bestandsdichte und vergesellschaftung brütender wasservögel südschwedens in beziehung zur...
TRANSCRIPT

Heft 1 ] 1985 ] 85
J. Orn. 126, 1985: S. 8 5 - 9 2
Bestandsdichte und Vergesellschaftung brütender Wasservögel Südschwedens in Beziehung zur Produktivität der Seen
Von Leif Nilsson
Im Unterschied zu Zählungen rastender Wasservögel sind bisher nur wenige Be- standsaufnahmen von brütenden Wasservogelbeständen publiziert worden (HAAPANeX & NlCSSON 1979). In den letzten Jahren wurden mehrere lokale Untersuchungen in Fennoskandien durchgeführt (z. B. DaNEtL & SJöBeRG 1978, 1979, HAAPANeN 1973, I-IAAeANEN & PAAS~VlRTA 1973, NILSSON 1978, NtLSSON & NILSSON 1978). Die meisten schwedischen Untersuchungen betrafen nur eine Lokalität als Beitrag zur Planung von Naturschutzmaßnahmen. 1975 wurde eine Untersuchung mehrerer Seen im Bezirk Stockholm durchgeführt, die nicht nur ornithologische, sondern auch botanische und limnologische Aspekte umfaßte. Die Ergebnisse sollen die Beziehungen zwischen Be- standsdichte und Zusammensetzung brütender Wasservogelbestände und verschiede- nen Habitatfaktoren beleuchten. Ein Bericht über die Bestandsaufnahme ist schon pu- bliziert (NILSSON 1979), doch ohne weiterführende Analysen.
Untersuchungsgebiet Der Bezirk von Stockholm ist zum großen Teil ein alter Schärenhof, der sich seit der letzten
Eiszeit aus dem Meer erhoben hat. Diese Hebung dauert noch an. Zwei Formationen lassen sich unterscheiden: die höheren Gebiete mit Moränenablagerungen und die Täler mit Meeressedimen- ten. Die Sedimente sind nährstoffreich , die Seen eutroph. Die Seen in den Moränengebieten sind dagegen von Natur aus oligotroph. Mehrere dieser oligotrophen Seen sind jedoch durch Abwäs- ser eutrophiert worden und müssen jetzt als mesotroph bezeichnet werden.
Bestandsaufnahmen liegen von 77 kleinen (< 5 km ~) und zwei großen eutrophen sowie von 13 kleinen und vier großen oligotrophen Seen vor. Ferner wurden 310 km 2 vom Mälarsee, dem drittgrößten See Schwedens, untersucht. Der Mälarsee umfaßt sowohl eutrophe Buchten als auch Schärenhöfe. Die meisten Teile des Mälarsees müssen als eutroph bezeichnet werden.
Methodik Alle brütenden Wasservögel (Taucher, Enten, Gänse, Schwäne und Blässhuhn) wurden nach
einer standardisierten Methode (Statens Naturvärdsverk 1978, NILSSON 1979) gezählt, bei brüten- den Enten die stationären Paare (Auswertungsmethodik s. NIcssoN 1979, 1981). Die anderen Ar- ten wurden durch eine Kombination von Beobachtungen revierbesitzender Paare mit Nestersuche ermittelt.
Die folgenden wasserchemischen Faktoren wurden nach Standardverfahren registriert: Total- gehalt Phosphor, Totalgehalt Stickstoff, Alkalinität, pH-Wert, Leitfähigkeit, Gehalt an a-Chloro- phyll. Die Ausdehnung der Ufervegetation wurde aus Vegetationskarten nach Luftfotos berech- net. Für jeden See wurden auch Uferlänge (nach den Vegetationskarten) und Wasserfläche ge- messen. Ein Uferindex wurde durch Division der Uferlänge mit der Wasserfläche berechnet. Voll- ständige Datenreihen liegen von 38 Seen vor und gingen in multiple Regressionsanalysen ein. Zu
[J. Orn. 86 L. NiLsso.x [ 126
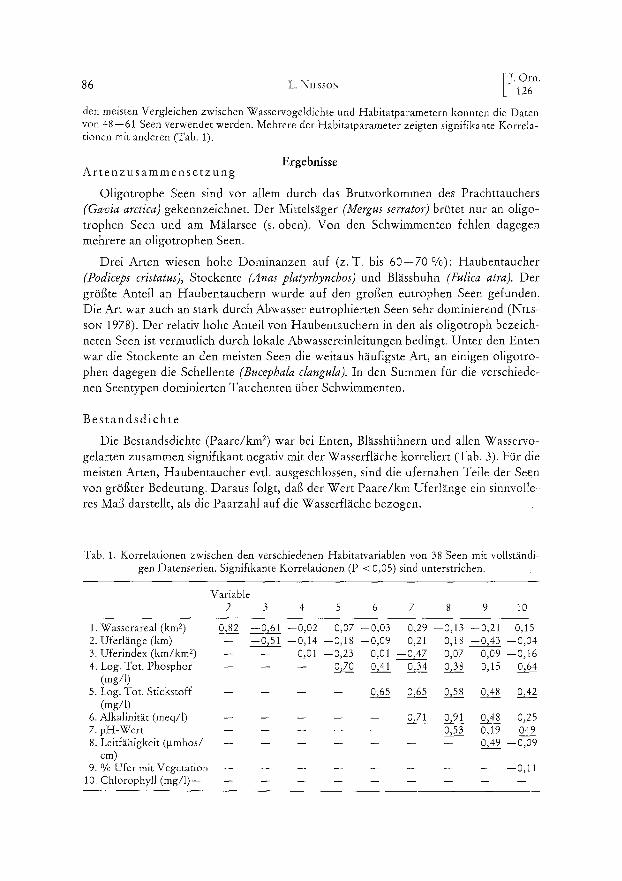
den meisten Vergleichen zwischen Wasservogeldichte und Habitatparametern konnten die Daten von 48--61 Seen verwendet werden. Mehrere der Habitatparameter zeigten signifikante Korrela- tionen mit anderen (Tab. 1).
Ergebnisse A r t e n z u s a m r n e n s e t z u n g
Oligotrophe Seen sind vor allem durch das Brutvorkornrnen des Prachttauchers (Gavia arctica) gekennzeichnet. Der Mittelsäger (Mergus serrator) brütet nur an oligo- trophen Seen und am Mälarsee (s. oben). Von den Schwimrnenten fehlen dagegen mehrere an oligotrophen Seen.
Drei Arten wiesen hohe Dorninanzen auf (z.T. bis 60--70 %): Haubentaucher (Podiceps cristatus), Stockente (Anas platyrhynchos) und Blässhuhn (Fulica atra). Der größte Anteil an Haubentauchern wurde auf den großen eutrophen Seen gefunden. Die Art war auch an stark durch Abwasser eutrophierten Seen sehr dorninierend (NILs- SON 1978). Der relativ hohe Anteil von Haubentauchern in den als oligotroph bezeich- neten Seen ist vermutlich durch lokale Abwassereinleitungen bedingt. Unter den Enten war die Stockente an den meisten Seen die weitaus häufigste Art, an einigen oligotro- phen dagegen die Schellente (Bucephala clangula). In den Summen für die verschiede- nen Seentypen dominierten Tauchenten über Schwirnmenten.
B e s t a n d s d i c h t e
Die Bestandsdichte (Paare/krn 2) war bei Enten, Blässhühnern und allen Wasservo- gelarten zusammen signifikant negativ mit der Wasserfläche korreliert (Tab. 3). Für die meisten Arten, Haubentaucher evtl. ausgeschlossen, sind die ufernahen Teile der Seen von größter Bedeutung. Daraus folgt, daß der Wert Paare /km Uferlänge ein sinnvolle- res Maß darstellt, als die Paarzahl auf die Wasserfläche bezogen.
Tab. 1. Korrelationen zwischen den verschiedenen Habitatvariablen von 38 Seen mit vollständi- gen Datenserien. Signifikante Korrelationen (P < 0,05) sind unterstrichen.
Variable 2 3 4 5 6 7 8 9 10
1. Wasserareal (km 2) 0,.8.2 --9,61 --0,02 0,07 --0,03 0,29 --0,13 --0,21 0,15 2. Uferlänge(km) -- --0»51 --0,14 --0,18 --0,09 0,21 0,18 --0,43 --0,04 3. Uferindex (km/km 2) -- -- 0,01 --0»23 0,01 --0,47 0,07 0,09 --0,16 4. Log. Tot. Phosphor -- -- -- 0»70 0»41 0»34 0~38 0,15 0»64
(mg/l) 5. Log. Tot. Stickstoff . . . . 0»65 0»65 0»58 0»48 0»42
(mg/l) 6. Alkalinität (meq/1) . . . . . 0»71 0~91 0»48 0,25 7. pH-Wert . . . . . . 0»53 0,19 04.__29 8. Leitfähigkeit (•mhos/ . . . . . . . 0,49 --0,09
cm) 9. % Ufer mit Vegetation . . . . . . . . . 0,11
10. Chlorophyll (mg/I) . . . . . . . . . .
Heft 1 ] 1985 ] Wasservögel und Produktivität der Seen 87
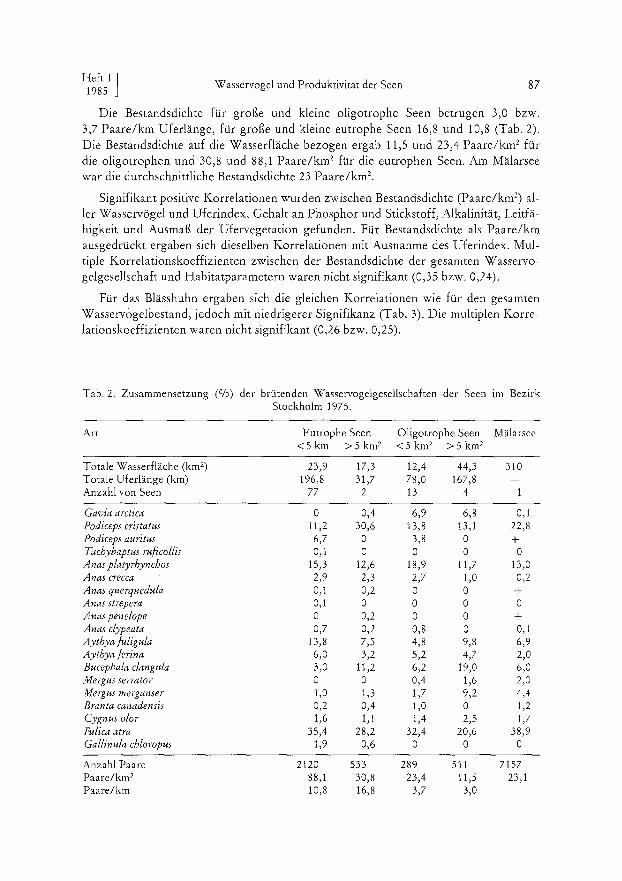
Die Bestandsdichte für große und kleine oligotrophe Seen betrugen 3,0 bzw. 3,7 Paare/km Uferlänge, für große und kleine eutrophe Seen 16,8 und 10,8 (Tab. 2). Die Bestandsdichte auf die Wasserfläche bezogen ergab 11,5 und 23,4 Paare/km 2 für die oligotrophen und 30,8 und 88,1 Paare/km 2 für die eutrophen Seen. Am Mälarsee war die durchschnittliche Bestandsdichte 23 Paare/km 2,
Signifikant positive Korrelationen wurden zwischen Bestandsdichte (Paare/km 2) al- ler Wasservögel und Uferindex, Gehalt an Phosphor und Stickstoff, Alkalinität, Leitfä- higkeit und Ausmaß der Ufervegetation gefunden. Für Bestandsdichte als Paare/km ausgedrückt ergaben sich dieselben Korrelationen mit Ausnahme des Uferindex. Mul- tiple Korrelationskoeffizienten zwischen der Bestandsdichte der gesamten Wasservo- gelgesellschaft und Habitatparametern waren nicht signifikant (0,35 bzw. 0,24).
Für das Blässhuhn ergaben sich die gleichen Korrelationen wie für den gesamten Wasservogelbestand, jedoch mit niedrigerer Signifikanz (Tab. 3). Die multiplen Korre- lationskoeffizienten waren nicht signifikant (0,26 bzw. 0,25).
Tab. 2. Zusammensetzung (%) der brütenden Wasservogelgesellschaften der Seen im Bezirk Stockholm 1975.
Art Eutrophe Seen Oligotrophe Seen Mälarsee < 5 k m >5km 2 <5km 2 >5km 2
Totale Wasserfläche (km 2) 23,9 17,3 12,4 44,3 310 Totale Uferlänge (km) 196,8 31,7 78,0 167,8 -- Anzahl von Seen 77 2 13 4 1
Gavia arctica 0 0,4 6,9 6, 8 O, 1 Podiceps cristatus 11,2 30,6 13,8 13,1 22,8 Podiceps auritus 6,7 0 3,8 0 + Tachybaptus ruficollis 0,1 0 0 0 0 Anasplatyrhynchos 15,3 12,6 18,9 11,7 13,0 Anas crecca 2,9 2,3 2,7 1,0 0,2 Anas querquedula 0,1 0,2 0 0 + A rias strepera O, 1 0 0 0 0 Anas penelope 0 0,2 0 0 + Anas clypeata 0,7 0,2 0,8 0 0,1 Aythyafuligula 13,8 7,5 4,8 9,8 6,9 Aythya/erina 6,0 3,2 5,2 4,7 2,0 Bucephala clangula 3,0 11,2 6,2 19,0 6,0 Mergus serrator 0 0 0,4 1,6 2,0 Mergus merganser 1,0 1,3 1,7 9,2 4,4 Branta canadensis 0,2 0,4 1,0 0 1,2 Cygnus olor 1,6 1,1 1,4 2,5 1,7 Fulica atra 35,4 28,2 32,4 20,6 38,9 Gallinula chloropus 1,9 0,6 0 0 0
Anzahl Paare 2t20 533 289 511 7157 Paare/km 2 88,1 30,8 23,4 11,5 23,1 Paare/km 10,8 16,8 3,7 3,0 --
88
-O
g o ~ d
~ V
~ o < d ~ z v
~ d
mp_.~
~ u
~.~
N ~
O
8 M
i
e~
8--.
L. NILSSON
o ~ o o ~ o o o * o ~ o * o
0o co
c:;" o o ~ o ~ o ~ o ~ c ; o ~ o ~ o
I
o ~ o o o ~ o ~ o ~ o ~ o ~ o o
I /
o o o o * o ~ o ~ o ~ o ~ o ~ o ~
I I I
(N
o ~ o o o ~ o ~ o ~ o ~ o ~ o ~ o ~ I
c ~ o o ~ o o ~ o ~ c ~ o ~ o ~ o
I I
o o o c ~ o o * c ~ o * o o
I I I
eq e~ eth ~ eq e~ o "~- er~
o ~ o o o ~ c~ o~d o ~ o ~ o ~ I I I
o c ~ c ~ c ? ~ c ~ ~ o ~ o I I
8 o
j o ~ o ~ o ~ o ~ o ~ o ~ o ~ c ~
I I
©
,1.} ;~ oc - -~00 -~ , -~ 0 . . ~
~ ~- ~0~~Do~o~,%D ~ - ~ - ~ ° ~0~ ~ ~ 3 3 ~ ' ~ ~ ' ~ ° ~0<~-o-- ~>{D~-~
J. Orn. 126

Heft 1 ] 1985 J Wasservögel und Produktivität der Seen 89
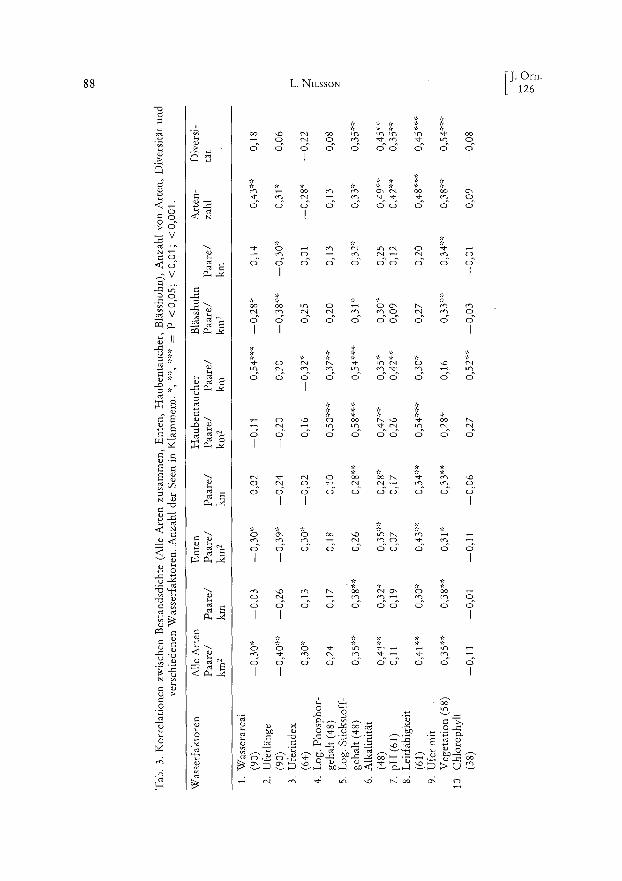
Für den Haubentaucher waren die meisten Korrelationen positiv und signifikant (Tab. 3) mit Ausnahme von Wasserfläche (Paare/km2), Uferlänge, Uferindex (Paare/ km2), Ausmaß der Ufervegetation (Paare/km) und Gehalt von Chlorophyll (Paare/ km2). Die meisten dieser Korrelationskoeffizienten waren jedoch positiv. Die multiplen Korrelationskoeffizienten zwischen Bestandsdichte der Haubentaucher (Paare/km 2, Paare/km) und Habitatparametern waren signifikant (0,36 ,:.:»* bzw. 0,62::'**) mit den Logarithmen der Totalgehalte von Phosphor und Stickstoff als signifikanten Variablen in beiden Fällen. Für die Bestandsdichte als Paare/km gemessen treten auch die Was- serfläche und die Uferlänge als signifikante Variable in die multiple Korrelation ein.
Für die Enten wurde eine hochsignifikante Korrelation zwischen Uferindex und Paare/km 2 für Seen > 0,10 km 2 gefunden, dazu signifikante Korrelationen mit Alkali- nität, Leitfähigkeit und Ausmaß der Ufervegetation. Die multiplen Korretationskoeffi- zienten waren nicht signifikant (0,31 bzw. 0,25).
A r t e n z a h l
19 Wasservogelarten wurden als Brutvögel auf den Seen nachgewiesen, zwischen 1 und 13 für Seen > 10 ha.
Die Artenzahl war positiv und signifikant mit der Größe der Seen korreliert. Signi- fikante Korrelationen wurden auch mit verschiedenen Wasserfaktoren gefunden, doch nicht mit dem Gehalt an Phosphor und Chlorophytl. Der multiple Korrelationskoeffi- zient war signifikant (0,53***) mit Wasserfläche, log. Totalgehalt Stickstoff, Alkalini- rät und Ausmaß der Ufervegetation als signifikante Variable in der Regressionsglei- chung.
Nimmt man Artenzahl und die Wasserfläche als Logarithmen, erhält man folgende Gleichung:
log (Artenzahl) = 1,76 + 0,16 log (Wasserfläche in km 2)
D ive r s i t ä t
Die Diversität der brütenden Wasservogelgesellschaften wurde mit der Shannon- funktion (H' = -- E loge Pi; Pi ~ der relative Anteil der Art i an der Gesamtheit) ge- messen. Sie war signifikant und positiv mit Totalgehalt am Stickstoff, Leitfähigkeit, A1- kalinität, pH-Wert und Ausmaß der Ufervegetation korreliert. Wenn man Seen < 10 ha ausscheidet, wird die Korrelation zur Wasserfläche signifikant, während die Korrelation mit dern Ausmaß der Ufervegetation nicht signifikant ist. Der multiple Korrelationskoeffizient war signifikant (0,34':) mit Totalgehalt an Stickstoff, Alkalini- tät und Ausmaß der Ufervegetation als Variable.
Diskussion
Diese Untersuchung wie auch jene von NILsso~N (1978) und NILSSON &; NILSSON (1978) zeigen deutlich, daß die Bestandsdichte von Wasservögeln mit zunehmendem Eutrophierungsgrad zunimmt (s. auch DANELL & SJö~ERG 1978, HAAPANEN & NILSSON

[j. Orn. 90 L. NILSSON [ 126
1979). Für den Haubentaucher gilt dies für das ganze Spektrum untersuchter Seen, während die positive Beziehung zwischen Eutrophierungsgrad und Bestandsdichte bei Entenvögeln nur für niedrige Eutrophierungsgrade vorhanden ist. So waren auch keine solchen Korrelationen für Entenvögel in den von NILSSON (1978) untersuchten hoch- eutrophen Seen vorhanden. In einer Untersuchung in Kanada (PATTERSSON 1976) wurde keine signifikanten Beziehungen zwischen Wasservogeldichte und wasserchemi- schen Variablen im Frühling gefunden, während die gleichen Beziehungen im Sommer signifikante Korrelationen aufwiesen. Für den Prachttaucher berichteten NILSSON & NILSSON (1978) von einer negativen Korrelation mit zunehmendem Phosphorgehalt.
Die zunehmende Eutrophierung von Seen führt zu einem Anwachsen der Produk- tion von Planktonalgen und zu einer Änderung der Primärproduktion von der Domi- nanz submerser Vegetation zur Dominanz des Phytoplanktons (WETZEL 1975). Mit zu- nehmender Eutrophierung und Dominanz von Phytoplankton wächst die Dominanz von Cypriniden in der Fischfauna (ANDERSSON 1981) und die Bestandsdichte der Fi- sche. Diese Verhältnisse sind für den Haubentaucher günstig und können die positive Korrelation zwischen der Bestandsdichte von Haubentauchern und Phosphorgehalt (als Indikator des Eutrophierungsgrades) erklären.
Wasserinsekten sind von großer Bedeutung für brütende Entenvögel besonders während der Eiablage, Bebrütung und als Jungennahrung (DANELL & SJöRERC 1980, ERIKSSON 1976, KRAFU 1981, PEHRSSON 1979). Die Reduktion der submersen Vegeta- tion ist für die Wasserinsekten ungünstig. Die Zunahme der Fischbestände führt dann zu einer Konkurrenzsituation zwischen Fischen und Entenvögeln (ANDERSSON 1982, EAmE & KEAST 1982, E~IKSSON 1979). ERIKSSON (1983) meint, daß eine solche Konkur- renz für die Schellente von Bedeutung ist, wenn sie Nahrung im freiem Wasser sucht, während die Konkurrenz zwischen Fischen und Stock- bzw. Krickente (Anas crecca) keine Bedeutung hat, weil diese Arten nur in seichtem Wasser Nahrung suchen. Ein zu hoher Eutrophierungsgrad mit dichtem Fischbestand ist also für die Enten, speziell für Tau&enten, ungünstig.
Die Bestandsdichte von rastenden und überwinternden Wasservögeln ist ebenfalls mit der Produktivität der Seen positiv korreliert (UTscHmK 1976, 1980), mit Hauben- taucher als geeignete Indikatorart für den Eutrophierungsgrad der Gewässer.
Hohe Artenzahl ist meistens charakteristisch für Ökosysteme von höherer Produkti- vität, weil diese die Koexistenz mehrerer Arten zulassen (MAcARTHUR 1970). In Über- einstimmung hiermit sind Korretationen zwischen Artenzahl und Diversität und ver- schiedenen produktivitätsindizierenden Wasserfaktoren signifikant (vgl. auch NILssoe,- & NILSSON 1978). Für abwassereutrophierte Seen Südschwedens wurde jedoch ab-
nehmende Diversität mit zunehmender Eutrophierung gefunden (vgl. UTSCHICK 1976 für tastende Wasservögel). Die Diversität nimmt also zunächst mit zunehmender Eu- trophierung zu und dann wieder in hocheutrophen Seen ab. Geringe Mengen von Ab- wasser in einem ursprünglich oligotrophen See führen zu einem Wasservogelbestand, der jenem natureutropher Seen ähnelt. Mit zunehmender Eutrophierung fallen die mei- sten Arten aus; Haubentaucher und ein bis zwei andere Arten, wie Stockente und Blässhuhn, erhalten hohe Dominanzwerte.

Heft19851 ] Wasservögel und Produktivität der Seen 91
Zusammenfassung 1975 wurden die Brutbestände von Wasservögeln (Seetaucher, Lappentaucher, Enten, Gänse,
Schwäne, Blässhuhn) in 79 eutrophen und 17 oligotrophen Seen verschiedener Größe im Bezirk Stockholm, Südschweden, gezählt. Bestandsdichten für den gesamten Wasservogelbestand, für Enten, Haubentaucher und Blässhuhn waren mit verschiedenen Wasserfaktoren signifikant und positiv korreliert. Ähnliche Korrelationen wurden zwischen Artenzahl, Diversität und verschiede- nen Wasserfaktoren gefunden. Signifikante multiple Korrelationen wurden zwischen Bestands- dichte des Haubentauchers, Artenzahl, Diversität und verschiedenen Wasserfaktoren gefunden. Entscheidende Bedeutung kommt dem Eutrophierungsgrad der stehenden Gewässer zu.
Summary C o m p o s i t i o n and d e n s i t y o f b r e e d i n g i n i a n d w a t e r f o w l c o m m u n i t i e s in s o u t h -
e rn S w e d e n i n r e l a t i o n t o l a k e p r o d u c t i v i t y
In 1975, breeding populations of waterfowl (i. e. divers, grebes, ducks, geese, swans, coot and moorhen) were censused in 79 eutrophic and 17 oligotrophic lakes of varying size within the county of Stockholm, south Sweden, in addition to 310 km 2 of Lake Mälaren, the third largest lake in Sweden. This report analyses the composition and density of the waterfowl communities of these lakes in relation to the size and configuration of the lakes and in relation to some factors indicative of the productivity. Total waterfowl density, the density of breeding Great Crested Grebes, ducks and coot were all found to be positively and significantly correlated with various factors indicative of lake productivity. Similar correlations were also found between the number of breeding waterfowl species and diversity and the same environmental factors. Significant mul- tiple correlations were found between the density of hreeding Great Crested Grebes and the water factors as weil as for number of breeding waterfowl species and diversity.
Literatur ANDERSSON, G. (1981): Fiskars inverkan p~t sjöfägel och fägelsjöar. Anser 20: 21--34. • ANDERS- SON, Ä. (1982): Competition between Fishes and Waterfowl. In: Scoa~r, D. A. (ed.) Managing Wetlands and Their Birds, 248--249. IWRB, Slimbridge, UK. • DANELL, K., & S3öBERG, K. (1978): Häbitat selection by breeding ducks in boreal lakes in northern Sweden. Swedish Wildl. Res. 10: 161--190. • Dies. (1979): Abundance and productivity of ducks on boreal lakes in nor- thern Sweden. Arm. Z ool. Fennici 16: 123--128. • Dies. (1980): Foods of Wigeon, Teal, Mallard and Pintail during the summer in a Northern Swedish Lake. Swedish Wildl. Res. 11 : 139-- 167. • EÆt)IE, J. McA., & KEAST, A. (t982): Do Goldeneye and Perch compete for food? Oecologia 55: 225--230. • ERIKSSON, M. O. G. (1976): Food and feeding habits of downy Goldeneye Bucephala clangula duck]ings. Ornis Scand. 7: 159--169. • Ders. (1979): Competition between freshwater fish and Goldeneye Bucephala clangula for common prey. Oecologia 41: 99-107. • Ders. (1983): The role of fish in the selection of lakes by non-piscivorous Anatids: Mallard, Teal and Golden- eye. Wildfowl 34: 89--98. • H~VANEN, A. (1973): Inland and coastal waterfowl census in Fin- land. A review. Finnish Garne Res. 33: 3--11. • H~PANEN, A., & NILSSON, L. (1979): Breeding waterfowl populafions in northern Fennoscandia. Ornis Scand. 10: 145--219. • H~PaNE~', A., & P~SlWRTa, O. (1973): The waterfowl in eutrophic waters of south Finland. Finnish Garne Res. 33: 13--26. • K~pu, G. L. (1981): The role of nutrient reserves in mallard reproduction. Auk 98: 29738. • MA¢A~tTHUR, R. (1970): Species packing and competitive equilibrium for many species. Theor. Popul. Biol. 1: 1--11. • N~LSSON, L (1978): Breeding waterfowl populations in eutrophicated waters of south Sweden. Wildfow129:101--110. • Ders. (1979): Fäglar i sjöar och vätmarker. Natur i Stockholms län. Naturresursinventeringen. Stockholms läns landsting. Re- gionplanekontoret. Stockholm. • NILSSON, S. G. (1981): Census methods for breeding ducks in south Sweden. Proc. Second Nordic Congr. Orn. 1979:118--126. Stavanger. • NILsso~,', S. G., & NILSSON, I. (1978): Breeding bird community densities and species richness in lakes. Oikos 31: 214--221. • PATrERSSON, J. H. (1976): The role of environmental heterogenity in the regulation of

[J. Orn. 92 L. NILSSON [ 126
duck populations. J. Wildl. Mgmt. 40: 22--32. • PEHRSSON» O. (1979): Feeding behaviour, feed- ing habitat utilization, and feeding efficiency of Mallard ducklings (Anas platyrhynchos L.) as guided by a domestic duck. Swedish Wildl. Res. 10: 191--218. • Statens Naturvärdsverk (1978): Biologiska Inventeringsnormer. Fäglar. Stockholm • UTSGHICK» H. (1976): Die Wasservögel als Indikatoren für den ökologischen Zustand von Seen. Verh. orn. Ges. Bayern 22: 393--438. • Ders. (1980): Wasservögel als Indikatoren für die ökologische Stabilität südbayerischer Stauseen. Verh. orn. Ges. Bayern 23: 273--345. • WSTZEL, R. G. (1975): Limnology. W. B. Saunders Com- pany. Philadelphia.
Anschrift des Verfassers: Ekologihuset, S-223 62 Lund, Schweden