assembly of new nucleosomal histones and new dna into chromatin
TRANSCRIPT

Assembly of New Nucleosomal Histones and New DNA into ChromatinAuthor(s): Ronald HancockSource: Proceedings of the National Academy of Sciences of the United States of America,Vol. 75, No. 5 (May, 1978), pp. 2130-2134Published by: National Academy of SciencesStable URL: http://www.jstor.org/stable/68398 .
Accessed: 08/05/2014 03:22
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
National Academy of Sciences is collaborating with JSTOR to digitize, preserve and extend access toProceedings of the National Academy of Sciences of the United States of America.
http://www.jstor.org
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions

Proc. Natl. Acad. Sci. USA Vol. 75, No. 5, pp. 2130-2134, May 1978 Biochemistry
Assembly of new nucleosomal histones and new DNA into chromatin (chromatin assembly/nucleosomes/DNA replication)
RONALD HANCOCK
Swiss Institute for Experimental Cancer Research, 1066-Epalinges/Lausanne, Switzerland
Communicated by James Bonner, February 2, 1978
ABSTRACT The assembly of chromatin from newly syn- thesized nucleosomal histones (labeled with [3H]arginine) and new DNA (density-labeled with [1251]iododeoxyuridine) was studied in growing cultured mouse cells. The nucleosomal his- tones were specifically examined by dissociating histone HI and nonhistone proteins from unsheared chromatin either by incu- bation in 0.6 M NaCl or by digestion with micrococcal nuclease to release nucleosomes. In both cases, the four nucleosomal histones (H2A, H2B, H3, and H4) are essentially the only pro- teins that remain bound to DNA and that are labeled by [3H]- arginine. After formaldehyde fixation, HI-depleted chromatin containing dense DNA can be completely resolved in CsCl buoyant density gradients from that containing unreplicated DNA; separation of nucleosomes is satisfactory although less complete.
New DNA and new histones are already assembled into chromatin possessing characteristic nucleosomal structure after 3 min of synthesis (the shortest time studied), as shown by the kinetics of digestion of new DNA by micrococcal nuclease, by the distribution of new DNA and new histones in nucleosomes and multimers, and by buoyant density analysis of nucleosonmes. However, after 3-30 min of synthesis most new nucleosomal histones are associated with unreplicated DNA rather than with new DNA. It is concluded that new nucleosomes are assembled on DNA at some distance from DNA replication sites, with concomitant migration of preexisting nucleosomes onto new DNA.
The process of replication of the eukaryotic chromosome in- cludes, among other steps, the assembly of new DNA and new histones into chromatin. After assembly, the four nucleosomal histones (H2A, H2B, H3, and H4) are conserved in chromatin for many cell generations (1-3), but the extranucleosomal his- tone Hi shows a turnover of about 15% per cell generation (2). New DNA rapidly acquires an approximately normal com- plement of proteins, as shown by buoyant density analysis of chromatin (4, 5). The simplest model of events that occur at the site of DNA replication (6-8) is that the nucleosomes of parental chromatin are distributed between the two daughter DNA molecules, which acquire a normal nucleosomal complement by the addition of new histones. During studies of chromatin assembly (9, 10), we have made observations that suggest that the situation in vivo may be more complex than that envisaged in these models.
MATERIALS AND METHODS
Cells. Cells of mouse line P815 (3), growing in stirred sus- pension cultures (30 ml) with a division time of 9-10 hr, were labeled with appropriate precursors during random exponential growth at 3-5 X 105 cells per ml.
Labeling of DNA and Histones. Parental DNA strands were prelabeled for at least two generations by [14C]dT (1.5 nCi/ml) in the presence of unlabeled dT (0.2 ,ug/ml). New DNA strands
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U. S. C. ?1734 solely to indicate this fact.
were density labeled by addition to the medium of 5-io- dodeoxyuridine (IdUrd) to 0.2 mM containing [5-'25I]IdUrd (1-30 ,Ci/ml), and new histones were simultaneously labeled with [3H]arginine (20-200 ACi/ml). Labeling periods of 30 min or less were terminated by pouring the culture onto 10 ml of crushed, frozen growth medium. Further manipulations were carried out on ice.
Preparation of Chromatin and Removal of HI. Chromatin was prepared in a low-ionic-strength environment, without shearing, in the form of chromatin structures (11). Hi and nonhistone proteins were dissociated by incubating the cen- trifuged chromatin structures with 0.6 M NaCl, sedimenting them through sucrose, and dialyzing them against 0.2 mM EDTA (pH 7.2), as described elsewhere (9). The Hl-depleted chromatin was then gently sheared (Dounce homogenizer) to a mass average fragment length of approximately 3000 DNA base pairs (11).
Preparation of Nucleosomes. Chromatin structures cen- trifuged in a Dounce homogenizer were gently resuspended (loose piston) in 1 mM Tris-HCl, pH 7.6/50 ,uM CaCl2 at a concentration equivalent to approximately 150 ,4g of DNA per ml. After 5 min of preincubation at 370, micrococcal nuclease (Worthington), in the same buffer, was added to 30 units/ml. Digestion of DNA was followed by withdrawing samples into 10% C13CCOOH; after addition of carrier bovine serum albu- min (100 ,ug), acid-precipitable radioactivity was collected on glass fiber filters (Whatman GF/C), which were washed with 5% C13CCOOH and ethanol, and radioactivity was measured as described below. To prepare nucleosomes, we stopped the digestion after 10 min (corresponding to approximately 20% acid-soluble DNA, see Fig. 2) by addition of EDTA to 1 mM and cooling in ice. The sample (0.4-0.8 ml) was layered onto a sucrose gradient (5-40% in 1 mM triethanolamine.HCI/l mM EDTA) (12) and centrifuged (Spinco rotor SW40, 32,000 rpm, 20 hr, 4?). Fractions (0.5 ml) were collected from the tube bottom and aliquots were precipitated for determination of radioactivity. The nucleosomes in the peak fraction (Fig. 2B) were dialyzed against 0.2 mM EDTA (pH 7.2). To examine the distribution of [3H]histones, I extracted the histones from gra- dient fractions with 0.2 M H2SO4 (30 min in ice) after addition of 1 mg of calf thymus DNA carrier; after centrifugation (Spinco rotor SW50.1, 30,000 rpm, 30 min, 40), the supernatant extracts were neutralized and their radioactivity was mea- sured.
Fixation and Buoyant Density Analysis. Hl-depleted chromatin and nucleosomes were fixed by formaldehyde and centrifuged in buoyant density CsCl gradients as described (9, 10). After fractions were collected on paper discs, 1251 was measured in a y spectrometer, and 14C and 3H were measured by liquid scintillation. The levels of radioactive precursors were chosen so that the combined spillover of 125I and 14C (measured with appropriate standards) was not more than 50% of the total counts in the 3H channel.
Electrophoresis. Sodium dodecyl sulfate/polyacrylamide
2130
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions
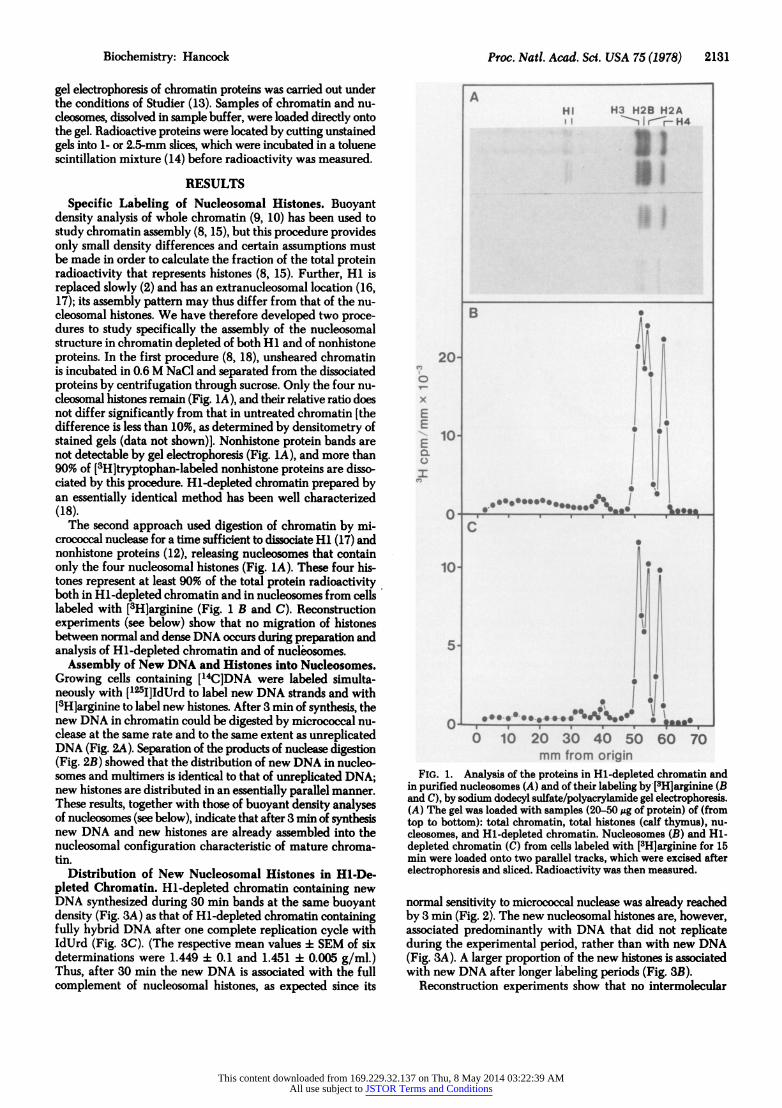
Biochemistry: Hancock Proc. Nati. Acad. Sci. USA 75 (1978) 2131
gel electrophoresis of chromatin proteins was carried out under the conditions of Studier (13). Samples of chromatin and nu- cleosomes, dissolved in sample buffer, were loaded directly onto the gel. Radioactive proteins were located by cutting unstained gels into 1- or 2.5-mm slices, which were incubated in a toluene scintillation mixture (14) before radioactivity was measured.
RESULTS
Specific Labeling of Nucleosomal Histones. Buoyant density analysis of whole chromatin (9, 10) has been used to study chromatin assembly (8, 15), but this procedure provides only small density differences and certain assumptions must be made in order to calculate the fraction of the total protein radioactivity that represents histones (8, 15). Further, Hi is replaced slowly (2) and has an extranucleosomal location (16, 17); its assembly pattern may thus differ from that of the nu- cleosomal histones. We have therefore developed two proce- dures to study specifically the assembly of the nucleosomal structure in chromatin depleted of both Hi and of nonhistone proteins. In the first procedure (8, 18), unsheared chromatin is incubated in 0.6 M NaCI and separated from the dissociated proteins by centrifugation through sucrose. Only the four nu- cleosomal histones remain (Fig. 1A), and their relative ratio does not differ significantly from that in untreated chromatin [the difference is less than 10%, as determined by densitometry of stained gels (data not shown)]. Nonhistone protein bands are not detectable by gel electrophoresis (Fig. 1A), and more than 90% of [3H]tryptophan-labeled nonhistone proteins are disso- ciated by this procedure. Hi-depleted chromatin prepared by an essentially identical method has been well characterized (18).
The second approach used digestion of chromatin by mi- crococcal nuclease for a time sufficient to dissociate Hi (17) and nonhistone proteins (12), releasing nucleosomes that contain only the four nucleosomal histones (Fig. 1A). These four his- tones represent at least 90% of the total protein radioactivity both in Hl-depleted chromatin and in nucleosomes from cells labeled with [3H]arginine (Fig. 1 B and C). Reconstruction experiments (see below) show that no migration of histones between normal and dense DNA occurs during preparation and analysis of Hl-depleted chromatin and of nucleosomes.
Assembly of New DNA and Histones into Nucleosomes. Growing cells containing [14C]DNA were labeled simulta- neously with [125I]IdUrd to label new DNA strands and with [3H]arginine to label new histones. After 3 min of synthesis, the new DNA in chromatin could be digested by micrococcal nu- clease at the same rate and to the same extent as unreplicated DNA (Fig. 2A). Separation of the products of nuclease digestion (Fig. 2B) showed that the distribution of new DNA in nucleo- somes and multimers is identical to that of unreplicated DNA; new histones are distributed in an essentially parallel manner. These results, together with those of buoyant density analyses of nucleosomes (see below), indicate that after 3 min of synthesis new DNA and new histones are already assembled into the nucleosomal configuration characteristic of mature chroma- tin.
Distribution of New Nucleosomal Histones in Hi-De- pleted Chromatin. Hl-depleted chromatin containing new DNA synthesized during 30 min bands at the same buoyant density (Fig. 3A) as that of Hl-depleted chromatin containing fully hybrid DNA after one complete replication cycle with IdUrd (Fig. 3C). (The respective mean values 4 SEM of six determinations were 1.449 :t 0.1 and 1.451 i 0.005 g/ml.) Thus, after 30 min the new DNA is associated with the full complement of nucleosomal histones, as expected since its
A HI H3 H23 H2A
E~ ~ ~~lf
'02 10' ir 4 B
20
10 . . t--
O 10 20: 9-0- :4-0- 50 W0 70 *m f'm orlol
FIG. 1. Analysis of the proteins in Hl-depleted chromatin and in purified nucleosomes (A) and of their labeling by [3H]arginine (B and C), by sodium dodecyl sulfate/polyacrylamide gel electrophoresis. (A) The gel was loaded with samples (20-50,gg of protein) of (from top to bottom): total chromatin, total histones (calf thymus), nu- cleosomes, and Hl-depleted chromatin. Nucleosomes (B) and Hl- depleted chromatin (C) from cells labeled with [3H]arginine for 15 min were loaded onto two parallel tracks, which were excised after electrophoresis and sliced. Radioactivity was then measured.
normal sensitivity to micrococcal nuclease was already reached by 3 min (Fig. 2). The new nucleosomal histones are, however, associated predominantly with DNA that did not replicate during the experimental period, rather than with new DNA (ig 3A). A lage proprtio of the new hisone is #.,.cate
wih ewDA ftrloge lblig eiodlFi.l3) Reontucio xprietsshwthtointrml eua
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions

2132 Biochemistry: Hancock Proc. Natl. Acad. Sci. USA 75 (1978)
100- A
0
3Z 50
E 0 .
0 10 20 30 40 50 60 120 min
6
o o 1O x 1 x 4i x o 0 0
4- 4- 4 4'
E E
0 ~~~~~~~~ 1A
0 10 20 30 4 Fraction
FIG. 2. Kinetics of digestion of new DNA and of unreplicated DNA in chromatin by micrococcal nuclease (A), and separation of the digestion products by sucrose gradient centrifugation (B). Cells prelabeled with [14C]dT were labeled for 3 min with [125IlIdUrd and [3H]arginine. Digestion of new 125I-labeled DNA (0) and of unrepli- cated [14C]DNA (0) in chromatin was followed by determining acid-precipitable radioactivity (A). Digestion of a portion of the chromatin was stopped after 10 min, and this sample was centrifuged through a 5-40% sucrose gradient (B). After fractionation of the gradient, the distribution of new 1251-labeled DNA (0), unreplicated ['14C]DNA (), and new [3H]histones (A) was determined.
migration of nucleosomal histones occurs during preparation of Hl-depleted chromatin; the distribution pattern of new histones (Fig. 3A) is not changed when an excess of unlabeled chromatin is present during all the experimental steps (see Fig. 5A). When simian virus 40 chromatin is exposed to 0.6 M NaCl, nucleosome migration is also undetectable (19). A probable explanation (9) for the discrepancy between these results and earlier work (20, 21) is that nucleosomes are tightly bound in the unsheared, constrained chromatin prepared here and in simian virus 40 chromatin, but that they can dissociate and migrate more easily when structural constraints are released by shearing.
Distribution of New Histones in Nucleosome Populations. The buoyant density of nucleosomes is more heterogeneous than that of Hl-depleted chromatin, probably as a result of hetero- geneity of DNA fragment length and smaller molecular size. The separation of nucleosomes that contain dense DNA (Fig. 4) is therefore less complete than that of Hl-depleted chromatin (Fig. 3), but the shorter DNA fragment length of nucleosomes allows them to be resolved after shorter labeling periods.
In agreement with the preceding results using Hl-depleted chromatin, new histones are found almost uniquely in nu- cleosomes that contain unreplicated DNA after 3 min of syn- thesis (Fig. 4A). At this time the new DNA is, however, in nu- cleosomes that possess a normal protein/DNA ratio, according to their buoyant density which is the same as that after one complete replication cycle with IdUrd (Fig. 5C). After longer
A pl.452 p1.430
2 - 4-
1 2 -: i * 8 2 o?0B | - r10
1 od< 1
00,0'..
0 - 0 I I 30
2 - 9
FIG. . Sepr_to of Hl-delete choaiotiignw
0 \ dense/A \ fo 5c
~~ E 1 ~~~~ 00 E EZe
0o 3 mnA)10in(B,r 0 h 0 w 4) C 0
deye and anlye. Th0itiuino e ulooa 3]
3- 4 o
#0 / \ -15
2- 0 2
101, % ~~~~~5
0 10 20 30
FIG. 3. Separation of Hi-depleted chromatin containing new, dense DNA from that containing unreplicated DNA in CsCl equi- librium density gradients. Cells prelabeled with [14C] dT were labeled for 30 min (A), 150 mimn (B), or 10 hr (C) with [125I]IdUrd and [3H]- arginine. Hi-depleted chromatin was prepared, fixed with formal- dehyde, and analyzed. The distribution of new nucleosomal [3H]- histones (0) between chromatin containing parental [14C]DNA strands (o) and new 1251-labeled DNA strands (A) was deter- mined.
synthesis periods (Fig. 4 B, C, and D), new histones appear progressively in nucleosomes containing new DNA. Recon- struction experiments, similar to those performed using Hl- depleted chromatin, show that no intermolecular migration of histones is detectable during preparation of nucleosomes (compare Fig. 5B with Fig. 4C). c
A trivial explanation of these results, that a significant fraction of chromosomal DNA replicates for a short period or a complete replication cycle without incorporating IdUrd, appears, to be excluded by the following observations (data not shown).. After one complete DNA replication cycle with IdUrd, all Hi-de- pleted chromatin incorporated the analog since it bands as a single dense peak and no material of normal density remains (Fig. DC). When cells were prelabeled for 3 hr with IdUrd to saturate precursor pools before labeling with [3H]arginine for 30 min, the distribution of new [3H]histones was similar to that in Fig. 3A. The maximum rate of incorporation of [ 2I]IdUrd (and of [3H]arginine) into trichloroacetic acid-insoluble material was reached within 30 sec of addition to the imedium. Results essentially identical with those in Fig. 4 were obtained when new DNA was labeled with [32p]orthophosphate in place of [125I]IdUrd and nucleosomes were treated with RNase before buoyannt de-nsity analysis., Thpnecpsib-ilityv of artifactual linear movement of new nucleosomes during experimental manipu- lations cannot be experimentally excluded; it would appear unlikely since to maintain histone/DNA ratios a compensatory,
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions

Biochemistry: Hancock Proc. Natl. Acad. Sci. USA 75 (1978) 2133
10 A p1.416 p1.396 2
5~~~ Q2 0e1
20 4! / A;s * 0 4C "A 2- *
0 * 0~~~~0
20 4 20
from ths cotinn 2nelctd N nCCleulbiu est
06
E2E4 A' * i \
0 0-- ~~/ 0~
20- 4-A 4
0
10n 2 120
A 0 IAA A / 5
orn 00(x) 0
4ppsit moemn of ol ulooe nt e N ol be reuied Duin dieto ofsma iu 0 hoa1wt
20 'COo* 1
Two cocuin maedanfrmteeepriet.Frt
2 0 E0- 0 12 A
A e
10 10,1 0 A~ U ,AA
S~~~~~ 0 0. 0~A
10 0- 10 2
FIG.e4.leparafte3mion of snucleosoms contohoaining new, densesDNA fro thosea nclontainin uonreplicatedn Daccring CsC eqiibiu dcensity
Chromtin watigsedb micrococcal rlc nuctltease,adithenresutiongan
groadint. Afernfixtipon,teso the distrbutonofne nrouclsoma SecHnd histonesw0 ewe nucleosomalhsoes contaeine uing parental [oCN
be requinared.oDuringmsionan asiians r with u
occurN(22,A23).
DICSSO Tw 1onlsin ma e rwnfo ths 5xeiet.Frt
ingrwig 81 mus cll, ewDN ad ewhitoesar assembld afte 3 m of sythsi inocrmai\ htosse
the. neweaatono nucleosomalhstnssycntheizenuing pe,eriose ofA 3-Somm aroe preomnatlaasocatdnit unreplicatedDNinCCeqlbrudnst grdinAs.Celprlbldwt 1Cd weelbldwt 15d d
o 10 20 30 II . I
20 A
40 10 A2 8 /A AO. 2
o - i o- | * , , , o O~~~~~~~~~~i 10- A 20
20 1 A ?0
AA od" "A'
co0000 '
40 A1 2 40 - ~ 20 B .0 20rac o x xA a of c
soe 0 A 15 .2 0 0- 0
j~~~20 2io ' ~~~~~10~ 2 ~~~~~~A\, 02 E
E A/
0 C -15 0 * ' 30
/ O'A0\2
10 9 ~ 0 * 3.
50 9
550 '0 '0 10 9,9/A
0 A'6t9
0 L 0-1I-- 0 10 20
Fraction FIG. 5. Analysis of Hi-depleted chromatin (A) and of nucleo-
somes (B) from mixed cell populations to examine possible inter- molecular migration of histones during preparative procedures. (A) Hl-depleted chromatin was prepared from a portion of the labeled cells used in the experiment of Fig. 3A, which were mixed with a 2-fold excess of unlabeled cells whose chromatin contained fully hybrid DNA after growth for 10 hr with IdUrd; this distribution pattern should be compared with Fig. 3A. (B) Nucleosomes were from a similar mixture of labeled cells from the experiment of Fig. 4C, with a 3-fold excess of unlabeled cells containing fully hybrid DNA; the distribution pattern should be compared with Fig. 4C. (C) Demonstration that new DNA, labeled during 3 min, is in nucleosomes containing a normal complement of histones. Nucleosomes were prepared from a mixture of cells prelabeled with [14C]dT and labeled for 3 min with 1125IJIdUrd, and cells containing fully hybrid DNA after growth for 10 hr with IdUrd and [3H]dT. The distribution of nucleosomes containing pa- rental [14CIDNA (i), 1251-labeled new DNA (A), and marker (dense) [3H]DNA (0) was determined.
The only process by which these two conclusions can be re- conciled is that the full nucleosomal complement of new DNA is acquired by migration of histones or nucleosomes from other regions of chromatin. In view of the very high affinity of nu- cleosomes for DNA, linear migration would appear much more probable than diffusion (dissociation-reassociation); an indi- cation that diffusional migration of nucleosomes does not occur in the nucleus is found in the observation (24, 25) that closed circular polyoma and simian virus 40 DNAs synthesized during cycloheximide inhibition of protein synthesis lack the su- p$rhelicity associatedwith the presence of nucleosomes (26). Linear migration on DNA of a large, multimeric DNA-un- winding protein is not an unusual phenomenon, for it occurs in the case of RNA polymerase (27, 28). One interesting aspect of a process in which new nucleosomes are formed in regions distant from replication sites is that their formation could, by imposing negative superhelicity (26) or other conformational changes in those regions, contribute towards the torque required for rotation and unwinding of the parental DNA. The observed slowing of 'DNA synthesis when histone synthesis is blocked (5, 24, 29) would be predicted from such a mechanism, which would require that new nucleosomes be formed before, rather than after, passage of the replication fork; the experiments
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions

2134 Biochemistry: Hancock Proc. Natl. Acad. Sci. USA 75(1978)
presented here cannot distinguish between these possibilities. Models in which nucleosomes of parental chromatin dissociate into two halves at the site of DNA replication, upon which new half-nucleosomes are assembled, appear to be incompatible with the data presented here. The possibility of linear migration of nucleosomes on DNA may also be relevant in the context of transcription.
When chromosomal DNA synthesis continues during inhi- bition of protein synthesis by cycloheximide (5, 29), simple models (6, 7) require that the daughter molecules should contain either regions of free DNA or internucleosomal DNA of twice the normal length. The kinetics of micrococcal nuclease di- gestion of new DNA in such chromatin are not, however, bi- phasic, (5-7), nor can regions of free DNA be detected by buoyant density analysis (ref. 5 and unpublished results). In- stead, the new DNA is in chromatin possessing a normal buoyant density (5) and a nucleosome spacing close to the normal value (6, 7). This paradox would be resolved if new DNA were fully covered by nucleosomes that had migrated from adjacent regions of DNA, as suggested by the present data.
I thank W. Garrard, B. Hirt, and N. Acheson for valuable discussions. This work, supported by the Fonds National Suisse, was presented in part at the Symposium on Chromatin of the British Society for Cell Biology, Glasgow, September 1975.
1. Byvoet, P. (1966) J. Mol. Biol. 17,311-318. 2. Gurley, L. R. & Hardin, J. M. (1969) Arch. Biochem. Biophys.
130, 1-6. 3. Hancock, R. (1969) J. Mol. Biol. 40, 457-466. 4. Fakan, S., Turner, G. N., Pagano, J. S. & Hancock, R. (1972) Proc.
Natl. Acad. Sci. USA 69,2300-2305. 5. Seale, R. L. & Simpson, R. T. (1975) J. Mol. Biol. 94,479-501. 6. Seale, R. L. (1976) Cell 9, 423-429. 7. Weintraub, H. (1976) Cell 9, 419-422.
8. Jackson, V., Granner, D. K. & Chalkley, R. (1975) Proc. Nati. Acad. Sci. USA 72,4440-4444.
9. Hancock, R. (1977) in Methods in Cell Biology, ed. Prescott D. M. (Academic, New York), Vol. 16, pp. 459-466.
10. Hancock, R. (1970) J. Mol. Biol. 48,357-60. 11. Hancock, R., Faber, A. J. & Fakan, S. (1977) in Methods in Cell
Biology, ed. Prescott, D. M. (Academic, New York), Vol. 15, pp. 127-147.
12. Bakayev, V. V., Melnickov, A. A., Osicka, V. D. & Varshavsky, A. J. (1975) Nucleic Acids Res. 2, 1401-1419.
13. Studier, F. W. (1973) J. Mol. Biol. 79,237-248. 14. Garrard, W. T. & Bonner, J. (1974) J. Biol. Chem. 249,5570-
5579. 15. Jackson, V., Granner, D. K. & Chalkley, R. (1976) Proc. Natl.
Acad. Sci. USA 73,2266-2269. 16. Baldwin, T. P., Boseley, P. G., Bradbury, E. M. & Ibel, K. (1975)
Nature 253,245-249. 17. Noll, M. & Kornberg, R. D. (1977) J. Mol. Biol. 109,393-404. 18. Oudet, P., Gross-Bellard, M. & Chambon, P. (1975) Cell 4,
281-300. 19. Germond, J.-E., Bellard, M., Oudet, P. & Chambon, P. (1976)
Nucleic Acids Res. 3, 3173-3192. 20. Varshavsky, A. J. & Ilyin, Yu, V. (1974) Biochim. Biophys. Acta
340,202-217. 21. Clark, R. J. & Felsenfeld, G. (1971) Nature New Biol. 229,
101-106. 22. Cremisi, C., Pignatti, P. F. & Yaniv, M. (1976) Biochem. Biophys.
Res. Commun. 73,548-554. 23. Polisky, B. & McCarthy, B. (1975) Proc. Natl. Acad. Sd. USA 72,
2895-2899. 24. Bourgaux, P. & Bourgaux-Ramoisy, D. (1972) Nature 235,
105-107. 25. White, M. & Eason, R. (1973) Nature New Biol. 241,46-49. 26. Germond, J. E., Hirt, B., Oudet, P., Gross-Bellard, M. & Cham-
bon, P. (1975) Proc. Natl. Acad. Sci. USA 72,1843-1847. 27. Wang, J. C., Jacobsen, J. H. & Saucier, J.-M. (1977) Nucleic Acids
Res. 4, 1225-1241. 28. Botchan, P., Wang, J. C. & Echols, H. (1973) Proc. Natl. Acad.
Sci. USA 70,3077-3081. 29. Weintraub, H. & Holzer, H. (1972) J. Mol. Biol. 66,1135.
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 03:22:39 AMAll use subject to JSTOR Terms and Conditions