avifauna in oak woodlands of the willamette valley, oregon
TRANSCRIPT

Avifauna in Oak Woodlands of the Willamette Valley, OregonAuthor(s): Joan C. Hagar and Mark A. SternSource: Northwestern Naturalist, Vol. 82, No. 1 (Spring, 2001), pp. 12-25Published by: Society for Northwestern Vertebrate BiologyStable URL: http://www.jstor.org/stable/3536642 .
Accessed: 16/06/2014 15:36
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Society for Northwestern Vertebrate Biology is collaborating with JSTOR to digitize, preserve and extendaccess to Northwestern Naturalist.
http://www.jstor.org
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

NORTHWESTERN NATURALIST 82:12-25 SPRING 2001
AVIFAUNA IN OAK WOODLANDS OF THE WILLAMETTE VALLEY, OREGON
JOAN C. HAGAR
Pacific Wildlife Research, 1521 NW Harrison Boulevard, Corvallis, Oregon 97330 USA
MARK A. STERN The Nature Conservancy of Oregon, 821 SE 14th Avenue, Portland, Oregon 97214 USA
ABSTRACT-Oregon white oak (Quercus garryana) woodlands are an important habitat for breeding birds that is threatened by development and altered disturbance regimes. We de- scribed habitat characteristics and examined species composition and abundance of breeding birds in 9 oak woodland sites in the Willamette Valley, Oregon during May and June, 1994 to 1996. Based on comparisons to earlier studies, our results suggest that avian community com- position has changed in recent decades. Changes in species composition likely have been as- sociated with a shift from open- to closed-canopy habitats, resulting from successional ad- vancement in the absence of fire. Although there is some evidence that avian species composi- tion has changed to more closely resemble that of closed-canopy, conifer-dominated habitats, oak woodlands in the Willamette Valley nonetheless support both neotropical migrant and res- ident bird species that are uncommon in or absent from coniferous habitats in western Oregon. We identified 12 species currently occurring in oak woodlands that may be negatively affected by the replacement of oak habitats with closed-canopy, conifer-dominated forest. Because co- nifers dominate much of the forested regions of western Oregon, Willamette Valley oak wood- lands provide an important element of regional diversity and should receive priority consid- eration in conservation planning. The conservation and management of semi-open woodlands with large-diameter oaks may be particularly critical for some species that have declined in abundance and/or do not occur in dense forests.
Key words: avifauna, birds, oak woodlands, succession, conservation of biodiversity, Quercus garryana, Willamette Valley, Oregon
Oregon white oak woodlands in western Oregon have been altered in extent and distri- bution over the past decades by human activi- ties. Prior to European settlement, prairie and oak savanna vegetation communities were maintained in the Willamette Valley by burning regimes practiced by Native Americans (Frank- lin and Dryness 1988). Regular burning ceased with the establishment of Euro-American set- tlements, and tree species not adapted to fire were able to encroach upon savanna habitats. In the continued absence of fire, succession has advanced from prairie and oak savanna to closed-canopy woodland (Franklin and Dry- ness 1988; Johannessen and others 1971), es-
pecially on the steep slopes of the valley mar-
gins (Towle 1982). Shade from the dense can-
opy that forms in closed oak stands prevents regeneration of oak, but more shade tolerant tree species, such as Douglas-fir (Pseudotsuga
menziesii) and bigleaf maple (Acer macrophyl- lum), are able to grow in the understory. Thus, succession is replacing oak with climax plant communities of conifer-types or bigleaf maple (Franklin and Dryness 1988). Furthermore, much of the land base on which Oregon white oak occurs or used to occur is privately owned and has been developed for agriculture or
housing. Where oak communities remain, they tend to take the form of closed-canopy forests.
Today, the valley is primarily agricultural land and urban developments, and open oak wood- lands are extremely rare (Towle 1982). Similar trends in conversion and loss of oak habitats also are occurring in western Washington (Lar- sen and Morgan 1998).
Changes in the extent and character of oak woodlands may represent a significant threat to regional biodiversity because oaks provide important and unique habitat for wildlife
12
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 13
(Ryan and Carey 1995), particularly birds (Ver- ner 1980). In 1 of the few published studies on wildlife in western Oregon oak habitats, An- derson (1972) found that avian species diver- sity was greater in oak than in Douglas-fir and hemlock habitats. In addition, he found that oak woodlands supported a higher proportion of neotropical migrant species during the breeding season than did the conifer types. There is widespread concern over declining populations of neotropical migrants (Finch and Stangel 1992), and oak woodlands in Oregon and Washington have been identified as high priority habitats for monitoring and manage- ment of this group of species (Partners In Flight 2000). Furthermore, several resident species, including acorn woodpecker (scientific names of birds observed in this study are given in Ap- pendix B) and white-breasted nuthatch, are strongly associated with oaks for foraging and nesting habitats in western Oregon (Cross and Simmons 1983). The extirpation of Lewis' woodpecker (Melanerpes lewis) from the Wil- lamette Valley may be related to the loss of ma- ture oak habitat (Marshall and others 1992) that this species used for nesting, roosting and acorn storage (Galen 1989).
In spite of the importance of oak habitat to birds, little work has been done to monitor avi- an communities in Willamette Valley wood- lands. The only systematic surveys of avian communities in oak habitats in the Willamette Valley of which we are aware were done in 1967 to 1968 by Anderson (1970, 1972), who sur- veyed birds in 5 oak stands in the west central Willamette Valley (Fig. 1). It is not known which species may have changed in abundance as a result of changes in the vegetative com- position and structure which oak woodlands have undergone through the process of succes- sion. In order to foresee and prevent further ex- tirpations of species such as Lewis' woodpeck- er, information on the current community com- position and on the abundance and habitat re- lationships of oak-associated species is critical. The primary objectives of this project were to describe the species composition of bird assem- blages currently occurring in oak woodlands in the Willamette Valley, and to qualitatively com- pare it to Anderson's (1970) recent historical as- semblage. An additional objective was to qual- itatively compare the species composition of bird communities in oak woodlands to that in
conifer forests in western Oregon in order to formulate hypotheses about the consequences of succession of oak woodlands to conifer- dominated forests.
METHODS
Study Areas
The Willamette Valley lies between the Oregon Coast and Cascade Ranges. Winters in the valley are mild and wet, and summers are hot and dry. Oregon white oak is the dominant tree species in the valley, occurring in forest stands, groves, and as scattered, individual trees in savannas and agricultural fields. Big- leaf maple, Douglas-fir, and Pacific madrone (Arbutus menziessi) also are common. California black oak (Q. kellogii) reaches the northern limit of its range in the southern portion of the Wil- lamette Valley, in Lane County. Poison oak (Rhus diversiloba) is the dominant understory component in most oak woodlands, especially on the driest sites (Franklin and Dryness 1988). Other shrubs typically associated with oak communities include snowberry (Symphocarpus albus), hazel (Corylus cornuta), serviceberry (Amelanchier alnifolia), and swordfern (Polysti- chum munitum). Because poison oak is less pal- atable to livestock than other understory plants, it typically replaces these species on heavily grazed sites and may maintain domi- nance even after grazing has stopped (Thilen- ius 1968).
Potential study sites were initially identified from a precursor of the Willamette Land Use/ Land Cover map (Klock and others 1998). We selected sites dominated by oak (>50% oak) and >20 ha in size to allow for sampling of in- terior stand condition, free from edge effect. We visited potential study sites identified from the map, and selected a final set that repre- sented the range of structure (for example, tree size and density) characteristic of Willamette Valley oak woodlands. Criteria for final selec- tion also included the ability to attain permis- sion to access a site. Most of the land in the val- ley is privately owned, making both the iden- tification of and access to potential study sites difficult. The majority of our study sites were therefore located on public land.
The 9 sites selected were located between 440 and 460 N latitude, roughly between the Co- lumbia River and Eugene (Fig. 1). All stands
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

14 NORTHWESTERN NATURALIST 82(1)
ft;
o0,. "..4? ?bo
' .?.?; AUB
. : .. . : " '
o,? I Irv
? 1 ,.
.
. . . .
? ". ,
"::.,.: ,, . -' " 0 5 10 Kiometers
EYQ ",,
... . ".? '
:fj. ".
:~,...?4 .--~ ?
rr"?.
FIGURE 1. Distribution of oak stands >5 ha as mapped by Oregon Department of Fish and Wildlife (Klock and others 1998) and the location of 8 south- ern-most oak stands sampled in 1994 to 1996 relative to sites studied in 1967 to 1968 by Anderson (1970) in the Willamette Valley. Inset shows location of cen- tral Willamette Valley in Oregon.
were dominated by Oregon white oak except the Eugene site, which had a significant com- ponent of California black oak in both the over- story and understory. The sites varied widely in other characteristics such as stand density and management history. The shapes and edge characteristics of the stands, as well as the land- scape setting around our study sites, varied widely. For example, Douglas-fir forests bor- dered 1 site at the western edge of the valley, whereas agricultural lands surrounded other sites, and another site was adjacent to a major river, the Columbia. Although we did not quan- tify habitat characteristics above the stand lev- el, the bird community we observed might have
been significantly influenced by the adjacent stands and surrounding landscapes.
Bird Counts
We used point counts (Ralph and others 1993) to record the species composition and to estimate relative abundance of individual spe- cies of breeding birds during at least 2 breeding seasons at each site. Point count stations within each stand were separated by -250 m, and were located -75 m from stand edges. We vis- ited each site 3 times per year, at evenly spaced intervals throughout the breeding season. At each point-count station all birds seen or heard during a 10-min count period were tallied by species and distance category (within or out- side of a 50-m radius of the observer). Surveys were conducted within 4 hr following sunrise, from the 3rd week in May through the end of June each year. We sampled 5 sites for 3 con- secutive breeding seasons, from 1994 through 1996. The remaining sites were sampled only in 1995 and 1996 (Appendix A).
Vegetation Sampling Data on habitat structure and composition
were collected at all point-count stations in all stands using the point-centered quarter meth- od (Mueller-Dumbois and Ellenberg 1974). Canopy cover, shrub cover, and basal area were estimated at 2 to 12 additional random plots in the largest stands. We estimated percent cover of canopy, subcanopy, and shrub layers of veg- etation, as well as of dominant shrub and tree species individually. In each quadrat of each plot we recorded the diameter of the tree clos- est to plot center that was >10 cm in diameter.
Data Analysis We calculated 2 community level indices: to-
tal abundance and species richness. The aver- age number of observations of all species com- bined per point-count station per visit served as our index of total abundance. We included only observations ?<50 m of observers. We com- bined data across years to calculate species richness using a rarefaction method to account for unequal sample sizes among sites (Ludwig and Reynolds 1988). We excluded non-native species from calculations of species richness. Species with only 1 observation in any stand over the entire period also were excluded from
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 15
calculations of species richness for those stands.
As a measure of the "commonness" of each bird species, we calculated frequency of occur- rence by summing the number of sites in which each bird species was observed in each year. We excluded the Sauvie Island WMA from our cal- culations of frequency of occurrence because this site was geographically separate from the sites in the interior Willamette Valley and spe- cies composition was notably different (see Re- sults and Discussion). The maximum frequen- cy of occurrence was 20 (4 sites sampled for 3 yr plus 4 sites sampled for 2 yr). We reported frequency of occurrence as a percentage of the maximum possible. We calculated an index of abundance (number of observations per point count station per visit, averaged over years) for each bird species with a frequency of occur- rence >25%.
We averaged habitat variables across all plots to derive a mean value for each variable in each stand. We examined normal probability plots and histograms of residuals to determine whether variables met statistical assumptions of normal distribution and constant variance (Sabin and Stafford 1990). Variables not meet- ing these assumptions were transformed with logarithmic- or square-root functions before being used in correlation analyses. Variables with badly skewed distributions that were not improved by transformations were not used in the analyses if the Shapiro-Wilk W test (Proc Univariate, SAS Institute Inc. 1990) indicated W < 0.80 (Sabin and Stafford 1990).
We limited our analysis of habitat relation- ships to species identified (a posteriori) as being associated with oak habitats, in order to reduce the probability of finding spurious significant correlations. We used 2 methods of analysis to relate variation in bird abundance and species richness among stands to differences in habitat structure and vegetation. For selected species with a frequency of occurrence ?75%, we cor- related abundance with habitat characteristics averaged at the stand level using Pearson cor- relation analysis (SAS Institute Inc. 1990). For species with 45 to 75% frequency of occurrence, we compared the average habitat characteris- tics between used and unused stands with the Kruskal-Wallis Chi-square approximation test (SAS Institute, Inc. 1990). Because this study is largely descriptive and exploratory, we report-
ed all habitat variables that were correlated with bird abundance at a level of significance of P < 0.10. Our relatively small sample size and the observational nature of this investiga- tion limit the interpretation of results from this analysis to the suggestion of hypotheses about bird-habitat relationships in oak woodlands.
Three of the sites we sampled corresponded very closely in location to 3 of those described by Anderson (1970, 1972; Fig. 1). In fact, An- derson's Stands 1 and 2 (1970) were the same stands as our Sites 3 and 4, and Anderson's Stand 4 was within /2 km of our Site 6. We be- lieved that a comparison between time periods, limited to these 3 sites, was valid because of correspondence in location and dominant tree species. We compared presence and absence of bird species in our study with those reported by Anderson (1970: Table 1) for his late spring and early summer sampling periods (aver- aged). In order to provide standard units for comparison, we calculated densities of bird species per 100 acres for those species that we observed but were not reported by Anderson (1970). However, we do not believe that the densities we calculated are precisely compa- rable to those calculated by Anderson (1970) because of different survey methods used. We calculated densities based on 50-m radius point counts (birds/visit/point-count station area), averaged over the 3 yr we sampled. Anderson calculated densities based on birds detected within 60 ft (18.3 m) of either side of transects, and may have underestimated densities rela- tive to our estimates. Therefore, we only report densities for bird species that were present in 1 study and not in the other for at least 2 sites.
RESULTS AND DISCUSSION
Vegetation and Habitat Oak canopy cover ranged from an average of
29.5% at the most open site to 84% at the dens- est site (Appendix A). One stand had a sub- stantial component of bigleaf maple in the can- opy and/or subcanopy, while 5 additional sites had minor maple components. About 2/3 of the sites had measurable (>1% cover) Douglas-fir in the canopy or subcanopy, but 2 sites had no measurable conifer component. Shrub cover ranged from 24% to 67% and was dominated by 5 species (in descending order of amount of cover: snowberry, poison oak, Rosa spp., ser- viceberry, and hazel).
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

16 NORTHWESTERN NATURALIST 82(1)
TABLE 1. Comparison of the density (birds/100 acres) of selected species between the present study (1994 to 1996) and a previous survey (1967 to 1968) by Anderson (1970; Anderson's site designations are given in parentheses) at 3 oak stands in the Willamette Valley, Oregon.
Site 3 (stand 1) Site 4 (stand 2) Site 6 (stand 4)
1967 to 1994 to 1967 to 1994 to 1967 to 1994 to 1968 1996 1968 1996 1968 1996
Species that have increased Mourning dove 0 1 0 0 0 4 Red-breasted sapsucker 0 19 0 7 0 6 Pacific-slope flycatcher 0 24 0 30 0 7 House wren 0 27 0 11 0 5 Winter wren 0 22 0 27 0 1 Cedar waxwing 1.5 14 0 3 0 4 European starling 0 13 0 0 0 1 Common yellowthroat 0 7 0 9 0 3 Black-headed grosbeak 0 19 0 7 0 10 Song sparrow 0 4 0 6 0 0 Purple finch 0 29 0 24 0 16 Steller's jay 0 1 0 0 0 14 Red-breasted muthatch 0 3 0 6 0 12 Swainson's thrush 0 67 0 54 0 5
Species that have decreased Yellow warbler 12 0 4 0 6 0 Chipping sparrow 12 0 8 0 24 0 Common bushtit 12 0 8 0 12 0
Bird Community The avian assemblage was comprised of 72
native and 3 non-native species (Appendix B). Total abundance averaged 13.8 birds/station/ visit (SE = 0.87, range: 9.8 to 16.3). The spotted towhee was the most abundant species ob- served, followed by western wood-pewee, or- ange-crowned warbler, and black-capped chickadee. Together these 4 species accounted for >30% of the total number of observations. European starlings, a non-native species, were observed at 5 sites, but Sauvie Island (Site 7) contributed the vast majority of observations (>65 %) of this species, probably because of the proximity to a large urban area. Sauvie Island also stood out as supporting a notably greater abundance of northern orioles, common yel- lowthroats, house wrens, scrub jays, and song sparrows than southern sites. In addition, black-throated gray warblers, dark-eyed jun- cos, Cassin's vireos, Swainson's thrushes, pur- ple finches, and western tanagers were abun- dant in most of the southern sites, but were ab- sent or rare on Sauvie Island (Appendix B). Most differences in bird community composi- tion between Sauvie Island and the southern sites were probably related to either low tree density on Sauvie Island (Appendix A) or ri-
parian influences, but these factors were con- founded in our study.
Based on a comparison of our work to that of Anderson (1970, 1972), assemblages of breed- ing birds in oak habitats have undergone changes in species composition over the past 3 decades. At the 3 sites that were common to our study and Anderson's, we detected 12 native and one introduced species that Anderson did not observe, and he detected 3 native species that we did not (Table 1). Although we were un- able to directly compare most habitat charac- teristics between our study sites and those of Anderson, it seems likely that an advancement of succession in oak woodland habitats has in- fluenced some of these changes in bird species composition. For example, using the same method (point quarter), we measured greater densities of trees than reported by Anderson (1970) at 2 sites (349 vs. 296 trees/ha and 715 vs. 138 trees/ha). Anderson (1970) did not re- port tree density for the 3rd common site. In- creased tree density may explain the relatively high abundance of Swainson's thrushes, Pacif- ic-slope flycatchers, Steller's jays, and winter wrens we observed, compared to their absence in Anderson's (1970, 1972) studies. These 4 spe- cies typically are associated with dense, closed-
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 17
TABLE 2. Habitat associations of selected bird species with a frequency of occurrence >45% in 9 Willamette Valley oak woodland sites, 1994 to 1996. Bird species listed are those that are most likely to be negatively affected by replacement of oak habitats with closed-canopy coniferous habitats based on a comparison of the results of this study with species lists from studies in coniferous habitats during the breeding season in western Oregon (Anderson 1972; Carey and others 1991; Gerig 1992; Chambers 1996; McGarigal and Mc- Comb 1992). R2 values are Pearson correlation coefficients.
Bird species Habitat descriptor R2
Mourning dove (No variables selected) - Downy woodpecker Subcanopy cover +0.60 0.09 Western wood-pewee Douglas-fir canopy cover -0.66 0.05 Scrub jay Canopy cover -0.65 0.06
Average tree diameter +0.63 0.07 Black-capped chickadee Oak subcanopy cover +0.65 0.06 White-breasted nuthatch Oak subcanopy cover -0.65 0.06
Average diameter of oak +0.65 0.06 Douglas-fir canopy cover -0.80 0.01
House wren Oak canopy cover -0.68 0.04 Average diameter of oak +0.60 0.08 Snowberry cover +0.70 0.04 Douglas-fir canopy cover -0.60 0.08
Bewick's wren (No variables selected) Cassin's vireo Average tree diameter -0.63 0.07 Cedar waxwing (No variables selected) Lazuli bunting Bigleaf maple cover - 0.02a American goldfinch (No variables selected) -
a Based on comparison of average cover of bigleaf maple between stands where Lazuli buntings were observed ( = 4.7%, SE = 2.9) and those where they were not (x = 0%).
canopy forests in western Oregon and are among the most abundant species in conifer- dominated forests of the Oregon Coast Range (Carey and others 1991; Chambers 1996; McGarigal 1993). Furthermore, species that are typically associated with semi-open wood- lands, such as chipping sparrows and common bushtits, were either absent from our study sites or were less abundant than in previous de- cades (Table 1).
In the absence of fire, the successional se- quence of vegetation in the Willamette Valley is likely to lead to a coniferous forest climax (Franklin and Dryness 1988:129). Hardwoods are relatively uncommon in the Tsuga hetero- phylla zone of western Oregon, and where they do occur, they are almost always subordinate (Franklin and Dryness 1988). Therefore, we hy- pothesize that bird communities in oak wood- lands will come to resemble those in conifer- dominated forests as succession proceeds. Our results suggest that bird species composition has indeed changed to more closely resemble that of closed-canopy coniferous habitats. However, oak woodlands still support a unique assemblage of both neotropical migrant and resident bird species. Twelve species that were
common at our sites were infrequently ob- served in closed-canopy coniferous forests in western Oregon (Table 2).
More species of breeding neotropical mi- grants occur in oak woodlands than in conif- erous habitats. We found more species of neo- tropical migrants in oak woodlands (26 spe- cies) than are listed from studies of avifauna in closed-canopy Douglas-fir forests in western Oregon at similar latitudes (Carey and others 1991, 17 species; Gerig 1992, 17 species). Neo- tropical migrant species that we observed in oak woodlands but that do not regularly occur in conifer forests include western wood-pewee, lazuli bunting, and Cassin's vireo. In addition, shrub-associated migrant species (for example, orange-crowned warbler), and those associated with mid-canopy deciduous trees (for example, black-throated gray warbler, black-headed grosbeak) were common on our study sites. Habitat for these species is limited in habitats dominated by closed-canopy conifer forest be- cause shrubs and hardwoods do not flourish under a dense canopy. The northern oriole is another migrant species that does not regularly occur in coniferous forests but is associated with semi-open oak and madrone woodlands.
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

18 NORTHWESTERN NATURALIST 82(1)
However, northern orioles also were rare on our study sites except on Sauvie Island. Their distribution in western Oregon is patchy, being limited mainly to the Rogue and Umpqua val- leys and to Multnomah and Columbia counties along the Willamette and Columbia rivers (Gil- ligan and others 1994), coincidental with con- centrations of open oak habitats.
Oak woodlands also provide primary habitat to several species of resident birds that are rare or absent from closed-canopy conifer forests. Species in this category include acorn wood- pecker, downy woodpecker, white-breasted nuthatch, black-capped chickadee, mourning dove, scrub jay, and American goldfinch. Other resident species in western Oregon that occur in conifer forests but which seem to occur more frequently in oak woodlands include: northern flicker, Bewick's wren, spotted towhee, cedar waxwing, purple finch, American robin, and song sparrow. Most of these species were com- mon in the oak stands we surveyed, except acorn woodpeckers, which typically use oak groves that are more open and have less un- derstory than was characteristic of our stands (Marshall and others 1992). Breeding Bird Sur- vey (BBS) data indicate significant population declines in Oregon over the period from 1966 to 1998 for mourning dove, American robin, ce- dar waxwing, song sparrow, and American goldfinch (Sauer and others 1999).
Six resident species (acorn woodpecker, downy woodpecker, white-breasted nuthatch, black-capped chickadee, northern flicker, Be- wick's wren) and one migrant species (house wren) that are associated with oak woodlands are cavity-nesting species. Large-diameter, open-grown Oregon white oak trees produce a higher density of cavities than do Douglas-fir of the same diameter class. However, oaks grown in dense stands are not as prone to cav- ity-development as are open-grown oaks (Gumtow-Farrior 1991). Cavity-nesting spe- cies, especially secondary cavity-nesters that cannot excavate their own cavities (for example, American kestrel and western bluebird (Sialia mexicana)), may therefore be particularly de- pendent on large, open-grown oaks for nesting habitat. Our study provides further evidence of the relationship between abundance of cavity- nesters and large trees because the abundance of northern flickers, house wrens, and white- breasted nuthatches was positively correlated
with the tree diameter (R2 = 0.60-0.65, P =
0.08). Of these species, white-breasted nut- hatches are of particular concern because in western Oregon they avoid dense coniferous forests (Csuti and others 1997), and seem to be strongly associated with oak-dominated habi- tats (Anderson 1980). In our study, abundance of white-breasted nuthatches also was nega- tively correlated with percent cover of Doug- las-fir (R2 = 0.80, P < 0.01). White-breasted nuthatches in oak woodlands in California also have been associated with low tree density and large tree diameter (Wilson and others 1991). Furthermore, white-breasted nuthatches have experienced significant population declines ac- cording to BBS data from Willamette Valley survey routes (Sauer and others 1999). Large, open-grown oaks mainly exist only as remnant trees in the Willamette Valley, and will likely become obsolete in the absence of disturbances that retard succession in oak woodlands. Fur- ther population declines can be expected for cavity-nesting bird species associated with large-diameter, open-grown oaks as this re- source diminishes.
In this study, we were unable to directly compare bird communities in valley wood- lands before and after conversion to conifer dominance through the process of succession. Therefore, we relied on qualitative compari- sons between bird species composition in co- niferous habitats of the Coast Ranges and that in our oak-dominated sites to formulate hy- potheses about changes in bird community composition that might accompany plant com- munity succession. Although other factors (for example, elevation) may influence differences in species composition between montane co- niferous forests and valley woodlands, the dif- ference in dominant vegetation type is likely among the most important (Wiens 1989:133- 134). Chambers (1996) surveyed birds along the western margin of the Willamette Valley, in an area where succession has advanced from oak savanna to closed-canopy conifers (Sprague and Hansen 1946). Although she observed some of the species that we categorize as oak associates (for example, lazuli bunting, Be- wick's wren, western wood-pewee, Cassin's vireo), she had relatively few observations of each of these species, and she did not observe any white-breasted nuthatches during the 3 yr of her study. The most abundant species in
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 19
Chamber's study were those that are character- istic of Coast Range conifer forests (for exam-
ple, hermit warbler, winter wren, Swainson's thrush, Pacific-slope flycatcher). This provides further support for our hypothesis that species associated with oak woodlands are likely to de- crease in abundance and perhaps be locally ex-
tirpated where oak habitats are replaced by co- nifers, but this hypothesis needs to be tested for each of the species we have categorized as oak- associated.
Conclusions and Conservation Strategies Conifer forests dominate much of the Pacific
Northwest, so the patches of Oregon white oak woodlands in western Oregon and Washington provide an important element of regional di-
versity (Larsen and Morgan 1998). We believe that the conservation of oak habitats is critical to the maintenance of populations of several avian species in the Willamette Valley. The
gradual but large-scale replacement of oak habitats by conifers, in combination with en- croachment upon oak woodlands by agricul- ture and urban development, has already co- incided with the loss of at least 1 species, Lewis' woodpecker (Marshall and others 1992). Fur- ther decreases in regional avian diversity are
likely as oak habitats continue to be eroded.
Species that may be of the greatest concern for land managers whose goals include conserva- tion of biodiversity are those that are uncom- mon in coniferous forest types and occur less
frequently in oak woodlands now than they did historically (Tables 1 and 2). Of these, the white-breasted nuthatch is perhaps the most
strongly associated with large-diameter oaks (Table 2) in semi-open woodlands (Pravosudov and Grubb 1993) and may, therefore, be one of the bird species most threatened by loss of
open oak woodlands. The multiple threats to oak habitats and the
predominant private ownership of the province historically dominated by oak (Puchy and Mar- shall 1993), makes oak woodlands 1 of the most threatened habitat types in western Oregon and Washington (Larsen and Morgan 1998). Private landowners should be encouraged to maintain oaks on their property by thinning and selectively harvesting conifers, bigleaf ma- ple, and non-native tree species where they are competing with oak. Selective harvesting of co- nifers in order to preserve oak habitats could
provide opportunities to test hypotheses that we have set forth, to educate the public about the importance of oak woodlands, and to per- haps produce some revenue for local govern- ments. Finally, because oak woodlands histor-
ically were maintained by fire, prescribed burning of understory vegetation may be an
appropriate and effective tool for preserving open stands of oak and for discouraging en- croachment by shade-tolerant tree species.
ACKNOWLEDGMENTS
This project was conducted under the auspices of the Oregon Natural Heritage Program of the Nature Conservancy of Oregon. Funding was provided by the U.S. Fish and Wildlife Service Migratory Birds Nongame Program. T. Zimmerman and M. Naugh- ton of the U.S. Fish and Wildlife Service and R. Gog- gans, S. Smith, W. Kinyoun and T. Dufour of the Oregon Department of Fish and Wildlife provided support and assistance. We thank S. Jorgensen, W. and J. Martinak, and H. Secrest for their generosity and cooperation in granting access to their property. R. Peck helped conduct bird surveys and J. Titus helped with vegetation sampling. D. Chiller created Fig. 1. J. Weikel, B. Altman, and J. Alexander provid- ed valuable comments on drafts of this manuscript.
LITERATURE CITED
ANDERSON SH. 1970. The avifaunal composition of Oregon white oak stands. Condor 72:417-423.
ANDERSON SH. 1972. Seasonal variations in forest birds of western Oregon. Northwest Science 46: 194-206.
ANDERSON SH. 1980. Habitat selection, succession, and bird community organization. In: DeGraff RM and Tilgham NG, editors. Management of western forests and grasslands for nongame birds. Ogden, UT: Intermountain Forest and Rangeland Experiment Station General Technical Report INT-86. p. 13-25.
CAREY AB, HARDT MM, HORTON SP, BISWELL BL. 1991. Spring bird communities in the Oregon Coast Range. In: Ruggiero LF, Aubry KB, Carey AB, Huff MH, technical coordinators. Wildlife and vegetation of unmanaged Douglas-fir forests. Portland, OR: USDA Forest Service General Tech- nical Report PNW-GTR-285. p 123-142.
CHAMBERS C. 1996. Response of terrestrial verte- brates to three silvicultural treatments in the cen- tral Oregon Coast Range [dissertation]. Corvallis, OR: Oregon State University. 213 p.
CROSS S, SIMMONS JK. 1983. Bird populations of the mixed-hardwood forests near Roseburg, Oregon. Portland, OR: Oregon Department of Fish and Wildlife Nongame Wildlife Program Technical Report 82-2-05. 42 p.
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

20 NORTHWESTERN NATURALIST 82(1)
CSUTI B, KIMERLING AJ, O'NEIL TA, SHAUGHNESSY MM, GAINES EP, Huso M. 1997. Atlas of Oregon wildlife: distribution, habitat, and natural histo- ry. Corvallis, OR: Oregon State University Press. 492 p.
FINCH DM, STANGEL PW 1992. Status and manage- ment of neotropical migratory birds. Fort Collins, CO: USDA Forest Service, Rocky Mountain Forest and Range Experiment Station General Technical Report RM-229. 422 p.
FRANKLIN JF, DRYNESS CT. 1988. The natural vege- tation of Oregon and Washington. Corvallis, OR: Oregon State University Press. 452 p.
*GALEN C. 1989. A preliminary assessment of the sta- tus of the Lewis' woodpecker in Wasco County, Oregon. Report No. 88-3-01, submitted to Oregon Department of Fish and Wildlife in fullfillment of Nongame Project. 23 p. Available from: Oregon Department of Fish and Wildlife, 2501 SW First Avenue, PO Box 59, Portland, OR 97207.
GERIG R. 1992. Breeding birds of the Coast Range: a comparison of the species mix in successional for- est communities in Polk County. Oregon Birds 18: 7-10.
GILLIGAN J, SMITH M, ROGERS D, CONTRERAS A. 1994. Birds of Oregon: status and distribution. Mc- Minnville, OR: Cinclus Press. 330 p.
GUMTOW-FARRIOR DL. 1991. Cavity resources in Oregon white oak and Douglas-fir stands in the mid-Willamette Valley, Oregon [thesis]. Corvallis, OR: Oregon State University. 89 p.
JOHANNESSEN CL, DAVENPORT WA, MILLET A, MCWILLIAMS S. 1971. The vegetation of the Wil- lamette Valley. Association of American Geogra- phers, Annals 61:286-302.
KLOCK C, SMITH S, O'NEIL T, GOGGANS R, BARRETT C. 1998. Willamette Valley map land use/land cover informational report. Internal research re- port for Oregon Department of Fish and Wildlife Corvallis, OR. Available at http://www.nwhi. org/nhiweb/nhi.html.
LARSEN EM, MORGAN JT. 1998. Management recom- mendations for Washington's priority habitats: Oregon white oak woodlands. Olympia, WA: Washington Department of Fish and Wildlife. 37 p.
LUDWIG JA, REYNOLDS JE 1988. Statistical ecology. New York, NY: John Wiley and Sons. 337 p.
MARSHALL DB, CHILCOTE M, WEEKS H. 1992. Sensi- tive vertebrates of Oregon. Portland, OR: Oregon Department of Fish and Wildlife. irregular pagi- nation.
MCGARIGAL K. 1993. Relationship between land- scape structure and avian abundance patterns in the Oregon Coast Range [dissertation]. Corvallis, OR: Oregon State University. 274 p.
MCGARIGAL K, MCCOMB WC. 1992. Streamside ver- sus upslope breeding bird communities in the central Oregon Coast Range. Journal of Wildlife Management 56:10-23.
MUELLER-DUMBOIS D, ELLENBERG H. 1974. Aims and methods of vegetation ecology. New York, NY: John Wiley and Sons. 545 p.
PARTNERS IN FLIGHT. 2000. Westside lowlands and valleys bird conservation plan. Available at http: / /community.gorge.net/natres/pif/conplans/ west-low /westside-low.html.
PRAVOSUDOv VV, GRUBB TC JR. 1993. White-breasted nuthatch (Sitta carolinensis). No. 54. In Poole A and Gill F, editors. The Birds of North America. Philadelphia, PA: The Birds of North America, Inc. 14 p.
PUCHY CA, MARSHALL DB, editors. 1993. Oregon wildlife diversity plan. 2nd edition. Portland, OR: Oregon Department of Fish and Wildlife. 413 p.
RALPH CJ, GEUPEL GR, PYLE P, MARTIN TE, DESANTE DE 1993. Handbook of field methods for moni- toring landbirds. Albany, CA: USDA Forest Ser- vice, Pacific Southwest Research Station General Technical Report PSW-144. 41 p.
RYAN LA, CAREY AB. 1995. Biology and management of the western gray squirrel and Oregon white oak woodlands: with emphasis on the Puget Trough. Portland, OR: USDA Forest Service Gen- eral Technical Report PNW-348. 36 p.
SABIN TE, STAFFORD SG. 1990. Assessing the need for transformation of response variables. Corvallis, OR: Forest Research Laboratory, Oregon State University. Special Publication 20. 31 p.
SAS INSTITUTE, INC. 1990. SAS/STAT user's guide, version 6. 3rd edition. Volume 2. Cary, NC: SAS Institute Inc. 846 p.
SAUER J R, HINES JE, THOMAS I, FALLON J, GOUGH G. 1999. The North American breeding bird survey, results and analysis 1966-1998. Version 98.1. <http://www.mbr-pwrc.usgs.gov/bbs/> Re- vised 3 December 1999, accessed 1 December 2000. Laurel, MD: USGS Patuxent Wildlife Re- search Center.
SPRAGUE FL, HANSEN HP. 1946. Forest succession in the McDonald Forest, Willamette Valley, Oregon. Northwest Science 20:89-98.
THILENIUS JE 1968. The Quercus garryana forests of the Willamette Valley, Oregon. Ecology 49:1124- 1133.
TOWLE JC. 1982. Changing geography of Willamette Valley woodlands. Oregon Historical Quarterly 83:66-87.
VERNER J. 1980. Birds of California oak habitats- management implications. Claremont, CA: USDA Forest Service General Technical Report PSW-44. p 246-264.
WIENS JA. 1989. The ecology of bird communities: * Unpublished.
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 21
processes and variations. Volume 2. Cambridge UK: Cambridge University Press. 316 p.
WILSON RA, MANLEY P, NOON BR. 1991. Covari- ance patterns among birds and vegetation in a California oak woodland. In Standiford RB, technical coordinator. Proceeding of the sym- posium on oak woodlands and hardwood
rangeland management. Berkeley, CA: USDA Forest Service General Technical Report PSW- 126. p. 126-135.
Submitted 18 March 2000, accepted 10 January 2001. Corresponding Editor: C. J. Ralph.
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

APPENDIX A Location, ownership, and habitat characteristics of oak woodlands in the Willamette Valley, Oregon.
Oak Average % cover (SE)
UTM coordinatesa diame- Oregon Doug- Trees/hac terd white Bigleaf las- Woody
Site Name County Easting Northing No.b Years surveyed (SE) (SE) oak maple fir shrubs
1 Baskett Slough NWR Polk 479605 4979294 12 1994 to 1996 670.0 46.90 69.0 2.2 1.0 49.6 (173.1) (3.6) (5.0) (1.2) (0.8) (6.1)
2 Fern Ridge Lane 480246 4880889 6 1995 to 1996 313.9 61.94 29.5 0.8 3.3 51.7 (188.0) (7.3) (7.5) (0.8) (3.3) (7.5)
3 Pigeon Butte, Finley NWR Benton 475006 4916410 8 1994 to 1996 349.1 62.37 50.4 19.3 0.0 57.2 (86.5) (4.5) (5.0) (3.9) (0.0) (6.3)
4 Mill Hill, Finley NWR Benton 473050 4916591 4 1994 to 1996 715.2 46.12 74.0 1.7 6.0 54.0 (334.79) (5.5) (10.0) (1.7) (4.9) (15.8)
5 Philomath (private) Benton 471912 4924679 4 1995 to 1996 782.2 38.57 83.7 0.0 0.0 67.5 (187.1) (4.1) (2.4) (0.0) (0.0) (9.7)
6 Oregon State University Benton 474970 4937142 12 1994 to 1996 622.1 42.87 57.1 2.5 2.2 24.0 (225.5) (5.0) (4.4) (2.1) (0.9) (5.0)
7 Sauvie Island Wildlife Management Multnomah 514217 5062995 12 1994 to 1996 141.4 76.5 60.6 0.0 0.0 65.0 Area (41.2) (3.3) (4.3) (0.0) (0.0) (5.3)
8 Airlie (private) Polk 478929 4955721 12 1995 to 1996 163.0 62.31 44.3 1.6 10.7 31.9 (23.8) (3.4) (4.3) (1.0) (3.2) (5.5)
9 Stayton (private) Marion 520510 4964064 4 1995 to 1996 550.2 49.85 61.2 0.0 3.7 33.7 (244.9) (5.7) (15.0) (0.0) (2.4) (10.3)
a Coordinates are for Zone 10. b Number of point-count stations at which birds were surveyed. c Average number of trees ?14 cm diameter at breast height/ha. d Average diameter at breast height (cm) of Quercus species.
2
;rr 0 H
(T3 H (TI
2 2 H
H
=71
GO
tO
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 23
APPENDIX B Frequency of occurrence (%), average abundance across all sites (standard error), and abundance index for each site for bird species observed in 9 oak woodland sites in the Willamette Valley, 1994 to 1996. Site-level abundance indices are given only for species with a frequency of occurrence >25%.
Percent Average Abundance indexc occur- abundance Site Site Site Site Site Site Site Site Site
Species rencea (SE)b 1 2 3 4 5 6 7 8 9
Sharp-shinned hawk 5 <0.01 - -- - - - P -
(Accipiter striatus) Red-tailed hawk 15 <0.01 - P - - P P P -
(Buteo jamaicensis) American kestrel <5 <0.01 -- -- -- - P -
(Falco sparverius) Ring-necked pheasant <5 <0.01 -- - - - -- P -
(Phasianus colchicus) Ruffed grouse 10 <0.01 - - P - -- P
(Bonasa umbellus) Wild turkey 10 <0.01 P
(Meleagris gallopavo) California quail 5 <0.01 - - - - --- - --- P
(Callipepla californica) Mountain quail 5 <0.01 -- - - - - -- P
(Oreortyx pictus) Band-tailed pigeon 10 <0.01 - - P P
(Columba fasciata) Mourning dove 50 0.04 (0.01) 0.11 0.03 0.01 - 0.04 0.08 0.06 0.01
(Zenaida macroura) Great horned owl 5 <0.01 - P- -
(Bubo virginianus) Rufous hummingbird 55 0.07 (0.03) 0.03 - 0.06 0.14 0.04 0.04 0.05 0.01 0.25
(Selasphorus rufus) Acorn woodpecker 5 <0.01 P - - -
(Melanerpes formicivorus) Northern flicker 50 0.06 (0.02) 0.01 0.08 0.03 0.03 - 0.11 0.19 0.01 0.08
(Colaptes auratus) Pileated woodpecker 5 <0.01 -- -- -- - P -
(Dryocopus pileatus) Red-breasted sapsucker 60 0.08 (0.04) 0.01 - 0.36 0.14 - 0.12 - - 0.08
(Sphyrapicus ruber) Hairy woodpecker 60 0.04 (0.01) 0.03 - 0.06 0.08 0.04 0.03 - 0.04 0.04
(Picoides villosus) Downy woodpecker 80 0.10 (0.03) 0.30 0.22 0.10 0.08 - 0.05 0.04 0.14
(Picoides pubescens) Pacific-slope flycatcher 85 0.24 (0.08) 0.09 0.08 0.46 0.58 0.63 0.13 - 0.07 0.08
(Empidonax difficilis) Olive-sided flycatcher 5 <0.01 - P - - - P
(Contopus cooperi) Western wood-pewee 100 0.95 (0.17) 1.44 0.42 1.24 0.36 1.92 0.57 0.94 0.89 0.75
(Contopus sordidulus) Willow flycatcher 10 <0.01 P P -- - P -
(Empidonax traillii) Violet-green swallow 10 <0.01 - P
(Tachycineta thalassina) Tree swallow 0 <0.01 ---- -- 0.33
(Tachycineta bicolor) Steller's jay 55 0.07 (0.03) 0.12 0.03 0.03 - - 0.27 - 0.11 0.04
(Cyanocitta stelleri) Scrub jay 50 0.14 (0.08) 0.09 0.61 0.01 - 0.04 0.03 0.45 0.01 0.04
(Aphelocoma californica) American crow 35 0.02 (0.01) 0.02 0.03 0.03 - - 0.06 0.03 0.03
(Corvus brachyrhynchos) Common raven 5 <0.01 P - - -
(Corvus corax)
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions
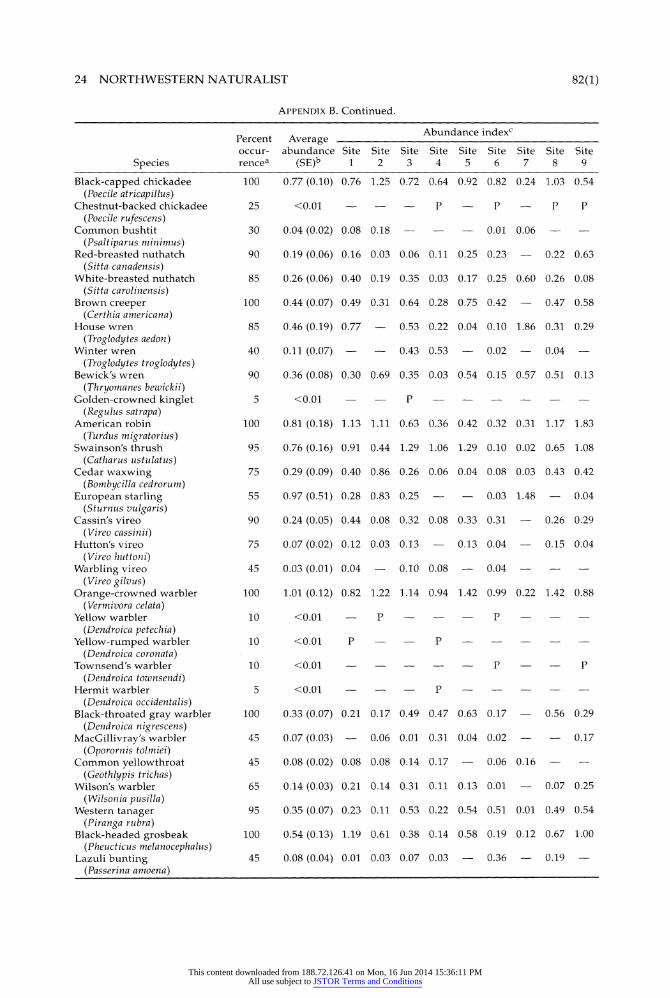
24 NORTHWESTERN NATURALIST 82(1)
APPENDIX B. Continued.
Percent Average Abundance indexc occur- abundance Site Site Site Site Site Site Site Site Site
Species rencea (SE)b 1 2 3 4 5 6 7 8 9
Black-capped chickadee 100 0.77 (0.10) 0.76 1.25 0.72 0.64 0.92 0.82 0.24 1.03 0.54 (Poecile atricapillus)
Chestnut-backed chickadee 25 <0.01 P - P - P P (Poecile rufescens)
Common bushtit 30 0.04 (0.02) 0.08 0.18 - - 0.01 0.06 - -
(Psaltiparus minimus) Red-breasted nuthatch 90 0.19 (0.06) 0.16 0.03 0.06 0.11 0.25 0.23 - 0.22 0.63
(Sitta canadensis) White-breasted nuthatch 85 0.26 (0.06) 0.40 0.19 0.35 0.03 0.17 0.25 0.60 0.26 0.08
(Sitta carolinensis) Brown creeper 100 0.44 (0.07) 0.49 0.31 0.64 0.28 0.75 0.42 - 0.47 0.58
(Certhia americana) House wren 85 0.46 (0.19) 0.77 - 0.53 0.22 0.04 0.10 1.86 0.31 0.29
(Troglodytes aedon) Winter wren 40 0.11 (0.07) - - 0.43 0.53 - 0.02 - 0.04 -
(Troglodytes troglodytes) Bewick's wren 90 0.36 (0.08) 0.30 0.69 0.35 0.03 0.54 0.15 0.57 0.51 0.13
(Thryomanes bewickii) Golden-crowned kinglet 5 <0.01 - - P
(Regulus satrapa) American robin 100 0.81 (0.18) 1.13 1.11 0.63 0.36 0.42 0.32 0.31 1.17 1.83
(Turdus migratorius) Swainson's thrush 95 0.76 (0.16) 0.91 0.44 1.29 1.06 1.29 0.10 0.02 0.65 1.08
(Catharus ustulatus) Cedar waxwing 75 0.29 (0.09) 0.40 0.86 0.26 0.06 0.04 0.08 0.03 0.43 0.42
(Bombycilla cedrorum) European starling 55 0.97 (0.51) 0.28 0.83 0.25 - - 0.03 1.48 - 0.04
(Sturnus vulgaris) Cassin's vireo 90 0.24 (0.05) 0.44 0.08 0.32 0.08 0.33 0.31 - 0.26 0.29
(Vireo cassinii) Hutton's vireo 75 0.07 (0.02) 0.12 0.03 0.13 - 0.13 0.04 - 0.15 0.04
(Vireo huttoni) Warbling vireo 45 0.03 (0.01) 0.04 0.10 0.08 0.04 - - -
(Vireo gilvus) Orange-crowned warbler 100 1.01 (0.12) 0.82 1.22 1.14 0.94 1.42 0.99 0.22 1.42 0.88
(Vermivora celata) Yellow warbler 10 <0.01 - P - - - P - - -
(Dendroica petechia) Yellow-rumped warbler 10 <0.01 P P
(Dendroica coronata) Townsend's warbler 10 <0.01 -- - - - - P - - P
(Dendroica townsendi) Hermit warbler 5 <0.01 - - - P- -
(Dendroica occidentalis) Black-throated gray warbler 100 0.33(0.07) 0.21 0.17 0.49 0.47 0.63 0.17 - 0.56 0.29
(Dendroica nigrescens) MacGillivray's warbler 45 0.07 (0.03) 0.06 0.01 0.31 0.04 0.02 - - 0.17
(Oporornis tolmiei) Common yellowthroat 45 0.08(0.02) 0.08 0.08 0.14 0.17 0.06 0.16 - -
(Geothlypis trichas) Wilson's warbler 65 0.14 (0.03) 0.21 0.14 0.31 0.11 0.13 0.01 - 0.07 0.25
(Wilsonia pusilla) Western tanager 95 0.35 (0.07) 0.23 0.11 0.53 0.22 0.54 0.51 0.01 0.49 0.54
(Piranga rubra) Black-headed grosbeak 100 0.54(0.13) 1.19 0.61 0.38 0.14 0.58 0.19 0.12 0.67 1.00
(Pheucticus melanocephalus) Lazuli bunting 45 0.08 (0.04) 0.01 0.03 0.07 0.03 0.36 - 0.19
(Passerina amoena)
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions

SPRING 2001 HAGAR AND STERN: AVIFAUNA IN OAK WOODLANDS 25
APPENDIX B. Continued.
Percent Average Abundance indexc occur- abundance Site Site Site Site Site Site Site Site Site
Species rencea (SE)b 1 2 3 4 5 6 7 8 9
Spotted towhee 100 1.65 (0.15) 1.59 2.39 1.65 1.31 2.33 1.04 1.36 1.44 1.71 (Pipilo maculatus)
Chipping sparrow 5 <0.01 - - - P- - -
(Spizella passerina) Song sparrow 55 0.15 (0.08) 0.22 0.07 0.11 0.08 0.01 0.76 - 0.13
(Melospiza melodia) White-crowned sparrow 35 0.13 (0.08) 0.28 - - 0.67 0.25
(Zonotrichia leucophrys) Dark-eyed junco 95 0.55 (0.18) 0.51 0.11 0.13 0.39 0.42 1.07 - 1.71 0.63
(Junco hyemalis) Brown-headed cowbird 80 0.50 (0.10) 0.39 0.47 0.43 0.14 0.96 0.25 0.25 0.83 0.79
(Molothrus ater) Northern oriole 10 <0.01 P - P 0.60 - -
(Icterus bullockii) Purple finch 95 0.44 (0.11) 1.06 0.03 0.56 0.47 0.67 0.31 0.51 0.38
(Carpodactus purpureus) House finch 5 <0.01 - --- P - P
(Carpodacus mexicanus) Pine siskin 5 <0.01 - - - P - -
(Carduelis pinus) American goldfinch 100 0.41 (0.09) 0.78 0.11 0.33 0.36 0.50 0.19 0.03 0.63 0.79
(Carduelis tristis) Evening grosbeak 5 <0.01
. .. P - -
(Coccothraustes vespertinus) a Frequency of occurrence = 100 X (the sum of the number of sites in which each bird species was observed/total number of sites); Sauvie
Island WMA was not included in calculations of frequency of occurrence. b Average of site-level abundance indices (see index of abundance, below). c Index of abundance = number of observations per point-count station per visit, averaged over years for each bird species with a frequency
of occurrence >25%. "P" indicates that the species was present on the site, but abundance index was <0.03; "-" indicates that the species was not observed on the site.
This content downloaded from 188.72.126.41 on Mon, 16 Jun 2014 15:36:11 PMAll use subject to JSTOR Terms and Conditions