characterization of the discontinuities in rice tungro bacilliform virus
TRANSCRIPT

Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Wed, 28 Mar 2018 15:28:42
Journal of General Virology (1992), 73, 1297-1301. Printed in Great Britain 1297
Characterization of the discontinuities in rice tungro bacilliform virus DNA
Yiming Bao and Roger Hull*
Department o f Virus Research, John Innes Institute, John Innes Centre for Plant Science Research, Colney Lane, Norwich NR4 7UH, U.K.
The dsDNA of rice tungro bacilliform virus (RTBV) has two discontinuities, one on each strand, each in a specific position as found in other pararetroviruses. The 5' end of discontinuity 1 was mapped to nucleotide 1 of the published RTBV DNA sequence which suggests that tRNA~ ~t serves as a primer for negative strand DNA synthesis. This 5' terminus contains up to
two ribonucleotides and the 3' terminus overlaps it by five to 25 nucleotides. The discontinuity 2 (D2) did not map to a purine-rich region as has been found in other similar viruses. Both the 5' and 3' termini of D2 were heterogeneous in position giving structures varying from a gap of 10 nucleotides to an overlap of 103 nucleotides.
Rice tungro bacilliform virus (RTBV) is one of the two viruses in the complex causing rice tungro, the most severe viral disease of rice in South East Asia (Hibino et al., 1978). RTBV has a circular dsDNA genome, and from the sequence of an 8002 bp viral genome clone, Hay et al. (1991) suggested that it is a plant pararetrovirus (Hull & Will, 1989; Temin, 1989) similar in its structure and DNA replication to caulimoviruses (see Covey, 1985; Mason et al., 1987). It is classified with the badnaviruses, along with Commelina yellow mottle virus (CoYMV) (Medberry et al., 1990) and cacao swollen shoot virus (Lot et al., 1991). These plant pararetro- viruses have two to three discontinuities located at specific sites, one in the negative strand and one or more in the other strand. These discontinuities are believed to be priming sites for DNA replication (Covey, 1985; Mason et al., 1987; Medberry et al., 1990). The sequence around the non-coding negative strand discontinuity, taken as the zero point on the map of these viral genomes, has complementarity to the 3' end of tRNAi M~t whereas the positive strand discontinuities are rich in purines (Franck et al., 1980; Hull et al., 1986; Richins et al., 1987; Verver et al., 1987; Medberry et al., 1990; Luo et al., 1990). The structure of the DNA around the three discontinuities of cauliflower mosaic virus (CaMV) has been shown to comprise a fixed 5' terminus with some ribonucleotides attached. The 3' terminus overlapped the 5' terminus by a variable amount of five to 35 nucleotides (Richards et al., 1981). In strand separation experiments, Jones et al. (1991) showed that there are two disconti- nuities in the RTBV genome, one (D1) in the negative strand at the zero point of the restriction map and the
other (D2) in the positive strand at around 0.58 map units. In this paper, we describe the structure of these two discontinuities.
RTBV (Philippines isolate) was propagated in rice (Oryza sativa) cv. TN1 after agroinoculation (Dasgupta et al., 1991). Virus particles and virion DNA were isolated as described by Jones et al. (1991). Cloned RTBV DNA pJIIS2 (Hay et al., 1991) was sequenced using [~-35S]dATP and a Sequenase Kit (United States Biochemical) as recommended by the manufacturer, or by a simplified method (Hsiao, 1991). Oligonucleotides used for DNA sequencing and primer extension experi- ments were: V36 (5774 GGCTTGTATCCATCC 5760), V104 (4383 ATATGAATGCAAATGAGG 4366), V105 (5192 TGCGGCTGAAGACTAATC 5175), V285 (7911 AGTCGACGGATGAGGTCA 7929), V305 (110 AATTCAAGTTTTTCGTAA 127), V361 (7937 GCA- CTAGTAGGTAACTAA 7954) and V403 (4111 GAC- GAAACTTTTGGAATA 4128) (the nucleotide num- bers refer to the sequence of Hay et al., 1991) (Fig. 1). Restriction fragments of virion DNA were recovered from low melting point agarose, or by the liquid nitrogen freeze-squeeze method (Gaastra et al., 1984). The recovered fragments were dephosphorylated and [y-32p]ATP 5' end-labelled using polynucleotide kinase as recommended by the manufacturer (Boehringer Mannheim).
Oligonucleotides V361 and V 104 were used for primer extension to map the 5' ends of DI and D2. These primers use the positive and negative strand of RTBV DNA as the template, respectively, and stop extending at the discontinuity position. Primer extension was
0001-0711 © 1992 SGM
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Wed, 28 Mar 2018 15:28:42
1298 Short communication
BamHI
HinclI EcoRI
R T B V
8002 bp
÷ 5'
5' I I ~ EcoRI
HinclI
Fig. 1. The position of oligonucleotides (V36, V104, V105, V285,
V305, V361 and V403) and restriction endonuclease cutting sites used to map the discontinuities (Q) of RTBV virion DNA. The orientations of the oligonucleotides and the plus and minus strand of RTBV are indicated by arrows.
performed as described by Medberry et al. (1990) except that an excess of primers was used and the virion DNA was denatured by heating in a boiling water bath for 5 min and then cooling immediately in ice. Plasmid pJIIS2 was sequenced using the same primer as in primer extension and the sequencing reactions were run in parallel to the primer extension samples on 6 ~ polyacryl- amide sequencing gels.
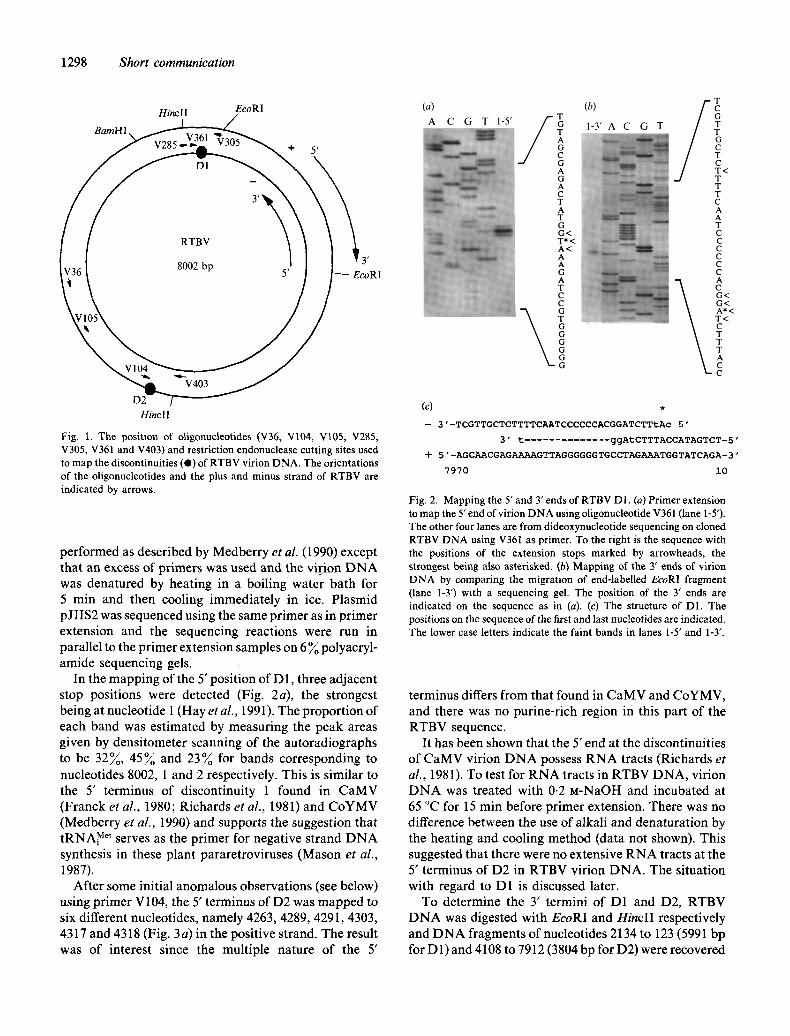
In the mapping of the 5' position of D1, three adjacent stop positions were detected (Fig. 2a), the strongest being at nucleotide 1 (Hay et al., 1991). The proportion of each band was estimated by measuring the peak areas given by densitometer scanning of the autoradiographs to be 32~, 4 5 ~ and 23~o for bands corresponding to nucleotides 8002, 1 and 2 respectively. This is similar to the 5' terminus of discontinuity 1 found in CaMV (Franck et al., 1980; Richards et al., 1981) and CoYMV (Medberry et al., 1990) and supports the suggestion that tRNAi Met serves as the primer for negative strand DNA synthesis in these plant pararetroviruses (Mason et al., 1987).
After some initial anomalous observations (see below) using primer V104, the 5' terminus of D2 was mapped to six different nucleotides, namely 4263, 4289, 4291, 4303, 4317 and 4318 (Fig. 3a) in the positive strand. The result was of interest since the multiple nature of the 5'
(a)
A C G T 1-5' / (b)
T T C 1 - 3 ' A C
A G C 13 A G A C T A T G G< T* < A< A A G A T C C
T G G G G G G
G T
T C G T T G C T C T < T T T C A A T C C C C C C
C G< (3<
T < C T T T A e c
( 3 .
-- 3'-TCGTTGCTCTTTTCAATCCCCCCACGGATCTTtAc 5'
3' t .............. ggAtCTTTACCATAGTCT-5'
+ 5"-AGCAACGAGAAAAGTTAGGGGGGTGCCTAGAAATGGTATCAGA-3'
7970 i0
Fig. 2. Mapping the 5' and 3' ends of RTBV D1. (a) Primer extension to map the 5' end of virion DNA using oligonucleotide V361 (lane 1-5').
The other four lanes are from dideoxynucleotide sequencing on cloned R T B V DNA using V361 as primer. To the right is the sequence with the positions of the extension stops marked by arrowheads, the strongest being also asterisked. (b) Mapping of the 3' ends of virion DNA by comparing the migration of end-labelled EcoRI fragment (lane 1-3') with a sequencing gel. The position of the 3' ends are indicated on the sequence as in (a). (c) The structure of D1. The positions on the sequence of the first and last nucleotides are indicated. The lower case letters indicate the faint bands in lanes 1-5' and 1-3".
terminus differs from that found in CaMV and CoYMV, and there was no purine-rich region in this part of the RTBV sequence.
It has been shown that the 5' end at the discontinuities of CaMV virion DNA possess RNA tracts (Richards et al., 1981). To test for RNA tracts in RTBV DNA, virion DNA was treated with 0.2 M-NaOH and incubated at 65 °C for 15 min before primer extension. There was no difference between the use of alkali and denaturation by the heating and cooling method (data not shown). This suggested that there were no extensive RNA tracts at the 5' terminus of D2 in RTBV virion DNA. The situation with regard to D1 is discussed later.
To determine the 3' termini of D1 and D2, RTBV DNA was digested with EcoRI and Hinc l I respectively and DNA fragments of nucleotides 2134 to 123 (5991 bp for D1) and 4108 to 7912 (3804 bp for D2) were recovered

Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Wed, 28 Mar 2018 15:28:42
Short communication 1299
(a) T G C C C T< T A T A T G G G T T G T C G G T C T G T A C T T G A< A T< T T A G T C T C A A A C* < G C G C T T T C G T A C C A * < C< C T O
x_g
(c) + 3 " -ACTCCTTAAAACAATATCCAAAGTCCTTACAAACTTTTATTACCGCCCGCA
3" t ............. a .......... AAA-TTt t a ...... c ..... 5" -TGAGGAATTTTGTTATAGGTTTCAGGAATGTTTGAAAATAATGGCGGGCGT
4370 4320
GgT ............. G ........... a-t ......................... A 5" ............. CGCGTTTGAGACTAAATTCAAGTACAGACCGACAACCCATATAAGGG - 5" CCACCATGCTTTCGCGCAAACTCTGATTTAAGTTCATGTCTGGCTGTTGGGTATATTCCC- 3 '
4 3 1 9 4 2 6 0
Fig. 3. Mapping the 5' and 3' ends of RTBV D2 as in Fig. 2. (a) Primer extension to map the 5' end using oligonucleotide V104 (lane 2-5'). (b) End-labelled HinclI fragment (lane 2-3') used to map the 3' ends. (c) The structure of D2.
from agarose gels. These fragments were dephosphoryl- ated and 5' end-labelled in the presence of 25 ~ DMSO according to Franck et al. (1980). The samples were incubated in a boiling water bath for 2 min just before loading onto a 6 ~ acrylamide gel. To determine the size of the end-labelled fragments, pJIIS2 was sequenced using oligonucleotides V305 and V403 as primers. These primers were designed to hybridize to the 5' end of the restriction fragment containing the 3' end of each discontinuity.
The 3' termini of D1 and D2 were mapped to nucleotides 7980, 7995 to 7998 in the negative strand and 4306, 4325, 4332 to 4336, 4338 to 4340, 4351 and 4365 in the positive strand respectively (Fig. 2b and 3b). (There might be one nucleotide change in the above data because the fragments from RTBV were end-labelled whereas the primers for the sequencing ladder were not.) This showed an overlap of five to 25 nucleotides between the 5' and 3' ends of D1. Thus, the structure of D1 closely resembles that of the negative strand discontinuity (G 1) of CaMV (Richards et al., 1981). At D2 the separation of the 5' and 3' ends varied from a gap of 10 nucleotides to an overlap of 103 nucleotides. The structure differs from the positive strand priming site of CaMV in that there is a gap in, at least, a portion of the molecules. The pararetrovirus hepatitis B virus (HBV) has a gap of up to 700 nucleotides in the positive strand (Delius et al., 1983).
Initial experiments on priming crude virion DNA templates with the oligonucleotide V36 indicated that the 5' end of D2 mapped to nucleotides 5681 to 5683, which is close to the purine-rich region identified by Hay et al. (1991). However, no 3' terminus of RTBV DNA could be found in this region. When further oligonucleo- tides hybridizing to different positions were used for primer extension, they all gave extension stop fragments corresponding to 91 to 93 nucleotides (Fig. 4). Even when no primer was added, the 91 to 93 nucleotide extension stop fragments were found. This suggested that the crude DNA preparation contained an endogenous primer. To test this possibility, virion DNA was further purified by agarose gel electrophoresis. It did not prove possible to recover intact virion DNA from either low melting point agarose gel or ordinary agarose by the liquid nitrogen freeze-squeeze method. Virion DNA was therefore linearized by BamHI digestion and recovered from the gel by the liquid nitrogen freeze-squeeze method. When this DNA was used as the template with primers V36, V104 and without primers, the 91 to 93 nucleotide fragments were not found (data not shown) indicating that there was an endogenous primer. If the crude DNA preparation was treated by alkali, the non-specific fragment was one to six nucleotides shorter indicating that the endogenous primer contained terminal ribo- nucleotides (data not shown). The nature of this endo- genous primer has yet to be determined.
As noted above, D1 has the same basic structure as GI of CaMV. The majority of the 5' ends were nucleotide 1 of the sequence which corresponds to the 3' nucleotide of the tRNA primer, a situation similar to that in CoYMV. If the molecule which actually primes the synthesis of the negative strand was the authentic tRNA with the 3'- terminal CCA, we would expect the 5' nucleotide of the negative strand DNA to be at the - 1 or 8002 position.

Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Wed, 28 Mar 2018 15:28:42
1300 Shor t communicat ion
A C G T 1 2 3 4 5
T* < A*< A A G A T C C G T G G G G G G A T T G A A A A G A
Fig. 4. Primer extension using a crude virion DNA preparation as the template and different oligonucleotides as the primer. Lanes I to 5 represent oligonucleotides V285, V36, V104, V105 and no oligonucleo- tide added respectively. The nucleotide coordinate of the asterisked T is 1 in the RTBV sequence.
Only a small portion of negative strand molecules appeared to prime at this site. There are two possible answers to this problem. Either the 3' end of t R N A was incomplete or a specific RNase H cleavage leaves the terminal rCA from the t R N A attached to the D N A strand as suggested by Whitcomb et al. (1990) for human immunodeficiency virus type 1. CCA at the 3' terminus of eukaryotic t R N A is added as individual nucleotides by t R N A nucleotidyltransferase to t R N A precursor (Deutscher, 1983). Thus, there might be some t R N A molecules having CC or even C at their 3" end, and such t R N A might be selected to serve as primers for D N A synthesis. When alkaline t reatment before pr imer extension was performed, the percentages for pr imer extension stop bands corresponding to nucleotides 8002, 1 and 2 are 22~ , 50~o and 2 8 ~ respectively. Thus, there are some changes in proportions of the three stop sites
when compared with heat denaturation, which indicates that some of the 5' termini contained one or two ribonucleotides. This suggests that both possibilities about the 3' end of t R N A existed. The 3' end of D1 overlaps the 5' end indicating that the reverse transcrip- tase strand displaces the 5' end by a certain amount.
The structure of D2 displays various unusual features when compared with the positive strand pr iming sites of other retroviruses and pararetroviruses. With the retro- viruses and the pararetroviruses, the priming site is specific and, except for HBV, is associated with a purine- rich region (Mason et al., 1987; Franck et al., 1980; Hull et al., 1986; Richins et al., 1987; Verver et al., 1987; Medberry et al., 1990; Luo et al., 1990). The 5' end of RTBV D2 ranged across 55 nucleotides though some were preferred over others. There was no obvious purine- rich region, and the 5' end did not fine map to a purine- rich region which had been previously identified (Hay et al., 1991). It is generally considered that the priming molecule for positive strand synthesis is R N A remaining after digestion of the reverse transcription template by viral RNase H (Varmus et al., 1982; Mason et al., 1987). Some specificity between the individual viral enzymes and the priming sequence has been suggested (Luo et al., 1990). I f this is so, the specificity of RTBV RNase H may differ from other retroviral and plant pararetroviral enzymes. Comparison of polypeptide sequences encod- ing the core consensus sequence of RNase H of RTBV and other plant pararetroviruses and retroviruses (Hay et al., 1991) revealed four amino acids in RTBV which differed from those common to similar viruses. This may account for RTBV RNase H having a different specific activity. Another possibility is that RTBV positive strand synthesis is not primed by R N A fragments left behind by RNase H, but by host R N A sequences.
We thank Mr M. Harvey for synthesizing oligonucleotides and for preparing Fig. 1, Mr A. Lucy for synthesizing oligonucleotides, and Professor J. W. Davies, Drs J. Hay, I. Dasgupta and G. Dahal, Mr G. Lee, S. Zhang and Z. Fan for helpful comments during the preparation of this manuscript. Y.B. was funded from a grant from the Rockefeller Foundation. Tungro is held under MAFF Plant Health licence no. PHF48A/116.
References
COVEY, S. N. (1985). Organization and expression of the cauliflower mosaic virus genome. In Molecular Plant Virology, vol. 2, pp. 121- 159. Edited by J. W. Davies. Boca Raton: CRC Press.
DAS6UPTA, I., HULL, R., EASTOP, S., PO66I-POLLINI, C., BLAKEBROUGH, M., BOULTON, M. I. & DAVIES, J. W. (1991). Rice tungro bacilliform virus DNA independently infects rice after Agro- bacterium-mediated transfer. Journal of General Virology '72, 1215-1221.
DELIUS, H., GouGI-I, N. M., CAMERON, C. H. & MURRAY, K. (1983). Structure of the hepatitis B virus genome. Journal of Virology 47, 337-343.

Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Wed, 28 Mar 2018 15:28:42
Shor t communicat ion 1301
DELrI~CHER, M. P. (1983). tRNA nucleotidyltransferase and the -C-C-A terminus of transfer RNA. In Enzymes of Nucleic Acid Synthesis and Modification, vol. 2, pp. 159-183. Edited by S. T. Jacob. Boca Raton: CRC Press.
FRANCK, A., GUILLEY, H., JONARD, G., RICHARDS, K. & HIRTH, L. (1980). Nucleotide sequence of cauliflower mosaic virus DNA. Cell 21, 285-294.
GAASTRA, W. 8/; JORGENSEN, P. L. (1984). The extraction and isolation of DNA from gels. In Methods in Molecular Biology, vol. 2, pp. 67-76. Edited by J. M. Walker. Clifton: Humana Press.
HAY, J. M., JONES, M. C., BLAKEBROUGH, M. L., DASGUPTA, I., DAVIES, J. W. & HULL, R. (1991). An analysis of the sequence of an infectious clone of rice tungro bacilliform virus, a plant pararetro- virus. Nucleic Acids Research 19, 2615-2621.
HIBINO, H., ROECHAN, M. & SUDARISMAN, S. (1978). Association of two types of virus particles with Penyakit Habang (tungro disease) of rice in Indonesia. Phytopathology 68, 1412-1416.
HSIAO, K. (1991). A fast and simple procedure for sequencing double stranded DNA with Sequenase. Nucleic Acid Research 19, 2787.
HULL, R. & WILL, H. (1989). Molecular biology of viral and nonviral retroelements. Trends in Genetics 5, 357-359.
HULL, R., SADLER, J. & LOI'~STAFF, M. (1986). The sequence of carnation etched ring virus DNA: comparison with cauliflower mosaic virus and retroviruses. EMBO Journal 5, 3083-3090.
JONES, M. C., GOUGH, K., DASGUPTA, 1., SUBBA RAO, B. L., CLIFFE, J., Qu, R., SHEN, P., KANIEWSKA, M., BLAKEBROUGH, M., DAVIES, J. W., BEACHY, R. N. & HULL, R. (1991). Rice tungro disease is caused by an RNA and a DNA virus. JournalofGeneral Virology 72, 757-761.
LOT, H., DJIEKPOR, E. & JACQUEMOND, M. (1991). Characterization of the genome of cacao swollen shoot virus. Journal of General Virology 72, 1735-1739.
Luo, G., SHARMEEN, U & TAYLOR, J. (1990). Specificities involved in the initiation of retroviral plus-strand DNA. Journal of Virology 64, 592-597.
MASON, W. S., TAYLOR, J. M. & HULL, R. (1987). Retroid virus genome replication. Advances in Virus Research 32, 35-96.
MEDBERRY, S. L., LOCKHART, B. E. L. & OLSZEWSKI, N. E. (1990). Properties of Commelina yellow mottle virus's complete DNA sequence, genomic discontinuities and transcript suggest that it is a pararetrovirus. Nucleic Acid Research 18, 5505-5513.
RICHARDS, K. E., GUILLEY, H. & JONARD, G. (1981). Further characterization of the discontinuities in cauliflower mosaic virus DNA. FEBS Letters 134, 67-70.
RICHINS, R. D., SCHOLTHOF, H. B. • SHEPHERD, R. J. (1987). Sequence of figwort mosaic virus DNA (caulimovirus group). Nucleic Acids Research 15, 8451-8466.
TEMIN, H. M. (1989). Retrons in bacteria. Nature, London 339, 254-255. VARMUS, H. & SWANSTROM, R. (1982). Replication of retrovirus. In
RNA Tumor Viruses, 2nd edn., pp. 369-512. Edited by R. Weiss, N. Teich, H. Varmus & J. Coffin. New York: Cold Spring Harbor Laboratory.
VERVER, J., SCHIJNS, P., HIBI, T. & GOLDBACH, R. (1987). Characteriz- ation of the genome of soybean chlorotic mottle virus. Journal of General Virology 68, 159-167.
WHITCOMB, J. M., KUMAR, R. & HUGHES, S. H. (1990). Sequence of circle junction of human immunodeficiency virus type 1: impli- cations for reverse transcription and integration. Journal of Virology 64, 4903--4906.
(Received 29 October 1991; Accepted 2 January 1992)