chapter 4 associated digestive glands
TRANSCRIPT

Part II – Chapter 4- 1
`
• Extramural glands of the digestive system include the major salivary glands
associated with the oral cavity (parotid, submandibular, and sublingual glands),
the pancreas, and the liver and gallbladder.
• Each of these glands has numerous functions aiding the digestive process, and their
secretory products are delivered to the lumen of the alimentary tract by a system of
ducts.
• Saliva produced by salivary glands facilitates the process of tasting food, initiate its
digestion, and permit its deglutition (swallowing). These glands also protect the
body by secreting the antibacterial agents lysozyme and lactoferrin as well as the
secretory immunoglobulin IgA.
• The pancreas manufactures a bicarbonate-rich fluid that buffers the acid chyme and
produces enzymes necessary for the digestion of fats, proteins, and carbohydrates.
The exocrine secretions of the pancreas are released into the lumen of the
duodenum as necessary.
• In addition, the pancreas synthesizes and releases endocrine hormones, including
insulin, glucagon, somatostatin, gastrin, and pancreatic polypeptide.
• Bile, the exocrine secretion of the liver, is required for proper absorption of lipids,
whereas many of the liver's endocrine functions are essential for life. These include
metabolism of proteins, lipids, and carbohydrates; synthesis of blood proteins and
coagulation factors; manufacture of vitamins; and detoxification of blood-borne
toxins.
• The gallbladder concentrates bile and stores it until its release into the lumen of the
duodenum.
Chapter 4 ASSOCIATED DIGESTIVE GLANDS

Part II – Chapter 4- 2
Salivary Glands MAJOR SALIVARY GLANDS
• There are three pairs of major salivary glands: parotid, sublingual, and
submandibular.
• The major salivary glands are the paired parotid, sublingual, and submandibular
glands.
• They are branched tubuloalveolar glands whose connective tissue capsule provides
septa that subdivide the glands into lobes and lobules. Individual acini are also
invested by thin connective tissue elements.
• The vascular and neural components of the glands reach the secretory units via the
connective tissue framework.
Anatomy of Salivary Glands
• Each of the major salivary glands has a secretory and a duct portion
• Salivon is the functional unit of a salivary gland, composed of the acinus,
intercalated duct, and striated duct.
Secretory Portions
• The secretory portions of salivary glands are composed of serous and/or mucous
secretory cells arranged in acini (alveoli) or tubules that are surrounded by
myoepithelial cells.
• The secretory portions, arranged in tubules and acini, are composed of three types
of cells:
1. Serous cells
• The plasmalemma basal to the tight junctions forms many processes that
interdigitate with those of neighboring cells.
• These cells are pyramids in shape and have single, round, basally located
nuclei, a well-developed rough endoplasmic reticulum (RER) and Golgi
complex, numerous basal mitochondria, and abundant apically situated
secretory granules (proteins and a considerable amount of polysaccharides).

Part II – Chapter 4- 3
• The secretory granules are rich in ptyalin (salivary amylase), some
kallikrein, lactoferrin, and lysozyme.
• The basal aspects of the lateral cell membranes form tight junctions with each
other.
• Apical to the tight junctions, intercellular canaliculi communicate with the
lumen.
2. Mucous cells
• They are similar in shape to the serous cells.
• Their nuclei are also basally located but are flattened instead of being round.
The organelle population of these cells differs from that of the serous cells in
that mucous secretory cells have fewer mitochondria, a less extensive RER,
and a considerably greater Golgi apparatus, indicative of the greater
carbohydrate component of their secretory product.
• Abundant secretory granules occupy the apical region of the cytoplasm.
• The intercellular canaliculi and processes of the basal cell membranes are
much less extensive than those of serous cells.
3. Myoepithelial cells
• They are called basket cells, share the basal laminae of the acinar cells.
• They have a cell body that houses the nucleus and several long processes that
envelop the secretory acinus and intercalated ducts.
• The cell body houses a small complement of organelles in addition to the
nucleus and makes hemidesmosomal attachments with the basal lamina.
• The cytoplasmic processes, which form desmosomal contacts with the acinar
and duct cells, are rich in actin and myosin; in electron micrographs these
processes resemble smooth muscle cells.
• As the processes of myoepithelial cells contract, they press on the acinus,
facilitating release of the secretory product into the duct of the gland.

Part II – Chapter 4- 4

Part II – Chapter 4- 5
Duct Portions • The ducts of major salivary glands are highly branched and range from very small-
intercalated ducts to very large principal (terminal) ducts. • The smallest branches of the system of ducts are the intercalated ducts, to which the
secretory acini (and tubules) are attached. • These small ducts are composed of a single layer of small cuboidal cells and
possess some myoepithelial cells. • Several intercalated ducts merge with each other to form striated ducts, composed
of a single layer of cuboidal to low columnar cells. • The basolateral membranes of these cells are highly folded, subdividing the
cytoplasm into longitudinal compartments that are occupied by elongated mitochondria.
• The basolateral cell membranes of these cells have sodium adenosine triphosphatase (Na+-ATPase) that pumps sodium out of the cell into the connective tissue, thus conserving these ions and reducing the tonicity of saliva.
• Striated ducts join with each other, forming intralobular ducts of increasing caliber, which are surrounded by more abundant connective tissue elements.
• Ducts arising from lobules unite to form interlobular ducts, which in turn form intralobar and interlobar ducts.
• The terminal (principal) duct of the gland delivers saliva into the oral cavity. Properties of Individual Salivary Glands Parotid Gland • Although physically the largest of the salivary glands, the parotid gland produces
only about 30% of the total salivary output. • The saliva produced by the parotid gland has high levels of the enzyme salivary
amylase (ptyalin) and secretory IgA. • Salivary amylase is responsible for digestion of most of the starch in food, and this
digestion continues in the stomach until the acidic chyme inactivates the enzyme. • Secretory IgA inactivates antigens located in the oral cavity. • The connective tissue capsule of the parotid gland is well developed and forms
numerous septa, which subdivide the gland into lobes and lobules. • By the 40th year of age, the gland becomes invaded by adipose tissue, which
spreads from the connective tissue into the glandular parenchyma.

Part II – Chapter 4- 6
Sublingual Gland • The sublingual gland is very small, is composed mostly of mucous acini with serous
demilunes, and produces mixed saliva. • The sublingual gland, the smallest of the three major salivary glands, is almond-
shaped, weighs only 2 to 3 g, and produces only about 5% of the total salivary output.
• The gland is composed of mucous tubular secretory units, many of which are capped by a small cluster of serous cells, known as serous demilunes .
• These serous cells have been shown to secrete lysozyme. • The sublingual gland produces a mixed, but mostly mucous, saliva. • The intercellular canaliculi are well developed between the mucous cells of the
secretory units. • Electron micrographs of the cells of the serous demilunes display apical
accumulations of secretory vesicles. • The sublingual gland has a scant connective tissue capsule, and its duct system does
not form a terminal duct. • Instead, several ducts open into the floor of the mouth and into the duct of the
submandibular gland. Submandibular Gland • The submandibular gland produces 60% of the total salivary output; although it
manufactures a mixed saliva, the major portion is serous. • About 90% of the acini produce serous saliva, whereas the remainder of the acini
manufacture a mucous saliva. • Electron micrographs of the apical aspects of the serous cells of this gland display
electron-dense secretory products, with a denser core, within membrane-bound secretory granules.
• The number of serous demilunes is limited. • The striated ducts of the submandibular gland are much longer than those of the
parotid or sublingual glands. • The connective tissue capsule of the submandibular gland is extensive and forms
abundant septa, which subdivide the gland into lobes and lobules. • Fatty infiltration of the connective tissue elements into the parenchyma is evident
by midlife.
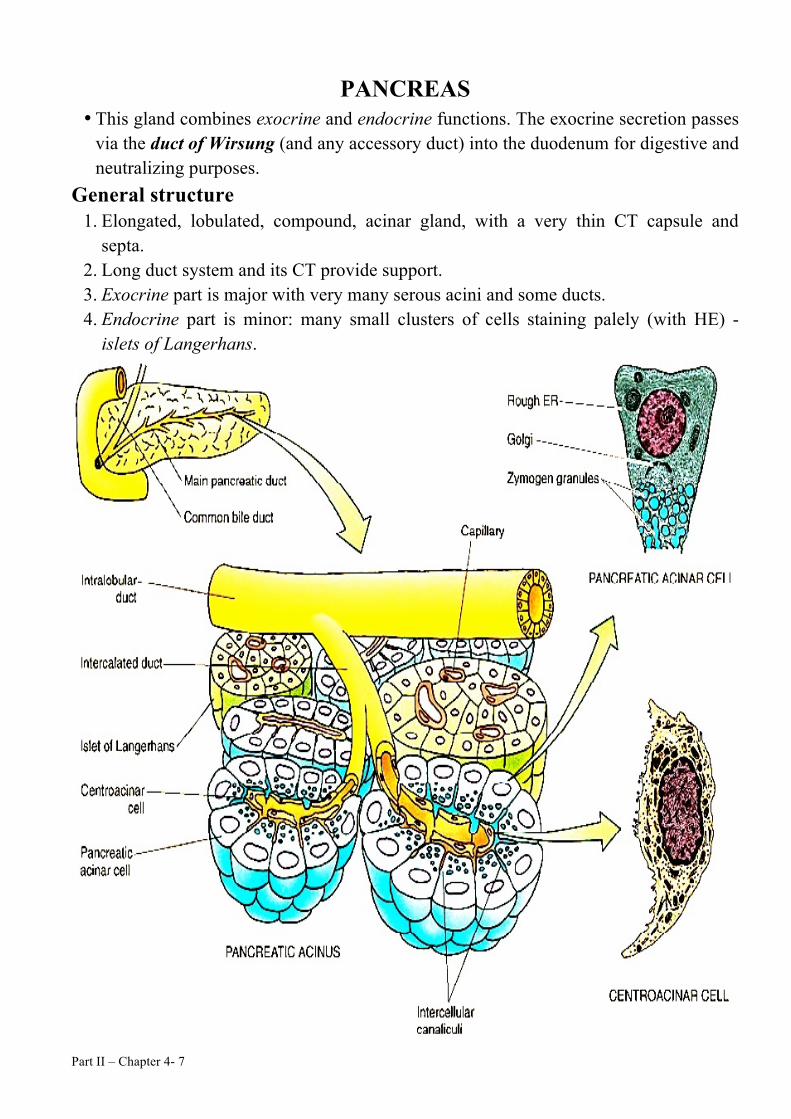
Part II – Chapter 4- 7
PANCREAS • This gland combines exocrine and endocrine functions. The exocrine secretion passes
via the duct of Wirsung (and any accessory duct) into the duodenum for digestive and neutralizing purposes.
General structure 1. Elongated, lobulated, compound, acinar gland, with a very thin CT capsule and
septa. 2. Long duct system and its CT provide support. 3. Exocrine part is major with very many serous acini and some ducts. 4. Endocrine part is minor: many small clusters of cells staining palely (with HE) -
islets of Langerhans.

Part II – Chapter 4- 8
Exocrine pancreas Acinar structure
• Forty to 50 acinar cells form a round to oval acinus whose lumen is occupied by three or four centroacinar cells, the beginning of the duct system of the pancreas. The presence of centroacinar cells in the center of the acinus is a distinguishing characteristic of this gland.
• A pale duct cell (or a pair) may be seen intruded into the center of the acinus as a centroacinar cell
• The acinar cells are rich in basal granular ER (deeply basophil); have a prominent supranuclear Golgi complex and apical zymogen granules (precursors of several digestive enzymes).
• The acinar cells synthesis alkaline secretions, which counter the gastric fluid's acidity, thereby activating pancreatic pro-enzymes for digestion.
• The release of alkaline and enzymatic secretions is under the hormonal control of secretin, and cholecystokinin/CCK, respectively.
Ducts
• The duct system of the pancreas begins within the center of the acinus with the
terminus of the intercalated ducts, composed of pale, low cuboidal centroacinar
cells.
• Centroacinar cells and intercalated ducts both have receptors on their basal
plasmalemma for the hormone secretin and possibly for acetylcholine, released by
postganglionic parasympathetic fibers.
• Intercalated ducts join each other to form larger intralobular ducts, several of which
converge to form interlobular ducts.
• These ducts are surrounded by a thick connective tissue and deliver their contents into
the main pancreatic duct, which joins the common bile duct before opening in the
duodenum at the papilla of Vater.
Note • Ducts are less in number than in the parotid gland, and are accompanied by less
connective tissue than in the salivary glands, which are exposed to masticatory forces.

Part II – Chapter 4- 9
Endocrine pancreas Islet structure and functions • The endocrine pancreas is composed of spherical aggregates of cells, known as islets
of Langerhans, which are scattered among the acini.Each islet of Langerhans is a richly vascularized spherical mass of approximately 3000 cells.
• The approximately 1 million islets distributed throughout the human pancreas constitute the endocrine pancreas. A somewhat greater number of islets are present in the tail than in the remaining regions.
• Each islet is surrounded by reticular fibers, which also enter the substance of the islet to encircle the network of capillaries that pervade it.
Cells Composing the Islets of Langerhans • Five types of cells compose the parenchyma of each islet of Langerhans:
Cell Total Location EM Granules Hormone Function
Alpha (α) 20% Periphery of the Islet
250 nm in diameter; dense core
granule with narrow
electron-lucent halo
Glucagon Increases blood glucose
Beta (β) 70%
Scattered throughout islet (but
concentrated in
center)
300 nm in diameter; dense core
granule surrounded by wide electron-
lucent halo
Insulin Decreases blood glucose
Delta (δ) (D& D1) 5%
Scattered throughout
islet
350 nm in diameter; electron-
lucent homogeneous
granule
(D) Somatostatin
(D1)
Vasoactive intestinal peptide (VIP),
Somatostatin: Paracrine: inhibits hormone release from
endocrine pancreas and enzymes from exocrine pancreas
Endocrine: reduces contractions of
alimentary tract and gallbladder smooth muscles
VIP: glycogenolysis; regulates smooth muscle tonus and
motility of gut; controls ion and water secretion by intestinal
epithelial cells PP
(pancreatic polypeptide-producing)
1% Scattered
throughout islet
180 nm in diameter
Pancreatic polypeptide
Inhibits exocrine secretions of pancreas
G (gastrin-
producing) 1%
Scattered throughout
islet
300 nm in diameter Gastrin
Stimulates production of hydrochloric acid by parietal
cells of stomach

Part II – Chapter 4- 10
LIVER AND GALL BLADDER • The liver is the largest gland in the body, weighing approximately 1500 g. • It is located in the upper right-hand quadrant of the abdominal cavity, just inferior to
the diaphragm. • The liver is subdivided into four lobes: right, left, quadrate, and caudate-the first two
of which constitute its bulk. • Similar to the pancreas, the liver has both endocrine and exocrine functions; unlike the
pancreas, the same cell (the hepatocyte) in the liver is responsible for the formation of bile-the liver's exocrine secretion-and its numerous endocrine products.
• In addition, hepatocytes convert toxic substances into nontoxic materials that are excreted in bile.
General Hepatic Structure and Vascular Supply • The inferior, concave aspect of the liver houses the porta hepatis, through which the
portal vein and hepatic artery bring blood into the liver and through which the bile ducts drain bile from the liver.
• The liver is completely enveloped by peritoneum (except bare area), which forms a simple squamous epithelium covering over the dense, irregular connective tissue capsule (Glisson's capsule) of the gland.
• Glisson's capsule is loosely attached over the entire circumference of the liver except at the porta hepatis, where it enters the liver, forming a channel for the blood and lymph vessels and bile ducts.
• The liver is unusual in that its connective tissue elements are sparse; thus, the bulk of the liver is composed of uniform parenchymal cells, the hepatocytes.
• The superior aspect of the liver is convex, whereas its inferior region presents a hilum-like indentation, the porta hepatis.
• The liver has a dual blood supply, receiving oxygenated blood from the left hepatic artery and the right hepatic artery (25%) and nutrient-rich blood via the portal vein (75%).
• Both vessels enter the liver at the porta hepatis. • Blood leaves the liver at the posterior aspect of the organ through the hepatic veins,
which deliver their contents into the inferior vena cava. • Bile also leaves the liver at the porta hepatis, by way of the right and left hepatic ducts,
to be delivered to the gallbladder for concentration and storage. • All nutrients (except for chylomicrons and lipids less than 12 carbons in length)
absorbed in the alimentary canal are transported directly to the liver via the portal vein.
• In addition, iron-rich blood from the spleen is routed, by way of the portal vein, directly to the liver for processing.
• Much of the nutritive material delivered to the liver is converted by the hepatocytes into storage products, such as glycogen, to be released as when required by the body.

Part II – Chapter 4- 11
• • • • Hepatocytes are arranged in hexagon-shaped lobules (classical lobules) about 2 mm in
length and 700 µm in diameter. • These lobules are clearly demarcated by slender connective tissue elements (known as
portal tracts) in animals such as the pig and the camel have excessive CT. • In humans, the boundaries of the classical lobules difficulty recognized • Where three classical lobules are in contact with each other, the connective tissue
elements are increased, and these regions are known as portal areas (triads). • In addition to lymph vessels, portal areas house the following three structures, each of
which follows the longitudinal axis of each lobule: 1. Branches of the hepatic artery 2. Tributaries of the relatively large portal vein 3. Interlobular bile ducts (recognized by their simple cuboidal epithelium).

Part II – Chapter 4- 12
• The portal areas are isolated from the liver parenchyma by the limiting plate, a sleeve of modified hepatocytes.
• A narrow space, the space of Möll, separates the limiting plate from the connective tissue of the portal area.
• Only three equally distributed portal areas are present in a random section. • Along the length of each slender branch of the hepatic artery within the portal area,
fine branches, known as distributing arterioles, arise; like outstretched arms, they reach toward their counterparts in the neighboring portal areas.
• Smaller vessels, known as inlet arterioles, branch from the distributing arterioles (or from the parent vessel).
• In addition, the interlobular bile ducts are vascularized by a peribiliary capillary plexus.
• Venules are of two sizes: the larger distributing veins and the smaller inlet venules. • The central vein, the initial branch of the hepatic vein, occupies the longitudinal axis
of each classical lobule. • Hepatocytes radiate, like spokes of a wheel, from the central vein, forming
anastomosing, fenestrated plates of liver cells, separated from each other by large vascular spaces known as hepatic sinusoids
• Inlet arterioles, inlet venules, and branches from the peribiliary capillary plexus pierce the limiting plate (of modified hepatocytes) to join the hepatic sinusoids
• As blood enters the sinusoids, its flow slows considerably and it slowly penetrates into the central vein.
• Because there is only one central vein in each lobule, it receives blood from every sinusoid of that lobule and its diameter increases as it progresses through the lobule. As the central vein leaves the lobule, it terminates in the sublobular vein.
• Numerous central veins deliver their blood into a single sublobular vein; sublobolar veins join each other to form collecting veins, which in turn form the right and left hepatic veins.
The Three Concepts of Liver Lobules • The three types of liver lobules are the classical lobules, portal lobules, and the
hepatic acinus (acinus of Rappaport). • There are three basic conceptualizations of the liver lobule. The classical liver lobule
was the first to be defined histologically because the connective tissue arrangement in the pig liver afforded an obvious rationale. In this concept, blood flows from the periphery to the center of the lobule into the central vein. Bile, manufactured by liver cells, enters into small intercellular spaces, bile canaliculi, located between hepatocytes, and flows to the periphery of the lobule to the interlobular bile ducts of the portal areas.

Part II – Chapter 4- 13
Liver's general features • Large, lobulated exocrine and blood-processing gland, with • Vessels and ducts entering and leaving at the porta. • Enclosed by a thin CT capsule, mostly covered by mesothelium. • CT of the branching vascular system provides gross support. • Parenchymal cells are supported by fine reticular fibers. • The internal structure is understandable in terms of the several vessels entering or
leaving the organ; a. Portal vein bringing food-rich blood from the gut. b.Hepatic artery bringing arterial blood. c. Hepatic veins taking away processed blood into the vena cava. d.Lymphatics taking away some lymph. e. Hepatic ducts removing bile to the gallbladder and gut.
Liver lobule • First impression is of a uniform mass of large
glandular cells throughout the liver substance. • Closer examination shows that the cells are arranged in
perforated plates, one cell wide. Between the plates are sinusoidal blood channels 9-l2 µm wide, lined by endothelial cells.
• Scattered in the glandular mass are blood vessels, alone and accompanied by other vessels.
• The distribution of these vessels defines or marks out the classic hepatic lobules.
LIVER ARCHITECTURE • The liver has several grossly visible lobes demarcated by deep fissures and prominent
CT septa. Each lobe contains many lobules (acini), which are the basic functional units of the liver. Since the liver parenchyma in humans appears continuous, with no distinct boundaries between lobules, histologists have imposed imaginary boundaries to define liver lobules in 3 ways: 1. Classical lobules are roughly hexagonal. They are centered on a central vein and
have a portal canal in each corner. This description is based on blood-flow in the lobule; blood enters the lobule through the portal canals and drains through the central vein.

Part II – Chapter 4- 14
2. Portal lobules are triangular. Each triangle has a portal canal in the center and a central vein at each apex. It does not describe the smallest unit of functional organization in the liver.
3. Rappaport's liver acini are rhomboidal and have a central vein at each end and a portal canal approximately in the middle of each side. This definition describes tissue that is supplied by a terminal branch of the portal vein and a terminal branch of the hepatic artery, and that is drained by a terminal branch of the bile duct.
Varieties of liver vessel • Central vein/terminal hepatic venule - very thin wall; lies in the center of a lobule,
with sinusoids converging towards and opening into it. • Sublobular/intercalated vein - thicker wall; lies alone at the periphery of the lobule. • Branch of portal vein - again at the periphery of the lobule, but accompanied by
one or more small hepatic arteries/arterioles, one or more bile ducts/ductules lined by cuboidal epithelium, and lymphatics.
• Vein, artery, and bile duct constitute a portal triad; the area in which they lie is a portal area/canal. (The lymphatics are ignored for this naming).
• In pig and camel, the lobules are separated from one another by CT and thus much more easily identified.
Hepatic lobular blood flow 1. From branches of the portal vein and hepatic artery; from the periphery towards the
center; 2.In the sinusoids, between the cell plates. 3.Blood collected in central veins goes to sublobular veins, thence to collecting veins,
and then hepatic veins leaving the liver.
Intralobular bile flow
• It is from the lobule's centre towards the peripheral bile ducts, and runs, within any one cell plate, between the liver cells in bile canaliculi.
• Rappaport's liver acinus was a functional unit comprising parts of three or so lobules. It sought to account for differences in exposure to the blood supply among various parts of lobules. Such differences are reflected in varied functional activities and degrees of susceptibility to toxic agents - a metabolic zonation.
• The territory of an acinus has, as its axis, one final branch of the portal vein, and is subdivided into: 1. Periportal, (zone 1) 2. Intermediate, (zone 2) 3. Perivenous (close to the central vein) (zone 3) zones, with the initial periportal zone being roughly spheroid, and isolated from periportal zones of adjacent acini.

Part II – Chapter 4- 15
• The concept is not easy for students to follow, nor, it seems, for hepatocytes, which, for many processes, heed different patterns. To best-fit events to the architecture, hepatologists are now more likely to employ the simpler concept of separately continuous periportal and perivenous/pericentral zones, than that of discrete acini.

Part II – Chapter 4- 16
Liver sinusoids 1. Are lined by fenestrated endothelial cells, loosely attached, and hold 2. Phagocytic Kupffer cells (larger, stellate, with a pale oval nucleus), demonstrated by
the vital intravascular injection of Trypan blue or carbon particles, or latex particles for microscopy in vivo.
3. Fenestrated lining cells are not tightly attached and rest on microvilli of underlying hepatic cells, without a BL intervening.
4. Plasma can thus pass through the sieve plate, formed by the lining cells, out into the perisinusoidal space of Disse to interact with the hepatocytes. Some of this fluid may pass to the periphery of the lobule to be collected as lymph.
5. Disse's 'space' contains ECM materials, but not a visible basal lamina. 6. Scarce, fat-storing, stellate cells of Ito lie outside the endothelial cells. They store
vitamin A. They respond to a variety of insults by making collagen and causing cirrhosis (fibrosis).
7. The sinusoidal wall provides for: a. Blood cleansing, e.g., of gut bacterial toxins; b. Haemopoiesis in the embryo; c. Bringing plasma into intimate contact with the hepatic cell for its many metabolic
functions of storage, transformations, syntheses, regulation of plasma concentrations, detoxifications, the production of bile, and assisting defense by producing acute-phase proteins.
Hepatocyte/hepatic cell • Large, polyhedral, 30 µm x
20 µm cell with: 1. large, spheroid nucleus
(sometimes two), with membrane pores, and ribosomes on the outer membrane;
2. Extensive granular ER (protein synthesis for enzymes, plasma proteins, etc.);
3. smooth ER (steroid hormone and cholesterol metabolism; lipids are taken in, processed, and secreted in a way very like the enterocyte's; SER carries enzymes for detoxifications);

Part II – Chapter 4- 17
4. mitochondria (oxidative and other enzymes); 5. Actin and other filaments, near the bile canaliculi and elsewhere. 6. cell membrane projecting microvilli into the space of Disse, and held firmly to
adjacent cells, especially around the channel, the bile canaliculus, formed by the separation of two or three cells' membranes and equipped with a few microvilli;
7. Golgi body lying near the canaliculus, as do the 8. lysosomes; both appear to help form bile; Peroxisomes with other enzymes, e.g.,
Catalase; 9. glycogen granules stored in association with smooth ER (an association seen
elsewhere); fat droplets occurring briefly after meals; 10. Lipofuscin or aging pigment, as another normal inclusion; and sometimes brown
haemosiderin, with its iron, may be seen. Bile pathways • System of canaliculi (seen easily only with EM or special impregnation) between the
hepatic cells leads to I. Canals of Hering/cholangioles, with both hepatocytes and pale duct cells in their
walls. Next come, in the portal areas, II. Bile ductules with only small, pale cuboidal cells, firmly held by membrane
interdigitations and junctional complexes, and having a few luminal microvilli. III. Bile ducts' epithelium changes to columnar mucous cells and, extrahepatically, the
ducts acquire smooth muscle as well as CT. IV. Cystic duct allows reflux into the gallbladder, when Boyden's sphincter choledochus
at the duodenal outlet of the common bile duct is closed. • Bile production starts with inward bile-acid pumping across the sinusoidal region of the
hepatocyte membrane. Then, transporters in the canalicular membrane send bile salts, phosphatidylcholine, and toxic metabolites out into the bile.
Gall bladder 1. Extensively folded mucosa of tall, simple, columnar epithelial cells with many
microvilli, lying on a loose lamina propria. 2. Goblet cells are absent, but in the neck there may be small glands of uncertain
function. 3. The middle layer has variously disposed (mainly circular) smooth muscle bundles. 4. Outermost is a serosa of mesothelium-covered areolar CT with vessels and
nerves, except where the gallbladder attaches to the liver. 5. Function - stores and concentrates the bile by actively absorbing sodium, coupled with
water and anions. The hormone - cholecystokinin - released from gut endocrine cells in response to fat or amino acids causes contraction of the muscle to expel the bile.