by herbert weissbach, betty g. redfield, and … · soluble monoamine oxidase; its properties and...
TRANSCRIPT

SOLUBLE MONOAMINE OXIDASE; ITS PROPERTIES AND ACTIONS ON SEROTONIN
BY HERBERT WEISSBACH, BETTY G. REDFIELD, AND SIDNEY UDENFRIEND
(Ir‘ om the Laboratory of Clinical Biochemistry, National Heart Institute, National Institutes of Health, Public Health Service, United States Department of
Health, Education, and Welfare, Bethesda, Maryland)
(Received for publication, July 24, 1957)
5-Hydroxyindoleacetic acid (5HIAA) has been shown to be the major metabolite of 5-hydroxytryptamine (serotonin) (1). In man 2 to 8 mg. of the acid are normally excreted in the urine per day (2). This suggests that the major route of metabolism of serotonin proceeds via oxidative deamination, catalyzed, presumably, by monoamine oxidase (MAO). Experiments carried out in vitro (3-6) and in viva demonstrate that MAO is a key substance in the physiology of serotonin and other amines.
MAO has been found in most organ tissues in which it is primarily localized in the mitochondria (5). The inability to make soluble the mito- chondrial enzyme has prevented studies concerning its cofactor require- ments and its mechanism of action. However, although the bulk of MAO activity is in mitochondria, appreciable activity is present in soluble form in some tissues. This *MAO lends itself to standard procedures of enzyme purification.
The present report deals with the purification of soluble MAO from guinea pig liver and with preliminary studies on its properties and mode of action . A comparison is also made with mitochondrial MAO.
Materials and Methods
Male guinea pigs (Hartley or Kational Institutes of Health strain) weighing about 250 gm. were used in these studies. The 5-hydroxyindoles employed were obtained from the California Foundation for Biochemical Research. Diphosphopyridine nucleotide (DPK) and triphosphopyridine nucleotide (TPN) were obtained from the Sigma Chemical Company.
Incubations were carried out in 20 ml. beakers by using an Aminco metabolic shaker at 37”, for 3 hour, in air. Serotonin was determined by the method of Udenfriend et al. by employing the 1-nitroso-2-naphthol color reaction (7).
The 5HIAA formed after incubation was assayed as follows: The incu- bation mixture was transferred to a 40 ml. glass-stoppered shaking tube containing 1 gm. of salt and 0.3 ml. of 6 N HCl. The incubation beaker was rinsed with 1 ml. of 0.2 N HCl, and the washing was added to the glass-
953
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

954 SOLUBLE MONOAMINE OXIDASE
stoppered tube. 15 ml. of ether were added, and the tube was shaken for 5 minutes. The phases were separated by centrifugntion, and 10 ml. of the ether layer were transferred to another glass-stoppered tube containing 1.5 ml. of 0.5 M phosphat’e buffer, pH 7.0. The tube was again shaken for 5 minutes and centrifuged. The ether layer was discarded, 1 ml. of the buffer was t,ransferred to another tube, and the 5HIAA was assayed colori- metrically by employing the l-nitroso-2-naphthol reaction (7). Standards and blanks were treated in the same manner. 25 y of 5HIAA, carried through the entire procedure, gave an optical density of 0.150 at 540 rnp.
Tissues were homogenized in water (1 part of tissue with 5 parts of water) by using either a ground glass homogenizer or a Waring blendor. The tissue homogenate was first centrifuged at 8000 X g for 20 minutes in an International angle head centrifuge (refrigerated), and the residue was discarded. The supernatant fluid was then centrifuged in a Spinco pre-
TABLE I
High speed supernatant fluid.. Ammonium sulfate.. Acid ppt..
‘21 ~ iii i ii ~
.____
‘f
I
* Micromoles of serotonin destroyed per mg. of protein per hour.
parative ultracentrifuge at 100,000 X g for 30 minutes. The supernatant fluid obtained after this centrifugation will be referred to as the high speed supernatant fluid. For further purification, the high speed supernatant fluid wa.s made 25 per cent sat.urated wit.h respect to ammonium sulfate by the addition of saturated ammonium sulfate solution adjusted to pH 8, and the precipitate was discarded. The supernatant fluid was then nd- justed to 40 per cent saturation with ammonium sulfate. The resulting precipitate was dissolved in one-third the volume of the initial high speed supernatant fluid with 0.01 M phosphate buffer, pH 7.4. The pH was then adjusted to 5.0 to 5.1 by the dropwise addition of 0.5 x acetic acid, and the pr&cipitate formed was quickly isolated by centrifugation and dissolved in 0.05 M phosphate buffer, pH 7.4. This precipitate contained the MAO. The relative activities of the various fractions are shown in Table I. The fractions at any stage could he dialyzed against phosphate buffer, pH 7.4, for 5 hours without any loss of activity, but dialysis against water resulted in rapid loss of activit,y in several hours. The MAO could also be kept at 3” for several days with little loss of activity. Specific activity is de-
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

H. WEISSBACH, B. G. REDFIELD, S. UDENFRIEND 955
fined as micromoles of serotonin destroyed or 5HIAA formed per mg. of protein per hour.
The mitochondria were isolated by differential centrifugation according to the method of Hogeboom el al. (8).
Protein was determined by the method of Warburg and Christian (9) as modified by Kalckar (10). MAO activity was also assayed manometri- tally essentially as described by Creasey (ll), and ammonia was assayed by the method of Johnson (12). Catalase was determined by the method of Beers and Sizer (13).
Guinea pig liver.. I‘ “ kidney
Rat liver. ‘( kidney......
Rabbit lung.. “ brain...... “ liver, “ kidney.....
TABLE II
Soluble MAO Activities in Various Tissues
Tissue pm& serotonin destroyed per hr. -
0.91 0.25 0.09 0 0.03 0 0.02 0
The’tissues were homogenized in 5 parts of HzO, and a high speed supernatant fluid was prepared. The complete system contained 1 ml. of supernatant fluid, 0.5 ml. of 0.5 M phosphate, pH 8.1, and 300 y of serotonin in a total volume of 3.5 ml.
EXPERIMENTAL
Occurrence of Soluble MAO-In all tissues examined the MAO activity was found mainly in the mitochondria, as measured by serotonin disap- pearance activity, but in certain tissues activity was also observed in the high speed supernatant fluid. Table II summarizes the relative activity of this soluble MAO in a variety of tissues tested. Guinea pig liver was the most active source and, therefore, was used in the studies presented here. Diamine oxidase has been reported to attack monoamines (14), but early studies showed that the enzyme under study here was inhibited by marsilid and other MAO inhibitors, while semicarhazide, a known dia- mine oxidase inhibitor, was ineffective.
Studies on SHIAA Formation-In previous studies with crude tissue preparations, it was observed that only 30 per cent of the serotonin metab- olized could be recovered as 5HIAA (4). It was noticed in this present system that, although the specific activity of the enzyme as measured by serotonin destruction indicated substantial purification, the specific activ- ity based on 5HIAA formed did not significantly increase (Table III). The
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

956 SOLUBLE MONOAMINE OXIDASE
addition of either acid filtrate or high speed supernatant fluid from guinea pig kidney to all the fractions, along with DPN, resulted in stoichiometric conversion of serotonin to 5HIAA. DPN was much more effective in this system than TPN, indicating that the second enzyme was an aldehyde dehydrogenase similar to that previously described (15). The acid filtrate and the high speed supernatant fluid were tested with a variety of alde-
TABLE III
Serolonin Destruction and 6HIAA Formation in Various Enzyme Fractions
Fraction Additions
High speed supernatant fluid “ “ ‘I I‘ “ “ “ “
Ammonium sulfate ‘I ‘I “ “
Acid ppt. I‘ I‘ “ “
None DPN
“ + aldehyde dehydrogen- ase
None DPN
“ + aldehyde dehydrogen- ase
None DPN
“ + aldehyde dehydrogen- as8
i
_-
-
0.05 0.05 0.05
0.16 0.16 0.16
0.60 0.60 0.60
-.
I
.-
-
rmole
E2 per mp. protem
er ?I r.
-~~ 0.02 0.04 0.05
0.04 0.14 0.16
0.04 0.06 0.60
The protein concentrations per ml. of the enzyme fractions were as follows: high speed supernatant fluid, 16.8 mg.; ammonium sulfate, 8.1; and acid precipitate, 3.3. 0.5 ml. of enzyme was used in each beaker. 0.1 ml. of high speed supernatant fluid from guinea pig kidney (13 mg. per ml.) was used as the source of aldehyde dehydro- genase. The incubation mixture contained 1 mg. of serotonin, 0.5 ml. of 0.5 M phosphate buffer, pH 8.1, in a total volume of 3.5 ml. 1 rmole of DPN was used where indicated.
hydes (butyraldehyde, etc.) and were shown to be active with respect to aldehyde dehydrogenase as determined by DPNH formation. The over- all scheme for SHIAA formation can, therefore, be summarized as follows:
MAO Serotonin A 5-hydroxyindoleacetaldehyde
aldehyde dehydrogenase
DPN ) 5HIAA
It appeared that MAO and the aldehyde dehydrogenase system were both present in the high speed supernatant fluid and the ammonium sul- fate fraction, while only MAO was present in the acid precipitate.
In the presence of DPN and an excess of aldehyde dehydrogenase it
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
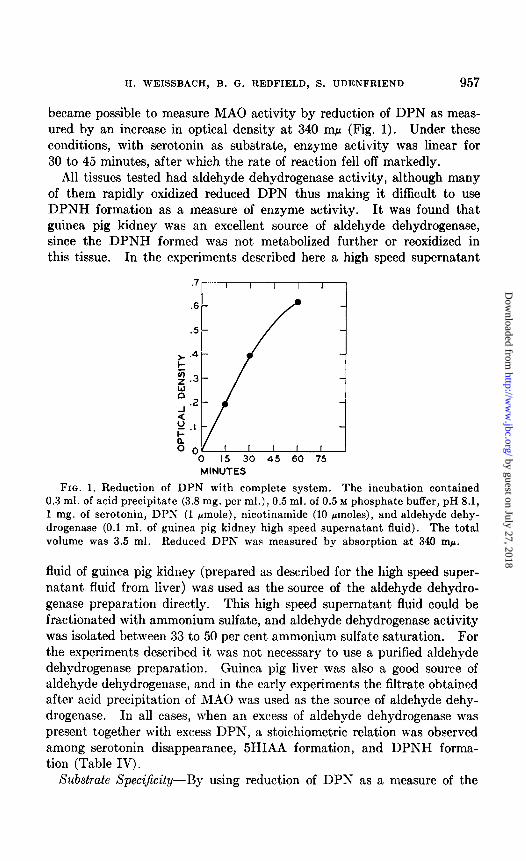
1%. WEISSBACH, B. G. REDFIELD, S. UDENFRIEND 957
became possible to measure MAO activity by reduction of DPN as meas- ured by an increase in optical density at 340 rnp (Fig. 1). Under these conditions, with serotonin as substrate, enzyme activity was linear for 30 to 45 minutes, after which the rate of reaction fell off markedly.
All tissues tested had aldehyde dehydrogenase activity, although many of them rapidly oxidized reduced DPN thus making it difficult to use DPNH formation as a measure of enzyme activity. It was found that guinea pig kidney was an excellent source of aldehyde dehydrogenase, since the DPNH formed was not metabolized further or reoxidized in this tissue. In the experiments described here a high speed supernatant
( ( , , , ,
I I I I I
0 15 30 45 60 75
MINUTES
FIG. 1. Reduction of DPN with complete system. The incubation contained 0.3 ml. of acid precipitate (3.8 mg. per ml.), 0.5 ml. of 0.5 M phosphate buffer, pH 8.1, 1 mg. of serotonin, DPN (1 rmole), nicotinamide (10 pmoles), and aldehyde dehy- drogenase (0.1 ml. of guinea pig kidney high speed supernatant fluid). The total volume was 3.5 ml. Reduced DPPI’ was measured by absorption at 340 rnp,
fluid of guinea pig kidney (prepared as described for the high speed super- natant fluid from liver) was used as the source of the aldehyde dehydro- genase preparation directly. This high speed supernatant fluid could be fractionated with ammonium sulfate, and aldehyde dehydrogenase activity was isolated between 33 to 50 per cent ammonium sulfate saturation. For the experiments described it was not necessary to use a purified aldehyde dehydrogenase preparation. Guinea pig liver was also a good source of aldehyde dehydrogenase, and in the early experiments the filtrate obtained after acid precipitation of MAO was used as the source of aldehyde dehy- drogenase. In all cases, when an excess of aldehyde dehydrogenase was present together with excess DPN, a stoichiometric relation was observed among serotonin disappearance, 5HIAA formation, and DPNH forma- tion (Table IV).
Substrate Specificity-By using reduction of DPN as a measure of the
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

958 SOLUBLE MONOAMINE OXIDASE
soluble MAO activity, it was possible to compare the activity of a variety of substrates, as shown in Table V. The best substrates were those amines
TABLE IV
Relationship a?nong Serotonin Destroyed, 5HIAA Formed, and DPN Reduced by Using Complete System
Specific activity, ~mole per mg. protein per hr.
Serotonin destroyed SHIAA formed DPN reduced
0.62 0.60 0.61
Results of a typical experiment with acid precipitate (1.9 mg.), 0.5 ml. of 0.5 M
phosphate buffer, pH 8.1, 1 @mole of DPN, 10 Mmoles of nicotinamide, and aldehyde dehydrogenase (0.1 ml. of high speed supernatant fluid from guinea pig kidney, 13.3 mg. of protein per ml.). The DPNH formed was measured by its absorption at 340 rnr by using 6.22 X lo3 optical density units as its molar extinction coefficient.
TABLE V
Relative Activity of Substrate
Oxygen uptake Substrate
Mitochondria Acid ppt. -
Serotonin................... 100 100 Tyramine................... 240 190 Tryptamine. 100 123 Heptylamine................ 20 9 Noradrenaline. 20 30 Benzylamine. 0 0 Phenylethylamine. 0 0 Histamine. 0 0 Spermine 0 0
-
DPN formation, acid ppt.
100 180
93 13 18
2 6 0 0
In the manometric experiments a final substrate concentration of 4 X 1OW M was employed. For these experiments 0.5 ml. of guinea pig mitochondrial suspen- sion, equivalent to 100 mg. of original liver, and a concentrated acid precipitate (1 ml. containing 18.2 mg. per ml.) were employed. The experiments with mito- chondria were buffered at pH 7.4, while buffer of pH 8.1 was used with the acid pre- cipitate. TheDPNH experiments were performed essentially as described in Fig. 1, with a final concentration of substrate of 1.6 X lo+.
which were also excellent substrates for the mitochondrial MAO. Dia- mines were not oxidized by this system. By assaying MAO in this manner, it was assumed that the aldehyde dehydrogenase, which converted the aldehyde intermediate to 5HIAA in the oxidation of serotonin, was also active on the aldehydes formed when other amines were oxidatively
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
H. WEISSBACH, B. G. REDFIELD, S. UDENFRIEND 959
deaminated by MAO. We, therefore, verified t.he substrate specificity by employing oxygen uptake as a criterion of enzyme activity (Table V). A good correlation was observed with all the substrates tested. The weak activity, about one-tenth of the soluble enzyme as compared with the mito- chondrial MAO, made manometric experiments impractical as a general method of measuring the activity of the soluble system.
Effect of pH and Substrate Concentration-The optimal pH was found to be about 8.1. Incubations were generally carried out at pH 8.1 with phosphate buffer.
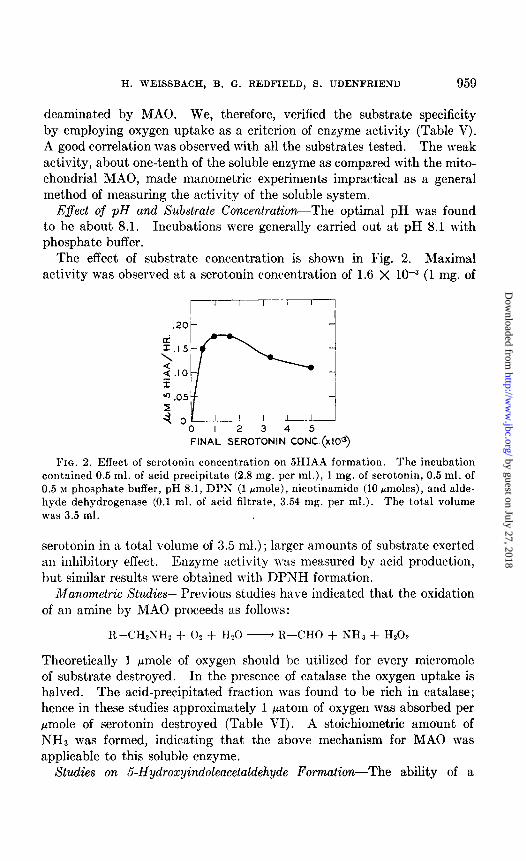
The effect of substrate concentration is shown in Fig. 2. Maximal activity was observed at a serotonin concentration of 1.6 X 10e3 (1 mg. of
012345
FINAL SEROTONIN CONC.(xlO-3)
FIG. 2. Effect of serotonin concent,ration on 5HIAA formation. The incubation contained 0.5 ml. of acid precipitate (2.8 mg. per ml.), 1 mg. of serotonin, 0.5 ml. of 0.5 M phosphate buffer, pH 8.1, DPS (1 pmole), nicotinamide (10 rmoles), and alde- hyde dehydrogenase (0.1 ml. of acid filtrate, 3.54 mg. per ml.). The total volume was 3.5 ml.
serotonin in a t,otal volume of 3.5 ml.); larger amounts of substrate exerted an inhibitory effect. Enzyme :tctivit,y was measured by acid production, but similar results were obtained with DPNH formation.
Manometric Studies-Previous studies have indicated that the oxidation of an amine by MAO proceeds as follows:
It--C’H?NH, + 02 + II& - R--CHO + lYH3 + H20.z
Theoretically 1 pmole of oxygen should be utilized for every micromole of substrate destroyed. In the presence of catalase the oxygen uptake is halved. The acid-precipitated fraction was found to be rich in catalase; hence in these studies approximately 1 patom of oxygen was absorbed per pmole of serotonin destroyed (Table VI). A stoichiometric amount of NH, was formed, indicating that the above mechanism for MAO was applicable to this soluble enzyme.
Studies on 5-Hydroxyindoleacetaldehyde Form&ion-The ability of a
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
960 SOLUBLE MONOAMINE OXIDASE
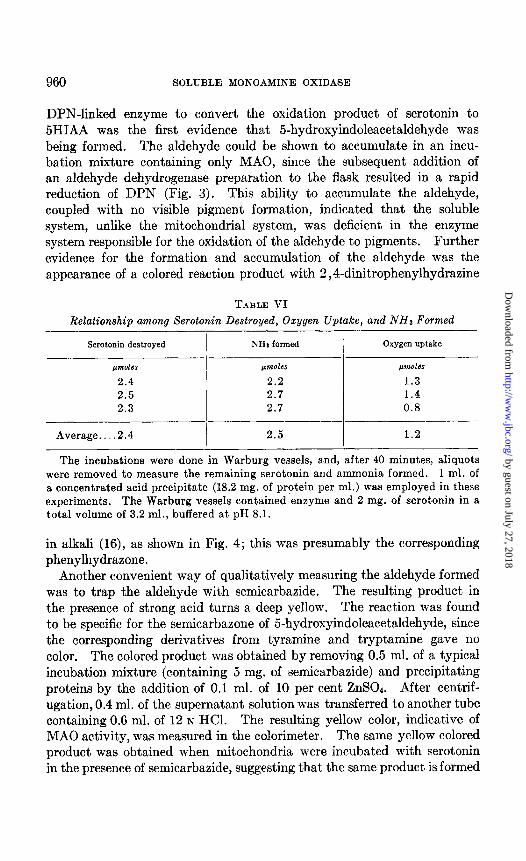
DPN-linked enzyme to convert the oxidation product of serotonin to 5HIAA was the first evidence that 5-hydroxyindoleacetaldehyde was being formed. The aldehyde could be shown to accumulate in an incu- bation mixture containing only MAO, since the subsequent addition of an aldehyde dehydrogenase preparation to the flask resulted in a rapid reduction of DPN (Fig. 3). This ability to accumulate the aldehyde, coupled with no visible pigment formation, indicated that the soluble system, unlike the mitochondrial system, was deficient in the enzyme system responsible for the oxidation of the aldehyde to pigments. Further evidence for the formation and accumulation of the aldehyde was the appearance of a colored reaction product with 2,4-dinitrophenylhydrazine
TABLE VI Relationship among Serotonin Destroyed, Oxygen Uptake, and NH3 Formed
Serotonin destroyed NH: formed Oxygen uptake
pmolcs ~moles pmoles
2.4 2.2 1.3 2.5 2.7 1.4 2.3 2.7 0.8
--- ---____---- Average....2.4 2.5 1.2
The incubations were done in Warburg vessels, and, after 40 minutes, aliquots were removed to measure the remaining serotonin and ammonia formed. 1 ml. of a concentrated acid precipitate (18.2 mg. of protein per ml.) was employed in these experiments. The Warburg vessels contained‘enzyme and 2 mg. of serotonin in a total volume of 3.2 ml., buffered at pH 8.1.
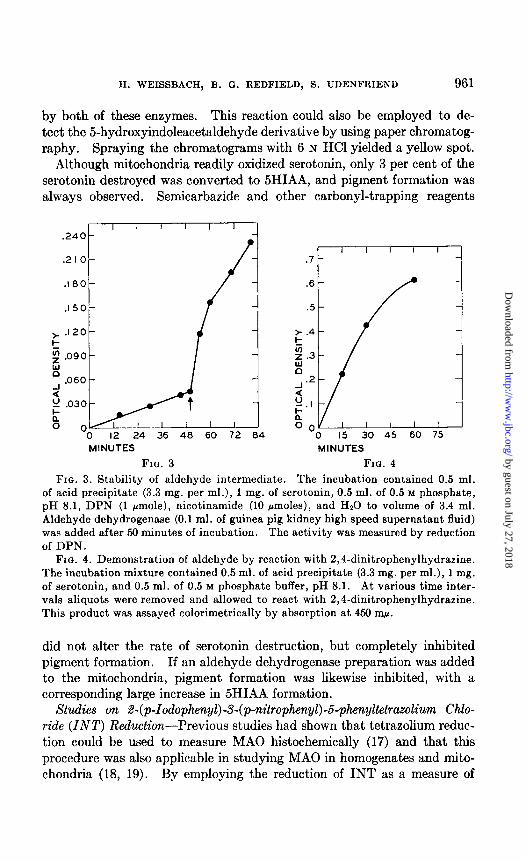
in alkali (16), as shown in Fig. 4; this was presumably the corresponding phenylhydrazone.
Another convenient way of qualitatively measuring the aldehyde formed was to trap the aldehyde with semicarbazide. The resulting product in the presence of strong acid turns a deep yellow. The reaction was found to be specific for the semicarbazone of 5-hydroxyindoleacetaldehyde, since the corresponding derivatives from tyramine and tryptamine gave no color. The colored product was obtained by removing 0.5 ml. of a typical incubation mixture (containing 5 mg. of semicarbazide) and precipitating proteins by the addition of 0.1 ml. of 10 per cent ZnSO1. After centrif- ugation, 0.4 ml. of the supernatant solution was transferred to another tube containing 0.6 ml. of 12 N HCl. The resulting yellow color, indicative of MAO activity, was measured in the calorimeter. The same yellow colored product was obtained when mitochondria were incubated with serotonin in the presence of semicarbazide, suggesting that the same product is formed
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
H. WEISSBACH, B. G. REDFIELD, S. UDENFRIEND 961
by both of these enzymes. This reaction could also be employed to de- tect the 5-hydroxyindoleacetaldehyde derivative by using paper chromatog- raphy. Spraying the chromatograms with 6 N HCl yielded a yellow spot.
Although mitochondria readily oxidized serotonin, only 3 per cent of the serotonin destroyed was converted to 5HIAA, and pigment formation was always observed. Semicarbazide and other carbonyl-trapping reagents
I I I I I I .240 -
.180-
l:;;;b, ,i 0 0 12 12 24 24 36 36 48 48 60 60 72 72 84 84
MINUTES MINUTES
FIG. 3 FIQ. 4
FIG+. 3. Stability of aldehyde intermediate. The incubation contained 0.5 ml. of acid precipitate (3.3 mg. per ml.), 1 mg. of serotonin, 0.5 ml. of 0.5 M phosphate, pH 8.1, DPN (1 pmole), nicotinamide (10 pmoles), and H*O to volume of 3.4 ml. Aldehyde dehydrogenase (0.1 ml. of guinea pig kidney high speed supernatant fluid) was added after 50 minutes of incubation. The activity was measured by reduction of DPN.
Fro. 4. Demonstration of aldehyde by reaction with 2,4-dinitrophenylhydrazine. The incubation mixture contained 0.5 ml. of acid precipitate (3.3 mg. per ml.), 1 mg. of serotonin, and 0.5 ml. of 0.5 M phosphate buffer, pH 8.1. At various time inter- vals aliquots were removed and allowed to react with 2,4-dinitrophenylhydrazine. This product was assayed calorimetrically by absorption at 450 mp.
did not alter the rate of serotonin destruction, but completely inhibited pigment formation. If an aldehyde dehydrogenase preparation was added to the mitochondria, pigment formation was likewise inhibited, with a corresponding large increase in 5HIAA formation.
Studies on d-(p-Iodop~yl)-S-(p-nitrophenyl)-5-phenyltetrazolium Chlo- ride (INT) Reduction-Previous studies had shown that tetrazolium reduc- tion could be used to measure MAO histochemically (17) and that this procedure was also applicable in studying MAO in homogenates and mito- chondria (18, 19). By employing the reduction of INT as a measure of
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

962 SOLUBLE MONOAMINE OXIDASE
mitochondrial MAO act,ivity (19), it was shown that only serotonin and tryptamine were suitable substrates. Inhibitor studies showed that an indole aldehyde was necessary for INT reduction, and it was concluded that the further oxidation of the indole aldehyde resulted in the INT re- duction. The soluble MAO studied here afforded a better opportunity to study INT reduct,ion. The acid-precipitated fraction readily reduced TNT in the presence of an indole amine. As reported earlier (19), a lag phase was always observed when one measured INT reduction with mito- chondria as t,he source of MAO. This lag phase was also observed with the soluble MAO. Similar to the mitochondrial system, indole amines were the best substrates for INT reduction.
DISCIJSSIOK
The physiologic significance of the soluble MAO described here remains to be determined. However, since it is similar t.o the mitochondrial MAO in other properties, it offers several advantages over the particulate enzyme. It can be purified and separated from aldehyde dehydrogenase. Being soluble, it should be possible to purify it, sufficiently to determine its prop- erties and requirements.
Substantial evidence has also been presented demonstrating that 5- hydroxyindoleacetaldehyde is the intermediate in the oxidation of sero- tonin to 5HIAA. The aldehyde has been demonstrated by the formation of a colored product with 2,4-dinitrophenylhydrazine and also by its reaction with semicarbazide. The studies in vitro have also indicated that the production of 5HIAA is dependent on a DPN-linked aldehyde dehy- drogenase. Since 5-hydroxyindoleacetaldehyde is formed in the oxidation of serotonin, the possibility that this compound may exert some pharma- cologic effects attributed to serotonin should not be overlooked.
It has also been possible to apply t,his system to the metabolism of the catechol amines. Preliminary experiments with epinephrine and norepi- nephrine indicate that 3,4-dihydroxymandelic acid is formed by this soluble MAO system, if sufficient DPN and aldehyde dehydrogenase are present.’ Further studies on the actions of MAO on the catechol amines are now in progress.
SUMMAIiY
1. A soluble monoamine oxidase (MAO) present in guinea pig liver has been described. Purification has been achieved by using ammonium sul- fate fractionation and acid precipitation.
2. The conversion of serotonin to 5-hydroxyindoleacetic acid (5HIAA) has been studied. The system has been divided into two part.s, thus mak-
1 Leeper, L. C., Weissbach, H., and IJdenfriend, S., unpublished observations.
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

H. WEISSBACH, B. G. REDFIELD, S. UDENFRIEND 963
ing it possible to accumulate and demonstrate 5-hydroxyindoleacetalde- hyde. The aldehyde has been shown to be converted to the acid by a diphosphopyridine nucleotide (DPN)-linked system. In the presence of the complete system, quantitative conversion of serotonin to 5HIAA could be obtained, with a stoichiometric reduction of DPN.
3. Manometric experiments showed that 1 rmole of oxygen was absorbed and 1 pmole of NH, formed per pmole of amine oxidized and that peroxide was produced. The substrate specificity of this soluble system was similar to that of the mitochondrial MAO.
4. Some characteristics of both this soluble MAO and the mitochondrial enzyme have been discussed.
The authors are indebted to Dr. C. Mitoma for his many helpful sug- gestions during t.he completion of this work.
BIBLIOGRAPHY
1. Udenfriend, S., Titus, E., Weissbach, H., and Peterson, R. E., J. Biol. Chem., 219, 335 (1956).
2. Udenfriend, S., Titus, E., and Weissbach, H., J. Biol. Chem., 216, 499 (1955). 3. Blaschko, H., Pharmacol. Rev., 4, 415 (1952). 4. Udenfriend, S., and Titus, E., in McElroy, W. D., and Glass, B., Amino acid
metabolism, Baltimore, 945 (1955). 5. Zeller, E. A., Barsky, J., and Berman, E. R., J. Biol. Chem., 214. 267 (1955). 6. Sjoerdsma, A., Smith, ‘I’. E., Stevenson, T. D., and Udenfriend, S., Proc. Sot.
Exp. Biol. and Med., 89, 36 (1955). 7. Udenfriend, S., Weissbach, H., and Clark, C. T., J. Biol. Chem., 216,337 (1955). 8. Hogeboom, G. H., Schneider, W. C., and Pallade, G. E., J. Biol. Chem., 172, 619
(1948). 9. Warburg, O., and Christian, W., Biochem. Z., 310,384 (1941-42).
10. Kalckar, H. M., J. Biol. Chem., 167, 461 (1947). 11. Creasey, N. H., Biochem. J., 64,178 (1956). 12. Johnson, M. J., J. Biol. Chem., 137,575 (1941). 13. Beers, R. F., Jr., and Sizer, I. W., J. Biol. Chem., 196, 133 (1952). 14. Fouts, J. R., Blanksma, L. A., Carbon, J. A., and Zeller, E. A., J. Biol. Chem., !226,
1025 (1957). 15. Racker, E., J. Biol. Chem., 177, 883 (1949). 16. Friedemann, T. E., and Haugen, G. E., J. Biol. Chem., 147,415 (1943). 17. Glenner, G. G., Burtner, H., and Brown, G. W., Jr., J. Histochem. and Cytochem.,
6, 591 (1957). 18. Lagnado, J. R., andsourkes, T. L., Canad. J. Biochem. and Physiol., 34,1095 (1956). 19. Weissbach, H., Redfield, B. G., Glenner, G. G., and Mitoma, C., J. Histochem.
and Cytochem., 6, 601 (1957).
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

Sidney UdenfriendHerbert Weissbach, Betty G. Redfield and
SEROTONINPROPERTIES AND ACTIONS ON
SOLUBLE MONOAMINE OXIDASE; ITS
1957, 229:953-963.J. Biol. Chem.
http://www.jbc.org/content/229/2/953.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/229/2/953.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on July 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from