circulation under basal conditions and after vasodilator
TRANSCRIPT

Br. J. Pharmacol. (1992), 106, 563-567
Effects of nitric oxide synthesis inhibition on the goat coronary
circulation under basal conditions and after vasodilatorstimulationJ.L. Garcia, N. Fernaindez, A.L. Garcia-Villalon, L. Monge, B. Gomez &1G. Dieguez
Departamento de Fisiologia, Facultad de Medicina, Universidad Autonoma, Arzobispo Morcillo 1, 28029 Madrid, Spain
1 The role of nitric oxide in the coronary circulation under basal conditions and when exposed tovarious vasodilator stimuli was studied in instrumented, anaesthetized goats, by examining the action ofinhibiting endogenous nitric oxide production with NG-nitro-L-arginine methyl ester (L-NAME).2 In 12 goats, left circumflex coronary blood flow (electromagnetically measured), systemic arterialblood pressure and heart rate were continuously recorded. L-NAME (3-4, or 8-10mg kg' injectedi.v.) decreased resting coronary blood flow by 20 and 28%, increased mean arterial pressure by 23 and30% and increased coronary vascular resistance by 47 and 65%, respectively, without affecting heartrate, or blood gases or pH. These haemodynamic effects were reversed by L-arginine (200- 300 mg kg-'by i.v. injection, 5 goats).3 Acetylcholine (0.001-0.1 lg), sodium nitroprusside (0.01-0.3 mg), and diazoxide (0.1-3 mg),injected intracoronarily in 6 goats, produced dose-dependent increases in coronary blood flow; sodiumnitroprusside (0.1-0.3 mg) also caused hypotension and tachycardia.4 During the effects of L-NAME, the coronary vasodilatation to acetylcholine was attenuated, tosodium nitroprusside was increased, and to diazoxide was unaffected, in comparison with controlconditions. The hypotensive effects of sodium nitroprusside were also increased during treatment withL-NAME.5 Graded coronary hyperaemic responses occurred after 5, 10 or 20 s of coronary occlusion. Themagnitude of hyerpaemia for each occlusion duration was increased during treatment with L-NAME, incomparison to control.6 The results suggest: (a) endogenous nitric oxide is involved in regulation of coronary circulation byproducing a basal vasodilator tone, (b) acetylcholine-induced coronary vasodilatation is mediated, inpart, by nitric oxide, and (c) inhibition of basal endogenous nitric oxide production induces supersen-sitivity of coronary vessels to nitrovasodilators and enhances hyperaemic responses after short periods ofischaemia of the myocardium.
Keywords: Endothelium; NG-nitro-L-arginine methyl ester (L-NAME); vasodilator tone; reactive hyperaemia; acetylcholine;nitrovasodilators; supersensitivity
Introduction
Since the original observation by Furchgott & Zawadzki(1980) it is now recognized that the endothelium plays amajor role in regulating vascular tone by releasing vasoactivefactors and by modulating vascular responses to variousstimuli (Furchgott & Vanhoutte, 1989). Experimentalevidence indicates that nitric oxide, or a closely related com-pound, is at least one type of endothelium-derived relaxingfactor that is synthesized from L-arginine (Palmer et al.,1987; 1988; Ignarro et al., 1987) and relaxes vascular smoothmusculature via the stimulation of guanylate cyclase(Rapoport & Murad, 1983; Ignarro et al., 1986). The syn-thesis of endothelial nitric oxide can be inhibited by severalL-arginine analogues, and this inhibition induces vascularcontraction in vitro and in vivo (Moncada & Higgs, 1990),increases resistance in several vascular beds (Gardiner et al.,1990a; 1991) and reduces blood flow in the brachial artery ofman (Vallance et al., 1989). Thus, the basal release of nitricoxide (Martin et al., 1986; Rees et al., 1989a) appears to beresponsible for maintaining a vasodilator tone in the cardio-vascular system (Rees et al., 1989b; Gardiner et al., 1990a).
Author for correspondence.
Endothelial function has been regarded as an importantfactor in the regulation of the coronary circulation and itmay be of pathophysiological significance in several diseasestates (Harrison, 1989). However, many questions remain tobe answered regarding the mechanisms by which theendothelium achieves this function. Endothelium-dependentvasodilatation has been found in coronary vessels from dogs(Rubanyi et al., 1986; Hayashi et al., 1988; Myers et al.,1989) and man (Hodgson & Marshall, 1989). More recentlyit has been reported that human coronary vessels in vitrorelease nitric oxide, which is cleaved from L-arginine (Chesteret al., 1990) and that nitric oxide plays a significant role inmodulating basal vasomotion and endothelial-dependentdilatation in the coronary circulation of dogs (Chu et al.,1991).The present experiments were performed to study the role
of nitric oxide in basal vascular tone and in vasodilatation ofthe coronary circulation of anaesthetized goats. To this end,the animals were instrumented and the effects of a potentinhibitor of the nitric oxide synthesis, N0-nitro-L-argininemethyl ester (L-NAME) (Moore et al., 1990; Rees et al.,1990), were investigated both on the basal coronary bloodflow and on coronary vasodilator responses to acetylcholine,sodium nitroprusside, diazoxide and ischaemia.
'." Macmillan Press Ltd, 1992

564 J.L. GARCIA et al.
Methods
Twelve female goats (32-49 kg) were anaesthetized with an
intramuscular injection of 10 mg kg-' ketamine hydro-chloride and i.v. administration of 2% thiopentone sodium;supplemental doses were given as necessary for maintenance.After orotracheal intubation, artificial respiration with room
air was instituted by use of a Harvard respirator. Adjust-ments in tidal volume or respiratory rate were used to main-tain blood gases and pH within the following range:
P02 = 85-110 mmHg; Pco2 = 26-40 mmHg; pH = 7.35-7.45.A polyethylene catheter was placed in one temporal artery
to measure arterial blood pressure (Statham transducer), andto obtain blood samples for gas and pH analysis(Radiometer ABL, Copenhagen, Denmark).A thoracotomy was performed through the fourth left
intercostal space, and the pericardium opened to gain access
to the left aspect of the heart. The proximal segment of theleft circumflex coronary artery was dissected and an electro-magnetic flow transducer (Biotronex) was placed on thisartery to measure blood flow. A snare type occluder was alsoplaced around this artery, just distal to the flow probe, toobtain zero flow baseline and to produce transient coronary
ischaemias.Coronary blood flow, systemic arterial pressure and heart
rate were continuously recorded on a Beckman recorderthroughout the experiments.
In this work the following experiments were performed: (1)after resting control measurements were recorded, 12 animalsreceived an i.v. bolus of L-NAME (3-4 mg kg-' within5 min), and 5 min after the haemodynamic variables hadreached a new steady state, animals received an additionali.v. infusion of L-NAME (0.05-0.08 mg kg-' min-1 over
70-90 min). L-NAME was dissolved in isotonic saline(5 mg ml'), and each animal received in total 8-Omg kg-'.In five of these goats 200-300mgkg-' of L-arginine was
given via i.v. route when the i.v. infusion of L-NAME was
stopped. L-Arginine, dissolved in isotonic saline (50mgml-'),was administered over 10-20min; (2) the effects of acetyl-choline (0.001, 0.003, 0.01, 0.03 and 0.1 fig), sodium nitro-prusside (0.01, 0.03, 0.1 and 0.3 mg), and diazoxide (0.1, 0.3,1 and 3 mg) were recorded in 6 animals before and duringthe i.v. infusion of L-NAME. All these substances were dis-solved in isotonic saline and given in volumes of 0.3 mldirectly- into the left circumflex coronary artery through a
27G needle; these substances were injected in randomsequence, and the administration of each dose was separatedat least by 5 min, and (3) in 5 animals hyperaemic responsesfollowing occlusions of the left circumflex coronary arterylasting 5, 10 and 20 s were recorded before and during thei.v. infusion of L-NAME. The order of occlusions was
randomized, and each observation was the average of 2occlusions for each occlusion duration. To determinehyperaemic responses, the following measurements and cal-culations were made (Marcus et al., 1981): (a) the control(basal) and the peak hyperaemic coronary blood flows were
measured and then the peak hyperaemic flow to control flowratio was calculated, and (b) the repayment-to-debt ratio was
calculated as reactive hyperaemia blood flow divided bytheoretical debt of blood flow. Reactive hyperaemia bloodflow is the blood flow in ml during the total hyperaemicresponse over the control blood flow and was determinedplanimetrically on the recordings. The debt of blood flow isthe blood flow in ml that theoretically would have occurredduring arterial occlusion and was calculated as the controlflow x the occlusion duration.
Resistance to blood flow through the left circumflex cor-
onary artery was calculated before and during the effects ofL-NAME as mean systemic arterial pressure in mmHgdivided by the coronary blood flow in ml min'.Drugs used were: NG-nitro-L-arginine methyl ester (L-
NAME, Sigma), acetylcholine chloride (Sigma), sodium
nitroprusside (Sigma) and diazoxide (Hyperstat, Schering).
Administration of isotonic saline alone intravenously orintracoronarily at the volumes employed in the present studyhad no systemic or coronary vascular effects.
Statistics: all haemodynamic measurements before and dur-ing treatment with L-NAME were compared using the sameanimal as its own control. Statistical analyses were performedusing an analysis of variance for repeated measures, followedby an individual Student's t test for paired data.
Results
Effects of NG-nitro-L-arginine methyl ester
Mean values for the circumflex coronary blood flow, meansystemic arterial pressure, coronary vascular resistance andheart rate obtained in 12 goats before and after treatmentwith L-NAME are summarized in Table 1. The i.v. bolus ofL-NAME (3-4mg kg-' within 5 min) increased systemicarterial pressure by about 23% (P<0.001), decreased cor-onary blood flow by about 20% (P<0.001), increasedcoronary vascular resistance by about 47% (P<0.001), butdid not change heart rate significantly (P> 0.05). Theseeffects occurred in all the animals except one in which cor-onary blood flow did not change; they were evident beforecompletion of L-NAME injection. These haemodynamicchanges were slightly accentuated during the additional i.v.infusion of L-NAME (0.05-0.08mgkg-1 over 70-90min);under these conditions, mean arterial pressure increasedabout 30%, coronary blood flow decreased about 28% andcoronary vascular resistance increased about 65% withrespect to control values. In these circumstances only thechanges in coronary vascular resistance were significantly(P< 0.05) different from the haemodynamic changes ob-tained after the i.v. bolus of L-NAME. Blood gases and pHduring the effects of L-NAME were not significantly differentfrom those obtained under control conditions. In 7 of theseanimals, the haemodynamic effects induced by L-NAMEremained, at least, for 1 h after stopping its administration.
In 5 goats, administration of L-arginine, after stopping thei.v. infusion of L-NAME, decreased systemic arterial pres-sure, increased coronary blood flow, decreased coronary vas-cular resistance but did not change heart rate. Thehaemodynamic changes induced by L-NAME returned tocontrol values within 5-10 min after stopping the i.v.administration of L-arginine (Table 1). This contrasted withthe haemodynamics in the 7 goats treated with L-NAME, butnot with L-arginine.
Effects of acetylcholine
Acetylcholine (0.001-0.1 pg), injected into the left circumflexcoronary artery, produced dose-dependent increases in cor-onary blood flow, but did not change systemic arterial pres-sure significantly; the doses of 0.03 and 0.1 pg also causedbradycardia. However, the increments in coronary blood flowproduced by acetylcholine during i.v. infusion of L-NAMEwere significantly lower than under control conditions(Figure 1).
Effects of sodium nitroprusside
Sodium nitroprusside (0.01-0.3 mg), injected into the leftcircumflex coronary artery, induced marked dose-dependentincreases in coronary blood blood flow; these effects weresignificantly greater during treatment with L-NAME thanunder control conditions. The doses of 0.1 and 0.3 mg ofsodium nitroprusside also induced decreases in systemicarterial pressure and caused tachycardia, and during treat-ment with L-NAME the doses of 0.03, 0.1 and 0.3 mg ofsodium nitroprusside induced systemic hypotension andtachycardia. The hypotensive effects of each dose of sodiumnitroprusside during L-NAME were significantly greater than
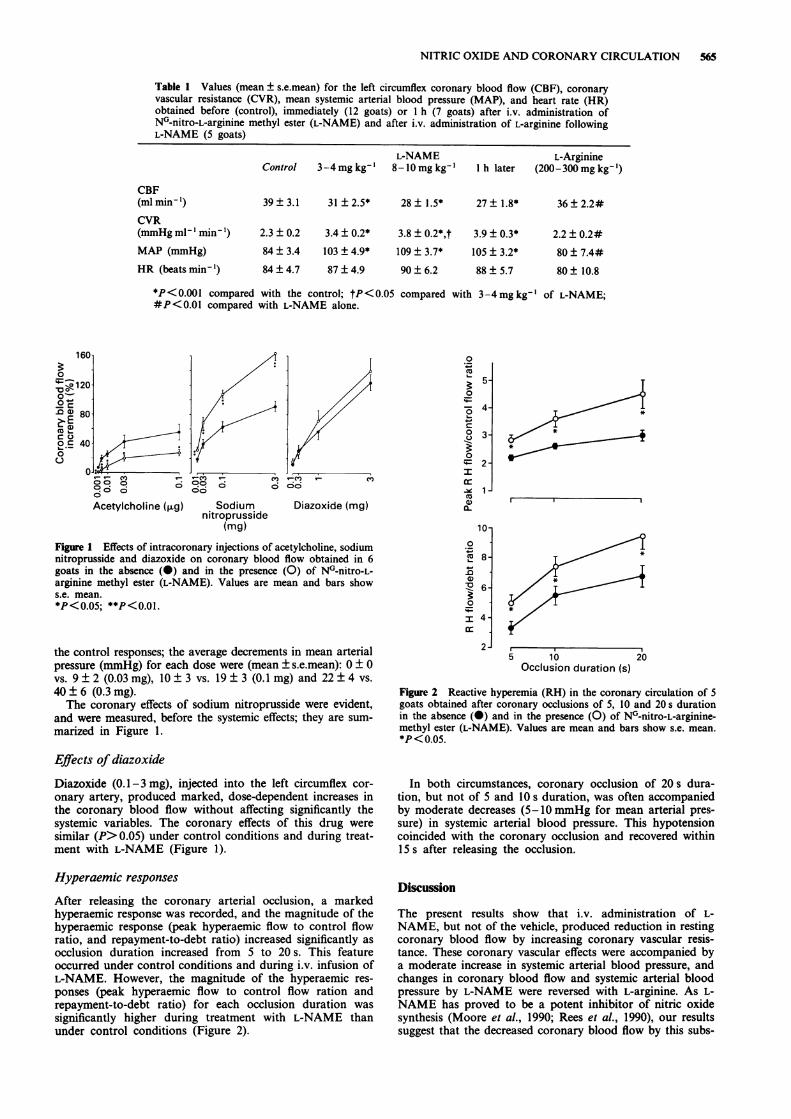
NITRIC OXIDE AND CORONARY CIRCULATION 565
Table 1 Values (mean ± s.e.mean) for the left circumflex coronary blood flow (CBF), coronaryvascular resistance (CVR), mean systemic arterial blood pressure (MAP), and heart rate (HR)obtained before (control), immediately (12 goats) or I h (7 goats) after i.v. administration ofN0-nitro-L-arginine methyl ester (L-NAME) and after i.v. administration of L-arginine followingL-NAME (5 goats)
L-NAMEControl 3-4mgkg-' 8-l0mgkg-'
CBF(ml minm-)CVR(mmHg ml-' min -')MAP (mmHg)
HR (beats min-')
L-Arginine1 h later (200-300 mg kg-')
39± 3.1 31 ±2.5* 28± 1.5* 27± 1.8*
2.3 ± 0.2 3.4 ± 0.2* 3.8 ± 0.21
84 ± 3.4 103 ± 4.9* 109 ± 3.71
84±4.7 87±4.9 90±6.2
'*,t 3.9 ± 0.3*
36 ± 2.2#
2.2±0.2#* 105 ± 3.2* 80 ± 7.4#
88 ± 5.7 80 ± 10.8
*P<0.001 compared with the control; tP<0.05 compared with 3-4mg kg-' of L-NAME;#P<0.01 compared with L-NAME alone.
0-0c) C') '-CY) c0)996C 6;6600
Sodiumnitroprusside
(mg)
Diazoxide (mg)
Figure 1 Effects of intracoronary injections of acetylcholine, sodiumnitroprusside and diazoxide on coronary blood flow obtained in 6goats in the absence (0) and in the presence (0) of N0-nitro-L-arginine methyl ester (L-NAME). Values are mean and bars shows.e. mean.*P<0.05; **P<0.01.
the control responses; the average decrements in mean arterialpressure (mmHg) for each dose were (mean ± s.e.mean): 0 ± 0vs. 9 ± 2 (0.03 mg), 10± 3 vs. 19 ± 3 (0.1 mg) and 22 ± 4 vs.
40 ± 6 (0.3 mg).The coronary effects of sodium nitroprusside were evident,
and were measured, before the systemic effects; they are sum-
marized in Figure 1.
Effects of diazoxide
Diazoxide (0.1-3 mg), injected into the left circumflex cor-
onary artery, produced marked, dose-dependent increases inthe coronary blood flow without affecting significantly thesystemic variables. The coronary effects of this drug weresimilar (P>0.05) under control conditions and during treat-ment with L-NAME (Figure 1).
Hyperaemic responses
After releasing the coronary arterial occlusion, a markedhyperaemic response was recorded, and the magnitude of thehyperaemic response (peak hyperaemic flow to control flowratio, and repayment-to-debt ratio) increased significantly as
occlusion duration increased from 5 to 20 s. This featureoccurred under control conditions and during i.v. infusion ofL-NAME. However, the magnitude of the hyperaemic res-
ponses (peak hyperaemic flow to control flow ration andrepayment-to-debt ratio) for each occlusion duration was
significantly higher during treatment with L-NAME thanunder control conditions (Figure 2).
0
5-
0
- 4-o
5 2-I
.v 1-
0)
10-
0uo
.0
4_,e
a)
6-
0
I 4-r-
2-
f ~~~~~~~~~*-
* X0 20
5 10 20Occlusion duration (s)
Figure 2 Reactive hyperemia (RH) in the coronary circulation of 5
goats obtained after coronary occlusions of 5, 10 and 20 s durationin the absence (0) and in the presence (0) of N0-nitro-L-arginine-methyl ester (L-NAME). Values are mean and bars show s.e. mean.*P<0.05.
In both circumstances, coronary occlusion of 20s dura-tion, but not of 5 and 10 s duration, was often accompaniedby moderate decreases (5-lO mmHg for mean arterial pres-sure) in systemic arterial blood pressure. This hypotensioncoincided with the coronary occlusion and recovered within15 s after releasing the occlusion.
Discussion
The present results show that i.v. administration of L-NAME, but not of the vehicle, produced reduction in restingcoronary blood flow by increasing coronary vascular resis-tance. These coronary vascular effects were accompanied bya moderate increase in systemic arterial blood pressure, andchanges in coronary blood flow and systemic arterial bloodpressure by L-NAME were reversed with L-arginine. As L-NAME has proved to be a potent inhibitor of nitric oxidesynthesis (Moore et al., 1990; Rees et al., 1990), our resultssuggest that the decreased coronary blood flow by this subs-
0g120-o ~ 80
40)_ c-0 'D 80-L.E
Co00
Acetylcholine (,ug)

566 J.L. GARCIA et al.
tance is related to reduction in basal nitric oxide production,and consequently with inhibition of nitric oxide-mediatedbasal vasodilator tone in the coronary vasculature in vivo.This agrees with observations in the dog coronary circulation(Chu et al., 1991) and supports the suggestion that a basalvasodilator tone mediated by nitric oxide is present in thecoronary circulation, as occurs in other vascular beds(Aisaka et al., 1989; Rees et al., 1989a; Gardiner et al.,1990b). In awake dogs N0-monomethyl-L-arginine (L-NMMA), another inhibitor of nitric oxide synthesis, inducedreductions in resting coronary blood flow of about 19% andincreases in systemic arterial blood pressure, withoutaffecting left ventricular contractility (Chu et al., 1991). Ourstudy shows that lower concentrations of L-NAME werenecessary to produce a reduction in coronary blood flow,comparable in magnitude to that found with L-NMMA (Chuet al., 1991). Heart rate decreases are frequently observedwhen inhibitors of nitric oxide are administered (Aisaka etal., 1989; Rees et al., 1989b; Gardiner et al., 1990b) but wedid not find this phenomenon, probably because anaesthesiamay block the baroreflex response (Widdop et al., 1992).Systemic hypertension to L-NAME has also been found byothers (Gardiner et al., 1990a; Moncada et al., 1991), and thehaemodynamic responses to this substance seems to differbetween vascular beds (Gardiner et al., 1991; Van Gelderenet al., 1991) and species (Van Gelderen et al., 1991).
Intracoronary administration of acetylcholine producedcoronary vasodilatation in anaesthetized goats, a feature alsoobserved in dogs (Cox et al., 1983; Reid et al., 1985; Chu etal., 1991) and man (Hodgson & Marshal, 1989). It appearsthat acetylcholine produces vasorelaxation by releasingendothelium-derived nitric oxide (Furchgott & Vanhoutte,1989; Moore et al., 1990; Rees et al., 1990; Gardiner et al.,1991) and we found that the acetylcholine-induced coronaryvasodilatation was attenuated during treatment with L-NAME. As the coronary vasodilator response to diazoxidewas preserved and those to sodium nitroprusside andischaemia were enhanced, the reduction in acetylcholine-induced vasodilatation after L-NAME was probably a con-sequence of inhibition of nitric oxide formation and notsecondary to a loss of ability of coronary vessels to dilate.This agrees with the observations in conscious dogs by Chuet al. (1991) and supports the idea that the coronaryvasodilatation by acetylcholine is mediated, at least in part,by release of endothelial nitric oxide.Both sodium nitroprusside and diazoxide produced in-
creases in coronary blood flow, and the effects of sodiumnitroprusside were increased, whereas those of diazoxide werenot modified, during the action of L-NAME. Sodium nitro-prusside has been used as a donor of exogenous nitric oxideand produces vasodilatation in a similar way to nitric oxide(Ignarro & Kadowitz, 1985). Enhanced vasorelaxant res-ponses to sodium nitroprusside and other nitrous compoundshave been found in the absence of a functional endotheliumor in the presence of L-arginine analogues (Shirasaki & Su,1985; Luischer et al., 1989; Forster et al., 1990; Gardiner etal., 1991; Moncada et al., 1991) and this has been related toan increased sensitivity of guanylate cyclase in vascular mus-culature to exogenous nitric oxide when production ofendogenous nitric oxide is reduced (Gardiner et al., 1991;Moncada et al., 1991). Therefore, it appears that the removalof endogenous nitric oxide in the vasculature could lead toincreases in sensitivity to vasodilators that act by stimulating
soluble guanylate cyclase (Gardiner et al., 1991; Moncada etal., 1991) and this phenomenon might also occur in thecoronary circulation as suggested from our study. Supersen-sitivity to exogenous nitric oxide also seems to develop in thesystemic circulation of the goat as indicated by the increasedhypotensive effects to sodium nitroprusside during treatmentwith L-NAME.
Reactive hyperaemia after short ischaemias of the myo-cardium is known to occur in the coronary circulation, andmetabolic and haemodynamic factors that contribute to thehyperaemic response have been explored (Feigl, 1983). How-ever, very little is yet known of the role of the endothelium inthe coronary hyperaemic response (Hayashi et al., 1988;Ueeda et al., 1990; Chu et al., 1991). Using guinea-pig heartLangendorff preparations, it was observed that N6-nitro-L-arginine (an inhibitor of nitric oxide synthesis) reduces cor-onary blood flow by 40%, but did not affect the peakhyperaemic response, although the duration of reactivehyperaemia to 30 s coronary occlusion was shortened (Ueedaet al., 1990). In dogs, endothelium denudation (Hayashi etal., 1988) or treatment with L-NMMA (Chu et al., 1991) didnot affect peak reactive hyperaemic flow, but reduced theflow-mediated vasodilatation after ischaemia following 20scoronary occlusion. Our observations show that in thepresence of L-NAME the hyperaemic responses (peakhyperaemic flow to control flow ratio, and repayment-to-debtratio) to 5, 10, and 20 s of coronary occlusion were increasedcompared to those found before administration of L-NAME.We have no immediate explanation for the differencesbetween our results and those of the above mentioned studies(Hayashi et al., 1988; Ueeda et al., 1990; Chu et al., 1991).The possibility that coronary vasodilator reserve to transientischaemias is augmented by an increased vascular tone dur-ing treatment with L-NAME may be excluded since an in-creased vasodilator reserve was not observed to diazoxide inthose conditions. Thus, inhibition of basal nitric oxide prod-uction with L-NAME augments the hyperaemic responsesafter short ischaemias in the coronary circulation as seenwith the vasodilator responses to sodium nitroprusside. Thisissue raises several questions that may deserve to be inves-tigated, for example (1) is the increased hyperaemic responseof the same nature as supersensitivity to nitrovasodilators,and (2) are the coronary vessels supersensitive to factorsreleased from the myocardium and/or the endothelium dur-ing ischaemia when endothelial nitric oxide production isreduced?
In conclusion, the results presented herein support the ideathat endogenous nitric oxide is involved in the regulation ofthe coronary circulation under basal conditions by producinga vasodilator tone. They also suggest that inhibition of basalendogenous nitric oxide production reduces the acetylcholine-induced coronary vasodilatation, induces supersensitivity ofcoronary vessels to nitrovasodilators, and enhances hyper-aemic responses after short ischaemias of the myocardium.These findings could be of relevance for understandingpathophysiology and for treatment of some coronary diseasesin which the endothelium is damaged.
The authors are grateful to Mrs H. Fernanez-Lomana and E.Martinez for technical assistance.
This work was supported, in part, by DGICYT (88/0058) and FIS(89/0639).
References
AISAKA, K., GROSS, S.S.,. GRIFFITH, O.W. & LEVI, R. (1989). NW-methylarginine, an inhibitor of endothelium-derived nitric oxidesynthesis, is a potent pressor agent in the guinea-pig: does nitricoxide regulate blood pressure in vitro? Biochem. Biophys. Res.Commun., 160, 881-886.
CHESTER, A.H., O'NEIL, G.S., TADJKARIMI, S., PALMER, R.M.J.,MONCADA, S. & YACOUB, M.H. (1990). The role of nitric oxidein mediating endothelium dependent relaxations in the humanepicardial coronary artery. Int. J. Cardiol., 29, 305-309.

NITRIC OXIDE AND CORONARY CIRCULATION 567
CHU, A., CHAMBERS, D.E., LIN, C.-C., KUEHL, W.D., PALMER,R.M.J., MONCADA, S. & COBB, F.R. (1991). Effects of inhibitionof nitric oxide formation on basal vasomotion and endothelium-dependent responses of the coronary arteries in awake dogs. J.Clin. Invest., 87, 1964-1968.
COX, D.A., HINTZE, T.H. & VATNER, S.F. (1983). Effects of acetyl-choline on large and small coronary arteries in conscious dogs. J.Pharmacol. Exp. Ther., 225, 764-769.
FEIGL, E.O. (1983). Coronary physiology. Physiol. Reg., 63, 1-205.FORSTER, C., MAIN, J.S. & ARMSTRONG, P.W. (1990). Endothelium
modulation of the effects of nitroglycerin on blood vessels fromdogs with pacing-induced heart failure. Br. J. Pharmacol., 101,109-114.
FURCHGOTT, R.E. & VANHOUTTE, P.M. (1989). Endothelium-derived relaxing and contracting factors. FASEB J., 3, 2007-2018.
FURCHGOTT, R.F. & ZAWADSKI, J.V. (1980). The obligatory role ofendothelial cells in the relaxation of arterial smooth muscle byacetylcholine. Nature, 228, 373-376.
GARDINER, S.M., COMPTON, A.M., BENNETT, T., PALMER, R.M.J. &MONCADA, S. (1990a). Control of regional blood flow byendothelium-derived nitric oxide. Hypertension, 15, 486-492.
GARDINER, S.M., COMPTON, A.M., KEMP, P.A. & BENNETT, T.(1990b). Regional and cardiac haemodynamic effects of NG-nitro-L-arginine methyl ester in conscious, Long Evans rats. Br. J.Pharmacol., 101, 625-631.
GARDINER, S.M., KEMP, P.A. & BENNETT, T. (1991). Effects ofN0-nitro-L-arginine methyl ester on vasodilator responses toacetylcholine. 5-N-ethylcarboxamidoadenosine or salbutamol inconscious rats. Br. J. Pharmacol., 103, 1725-1732.
HARRISON, D.G. (1989). From isolated vessels to the catheterizationlaboratory. Studies of endothelial function in the coronary cir-culation of humans. Circulation, 80, 703-706.
HAYASHI, Y., TOMOIKE, H., NAGASAWA, K., YAMADA, A., NISHI-JIMA, H., ADACHI, H. & NAKAMURA, M. (1988). Functional andanatomical recovery of endothelium after denudation of coronaryartery. Am. J. Physiol., 254, H1081 -H1090.
HODGSON, J.McB. & MARSHALL, J.J. (1989). Direct vasoconstrictionand endothelium-dependent vasodilation. Mechanisms of acetyl-choline effects on coronary flow and arterial diameter in patientswith nonstenotic coronary arteries. Circulation, 79, 1043-1051.
IGNARRO, L.J. & KADOWITZ, P.J. (1985). The pharmacological andphysiological role of cyclic GMP in vascular smooth musclerelaxation. Annu. Rev. Pharmacol. Toxicol., 25, 171-191.
IGNARRO, L.J., BUGA, G.M., WOOD, K.S., BYRNS, R.E. & CHAUD-HURI, G. (1987). Endothelium-derived relaxing factor producedand released from artery and vein is nitric oxide. Proc. Natl.Acad. Sci. U.S.A., 84, 9265-9269.
IGNARRO, L.J., HARBISON, R.G., WOOD, K.S. & KADOWITZ, P.J.(1986). Activation of purified soluble guanylate cyclase byendothelium-derived relaxing factor from intrapulmonary arteryand vein: stimulation by acetylcholine, bradykinin and arachi-donic acid. J. Pharmacol. Exp. Ther., 237, 893-900.
LOSCHER, T.F., RICHARD, V. & YANG, Z. (1989). Interactionbetween endothelium-derived nitric oxide and SIN-1 in humanand porcine blood vessels. J. Cardiovasc. Pharmacol., 14, Suppl.11, S76-S80.
MARCUS, M., WRIGHT, C., DOTY, D., EASTHAM, C., LAUGHLIN, D.,KRUMM, P., FASTENOW, C. & BRODY, M. (1981). Measurementsof coronary velocity and reactive hyperemia in the coronarycirculation of humans. Circ. Res., 49, 877-891.
MARTIN, W., FURCHGOTT, R.F., VILLANI, G.M. & JOTHIANAN-DAN, D. (1986). Depression of contractile responses in rat aortaby spontaneously released endothelium-derived relaxing factor. J.Pharmacol. Exp. Ther., 237, 529-538.
MONCADA, S. & HIGGS, E.A. (1990). Nitric Oxide from L-arginine: ABioregulatory System. Amsterdam: Excepta Medica.
MONCADA, S., REES, D.D., SCHULZ, R. & PALMER, R.M.J. (1991).Development and mechanism of a specific supersensitivity tonitrovasodilators after inhibition of vascular nitric oxide synthesisin vivo. Proc. Nati. Acad. Sci. U.S.A., 88, 2166-2170.
MOORE, P.K., AL-SWAYEH, O.A., CHONG, N.W.S., EVANS, R.A. &GIBSON, A. (1990). L-N0-nitro arginine (L-NOARG), a novelL-arginine-reversible inhibitor of endothelium-dependentvasodilatation in vitro. Br. J. Pharmacol., 99, 408-412.
MYERS, P.R., BANITT, P.F., GUERRA, F. Jr. & HARRISON, G. (1989).Characteristics of canine coronary resistance arteries: importanceof endothelium. Am. J. Physiol., 257, H603-H610.
PALMER, R.M.J., FERRIGE, A.G. & MONCADA, S. (1987). Nitricoxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature, 327, 524-526.
PALMER, R.M.J., ASHTON, D.S. & MONCADA, S. (1988). Vascularendothelial cells synthesize nitric oxide from L-arginine. Nature,333, 664-666.
RAPOPORT, R.M. & MURAD, F. (1983). Agonist-induced endotheliumdependent relaxation in rat thoraci aorta may be mediatedthrough cGMP. Circ. Res., 52, 352-357.
REES, D.D., PALMER, R.M.J., HODSON, H.F. & MONCADA, S.(1989a). A specific inhibitor of nitric oxide formation from L-arginine attenuates endothelium-dependent relaxation. Br. J.Pharmacol., 96, 418-424.
REES, D.D., PALMER, R.M.J. & MONCADA, S. (1989b). Role ofendothelium-derived nitric oxide in the regulation of blood pres-sure. Proc. Natl. Acad. Sci. U.S.A., 86, 3375-3378.
REES, D.D., PALMER, R.M.J., SCHULZ, R., HODSON, H.R. & MON-CADA, S. (1990). Characterization of three inhibitors of endothelialnitric oxide synthase in vitro and in vivo. Br. J. Pharmacol., 101,746-752.
REID, J.V.O., ITO, B.R., HUANG, A.H., BUFFINGTON, C.W. & FEIGL,E.O. (1985). Parasympathetic control of transmural coronary bloodflow in dogs. Am. J. Physiol., 249, H337-H343.
RUBANYI, G.M., ROMERO, J.C. & VANHOUTTE, P.M. (1986). Flow-induced release of endothelium-derived relaxing factor. Am. J.Physiol., 250, H1145-H1149.
SHIRASAKI, Y. & SU, C. (1985). Endothelium removal augmentsvasodilatation by sodium nitroprusside and sodium nitrite. Eur. J.Pharmacol., 114, 93-96.
UEEDA, M., ARROYO, L.H. & OLSSON, R.A. (1990). Role of nitricoxide (N=O) in coronary reactive hyperemia (RH) and theactive hyperemic response to pacing (AG). (Abstract). ThePhysiologist, 33, A-78.
VALLANCE, P., COLLIER, J. & MONCADA, S. (1989). Effects ofendothelium-derived nitric oxide on peripheral arteriolar tone inman. Lancet, ii, 997-1000.
VAN GELDEREN, E.M., HEILIGERS, J.P.C. & SAXENA, P.R. (1991).Haemodynamic changes and acetylcholine-induced hypotensiveresponses after N0-nitro-L-arginine methyl ester in rats and cats.Br. J. Pharmacol., 103, 1899-1904.
WIDOPP, R.E., GARDINER, S.M., KEMP, P.A. & BENNETT, T. (1992).The influence of atropine and atenolol on the cardiachaemodynamic effects of NG-nitro-L-arginine methyl ester in con-scious, Long Evans rats. Br. J. Pharmacol., 105, 653-656.
(Received February 7, 1992Revised March 2, 1992
Accepted March 4, 1992)