blood functional assay for rapid clinical interpretation
TRANSCRIPT

796 Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Original research
Blood functional assay for rapid clinical interpretation of germline TP53 variantsSabine Raad,1 Marion Rolain,1 Sophie Coutant,1 Céline Derambure,1 Raphael Lanos,1 Françoise Charbonnier,1 Jacqueline Bou,1 Emilie Bouvignies,1 Gwendoline Lienard,1 Stéphanie Vasseur,1 Michael Farrell,2 Olivier Ingster,3 Stéphanie Baert Desurmont,1 Edwige Kasper,1 Gaëlle Bougeard,1 Thierry Frébourg,1 Isabelle Tournier 1
Cancer genetics
To cite: Raad S, Rolain M, Coutant S, et al. J Med Genet 2021;58:796–805.
► Additional material is published online only. To view, please visit the journal online (http:// dx. doi. org/ 10. 1136/ jmedgenet- 2020- 107059).
1Normandie University, UNIROUEN, Inserm U1245 and Rouen University Hospital, Department of Genetics, F76000, Normandy Centre for Genomic and Personalized Medicine, University of Rouen Faculty of Medicine and Pharmacy, Rouen, France2Cancer Genetics Service, Mater Private Hospital, Dublin, Leinster, Ireland3Department of Genetics, University Hospital Centre Angers, Angers, Pays de la Loire, France
Correspondence toProfessor Thierry Frébourg, Normandie Univ, UNIROUEN, Inserm U1245 and Rouen University Hospital, Department of Genetics, F76000, Normandy Centre for Genomic and Personalized Medicine, University of Rouen Faculty of Medicine and Pharmacy, Rouen 76183, France; thierry. frebourg@ chu- rouen. fr
SR and MR contributed equally.
Received 1 April 2020Revised 5 August 2020Accepted 1 September 2020Published Online First 13 October 2020
© Author(s) (or their employer(s)) 2021. Re- use permitted under CC BY- NC. No commercial re- use. See rights and permissions. Published by BMJ.
ABSTRACTBackground The interpretation of germline TP53 variants is critical to ensure appropriate medical management of patients with cancer and follow- up of variant carriers. This interpretation remains complex and is becoming a growing challenge considering the exponential increase in TP53 tests. We developed a functional assay directly performed on patients’ blood.Methods Peripheral blood mononuclear cells were cultured, activated, exposed to doxorubicin and the p53- mediated transcriptional response was quantified using reverse transcription–multiplex ligation probe amplification and RT- QMPSF assays, including 10 p53 targets selected from transcriptome analysis, and two amplicons to measure p53 mRNA levels. We applied this blood functional assay to 77 patients addressed for TP53 analysis.Results In 51 wild- type TP53 individuals, the mean p53 functionality score was 12.7 (range 7.5–22.8). Among eight individuals harbouring likely pathogenic or pathogenic variants, the scores were reduced (mean 4.8, range 3.1–7.1), and p53 mRNA levels were reduced in patients harbouring truncating variants. We tested 14 rare unclassified variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Phe134Leu), p.(Arg158Cys), p.(Pro191Arg), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr), p.(Gly108_Phe109delinsVal), p.(Asn131del), p.(Leu265del), c.-117G>T) and 12 yielded functionally abnormal scores. Remarkably, the assay revealed that the c.*1175A>C polymorphic variant within TP53 poly- adenylation site can impact p53 function with the same magnitude as a null variant, when present on both alleles, and may act as a modifying factor in pathogenic variant carriers.Conclusion This blood p53 assay should therefore be a useful tool for the rapid clinical classification of germline TP53 variants and detection of non- coding functional variants.
INTRODUCTIONIdentification of a germline pathogenic TP53 (MIM: *191170) variant in a patient with cancer has drastic medical impacts.1 Indeed, in TP53 variant carriers, chemotherapy and radiotherapy have been shown to contribute to the development of subsequent primary cancers, the incidence of which is remarkably high (above 40%).1–4 There-fore, in these patients, surgical treatment should
be prioritised and radiotherapy and chemotherapy avoided, if possible, or at least carefully discussed in terms of benefit:risk ratio between risk of recur-rence and risk of inducing second primary tumours. Furthermore, TP53 variant carriers should have specific surveillance protocols, including annual whole- body MRI,5 6 whose efficiency for early tumour detection has recently been shown by numerous studies.5–14
Interpretation of germline TP53 variants, which are mainly missense variants, remains particularly complex. Whereas germline variants of TP53 were initially detected in Li- Fraumeni syndrome (LFS, MIM#151623),15–17 our perception of cancers related to germline alterations of TP53 has dras-tically evolved over time.1 2 18 19 The presence of a disease- causing germline variant should be considered in patients fulfilling Chompret criteria, which were sequentially updated and extended.1 The question of germline TP53 variant interpre-tation is becoming a growing concern in the field because the TP53 gene is currently included in many cancer gene panels, and the number of TP53 tests performed in patients not fulfilling the criteria mentioned earlier has increased exponentially. 20 21
Classification of TP53 variants, in agreement with the American College of Medical Genetics and Genomics/Association for Molecular Pathology guidelines, is based on several items, including frequency of the variant in the general popula-tion (gnomAD; https:// gnomad. broadinstitute. org/), segregation data, bioinformatics predictions and functional assays developed in yeast or human cancer cell lines.22 One of the first assays commonly used for TP53 missense variant interpretation was developed in yeast and is based on the expression of TP53 cDNA in strains containing reporter plasmids with different p53 binding sites.23 In this assay, p53 variants are classified as functional, not functional or partially functional if the transcriptional activity is conserved for some but not all yeast reporter plasmids (http:// p53. iarc. fr/). More recently, two teams have developed in human cancer cell lines high throughput p53 functional assays.24 25 Kotler et al24 generated a synthetic library of TP53 vari-ants located within the p53 DNA- binding domain and quantified the antiproliferative activity of these variants in the p53- null H1299 cancer cell line. In this assay, TP53 variants are categorised as ‘wild- type TP53- like variant’ (functional) or ‘disrupting’
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
on F
ebruary 4, 2022 by guest. Protected by copyright.
http://jmg.bm
j.com/
J Med G
enet: first published as 10.1136/jmedgenet-2020-107059 on 13 O
ctober 2020. Dow
nloaded from
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
on F
ebruary 4, 2022 by guest. Protected by copyright.
http://jmg.bm
j.com/
J Med G
enet: first published as 10.1136/jmedgenet-2020-107059 on 13 O
ctober 2020. Dow
nloaded from
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
on F
ebruary 4, 2022 by guest. Protected by copyright.
http://jmg.bm
j.com/
J Med G
enet: first published as 10.1136/jmedgenet-2020-107059 on 13 O
ctober 2020. Dow
nloaded from
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
on F
ebruary 4, 2022 by guest. Protected by copyright.
http://jmg.bm
j.com/
J Med G
enet: first published as 10.1136/jmedgenet-2020-107059 on 13 O
ctober 2020. Dow
nloaded from
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
on F
ebruary 4, 2022 by guest. Protected by copyright.
http://jmg.bm
j.com/
J Med G
enet: first published as 10.1136/jmedgenet-2020-107059 on 13 O
ctober 2020. Dow
nloaded from

797Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
(non- functional). In another assay, Giacomelli et al25 generated by saturation mutagenesis a TP53 library and tested the ability of the variants (1) to restore the survival of the p53- null A459 cell line exposed to high doses of DNA damaging agents, in order to detect loss of function (LOF) variants and (2) to induce in p53- wild- type A459 cells resistance to Nutlin- 3, in order to detect variants with dominant negative effect (DNE).
We previously developed, in Epstein- Barr virus- immortalised lymphocytes, a p53 functional assay exploring the transcrip-tional activity of the protein underlying its tumour suppressor activity.26 This assay is based on the exposure of cells to DNA damaging agents followed by the measurement of the p53 tran-scriptional response.27 28 With this assay, we showed that patho-genic TP53 variant carriers exhibit a constitutive defect in the transcriptional response to DNA damage, establishing a biolog-ical endophenotype associated with germline pathogenic vari-ants.27 28 Compared with the other assays, its main advantage is to evaluate the impact of heterozygous variants in the genetic context of the patients. Its main disadvantage is that it requires EBV immortalisation, which is time- consuming and, therefore, not suited for a rapid classification and interpretation of TP53 variants in medical practice.
Therefore, despite the different tools indicated previously and before the completion in the future of curated international databases, interpretation of germline TP53 variants remains chal-lenging in clinical practice. This prompted us to develop a p53 functional assay derived from the previous one but performed on fresh blood samples and suitable for rapid interpretation and medical management of patients. We show here that this assay can accurately detect pathogenic variants and can be used to reallocate unclassified variants by integrating the results to the classification strategy.22 Furthermore, this assay revealed that a TP53 polymorphism (rs78378222), present in 1.7% of the Euro-pean population, compromises p53 functional activity with the same magnitude as a heterozygous null variant, when carried on both alleles.
METHODSCell culture and treatmentEBV- immortalised cell lines were maintained in RPMI 1640 medium (GIBCO; Life Technologies, Carlsbad, California, USA) with 10% fetal calf serum (Invitrogen, Life Technologies) and 1% L- glutamine (Invitrogen) at 37°C with 5% CO2. Cells were seeded in duplicate in 12- well plates (Corning, New York, USA) at a density of 106 cells/well. Cells were treated or not with 200 ng/mL (0.3 µM final concentration) of doxorubicin (Sigma Aldrich, St. Louis, Missouri, USA) for 8 hours. Cells were washed with 1× PBS and harvested for RNA extraction.
Peripheral blood mononuclear cell (PBMC) isolation and cultureBlood samples were collected in EDTA tubes and kept for 2 days at room temperature before PBMC isolation on a lympho-cyte separation medium (Eurobio, Evry, France). From 2.5 to 10.0 mL of blood per patient was used for PBMC isolation. Cell number and cell viability were assessed on a NanoEnTek Adam automatic cell counter with the AccuChip Kit (ScienceTEC, Villebon- sur- Yvette, France). One million cells were seeded per well in a 24- well plate and were let to grow for 48 hours in a lymphocyte activating medium (Chromosome Medium P, Ampli-Tech, Compiègne, France). At least two wells were seeded per patient (treated and untreated) and duplicates or triplicates were performed whenever possible. Cells were treated with 800 ng/
mL of doxorubicin for 8 hours, washed with 1× PBS, harvested and RNA extraction was performed using the NucleoSpin RNA XS kit (Macherey Nagel, Düren, Germany) according to the manufacturer’s instructions and quantified using a UV- VIS ND- 1000 spectrophotometer (Biocompare, Nanodrop Technol-ogies, USA).
RNA-SeqFour control EBV cell lines wild- type for TP53 and four hetero-zygous TP53- mutant cell lines, corresponding to three canon-ical dominant negative missense variants (p.(Arg175His), p.(Arg248Trp) and p.(Arg273His)) and one complete dele-tion of the TP53 locus, were treated or not with doxorubicin. RNA was extracted using the Nucleospin RNAII kit (Mach-erey Nagel). Libraries were prepared using the NEBNext Ultra Directional RNA Library Kit for Illumina (NEB, Ipswich, USA) and NGS sequencing of the libraries was performed on an Illu-mina NextSeq500 (Illumina, San Diego, USA) using 2*75 bp sequencing to generate 50M read pairs on average per sample. Experiments were performed in triplicates. Bioinformatic analysis was carried out using an in- house automated pipeline AURIGA that uses the STAR V.2.5.3a tool for alignment, Featu-reCounts tool V.1.5.2 for read counting and DESeq2 V.1.18.1 for statistical analysis.
Selection of biomarkers indicative of p53-transcriptional activityNew biomarkers were selected among the transcripts strongly up- egulated by doxorubicin in control cells but not in the cells harbouring heterozygous TP53 alterations: CEP170B (NM_015005), PODXL (MIM*602632, NM_001018111), RRAD (MIM*179503, NM_004165), GLS2 (MIM*606365, NM_013267), CABYR (MIM*612135, NM_012189), TP53I3 (MIM*605171, NM_004881), EPS8L2 (MIM*614988, NM_022772), SULF2 (MIM*610013, NM_001161841), SESN1 (MIM*606103, NM_014454) and FHL2 (MIM*602633, NM_201555). Three control transcripts with a steady expres-sion across all conditions and genotypes and expressed at the same level as the selected targets were also selected: TBP (MIM*600075, NM_003194), RIC8B (MIM*609147, NM_001330145) and MPP5 (MIM*606958, NM_022474.3). An internal control of treatment efficacy was included: PLK1 (MIM*602098, NM_005030.5), whose transcript is downreg-ulated by doxorubicin treatment both in wild- type and mutant cells.
Reverse transcription–quantitative multiplex PCR of short fluorescent fragment (RT-QMPSF)Reverse transcription (RT) was performed on 100 ng of total RNA using the Verso cDNA Synthesis Kit (Thermo Scientific, Waltham, USA). RT- QMPSF was performed on 1.5 µL of RT using Diamond Taq DNA polymerase (Kaneka Eurogentec, Seraing, Belgium), 6% Dymethyl sulfoxide and 26 PCR cycles (94°C: 30 s/58°C: 1 min/72°C: 30 s). Primer sequences are listed in online supplemental table 1. Amplicons were analysed on an ABI Prism 3500 Genetic Analyzer (Applied Biosystems, Foster City, California, USA) using GeneScan 3.7 software.
Reverse transcription–multiplex ligation probe amplification (RT-MLPA)RT- MLPA probes were pooled at a concentration of 1 fmol/µL each in 10 mM Tris/1 mM EDTA. Probe sequences are given in online supplemental table 1. RT (6.5 µL), probe mixture (1.5 µL)
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

798 Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
and SALSA- MLPA buffer (1.5 µL, MRC- Holland, Amsterdam, The Netherlands) were mixed before denaturation (95°C, 2 min) and hybridisation (60°C, 1 hour). Ligation was performed at 54°C for 15 min, adding 32 µL of ligation mixture, and heated 5 min at 98°C. Then, 2.5 µL of the ligation was added to 7.5 µL of a Q5Hot Start High- Fidelity 2X Master Mix (NEB) supplemented with universal fluorescent PCR primers. PCR was performed using 35 cycles (94°C: 30 s/58°C: 30 s/72°C: 30 s). Amplicons were analysed on an ABI Prism 3500 Genetic Analyzer using GeneScan V.3.7 software.
Calculation of p53 functionality score and p53 mRNA ratioThe RT- MLPA or RT- QMPSF profiles of doxorubicin- treated and untreated cells were superimposed after adjusting the control amplicons to the same height. In the treated condition, the peak height of each of the 10 p53 target genes was measured and divided by the sum of the heights of the three control genes. This value was then divided by the same ratio calculated in the untreated condition. In the assay, the mean of the 10 values defines the p53 functionality score. The final p53 function-ality score is the mean of the scores obtained in RT- MLPA and RT- QMPSF assays. The p53 mRNA levels were expressed as a ratio of the normal values obtained for 3 TP53 wild- type control individuals. The efficacy of the genotoxic treatment was assessed by calculating a PLK1 (MIM*602098) ratio (treated/untreated) normalised with the three controls, which should be less than 0.5.
RESULTSDevelopment of a rapid p53 functional assay performed on blood The rationale of the assay is that p53 acts as a powerful tran-scriptional inductor when DNA damage occurs and that the common deleterious impact of pathogenic variants is the alter-ation of this transcriptional activity.26 To develop a functional assay directly performed on patient’s fresh blood, we first opti-mised the quantitative assay that we had previously developed in EBV- immortalised cell lines.27 28 To this aim, we performed a new comparative transcriptomic analysis using RNA- Seq, including non- polyadenylated RNAs. Four control EBV cell lines wild type for TP53 and four patients with LFS EBV cell lines were compared in the context of genotoxic stress induced by doxorubicin treatment. We selected 10 biomarkers corre-sponding to p53 targets involved in different biological pathways controlled by p53, such as cell adhesion and migration, cellular response to stress, apoptosis, cytoskeleton organisation, glycol-ysis or regulation of other metabolic pathways. To normalise the results, we selected three transcripts with a steady expression across all conditions and genotypes. All these biomarkers were then included in two quantitative assays based on RT- MLPA and RT- QMPSF. To detect in the same assay the potential effect of variants on the TP53 transcript levels, we added different ampl-icons or probes corresponding to TP53 cDNA. As a defect in treatment efficacy would result in a low functionality score leading to the misinterpretation of a wild- type genotype as a mutant one, we also integrated in the assays an internal control of treatment efficacy. After exposure to doxorubicin, cells were harvested and the RT- MLPA and RT- QMPSF assays were performed in parallel for each sample to increase the robustness of the assay. An arbitrary functionality score was calculated from the induction score of the 10 p53 targets; the p53 RNA levels were evaluated and expressed as a percentage of the mean levels
obtained for three wild- type TP53 individuals. This new quanti-tative assay, based on both RT- QMPSF and RT- MLPA, was first validated on 31 lymphoblastoid cell lines derived from patients with LFS harbouring different germline heterozygous TP53 vari-ants (online supplemental table 2).
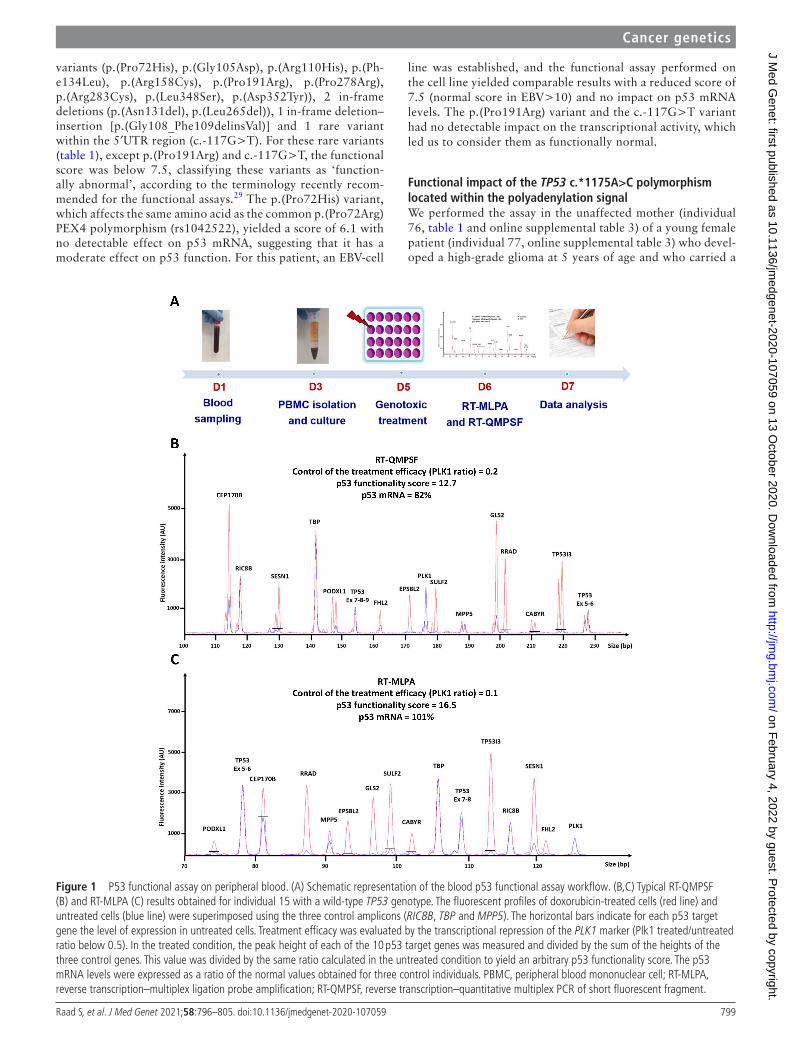
We then set up the conditions allowing the assay to be performed directly on the patients’ peripheral blood. Blood was collected in conventional EDTA tubes and kept at room tempera-ture for 2 days to mimic sample shipping delays. PBMCs were isolated and cultured for 48 hours in a lymphocyte activating medium. Under these conditions, a strong p53 transcriptional response could be monitored in wild- type individuals (figure 1), indicating that testing p53 function directly on patients’ blood cells was feasible.
p53 functional analysis of patient’s blood cells with different TP53 genotypesWe then applied the p53 functional assay on blood samples sent to our laboratory for TP53 molecular analysis (NGS screening of the 11 exons complemented by QMPSF). Molecular and functional analyses were performed in parallel, in double blind conditions. We analysed a total of 82 blood samples derived from 77 individuals (online supplemental table 3). These 77 individuals corresponded either to new index cases suspected to harbour a pathogenic TP53 variant or to relatives of index cases harbouring TP53 variants. This sample reflects the real- life recruitment of our diagnostic laboratory as it includes unaffected individuals as well as individuals affected by cancer who may have undergone different chemotherapy treatments. Molecular analyses revealed that 51 individuals had no detect-able germline TP53 variant. For these 51 individuals, the mean p53 functionality score measured was 12.7 (13.6 for the RT- QMPSF assay and 11.9 for the RT- MLPA assay) with a range of 7.5–22.8 (online supplemental table 3 and figure 2). The mean observed p53 mRNA levels were 93% with a range of 74%–125% (online supplemental table 3). In eight tested individuals, molecular analysis revealed seven distinct TP53 variants which could be considered as likely pathogenic or pathogenic based on their ClinVar classification or their trun-cating nature (table 1). All the variants tested were confirmed to be germline heterozygous variants. For these eight patients, the assay yielded a reduced score compared with the wild- type individuals (mean 4.8, range 3.1–7.1; table 1 and figure 2). In the patients with missense variants, p53 mRNA levels were above 75%. In contrast, p53 mRNA was clearly reduced in patients harbouring frameshift or splice variants (mean 58%, table 1 and figure 2) probably reflecting the activity of the nonsense- mediated mRNA decay.
Functional evaluation of TP53 variants of unknown biological significanceBased on these results, we refined the experimental thresh-olds for the analysis of unclassified variants. A functionality score above 7.5 was considered as indicative of a wild- type TP53 genotype, a score below 7.5 indicative of a variant impacting p53 function. A score below 5.5 is associated with a strong impact, and a score between 5.5 and 7.5 is associated with an intermediate effect. A reduction in p53 mRNA levels (<65%) was, by itself, suggestive of a defect independently of the functional score obtained. Using these criteria, we undertook the functional interpretation of 14 rare TP53 variants which remained unclassified despite the avail-able tools (table 1). These variants included 10 rare missense
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from
799Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Ph-e134Leu), p.(Arg158Cys), p.(Pro191Arg), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr)), 2 in- frame deletions (p.(Asn131del), p.(Leu265del)), 1 in- frame deletion–insertion [p.(Gly108_Phe109delinsVal)] and 1 rare variant within the 5′UTR region (c.-117G>T). For these rare variants (table 1), except p.(Pro191Arg) and c.-117G>T, the functional score was below 7.5, classifying these variants as ‘function-ally abnormal’, according to the terminology recently recom-mended for the functional assays.29 The p.(Pro72His) variant, which affects the same amino acid as the common p.(Pro72Arg) PEX4 polymorphism (rs1042522), yielded a score of 6.1 with no detectable effect on p53 mRNA, suggesting that it has a moderate effect on p53 function. For this patient, an EBV- cell
line was established, and the functional assay performed on the cell line yielded comparable results with a reduced score of 7.5 (normal score in EBV>10) and no impact on p53 mRNA levels. The p.(Pro191Arg) variant and the c.-117G>T variant had no detectable impact on the transcriptional activity, which led us to consider them as functionally normal.
Functional impact of the TP53 c.*1175A>C polymorphism located within the polyadenylation signalWe performed the assay in the unaffected mother (individual 76, table 1 and online supplemental table 3) of a young female patient (individual 77, online supplemental table 3) who devel-oped a high- grade glioma at 5 years of age and who carried a
Figure 1 P53 functional assay on peripheral blood. (A) Schematic representation of the blood p53 functional assay workflow. (B,C) Typical RT- QMPSF (B) and RT- MLPA (C) results obtained for individual 15 with a wild- type TP53 genotype. The fluorescent profiles of doxorubicin- treated cells (red line) and untreated cells (blue line) were superimposed using the three control amplicons (RIC8B, TBP and MPP5). The horizontal bars indicate for each p53 target gene the level of expression in untreated cells. Treatment efficacy was evaluated by the transcriptional repression of the PLK1 marker (Plk1 treated/untreated ratio below 0.5). In the treated condition, the peak height of each of the 10 p53 target genes was measured and divided by the sum of the heights of the three control genes. This value was divided by the same ratio calculated in the untreated condition to yield an arbitrary p53 functionality score. The p53 mRNA levels were expressed as a ratio of the normal values obtained for three control individuals. PBMC, peripheral blood mononuclear cell; RT- MLPA, reverse transcription–multiplex ligation probe amplification; RT- QMPSF, reverse transcription–quantitative multiplex PCR of short fluorescent fragment.
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

800 Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
frameshift TP53 variant [c.723del, p.(Cys242Alafs*5)], trans-mitted by her father (individual 58, table 1 and online supple-mental table 3). Unexpectedly, the mother had a reduced score of 5.5 and a moderate decrease in p53 mRNA levels (69%). A second blood sample yielded the same results (5.7; 73%). To explore this discrepancy, we reanalysed by NGS the whole TP53 locus in this family. This analysis revealed that the affected daughter carried the rs78378222 c.*1175A>C variant located within the polyadenylation signal, in addition to the frameshift TP53 variant. This polymorphic variant, present in 1.7% of non- Finnish Europeans, had previously been shown to be a risk factor for glioma and to impair 3′-end processing of p53 mRNA.30–33 Remarkably, the reanalysis of the TP53 locus in the mother revealed that she was homozy-gous for the c.*1175A>C variant. The p53 functional score in the index case harbouring the p.(Cys242Alafs*5) variant and the c.*1175A>C polymorphism (individual 77, online supplemental table 3) was lower than that obtained in her father harbouring only the frameshift variant (3.1 vs 7.1 and 6.0; see table 1 and online supplemental table 3). We analysed the impact of the c.*1175A>C variant on p53 mRNA 3’end processing using a dedicated RT- QMPSF with two amplicons located upstream and downstream of the polyadenylation site (‘exon 11’ and ‘postpoly- A’ respectively). This analysis revealed a reduction in p53 normal transcripts and the appearance of longer transcripts in the mother’s and daughter’s blood cells (figure 3).
DISCUSSIONThe interpretation of germline TP53 variants in patients with cancer is critical and should be performed before starting treat-ment considering their medical impact. The main objective of our assay was to provide a fast functional classification of rare uncharacterised variants in order to help clinicians with decision- making. Compared with the previous assay that we developed in EBV- immortalised lymphocytes,27 28 this blood assay does not require long- term cell culture and the results can be obtained within 1 week, fulfilling the timing required for diagnostic prac-tice. The only constraint is to perform it within 48 hours after blood sampling in order to obtain robust results. Under these
conditions, we were able to successfully analyse samples sent from other European countries.
Our assay fulfils most of the recommendations recently published by the Clinical Genome Resource Sequence Variant Interpretation working group regarding the clinical validity of functional assays29: (1) compared with the previously described p53 functional assays that test in vitro either cloned cDNA in yeast or artificial mutant libraries in cancer cell lines,23–25 this blood assay is performed in clinical samples in the patients’ genetic context; (2) the assay evaluates the transcriptional activity of p53 and not a specific domain of the protein; (3) it analyses simultaneously the impact of the variant on protein function and mRNA levels; (4) it was validated using 51 wild- type TP53 controls and 8 patients with seven distinct patho-genic or likely- pathogenic TP53 variants; and finally, (5) results show the robustness of the assay. Indeed, as shown in table 1, for 12 tested variants, we were able to perform the assay on EBV- immortalised cell lines and the results were very similar. Moreover, for five individuals, two different blood samples were tested and yielded similar results (table 1), and two vari-ants (c.844C>T, p.(Arg282Trp); c.847C>T, p.(Arg283Cys)) were tested on two different individuals’ blood with concordant results (4.8 vs 5.0 and 5.3 vs 6.4).
We observed among the wild- type TP53 individuals a wide range of functionality scores (7.5–22.8). This probably suggests that there is a variability of the p53- mediated tran-scriptional response to DNA damage in the general popula-tion, although no obvious impact of age, clinical status or sex could be observed. The thresholds used in this study could be refined by testing additional deleterious variants. Despite this variability, all pathogenic/likely pathogenic variants generated low p53 functionality scores, and variants resulting in prema-ture stop codons were also detected by a clear reduction of p53 mRNA levels. In addition, our assay allows testing of non- missense variants such as in frame indels. It should be highlighted that none of the previously published functional assays can be considered as a gold- standard method to classify germline TP53 variants.23–25 Therefore, no available p53 func-tional assay can be used to calibrate the blood assay. Indeed, as illustrated in table 1, discordant results were obtained for
Figure 2 p53 functional scores and mRNA level ratios in individuals with wild- type TP53 or with germline TP53 variants. (A) p53 functionality scores obtained in 51 wild- type TP53 individuals, compared with the scores obtained for nine samples from eight individuals carrying a classified TP53 variant (online supplemental table 3) using a Mann- Whitney non- parametric test. (B) Comparison of the p53 mRNA ratios obtained in 51 wild- type TP53 individuals and in samples carrying a missense (five samples) or a truncating variant of TP53 (four samples), using a Kruskal- Wallis test with Dunns post- test (p=0.0031). ***P<0.01.
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

801Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
Tabl
e 1
Inte
rpre
tatio
n of
ger
mlin
e TP
53 v
aria
nts
inte
grat
ing
the
bloo
d p5
3 fu
nctio
nal a
ssay
Vari
ant*
MA
F†
Bioi
nfor
mat
ic p
redi
ctio
ns‡
Func
tion
al s
tatu
s of
the
var
iant
s as
sess
ed in
yea
st a
nd h
uman
can
cer
cell
lines
Clin
Var c
lass
ifica
tion§
Bloo
d p5
3fu
ncti
onal
ass
ay¶
in p
53
scor
e, m
RNA
%
p53
func
tion
al a
ssay
in E
BV c
ell l
ines
sc
ore;
mRN
A%
Inte
rpre
tati
on
acco
rdin
g to
the
ass
ayA
lign
GVG
DPo
lyPh
en-2
SIFT
Baye
sD
elKa
toas
say*
*Ko
tler
assa
y††
Gia
com
elli
assa
y‡‡
Path
ogen
ic/li
kely
pat
hoge
nic
vari
ants
c.52
4G>
Ap.
(Arg
175H
is)
0.00
0004
C25
PD
0.54
62N
on- f
unct
iona
lN
on- f
unct
iona
lLO
FD
NE
Path
ogen
icIn
d. 5
2: 3
.2; 9
6%2.
9; 9
3%4.
1; 8
6%Fu
ncti
onal
ly a
bnor
mal
c.54
2G>
Ap.
(Arg
181H
is)
0.00
0014
C25
DD
0.25
84Pa
rtia
lly fu
ncti
onal
Func
tiona
lU
ncla
ssifi
edPa
thog
enic
/like
ly
path
ogen
icIn
d. 5
3: 3
.8; 8
1%Fu
ncti
onal
ly a
bnor
mal
c.84
4C>
Tp.
(Arg
282T
rp)
0.00
0004
C65
DD
0.54
18N
on- f
unct
iona
lN
on- f
unct
iona
lLO
FD
NE
Path
ogen
ic/li
kely
pa
thog
enic
Ind.
54:
4.8
; 102
%In
d. 5
5: 5
.0; 9
8%2.
0; 1
04%
2.7;
116
%4.
6; 7
6%
Func
tion
ally
abn
orm
al
c.10
10G
>A
p.(A
rg33
7His
)0.
0000
12C2
5D
D0.
1777
Part
ially
func
tion
alN
AN
ot L
OF
Not
DN
EPa
thog
enic
Ind.
56:
5.4
; 101
%5.
8; 8
1%6.
0; 8
5%6.
5; 8
1%6.
9; 1
01%
Func
tion
ally
abn
orm
al
c.37
5G>
Ap.
?Sp
licin
g Al
tera
tion
NR
//
//
/Fu
nctio
nal
NA
Path
ogen
icIn
d. 5
7: 3
.1; 6
1%5.
5; 6
1%§§
Func
tion
ally
abn
orm
al
c.72
3del
p.(C
ys24
2Ala
fs*5
)N
R/
//
/N
AN
AN
AN
RIn
d. 5
8:S1
: 7.1
; 56%
S2: 6
.0; 5
2%
4.2;
52%
§§Fu
ncti
onal
ly a
bnor
mal
c.77
0del
p.(L
eu25
7Arg
fs*8
8)N
R/
//
/N
AN
on- f
unct
iona
lN
AN
RIn
d. 5
9: 4
.7; 6
2%Fu
ncti
onal
ly a
bnor
mal
Beni
gn v
aria
nt
c.70
4A>
Gp.
(Asn
235S
er)
0.01
945
C0B
T−
0.03
42Fu
nctio
nal
Func
tiona
lN
ot L
OF
Not
DN
EBe
nign
Ind.
60:
12.
9; 1
18%
Func
tiona
lly n
orm
al
Unc
lass
ified
rar
e va
rian
ts
c.21
5C>
Ap.
(Pro
72Hi
s)N
RC0
PD
−0.
1017
Func
tiona
lN
AN
ot L
OF
Not
DN
EU
ncer
tain
sig
nific
ance
Ind.
61:
6.1
; 86%
7.5;
107
%§§
Func
tion
ally
abn
orm
al
c.31
4G>
Ap.
(Gly
105A
sp)
NR
C65
DD
0.56
83Pa
rtia
lly fu
ncti
onal
Func
tiona
lLO
FD
NE
Confl
ict.
int¶
¶: L
ikel
y pa
thog
enic
/ unc
erta
in
sign
ifica
nce
Ind.
62:
2.9
; 82%
2.1;
85%
3.6;
70%
Func
tion
ally
abn
orm
al
c.32
9G>
Ap.
(Arg
110H
is)
0.00
0046
C0B
T0.
0728
Part
ially
func
tion
alFu
nctio
nal
Not
LO
FN
ot D
NE
Unc
erta
in s
igni
fican
ceIn
d. 6
3: 4
.1; 8
3%3.
7; 9
2%4.
7; 9
1%§§
5.2;
94%
Func
tion
ally
abn
orm
al
c.40
2T>
Gp.
(Phe
134L
eu)
NR
C0D
D0.
3166
Non
- fun
ctio
nal
Non
- fun
ctio
nal
LOF
DN
EN
RIn
d. 6
4: 3
.5; 7
8%3.
5; 9
4%§§
Func
tion
ally
abn
orm
al
c.47
2C>
Tp.
(Arg
158C
ys)
0.00
0008
C65
DD
0.51
45Pa
rtia
lly fu
ncti
onal
Func
tiona
lU
ncla
ssifi
edCo
nflic
t. in
t:Li
kely
pa
thog
enic
/unc
erta
in
sign
ifica
nce
Ind.
65:
3.2
; 86%
Func
tion
ally
abn
orm
al
c.57
2C>
Gp.
(Pro
191A
rg)
0.00
0012
C35
PD
0.09
42Fu
nctio
nal
Func
tiona
lN
ot L
OF
Not
DN
EU
ncer
tain
sig
nific
ance
Ind.
66:
17.
1; 9
0%Fu
nctio
nally
nor
mal
c.83
3C>
Gp.
(Pro
278A
rg)
NR
C65
DD
0.60
78N
on- f
unct
iona
lN
on- f
unct
iona
lLO
FD
NE
Unc
erta
in s
igni
fican
ceIn
d. 6
7: 5
.1; 7
8%3.
9; 1
08%
§§Fu
ncti
onal
ly a
bnor
mal
c.84
7C>
Tp.
(Arg
283C
ys)
0.00
0074
C55
BD
0.33
09Fu
nctio
nal
Func
tiona
lN
ot L
OF
Not
DN
EU
ncer
tain
sig
nific
ance
Ind.
68:
5.3
; 79%
Ind.
69:
6.4
; 88%
3.1;
96%
2.5;
89%
§§5.
6; 8
6%5.
4; 8
8%
Func
tion
ally
abn
orm
al
c.10
43T>
Cp.
(Leu
348S
er)
NR
C65
DD
0.54
39N
on- f
unct
iona
lN
AN
ot L
OF
Not
DN
EN
RIn
d. 7
0:S1
: 5.8
; 89%
S2 :
6.1;
90%
Func
tion
ally
abn
orm
al
c.10
54G
>T
p.(A
sp35
2Tyr
)N
RC3
5D
D0.
0844
Func
tiona
lN
AN
ot L
OF
Not
DN
EU
ncer
tain
sig
nific
ance
Ind.
71*
**:
S1: 4
.0; 1
01%
S2: 5
.0; 1
06%
2.3;
90%
Func
tion
ally
abn
orm
al
c.32
3_32
5del
p.(G
ly10
8_Ph
e109
delin
sVal
)N
R/
//
//
//
NR
Ind.
72:
3.8
; 94%
Func
tion
ally
abn
orm
al
Cont
inue
d
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

802 Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
variants unambiguously classified in ClinVar as pathogenic or likely pathogenic. In particular, the founder Brazilian p.(Ar-g337His), an example of a variant with low penetrance, high-lights the limits of the available tools. Whereas segregation data performed on large Brazilian pedigrees have clearly shown that this variant is pathogenic,34 bioinformatic predic-tions and functional analyses35 are conflicting (table 1). Our blood functional assay clearly shows that this variant alters the transcriptional activity of p53, although to a lesser extent than DNE missense variations, highlighting the limits of func-tional assays based on overexpression of cDNA. This result was confirmed in four additional patients carrying this variant using EBV cell lines (table 1).
The blood functional assay performed on PBMC harbouring unclassified variants led us to consider 12 variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Phe134Leu), p.(Arg158Cys), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr), p.(Gly108_Phe109delinsVal), p.(Asn131del), p.(Leu265del)) as ‘functionally abnormal’, some with high impact. The inter-pretation is particularly challenging for p.(Pro72His), p.(Ar-g110His), p.(Arg158Cys), p.(Arg283Cys) and p.(Asp352Tyr) variants, as they were considered in yeast assays as functional or partially functional, and the Giacomelli assay classified them as not LOF_not DNE or was not conclusive. The low function-ality score observed for p.(Arg110His) was confirmed in an EBV cell line derived from the patient and confirmed in two EBV cell lines from other patients carrying this variant. The result for the p.(Asp352Tyr) variant was confirmed on a second blood sample and with an EBV cell line derived from another patient also carrying this variant. The effect of p.(Arg283Cys) was also confirmed in EBV cell lines derived from the patient and from three additional patients with the same variant (table 1).
The clinical utility of the p53 functional assay is high-lighted by the p.(Pro191Arg) variant. This variant was initially detected in a child with medulloblastoma at 2 years of age and whose brother died from a fibrosarcoma. Presymptomatic testing revealed that an unaffected brother (18 months), the mother and two maternal aunts were also carriers. We were then requested to evaluate this variant, and the functional assay performed in the maternal aunt (individual 65, online supplemental table 3) clearly showed that this variant does not alter the p53 transcriptional activity (table 1 and online supplemental table 3). Considering this result, segregation analysis was performed on the brother’s fibrosarcoma sample, revealing the absence of the variant and consolidating the conclusion of a non- pathogenic variant.
Our results show that this blood functional assay is also able to detect TP53 variations outside the coding regions, which are the only regions commonly analysed. Thanks to this assay, we discovered that the unaffected mother of an index case was homozygous for the polymor-phic c.*1175A>C variant, and we show that this variant decreases p53 mRNA by altering the polyadenylation signal and produces longer transcripts extending beyond the poly- A site, as previously reported.30 When present on both alleles, this variant impacts p53 functionality with the same magni-tude as a germline pathogenic TP53 variant. This prompted us to recommend breast MRI every year for this unaffected adult relative. We had the opportunity to perform the assay on EBV- immortalised lymphocytes harbouring only this heterozygous variant, and we observed a normal score (data not shown), suggesting that the heterozygous c.*1175A>C variant alone is insufficient to alter p53 function. The comparison of the p53 functional scores observed in the Va
rian
t*M
AF†
Bioi
nfor
mat
ic p
redi
ctio
ns‡
Func
tion
al s
tatu
s of
the
var
iant
s as
sess
ed in
yea
st a
nd h
uman
can
cer
cell
lines
Clin
Var c
lass
ifica
tion§
Bloo
d p5
3fu
ncti
onal
ass
ay¶
in p
53
scor
e, m
RNA
%
p53
func
tion
al a
ssay
in E
BV c
ell l
ines
sc
ore;
mRN
A%
Inte
rpre
tati
on
acco
rdin
g to
the
ass
ayA
lign
GVG
DPo
lyPh
en-2
SIFT
Baye
sD
elKa
toas
say*
*Ko
tler
assa
y††
Gia
com
elli
assa
y‡‡
c.39
3_39
5del
p.(A
sn13
1del
)N
R/
//
/N
AN
on- f
unct
iona
lN
AU
ncer
tain
sig
nific
ance
Ind.
73:
S1: 2
.6; 8
0%S2
: 4.8
; 99%
Func
tion
ally
abn
orm
al
c.79
2_79
4del
p.(L
eu26
5del
)N
R/
//
//
NA
NA
NR
Ind.
74:
3.4
; 95%
Func
tion
ally
abn
orm
al
c.-1
17G
>T
p.(=
) (5'
UTR
)N
R/
//
/N
AN
AN
AN
RIn
d. 7
5: 2
0.5;
100
%Fu
nctio
nally
nor
mal
Poly
mor
phis
m
c.*1
175A
>C
p.(=
)(H
omoz
ygou
s)
0.01
189
//
//
NA
NA
NA
Confl
ict.
int:
likel
y be
nign
/unc
erta
in
sign
ifica
nce
Ind.
76:
S1: 5
.5; 6
9%S2
: 5.7
; 73%
Func
tion
ally
abn
orm
al
whe
n ho
moz
ygou
s
*Des
crib
ed a
ccor
ding
to th
e re
fere
nce
tran
scrip
t NM
_000
546.
5.†A
llele
freq
uenc
y in
the
gene
ral p
opul
atio
n, a
s in
dica
ted
in g
nom
AD (h
ttps
://gn
omad
.bro
adin
stitu
te.o
rg/).
‡Pre
dict
ion
of th
e va
riant
impa
ct o
n pr
otei
n, a
ccor
ding
to th
e Al
ign
GVG
D (C
0: to
lera
ted,
oth
er c
lass
es: d
amag
ing)
, Pol
yPhe
n- 2
(D: p
roba
bly
dam
agin
g, P
: pos
sibl
y da
mag
ing,
B: b
enig
n), S
IFT
(D: D
amag
ing,
T: to
lera
ted)
and
Bay
esDe
l (sc
ore≥
0.16
: dam
agin
g/sc
ore
<0.
16: t
oler
ated
, as
indi
cate
d in
For
tuno
et a
l, 20
1820
) alg
orith
ms.
§The
Clin
Var c
lass
ifica
tion
of th
e va
riant
(htt
ps://
ww
w.n
cbi.n
lm.n
ih.g
ov/c
linva
r/) is
indi
cate
d.¶F
or e
ach
varia
nt a
re in
dica
ted
the
indi
vidu
al w
ho w
as te
sted
and
the
mea
n va
lues
of t
he fu
nctio
nalit
y sc
ore
and
of th
e m
RNA
expr
essi
on, a
s de
term
ined
usi
ng th
e RT
- QM
PSF
and
RT- M
LPA
assa
ys (s
ee ta
ble
1). F
or s
ome
indi
vidu
als,
two
inde
pend
ent s
ampl
es (S
1 an
d S2
) wer
e te
sted
.**
As c
lass
ified
in th
e In
tern
atio
nal A
genc
y fo
r Res
earc
h on
Can
cer (
IARC
) dat
abas
e (h
ttp:
//p53
.iarc
.fr/),
acc
ordi
ng to
the
resu
lts o
btai
ned
in th
e ye
ast t
rans
activ
atio
n as
say
(Kat
o et
al23
)††
As d
eter
min
ed in
the
assa
y de
velo
ped
by K
otle
r et a
l24 a
nd b
ased
on
the
quan
tifica
tion
of th
e an
tipro
lifer
ativ
e ac
tivity
of t
he v
aria
nt in
the
p53-
null
H129
9 ca
ncer
cel
l lin
e.‡‡
As d
eter
min
ed in
the
assa
ys d
evel
oped
by
Gia
com
elli
et a
l25 a
nd b
ased
on
the
abili
ty o
f the
var
iant
to re
stor
e th
e su
rviv
al o
f the
p53
- nul
l A45
9 ce
ll lin
e af
ter e
xpos
ure
to h
igh
dose
s of
eto
posi
de D
NA
brea
king
age
nts
and
to in
duce
in p
53- w
ild- t
ype
A459
cel
l lin
e re
sist
ance
to N
utlin
- 3.
§§p5
3 fu
nctio
nal a
ssay
was
als
o ca
rrie
d ou
t on
an E
BV c
ell l
ine
deriv
ed fr
om th
e sa
me
patie
nt a
nd le
d to
the
sam
e in
terp
reta
tion
as th
e bl
ood
p53
func
tiona
l ass
ay (n
orm
al s
core
for E
BV>
10).
Item
s in
bol
d in
dica
te re
sults
sug
gest
ive
of a
del
eter
ious
effe
ct o
f the
var
iant
.¶¶
Confl
ictin
g in
terp
reta
tions
of p
atho
geni
city
.**
*For
this
indi
vidu
al, a
sec
ond
TP53
var
iatio
n w
as id
entifi
ed in
cis
: c.1
021T
>A,
p.(P
he31
4Ile
), w
hich
is c
lass
ified
as
beni
gn.
DNE,
dom
inan
t neg
ativ
e ef
fect
; LO
F, lo
ss o
f fun
ctio
n; N
A, n
ot a
naly
sed;
NR,
not
repo
rted
; RT-
MLP
A, re
vers
e tr
ansc
riptio
n–m
ultip
lex
ligat
ion
prob
e am
plifi
catio
n; R
T- Q
MPS
F, re
vers
e tr
ansc
riptio
n–qu
antit
ativ
e m
ultip
lex
PCR
of s
hort
fluo
resc
ent f
ragm
ent.
Tabl
e 1
Cont
inue
d
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

803Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
index case who developed a high- grade glioma at 5 years of age and harbours the null c.723del, p.(Cys242Alafs*5) variant and in trans the polymorphic c.*1175A>C variant, and in her father carrying only the TP53 null variant suggests
that the c.*1175A>C variant may act as a genetic modifier in pathogenic TP53 variant carriers and could increase the risk of glioma in carriers, as previously shown in the general population.30–33
Figure 3 Impact of the heterozygous and homozygous TP53 c.*1175A>C variation on p53 pre- mRNA 3′ end processing. (A) Schematic representation of the TP53 3′ end region. The c.*1175A>C variant is predicted to yield at least two different transcripts; the upper one corresponds to the normal transcript with pre- mRNA cleavage and polyadenylation, and the lower one to longer transcript that extends after the poly- A signal. ‘Exon 11’ primers amplify both transcripts, while ‘postpoly- A’ primers specifically amplify the longer transcripts. As postpoly- A primers could also amplify gDNA, primers ‘exon 7’ and ‘exon 10’, which are specific to gDNA, were added to the reaction in order to monitor DNA contamination. (B) RT- QMPSF result obtained for the index case’s father (individual 58, S1; table 1 and online supplemental table 3) carrying the variant TP53 c.723del, p.(Cys242Alafs*5). The profile (in red) was superimposed on the profile of a control individual wild type for TP53 (in blue), using the control amplicons RIC8B and TBP. (C) RT- QMPSF result obtained for the index case’s mother (individual 76, S1; table 1 and online supplemental table 3) carrying the c.*1175A>C variant at the homozygous state. (D) RT- QMPSF result for the index case (individual 77, online supplemental table 3) carrying the c.723del, p.(Cys242Alafs*5) variant and the c.*1175A>C in trans. Red arrows indicate the appearance of longer p53 transcripts. The horizontal bars show the reduction of the normal p53 transcript level, as compared with the control. RT- QMPSF, reverse transcription–quantitative multiplex PCR of short fluorescent fragment.
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

804 Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
In summary, we suggest that our blood p53 functional assay should be a useful tool not only for the rapid interpretation of germline TP53 variants of unknown significance in clinical practice, in complement to the previously developed assays, but also for the indirect detection of cryptic alterations within regulatory regions impacting p53 function.
Acknowledgements The authors are grateful to their French and European colleagues for providing clinical information and sending blood samples for TP53 analysis. The authors are indebted to Philippe Ruminy (Inserm U1245, Comprehensive Cancer Centre Becquerel, Rouen) for advices on the reverse transcription–multiplex ligation probe amplification experiments and to Nikki Sabourin- Gibbs (Rouen University Hospital) for her assistance in editing the manuscript.
Contributors IT, GB and TF formulated research goals and aims. Functional analyses were performed by SR and MR. RNA- Seq was performed by SR, CD and IT and bioinformatic analysis by RL and SC. Molecular analyses were performed by JB, EB, FC, GL, SV and supervised by GB, EK and SBD. TF, IT and GB supervised the overall progress and manuscript writing. SR and MR contributed equally to this work.
Funding This work was supported by the North- West Canceropole, the European Regional Development Fund (European Union and Région Normandie), the Association pour la Recherche contre le Cancer and the Ligue Contre le Cancer.
Competing interests None declared.
Patient consent for publication Not required.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information. Deidentified participant data are available from thierry. frebourg@ chu- rouen. fr.
Open access This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY- NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non- commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non- commercial. See: http:// creativecommons. org/ licenses/ by- nc/ 4. 0/.
ORCID iDIsabelle Tournier http:// orcid. org/ 0000- 0002- 5544- 048X
REFERENCES 1 Frebourg T, Bajalica Lagercrantz S, Oliveira C, Magenheim R, Evans DG, European
Reference Network GENTURIS. Guidelines for the Li- Fraumeni and heritable TP53- related cancer syndromes. Eur J Hum Genet 2020. doi:10.1038/s41431-020-0638-4. [Epub ahead of print: 26 May 2020].
2 Bougeard G, Renaux- Petel M, Flaman J- M, Charbonnier C, Fermey P, Belotti M, Gauthier- Villars M, Stoppa- Lyonnet D, Consolino E, Brugières L, Caron O, Benusiglio PR, Bressac- de Paillerets B, Bonadona V, Bonaïti- Pellié C, Tinat J, Baert- Desurmont S, Frebourg T. Revisiting Li- Fraumeni syndrome from TP53 mutation carriers. J Clin Oncol 2015;33:2345–52.
3 Mai PL, Best AF, Peters JA, DeCastro RM, Khincha PP, Loud JT, Bremer RC, Rosenberg PS, Savage SA. Risks of first and subsequent cancers among TP53 mutation carriers in the National cancer Institute Li- Fraumeni syndrome cohort. Cancer 2016;122:3673–81.
4 Kasper E, Angot E, Colasse E, Nicol L, Sabourin J- C, Adriouch S, Lacoume Y, Charbonnier C, Raad S, Frebourg T, Flaman J- M, Bougeard G. Contribution of genotoxic anticancer treatments to the development of multiple primary tumours in the context of germline TP53 mutations. Eur J Cancer 2018;101:254–62.
5 Villani A, Shore A, Wasserman JD, Stephens D, Kim RH, Druker H, Gallinger B, Naumer A, Kohlmann W, Novokmet A, Tabori U, Tijerin M, Greer M- LC, Finlay JL, Schiffman JD, Malkin D. Biochemical and imaging surveillance in germline TP53 mutation carriers with Li- Fraumeni syndrome: 11 year follow- up of a prospective observational study. Lancet Oncol 2016;17:1295–305.
6 Kratz CP, Achatz MI, Brugières L, Frebourg T, Garber JE, Greer M- LC, Hansford JR, Janeway KA, Kohlmann WK, McGee R, Mullighan CG, Onel K, Pajtler KW, Pfister SM, Savage SA, Schiffman JD, Schneider KA, Strong LC, Evans DGR, Wasserman JD, Villani A, Malkin D. Cancer screening recommendations for individuals with Li- Fraumeni syndrome. Clin Cancer Res 2017;23:e38–45.
7 Ballinger ML, Best A, Mai PL, Khincha PP, Loud JT, Peters JA, Achatz MI, Chojniak R, Balieiro da Costa A, Santiago KM, Garber J, O’Neill AF, Eeles RA, Evans DG, Bleiker E, Sonke GS, Ruijs M, Loo C, Schiffman J, Naumer A, Kohlmann W, Strong LC, Bojadzieva J, Malkin D, Rednam SP, Stoffel EM, Koeppe E, Weitzel JN, Slavin TP, Nehoray B, Robson M, Walsh M, Manelli L, Villani A, Thomas DM, Savage SA. Baseline
surveillance in Li- Fraumeni syndrome using whole- body magnetic resonance imaging: a meta- analysis. JAMA Oncol 2017;3:1634–9.
8 Caron O, Frebourg T, Benusiglio PR, Foulon S, Brugières L. Lung adenocarcinoma as part of the Li- Fraumeni syndrome spectrum: preliminary data of the LIFSCREEN randomized clinical trial. JAMA Oncol 2017;3:1736–7.
9 Ruijs MWG, Loo CE, van Buchem CAJM, Bleiker EMA, Sonke GS. Surveillance of Dutch patients with Li- Fraumeni syndrome: the LiFe- Guard study. JAMA Oncol 2017;3:1733–4.
10 Saya S, Killick E, Thomas S, Taylor N, Bancroft EK, Rothwell J, Benafif S, Dias A, Mikropoulos C, Pope J, Chamberlain A, Gunapala R, Izatt L, Side L, Walker L, Tomkins S, Cook J, Barwell J, Wiles V, Limb L, Eccles D, Leach MO, Shanley S, Gilbert FJ, Hanson H, Gallagher D, Rajashanker B, Whitehouse RW, Koh D- M, Sohaib SA, Evans DG, Eeles RA, SIGNIFY Study Steering Committee. Baseline results from the UK signify study: a whole- body MRI screening study in TP53 mutation carriers and matched controls. Fam Cancer 2017;16:433–40.
11 Bojadzieva J, Amini B, Day SF, Jackson TL, Thomas PS, Willis BJ, Throckmorton WR, Daw NC, Bevers TB, Strong LC. Whole body magnetic resonance imaging (WB- MRI) and brain MRI baseline surveillance in TP53 germline mutation carriers: experience from the Li- Fraumeni syndrome education and early detection (lead) clinic. Fam Cancer 2018;17:287–94.
12 O’Neill AF, Voss SD, Jagannathan JP, Kamihara J, Nibecker C, Itriago- Araujo E, Masciari S, Parker E, Barreto M, London WB, Garber JE, Diller L. Screening with whole- body magnetic resonance imaging in pediatric subjects with Li- Fraumeni syndrome: a single institution pilot study. Pediatr Blood Cancer 2018;65. doi:10.1002/pbc.26822. [Epub ahead of print: 27 Oct 2017].
13 Paixão D, Guimarães MD, de Andrade KC, Nóbrega AF, Chojniak R, Achatz MI. Whole- Body magnetic resonance imaging of Li- Fraumeni syndrome patients: observations from a two rounds screening of Brazilian patients. Cancer Imaging 2018;18.
14 Bancroft EK, Saya S, Brown E, Thomas S, Taylor N, Rothwell J, Pope J, Chamberlain A, Page E, Benafif S, Hanson H, Dias A, Mikropoulos C, Izatt L, Side L, Walker L, Donaldson A, Cook JA, Barwell J, Wiles V, Limb L, Eccles DM, Leach MO, Shanley S, Gilbert FJ, Gallagher D, Rajashanker B, Whitehouse RW, Koh D- M, Sohaib SA, Evans DG, Eeles RA, Walker LG. Psychosocial effects of whole- body MRI screening in adult high- risk pathogenic TP53 mutation carriers: a case- controlled study (SIGNIFY). J Med Genet 2020;57:226–36.
15 Li FP, Fraumeni JF. Soft- Tissue sarcomas, breast cancer, and other neoplasms. A familial syndrome? Ann Intern Med 1969;71:747–52.
16 Malkin D, Li FP, Strong LC, Fraumeni JF, Nelson CE, Kim DH, Kassel J, Gryka MA, Bischoff FZ, Tainsky MA, Nelson CE, Nelson CE, Friend SH. Germ line p53 mutations in a familial syndrome of breast cancer, sarcomas, and other neoplasms. Science 1990;250:1233–8.
17 Srivastava S, Zou ZQ, Pirollo K, Blattner W, Chang EH. Germ- Line transmission of a mutated p53 gene in a cancer- prone family with Li- Fraumeni syndrome. Nature 1990;348:747–9.
18 Gonzalez KD, Noltner KA, Buzin CH, Gu D, Wen- Fong CY, Nguyen VQ, Han JH, Lowstuter K, Longmate J, Sommer SS, Weitzel JN. Beyond Li Fraumeni syndrome: clinical characteristics of families with p53 germline mutations. J Clin Oncol 2009;27:1250–6.
19 Ruijs MWG, Verhoef S, Rookus MA, Pruntel R, van der Hout AH, Hogervorst FBL, Kluijt I, Sijmons RH, Aalfs CM, Wagner A, Ausems MGEM, Hoogerbrugge N, van Asperen CJ, Gomez Garcia EB, Meijers- Heijboer H, Ten Kate LP, Menko FH, van ’t Veer LJ. Tp53 germline mutation testing in 180 families suspected of Li- Fraumeni syndrome: mutation detection rate and relative frequency of cancers in different familial phenotypes. J Med Genet 2010;47:421–8.
20 Fortuno C, James PA, Young EL, Feng B, Olivier M, Pesaran T, Tavtigian SV, Spurdle AB. Improved, ACMG- compliant, in silico prediction of pathogenicity for missense substitutions encoded by TP53 variants. Hum Mutat 2018;39:1061–9.
21 Fortuno C, Cipponi A, Ballinger ML, Tavtigian SV, Olivier M, Ruparel V, Haupt Y, Haupt S, Study ISK, Tucker K, Spurdle AB, Thomas DM, James PA. A quantitative model to predict pathogenicity of missense variants in the TP53 gene. Hum Mutat 2019;40:788–800.
22 ClinGen. TP53- specific ACMG/AMP guidelines. Available: https:// clinicalgenome. org/ site/ assets/ files /3876/ clingen_ tp53_ acmg_ specifications_ v1. pdf
23 Kato S, Han S- Y, Liu W, Otsuka K, Shibata H, Kanamaru R, Ishioka C. Understanding the function- structure and function- mutation relationships of p53 tumor suppressor protein by high- resolution missense mutation analysis. Proc Natl Acad Sci U S A 2003;100:8424–9.
24 Kotler E, Shani O, Goldfeld G, Lotan- Pompan M, Tarcic O, Gershoni A, Hopf TA, Marks DS, Oren M, Segal E. A systematic p53 mutation library links differential functional impact to cancer mutation pattern and evolutionary conservation. Mol Cell 2018;71:178–90.
25 Giacomelli AO, Yang X, Lintner RE, McFarland JM, Duby M, Kim J, Howard TP, Takeda DY, Ly SH, Kim E, Gannon HS, Hurhula B, Sharpe T, Goodale A, Fritchman B, Steelman S, Vazquez F, Tsherniak A, Aguirre AJ, Doench JG, Piccioni F, Roberts CWM, Meyerson M, Getz G, Johannessen CM, Root DE, Hahn WC. Mutational processes shape the landscape of TP53 mutations in human cancer. Nat Genet 2018;50:1381–7.
26 Hafner A, Bulyk ML, Jambhekar A, Lahav G. The multiple mechanisms that regulate p53 activity and cell fate. Nat Rev Mol Cell Biol 2019;20:199–210.
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

805Raad S, et al. J Med Genet 2021;58:796–805. doi:10.1136/jmedgenet-2020-107059
Cancer genetics
27 Zerdoumi Y, Aury- Landas J, Bonaïti- Pellié C, Derambure C, Sesboüé R, Renaux- Petel M, Frebourg T, Bougeard G, Flaman J- M. Drastic effect of germline TP53 missense mutations in Li- Fraumeni patients. Hum Mutat 2013;34:453–61.
28 Zerdoumi Y, Lanos R, Raad S, Flaman J- M, Bougeard G, Frebourg T, Tournier I. Germline TP53 mutations result into a constitutive defect of p53 DNA binding and transcriptional response to DNA damage. Hum Mol Genet 2017;26:2591–602.
29 Brnich SE, Abou Tayoun AN, Couch FJ, Cutting GR, Greenblatt MS, Heinen CD, Kanavy DM, Luo X, McNulty SM, Starita LM, Tavtigian SV, Wright MW, Harrison SM, Biesecker LG, Berg JS, Clinical Genome Resource Sequence Variant Interpretation Working Group. Recommendations for application of the functional evidence PS3/BS3 criterion using the ACMG/AMP sequence variant interpretation framework. Genome Med 2019;12:3.
30 Stacey SN, Sulem P, Jonasdottir A, Masson G, Gudmundsson J, Gudbjartsson DF, Magnusson OT, Gudjonsson SA, Sigurgeirsson B, Thorisdottir K, Ragnarsson R, Benediktsdottir KR, Nexø BA, Tjønneland A, Overvad K, Rudnai P, Gurzau E, Koppova K, Hemminki K, Corredera C, Fuentelsaz V, Grasa P, Navarrete S, Fuertes F, García- Prats MD, Sanambrosio E, Panadero A, De Juan A, Garcia A, Rivera F, Planelles D, Soriano V, Requena C, Aben KK, van Rossum MM, Cremers RGHM, van Oort IM, van Spronsen D- J, Schalken JA, Peters WHM, Helfand BT, Donovan JL, Hamdy FC, Badescu D, Codreanu O, Jinga M, Csiki IE, Constantinescu V, Badea P, Mates IN, Dinu DE, Constantin A, Mates D, Kristjansdottir S, Agnarsson BA, Jonsson E, Barkardottir RB, Einarsson GV, Sigurdsson F, Moller PH, Stefansson T, Valdimarsson T, Johannsson OT, Sigurdsson H, Jonsson T, Jonasson JG, Tryggvadottir L, Rice T, Hansen HM, Xiao Y, Lachance DH, O Neill BP, Kosel ML, Decker PA, Thorleifsson G, Johannsdottir H, Helgadottir HT, Sigurdsson A, Steinthorsdottir V, Lindblom A, Sandler RS, Keku TO, Banasik K, Jørgensen T, Witte DR, Hansen T, Pedersen O, Jinga V, Neal DE, Catalona WJ, Wrensch M, Wiencke J, Jenkins RB, Nagore E, Vogel U, Kiemeney LA, Kumar R, Mayordomo JI, Olafsson JH, Kong A, Thorsteinsdottir U, Rafnar T, Stefansson K, Swedish Low- risk
Colorectal Cancer Study Group. A germline variant in the TP53 polyadenylation signal confers cancer susceptibility. Nat Genet 2011;43:1098–103.
31 Egan KM, Nabors LB, Olson JJ, Monteiro AN, Browning JE, Madden MH, Thompson RC. Rare TP53 genetic variant associated with glioma risk and outcome. J Med Genet 2012;49:420–1.
32 Enciso- Mora V, Hosking FJ, Di Stefano AL, Zelenika D, Shete S, Broderick P, Idbaih A, Delattre J- Y, Hoang- Xuan K, Marie Y, Labussière M, Alentorn A, Ciccarino P, Rossetto M, Armstrong G, Liu Y, Gousias K, Schramm J, Lau C, Hepworth SJ, Schoemaker M, Strauch K, Müller- Nurasyid M, Schreiber S, Franke A, Moebus S, Eisele L, Swerdlow A, Simon M, Bondy M, Lathrop M, Sanson M, Houlston RS. Low penetrance susceptibility to glioma is caused by the TP53 variant rs78378222. Br J Cancer 2013;108:2178–85.
33 Wang Z, Rajaraman P, Melin BS, Chung CC, Zhang W, McKean- Cowdin R, Michaud D, Yeager M, Ahlbom A, Albanes D, Andersson U, Freeman LEB, Buring JE, Butler MA, Carreón T, Feychting M, Gapstur SM, Gaziano JM, Giles GG, Hallmans G, Henriksson R, Hoffman- Bolton J, Inskip PD, Kitahara CM, Marchand LL, Linet MS, Li S, Peters U, Purdue MP, Rothman N, Ruder AM, Sesso HD, Severi G, Stampfer M, Stevens VL, Visvanathan K, Wang SS, White E, Zeleniuch- Jacquotte A, Hoover R, Fraumeni JF, Chatterjee N, Hartge P, Chanock SJ. Further confirmation of germline glioma risk variant rs78378222 in TP53 and its implication in tumor tissues via integrative analysis of TCGA data. Hum Mutat 2015;36:684–8.
34 Achatz MIW, Olivier M, Le Calvez F, Martel- Planche G, Lopes A, Rossi BM, Ashton- Prolla P, Giugliani R, Palmero EI, Vargas FR, Da Rocha JCC, Vettore AL, Hainaut P. The TP53 mutation, R337H, is associated with Li- Fraumeni and Li- Fraumeni- like syndromes in Brazilian families. Cancer Lett 2007;245:96–102.
35 Wasserman JD, Novokmet A, Eichler- Jonsson C, Ribeiro RC, Rodriguez- Galindo C, Zambetti GP, Malkin D. Prevalence and functional consequence of TP53 mutations in pediatric adrenocortical carcinoma: a children’s Oncology Group study. J Clin Oncol 2015;33:602–9.
on February 4, 2022 by guest. P
rotected by copyright.http://jm
g.bmj.com
/J M
ed Genet: first published as 10.1136/jm
edgenet-2020-107059 on 13 October 2020. D
ownloaded from

Supplementary table 1 RT-QMPSF primer sequences and RT-MLPA probe sequences used for the blood p53 functional assay
RT-QMPSF RT-MLPA Gene Primer sequences a,b Length
(bp) Probe sequences a,c,d Length (bp)
CEP170B F 5’-6FAM-TCAACGCCGAGAACGAGG-3’ R 5’-GGTCCACGATGGCATTGAT-3’ 170 F 5’-GTGCCAGCAAGATCCAATCTAGAGGGTGCAGAAACAGCTGGAAG-3’
R 5’-5Phos-TTATCAATGCCATCGTGGACCCCACTCCAACCCTTAGGGAACCC-3’ 88
PODXL F 5’-6FAM-CATCATTCCTGCTCCTCGTG-3’ R 5’-AAGAGGTCTCCATCACTTCCAG-3’ 150 F 5’-GTGCCAGCAAGATCCAATCTAGACTCTCCCAGAGGAAGGACCAG-3’
R 5’-5Phos-CAGCGGCTAACAGAGGAGCTCCAACCCTTAGGGAACCC-3’ 82
RRAD F 5’-6FAM-ATCTTCGGCGGTGTGGAG-3’ R 5’-GCTGCCCTTGTCCGTCACT-3’ 198 F 5’-GTGCCAGCAAGATCCAATCTAGACATCGCCTGAAGCAGAGGCAGCAG-3’
R 5’-5Phos-GGCACACCTATGATCGCTCCATTGCATCTCCAACCCTTAGGGAACCC-3’ 94
GLS2 F 5’-6FAM–AGGACAGGTGGGGCAACATT-3’ R 5’-GCTTTTTCTTGAGCAGGGGC-3’ 190 F 5’-GTGCCAGCAAGATCCAATCTAGAGCAAAGTGAATCCTTTTGCCAAGGACAG-3’
R 5’-5Phos-GTGGGGCAACATTCCCCTGGCATCATCATCATCTCCAACCCTTAGGGAACCC-3’ 103
CABYR F 5’-6FAM–TTCAGGAAGCACAGGGATGG-3’ R 5’-TGGGAAAGCAACAGAAAGGA-3’ 212 F 5’-GTGCCAGCAAGATCCAATCTAGACACAAACGTCGCAAAGCAGAAACTGAAAACTG-3’
R 5’-5Phos- ATCCAGAAATGACGCTGTCTGGGCATCATCATCATTCCAACCCTTAGGGAACCC-3’ 109
TP53I3 F 5’-6FAM–ATGGTCTGATGGGAGGAGGT-3’ R 5’-TGGATTTCGGTCACTGGGTA-3’ 220 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCATCCGCCTTCCAGCTGTTACATCTTGTGG-3’
R 5’-5Phos- GAAATGTTCAGGCTGGAGACTATGTGCTAATCATCATCATTCCAACCCTTAGGGAACCC-3’ 118
EPS8L2 F 5’-6FAM–CCAGCACCTGGCCAC-3’ R 5’-GCTCCTCCTGTGACTCGAT-3’ 173 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCCCTCGCAGTACCACGTCCAG-3’
R 5’-5Phos-CACCTGGCCACATTCATCATGGACATCATCATCCAACCCTTAGGGAACCC-3’ 100
SULF2 F 5’-6FAM–CTCCAAGGATTACCTCACAGAC-3’ R 5’-GGCGTGATGTGCTGAGA-3’ 183 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCAGTGGACGACTCCATGGAGACG-3’
R 5’-5Phos-ATTTACAACATGCTGGTTGAGACGGGCATCATCATCCAACCCTTAGGGAACCC-3’ 105
SESN1 F 5’-6FAM–CTTCTCTTCAGATGATGAAGAAGTTAC-3’ R 5’-AATAGTCCTGGACACGAAATGTT-3’ 131 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCAGATGCCGGTCAACTCAGCAGAAAATGTTTCT-3’
R 5’-5Phos-GTAAGTGATTCTTTCTTTGAGGTTGAAGCCCTCATGCATCATCTCCAACCCTTAGGGAACCC-3’ 124
FHL2 F 5’-6FAM–AAGCCCATCACCACGG-3’ R 5’-ACTTCTTGGCATACAAGTCACAG-3’ 163 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCATCATCCAGGAATGCAAGAAGACCATCATGCCAG-3’
R 5’-5Phos-GTACCCGCAAGATGGAGTACAAGGGCATCATCATCATCATCATCTCCAACCCTTAGGGAACCC-3’ 127
TBP F 5’-GCACCACTCCACTGTATCCCT-3’ R 5’-6FAM–TTTAGGTCAAGTTTACAACCAAGATT-3’ 142 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCCCCGAAACGCCGAATATAATCCCAAG-3’
R 5’-5Phos-CGGTTTGCTGCGGTAATCATGAGGATAAGCATCATCATCCAACCCTTAGGGAACCC-3’ 112
RIC8B F 5’-6FAM–AAGGTGCATAAAGAGAGTGATTCT-3’ R 5’-GACTGCATTGCTGTGTAGCTC-3’ 121 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATGACGGTAGACAGTTGGAAGGTGCATAAAGAG-3’
R 5’-5Phos-AGTGATTCTCATCAGTTCCGTGTAATGGCAGCATCATCATCTCCAACCCTTAGGGAACCC-3’ 120
MPP5 F 5’-6FAM–AATGATGATTATGACAACGAGGA-3’ R 5’-TACTCCGGGTTGTATGAGGA-3’ 190 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCGCTTTGCATCTGCAGTTCCTC-3’
R 5’-5Phos-ATACAACCCGGAGTAGGCGAGACATCATCCAACCCTTAGGGAACCC-3’ 97
PLK1 F 5’-6FAM–AGTCCTCAATAAAGGCTTGGA-3’ R 5’-AGCCTCCTCTTGCCTGAC-3’ 178 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCATCATCATCATCATCACGTGGGCTGGTCAGGCAAG-3’
R 5’-5Phos-AGGAGGCTGAGGATCCTGCCATCATCATCATCATCATCATCATCATTCCAACCCTTAGGGAACCC-3’ 130
TP53 ex5-6
F 5’-6FAM–CTGGCCAAGACCTGCC-3’ R 5’-GAAAAGTGTTTCTGTCATCCAAATAC-3’ 229 F 5’-GTGCCAGCAAGATCCAATCTAGAGAGCGCTGCTCAGATAGCGATG-3’
R 5’-5Phos-GTCTGGCCCCTCCTCAGCATCTCCAACCCTTAGGGAACCC-3’ 85
TP53 ex7-9 e
F 5’-6FAM–TGGAAGACTCCAGTGGTAATCTAC-3’ R 5’-GCAGTGCTCGCTTAGTGC-3’ 156 F 5’-GTGCCAGCAAGATCCAATCTAGACATCATCTCACCATCATCACACTGGAAGACTCCAG-3’
R 5’-5Phos-TGGTAATCTACTGGGACGGAACAGCTTTCATCATCATCTCCAACCCTTAGGGAACCC-3’ 115 a F and R indicate forward and reverse primers, respectively. b Fluorescent 6-FAM-labeled primers are indicated. c Probes with a 5’phosphate are indicated (5Phos). d The 5’and 3’ tails in bold correspond to primers U1 (TCCAACCCTTAGGGAACCC) and U2 (GTGCCAGCAAGATCCAATCTAGA) used for the final PCR amplification step. Spacers (underscored) were added between these tails and the gene-specific regions to allow the separation and identification of the PCR products based on size. e For RT-MLPA, probes are targeting exons 7 and 8.
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S

Supplementary table 2 Results of the new version of the p53 functional assay on 34 lymphoblastoid cell lines derived from control individuals or individuals with germline TP53 variants and comparison with the primary version of the assay
TP53 varianta Clinical presentationb
Primary version of the p53 functional
assay, (Zerdoumi et
al., 2017)
New version of the p53 functional assay
RT-QMPSF RT-QMPSF RT-MLPA Mean values p53
score mRNA
(%) p53
score mRNA
(%) p53
score mRNA
(%) p53
score mRNA
(%) Individuals with wild-type TP53 genotype
Control 1 Unaffected 13.1 100 16.7 100 10.4 100 13.6 100 Control 2 Unaffected 11.0 79 11.1 84 10.5 105 10.8 95 Control 3 Unaffected 14.6 100 12.0 79 10.9 97 11.5 88
Individuals with dominant-negative TP53 missense variants c.524G>A, p.(Arg175His)
RMS (1) 4.1
86 6.4 107 5.1 102 5.8 105
c.535C>T, p.(His179Tyr)
Glioblastoma (14), Osteosarcoma (16)
3.8 78 3.8 97 3.2 95 3.5 96
c.638G>A, p.(Arg213Gln)
Choroïd plexus carcinoma (0.3) 4.1 98 5.1 101 6.1 95 5.6 98
c.736A>G, p.(Met246Val)
Bilat. breast cancer (25) 2.7 90 2.9 99 3.2 95 3.1 97
c.742C>T, p.(Arg248Trp)
Breast cancer (29, 34) 3.2 73 2.4 89 3.1 98 2.8 94
c.743G>A, p.(Arg248Gln)
Osteosarcoma (16), Breast cancer (28)
2.2 76 2.1 89 1.9 92 2.0 91
c.818G>A, p.(Arg273His)
Choriocarcinoma (17, 19) 1.9 104 2.9 75 2.7 92 2.8 84
c.844C>T, p.(Arg282Trp)
Unaffected 4.6 76 5.7 80 5.3 85 5.5 83
Individuals with null TP53 variants c.(?_-202)_(*1207_?)del, p.0
Unaffected 7.2 57 6.0 38 8.1 59 7.1 49
c.(?_-202)_(29+1_-28+1)del, p.?
Unaffected 5.4 53 5.9 51 5.7 65 5.8 58
c.216dup, p.(Val73Argfs*76)
Breast cancer (28) 6.5 44 5.8 35 4.9 55 5.4 45
c.323_329dup, p.(Leu111Phefs*40)
Breast cancer (27) 6.9 59 5.3 59 6.6 73 6.0 66
c.455del, p.(Pro152Argfs*18)
Breast cancer (26, 29) 4.7 63 4.3 54 3.7 76 4.0 65
c.491_494del, p.(Lys164Serfs*5)
Osteosarcoma (11) 8.6 50 9.1 51 8.0 68 8.6 60
c.632_641del, p.(Thr211Ilefs*33)
Bilat. breast cancer (25) 5.7 50 5.4 54 4.4 77 4.9 66
c.673-2A>G, p. ?
Unaffected 5.5 42 5.6 40 4.2 61 4.9 51
c.690del, p.(Thr231Profs*16)
Unaffected 9.4 52 10.4 53 11.1 68 10.8 61
c.820del p.(Val274Phefs*71)
ACC (11) 7.0 53 7.1 41 6.2 71 6.7 56
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S

Individuals with other TP53 missense variants c.31G>C, p.(Glu11Gln)
Neuroblastoma (2) 3.0 120 4.9 75 5.8 97 5.4 86
c.323G>A, p.(Gly108Asp)
Unaffected 4.2 81 5.2 81 4.5 100 4.9 91
c.523C>G, p.(Arg175Gly)
Osteosarcoma (18), Breast cancer (27), Lung cancer (43)
3.2 101 3.6 83 3.4 82 3.5 83
c.577C>G, p.(His193Asp)
Liposarcoma (24), Breast cancer (26)
3.7 91 4.0 109 4.1 88 4.1 99
c.578A>C, p.(His193Pro)
Leiomyosarcoma (49), ACC (54), Melanoma (55)
3.7 109 3.7 104 4.6 104 4.2 104
c.646G>A, p.(Val216Met)
RMS (3), Sarcoma (17) 4.3 94 4.3 93 4.4 98 4.4 96
c.685T>C, p.(Cys229Arg)
Breast cancer (34) 4.3 82 5.3 110 4.7 99 5.0 105
c.761T>C, p.(Ile254Thr)
Unaffected 2.8 72 2.6 65 3.6 88 3.1 77
c.845G>C, p.(Arg282Pro)
Choroid plexus carcinoma (17), Sarcoma (24)
4.7 105 3.7 98 3.3 98 3.5 98
c.869G>A, p.(Arg290His)
Breast cancer (47) 10.9 85 8.6 91 6.8 106 7.7 99
c.904G>A, p.(Gly302Arg)
Breast cancer (41) 10.0 71 8.1 61 6.1 76 7.1 69
c.910A>G, p.(Thr304Ala)
Teratocarcinoma (28) 8.3 78 7.1 69 6.5 90 6.8 80
c.1010G>A, p.(Arg337His)
ACC (2) 5.8 81 6.7 84 6.0 80 6.4 82
aFor each individual, the TP53 variant is indicated: cDNA numbering is given with the first nucleotide corresponding to the A of the ATG translation initiation codon in the reference sequence (GenBank RefSeq-file accession number NM_000546.5) and protein numbering with the initiation codon numbered as codon 1 (accession number NP_000537.3).bAge at diagnosis is indicated in years, if not specified. ACC: Adrenocortical carcinoma; Bilat.: bilateral; RMS: rhabdomyosarcoma.
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S

Supplementary table 3 Results of the blood p53 functional assay in individuals with wild-type TP53 genotype or with germline TP53 variants
Individual (sex, age)
TP53 variationa
Index case / relative Clinical presentationb
Blood p53 functional assay RT-QMPSF RT-MLPA Mean values
p53 score
mRNA (%)
p53 score
mRNA (%)
p53 score
mRNA (%)
Individuals with wild-type TP53 genotype 1. (F, 45) Index case Bilat. breast cancer (30, 45) 13.9 89 11.1 79 12.5 84 2. (F, 32) Index case ACC (31) 10.5 84 10.6 100 10.6 92 3. (F, 31) Index case Glioblastoma (31) 13.8 99 10.4 92 12.1 96 4. (F, 34) Index case Breast cancer (34) 9.8 94 8.0 80 8.9 87 5. (M, 17) Relative Unaffected 11.0 110 10.2 114 10.6 112 6. (F, 61) Relative Unaffected 15.5 81 13.7 90 14.6 86 7. (F, 2) Index case ACC (2) 12.8 126 12.8 108 12.8 117 8. (F, 46) Index case Breast cancer (34) 8.2 114 7.3 85 7.8 100 9. (F, 47) Index case RMS (12), Breast cancer (47) 9.8 81 10.2 83 10.0 82 10. (F, 29) Index case Breast cancer (27) 12.1 90 12.1 100 12.1 95 11. (H, 63) Relative Unaffected 8.4 81 6.5 87 7.5 84 12. (F, 36) Index case Breast cancer (34) 11.7 81 8.9 84 10.3 83 13. (F, 44) Index case Lung cancer (30,43) 16.6 70 11.5 77 14.1 74 14. (F, 31) Index case Lung cancer (31) 22.5 95 10.5 95 16.5 95 15. (M, 8) Relative Unaffected 12.7 82 16.5 101 14.6 92 16. (F, 16) Index case Lymphoma (7), CNS sarcoma (16) 31.4 111 14.1 110 22.8 111 17. (M, 41) Index case Liposarcoma (31) 13.6 88 12.4 89 13.0 89 18. (M, 14) Relative Unaffected 18.2 94 12.6 96 15.4 95 19. (F, 66) Index case Breast cancer (48), Liposarcoma (65) 9.7 74 7.0 81 8.4 78 20. (F, 34) Index case Breast cancer (33) 12.1 78 13.9 93 13.0 86 21. (F, 36) Index case Breast cancer (34) 11.5 99 10.0 97 10.8 98 22. (F, 31) Index case Breast cancer (31) 14.9 100 10.9 87 12.9 94 23. (F, 68) Relative Unaffected 9.4 73 9.3 85 9.4 79 24. (F, 31) Index case Breast cancer (31) 13.5 64 9.9 92 11.7 78 25. (M, 18) Relative Unaffected 10.1 89 12 105 11.1 97 26. (M,19) Relative Unaffected 11 90 8.9 78 10.0 84 27. (F, 57) Relative Head Neck cancer (52) 18.2 131 11.2 113 14.7 122 28 (F, 71) Relative Lung cancer (71) 10.8 94 19.4 106 15.1 100 29. (F, 34) Index case Breast cancer (34) 21.0 81 15.1 96 18.1 89 30. (F, 61) Index case Breast cancer (35) 9.4 91 6.6 114 8.0 103 31. (F, 4) Index case RMS (8 months) 16.2 118 12.1 113 14.2 116 32. (F, 72) Relative Unaffected 9.0 94 7.6 85 8.3 90 33. (F, 46) Relative Unaffected 10.3 87 13.6 83 12.0 85 34. (M, 14) Index case Medulloblastoma (4) 12.0 89 11.4 90 11.7 90 35. (F, 20) Index case Colon cancer (19) 17.3 81 17.4 79 17.4 80 36. (M, 16) Index case Sarcoma (15) 10.6 89 11.5 115 11.1 102 37. (M, 13) Index case RMS (4) 20.8 81 18.6 91 19.7 86 38. (F, 28) Index case Breast cancer (28) 17.0 96 17.3 85 17.2 91 39. (F, 29) Index case Breast cancer (28) 10.1 78 12.1 89 11.1 84 40. (F, 24) Index case ACC (24) 9.6 77 11.1 80 10.4 79 41. (F, 30) Index case Breast cancer (30) 13.1 87 17.1 88 15.1 88 42. (F, 39) Index case Breast cancer (31) 8.3 71 9.0 78 8.7 75 43. (F, 33) Index case Breast cancer (33) 9.0 75 9.1 92 9.1 84 44. (F, 33) Index case Breast cancer (31) 11.7 115 7.8 114 9.8 115 45. (F, 36) Index case Breast cancer (29) 20.4 114 18.1 135 19.3 125 46. (F, 47) Index case Breast cancer (25) 18.4 105 12.4 93 15.4 99
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S
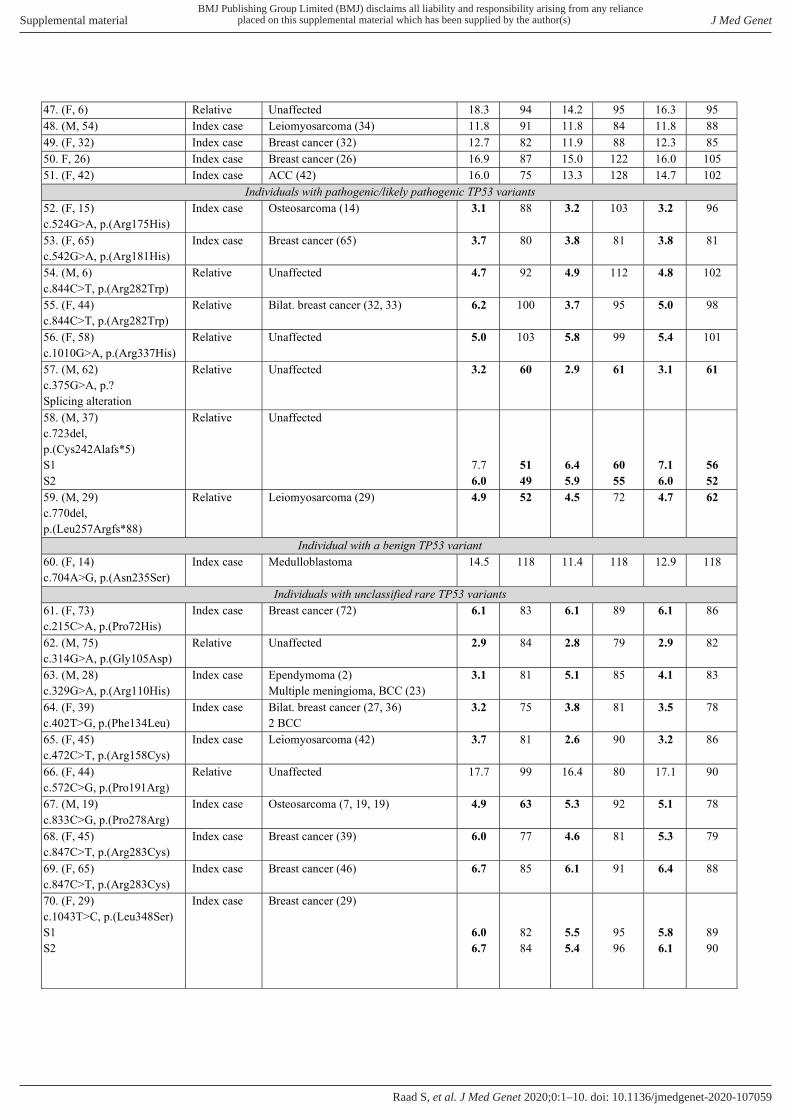
47. (F, 6) Relative Unaffected 18.3 94 14.2 95 16.3 95 48. (M, 54) Index case Leiomyosarcoma (34) 11.8 91 11.8 84 11.8 88 49. (F, 32) Index case Breast cancer (32) 12.7 82 11.9 88 12.3 85 50. F, 26) Index case Breast cancer (26) 16.9 87 15.0 122 16.0 105 51. (F, 42) Index case ACC (42) 16.0 75 13.3 128 14.7 102
Individuals with pathogenic/likely pathogenic TP53 variants 52. (F, 15) c.524G>A, p.(Arg175His)
Index case Osteosarcoma (14) 3.1 88 3.2 103 3.2 96
53. (F, 65) c.542G>A, p.(Arg181His)
Index case Breast cancer (65) 3.7 80 3.8 81 3.8 81
54. (M, 6) c.844C>T, p.(Arg282Trp)
Relative Unaffected 4.7 92 4.9 112 4.8 102
55. (F, 44) c.844C>T, p.(Arg282Trp)
Relative Bilat. breast cancer (32, 33) 6.2 100 3.7 95 5.0 98
56. (F, 58) c.1010G>A, p.(Arg337His)
Relative Unaffected 5.0 103 5.8 99 5.4 101
57. (M, 62) c.375G>A, p.? Splicing alteration
Relative Unaffected 3.2 60 2.9 61 3.1 61
58. (M, 37) c.723del, p.(Cys242Alafs*5) S1 S2
Relative Unaffected
7.7 6.0
51 49
6.4 5.9
60 55
7.1 6.0
56 52
59. (M, 29) c.770del, p.(Leu257Argfs*88)
Relative Leiomyosarcoma (29) 4.9 52 4.5 72 4.7 62
Individual with a benign TP53 variant 60. (F, 14) c.704A>G, p.(Asn235Ser)
Index case Medulloblastoma 14.5 118 11.4 118 12.9 118
Individuals with unclassified rare TP53 variants 61. (F, 73) c.215C>A, p.(Pro72His)
Index case Breast cancer (72) 6.1 83 6.1 89 6.1 86
62. (M, 75) c.314G>A, p.(Gly105Asp)
Relative Unaffected 2.9 84 2.8 79 2.9 82
63. (M, 28) c.329G>A, p.(Arg110His)
Index case Ependymoma (2) Multiple meningioma, BCC (23)
3.1 81 5.1 85 4.1 83
64. (F, 39) c.402T>G, p.(Phe134Leu)
Index case Bilat. breast cancer (27, 36) 2 BCC
3.2 75 3.8 81 3.5 78
65. (F, 45) c.472C>T, p.(Arg158Cys)
Index case Leiomyosarcoma (42) 3.7 81 2.6 90 3.2 86
66. (F, 44) c.572C>G, p.(Pro191Arg)
Relative Unaffected 17.7 99 16.4 80 17.1 90
67. (M, 19) c.833C>G, p.(Pro278Arg)
Index case Osteosarcoma (7, 19, 19) 4.9 63 5.3 92 5.1 78
68. (F, 45) c.847C>T, p.(Arg283Cys)
Index case Breast cancer (39) 6.0 77 4.6 81 5.3 79
69. (F, 65) c.847C>T, p.(Arg283Cys)
Index case Breast cancer (46) 6.7 85 6.1 91 6.4 88
70. (F, 29) c.1043T>C, p.(Leu348Ser) S1 S2
Index case Breast cancer (29)
6.0 6.7
82 84
5.5 5.4
95 96
5.8 6.1
89 90
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S

71. (F, 15) c.1054G>T, p.(Asp352Tyr) S1 S2
Relative Unaffected
3.5 4.9
110 103
4.5 5.0
92 108
4.0 5.0
101 106
72. (H, 31) c.323_235del p.(Gly108_Phe109delinsVal)
Index case Osteosarcoma (31) 2.9 88 4.7 100 3.8 94
73. (F, 22) c.393_395del, p.(Asn131del) S1 S2
Relative
Unaffected
2.2 4.3
81 97
3.0 5.2
78 100
2.6 4.8
80 99
74. (F, 22) c.792_794del, p.(Leu265del)
Index case Breast cancer (31) 3.6 98 3.2 91 3.4 95
75. (F, 32) c.-117G>T (5'UTR)
Index case Breast cancer (32) 23.7 102 17.3 97 20.5 100
Individual with the homozygous c.*1175A>C polymorphic variant
76. (F, 35) c.*1175A>C (homozygous)
S1 S2
Relative Unaffected
5.1 6.1
61 67
5.8 5.2
76 78
5.5 5.7
69 73
Individual with a pathogenic TP53 variant and the c.*1175A>C polymorphic variant 77. (F, 6) c.723del p.(Cys242Alafs*5) c.*1175A>C (heterozygous)
Index case Glioma (5) 2.4 47 3.8 56 3.1 52
aFor each individual, the identification number is indicated. For some individuals, two blood samples (S1 and S2) were collected. In brackets are indicated the sex (M, male; F, female) and age (in years) at blood sampling. TP53 variation is indicated for carriers: cDNA numbering is given with the first nucleotide corresponding to the A of the ATG translation initiation codon in the reference sequence (GenBank RefSeq-file accession number NM_000546.5) and protein numbering with the initiation codon numbered as codon 1 (accession number NP_000537.3).bAge at diagnosis is indicated in years, if not specified. For this individual, a second TP53 variant was identified in cis: c.1021T>A, p.(Phe341Ile), which is classified as benign. ACC: Adrenocortical carcinoma; BCC: basocellular carcinoma; Bilat.: bilateral; CNS: central nervous system; RMS: rhabdomyosarcoma. Results in bold are suggestive of a deleterious effect of the variant.
BMJ Publishing Group Limited (BMJ) disclaims all liability and responsibility arising from any relianceSupplemental material placed on this supplemental material which has been supplied by the author(s) J Med Genet
doi: 10.1136/jmedgenet-2020-107059–10.:10 2020;J Med Genet, et al. Raad S