binding of 3h-catecholamines to a-noradrenergic receptor sites in
TRANSCRIPT

THE JOURNAL OF BIOLOIXCAL CHEMISTRY Vol 252, No. 19, Issue of September 25, pp. 6450-6463, 1977
Prmfed in U.S.A.
Binding of 3H-Catecholamines to a-Noradrenergic Receptor Sites in Calf Brain*
(Received for publication, April 25, 1977)
DAVID C. U'PRICHARD AND SOLOMON H. SNYDER
From the Departments of Pharmacology and Experimental Therapeutics and Psychiatry and Behavioral Sciences, Johns Hopkins University School of Medicine, Baltimore, Maryland 21205
The binding of (+ )-L3Hlepinephrine and (-PHlnorepi- nephrine to calf cortical membranes in the presence of 1.0 mM pyrocatechol has properties indicating an association with a-noradrenergic receptors. Binding of the “H-catechol- amines associated with /3-noradrenergic receptors has not been detected, even at low concentrations of pyrocatechol. Chromatographic studies reveal that radioactivity bound to membranes is authentic catecholamine. Binding of both “H- catecholamines is saturable with dissociation constants at 37” of 18 nM for (?)-[3Hlepinephrine and 26 nM for c-j- [“Hlnorepinephrine. The Hill coefficients for binding of both “H-ligands are 1.0. At 37, both “H-catecholamines asso- ciate rapidly to equilibrium, and binding is completely re- versible. Kinetically determined dissociation constants cor- respond well with values obtained from equilibrium studies. Binding is stereoselective for both ligands, with (-)-norepi- nephrine displaying about 40 times greater affinity than (+)-norepinephrine. The structure-activity relationship of a series of phenylethanolamine derivatives in competing for “H-catecholamine binding sites parallels the known relative potencies of these compounds at peripheral a-receptors. (Y- Antagonists and partial agonists are potent competitors, whereas /&antagonists and catechol derivatives are ex- tremely weak. The structurally dissimilar a-agonist, [“Hlclonidine, seems to bind to the same calf cortical mem- brane sites as the “H-catecholamines, whereas the a-antago- nist, “H-labeled WB-4101, binds to sites with the same LY- receptor characteristics, but which have greater affinity for antagonists and lesser affinity for agonists. At 25” and 4”, the “H-catecholamines and agonist competitors have 2 to 4 times greater affinity for the binding sites than at 37”, whereas antagonists are 2 to 10 times weaker, and partial agonists have approximately the same affinity. At 4” and 25”, “H-catecholamines dissociate in a biphasic manner which is time-dependent.
In binding studies of neurotransmitter receptors, direct la- beling of receptor sites with the transmitter itself can provide
* This work was supported by United States Public Health Service grant MH-18501, Research Scientist Development Award MH-33128 to S.H.S., and United States Public Health Service Fellowship Award MH-05105 to D.C.U’P. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “‘uduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
valuable information. Characteristics of the dopaminergic and serotonergic receptors in the brain have been elucidated through binding studies with L3H]dopamine and [“Hlserotonin (l-3). However, numerous attempts, primarily in P-receptor tissues, to label noradrenergic receptors with the transmitter IgH]norepinephrine and other 3H-catecholamines have met with only limited success, because in most of these studies 3H- catecholamine binding has failed to demonstrate (a) normal attributes of ligand-receptor binding, and (b) stereospecificity and other structure-activity features characteristic of noradre- nergic receptors (4-11). Some success according to these crite- ria has been met in the binding of [“Hlisoproterenol to p- receptors in avian erythrocytes (12, 13). In the past year, (Y- noradrenergic receptor sites have been labeled with the ago- nist [“Hlclonidine and the antagonist “H-labeled WB-4101 (2- (12’,6’-dimethoxylphenoxyethylamino)methyl benzodioxan) in the brain (14, 151, and with the mixed agonist-antagonist [“Hldihydroergokryptine in rabbit uterine (16, 17) and rat cerebral cortical (18) membranes. Recently we have described conditions under which the binding of “H-catecholamines to central nervous system tissue, in the presence of high concen- trations of pyrocatechol, seems to have a-receptor specificity (19), and binding of a potent catecholamine derivative to /3- receptors in frog erythrocytes has been observed also in the presence of pyrocatechol(20). In the present paper we examine in more detail the characteristics of (+)-[“Hlepinephrine and (-)-[YH]norepinephrine binding to calf brain membranes with properties indicating a selective labeling of cu-noradrenergic receptors.
MATERIALS AND METHODS
Methods were essentially as described previously (19). Gray mat- ter from the frontal lobe of fresh or frozen calf cerebral cortices was homogenized in 20 volumes of ice-cold 50 rnM Tris/HCl buffer, pH 7.7 at 25”, with a Tekmar model SDT homogenizer (setting No. 80,30 s). The homogenates were centrifuged twice at 50,000 x g for 10 min, with rehomogenization in fresh buffer between centrifugations. For “H-catecholamine binding assays, the final pellets were resuspended in 46.5 volumes of ice-cold 50 mM Tris/HCl buffer (pH 8.0 at 25” for ( +-)-[3H]epinephrine; pH 7.7 at 25” for (-)-YHlnorepinephrine). The tissue suspensions were incubated with 1.0 pM pargyline for 15 min at 25” and returned to ice. For L3H1clonidine and 3H-labeled WB-4101 binding assavs. final pellets were resuspended in 49 volumes of ice- cold 5crnM %&/HCl buffer (pH 7.5 at i5” for clonidine; pH 7.7 for WB-4101), and the pargyline preincfibation step was omitted.
Ligunds- DL-[7-“H]Eiinephrine L-bitartrate, 9.9 to 11.0 Ci/mmol, and L-[7,8-3H]norepinephrine, 24.4 to 35.6 Ci/mmol, were obtained from New England Nuclear Corp., and stored under nitrogen at 5”. Immediately before use, each 3H-catecholamine ligand was diluted
6450
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Catecholamine a-Receptor Binding in Brain 6451
with 0.1% ascorbic acid to a concentration of 250 no or 125 no ((i)- L3H1epinephrine), or 100 nM ((-)-[3H1norepinephrinel. Under these storage conditions both 3H-catecholamines retained their stability, without loss of specific binding, for about 1 month. WB-4101, pro- vided by WB Pharmaceuticals, Wembley, England, was custom- tritiated to a specific activity of 11.7 Ci/mmol at New England Nuclear by catalytic exchange with 25 Ci of tritiated water (15). [3H1Clonidine hydrochloride, 1.6 Ci/mmol, was the gift of Boehringer Ingelheim, Germany. The radiochemical purity of these ligands was ascertained by thin layer chromatography (151.
Standard Binding Assays- To triplicate incubation tubes on ice were added 20 ~1 of 250 nM or 125 nM (e)-[3H]epinephrine, or 100 nM (-)-13Hlnorepinephrine in 0.1% ascorbic acid, 10 /*l of various con- centrations of drugs dissolved in 0.1% ascorbic acid, 20 ~1 of a solution containing 50 rnM pyrocatechol, 0.05% ascorbic acid, 5 rnM disodium EDTA, and 0.5 rnM dithiothreitol, and 0.95 ml of freshly resuspended tissue. The final incubation concentrations were: (i)- 13Hlepinephrine, 5 nM or 2.5 nM, or (-)-[3H1norepinephrine, 2 nM; ascorbic acid, 0.004%; pyrocatechol, 1 mM; disodium EDTA, 0.1 mM; dithiothreitol, 10 PM; tissue, 20 mg, original wet weight. The tubes were incubated at 37” for 5 min with 5 nM (-+I-lSH]epinephrine, 25”, for 60 min with 2.5 nM (?)-[3H]epinephrine, or 37” for 5 min, or 25” for 40 min with 2 nM (-)-13H]norepinephrine. Following incubation, the tubes were immediately filtered under vacuum through Whatman GF/B filters presoaked in 1 rnM pyrocatechol. The filters were rinsed with 16 ml of ice-cold 50 mM TrisiHCl buffer, pH 7.7, at 25”, contain- ing 1 rnM pyrocatechol and 0.1% ascorbic acid, and were subse- quently counted by liquid scintillation spectrometry in 10 ml of Formula 947 (New England Nuclear) at 37% efficiency. PHlClonidine and 3H-labeled WB-4101 binding assays were conducted as described previously (14, 15). Saturable or specific binding of (*I- [3H]epinephrine and (-)-[“Hlnorepinephrine in different incubation conditions was defined as the excess over blanks containing 0.1 PM oxymetazoline.
Chromatography of “Free” and “Bound” ( e))-[3H]Epinephrine- (?)-13HlEpinephrine (10 nM) was incubated with 500 mg of calf cortical tissue, wet weight, and the standard concentrations of EDTA and dithiothreitol, in a volume of 10 ml for varying times at 37’ or 25”. Some incubations were conducted in the presence of 0.1 pM oxymetazoline and some in the absence of tissue. The binding reac- tion was stopped by centrifugation at 4” and 50,000 x g for 10 min. Pellets were kept cold, resuspended rapidly in 2.0 ml of ice-cold Tris/ HCl buffer (pH 8.0 at 25’1, and recentrifuged. The final pellets were resuspended in 3.0 ml of a 90% ethanol plus 1.0% ascorbic acid solution and incubated under nitrogen for 20 min at 37” in a Dubnoff incubator to elute the bound radioactivity. Following centrifugation at 50,000 x g for 10 min, the ethanolic supernatant was reduced in volume to 300 to 500 ~1 under nitrogen. Ascending thin layer chro- matography of pellet extracts and supernatants from the original centrifugation, alongside stock (f )-PHlepinephrine and authentic (-I-epinephrine, was performed on prerun cellulose sheets (East- man Kodak, Rochester, N. Y.) using an acidic solvent system con- taining 1-butanol:acetic acid:water (4:1:5), upper phase. Iodine vapor was used to visualize authentic epinephrine. Rebinding experiments were conducted by adding aliquots of pellet extracts or supernatants from the original centrifugation to standard assay incubation tubes.
Drugs- Catecholamine compounds were donated by Sterling- Winthrop, (z)-a-methyl-(+)-norepinephrine by Dr. P. N. Patil (Cal- lege of Pharmacy, The Ohio State University), indoramin by Wyeth, and ergot alkaloids by Sandoz. Other drugs were obtained from the pharmaceutical company of origin or commercial sources.
RESULTS
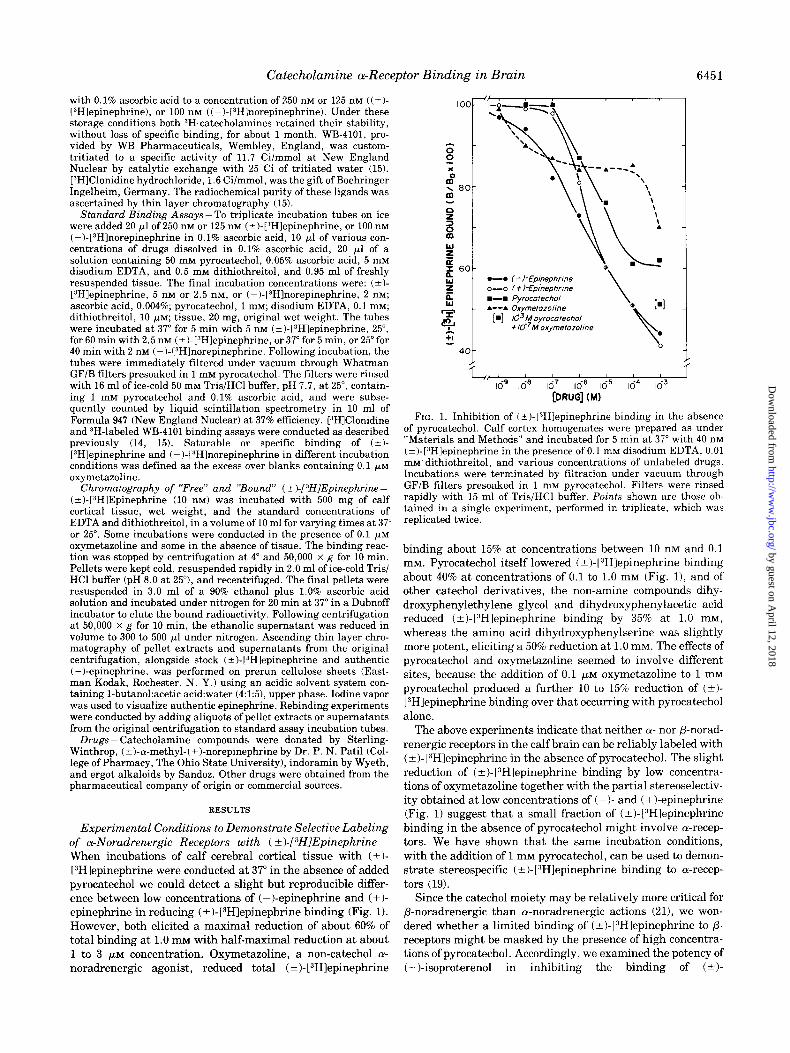
Experimental Conditions to Demonstrate Selective Labeling of a-Noradrenergic Receptors with ( k)-[DHIEpinephrine- When incubations of calf cerebral cortical tissue with (+I- [3H]epinephrine were conducted at 37” in the absence of added pyrocatechol we could detect a slight but reproducible differ- ence between low concentrations of (-)-epinephrine and (+I- epinephrine in reducing (? )- [3H]epinephrine binding (Fig. 1). However, both elicited a maximal reduction of about 60% of total binding at 1.0 mM with half-maximal reduction at about 1 to 3 PM concentration. Oxymetazoline, a non-catechol cy- noradrenergic agonist, reduced total ( +)- 13H1epinephrine
-. I-l-Epineptme o-o (t)-EomeDhrme \
L / L, 1 1
lo- Id6 167 10-j lo-5 Id4 l63
[DRUG] (MI
FIG. 1. Inhibition of (?I-[“Hlepinephrine binding in the absence of pyrocatechol. Calf cortex homogenates were prepared as under “Materials and Methods” and incubated for 5 min at 37” with 40 nM (?)-[3Hlepinephrine in the presence of 0.1 mM disodium EDTA, 0.01 mM’ dithiothreitol, and various concentrations of unlabeled drugs. Incubations were terminated by filtration under vacuum through GF/B filters presoaked in 1 mM pyrocatechol. Filters were rinsed rapidly with 15 ml of Tris/HCl buffer. Points shown are those ob- tained in a single experiment, performed in triplicate, which was replicated twice.
binding about 15% at concentrations between 10 nM and 0.1 mM. Pyrocatechol itself lowered (?I-1”Hlepinephrine binding about 40% at concentrations of 0.1 to 1.0 mM (Fig. 11, and of other catechol derivatives, the non-amine compounds dihy- droxyphenylethylene glycol and dihydroxyphenylacetic acid reduced (&)-[“Hlepinephrine binding by 35% at 1.0 mM, whereas the amino acid dihydroxyphenylserine was slightly more potent, eliciting a 50% reduction at 1.0 mM. The effects of pyrocatechol and oxymetazoline seemed to involve different sites, because the addition of 0.1 FM oxymetazoline to 1 mM pyrocatechol produced a further 10 to 15% reduction of (+I- VHlepinephrine binding over that occurring with pyrocatechol alone.
The above experiments indicate that neither OI- nor P-norad- renergic receptors in the calf brain can be reliably labeled with (? )- [“Hlepinephrine in the absence of pyrocatechol. The slight reduction of ( +)-(3H]epinephrine binding by low concentra- tions of oxymetazoline together with the partial stereoselectiv- ity obtained at low concentrations of ( - )- and ( + )-epinephrine (Fig. 1) suggest that a small fraction of (-cl-[“Hlepinephrine binding in the absence of pyrocatechol might involve a-recep- tors. We have shown that the same incubation conditions, with the addition of 1 mM pyrocatechol, can be used to demon- strate stereospecific (-+ )- l:‘H]epinephrine binding to cu-recep-
tors (19). Since the catechol moiety may be relatively more critical for
P-noradrenergic than a-noradrenergic actions (21), we won- dered whether a limited binding of (?)-[3Hlepinephrine to /3- receptors might be masked by the presence of high concentra- tions of pyrocatechol. Accordingly, we examined the potency of ( - )-isoproterenol in inhibiting the binding of (k+)-
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
6452 Catecholamine a-Receptor Binding in Brain
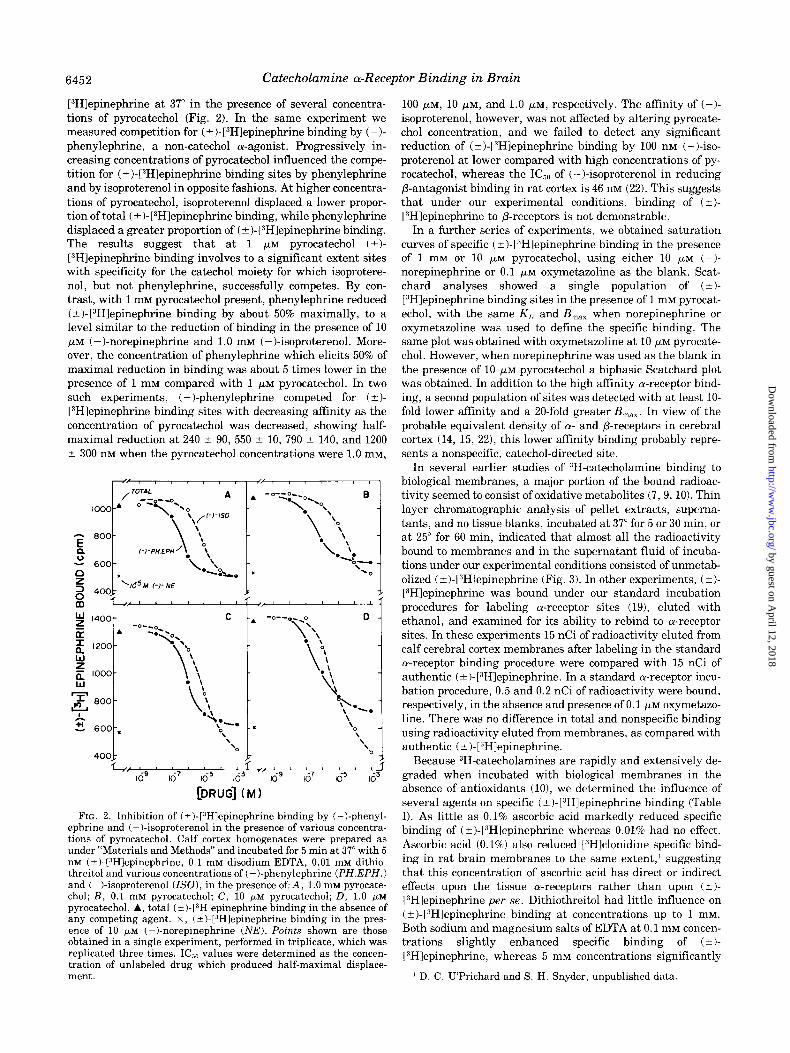
[“Hlepinephrine at 37” in the presence of several concentra- tions of pyrocatechol (Fig. 2). In the same experiment we measured competition for ( + )- [“Hlepinephrine binding by ( - )- phenylephrine, a non-catechol a-agonist. Progressively in- creasing concentrations of pyrocatechol influenced the compe- tition for (?I-[“Hlepinephrine binding sites by phenylephrine and by isoproterenol in opposite fashions. At higher concentra- tions of pyrocatechol, isoproterenol displaced a lower propor- tion of total ( ?)-[3H]epinephrine binding, while phenylephrine displaced a greater proportion of (?)-[3H1epinephrine binding. The results suggest that at 1 pM pyrocatechol (?)- [“Hlepinephrine binding involves to a significant extent sites with specificity for the catechol moiety for which isoprotere- nol, but not phenylephrine, successfully competes. By con- trast, with 1 mM pyrocatechol present, phenylephrine reduced ( a)-[3H]epinephrine binding by about 50% maximally, to a level similar to the reduction of binding in the presence of 10 pM (-)-norepinephrine and 1.0 mM (-)-isoproterenol. More- over, the concentration of phenylephrine which elicits 50% of maximal reduction in binding was about 5 times lower in the presence of 1 mM compared with 1 pM pyrocatechol. In two such experiments, (-)-phenylephrine competed for (t)- VHlepinephrine binding sites with decreasing affinity as the concentration of pyrocatechol was decreased, showing half- maximal reduction at 240 & 90, 550 2 10, 790 -+ 140, and 1200 t 300 nM when the pyrocatechol concentrations were 1.0 mM,
[DRUG] (MI
FIG. 2. Inhibition of (t-)- p?H]epinephrine binding by (-)-phenyl- ephrine and (-)-isoproterenol in the presence of various concentra- tions of pyrocatechol. Calf cortex homogenates were prepared as under “Materials and Methods” and incubated for 5 min at 37” with 5 nrvr (-cl-13H]epinephrine, 0.1 rn~ disodium EDTA, 0.01 rn~ dithio- threitol and various concentrations of (-Lphenylephrine (PH.EPH.1 and (-)-isoproterenol (ISO), in the presence of: A, 1.0 mM pyrocate- chol; B, 0.1 rnM pyrocatechol; C, 10 pM pyrocatechol; D, 1.0 /*M
pyrocatechol. A, total (i)-[3Hlepinephrine binding in the absence of any competing agent. x, (k)-[“Hlepinephrine binding in the pres- ence of 10 PM (-)-norepinephrine (NE). Points shown are those obtained in a single experiment, performed in triplicate, which was replicated three times. IC,, values were determined as the concen- tration of unlabeled drug which produced half-maximal displace- ment.
100 PM, 10 PM, and 1.0 PM, respectively. The affinity of (-)- isoproterenol, however, was not affected by altering pyrocate- chol concentration, and we failed to detect any significant reduction of (?I-[“Hlepinephrine binding by 100 nM (-)-iso- proterenol at lower compared with high concentrations of py- rocatechol, whereas the IC,, of (-)-isoproterenol in reducing P-antagonist binding in rat cortex is 46 nM (22). This suggests that under our experimental conditions, binding of (k)- VHlepinephrine to P-receptors is not demonstrable.
In a further series of experiments, we obtained saturation curves of specific ( f)- l”H]epinephrine binding in the presence of 1 mM or 10 PM pyrocatechol, using either 10 FM (-I- norepinephrine or 0.1 pM oxymetazoline as the blank. Scat- chard analyses showed a single population of (*l- [3H]epinephrine binding sites in the presence of 1 mM pyrocat- echol, with the same K,, and B,,, when norepinephrine or oxymetazoline was used to define the specific binding. The same plot was obtained with oxymetazoline at 10 PM pyrocate- chol. However, when norepinephrine was used as the blank in the presence of 10 PM pyrocatechol a biphasic Scatchard plot was obtained. In addition to the high affinity a-receptor bind- ing, a second population of sites was detected with at least lo- fold lower affinity and a 20-fold greater B,,,. In view of the probable equivalent density of w and P-receptors in cerebral cortex (14, 15, 221, this lower affinity binding probably repre- sents a nonspecific, catechol-directed site.
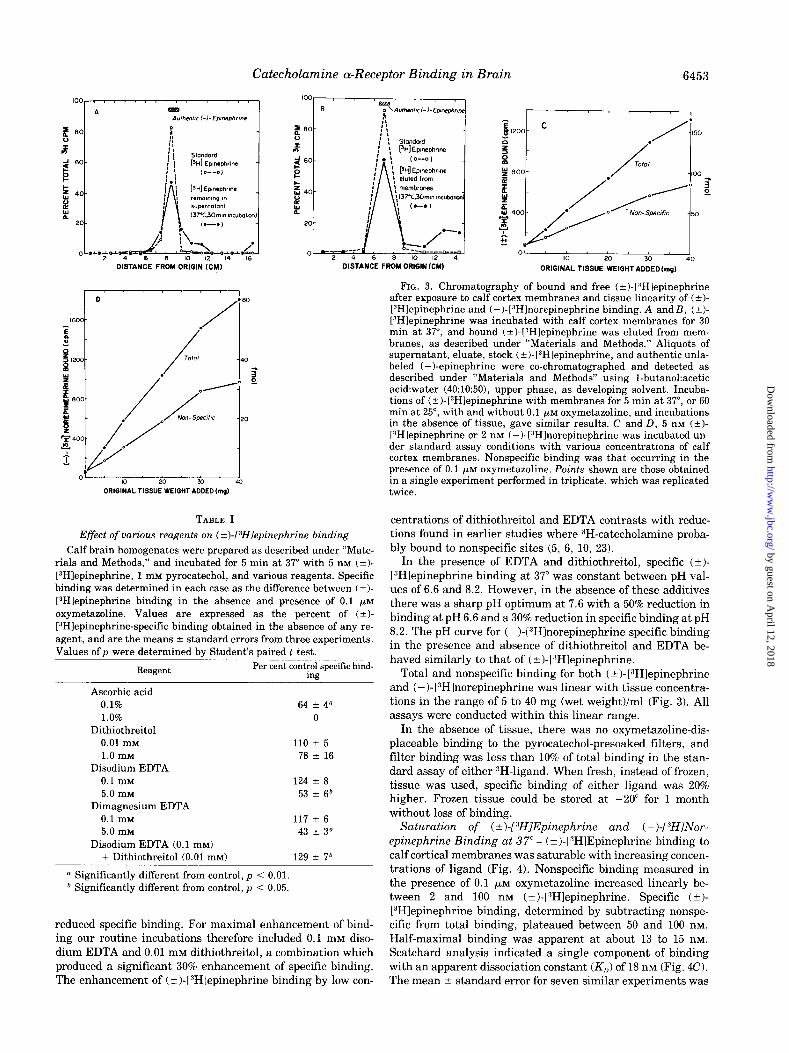
In several earlier studies of “H-catecholamine binding to biological membranes, a major portion of the bound radioac- tivity seemed to consist of oxidative metabolites (‘7, 9, 101. Thin layer chromatographic analysis of pellet extracts, superna- tants, and no tissue blanks, incubated at 37” for 5 or 30 min, or at 25” for 60 min, indicated that almost all the radioactivity bound to membranes and in the supernatant fluid of incuba- tions under our experimental conditions consisted of unmetab- olized (+)-[“Hlepinephrine (Fig. 3). In other experiments, (?I- [“Hlepinephrine was bound under our standard incubation procedures for labeling a-receptor sites (191, eluted with ethanol, and examined for its ability to rebind to a-receptor sites. In these experiments 15 nCi of radioactivity eluted from calf cerebral cortex membranes after labeling in the standard a-receptor binding procedure were compared with 15 nCi of authentic (*I-[“Hlepinephrine. In a standard a-receptor incu- bation procedure, 0.5 and 0.2 nCi of radioactivity were bound, respectively, in the absence and presence of 0.1 PM oxymetazo- line. There was no difference in total and nonspecific binding using radioactivity eluted from membranes, as compared with authentic (i)-[“Hlepinephrine.
Because “H-catecholamines are rapidly and extensively de- graded when incubated with biological membranes in the absence of antioxidants (lo), we determined the influence of several agents on specific (t)-[“Hlepinephrine binding (Table I). As little as 0.1% ascorbic acid markedly reduced specific binding of (?I-[“Hlepinephrine whereas 0.01% had no effect. Ascorbic acid (0.1%) also reduced [“Hjclonidine specific bind- ing in rat brain membranes to the same extent,’ suggesting that this concentration of ascorbic acid has direct or indirect effects upon the tissue a-receptors rather than upon (?)- [“Hlepinephrine per se. Dithiothreitol had little influence on (?I-[“Hlepinephrine binding at concentrations up to 1 mM. Both sodium and magnesium salts of EDTA at 0.1 mM concen- trations slightly enhanced specific binding of (?I- 13H1epinephrine, whereas 5 mM concentrations significantly
1 D. C. U’Prichard and S. H. Snyder, unpublished data.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Catecholamine a-Receptor Binding in Brain 6453
DISTANCE FROM DRIQIN KM)
01 ’ 10 20 Y) 40
ORIGINALTISSUE WEIGHTADDEDh,,$
TABLE I
Effect of various reagents on (k)-PHlepinephrine binding
Calf brain homogenates were prepared as described under “Mate- rials and Methods,” and incubated for 5 min at 37” with 5 nM (t)- [3H]epinephrine, 1 rnM pyrocatechol, and various reagents. Specific binding was determined in each case as the difference between (?)- PHlepinephrine binding in the absence and presence of 0.1 pM
oxymetazoline. Values are expressed as the percent of (?)- [3H]epinephrine-specific binding obtained in the absence of any re- agent, and are the means 2 standard errors from three experiments. Values of p were determined by Student’s paired t test.
Reagent
Ascorbic acid 0.1% 1.0%
Dithiothreitol 0.01 rnM 1.0 mM
Disodium EDTA 0.1 rnM
5.0 rnM Dimagnesium EDTA
0.1 rnM 5.0 rnM
Disodium EDTA (0.1 mM)
Per cent control specific bind- ing
64 f 4” 0
110 f 5
‘78 k 16
124 f 8
53 t 6b
117 f 6 43 T 3”
+ Dithiothreitol (0.01 mM) 129 rfl 76
a Significantly different from control, p < 0.01. b Significantly different from control, p i 0.05.
reduced specific binding. For maximal enhancement of bind- ing our routine incubations therefore included 0.1 mM diso- dium EDTA and 0.01 mM dithiothreitol, a combination which produced a significant 30% enhancement of specific binding. The enhancement of (t)-[“Hlepinephrine binding by low con-
01 * 1 I I 10 2-0 33 40
ORIGINAL TISSUE WEIGHT ADDED
FIG. 3. Chromatography of bound and free (?I-[“Hlepinephrine after exposure to calf cortex membranes and tissue linearity of (?I- L”Hlepinephrine and (-)-[“Hlnorepinephrine binding. A andB, (*I- L”Hlepinephrine was incubated with calf cortex membranes for 30 min at 37”, and bound (?I-13Hlepinephrine was eluted from mem- branes, as described under “Materials and Methods.” Aliquots of supernatant, eluate, stock (?)-[“Hlepinephrine, and authentic unla- beled (-)-epinephrine were co-chromatographed and detected as described under “Materials and Methods” using 1-butanol:acetic acid:water (40:10:50), upper phase, as developing solvent. Incuba- tions of (?I-l?H]epinephrine with membranes for 5 min at 37”, or 60 min at 25”, with and without 0.1 pM oxymetazoline, and incubations in the absence of tissue, gave similar results. C and D, 5 nM (?)- 13Hlepinephrine or 2 nM (-)-1:‘Hlnorepinephrine was incubated un- der standard assay conditions with various concentrations of calf cortex membranes. Nonspecific binding was that occurring in the presence of 0.1 yM oxymetazoline. Points shown are those obtained in a single experiment performed in triplicate, which was replicated twice.
centrations of dithiothreitol and EDTA contrasts with reduc- tions found in earlier studies where “H-catecholamine proba- bly bound to nonspecific sites (5, 6, 10, 23).
In the presence of EDTA and dithiothreitol, specific (?)- [“Hlepinephrine binding at 37” was constant between pH val- ues of 6.6 and 8.2. However, in the absence of these additives there was a sharp pH optimum at 7.6 with a 50% reduction in binding at pH 6.6 and a 30% reduction in specific binding at pH 8.2. The pH curve for (-)-lRHlnorepinephrine specific binding in the presence and absence of dithiothreitol and EDTA be- haved similarly to that of C-t)-1”Hlepinephrine.
Total and nonspecific binding for both (?)-[“Hlepinephrine and (-)-l”H]norepinephrine was linear with tissue concentra- tions in the range of 5 to 40 mg (wet weight)/ml (Fig. 3). All assays were conducted within this linear range.
In the absence of tissue, there was no oxymetazoline-dis- placeable binding to the pyrocatechol-presoaked filters, and filter binding was less than 10% of total binding in the stan- dard assay of either JH-ligand. When fresh, instead of frozen, tissue was used, specific binding of either ligand was 20% higher. Frozen tissue could be stored at -20” for 1 month without loss of binding.
Saturation of (*I-[sH]Epinephrine and (-I-[“HINor- epinephrine Binding at 37”- (*I-[“HlEpinephrine binding to calf cortical membranes was saturable with increasing concen- trations of ligand (Fig. 4). Nonspecific binding measured in the presence of 0.1 ~.LM oxymetazoline increased linearly be- tween 2 and 100 nM (*I-1”Hlepinephrine. Specific (*I- lYH]epinephrine binding, determined by subtracting nonspe- cific from total binding, plateaued between 50 and 100 nM. Half-maximal binding was apparent at about 13 to 15 nM. Scatchard analysis indicated a single component of binding with an apparent dissociation constant (K,,) of 18 nM (Fig. 4C). The mean t standard error for seven similar experiments was
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6454 Catecholamine a-Receptor Binding in Brain
18.4 + 1.7 nM. The calculated maximal number of binding sites (B,,,,) from seven experiments was 11 * 1 pmol/g of cerebral cortex tissue, wet weight. A plot of these data accord- ing to the Hill equation revealed a single straight line with a Hill coefficient (n,,) of 1, indicating the absence of positively or negatively cooperative interactions. The ratio of total (+I- L3H]epinephrine binding to nonspecific binding was about 2 at a concentration of 5 nM (*I-I”Hlepinephrine, which was used for routine binding assays.
For (-)-I JH]norepinephrine, nonspecific binding, measured in the presence of 0.1 PM oxymetazoline, increased almost linearly between 1 and 100 nM (-)-[“Hlndrepinephrine. Spe- cific (-)-I “Hlnorepinephrine binding, taken as the difference between total and nonspecific binding, was saturable with a plateau apparent between about 50 and 100 nM [:$H]norepinephrine. Half-maximal binding was apparent at about 20 nM. Scatchard analysis indicated the presence of a single population of binding sites; the dissociation constant (K,,) determined was about 31 nM (Fig. 5C). The mean * standard error for three similar experiments was 26.0 +- 3.6 nM. The maximal number of binding sites was calculated from three experiments as 7.3 t 0.2 pmol/g of cerebral cortex, wet weight. Thus, the maximal numbers of (-)-[“HInor- epinephrine and (f)-[“Hlepinephrine binding sites were simi- lar, although the (-)-[“Hlnorepinephrine B,,, value was defi- nitely lower. The affinity of (t)-[“Hlepinephrine for binding sites appeared to be about 1.5 times the affinity of (-)-[“HI- norepinephrine, and since (2)~[“Hlepinephrine was used in these studies and binding in this system is stereoselective for the biologically active isomer (-)-epinephrine (191, the K,) for (-)-epinephrine would be about 9 nM, indicating a 3-fold greater affinity than (-)-[“Hlnorepinephrine. As was observed
with ( ? )- I 3H]epinephrine binding, a plot of ( - )- [ “Hl-norepi- nephrine binding according to the Hill equation revealed one linear component with a Hill coefficient of 1. The ratio of total to nonspecific (-)-[“Hlnorepinephrine binding was about 2 at a concentration of 2 nM (-)-[“HI-norepinephrine, which was used for routine binding assays.
Kinetics of ( +)-[3H]Epinephrine and ( -)-[8H]Norepi- nephrine Binding- In some earlier studies of 3H-catechola- mine binding to tissue membranes binding did not reach equi- librium even with long incubation intervals but continued to increase gradually, apparently because binding involved irre- versible membrane interactions of catecholamine metabolites (6, 7). Under our experimental conditions 3H-catecholamine binding did reach equilibrium. At 37” (?)-[3H]epinephrine binding increased rapidly to a plateau at about 5 min (Fig. 6). At this temperature equilibrium levels of binding remained stable at least until 20 min, with some slight decrease in specific binding at 30 min. Half-maximal binding at 37” was apparent at about 1 min. At 25” binding did not reach equilib- rium until 30 min and values remained constant until 180 min. Half-maximal binding at 25” was attained at about 5 min. Binding proceeded even more slowly at 4” with equilibrium being reached at 240 min and half-maximal values at about 30 min. Strikingly, equilibrium levels of specific (*I- [“Hlepinephrine binding at 37” were only half of values at 4 and 25”, whereas the a-agonist ligand [“Hlclonidine in rat brain membranes associated to the same levels of specific binding at 4”, 25”, and 37” (15). Nonspecific ( t)-[JH]epi- nephrine binding did not associate instantaneously, but at a rate which was independent of temperature, reaching maxi- mum levels at 3 min with a half-time of 40 s. The maximum level of nonspecific binding was also independent of tempera-
FIG. 4. (?I-[“HlEpinephrine bind- ing as a function of increasing concen- trations of (f)-[“Hlepinephrine. Calf cortex homogenates (20 mg of tissue, original wet weight) were incubated for 5 min at 37”, as described under “Mate- rials and Methods,” with various con- centrations of (-c)-[3H]epinephrine. Nonspecific binding was determined by the addition of 0.1 ELM oxymetazoline. Points shown are those obtained in a single experiment, performed in tripli- cate, which was replicated seven times. A, total and nonspecific binding. B, specific binding, defined as difference between total and nonspecific. C, Scat- chard plot, showing a K,] value of 18.4 nM and receptor density bf 13 pmol/g of tissue, wet weight. D, Hill plot, with nH = 0.97.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Catecholamine a-Receptor Binding in Brain 6455
FIG. 5. (-I- [ 3 H] Norepinephrine binding as a function of increasing con- centrations of (-I-[“Hlnorepinephrine. Calf cortex homogenates (20 mg of tis- sue, original wet weight) were incu- bated for 5 min at 3’7”, as described under “Materials and Methods,” with various concentrations of Cc)- t3H]norepinephrine. Nonspecific bind- ing was determined by addition of 0.1 ELM oxymetazoline. Points shown are those obtained in a single experiment, performed in triplicate, which was rep- licated three times. A, total and non- specific binding. B, specific binding de- fined as difference between total and nonspecific. C, Scatchard plot, showing a K,, value of 31.4 nM and receptor den- sity of 7 pmol/g of tissue, wet weight. D, Hill plot, with nH = 0.99.
[t-I-f%ii NOREPINEPHRINE] (nM)
(pm01 /a tissur)
I-I-[3H] NOREPINEPHAINE (n)rl)
+05
D .
P : n,=osB o
f4D a! .
=
-05-
. /
3 : / .
-1 56 -90 -85 -80 -75
L.,~ [(-I-[34 NOREPINEPHRINE] CM)
ture, and nonspecific binding remained constant at each tem- perature for long periods of time. (&I-[“HlEpinephrine did not associate to a-receptors in a truly hyperbolic manner, and the semilogarithmic plots of association were linear for only the initial phase of association (Fig. 6A, inset).
In certain earlier studies, 3H-catecholamine binding to bio- logical membranes was irreversible (7, 9, lo), or only partially reversible in the absence of guanyl nucleotides (201. In the present investigation, binding of both (+I-VHlepinephrine and (-)-[“Hlnorepinephrine was completely reversible at 37”. The rate of dissociation of (+)-[3H]epineIjhrine binding was examined by incubating calf cortical membranes to equilib- rium at either 37” or 25”, equilibrating to the desired tempera- ture of dissociation for 2 min, and then adding 0.1 PM oxyme- tazoline to prevent rebinding of dissociated ( f )- [“Hlepi- nephrine (Fig. 6B). The ( ?)-[3H]epinephrine remaining bound was then assessed at three temperatures after various time intervals. During 2 min equilibrations at 37”, the high specific binding of (+-)-l”H]epinephrine associated to membranes at 25 reverted almost completely back to the specific binding level
after 37” association. When plotted on a semilogarithmic scale the dissociation of
(+)-[:‘H]epinephrine binding, which had attained equilibrium at 37” after 5 min, was monophasic at 37”, 25”, and 4”, with mean half-times from three experiments, for dissociation at each temperature, of 50 s, 3% min, and 18 min, respectively. The dissociation of (t)-[“Hlepinephrine that had been incu-
bated to equilibrium at 25” for 60 min was monophasic at 37”, but biphasic at 25” and 4”, with the initial rates of dissociation at the three temperatures being identical to the dissociation rates obtained when the receptor was labeled at 37”.
The means * standard errors from five experiments of the rate constants for dissociation of ( *)-[3H]epinephrine binding (km,), determined from the linear dissociation plots obtained after 37” association, or the initial dissociation slope after 25” association, were 0.84 2 0.04, 0.24 ? 0.04, and 0.04 ? 0.01 min-’ at 37”, 25”, and 4”, respectively. The calculation of the rate constants of association at the three different tempera- tures was based on pseudo-first order kinetics dependent on the binding site concentration, which can be described by the equation In@,/@, - B)] = k,, t where B is the amount of (*I- [3H]epinephrine bound at each time (t) and B, is the amount bound at equilibrium. The slopes of the lines in Fig. 6A, inset, represent the observed initial rate constants (k,,) at 37”, 25”, and 4” in one experiment. The second order rate constant (k,) of (&)-[3H]epinephrine binding was then determined from the equation k, = (!z,~ - km,)/[EPI] where [EPI] is the concentra- tion of (t)-[YH]epinephrine used in the association experi- ments. Thus kob should be dependent on the amount of C&l- [3H]epinephrine used, and in a series of experiments the ob- served rate of association increased with increasing (f)- [3H]epinephrine concentration.
The means ? standard errors, from four association experi- ments, of the rate constants for association of (*I-
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6456 Catecholamine a-Receptor Binding in Brain
[“Hlepinephrine binding at 37”, 25”, and 4”, were, respectively, 0.045 ? 0.009, 0.040 ? 0.024, and 0.012 + 0.002 nM-’ min-I. The kinetically derived estimate for the KD of (?)-[“Hlepinephrine binding at 37” (k-,/k,) was 18.7 2 3.8 nM, in close agreement with the value derived from equilibrium experiments (Fig. 4). However, the kinetically derived K,) values at 25”, and 4”, were 6.2 + 3.9 nM and 3.3 ? 0.8 nM, respectively. The lower K,, values at 25” and 4” suggest that the higher equilibrium levels of binding observed at these temperatures compared to 37 reflect a higher affinity of (+-)-[“Hlepinephrine for the receptor at lower temperatures, and that this higher affinity is due to slower dissociation of the ligand from a portion of the receptor binding sites. Preincubation of the tissue with pyrocatechol, as opposed to the standard simultaneous incubation with pyro- catechol and (?I-[“Hlepinephrine, lowered the equilibrium specific binding attained in 25” association experiments by 20%, but did not alter the rate of association of specific or nonspecific binding.
The kinetics of (-)-[“Hlnorepinephrine binding were very similar to those of (?I-[“Hlepinephrine (Fig. 7). At 37” the association of (- )-[“Hlnorepinephrine to specific binding sites occurred rapidly with equilibrium attained in 2 to 3 min. At 37 binding levels remained constant for about 20 min with some decrease at 30 min. Equilibrium was reached at 25” and 4 after about 20 min and 4 h, respectively. Half-maximal bind- ing at 25” and 4” occurred at 5 min and 30 min, respectively. As was observed with (t)-[“Hlepinephrine, equilibrium levels of specific (-)-[“Hlnorepinephrine binding were substantially higher at 25” and 4” than at 37”, and (-)-[“Hlnorepinephrine association at all three temperatures was not strictly hyper-
TIME (mm1
FIG. 6. Time course of association and dissociation of (?I- [3H]epinephrine specific binding. A, association of (k)-PHlepi- nephrine specific binding to calf cortex homogenates (20 mg, original tissue weight) at 4”, 25”, and 37” was measured at various time intervals following the addition of 5 nM (+-)-[3Hlepinephrine. Specific binding was defined as the difference between binding obtained in the absence and presence of 0.1 FM oxymetazoline. Incubation condi- tions were as described under “Materials and Methods.” Points shown are those from a single experiment performed in triplicate, which was replicated twice. Inset, pseudo-first order kinetic plots of initial (2)-[“Hlepinephrine specific binding at 4” (x),25” (WI, and 3’7 (0). B is the amount of (i)-[3H]epinephrine specifically bound at time t, and B, is the amount of (?I-lSHlepinephrine specifically bound at equilibrium. The slopes of these lines (shown in figure) are k “b> equal to the observed rate constant for the pseudo-first order
bolic. The characteristics of association of nonspecific (-)- [3H]norepinephrine binding were the same as was found with ( ? )- [ “Hlepinephrine.
The rates of dissociation of (-)-[3H]norepinephrine binding at different temperatures were examined in the same way as for (*I-[“Hlepinephrine. A semilogarithmic plot gave linear slopes for (-)-[JH]norepinephrine dissociation at 37” and 4”, and an almost linear slope at 25”, when the receptors were initially labeled at 37”. The mean half-times from 2 to 4 experiments for (-)-[“Hlnorepinephrine dissociation were 20 to 30 s, 1 to 2 min, and 15 min at 37”, 25”, and 4”, respectively. After initial labeling of the receptor at 25” for 40 min with (-)- [“Hlnorepinephrine, dissociation at 37” was monophasic, but dissociation at 25” and 4” was biphasic, with the initial phases coinciding with the slopes obtained following 37” labeling. Thus, closely similar results were obtained for (-)- [“Hlnorepinephrine and (+)-[“Hlepinephrine dissociation. Again equilibration at 37” for 2 min of membranes initially labeled to equilibrium with (-I-[“Hlnorepinephrine at 25” al- most totally reversed the increased binding obtained with 25” association.
From three to four experiments the mean * standard error of the kinetic constants for (-)-[“Hlnorepinephrine binding at 37”, 25”, and 4” were k-,: 1.40 2 0.19, 0.24 2 0.02, 0.05 2 0.01 min-I; andk,: 0.09 + 0.02, 0.06 & 0.02, 0.02 ? 0.01 nM-’ min-I, respectively. The kinetically derived K,, for (-)-[“Hlnorepi- nephrine binding (h-,/k,) at 37” was 15 2 4 nM, somewhat lower than the value of 26 nM derived from equilibrium ex- periments (Fig. 5). As with (t)-[3H]epinephrine, the K, values for (-)-[“Hlnorepinephrine obtained kinetically at 25
TIME (min)
reaction. The second order association rate constant, k,, is calculated from k, = (k,, - k-,)/[(i-)-[3H]epinephrine] where k-, is the first order rate constant for dissociation and I(i)-L:‘Hlepinephrinel is the concentration of (?I-[“Hlepinephrine used in the experiment (5 nM). Points shown are those from a single experiment replicated four times. B, dissociation ofbound (?I-[“Hlepinephrine was measured at 4”, 25”, and 37” following incubation with (?I-[“Hlepinephrine to equilibrium at 25” (open symbols) or 37” (solid symbols) under stan- dard assay conditions, and subsequent equilibration for 2 min to the indicated temperatures. At time zero, 0.1 PM oxymetazoline was added to the incubation mixtures, and the reactions terminated by filtration at various time intervals. Points shown are those from a single experiment, performed in triplicate, which was replicated three times.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
CatechoLamine a-Receptor Binding in Brain 6457
and 4” were much lower, 4 * 1 nM and 3 f 1 nM, respectively. Characteristics of Zncreased “H-Catecholamine Binding at
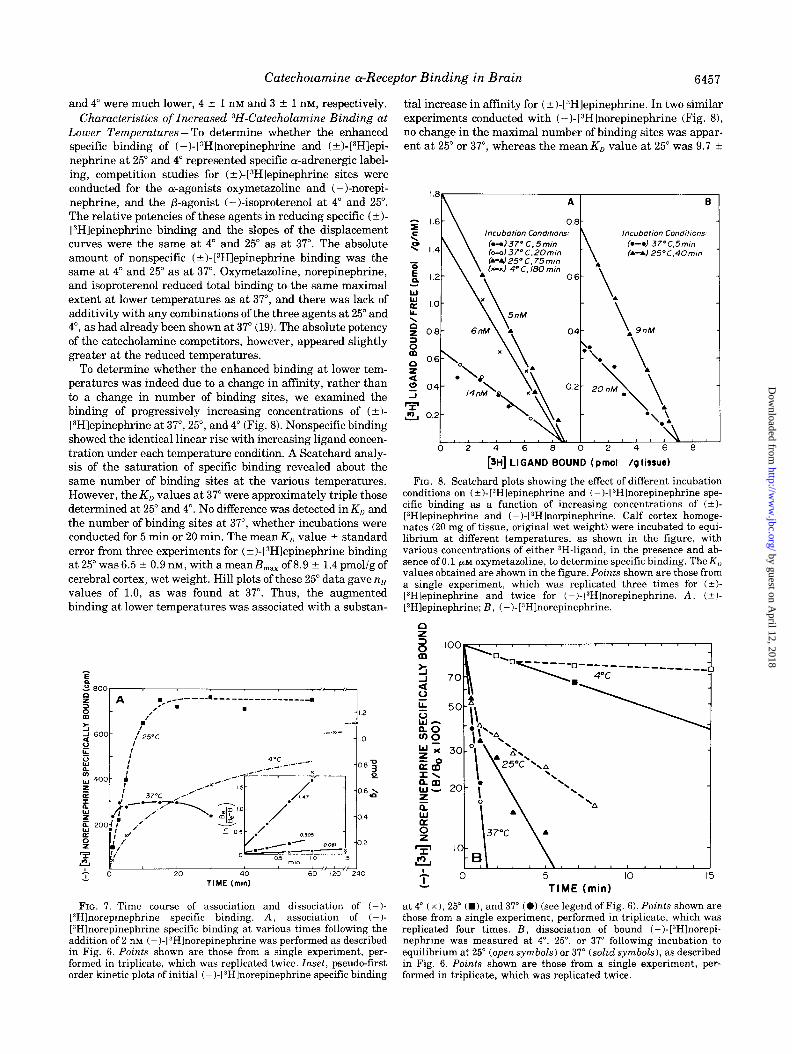
Lower Temperatures - To determine whether the enhanced specific binding of ( -)-[3H]norepinephrine and (?)-[3Hlepi- nephrine at 25” and 4” represented specific cY-adrenergic label- ing, competition studies for (+)-[3Hlepinephrine sites were conducted for the a-agonists oxymetazoline and (-)-norepi- nephrine, and the P-agonist (-)-isoproterenol at 4” and 25”. The relative potencies of these agents in reducing specific (*I- [“Hlepinephrine binding and the slopes of the displacement curves were the same at 4” and 25” as at 37”. The absolute amount of nonspecific (+)-[3H1epinephrine binding was the same at 4” and 25” as at 37”. Oxymetazoline, norepinephrine, and isoproterenol reduced total binding to the same maximal extent at lower temperatures as at 37”, and there was lack of additivity with any combinations of the three agents at 25” and 4”, as had already been shown at 37” (19). The absolute potency of the catecholamine competitors, however, appeared slightly greater at the reduced temperatures.
To determine whether the enhanced binding at lower tem- peratures was indeed due to a change in affinity, rather than to a change in number of binding sites, we examined the binding of progressively increasing concentrations of (*I- [3H]epinephrine at 37”, 25”, and 4” (Fig. 8). Nonspecific binding showed the identical linear rise with increasing ligand concen- tration under each temperature condition. A Scatchard analy- sis of the saturation of specific binding revealed about the same number of binding sites at the various temperatures. However, the K,, values at 37” were approximately triple those determined at 25” and 4”. No difference was detected in K,, and the number of binding sites at 37”, whether incubations were conducted for 5 min or 20 min. The mean KD value -r- standard error from three experiments for (+)-[3H]epinephrine binding at 25” was 6.5 ? 0.9 nM, with a meanB,,, of 8.9 ? 1.4 pmol/g of cerebral cortex, wet weight. Hill plots of these 25” data gave nH values of 1.0, as was found at 37”. Thus, the augmented binding at lower temperatures was associated with a substan-
P i?a I
A c------ ,:* .
.-------- -------- ~ .
I’
i TIME (mid
FIG. 7. Time course of association and dissociation of (-I- [“Hlnorepinephrine specific binding. A, association of (-)- [E’Hlnorepinephrine specific binding at various times following the addition of 2 nM (-)-[“Hlnorepinephrine was performed as described in Fig. 6. Points shown are those from a single experiment, per- formed in triplicate, which was replicated twice. Inset, pseudo-first order kinetic plots of initial (-)-[3H]norepinephrine specific binding
tial increase in affinity for (k)-[“Hlepinephrine. In two similar experiments conducted with (-)-[3Hlnorepinephrine (Fig. 81, no change in the maximal number of binding sites was appar- ent at 25” or 37”, whereas the mean K, value at 25” was 9.7 2
A
0.8 - tncubotton Condftmns: h--137* C.Smin b-ol37~C.2Omln
B
Incub(Itmn Conditmns (*-*I 37°C.5mln (r-r) 25’C,40mm
0 2 4 6 8 0 2 4 6 0
[3H] LIGAND BOUND (pmol /g tissue)
FIG. 8. Scatchard plots showing the effect of different incubation conditions on (?)-[3Hlepinephrine and (-)-13Hlnorepinephrine spe- cific. binding as a function of increasing concentrations of (?I- [“Hlepinephrine and (-)-[3Hlnorpinephrine. Calf cortex homoge- nates (20 mg of tissue, original wet weight) were incubated to equi- librium at different temperatures, as shown in the figure, with various concentrations of either 3H-ligand, in the presence and ab- sence of 0.1 PM oxymetazoline, to determine specific binding. The K,] values obtained are shown in the figure. Points shown are those from a single experiment, which was replicated three times for (?I- [3Hlepinephrine and twice for (-I-[“Hlnorepinephrine. A, (iI- [3Hlepinephrine; B, (-)-[“Hlnorepinephrine.
37°C
o- ’ -B
0 5 IO 15
TIME (min)
25” (WI, and 37” (0) (see legend of Fig. 6). Points shown are those from a single experiment, performed in triplicate, which was replicated four times, B, dissociation of bound (-)-[3Hlnorepi- nephrine was measured at 4”, 25”, or 37” following incubation to equilibrium at 25” (open symbols) or 37” (solid symbols), as described in Fig. 6. Points shown are those from a single experiment, per- formed in triplicate, which was replicated twice.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6458 Catecholamine a-Receptor Binding in Brain
0.8 nM, 3 times lower than at 37”. The Hill coefficient (q,) at 25” was 1.0, as at 37”.
[3HIClonidine and 3H-Labeled WB-4101 Binding in Calf Cerebral Cortex - The a-agonist [“Hlclonidine and the cw-an- tagonist 3H-labeled WB-4101 have been used to label cY-recep- tors in rat brain membranes (14, 15). In the present study with calf frontal cortex membranes, both ligands also labeled (Y- receptors, and were approximately twice as potent as in the rat brain. When increasing concentrations of [3H]clonidine or 3H-labeled WB-4101 were incubated with calf cortical mem- branes in the presence or absence of high concentrations of (-)-norepinephrine, specific binding of each 3H-ligand was saturable, with apparent K,) values of 2.6 nM and 0.35 nM for [;Hlclonidine and 3H-labeled WB-4101, respectively. The max-
imum number of binding sites in calf cortical membranes was 10 pmol/g for [“Hlclonidine and 7 pmol/g for “H-labeled WB- 4101, values which are very similar to those for (+I- 13Hlepinephrine and (-)-[“Hlnorepinephrine.
Influence of Various Drugs on Specific ( +)-[3HIEpi- nephrine and ( -)-13H]Norepinephrine Binding at 37’- Eval- uation of a limited number of drugs in preliminary studies established that specific (2 )-I “Hlepinephrine and ( -)- L3Hlnorepinephrine binding to calf cerebral cortex membranes was stereospecific and inhibited by a-noradrenergic agonists and antagonists but not by some other drugs (19). In the present study we have examined a much larger series of agents (Tables II and III). In previous studies of “H-catechola- mine binding, numerous catechol compounds lacking an
TABLE II Inhibition of ( k)-[3Hlepinephrine, 13Hlclonidine, and 3H-labeled WE-4101 a binding to calf frontal cortex membranes
Calf cortex homogenates were incubated with 5 no (k)- by log-probit analysis, and apparent K, values calculated from the PHlepinephrine for 5 min at 37”, 2.5 nM (+)-L3Hlepinephrine for 60 equation K, = I&,/(1 + [3H-ligandl/K,) for each ligand. The KI, min at 25”, 4 nM [3Hlclonidine for 30 min at 25”, or 0.25 nM 3H-labeled values used for (t)-[3Hlepinephrine were 18.4 nM at 37” and 6.5 nM at WB-4101 for 20 min at 25”, together with three to five concentrations 25”. Values given are means ? standard errors for the numbers of of unlabeled drugs, under standard assay conditions for each 3H- experiments indicated in parentheses, each conducted in triplicate. ligand (see “Materials and Methods”). IC,, values were determined Values of p were determined by Student’s t test (two-tailed).
Drug K, 13Hlepinephrine/37 K, [‘Hlepinephrine/25 K, [“Hlclonidine K, “H-labeled WB- 4101
a-Agonists (-)-Epinephrine (+)-Epinephrine (k)-a-Me-(?I-Norepinephrine (r)-a-Me-(+)-Norepinephrine (-)-Norepinephrine (+)-Norepinephrine Dopamine (-)-Phenylephrine Clonidine Naphazoline Oxymetazoline
a-Partial agonists Ergotamine Dihydroergotamine a-Ergokryptine Dihydro-a-ergokryptine n-Lysergic acid diethylamide Ergonovine Serotonin Piperoxan
a-Antagonists Phentolamine WB-4101” Phenoxybenzamine Dibenamine Indoramin Chlorpromazine Haloperidol
P-Agonists and antagonists (-)-Isoproterenol (-)-Alprenolol (-)-Propranolol
Catechol derivatives Dihydroxyphenylethylene glyc Dihydroxyphenylacetic acid Dihydroxyphenylserine
6.5 t 1.9 (3) 97 f 16 (5) 28 k 5 (3)
16,000 (1) 24 f 3 6)
1,000 f 100 (4) 310 f 30 (4)
370 k 50 3.3 + - 0.7 iii 1.4 k 0.4 (4) 3.3 k 0.8 (4)
5.8 f 0.6 (4) 9.6 t 1.1 (4)” 3.2 k 0.9 (2) 6.4 + 0.3 (3)” 13 t 2 (4) 13 k 1 (3)
8.3 t 1.5 (4) 5.4 r 0.6 (3)” 330 f 10 (2) 250 k 90 (2)
3,800 k 900 (3) 5,600 + 1,300 (2) 5,700 f 800 (2) 6,600 f 800 (2)
22 f 3 (3) 18 f 6 (3)
1.2 f 0.3 (5) 16 k 4 (5)
200 k 30 (4) 1,500 f 400 (3)
12,000 k 2,000 (3) 1,300 i- 300 (2) 5,800 ? 3,000 (2)
3,900 k 900 (4) 21,000 t 3,000 (2) 35,000 ? 6,000 (2)
:ol >lOfi 1106 -10”
2.1 t 0.2 (3)” 26 f 4 (3)’
7.7 k 1.0 (3)” 4,600 (1)
6.4 t 0.6 (5)l 260 i 60 (3Y
87 f 13 140 f 20 ii;:: 1.3 f 0.3 (4)” 0.9 t 0.1 (4)b 4.7 ?I 1.0 (5)
2.0 k 0.2 (5)’
11,000 * 3,000 (3) 3,400 i 2,000(2) 8,000 + 4,000 (2)
1,100 ‘- 200 (3)” 19,000 f 3,000(2)
>70,000 (2)
>lO” >lO”
-2 x lo”
4
16 1.300 360 49:ooo 230 260
1.2 450 0.9 1.6
9 3.1 12 2.3
1.2 1.2 27 0.3
150 -16
2,700
16,000
a WB-4101 = 2-~l2’.6’-dimethoxvlnhenoxvethvlamino)methvlbe~odioxan. I -_
b Significantly different from K, at 37”, p” < d.05. I’ Significantly different from Ki at 37”, p < 0.001. d Significantly different from K, at 37”, p < 0.01.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Catecholamine a-Receptor Binding in Brain 6459
a-Agonists ( - f-Epinephrine (+)-Epinephrine (k)-a-Me-C-C)-Norepi-
nephrine (%I-c~-Me-(+l-Norepi-
nephrine (-)-Norepinephrine (+)-Norepinephrine Dopamine (-)-Phenylephrine Clonidine Naphazoline Oxymetazoline
a-Partial agonists Ergotamine Dihydroergotamine a-Ergokryptine Dihydro-cr-ergokryp-
tine n-Lysergic acid diethyl-
amide Ergonovine Serotonin Piperoxan
a-Antagonists Phentolamine WB-4101’ Phenoxybenzamine Dibenamine Indoramin Chlorpromazine Haloperidol
P-Agonists and antago- nists
(-)-Isoproterenol (-)-Alprenolol ( - )-Propranolol
Catechol derivatives Dihydroxyphenyleth-
ylene glycol Dihydroxyphenylacetic
acid Dihydroxyphenylserine
TABLE III
Inhibition of (-)-[3Hlnorepinephrine binding to calf frontal cortex membranes
Calf cortex membranes were incubated with 2 nM c-l- L”HInorepinephrine, either for 5 min at 37” or for 40 min at 25”. Apparent K, values were determined as described in Table I. The K,, values used for (-)-[“Hlnorepinephrine were 26 nM at 37” and 9.7 nM at 25”. Values given are means ? standard errors for the numbers of experiments indicated in parentheses, each conducted in triplicate. Values ofp were determined by Student’s t test (two-tailed).
Drug [“Hlnore n+ephrine/ f
ILM
4.7 f 1.2 (51 34 k 3 (3) 36 -t 8 (3)
35,000 (1)
19 f 5 U-5) 710 t 170 (4) 210 t 20 (2) 220 t 20 (2) 3.2 T 1.3 (4) 2.7 ‘- 0.9 (4) 4.3 i- 2.0 (3)
7.4 t 2.2 (4) 5.2 -t 1.7 (3) 15 f 6 (31 11 Y!Z 2 (3)
370 f 150 (2)
3,600 k 1,400 (2) 9,800 zk 1,200 (2)
30 f 10 (3)
1.4 ? 0.3 (4) 17 i- 4 (3)
240 2 60 (4) 2,200 f 400 (3)
20,000 t 4,000 (3) 740 f 40 (2)
6,900 f 1,100 (2)
3,700 (1) 19,000 + 3,000 (2) 36,000 k 9,000 (2)
>lO”
>lO”
1.0 % 0.1 (3)” 16 2 5 w
7.6 i 1.3 (31”
4,800 (1)
5.7 f 0.7 (3)” 150 k 10 (3)” 75 f 9 (3)”
110 ‘- 10 (31” 1.4 t 0.6 (31 1.3 -t 0.3 (410 3.7 It 0.9 (3)
11.3 t 4.8 (4) 5.7 -t 1.8 (3)
34 k 13 (3) 24 t 9 (3)
240 -t 150 (2)
3,800 +- 600 (2) 9,800 -t 3,000 (21
14 * 2 (3)
3.0 t 0.6 (31” 26 t 5 (3)
1,500 i 500 (3)” 11,000 * 5,000 (3lh
9,600 f 1,200 (3tb 1,400 i 100 (2)”
12,000 * 4,000 (21
1,300 (1) 11,000 * 4,000 (2)
>60,000 (2)
>lO”
>lO”
-3 x lo”
” Significantly different from K, at 37”, p < 0.01. b Significantly different from K, at 37”, p < 0.05. c WB-4101 = 2-([2’,6’-dimethoxylphenoxyethylaminofmethylbenzo-
dioxan.
amine moiety were as potent as catecholamines in reducing binding (6, 7, 10, 23). In the present study, catechol agents such as dihydroxyphenylacetic acid and dihydroxyphenylethy- lene glycol had negligible influence on specific (k)- [“Hlepinephrine or (-)-IJHlnorepinephrine binding in concen-
trations exceeding 1 mM. Dihydroxyphenylserine, a catechol amino acid, was slightly more potent than dihydroxyphenyl- ethylene glycol or dihydroxyphenylacetic acid, probably be- cause of the side chain nitrogen, but was still very weak. By contrast (-)-epinephrine displayed an apparent K, value of about 7 nM in competition with ( t)-l:SH]epinephrine and 5 nM in competition with (-)-l”H]norepinephrine. These values are very close to the apparent Ki of 4 nM for (-)-epinephrine competing with l”H]clonidine, a noncatecholamine a-agonist. “H-Catecholamine binding was markedly stereospecific with (-)-epinephrine being about 16 and 8 times more potent than (+)-epinephrine, and (-)-norepinephrine being about 42 and 37 times more potent than (+)-norepinephrine in reducing (+-)-[“Hlepinephrine and (-)-[“Hlnorepinephrine binding, re- spectively. The apparent Ki values determined at 37” for (-I- epinephrine and (- )-norepinephrine resembled the dissocia- tion constants determined for tritiated forms of these drugs in equilibrium and kinetic experiments also performed at 37”.
The structure-activity relationships of a series of phenyleth- ylamine compounds in competing for ( 2 )- 1 “Hlepinephrine and (-)-[:‘H]norepinephrine binding at 37”, and [:‘H]clonidine bind- ing at 25”, were very similar to each other, and corresponded closely to known physiological affinities at peripheral a-recep- tor sites (241, and affinities at [“Hlclonidine binding sites in the rat brain (15). The relative affinities of (-)-epinephrine, (- )-norepinephrine, dopamine, (- )-phenylephrine, and (- )- isoproterenol (lower numbers having greater affinity) were 1:4:48:57:600 for (-+I-l”H]epinephrine, 1:4:45:47:790 for (-I- [“Hlnorepinephrine, and 1:4:58:64:680 for [“Hlclonidine. cy- Methyl-norepinephrine was intermediate in potency between epinephrine and norepinephrine for “H-catecholamine binding sites, and showed the greatest stereospecificity, the racemic form being 600 and 1000 times more potent than the (+)-form in competing for (2 )-[“Hlepinephrine and (-1-I “Hlnorepi- nephrine binding, respectively.
The imidazoline a-agonists clonidine, oxymetazoline, and naphazoline displayed apparent K, values of 1 to 4 nM in competing with (?I-[:‘H]epinephrine, (-)-[:‘H]norepinephrine, and [“Hlclonidine, and hence were even more potent than (-I- epinephrine. The affinities of the imidazolines in reducing catecholamine binding were slightly greater than their afini- ties for a-receptor sites labeled by [“Hlclonidine in the rat (15). In the calf, the affinities of phenylethylamine and imidazoline a-agonist competitors, such as (-)-norepinephrine and cloni- dine, were essentially the same whichever of the three “H- agonist ligands was used. However, the affinities of these two agents in displacing the a-antagonist ligand :‘H-labeled WB- 4101 in the same tissue were 60 to 100 times less (Table II). This is in agreement with results for 1”Hlclonidine and “H- labeled WB-4101 in the rat brain (151, and suggests that, in the calf cerebral cortex, while structurally dissimilar a-agonist ligands label the same site, the a-antagonist ligand labels a different site.
The a-antagonist phentolamine was extremely potent in competing for 1”Hlcatecholamine binding with an apparent K, value of about 1.2 nM against (+I-[:‘H]epinephrine and 1.4 nM against (-)-l:lHJnorepinephrine. Phentolamine was about 15, 170, and 1200 to 1500 times more potent than WB-4101, phe- noxybenzamine, and dibenamine, other well known cu-norad- renergic antagonists. Of the haloalkylamines, dibenamine was much weaker than phenoxybenzamine, in concurrence with pharmacologic data (25). Antagonist drugs had the same affinity in competing with l:‘H]clonidine or the “H-catechol- amines, but, with the exception of phentolamine, were much
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6460 Catecholamine a-Receptor Binding in Brain
more potent in competing with “H-labeled WB-4101. Phentol- amine was equipotent against the “H-agonist or “H-antagonist ligands, suggesting that in the calf cortex it has mixed ago- nist-antagonist properties. In general, the antagonist competi- tors showed the same potencies against “H-agonists in the calf cortex as against I”Hlclonidine in the rat brain (E), except that imidazoline (phentolamine) and benzodioxan (WB-4101 and piperoxan) derivatives were much more potent in the calf brain. The neuroleptic agents chlorpromazine and haloperidol are very potent a-antagonists physiologically (261, and highly effective competitors for 3H-labeled WB-4101 binding in rat brain (15, 27). They were relatively weak competitors for (tl- [“Hlepinephrine (Table II) and (-)-[3Hlnorepinephrine bind- ing (Table III), with about the same affinity as they have against [‘~Hlclonidine in the rat brain (15).
Several ergot alkaloids with mixed agonist-antagonist ac- tivity at a-receptors (25, 28) were potent inhibitors of (*I- [ “Hlepinephrine and ( - )- L3H]norepinephrine binding. Appar- ent K, values for dihydroergotamine, ergokryptine, dihydroer- gokryptine, and ergotamine ranged from about 3 to 15 nM. The non-peptide ergots d-lysergic acid diethylamide and ergonov- ine, which display little direct a-receptor activity in uiuo, had respective apparent Ki values of 350 and 4000 nM against both catecholamine ligands. The indoleamine transmitter, sero- tonin, displayed very weak affinity for both :‘H-catecholamine binding sites. Piperoxan, a physiological a-antagonist, which in binding studies appears to have mixed agonist-antagonist properties (151, was quite potent in displacing (*I- [ “Hlepinephrine and (-)-[“Hlnorepinephrine, with apparent Kj values of 22 and 30 nM, respectively. The P-receptor antago- nists propranolol and alprenolol competed only weakly for c-t)- [“Hlepinephrine and (-)-[“Hlnorepinephrine binding with ap- parent Ki values of about 35 pM for (-)-propranolol and 20 PM
for (-)-alprenolol. Competition for :‘H-Catecholamine Binding Sites at 25”-
Because of the enhanced affinity of RH-catecholamine binding to a-receptors at lower temperatures, we evaluated the influ- ence of drugs on “H-catecholamine binding at 25” (Tables II and III). Most agonist agents displayed significantly lower apparent Ki values at 25” than at 37”. The apparent K, values of (-)-epinephrine at 37”, 25”, and 4” corresponded closely to what would be expected from the apparent K,, values deter- mined at each temperature for the racemic form of [“Hlepinephrine. Thus (-)-epinephrine is 3 and 5 times more potent in competing with (*I-IsHlepinephrine at 25” and 4” than at 37” (Table II). Similar findings were obtained with (-)- [“Hlnorepinephrine competition experiments at 25” (Table III). Thus the apparent Ki value of (-)-norepinephrine against (-I- [“Hlnorepinephrine, 5.7 nM, was very similar to the apparent
K,] value for (-)-[“Hlnorepinephrine of 9.7 nM obtained at the same temperature. All phenylethylamine and imidazoline ag- onists exhibited significant 2- to 4-fold increases in affinity with either :‘H-catecholamine ligand at 25” compared to 37”. One exception was oxymetazoline, which showed no compara- ble increase in affinity. Oxymetazoline is a potent competitor of :%H-labeled WB-4101 binding in rat brain and would seem to have more mixed agonist-antagonist characteristics than other a-agonists (15).
An opposite pattern was detected for the u-antagonists. Phentolamine, phenoxybenzamine, WB-4101, and dibenamine were significantly more potent against either 3H-catechola- mine ligand at 37” than at 25”. The potency differences ranged from 2-fold for phentolamine, which seems relatively mixed in this system, to 5- to lo-fold for the irreversible antagonists
phenoxybenzamine and dibenamine. Chlorpromazine and haloperidol, which possess known a-antagonist activity, also were more potent at 37” than at 25”.
The mixed agonist-antagonists showed little significant dif- ference in affinities at 37” and 25” for binding sites labeled by both “H-catecholamine ligands.
In studies of receptor binding for dopamine and serotonin, marked differences between agonists and antagonists have been observed in slopes of competition curves (3, 29,301. Thus, whereas Hill coefficients for reduction of [“Hlserotonin and [“Hldopamine binding by agonists are about 1.0, antagonists display Hill slopes substantially lower than 1.0 in reducing [3H]serotonin and [“Hldopamine binding (29). To evaluate the properties of inhibition of “H-catecholamine binding by var- ious drugs, we have examined the slopes for reduction of binding in some detail (Fig. 9). Agonists, partial agonists, and antagonists caused parallel reductions of (&I-VHlepinephrine and ( -)-L3H]norepinephrine binding at 37”. The extent of max- imal reduction was the same for all drugs evaluated. More- over, there was no additivity in reduction of binding between maximally inhibitory concentrations of agonists, partial ago- nists, and antagonists. The slopes of competition curves for agonists, partial agonists, and antagonists were the same at 25 and 37”. The Hill coefficients (n,) of any agent competing with either 3H-catecholamine ligand at 37” and 25” were all about 1.0 (Fig. 9). Thus, contrary to the results obtained with seroto- nergic and dopaminergic binding, and in agreement with ear- lier studies of I”H]clonidine and 3H-labeled WB-4101 binding to rat brain a-receptors, agonists and antagonists exhibited qual- itatively similar competition at a-receptor sites. In addition, the enhanced affinity of agonists and decreased affinity of antagonists at 25” compared to 37” was not attributable to any change in the slope of displacement curves.
DISCUSSION
The present study demonstrates that ( 5 )- 13Hlepinephrine and ( - )- [“Hlnorepinephrine can be utilized to label a-noradre- nergic receptors in calf cerebral cortex membranes. Several aspects of the binding data indicate that physiological (Y- receptors are being labeled. For both :‘H-catecholamines, bind- ing is saturable and shows high affinity. The maximum num- ber of sites labeled is much lower than was found in previous reports on catecholamine binding (9, II), and approximates the receptor density seen in central nervous system binding studies using other amine transmitters as 3H-ligands (3, 29). Furthermore, the B,,, for (+I-[“Hlepinephrine and (-I- I”Hlnorepinephrine is very close to the B,,, for the non-cate- cholamine a-agonist [“Hlclonidine in calf frontal cortex. Bind- ing of both 3H-catecholamines is stereoselective with respect to the optical isomers of several phenylethanolamine deriva- tives, the biologically active (-)-isomer being considerably more potent in each case. The stereoselectivity is less than has been observed in P-receptor binding studies (20, 22, 31, 321, and whole tissue pharmacological studies suggest that optimal conformation at the /3 carbon on the phenylethanolamine side chain is slightly less paramount at the a-receptor (21).
Binding of “H-catecholamines in the presence of 1.0 mM pyrocatechol is essentially catethol-independent, and non- amine catechol derivatives such as dihydroxyphenylethylene glycol and dihydroxyphenylacetic acid are extremely weak. Catechol-independent binding studies of L3Hlnorepinephrine have been conducted previously (9), but mainly in P-receptor containing tissues. Probably with both the CY- and p-noradre- nergic receptor, the three attachment points on the catechola-
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Catecholamine a-Receptor Binding in Brain 6461
FIG. 9. Inhibition of (?I-l”Hlepi- nephrine and (-)-1:‘Hlnorepinephrine binding at 37” and 25” by a-agonists, partial agonists, and antagonists. A, Calf cortex homogenates were incu- bated with 5 nM (-t)-l:‘Hlepinephrine for 5 min at 37” (solid symbols) or 2.5 nM (t)-[3Hlepinephrine for 60 min at 25” (open symbols), together with var- ious concentrations of (-)-norepineph- rine ((-)-NE), ergotamine (EA) or phentolamine (PHE), under standard assay conditions (see “Materials and Methods”). B, homogenates were incu- bated with 2 nM (-I-[“Hlnorepineph- rine either for 5 min at 37” (solid sym- bols) or for 40 min at 25” (open symbols) together with various concentrations of the above drugs. C, Hill plots for the displacement of (-t)-l:<Hlepinephrine. D, Hill plots for the displacement of (-)-[“Hlnorepinephrine.
mine molecule, namely the catechol , side chain hydroxyl , and terminal amino moieties are necessary, but the terminal amino group is more significant at the a-receptor (21). There- fore non-amine catechol compounds in high concentration would still be relatively potent at the P-receptor, but very weak at the a-receptor, preferentially leaving o-receptors un- occupied. Our experiments utilizing decreasing concentrations of pyrocatechol (Fig. 2) demonstrate nonequivalent maximum inhibition of ( t)-lYH]epinephrine binding by catechol and non- catechol competitors even at 0.1 mM catechol, thereby showing nonspecific binding with selectivity for the catechol moiety. With decreasing pyrocatechol concentrations, we fail to find any evidence for the binding of ( t)-13H]epinephrine to /3- noradrenergic sites.
Previous 3H-catecholamine binding studies have empha- sized the slow rate of association of the ligand to the binding sites, and in some cases the nonattainment of equilibrium in contrast to the rapid stimulation of cyclic nucleotide generat- ing systems (4, 11). In our studies, both “H-catecholamines associate very rapidly to a-receptors at 37”, with association constants of 0.8 x lo7 M-’ S-I and 1.5 x lo7 Mm’ S’ for (t)- 13Hlepinephrine and (-)-13Hlnorepinephrine, only slightly be-
P”ELc2 , I
-8 -10 -9 -8 -7
Log [DRUG] (MI
low the theoretical association limit if association is dependent solely on molecular diffusion, with very little geometric re- striction (33). The “H-catecholamine binding we examined is reversible, and the rate of dissociation is temperature-depend- ent. Kinetically determined dissociation constants agree closely with values obtained in equilibrium experiments.
The most convincing evidence that (%I-[:‘H]epinephrine and (-)-[:‘H]norepinephrine are binding to o-receptors lies in the close correlation of the physiological affinities of agents active and inactive at a-receptor sites and their potencies in compet- ing with the “H-ligands. The sites labeled by the :‘H-catechol- amines behave similarly to those labeled by [“Hlclonidine, an a-agonist with a different molecular structure.
Since the number of sites labeled by (?I-[“Hlepinephrine appears to be somewhat greater than the number labeled by (-)-[“Hlnorepinephrine in the same tissue, one might specu- late that (?)-(:‘H]epinephrine is labeling, in addition to cy- noradrenergic receptors, receptors postsynaptic to neurons which use epinephrine itself as the transmitter. Such “adre- nergic” neurons have been postulated to exist in the brain stem (34, 35), but the lack of the epinephrine-synthesizing enzyme, phenylethanolamine N-methyltransferase, in the
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6462 Catecholamine a-Receptor Binding in Brain
cerebral cortex of several mammalian species (361, indicates that (* )-[“Hlepinephrine in our hands is labeling only cortical a-noradrenergic receptors.
At lower temperatures (25” and 4”), (-t)-[“Hlepinephrine and i-)-[“Hlnorepinephrine display higher affinity than at 37”. In addition, competition studies indicate that agonists are more potent at the lower temperatures, whereas antagonists are weaker, and the affinities of partial agonists are essentially unchanged. One possible explanation is that the alterations in affinity are due to a phase change in the membrane suspen- sion at lower temperatures, primarily involving membrane phospholipids as in the case of the membrane-bound Ca’+- ATPase system (37). A more attractive hypothesis is that the interaction of agonist ligand and receptor induces a conforma- tional change to a “desensitized” receptor state, which exhibits higher affinity for agonists. P-Receptors in frog erythrocytes show a reduced number of binding sites for a JH-antagonist ligand and a reduced maximal stimulation of adenylate cy- clase by P-agonists upon prior exposure to a /?-agonist in viva (38) or in vitro (39). Similar findings have been demonstrated for P-receptors in the pineal gland (40). These results were explained on the basis of a loss of free receptor sites available for antagonist binding due to the exceedingly slow dissociation of agonist from desensitized receptors. The desensitized recep- tors, tightly bound to agonist, also cannot activate adenylate cyclase. In erythrocyte membrane preparations, desensitiza- tion is much more rapid, and the preincubated agonist pro- duces half-maximal desensitization at a concentration where it occupies half of the receptors (41). Guanyl nucleotides re- verse this phenomenon (411, and seem to decrease the affinity of a P-agonist ligand for the receptor by increasing the rate of dissociation (20).
The higher affinities of agonist ligands and competitors at lower temperatures could conceivably be due to a conforma- tional change leading to desensitized receptors caused by the lower temperature itself or the longer duration of incubation required to attain equilibrium at these temperatures. The period of contact between agonist and receptor in our studies at 25” is equivalent to times when desensitization has occurred in other isolated membrane studies (41). I f the 1:l correspond- ence in binding affinities of agonists and their desensitizing potency holds for the a-receptor, then significant desensitiza- tion may occur with the “H-catecholamine concentrations used here at 25”. A study of [“Hlepinephrine binding to turkey erythrocytes also shows greater displacement by agonists at lowered temperatures (42).
Dissociation at lower temperatures after attainment of equi- librium over 40 to 60 min at 25” is biphasic for both 3H- catecholamines. The faster and slower phases may conceiva- bly represent dissociation of the agonist from sensitized and desensitized receptors, respectively. The slower rate of disso- ciation may involve receptors which account for the apparent increase in affinity at lower temperatures. The nonlinearity of dissociation appears to depend upon the time of receptor- ligand contact, since “H-catecholamines incubated to equilib- rium for only 5 min at 37” show monophasic dissociation curves at all temperatures. Similar kinetic data have also been ob- tained in the binding of [“Hlprostaglandin E, to sites in cul- tured mammalian cells (43). Such results appear to fit a “cyclic” model of receptor desensitization following agonist interaction (44, 45).
In calf cerebral cortex membranes, the agonists (k)- 13H1epinephrine, (-)-[“Hlnorepinephrine, and [“Hlclonidine
bind to the same site, which has the characteristics of the physiological a-receptor. The a-antagonist “H-labeled WB- 4101 binds to a site which also has a-receptor characteristics, but where antagonists are much more potent, and agonists much less potent. The binding of [“Hlclonidine and “H-labeled WB-4101 in terms of a “two-state” model of the a-receptor has been discussed (15). The absence of any cooperativity in com- petition experiments (Fig. 9) militates against the possibility of interconversion between the two sites. It has been suggested that a-antagonists, which display no structural similarity to the catecholamine transmitters, bind to a regulatory site near the a-receptor and so modulate agonist-receptor binding, as if by direct competition (21). Further studies are in progress to clarify agonist and antagonist sites, and the phenomenon of desensitization, at the a-receptor in the central nervous sys- tem.
Acknowledgments-We thank Dr. David A. Greenberg for his interest and advice, and Messrs. G. Mack and P. Sheehan for their superb technical assistance.
REFERENCES
1. Burt, D. R., Enna, S. J., Creese, I., and Snyder, S. H. (1975) Proc. Natl. Acad. Sci. U. S. A. 72, 4655-4659
2. Seeman. P.. Chau-Wane. M.. Tedesco. J.. and Wang, K. (1975) Proc. tic&. Acad. Sci:U. S. A. 72, 4376-4380 -
3. Bennett, J. P., Jr., and Snyder, S. H. (1976) Mol. Pharmacol. 12, 373-389
4. Lefkowitz, R. J., and Haber, E. (1971) Proc. Natl. Acad. Sci. U. S. A. 68, 1’773-1777
5. Lefiowitz, R. J., Sharp, G. W. G., and Haber, E. (1973) J. Biol. Chem. 248, 342-349
6. Maguire, M. E., Goldmann, P. H., and Gilman, A. G. (1974) Mol. Pharmacol. 10, 563-581
7. Wolfe. B. B.. Zirrolli. J. A.. and Molinoff. P. B. (1974) Mol. Phai-macol.’ 10, 582-j96
8. Tell, G. P. E., and Cuatrecasas, P. (1974)Biochem. Biophys. Res. Commun. 57, 793-800
9. Cuatrecasas, P., Tell, G. P. E., Sica, V., Parikh, I., and Chang, K. J. (1974) Nature 247, 92-97
10. Valieres, J., Drummond, M., and Drummond, G. I. (1975) Can. J. Biochem. 53. 458-469
11. B&r, H. P. (1976)‘in The Mechanism ofNeurona1 and Extraneu- ronal Transport of Catecholamines (Paton, D. M., ed) pp. 247- 257, Raven Press, New York
12. Levitzki, A., Atlas, D., and Steer, M. L. (1974) Proc. Natl. Acad. Sci. U. S. A. 71, 2773-2776
13. Malchoff, C. D., and Marinetti, G. V. (1976) Biochim. Biophys. Actu 436, 45-52
14. Greenberg, D. A., U’Prichard, D. C., and Snyder, S. H. (1976) Life Sci. 19, 69-76
15. U’P&hard, D. C., Greenberg, D. A., and Snyder, S. H. (1977) Mol. Pharmacol., 13, 454-473
16. Williams, L. T., and Lefkowitz, R. J. (1976) Science 192, 791-793 17. Williams, L. T., Mullikin, D., and Lefkowitz, R. J. (1976) J. Biol.
Chem. 251. 6915-6923 18. Greenberg, c. A., and Snyder, S. H. (1977) Life Sci. 20, 927-932 19. U’Prichard. D. C.. and Snyder, S. H. (1977) Life Sci. 20. 527-534 20. Lefkowitz, R. J., and Wiiliams, L. T. (1977) Proc. Natl. Acad.
Sci. U. S. A. 74, 515-519 21. Triggle, D. J. (1976) in Chemical Pharmacology of the Synapse
(Triggle, D. J., and Triggle, C. R., eds) pp. 233-430, Academic Press, New York
22. Bylund, D. B., and Snyder, S. H. (1976) Mol. Pharmacol. 12,568- 580
23. Ruffolo, R. R., McCreery, R. L., and Patil, P. N. (1976) Eur. J. Pharmacol. 38, 221-232
24. Furcheott. R. F. (1970) Fed. Proc. 29. 1352-1361 25. Nickerson; M., and Hollenberg, N. K. (1967) in Physiological
Pharmacology (Root, W. S., and Hoffman, F. G., eds) Vol. 4, pp. 243-305, Academic Press, New York
26. van Rossum, J. M. (1965) J. Pharm. Pharmacol. 17, 202-216
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27.
28. 29.
30.
31.
32.
33.
34.
35.
Catecholamine a-Receptor Binding in Brain 6463
Peroutka, S. J., U’Prichard, D. C., Greenberg, D. A., and Sny- der, S. H. (1977) Neuropharmacology, in press
Innes, I. R. (1962) Br. J. Pharmacol. 19, 120-128 Burt, D. R., Creese, I., and Snyder, S. H. (1976) Mol. Pharmacol.
12, 800-812 Seeman, P., Lee, T., Chau-Wong, M., Tedesco, J., and Wong, K.
(1976) Proc. Natl. Acad. Sci. U. S. A. 73, 4354-4358 Lefkowitz, R. J., Mukherjee, C., Coverstone, M., and Caron, M.
G. (1974) Biochem. Biophys. Res. Commm 60, 703-710 Aurbach, G. D., Fedak, S. A., Woodard, C. J., Palmer, J. S.,
Hauser, D., and Troxler, F. (1974) Science 186, 1223-1224 Hammes, G. G., and Schimmel, P. R. (1970) in The Enzymes,
Vol. 2, (Bayer, P. D., ed) pp. 67-114, Academic Press, New York
Saavedra, J. M., Palkovitz, M., Brownstein, M. J., and Axelrod, J. (1974) Nature 248, 695-696
Cedarbaum, J. M., and Aghajanian, G. K. (1976)Brain Res. 112, 413-419
36. Lew, J. Y., Matsumoto, Y., Pearson, J., Goldstein, M., Hokfelt, T., and Fuxe, K. (1977) Brain Res. 119, 199-210
37. Moreno, H., Sineriz, F., and Farias, R. N. (1974) J. Biol. Chem. 249, 7701-7706
38. Mukherjee, C., Caron, M. G., and Lefkowitz, R. J. (1975) Proc. Natl. Acad. Sci. U. S. A. 72, 1945-1949
39. Mickey, J., Tate, R., and LeIkowitz, R. J. (1975) J. Biol. Chem. 250, 5727-5729
40. Kebabian, J. W., Zatz, M., Romero, J. A., and Axelrod, J. (1975) Proc. Natl. Acad. Sci. U. S. A. 72, 3735-3739
41. Mukherjee, C., and Lefkowitz, R. J. (1976) Proc: Natl. Acad. Sci. U. S. A. 73, 1494-1498
42. Schramm, M., Feinstein, H., Naim, E., Lang, M., and Lasser, M. (1972) Proc. Natl. Acad. Sci. U. S. A. 69, 523-567
43. Brunton, L. L., Wiklund, R. A., Van Arsdale, P. M. and Gilman, A. G. (1976) J. Biol. Chem. 251, 3037-3044
44. Whitehead, E. P. (1970) Prog. Biophys. Mol. Biol. 21, 321-397 45. Rang, H. P., and Ritter, J. M. (1970) Mol. Pharmacol. 6,357-382
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D C U'Prichard and S H SnyderBinding of 3H-catecholamines to alpha-noradrenergic receptor sites in calf brain.
1977, 252:6450-6463.J. Biol. Chem.
http://www.jbc.org/content/252/18/6450.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/252/18/6450.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from