caldasia 23(1): 21-41 historical biogeography … · las distribuciones de estos taxones fueron...
TRANSCRIPT

Caldasia 23(1): 21-41
HISTORICAL BIOGEOGRAPHY OF THEASTERACEAE FROM TANDILIA AND VENTANIA
MOUNTAIN RANGES (BUENOS AIRES, ARGENTINA)
A nuestra amiga Pilar
JORGE CRISCI- V.Departamento Científico de Plantas Vasculares, Museo de La Plata, Paseo del Bosque s.n.,1900. La Plata, Argentina. [email protected]
SUSANA FREIRE-E.
Departamento Científico de Plantas Vasculares, Museo de La Plata, Paseo del Bosque s.n.,1900. La Plata, Argentina.
GISELA SANCHO
Departamento Científico de Plantas Vasculares, Museo de La Plata, Paseo del Bosque s.n.,1900. La Plata, Argentina.
LILlANA KATlNAS
Departamento Científico de Plantas Vasculares, Museo de La Plata, Paseo del Bosque s.n.,1900. La Plata, Argentina.
ABSTRACTTandilia and Ventania are the only systems ofmountain ranges situated in a grassysteppe or "pampas" in the political province ofBuenos Aires in Argentina. Tandiliaand Ventania have a high taxa diversity and endemicity. A historical biogeographicanalysis was carried out on the basis of distributional patterns of species andinfraspecific taxa of Asteraceae inhabiting Tandilia and Ventania in orderto establishthe relationships ofthese mountain ranges with other areas. Two methods were appliedfor the analysis: panbiogeography using the compatibility track method and parsimonyanalysis of endemicity (PAE). Thirteen areas were delimited for the study: SouthernNorth America and Central América, Southern Brazil, Uruguay, Pampa, Tandilia,Ventania, Chaco, Sierras Pampeanas, Sierras Subandinas, Mahuidas, Patagonia, Cen-tral Chile, and Northern Andes. The units of the study were 112 taxa inhabitingTandilia and Ventania (endernic, naturalized, and adventicious species were notincluded). Both methods connect southern Brazil, Uruguay, Pampa, Tandilia, Ventania,and Sierras Pampeanas, showing that the Asteraceae biota ofTandilia and Ventaniahave closer relationships with the biota ofthese are as rather than with that ofSierrasSubandinas, North Andean, Chaco, Patagonia, Mahuidas, and Central Chile. TheAsteraceae ofCentral and North America appear less related to Tandilia and Ventaniain regard ofthe rest ofthe areas. The evolution of Asteraceae in Tandilia and Ventaniaand related areas is hypothetized to have been affected mainly by Tertiary and

Asteraccae from Tandilia and Ventania
Quaternary geologic events. The discontinuous pattern found, occurring mostly inelevated areas, is explained principally by vicariance under dry conditions.
Key words. Asteraceae; biogeography; Tandilia; Ventania.
RESUMENLas sierras septentrionales, o de Tandilia y las australes, o de Ventania, caracteriza-das por su gran diversidad orgánica y su alto número de endemismos, están ubicadasen la planicie bonaerense de pastizales denominada "pampa". Con el objeto de esta-blecer las relaciones entre Tandilia y Ventania con otras áreas, se analizaron las dis-tribuciones de los taxones específicos e infraespecíficos de Asteraceae que habitanestas sierras. Se delimitaron trece áreas: sur de América del Norte y América Central,sur de Brasil, Uruguay, Pampa, Tandilia, Ventania, Chaco, Sierras Pampeanas, Sie-rras Subandinas, Mahuidas, Patagonia, Chile Central, y Andes del Norte. Las unida-des de estudio fueron 112 taxones de Asteraceae presentes en Tandilia y Ventania(taxones endémicos, adventicios y naturalizados no fueron incluidos en el análisis).Las distribuciones de estos taxones fueron analizadas utilizando dos métodos:panbiogeografía (compatibilidad de trazos) y análisis de simplicidad de endem ismos.Ambos métodos conectan sur de Brasil, Uruguay, Pampa, Tandilia, Ventania y Sie-rras Pampeanas. Las Asteraceae de Tandilia y Ventania estarían más relacionadas conestas áreas que con la de las Sierras Subandinas, Andes del Norte, Chaco, Patagonia,Mahuidas y Chile Central. El área de América del Norte y América Central aparecemenos relacionada a Tandi1ia y Ventania en relación a las otras áreas. La evolución delas Asteraceae de Tandilia y Ventania y las áreas relacionadas ha sido probablementeafectada por los eventos geológicos de los períodos Terciario y Cuaternario. La dis-continuidad actual en el patrón de distribución de las Asteraceae, restringidas engeneral a las zonas elevadas, es explicada principalmente por eventos vicariantesdebidos a condiciones áridas.
Palabras clave. Asteraceae; biogeografía; Tandilia; Ventania.
INTRODUCfION
Since Darwin'sjourney in Argentina in 1835the geology ofTandilia and Ventania and theirinhabiting biota have attracted severalgenerations of geologists and biologists (e.g.,Darwin 1846, Alboff 1895, Hauthal190 1, Ca-brera 1938, Sota 1967, Teruggi & Kilmurray1975). The political province ofBuenos Aires,situated in central eastern Argentina, is coveredin most of its surface by an herbaceous, grassysteppe, ca\led "pampas". The pampas are a plainsituated at the sea level, or even below the sealevel (like the big depression ofthe Salado river),with the exception of two major systems of
22
mountains ranges or"sierras", namely Tandiliaand Ventania.From a geological point ofview, Tandilia andVentania do not appear to be very closelyrelated (Rolleri 1975). Tandilia, southeasternof Buenos Aires province, constitutes theoldest nucleus in the country originated inthe Proterozoic age. Ventania is locatedsouthwest of the province and has a morerecent origin dated from the Paleozoic age.Gondwanaland and Pre-Gondwanalandevents seem to have played a major role in thebuilding ofthese mountain ranges.The biodiversity ofTandilia and Ventania isparticularly interesting because oftheir high

number of endemic plantae and animal taxa(Table 1), and led to consider the ranges as"orcgraphic islands" (Kristensen & Frangi1995). When compared with the remainingvegetation of Buenos Aires, Tandilia andVentania have not only a characteristicpetrophylous flora, but they are the on lyregions in the province with endemic planttaxa (Parodi 1947). There have been numerousfloristic (Alboff 1895, Spegazzini 1896, 1901,Cabrera 1938, 1940, 1963) and ecological(Frangi 1973, Ponce 1982, 1986, Kristensen &Frangi 1995) studies in the area. The differentcommunities were explained by the variationin the local climate and soil, which is causedby aspects of the topography of the ranges(Kristensen & Frangi 1995).Within the diverse plant families inhabitingTandilia and Ventania, Asteraceae have thesecond place (after Poaceae, the grass family)in number of genera and species representedin the area (Parodi 1947). Approximately 30 %(112 species and infraspecific taxa) out of the356 Asteraceae known from the Buenos Ai-res province (Cabrera 1963, Zuloaga &Morrone 1999, Cabrera et al. 2000) occur inVentania and Tandilia. Furthermore, there havebeen found until the present four species ofAsteraceae endemic to Tandilia, three speciesand two varieties endemic to Ventania, andone species endemic to both, Tandilia andVentania.The current understanding of the biotichistory ofTandilia and Ventania is based onseveral lines of evidence. Systematic studieshave documented much of the taxonomicdiversity and distribution patterns of ferns(Sota 1967, 1972, 1973, 1985) and scorpions(Maury 1973) and some hypotheses werepostulated to explain those distributions.
It would be interesting to seek the historicalexplanations that led to the high diversity andendemicity of Asteraceae in Tandilia andVentania and the relationships of the biotainhabiting these mountain ranges with other
Crisci el al.
areas. In order to achieve this goal, we applyherein two methods: a panbiogeographicapproach (compatibility track analysis) andparsimony analysis ofendemicity (Morrone &Crisci 1990, 1995, Crisci et al. 2000) based ondistributional patterns exhibited by Asteraceae.
MATERIAL AND METHODS
Areas
Except for a few weedy introductions (e.g.,Facelis retusa, Gamochaeta subfalcatai, theAsteraceae of this region shows no linksoutside of the Americas. For these reason,areas such as Palearctic, Afrotropical, Orien-tal, Australian, Cape subregions, were nottaken into consideration for the delimitationareas.On the basis of several over1apping taxadistribution and phytogeographical andgeological criteria the areas delimited here are(Fig.I):
(1) Central Chile (CHI): Chile between 30°and 37° south latitude (Cabrera & Willink 1973,Morrone et al. 1997).(2) Central and North America (CNA):Southern North America and Central America.(3) Chaco (CHA): xerophyllous forestextending from southern Bolivia, western Pa-raguay, and northern and central Argentina(Cabrera & Willink 1973).(4) Mahuidas (MA): group of mountainranges in the political province ofLa Pampa,in central Argentina. They are 300-500 m high,and they are dated from the Cambric age (500m. y. ago)(Zambrano 1980).(5) Northern Andes (NA): corresponds to theParamo and Puna provinces, and comprises thehighlands over 3000 m altitude from Venezuelato northwestern Argentina (Morrone 1994).(6) Pampa (PAM): grassy steppe ranging fromsouthern Brazil, Uruguay, and eastern Argen-tina, from 30° south latitude to 39° southlatitude (Cabrera & Willink 1973).
23
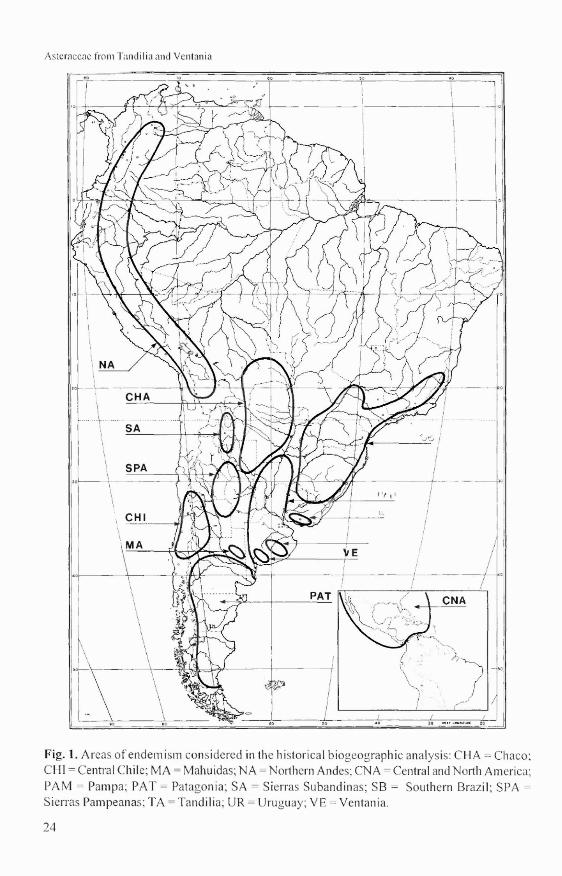
Asteraccac frorn Tandilia and Ventania
Fig. 1. Areas of endemism considered in the historical biogeographic analysis: CHA = Chaco;CHI =Central Chile; MA =Mahuidas; NA =Northern Andes; CNA =Central and North America;PAM = Pampa; PAT = Patagonia; SA = Sierras Subandinas; SB = Southern Brazil; SPA =
Sierras Pampeanas; TA = Tandilia; UR =Uruguay; VE =Ventania.
24

(7) Patagonia (PAT): semidesert east of thesouthern Andes to the Atlantic coast in Ar-gentina, approximately between 35° and 55°south latitude (Cabrera & Willink 1973).(8) Sierras Pampeanas (SPA): group of moun-tain ranges in central Argentina, extending inthe political provinces of Córdoba, Santiagodel Estero, eastern Catamarca, and San Luis.They are 3500 m high and their Precambricrocks were uplifted during Tertiary andQuaternary times (Gordillo & Lencinas 1980,Michaut 1980).(9) Sierras Subandinas (SA): a group ofmountain ranges in northwestern Argentina,extending in the political provinces of Saltaand Jujuy. They are 3000-5000 m high; themost ancient rocks be long to the Precambricage (Mingramm et al. 1980) and were upliftedin the Tertiary and Quaternary periods.(10) Southern Brazil (SB): subtropical forestthat corresponds to the Paran ense provinceofCabrera & Willink (1973) in southern Brazil(ranging west of Serra do Mar to central RioGrande do Sul), northeastern Argentina, andeastern Paraguay.(1 1) Tandilia (TA): or "Sierras Septentriona-les", a group ofmountain ranges east ofBue-nos Aires province. They are 500 m high, andrepresent the most ancient mountains in Ar-gentina dated from 2200 m. y. ago, in theProterozoic (Teruggi & Kilmurray 1975, 1980).(12) Uruguay (UR): prairie with endemic taxa,and low mountain ranges. It corresponds tothe eastern Uruguayan province (Castellanos& Pérez Moreau 1944,Chebataroff 1960), situa-ted in the south of Brazil (southern Rio Gran-de do Sul) and Uruguay.(13) Venlania (VE): or "Sierras Australes" agroup of moutain ranges southwest of Bue-nos Aires province. They are 900-1000 m high,and the geological age corresponds to 490 m.y. ago, in the Paleozoic (Llambías & Prozzi1972, Harrington 1980).
Crisci el al.
Taxa analyzed
The un its of study (Table 2) were species andinfraspecific taxa of Asteraceae inhabitingTandilia and Ventania. The distributional datawere taken from the specimens' labels oftheherbarium ofDepartamento Científico de Plan-tas Vasculares (Herbarium LP), Museo de LaPlata, Argentina and from the literature. Thenumber of specimens analyzed are ca. 2500.Each specimen was correctly identified priorto the extraction of data. Checklists (Troiani& Steibel 1999, Zuloaga & Morrone 1999, Ca-brera et al. 2000, Peter 2000) and floristictreatments of the province of Buenos Aires(Cabrera 1963, Cabrera & Zardini 1978), andrevisionary studies were taken al so intoconsideration. For nomenclature purposes wefollowed Zuloaga & Morrone (1999). Adven-ticious (e.g., Anthemis cotula, Matricariarecutita, Sonchus asper) , cultivated (e.g.,Ci-chorium intybus), naturalized (e.g.,Artemisia velortorum, Sonchus oleraceous)or cosmo-politan (e.g., Solidago eh ilensis,Taraxacum officinale¡ species were not takeninto consi-deration.
Panbiogeography (compatibility trackmethod)
Croizat postulated that earth and life evolvetogether (Croizat 1958), meaning that geo-graphic barriers evolve together with biotas.The panbiogeographic approach (Croizat 1958,1981) consists of plotting distributions oforganisms on maps, connecting their separatedistributional areas together with lines calledindividual tracks. As an example, some ofthe112 individual tracks produced in this study ofAsteraceae inhabiting Tandilia and/or Ventaniaand other areas are illustrated (Figs. 2-3). Whenindividual tracks coincide, the resultingsummary lines are considered gene-ralizedtracks, which indicate the pre-existence of an-cestral biotas, that become fragmented bytectonic and/or climatic changes.
25

Asteraceae from Tandilia and Ventania
Table 1. Taxa endemic to Tandilia and Ventania with the main references.
Taxa Tandilia Ventania References Tuxa Tandilia Vcntania ReferencesI'LANTAE IRIDACEAE
ASTERACEAE Cypctto herbertlí (Lindl.) Herb X X Ravenna 1968Bacchans rufesccns Sprcng val' X Cabrera 1963 subsp. l1'O/jjhucj.!,eli (1 Iauman)
vcntctnícula Cabrera RavennaBacchans tondttens¡s Speg. X Cabrera 1963 Sisyrinchinrnjunccnm i:. i\1ey X X Ravenna 1968l Iícrctcínm burlcartií Sleumer X X Cabrera 1963 subsp. lainezií {Hicken ) Ravenna
Hícrctcinm chacoense (Zahn ) X Cabrera 1963
Sleumet PLANTAGINACEAE
Hícrocinm tandttensc Sleumer X Cabrera 1963 ¡l/al/lago btsmarckt¡ Niederl. X Pontiroli 1965,)'1.'11(,(,;0 tcncopcptus Cabrera X Cabrera 1963 !J/onfuj.!,() tundilcnsis (Pilg.) X Pontiroli \065
S tandttcnsís Cabrera X Cabrera 1963 Rahn.'J'. twavcnsis Cabrera X Cabrera 1963 POACEAE
S vcníancnsis Cabrera X Cabrera 1963 Bromus bonarícnsis Parodi & J. X Cámara H
Stcvict satureiafotía (Lam.) Lam X Cabrera 1963 A.Cámara 1970
vnr. \'CI1ICII/{.:nsisCabrera Fcstnca ventanico/a Speg X Cabrera 1970
BRASSICACEAE l'íptochactíum ca/J'cSCL'I1S Parodi X Tones 1970a
!?o";ppa vcntanensis Cabrera X Iloelcke 1967
Stsvmbriurn ventancnsc Boelcke X Boelcke 1967 t'oa irtdfolia Ilauman X X Torres 1970bSetar¡a vaginata Spreng val' X Nicora 1970
BROMELlACEAE tcmdtlcnsis Nicora
ñyck¡a nstnotiilora Otto & DictrX Cabrera 1968 Slipa vcntunícolu Cabrera & X Cabrera &
Tones TO!Tes 1970var. 10I1dí/CIIS;S (Speg.) Cabrera
POLYGALACEAE
Titíondsío hergcri Mcz X Cabrera 1968 Pulygatu vcntancns¡s Grandona X Fnbris 1965
CACTACEAE
(~Vl1ln()caZ)'c;1/11I platense (Speg.)X Zuloaga & SOLANACEAE
Morrone 1999 Nicrcmbervia íandilcnsts X X Cabrera 1965Britton & Rose var. platense
(Kuntze) Cabrera
X Zuloaga & ANIMALIAGyrnnocctivcíum platense (Speg.)
Morronc 1999 BOTIIRIURIDAEBritton & Rose var, vcntantcotu
Bnthríurns vovan Mauri X Maury 1973(Speg.) R. Kiesling
IGUANIDAE
FAIl¡\CI:AE Líolaennrs gracitis spp. X Vega&
Adcsmto t-onarícnsts Burkart X Burkart 1967 13cllagamba
A. pampeana Speg. X Burkart 1967 1990
A. pselldogrisea Burk¡1I1 X Burkart 1967 Liolaemus sp X Vega &
Asfragalus argef1lÍ!lIIs X X Burkart 1967 Bellagamba
Manganaro 1990
Lcühvrus hookcri G. Don f. X Burkart 1967 l'rvstidactylus cctshnaíícnsís X Kristensen &
otbtttora (Kuntze) Burkart Frangi 1995
Mimosa rocac Lorentz & X X Burkart 1967 I.EPTOTYPIILOPI DAE
Nicdcrt. IJeJ7I()~VJ7/¡loJ7smunoaí Orejas X X Vega 8:
Vicia muntevidensis Vogel X Burkart 1967 Miranda Bcllagamba
oblonga Burkart 1990
V scütotía Kunth Val X X Burkart 1967 Mclnnophryníscus stclzncrí X Vega &
honaricnsís Burkart subsp. numtcvidensis Philippi Bcllagamba
1990
The compatibilitytrack method herein applied,developed by Craw (1988), is based on theconcept of distributional compatibility (Co-nnor 1988, Craw 1989). In this method, indivi-dual tracks are treated as biogeograph ichypotheses of locality or distribution arearelationship. Two or more individual tracksare regarded as being compatible with eachother only if they are the same pairwisecomparison or if one track is a subset ofthe other
26
(i.e., tracks are either included within orreplicated by one another). It basically consistsof constructing a matrix (areas vs. tracks),where each entry is 1 or O depending onwhether the track is present or absent in eacharea, and using a compati-bility analysisprogram to find the largest clique(s) of com-patible tracks. The method involves finding asimple form ofspanning tree linking localitiesor distribution areas. This tree is constructed

from the largest clique of compatibledistributions in a distributional compatibilityrnatrix and is identified as a generalized track.If more than one largest clique or severalcliques of considerable size are found, then ahypothesis of existence of several generalizedtracks linking the localities or distributionareas in more than one way can be considered.Alternatively, the intersection (i.e., thosetracks cornrnon to all the largest cliques) ofthe largest el iques can be identified as ageneralized track (Craw 1990). For more detailsand other applications ofthis method see Craw(1988, 1989),Morrone&Crisci(1990,1995),Crawet al. (1999) and Crisci et al. (2000). The analysisof the data matrix of 13 areas of endemisrn vs.distributional data of I 12 taxa (Table 3) wascarried out with SECANT 2.2 (Salisbury 1999).SECANT isa program for identifying all groups
Crisci el al.
of eladis-tically compatible characters. Wetreated our tracks as binary characters orderedwith absence as the "ancestral" state for each(= outgroup with all zeros).
Parsimony analysis of endernicity (PAE)
Parsimony analysis of endernicity or PAE(Rosen 1988, Rosen & Smith 1988) is anapproach of historical biogeography that ledto identify the distributional pattern oforganisms. It classifies localities, quadrats orareas (which are analogous to taxa) by theirshared taxa (wh ich are analogous to characters)according to the most parsirnonious solution(Morrone & Crisci 1995, Crisci et al. 2000).The rnethod was originally proposed by Rosen(1988) using localities as study units.
Fig.2 A-B. Individual tracks. A, Achyroc!ine satureioides ce), Baccharis crispa C-); B, B.ulicina (e), Conyza bonariensis (-).
27

Astcraccae trom Tandilia and Vcntania
Table 2. Species of Asteraceae from Tandilia and Ventania and other areas of endemism consideredin the study. Taxa with the same areas of endemism have been indented. Areas ofendemism foreach taxa are indicated in Table 3.
1 Achvrocline satnreioidcs (Lam.) De.2. Acmella decumbens (Sw.) R. K. Jansen var. affinis (Hook. &Arn.) R. K. Jansen
3. As/er squamatus (Spreng.) Hieran.4. Baccharis artetnisioides I Iook. & Arn.5. Ji articutata (Lam.) Pers.6. H crispo Spreng.7. Ji gilliesii A. Gray8. Ji spicat« (Lam.) Bail!.9. H. sleJ/opl1yl/a Ariza10. n triangularis Hauman11 n tridentcua Vahl var. suboppositc: (DC) Cabrera12. 1I tritnera (Less.) DC.13. B. nlicina Hook. & Arn.14. lsidens laevis (L.) Britton, Sterns & Poggenb.15. Chuptalia exscapa (Pers.) Bakcr var. cxscapa16. C. i;;J1ola Burkart17. C. integcrrinta (Vel!.) I3urkar118. (' I'illfselllfides (Vahl) Baker19. r: runcinata Kunth20. (' sinucna (Less.) Baker
21 Baccharis corídifola OC.22. Conyzu sumcurensís (Reitz.) E. Walkcr
23. Clw(juiroKu erinacea D. Don subsp. crinacea24. Conv:a honariensis (L.) Cronq. var. bonaricnsis25. e: floribunda Kunth26. (.'. tnonorchis (Griseb.) Cabrera
27. Hnlochetlns brasiliensís (L.) Cabrera28. Mikania pcriplocífolia Ilook. & Arn.20. Panphalea heterophvlla Less.30. Pndncoma hierac!fália (Poir.) Cass.
31 Conv:a prinmlifolia (Lam.) Cuatree. & l.ourteig32. C. serrana CabreraXL Enpcuoriutn bnníifolirnn I Iook. & Arn. var. Ivnniifolinrn34. lo'. bnpleurifolimn DC35. h'. macrocrphalntn Less.
36. Chevreulia sarmentosa (Pcrs.) S. r. Blake37. Enpcnoriuin squarrnlosum Hook. & Arn,
38. Bctccharis gnapbcüíoidcs Sprcng.39. Micropsis spastudata (Pers.) Cabrera
40. Facelís renrsa (Lam.) Seh. Bip. subsp, pcttulü Beauv.41 Jo'. re/lisa (Larn.) Sch. Bip. subsp. re/lisa
42. Guilla nt¡« meg apot amtca (Spreng.) Bak cr val'scabiosoidca (Arn. ex DC) Ilaker
43. Flavcria hcnunanii Dimitri & Orfila44. Stuckcrticlla peregrina Beauv.
45, Guillardia nusgapotamica (Sprcng.) Baker var. tueg«:po/{flJllca
46. Berrea gnaphalioídes (Less.) Bcauv.47. Grnnochaeta americana (Mill.) Wedd.48. (j. argentina Cabrera49. (; fatccua (Lam.) Cabrera
50. (i subfulcato (Cabrera) Cabrera51. Gienochactofituginea (DC.) Cabrera52. U plutensis (Cabrera) Cabrera53. r; stachvdifolío (Larn.) Cabrera54, Gnuphctlium cabrcrcu: S. E. Freire
55. Hypochucrts pcnnpasíca Cabrera56, Gnaphaliun) cheironthifolnnn Lam.57. c. gaudic/wudiomfm DC.58. (i lelfcopep!tflll Cabrera
28
59. Grindelia aegialítís Cabrera60. U. brachystephana Griseb.61. U./JII/chella Dunal var. discoidea (Hook & Am.) A. Bartoli& Tortosa
62. c. vcntancnsis A. Bartoli & Tortosa63. Gutierrezia gilliesii Griseb.64. Helenium radiotum (Lcss.) Seckt65. Hieracium palezienxii Zahn66. l Iyalis argenten D. Don ex Hook. & Arn, val'. latisouama
Cabrera67. Trichoctine sínucüa (D. Don) Cabrera
68. Hypochaeris pctiolaris (Ilook. & Arn.) Griseb,69. Hystenonicojasionotdcs Willd.70. Cony:a bnnoríens¡s (L.) Cronq. var. allglfsli/olia (Cabrera)l. Cabrera71.I(vs/eriolliea pinifotia (Poir.) l3aker
72. tIvnochaerís roscnguruii Cabrera var ptnncülfidu(Speg.) Cabrera
73. l.ncilia acntifuli« (Poir.) Cass.74. Microgyncl!a trifurcata (Less.) Grau75. Noticastrum marginantm (Kunth) Cuatrcc.76.I'crezia multiflora (Humb. & Bonpl.) Lcss. subsp. sonctnfotiu
(I3aker) Vuillcum.77. liupatorintn connnersanii (Cass.) l Iieron.78. llypuchuerís grisehachii Cabrera
79. Pluchea sagi//alls (Larn.) Cabrera80. ,)'enecio grisebctchii Bakcr val'. grisebachii
81. Podocoma hírsrüa (Hook. & Am.) Bakcr82. Cony:a hlukei (Cabrera) Cabrera83. Eupatortnm snbhastut um lIook. & Arn.84. Noticastrum diffusnn: (Pcrs.) Cabrera
85. Senecto arechavaletae Bakcr86. S bonaricnsis Hook. & Arn.87. S. griselwchii Baker val'. subincanns Cabrera
88. S osteni¡ Mattf.89. Senecto ntontevidensis (Sprcng.) Bakcr
90. Micropsis australts Cabrera91 Sencc¡o panrpeanus Cabrera92. S pulcher Ilook. & Arn.
93. liupatoriiun tanacctifolunn Gillics ex Ilook. & Arn94, Grindelia tniphthahnouk:s DC.95 l lvpochaeris l'oriegu/o (Lam.) Baker96. Sonunerfeltia spinulosa (Spreng.) I.css.
97. Senecto subnlan,» O. Don ex Ilook. & Arn. varo crcctus Ilook.&Am.
98. So/ira pterosperm« (Juss.) Less.99. Stevia saturciijolia (Lam.) Larn. var. pcuaganica lIieron.100. S. sotureütot¡« (Lam.) Lam, varo satureiifolia101 Ihclesperma megajJotmJ/iclflll (Sprcng.) Kuntzc
102. Noticastrum argentinensc (Cabrera) Cuatrcc.103. Vernonia echioides Lcss.
104. Actnella decumhcns (Sw.) R. K. Jansen val' decumbcns105. líaccharis gel1;sli!o/ia DC.106. Criscía stricta (Spreng.) Katinas107. Enpatorinm twccdiamnn Hook. & Arn.108. Hypochacris megapotomíca Cabrera109 Senecto selloí (Spreng.) DC110. Stcvia nntltiüristüta Sprcng.
111 Vernoniaflexuosa Sims.\12. Zcxtncniu buphthahniflom (Lorcntz) Ariza

Craw ( 1988) and Cracraft (1991) presented avariation of the method using are as ofendem ism as study un its to identify thehierarchical information contained in thegeographical distribution of organisms toestablish are a relationships. In this methoda data matrix of endemism areas by taxa isconstructed, and the presence of a taxon inan area is coded as 1 and its absence as O. Aparsimony algorithm is applied to the matrixto obtain a cladogram(s). PAE cladogramsrepresent nested sets of areas, in which ter-minal dichotomies represent two areasbetween which the most recent bioticinterchange has occurred (Morrone & Crisci1995). In this analysis the same are as anddata matrix (Table 3) used in thepanbiogeographic approach were taken intoconsideration. The analysis was carried outwith PAUP* version 4.0 beta 2 (Swofford1999), applying the branch-and-boundoption. A strict consensus tree wasconstructed based on the results generatedfrom PAUP*. As proposed by Rosen (1988),the cladogram was rooted with a hypotheticalarea coded all zeros.
RESULTS
Pan biogeography (compatibility track method)Based on the data matrix (Table 3), thecombination ofthe individual tracks resultedin four largest cliques (generalized tracks) of30 individual tracks each one (Fig. 4 A-O).The intersection of 27 individual tracks(identifíed with the numbers 3, 20, 21,23,25,26,27,28,29,30,36,37,38,81,82,83,84,87,88,103,104,105,106,107,108,109,110 inTable2)common to the four largest cliques is identifíedas the fifth clique or generalized track (Fig. 4E). In this way, six individual tracks (5, 40, 41,63, 64, and 98 in Table 2) are combined ingroups ofthree with the 27 individual tracksto form the four cliques of30 individual tracksmentioned above.Fivetrees connecting the areas can be constructed
Crisci el al.
from the fíve cliques, as follows (Fig.4):Clique A: ««««(T A, UR), SB), PAM), VE),SPA), CHA), SA), NA), PAT, CHI), supportedby the 30 individual tracks 3, 5, 20, 21,23,25,26,27,28,29,30,36,37,38,40,41,81,82,83,84,87,88,103,104,105,106,107,108,109,110(Fig.4A).Clique B: ««««(T A, UR), SB), PAM), VE),SPA), SA), CHA), NA), CHI, PAT), supportedby the 30 individual tracks 3,5,20,21,23,25,26,27,28,29,30,36,37,38,63,64,81,82,83,84,87,88,103,104,105,106,107,108,109,110(Fig. 4 B).Clique C: ««««(T A, UR), SB), PAM), VE),SPA), CHA), SA), CHI) NA, PAT), supportedby the 30 individual tracks 3, 20, 21,23,25,26,27,28,29,30,36,37,38,40,41,81,82,83,84,87,88,98,103,104,105,106,107,108,109,110(Fig.4C).Clique O: ««««(TA, UR), SB), PAM), VE),SPA), SA), CHA), CHI), NA, PAT), supportedby the 30 individual tracks 3, 20, 21,23,25,26,27,28,29,30,36,37,38,63,64,81,82,83,84,87,88,98,103,104,105,106,107,108,109,110(Fig.4 O).Clique E: ««««(TA, UR), SB), PAM), VE),SPA), SA, CHA) NA, PAT, CHl), supportedby the 27 individual tracks 3, 20, 21,23,25,26,27,28,29,30,36,37,38,81,82,83,84,87,88,103, 104,105,106,107, 108, 109, 110(Fig. 4 E).Mahuidas and Central and North America arethe only areas that appear in none of thesepattern of relationships in the fíve generalizedtracks obtained.Oue to the fifth clique (E) represents theintersection of27 individual tracks commonto the four largest cliques, it was selected toshow the taxa supporting the areasrelationship (Fig. 4 E):(1) (TA,UR). Based on Senecio grisebachiivaro subincanus (individual track 87) and S.ostenii (track 88).(2) «TA,UR), SB). Based on Eupatoriumsquarrulosum (track 37), Baccharis gnapha-lioides (track 38), and Micropsis spathulata(track 39).
29

Asteraceae from Tandilia and Ventania
Fig.3 A-F. Individual tracks. A, Podocoma hirsuta ce), Senecio arechavaletae C-); B,Hieraciumpalezieuxii ce), Hyalis argentea var. latisquama C-). e, Hysterionica pinifolia ce), Pereziamultiflora subsp. sonchifolia C-); D, Senecio pulcher ce), Sommerfeltia spinulosa C-); E,Senecio bonariensis ce), Senecio pampeanus C-); F, Vernonia echioides ce), Zexmeniabuphtalmiflora C-).
30

,----------- CHIf---------- PAT~--------- NA,--------- SA....-------- CHA~-----SPA.------VE.-----PAM.----SB
TAUR
,.--------- PATf----------- NA....--------------CHI~--------- SA,----------- CHAr------SPA.------VE,.----PAM,----SB
TAUR
INDIVIDUAL TRACKS
$ 3.24.31Q5X 20.21.98<:> 22.64• 41.42* 81.82.83.84$ 26.27.28.29.30* 103.104.105.106
107.108.109.110* 37.38.39~ 87.88
Crisci el al .
.------------------ PAT~----------------CHI.-----------NA.--------- CHAr---------- SA,-------- SPA,------- VE~---PAM~---SB
TAUR
~------------PAT~-------------NA~---------------CHI~---------- CHA....--------- SA,-------- SPA.------VE,.-----PAM.----SB
TAUR
....--------------- CHI~--------PAT
~~------------- NAr-------- CHA~-------- SA....------- SPA,----- VE.------ PAM~--SB
TAURE
Fig. 4. A-E. Cliques obtained in the compatibility track method after applying SECANT 2.2,showed as cladograms. A-D, Four largest c\iques each supported by 30 individual tracks. E,Clique obtained by the intersection of27 individual tracks common to the four largest c\iques.The table shows the individual tracks or the sets of individual tracks, represented by syrnbols,that are superimposed onto the cladograms (see Table 2 for numbers corresponding to taxanames and Table 3 for individual tracks). CHA = Chaco; CHI = Central Chile; NA = NorthernAndes; PAM = Pampa; PAT= Patagonia; SA = Sierras Subandinas; SB = Southem Brazil; SPA= Sierras Pampeanas; TA = Tandilia; UR = Uruguay; VE = Ventania.
31

Asteraccac from TandiJia and Ventania
(3) (((TA,UR), SB), PAM). Based on Vernoniaechioides (track 103), Acmella decumbens(track 104), Baccharis genistifolia (track 105),Criscia stricta (track 106), Eupatoriumtweedianum (track 107), Hypochaerismegapotamica (track 108), Senecio selloi (track109), and Stevia multiaristata (track 110).(4) ((((TA,UR), SB), PAM), VE). Based onConyza monorchis (track 26), Holocheilusbrasiliensis (track 27), Mikania periplocifolia(track 28), Panphalea heterophylla (track 29),and Podocoma hieracifolia (track 30).(5) (((((TA,UR), SB), PAM), VE), SPA). Basedon Podocoma hirsuta (track 81), Conyza blakei(track 82), Eupatorium subhastatum (track 83),and Noticastrum diffusum (track 84).
Parsimony analysis of endemicity
The analysis ofthe data matrix (Table 3) withPAUP* generated four area cladograms (Fig.5 A-D) with 263 steps, c.i.= 0.426, and r.i. =0.663. The strict con sen sus ofthe four trees(Fig. 5 E) shows the following area relation-ships: (Mahuidas (Patagonia (Chile (SierrasSubandinas (Sierras Pampeanas (Ventania(Pampa (Southern Brazil - Tandilia - Uru-guay)))))))). The remaining areas: Chaco(CHA), Central and North America (CNA), andnorthern Andes (NA) do not show resolvedrelationships with the other areas in theconsensus.
Comparison ofboth methods
The comparison of the results obtainedapplying panbiogeography (compatibilitytrack rnethod) and parsimony analysis ofendernicity to the areas under study shows acoincidence in the relationships of SierrasPampeanas (SPA), Ventania (VE), Pampa(PAM), Southern Brazil (SB), Tandilia (T A),and Uruguay (UR).The areas relationship PAM-SB-SPA-TA-UR-VE conform a rnain distributional patternstrongly supported by the two approaches
32
applied in this study, and represented in Fig.6 as a generalized track. Both methods showthat Tandilia is closer to Uruguay, southernBrazil, and Pampa than to Ventania. On theother hand, PAE and panbiogeographyresulted in different relationships of theremaining other areas with the m a indistributional pattern. PAE shows that Sie-rras Subandinas (SA) (Fig. 5 A-E) are relatedto this pattern, whereas panbiogeo-graphyshows that Sierras Subandinas can be directlyrelated to it (Fig. 4 B, D) or cannot be directlyrelated (Fig. 4 A, C); PAE shows Chaco (CHA)as not closely related to the pattern (Fig. 5 A-E), whereas panbiogeography shows CHAconnected to it (Fig. 4 A-E); PAE showsMahuidas (MA) and Central and NorthArnerica (CNA) not closely related to thepattern (Fig. 5 A-E), and panbiogeographyshows these last two areas very distantlyrelated to it (for this reason they are notindicated in the results).
DISCUSSION
The main distributional pattern connectingTandilia and Ventania with southern Brazil,Pampa, Uruguay, and Sierras Pampeanasobtained through the cornpatibility trackmethod and PAE partially agree with othersauthors' hypotheses.A biotic rnigratorial route frorn the Andes toBrazil, through Sierras Pampeanas, sierras ofBuenos Aires, sierras ofUruguay, and Planaltoand sierras of Brazil was postulated rnany yearsago by Hicken (1918-1919) and Brade (1942).Frenguelli (1950) gavethe name of periparnpasicorogenic are (arco peripampásico serrano) tothe biotic corridor constituted by SierrasPampeanas, Mahuidas, and Tandilia. Ringuelet(1956) postulated that the province ofBuenosAires constitutes a cornposite area due to thepresence oftwo types offauna: one related tothe Subtropical domain (Guaianian-Brazilian),and the other related to the Austral region(Andean-Patagonic, or Chilean-Patagonic).

Crisci el al.
o 11 Q) ~l o o ,.... o o o o o ,.... o ,.... ,.... o ,....u f- u ,.... o ,.... o o o o ,.... o o ,.... o ,.... ,.... oe~ « Q) o6 VJ o o o o o o ,.... o o ,.... o ,.... ,.... oe, .D 0-, o o o o o o ,.... o o ,.... o ,.... ,.... o
f'? 8 8 8 8 8 8 ,.... 8 8 ,.... 8 ,.... ,.... 811 (.:j « ,.... ,.... •....• ,....,.... 'O o o o o o o ,.... o o ,.... o ,.... ,.... o« o. .s:;j ,.... or, 8 8 8 8 8 8 ,.... 8 8 ,.... 8 ,.... ,.... 86 "1' ,.... ,.... ,.... ,....::c e Ñ 8 8 o 8 o 8 ,.... o 8 ,.... o ,.... ,.... oU ~ co ,.... ,.... ,.... ,.... o ,.... o ,.... ,....e, ~
,.... o o ,.... o ,.... o ,.... ,.... o o ,.... o ,.... ,....
.~11 Q) o o o o o o o ,.... o o o ,.... ,.... o ,....;¿ > 0-, o o ,.... o ,.... o o ,.... o o ,.... ,.... o ,....
.~ f'? 8 ,.... ,.... 8 8 8 ,.... o ,.... ,.... ,.... ,.... ,.... ,....11 o ,.... ,.... ,.... ,.... ,.... ,.... o o o
6 « o 'O o o o o o o o o o ,.... o ,.... ,.... ,....e, [..Ll ,.... 'n 8 8 8 8 8 8 8 8 8 ,.... 8 ,.... ,.... ,....
Q) "1' ...... ...... ...... ,....-o > Ñ 8 8 8 8 8 8 8 8 8 ...... 8 ,.... ...... ......e o. ...... ,.... ,.... ......Q) g c-, ...... o o o o ,.... o ...... ...... o o ...... •....• o ......4- ~o :.... ::l o o o o o o o ...... ...... o ...... ...... o ...... ,....VJ bl) bl) O'. o o o o o o ...... ,.... o ,.... ,.... o ...... ,....S f'? 8 8 8 8 8 8 8 8 8 8 8 ...... ...... 8'Vi ::l •....• ,....
>. o :....'O o o o o o o ...... ...... o o ,.... ...... o ,....
~ ~ 0-, 'n 8 8 8 8 8 8 o 8 8 o o o ...... ......11 "1' ,.... ...... ...... ...... ...... ,....
11 Ñ 8 8 8 8 8 8 ...... 8 8 ...... ,.... ,.... ...... ,....¡¡¡ f- ~
...... ,.... ,.... •....• ...... ,....>,~
...... o o o o o o ...... o o ,.... ...... •....• ,.... ,....§ O ~ o o ...... o o o o ...... o ...... ...... ...... ,.... ...... o¿ O'. o ...... o o o o ...... o ...... ...... ...... ,.... ,.... or f!2 8 8 8 8 8 8 ,.... 8 8 8 8 •....• ,.... .......5 VJ .- ...... •....• ,.... ......Vl
v;-:::::'O o o o o o o ...... o o o o ...... ...... ......
'- -o -o 00 'ro 8 8 8 8 8 ...... o 8 ...... o ...... 8 o ......~ e e "1' o ...... o ...... ...... ...... ......0.« ~ Ñ 8 8 ,.... 8 8 ...... ...... 8 ...... ,.... ...... •....• ...... ,....f- o o o o o o •....• ...... ,....
Q) ...... o o o o o o o o o o o ...... ...... ......-5 e11'- ......•... Q) < o o ...... ,.... o o ...... ...... ...... o ,.... o o ,....
t.8 ..c: O'. o ...... ,.... o ...... o ...... ...... ,.... o ...... o o ......t f- f!2 8 8 8 8 8 8 ...... 8 8 8 o 8 ...... ......,.... ,.... o ......-o o
Vl t ..· 'O o o o o o o ,.... o o o ...... o o ,....e z; oro 8 8 8 8 8 8 ...... 8 o o ...... ...... ...... ,....~ ~ "1' ...... ,.... ,.... ...... •....• ...... ,....VJ e Ñ 8 8 ...... 8 ,.... 8 ,.... ...... ...... 8 ...... o 8 ,....~ o o o o o o ,.... ......Vl Q) ...... o o ,.... o o o ...... ...... o ...... ,.... ,.... ,.... o>.« o."'§:z 6 o o ...... o o o o ...... ...... o ...... o ,.... o ......
O'. o o o o o o ,.... o o o o •....• o o~ f!2 8 o ...... 8 ...... 8 o ...... ...... ...... ...... 8 ,.... ,....~ Vl e, ,.... ...... o ,.... ,.... ...... ...... ...... ...... ,....~ ce VJ 'O 'O o o ...... o ...... o o o o ...... ,.... ...... ...... ......
~ 'n 8 8 ...... 8 8 8 ...... ...... 8 8 ...... 8 8 ......u ce "1' ...... ...... ...... ,.... ......~ ::l t:: Ñ 8 o 8 8 ,.... 8 ...... ...... 8 ...... ,.... ,.... ,.... ,.....t:: ..c: .~ ...... o ...... o o o o o .......~\ co ...... o o ...... o ...... o ...... ...... o ...... ...... ,.... ...... ......
;¿ VJ11
o o o o o o o ...... o ,.... ,.... ...... o ...... ......:.o 11 N 0-, o o o o o o ...... o ...... ,.... ,.... o ...... ......
<' f'? 8 8 ...... o ...... o ...... o o o ...... ,.... ...... .......~< ~ ...... ,.... o ...... ...... ...... ...... ...... ...... ,.... ,.... ......e, 'O o o ...... o o o ...... ...... o ...... ...... o ,.... ,....;::-;¿ VJ .D 'ro 'ro 8 8 ...... 8 8 8 ...... ...... o ...... ...... 8 ,.... ......
ro "1' ...... ...... ...... ...... o ...... o ,....§ f- Ñ 8 o ...... 8 8 8 ,.... ,.... ...... o ...... o o ,....
~ 'Ñ...... o ...... o o ...... ...... ...... ...... ......
u u 4- ...... o ...... o o o o ...... o o ,.... ...... •....• ...... .......¡:: ~ o
Q) :....Q) o o o o o ...... o o ...... o o ...... ,.... o ......-5 Q) ca VJ O'. o o o o o o o o o ...... o ,.... ,.... o
6 f'? 8 8 8 8 8 8 8 8 8 ...... 8 ,.... ,.... 8'- e o ...... •....• ......t.8 <' •... -5 'O o o o ,.... o o ...... o ...... ...... ...... ,.... o ......
~ "1' 'n 8 8 8 ,.... 8 8 ...... 8 ...... ...... ...... ,.... o ,....~ . ..c 8 "1' o ...... ...... ...... o ,.... ...... ,....VJ t Ñ 8 8 ,.... 8 ,.... 8 ,.... 8 ...... ,.... ...... ,.... ,.... ,....~ g -o o o o ...... ...... o •....• ,.... ......u o e ...... o ...... ,.... o o ...... ...... ...... ...... ...... ...... ,.... ,.... ......~ Z VJ ob -o 11 o. o o o o o o o ...... o o ...... o ...... ...... ......
0-, o o o o o o ...... o o ,.... o •....• ...... ......C;; e ro Vl f'? 8 8 8 8 8 8 ...... 8 8 ...... 8 •....• ,.... ......
Q)::l ~ VJ •... ...... ...... ...... ,.... ......~ '- 'O o o o o o o ,.... o o ...... o ...... ,.... ,....
~ o M 'n 8 o ...... 8 ...... ,.... ,.... ,.... ,.... ...... ...... ...... ...... ,.....2: ~
VJ u "1' ...... ...... o ,.... ...... ...... ...... ...... ...... ...... ...... ,....~ Ñ 8 8 ...... 8 8 8 o ,.... ,.... o ...... o o ......-o .::: C"J o ...... o ,.... ,.... ,.... ...... ...... .......::: (jj x ...... o ,.... o o o o ...... o ...... ,.... ...... ,.... ...... ......
U -o Evi e o o ...... o o o o ...... o ...... ...... ,.... ,.... ,.... ......11 ~ 4- O'. o ...... o ...... o ...... ,.... o o ...... ...... •....• ,.... o> .D o f!2 8 8 8 8 8 o ,.... 8 o ...... o o ...... ......
Vl « ::l :.... ...... ,.... ...... ...... ...... ...... o ......~ 'O o o o o o o ...... o o o o ...... ...... oQ) Z VJ Q) """'1 Ir, 8 O O O 8 ...... ...... 8 O ...... ,.... ,.... ,.... ......•... VJ
.D "1' ,.... ,.... ,.... ...... ...... ...... ,.... ,.... ,.... ...... O~ U ~ E Ñ 8 ...... 8 8 ...... ...... ...... ...... ...... O ,.... 8 O ,....•... O O ...... ...... O ...... ...... O ...... ......X ~ '- ;::l ...... O ...... o· O O O ...... O ...... ...... O ,.... ,.... ......"5 .~ z..c: VJ O O O O O O O O ...... O O ...... O O ......~ U
O'. O O O O O O ,.... ,.... O O ...... O ...... ,....E 11 f'? 8 ...... O 8 8 8 ...... O O ,.... ...... •....• ...... O
Cij...... ...... ,.... ...... ...... O ...... O O ,....
~ < Q) ...... 'O O O ...... O O O ,.... ...... O O ...... O ...... ......'i:j b VJ 'n 8 ...... 8 8 O ,.... ,.... 8 ...... ...... ...... ...... ...... ......
U "1' O ...... O ...... ,.... O ...... ...... ...... ......O e
.~ e Ñ 8 ,.... ...... 8 o ,.... ...... ,.... ,.... ,.... ...... •....• ...... ......Q) Q) o o ...... o o o ,.... o ,.... ,.... o o...-i U e Vl
...... o o o o o ...... ...... ...... ...... ...... ...... ,.... ...... ......Q)
Q) o :....
:c bl) o.5 ~
~ "'" E-<~ § ~ ~ ~~ ::c 'i:j o G :i ~ ~ ~ ~ ~ g;f- U e, 11 O p... p... <JJ <JJ <JJ
33

Asteraceae frorn Tandilia and Vcntania
Sota (1967,1972, 1973, 1985) in his analyses ofthe ferns growing in Tandilia and Ventania,concluded that the pteridological flora ofthesemountain ranges have four origins: Austral-Antarctic, Andean-Pampean, Austrobrazilian,and Endemic. According to his studies the Bue-nos Aires' mountain ranges constitute anintermediate orofilous station in the migratorialroutes between the Andean-Pampean or theAustral-Antarctic flora, and the Austrobrazilianflora. Maury (1973) established, on the basis ofthe scorpions inhabiting Tandilia and Ventania,nearly the same kind ofrelationships with otherareas as showed by Sota: with the Subtropicaldomain, Central domain (Sierras Pampeanas),and Patagonian and Central Chilean domains.The studies based on ferns and scorpionsshow a link between Tandilia and Ventaniawith the Austral region, a kind ofrelationshipnot found here for Asteraceae. Althoughsorne Asteraceae inhabiting Tandilia andVentania occur in Patagonia, Central Chile andMahuidas, they do not constitute or areincluded in a generalized track. It is possiblethat different taxonomic groups such asscorpions, ferns and Asteraceae haveundergone different historical processes thatled to their actual distributional pattern.The relationships among the areas ofthe maindistributional pattern found in this studyshow that Tandilia is closer to Uruguay andsouthern Brazil than to Ventania. AlthoughTandilia and Ventania are geographically closeto each other, the geological evidencesuggests that both mountain ranges resultedfrom independent geological processes atdifferent geological times. According to Ra-mos (1989), Ventania was the result of thecollision of a Patagonian block that driftedfrom the south and accreted to Gondwanaland(ca. 360-440 m.y. ago). Tandilia, on the otherhand, was a part of the Río de La Plata craton(together with northeast Argentina, east Pa-raguay, Uruguay and southeast Brazil) beforethe end ofthe Precambrian (530-570 m.y. ago).Due to the family Asteraceae is believed to
34
have originated in South America in theTertiary, during the Oligocene (38 m.y.ago)(Stuessy et al. 1996), two geological eventsmay have played a significant role in theAsteraceae evolution and distribution inTandilia and Ventania and related areas, theuplift of the Andes and the Pleistoceneglaciations.During late Cenozoic (10-5 m.y. ago)the upliftof the Andes together with the developmentof cold offshore currents have beenassociated with a diversification of habitatsand evolutionary opportunities for the biota(Darlington 1968, Axelrod et al. 1991)especially for the Asteraceae (Funk et al. 1995,Stuessy et al. 1996).From the Miocene (25-5 m.y. ago) to Pliocene(5-1 m.y. ago) the climate was drying in southemSouth America caused by the slowly raising ofthe Andean chain and the cold Humboldtcurrent that intensified aridity. The last phaseofthe Andean orogeny in the upper Pliocenecaused additional uplift ofSierras Pampeanasand Sierras Subandinas (Taylor 1991), that hadstarted their uplift with the climax of theAndean cordillera raising during the Tertiary.Finally, during the Pliocene and Pleistocene inthe Quaternary period, glaciations caused dryand wet cycles that caused fragmentation anddifferentiation of the populations (SimpsonVuilleumier 1971, Simpson 1975). Semiaridvegetation extended into the continent, e.g., tothe south Patagonia, north of Atacaman desert,Chaco, and Monte, in the core of SouthArnerica, and the cerrado and caatingas incentral and northeastern Brazil, respectively(Prado & Gibbs 1993). This may resulted in aneventual separation of populations in moreelevated regions, restricted now to rocky andloose soils. The main distributional patternfound in this study connecting the mountainranges ofsouthern Brazil, Uruguay, Tandilia,Ventania and Sierras Pampeanas, and theAsteraceae endemicity found in Tandilia andVentania could be explained as a consequenceof these events.

OUTCNANACHACHIPAMS8URTAVESPASA
DPATMA
OUTCHACHIPAMS8TAURVESPASAPAT
E MACNANA
OUTCNANACHACHIPAMURS8TAVESPASA
A PATMA
Crisci el al.
r--r- -~- lr=r-e-
,.--'--
'--
OUTCHACHIPAMURS8TAVESPASAPATMANACNAB
,.-------------------------OUT,-------------------CHA
.------------CHI,-----PAM
S8URTA
L-.-----VE
L--------SPAL------SA
L--------------PATL-.--------MA
L-----------NAL-.. CNA
Fig. 5. A-E, Cladograms obtained after applying parsimony analysis of endemicity after applyingPAUP*. A-O, Four area cladograms. E, Strict consensus ofthe four cladograms. CHA =Chaco;CHI = Central Chile; MA = Mahuidas; NA =Northem Andes; CNA = Central and North America;PAM = Pampa; PAT = Patagonia; SA = Sierras Subandinas; SS = Southem Brazil; SPA =Sierras Pampeanas; TA = Tandilia; UR = Uruguay; VE = Ventania.
35

Asteraceae from Tandilia and Ventania
--1'\
-J_o\~
re \
\
\\- -\---5PA----
\ PAM
\--\\\\
Fig. 6. Distributional pattern of relationships among areas based on Asteraceae taxa, andrepresented as a generalized track. PAM = Pampa; SS = Southern Brazil; SPA = SierrasPampeanas; TA = Tandilia; UR = Uruguay; VE =Ventania.
36

In surnmary, it is probable that the evolutionof Asteraceae in Tandilia and Ventania wasmainly affected by Tertiary and Quaternarygeologic events. As a result, this biota showsa high level of diversity, many endem ic taxa,and closer relationships with southern Brazil,Uruguay, Pampa, and Sierras Pampeanasrather than with other arcas of América. Thediscon-tinuous pattern, i.e., occurring inelevated areas surrounding a core area ofCha-co and Monte vegetation, is what one wouldexpect under vicariance due to dry conditions,although dis-persal is taken also as a commonand impor-tant distributional event. Furthercorrelations with analyses of other plant andanimal taxa will determine the generality ofthe pattern shown by Asteraceae.
ACKNOWLEDGEMENTS
We thank Paul Berry, John Grehan, EdgardoOrtiz Jaureguizar, Mario Teruggi, RosendoPascual, and Elías de la Sota for helpfulcomments on the manuscript. We thank JorgeFrangi for checking the Iist of species. Specialthanks to Benjamin A. Salisbury for his gene-rous assistance with his program SECANT.This study was supported by grant 5776-96of the National Geographic Society. Support11'0111the Consejo Nacional de Investigaciones Cien-tíficas y Técnicas (CONICET), Argentina, towhichthe authors belong, is gratefully acknowledged.
L1TERATURE ClTED
AI.BOIF,N. 1895. Rapport préliminaire sur uneexcursion botanique dans la Sierra Ventana.Revista del Musco de La Plata 7: 183-187.
AXFLROU,D. i.M. T. K¡\L1NARROYO& P. H.R¡\VEN. 1991. Historical development oftemperate vegetation in the Americas. Revistachilena de Historia Natural 64:413-446.
BOLLCKE,O. 1967.Cruciferae, Vol. 4 (3), pp. 281-371 en: A. L. Cabrera (ed.). Flora de la pro-vincia de Buenos Aires. Colección Cientí-fica INT A, Buenos Aires, Argentina.
Crisci el al,
BR¡\DE,A. C. 1942. A composicáo da flora doItatiaia, Rodriguesia 6 (15): 29-43.
BURK¡\RT,A. 1967. Leguminosae. Vol. 4 (3), pp.394-647 en: A. L. Cabrera (ed.). Flora de laprovincia de Buenos Aires. ColecciónCientífica INTA, Buenos Aires, Argenti-na.
C¡\BRER¡\,A. L. 1938. Excursión botánica porlas Sierras Australes de la provincia deBuenos Aires. Revista del Museo de LaPlata (nueva serie) Sección Oficial: 60-69.
C¡\BRERA,A. L. 1940. Notas sobre la vegeta-ción del parque provincial Sierra de la Ven-tana. Anuario Rural de la Dirección deAgricultura, Ganadería e Industria 8: 3-16.
C¡\BRERA,A. L. 1963. Compuestas. Vol. 4 (6),pp. 1-443 en: A. L. Cabrera (ed.). Flora dela provincia de Buenos Aires. ColecciónCientífica INT A, Buenos Aires, Argenti-na.
C¡\BRER¡\,A. L. 1965. Solanaceae. Vol. 4 (5),pp. 190-250 en: A. L. Cabrera (ed.). Florade la provincia de Buenos Aires. Colec-ción Científica INT A, Buenos Aires, Ar-gentina.
C¡\BRER¡\,A. L. 1968. Bromeliaceae. Vol. 4 (1),pp. 448-459 en: A. L. Cabrera (ed.). Flora dela provincia de Buenos Aires. ColecciónCientífica INT A, Buenos Aires, Argentina.
C¡\BRERA,A. L. 1970. Festuca L. (Poaceae).Vol. 4 (2), pp. 125-128 en: A. L. Cabrera(ed.). Flora de la provincia de Buenos Ai-res. Colección Científica INT A, Buenos Ai-res, Argentina.
C¡\BRER¡\,A. L. & M. A. TORRES.1970. Stipa L.(Poaceae). Vol. 4 (2), pp. 255-290 en: A. L.Cabrera (ed.). Flora de la provincia de Bue-nos Aires. Colección Científica INTA,Buenos Aires, Argentina.
C¡\BRERA,A. L. & A. WILLlNK. 1973. Biogeo-grafía de América Latina. Monografía 13,Serie de Biología, OEA, Washington, D. C.
C¡\I3RER¡\,A. L. & E. M. Z¡\RDINI.1978. Manualde la flora de los alrededores de BuenosAires. Editorial ACME, S.A.C.I., BuenosAires, Argentina.
37

Asteraccae lrom Tandilia and Ventania
CABRERA,A. L., 1. V. CRISCI,G. DELUCCHI,S. E.FREIRE, D. GIULlANO, L. rHARLEGIII,L.KATlNAS, A. SÁENZ, G. SANCHO& E.URTUBEY.2000. Catálogo ilustrado de lascompuestas de la provincia de Buenos Ai-res. COBrOBO (Comisión de Biodiver-sidad Bonaerense). Convenio Secretaríade Política Ambiental-UNLP, Provincia deBuenos Aires, Argentina.
CÁMARA HERNÁNDEZ, J. 1970. Bromus L.(Poaceae). Vol. 4 (2), pp. 85-101 en: A. L.Cabrera (ed.). Flora de la provincia de Bue-nos Aires. Colección Científica INT A,Buenos Aires, Argentina.
CASTELLANOS,A. & R. PÉREZMOREAU.1944.Los tipos de vegetación de la RepúblicaArgentina. Universidad de Buenos AiresPublicaciones 3.
CONNOR,E. F. 1988. Fossils, phenetics andphylogenetics: inferring the historicaldynamics ofbiogeographic distributions.Pp. 254-269 in: J. K. Liebherr (ed.),Zoogeography of Caribbean insects.Comell University Press, Ithaca, New York.
CHEBATAROFF,J. 1960. Algunos aspectos evo-lutivos de la vegetación de la ProvinciaFitogeográfica Uruguayense. Revista Na-cional (Uruguay) 20 1: 3-18.
CRACRAFT,J. 1991. Patterns of diversificationwithin continental biotas: hierarchicalcongruence among the are as of endemismof Australian vertebrates. AustralianSystematic Botany 4: 211-427.
CRAW, R. C. 1988. Continuing the synthesisbetween panbiogeography, phylogeneticsystematics and geology as illustrated bystudies on the biogeography of NewZealand and the Chatham islands.Systematic Zoology 37: 291-310.
CRAW, R. C. 1989. Quantitative panbio-geography: introduction to methods. NewZealand Journal ofZoology 16: 485-494.
CRAW,R. C. 1990. New Zealand biogeography:a panbiogeographic approach. New Zea-land Journal ofZoology 16: 527-547.
38
CRAW,R. c., 1. R. GREHAN& M. 1. HEADS.1999.Panbiogeography: Tracking the his-toryoflife. Oxford University Press, New York,Oxford.
CRISCI,1. V., L. KATINAS& P. POSADAS.2000.Introducción a la teoría y práctica de labiogeografía histórica. Sociedad Argenti-na de Botánica, Buenos Aires, Argentina.
CROIZAT,L. 1958. Panbiogeography. Vols. 1,2a,and 2b. Published by the author, Caracas,Venezuela.
CROIZAT,L. 1981. Biogeography: past, present,and future. Pp. 501-523 in: G. Nelson & D.Rosen (eds.), Vicariance biogeography: Acritique. Columbia University Press, NewYork.
DARLlNGTON,P. 1. JR. 1968. Biogeography ofthe southem end of the world. HarvardUniversity Press, Cambridge, Massachu-ssets.
DARWIN,C. 1846. Geological observations ofSouth America, being the third part ofthegeology ofthe voyage ofthe Beagle during1832-1836. London.
FABRIS,H. A. 1965. Polygalaceae. Vol. 4 (4),pp. 58-69 en: A. L. Cabrera (ed.). Flora dela provincia de Buenos Aires. ColecciónCientífica INT A, Buenos Aires, Argenti-na.
FRANGI,1. L. 1973. Las comunidades vegeta-les de las Sierras de Tandil (provincia deBuenos Aires). Tesis, Universidad Nacio-nal de La Plata, La Plata, Argentina.
FRENGUELLI,1. 1950. Rasgos generales de lamorfología y la geología de la provinciade Buenos Aires. Serie 2, n. 33, pp. 72. L.E. M. 1.T., La Plata, Argentina.
FUNK,V. A., H. ROI3INSON,G. S. MCKEE& 1. F.PRUSKI. 1995. Neotropical montaneCompositae with an emphasis on the An-des. Pp. 451-471 in: S. P. Churchill, H.Balslev, E. Forero & J. L. Luteyn (eds.).Biodiversity and Conservation inNeotropical Montane Forests. The NewYork Botanical Garden, New York.

GORDILLO,C. E. & A. N. LENCINAS.1980. Sie-rras Pampeanas de Córdoba y San Luis.Vol. 1, pp. 577-638 en: Segundo Simposiode Geología Regional Argentina. Córdo-ba, Argentina.
HARRINGTON,H. J. 1980. Sierras Australes dela provincia de Buenos Aires. Vol. 2, pp.967-983 en: Segundo Simposio de Geolo-gía Regional Argentina. Córdoba, Argen-tina.
HAUTHAL,R. 190 l. Contribución al conocimien-to de la geología de la Provincia de Bue-nos Aires. I. Excursión de la Sierra de laVentana. II: Apuntes geológicos de las Sie-rras de Olavarría. Publicaciones de la Uni-versidad Nacional de La Plata, Facultadde Ciencias Naturales y Museo l.
HICKEN,C. M. 1918-1919. Lamigración de loshelechos en la flora de Tucumán. Pp. 187-209. Primera Reunión Nacional de la So-ciedad Científica Argentina de CienciasNaturales, Tucumán, Argentina.
KRISTENSEN,M. J. & J. L. FRANG!.1995. La Sie-rra de la Ventana: una isla de biodiversi-dad. Ciencia Hoy (Argentina) 5(30): 25-34.
LLAMBÍAS,E. & C. E. PROZZ!.1972. Ventania.Pp. 79-10 1. Relatorio del Sexto Congresode Geología Argentina. Buenos Aires, Ar-gentina.
MAURY,E. A. 1973. Los escorpiones de lossistemas serranos de la provincia de Bue-nos Aires. Physis (Buenos Aires) 32 (85):351-371.
MICHAUT,H. N. L. 1980. Sierras Pampeanas delnorte de Córdoba, sur de Santiago del Es-tero, borde oriental de Catamarca y ángu-lo sudeste de Tucumán. Vol. 1, pp. 293-347en: Segundo Simposio de Geología Regio-nal Argentina, Córdoba, Argentina.
MINGRAMM,A., A. Russo, A. Pozzo & L.CAZAU.1980. Sierras Subandinas. Vol. 1,pp. 95-137 en: Segundo Simposio de Geo-logía Regional Argentina, Córdoba, Ar-gentina.
MORRONE,J. J. 1994. Distributional patternsof species of Rhytirrhinini (Coleoptera:
Crisci el al.
Curculionidae) and the historicalrelationships ofthe Andean province. Glo-bal Ecology and Biogeography Letters 4:188-194.
MORRONE,J. J. & J. V. CRISC!. 1990. Panbio-geografía: fundamentos y métodos. Evo-lución Biológica (Bogotá) 4: 119-140.
MORRONE,J. J. & J. V. CRISC!.1995. Historicalbiogeography: introduction to methods.An nu a l Review of Ecology andSystematics 26: 373-40 l.
MORRONE,J. J., L. KATINAS& J. V. CRISC!.1997.A c1adistic biogeographic analysis of Cen-tral Chile. Joumal ofComparative Biology2 (1): 25-41.
Nrconx, E. G. 1970. Setaria Beauv. (Poaceae).Vol. 4 (2), pp. 439-460 en: A. L. Cabrera(ed.). Flora de la provincia de Buenos Ai-res. Colección Científica INT A, BuenosAires, Argentina.
PARODl,L. R. 1947. La estepa pampeana. Vol.8, pp. 143-207 en: L. Hauman, A. Burkart,L. R. Parodi & A. L. Cabrera (eds.), Lave-getación de la Argentina, Geografía de la Re-pública Argentina. Buenos Aires, Argentina.
PETER,G. 2000. Las especies silvestres de lafamilia de las Compuestas que habitan enel partido de Saavedra (provincia de Bue-nos Aires). Tesina, Universidad Nacionaldel Sur, Bahía Blanca, Argentina.
PONCE,M. M. 1982. Morfologíaecológicacom-parada de las Filicópsidas de las SierrasAustrales de Buenos Aires (República Ar-gentina). Boletín de la Sociedad Argenti-nade Botánica21 (1-4): 187-211.
PONCE,M. M. 1986. Morfología ecológica deplantas saxícolas del Cerro de la Ventana,Prov. de Buenos Aires, Argentina. Darwi-niana27 (1-4): 237-271.
PONTIROLl,A. 1965. Plantaginaceae. Vol. 4 (S),pp. 331-342 en: A. L. Cabrera (ed.). Florade la provincia de Buenos Aires. Colec-ción Científica lNT A, Buenos Aires, Ar-gentina.
PRADO,D. E. & P. E. GIBBS. 1993. Patterns ofspecies distributions in the dry seasonal
39

Asteraccac Irom Tandilia and Vcntania
forests of South America. Annals of theMissouri Botanical Garden 80: 902-927.
RAMos, V. A. 1989. The birth ofSouthern SouthAmerica. American Scientist 77 (5): 444-450.
RAVENNA,P. F. 1968. Iridaceae. Vo\. 4 (1), pp.539-565 en: A. L. Cabrera (ed.). Flora de laprovincia de Buenos Aires. Colección Cien-tífica INT A, Buenos Aires, Argentina.
RINGUELET,R. A. 1956. Panorama zoogeográ-fico de la provincia de Buenos Aires. No-tas del Museo de La Plata 18: 1-15.
ROLLERI, E. O. 1975. Provincias geológicasbonaerenses. Pp. 29-53 en: Relatorio delSexto Congreso de Geología Argentina.Buenos Aires, Argentina.
ROSEN,B. R. 1988. Frorn fossils to earth history:applied historical biogeography. Pp. 437-481 in: A. A. Myers & P. S. Giller (eds.).Analytical biogeography: an integratedapproach to the study of animal and plantdistributions. Chapman & Hall, London &NewYork.
ROSEN, B. R. & A. B. SMIlII. 1988. Tectonicfrorn fossils? Analysis of reef-coral andsea-urchin distributions from lateCretaceous to Recent, using a newmethod. Geological Society SpecialPublication n° 37, pp. 275-306 in: M. G.Audley & A. Hallam (eds.). Gondwana andTethys. Oxford Univ. Press, Oxford.
SAUSllURY,B. A. 1999. SECANT: strongest evi-den ce compatiblity analytic too\. Version2.2. DepartmentofEcology and EvolutionaryBiology, Yale University, New Haven.
SIMPSON,B. B. 1975. Pleistocene changes inthe flora of high tropical Andes. Paleo-biology 1: 273-294.
SIMPSONVUILlXUMIER,B. B. 1971. Pleistocenechanges in the fauna and flora of SouthAmérica. Science 173:771-774.
SOTA, E. A. DELA. 1967. Composición, origeny vinculación de la flora pteridológica delas sierras de Buenos Aires (Argentina).Boletín de la Sociedad Argentina de Botá-nica I1 (2-3): 105-128.
40
SOTA, E. A. DE I.A. 1972. Sinopsis de lasPteridófitas del Noroeste de la Argentina,I. Darwiniana 17: 11-27.
SOTA, E. A. 1)1' LA. 1973. La distribución geo-gráfica de las Pteridófitas en e Iconosur de América meridional. Boletín dela Sociedad Argentina de Botánica 15(1): 23-34.
SOTA,E. A. DELA. 1985. Las Pteridófitas de laprovincia de La Pampa, Argentina. Revis-ta de la Facultad de Agronomía, Universi-dad Nacional de La Pampa I (1-2): 23-34.
SPEGAZZlNI,C. L. 1896. Algunas observacio-nes sobre la flora de la Ventana. Revistade la Facultad de Agronomía y Veterinariade La Plata 2 (23-24): 347-349.
SPIGA77INI, C. L. 1901. Contribución al estu-dio de la flora de Tandil. Ed. Sessé,Larrañaga y Renovales, La Plata y Bue-nos Aires, Argentina.
SnJESSY,T. F., T. SANG& M. L. DI' VORE.1996.Phylogeny and biogeography 01' thesubfamily Barnadesioideae with impli-cations for early evolution of the Com-positae. Vo\. 1, pp, 463-490 in: D. 1. N. Hind& H. J. Beentje (eds.). Compositae:Systematics. Proceedings of the Interna-tional Compositae Conference Kew, 1994.Royal Botanic Gardens Kew.
SworFORD, D. L. 1999. PAUP*: Phylogeneticanalysis using parsimony. Version 4.0 beta 2.Sinauer Associates, Sunderland, Massachusetts.
TAYLOR, D. W. 1991. Paleobiogeographicrelationships of Andean angiosperms ofCretaceous to Pliocene age. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology88: 69-84.
TERUGGI, M. E. & 1. O. KILMlJRRAY. 1975.Tandilia. Pp. 55-77 en: Relatorio del SextoCongreso de Geología Argentina. BuenosAires, Argentina.
TERlJGGI,M. E. & J. O. KILMURRAY.1980. Sie-rras septentrionales de la provincia deBuenos Aires. Vo\. 2, pp. 918-965 en: Se-gundo Simposio de Geología Regional Ar-gentina, Córdoba, Argentina.

TORRES,M. A. 1970a. Piptochaetium Presl(Poaceae). Vol. 4 (2), pp. 234-255 en: A. L.Cabrera (cd.). Flora de laprovinciade Bue-nos Aires. Colección Científica INT A,Buenos Aires, Argentina.
TORRES,M. A. 1970b. roa L. (Poaceae). Vol. 4(2), pp. 102-125 en: A. L. Cabrera (ed.). Flo-ra de la provincia de Buenos Aires. Colec-ción Científica INTA, Buenos Aires, Ar-gentina.
TROfANf,H. O. & P. E. STEIBEL.1999. Sinopsisde las compuestas (Compositae Giseke)de la provincia de La Pampa, República Ar-gentina. Revista de la Facultad de Agrono-m ía de La Pampa, serie supl. 1, 10: 1-86.
Recibido: noviembre 1/2000Aceptada: febrero 2012001
Crisci el al.
VEGA, L. & P. BELLAGAMBA.1990. Lista co-mentada de la herpetofauna de las Sierrasde Balcarce y Mar del Plata, Buenos Ai-res, Argentina. Cuadernos de Herpetología5(2): 10-14.
ZAMBRANO,1. 1. 1980. Comarca de la cuencacretácica de Colorado. Vol. 2, pp. 1033-1070en: Segundo Simposio de Geología Regio-nal Argentina, Córdoba, Argentina.
ZUIOAGA,F. O. & O. MORRONE(eds.). 1999.Catálogo de las plantas vasculares de laRepública Argentina 11. Acanthaceae-Euphorbiaceae (Dicotyledoneae).Monographs in Systematic Botany fromthe Missouri Botanical Garden 74: 1-621.
41