body part representation, fragmentation and patterns of ertebølle deer exploitation in northwest...
TRANSCRIPT

Body Part Representation, Fragmentationand Patterns of Ertebølle Deer Exploitationin Northwest Zealand, DenmarkK. J. GRON*Department of Anthropology, University of Wisconsin-Madison, Madison, WI, USA
ABSTRACT The faunal assemblages from Asnæs Havnemark, Fårevejle and Trustrup, located in northwest Zealand,Denmark, yielded extensive yet substantially fragmented bone material dating predominantly to the lateMesolithic Ertebølle culture, ca. 5400–3950 cal BC. This paper presents a combined assessment of skeletalpart representation of red deer (Cervus elaphus) and roe deer (Capreolus capreolus) from these three sites inconjunction with quantification of overall fragmentation data for the assemblages. These data indicate that theobserved pattern of skeletal part abundance is most consistent with a pattern of density-mediated skeletal at-trition, and therefore the whole carcasses were initially deposited. Deer were not part of a logistic exploitationstrategy at these sites. This indicates variability during the Ertebølle period not only in which resources wereexploited, but in how these resources were processed. Copyright © 2013 John Wiley & Sons, Ltd.
Key words: body part representation; bone density; deer; Ertebølle; fragmentation; Mesolithic; resourcemovement
Introduction
The Ertebølle culture (hereafter EBK) of southernScandinavia represents some of the last hunter-gatherersin Europe whose hunting and gathering economy wasreplaced by an agricultural way of life starting around3950 B.C. (Price and Gebauer, 2005). SouthernScandinavia at the time of the Late Atlantic period(ca. 7000–3900 cal BC) was one of extensive environ-mental variability, where many different types ofresource zones were visited to obtain a broad spectrumof resources from the land, sea and air (Paludan-Müller, 1978; Aaris-Sørensen, 2009). Faunal assem-blages recovered from EBK sites can be exceedinglyrich and variable in species, reflecting a complexeconomic base for EBK hunters, gatherers and fishers(Ritchie et al., in press).The EBK hunter-gatherers utilised a toolkit well suited
to the securing of these resources, including specialisedhunting and fishing equipment (Price and Gebauer,2005; Price and Noe-Nygaard, 2009). Examples of thesetechnologies include the bow and arrow, equipped withboth flint-tipped and blunt-tipped arrows, boats, leisters,
fishhooks, fishing structures and other equipmentspecifically for the procurement of particular resources(Andersen, 1985, 1987, 2011; Pedersen, 1997). If itwas available in the environment, then EBK groupshad developed means to obtain it.Under the general model of resource procurement
as proposed by Rowley-Conwy (1983, 1999), basedon Binford (1980) and elaborated by Price andGebauer (2005), EBK collectors ventured out frombase camps to smaller, short-term collecting sites,and then brought these resources back to the moregeneralised base camps. The model involves bothradial and logistic components (Rowley-Conwy,1999). This perspective emphasises an exploitativestrategy consistent with a collector model of hunter-gatherer resource exploitation (Binford, 1980;Rowley-Conwy, 1983; Price and Gebauer, 2005).However, this general strategy may have variedwithin southern Scandinavia during the last centuriesof the Atlantic Period. Possible indicators includeobserved differences in material culture and resourceexploitation between regions as well as variation inthe available fauna (Aaris-Sørensen, 1980, 2009;Petersen, 1984; Ritchie, 2010).In this paper, faunal evidence is presented from
several localities in northwest Zealand, Denmark,which indicate a similarity in different body part repre-sentation in deer recovered from excavations. The data
* Correspondence to: Kurt J. Gron, Department of Anthropology, University ofWisconsin-Madison, Madison, WI, USA.e-mail: [email protected]
Copyright © 2013 John Wiley & Sons, Ltd. Received 8 January 2013Revised 28 June 2013Accepted 29 July 2013
International Journal of OsteoarchaeologyInt. J. Osteoarchaeol. (2013)Published online in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/oa.2339

derives from three Ertebølle sites, all located in differ-ent geographic settings (Figure 1) in the same generalregion of the island. Relative abundance of differentbody parts is presented in conjunction with overallassemblage fragmentation data, which illustrates apattern of density-mediated attrition of axial skeletalelements. This means that nearly all of the carcasseswere brought to the site intact by EBK hunters.Furthermore, the evidence demonstrates a correlationbetween the observed pattern of skeletal attritionand bone density, indicating that conclusionsconcerning faunal data hold little weight withoutdetailed consideration of overall fragmentation. Theobserved similarity between the three sites indicatesan absence of large-scale butchery and movement ofselected parts of deer across the landscape. Roe deer(Capreolus capreolus) and red deer (Cervus elaphus) weretreated the same at multiple locations in northwestZealand. This pattern is possibly indicative of theubiquity of these species at all locations, precludinga need for transport. It may be that deer were not partof a logistic resource extraction strategy in this regionof Denmark, pointing to possible differences betweenEBK regions in this regard. Finally, these data under-score the importance of careful and consistent recov-ery of faunal remains in the field, and detailedquantification of fragmentation during analysis.
Materials
Three faunal assemblages from Asnæs Havnemark,Fårevejle and Trustrup (Figure 1) were analysed as apart of this study. Some of the data presented herehave elsewhere been discussed in a different form(Gron, 2013; Ritchie et al., in press). All sites are datedthrough associated diagnostic artefacts or acceleratormass spectrometry to the late Mesolithic Ertebølleperiod (Ritchie et al., in press; T.D. Price, personalcommunication), and all sites were excavated underthe direction of T. Douglas Price by using similarmethodology, which included the water screening ofall materials through a 4mm mesh. Every single boneincluded in this study was recovered either prior to,or during sieving. Therefore, the assemblages reportedhere explicitly include only bones recovered fromsieving and those large enough to be recognised andcollected prior to water sieving. A summary of detailsconcerning the three sites is presented in Table 1.Some middle Neolithic dates (Funnel Beaker Culture,
TRB) were obtained from Fårevejle, and some clearlyNeolithic material was also recovered from all three sites.In all cases, however, these finds derived from upperlayers, were limited in number, and/or were from layersyielding very little faunal material. Therefore, themajorityof the collections reported here are almost certainlyErtebølle in date and predominantly from the late EBK.Possible interpretations of the sites presently indi-
cate different patterns of human occupation. AsnæsHavnemark may have been a residential site with visitsor occupation over much of the year. Fårevejle is anon-residential shell midden probably visited in onlylimited periods of the year. Trustrup has yieldedinconclusive seasonality information but may be bestdescribed as a seasonal residential area. In all cases,these interpretations must only be considered prelimi-nary at this stage (T.D. Price, personal communication).This study focuses only on the remains of roe deer and
red deer, two of the most abundant game mammalsutilised in northwest Zealand during the EBK. Deer rep-resent the majority of the identifiable sample in terms ofNISP at these three sites (Table 2) (Ritchie et al., in press).Significant use of these species is typical at Ertebølle sites,even though human diets were dominated by aquaticfoods according to isotopic studies (Tauber, 1981;Noe-Nygaard, 1988; Price and Gebauer, 2005; Fischeret al., 2007; Enghoff, 2011). Wild boar are omitted inthis study because of morphological differences withdeer and reported differences in butchery and transportof this species with others in EBK contexts (Rowley-Conwy, 1998). At Ringkloster, for example, al-though butchery and subsequent transport of bodyFigure 1. Locations of Danish archaeological sites discussed in the text.
K. J. Gron
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)
parts from the site is evident, the specific elementsapparently being extracted from the site differ betweendeer and boar (Rowley-Conwy, 1998). In addition, thespecies is rare compared with deer at EBK sites onZealand (Møhl, 1971; Noe-Nygaard, 1995; Gotfredsen,1998; Magnussen, 2007).
Methods
Of interest for understanding the purpose of a hunter-gatherer archaeological site is whether the bones ofthe predominant taxa found at the site were selectivelytransported to that site from elsewhere, butchered atthat site and then selectively transported elsewhere,utilised whole or any permutation of the preceding.Of further interest is whether there are instances ofselective butchery and transport reflected in comple-mentary assemblages. Therefore, body part representa-tion studies have the potential to inform about the roleof any given EBK locality. There are only a few studiesof skeletal part representation in EBK contexts, each withdiffering results and methods (Jonsson, 1988; Trolle-Lassen, 1992; Rowley-Conwy, 1998; Eriksson andMagnell, 2001).The minimum number of individual (MNI) counts
were established using the simplest method possiblefollowing Richter and Noe-Nygaard (2003). Thismethod involves side-by-side comparisons of bonesand bone fragments in order to best estimate theoriginal number of individuals (MNI) that would have
been present in the assemblages (Bunn, 1982). Theminimum number of element (MNE) values were cal-culated by ignoring symmetry and using side-by-sidecomparisons to establish the minimum number ofactual complete bone elements required to accountfor what is observed in the assemblage. These MNEvalues were then used to calculate per cent minimumanimal units (%MAU) by dividing by the elements’occurrence in the mammalian skeleton, and thendividing by the most common value therein. Thismethod is that used by Rowley-Conwy (1998) andonly slightly modified, particularly in terms of whichskeletal elements were included in the analysis. Datamanipulations are shown in Table 3.Carpals, the external cuneiform, and similar or
smaller bones such as sesamoids are omitted in thisanalysis. Their size, shape and compactness results inthese bones not being subject to the same taphonomicprocesses, and especially the human-mediated pro-cesses, affecting the bulk of the faunal material(Marean, 1991). As evidence of this, the carpals and ex-ternal cuneiform are almost always complete, althoughthe elements that are included here are almost alwaysincomplete (Table 4). Therefore, it is highly likely thattaphonomic processes, regardless of origin, affectedthese bones differently, than the bulk of the materialand in ways unrelated to their bone mineral densities.The %MAU values measure the relative frequencies
of different kinds of bones normalised for intersitecomparison. This is performed in order to assesswhether differential transport or destruction is evident
Table 1. Site details
Site Approximate dates Based on Culture Location Type of siteSeasons with
evidence of occupation
AsnæsHavnemark
4500-4100 BC C14 Ertebølle Coast Coastal settlement Summer/fall/winter
Fårevejle 4350-3950,3500-3200 BC
C14 Ertebølle-TRB Near coast Shell midden Spring/summer
Trustrup 4800-4000 BC Diagnosticartefacts
Ertebølle Inland Lakeshoresettlement
Unclear
Table 2. The three assemblages and their deer component
Whole assemblage Deer component
Total bones % Identified Total NISP % of NISP that are deer NISP MNI MNE NISP/MNE ratio
Asnæs Havnemark 12 202 18.14% 2214 72.94% 1615 22 667 2.421289Fårevejle 14 343 8.84% 1268 52.52% 666 13 256 2.601563Trustrup 20790 5.96% 1240 76.69% 951 23 348 2.732759
NISP, number of identified specimens; MNI, minimum number of individuals; MNE, minimum number of elements.
Ertebølle Deer Exploitation in Northwest Zealand, Denmark
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)
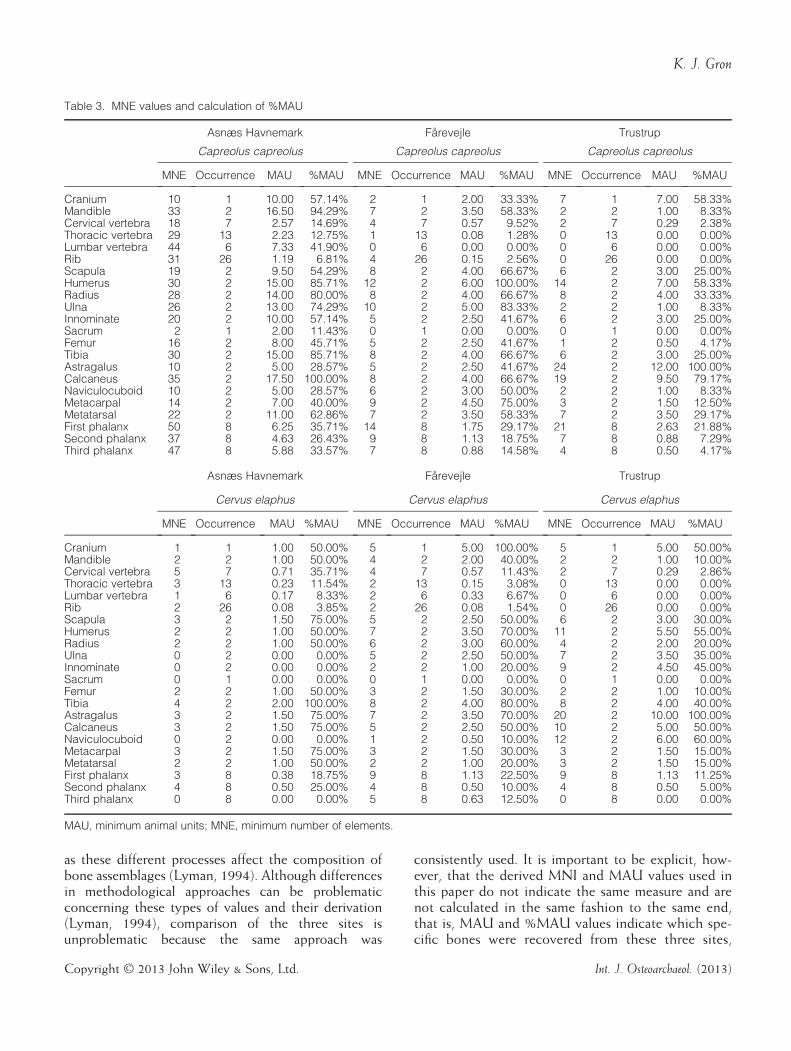
as these different processes affect the composition ofbone assemblages (Lyman, 1994). Although differencesin methodological approaches can be problematicconcerning these types of values and their derivation(Lyman, 1994), comparison of the three sites isunproblematic because the same approach was
consistently used. It is important to be explicit, how-ever, that the derived MNI and MAU values used inthis paper do not indicate the same measure and arenot calculated in the same fashion to the same end,that is, MAU and %MAU values indicate which spe-cific bones were recovered from these three sites,
Table 3. MNE values and calculation of %MAU
Asnæs Havnemark Fårevejle Trustrup
Capreolus capreolus Capreolus capreolus Capreolus capreolus
MNE Occurrence MAU %MAU MNE Occurrence MAU %MAU MNE Occurrence MAU %MAU
Cranium 10 1 10.00 57.14% 2 1 2.00 33.33% 7 1 7.00 58.33%Mandible 33 2 16.50 94.29% 7 2 3.50 58.33% 2 2 1.00 8.33%Cervical vertebra 18 7 2.57 14.69% 4 7 0.57 9.52% 2 7 0.29 2.38%Thoracic vertebra 29 13 2.23 12.75% 1 13 0.08 1.28% 0 13 0.00 0.00%Lumbar vertebra 44 6 7.33 41.90% 0 6 0.00 0.00% 0 6 0.00 0.00%Rib 31 26 1.19 6.81% 4 26 0.15 2.56% 0 26 0.00 0.00%Scapula 19 2 9.50 54.29% 8 2 4.00 66.67% 6 2 3.00 25.00%Humerus 30 2 15.00 85.71% 12 2 6.00 100.00% 14 2 7.00 58.33%Radius 28 2 14.00 80.00% 8 2 4.00 66.67% 8 2 4.00 33.33%Ulna 26 2 13.00 74.29% 10 2 5.00 83.33% 2 2 1.00 8.33%Innominate 20 2 10.00 57.14% 5 2 2.50 41.67% 6 2 3.00 25.00%Sacrum 2 1 2.00 11.43% 0 1 0.00 0.00% 0 1 0.00 0.00%Femur 16 2 8.00 45.71% 5 2 2.50 41.67% 1 2 0.50 4.17%Tibia 30 2 15.00 85.71% 8 2 4.00 66.67% 6 2 3.00 25.00%Astragalus 10 2 5.00 28.57% 5 2 2.50 41.67% 24 2 12.00 100.00%Calcaneus 35 2 17.50 100.00% 8 2 4.00 66.67% 19 2 9.50 79.17%Naviculocuboid 10 2 5.00 28.57% 6 2 3.00 50.00% 2 2 1.00 8.33%Metacarpal 14 2 7.00 40.00% 9 2 4.50 75.00% 3 2 1.50 12.50%Metatarsal 22 2 11.00 62.86% 7 2 3.50 58.33% 7 2 3.50 29.17%First phalanx 50 8 6.25 35.71% 14 8 1.75 29.17% 21 8 2.63 21.88%Second phalanx 37 8 4.63 26.43% 9 8 1.13 18.75% 7 8 0.88 7.29%Third phalanx 47 8 5.88 33.57% 7 8 0.88 14.58% 4 8 0.50 4.17%
Asnæs Havnemark Fårevejle Trustrup
Cervus elaphus Cervus elaphus Cervus elaphus
MNE Occurrence MAU %MAU MNE Occurrence MAU %MAU MNE Occurrence MAU %MAU
Cranium 1 1 1.00 50.00% 5 1 5.00 100.00% 5 1 5.00 50.00%Mandible 2 2 1.00 50.00% 4 2 2.00 40.00% 2 2 1.00 10.00%Cervical vertebra 5 7 0.71 35.71% 4 7 0.57 11.43% 2 7 0.29 2.86%Thoracic vertebra 3 13 0.23 11.54% 2 13 0.15 3.08% 0 13 0.00 0.00%Lumbar vertebra 1 6 0.17 8.33% 2 6 0.33 6.67% 0 6 0.00 0.00%Rib 2 26 0.08 3.85% 2 26 0.08 1.54% 0 26 0.00 0.00%Scapula 3 2 1.50 75.00% 5 2 2.50 50.00% 6 2 3.00 30.00%Humerus 2 2 1.00 50.00% 7 2 3.50 70.00% 11 2 5.50 55.00%Radius 2 2 1.00 50.00% 6 2 3.00 60.00% 4 2 2.00 20.00%Ulna 0 2 0.00 0.00% 5 2 2.50 50.00% 7 2 3.50 35.00%Innominate 0 2 0.00 0.00% 2 2 1.00 20.00% 9 2 4.50 45.00%Sacrum 0 1 0.00 0.00% 0 1 0.00 0.00% 0 1 0.00 0.00%Femur 2 2 1.00 50.00% 3 2 1.50 30.00% 2 2 1.00 10.00%Tibia 4 2 2.00 100.00% 8 2 4.00 80.00% 8 2 4.00 40.00%Astragalus 3 2 1.50 75.00% 7 2 3.50 70.00% 20 2 10.00 100.00%Calcaneus 3 2 1.50 75.00% 5 2 2.50 50.00% 10 2 5.00 50.00%Naviculocuboid 0 2 0.00 0.00% 1 2 0.50 10.00% 12 2 6.00 60.00%Metacarpal 3 2 1.50 75.00% 3 2 1.50 30.00% 3 2 1.50 15.00%Metatarsal 2 2 1.00 50.00% 2 2 1.00 20.00% 3 2 1.50 15.00%First phalanx 3 8 0.38 18.75% 9 8 1.13 22.50% 9 8 1.13 11.25%Second phalanx 4 8 0.50 25.00% 4 8 0.50 10.00% 4 8 0.50 5.00%Third phalanx 0 8 0.00 0.00% 5 8 0.63 12.50% 0 8 0.00 0.00%
MAU, minimum animal units; MNE, minimum number of elements.
K. J. Gron
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

whereas MNI values simply give an impression of thenumber of actual animals represented.As the primary concerns of this study rely on quan-
titative recording of the degree of fragmentation ofthree faunal assemblages, the following methods wereapplied. Fish bones and recognisable human remainswere removed from the assemblages and were notincluded in the analysis. In most cases, fish bones werecomplete elements, were not fragmented and were soabundant, in some cases they had to be selectivelysampled during excavation. Human remains wereexceedingly rare or absent. For those reasons, theseclasses of remains are excluded, as they would haveeither introduced bias (fish sampling) or were so rarethat the overall impression is unaffected. Therefore,with the exception of those categories, all bonesmeasured to quantify overall fragmentation were frommammals, birds or amphibians. Bird and amphibianbones were so rare or fragmentary in these assem-blages that meaningful sorting from the mammalsample was simply not possible.The degree of fragmentation is perhaps one of the
most important factors influencing quantitative record-ing of zooarchaeological data (in particular taxonomicidentifications). Several easily-calculated indices offragmentation were applied for comparison, includinga NISP to MNE ratio for the deer from all three sites,and the ratio of the identified to unidentified sampleof all bones recovered (Table 2) (Richardson, 1980;Gifford-Gonzalez, 1989). However, the only realmethod of quantifying it directly is to measure andassign all bones to size classes (Lyman and O’Brien,1987; Marshall and Pilgram, 1993; Outram, 2004).Outram (2004) recommends that only non-completebones be measured in order to quantify fragmentationto account for the variable meaning behind the pres-ence of whole bones versus fragmentary bones(Outram, 2001). However, I have measured all thebones, complete or fragmented, and this requires somejustification. Most importantly, very few of the boneswere assessed as complete, with such examples com-prising only 4.8%, 2.7% and 1.2% of the total numberof all identified and unidentified recovered bones from
Asnæs Havnemark, Fårevejle and Trustrup, respec-tively. Any effect of complete bones on the overallsize distribution is negligible. Second, the meaningof a whole bone versus a fragmentary bone in anassemblage will differ depending on the individualtaphonomic or behavioural process involved. In mea-suring the bones, my goal is to identify the effect ofthe sum total of taphonomic processes affecting thesethree assemblages so that the bone material can becompared among them without any presumption ofnecessarily being able to differentiate precisely eachfactor contributing to its current state. By includingcomplete and fragmentary bones, and given the lowoccurrence of complete bones, I am recording boththe degree of fragmentation and the overall characterof the assemblages. Ultimately, it is not the purpose ofthis paper to pinpoint specific taphonomic factors, butto use fragmentation data in a comparative sense tounderstand the relationship between fragmentationand body part representation.The measurements taken were very simple. Materials
were measured by hand to maximum dimension,assigned to classes of one centimetre increments (e.g.0-1 cm, 1-2 cm, 2-3 cm, etc.) and counted by physicallyplacing and counting each fragment and whole bone,regardless of its ability to be identified, on 1 cm graphpaper. At the low end of the smallest category, frag-ments were included if they were identifiable as boneand were moveable by hand without the use of tools.All of the assemblages from Fårevejle (N=14343), AsnæsHavnemark (N=12202) and Trustrup (N=20790) wererecorded in this fashion.
Results
Overall fragmentation data were obtained for the entireAsnaes Havnemark, Fårevejle and Trustrup assem-blages. The results are displayed in Figure 2. AsnæsHavnemark is the least fragmented of the assemblages,with over 20% of the material over 3 cm in overalllength. Fårevejle is a more fragmented assemblage,with just over 14% of the material over 3 cm in overall
Table 4. Sample completeness
Carpals and external cuneiform Bones in this study
Total Complete Total MNE % Complete Total complete Total MNE % Complete
Asnæs Havnemark 54 61 88.52% 122 606 20.20%Fårevejle 23 26 88.46% 39 230 16.96%Trustrup 72 90 80.00% 26 258 10.08%
MNE, minimum number of elements.
Ertebølle Deer Exploitation in Northwest Zealand, Denmark
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

length. Trustrup is also more fragmented, with lessthan 10% of the material greater than 3 cm in overalllength. Trustrup has a higher proportion of materialsthat are in the 2–3 cm size class than Fårevejle andfewer materials that are of 0–1 cm in maximum length.However, in terms of overall extent of fragmentationpertaining to identifiably and utility in determiningthe past economy of the sites, the larger fragmentsare more often useful. This means that althoughTrustrup has a higher proportion of larger, 2-3 cm frag-ments, it has far fewer fragments larger than that, and ittherefore may be considered the most fragmented ofthe group.The fragmentation data are also significant in light
of the proportion of the materials that are determin-able to species relative to the entire assemblage andthe relationship between the total NISP for deerversus the MNE for deer at each site. Size is obviouslynot the only factor determining identifiability offaunal remains, but it is a factor. For example, a toothfragment from a wild boar with dimensions well underone centimetre can confidently be identified as suchwhen coming from Mesolithic contexts, whereas amuch larger fragment of long bone is often requiredto identify the same taxon. However, as all three ofthese sites from northwest Zealand yielded assem-blages that are at least half deer remains based onNISP values, such a comparison may be valid. Takingall classes of fauna (mammals, birds and amphibians)identified to species or class of species together interms of NISP, 18.62% of the Asnæs Havnemarkassemblage was confidently identified to species,8.93% of the Fårevejle assemblage was similarlyidentified and only 5.98% of the Trustrup assemblagewas identifiable. Similarly, the deer NISP/MNE ratioincreases from Asnæs Havnemark, to Fårevejle, toTrustrup (Table 2). Accordingly, the Asnæs Havnemarkassemblage has far more utility for determining the pastactivities of EBK hunters than then more fragmentedones from Fårevejle and Trustrup. As shown earlier,the Fårevejle assemblage was nominally less fragmented
than Trustrup, and each of the pair are much morefragmented than the material from Asnæs Havnemark.The %MAU for roe deer and red deer were calcu-
lated for a number of elements in the skeleton andare shown graphically in Figure 3. Roe deer are the mostabundant species at each site under consideration interms of numbers of actual animals. There are 19 indi-viduals at Asnæs Havnemark (Ritchie et al., in press),8 individuals at Fårevejle and 14 individuals at Trustrup.The red deer sample at Asnæs Havnemark is threeindividuals (Ritchie et al., in press), with five at Fårevejleand nine at Trustrup. For both species, animals are bestrepresented by elements of the head, forelimb andhindlimb with the axial skeleton largely absent.
Discussion
The most relevant example that warrants comparisonwith my study is the inland hunting lakeside site atRingkloster on Jutland. In his study of the fauna,Rowley-Conwy (1998) quantified auroch (Bosprimigenius), red deer and wild boar (Sus scrofa) bonesin much the same way. Axial elements dominate theauroch and red deer components of the Ringklosterassemblage, with other elements relatively underrepre-sented (Rowley-Conwy, 1998). Wild boar at Ringklostershowed a somewhat different pattern. These data wereargued to indicate on-site butchery of animals with thetransport of limbs elsewhere, presumably to sites on thecoast (Rowley-Conwy, 1998). Rowley-Conwy thencompared the observed patterns to the ethnographically-studied and archaeologically-studied inland summercaribou hunting camp of Aasivissuit in west Greenlandand notes the general similarities with Ringkloster(Grønnow et al., 1983; Rowley-Conwy, 1998). The argu-ment is effective in demonstrating that carcasses were butch-ered at Ringkloster and parts were selectively transported.Therefore, studies of this type can be a useful tool forunderstanding the mechanisms of resource exploitation.In this study, the observed pattern of skeletal part repre-
sentation of roe deer is consistent with two possibleoptions. The first is that these three sites represent the coun-terpoint to the observed pattern at Ringkloster andAasivissuit (Grønnow et al., 1983; Rowley-Conwy, 1998),indicating that all three were localities to which roe deerheads, forelimbs and hindlimbs were brought. The skeletalpattern observed does appear to be an exact fit for what ismissing from the larger game specimens at hunting sites likeRingkloster and Aaivissuit, and, therefore, these three sitescould be locations to which the missing elements such asthose absent from the two comparative sites could have beentransported (Grønnow et al., 1983; Rowley-Conwy, 1998).
Figure 2. Overall fragmentation data for three Ertebølle culture sites.
K. J. Gron
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

Following, these three EBK sites in Zealand mostcertainly do not represent locations from which deerare exported elsewhere.Another option is that the observed pattern is the result
of density-mediated skeletal attrition (Lyman, 1993). Lessdense skeletal elements, particularly elements of the axialskeleton such as the neck and spine, are expected to bemore fragile than more dense elements (Lyman, 1993).By virtue of their relative fragility, these elements areexpected to be more affected by taphonomic processes.Therefore, more highly fragmented assemblages can beexpected to have fewer fragile bones compared with morerobust elements. In highly fragmented assemblages, evenwhen whole carcasses were originally deposited, thereshould by far be more head, forelimb and hindlimbelements than parts of the axial skeleton because ofrelative skeletal bone densities (Lyman, 1993).From these options, the more likely scenario at Asnæs
Havnemark, Fårevejle and Trustrup is the second, giventhe presence of some axial elements, significant correlation
between published bone density values and obtained%MAU values, the types of sites these locations repre-sent, available seasonality data and considerations oftransport. Percentage MAU values actually show someaxial elemental representation in the lesser-fragmentedassemblages. That is, the least fragmented site, AsnæsHavnemark, contains more axial elements (includingca. 42% MAU lumbar vertebrae) than its morefragmented counterparts. As fragmentation increasesin the Fårevejle and Trustrup assemblages, these valuesdecline sharply. This strongly suggests that fragmenta-tion is responsible for lower %MAU values in theseless dense elements. As a test of the relationshipbetween bone density and the observed body partrepresentation, calculated %MAU values were corre-lated with published bone density values for reindeer(Rangifer tarandus). Because of the extensive similaritiesin bone density patterns between different bovids,equids and cervids (Lam et al., 1999), it is appropriateto compare cervid density values with each other.
Figure 3. Per cent minimum animal unit Capreolus capreolus and Cervus elaphus by site.
Ertebølle Deer Exploitation in Northwest Zealand, Denmark
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)
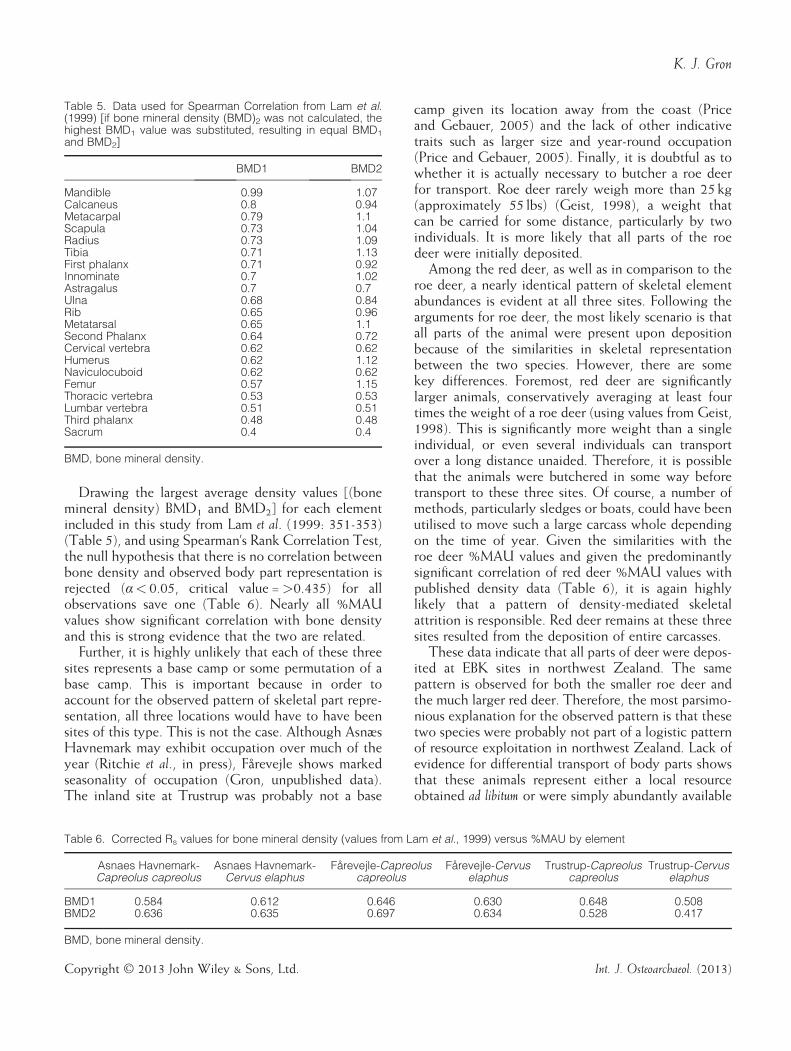
Drawing the largest average density values [(bonemineral density) BMD1 and BMD2] for each elementincluded in this study from Lam et al. (1999: 351-353)(Table 5), and using Spearman’s Rank Correlation Test,the null hypothesis that there is no correlation betweenbone density and observed body part representation isrejected (α< 0.05, critical value =>0.435) for allobservations save one (Table 6). Nearly all %MAUvalues show significant correlation with bone densityand this is strong evidence that the two are related.Further, it is highly unlikely that each of these three
sites represents a base camp or some permutation of abase camp. This is important because in order toaccount for the observed pattern of skeletal part repre-sentation, all three locations would have to have beensites of this type. This is not the case. Although AsnæsHavnemark may exhibit occupation over much of theyear (Ritchie et al., in press), Fårevejle shows markedseasonality of occupation (Gron, unpublished data).The inland site at Trustrup was probably not a base
camp given its location away from the coast (Priceand Gebauer, 2005) and the lack of other indicativetraits such as larger size and year-round occupation(Price and Gebauer, 2005). Finally, it is doubtful as towhether it is actually necessary to butcher a roe deerfor transport. Roe deer rarely weigh more than 25 kg(approximately 55 lbs) (Geist, 1998), a weight thatcan be carried for some distance, particularly by twoindividuals. It is more likely that all parts of the roedeer were initially deposited.Among the red deer, as well as in comparison to the
roe deer, a nearly identical pattern of skeletal elementabundances is evident at all three sites. Following thearguments for roe deer, the most likely scenario is thatall parts of the animal were present upon depositionbecause of the similarities in skeletal representationbetween the two species. However, there are somekey differences. Foremost, red deer are significantlylarger animals, conservatively averaging at least fourtimes the weight of a roe deer (using values from Geist,1998). This is significantly more weight than a singleindividual, or even several individuals can transportover a long distance unaided. Therefore, it is possiblethat the animals were butchered in some way beforetransport to these three sites. Of course, a number ofmethods, particularly sledges or boats, could have beenutilised to move such a large carcass whole dependingon the time of year. Given the similarities with theroe deer %MAU values and given the predominantlysignificant correlation of red deer %MAU values withpublished density data (Table 6), it is again highlylikely that a pattern of density-mediated skeletalattrition is responsible. Red deer remains at these threesites resulted from the deposition of entire carcasses.These data indicate that all parts of deer were depos-
ited at EBK sites in northwest Zealand. The samepattern is observed for both the smaller roe deer andthe much larger red deer. Therefore, the most parsimo-nious explanation for the observed pattern is that thesetwo species were probably not part of a logistic patternof resource exploitation in northwest Zealand. Lack ofevidence for differential transport of body parts showsthat these animals represent either a local resourceobtained ad libitum or were simply abundantly available
Table 5. Data used for Spearman Correlation from Lam et al.(1999) [if bone mineral density (BMD)2 was not calculated, thehighest BMD1 value was substituted, resulting in equal BMD1and BMD2]
BMD1 BMD2
Mandible 0.99 1.07Calcaneus 0.8 0.94Metacarpal 0.79 1.1Scapula 0.73 1.04Radius 0.73 1.09Tibia 0.71 1.13First phalanx 0.71 0.92Innominate 0.7 1.02Astragalus 0.7 0.7Ulna 0.68 0.84Rib 0.65 0.96Metatarsal 0.65 1.1Second Phalanx 0.64 0.72Cervical vertebra 0.62 0.62Humerus 0.62 1.12Naviculocuboid 0.62 0.62Femur 0.57 1.15Thoracic vertebra 0.53 0.53Lumbar vertebra 0.51 0.51Third phalanx 0.48 0.48Sacrum 0.4 0.4
BMD, bone mineral density.
Table 6. Corrected Rs values for bone mineral density (values from Lam et al., 1999) versus %MAU by element
Asnaes Havnemark-Capreolus capreolus
Asnaes Havnemark-Cervus elaphus
Fårevejle-Capreoluscapreolus
Fårevejle-Cervuselaphus
Trustrup-Capreoluscapreolus
Trustrup-Cervuselaphus
BMD1 0.584 0.612 0.646 0.630 0.648 0.508BMD2 0.636 0.635 0.697 0.634 0.528 0.417
BMD, bone mineral density.
K. J. Gron
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

locally in most areas of Zealand and did not require anelaborate exploitation strategy.This contrast with the site of Ringkloster and ethno-
graphic examples of extraction sites where at least somelarge game are being processed for transport hints at a de-gree of variability in resource use within the EBK. AtRingkloster, the meat from large game may have beenmoved to the coast (Rowley-Conwy, 1998) whereas innorthwest Zealand, there is less evidence for such apractice. Whether or not this relates to regional differ-ences, with relatively more sedentism in one region orthe other, or simply the fact that there was so much foodavailable on Zealand that there was no need for extractionis open to interpretation. The point is that not all re-sources were part of a logistical system of resource move-ment. Finally, results presented here are consistent withresults reported at Bökeberg III in Scania and TybrindVig on Fyn for animal butchery and body part representa-tion, possibly indicating similarity in butchery practicesbetween Zealand, and these adjacent regions (Trolle-Lassen, 1992; Eriksson and Magnell, 2001).These data are of interest for both methodological
reasons as well as for their importance for reconstructingthe past. In this case, the ability to discern the reasonsfor apparent differential body part representation restedsolely on the fact that overall assemblage fragmentationdata was available. Alone, the only conclusion followingfrom the %MAU data would be that selective body parttransport from logistical extraction sites is in evidence atall three localities. However, when combining both linesof evidence the interpretation changes completely.Therefore, in the absence of quantitative data describingthe degree of fragmentation, comparisons of differentialbody part representation and overall relative abundancedata between sites may be quite biased. This is not anew observation (Marshall and Pilgram, 1993), but ishighly germane to studies of the EBK.
Conclusions
Data have been presented, which show that there is noselective transport of deer to or from the three EBKsites of Asnæs Havnemark, Fårevejle and Trustrup innorthwest Zealand, a region of high environmentalvariability during the Atlantic Period (Paludan-Müller,1978). The observed pattern reflects bone density-mediated skeletal attrition and not differential transportof specific body parts. Therefore, movement of hunteddeer from site to site is not indicated. Without detailedconsiderations of patterns of assemblage fragmentation,such an assessment is not possible. Therefore, it is
advisable to include quantification of fragmentation inzooarchaeological studies in the region.These data contrast with observed butchery pat-
terns elsewhere in the EBK area, and may indicatecultural differences between different regions duringthe late Mesolithic period. The data also suggest thatdeer may not have been part of a logistic movement ofresources, but instead were processed and consumedclose to where they were killed, possibly indicatingvariability in the degree of sedentism among regions.Ultimately, this paper serves to illustrate that not onlyis there variability among the EBK in terms of theresources utilised (Ritchie et al., in press), but also inhow resources are processed, an important finding inlight of other observed cultural differences betweenregions of this affluent hunter-gatherer-fisher culture(Petersen, 1984; Ritchie, 2010).
Acknowledgements
This research was funded by the Danish-AmericanFulbright Commission and the National Science Founda-tion (DDIG #1135155). The Department of Geographyand Geology at Copenhagen University as well asthe Zoological Museum at the University of Wisconsin-Madison provided laboratory facilities. Excavationsyielding this faunal material were funded by the NationalScience Foundation and the Carlsberg Foundation.Thanks in particular are owed to T. Douglas Price whoput the material at my disposal. In addition, Dr Price,Nanna Noe-Nygaard and Travis Pickering providedhelpful advice. Finally, I would like to thank Ken Ritchie,Harry Robson and an anonymous reviewer who carefullyread and commented on earlier versions of this paper.
References
Aaris-Sørensen K. 1980. Depauperation of the mammalianfauna of the island of Zealand during the Atlantic Pe-riod. Videnskabelige Meddelelser Dansk Naturhistorisk Forening142: 131–138.
Aaris-Sørensen K. 2009. Diversity and dynamics of the mam-malian fauna in Denmark Throughout the last glacial-interglacial cycle, 115-0 kyr BP. Fossils and Strata 57: 1–59.
Andersen SH. 1985. Tybrind Vig: A preliminary report on asubmerged Ertebølle settlement on the west coast of Fyn.Journal of Danish Archaeology 4: 52–69.
Andersen SH. 1987. Mesolithic dug-outs and paddles fromTybrind Vig, Denmark. Acta Archaeologica 57: 97–106.
Andersen SH. 2011. Ertebølle canoes and paddles from thesubmerged habitation site of Tybrind Vig, Denmark. In
Ertebølle Deer Exploitation in Northwest Zealand, Denmark
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

Submerged Prehistory, J Benjamin, C Bonsall, C Pickard,A Fischer (eds). Oxbow Books: Oxford; 1–14.
Binford L. 1980. Willow smoke and dogs’ tails: hunter-gatherer settlement systems and archaeological site forma-tion. American Antiquity 45(1): 4–20.
Bunn H. 1982. Meat eating and human evolution: studies onthe diet and subsistence patterns of plio-pleistocenehominids in east Africa. PhD Dissertation, University ofCalifornia, Berkeley.
Enghoff IB. 2011. Regionality and Biotope Exploitation inDanish Ertebølle and Adjoining Periods. The RoyalDanish Academy of Sciences and Letters: Copenhagen.
Eriksson M, Magnell O. 2001. Jakt och Slakt. In DansarmaFrån Bökeberg: Om Jakt, Ritualer och Inlandsbos ttning vidJ garstenålderns Slut, P Karsten (ed.). Riksantikvarie mbetet:Lund; 49–78.
Fischer A, Olsen J, Richards M, Heinemeier J, SveinbjörnsdóttirÁ, Benike P. 2007. Coast-inland mobility and diet in theDanish Mesolithic and Neolithic: evidence from stableisotope values of humans and dogs. Journal of ArchaeologicalScience 34: 2125–2150.
Geist V. 1998. Deer of the World: Their Evolution, Behav-iour, and Ecology. Stackpole Books: Mechanicsburg.
Gifford-Gonzalez D. 1989. Ethnographic Analogues forInterpreting Modified Bones: Some Cases from EastAfrica. In Bone Modification, R Bonnichesen, MH Sorg(eds). Center for the Study of the First Americans: Orono;179–246.
Gotfredsen AB. 1998. En rekonstruktion af PalæomiljøetOmkring tre Senmesolitske Bopladser i store åmose,vestsjælland-Baseret på Pattedyr- og Fugleknogler. GeologiskTidsskrift 1998(2): 92–103.
Gron K. 2013. The Ertebølle faunal economy and the transi-tion to agriculture in southern Scandinavia. PhD Disserta-tion, University of Wisconsin-Madison.
Grønnow B, Meldgaard M, Nielsen JB. 1983. Aasivissuit-Thegreat summer camp: Archaeological, ethnographical andzoo-archaeological studies of a caribou-hunting site inWest Greenland. Meddelelser om Grønland. Man and Society5: 1–96.
Jonsson L. 1988. The vertebrate faunal remains from the lateAtlantic settlement Skateholm in Scania, south Sweden. InThe Skateholm Project. I. Man and Environment, L Larsson(ed.). Almqvist and Wiksell International: Lund; 56–88.
Lam YM, Chen X, Pearson OM. 1999. Intertaxonomicvariability in patterns of bone density and the differen-tial representation of Bovid, Cervid, and Equid ele-ments in the archaeological record. American Antiquity64(2):343–362.
Lyman RL. 1993. Density-mediated attrition of bone assem-blages: new insights. In From Bones to Behavior:Ethnoarchaeological and Experimental Contributions tothe Interpretation of Faunal Remains, J Hudson (ed.).Center for Archaeological Investigations: Carbondale;324–341.
Lyman RL. 1994. Quantitative units and terminology inzooarchaeology. American Antiquity 59(1): 36–71.
Lyman RL, O’Brien MJ. 1987. Plow-zone zooarchaeology:fragmentation and identifiability. Journal of Field Archaeology14(4): 493–498.
Magnussen BI. 2007. En geologisk og zooarkæologisk analyse ofkystbopladsen; lollikhuse på overgangenmellemmesolitikumog neolitikum. MA thesis, Copenhagen University.
Marean CW. 1991. Measuring post-depositional destructionof bone in archaeological assemblages. Journal of Archaeo-logical Science 18: 677–694.
Marshall F, Pilgram T. 1993. NISP vs. MNI in quantifi-cation of body-part representation. American Antiquity58(2): 261–9.
Møhl U. 1971. Oversigt over dyreknoglerne fra ølby lyng.En østsjællandsk kystboplads med ertebøllekultur. Aarbøgerfor nordisk Oldkyndighed og Historie 1970: 43–77.
Noe-Nygaard N. 1988. δ13C-Values of dog bones reveal thenature of changes in man’s food resources at theMesolithic-Neolithic transition, Denmark. Chemical Geol-ogy (Isotope Geoscience Section) 73: 87–96.
Noe-Nygaard N 1995. Ecological, Sedimentary, andGeochemical Evolution of the Late-glacial to PostglacialÅmose Lacustrine Basin, Denmark. Scandinavian UniversityPress: Oslo.
Outram AK. 2001. A new approach to identifying bone mar-row and grease exploitation: why the ‘indeterminate’ frag-ments should not be ignored. Journal of Archaeological Science28: 401–410.
Outram AK. 2004. Applied models and indices vs. high-resolution, observed data: detailed fracture and frag-mentation analyses for the investigation of skeletalpart abundance patterns. Journal of Taphonomy 2(3):167–184.
Paludan-Müller K. 1978. High Atlantic food gathering innorthwestern Zealand: ecological conditions and spatialrepresentation. New Directions in Scandinavian Archaeology1: 120–57
Pedersen L. 1997. They put Fences in the sea. In The DanishStorebælt Since the Ice Age-Man, Sea and Forest, L Pe-dersen, A Fischer, B Aaby (eds). A/S Storebælt Fixed Link:Copenhagen; 124–143.
Petersen PV. 1984. Chronological and regional variation inthe late Mesolithic of eastern Denmark. Journal of DanishArchaeology 3:7–18.
Price TD, Gebauer AB. 2005. Smakkerup Huse: A LateMesolithic Coastal Site in Northwest Zealand, Denmark.Aarhus University Press: Aarhus.
Price TD, Noe-Nygaard N. 2009. Early Domestic Cattle inSouthern Scandinavia and the Spread of the Neolithic inEurope. In From Bann Flakes to Bushmills: Papers in Hon-our of Professor Peter Woodman, N Finlay, S McCartan,N Milner, C Wickham-Jones (eds). Oxbow Books:Oxford; 198–210.
Richardson PRK. 1980. Carnivore damage to antelope bonesand its archaeological implications. Paleontolgica Africana23: 109–125.
Richter J, Noe-Nygaard N. 2003. A late Mesolithic huntingstation at Agernæs, Fyn, Denmark: Differentiation and
K. J. Gron
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)

specialization in the late Ertebølle-culture, heralding theintroduction of agriculture? Acta Archaeologica 74: 1–64.
Ritchie K. 2010. The Ertebølle Fisheries of Denmark, 5400-4000 B.C. PhD Dissertation, University of Wisconsin-Madison.
Ritchie K, Gron K, Price TD. In press. Flexibility and diversityin subsistence during the late Mesolithic: faunal evidencefrom Asnæs Havnemark. Danish Journal of Archaeology.
Rowley-Conwy P. 1983. Sedentary Hunters: The Ertebølle Ex-ample. In Hunter-Gatherer Economy in Prehistory, G Bailey(ed.). Cambridge University Press: Cambridge; 111–126.
Rowley-Conwy P. 1998. Meat, furs, and skins: Mesolithicanimal bones from Ringkloster, a seasonal huntingcamp in Jutland. Journal of Danish Archaeology 12: 87–98.
Rowley-Conwy P. 1999. Economic prehistory in southernScandinavia. Proceedings of the British Academy 99: 125–159.
Tauber H. 1981. 13C Evidence for dietary habits of prehis-toric man in Denmark. Nature 292: 332–333.
Trolle-Lassen T. 1992. Butchering of red deer (Cervuselaphus L.)—a case study from the late Mesolithic set-tlement of tybrind Vig, Denmark. Journal of DanishArchaeology 9: 7–37.
Ertebølle Deer Exploitation in Northwest Zealand, Denmark
Copyright © 2013 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2013)