the oxygen was due both to the presence of larger numbers ofbacterial cells and to greater concentration of the unconsumednutrients. In practically all cases, the ratio for oxygen consump-tion in the bottles receiving 0.3 and 1.0 cc. of the inoculum wassimilar to the ratio of the bacterial numbers in the originalinocula. However, after 24 hours incubation, the increasingoxygen consumption was quite independent of the numbers ofbacteria since the latter decreased rapidly, while the former,being cumulative, continued to increase. In the heated seawater, the bacterial numbers remained very high even after 6 daysincubation, the total number per bottle with 1 cc. inoculum being1,760,000,000 or four times as great as the number of bacteriaoriginally introduced. The oxygen consumption was still con-siderably less than that in the fresh sea water.The water from several of the bottles was centrifuged at the end
of the incubation periods and the resulting sediments examinedmicroscopically for the presence of protozoa and other animals.This examination revealed the fact that the heated water con-tained a large number of bacteria in addition to some deaddiatomaceous material. The fresh sea water, on the other hand,contained many protozoa, a number of copepods (12 living formshaving been counted in one bottle) and various other lower animalforms.One may draw from these experiments two broad conclusions,
namely, (1) that sea water contains an agent or agents destructiveto marine bacteria, and (2) this agent does not interfere with theprocesses of marine organic matter decomposition, as measured byoxygen consumption. These seemingly contradictory conclusionsare possible only by assuming that the agents which are largelyresponsible for the destruction of the bacteria form a part of thesystem which is responsible for the decomposition of the organicmatter in the sea water. This explanation can help in under-standing the reasons for the low bacterial population in naturalsea water; the bacterial population is largely held down throughthe activities of the numerous animals inhabiting the sea.
397
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

398 SELMAN A. WAKSMAN AND MARGARET HOTCHKISS
DISCUSSION
The abundance of bacteria in sea water is controlled by a num-ber of factors, chief among which are (1) the amount and natureof available food material; (2) the rate of bacterial multiplication,leading to the formation of substances injurious to bacterialdevelopment; (3) the nature and abundance of the animal popu-lation, ranging from minute protozoa to larger forms; (4) theenvironmental conditions, as exemplified by temperature, andaeration or stagnation of water.
It has been amply demonstrated that extensive bacterialmultiplication takes place in sea water, either as a result of (a)adding fresh organic matter in the form of algal or of animalresidues; (b) keeping the water in an undisturbed condition; (c)warming the water. Rapid bacterial growth is sooner or laterfollowed by a drop in numbers; the extent of the drop and theequilibrium which is finally established depend on the natureof the stimulus and on the amount of residual organic matter inthe water.The drop in bacterial numbers is greatly delayed and its rate
is much slower, when the bacteria are added to sterile sea waterinstead of to the same water in a fresh state. The bacterialproducts and the exhaustion of available nutrients cannot,therefore, be alone responsible for the low numbers of bacteriawhich remain in fresh water in a living and active state. It isbelieved that another factor, namely the animal population,largely the protozoa and other members of the nannoplanktonin the water, is also responsible for bringing about the reductionin the numbers of bacteria. This assumption is substantiated bytwo facts, first, the multiplication of these organisms in freshwater in which bacterial development has taken place, andsecondly, the greater oxygen consumption in the fresh sea wateras compared with the sterile sea water, in spite of the greaternumbers of bacteria in the latter.Consumption of oxygen is a measure of the respiratory processes
of living systems. When light is excluded, especially in the caseof water containing only a limited number of diatoms, the process
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VIABILITY OF BACTERIA IN SEA WATER
of oxygen consumption is carried out by the bacteria and theanimal population. The reduction in bacterial numbers accom-panied by the growth of animal forms can only mean that thelatter are partly responsible for both the destruction of the bac-teria and the consumption of the oxygen.The limited numbers of bacteria under natural conditions in
the sea can thus be explained by a state of equilibrium betweenbacterial multiplication and bacterial destruction. A change inthis equilibrium is brought about by a change in food supply,environmental conditions and modification of agents unfavorableto bacteria, whether these are simple organic substances resultingfrom bacterial metabolism, bacteriolytic substances, bacteri-ophagic agents, or organisms feeding directly upon the bacteria.
SUMMARY
1. A study has been made of the survival of a marine bacteriumadded to fresh sea water, to sea water sterilized by heat or by fil-tration through a Berkefeld filter, and to prepared salt water.
2. The fresh sea water exerted a marked destructive effect uponthe cells of the marine bacterium added; this effect followed aninitial rise in numbers. Sterilized sea water and prepared saltwater had no destructive effect upon the bacteria.
3. The destructive effect of fresh sea water upon the bacteriawas not accompanied by a similar effect upon the rate of decom-position of the organic matter in the water, as measured byoxygen consumption.
4. It is suggested that the activities of the animal populationof the sea, especially the nannoplankton, can explain, at leastpartly, both the destruction of the bacteria and the stimulationof the processes of organic matter decomposition.
5. These facts are used as a basis for explaining the low numbersof bacteria usually found in natural sea water.
The authors are indebted to Dr. M. Butler for assistance inmaking the oxygen determinations.
399
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

400 SELMAN A. WAKSMAN AND MARGARET HOTCHKISS
REFERENCESALLEN, E. J. 1914 Jour. Mar. Biol. Ass., 10, 417-439.ARLOING, F., SEMPE, AND CHAVANNE 1925 Bull. Acad. Med. (Paris), 93,
184-187.GAARDER, T., AND SPXRCK, R. 1931 Rapports et Proces-Verbaux, 75, 47.GIAXA, D. 1889 Ztschr. Hyg., 6, 162-224.GREIG-SMITH, R. 1914 Proc. Linn. Soc. N. S. Wales, 39, 533-537.KORINEK, J. 1927 Centrbl. Bakt. II, 66, 500-505, 1926; 71, 73-79.MIQUEL 1891 Revue d'Hygiene, 9, 737; Manuel pratique d'Analyse bacteri-
ologique des eaux. Paris. (Ref. Frankland. Microorganisms inWater. London, 1894.)
RUBENTSCHIK, L., ROIsIN, M. B., AND BIELJANSKY, F. M. 1936 Jour. Bact.,32, 11-31.
WAKSMAN, S. A. AND CAREY, C. L. 1935 Jour. Bact., 29, 531-544, 545-561.ZOBELL, C. E. 1936 Proc. Soc. Exp. Biol. Med., 34, 113-116.
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from