biologic and biochemic properties of recombinant platelet - blood
TRANSCRIPT

Biologic and Biochemic Properties of Recombinant Platelet Factor 4 Demonstrate Identity With the Native Protein
By Kwang Sook Park, Salahaldin Rifat, Howard Eck, Kazuhiko Adachi, Saul Surrey, and Mortimer Poncz
Platelet factor 4 (PF4) is a 70 amino acid protein released from the a-granules of platelets after activation. The exact biologic function of this protein is unknown. We have constructed an expression vector for recombinant PF4 (rPF4) in the T7-based promoter vector pT7-7 to better study the relationship between PF4 structure and function. The protein was expressed in Escherichia coli and purified to homogeneity by heparin-agarose affinity chromatogra- phy and reverse-phase high-performance liquid chromatog- raphy. Purity of protein was confirmed by immunoblot analysis and sodium dodecyl sulfate-polyacrylamide gel electrophoresis, which resulted in a single component with a molecular weight of 8,000 daltons. The amino acid composition and sequence of the N-terminal 20 residues showed that rPF4 is identical to PF4 prepared from human platelets (hPF4). except for an N-terminal initiating methio-
LATELET FACTOR 4 (PF4) is synthesized by bone P marrow megakaryocytes and stored in a-granules’.2 as a noncovalent-bound tetramer composed of identical 7,800 dalton monomers of 70 amino acids.’” PF4 has been purified6-” and is a heat-stable protein that is insoluble a t low ionic strength, but soluble in solutions of high ionic strength or a ~ i d i t y . ~ . ~ The complete amino acid sequence of human platelet factor 4 (hPF4) has been determined.’” A number of biologic features have been noted for this protein. PF4 binds to heparin with high affinity.6 This property allowed initial purification of PF4 from platelets, and may reflect its key biologic role. By neutralizing heparin, PF4 may inhibit heparin-induced anti-thrombin I11 activation and may per- mit platelets to bind to endothelial cells, thus supporting the early phases of coagulation."^" The heparin-binding activity appears to reside in the lysine-rich carboxyl-terminal domain of the molecule.“
PF4 also appears to have a number of other properties in vitro whose biologic significance is unknown. Since some of these properties require fairly high concentrations of PF4, it is also possible that some of these activities may be due to contaminating proteins. One of the more noted properties of PF4 is its potency as a chemoattractant for neutrophils,
From the Divisions of Hematology and Immunology. The Chil- dren’s Hospital of Philadelphia; and the Departments of Pediatrics and Human Genetics, the University of Pennsylvania. Philadelphia, PA.
Submitted August 24,1989; accepted November 24,1989. Supported in part by Grants No. HL37419. HL40387, and
HL281.57 from the National Institutes of Health, and the March of Dimes (Basic Research Grant to M.P.).
Address reprint requests to Mortimer Poncz, MD. Division of Hematology, The Children’s Hospital of Philadelphia, 34th St and Civic Center Blvd, Philadelphia, PA 19104.
The publication costs ofthis article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 8 1990 by The American Society of Hematology. ~6-4971/90/7506-0012$3.00/0
nine residue. lmmunoblots revealed that rPF4 and hPF4 bound polyclonal anti-hPF4 equally well, while chemotaxis experiments demonstrated similar potencies as neutrophil attractants. Dose-dependent neutrophil chemotactic re- sponses and competitive studies with polyclonal anti-hPF4 antiserum further demonstrate similar chemotactic proper- ties of t h e two PF4 species. In conclusion, our data show that this recombinant protein and t h e native protein appears to have similar immunologic, heparin-binding, and chemotactic properties. The chemotactic properties of hPF4 appear to be entirely intrinsic to the protein and not due, in part, to any contaminating protein. Furthermore, our expression vector should prove useful for t h e construc- tion of recombinant forms of PF4 to investigate structure/ function relationships of this biologically important protein. 0 1990 by The American Society of Hematology.
monocytes, and fibroblast^.'^-'^ This property appears to be related to the carboxyl-terminal domain of PF4.I6 It has also been demonstrated that hPF4 has immunoregulatory activity.’9s20 Recently, PF4 has been shown to behave as a negative autocrine in regulating megakaryocyte differentia- tion.”
PF4 is a member of a multigene family, the small inducible genes (SIC), which appear to be important in coagulation, inflammation, and cell g r o ~ t h . ~ ~ - ~ ~ Members of this family are either induced or released after external stimulation. The cloned SIC proteins share amino acid homology and contain four highly-conserved cysteine residues. In addition, mem- bers of this. family have a similar genomic exon organization, suggesting that they were derived from a common ancestral gene.
Other members of the SIC family include platelet P- thromboglobulin (BTG) and its precursors, which are also contained in platelet a-granules and released during platelet a c t i v a t i ~ n . ~ ~ ~ ~ ~ PTG has a glutamine residue immediately preceding the second dilysine repeat unit, which may explain why it binds heparin with less affinity than PF4. MGSA or GRO is secreted by melanoma cells and is another member of this family with potent mitogenic properties.25 A number of S I C proteins are synthesized in various cell lines after stimulation with an appropriate inducer: y I P - 1 0 is induced in the human monocyte cell line U937 and other cell lines after stimulation with gamma interferonz6; JE is synthesized in fibroblasts and smooth muscle cells that have been exposed to platelet-derived growth factorz7; 3- 1OC is induced by staphylococcal enterotoxin in peripheral human lympho- cytesZ8; and LD78 is induced in human tonsillar lymphocytes after phytohemagglutinin or 12-0-tetradecanoylphorbol- 1 3- acetate stim~lation.’~
Recently, a recombinant PF4 (rPF4) protein was de- scribed that has an additional seven amino acids a t the N terminus, four additional amino acid residues at the C terminus, and an Asn to Asp substitution a t position 47 of the mature protein.)’ While this recombinant protein demon- strated biologic activity in an immunoregulatory assay, it is less than ideal for further structural/functional studies. The
1290 Blood, Vol75, No 6 (March 15). 1990: pp 1290-1295
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom

RECOMBINANT PLATELET FACTOR 4 1291
presence of differences at both termini and near the center of this small protein would make it difficult to correlate struc- tural changes with changes in biologic function. We report here the expression of rPF4 in Escherichia coli using a T7- promoter-based vector system. Except for a retained initiat- ing methionine residue, the recombinant protein is identical to hPF4. Chemotactic activity for neutrophils by the newly constructed rPF4 molecule is the same as the native protein.
MATERIALS AND METHODS
Heparin-agarose, hPF4, and E-toxate (Limulus Ame- bocyte lysate) were obtained from Sigma, St Louis, MO. Vectastain ABC kit was from Vector Laboratory Inc, Burlingame, CA. Anti- hPF4 was from Atlantic Antibodies, Scarborough, ME. Polyvinyli- denedifluoride (PVDF) transfer membrane (Immobilon) was from Millipore, Bedford, MA. Restriction endonucleases and other en- zymes were obtained from Bethesda Research Laboratory, Be- thesda, MD.
The rPF4 expression vector is shown diagrammatically in Fig 1. This vector was con- structed from the pT7-7 expression vector’’ (kindly provided by Drs Tabor and Richardson, Massachusetts Institute of Technology, Boston), which had been linearized by double digestion with Nde I and EcoRI. The linearized vector was ligated with equal molar amounts of the 33 1 bp Psi IIEcoRI PF4 cDNA fragment previously described,” and the two phosphorylated oligonucleotides, PI (5’- dTATGGAAGCTGAAGAAGATGGGGACCTGCA-3’) and P2 (5’-dGGTCCCCATCTTCTTCAGCTTCCA-3’), which supply the missing seven amino acids of the mature PF4 protein and the
Materials.
Construction of rPF4 expression vector.
Amp‘
Fig 1. Diagram of the rPF4 expression vector in PT7-7. The PF4 cDNA region incorporated into’the construct is shown as the stripped region between the Pot I and EcoRl sites. The T7 promoter, ribosomal binding sites (RES), N d e I site, and ampicillin resistant (Amp‘) regions are shown. The stippled region between the N d e I and Pst I sites was filled in using two dgonucleotides, P1 and P2.
initiating methionine residue. They also generate an Nde I compati- ble sequence at the 5’ end and P Pst I sequence at the 3‘ end. Correct rPF4/pT7-7 construction was verified by appropriate enzyme diges- tions using a combination of Nde I, Psi I , and EcoRI, and by sizing the resulting fragments on an agarose gel.
Expression and purification of rPF4. rPF4 was expressed in E coli by a modification of a previously described techniq~e.”.’~ E coli K38” transformed with plasmid pGpl-2 and pT7-7” was inoculated into a 10 mL aliquot of enriched medium (2% [wt/vol] tryptone, 1% [wt/vol] yeast extract, 0.2% [VOl/VQl] glycerol, 0.5% [wt/vol] NaCl, 40 pg/mL ampicillin, and 50 pg/mL kanamycin). The culture was incubated overnight at 3OoC, and 0.5 mL was then inoculated into 500 mL of enriched medium at 3OOC. At approximately OD,, = 1.3, the T7 RNA polymerase activity was induced by raising the temperature to 42OC for 30 minutes, after which 2.5 mL of a rifampicin solution (20 mg/mL in methanol) was added to.inhibit bacterial RNA polymerase activity. The culture was grown for an additional 60 minutes at 37OC. E coli was harvested by centrifuga- tion at 5,OOQg for 10 minutes at 4OC in a Sorvall GSA rotor (Dupont, Wilmington, DE). Cell pellets were washed with TED (50 mmol/L Tris-CI, pH 8.0; 1 mmol/L EDTA, 1 mmol/L DTT), repelleted, resuspended, and incubated for 30 minutes in %O of the original culture volume in TED to which 0.1 mg/mL of lysozyme was added. NaCl was then added to a final concentration of 0.4 mol/L, and the lysate was sonicated at 4OC three times with a Branson Sonifier (Branson Ultrasonics, Danbury, CT) using a microtip at a power level of 6 for 1 minute each. The sample was centrifuged at 100,OOOg for 30 minutes in a Beckman Type Ti 75 rotor (Fullerton, CA), and the supernatant was collected, mixed with 5 mL of packed heparin- agarose, and rocked at 4OC overnight. The next day, a column was poured with the agarose and washed with TED buffer containing 0.5 mol/L NaCl until the eluate absorption at 280 nm had returned to baseline. The bound rPF4 was then eluted with TED buffer containing 1.5 mol/L NaCl.
To further purify the rPF4, the eluate was chromatographed on a Waters Associates liquid chromatography system (Millipore, Mil- ford, MA) consisting of an M450 variable wavelength detector, two model 6000 solvent delivery systems, and a p Bondpak C18 column (30 cm x 3.9 mm ID; Millipore). The rPF4 was eluted from this column with a convex gradient of 0 to 60% (vol/vol) acetonitrile, containing 0.1% (vol/vol) trifluoroacetic acid in water. Elution was performed for 40 minutes at a flow rate of 1 mL/min, and protein detection was monitored at 229 nm. The desired fractions were collected and lyophilized. Final rPF4 yield was measured with a Coomassie assay reagent (Pierce, Rockford, IL) following the manufacturer’s protocol and using a modified Bradford method.’6 Bovine serum albumin was used as a protein standard. Endotoxin contamination of rPF4 was tested with E-toxate as described by the manufacturer.
Protein sizing, sequencing, amino acid analysis, and immunoblot- ting. Approximately 10 pg of purified rPF4 was electrophoresed on a 17.5% (wt/voQ sodium dodecyl sulfate (SDS)-polyacrylamide gel (8 cm x 6 cm x 0.75 mm) as described by Laemmli.3’ After electrophoresis, the protein was blotted onto a PVDF membrane.’* The PVDF membrane was then stained with 0.1% (wt/vol) Coomassie blue R-250 in 50% methanol and destained in 50% methanol, 10% acetic acid. The membrane was then rinsed in deionized H,O, and the rPF4 baud was excised with a clean razor blade for sequence analysis. Approximately half of the initial 10 pg of rPE4 was present in the excised membrane, and the band was analyzed on an Applied Biosystems 477A pulsed liquid phase protein sequenator (Foster City, CA) equipped with an on-line 120A FTH analyzer.
Immunoblotting was performed after electrotransfer of proteins from SDS gels to a PVDF membrane at 270 mA for 15 minutes at
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom
1292 PARK ET AL
4% in CAPS buffer (IO mmol/L 3-(cyclohexylamino)-l-~ne sulfonic acid. pH I I . 10% [vol/vol] methanol). The membrane was blocked with 20% (wt/vol) fetal bovine serum in Tris-buffered saline containing 0.05% (vol/vol) Tween-20 (TRSf) for 30 minutes a t m m temperature. The blocked membrane was then incubated for I hour at room temperature with goat anti-hPF4 (Atlantic Antibody. h r b o r o u g h . ME) in TRST. After washing twice with TRST, the membrane was treated with the Vectastain ABC biotin/peroxidase." and finally developed by the addition of 4-chloto-I-naphthol in the presence of 0.01 Q (vol/vol) H,O,.
For amino acid analysis. 600 pmol of the purified rPF4 was hydrolyzed in 6N HCI for 24 hours at I IOOC in a Pierce amino acid hydrolysis tube under vacuum. After hydrolysis, the samples were lyophilyzed and washed 3 to 4 times. and the resultant amino acids were derivatized and chromatographed on an Applied Biosystems 420A amino acid analytrr. Free amino acids converted to phenol- thiocarbonyl (PTC) derivatives were separated and quantitated by microbore high-performance liquid chromatography (HPLC) on a C I8 reverse-phase (RP) column.
Neutrophils w m prepared from heparinized whole blood by Ficoll-Hypaque density gradient centrifugation. followed by dextran sedimentation and hypotonic lysis of residual erythrocytes." Purity was greater than 85% as demonstrated by light microscopy examination of Wright-Giemsa stained cells. Viability exceeded 95% as determined by trypan blue exclusion. Cells were plated at a density of 2.5 x I@/mL in Hank's balanced salt solution (HRSS) containing 0.15% (wl/vol) bovine serum albumin. The raft technique of Addison and Babage" and the leading front method were used to measure chemotactic ruponsc. A membrane filter with a mean pore sire of 2 pm (Nucleoport. Pleasanton. C h ) was laid on a flat absorbant pad (2 layers of 3 cm diameter Whatman filter paper placed in a Petri dish of similar diameter) saturated with HRSS only or 2.54; (wt/vol) zymosan-activated serum," IO " mol/L N- formylmethionyl-leucyl-phenylalanine (FMLP). I % (wt/wt) casein. or different concentrations of either rPF4 or hPF4. Plastic caps (8 mm diameter) containing 500 to 600 p L of neutrophil suspension were inverted and gently placed on the membrane. After incubation in a moist chamber at 37OC for 90 minutes. the caps were removed and discarded. the membrane was washed in phosphate-buffered saline (PRS). fixed in isopropanol. and then rehydrated and stained with Harris hematoxylin. The membranes were cleared in xylene and oil of cedar. The vmical distance traveled by the leading neutrophils into the membrane was measured and compared with
Chcmofuxb.
that in HBSS alone. w h m the filter paper raft contained no chemoattractant. Use of the latter gave a random or background migration baseline that could be subtracted from the total distance traveled by neutrophils in each experiment. The distance between the base layer and the leading cells was measured in a t least five random fields, and the mean distance migrated (in microns) was calculated. To study competitive inhibition between the two PF4 species. different aliquots of neutrophils were simultaneously prein- cubated with different concentrations of either rPF4 (0.5. 2. 5 pg/mL). hPF4 (0.5.2.5 pg/mL). 2.5% (wt/vol) zymosan-activated serum. IO * mol/L FMLP.or HBSS alone for 30 minutes a t 37% in 5% CO,. After this preincubation. the cells were washed twice with PRS and immediately tested against each of the above mentioned chemoattractants. Neutralizing studies using goat polyclonal anti- hPF4 antibody ( I 0 0 pglml.) were also used to test the specificity of the chemoattractant. Neutrophils were preincubated with anti-hPF4 antibodies at room temperature for 30 minutes. and cells were then tested in the continued presence of the antibody for chemotaxis with both rPF4 or hPF4.
RESULTS
After t h e first heparin-binding column, approximately 700 rg of partially purified PF4 was isolated f rom I L of bacterial growth (3.6 g of wet cells) using t h e expression vector system a n d purification scheme described above. Final yield of rPF4 af te r RP-HPLC was approximately 450 rg. Endotoxin contaminat ion of rPF4 tested with t h e E-toxate kit w a s est imated t o be lcss t h a n IO n g / m L (da ta not shown).
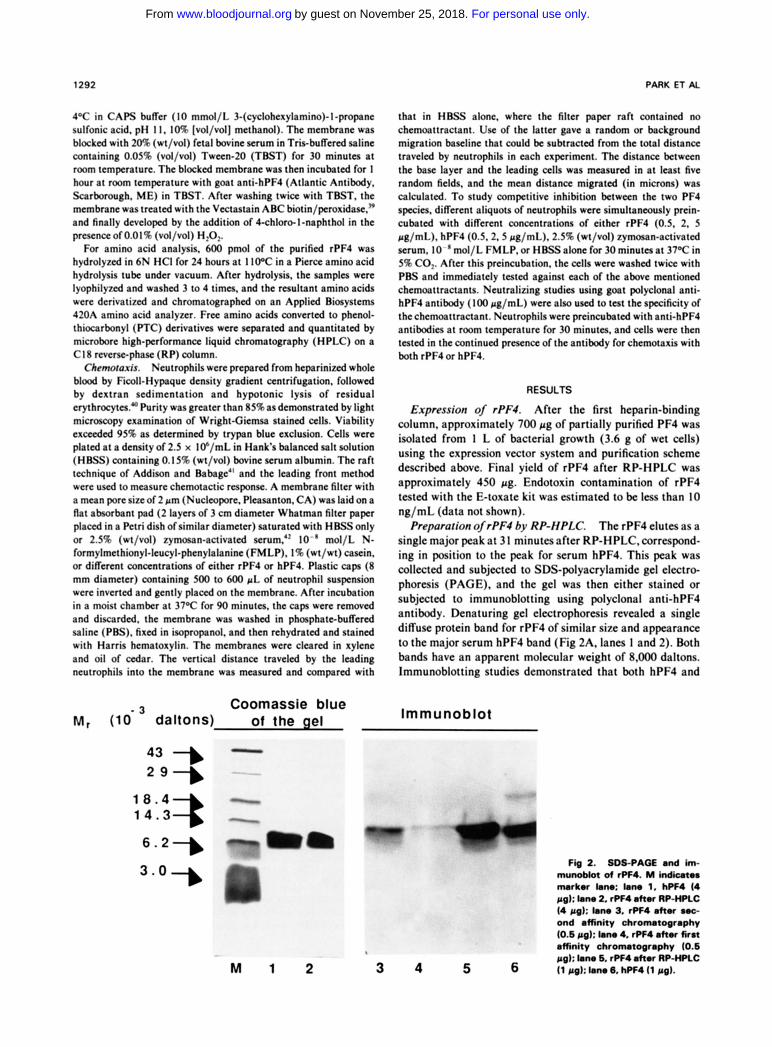
Prepororion of rPF4 by RP-HPLC. The rPF4 elutes as a single major peak a t 31 minutesaf te r RP-HPLC.correspond- ing in position t o t h e peak for s e r u m hPF4. This peak w a s collected and subjected t o SDS-polyacrylamide gel electrcF phoresis (PAGE) . and t h e gel was then ei ther s ta ined or subjected t o immunoblot t ing using polyclonal an t i -hPF4 antibody. Denatur ing gel electrophoresis revealed a single diffuse protein band for r P F 4 of similar site and appearance t o t h e major serum h P F 4 band (Fig 2A. lanes 1 a n d 2). Both bands have an apparent molecular weight of 8,000 daltons. lmmunoblot t ing s tudies demonstrated t h a t both h P F 4 and
Expression of rPF4.
.) Coomassle blue M, (IO-' daltons) of the gel lmmunoblot
6.2-
3.0- Fb2 . 8 0 8 9 A Q E n d h - munobkt of rPF4. M k d i a t r nurkor lmw km 1, hPF4 (4 ccg): lam 2, rPF4 dtu RP-HPLC (4 pa): (.no 3. rPF4 oftor uc- ond amnity chromatography (0.6 pal: Ism 4. rPF4 ahor first affinity chromatography (0.6 f ig): bm 6. rPF4 ahor RP-HPLC
M 1 2 3 4 5 6 (1 BO): Ian. 6, hPF4 (1 fig).
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom
RECOMBINANT PLATELET FACTOR 4 1293
rPF4 react with goat anti-hPF4 (Fig 2B, lanes 4 and 5). It should be noted that additional minor bands are present in the commercial hPF4 preparation, but were absent in the purified rPF4.
Protein sequence analysis and amino acid content. N-terminal amino acid sequence analysis was determined in two separate studies using 10 r g of blotted HPLC-purified rPF4. Aside from the two cysteine residues in PF4 that could not be determined by the sequencing technique used, the sequence of the first 20 N-terminal amino acid residues was clearly determined and was identical to hPF4 except for the N-terminal methionine residue in rPF4. This sequence in- cludes the seven-amino acid region added at the N-terminus of the PF4 Pst IIEcoRI cDNA insert with the two oligonucle- otides, P1 and P2, used to provide the N-terminus of the recombinant protein (Fig 1). Further confirmation of the correct composition of rPF4 was obtained from the determi- nation of the amino acid content using HPLC analysis after vacuum hydrolysis (Table 1). Within the range of variability inherent in this technique, the amino acid analysis is consis- tent with rPF4 being identical to hPF4, except for the initiating methionine residue.
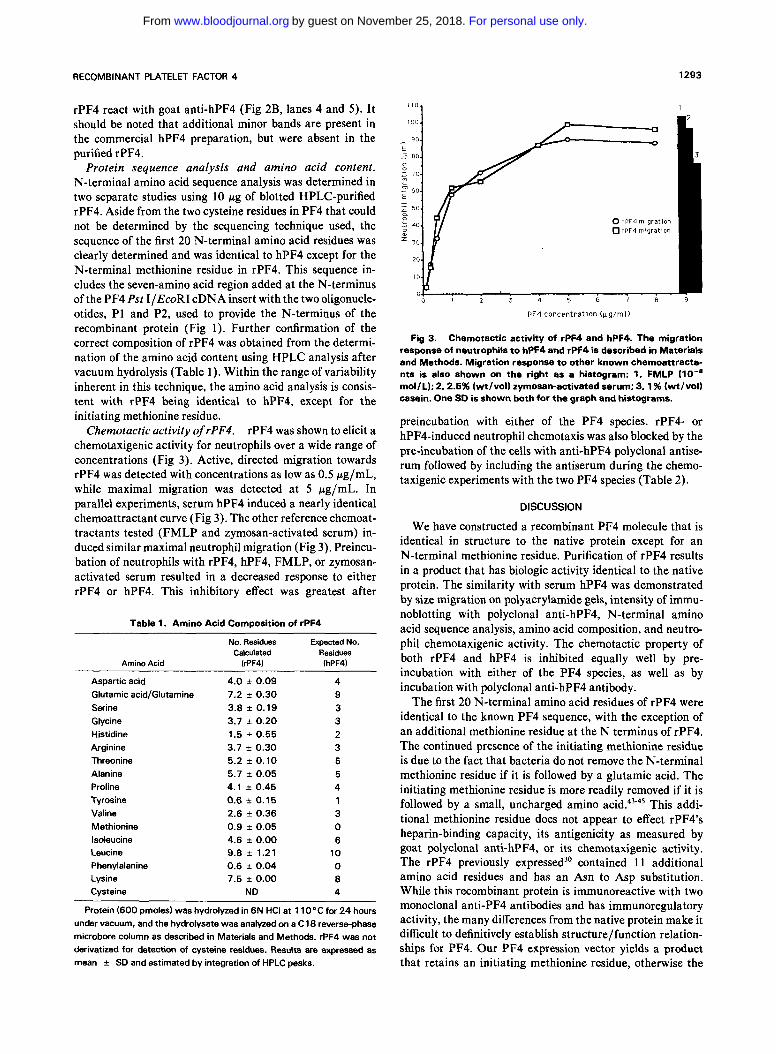
rPF4 was shown to elicit a chemotaxigenic activity for neutrophils over a wide range of concentrations (Fig 3). Active, directed migration towards rPF4 was detected with concentrations as low as 0.5 pg/mL, while maximal migration was detected a t 5 pg/mL. In parallel experiments, serum hPF4 induced a nearly identical chemoattractant curve (Fig 3). The other reference chemoat- tractants tested (FMLP and zymosan-activated serum) in- duced similar maximal neutrophil migration (Fig 3). Preincu- bation of neutrophils with rPF4, hPF4, FMLP, or zymosan- activated serum resulted in a decreased response to either rPF4 or hPF4. This inhibitory effect was greatest after
Chemotactic activity of rPF4.
Table 1. Amino Acid Composition of rPF4
No. Residues Calculated
Amino Acid (rPF4)
Aspartic acid 4.0 f 0.09 Glutamic acid/Glutamine 7.2 f 0.30 Serine 3.8 f 0.19 Glycine 3.7 f 0.20 Histidine 1.5 f 0.55 Arginine 3.7 f 0.30 Threonine 5.2 * 0.10 Alanine 5.7 f 0.05 Proline 4.1 f 0.45 Tyrosine 0.6 f 0.15 Valine 2.6 f 0.36 Methionine 0.9 f 0.05 Isoleucine 4.6 f 0.00 Leucine 9.8 & 1.21 Phenylalanine 0.6 f 0.04 Lysine 7.5 f 0.00 Cysteine ND
Expected No. Residues
(hPF4)
4 9 3 3 2 3 5 5 4 1 3 0 6
10 0 8 4
Protein (600 pmoles) was hydrolyzed in 6N HCI at 1 1 0 ° C for 24 hours under vacuum, and the hydrolysate was analyzed on a C 18 reverse-phase microbore column as described in Materials and Methods. rPF4 was not derivatized for detection of cysteine residues. Results are expressed as mean f SD and estimated by integration of HPLC peaks.
I I O 1 I
I 0 0
90 - E 2 80 c
E 70 L
? 60
50
2 40
30
20
I 0
0
- a 0
O h P F 4 migrat ion 3 aj u r P F 4 mlgrat lon
0 1 2 3 4 5 6 1 8 9
PF4 concent ra t ion (pg /m l )
Fig 3. Chemotactic activity of rPF4 and hPF4. The migration response of neutrophils to hPF4 and rPF4 is described in Materials and Methods. Migration response to other known chemoattracta- nts is also shown on the right as a histogram: 1. FMLP (lo-' mol/L); 2.2.5% (wt/vol) zymosan-activated serum; 3.1% (wt/vol) casein. One SO is shown both for the graph and histograms.
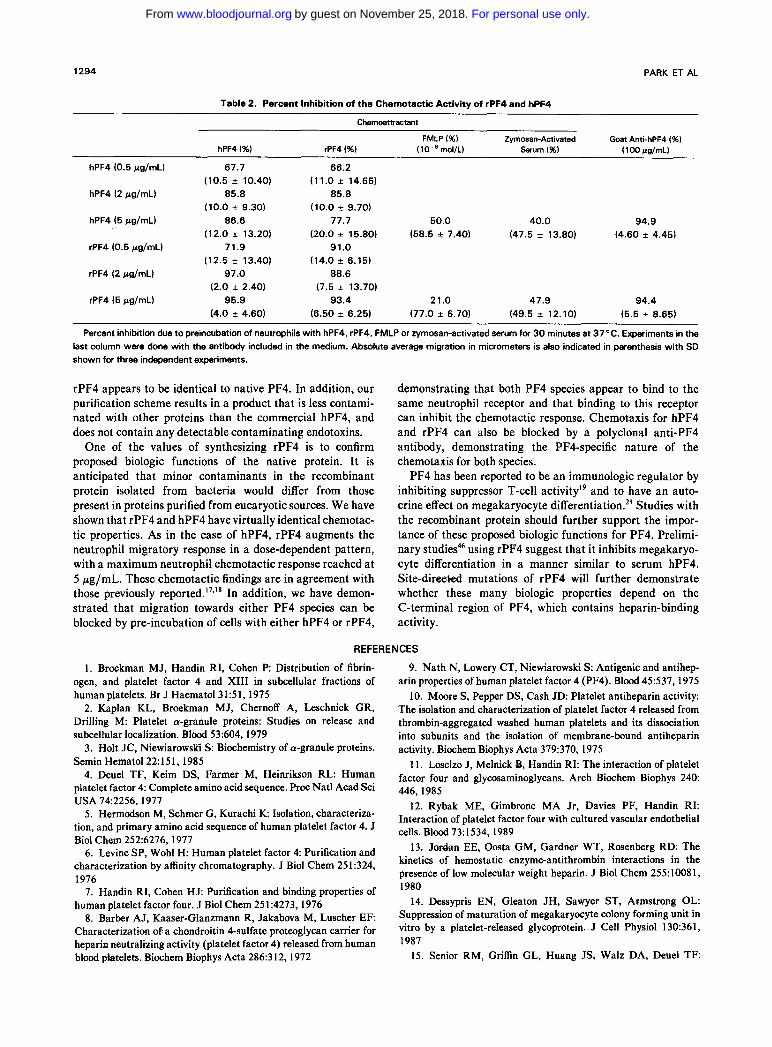
preincubation with either of the PF4 species. rPF4- or hPF4-induced neutrophil chemotaxis was also blocked by the pre-incubation of the cells with anti-hPF4 polyclonal antise- rum followed by including the antiserum during the chemo- taxigenic experiments with the two PF4 species (Table 2).
DISCUSSION
We have constructed a recombinant PF4 molecule that is identical in structure to the native protein except for an N-terminal methionine residue. Purification of rPF4 results in a product that has biologic activity identical to the native protein. The similarity with serum hPF4 was demonstrated by size migration on polyacrylamide gels, intensity of immu- noblotting with polyclonal anti-hPF4, N-terminal amino acid sequence analysis, amino acid composition, and neutro- phil chemotaxigenic activity. The chemotactic property of both rPF4 and hPF4 is inhibited equally well by pre- incubation with either of the PF4 species, as well as by incubation with polyclonal anti-hPF4 antibody.
The first 20 N-terminal amino acid residues of rPF4 were identical to the known PF4 sequence, with the exception of an additional methionine residue a t the N terminus of rPF4. The continued presence of the initiating methionine residue is due to the fact that bacteria do not remove the N-terminal methionine residue if it is followed by a glutamic acid. The initiating methionine residue is more readily removed if it is followed by a small, uncharged amino a ~ i d . ~ ~ - ~ ~ This addi- tional methionine residue does not appear to effect rPF4's heparin-binding capacity, its antigenicity as measured by goat polyclonal anti-hPF4, or its chemotaxigenic activity. The rPF4 previously expressed" contained 1 1 additional amino acid residues and has an Asn to Asp substitution. While this recombinant protein is immunoreactive with two monoclonal anti-PF4 antibodies and has immunoregulatory activity, the many differences from the native protein make it difficult to definitively establish structure/function relation- ships for PF4. Our PF4 expression vector yields a product that retains an initiating methionine residue, otherwise the
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom
1294 PARK ET AL
Table 2. Percent Inhibition of the Chemotactic Activitv of rPF4 and hPF4
Chemoamactant
FMLP (%) Zymosan-Activated Goat Anti-hPF4 (%) hPF4 (%I rPF4 (%) (lo-* mol/L) Serum (%I ( 100 pg/mL)
hPF4 (0.5 pg/mL) 67.7 66.2
hPF4 (2 pg/mL) 85.8 85.8
hPF4 (5 pg/mL) 86.6 77.7 50.0 40.0
(10.5 f 10.40)
(10.0 f 9.30)
(12.0 f 13.20) (20.0 f 15.80) (58.5 f 7.40) (47.5 f 13.80)
(12.5 f 13.40)
(2.0 f 2.40)
(4.0 f 4.60) (6.50 f 6.25) (77.0 f 6.70) (49.5 f 12.10) (5.5 f 8.65)
(11.0 f 14.65)
(10.0 f 9.70)
rPF4 (0.5 pg/mL) 71.9 91.0 (14.0 f 6.15)
rPF4 (2 pg/mL) 97.0 88.6 (7.5 f 13.70)
rPF4 (5 pg/mL) 95.9 93.4 21.0 47.9 94.4
94.9 (4.60 f 4.45)
Percent inhibition due to preincubation of neutrophils with hPF4, rPF4, FMLP or zymosan-activated serum for 3 0 minutes at 37°C. Experiments in the last column were done with the antibody included in the medium. Absolute average migration in micrometers is also indicated in parenthesis with SD shown for three independent experiments.
rPF4 appears to be identical to native PF4. In addition, our purification scheme results in a product that is less contami- nated with other proteins than the commercial hPF4, and does not contain any detectable contaminating endotoxins.
One of the values of synthesizing rPF4 is to confirm proposed biologic functions of the native protein. It is anticipated that minor contaminants in the recombinant protein isolated from bacteria would differ from those present in proteins purified from eucaryotic sources. We have shown that rPF4 and hPF4 have virtually identical chemotac- tic properties. As in the case of hPF4, rPF4 augments the neutrophil migratory response in a dose-dependent pattern, with a maximum neutrophil chemotactic response reached at 5 rg/mL. These chemotactic findings are in agreement with those previously reported.I7." In addition, we have demon- strated that migration towards either PF4 species can be blocked by pre-incubation of cells with either hPF4 or rPF4,
demonstrating that both PF4 species appear to bind to the same neutrophil receptor and that binding to this receptor can inhibit the chemotactic response. Chemotaxis for hPF4 and rPF4 can also be blocked by a polyclonal anti-PF4 antibody, demonstrating the PFCspecific nature of the chemotaxis for both species.
PF4 has been reported to be an immunologic regulator by inhibiting suppressor T-cell activity" and to have an auto- crine effect on megakaryocyte differentiation.2' Studies with the recombinant protein should further support the impor- tance of these proposed biologic functions for PF4. Prelimi- nary using rPF4 suggest that it inhibits megakaryo- cyte differentiation in a manner similar to serum hPF4. Site-directed mutations of rPF4 will further demonstrate whether these many biologic properties depend on the C-terminal region of PF4, which contains heparin-binding activity.
REFERENCES
I . Broekman MJ, Handin RI, Cohen P Distribution of fibrin- ogen, and platelet factor 4 and XI11 in subcellular fractions of human platelets. Br J Haematol31:51, 1975
2. Kaplan KL, Broekman MJ, Chernoff A, Leschnick GR, Drilling M: Platelet a-granule proteins: Studies on release and subcellular localization. Blood 53:604, 1979
3. Holt JC, NiewiarowsGi S: Biochemistry of a-granule proteins. Semin Hematol22: 15 1, 1985
4. Deuel TF, Keim DS, Farmer M, Heinrikson R L Human platelet factor 4: Complete amino acid sequence. Proc Natl Acad Sci USA 74:2256,1977
5. Hermodson M, Schmer G, Kurachi K Isolation, characteriza- tion, and primary amino acid sequence of human platelet factor 4. J Biol Chem 252~6276,1977
6. Levine SP, Wohl H: Human platelet factor 4 Purification and characterization by affinity chromatography. J Biol Chem 251:324, 1976
7. Handin RI, Cohen HJ: Purification and binding properties of human platelet factor four. J Biol Chem 251:4273, 1976
8. Barber AJ, Kaaser-Glanzmann R, Jakabova M, Luscher E F Characterization of a chondroitin 4-sulfate proteoglycan carrier for heparin neutralizing activity (platelet factor 4) released from human blood platelets. Biochem Biophys Acta 286:312, 1972
9. Nath N, Lowery CT, Niewiarowski S: Antigenic and antihep- arin properties of human platelet factor 4 (PF4). Blood 45537, 1975
10. Moore S, Pepper DS, Cash JD: Platelet antiheparin activity: The isolation and characterization of platelet factor 4 released from thrombin-aggregated washed human platelets and its dissociation into subunits and the isolation of membrane-bound antiheparin activity. Biochem Biophys Acta 379:370, 1975
11. Losclzo J, Melnick B, Handin RI: The interaction of platelet factor four and glycosaminoglycans. Arch Biochem Biophys 240 446,1985
12. Rybak ME, Gimbrone MA Jr, Davies PF, Handin RI: Interaction of platelet factor four with cultured vascular endothelial cells. Blood 73:1534, 1989
13. Jordan EE, Oosta GM, Gardner WT, Rosenberg RD: The kinetics of hemostatic enzyme-antithrombin interactions in the presence of low molecular weight heparin. J Biol Chem 255:10081, 1980
14. Dessypris EN, Gleaton JH, Sawyer ST, Armstrong OL: Suppression of maturation of megakaryocyte colony forming unit in vitro by a platelet-released glycoprotein. J Cell Physiol 130:361, 1987
15. Senior RM, Griffin GL, Huang JS, Walz DA, Deuel T F
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom

RECOMBINANT PLATELET FACTOR 4 1295
Chemotactic activity of platelet a granule proteins for fibroblasts. J Cell Biol96:382, 1981
16. Osterman DG, Griffin GL, Senior RM, Kaiser ET, Deuel T F The carboxyl-terminal tridecapeptide of platelet factor 4 is a potent chemotactic agent for monocytes. Biochem Biophys Res Commun 107:130, 1982
17. Bebawy ST, Gorka J, Hyers TM, Webster RO: In vitro effects of platelet factor 4 on normal human neutrophil functions. J Leukocyte Biol39:423, 1986
18. Deuel TF, Senior RM, Chang D, Griffin GL, Heinrikson RL, Kaiser E T Platelet factor 4 is chemotactic, for neutrophils and monocytes. Proc Natl Acad Sci USA 78:4584, 1981
19. Katz IR, Hoffmann MK, Zucker MB, Bell MK, Thorbecke GJ: A platelet-derived immunoregulatory serum factor with T cell affinity. J Immunol 134:3199, 1985
20. Katz IR, Thorbecke GJ, Bell MK, Yin Z, Clarke D, Zucker MB: Protease-induced immunoregulatory activity of platelet factor 4. Proc Natl Acad Sci USA 83:3491,1986
21. Gewirtz AM, Calabretta B, Rucinski B, Niewiaroski S, Xu WY: Inhibition of human megakaryocytopoiesis in vitro by platelet factor 4 (PF4) and a synthetic COOH-terminal PF4 peptide. J Clin Invest 83:1477, 1989
22. Begg GS, Pepper DS, Chesterman CN, Morgan FJ: Com- plete covalent structure of human 0-thromboglobulin. Biochemistry 17:1739, 1978
23. Holt JC, Rabellino EM, Gewirtz AM, Gunkel LM, Rucinski B, Niewiarowski S: Occurrence of platelet basic protein, a precursor of low affinity platelet factor 4 and P-thromboglobulin, in human platelets and megakaryocytes. Exp Hematol 16:302, 1988
24. Castor AR, Miller JW, Walz DA: Structural and biological characteristics of connective tissue activating peptide (CTAP-III), a major human platelet derived growth factor. Proc Natl Acad Sci USA 80:765,1983
25. Richmond A, Balentien E, Thomas HG, Flaggs G, Barton DE, Spiess J, Bordoni R, Franke U, Derynck R: Molecular charac- terization and chromosomal mapping of melanoma growth stimula- tory activity, a growth factor structurally related to 8-thromboglobu- lin. EMBO J 7:2025, 1988
26. Luster AD, Unkeless JC, Ravetch JV: y-interferon transcrip- tionally regulates an early-response gene containing homology to platelet proteins. Nature 3 15:324, 1985
27. Kawahara RS, Deuel T F Platelet-derived growth factor- inducible gene JE is a member of small inducible genes related to platelet factor 4. J Biol Chem 264:679, 1989
28. Schmid J, Weissman C: Induction of mRNA for a serine protease and a 0-thromboglobulin-like protein in mitogen stimulated human leukocytes. J Immunol 139:250, 1987
29. Obaru K, Hattori T, Yamamura Y, Takatsuki K, Nomiyama H, Schuichiro M, Shimada K: A cDNA clone inducible in human tonsillar lymphocytes by a tumor promoter codes for a novel protein of the 8-thromboglobulin superfamily. Mol Immunol26:423, 1989
30. Barone AD, Ghrayeb J, Hammerling U, Zucker MB, Thor- becke G: The expression in Escherichia coli of recombinant human
platelet factor 4, a protein with immunoregulatory activity. J Biol Chem 263:8710,1988
31. Tabor .S, Richardson CC: A bacteriophage T7 RNA poly- merase/promoter system for controlled exclusive expression of specific genes. Proc Natl Acad Sci USA 82:1074,1985
32. Poncz M, Surrey S, LaRocco P, Weiss MJ, Rappaport EF, Conway TM, Schwartz E: Cloning and characterization of platelet factor 4 cDNA derived from a human erythroleukemic cell line. Blood 69:219,1987
33. Graziano MP, Casey PJ, Gilman AG: Expression of cDNA for G proteins in Escherichia coli. J Biol Chem 262:11375,1987
34. Lobb R, Sasse J, Sullivan R, Shing Y., D’Amore P, Jacobs J: Purification and characterization of heparin-binding endothelial cell growth factors. J Biol Chem 261:1924, 1986
35- Russel M, Model P Replacement of the fip gene of Escheri- chia coli by an inactive gene cloned on a plasmid. J Bacteriol 159:1034,1984
36. Bradford MM: A rapid and sensitive method for the quantita- tion of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248, 1976
37. Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680, 1970
38. Batteiger B, Newhall J, Jones RB: The use of tween 20 as a blocking agent in the immunological detection of proteins trans- ferred to nitrocellulose membranes. J Immunol Methods 55:297, 1982
39. Hsu S, Raine L, Fanger H: Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques. J Histochem Cytochem 29:577,1981
40. Boyum A Isolation of mononuclear cells and granulocytes from human blood. Scand J Clin Lab Invest 97:77, 1968
41. Addison LE, Babage J-W: A raft technique for chemotaxis: A versatile method suitable for clinical studies. J Immunol Methods 10385,1976
42. Nelson RD, Quie DG, Simmons RC: Chemotaxis under agarose: A new and simple method for measuring chemotaxis and spontaneous migration of human polymorphonuclear leukocytes and monocytes. J Immunol 115:1650, 1975
43. Tsunasawa S, Stewart JW, Sherman F Amino-terminal processing of mutant forms of yeast iso-1-cytochrome c: The specificities of methionine amino peptidase and acetyltransferase. J Biol Chem 2605382,1985
44. Boissel J-P, Kasper TJ, Shah SC, Malone JI, Bunn HF: Amino-terminal processing of proteins: Hemoglobin South Florida, a variant with retention of initiator methionine and Na-acetylation. Proc Natl Acad Sci USA 82:8448,1985
45. Flinta C, Persson B, Jornvall H, Von Heijne G: Sequence determinants of cytosolic N-terminal protein processing. Eur J Biochem 154:193,1986
46. Park KS, DeFeo P, Poncz M, Gewirtz A: Recombinant human platelet factor 4 inhibits human megakaryopoiesis in vitro. Blood 74:288a, 1989 (suppl 1)
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom

1990 75: 1290-1295
KS Park, S Rifat, H Eck, K Adachi, S Surrey and M Poncz demonstrate identity with the native proteinBiologic and biochemic properties of recombinant platelet factor 4
http://www.bloodjournal.org/content/75/6/1290.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on November 25, 2018. by guest www.bloodjournal.orgFrom