caracterizaÇÃo hidroquÍmica e distribuiÇÃo das...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE SANTA CRUZ DEPARTAMENTO DE CIENCIAS BIOLÓGICAS PROGRAMA PÓS-GRADUAÇÃO EM SISTEMA ÁGUATICOS TROPICAIS ÁREA DE CONCENTRAÇÃO ECOLOGIA
CARACTERIZAÇÃO HIDROQUÍMICA E DISTRIBUIÇÃO DAS ASSEMBLÉIAS MACROBÊNTICAS NA PORÇÃO CENTRAL DO RECIFE DE COROA VERMELHA, SANTA CRUZ DE CABRÁLIA, BAHIA.
THAÍS BOMFIM DE SANTANA
ILHÉUS 2006

THAIS BOMFIM DE SANTANA
CARACTERIZAÇÃO HIDROQUÍMICA E DISTRIBUIÇÃO DAS ASSEMBLÉIAS MACROBÊNTICAS NA PORÇÃO
CENTRAL DO RECIFE DE COROA VERMELHA, SANTA CRUZ DE CABRÁLIA, BAHIA.
Dissertação apresentada, para obtenção do título de Mestre em Sistemas Aquáticos Tropicais, à Universidade Estadual de Santa Cruz. Área de Concentração: Ecologia e Meio Ambiente. Orientador: Marcelo Friederichs Landim de Souza Co-orientadora: Erminda da Conceição Guerreiro Couto
ILHÉUS 2006

S231 Santana, Thaís Bomfim de Caracterização hidroquímica e distribuição das
assembléias macrobênticas na porção central do recife de Coroa Vermelha, Santa Cruz Cabrália, Bahia / Thaís Bomfim de Santana. – Ilhéus, BA: UESC, 2006.
66 f.: il. Orientador: Marcelo Friederichs Landim de Souza. Dissertação (Mestrado) – Universidade Estadual de
Santa Cruz. Programa de Pós-Graduação em Sistemas Aquáticos Tropicais.
Inclui referências.
1. Biologia dos recifes de coral – Porto Seguro (BA). 2. Corais
– Porto Seguro (BA). 3. Ecossistemas – Bahia. 4. Química da
água. 5. Biodiversidade marinha. I. Título.
CDD 593.6098142

THAÍS BOMFIM DE SANTANA
CARACTERIZAÇÃO HIDROQUÍMICA E DISTRIBUIÇÃO DAS ASSEMBLÉIAS MACROBÊNTICAS NA PORÇÃO CENTRAL DO RECIFE DE COROA
VERMELHA, SANTA CRUZ DE CABRÁLIA, BAHIA.
APROVADA EM: ___/___/___.
Comissão Examinadora
_________________________________________________________________
Dr. Marcelo Friederichs Landim de Souza (Orientador)
_________________________________________________________________ Dra. Erminda da Conceição Guerreiro Couto (Co-Orientadora)
__________________________________________________________________ Dra. Fernanda Maria Duarte Amaral (UFRPE)
__________________________________________________________________ Dra. Daniela Mariano Lopes da Silva (UESC- DCET)
Suplente:
____________________________________________________________________ Dra. Gecely Rodrigues Rocha (UESC - DCB)

A minha família, razão de meus esforços e por quem busco a
todo o momento ser motivo de orgulho e nunca decepcionar.
Aqui está mais um fruto de todo o incentivo e apoio que
tenho recebido em minha jornada.

AGRADECIMENTOS
O presente trabalho é o resultado da integração dos esforços de um conjunto de pessoas
que direta ou indiretamente comprometidas, subsidiaram apoio científico, técnico, financeiro,
espiritual, sentimental e familiar ao meu plano de investigação e acima de tudo, de vida. Desta
forma, gostaria de agradecer:
Aos meus pais que me financiaram uma boa educação e por estarem sempre ao meu
lado, me dando forças mesmo quando estas me faltavam.
Aos meus familiares pelo apoio moral e pela compreensão da minha ausência durante
longos períodos.
Ao meu namorado e melhor amigo por toda paciência, incentivo em todos os
momentos, apesar dos desentendimentos por não ter dado a atenção que ele merecia neste
tempo.
Ao professor Marcelo Landim pela oportunidade de trabalhar com a área que eu
sempre almejei, e graças a ele, eu alcancei. Por ter aceitado o desafio da orientação e confiado
no meu potencial para a realização deste trabalho, pela dedicação e eficiente orientação e
principalmente, pela amizade gratificante.
A Erminda Couto, pelos incentivos, apoio e “puxões de orelhas” na hora certa que
ajudaram e vem ajudando a construção do meu perfil científico e por ter sido mais que uma
professora, uma amiga que me ajudou e me aconselhou num dos momentos mais difíceis da
minha vida.
A Maria Aparecida e Gilmara pela paciência e orientação dentro do laboratório,
estando sempre dispostas a ajudar nas análises das amostras.
A minha melhor amiga, Dani, pelo apoio, conselhos e um ombro amigo para desabafar
e me apoiar nas horas difíceis.
Aos professores do mestrado que ajudaram na orientação e construção do meu
pensamento científico.

RESUMO
Os recifes de coral são ecossistemas de elevada biodiversidade formados pelo acúmulo
de carbonato de cálcio produzido pelos corais escleratinídeos e pelas algas calcáreas .
Ocorrem, de forma geral, em zonas tropicais localizadas próximas às praias e distantes dos
estuários, entre as isotermas de 20°C nos hemisférios norte e sul. Este trabalho foi realizado no
Recife de Coroa Vermelha (16º19’53”S e 39º00’00”W), situado em Porto Seguro
(Bahia,Brasil), com o objetivo de avaliar as características físico-químicas e biológicas do
Recife de Coroa Vermelha, e taxas de calcificação e metabolismo a nível de ecossistema. O
sistema estudado apresentou ampla variação nas características hidroquímicas tanto espacial
quanto temporal. Pode-se ressaltar que o sistema apresenta uma elevada taxa de processos
biogeoquímicos e com elevada taxa de resposta a mudanças temporais. E quanto as
características bióticas, o grupo de maior representatividade no sistema é o de corais pétreos .
Palavras-Chave: Sistema, Recifes de Coral, Processos e Hidroquímica

SUMÁRIO 1. INTRODUÇÃO ................................................................................................................ 11 2. OBJETIVOS ..................................................................................................................... 21 Objetivo geral ....................................................................................................................... 21 Objetivos Específicos ........................................................................................................... 21 3. ÁREA DE ESTUDO ......................................................................................................... 21 4. MATERIAIS E MÉTODOS ............................................................................................ 25 Caracterização biológica do ambiente e estabelecimento das estações .......................... 25 Diagnose dos hidróides calcáreos e dos corais pétreos ..................................................... 25 Caracterização hidroquímica do ambiente recifal ........................................................... 25 Cálculo das taxas de calcificação e do metabolismo líquido das poças de maré ............ 27 5. RESULTADOS E DISCUSSÃO ..................................................................................... 28 Diagnose dos hidróides calcáreos e dos corais pétreos ..................................................... 28
Distribuição espacial das associações macrobênticas ................................................... 39 Caracterização hidroquímica das poças ............................................................................ 46 Temperatura ......................................................................................................................... 46 Salinidade .............................................................................................................................. 47 pH ........................................................................................................................................... 49 Oxigênio dissolvido ............................................................................................................... 50 Nutrientes inorgânicos dissolvidos ...................................................................................... 51 Alcalinidade total .................................................................................................................. 55 CO2 Total (ou CID, ou ΣΣΣΣCO2, padronizar) e pCO2 ........................................................... 56 Saturação da Aragonita (ΩΩΩΩarag.) ........................................................................................... 58 Calcificação e metabolismo líquido ..................................................................................... 58 6. CONCLUSÃO ................................................................................................................... 60 7. REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 61

LISTA DE FIGURAS Figura 1. Distribuição mundial dos ambientes recifais a nível mundial. Informação de Mitsuaki Takata.........................................................................................................................12 Figura 2- Sistema de recifes de Coroa Vermelha.....................................................................22 Figura 3: Imagem retratando a borda externa ( próxima ao mar) do recife de Coroa Vermelha....................................................................................................................................23 Figura 4- Vista geral do tômbolo que facilita o acesso de pessoas ao platô recifal de Coroa vermelha.....................................................................................................................................24 Figura 5: Vista geral do platô do recife de Coroa Vermelha....................................................24 Figura 6: Foto tirada de cima de um espécime de Agaracia agarites coletado no recife de Coroa Vermelha.........................................................................................................................31 Figura 7: Fotos tiradas nos Recifes de Coroa Vermelha. (a) Imagem de um espécime de Siderastea stellata em detalhes.(b) Imagem de uma poça de maré tirada com um quadract de 25X25 retratando a abundância de Siderastea Stellata..............................................................32 Figura 8: Foto de uma colônia de Favia grávida dominada por cianobactérias.............................................................................................................................34
Figura 9: Imagem de uma colônia de Mussismilia harttii encontrada no Recife de Coroa Vermelha....................................................................................................................................37
Figura 10: Distribuição espacial das espécies de Siderastrea stellata e Favia gravida somadas ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo......................................................................................................................................40 Figura 11: Distribuição espacial das espécies de Siderastrea stellata e Favia gravida ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo..............................................40 Figura 12: Distribuição espacial das macroalgas crustosas mais as frondosas ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo..................................................................41 Figura 13: Distribuição espacial do microfitobentos ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.........................................................................................................41

Figura 14: Distribuição espacial dos poríferos ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.........................................................................................................42 Figura 15: Distribuição espacial do grupo Ascidiaceae ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo............................................................................42 Figura 16: Distribuição espacial das áreas sem cobertura animal ou vegetal (desnudas) e recobertas por areia fina a média ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo......................................................................................................................................43 Figura 17: Distribuição espacial das espécies Zoanthus sociates e Palythoa caribaeorum ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo......................................................................................................................................43 Figura 18: Distribuição espacial dos principais componentes do recife de Coroa Vermelha ao longo de um transecto perpendicular ao mesmo, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo............................................................................44 Figura 19 - Distribuição da temperatura nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul................................47 Figura 20 - Distribuição da salinidade nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul................................48 Figura 21 - Distribuição do pH nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul....................................................49 Figura 22 - Distribuição da concentração de oxigênio dissolvido nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................50 Figura 23 - Distribuição da concentração de amônio nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................51 Figura 24 - Distribuição da concentração de nitrito nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................52 Figura 25 - Distribuição da concentração de nitrato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................53

Figura 26 - Distribuição da concentração de fosfato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................54 Figura 27 - Distribuição da concentração de silicato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul; * = não analisado....................................................................................................................................55 Figura 28: Distribuição da alcalinidade total nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul................................56 Figura 29: Distribuição da concentração de carbono inorgânico dissolvido (CID) nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.............................................................................................................................57 Figura 30: Distribuição da pressão parcial de dióxido de carbono (pCO2) na água das poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.............................................................................................................................57 Figura 31: Distribuição do estado de saturação da aragonita (Ωarag.) nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul...............................................................................................................................................58 Figura 32: Taxas de calcificação/dissolução de carbonatos e metabolismo líquido (P – R) nas poças ao longo das campanhas. ................................................................................................59

11
1. INTRODUÇÃO
Os recifes de coral são ecossistemas de elevada biodiversidade formados pelo acúmulo
de carbonato de cálcio produzido pelos corais escleratinídeos e pelas algas calcáreas (Bates,
2002). A diversidade de espécies de corais nos recifes de coral difere de acordo com sua
história geológica e biológica. Os recifes do Indo-Pacífico apresentam a maior diversidade,
apresentando cerca do dobro de espécies de corais pétreos existentes nos ricos recifes do
Caribe, que por sua vez são mais diversificados que os brasileiros (Paulay, 1997). Pode-se
considerar, que de forma geral, os ambientes recifais brasileiros apresentam baixa diversidade
de corais, distribuição descontínua e localização, em sua maior parte, em estreitas linhas
próximas à costa (Castro, 1999), ocorrendo desde o Parcel Manoel Luiz (Ma) até Arraial do
Cabo (RJ), além do Atol das Rocas (RN) e Fernando de Noronha (Pe) (Leão et al., 2003).
Entretanto, as características do sistema fazem com que mesmo os recifes brasileiros
apresentem uma altíssima diversidade biológica, quando comparados com outros ambientes.
Um recife de coral, sob o ponto de vista geomorfológico, é uma estrutura rochosa,
rígida, resistente à ação mecânica das ondas e das correntes marinhas, construída por
organismos marinhos que são capazes de secretar um exoesqueleto carbonático (Leão, 1994).
Em geral são denominados “recifes de coral” devido ao papel preponderante desempenhado
por estes organismos na construção destes ambientes em diversas partes do mundo. Sob o
ponto de vista biológico, recifes coralíneos são formações criadas pela ação de comunidades
de organismos denominados genericamente "corais". Embora a estrutura básica de recifes
biogênicos seja em geral formada pelo acúmulo dos exoesqueletos destes animais, para sua
formação é necessária a atuação conjunta de uma infinidade de seres, formando uma complexa
teia de associações e de eventos em sucessão. Estudos desenvolvidos a partir da década de 80
têm demonstrado que, em muitos casos, outros organismos podem desempenhar um papel de
igual ou maior relevância. Em alguns recifes, inclusive do Brasil, o crescimento de outros
organismos, como algas calcárias, pode assumir uma relevância igual ou superior a dos
próprios corais (Kikuchi & Leão, 1997).
Estão presentes, de forma geral, em zonas tropicais localizadas próximas às praias e
distantes dos estuários, entre as isotermas de 20°C nos hemisférios norte e sul (Kaplan, 1982)
(Fig. 1). A faixa de temperatura ideal para sua existência estaria entre 25 e 29°C, salinidade

12
em torno de 36S, águas de elevada transparência, baixa densidade planctônica e baixas
concentrações de nutrientes (Belém et al., 1986). O excesso de nutrientes pode diminuir a
transparência da água e estimular o crescimento de macro e microalgas e a bioerosão (Hallock,
2002).
Figura 1. Distribuição mundial dos ambientes recifais a nível mundial. Informação de Mitsuaki Takata.
Ambientes recifais são ainda importantes para o homem em diversos aspectos. Em
termos físicos, protegem as regiões costeiras da ação do mar em diversas áreas do litoral. A
grande diversidade e quantidade de organismos presentes associam-se em uma teia alimentar
de grande complexidade. Esta teia culmina nos grandes predadores, que incluem muitos dos
peixes utilizados para alimentação humana. Os recifes funcionam como verdadeiros
criadouros de peixes, renovando estoques e, principalmente, no caso de áreas protegidas,
favorecendo a reposição de populações em áreas densamente explotadas. Devido à
complexidade das suas cadeias alimentares e à intensa competição por espaço entre os
organismos sésseis, muitos organismos presentes produzem substâncias que são utilizadas
para proteção contra predadores, inibição da ocupação do espaço por competidores e outras
funções (Pennings, 1997). Pesquisadores em farmacologia buscam extrair e isolar estas

13
substâncias, testando suas propriedades no tratamento de doenças e disfunções no homem
(Rinehart, 1988), desta forma, estes sistemas fornecem matéria prima para pesquisas na área
farmacológica.
Entre os principais representantes da fauna coralínea, habitando estes ambientes,
encontram-se os corais e os hidróides calcáreos, pertencentes ao Filo Cnidaria (Brusca &
Brusca, 2003). Os primeiros pertencem à Classe Anthozoa e a Ordem Scleractinia e possuem
um esqueleto calcáreo externo composto, principalmente, por carbonato de cálcio. Podem ser
zooxantelados, quando apresentam dinoflagelados simbiontes (zooxantelas) ou
azooxantelados, na ausência desta simbiose. Os zooxantelados participam ativamente da
construção de ambientes recifais enquanto os azooxantelados raramente participam desta
formação (Schumacher & Zibrowius, 1985). O incremento na produção de esqueleto
associado à presença das zooxantelas é considerado um dos principais fatores que restringem a
presença de recifes de coral a baixas profundidades, até cerca de 50m, uma vez que estes
protistas necessitam de luminosidade para realizar a fotossíntese. A quantidade de luz
presente no mar decresce à medida que aumenta a profundidade. A coluna de água funciona
como um filtro azul cada vez mais intenso, até bloquear totalmente a passagem da luz,
inibindo este processo. Outro fator determinante no desenvolvimento dos recifes de coral é a
temperatura da água. Este fator limita a presença de recifes apenas às áreas onde a temperatura é
mais elevada, em geral áreas tropicais com correntes de águas quentes, como é o caso da costa
brasileira.
Os segundos pertencem a Classe Hydrozoa e à Ordem Capitata, possuindo o
polimorfismo típico dos hidrozoários (Ruppert & Barnes, 1986). Também secretam um
esqueleto externo de carbonato de cálcio com poros, contribuindo para a construção dos
recifes (Amaral, 1997).
Estes ecossistemas, além da sua elevada biodiversidade têm despertado recentemente
grande interesse, uma vez que processos biológicos que neles ocorrem, como a calcificação e
fotossíntese, estão intimamente relacionados com as mudanças climáticas globais (Gattuso et
al, 1999).
A fotossíntese e a calcificação consomem carbono inorgânico dissolvido e respondem,
simultaneamente, a mudanças em parâmetros ambientais tais como, luz e temperatura (Leclerq
et al, 2002). O acompanhamento destes fatores pode ajudar num melhor entendimento do

14
sistema e responder questões relacionadas ao aumento na pressão parcial do CO2(pCO2) na
atmosfera.
Leclercq et al (2000) e Gattuso et al (1998) sugeriram e recentemente demonstraram
que um aumento na pressão parcial de CO2 (pCO2) tem um efeito negativo em corais e
comunidades recifais calcificantes como um resultado do decréscimo do estado de saturação
da aragonita. Estima-se que pCO2 irá alcançar o dobro do nível pré-industrial no ano de 2065
(Gattuso et al, 1999), podendo reduzir os níveis de saturação do carbonato na superfície do
mar tropical em torno de 2/3 dos valores atuais (Gattuso et al, 1998). O aumento na pCO2
atmosférica, previsto para este século, provocará um significativo decréscimo na calcificação
marinha (Kleypas et al,1999; Leclerq et al, 2002).
O aumento na concentração de CO2 na atmosfera age sinergicamente com outros
estresses antropogênicos, como a eutrofização, fluxo turístico e aumento de taxas de
sedimentação. Em escala regional estes fatores podem ser mais importantes que as mudanças
globais, especialmente em ambientes recifais costeiros. Além de constituírem uma ameaça a
biodiversidade, podem afetar drasticamente o metabolismo dos corais e suas relações
ecológicas (Reaser et al, 2000; Abramovitch-Gottlib et al, 2002).
A proximidade de grandes cidades e a facilidade de acesso aos ambientes recifais os
tornam cada vez mais vulneráveis à influência humana (Castro & Pires, 2001), sendo, na
maioria das vezes, destruídos pelas mesmas atividades humanas econômicas que os sustentam
(Wilkinson, 1992; Richmond, 1993).
Entre os problemas relacionados ao desenvolvimento costeiro, a produção de lixo e de
esgotos “in natura” podem causar alterações na composição das comunidades recifais, por
exemplo, através de um favorecimento ao crescimento de algas em detrimento dos corais. Este
tipo de problemas é especialmente grave em áreas recifais fronteiriças a grandes concentrações
humanas, como é o caso da maioria dos recifes costeiros da Paraíba até a Bahia. Na costa dos
arrecifes (Paraíba até Alagoas), foi observado que os recifes localizados próximos a grandes
cidades (Recife e Maceió) encontram-se bastante modificados (Maida & Ferreira, 1997).
O turismo e recreação marinhos representam um setor em franca expansão em várias
regiões do Brasil,. Leão et al. (1994) relataram um aumento de 400% no número de visitantes
ao Parque Nacional Marinho dos Abrolhos em período de cinco anos (1988-1992). Após este

15
período, o número de visitantes continuou a crescer, ampliando-se também a sua distribuição
ao longo do ano. Em parte, esta maior distribuição ocorreu com o advento do “turismo das
baleias”, cuja presença ocorre de julho a novembro. Em contrapartida, o “turismo de
mergulho” ocorre principalmente no verão. O aumento da visitação a áreas recifais traz
problemas, entre eles os danos físicos à biota, como pisoteamento e ancoragem de barcos e a
pesca predatória por mergulhadores amadores. Esta pesca é altamente seletiva, direcionada a
algumas poucas espécies, podendo diminuir drasticamente os estoques das espécies visadas,
como os Serranidae (meros, badejos e garoupas).
O Brasil possui os únicos recifes coralíneos do Atlântico Sul. Sua distribuição real
ainda é pouco conhecida. O trabalho mais abrangente sobre o assunto foi realizado na década
de setenta (Laborel, 1970) que, em muitas áreas, enfrentou sérios problemas logísticos.
Grandes comunidades coralíneas foram registradas desde o Parcel Manuel Luís - Ma
(~01°S) até os recifes de Viçosa, na área de Abrolhos - Ba (~18°S), em ilhas oceânicas como
Atol das Rocas e Fernando de Noronha. Algumas comunidades expressivas também foram
observadas em direção ao sul, estendendo-se até Cabo Frio – RJ, no que foi chamada “zona de
desaparecimento das comunidades coralíneas” (Laborel, 1970).
Como pode ser visto os recifes brasileiros não formam uma unidade homogênea,
entretanto, em mais de 2.000km de litoral, existem apenas 7 unidades de conservação
marinhas que englobam comunidades recifais significativas. Duas delas em ilhas oceânicas
(Reserva Ecológica do Atol das Rocas e Parque Nacional Marinho de Fernando de Noronha),
quatro nos limites de distribuição de recifes (Parque Estadual Pedra da Risca do Meio - Ce,
Parque Estadual do Parcel Manoel Luís - Ma, Parque Nacional Marinho dos Abrolhos e Área
de Proteção Ambiental da Ponta da Baleia - ambas localizadas no Banco dos Abrolhos, Ba),
duas em áreas mais próximas à costa (APA Costa dos Corais, Pe/Al e Parque Municipal do
Recife de Fora - Porto Seguro, BA). A APA da Costa dos Corais e o PM do Recife de Fora se
encontram em áreas altamente ameaçadas pela grande acessibilidade, pela proximidade de
grandes populações humanas e pela expressiva visitação.
A história geológica de recifes brasileiros é mais bem conhecida para a costa baiana,
graças ao trabalhos desenvolvidos por Leão e colaboradores, da Universidade Federal da

16
Bahia, a partir do início da década de 1980 (Leão, 1982, 1994, 1996; Leão et al., 1988, 1997;
Kikuchi & Leão, 1997, 1998; e outros). A história geológica do período Quaternário indica
que os recifes de coral atuais só começaram a crescer a 7.000 anos atrás, quando o nível do
mar elevou-se e, desta forma, inundou toda a atual plataforma continental. Os corais,
hidrocorais e outros organismos foram crescendo em direção à superfície do mar construindo,
com os seus exoesqueletos, uma estrutura rochosa. Como o nível do mar, durante os últimos
7.000 anos, subiu e desceu várias vezes, estas informações foram comparadas com os registros
das idades de corais obtidos em um testemunho retirado do Recife de Coroa Vermelha - Ba.
Este trabalho revelou que os recifes cresceram acompanhando a subida do mar, e alcançaram o
seu clímax, a aproximadamente 5.000 anos antes do presente, coincidindo com a altura
máxima do mar que, nessa época, estava cerca de 5m acima do nível atual.
Uma fase de estabilidade do nível do mar, que durou cerca de 1.000 anos, deve ter
favorecido o crescimento de numerosas estruturas recifais ao longo de todo o litoral. Porém
desde há 4.000 anos, devido a várias descidas bruscas do nível do mar, as quais deixaram os
topos de diversos recifes emersos e sujeitos à erosão, as estruturas recifais só puderam crescer
lateralmente, o que deve ter provocado a coalescência dos chapeirões baianos e a formação de
bancos recifais. Além disso, em diversos estados do Brasil, comunidades recifais se instalaram
sobre bancos de arenito de praia, em geral estreitos, alongados e localizados adjacentes à praia
(Leão, 1996).
Estudos quantitativos de comunidades coralíneas iniciaram-se principalmente com
pesquisadores do Rio de Janeiro, especialmente na área de Abrolhos (Pitombo et al., 1988;
Coutinho et al., 1993; Maida et al., 1995; Villaça & Pitombo, 1997; Figueiredo, 1997; e
outros). Porém, ainda são poucos os estudos estruturais e funcionais desenvolvidos em áreas
recifais.
No passado, diversos recifes costeiros sofreram a retirada em larga escala de blocos
calcários para uso na construção civil e para suprimento de calcário para a indústria do açúcar
(Maida & Ferreira, 1997). Esta prática, já descrita desde o século passado (Hartt, 1870;
Branner, 1904), só foi banida na década de 1970 (Maida & Ferreira, 1997). A coleta comercial
de exemplares da fauna e flora para souvenirs e para aquariofilia é outra atividade
potencialmente prejudicial às comunidades marinhas. Esta prática torna-se predatória se não
levar em consideração características biológicas das espécies a serem exploradas (estoques

17
disponíveis, período reprodutivo, tamanho mínimo para reprodução, tempo de reposição de
estoques – recrutamento e crescimento, etc.). Há relatos de coleta de peixes ornamentais na
Baía de Todos os Santos - Ba, e de invertebrados marinhos na região de Arraial do Cabo - RJ,
com conseqüências danosas para algumas espécies, como anêmonas-do-mar (Condylactis
gigantea), gorgônias orelha-de-elefante (Phyllogorgia dilatata) e corais-de-fogo (Millepora
alcicornis) (Belém et al., 1986). Entretanto, ainda existem poucas avaliações formais
publicadas sobre as pressões exercidas em ambientes recifais brasileiros.
Impactos naturais geralmente não destroem os recifes quando isolados e pontuais no
tempo e/ou no espaço. Entre estes impactos estão incluídos efeitos de tempestades, variação
do nível do mar, epidemias de doenças em organismos recifais, aumentos massivos nas
populações de predadores. Entre os impactos antropogênicos, destaca-se o aumento na
turbidez da água como o que mais tem afetado regiões coralíneas naturais, em nível global.
Estes em geral estão associados a atividades na costa que aumentam o carreamento de
sedimentos para os oceanos, atividades turísticas e de recreação, contaminação por substâncias
tóxicas e exploração de petróleo.
O Estado da Bahia apresenta recifes por mais de 900 km ao longo de sua costa. A área
mais extensa que, aparentemente, não os possui localiza-se na parte norte do Estado, devido à
influência da desembocadura do Rio São Francisco (Laborel, 1970). A plataforma da Bahia
apresenta largura média de 50km, alcançando cerca de 200km ao sul (Leão et al., 1988). Leão
(1996) revisou o conhecimento de sua morfologia e distribuição e descreveu os principais
impactos ambientais sofridos por estes recifes, dividindo-os em cinco setores principais. Dois
destes setores ainda são quase praticamente desconhecidos; enquanto para os outros três já
foram estudados aspectos de sua história geológica, geomorfologia e estrutura da comunidade.
A localização e área dos recifes submersos de todos os setores ainda são pouco conhecidas.
A parte norte da Bahia (setor 1) apresenta bancos recifais descontínuos, localizados,
geralmente, em profundidades inferiores a 10m e formados por colunas isoladas que podem se
fundir nos seus topos (Leão, 1996). Outras comunidades de corais-algas ocorrem nestes recifes
como camadas finas (< 1m de espessura) sobre arenitos de praia que ocorrem paralelos à linha
de costa (Leão, 1996). A diversidade de corais é menor do que a existente ao sul, sendo
constituída por oito espécies de corais pétreos e um hidrocoral (Leão et al., 1997).

18
Estudos apontaram um forte declínio na cobertura de corais e mudança na estrutura das
comunidades dos recifes de coral localizados na parte norte da Bahia (Leão et al., 1997;
Kikuchi e Leão, 1998). Nos últimos 3.000 anos houve um forte declínio da cobertura da
principal espécie de coral construtora (Mussismilia braziliensis), e sua substituição por
Siderastrea stellata ou Mussismilia hispida como espécies principais. Este fato foi associado a
queda do nível do mar de 5 m. Estes mesmos autores registraram, neste trecho da costa, uma
cobertura de coral atual de menos de 6%, comparando esta taxa com a de 20% estimada para
os topos expostos antigos.
A área da Baía de Todos os Santos (setor 2), adjacente a Salvador, apresenta recifes em
franja ao longo da maior parte das costas leste e sudeste da Ilha de Itaparica e na costa leste
das ilhas situadas ao norte da Baía (Laborel, 1970; Leão, 1996). Perfis sísmicos e de
perfuração realizados na Ilha de Itaparica mostraram um recife biogênico de 10m de espessura
(Leão et al., 1988). Laborel (1970) tinha classificado os recifes da Ilha de Itaparica como
aparentemente mortos no início da década de 60. Ele associou este fato a extração e queima de
corais para a produção de calcário, como já havia sido previamente registrado por Hartt
(1870).
Entre Salvador e Abrolhos (setores 3-4) existiriam zonas diversificadas, com recifes
costeiros (biogênicos e comunidades desenvolvidas sobre bancos arenito) e bancos isolados ao
largo. Alguns destes recifes estão entre os menos conhecidos do Brasil, como os recifes da
Baía de Camamu (setor 3) e os Recifes de Itacolomis e grupos de pináculos recifais que os
circundam (setor 4).
A área de Abrolhos (setor 5), localizada no sul da Bahia, está localizada num
alargamento da plataforma continental. É considerada a maior e mais rica área de recifes de
coral do Atlântico Sul (Laborel, 1970; Leão, 1982, 1994, 1996; Leão et al., 1988; Castro,
1994), com recifes relativamente bem preservados que apresentam todas as espécies de corais
recifais já descritas para o Brasil (Laborel, 1969, 1970). Estes recifes são também os melhor
conhecidos da costa baiana (Leão, 1996), encontrando-se em profundidades normalmente
inferiores a 20 m. Esta área apresenta os recifes de coral verdadeiros conhecidos localizados
mais ao sul no Atlântico (~18° S) (Hartt, 1870; Laborel, 1970; Leão, 1994). É delimitada ao

19
norte por um golfo submarino profundo (“Poço do Jucurussú”) (Leão, 1994) e ao sul pela
desembocadura do Rio Doce (Laborel, 1970).
Abrolhos apresenta a maior diversidade de corais recifais do Brasil (Laborel, 1970;
Castro, 1994; Leão, 1996), com 17 escleractíneos (incluindo dois não formadores de recifes ou
ahermatípicos) e três mileporídeos. Leão & Ginsburg (1997) enfatizaram a ocorrência de altos
níveis de sedimentos siliciclásticos nestes ambientes recifais (como parece ser o caso de vários
outros recifes brasileiros), sugerindo que a baixa diversidade de corais nos recifes de Abrolhos
seria uma conseqüência desta condição. Entretanto, os corais escleractíneos brasileiros
necessitam de revisões taxonômicas que verifiquem realmente sua condição endêmica em
relação aos seus aparentados do Caribe, e a ocorrência de espécies “sibling” dentre as espécies
nominais de hoje.
Foram detectadas em alguns recifes de Abrolhos coberturas de corais de até 39%
(recifes localizados mais ao largo), com uma média de cobertura nos bancos circundantes de
cerca de 15% (Villaça & Pitombo, 1997, dados restritos a estações onde ocorreram corais). A
maior parte da cobertura de corais está localizada somente nos topos dos recifes, em
profundidades inferiores a 10m. Figueiredo (1997) observou coberturas de algas coralíneas de 32
a 79%, em águas extremamente rasas (0,5m). A autora indicou que esta cobertura era similar às
observadas em outros recifes de outras regiões do mundo, porém as médias absolutas das taxas de
crescimento de algas coralíneas incrustantes eram superiores as registradas em outras áreas do
Atlântico tropical. Leão (1996) sugeriu que os agentes antropogênicos mais comuns atuando
sobre os recifes da Bahia estão relacionados ao desenvolvimento da zona costeira, turismo,
pesca predatória, instalação de projetos industriais e exploração de petróleo. A autora indicou
que os recifes localizados mais próximos à costa encontram-se sob estresse, devido ao
aumento do carreamento de sedimentos causados pelo desmatamento das bacias hidrográficas
costeiras. Baseado na alta densidade de macroalgas nos recifes mais próximos da costa,
Coutinho et al. (1993) sugeriram a ocorrência de pesca predatória e/ou aumento nas taxas de
sedimentação. Não há registros conclusivos da interferência do desenvolvimento costeiro nos
recifes da área de Abrolhos. Estudos preliminares sugerem que existe uma interação
relativamente pequena entre a costa adjacente aos recifes e os sistemas recifais propriamente
ditos (Knoppers, 1996; Lopez & Dutz, 1996; Marone & Camargo, 1996). Entretanto, o
aumento de sedimentação pode estar relacionado a atividades costeiras realizadas em pontos

20
distantes da face do continente localizada em frente aos recifes. Foi sugerido que os
sedimentos do sul da Bahia são originados em grandes rios, como Jequitinhonha, localizados a
cerca de 200km ao norte das áreas recifais (Melo et al., 1975; Leão & Ginsburg, 1997). Em
outros trechos da costa o aporte de sedimentos carreados do continente também aproximou da
linha costeira muitos recifes e os submeteu à influência de uma forte sedimentação
siliciclástica. A maior parte destes sedimentos é composta por areias inconsolidadas com
camadas lamosas, sendo facilmente transportado para os ambientes recifais. Efeitos nocivos de
uma maior sedimentação sobre ambientes coralíneos incluem, por exemplo, a inibição da
fotossíntese realizada pelos protistas simbiontes dos corais e maior gasto metabólico para
remoção de partículas que se depositam sobre estes organismos. Embora os recifes costeiros
baianos coexistam com altas taxas de sedimentação há muito tempo, é sugerido (Leão, 1996;
Leão et al., 1997) que estejam passando por maior carga de estresse na atualidade, devido
principalmente a uma maior entrada de sedimento no sistema que parece estar associado a
aumento da perda de mata atlântica para práticas agrícolas e industriais ao longo das últimas
décadas (cana-de-açúcar, coqueirais, extração de madeira e cultivo de eucalipto) (Leão, 1996;
Maida & Ferreira, 1997).
Outros impactos estão associados a atividades industriais como a indústria de celulose
e a exploração de petróleo “offshore” (Leão, 1996). Leão et al. (1994) realizaram uma
descrição qualitativa dos distúrbios ocorridos na área de Abrolhos relacionados ao turismo.
Posteriormente, Creed & Amado Filho (1999) estudaram os distúrbios causados por ancôras e
o posterior recobrimento dos bancos de gramíneas. Os autores observaram que barcos de
turismo danificaram anualmente 0,5% da área recoberta pelos bancos.
Um evento de branqueamento massivo também foi também registrado, tendo sido
relacionado a um fenômeno de escala maior (Castro & Pires, 1999). Branqueamento de corais
em conjunção com estas anomalias vem sendo associado por muitos pesquisadores as
mudanças climáticas globais (UNEP/AIMS, 1993).
Este estudo visa caracterizar o ambiente do Recife de Coroa Vermelha (Porto Seguro,
Bahia) através de variáveis físico-químicas da água, do levantamento das principais espécies
de macroinvertebrados bênticos presentes, da diagnose dos corais pétreos e hidróides calcáreos
e da estimativa de taxas de calcificação/dissolução de carbonatos e metabolismo do carbono
nas poças de maré.

21
2. OBJETIVOS Objetivo geral
Avaliar as características físico-químicas e biológicas do Recife de Coroa Vermelha, e
taxas de calcificação e metabolismo a nível de ecossistema.
Objetivos Específicos
• Levantar os hidroides calcáreos e corais pétreos do sistema
• Determinar a distribuição dos principais componentes biológicos
• Caracterizar as variáveis físico-químicas da água do mar nas poças de maré e
água do mar adjacente ao Recife de Coroa Vermelha;
• Medir as taxas de calcificação/dissolução líquidas nas poças de maré;
• Medir as taxas metabólicas líquidas nas poças de maré;
3. ÁREA DE ESTUDO
O sistema de recifes de Coroa Vermelha (16º19’53”S e 39º00’00”W)(Fig.2) é formado
por estruturas descontínuas paralelas à costa, unidas ao litoral por um tômbolo (Fig.4).
O recife de Coroa Vermelha situa-se próximo ao povoamento de mesmo nome. A área
destaca-se pela associação de uma extensa estrutura recifal de natureza coralínea e arenítica.
Durante a maré vazante a superfície do recife é exposta sendo possível observar a formação do
arco recifal. As bordas do platô recifal encontram-se bastante fragmentadas em resposta à
presença de organismos bioerosionadores (Fig. 3).

22
Figura 2- Sistema de recifes de Coroa Vermelha.
Na borda externa superior do platô recifal (Fig. 3) há dominância dos antozoários
coloniais Palythoa caribaeorum e Zoanthus sociatus e de magroalgas clorofíceas (Caulerpa
racemosa) e rodofícias calcáreas (Jania sp), além da presença de organismos
bioerosionadores, como o ouriço-negro (Echinometra lucunter), propiciando uma grande
heterogeneidade do substrato. Na porção inferior predominam os hidrocorais do gênero
Millepora, que compõem uma franja quase contínua e, abaixo de 1m, os corais pétreos do
gênero Mussismilia. Nas suas bordas interna e externa há o desenvolvimento de pradarias
extensas dominadas pela fanerógama Halodule wrightii Aschers.

23
Figura 3: Imagem retratando a borda externa ( próxima ao mar) do recife de Coroa Vermelha.
O tempo de emersão do platô varia entre 6 e 9 horas por dia (Fig. 5). Sua porção
interna, mais elevada, apresenta densa cobertura macro e microfitobêntica, além da presença
de extensos agregados de Palythoa caribaeorum e Zoanthus sociatus cobrindo os esqueletos de
coral (Costa et al., 2001). Os corais hermatípicos Favia gravida e Siderastrea stellata estão
presentes no interior de poças do platô em vários estágios de crescimento. Nas últimas três
décadas o atrativo turístico tem promovido o crescimento da economia e da atividade de
construção civil no litoral de Santa Cruz de Cabrália. Devido à facilidade de acesso por um
tômbolo durante a maré baixa (Fig. 4), atualmente o recife de Coroa Vermelha encontra-se
severamente impactado pelo trânsito de turistas, remoção de organismos e assoreamento.

24
Figura 4- Vista geral do tômbolo que facilita o acesso de pessoas ao platô recifal de Coroa vermelha.
Além disso, a construção do assentamento para a aldeia indígena de Coroa Vermelha
gerou um aumento no volume de esgoto e lixo. É provável que a água subterrânea já esteja
contaminada pelas fossas sépticas e pelo lixiviamento do lixo disposto a céu aberto. Um canal
de escoamento fluvial, localizado a cerca de 30 metros do trecho de recife estudado, apresenta
águas fortemente eutrofizadas que podem estar influenciando o aporte de nutrientes para o
recife.
Figura 5: Vista geral do platô do recife de Coroa Vermelha.

25
4. MATERIAIS E MÉTODOS
Caracterização biológica do ambiente e estabelecimento das estações
Censos visuais foram realizados para registro das espécies em dezembro de 2004,
junho de 2005, agosto 2005 e setembro 2005. As amostragens foram realizadas no platô recifal
(porções central, norte e sul) e na borda até cerca de um metro de profundidade. Como são os
principais construtores da estrutura dos recifes foram enfatizados, neste trabalho, os hidróides
calcáreos e os corais pétreos. Após esta etapa foi delimitada a porção central do arco recifal
para o detalhamento da distribuição espacial dos principais organismos macrobênticos e o
levantamento das características hidroquímicas, cálculo das taxas de calcificação e do
metabolismo líquido das poças de maré.
Diagnose dos hidróides calcáreos e dos corais pétreos
Durante a realização do levantamento para caracterização do ambiente exemplares
foram coletados para depósito na coleção de referência do Laboratório de Oceanografia
Biológica da Universidade Estadual de Santa Cruz e envio a especialistas para
confirmação/identificação.
Para a distribuição espacial foi empregada uma modificação do método pontual como
sugerido em LOYA (1978), adaptado por SEGAL & CASTRO (2000). Dois transectos
perpendiculares à costa foram estabelecidos na porção central do platô. Os pontos para registro
foram estabelecidos a cada 0,25m, perfazendo um total de aproximadamente 200 pontos em
cada transecto. Em cada ponto foram registrados os organismos macrobênticos presentes, com
ênfase nos corais pétreos e hidróides calcáreos.
Caracterização hidroquímica do ambiente recifal
Foram coletadas amostras de água das poças a cada 10 metros, ao longo de um
transecto estabelecido entre a borda interna do tômbolo e a borda externa do recife, nos
períodos de dezembro de 2004, junho de 2005, agosto 2005 e setembro 2005. Esta
amostragem foi repetida após um intervalo de tempo, no trajeto de retorno. Adicionalmente

26
foram coletadas amostras de água dos dois lados do tômbolo (Norte e sul), e da água do mar
na borda externa.
As amostras foram recolhidas com uma seringa de 60mL, e filtradas com um suporte
Swinnex Millipore e filtros de fibra de vidro tipo GF/C de 25mm de diâmetro. O filtrado foi
preservado para determinação da concentração de nutrientes inorgânicos dissolvidos.
Durante as amostragens foram medidos a temperatura, condutividade, salinidade, pH
(escala NBS) e oxigênio dissolvido com medidores digitais portáteis. Amostras para análise da
alcalinidade total foram coletadas em seringas de 20mL, previamente lavadas com solução
saturada de cloreto de mercúrio, de forma a cessar a atividade biológica (DOE, 1994).
No filtrado foram realizadas análises de nitrato, nitrito, amônia, fosfato e silicato
utilizando-se método espectrofotométrico (Grasshoff et al., 1983). A alcalinidade total foi
determinada através de titulação com HCl 0,0100 M e cálculo da Função de Gran utilizando-
se o programa ALCAGRAN.BAS (Carmouze, 1994). A concentração de carbono inorgânico
total (TCO2) e demais variáveis do sistema carbonato foi calculada com base nos dados de pH,
temperatura, salinidade, alcalinidade total e nutrientes, utilizando-se o programa
CO2SYS.EXE (Lewis & Wallace, 1994). Neste programa foram empregadas as constantes de
Dickson (KSO4-2) e Merhbach (K1 e K2) replotadas por Dickson & Millero.
As taxas de calcificação e de metabolismo líquido (Produção primária bruta –
Respiração; P-R) nas poças durante as campanhas foram determinadas utilizando-se a variação
da alcalinidade total e da concentração de TCO2. As taxas de calcificação foram calculadas
através da técnica da anomalia da alcalinidade (Smith & Kinsey, ),
2. totalAlc
Calc =
e o metabolismo de acordo com a equação
.2 CalcTCORP −∆=−

27
Nestes cálculos foram utilizadas as alcalinidades e TCO2 médias obtidas em cada
percurso do transecto. As taxas de troca com a atmosfera foram desprezadas devido ao
pequeno intervalo de tempo transcorrido entre cada transecto.
Cálculo das taxas de calcificação e do metabolismo líquido das poças de maré
Foram selecionadas três poças localizadas próximas umas das outras e possuindo um
maior tempo de exposição ao ar. Nestas poças foram feitas coletas em dois momentos, após a
emersão do recife (tempo inicial) e antes da imersão (tempo final).
Para cada momento foram coletados dados de salinidade, temperatura, condutividade,
pH e oxigênio dissolvido com aparelho digital e amostras em seringas de 20mL para análise
de alcalinidade.
Em cada poça foi coletada amostra de água para análise de nitrato, nitrito, amônia e
fosfato.
A atividade metabólica das poças de maré foi inferida através da variação temporal da
concentração de O2 e CO2 total, sendo esta última calculada a partir da alcalinidade, pH e
salinidade (Carmouze, 1994).
Após o término dos experimentos, as poças foram delineadas quanto à altura, largura e
profundidade para posterior construção do perfil e estimativa do volume. O número de
medidas tiradas variou em função do tamanho das poças. Faixas foram percorridas da
superfície até o fundo obedecendo às reentrâncias da poça, utilizando em cada faixa,
quadrados de 25 X 25 cm para estimar o percentual de cobertura das espécies sésseis
existentes.
Além disso, levantamentos qualitativos foram feitos nas poças utilizando o método de
pontuação (Silva, 2004) para análise da distribuição e composição da biota.

28
5. RESULTADOS E DISCUSSÃO Diagnose dos hidróides calcáreos e dos corais pétreos
Os corais pétreos e os hidrocorais são de particular importância para os recifes de
coral, por formarem parte da estrutura física destes ambientes. Os recifes brasileiros apresentam
um número relativamente pequeno de espécies de corais-pétreos (somente 17). Apesar de sua
taxonomia ainda necessitar de maiores estudos, outras regiões do mundo apresentam,
nitidamente, uma maior diversidade. No Caribe, por exemplo, podem ser encontradas mais de
cerca de 70 espécies.
No Brasil, destaca-se a importância do Banco dos Abrolhos, o qual abriga todas as
espécies de corais-pétreos recifais até hoje descritos para o Brasil. Os recifes de outras áreas do
Brasil possuem apenas um sub-conjunto destas espécies.
A fauna brasileira de corais, e a do Sul da Bahia em particular, possui diversas espécies
endêmicas. Como os recifes ainda são pouco estudados, acredita-se que neles possam ainda
ocorrer diversas outras espécies ainda desconhecidas. Por exemplo, espécies como os octocorais
Plexaurella regia e Muricea flamma, que atingem mais de 30cm de altura e são muito visíveis
no ambiente, só foram descritos na década de 90.
Diversas espécies de corais-pétreos ou corais-verdadeiros de nossos recifes ocorrem
unicamente no Brasil (Veron, 1995) ou, mais especificamente, apenas na Bahia. O gênero
Mussimilia só possui espécies "vivas" no Brasil, com Mussimilia braziliensis ocorrendo apenas
na costa baiana. Este gênero é registrado como fóssil na Europa (bacia do Mediterrâneo) e na
América do Norte (Flórida). Favia leptophylla também só é encontrada na Bahia, enquanto que
Porites branneri e Siderastrea stellata só foram encontradas em águas brasileiras. Favia gravida
foi registrada no Atlântico Sul (Laborel, 1974).
No platô do Recife de Coroa Vermelha foram registradas dez espécies de corais
escleractíneos distribuídos em cinco famílias. Isto representa 58,8% das espécies descritas
para a costa brasileira. A família com maior número de espécies foi Poritodae com quatro
espécies em três gêneros. Foram ainda registradas duas espécies de hidróides calcáreos
pertencentes a uma família.

29
Filo Cnidaria Verril, 1865
Classe Hydrozoa
Ordem Capitata
Família Milleporidae
Millepora alcicornis Linnaeus, 1758
Millepora sp
Classe Anthozoa Ehrenberg,1834
Subclasse Hexacorallia Haeckel, 1866
Ordem Scleratinia Bouner, 1900
Subordem Fungiina Verril, 1865
Família Agaricidae Gray, 1847
Agaricia agaricites (Linnaeus,1758)
Família Siderastreidae Vaughan and Wells, 1943
Siderastrea stellata Verrill,1868
Família Poritodae gray, 1842
Porites astreoides Lamark,1816
Porites branneri Rathbun,1887
Favia gravida, Verril, 1868
Montastrea caverrnosa (Linnaeus, 1767)
Família Mussidae Ortmann, 1890
Mussismilia barziliensis Verril, 1869
Mussismilia harttii (Verril, 1902)
Mussismilia hispida (Verril, 1902)
Família Meandriniidae, Gray, 1857
Meandrina braziliensis (Milne Edwards & Haime, 1849)

30
Diagnose do grupo
Millepora alcicornis Linnaeus, 1758
A forma das colônias apresenta, geralmente, forma de crescimento ramificado. Os
ramos possuem diferentes espessuras. Textura da superfície lisa. Gastróporos rodeados por
dactilóporos, ambos com diâmetros relativamente grandes em relação às outras espécies do
gênero (Amaral, 1997). Podem ocorrer colônias incrustantes, formando prolongamentos
achatados (Hetzel & Castro, 1994; Castro, 1994).
É freqüente nas bordas e laterais dos recifes, desenvolvendo-se muito bem na face de
maior incidência de ondas. Coloração padrão marrom clara a marrom escura (Hetzel & Castro,
1994). Distribuição no Brasil: desde o Parcel do Manuel Luís (Ma) até a costa do Rio de
Janeiro (Laborel, 1969/70; Leão, 1986; Hetzel & Castro, 1994; Amaral et al., 2000).
No Recife de Coroa Vermelha foram observadas colônias arborescentes bem
desenvolvidas com coloração creme claro / marrom claro ocupando a maior parte da franja
externa do arco.
Millepora sp
A forma das colônias apresenta, geralmente, forma de crescimento ramificado. Os
ramos possuem diferentes espessuras. Textura da superfície levemente áspera. Gastróporos
rodeados por dactilóporos, ambos com diâmetros relativamente menores em relação à M.
alcicornis.
No Recife de Coroa Vermelha foram observadas algumas colônias arborescentes
pouco desenvolvidas com coloração cinza claro ocupando a borda externa do recife abaixo de
1m de profundidade.

31
Agaricia agaricites (Linnaeus, 1758 (Fig.6)
A forma das colônias varia de acordo com a disponibilidade de luz e com o
batimento das ondas. Sua superfície tem aparência irregular acompanhando o substrato. Os
cálices formam vales fundos, curtos e irregulares, de forma colonial, estrutura foliácea
incrustante com superfície bifacial. Colônias com até 15cm de diâmetro. Cálices com tamanho
inferior a 1,6mm. Coloração entre o marrom-claro e o marrom-esverdeado (Leão,1986; Neves
et al., 2002).
Espécie comum em águas rasas. Distribuição no Brasil: Parcel do Manuel Luís – Ma;
Nova Almeida - ES, Atol das Rocas - RN, Fernando de Noronha - Pe, Abrolhos - Ba. (Almy &
Carrión–Torres, 1963, Gorreus & Wells, 1967, Olivares, 1973, Belém et al., 1986, apud
Neves, 2002; Leão, 1986; Hetzel & Castro, 1994).
No recife de Coroa Vermelha a espécie foi rara sendo observadas apenas três
colônias em fendas da borda externa do recife.
Figura 6: Foto de um espécime de Agaracia agarites coletado no recife de Coroa Vermelha
Siderastrea stellata Verrill, 1868 (Fig. 7a)
Forma colônias maciças, de forma esférica e achatada, não ultrapassando 15cm de
diâmetro. Apresenta coralitos cerióides de crescimento extracelular. Os cálices podem
compartilhar a mesma parede (Vaughan & Wells, 1943; Veron, 2000). A coloração é variada,

32
sendo comuns os tons de amarelo, rosa e azul. A espécie é endêmica da costa brasileira, sendo
mais abundante ao norte da Foz do Rio São Francisco (Al / Se) e podendo formar construções
recifais monoespecíficas como as observadas na costa do Rio Grande do Norte. Embora
apresente ampla distribuição no sul da Bahia, as colônias de S. stellata têm papel secundário
na formação dos recifes baianos. São comuns em poças de maré, no topo dos recifes e em
regiões submersas rasas e bem iluminadas (Leão,1994; Neves et al., 2002). Variações
morfológicas permitem distinguir exemplares de águas rasas daqueles provenientes de
profundidades superiores a 5m. Segundo Laborel (1970), as colônias situadas em águas muito
rasas são geralmente pequenas com cálices apresentando entre 2-3mm de diâmetro, pólípos
profundos e quarto ciclo de septo incompleto enquanto as presentes em maiores profundidades
possuem cálices com diâmetro em torno de 5mm, pólipos pouco profundos e septos soldados
ao ciclo anterior. Neves (2004), descreve duas formas para a espécie, denominadas como
forma A e B. Distribuição no Brasil: do Parcel do Manuel Luís – Ma até o Rio de Janeiro,
Atol das Rocas - RN, Fernando de Noronha – Pe e Arquipélago de Abrolhos – Ba (Laborel,
1969/70; Leão, 1986; Belém et al., 1986 apud Neves et al, 2002; Hetzel & Castro, 1994;
Amaral et al., 2000b).
No Recife de Coroa Vermelha foram observadas colônias com variadas formas e
tamanhos. A coloração predominante foi rósea a vinho. Ao longo do período de trabalho
foram observadas colônias branqueadas. A espécie foi dominante nas poças de maré presentes
no platô (Fig. 7b).

33
Figura 7: Fotos tiradas nos Recifes de Coroa Vermelha. (a) Imagem de um espécime de Siderastea stellata em detalhes.(b) Imagem de uma poça de maré tirada com um quadract de 0,25X0,25m retratando a abundância de Siderastea Stellata.
Porites astreoides Lamark, 1816.
As colônias apresentam forma maciça. Podem ser esféricas ou incrustantes com
nódulos devidos às diferenças de nivelamento do substrato. Os coralitos têm aparência de
favos de mel. Coloração tipicamente amarelada, podendo ser amarronzada ou esverdeada.
Encontra-se no infralitoral, principalmente no topo ou na lateral dos recifes, geralmente
inseridas em fendas ou valas, parecendo tolerar condições de turbidez (Leão, 1994; Leão,
1986; Neves et al. 2002).
Distribuição no Brasil: desde o Parcel Manuel Luís – Ma até a costa baiana, Atol das
Rocas - RN, Fernando de Noronha - Pe, Abrolhos - Ba. (Belém et al, 1986, Hetzel & Castro,
1994; Echeverría et al., 1997; Neves et al, 2002 Amaral et al., 2000b; Leão, 1986).
No recife de Coroa Vermelha a espécie foi pouco comum sendo observada em fendas
da borda externa do recife. As maiores colônias registradas apresentavam cerca de 20cm de
diâmetro encontrando-se, freqüentemente, infestadas por cirripédios.
b a

34
Porites branneri Rathbun, 1887.
As colônias apresentam formas maciças, incrustantes e nodulares. Os coralitos
apresentam crescimento extratentacular, sem columelas. Os cálices são minúsculos, com
diâmetros de 1,0 a 1,5 milímetros. Apresentam coloração marrom-escura. São mais comuns
no infralitoral, principalmente no topo ou lateral dos recifes, geralmente inseridos em fendas
ou valas. Têm preferência por áreas bem iluminadas (Leão, 1994; Neves et al. 2002).
Distribuição no Brasil: Fernando de Noronha – Pe até o norte do Rio de Janeiro.
No recife de Coroa Vermelha a espécie foi pouco comum sendo observada em fendas
da borda externa do recife.
Favia gravida Verril, 1868.
Colônias maciças de forma ovóide ou esférica, medindo, em geral menos de dez
centímetros de diâmetro. Cálices assimétricos, arredondados, ovais ou alongados e
meandrinizados. Os septos são compactos e denteados. Apresentam crescimento
intratentacular. Coloração variando entre o marrom-claro e amarelo-claro (Leão, 1994; Leão,
1986; Neves et al., 2002).
Espécie endêmica do Brasil. Distribuindo-se do Parcel Manuel Luís – Ma até o Rio
de Janeiro, Atol das Rocas - RN, Fernando de Noronha – Pe e Arquipélago de Abrolhos – Ba
(Laborel, 1969/70; Leão, 1986; Belém et al., 1986 apud Neves et al, 2002; Hetzel & Castro,
1994; Amaral et al., 2000b).
Esta espécie é também considerada como muito resistente à variação das condições
ambientais, como temperatura, salinidade e turbidez da água, sendo comum encontrá-la nas
poças de maré e em locais bem iluminados nas paredes dos recifes, o que pode explicar a
acentuada variação nas características morfológicas como já reportado por Amaral (1991).
No Recife de Coroa Vermelha a espécie foi a segunda mais abundante,
predominando pequenas colônias de formato oval a esférico, com pólipos meandróides e

35
coloração marrom clara. Ao longo do período de trabalho foram ainda observadas colônias
branqueadas e colônias recobertas por cianobactérias (Fig. 8).
Figura 8: Foto de uma colônia de Favia grávida dominada por cianobactérias
Montastrea cavernosa (Linnaeus, 1767)
O formato das colônias varia de acordo com o local e profundidade, podendo ser
maciças, ovóides, esféricas, foliáceas (nas bordas dos chapeirões) ou em forma de lâmina
incrustante. Podem chegar a atingir dois metros de diâmetro. Os cálices são regulares e
apresentam pardes laterais espessas, conferindo à colônia a forma placóide. O diâmetro varia
entre 7 e 10mm. A teca externa e a columela tem altura de aproximadamente 2,2mm. O
número de septos por coralito atinge o valor máximo de 50 (Amaral, 1991). Coloração
geralmente verde ou parda. Preferência por águas claras e calmas (Leão, 1994; Leão, 1986;
Neves et al. 2002, Laborel, 1970).
Distribuição no Brasil: litoral de Pernambuco até o Espírito Santo, Atol das Rocas –
RN, Fernando de Noronha - Pe, Arquipélago de Abrolhos - Ba (Leão, 1986).

36
No recife de Coroa Vermelha a espécie foi pouco comum sendo observadas
pequenas colônias em fendas da borda externa do recife.
Mussismilia barziliensis Verril, 1869.
O gênero possui características arcaicas, próximas de espécies da idade Miocênica
encontradas em fósseis na bacia do Mediterrâneo. Nesta espécie a forma das colônias é
maciça, arredondada ou esférica, com pólipos lamelares de crescimento intracelular formando
sempre colônias que podem atingir um metro de diâmetro. Os coralitos são compactos,
serióides, com formato irregular, e 8 a 10 mm de diâmetro (Laborel, 1969). O tipo de
crescimento dá a colônia o aspecto de cogumelo (coral cérebro), uma vez que a base da
colônia, geralmente mais fina, é constituída apenas pelo esqueleto calcário, estando o tecido
vivo presente apenas na parte superior da colônia. A base se torna freqüentemente mais fina
devido à ação dos organismos bioerosivos, como as esponjas e os bivalves, que destroem as
camadas do esqueleto calcário dos corais. Coloração que varia entre o cinza amarelado e o
verde podendo apresentar tons misturados dessas cores. São encontrados, principalmente, em
áreas bem iluminadas, no topo e borda dos recifes (Leão, 1994; Neves et al. 2002).
Distribuição no Brasil: Endêmico da costa baiana. Encontrada principalmente no
Arquipélago de Abrolhos (Leão, 1986; Segal & Castro, 2000).
No recife de Coroa Vermelha a espécie foi comum sendo observadas colônias apenas
nas bordas externa do recife, abaixo de 1m de profundidade.
Mussismilia harttii (Verril, 1902) (Fig. 9).
Colônias ramificadas, facelóides, com isolamento de pólipos na extremidade dos
ramos. Em geral a conexão basal encontra-se morta (Veron, 2000). Os coralitos tubulares

37
variam de 15 a 20mm de diâmetro. Três diferentes variações morfólogicas são descritas por
Laborel (1970): laxa, intermédia e confertifolia. A variedade laxa possui cálices bastante
separados sendo encontrada em ambientes de águas mais calmas. Já a variedade confertifolia
apresenta os cálices pouco separados e é comumente encontrada em águas mais agitadas
enquanto a variedade intermédia é mais freqüente em águas com características intermediárias.
Coloração ventre tons de cinza, amarelo e verde. Preferem áreas rasas, estando comumente
presente nas bordas dos recifes. Tolera águas túrbidas. Esta espécie apresenta grande
importância para a formação dos recifes em Pernambuco sendo, aparentemente, substituída
por M. braziliensis no sul da Bahia. Pode formar grandes agrupamentos, tendo sido registradas
colônias com mais de quatro metros de comprimento e três metros de altura (Leão, 1994;
Neves et al., 2002).
Distribuição no Brasil: Endêmica. Atol das Rocas - RN, Fernando de Noronha – Pe,
Arquipélago de Abrolhos – Ba (Leão, 1986; Barros et al., 2000).
No recife de Coroa Vermelha a espécie foi comum sendo observadas colônias apenas
nas bordas externa do recife, abaixo de 1m de profundidade. Foi observada a presença da
variedade laxa sugerindo o predomínio, na área, de condições de menor energia de ondas.
Figura 9: Imagem de uma colônia de Mussismilia harttii encontrada no Recife de Coroa Vermelha

38
Mussismilia hispida (Verril, 1902).
As colônias são maciças e hemisféricas com pólipos lamelares de crescimento
intracelular formando sempre colônias baixas, sendo comum que a base represente seu maior
diâmetro, que pode chegar a 40cm. Coralitos geralmente arredondados, com 10-15mm de
diâmetro, com paredes grossas (Laborel, 1969). Seus cálices são maiores, quando comparados
a M. braziliensis, e irregulares, porém de forma mais circular. Segundo Laborel (1970), o
diâmetro dos cálices e o número de septos são muito variáveis. Podem apresentar coloração
listrada, variando entre amarelo, creme, cinza-claro, e verde. Tem preferência por áreas bem
iluminadas, sendo muito comuns nas áreas rasas, topos e bordas dos recifes. A plasticidade
notória deve-se possivelmente às variações ambientais dos parâmetros físicos, os quais
influenciam na variação da relação simbiótica coral-zooxantela (Antônio-de–Souza & Amaral,
2000).
Espécie endêmica do Brasil distribuindo-se desde o Parcel Manuel Luís – Ma até São
Paulo, Atol das Rocas - RN, Fernando de Noronha - Pe, Arquipélago de Abrolhos – Ba
(Laborel, 1969/70; Leão, 1986; Belém et al., 1986; Hetzel & Castro, 1994; Leão, 1994;
Migotto, 1997; Neves et al., 2002 ).
No Recife de Coroa Vermelha foram observadas colônias com formato hemisférico,
pólipos verde acinzentados e grandes. A espécie esteve restrita à borda externa do recife,
sendo mais freqüente abaixo de 1m.
Meandrina braziliensis Milne Edwards & Haime, 1849)
Suas colônias apresentam formas ovaladas ou meandróides, com columelas
lamelares. Atingem, no máximo, 15cm de comprimento. Encontram-se preferencialmente em
águas calmas e claras, podendo ocorrer fixas ao substrato duro ou soltas na areia (Leão, 1994;
Neves et al., 2002).
Distribuição no Brasil: Ceará ao Espírito Santo.

39
No Recife de Coroa Vermelha foram observadas colônias com formato ovalado. A
espécie esteve restrita a área arenosa recoberta por uma pradaria dominada por Halodule
wrightii Aschers, localizada na borda interna do arco recifal.
Distribuição espacial das associações macrobênticas
As espécies de Siderastea stellata e Favia grávida cobriram em média 30% do
transecto realizado (Fig. 10 e 11). Dessa distribuição registrada Siderastea Stellata contribui
com 70%, sendo o coral pétreo dominante em todo o transecto.
As macroalgas foram mais abundantes da borda externa até o meio do recife, áreas
de menor exposição e com menor grau de soterramento (Fig. 12). As macroalgas crustosas
foram mais dominantes nas bordas e as macroalgas frondosas no meio do platô. Na borda
externa, existem constantes ações das ondas, para sobreviver tais condições é necessária uma
estrutura mais rígida como a encontradas em macroalgas crustosas.
O microfitobentos apesar da baixa cobertura foi encontrado distribuído por todo o
transecto analisado (Fig. 13).
Os poríferos e ascidiáceos, por serem organismos filtradores, foram encontrados no
meio do recife, área na qual não existe forte ação de ondas e pouco material em suspensão
(Fig. 14 e 15).
As áreas desnudas de cobertura animal e vegetal foram aumentando sua significância
á medida que se aproximava do tômbolo do recife (Fig. 16).

40
.
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Figura 10: Distribuição espacial das espécies de Siderastrea stellata e Favia gravida somadas ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Siderastrea stellata Favia gravida
Figura 11: Distribuição espacial das espécies de Siderastrea stellata e Favia gravida ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.

41
0
10
20
30
40
50
60
70
80
90
100
d ist ância da b ord a ext erna ( m)
f rondosas crustosas
Figura 12: Distribuição espacial das macroalgas crustosas mais as frondosas ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Figura 13: Distribuição espacial do microfitobentos ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
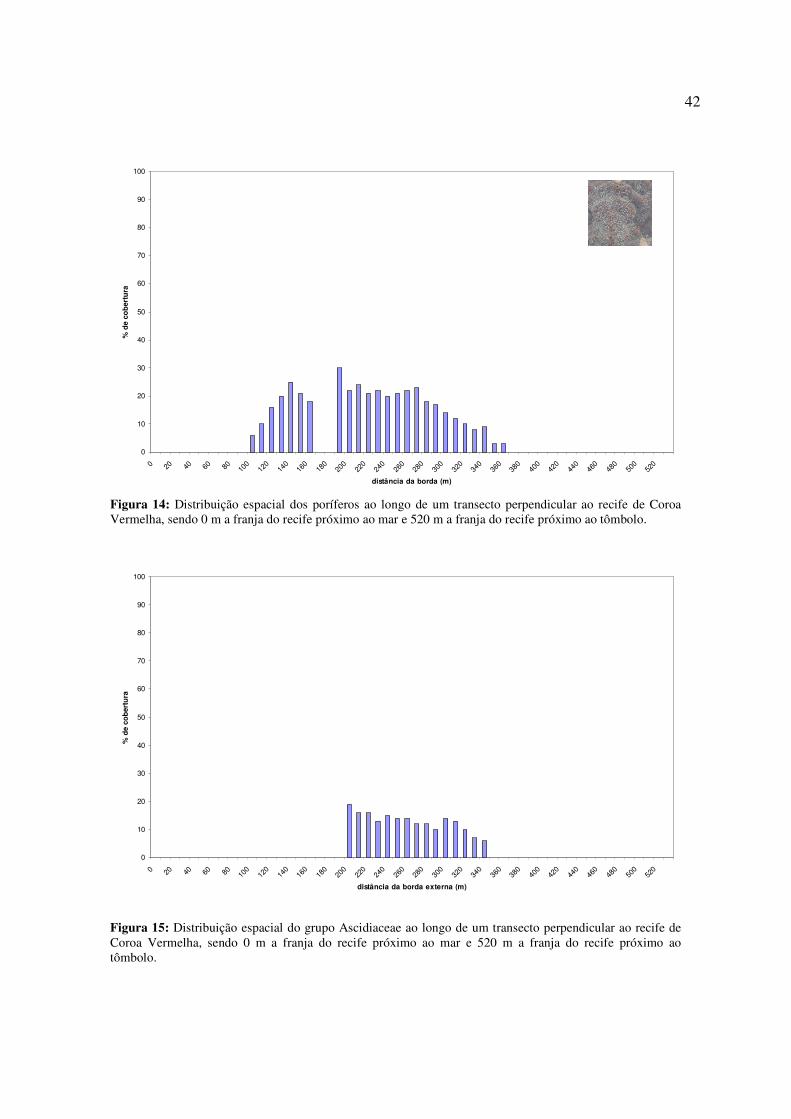
42
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda (m)
% d
e c
ob
ert
ura
Figura 14: Distribuição espacial dos poríferos ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Figura 15: Distribuição espacial do grupo Ascidiaceae ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
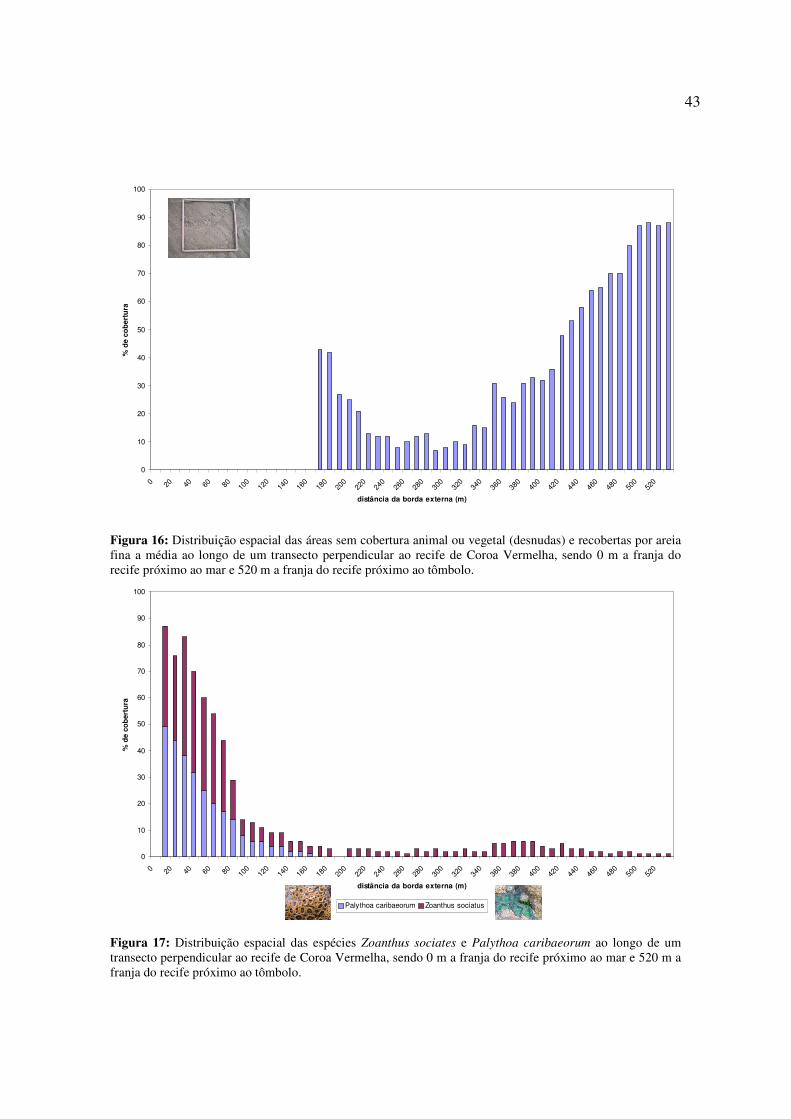
43
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Figura 16: Distribuição espacial das áreas sem cobertura animal ou vegetal (desnudas) e recobertas por areia fina a média ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Palythoa caribaeorum Zoanthus sociatus
Figura 17: Distribuição espacial das espécies Zoanthus sociates e Palythoa caribaeorum ao longo de um transecto perpendicular ao recife de Coroa Vermelha, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.

44
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
distância da borda externa (m)
% d
e c
ob
ert
ura
Corais pétreos Macroalgas Porifera marrom Ascidiacea negra filme microbêntico "desnudo"
Figura 18: Distribuição espacial dos principais componentes do recife de Coroa Vermelha ao longo de um transecto perpendicular ao mesmo, sendo 0 m a franja do recife próximo ao mar e 520 m a franja do recife próximo ao tômbolo.
O fato desta espécie se adaptar bem em áreas de sedimentação parece estar
relacionado com seu comportamento alimentar: além dos produtos derivados das zooxantelas,
ele utiliza matéria orgânica dissolvida no sedimento como fonte de nutrientes. Bem, espero de
alguma forma poder ajudá-los.
Áreas de concentração de ouriços-do-mar apresentam-se freqüentemente erodidas
pela ação escavadora destes animais (bioerosão), que raspam o substrato para remover as algas
que usam como alimento. Além disso, os ouriços-do-mar possuem um papel importante como
consumidores de algas dos recifes, que são competidoras por espaço com os corais.
A fauna de esponjas recifais é ainda pouco estudada. Muricy & Moraes (1998)
estudaram duas áreas aqui discutidas (Tamandaré, PE, na costa dos arrecifes, e a ilha oceânica
de Fernando de Noronha, PE). Estes autores elevaram o número de espécies registradas no
Estado de Pernambuco para 75. Entretanto, o grande número de novos registros e novas
espécies (34) mostra que a real diversidade de esponjas nestas áreas deve ser muito maior

45
(Muricy & Moraes, 1998). Em relação a Abrolhos, apenas 34 espécies foram incluídas na
listagem em anexo, obtidas dos trabalhos de Boury-Esnault (1973) e Mothes (1996) e de
exemplares depositados na Coleção de Porifera da UFRJ – todas fontes que não focam
adequadamente a área. Espera-se que a diversidade neste banco possa ser dez vezes maior que
este número (Hajdu et al., 1996).
As macroalgas - filamentosas, frondosas, articuladas e crustosas recobriram
praticamente toda a superfície do platô. Algas coralíneas crostosas foram muito abundantes
nos locais próximos às bordas, dominando o gênero Jania. Próximo ao funil há uma nítida
dominância de Hypnea musciformis e Acantophora spicifera. Além destas as espécies mais
abundantes e constantes foram Caulerpa racemosa e C. sertularioides, Halimeda spp.,
Dyctiota dichotoma, D. cervicornis, e D. mertensi, e Sargassum sp. Em alguns pontos onde a
profundidade era maior, o substrato duro encontrava-se recoberto por areia e algas frondosas
principalmente do gênero Padina.

46
Caracterização hidroquímica das poças
Temperatura
A temperatura nas poças oscilou entre 34,2 ºC em Novembro de 2004 e 26,5 ºC em
Junho de 2005 (Fig. 19). Com exceção deste último mês, em que a temperatura na borda
externa foi igual à máxima, os valores observados para os dois lados do tômbolo e na borda
externa foram bem inferiores aos medidos nas poças. Estes resultados demonstram o grau de
aquecimento da água retida nas poças durante grande parte do ano. Apenas no período de
“inverno” (Junho de 2005) o maior aporte de água doce pluvial e continental torna suas águas
mais frias que a água do mar adjacente. Os valores máximos estão compreendidos entre os
observados por Silva (2002) em poças de maré em um recife na Barra de Mamanguape (PB),
porém os valores mínimos são mais de 2 graus inferiores. Barradas (2005) relata em seu
estudo temperaturas entre 27,5 e 28,5 ºC em um ambiente recifal de Pernambuco (Porto de
Galinhas), porém em áreas de maior profundidade e comunicação com o mar. Os valores de
temperatura registrados por Souza (2002) para o mesmo ambiente foi de 26,9±0,3, valores
estes situados dentro do encontrado neste trabalho.

47
Figura 19 - Distribuição da temperatura nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
Salinidade
A salinidade nas poças evidencia a prevalência de água de mistura (AM; S < 37, T >
20 ºC; Fig. 20). Apenas na campanha de Novembro de 2004 a influência da água tropical (AT;
S > 37, T > 20 ºC) se mostra perceptível na faixa superior de salinidade. A evaporação líquida
também deve ser um fator preponderante na maior salinidade observada durante os outros
meses. Em Junho de 2005 a salinidade reflete o maior aporte da água doce, provocando uma
salinidade nas poças menor que a da água do mar adjacente. Na campanha de Setembro de
2005 observou-se uma variação de mais de 1 S entre as amostras. Ao longo das quatro
campanhas a salinidade varia cerca de 2 – 3 S entre as amostras do tômbolo, borda externa e
as poças. A distribuição da salinidade entre os meses de Junho – Setembro de 2005 demonstra
a grande variabilidade temporal e espacial (Setembro 2005) desta variável.
Silva (2002) não observou em seu trabalho uma relação entre a salinidade das poças
de maré e a sazonalidade. A salinidade em poças de maré recifais depende basicamente da

48
salinidade da água retida (características da água em situação de vazante), do balanço
precipitação/evaporação durante o período de emersão. Como a área de estudo deste trabalho
situou-se na barra de um estuário, a combinação dos efeitos da amplitude de maré, aporte
fluvial, balanço hídrico (P-E) e tempo de emersão ao longo do ano podem resultar neste
resultado aparentemente anômalo. Na região em torno do recife de Coroa Vermelha ocorrem
apenas pequenas drenagens superficiais (uma das quais altamente eutrofizadas) e o aporte
subsuperficial. Souza (2002) encontrou valores de salinidade médio (34,8±2,4) maiores do que
neste trabalho para todas as campanhas, evidenciando que nos últimos tempos a influência de
água doce sobre o sistema pode ter aumentado.
Figura 20 - Distribuição da salinidade nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.

49
pH
A faixa extrema de pH na água das poças foi superior à da água do mar (Pilson, 1998;
Wolf & Gladrow, 2001; Fig 21). Os valores médios de pH encontrados aqui foram
semelhantes aos encontrados para o mesmo ambiente há três anos atrás (Souza, 2002)
A mediana das campanhas de Novembro de 2004 e Setembro de 2005 situa-se bem
acima de seu limite superior. Estes resultados podem explicados pela ocorrência de altas taxas
de autotrofia líquida. Com exceção da campanha de Junho de 2005, na qual a mediana situa-se
um pouco abaixo do valor observado na borda externa, os valores de borda e tômbolo são bem
inferiores aos observados nas poças. Esta é mais uma evidência da influência de processos
biogeoquímicos ocorrendo no platô recifal e modificando uma variável sujeita a forte
tamponamento na água do mar devido ao sistema CO2-H2O (Pilson, 1998; Wolf & Gladrow,
2001).
Figura 21 - Distribuição do pH nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.

50
Oxigênio dissolvido
Os valores observados para o oxigênio dissolvido foram bastante elevados (Fig.22).
Estudos anteriores em recifes costeiros da região Nordeste já registraram concentrações
elevadas (Mucio, 1993). Este autor atribuiu estas concentrações a problemas nas técnicas
analíticas (oximetro e titulação de Winkler). Na realidade, as duas técnicas são inadequadas à
determinação deste gás dissolvido nas águas das poças, em virtude do alto grau de
supersaturação apresentado. Foram freqüentemente observadas bolhas formadas na superfície
do eletrodo, logo após a sua imersão. Este fato, aliado ao seu elevado coeficiente de troca com
a atmosfera (Carmouze, 1994) torna a concentração deste gás de pouco uso para a aplicação da
técnica de “água livre” (Lieth & Whittaker, 1975) para a avaliação do metabolismo aquático.
Figura 22 - Distribuição da concentração de oxigênio dissolvido nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.

51
Nutrientes inorgânicos dissolvidos
As concentrações de nutrientes inorgânicos dissolvidos foram bastante elevadas
(Figs.23,24,25,26 e 27), comparadas a outros ambientes recifais costeiros (Mucio, 1993;
Barradas, 2005), inclusive Taipús de Fora (BA; dados não publicados).
A mediana da concentração de amônio foi superior a 1,5 µM, exceto em Junho de
2005, quando se situou próximo ao limite de detecção do método. Em Junho de 2005 e
especialmente em Novembro de 2004 a concentração no tômbolo norte foi elevada, sugerindo
a influência do aporte da drenagem eutrofizada ao norte do tômbolo.
Figura 23 - Distribuição da concentração de amônio nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
A concentração de nitrito (Fig.24) foi especialmente alta nas poças em Setembro de
2005, assim como nas amostras do tômbolo e da borda externa. Nesta campanha a
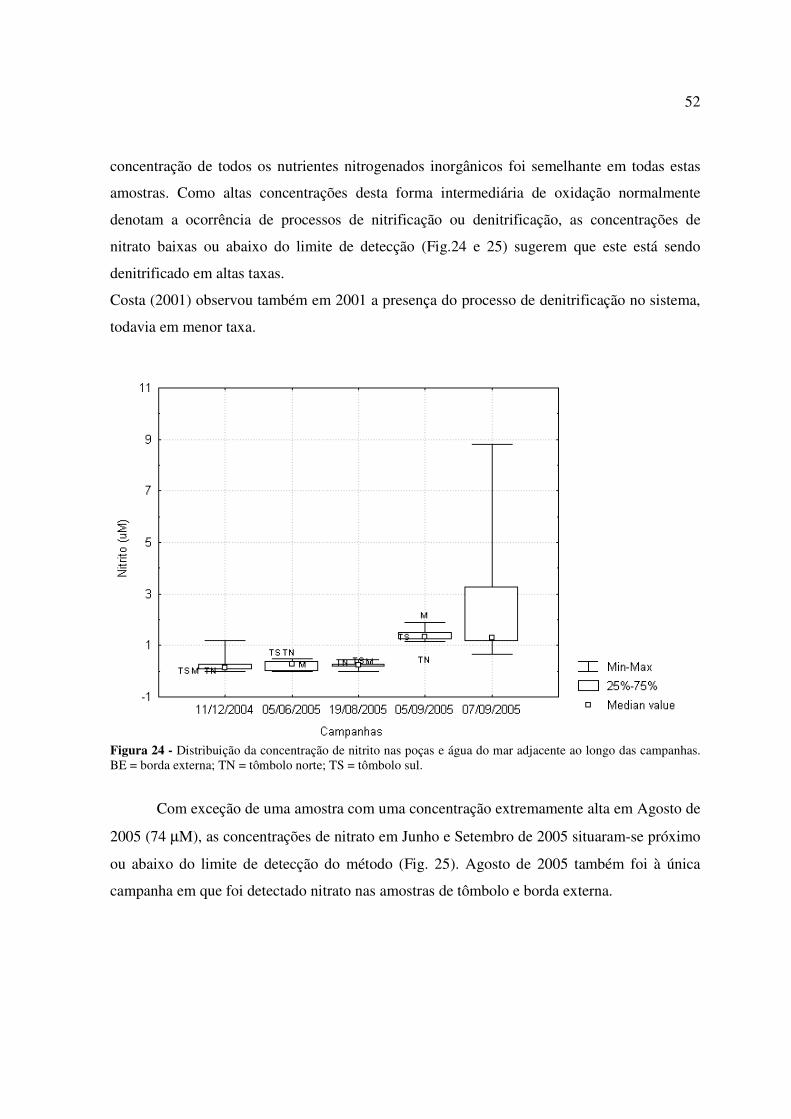
52
concentração de todos os nutrientes nitrogenados inorgânicos foi semelhante em todas estas
amostras. Como altas concentrações desta forma intermediária de oxidação normalmente
denotam a ocorrência de processos de nitrificação ou denitrificação, as concentrações de
nitrato baixas ou abaixo do limite de detecção (Fig.24 e 25) sugerem que este está sendo
denitrificado em altas taxas.
Costa (2001) observou também em 2001 a presença do processo de denitrificação no sistema,
todavia em menor taxa.
Figura 24 - Distribuição da concentração de nitrito nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
Com exceção de uma amostra com uma concentração extremamente alta em Agosto de
2005 (74 µM), as concentrações de nitrato em Junho e Setembro de 2005 situaram-se próximo
ou abaixo do limite de detecção do método (Fig. 25). Agosto de 2005 também foi à única
campanha em que foi detectado nitrato nas amostras de tômbolo e borda externa.

53
Figura 25 - Distribuição da concentração de nitrato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
Com exceção de poucas amostras, as concentrações de fosfato nas poças foram baixas
(Fig. 26). Apenas em Setembro de 2005 as concentrações nas águas em torno do tômbolo
foram mais elevadas que nas poças, porém bem abaixo do valor máximo observado (cerca de
30 µM).
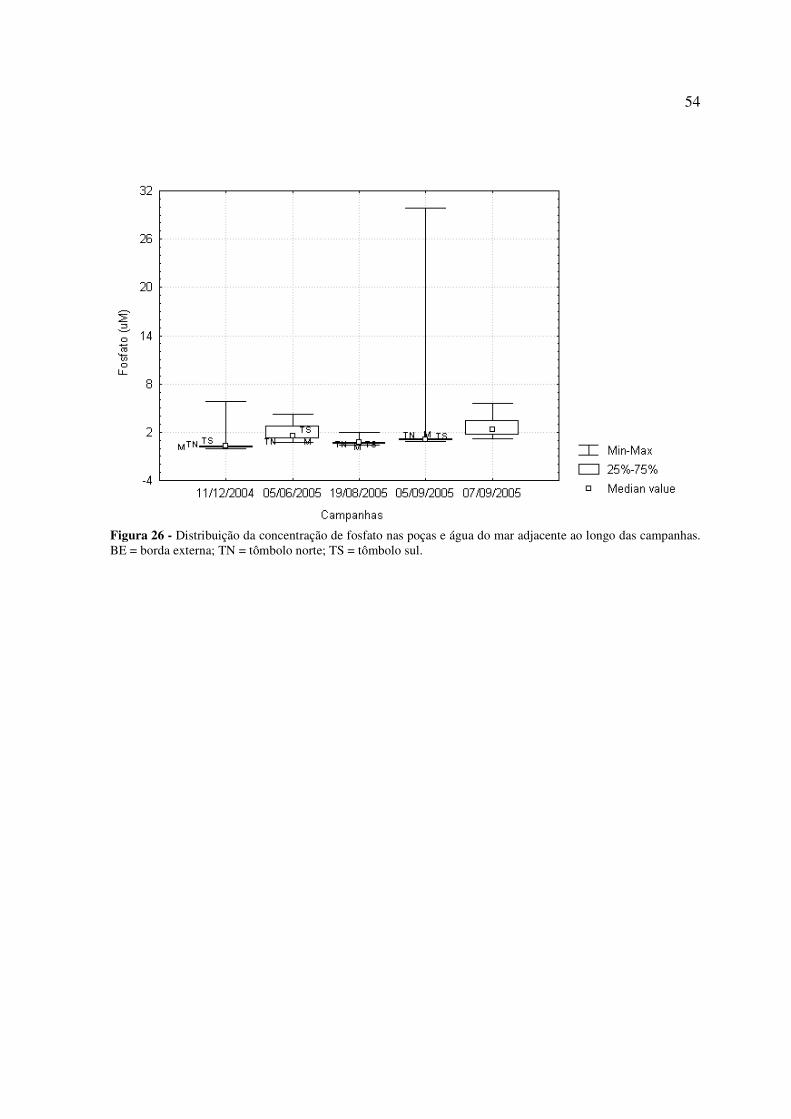
54
Figura 26 - Distribuição da concentração de fosfato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
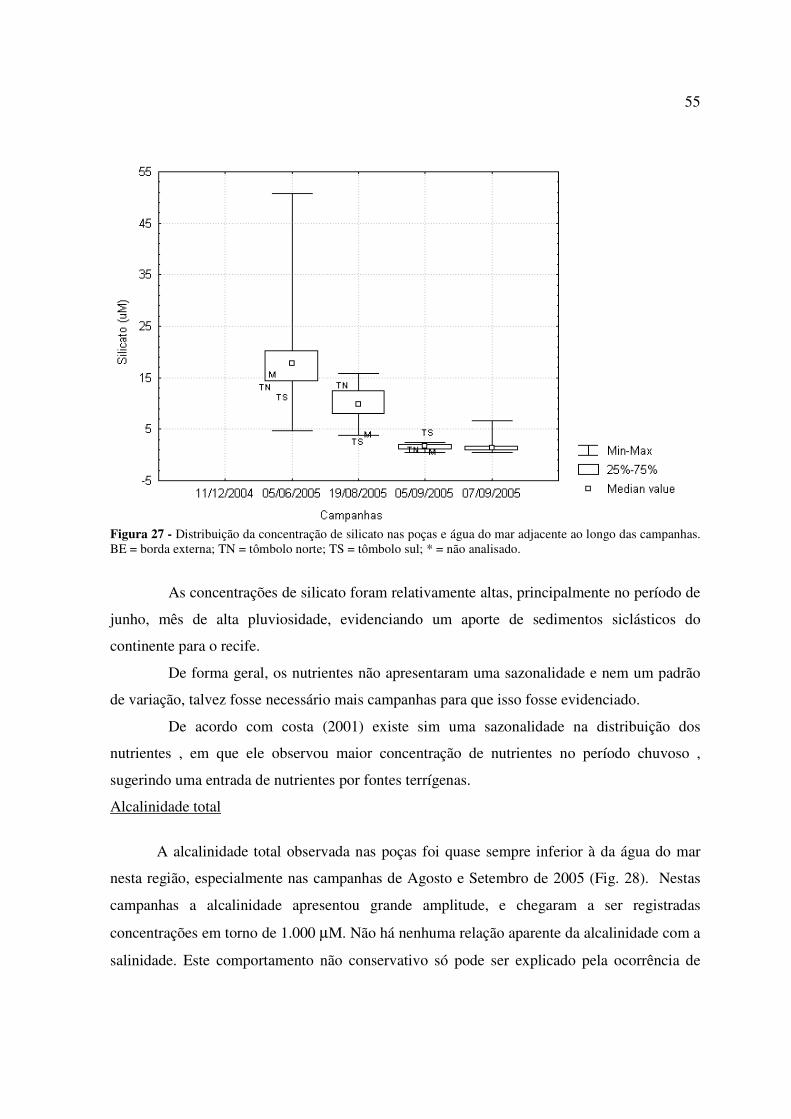
55
Figura 27 - Distribuição da concentração de silicato nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul; * = não analisado.
As concentrações de silicato foram relativamente altas, principalmente no período de
junho, mês de alta pluviosidade, evidenciando um aporte de sedimentos siclásticos do
continente para o recife.
De forma geral, os nutrientes não apresentaram uma sazonalidade e nem um padrão
de variação, talvez fosse necessário mais campanhas para que isso fosse evidenciado.
De acordo com costa (2001) existe sim uma sazonalidade na distribuição dos
nutrientes , em que ele observou maior concentração de nutrientes no período chuvoso ,
sugerindo uma entrada de nutrientes por fontes terrígenas.
Alcalinidade total
A alcalinidade total observada nas poças foi quase sempre inferior à da água do mar
nesta região, especialmente nas campanhas de Agosto e Setembro de 2005 (Fig. 28). Nestas
campanhas a alcalinidade apresentou grande amplitude, e chegaram a ser registradas
concentrações em torno de 1.000 µM. Não há nenhuma relação aparente da alcalinidade com a
salinidade. Este comportamento não conservativo só pode ser explicado pela ocorrência de
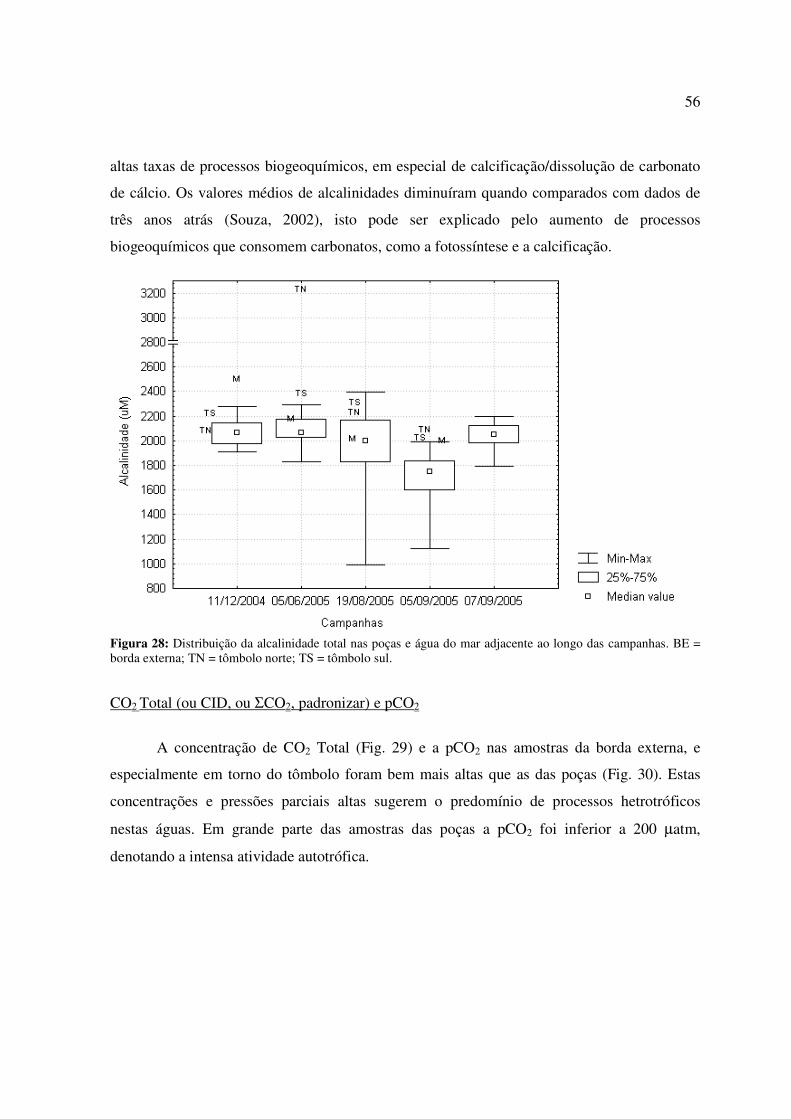
56
altas taxas de processos biogeoquímicos, em especial de calcificação/dissolução de carbonato
de cálcio. Os valores médios de alcalinidades diminuíram quando comparados com dados de
três anos atrás (Souza, 2002), isto pode ser explicado pelo aumento de processos
biogeoquímicos que consomem carbonatos, como a fotossíntese e a calcificação.
Figura 28: Distribuição da alcalinidade total nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
CO2 Total (ou CID, ou ΣCO2, padronizar) e pCO2
A concentração de CO2 Total (Fig. 29) e a pCO2 nas amostras da borda externa, e
especialmente em torno do tômbolo foram bem mais altas que as das poças (Fig. 30). Estas
concentrações e pressões parciais altas sugerem o predomínio de processos hetrotróficos
nestas águas. Em grande parte das amostras das poças a pCO2 foi inferior a 200 µatm,
denotando a intensa atividade autotrófica.

57
Figura 29: Distribuição da concentração de carbono inorgânico dissolvido (CID) nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
Figura 30: Distribuição da pressão parcial de dióxido de carbono (pCO2) na água das poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.

58
Saturação da Aragonita (Ωarag.)
O estado de saturação da aragonita foi mais elevado que a média da água do mar na
maior parte das amostras (Fig. 31), atingindo valores iguais ou superiores a 7 em todas as
campanhas. Estes resultados indicam condições termodinâmicas favorecendo fortemente à
calcificação. Na campanha de Agosto de 2005 foram observados os menores valores nas
poças, atingindo um mínimo de 2,2.
Figura 31: Distribuição do estado de saturação da aragonita (Ωarag.) nas poças e água do mar adjacente ao longo das campanhas. BE = borda externa; TN = tômbolo norte; TS = tômbolo sul.
Calcificação e metabolismo líquido
As taxas de calcificação e metabolismo líquido das poças foram mais elevadas em
Novembro de 2005 (Fig.32), atingindo valores equivalentes à cerca de 3 mmoles . m2 . hora-1.
Esta campanha também apresentou valores mais elevados de Ωarag. na maioria das poças
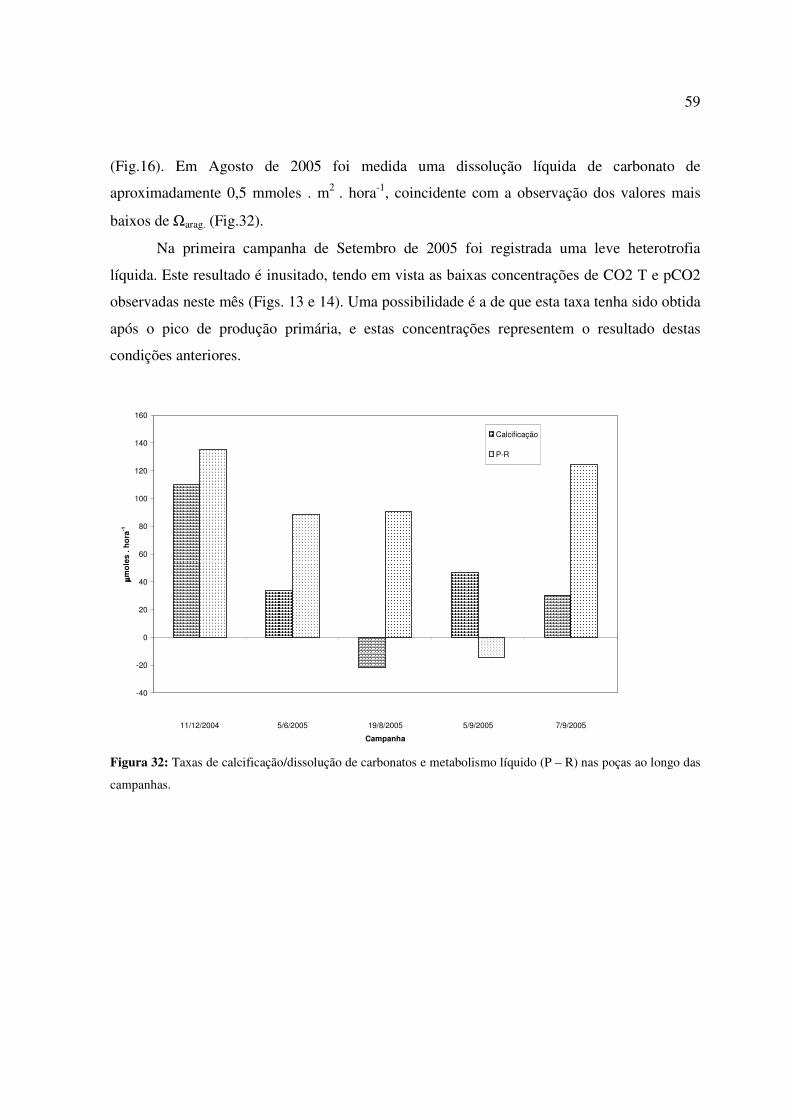
59
(Fig.16). Em Agosto de 2005 foi medida uma dissolução líquida de carbonato de
aproximadamente 0,5 mmoles . m2 . hora-1, coincidente com a observação dos valores mais
baixos de Ωarag. (Fig.32).
Na primeira campanha de Setembro de 2005 foi registrada uma leve heterotrofia
líquida. Este resultado é inusitado, tendo em vista as baixas concentrações de CO2 T e pCO2
observadas neste mês (Figs. 13 e 14). Uma possibilidade é a de que esta taxa tenha sido obtida
após o pico de produção primária, e estas concentrações representem o resultado destas
condições anteriores.
Figura 32: Taxas de calcificação/dissolução de carbonatos e metabolismo líquido (P – R) nas poças ao longo das
campanhas.
-40
-20
0
20
40
60
80
100
120
140
160
11/12/2004 5/6/2005 19/8/2005 5/9/2005 7/9/2005
Campanha
µµ µµm
ole
s .
ho
ra-1
Calcificação
P-R

60
6. CONCLUSÃO
As características hidroquímicas no sistema recifal variaram tanto espacialmente
quanto temporalmente, isto se deve altas taxas de processos biogeoquímicos do sistema . O
sistema responde rapidamente a mudanças nas condições fisico-quimicas. Além disso, dentro
sistema em poças próximas pode-se encontrar diferentes processos dominantes
(calcificação/dissolução e fotossistese/respiração), denotando que dentro deste sistema,
existem microcompartimentos formando uma teia de processos, ou seja, seriam necessárias
inúmeras amostragens ao longo de todo o recife para entender e descrever os processos
dominantes do sistema, ressaltando que essas amostras teriam que ser realizadas no mesmo
dia, devido à rápida resposta do sistema a mudanças temporais.
O grupo de maior dominância e distribuição no recife foram os corais pétreos com uma
maior contribuição das espécies de Sideratrea stelata e Favia gravida sendo sua distribuição e
abundancia influenciada pela presença de sedimentos e influências continentais.

61
7. REFERÊNCIAS BIBLIOGRÁFICAS AMARAL, F. D. 1991. Variação morfológica de Montastrea cavernosa (Linnaeus, 1767) do Brasil e do México e Favia gravida Verril, 1868 do Brasil (Cnidaria, Scleractinia). Dissertação de mestrado. Programa de Pós-Graduação em Zoologia, Museu Nacional do Rio de Janeiro, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 120pp AMARAL, F. D. 1997. Milleporidae (Cnidaria, Hydrozoa) do litoral brasileiro. Tese de doutorado. Instituto de Biociências, Universidade de São Paulo, 204p AMARAL, F. D. 1998. Recifes de corais de arenito do Brasil. Resumos do XIII Simpósio do CEBIMAR. São Sebastião, SP. Cebimar - Universidade de São Paulo, p. 05. AMARAL, F. D.; HUDSON, M.M & COURA M.F. 2000. New findings on corals end hydrocoral from the Marine state Park of the Manoel Luiz Parcel (Maranhão State). Abstracts of International Reef Symposium Bali. Camberra: Australian Agency for internacinal Development, p. 294. AMARAL, F. D.; HUDSON, M.M.; SILVEIRA, F. L.; MIGOTTO, A. E.; PINTO, S. M. & LONGO, L. L. 2002a. Cnidarians of Saint Peter and St. Paul Archipelago, Northeast Brazil. Proceedings of 9th International Reef Symposium 1: 567 - 572 . ANTONIO-DE-SOUZA, C.; Amaral, F.D. 2000. Variação intrapopulacional do coral Mussismilia brasiliensis (Cnidaria, Scleractinia) de Abrolhos (Ba). Resumos XXIII Cong. Bras. Zool. , p 46 BAK, R.P.M. & STEWARD-VAN ES, Y., 1980. Regeneration of superfitial damage in the Scleractinian corals Agaricia agaricides f. purpurea and Porites astreoides. Bull. Mar. Sci. Coral Gable, 30 (4): 883 - 887. BARRADAS, J. I. 2005. Avaliação das condições do ambiente recifal da Praia de Porto de Galinhas (Pe), com ênfase nos corais (Cnidaria : Anthozoa) e hidróides calcáreos (Cnidaria : Hydrozoa). Dissertação de mestrado. Programa de Pós-Graduação em Zoologia, Universidade Federal da Paraíba, 73pp + apêndices BARROS, M.M.L; CASTRO, C.L; PIRES, D.O & SEGAL, B. 2000. Coexistence of reef organisms the Abrolhos Archipelago, Brazil. Rev. Biol. Trop, 48 (4): 741 – 747. BELÉM, M. J. C.; ROHLFS, C.; PIRES, D. O. & CASTRO, C B. 1986. SOS Corais. Ciência Hoje, 5 (26): 34 – 42 BOSCOLO, H. K. 2000. Estimativa do comportamento reprodutor de zoantídeos (Cnidaria, Anthozoa, Hexacorallia) do Canal de São Sebastião - SP. Dissertação de

62
mestrado. Programa de Pós-Graduação em Zoologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, 168pp CALDERON, E. N., CASTRO, C. B. & PIRES, D. O. 2000. Natação, assentamento e metamorfose de plânulas do coral Favia gravida Verril, 1868 (CNIDARIA, SCLERACTINIA ). Boletim do Museu Nacional, Nova Série, Zoologia, Rio de janeiro, 429: 1 - 12. CARMOUZE, J-P. 1994. O Metabolismo dos Ecossistemas Aquáticos. Fundamentos teóricos, métodos de estudo e análises químicas. São Paulo, Ed. Edgard Blücher/FAPESP. 253 p. CASTRO, C B. 1994. Distribution and diversity of coral communities in the Abrolhos reef complex. CASTRO, C B.; SEGAL, B.; PIRES, D. O. & MEDEIROS, M. S. 2006. Distribution and diversity of coral communities in the Abrolhos reef complex. In: Dutra, G. F.; Allen, G. R.; Werner, T. & McKenna, S. A. (eds.) A Rapid Marine Biodiversity Assessment of the Abrolhos Bank, Bahia, Brazil. RAP Bulletin of Biological Assessment, 38: 19 – 39 + apêndice. CASTRO, C. B. 1986. Revisão sistemática dos Octacorallia (Coelenterata, Anthozoa ) dos recifes de Abrolhos, Ba, Brasil. Dissertação de Mestrado, Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro. p. 98. CORNELL , H.V. & R. H. KARLSON.1996. Species richness of reef building coral determinate by local and regions processes . J, Anim. Ecol 65:233-241. COSTA Jr., O.S.; ATRILL M.J.; PEDRINI, A.G.; DE-PAULA, J.C. 2001. Benthic Macroalgal Distribution in Coastal and Offshore Reefs at Porto Seguro Bay, Brazilian Discovery Coast. Proc. 9th Int. Coral Reef Symp. COUTINHO, R., VILLACA, R., MAGALHÃES, C., GUIMARAENS, M. A., APOLINARIO, M., MURICY, G, 1993. Influência antrópica nos ecossistemas coralinos da região de abrolhos, Bahia, Brasil. Acta Lepoldensis, 15 (1): 133 – 144. COUTO, E.C.G & LANA, P.C. da 2004. O contexto brasileiro do conhecimento da biodiversidade marinha In: COUTO, E.C.G. & ROCHA, G.R.A. (edts). Biodiversidade marinha na costa brasileira: o estado da arte. EDITUS. (in press). COUTO, E.C.G & SOUZA, M.F.L, 2003. Resposta de cinco espécies de Cnidaria: Anemonia sargassensis (Actinaria), Siderastrea stellata (Scleractinea), Isaurus tuberculatus, Zoanthus sociatus e Palythoa caribaeorum (Coralliomorpharia) a eventos de soterramento em poças de maré (Praia do Back Door, Ilhéus – Ba). Resumo da SBPC_ Sociedade Brasileira Para o Progresso da Ciência, Recife - Pernambuco, CDROM.

63
DOE. 1994. Handbook of Methods for the Analysis of the Various Parameters of the Carbon Dioxide System in Sea Water. Version 2, A.G. Dickson & C. Goyet, eds. ORNL/CDIAC-74. DUTRA, L. X. C., KIKUCHI, R. K. P., LEÃO, Z. M. A. N. 2003. Effects of sediment accumulation on coral reefs in Abrolhos, Bahia, Brazil. In: Anales del X Congreso Latinoamericano en Ciencias del Mar, 1 – 4. ECHEVERRIA, C. A.; PIRES, D. O.; MEDEIROS, M. S.& CASTRO, C B. 1997. Cnidarians of the Atol das Rocas, Brazil. Proceedings of 8th International Coral Reef Symposium 2: 443 – 446. FIGUEIREDO, M.A . O. 2000. Recife de corais ou recife de algas. Ciência Hoje, 28 (166): 74-76. GATTUSO, J-P.; ALLEMAND, D.; FRANKIGNOULE, M., 1999. Photosynthesis and calcification at cellular, organismal and community levels in coral: a review on interactions and control by carbonate chemistry. Amer. Zool., 39: 160-183. GATTUSO, J-P.; FRANKIGNOULE, M. ; BOURGE, I.; ROMAINE, S. & BUDDEMEIER, R. W. 1998. Effect of Calcium Carbonate Saturation of Seawater on Coral Calcification. Global and Planetary Change 18: 37 - 46. GATTUSO, J-P; FRANKIGNOULE, M. & SMITH, S. 1999. Measurement of Community Metabolism and Significance in the Coral Reef CO2 Source-Sink Debate. Proc. Natle. Acad. Sci. USA 23: 13017 - 13022. GRASSHOFF, K.; EHRARDT, M. KREMLING, K. 1983. Methods of Seawater Analisys. Weinhein : Verlag Chemie. 419 p. HETZEL, B. & CASTRO, C. B. 1994. Corais do sul da Bahia. Editora Nova Fronteira. 189p. HUSTON, M.A. 1985. Changes in coral growth with depth at Discovery Bay, Jamaica. Coral reefs, 4: 19 – 25. KAPLAN, E. H. 1982. Field guide to coral reef of the Caribbean and Florida. Boston: Houghton-Mifflin Company. 289 p. KIKUCHI, R.K.P. & LEAO, Z.M.A. N.1998. The effects of Holocene sea level fluctuation on reef development and coral community structure, northern Bahia, Brasil. An. Acad.Bras. Ci., 70 (2): 159 – 171. LABOREL, J.L., 1969/70. Madreporaires te hidrocoralliaires recifaux des côtwes bréssiliennes: systematique, ecologie, repartition verticale et geografie. Ann. Inst. Oceogr., Paris, 47: 171 - 229 .

64
LEAO, Z. M. A. N. 1986. Guia para identificação dos corais do Brasil. Programa de Pesquisa e Pós-Graduação em Geofísica. Instituto de Geociências – UFBA, Programa de Pesquisa e Pós Graduação em Geofísica, Salvador. 57p. LEÃO, Z. M. A. N., KIKUCHI, R. K. P., MAIA, M. P., LAGO, R. L. 1996. A catastrophic coral cover decline since 3,000 years B.P., northern Bahia, Brazil In: Proceedings of 8th. International Coral Reef Symposium, Panamá. 1: 583 – 588. LEÃO, Z.M. A. N. 1994. The coral reefs of Southern Bahia. In: Hetzel, B. & Castro C.B (eds). Coral of Southern Bahia. Ed. Nova fronteira, RJ, 159p. LEÃO, Z.M. A. N. 1999. Abrolhos - O complexo recifal mais extenso do Oceano Atlântico Sul. In: Schobbenhaus,C.; Campos,D.A.; Queiroz,E.T.; Winge,M.; Berbert-Born,M. (Edts.) Sítios Geológicos e Paleontológicos do Brasil. Publicado na Internet no endereço http://www.unb.br/ig/sigep/sitio090/sitio090.htm. LECLERCQ, N. & GATTUSO, J-P. 2002. Primary Production, Respiration, and Calcification of a Coral Reef Mesocosm under Increased CO2 Partial Pressure. Limnol. Oceanogr. 47(2): 558 -564. LECLERCQ, N; GATTUSO, J-P. & Jaubert, J. 2000. CO2 Partial Pressure Controls the Calcification Rate of a Coral Community. Global Change Biology 6: 329 - 334. LEWIS, J.B.,1974 Settlement and growth factors influencing the contagius distribution of some atlantic reefs coral. Proc. and. Int. Coral Reef Simp., Brisbane, 2: 201 – 206. LIRMAN, D; MAZNZELLO, D.; MACIÁ, S. 2002. Back from the dead: the resilence of Siderastrea radians to several stress. Coral Reefs, 21:291-292. LONGO, L. L. 1997. Repartição espaço-temporal da cnidofauna em arenito de praia em Santa Cruz (Aracruz – ES). Dissertação de mestrado. Programa de Pós-Graduação em Zoologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, 217pp. LONGO, L. L.; ARIA, M. C. & SCHLENZ, E. 2000. PCR amplification of 16S mithocondrial gene of Zoanthus sociatus (Zoanthidae : Anthozoa) using heterologous primers. Absracts of 9th International Coral Reef Symposium, 19. MIGOTTO, A. E. 1997. Anthozoan bleaching on the southeastern coast of Brazil in the summer of 1994. Proceedings of 6th International Conference Coel. Biol.: 329 – 335. NEVES, E. G; SILVEIRA, F. L; JOHNSSON, R; LONGO, L. L. 2002. Shallow-Water sleractinian corals and zoanthids from reefs of Coroa Grande, Pernambuco state, Brazil. Avanços em Biologia celular - Biociências, 10( 2): 127 – 145.

65
NYBAKKEN, J.W.1993. Marine biology – un ecological approach. HaperCDollins Colleger Publishers. 3º ed. 462p. PARSONS, T. R.; MAITA, Y & LALLI, C. M. 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. Pergamon Press,173 p. PIRES, D. O.; CASTRO, C. B.; MIGOTTO, A. E. & MARQUES, A. C. 1992. Cnidários bentônicos de Fernando Noronha, Brasil. Bol. Mus. Nac., Série Zoologia, 354: 1 – 21. REASER, J. K. ; POMERANCE, R. & THOMAS, P. O. 2000. Coral Bleaching and Global Climate Change: Scientific Findings and Policy Recommendations. Conservation Biology (14) 5, 1500-1511. RIVERA, G.L. 1994. Effects of water movement and sedimentation on the comunity structure of scleractinian corals, St. John, Virgin Islands (United States). Nerítica, 8 (1-2): 7 – 19 ROHLFS, C. & BELÉM, M. J. C. 1994. O gênero Zoanthus no Brasil. I – Caracterização e revisão anatômica de Zoanthus sociatus (Cnidaria: Zoanthiniaria: Zoanthidae). Iheringia, Série Zoologia, 77: 135 – 144. RUPPERT, E. E.; BARNES, R. D. 1996. Zoologia dos Invertebrados. São Paulo: Ed. Roca Ltda., 1029p. SANTANA, T.B. 2004. Determinação da Biomassa de Zooxantelas e Cianobactérias Euendolíticas nos corais do Gênero Siderastrea. Monografia de Especialização em Oceanografia, UESC. SEGAL, B.; CASTRO, B. 2000. Slope preferences of reefs corals (Cnidaria, Scleractinia) in the Abrolhos Archipelago, Brazil. Boletim do Museu Nacional , Nova Série, Zoologia, Rio de janeiro. 418 p. SHLESINGER, Y. & LOYA, Y.,1999. Larval development and surviveship in the corals Favia favus and Patygyra lamelina. Hydrobiol., 216/217 : 101 - 108. SILVA, A. M., CUEVAS, J.M.; VASQUES, R.R.; COUTO, E.C.G. 2003a. Distribuição espacial da comunidade macrofitobêntica no platô do recife de Taipús de Fora (Maraú, Ba, Brasil ). Resumos Expandido do X COLACMAR – Congreso Latinoamericano de Ciencias del Mar, San Jose – Costa Rica (resumo expandido). p 214. SILVA, A.M. 2004. Corais Pétreos (Cnidaria: Anthozoa: Hexacorallia) no ambiente recifal de Taipus de Fora, Península de Maraú, Bahia, Brasil. Monografia de Graduação. Curso de Ciências Biológicas - Bacharelado. UESC.

66
SILVA, A; CUEVAS, J.M; VASQUES, R.R & COUTO, E.C.G. 2003b. Echinodermata do ambiente recifal de Taipús de Fora, Península de Maraú, Sul da Bahia. Resumos da XIV Reunião Nordestina de Zoología. Maceió-Al. p. 259. SOUZA, M.F.L.; COIMBRA, C.S.; COUTO, E.C.G. 2002. Efeitos do aumento do CO2 atmosférico e da temperatura sobre o metabolismo de corais e algas calcáreas. Relatório final de projeto. UESC/Fundação O Boticário de Proteção à Natureza. 46 p. STEINER, A. Q. 2005. Cnidofauna de recifes emersos da área de proteção ambiental Costa dos Corais (estados de Pernambuco e Alagoas) e percepção do ambiente por moradores locais. Dissertação de mestrado. Programa de Pós-Graduação em Zoologia, Universidade Federal da Paraíba, 88pp + apêndices. STRICKLAND, J.D.H.; Parsons, T.R. 1972. A Practical Handbook of Seawater Analisys. Bull. Fish Res. Board Can. No. 167. 311 p. VAUGHAN, T. W. & Wells, J. W. 1943. Revision of the sub orders, families and genera of the Scleractinia. Spec. Pap. Geol. Soc. Am., 44: 1 – 363. VERON, J. E. N. 2000. Coral of the world. Australian Institute of Marine Science,473pp. YOUNG, P.S. 1988. Recent cnidarian – associated barnacles (Cirripedia, Balanomorpha) from the brasilian coast. Rev. Bras. Zool. 5 (3): 353 – 369