application for commodity clearance of genetically modified organisms … for commodity... ·...
TRANSCRIPT

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 1
APPLICATION FOR COMMODITY CLEARANCE OF GENETICALLY MODIFIED ORGANISMS (GMO)
MON 87460 x MON 89034 x MON 88017
Data protection. This application contains scientific data and other information which are protected in accordance with Section 18 of the Genetically Modified Organisms Act, 1997 (Act No. 15 of 1997), including all amendments. © 2014 Monsanto Company. All Rights Reserved. This document is protected under national and international copyright law and treaties. This document and any accompanying material are for use only by the regulatory authority to which it has been submitted by Monsanto Company and its affiliates, collectively “Monsanto Company”, and only in support of actions requested by Monsanto Company. Any other use, copying, or transmission, including internet posting, of this document and the materials described in or accompanying this document, without prior consent of Monsanto Company, is strictly prohibited; except that Monsanto Company hereby grants such consent to the regulatory authority where required under applicable law or regulation. The intellectual property, information and materials described in or accompanying this document are owned by Monsanto Company, which has filed for or been granted patents on those materials. By submitting this document and any accompanying materials, Monsanto Company does not grant any party or entity any right or license to the information, material or intellectual property described or contained in this submission.
DIRECTORATE GENETIC RESOURCES Private Bag X973, Pretoria, 0001
Harvest House Room 167, 30 Hamilton Street, Arcadia, Pretoria, 0002
Tel: (+27) 12 319 6382, Fax: (+27) 12 319 6298, E-mail: [email protected]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 2
Table of Contents
1. BRIEF DESCRIPTION OF THE GENETICALLY MODIFIED ORGANISM ............ 9
1.1 Include specific and common names of the organism, the country of origin of the plant and a description of the genetically modified trait. ........................................ 9
2. COMMODITY CLEARANCE ............................................................................... 11
2.1 Please indicate the type of clearance requested. ................................................ 11
2.2 Detail specific instructions for the storage and handling of the plant or plant parts. . ........................................................................................................................... 11
2.3 When will commodity import take place? ............................................................ 11
2.4 Where will commodity import take place? ........................................................... 11
2.5 Detail the type of environment and the geographical areas for which the plant is suited. ................................................................................................................. 12
2.6 Who will undertake the commodity import? ......................................................... 12
2.7 Estimate the amount of production of the genetically modified plant within South Africa per annum, or the amount that will be imported into South Africa per annum. ............................................................................................................... 12
3. DESCRIPTION OF ANY PRODUCT DERIVED FROM THE PLANT .................. 12
3.1 Identify the part of the plant to be used for the product, the type of product, and the use of the product, the market sector in which the product will be marketed and the trade name of the product. ..................................................................... 12
3.2 Specify the exact conditions of use of the product. ............................................. 13
3.3 Provide information on the proposed labelling of the product for marketing. ....... 13
3.4 State whether the benefits of the product are available in any other non-genetically modified form. If so, state why the genetically modified form should be approved for general release when other, non-modified products are available. . 13
3.5 Detail specific instructions for the storage and handling of GMO’s that will avoid misuse or escape of the genetically modified plant into an environment for which it was not intended. ............................................................................................... 13
3.6 Detail the likelihood of the GMO being exported from South Africa, particularly if such export could result in the introduction of the plant into its centre of origin. .. 14
4. FOREIGN GENES AND GENE PRODUCTS...................................................... 14
4.1 Identify all foreign genes in the genetically modified plant. .................................. 14
4.2 Describe the gene products that are derived from the foreign genes. ................. 30
4.3 Describe the biological activity associated with the foreign gene products. ......... 30
4.4 Provide information on the rate and level of expression of the foreign genes and the sensitivity of the measurement of the rate and level. State whether expression is constitutive or inducible. Are foreign genes expressed throughout the plant or only in certain organs or tissues? ..................................................... 31
4.5 Provide protocols for the detection of the foreign genes in the environment including sensitivity, reliability and specificity of the techniques. ......................... 39

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 3
5. RESISTANCE ..................................................................................................... 39
5.1 Detail whether the genetically engineered plant is able to initiate resistance, in any biotic component of the environment, to any biologically active foreign gene product. .............................................................................................................. 39
5.2 Detail what methods are available to minimise the risk of resistance developing in the environment. ................................................................................................. 39
5.3 Detail how resistance will be managed during release of the genetically modified plant. .................................................................................................................. 39
6. HUMAN AND ANIMAL HEALTH ......................................................................... 40
6.1 State whether the genetically modified plant or its products will enter human or animal food chains. ............................................................................................. 40
6.2 Detail the results of experiments undertaken to determine the toxicity of the foreign gene products (including marker genes) to humans and animals. ........... 40
6.3 If the foreign gene products are toxic or allergenic in any way, detail how the commodity clearance will be managed to prevent contact with animals or humans that will lead to discomfort or toxicity. ................................................................ 104
6.4 What are the common/major allergens present in the recipient organism before modification? .................................................................................................... 104
6.5 What evidence is there that the genetic modification described in this application did not result in over-expression of the possible allergens indicated in 6.4 i.e. is the expression of the possible allergens in the non-GM counterpart substantially equivalent to that in the GM organism? ............................................................. 104
6.6 What are the implications of the proposed activity with regard to the health and safety of the workers, cleaning personnel and any other person that will be directly or indirectly involved in the activity? Please take into consideration the provisions of the Occupational Health and Safety Act, 1993 (Act No. 181 of 1993) and accompanied regulations. .......................................................................... 104
6.7 Indicate the proposed health and safety measures that would be applied to safeguard employees during the proposed activity. .......................................... 104
7. ENVIRONMENTAL IMPACT AND PROTECTION ............................................ 105
7.1 Detail any long-term effect the commodity clearance of the genetically modified organism is likely to have on the biotic and abiotic components of the environment. ..................................................................................................... 105
7.2 Provide data and information on ecosystems that could be affected by use of the plant or its products. ......................................................................................... 105
7.3 Specify what effect the general release of the genetically modified plant will have on biodiversity. .................................................................................................. 105
7.4 Specify the measures to be taken in the event of the plant or product being misused or escaping into an environment for which it is not intended. .............. 105
7.5 If the foreign genes give rise to crops resistant to agrochemicals, provide information on the registration of the agrochemicals to be used on the crop. .... 105
8. SOCIO-ECONOMIC IMPACTS......................................................................... 106

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 4
8.1 Specify what, if any, positive or negative socio-economic impacts the genetically modified plant will have on communities in the proposed region of release. ..... 106
9. WASTE DISPOSAL .......................................................................................... 106
9.1 Where only a portion of the genetically modified plant will be used for the product, how will the unused plant parts be disposed of? ............................................... 106
10. MONITORING AND ACCIDENTS .................................................................... 106
10.1 Indicate the methods and plans for monitoring of the GMO (also refer to Environmental Risk Assessment Framework for genetically modified organisms) in a comprehensive compliance plan ................................................................ 106
10.2 Indicate any emergency procedures that will be applied in the event of an accident in a comprehensive contingency plan ................................................. 106
11. PATHOGENIC AND ECOLOGICAL IMPACTS ................................................. 107
11.1 Submit an evaluation of the foreseeable impacts, in particular any pathogenic and ecologically disruptive impacts. ......................................................................... 107
12. RISK MANAGEMENT ....................................................................................... 107
12.1 Please indicate any risk management measures that would be required for commodity clearance. ....................................................................................... 107
13. COMPLETE THE AFFIDAVIT ........................................................................... 108
14. AFFIDAVIT/VERKLARING/STATEMENT ......................................................... 115
15. REFERENCES ................................................................................................. 116

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 5
List of Tables Table 1. Summary of inserted DNA inherited from MON 87460 ........................... 18 Table 2. Summary of inserted DNA inherited from MON 89034 ........................... 18 Table 3. Summary of inserted DNA inherited from MON 88017 ........................... 18 Table 4. Summary of CspB Protein Levels in Maize Tissues Collected from MON
87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions ................................................................ 33
Table 5. Summary of NPTII Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions ................................................................ 34
Table 6. Summary of CP4 EPSPS Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions .................................................. 35
Table 7. Summary of Cry1A.105 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions .................................................. 36
Table 8. Summary of Cry2Ab2 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions .................................................. 37
Table 9. Summary of Cry3Bb1 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions .................................................. 38
Table 10. N-terminal amino acid sequence analysis of the CspB protein purified from grain tissue of MON 87460 ............................................................. 45
Table 11. CspB functional assay ............................................................................ 48 Table 12. Amino acid sequence identity between MON 87460-produced CspB
protein and other cold shock domain containing proteins present in foods ............................................................................................................... 56
Table 13. Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Well-Watered Conditions .............................................................................................. 74
Table 14. Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Water-limited Conditions .............................................................................................. 76
Table 15. Literature and ILSI Ranges for Components in Maize Forage and Grain 79 Table 16. Combined-Site Analysis of Phenotypic Characteristics of Test Substance
MON 87460 × MON 89034 × MON 88017 Compared to the Control in the Well-Watered Treatment in 2007 ............................................................ 84
Table 17. Individual Site Analysis of Phenotypic Characteristics of Test Substance MON 87460 × MON 89034 × MON 88017 Compared to the Control in the Well-Watered Treatment in 2007 ............................................................ 85

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 6
Table 18. Individual Site Analysis of Abiotic Stressors Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007 ........................ 86
Table 19. Individual Site Analysis of Disease Damage Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007 ........................ 87
Table 20. Individual Site Analysis of Arthropod Damage Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007 ........................ 88
Table 21. Estimates of Maize Consumption by the WHO GEMS/Food Programme1 ...
............................................................................................................... 93 Table 22. Mean Expression of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2,
and CP4 EPSPS Proteins in Grain from MON 87460 × MON 89034 × MON 88017 Grown Under Well-Watered Conditions in a 2006-2007 Chilean Field Trial1 ................................................................................. 95
Table 23. Intake of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS Proteins from Consumption of Foods Derived from MON 87460 × MON 89034 × MON 88017 in South Africa ............................................. 95
Table 24. Margins of Exposure for the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS Proteins from Consumption of Foods Derived from MON 87460 × MON 89034 × MON 88017 in South Africa .............. 97
Table 25. The estimated daily consumption of maize in poultry and livestock.1 .... 100 Table 26. The mean and highest (max) levels of CSPB, NPTII, Cry1A.105,
Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein levels in the maize tissues collected from MON 87460 × MON 89034 × MON 880171 ................... 101
Table 27. Mean and highest (Max) daily dietary intakes (DDIs) of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in poultry and livestock (g/kg bwt/d)1 .......................................................................... 102

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 7
List of Figures Figure 1. Taxonomic classification of maize and its close relatives ......................... 9 Figure 2. Map of the Insert in MON 87460 ............................................................ 16 Figure 3. Map of the Insert in MON 89034 ............................................................ 16 Figure 4. Map of the Insert in MON 88017 ............................................................ 17 Figure 5. Map of Plasmid Vector PV-ZMAP595 ..................................................... 23 Figure 6. Map of Plasmid Vector PV-ZMIR245 ...................................................... 24 Figure 7. Map of Plasmid Vector PV-ZMIR39 ........................................................ 25 Figure 8. Southern Blot Analysis to Verify the Presence of MON 87460 in
MON 87460 × MON 89034 × MON 88017: CS-cspB Probe .................. 26 Figure 9. Southern Blot Analysis to Verify the Presence of MON 87460 in
MON 87460 × MON 89034 × MON 88017: CS-nptII Probe ................... 27 Figure 10. Southern Blot Analysis to Verify the Presence of MON 89034 in
MON 87460 × MON 89034 × MON 88017: CS-cry1A.105 and CS-cry2Ab2 Probes...................................................................................... 28
Figure 11. Southern Blot Analysis to Verify the Presence of MON 88017 in MON 87460 × MON 89034 × MON 88017: TS-CTP2/CS-cp4 epsps and CS-cry3Bb1 Probes ............................................................................... 29
Figure 12. MALDI-TOF MS coverage map of the CspB protein isolated from MON 87460 ............................................................................................ 45
Figure 13. Western Blot analysis of MON 87460- and E. coli-produced CspB proteins .................................................................................................. 45
Figure 14. SDS-PAGE of E. coli-and MON 87460-produced CspB proteins ............ 46 Figure 15. Glycosylation analysis of the MON 87460-produced CspB protein ......... 47 Figure 16. Western Blot analysis of the MON 87460- and E. coli-produced NptII
protein .................................................................................................... 51 Figure 17. Phylogram establishing that the Cry1A.105 and Cry1Ac proteins are
located in the same cluster based on amino acid sequence identity {Crickmore, 1998 #8153}. ....................................................................... 64

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 8
List of Attachments
Attachment A: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment B: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment C: Event-specific Method for the Quantification of Maize MON 87460 Using Real-time PCR
Attachment D: Event-specific Method for the Quantification of Maize line MON 89034 Using Real-time PCR
Attachment E: Event-specific Method for the Quantification of Maize line MON 88017 Using Real-time PCR
Attachment F: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment G: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment H: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment I: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment J: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Attachment K: DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 9
PART I 1. BRIEF DESCRIPTION OF THE GENETICALLY MODIFIED ORGANISM 1.1 Include specific and common names of the organism, the country of origin
of the plant and a description of the genetically modified trait. Maize (Zea Mays L.) is a member of the tribe Maydae, which is included in the subfamily Panicoideae of the grass family Poaceae (formerly Gramineae). Figure 1 summarizes the taxonomic classification of maize and its close relatives. The genera included in the tribe Maydae include Zea and Tripsacum in the Western Hemisphere, and Coix, Polytoca, Chionachne, Schlerachne, and Trilobachne in Asia. Although some researchers have implicated the Asian genera in the origin of maize, the evidence for them is not as extensive and convincing as for the genera located in the Western Hemisphere.
Family - Poaceae Subfamily - Panicoideae Tribe - Maydae Western Hemisphere: I. Genus - Zea A. Subgenus - Luxuriantes 1. Zea luxurians (2n = 20) 2. Zea perennis (2n = 40) 3. Zea diploperennis (2n = 20) B. Subgenus - Zea 1. Zea mays (2n = 20) Subspecies 1. Z. mays parviglumis (2n = 20) 2. Z. mays huehuetenangensis (2n = 20)
3. Z. mays mexicana (Schrad.) (2n = 20) II. Genus – Tripsacum
A. Section – Tripsacum B. Section – Fasciculata Species Species 1. T. andersomii (2n = 64) 1. T. jalapense (2n = 72) 2. T. australe (2n = 36) 2. T. lanceolatum (2n = 72)
Varieties 3. T. fasciculatum (2n = 36) a) T. australe var. australe 4. T. maizar (2n = 36, 72) b) T. australe var. hirstum 5. T. pilosum (2n = 72)
3. T. bravum (2n = 36, 72) Varieties 4. T. cundinamarce (2n = 36) a). T. pilosum var. guatemalense 5. T. dactyloides (2n = 72) b). T. pilosum var. pilosum
Varieties a) T. dactyloides var. hispidum b) T. dactyloides var. dactyloides c) T. dactyloides var. meridonale d) T. dactyloides var. mexicanum
6. T. floridanum (2n = 36) 7. T. intermedium (2n = 72 8. T. manisuroides (2n = 72) 9. T. latifolium (2n = 36) 10. T. percuvianum (2n = 72, 90, 108) 11. T. zopilotense (2n = 36, 72)
Asia: I. Genera— Chionachne (2n = 20) Schlerachne (2n = 20) Coix (2n = 10, 20) Trilobachne (2n = 20) Polytoca (2n = 20) Tribe—Andropogoneae I. Genus - Manisuris
Figure 1. Taxonomic classification of maize and its close relatives

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 10
The genus Zea includes two sections: Luxuriantes and Zea. Maize (Zea mays L.) is a separate species within the subgenus Zea, along with three subspecies. All species within the genus Zea, except maize/corn, are different species of teosinte. Until recently, the teosinte species were included in the genus Euchlaena rather than the genus Zea. The other genus included in the Maydae tribe is Tripsacum. Tripsacum includes 16 species with a basic set of 18 chromosomes (n = 18); the different species of Tripsacum contain multiples of 18 chromosomes ranging from 2n = 36 to 2n = 108 (Figure 1). An additional species, Tripsacum hermaphrodita (Anthephora hermaphrodita), has been described in the literature but has not been formally included in this taxonomic classification1. Five genera are included in the tribe Maydeae that originated in Asia. Except for Coix, the basic chromosome number is n = 10. Within Coix, n = 5 and n = 10 have been reported. This is an application for commodity clearance approval of the multi-event stack MON 87460 × MON 89034 × MON 88017. Monsanto Company has used conventional breeding techniques to develop the combined trait maize product MON 87460 × MON 89034 × MON 88017 that confers drought tolerance, insect resistance and herbicide tolerance. MON 87460 × MON 89034 × MON 88017 provides insect protection against certain lepidopteran insect pests and corn rootworm larval feeding. Each biotechnology-derived trait contributes to the specific benefits of the final combined product as follows: MON 87460 is a drought tolerant maize that can exhibit reduced yield loss under water-limited conditions compared to conventional maize. Efficacy in MON 87460 is derived by expression of the inserted Bacillus subtilis (B. subtilis) cold shock protein B (CspB). MON 87460 also expresses neomycin phosphotransferase II (NPTII) from Escherichia coli (E. coli) as a selectable marker. MON 87460 was developed through Agrobacterium-mediated transformation of conventional maize embryos. The CspB protein expressed by MON 87460 binds RNA and helps to maintain plant cellular functions, thereby reducing yield loss under water-limited conditions by minimizing the effect of water limitation on photosynthesis, stomatal conductance, and carbon fixation. MON 89034 produces two insecticidal proteins that protect against feeding damage caused by European corn borer (Ostrinia nubilalis) and other lepidopteran insect pests, including Busseola fusca and Chilo partellus in South Africa. Cry1A.105 is a modified Bacillus thuringiensis (Bt) Cry1A protein and Cry2Ab2 is a Bt (subsp. kurstaki) protein. The combination of these two insecticidal proteins provides enhanced insect control and offers an additional insect-resistance management tool. MON 89034 was produced by Agrobacterium-mediated transformation of maize cells. MON 88017 produces a modified Bt (subsp. kumamotoensis) Cry3Bb1 protein to protect
against corn rootworm larval feeding. In addition, MON 88017 is a Roundup Ready®
maize that produces 5-enolpyruvylshikimate-3phosphate synthase protein from Agrobacterium sp. strain CP4 (CP4 EPSPS), which confers tolerance to glyphosate, the
active ingredient in Roundup®
agricultural herbicides. MON 88017 was produced by Agrobacterium-mediated transformation of maize cells. As mentioned previously it
1 The Plants Database on http://plants.usda.gov .

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 11
produces a modified Bacillus thuringiensis (subsp. kumamotoensis) MON 88017 Cry3Bb12 insecticidal protein that provides protection against certain coleopteran insect pests belonging to the Chrysomelidae family (corn rootworm, Diabrotica spp.). MON 88017 also produces the CP4 5-enolpyruvyl shikimate-3-phosphate synthase (CP4 EPSPS) protein that confers tolerance to glyphosate3. 2. COMMODITY CLEARANCE 2.1 Please indicate the type of clearance requested. This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 maize. The scope of the application is for all uses of MON 87460 × MON 89034 × MON 88017 maize derived grain, i.e. in food, feed and for processing. MON 87460 × MON 89034 × MON 88017 grain segregates for all genes, including those introduced by genetic modification. Thus, MON 87460 × MON 89034 × MON 88017 F2 grain includes the combined trait product and all sub-combinations of these events. 2.2 Detail specific instructions for the storage and handling of the plant or
plant parts. Information and data provided in this application support the conclusion that, except for the specifically introduced traits, MON 87460 × MON 89034 × MON 88017 is equivalent to conventional maize. Furthermore, South Africa is not the centre of origin for Zea mays and there are no wild relatives in South Africa with which maize can outcross. MON 87460 × MON 89034 × MON 88017 grain will therefore be stored and handled in the same manner as current commercial maize grain, both conventional and genetically modified. No specific measures are required for the storage and handling of MON 87460 × MON 89034 × MON 88017 derived maize grain. 2.3 When will commodity import take place? MON 87460 × MON 89034 × MON 88017 was approved for commercial planting in the USA and Canada at the end of 2011. Considering cultivation of this product in these countries, grain that may contain MON 87460 × MON 89034 × MON 88017 and its derivative segregating grain have been exported since 2013. Grain imports are made by international grain traders, with the time of importation dependent on the local or regional need for grain. The grain traders should, as per the requirements in terms of the Genetically Modified Organisms Act, 1997 (Act No. 15 of 1997), obtain the necessary permits from the Registrar prior to importation. 2.4 Where will commodity import take place? As indicated in section 2.3, grain imports are made by international grain traders and could potentially enter South Africa through any of the ports of entry.
2The Cry3Bb1 protein produced in MON 88017 and in MON 89034 × 1507 × MON 88017 × 59122. Hereafter referred to as Cry3Bb1.
3 Glyphosate is the active ingredient of Monsanto’s Roundup
® family of agricultural herbicides. Roundup is a registered
trademark of Monsanto Technology LLC.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 12
2.5 Detail the type of environment and the geographical areas for which the
plant is suited. Except for the introduced insect protection and herbicide tolerant traits, MON 87460 × MON 89034 × MON 88017 maize is equivalent to conventional maize and would therefore, in countries where this product is approved for commercial use, be grown in all the maize producing areas. However, this is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 and is not intended for use in the environment of South Africa. 2.6 Who will undertake the commodity import? As indicated in section 2.3, grain imports are made by international grain traders. 2.7 Estimate the amount of production of the genetically modified plant within
South Africa per annum, or the amount that will be imported into South Africa per annum.
This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 and is not intended for use in the environment of South Africa. Statistics from the South African Grain Information Services indicated that only 27 432 tons of maize was imported into South Africa in 2008-2009, 27 357 tons in 2009-2010 , no maize was imported in 2010-2011, 422 075 tons in 2011-2012 , 10 562 tons in 2012-2013 and 79 673 tons in 2013-2014 (http://www.sagis.org.za/). 3. DESCRIPTION OF ANY PRODUCT DERIVED FROM THE PLANT 3.1 Identify the part of the plant to be used for the product, the type of product,
and the use of the product, the market sector in which the product will be marketed and the trade name of the product.
The principal product of maize is the grain derived from the maize ear. The whole plant and immature ears are also used to make silage as cattle fodder. In addition, immature green maize ears are consumed by humans after a boiling or grilling process and are sold as “green maize” by supermarkets, hawkers and roadside vendors. Dry maize ears (12.5% moisture) can be processed in two separate ways:
Dry milling from which the following products are derived - o Maize meal (combination of germ, carbohydrate, fibre and fat) o Samp (pure carbohydrate with very little fibre) o Mielie rice (fat, mineral and vitamins – no protein) o Mielie grits (used by breweries to assist the fermentation process) o Mieliemeal flour (thickener in the canning industry)
Wet milling from which maize starch, syrup and dextrose is produced - o These products have a very wide range of uses in the industrial, food,
drug and cosmetics sectors. Considering the vast number of the trade

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 13
names of products containing starches, syrups and dextrose derived from the endosperm of maize available, a list is not provided.
Maize products have a very wide range of uses in the industrial, food; drug and cosmetics sectors. 3.2 Specify the exact conditions of use of the product. Maize consumption in South Africa can be divided into three categories:
Green maize for human consumption and silage production on a limited basis for cattle fodder.
Maize products derived from the dry milling process, which are maize meal, samp, mielie rice, grits and flour used for human consumption and the brewing and canning industry.
Maize products derived from the wet milling process. Only endosperm is used to produce starch, maize syrup and dextrose. These basic ingredients are used in the production of a multitude of end products.
3.3 Provide information on the proposed labelling of the product for marketing. This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 and is not intended for use in the environment of South Africa. The commercially cultivated maize, including any genetically modified (GM) events in the country of export, would determine which GM events would be in a consignment of grain destined for South Africa. As per the requirements of the Cartagena Protocol on Biosafety, if the consignment contains any genetically modified maize events, such as MON 87460 × MON 89034 × MON 88017, the consignment would need to be accompanied by documentation stating that it may contain GM maize. 3.4 State whether the benefits of the product are available in any other non-
genetically modified form. If so, state why the genetically modified form should be approved for general release when other, non-modified products are available.
There are no commercial maize hybrids that have been developed through conventional breeding practices in South Africa, which exhibit the drought tolerance, insect protection and herbicide tolerant traits present in MON 87460 × MON 89034 × MON 88017 maize. 3.5 Detail specific instructions for the storage and handling of GMO’s that will
avoid misuse or escape of the genetically modified plant into an environment for which it was not intended.
This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 and is not intended for use in the environment of South Africa. As indicated previously, grain imports are made by international grain traders, with the time of importation dependent on the local or regional need for grain. The grain traders would, as per the requirements in terms of the Genetically Modified Organisms Act, 1997 (Act No. 15 of 1997), obtain the necessary permits from the Registrar prior to

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 14
importation. Importation activities would thus be subjected to the conditions prescribed in the relevant import permits. 3.6 Detail the likelihood of the GMO being exported from South Africa,
particularly if such export could result in the introduction of the plant into its centre of origin.
According to information available from the South African Grain Information Services (http://www.sagis.org.za/), South Africa imports and exports maize. It is however unlikely that grain imported into South Africa would be exported again. South Africa does export to Mexico, the centre of origin of maize, where MON 87460 × MON 89034 × MON 88017 was approved for Import for food and feed use on 16 April 2012, approval ID 113300913X0014, by the Ministry of Health – Federal Commission for Protection from Sanitary Risks. 4. FOREIGN GENES AND GENE PRODUCTS 4.1 Identify all foreign genes in the genetically modified plant. MON 87460 × MON 89034 × MON 88017 maize was obtained by conventional breeding of three single maize event products: MON 87460, MON 89034 and MON 88017.
Like MON 87460, MON 87460 × MON 89034 × MON 88017 expresses cold shock protein B (CspB) and neomycin phosphotransferase II (NPTII), isolated from Bacillus subtilis and Escherichia coli, respectively.
Like MON 89034, MON 87460 × MON 89034 × MON 88017 expresses the Cry1A.105 and Cry2Ab2 insecticidal proteins, providing tolerance to inter alia maize stalk borers in South Africa.
Like MON 88017, MON 87460 × MON 89034 × MON 88017 expresses the Cry3Bb1 insecticidal protein that provides protection against certain coleopteran insect pests belonging to the Chrysomelidae family (corn rootworm, Diabrotica spp.) and the CP4 5-enolpyruvyl shikimate-3-phosphate synthase (CP4 EPSPS) protein that confers tolerance to glyphosate.
Detailed molecular information pertaining to MON 87460, MON 89034 and MON 88017 was provided in Monsanto’s applications for General Release of MON 87460, submitted on 14 July 2014; MON 89034 submitted on 8 June 2009 and Commodity Clearance of MON 88017 submitted on 20 December 2006. MON 87460 is currently under review by the Advisory Committee/Executive Council, MON 89034 was approved for general release on 19 October 2010 (Permit number 39.4(5/10/304) and MON 88017 was approved for commodity import on 26 September 2011 (Permit number 39.4(6/11/280). Thus, molecular information pertaining to the transformation of MON 87460, MON 89034 and MON 88017 was previously provided and has been or is in the process of been assessed by the Advisory Committee and Executive Council. However, for ease of reference, a summary of the information previously submitted for these three single events, is provided below. Molecular information on MON 87460 × MON 89034 × MON 88017 is provided at the end of this section.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 15
MON 87460 MON 87460 was produced by stable insertion of the coding sequence for cold shock protein B (CspB) from B. subtilis and a selectable marker, neomycin phosphotransferase II (NPTII) from E. coli making use of a binary plasmid vector, PV-ZMAP595. Plasmid vector PV-ZMAP595 contains a single T-DNAs delineated by left and right border regions that contains two expression cassettes: a cspB gene expression cassette, which contains the coding sequence for CspB from B. subtilis and a neomycin phosphotransferase II (nptII) expression cassette from E. coli, which confers resistance to kanamycin. The results of the detailed molecular characterization of MON 87460 maize support the following conclusions:
The DNA is inserted into the maize genome at a single locus.
The insert contains one functional copy of the cspB and nptII expression cassettes (Figure 2).
No elements were detected other than those that present in the T-DNA.
No backbone plasmid sequences were detected.
PCR and DNA sequence analyses supported the conclusion that only the two expected proteins, CspB and NPTII, are encoded by the DNA insert present in MON 87460.
Stability of the integrated DNA was demonstrated by the fact that the Southern blot fingerprint of MON 87460 was maintained in seven tested generations. These generations were also shown not to contain any detectable backbone sequence from plasmid PV-ZMAP595. Segregation analyses show inheritability and stability of the cspB and nptII genes occurred as expected across multiple generations, which corroborates the molecular insert stability analysis and establishes the genetic behaviour of the DNA insert at a single chromosomal locus.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 16
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Figure 2. Map of the Insert in MON 87460
MON 89034 MON 89034 was developed through Agrobacterium-mediated transformation of maize cells to produce the Bt insecticidal proteins Cry1A.105 and Cry2Ab2, making use of a binary plasmid vector, PV-ZMIR245. Plasmid vector PV-ZMIR245 contains two separate T-DNAs. The first T-DNA, designated as T-DNA I, contains the cry1A.105 and the cry2Ab2 expression cassettes. The second T-DNA, designated as T-DNA II, contains the nptII expression cassette that encodes the neomycin phosphotransferase enzyme (Rice et al., 2006). The results of the detailed molecular characterization of MON 89034 maize support the following conclusions:
The DNA is inserted into the maize genome at a single locus.
The insert contains one functional copy of the cry1A.105 and cry2Ab2 expression cassettes (Figure 3).
No elements were detected other than those that are present in T-DNA I.
No backbone plasmid DNA, nptII or other T-DNA II derived sequences were detected.
PCR and DNA sequence analyses supported the conclusion that only the two expected proteins, Cry1A.105 and Cry2Ab2, are encoded by the DNA insert present in MON 89034.
Stability of the integrated DNA was demonstrated by the fact that the Southern blot fingerprint of MON 89034 was maintained in four tested generations. Additionally, T-DNA II analysis of multiple generations indicated that there were no T-DNA II elements present other than those associated with T-DNA I. Furthermore, these generations have been shown not to contain any backbone sequence from plasmid PV-ZMIR245. This stability was further confirmed by demonstration of Mendelian inheritance of the Cry1A.105 and Cry2Ab2 proteins across generations.
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Figure 3. Map of the Insert in MON 89034
MON 88017

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 17
MON 88017 was produced by Agrobacterium-mediated transformation to produce the CP4 EPSPS and Cry3Bb1 proteins, making use of the plasmid vector PV-ZMIR39. Plasmid vector PV-ZMIR39 contains two expression cassettes: one coding for the native CP4 EPSPS protein from Agrobacterium sp. strain CP4 and the other coding for a variant of the wild-type Cry3Bb1 protein from Bacillus thuringiensis subsp. kumamotoensis. The results of the detailed molecular characterization of MON 88017 maize support the following conclusions:
The DNA is inserted into the maize genome at a single locus
The insert contains one functional copy of the cp4 epsps and cry3Bb1 gene expression cassettes (Figure 4).
No backbone plasmid DNA sequences were detected.
PCR and DNA sequence analyses supported the conclusion that only the two expected proteins, CP4 EPSPS and Cry3Bb1, are encoded by the DNA insert present in MON 88017.
Stability of the integrated DNA was demonstrated by the fact that the Southern blot fingerprint of MON 88017 was maintained across the branches of the breeding tree that were tested, confirming the stability of the integrated DNA over multiple generations.
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Figure 4. Map of the Insert in MON 88017

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 18
MON 87460 × MON 89034 × MON 88017 MON 89034 × MON 89034 × MON 88017 maize was obtained by conventional breeding of three single maize event products: MON 87460, MON 89034 and MON 88017. The inserted DNA fragments from the three single event maize products are inherited in MON 87460 × MON 89034 × MON 88017. The individual components and the size, source and function of these inherited DNA sequences are given in Table 1, Table 3 and Table 3. Schematic representations of MON 87460, MON 89034 and MON 88017 inserts are given in Figure 2, Figure 3 and Figure 4, respectively.
Table 1. Summary of inserted DNA inherited from MON 87460
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Table 2. Summary of inserted DNA inherited from MON 89034
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Table 3. Summary of inserted DNA inherited from MON 88017
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 19
MON 89034 × MON 89034 × MON 88017 contains three different inserts derived from the respective singles. There is low likelihood of molecular interactions between the different inserts and, therefore, low likelihood of any changes in the molecular characteristics of the inherited inserts in MON 87460 × MON 89034 × MON 88017 (e.g. insert number, copy number, absence of backbone DNA and integrity of the individual inserts). Therefore, it is not considered necessary to repeat the laboratory analyses of copy number and insert integrity for MON 87460 × MON 89034 × MON 88017. An analytical confirmation of the presence of the three inserts in the hybrid (insert-specific fingerprint by Southern blot analysis) is considered sufficient. Genomic DNA from MON 87460 × MON 89034 × MON 88017, MON 87460, MON 89034, MON 88017, and conventional corn were digested with the appropriate restriction enzymes and subjected to Southern blot analyses. The enzymes were chosen to produce two border fragments that together spanned the entire insert. The presence of these border fragments in the combined-trait product indicates that the intact insert was successfully inherited during breeding. Maps of plasmid vectors PV-ZMAP595, PV-ZMIR245, and PV-ZMIR39, annotated with the probes used for Southern analyses are presented in Figure 5, Figure 6 and Figure 7, respectively. Linear maps depicting restriction sites within the inserted DNA, as well as within the corn genomic DNA immediately flanking the inserts in MON 87460, MON 89034, and MON 88017 are presented in Figure 2, Figure 3 and Figure 4, respectively. For estimating the sizes of bands present, the molecular weight markers on the left of each Southern blot were used. Southern Blot Analysis to Verify the Presence of MON 87460 in MON 87460 × MON 89034 × MON 88017 (Attachment A) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act] cspB Coding Sequence Probe Test and control DNA samples were digested with the enzyme EcoR V. Approximately one genomic equivalent of PV-ZMAP595, previously digested with Blp I and Xba I, was mixed with conventional maize DNA digested with EcoR V to serve as the positive hybridization control. The blot was hybridized with a 32P-labeled probe containing the cspB coding sequence (Probe 1, Figure 5). The results are shown in Figure 8. The plasmid PV-ZMAP595 DNA digested with Blp I and Xba I produced one band of ~3.3 kb (Figure 8, Lane 1), consistent with the expected band of ~3.2 kb (Figure 5). The migration of the band was slightly higher than indicated by the molecular weight marker, possibly due to a difference in salt concentration between the DNA sample and the molecular weight marker (Sambrook and Russell, 2001). As expected, genomic DNA from conventional maize, MON 89034, and MON 88017 showed no detectable hybridization bands (Figure 8, Lanes 2, 3, and 4, respectively). MON 87460 produced a single band of ~7.2 kb (Figure 8, Lane 5), consistent with the expected band of > 2.2 kb, and consistent with the fingerprint band reported in Figure 4 by Skipwith (2007). MON 87460 × MON 89034 × MON 88017 also produced a band of ~7.2 kb (Figure 8, Lane 6), consistent with the expected band of > 2.2 kb (~7.2 kb). The data reported in Figure 7 verify the presence of the 5'-sequenceof MON 87460 in the combined-trait product MON 87460 × MON 89034 × MON 88017. nptII Coding Sequence Probe

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 20
Test and control DNA samples were digested with the enzyme EcoR V. Approximately one genomic equivalent of PV-ZMAP595, previously digested with Blp I and Xba I, was mixed with conventional maize DNA digested with EcoR V to serve as the positive hybridization control. The blot was hybridized with a 32P-labeled probe containing the nptII coding sequence (Probe 2, Figure 5). The results are shown in Figure 9. The plasmid PV-ZMAP595 DNA digested with Blp I and Xba I produced one band of ~6.0 kb (Figure 9, Lane 1), consistent with the expected band of ~6.1 kb (Figure 5). The migration of the band was slightly lower than indicated by the molecular weight marker, possibly due to a difference in salt concentration between the DNA samples and the molecular weight marker (Sambrook and Russell, 2001). As expected, genomic DNA from conventional maize, MON 89034, and MON 88017 showed no detectable hybridization bands (Figure 9, Lanes 2, 3, and 4, respectively). MON 87460 produced a single band of ~2.7 kb (Figure 9, Lane 5), consistent with the fingerprint band reported in Figure 4 by Skipwith (2007). MON 87460 × MON 89034 × MON 88017 also produced the expected band of ~2.7 kb (Figure 9, Lane 6). The data reported in Figure 9 verify the presence of the 3'-sequence of MON 87460 in the combined-trait product MON 87460 × MON 89034 × MON 88017. Together, the data reported in Figure 8 and Figure 9 verify the presence of MON 87460 in the combined-trait product MON 87460 × MON 89034 × MON 88017. Southern Blot Analysis to Verify the Presence of MON 89034 in MON 87460 × MON 89034 × MON 88017 (Attachment A) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act] Test and control DNA samples were digested with the enzyme Ssp I. Approximately one genomic equivalent of PV-ZMIR245, previously digested with Ssp I, was mixed with conventional maize DNA digested with Ssp I to serve as the positive hybridization control. The blot was hybridized simultaneously with two 32P-labeled probes; one containing a portion of the cry1A.105 coding sequence (Probe 3, Figure 6) and the other containing a portion of the cry2Ab2 coding sequence (Probe 4, Figure 6). The results are shown in Figure 10. The plasmid PV-ZMIR245 DNA digested with Ssp I produced two bands of ~ 7.0 kb and ~10.1 kb (Figure 10, Lane 1), consistent with the expected bands of ~7.2 kb and ~10.4 kb (Figure 6). As expected, genomic DNA from conventional maize, MON 87460 and MON 88017 showed no detectable hybridization bands (Figure 10, Lanes 2, 3, and 4, respectively). MON 89034 produced two bands of ~7.1 kb and ~8.0 kb (Figure 10, Lane 5), consistent with the fingerprint bands of ~8.2 kb and ~7.4 kb in Figure 3 as reported by Rice (2006). MON 87460 × MON 89034 × MON 88017 also produced two bands of ~7.1 kb and ~8.0 kb (Figure 10, Lane 6). The migration of the bands seen in lanes 1, 5, and 6 was slightly lower than indicated by the molecular weight marker, possibly due to a difference in salt concentration between the DNA samples and the molecular weight marker (Sambrook and Russell, 2001). The data reported in Figure 10 verify the presence of MON 89034 in the combined-trait product MON 87460 × MON 89034 × MON 88017. Southern Blot Analysis to Verify the Presence of MON 88017 in MON 87460 × MON 89034 × MON 88017 (Attachment A) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act] Test and control DNA samples were digested with the enzyme Xba I. Approximately one genomic equivalent of PV-ZMIR39, previously digested with Xba I, was mixed with

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 21
conventional maize DNA digested with Xba I to serve as the positive hybridization control. The blot was hybridized simultaneously with two 32P-labeled probes; one containing the TS-CTP2/CS-cp4 epsps coding sequence (Probe 5, Figure 7) and the other one containing the cry3Bb1 coding sequence (Probe 6, Figure 7). The results are shown in Figure 11. The plasmid PV-ZMIR39 DNA digested with Xba I produced one band of ~12.4 kb Figure 11, Lane 1), as expected (Figure 7). Genomic DNA from conventional maize, MON 87460, and MON 89034 showed no detectable hybridization bands (Figure 11, Lanes 2, 3, and 4, respectively). MON 88017 produced two bands of ~5.5 kb and ~7.4 kb (Figure 11, Lane 5), consistent with the fingerprint bands of ~5.5 kb and ~7.4 kb reported in Figure 6 by Groat (2006). MON 87460 × MON 89034 × MON 88017 also produced two bands of ~5.5 kb and ~7.4 kb (Figure 11, Lane 6), consistent with the fingerprint bands of ~5.5 kb and ~7.4 kb reported by Groat (2006). The data reported in Figure 11 verify the presence of MON 88017 in the combined-trait product MON 87460 × MON 89034 × MON 88017. Conclusions The results of this study show that the fingerprints obtained by Southern blot analysis of MON 87460 × MON 89034 × MON 88017 are the same as the corresponding fingerprints obtained with MON 87460, MON 89034, and MON 88017. These results demonstrate that MON 87460, MON 89034, and MON 88017 are present in the combined-trait maize product MON 87460 × MON 89034 × MON 88017.
In summary; MON 87460 × MON 89034 × MON 88017 contains:
The single DNA insert from the genome of MON 87460 (see Table 1 and Figure 2) containing the following elements:
o the cspB expression cassette consisting of the right border region, promoter and leader from the rice actin gene (P-Ract1), a non-translated intron from the rice actin gene (I-Ract1), the cspB coding sequence (CS-cspB) and a polyadenylation sequence from the transcript 7 gene (T-tr7).
o the nptII expression cassetteconsisting of two loxP sites and a transcriptional promoter (P-35S), the nptII coding sequence (CS-nptII), and a polyadenylation sequence from the nopaline synthase gene (T-nos).
The single DNA insert from the genome of MON 89034 (see Table 2 and Figure 3) composed of the following elements:
o the cry1A.105 expression cassette consisting of the P-e35S89 promoter and leader (0.30 kb), the L-Cab leader (0.06 kb), the I-Ract1 intron (0.48 kb), the CS-cry1A.105 coding sequence (3.53 kb) and the T-Hsp17 3’ transcript termination sequence (0.21 kb).
o the cry2Ab2 expression cassette consisting of the P-FMV promoter (0.56 kb), the I-Hsp 70 intron (0.80 kb), the CS-cry2Ab2 coding sequence (1.91 kb), the TS-SSU-CTP targeting sequence (0.40 kb) and the T-nos transcript termination sequence (0.25 kb).
The single DNA insert from the genome of MON 88017 (see Table 3 and Figure 4) composed of the following elements:
o the cp4 epsps expression cassette consisting of the P-Ract1 promoter (0.93 kb), the I-Ract1 intron (0.48* kb) enhancing the transcription, the TS-CTP 2 (0.23 kb) targeting sequence, the CS-cp4 epsps coding sequence (1.37 kb), the T-nos transcript termination sequence (0.25* kb).

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 22
o the cry3Bb1 expression cassette consisting of the P-e35 S promoter (0.61 kb), the L-Cab leader (0.06* kb) enhancing the translation, the I-Ract1 intron (0.48* kb) enhancing the transcription, the CS-cry3Bb1 coding sequence (1.96 kb), and the T-Hsp17 transcript termination sequence (0.21* kb). * The approximation of these genetic elements has been refined since the submission made in December 2006.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 23
Figure 5. Map of Plasmid Vector PV-ZMAP595
Circular map of the plasmid vector PV-ZMAP595 used to generate MON 87460. Genetic elements are depicted as arrows and are annotated on the exterior of the map. Restriction sites (with positions relative to the size of the plasmid vector) are shown for enzymes used in the Southern blot analysis on the exterior of the map. Probes used in the Southern blot analysis are shown as bold arcs on the interior of the map. Specific descriptions of the probes are given in the accompanying table.
Probe DNA Probe Start Position
End Position
Total Length (~kb)
1 CS-cspB Probe 4608 4811 0.2 2 CS-nptII Probe 5839 6635 0.8
PV-ZMAP5959379 bp
Blp I 2140
EcoR V 5041
Ssp I 4935
Ssp I 5016
Ssp I 7618
Xba I 5376
CS-rop
OR-ori-pBR322
aadA
B-Right Border
P-Ract1
I-Ract1
CS-cspB
T-tr7
loxP
P-35S
CS-nptII
T-nos
loxP
B-Left Border
OR-ori V
2
1

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 24
Figure 6. Map of Plasmid Vector PV-ZMIR245
Circular map of the plasmid vector PV-ZMIR245 used to generate MON 89034. Genetic elements are depicted as arrows and are annotated on the exterior of the map. Restriction sites (with positions relative to the size of the plasmid vector) are shown for enzymes used in the on the exterior of the map. Probes used in the are shown as bold arcs on the interior of the map. Specific descriptions of the probes are given in the accompanying table.
Probe DNA Probe Start Position
End Position
Total Length (~kb)
3 CS-cry1A.105 2942 4923 1.3 4 CS-cry2Ab2 9296 10509 1.2
PV-ZMIR24517600 bp
EcoR V 1714
EcoR V 2250
EcoR V 10783
EcoR V 12942
Ssp I 7921
Ssp I 15077
CS-aadA
B-Right Border
P-e35S
L-Cab
I-Ract1
CS-cry1A.105
T-Hsp17P-FMV
I-Hsp70
TS-SSU -CTP
CS-cry2Ab2
T-nos
B-Left Border
B-Right Border
T-nos
CS-nptII
P-35S
B-Left Border
OR-ori V
CS-rop
OR-ori-pBR322
Xba I 8176
Xba I 7471
Xba I 6490Xba I 12892
Xba I 1664
Xba I 2371
Xba I 3608
4
3

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 25
Figure 7. Map of Plasmid Vector PV-ZMIR39
Circular map of the plasmid vector PV-ZMIR39 used to generate MON 88017. Genetic elements are depicted as arrows and are annotated on the exterior of the map. Restriction sites (with positions relative to the size of the plasmid vector) are shown for enzymes used in the on the exterior of the map. Probes used in the are shown as bold arcs on the interior of the map. A detailed description of the probes is given in the accompanying table.
Probe DNA Probe Start Position
End Position
Total Length (~kb)
5 TS-CTP2/ CS-cp4 epsps 1423 3019 1.6 6 CS-cry3Bb1 4534 6510 2.0
PV-ZMIR3912368 bp
EcoR V 3842
Ssp I 11732
Xba I 3963
P-Ract1
I-Ract1
TS-CTP2
CS-cp4 epsps
T-nos
P-e35S
L-Cab
I-Ract1
CS-cry3Bb1
T-Hsp17
B-Right Border
aadA
OR-ori-pBR322
CS-rop
OR-ori V
B-Left Border
6

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 26
Figure 8. Southern Blot Analysis to Verify the Presence of MON 87460 in
MON 87460 × MON 89034 × MON 88017: CS-cspB Probe The blot was hybridized with a 32P-labeled probe that spanned the sequence of the CS-cspB. Each lane contains ~10 µg of genomic DNA digested with EcoR V. Lane designations are as follows: Lane 1: Conventional maize spiked with PV-ZMAP595 (Blp I/Xba I) [~1.0 genomic equivalent] 2: Conventional maize 3: MON 89034 4: MON 88017 5: MON 87460 6: MON 87460 × MON 89034 × MON 88017 Symbol denotes sizes of DNA, in kb, obtained from molecular weight markers on ethidium bromide stained gel.
654321
0.5
1.0
1.6
2.0
3.1
4.1
5.16.1
7.18.1
1520
40
10

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 27
Figure 9. Southern Blot Analysis to Verify the Presence of MON 87460 in
MON 87460 × MON 89034 × MON 88017: CS-nptII Probe The blot was hybridized with a 32P-labeled probe that spanned the sequence of the CS-nptII. Each lane contains ~10 µg of genomic DNA digested with EcoR V. Lane designations are as follows: Lane 1: Conventional maize spiked with PV-ZMAP595 (Blp I/Xba I) [~1.0 genomic equivalent] 2: Conventional maize 3: MON 89034 4: MON 88017 5: MON 87460 6: MON 87460 × MON 89034 × MON 88017 Symbol denotes sizes of DNA, in kb, obtained from molecular weight markers on ethidium bromide stained gel.
654321
0.5
1.0
1.6
2.0
3.1
4.1
5.16.1
7.18.1
1520
40
10

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 28
Figure 10. Southern Blot Analysis to Verify the Presence of MON 89034 in MON 87460 × MON 89034 × MON 88017: CS-cry1A.105 and CS-cry2Ab2 Probes
The blot was hybridized with 32P-labeled probes containing a portion of cry1A.105 and a portion of cry2Ab2. Each lane contains ~10 µg of genomic DNA digested with Ssp I. Lane designations are as follows: Lane 1: Conventional maize spiked with PV-ZMIR245 (Ssp I) [~1.0 genomic
equivalent] 2: Conventional maize 3: MON 87460 4: MON 88017 5: MON 89034 6: MON 87460 × MON 89034 × MON 88017 Symbol denotes sizes of DNA, in kb, obtained from molecular weight markers on ethidium bromide stained gel.
6 5 4 3 2 1
0.5
1.0
1.6
2.0
3.1
4.1
5.1
6.1 7.1
8.1
15 20
40
10

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 29
Figure 11. Southern Blot Analysis to Verify the Presence of MON 88017 in MON 87460 × MON 89034 × MON 88017: TS-CTP2/CS-cp4 epsps and CS-cry3Bb1 Probes
The blot was hybridized with 32P-labeled probes that spanned the sequence of the TS-CTP2/CS-cp4 epsps and CS-cry3Bb1. Each lane contains ~10 µg of genomic DNA digested with Xba I. Lane designations are as follows: Lane 1: Conventional maize spiked with PV-ZMIR39 (Xba I) [~1.0 genomic equivalent] 2: Conventional maize 3: MON 87460 4: MON 89034 5: MON 88017 6: MON 87460 × MON 89034 × MON 88017
Symbol denotes sizes of DNA, in kb, obtained from molecular weight markers on ethidium bromide stained gel.
654321
0.5
1.0
1.6
2.0
3.1
4.1
5.16.1
7.18.1
10
1520
40

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 30
4.2 Describe the gene products that are derived from the foreign genes. MON 87460 × MON 89034 × MON 88017 is produced by crossing MON 87460, MON 89034 and MON 88017 pants using conventional breeding methods. Like MON 87460, MON 87460 × MON 89034 × MON 88017 expresses cold shock protein B (CspB) and neomycin phosphotransferase II (NPTII) which were isolated from B. subtilis and E. coli, respectively. Like MON 89034, MON 87460 × MON 89034 × MON 88017 produces two distinct Bt proteins, Cry1A.105 and Cry2Ab2, which provide a dual effective dose against feeding damage caused by the key lepidopteran pest complex in maize: European corn borer (ECB, Ostrinia nubilalis), southwestern corn borer (SWCB, Diatraea grandiosella), African maize stalk borer (Busseola fusca) and corn earworm (CEW, Helicoverpa zea); other lepidopteran insect pests such as the fall armyworm (FAW, Spodoptera frugiperda) and sugarcane borer (SCB, Diatraea saccharalis). Cry1A.105 is a modified Bt Cry1A-type protein with overall amino acid sequence identity of 93.4%, 90%, and 76.7% to the Cry1Ac (subsp. kurstaki), Cry1Ab (subsp. kurstaki) and Cry1F (subsp. aizawai) proteins, respectively. Cry2Ab2 is a B. thuringiensis subsp. kurstaki protein. Like MON 88017, MON 87460 × MON 89034 × MON 88017 produces the modified Cry3Bb1 protein, derived from B. thuringiensis subsp. kumamotoensis that provides protection against corn rootworm (Diabrotica spp.) larval feeding and the CP4 EPSPS protein, derived from Agrobacterium sp. strain CP4, which provides tolerance to glyphosate. 4.3 Describe the biological activity associated with the foreign gene products. MON 87460 × MON 89034 × MON 88017 will combine multiple effective doses in multiple traits and provides season-long yield protection in four areas:
Abiotic stress tolerance, the MON 87460 component in MON 87460 × MON 89034 × MON 88017 provides a yield benefit when limited by water availability.
Above-ground insect control (lepidopteran protection), the MON 89034 component in MON 87460 × MON 89034 × MON 88017 provides protection against maize (corn) borers.
Below-ground insect control (coleopteran protection), the MON 88017 in MON 87460 × MON 89034 × MON 88017 provides protection against corn rootworm larval feeding.
Weed control, the MON 88017 component in
MON 87460 × MON 89034 × MON 88017 is a Roundup Ready event that produces a 5-enolpyruvylshikimate-3-phosphate synthase protein which confers tolerance to glyphosate, the active ingredient in Roundup® agricultural herbicides.
Roundup and Roundup Ready are registered trademarks of Monsanto Technology LLC.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 31
4.4 Provide information on the rate and level of expression of the foreign genes and the sensitivity of the measurement of the rate and level. State whether expression is constitutive or inducible. Are foreign genes expressed throughout the plant or only in certain organs or tissues?
The levels of CspB, NPTII, CP4 EPSPS, Cry1A.105, Cry2Ab2, and Cry3Bb1 proteins were assessed by validated enzyme-linked immunosorbent assays (ELISA) in maize tissues collected from MON 87460 × MON 89034 × MON 88017 produced during 2006-2007 in Chilean field trials under well-watered and water-limited conditions. Tissue samples were collected from plants grown at four field sites [Calera de Tango (CT), Colina (CL), Lumbreras (LUM) and Quillota (QUI)] in Chile under Production Plan 06-45-B3-02 (Adu-Tutu, 2008). The experiment was arranged in a strip plot design with three replicates per site, with two irrigation treatments (well-watered or water-limited). The well-watered irrigation treatment was managed to provide optimal grain yield. The water-limited irrigation treatment was managed to impose a drought stress by withholding irrigation during the late vegetative through early grain fill growth stages (i.e., approximately V10 through R2). Within each treatment, the CspB, NPTII, CP4 EPSPS, Cry1A.105, Cry2Ab2 and Cry3Bb1 protein levels were determined across sites (combined site analysis) and within site (individual site analysis). For the combined site analysis, each site had to meet a series of agronomic acceptance criteria to be included in the combined site average for protein expression of each tissue type. These criteria were: 1) water had to be managed appropriately to impose differential water treatments, 2) monitoring data had to confirm that the desired treatments were achieved, and 3) the reference materials had to exhibit phenotypic responses appropriate to the intended treatment. Three sites CT, CL and LUM met all of these criteria (Whitsel and Sammons, 2008). The fourth site, QUI, met the first two criteria but not the third and therefore, it was not possible to include this site in the combined-site analysis (Attachment B) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. CspB, CP4 EPSPS, Cry1A.105, Cry2Ab2, and Cry3Bb1 protein levels were determined in seven tissue types, including over season leaf (OSL-1), over season root (OSR-1), over season whole plant (OSWP-3), pollen, forage, forage root and grain; NPTII was determined only in OSL-1, OSR-1, forage and grain. The OSL-1 and OSR-1 samples were collected at growth stage V2-V4 and OSWP-3 samples were collected at growth stage V10-V12. The forage and forage root samples were collected at early dent stage and pollen samples were collected during pollen shedding. Grain samples were collected at crop maturity stage. All protein levels for all tissue types were calculated on a microgram (µg) per gram (g) fresh weight (fwt) basis. All µg/g fwt protein levels >LOQ were converted and reported on a dry weight (dwt) basis using dry weight conversion factors (DWCF). Results showed that the mean CspB protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 2.0 μg/g dwt and 1.8 μg/g dwt in OSL-1, 1.4 μg/g dwt and 1.5 μg/g dwt in OSR-1, 0.11 μg/g dwt and 0.17 μg/g dwt in forage, 0.050 μg/g dwt and 0.034 μg/g dwt in grain, 0.055 μg/g dwt and 0.10 μg/g dwt in forage root, 0.64 μg/g dwt and 0.50 μg/g dwt in OSWP-3, and 22 μg/g dwt and 23 μg/g dwt in pollen, respectively (Table 4).

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 32
Results showed that the mean NPTII protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 3.6 μg/g dwt and 3.7 μg/g dwt in OSL-1, 0.47 μg/g dwt and 0.51 μg/g dwt in OSR-1, 0.12 μg/g dwt and 0.14 μg/g dwt in forage, respectively. The mean NPTII protein levels in MON 87460 × MON 89034 × MON 88017 were below the assay limit of quantitation (0.0047 μg/g fwt) in grain across three sites under both well-watered and water-limited conditions (Table 5). Results showed that the mean CP4 EPSPS protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 210 μg/g dwt and 210 μg/g dwt in OSL-1, 190 μg/g dwt and 200 μg/g dwt in OSR-1, 61 μg/g dwt and 72 μg/g dwt in forage, 4.8 μg/g dwt and 4.8 μg/g dwt in grain, 85 μg/g dwt and 110 μg/g dwt in forage root, 130 μg/g dwt and 110 μg/g dwt in OSWP-3, and 260 μg/g dwt and 290 μg/g dwt in pollen, respectively (Table 6). Results showed that the mean Cry1A.105 protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 330 μg/g dwt and 360 μg/g dwt in OSL-1, 74 μg/g dwt and 93 μg/g dwt in OSR-1, 25 μg/g dwt and 30 μg/g dwt in forage, 5.4 μg/g dwt and 5.6 μg/g dwt in grain, 23 μg/g dwt and 22 μg/g dwt in forage root, 60 μg/g dwt and 64 μg/g dwt in OSWP-3, and 11 μg/g dwt and 12 μg/g dwt in pollen, respectively (Table 7). Results showed that the mean Cry2Ab2 protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 370 μg/g dwt and 340 μg/g dwt in OSL-1, 61 μg/g dwt and 69 μg/g dwt in OSR-1, 82 μg/g dwt and 94 μg/g dwt in forage, 3.0 μg/g dwt and 2.6 μg/g dwt in grain, 40 μg/g dwt and 41 μg/g dwt in forage root, 93 μg/g dwt and 82 μg/g dwt in OSWP-3, and 0.73 μg/g dwt and 0.89 μg/g dwt in pollen, respectively (Table 8). Results showed that the mean Cry3Bb1 protein levels in MON 87460 × MON 89034 × MON 88017 across three sites (CL, CT, and LUM) under well-watered and water-limited conditions were 310 μg/g dwt and 350 μg/g dwt in OSL-1, 260 μg/g dwt and 310 μg/g dwt in OSR-1, 45 μg/g dwt and 58 μg/g dwt in forage, 7.6 μg/g dwt and 7.5 μg/g dwt in grain, 37 μg/g dwt and 57 μg/g dwt in forage root, 130 μg/g dwt and 130 μg/g dwt in OSWP-3, and 13 μg/g dwt and 15 μg/g dwt in pollen, respectively (Table 9).

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 33
Table 4. Summary of CspB Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
0.35 (0.15) 2.0 (0.86) 0.32 (0.16) 1.8 (0.80)
0.18 - 0.60 1.0 - 3.3 0.17 - 0.56 1.1 - 2.9
OSR-1
0.14 (0.029) 1.4 (0.29) 0.14 (0.036) 1.5 (0.32)
0.11 - 0.19 1.1 - 1.9 0.083 - 0.19 0.92 - 1.8
Forage
0.026 (0.0041) 0.11 (0.019) 0.038 (0.0094) 0.17 (0.047)
0.020 - 0.035 0.084 - 0.15 0.022 - 0.050 0.095 - 0.23
Grain
0.043 (0.017) 0.050 (0.020) 0.030 (0.0094) 0.034 (0.011)
0.023 - 0.074 0.026 - 0.086 0.015 - 0.045 0.017 - 0.053
Forage Root
0.0075 (0.0018) 0.055 (0.014) 0.014 (0.0045) 0.10 (0.038)
0.0046 - 0.010 0.035 - 0.075 0.0067 - 0.022 0.042 - 0.17
OSWP-3
0.075 (0.026) 0.64 (0.21) 0.065 (0.024) 0.50 (0.19)
0.053 - 0.11 0.44 - 0.95 0.048 - 0.11 0.32 - 0.84
Pollen
16 (1.6) 22 (3.4) 16 (4.7) 23 (7.9)
13 - 19 17 - 26 8.9 - 24 12 - 36
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. OSWP-3:V10-V12 c. Grain: crop maturity d. Forage/Forage Root: early dent e. Pollen: during pollen shedding
2. The mean and standard deviation across three sites were calculated using the sample size (n=9).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a 5. fresh weight (fwt) basis.
Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 34
Table 5. Summary of NPTII Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
0.63 (0.16) 3.6 (1.0) 0.64 (0.23) 3.7 (1.4)
0.43 - 0.92 2.4 - 5.4 0.36 - 0.99 1.9 - 5.8
OSR-1
0.047 (0.0073) 0.47 (0.073) 0.049 (0.015) 0.51 (0.16)
0.037 - 0.058 0.37 - 0.58 0.025 - 0.066 0.22 - 0.71
Forage
0.029 (0.0049) 0.12 (0.020) 0.031 (0.0048) 0.14 (0.027)
0.021 - 0.034 0.091 - 0.14 0.025 - 0.037 0.099 - 0.17
Grain
<LOQ6 (N/A
7) N/A (N/A) <LOQ (N/A) N/A (N/A)
<LOD8 - 0.0051 N/A <LOD-<LOQ N/A
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. Grain: crop maturity c. Forage: early dent
2. The mean and standard deviation across three sites were calculated using the sample size (n=9).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a
fresh weight (fwt) basis. 5. Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values
were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.
6. NPTII protein levels were below the assay Limit of Quantitation (LOQ; 0.0047 μg/g fwt) in grain tissues.
7. Not applicable. The values <LOQ (μg/g fwt) were not converted to μg/g dwt. 8. NPTII protein levels were below the assay Limit of Detection (LOD; 0.0024 μg/g fwt) in
grain tissues.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 35
Table 6. Summary of CP4 EPSPS Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
37 (5.1) 210 (32) 36 (7.7) 210 (53)
29 - 47 160 - 280 27 - 48 140 - 300
OSR-1
19 (3.5) 190 (35) 20 (4.7) 200 (42)
13 - 26 130 - 260 13 - 28 150 - 250
Forage
14 (1.9) 61 (8.4) 17 (7.1) 72 (34)
12 - 17 51 - 72 11 - 33 44 - 150
Grain
4.2 (0.40) 4.8 (0.49) 4.2 (0.56) 4.8 (0.64)
3.7 - 4.9 4.3 - 5.7 3.0 - 4.9 3.5 - 5.6
Forage Root
11 (1.9) 85 (17) 15 (6.9) 110 (58)
8.0 - 13 54 - 100 8.4 - 32 53 - 240
OSWP-3
16 (3.6) 130 (34) 14 (3.6) 110 (23)
12 - 24 110 - 220 11 - 22 77 - 150
Pollen
190 (31) 260 (26) 200 (36) 290 (65)
150 - 230 230 - 310 140 - 250 220 - 400
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. OSWP-3:V10-V12 c. Grain: crop maturity d. Forage/Forage Root: early dent e. Pollen: during pollen shedding
2. The mean and standard deviation across three sites were calculated using the sample size (n=9).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a
fresh weight (fwt) basis. 5. Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values
were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 36
Table 7. Summary of Cry1A.105 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
59 (9.9) 330 (58) 62 (9.5) 360 (59)
39 - 68 220 - 390 44 - 78 280 - 490
OSR-1
7.4 (1.6) 74 (16) 8.8 (6.3) 93 (71)
5.6 - 11 56 - 110 3.1 - 25 34 - 270
Forage6
6.0 (1.1) 25 (4.8) 7.0 (2.3) 30 (11)
4.1 - 7.6 17 - 33 4.3 - 11 19 - 48
Grain
4.7 (0.82) 5.4 (0.92) 4.9 (0.89) 5.6 (1.0)
3.7 - 5.8 4.3 - 6.6 3.2 - 5.7 3.7 - 6.6
Forage Root7
3.1 (0.32) 23 (3.5) 3.0 (0.63) 22 (6.0)
2.5 - 3.5 17 - 27 2.4 - 4.4 15 - 34
OSWP-3
7.0 (1.4) 60 (10) 8.2 (2.2) 64 (14)
5.6 - 9.8 46 - 82 6.2 - 12 47 - 91
Pollen
8.2 (0.35) 11 (0.97) 8.4 (0.48) 12 (1.5)
7.7 - 8.9 10 - 13 7.7 - 9.2 10 - 15
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. OSWP-3:V10-V12 c. Grain: crop maturity d. Forage/Forage Root: early dent e. Pollen: during pollen shedding
2. The mean and standard deviation across three sites were calculated using the sample size (n=9, except n=8 for forage and forage root under water-limited condition).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a
fresh weight (fwt) basis. 5. Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values
were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.
6. One forage sample at site LUM under water-limited condition was < LOD (0.37 μg/g fwt). This sample is not included in the average, range and SD calculation.
7. One forage root sample at site LUM under water-limited condition was <LOD (0.25 μg/g fwt). This sample is not included in the average, range and SD calculation.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 37
Table 8. Summary of Cry2Ab2 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
65 (18) 370 (97) 59 (16) 340 (77)
46 - 98 270 - 550 37 - 82 220 - 430
OSR-1
6.1 (2.6) 61 (26) 6.6 (2.0) 69 (24)
3.5 - 11 35 - 110 3.7 - 9.8 41 - 110
Forage6
19 (5.4) 82 (23) 22 (3.8) 94 (21)
12 - 26 49 - 110 19 - 31 75 - 140
Grain
2.6 (0.51) 3.0 (0.61) 2.3 (0.64) 2.6 (0.73)
1.6 - 3.3 1.9 - 3.9 1.4 - 3.2 1.6 - 3.6
Forage Root7
5.5 (0.77) 40 (4.8) 5.7 (1.4) 41 (12)
4.6 - 7.0 36 - 47 3.7 - 7.6 23 - 59
OSWP-3
11 (2.1) 93 (17) 11 (3.5) 82 (24)
8.7 - 15 72 - 120 7.2 - 18 48 - 140
Pollen
0.53 (0.10) 0.73 (0.081) 0.61 (0.13) 0.89 (0.18)
0.40 - 0.65 0.64 - 0.87 0.46 - 0.88 0.70 - 1.1
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. OSWP-3:V10-V12 c. Grain: crop maturity d. Forage/Forage Root: early dent e. Pollen: during pollen shedding
2. The mean and standard deviation across three sites were calculated using the sample size (n=9, except n=8 for forage under water-limited condition and forage root under water-limited condition).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a
fresh weight (fwt) basis. 5. Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values
were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.
6. One forage sample at site LUM under water-limited condition had a low expression level of Cry2Ab2 (<10% of the average). This sample is not included in the average, range and SD calculation.
7. One forage root sample at site LUM under water-limited condition was <LOQ (0.22 μg/g fwt). This sample is not included in the average, range and SD calculation.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 38
Table 9. Summary of Cry3Bb1 Protein Levels in Maize Tissues Collected from MON 87460 × MON 89034 × MON 88017 Produced in a Chilean Field Trial (CL, CT, and LUM Sites) Conducted in 2006-2007 under Well-Watered and Water-Limited Conditions
Tissue Type1
MON 87460 × MON 89034 × MON 88017
Well-Watered Water-Limited
Mean (SD)2
Range3
(μg/g fwt)4
Mean (SD) Range
(μg/g dwt)5
Mean (SD) Range
(μg/g fwt)
Mean (SD) Range
(μg/g dwt)
OSL-1
55 (13) 310 (76) 61 (15) 350 (85)
39 - 79 220 - 440 38 - 81 220 - 470
OSR-1
26 (3.4) 260 (34) 30 (8.3) 310 (64)
21 - 32 210 - 320 16 - 46 180 - 410
Forage6
11 (2.0) 45 (8.9) 14 (3.9) 58 (19)
7.6 - 14 32 - 60 9.9 - 21 43 - 96
Grain
6.6 (1.2) 7.6 (1.4) 6.5 (0.98) 7.5 (1.1)
4.7 - 8.6 5.4 - 9.8 5.1 - 8.1 5.9 - 9.2
Forage Root7
5.1 (0.89) 37 (7.5) 7.7 (4.0) 57 (33)
3.8 - 6.3 25 - 47 4.1 - 15 26 - 110
OSWP-38
15 (1.5) 130 (13) 17 (3.8) 130 (32)
13 - 18 110 - 150 13 - 24 88 - 190
Pollen
9.6 (1.6) 13 (1.1) 10 (1.2) 15 (2.2)
7.5 - 12 12 - 15 8.8 - 12 13 - 18
1. Tissues were collected at the following growth stages (Ritchie et al., 1997): a. OSL-1/OSR-1: V2-V4 b. OSWP-3:V10-V12 c. Grain: crop maturity d. Forage/Forage Root: early dent e. Pollen: during pollen shedding
2. The mean and standard deviation across three sites were calculated using the sample size (n=9, except n=8 for forage under water-limited condition, forage root under water-limited condition, and OSWP-3 under well-watered condition).
3. Minimum and maximum values were determined for each tissue type across sites. 4. Protein levels are expressed as microgram (μg) of protein per gram (g) of tissue on a
fresh weight (fwt) basis. 5. Protein levels are expressed as μg/g on a dry weight (dwt) basis. The dry weight values
were calculated by dividing the “μg/g fwt” values by the dry weight conversion factors obtained from moisture analysis data.
6. One forage sample at site LUM under water-limited condition had a low expression level of Cry3Bb1 (<5% of the average). This sample is not included in the average, range and SD calculation.
7. One forage root sample at site LUM under water-limited condition was <LOD (0.28 μg/g fwt). This sample is not included in the average, range and SD calculation.
8. One OSWP-3 sample at site CT under well-watered condition had a low expression level of Cry3Bb1 (<10% of the average). This sample is not included in the average, range and SD calculation.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 39
4.5 Provide protocols for the detection of the foreign genes in the environment including sensitivity, reliability and specificity of the techniques.
MON 87460 × MON 89034 × MON 88017 was produced by crossing plants containing MON 87460, MON 89034 and MON 88017 by use of conventional breeding methods. Therefore, MON 87460 × MON 89034 × MON 88017 is detectable using the combination of the single event-specific PCR method for detecting the introduced DNA present in the single events MON 87460, MON 89034 and MON 88017. For all plants in which two or more events are combined by conventional breeding, the events involved will segregate in the grain; therefore such detection methods when applied to individual grains harvested from fields of MON 87460 × MON 89034 × MON 88017 will detect any combination of the three events. Event specific detection methods for detection of MON 87460, MON 89034 and MON 88017 DNA have been validated by the European Commission Joint Research Centre (EU JRC) and are available on the EU JRC website at: http://gmo-crl.jrc.ec.europa.eu/ For ease of reference a copy of the three detection methods are provided in Attachments C, D and E (DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]).
5. RESISTANCE 5.1 Detail whether the genetically engineered plant is able to initiate resistance,
in any biotic component of the environment, to any biologically active foreign gene product.
This is an application for commodity clearance of MON 87460 × MON 89034 × MON 88017, i.e. use as food, feed or for processing. This is not an application for release of MON 87460 × MON 89034 × MON 88017 into the environment of South Africa and therefore it is unlikely that the genetically engineered plant is able to initiate resistance in any biotic component of the environment or to any biologically active foreign gene product. 5.2 Detail what methods are available to minimise the risk of resistance
developing in the environment. Considering that this is not an application for environmental release, this question is not applicable. 5.3 Detail how resistance will be managed during release of the genetically
modified plant. Considering that this is not an application for environmental release, this question is not applicable.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 40
6. HUMAN AND ANIMAL HEALTH
6.1 State whether the genetically modified plant or its products will enter human or animal food chains.
This is an application for commodity clearance of MON 87460 × MON 89034 × MON 88017 and will therefore enter the human and animal food chains. 6.2 Detail the results of experiments undertaken to determine the toxicity of the
foreign gene products (including marker genes) to humans and animals. The safety assessment of biotechnology-derived crops addresses two major areas viz. (i) the safety of the introduced trait and (ii) the safety of the derived food and feeds. MON 87460 × MON 89034 × MON 88017 was produced by crossing plants containing MON 87460, MON 89034 and MON 88017 using conventional breeding methods. Full details pertaining to the potential toxicity and allergenicity of the CspB and NPTII proteins in MON 87460; Cry1A.105 and Cry2Ab2 proteins in MON 89034 and the Cry3Bb1 and CP4 ESPSPS proteins in MON 88017, was previously submitted in terms of Monsanto’s applications for General Release of MON 87460 (Submitted to the Registrar’s office for review on 14 July 2014), MON 89034 (17/3/1-Monsanto-09-649 on 8 June 2009) and Commodity Clearance of MON 88017 (17/3/1-Monsanto-07/649 on 20 December 2006). Section 6.2 is divided into three main parts:
Toxicological assessment
Allergenicity assessment
Nutritional assessment Toxicological Assessment The assessment of potential toxicity of a protein is based on the established premise that a protein is not likely to have a toxic effect if:
The protein has a demonstrated history of safe use.
The protein has no structural similarity to known toxins or other biologically active proteins that could cause adverse effects in humans or animals.
The protein does not exert any acute toxicity to mammals.
The protein is rapidly digested in mammalian gastrointestinal systems. MON 87460 Data regarding the toxicology of the CspB and NPTII proteins expressed in MON 87460 × MON 89034 × MON 88017 has been previously submitted as part of Monsanto’s application for General Release of MON 87460 (dossier submitted 14 July 2014). As the data is still under review and for ease of reference the relevant section is repeated below: In the following sections, the mode of action, the history of safe use and potential mammalian toxicity of the CspB and NPTII proteins produced in MON 87460 are discussed. General information regarding the methods used to evaluate the structural similarity to known toxins and acute toxicity is also provided below. The results of the mammalian toxicity studies conducted with these proteins are compared to the anticipated levels of human exposure. Potential health risks were evaluated by calculating a Margin of Exposure (MOE) between the No Observable

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 41
Adverse Effect Level (NOAEL) from acute mouse gavage studies with the CspB and NPTII proteins and a conservative estimate of human dietary exposure. Characterization of the CspB and NPTII proteins in MON 87460 and equivalence of the plant- and E. coli-produced CspB and NPTII proteins The safety assessment of crops improved through biotechnology includes characterization of the introduced protein(s), confirmation of their functional and physicochemical properties, and confirmation of the safety of each protein produced from the inserted DNA. Levels of introduced proteins in planta are usually too low to allow purification of sufficient quantities for use in safety assessment studies. Therefore, it is necessary to produce the proteins in high-expressing recombinant host systems (such as bacteria) in order to obtain sufficient quantities of the protein(s) of interest. The approach of using a heterologously produced protein as a surrogate for a plant-expressed protein in safety testing is a well-established and widely accepted strategy (Codex Alimentarius Commission, 2003; FAO/WHO, 2000; König et al., 2004). Proteins produced by bacterial systems are engineered to match the amino acid sequences of their counterparts expressed in the biotechnology-improved plants. Prior to use of the heterologously produced protein in safety assessment, additional studies on the physicochemical and functional equivalence of plant- and bacteria-produced proteins need to be performed. Characterization and equivalence of the plant- and E. coli-produced CspB proteins The CspB protein was produced in E. coli and subsequently purified, and characterized. A small quantity of the CspB protein was also purified from grain of MON 87460. The identity of the MON 87460-derived protein was confirmed and its physicochemical properties and functional activity were compared to those of the bacteria-produced protein standard (Burzio et al., 2008b)(Attachment F) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. The identity of the plant-produced CspB protein was verified by:
1. N-terminal sequencing by Edman degradation chemistry; 2. Tryptic peptide mapping followed by MALDI-TOF MS analysis; 3. Western blot analysis with CspB specific antibodies; 4. SDS-PAGE analysis to assess the apparent molecular weight of the protein; 5. Functional dsDNA-destabilizing (“melting”) assay to demonstrate biological activity
of the CspB protein.
Equivalence between the E. coli- and the MON 87460-produced proteins was established by: 1. SDS-PAGE analysis to confirm equivalent molecular weight; 2. Western blot analysis to confirm equivalent immunoreactivity; 3. Glycosylation analysis to confirm the presence or absence of covalently linked
carbohydrates in both proteins; 4. A functional ds DNA-destabilizing (“melting”) assay to confirm equivalent biological
activity. N-terminal sequence analysis N-terminal sequencing of the first 15 amino acids performed on MON 87460-produced CspB protein resulted in the sequence expected for the CspB protein (Table 10) with the exception of the N-terminal methionine, which was not detected. This result is expected as removal of the N-terminal methionine, catalyzed by methionine aminopeptidase, is a common modification that occurs co-translationally before completion of the nascent protein chain and has no effect on protein structure or activity (Arfin and Bradshaw, 1988; Polevoda and Sherman, 2000). The N-terminal sequence information, therefore, confirms the identity of the CspB protein isolated from MON 87460 and the intactness of its N-terminus.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 42
MALDI-TOF mass spectrometry analysis The identity of the MON 87460-produced CspB protein was further confirmed by tryptic peptide mass mapping analysis using matrix assisted laser desorption ionization - time of flight (MALDI-TOF) mass sprectometry (MS). In general, protein identification made by peptide mapping is considered to be reliable if the measured coverage of the sequence is 15% or higher with a minimum of five matched peptides (Jensen et al., 1997). Observed tryptic peptides were considered a match to the expected tryptic mass when differences in molecular weight of less than one Dalton (Da) were found between the observed and predicted fragment masses. Such matches were made without consideration for potential natural amino acid modifications such as glycosylation. The protein sample was heat-denatured, chemically reduced, alkylated, digested with trypsin, guanidinated, and the masses of the tryptic peptides were measured. CspB is a small protein with a limited number of the trypsin-digested peptides that are amenable to the identification by MALDI-TOF. There were four unique peptide fragments identified that corresponded to the expected masses of the CspB trypsin-digested peptides. The identified masses were used to assemble a coverage map indicating the matched peptide sequences for the entire CspB protein (Figure 12), resulting in ~88% (58 out of 66 amino acids) coverage of the total protein. This analysis confirmed the identity of the MON 87460-produced CspB protein. CspB protein immunoreactivity A western blot analysis using goat anti-CspB serum was conducted to determine the relative immunoreactivity of the MON 87460-produced CspB protein and the E. coli-produced CspB reference standard. The results demonstrated that the anti-CspB antibody recognized the MON 87460-produced CspB protein that migrated to a similar position as the E. coli-produced reference standard protein (Figure 13). Furthermore, the immunoreactive signal increased with increasing levels of CspB protein loading. The observed immunoreactivities between the MON 87460- and E. coli-produced proteins were similar based on densitometric analysis of the western blot. Based on the above analysis, the MON 87460- and E. coli-produced CspB proteins demonstrated equivalent immunoreactive properties, which confirmed the identity and equivalence of the two proteins. CspB protein molecular weight equivalence The equivalence in apparent molecular weight of the purified MON 87460- and the E. coli-produced CspB proteins was demonstrated using SDS-PAGE (Figure 14). The MON 87460-produced CspB protein migrated with a molecular weight indistinguishable to that of the E. coli-produced protein standard analyzed concurrently (Figure 14). Based on comparable electrophoretic mobilities, the MON 87460- and E. coli-produced CspB proteins were determined to have equivalent apparent molecular weights. The predicted mass of the MON 87460-produced CspB protein was also confirmed by MALDI-TOF MS (Figure 12). The average mass obtained for CspB was 7220 Da. This experimentally obtained mass differs from the theoretical mass calculated for the CspB reference standard protein by 131 Da. The difference between the expected and the observed mass for MON 87460-produced CspB corresponds to the mass of methionine (131 Da). The absence of the N-terminal methionine was confirmed by N-terminal sequencing (Table 10). CspB protein glycosylation equivalence Some eukaryotic proteins are post-translationally modified by the addition of carbohydrate moieties (Rademacher et al., 1988). These carbohydrate moieties may be complex, branched polysaccharide structures, simple oligosaccharides or monosaccharides. In contrast, prokaryotic

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 43
organisms such as non-virulent E. coli strains used for cloning and expression purposes, lack the necessary biochemical synthetic capacity required for protein glycosylation. Therefore, determining whether the MON 87460-produced CspB protein is equivalent to the E. coli-produced CspB protein requires an investigation of its glycosylation status. To assess whether potential post-translational glycosylation of the MON 87460-produced CspB protein occurred, the purified protein sample was subjected to glycosylation analysis. The E. coli-produced CspB reference standard represented a negative control. The positive controls were the transferrin and horseradish peroxidase (HRP) proteins which are known to have multiple covalently-linked carbohydrate modifications. The transferrin protein and HRP, as well as the purified CspB protein isolated from MON 87460 and E. coli were separated on SDS-PAGE, transferred to a PVDF membrane, and glycosylation analysis was performed to detect carbohydrate moieties on the proteins. The results of this analysis are shown in Figure 15. The positive controls, transferrin and HRP, were detected at the expected molecular weights of ~75 and ~50 kDa, respectively, in a concentration-dependent manner (Figure 15, Panel A, Lanes 4-5 and 2-3). No detectable signal was observed for the MON 87460- and E. coli-produced CspB proteins (Figure 15, Panel A, Lanes 6-7 and 8-9). To confirm that sufficient MON 87460- and E. coli-produced CspB proteins were present for carbohydrate detection and glycosylation analysis, the membrane was stained with SYPRO Ruby stain to detect proteins (Figure 15, Panel B). Both MON 87460- and E. coli-produced CspB were clearly detected on the membrane (Figure 15, Panel B, Lanes 6-9). These results indicate that the MON 87460-produced CspB protein is not glycosylated and, thus is equivalent to the E. coli-produced CspB reference standard . CspB protein functional activity equivalence The functional activities of the E. coli- and MON 87460-produced CspB proteins were estimated using an assay where the protein destabilizes or “melts” a DNA-hairpin structure. In this assay protein specific activity is expressed as the amount (pmol) of open Dual Labeled Probe (DLP)
that is induced by a microgram (g) of CspB. The E. coli- and MON 87460-produced CspB proteins were considered functionally equivalent if the specific activity of one protein was within 25% (assay variability) of the other. Assay activity is expressed as the amount of an DLP that is destabilized by CspB. DLP consists of a custom synthesized 35-base oligonuclotide DNA fragment with a fluorescein amidite derived from 6-carboxyfluorescein (6-FAM) label at the 5’ end and a black hole quencher at the 3’ end. The oligonucleotide probe forms a double strand stem of six base pairs due to the complementary bases located at the 5’ and 3’ ends. The 23 nucleotides (dT) in the middle form a loop. CspB has been shown to have a high affinity for poly dT sequences (Max et al., 2006) and its binding to the loop will destabilize the double strands of the probe, which separates the fluorophore from the quencher, allowing fluorescence to be emitted and measured.
Results indicated that the specific activity was 0.660 pmole open DLP/g protein for the
MON 87460-produced CspB, and 0.757 pmole open DLP/g protein for the E. coli-produced reference standard. The difference in specific activities was 12.8% (Table 11) which is within assay variability. These results clearly demonstrate that the CspB proteins derived from MON 87460 and E. coli have equivalent functional activities. Conclusions on the characterization and equivalence of the MON 87460-produced and E. coli-produced CspB proteins A comparison of the MON 87460-produced CspB to the E. coli-produced CspB reference protein standard confirmed the identity of the MON 87460-produced CspB protein and established the

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 44
equivalence of the plant produced protein to the E. coli-produced CspB reference protein standard (Burzio et al., 2008b)(Attachment F) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. The molecular weight of the MON 87460- and E. coli-produced CspB proteins was estimated by SDS-PAGE. The SDS-PAGE demonstrated that the proteins migrated at the same molecular weight, indicating that the CspB proteins from both sources are equivalent in their molecular weight. The electrophoretic mobility and immunoreactive properties of the MON 87460-produced CspB protein were shown to be equivalent to those of the E. coli-produced CspB reference standard. The N-terminus of the MON 87460-produced CspB was consistent with the predicted amino acid sequence translated from the cspB coding sequence, and the MALDI-TOF mass spectrometry analysis yielded peptide masses consistent with the expected peptide masses from the translated cspB coding sequence. The MON 87460- and the E. coli-produced CspB reference standard were also found to be equivalent based on the functional activities and the lack of glycosylation. Taken together, these data provide a detailed characterization of the CspB protein isolated from MON 87460 and established its equivalence to the E. coli-produced CspB reference protein standard.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 45
Table 10. N-terminal amino acid sequence analysis of the CspB protein purified from grain tissue of MON 87460
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]
Figure 12. MALDI-TOF MS coverage map of the CspB protein isolated from MON 87460
Figure 13. Western Blot analysis of MON 87460- and E. coli-produced CspB proteins Aliquots of the purified, MON 87460- and E. coli-produced CspB proteins were separated by SDS-PAGE, and electrotransferred to a PVDF membrane. The membrane was probed with goat anti-CspB serum and developed using an ECL system (GE Healthcare). Approximate molecular weights (kDa) of markers loaded in Lane 1 are shown on the left side of the blot. The 1 min exposure is shown.
Lane Sample Amount
Loaded (ng) 1 See Blue® Plus2 Pre-Stained molecular weight markers 2 E. coli-produced CspB reference standard 3 3 E. coli-produced CspB reference standard 3 4 E. coli-produced CspB reference standard 6 5 E. coli-produced CspB reference standard 6 6 E. coli-produced CspB reference standard 9 7 E. coli-produced CspB reference standard 9 8 MON 87460-produced CspB protein 3 9 MON 87460-produced CspB protein 3 10 MON 87460-produced CspB protein 6 11 MON 87460-produced CspB protein 6 12 MON 87460-produced CspB protein 9 13 MON 87460-produced CspB protein 9
CspB protein
210
105
78 55
45
34
17 16
7
4
1 2 3 4 5 6 7 8 9 10 11 12 13

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 46
Figure 14. SDS-PAGE of E. coli-and MON 87460-produced CspB proteins
Aliquots of the MON 87460-produced CspB and the E. coli-produced CspB reference standard were separated by a tricine 10-20% polyacrylamide gradient gel and stained with an Owl Silver Staining kit. Approximate molecular weights (kDa) of markers loaded in Lanes 1 and 18 are shown on the left side of the gel.
Lane Sample Amount loaded (ng) (µl)
1 See Blue® Plus2 Pre-Stained molecular weight markers 15
2 E. coli-produced CspB standard 10
3 E. coli-produced CspB standard 10
4 E. coli-produced CspB standard 20
5 E. coli-produced CspB standard 20
6 E. coli-produced CspB standard 30
7 E. coli-produced CspB standard 30
8 E. coli-produced CspB standard 40
9 E. coli-produced CspB standard 40
10 E. coli-produced CspB standard 60
11 E. coli-produced CspB standard 60
12 MON 87460-produced CspB protein 10
13 MON 87460-produced CspB protein 10
14 MON 87460-produced CspB protein 20
15 MON 87460-produced CspB protein 20
16 MON 87460-produced CspB protein 30
17 MON 87460-produced CspB protein 30
18 See Blue® Plus2 Pre-Stained molecular weight markers 15
kDa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
210
105
78
55
45
34
17
16
7
4
CspB protein
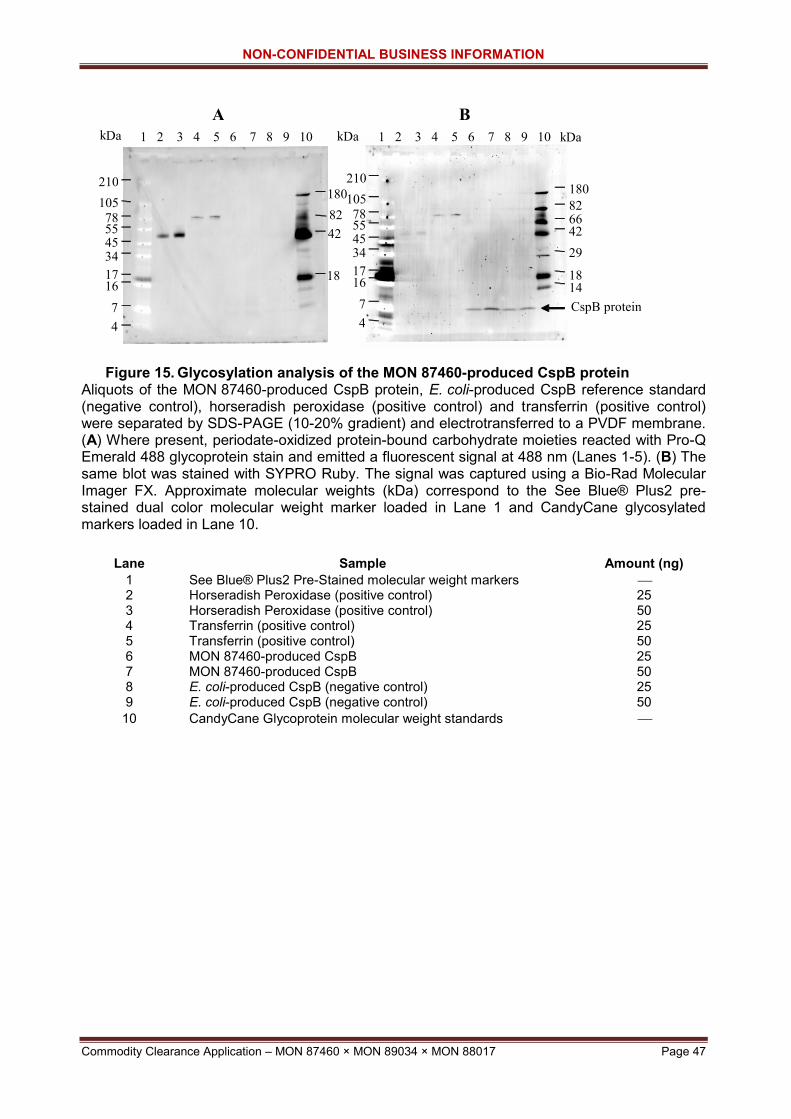
NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 47
Figure 15. Glycosylation analysis of the MON 87460-produced CspB protein Aliquots of the MON 87460-produced CspB protein, E. coli-produced CspB reference standard (negative control), horseradish peroxidase (positive control) and transferrin (positive control) were separated by SDS-PAGE (10-20% gradient) and electrotransferred to a PVDF membrane. (A) Where present, periodate-oxidized protein-bound carbohydrate moieties reacted with Pro-Q Emerald 488 glycoprotein stain and emitted a fluorescent signal at 488 nm (Lanes 1-5). (B) The same blot was stained with SYPRO Ruby. The signal was captured using a Bio-Rad Molecular Imager FX. Approximate molecular weights (kDa) correspond to the See Blue® Plus2 pre-stained dual color molecular weight marker loaded in Lane 1 and CandyCane glycosylated markers loaded in Lane 10.
Lane Sample Amount (ng)
1 See Blue® Plus2 Pre-Stained molecular weight markers 2 Horseradish Peroxidase (positive control) 25 3 Horseradish Peroxidase (positive control) 50 4 Transferrin (positive control) 25 5 Transferrin (positive control) 50 6 MON 87460-produced CspB 25 7 MON 87460-produced CspB 50 8 E. coli-produced CspB (negative control) 25 9 E. coli-produced CspB (negative control) 50
10 CandyCane Glycoprotein molecular weight standards
1 2 3 4 5 6 7 8 9 10
180
82
42
18
k Da k Da 1 2 3 4 5 6 7 8 9 10 k Da
210
105 78 55 45 34
17 16
7
4
210
105 78 55 45 34
17 16
7
4
180
82
42
18
66
29
14
CspB protein
A B

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 48
Table 11. CspB functional assay Assay activity is expressed as the amount (pmol) of open Dual Labeled Probe (DLP) that is
induced by a microgram (g) of CspB. The probe consists of a custom synthesized 35-base oligonuclotide DNA fragment with a 6-FAM fluorescent label at the 5’ end and a black hole quencher at the 3’ end. The fluorescence originated from the open DLP was determined with an excitation wavelength of 485 nm and an emission wavelength of 520 nm. The amount of open probe was determined relative to the standard curve prepared from serial dilutions of 6-FAM. The opening of the DLP was measured for both proteins, using 3 µg of CspB for each replicate. The activity represents the average of three replicates.
Specific Activity (pmoles opened DLP/µg CspB)
% Difference 1
(MON 87460 vs. E. coli) CspB MON 87460 - CspB E. coli - CspB
0.660 0.757 12.8
1 Percent difference was calculated as follows:
DifferencecoliEActivity
PlantActivitycoliEActivity%100
._
_._

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 49
Characterization and equivalence of the plant- and E. coli-produced NPTII proteins Protein safety assessment that was previously conducted using E. coli-produced NptII protein established safety of the NptII protein for human and animal consumption (Fuchs et al., 1993a; Fuchs et al., 1993b). Since the same NptII protein is expressed in MON 87460, the previously generated safety data support safety of the MON 87460-produced NptII protein. To use the existing protein safety data for the MON 87460-produced NptII protein, equivalence between E. coli-produced and MON 87460-produced proteins has to be demonstrated. The level of expression of the NptII protein in the grain of MON 87460 was below the limit of detection of the current validated ELISA. Such low level expression prevents the purification of sufficient amounts of the NptII protein from grain and the ability to detect the protein in grain extracts. However, leaves from MON 87460 express levels of NptII that are compatible with the detection of the NptII protein by western blot analysis. Hence, a western blot bridging strategy was utilized to demonstrate equivalence of the MON 87460-produced NptII protein and E. coli-produced NptII protein (Gu et al., 2008). This strategy is based on the demonstration of equivalent immunoreactivity with NptII-specific antibodies and equivalent apparent molecular weights of NptII protein in plant extracts compared to the E. coli-produced NptII reference protein standard. Immunoreactivity of the NptII protein The expression levels of NptII protein in leaf tissue of MON 87460 allowed detection of the protein with an NptII-specific antibody directly in leaf extracts without additional enrichment. An extract was also prepared from a leaf sample of conventional maize with a similar genetic background to MON 87460, which served as a negative control for the presence of the NptII protein. To ensure that the electromobility of the NptII protein had not been altered due to a matrix effect, the reference substance was spiked into the leaf extract from conventional maize and analyzed alongside the leaf extract from MON 87460. The leaf extracts from MON 87460, E. coli-produced NptII protein, and NptII-spiked conventional maize were subjected to reducing and denaturing SDS-PAGE and then transferred to a nitrocellulose membrane for detection using an anti-NptII antibody. A co-migrating immunoreactive band was observed in the leaf extract from MON 87460 (Figure 16, Lanes 4-6), leaf NptII-spiked conventional maize leaf extract (Figure 16, Lanes 7-9), and pure E. coli-produced NptII protein (Figure 16, Lane10). As expected, the immunoreactive signal increased with increased loading levels of the leaf extract from MON 87460 and increased amount of the leaf extract from conventional maize spiked with the E. coli-produced NptII protein. No immunoreactive bands were observed in the leaf extract from conventional maize (Figure 16, Lane 3). Based on this analysis, the MON 87460- and E. coli-produced NptII proteins demonstrated equivalent immunoreactive properties, which confirmed both the identity and equivalence of the two proteins. NptII protein molecular weight equivalence The molecular weight of the MON 87460-produced NptII protein and its equivalence to the molecular weight of the E. coli-produced NptII reference standard were demonstrated using densitometric analysis of the western blot (Figure 16). The electromobility of the MON 87460-produced protein was indistinguishable from the electromobility of the E. coli-produced NptII protein. The estimated molecular weight of the MON 87460-produced NptII protein was determined to be 27.4 kDa, which was similar to the previously determined molecular weight of the E. coli-produced NptII reference standard (27.1 kDa). Based on the identical electrophoretic mobility and apparent molecular masses, the MON 87460- and E. coli-produced NptII proteins were determined to have equivalent molecular weights. Conclusions on characterization and equivalence of the MON 87460-produced and E. coli-produced NptII proteins

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 50
The NptII protein from MON 87460 was characterized and compared to the E. coli-produced NptII reference protein standard. The results of this analysis confirmed the identity of the MON 87460-produced NptII protein and established the equivalence of the plant produced protein to the E. coli-produced NptII reference protein standard (Gu et al., 2008). A western blot analysis was utilized to compare the immunoreactivity and apparent molecular weight of the MON 87460-produced NptII protein to that of the previously characterized E. coli-produced NptII reference protein standard. The MON 87460- and E. coli-produced NptII proteins displayed similar immunoreactivity with NptII-specific antibody and had identical electromobility on SDS-PAGE. Taken together, these data establish equivalence between the MON 87460-produced and E. coli-produced NptII reference protein standard.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 51
Figure 16. Western Blot analysis of the MON 87460- and E. coli-produced NptII protein
Maize leaf extracts from MON 87460 and conventional maize. E. coli-produced NptII and E. coli-produced NptII spiked into leaf extract of conventional maize were separated by SDS-PAGE and transferred to a nitrocellulose membrane. The membrane was probed with rabbit anti-NptII antibody and an HPR-conjugated secondary antibody and visualized using an ECL system. Approximate molecular weights (kDa) are shown on the left and correspond to the protein marker loaded in Lanes 2, 11 and 12. The 10 s exposure is shown and is representative of the bands observed in the other exposures.
Lane Sample Amount loaded (ng) (µl)
1 Empty
2 Precision Plus Protein WesternC markers 5
3 Leaf extract from conventional maize 10
4 Leaf extract from maize MON 87460 5
5 Leaf extract from maize MON 87460 10
6 Leaf Extract from maize MON 87460 15 7 E. coli-produced NptII spiked* 0.25 5 8 E. coli-produced NptII spiked* 0.5 10 9 E. coli-produced NptII spiked* 0.75 15 10 E. coli-produced NptII 0.5 10
11 Precision Plus Protein WesternC markers 5
12 Precision Plus Protein WesternC markers 5
* E. coli-produced NptII spiked in leaf extract from conventional maize
kDa
250
150
100
75
50
37.5
25
20
15
10
1 2 3 4 5 6 7 8 9 10 11 12

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 52
Biochemistry and mode of action of the introduced CspB and NptII proteins in MON 87460 Identity and function of the CspB protein The CspB protein in MON 87460 belongs to the cold-shock protein (Csps) family and is identical in amino acid sequence to the native CspB protein produced in B. subtilis with the exception of one amino acid change in the second position from leucine to valine (L2V) that was necessary for cloning purposes. Bacterial Csps are composed of approximately 67-73 amino acid residues (Graumann et al., 1997) and although typically acidic in nature, contain several positively charged amino acid residues that may facilitate binding to the negatively charged backbones of polynucleotides. Accumulation of the CspB protein in B. subtilis cells occurs after transition from exponential growth to stationary phase (Graumann et al., 1997; Graumann and Marahiel, 1999), indicating that CspB accumulation in cells can be triggered under several stress conditions that share a common signal such as inactivation of ribosomes (Graumann et al., 1997; Schindler et al., 1999). Stability of the protein both in vivo and in vitro depends on the protein’s ability to form a complex with nucleic acids, most likely mRNAs (Schindler et al., 1999). In the absence of polynucleic acids, the CspB protein has a very low thermodynamic stability and is susceptible to rapid proteolytic degradation (Schindler et al., 1999). The structure of the CspB protein has been previously described (PDB accession number 1NMF) (Schindelin et al., 1994; Schindelin et al., 1993). The CspB protein in MON 87460 consists of 66 amino acids and has an isoelectric point of 4.31. The protein is composed of five antiparallel β-strands forming a five-strand β-barrel similar to the structure of the CspA protein from E. coli (PDB accession number 1MJC) (Newkirk et al., 1994; Schindelin et al., 1993). All Csps possess binding sites for single stranded nucleic acids called RNA-binding ribonucleoprotein (RNP) motifs (Newkirk et al., 1994; Schröder et al., 1995). The CspB protein, like other CSPs, contains two conserved RNP motifs: RNP1 and RNP2. Within the CspB RNP domains four aromatic amino acids F15, F27, H29, and F30 are required for the double-stranded polynucleotide unwinding or “melting” activity exhibited by CspB. These amino acids are conserved in all CSPs and thought to be essential for protein function in bacteria (Phadtare et al., 2002). In vitro studies suggest that by binding to RNA secondary structures, CSPs reduce the free energy required for wrongly folded RNA to unfold and adopt the correct configuration (Herschlag, 1995). Experimental evidence suggests that CSPs bind at the single-stranded mRNA loop and then progressively cover this region, forcing the stem to open (Phadtare et al., 2002). It was suggested that CSPs bind to single stranded nucleic acids, RNA and ssDNA, but do not appear to bind to dsDNA (Max et al., 2006). The stable association of CSPs with nucleic acids has been confirmed by co-crystallization of the B. subtilis CspB protein in a complex with single stranded polynucleotides (Bienert et al., 2004; Max et al., 2007). The crystal structure data revealed the stoichiometry and sequence determinants of the binding of single-stranded nucleic acids to a preformed site on CspB. These findings together with the described mechanism of RNA destabilization led to the classification of CSPs as RNA chaperones. The nucleic acid destabilizing (“melting”) function of CspB can be demonstrated in an in vitro assay using a molecular beacon system (Phadtare and Severinov, 2005). The hairpin-shaped molecule beacon is labeled with a fluorophore at the 5’- and quencher at the 3’-terminus. Due to the close proximity of the fluorescent tag and quencher in the hairpin conformation, the fluorescence is efficiently quenched. When a CspB protein “melts” the hairpin conformation, the fluorescent tag and quencher are spatially separated which permits fluorescence. This assay has been broadly utilized to characterize the specificity of a variety of CSPs including cold-shock domain (CSD)-containing proteins identified in bacteria and plants (Karlson et al., 2002; Kim et al., 2007; Phadtare et al., 2002).

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 53
Identity and function of the NptII protein The NptII protein functions as a selectable marker in the initial laboratory stages of plant cell selection following transformation (De Block et al., 1984; Horsch et al., 1984). The NptII enzyme uses ATP to phosphorylate neomycin and related aminoglycoside antibiotics, thereby inactivating them. Cells that produce the NptII enzyme selectively survive exposure to these aminoglycosides. The nptII coding sequence is derived from the prokaryotic E. coli transposon Tn5 (Beck et al., 1982). The purpose of inserting the gene encoding the NptII protein into maize cells along with CspB was to have an effective method for selecting cells after transformation. In general, the efficiency of plant cells transformation is often low, ranging from 1 × 10-5 to 1 × 10–4 of cells treated (Fraley et al., 1983). Therefore, the selectable marker, NptII, was used to facilitate the screening process. History of safe use of CspB protein The history of safe use for CspB is established by demonstrating (i) the safety of the CspB donor organism, B. subtilis; (ii) the CspB protein has a common domain with high homology to several proteins found in bacteria and plants used in feed and food production, and (iii) the history of safe consumption of cold shock proteins. Together, these data demonstrate that the CspB protein has a history of safe consumption. Safety of CspB donor organism Bacillus subtilis The safety of and products derived from B. subtilis for use in food has been confirmed consistently by numerous reviews. In 1999, the US Food and Drug Administration (FDA) designated enzyme preparations from this organism as ‘generally recognized as safe’ (GRAS) (FDA, 1999). EFSA's Scientific Committee proposed B. subtilis for a qualified presumption of safety in December 2007 based on the extensive body of knowledge available about this species (EFSA, 20074). The US Environmental Protection Agency (EPA) also exempted B. subtilis from further review under the Toxic Substances Control Act (TSCA) (EPA, 19975). A detailed safety profile of B. subtilis was reviewed as a part of the safety assessment of several
enzymes used in food preparation including -acetolactate, decarboxylase,-amylase, maltogenic amylase, and pullulanase (de Boer and Diderichsen, 1991; Olempska-Beer et al., 2006; Pedersen et al., 2002). In addition, B. subtilis was tested for cytotoxicity in Chinese hamster ovary K1 (CHO-K1) cells, for production of hemolytic and nonhemolytic enterotoxins (Pedersen et al., 2002), acute toxicity in BALB/c mice, and chronic toxicity in mice, rabbits, and pigs (Sorokulova et al., 2008). No toxic effects were attributed to B. subtilis in these studies and no similarities to pathogenic bacteria were found, which led to the conclusion that B. subtilis is non-pathogenic and safe for human consumption (Pedersen et al., 2002; Sorokulova et al., 2008). As further confirmation of the safety of B. subtilis, spores and cultures of this organism and other Bacillus species have been sold worldwide as probiotics, including in the US, Mexico, Europe and South Asia (Sanders et al., 2003). Probiotics are dietary supplements containing potentially beneficial bacteria or yeasts. FAO/WHO (2001) defined probiotics as “live microorganisms which when administered in adequate amounts confer a health benefit on the host”. Examples of commercial probiotic products intended for human or animal use and containing Bacillus spores
include Bidisubtilis (Bidiphar, Vietnam), BioGrow (Provita Eurotech Ltd., UK), BioPlus 2B (Christian Hansen Hoersholm, Denmark), Biosporin (Biofarm, Ukraine), Biostart™ (Microbial Solutions, South Africa and Advanced Microbial Systems, USA), Lactipan Plus (Istituto Biochimico Italiano SpA, Italy), Liqualife™ (Cargill, USA), Medilac (Hanmi Pharmaceutical Co., 4 http://www.efsa.europa.eu/cs/BlobServer/Scientific_Opinion/sc_appendixb_qps_en.pdf?ssbinary=true
5 http://www.epa.gov/biotech_rule/pubs/fra/fra009.htm Registered trademark

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 54
China), Nature’s First Food™ (Nature’s First Law, USA) (reviewed in Hong et al. (2005)). Human probiotic products are used for oral bacteriotherapy of gastrointestinal disorders since the ingestion of large amounts of B. subtilis is thought to restore the normal microbial flora following extensive antibiotic use or illness (Mazza et al., 1992). B. subtilis has been extensively tested by numerous agencies and found to be non-pathogenic and safe for human consumption. In addition, B. subtilis has been used for many years in probiotics without any adverse health effect. Thus, the safety of the donor organism for the CspB protein in MON 87460 is well established and documented. The CspB protein contains a common domain found in bacteria and plants used in feed and food production The CspB protein from B. subtilis is homologous to several bacterial proteins that are ubiquitous in the human diet and directly consumed in many common foods. CspB is a member of the cold shock protein (Csp) family (Karlson and Imai, 2003; Nakaminami et al., 2006). This family includes bacterial and plant proteins possessing a cold shock domain (CSD) that has high amino acid sequence similarity to the CspB protein (Graumann et al., 1997; Karlson and Imai, 2003; Nakaminami et al., 2006). The cold shock domain database currently contains 547 entries representing cold shock proteins found in bacterial and plant species (http://www.chemie.uni-marburg.de/~csdbase/). A search of publicly available databases using the CspB protein sequence has revealed that the CspB protein present in MON 87460 shares amino acid identity to other naturally occurring CSD-containing proteins found in food and food products (Table 12). The amino acid identity ranges from 35% to 98.5% across different plant and bacterial species. CspB from B. subtilis is homologous to the CSP proteins found in E. coli, Lactobacillus, Lactococcous, and Bifidobacterium species (Table 12), the most common types of bacteria used in the dairy industry to prepare cheese, sour cream, buttermilk, yogurt and probiotic products containing live bacterial cultures (http://www.nationaldairycouncil.org; Morea et al. (1999); Ogier et al. (2002)). These bacteria are commonly present in gastrointestinal flora. In addition, Bacillus, Lactobacillus and Lactoccoccus species containing CSPs are involved in many fermentation processes of milk, meats, cereals and vegetables. Food fermentation is a widely practiced and ancient technology to preserve food or to make it more digestible (Caplice and Fitzgerald, 1999). Examples of fermented foods that are popular in different countries and prepared with the help of bacteria include European sausages prepared by fermentation of the raw meat with Lactobacilli cultures (Lactobacillus sakei and/or Lactobacillus curvatus) (Leroy et al., 2006), pickled vegetables, kefir, yogurt (Lopitz-Otsoa et al., 2006), products of soy fermentation including soy sauce (Tanasupawat et al., 2002) and okpehe (Oguntoyinbo et al., 2007), a traditional African fermented soup condiment produced by fermentation of cooked Prosopis africana seeds. Other examples include Dawadawa (made from soy or Locust beans), a popular condiment used in Africa and Asia to add flavour to soups and stews (Terlabie et al., 2006), thua nao (made from soy), popular in Asian food as a condiment for enhancing flavour in soups and curries (Inatsu et al., 2006) and natto, a commonly consumed food in Japan, made by fermenting cooked soybeans with B. subtilis ssp. natto (Ashikaga et al., 2000). Cold shock domains (CSDs) with high similarity to the CspB protein expressed in MON 87460 are also present in many crops and, therefore, are ubiquitous in the human diet and directly consumed in many common foods. Proteins that are homologous to B. subtilis CspB were identified in rice (Oryza sativa L.) and wheat (Triticum aestivum L.) (Table 12). The wheat cold shock protein, WCSP1 (Karlson et al., 2002), has been shown to be functionally similar to CspB, as demonstrated by complementation of the E. coli cold-sensitive phenotype with WCSP1 and in vitro functional melting assays (Karlson et al., 2002; Nakaminami et al., 2006). It has been suggested that WCSP1 protein functions in planta as a RNA chaperone by unfolding the secondary structure of the nucleic acids and is involved in the regulation of translation at low temperatures (Nakaminami et al., 2006). Recently two CSD-containing proteins were described in rice (Chaikam and Karlson, 2008). Rice CSPs contain two distinct domains: an N-terminal

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 55
CSD region and a glycine-rich domain. These CSD-containing proteins possess similar in vitro and in vivo functions as those of WCSP1 and bacterial CSPs (Chaikam and Karlson, 2008). Furthermore, the cold shock domains of CSD-containing proteins from wheat and rice CSPs share 40-50% identity to the entire sequence of CspB from B. subtilis. Recently, Karlson and Imai (2003) , showed that, by use of bioinformatic analyses of a GenBank expressed sequence tag (EST) database, proteins containing highly conserved CSDs are likely present in food crops such as maize, barley and soy. Digestion of plant CSD-containing proteins would be expected to release these highly conserved CSD portions or fragments thereof into the human digestive tract. It is recognized that human consumption of plant proteins with significant homology to CspB occurs on a daily basis. The CspB protein in MON 87460 is similar to several bacterial CSPs and CSD-containing proteins present in food crops that are ubiquitous in the human diet and directly consumed in common foods establishing a history of safe exposure for this protein.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 56
Table 12. Amino acid sequence identity between MON 87460-produced CspB protein and other cold shock domain containing proteins present in foods
Sequences of CSPs were extracted from publicly available database of cold shock proteins, CDBAse (http://www.chemie.uni-marburg.de/~csdbase/; Weber et al. (2002)). Sequences were aligned and sequence identity was calculated using MegAlign module of Lasergene software suit for sequence analysis (version 7.1.0(44)) (DNASTAR Inc., Madison, WI, USA).
# Proteins Sequence Identity (%)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
1 MON 87460 CspB 100 97 44 61 64 58 66 66 64 40 69 79 49 61 59 37 43 47
2 B. subtilis CspB 100 45 61 64 58 66 66 64 40 69 79 49 61 59 38 42 46
3 Bifidobacterium longum CSPA 100 52 49 46 38 38 54 39 54 50 36 44 52 45 49 45
4 E. coli CSPA 100 68 77 59 59 39 65 68 57 44 58 57 44 54 53
5 E. coli CSPE 100 66 47 47 37 60 62 64 47 58 61 41 45 53
6 E. coli CspB 100 47 47 34 60 62 55 46 58 58 43 48 50
7 Lactobacillus acidophilus CPSL 100 100 47 69 75 63 44 59 41 32 31 41
8 Lactobacillus casei CSPL 100 47 69 75 63 44 59 41 32 31 41
9 Lactobacillus delbrueckii CSPA 100 42 38 63 55 69 57 48 39 51
10 Lactobacillus delbrueckii CspB 100 69 39 26 34 33 29 22 56
11 Lactobacillus plantarum CSPC 100 71 53 73 57 41 42 55
12 Lactobacillus sakei CSPA 100 51 65 42 32 33 36
13 Lactococcus lactis CSPA 100 62 52 42 44 42
14 Lactococcus lactis CspB 100 60 33 32 53
15 Corynebacterium glutamicum CSP 100 38 54 60
16 Agrobacterium tumefaciens YSP 100 37 28
17 Oryza sativa P0582D05 100 79
18 Triticum aestivum WCSP1 100

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 57
History of safe consumption of cold shock proteins Proteins that are highly similar to MON 87460 CspB are consumed in a wide variety of foods and are utilized in numerous food processes that have been in use for long periods of time. Lactobacillus sp. in dairy foods serves as an example to estimate the consumption of CSP proteins. Lactobacillus sp. is the most common type of bacteria used in the dairy industry for preparation of products containing live bacterial cultures, including buttermilk, sour cream, cheddar cheese, mozzarella cheese and yogurt (http://www.nationaldairycouncil.org). Lactobacillus species contain Csps that share 40-79% sequence identity with the CspB in MON 87460 (Table 12). These bacteria resist gastric acid, bile salts and pancreatic enzymes, and, thus, readily colonize the intestinal tract (Rolfe, 2000). In the US during 2006, per capita daily availabilities of buttermilk, sour cream, cheddar cheese and mozzarella cheese were 2.1 g, 5.2 g, 12.8 g and 12.9 g, respectively (http://www.ers.usda.gov). In addition, 2006 US per capita yogurt availability was 15.3 g per day. In other countries such as the United Kingdom (UK), per capita daily yogurt consumption is similar (19 to 24 g; Henderson et al. (2002)). Yogurt is unique among dairy products due to the fact that standards exist for the number of colony-forming units (CFUs) that must be present at the time of manufacture. Codex standards specify that each gram of yogurt should contain a minimum of 107 Lactobacilli CFUs, a number reflected in a 2003 FDA advance notice of public rulemaking (Codex Standard, 2003; FDA, 2003). Applying this level of Lactobacilli CFUs to an estimated per capita daily yogurt consumption of 19 g per day equates to approximately 190 million CFU per person per day. Expression of CSPs in Lactobacillus varies depending on temperature (Derzelle et al., 2000), making it difficult to estimate an average consumption rate based on bacterial exposure. It is known, however, that levels of bacterial CSPs can increase several folds in response to cold shock (Etchegaray and Inouye, 1999; Phadtare, 2004). CSPs can be present in E. coli, an organism with a cold shock response that is similar to that observed in Lactobacillus, at levels of 106 copies per cell (Derzelle et al., 2000; Phadtare, 2004; Sauvageot et al., 2006; Thieringer et al., 1998). Assuming 106 copies of a 7 kDa Lactobacillus CSP is present in each cell, per
capita daily CSP consumption through yogurt alone would be approximately 3 g. Considering the prevalence of other dairy sources of bacterial CSPs such as buttermilk, sour cream and cheese and assuming they contain similar CSP levels as yogurt, it is likely that average consumption rates of just a few varieties of dairy foods a day would provide amounts of bacterial CSPs that far exceed the amount of CspB a person would consume from MON 87460 even if it were the sole source of maize in the diet. Conclusions on the history of safe use of CspB protein MON 87460 represents a minimal contribution to human CSP consumption and poses no safety risk. Proteins homologous to CspB are present in many widely consumed foods and human consumption of CspB itself is also well documented. Bacterial and plant sources of CSPs and other CSD-containing proteins are common in human diets throughout the world. An estimate of human CSP consumption from yogurt is ten times greater than estimates of CspB consumption from MON 87460. Plant sources such as wheat and rice represent still more dietary sources of CSD-containing proteins. The donor organism of the CspB protein, B. subtilis, is not pathogenic, is often used in probiotic formulations with purported health benefits, is present in many fermented foods, has a history of safe consumption and is the source of enzyme preparations with

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 58
GRAS status. CSD-containing proteins are ubiquitous in nature, being present in many plants and common bacteria including species that are normally present in gastrointestinal flora. Cold shock proteins have no known toxicity and are not associated with pathogenicity. The B. subtilis CspB shares a high percent of identity with CSPs present in other bacterial species broadly used by the food industry and with CSD-containing proteins in plant species used as a food. Foods prepared with the help of B. subtilis have been consumed for centuries with no documented history of any adverse effects on human health. Taken together, this information demonstrates that CspB from B. subtilis has a history of safe exposure and a history of safe consumption. History of safe use of the NptII protein The history of safe use for NptII is established by demonstrating (i) the safety of the NptII donor organism, E. coli; (ii) the history of safe use of the NptII protein, and (iii) the presence of NptII in commercial food and feed crops. Together, these data demonstrate that the CspB protein has a history of safe consumption. Safety of NptII donor organism Escherichia coli The nptII gene was isolated from E. coli K-12 transposon Tn5. E. coli bacterium is ubiquitous in the environment and is found in the digestive tracts of vertebrate species, including humans (Jefferson et al., 1986). The safety of E. coli has been previously assessed by the FDA as part of the safety evaluation of the chymosin enzyme preparation derived from E. coli K-12 (Flamm, 1991). As a result of the FDA safety evaluation, the donor organism was determined to be safe and the FDA affirmed the chymosin preparation as GRAS. E. coli is one of the most studied laboratory organisms, with a long history of safe use in the laboratory environment. E. coli represents one of the best understood living organisms whose full genome has been sequenced (Blattner et al., 1997). It is classified in all major national and international safety guidelines as a biologically safe organism for the propagation of a broad range of gene cloning and expression vectors and has been used as such for protein production in many commercial applications (Bogosian and Kane, 1991). E. coli K-12 is apathogenic (Gray and Tatum, 1944). To date, no virulence genes have been found in E. coli K-12 derivatives, in contrast to the pathogenic E. coli strains (Mühldorfer and Hacker, 1994). Safety of NptII protein The enzyme NptII is the most commonly used antibiotic resistance marker, inactivating neomycin and related antibiotics, such as kanamycin. The NptII protein is ubiquitous in E. coli, and, therefore, is normally present in the human gastrointestinal tract (Fuchs et al., 1993a; Jefferson et al., 1986). The NptII protein has been used as a selectable marker in a variety of crops including tomato, cotton, oilseed rape and maize and its safety has been thoroughly evaluated by the US FDA (FDA, 1994, 1998), that has approved its use as a food additive (FDA, 1994). Also the EPA has established an exemption from the requirement of a tolerance for NptII for use as a selectable marker in raw agricultural commodities (40 CFR Part 180.1134). Numerous studies have suggested that the presence of this antibiotic-resistance gene in any crop or crop product will have no impact on food safety (reviewed in Miki and McHugh (2004)). Studies using purified NptII protein revealed that NptII degrades rapidly in simulated gastric and intestinal fluids, suggesting that the protein is unlikely to cause an allergic response (Fuchs et al., 1993b). An assessment of the ecological impact of the use of the

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 59
NptII protein in crops has been discussed by Nap et al. (1992). It was suggested that the amount of free kanamycin accumulating in soils by the action of micro-organisms or via animal feces, is restricted by absorption to soil components so that no direct selection pressure for kanamycin resistant plants can occur (Nap et al., 1992). Further, enhancement of physiological fitness resulting from pleiotropic effects of nptII gene expression has not been documented (Nap et al., 1992). Based on all the available evidence, it can be concluded that the NptII protein is safe for use as a selectable marker in genetically modified plants (FDA, 1994; Flavell et al., 1992; Fuchs et al., 1993a; Fuchs et al., 1993b; Huppatz and Fitzgerald, 2000; Miki and McHugh, 2004; Nap et al., 1992; WHO, 1993). NptII was used as an antibiotic resistance marker in the initial selection process of MON 87460 and given the performance of the product in early trials and a lack of safety concerns around the NptII protein, it was deemed unnecessary to excise the nptII gene from the plant using the loxP sites that flank this gene. Presence of the NptII protein in commercial food and feed crops NptII-containing crops, such as MON 863 maize and MON 863-containing stacks6 have been commercialized on a broad acreage and safely consumed as foods and feeds since their initial introduction in 2003. The NptII protein produced in these maize crops is identical to NptII produced in MON 87460. The safety assessment on MON 863 by EFSA concluded that MON 863 maize and MON 863-containing stacks will not have any adverse effect on human and animal health or the environment7. Conclusions on history of safe use of NptII protein The safety of the NptII protein and its donor organism, E. coli, has been recognized by regulatory agencies and well documented. All existing data suggest that the NptII protein represents a negligible hazard to human health and is safe for consumption. Structural similarity of CspB and NptII proteins to known toxins or other biologically active proteins The assessment of the potential for protein toxicity includes a bioinformatic analysis of the amino acid sequence of the introduced protein. The goal of the bioinformatic analysis is to ensure that the introduced protein does not share homology to known toxins or anti-nutritional proteins associated with adverse health effects.
Potential structural similarities shared between the CspB and NptII proteins and sequences in a protein database were evaluated using the FASTA sequence alignment tool. The FASTA program directly compares amino acid sequences (i.e., the primary, linear protein structure) and the alignment data may be used to infer shared higher order structural similarities between two sequences (i.e., secondary and tertiary protein structures). Proteins that share a high degree of similarity throughout the entire sequence are often homologous. Homologous proteins usually have common secondary
6 MON 863 × MON 810, MON 863 × NK603 and MON 863 × MON 810 × NK603
7 http://www.efsa.europa.eu/EFSA/Scientific_Opinion/gmo_opinion_ej256_mon863xmon810xnk603_en1,0;
http://www.efsa.europa.eu/EFSA/Scientific_Opinion/gmo_opinion_ej252_mon863x810_2_en1.pdf?; http://www.efsa.europa.eu/EFSA/Scientific_Opinion/gmo_opinion_ej255_mon863xnk603_en1,0.pdf; http://www.efsa.europa.eu/EFSA/Scientific_Opinion/opinion_gmo_07_en1.pdf?ssbinary=true

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 60
structures, common three-dimensional configuration and, consequently, may share similar functions. FASTA bioinformatic alignment searches using the CspB and NptII protein sequences were performed with the TOX_2009 and PRT_2009 databases to identify possible homology with proteins that may be harmful to human and animal health (Tu, 2009; Tu and Silvanovich, 2009a, 2009b) (Attachments G & H) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. The TOX_2009 database is a subset of 7,651 sequences derived from the protein database (PRT_20098) consisting of publicly available protein sequences from GenBank (GenBank protein database, release 169.0, January 5, 2009). The sequences in TOX_2009 were selected using a keyword search and filtered to remove non-toxin proteins. Initially, all sequence descriptions contained in header lines and the associated protein sequence derived from the PRT_2009 database were keyword screened using all possible combinations of upper and lower case characters spelling the words “toxic” and “toxin”. The resulting sequences and their respective descriptions were then filtered to exclude several terms used in combination with “toxic” or ”toxin”; the exclusion terms “synthetic”, “anti”, “putative”, “like”, “insect”, “Cry”, “thuringiensis” and “toxin-reductase” were used to remove non-toxin protein sequences.
An E-score acceptance criterion of 1 × 10-5 for any alignments was used to identify proteins from the TOX_2009 and PRT_2009 databases with potential for significant shared structural similarity and function with CspB and NptII proteins. The results of the search with TOX_2009 produced no alignments below an E-score of 1 × 10-5 and therefore, demonstrated no significant similarity with known toxins. When CspB was compared to the PRT_2009 database, 4738 alignments were yielded with an E-score of less than or equal to 10-5. Visual inspection of these alignments revealed that most alignments were with naturally occurring or theoretical proteins described as cold shock proteins; these are homologous sequences to the CspB protein. The top alignment demonstrated 100% identity over the full length of both the query and aligned proteins with an E-score of 1.2 × 10-26 to an unnamed protein product (GI-63092813). The GI-63092813 sequence is associated with a patent application related to the enhancement of stress tolerance in transgenic plants and is not a toxin. The 2nd and 3rd best aligned proteins (GI-63092738 and -297761) displayed 100% identity with the query and one of these, GI-297761, was identified as being the CspB protein from B. subtilis; the same sequence expressed in MON 87460. The results of the bioinformatic analyses demonstrated that no structurally relevant similarity exists between the CspB and NptII proteins and any known toxic or other biologically active proteins that would be harmful to human or animal health (Tu, 2009; Tu and Silvanovich, 2009a, 2009b) (Attachments G & H) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. 28-day oral toxicity study for the CspB protein
8 A detailed description on how the TOX_2009 and PRT_2009 databases were constructed can be found in
Silvanovich Silvanovich A, 2009. The assembly of AD_2009, TOX_2009 and PRT_2009. Monsanto Technical Report, MSL0021840, 1-309..

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 61
A 28-day oral toxicity study with CspB is not necessary to confirm its safety. Bioinformatics analyses (Burzio et al., 2008a; Tu and Silvanovich, 2009a, 2009b) (Attachments F, G & H) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act], demonstrate that CspB expressed in MON 87460 is not structurally or functionally related to proteins that adversely effect human or animal health. As discussed in Section 8.4.1 and Section 8.5.2, both the CspB protein and its source organism have a history of safe consumption. CspB was derived from B. subtilis, a soil microorganism with a history of safe use in fermented foods and probiotics (de Boer and Diderichsen, 1991; Hosoi et al., 2003; Sanders et al., 2003). B. subtilis has more than 300 years of safe consumption through its use in production of natto, a popular fermented soybean food eaten primarily in Japan (Schaffer et al., 2007). Furthermore, CspB is homologous to other cold shock proteins and proteins containing a cold shock domain found in bacteria and plants including those used for food (Graumann and Marahiel, 1998; Karlson and Imai, 2003; Nakaminami et al., 2006). CspB expression in grain was very low (Mean expression was 0.063 μg/g (ppm) fw) in whole MON 87460 grain grown in the US (Mozaffar and Silvanovich, 2008), indicating that potential exposure to this protein from MON 87460 derived foods will be low. Maize is, with the exception of sweet maize, extensively processed prior to human consumption including processes such as nixtimalization (alkaline cooking for masa, tortillas, etc.) and wet milling (for oil, starch, syrups, etc.) that employ varying degrees of mechanical, enzymatic, solvent, heat, extreme pH conditions and pressure treatments (May, 1987; Milan-Carrillo et al., 2004; White and Pollak, 1995). CspB was also shown to be completely digested following sequential in vitro digestion in simulated gastric fluid and simulated intestinal fluid assays (Kapadia et al., 2008), indicating that if any CspB protein survives processing and cooking, it would be digested and degraded in the gastrointestinal tract, thus minimizing the potential for systemic absorption. Lastly, the CspB protein and grain from MON 87460 did not elicit toxicity in an acute mouse toxicity study (CRO-2007-182, 2008) (Attachment I) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act] or a subchronic rat feeding study (WI-2007-064, 2008) at doses several orders of magnitude higher than anticipated dietary exposure in the South African population. A rodent 28-day repeat dose oral toxicity study with CspB was therefore not considered necessary to demonstrate the safety of this protein, based on: 1) The lack of structural or functional relationship of CspB to proteins that adversely effect human or animal health; 2) The history of safe use for CspB and its source organism, B. subtilis; 3) The low expression level of CspB in grain; 4) The digestibility of the CspB protein; 5) The lack of toxicity of CspB or grain from MON 87460 at doses several orders of magnitude higher than anticipated human exposure. Conclusions regarding CspB and NptII The CspB and NptII proteins have been assessed for their potential toxicity according to the recommendations of Codex. The genes encoding the newly expressed proteins were isolated from donor organisms considered to be safe and both proteins have a long history of safe consumption in foods and feeds. The proteins lack structural similarity to known toxins or biologically active proteins known to have adverse effects to mammals. In addition, no indications of toxicity were reported in mice administered the CspB and NptII proteins by oral gavage. All these data indicate that the CspB and NptII proteins are safe for human and animal consumption.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 62
MON 89034 The safety of the Cry1A.105 and Cry2Ab2 proteins has been previously assessed during Monsanto’s application for general release of MON 89034 and MON 89034 × NK603. Additionally the proteins were also assessed during the application for Commodity Import of MON 89034 × 1507 × MON 88017 × 59122 (SmartStax). For ease of reference the relevant section is provided below: Safety of the donor organism: Bacillus thuringiensis Bacillus thuringiensis (Bt) is a spore-forming, gram-positive bacterium that is found naturally in soil. Bt strains have been used commercially in the US since 1958 to produce microbial-derived products with insecticidal activity (US EPA, 1988). Many strains of Bt have been shown to produce protein crystals or inclusion bodies that are selectively toxic to certain orders and species of insect pests. The current nomenclature, based on amino acid identity, allows closely related proteins to be ranked together. Each protein is assigned a unique name incorporating four hierarchical ranks consisting (in order) of an Arabic numeral, upper case letter, lower case letter, and an Arabic numeral (e.g., Cry2Ab2 protein). Thus, proteins with identity of:
<45% differ in a primary rank (e.g., Cry2, Cry3, etc.), >45% but <78% differ in secondary rank (e.g., Cry3A, Cry3B), >78% but <95% differ in tertiary rank (e.g., Cry3Ba, Cry3Bb), and >95% differ in quaternary rank (e.g., Cry2Ab1, Cry2Ab2) and are considered allelic variants.
In general, the primary rank of the Bt proteins denotes its specific insecticidal activity; for example, Cry1, Cry2, Cry3, and Cry4 proteins are toxic to lepidopteran, lepidopteran/dipteran, coleopteran, and dipteran pests, respectively (Bravo, 1997; Höfte and Whiteley, 1989). It has been established that Cry proteins have a narrow range of insecticidal activity against one or, less commonly, two orders of insects (Crickmore et al., 1998). In the US, an exemption from the requirement of a tolerance for the first microbial Bt product was granted in 1960 by the FDA after an extensive toxicity and infectivity evaluation program. The testing program consisted of acute, subchronic, and chronic studies, which resembled the testing required for conventional chemical pesticides. Registration was granted by the USDA later the same year. In 1971, EPA assumed responsibility for all pesticide tolerance exemptions. Since then, a variety of naturally occurring and genetically modified microbial Bt products have been registered and included under this tolerance exemption. EPA has established separate tolerance exemptions by amendment for various Cry proteins (e.g., Cry1Ab, Cry1Ac, Cry2Ab2 and Cry3Aa, Cry3Bb1) expressed in biotechnology-derived crops (US EPA, 1995, 1996, 1997, 2001, 2002). The conclusion of reasonable certainty of no harm and the resultant tolerance exemptions for this wide array of Bt mixtures and Cry proteins in food or feed were based on the lack of adverse effects to mammals in numerous toxicological studies. This conclusion is supported by a history of safe use in agriculture for over 40 years (McClintock et al., 1995). There are no adverse effects known to have occurred in humans during this prolonged period of use (US EPA, 1988). An extensive review of the

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 63
safety of the various Cry proteins that have been expressed in biotechnology-derived plants is available (Betz et al., 2000). Similarity to known toxins
Cry1A.105 protein Similarity of the Cry1A.105 protein with Cry1A class of Bt proteins The Cry1A.105 protein is a chimeric protein that consists of domains I and II from Cry1Ac or Cry1Ab, domain III from Cry1F, and of the C-terminal domain from Cry1Ac. Cry1Ab and Cry1Ac proteins share 100% amino acid sequence identity in domains I and II. Overall amino acid sequence identity of the Cry1A.105 protein to the Cry1Ac, Cry1Ab, and Cry1F proteins is 93.6%, 90.0%, and 76.7%, respectively. Domain exchange is a well known naturally occurring mechanism for generating diversity and has been described in detail in the literature (de Maagd et al., 2001b; Masson et al., 2002). To better understand the structural relatedness of the Cry1A.105 protein to other Cry proteins, a phylogenetic tree (Figure 17) was produced using the Cry1A.105 amino acid sequence and the methods employed to define the Bt protein nomenclature (Crickmore et al., 1998). The analysis demonstrates that Cry1A.105 belongs to the Cry1A class of Bt proteins and is most closely related (93.6% identical) to the Cry1Ac protein. This study is attached as Attachment 9 of the application for General Release of MON 89034. Recombinant DNA techniques have been used to generate Bt strains with enhanced insecticidal activity for use as biopesticides (Baum, 1998; Baum et al., 1999). For example, the microbial pesticide Lepinox™9 WDG (Ecogen Inc.) that contains a Cry1Ac/Cry1F chimeric protein has been approved and used for control of lepidopteran pests since 1997 (Baum, 1998; Baum et al., 1999; US EPA, 1997). Cry1A proteins are widely used as topical bioinsecticides and in biotechnology-derived crops. The FDA completed consultations for the safety assessment of maize that produces the Cry1Ab (YieldGard, Bt11) and Cry1F (Herculex™1 I) proteins, as well as cotton producing the Cry1Ac (Bollgard™1 and Bollgard II, WideStrike™1) and Cry1F (WideStrike™2) proteins. The EPA, which has responsibility of assessment the safety of pesticidal proteins, issued exemptions from the requirement of a tolerance for the Cry1Ab protein in maize YieldGgard and Bt11, the Cry1Ac protein in cotton Bollgard and WideStrike, and the Cry1F protein for Herculex I maize and WideStrike cotton. The detailed human and animal safety assessments and a history of safe use conducted on Cry1Ab, Cry1Ac, and Cry1F proteins confirm their safety. Due to the chimeric nature of Cry1A.105, derived from domains of Cry1Ab, Cry1Ac, and Cry1F, a similar safety profile is expected for Cry1A.105.
9 Lepinox
TM is a registered trademark from Ecogen Inc. Bollgard
TM is a registered trademark of
Monsanto Company. HerculexTM
and WidestrikeTM
are registered trademarks of Dow AgroSciences LLC.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 64
Figure 17. Phylogram establishing that the Cry1A.105 and Cry1Ac proteins are located in the same cluster based on amino acid sequence identity (Crickmore et al., 1998).
Having established the high structural similarity between the Cry1A.105 protein and the Cry1A class of proteins, it is important also to establish the functional similarity. To assess the function of this protein, the biological activity of Cry1A.105 protein was compared to the related proteins Cry1Ab, Cry1Ac, and Cry1F. Amino acid sequence identity for insecticidal proteins predicts similarity in biological function, i.e., activity towards a similar spectrum of insects (Attachment 9 of the application for General Release of MON 89034). It has been established that Cry proteins have a defined spectrum of insecticidal activity within a particular insect order (Crickmore et al., 1998; de Maagd et al., 2001b). This high degree of specificity is governed by four levels of selectivity, which collectively lead to intoxication (Federici, 2002). The levels of selectivity include: 1) the route by which the insect is exposed to Cry proteins (i.e., ingestion of plant tissues); 2) protein toxin activation by specific proteolytic enzymes (determined by differences in gut physiology between insects); 3) toxin binding to available specific midgut receptors; and 4) changes in the protein configuration. The reconfigured protein then is able to enter the midgut membrane and form channels. This activity affects larval ability to feed and develop, and eventually leads to death of the susceptible insect. As a consequence, only insects with specific receptors are affected and no toxicity is observed in species that lack these receptors. For example, the Cry1Ab, Cry1Ac and Cry1F proteins are active against lepidopteran but not coleopteran insects, and the Cry3Bb1 protein is active against coleopteran insects such as corn rootworms but not lepidopteran insects.
}}

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 65
Studies were conducted to evaluate the spectrums of the insecticidal activities of the Cry1A.105 protein with a variety of agronomically important insects from three major taxa. Insect species tested included four representative lepidopterans: black cutworm, corn earworm, fall armyworm, and European corn borer; two representative coleopterans: boll weevil (Anthonomus grandis grandis) and southern corn rootworm (Diabrotica undecimpunctata howardi); and two representative hemipterans: western tarnished plant bug (Lygus hesperus) and green peach aphid (Myzus persicae). The insects were exposed to high Cry1A.105 concentrations of (up to 80 or 100 µg of protein per ml of insect diet in diet-incorporation bioassays). Mortality and a reduction in weight or honeydew production over the exposure period were the endpoints used to indicate insecticidal activity. The results showed that Cry1A.105 had activities on all four representative lepidopteran insects. However, there was no indication of Cry1A.105 activity against the two coleopteran or the two hemipteran representatives. This activity spectrum of Cry1A.105 is similar to the activities of the Cry1Ac, Cry1Ab, and Cry1F proteins described in the literature (De Maagd et al., 2003; de Maagd et al., 2001b). Similarity in biochemical and functional characteristics between proteins with established safety is a good indication of the general safety of that particular class. Absence of similarity of Cry1A.105 to known toxins or other biologically active proteins The safety assessment of proteins expressed in biotechnology-derived crops evaluates potential health effects through a comprehensive approach, which includes bioinformatic analysis of the amino acid sequence of the introduced protein to ensure the protein is not similar to toxic proteins that are known to cause adverse health effects. The FASTA sequence alignment tool was used to assess structural similarity. Although the FASTA program directly compares amino acid sequences (i.e., the primary protein structure), the alignment data may be used to infer secondary and tertiary protein structural similarities. Proteins that share a high degree of similarity throughout their entire lengths are often homologous. Homologous proteins share secondary structure and common three-dimensional configuration. Structural similarities between the Cry1A.105 protein sequence and the toxin (TOXIN5) and public domain (ALLPEPTIDES) database sequences were examined. The extent of each similarity was evaluated by visual inspection of the alignment, the calculated percent identity, and the E score value for that alignment. The results showed that no biologically relevant structural similarities to human and animal toxins were observed for the Cry1A.105 protein sequence. Cry2Ab2 protein Similarity of the Cry2Ab2 protein with Cry2Ab2 class of Bt proteins The amino acid sequence of the Cry2Ab2 protein expressed in MON 89034 is 88% identical to the Cry2Aa protein produced by the B. thuringiensis subsp. kurstaki. The B. thuringiensis subsp. kurstaki strain controls insect pests by the production of crystalline insecticidal proteins known as delta-endotoxins. These proteins are produced as the bacterium enters the sporulation phase and can account for approximately one-third of

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 66
the weight of the bacterial cell. These spores are commonly used in agriculture to control insect pests. In addition to the history of safe use in bacterial sprays, the Cry2Ab2 protein produced in MON 89034 is identical in amino acid sequence to the Cry2Ab2 produced in insect protected cotton Bollgard II. Bollgard II has been authorized by several regulatory agencies around the world which have confirmed the safety of the Cry2Ab2 protein. Bollgard II has also been approved for commercial use in South Africa in 2003. Absence of similarity of Cry2Ab2 to known toxins or other biologically active proteins The comparison of the Cry2Ab2 sequence was performed with the toxin (TOXIN5), and public domain (ALLPEPTIDES) database sequences using bioinformatic tools. Sequence identity and amino acid similarity to toxins and other proteins was assessed using the sequence alignment tool FASTA. Identified proteins were ranked according to their degree of similarity. The highest similarity observed was to pesticidial crystal protein Cry2Ab, demonstrating 100.0% identity over 632 aa with and an E score of zero. All remaining alignments with significant E scores are to Cry protein homologues derived from either B. thuringiensis, Paenibacillus popilliae or Paenibacillus lentimorbus. Based on these data, the Cry2Ab2 protein does not share structural congruence with any proteins that may cause adverse effects in humans and animals. Acute oral toxicity study Most known protein toxins, including the insecticidal Cry proteins, act through acute mechanisms to exert toxicity (Hammond and Fuchs, 1998; Pariza and Johnson, 2001; Sjoblad et al., 1992). The primary exceptions to this rule consist of certain anti-nutritional proteins such as lectins and protease inhibitors, which typically require a short-term (2-4 week) feeding study to manifest toxicity (Liener, 1994). The amino acid sequences of the Cry1A.105 and Cry2Ab2 proteins produced in MON 89034 are not similar to any of these anti-nutritional proteins or to any other known protein toxin. In addition, since Cry proteins act through acute mechanisms to control insect pests, and have no activity against non-target organisms such as mammals, the EPA has determined that a high dose acute test is sufficient to confirm their absence of toxicity towards mammals (McClintock et al., 1995). Therefore, an acute oral mouse toxicity study was considered appropriate and adequate to confirm the lack of mammalian toxicity of the Cry1A.105 and Cry2Ab2 proteins. Cry1A. 105 protein The acute oral toxicity with the Cry1A.105 protein was evaluated in CD-1 mice. Cry1A.105 protein was administered by gavage (as two doses about 4 hours apart) to 10 male and 10 female CD-1 mice at a total dose level of 2072 mg/kg body wt (bw). The Cry1A.105 protein used in this study was obtained from a recombinant E. coli strain. This protein was demonstrated to be biochemically and functionally equivalent to the Cry1A.105 protein produced in MON 89034. There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOEL for Cry1A.105 was

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 67
considered to be 2072 mg/kg bw, the highest dose tested. This study was provided as Attachment 10 of the application for General Release of MON 89034. Cry2Ab2 protein In a similar study, Cry2Ab2 protein was administered by gavage (as two doses about 4 hours apart) to 10 male and 10 female CD-1 mice at a total dose level of 2198 mg/kg bw. The Cry2Ab2 protein used in this study was obtained from another recombinant E. coli strain. This protein was demonstrated to be biochemically and functionally equivalent to the Cry2Ab2 protein produced in MON 89034. There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOEL for Cry2Ab2 was considered to be 2198 mg/kg bw, the highest dose tested. This study was provided as Attachment 11 of the application for General Release of MON 89034. In addition, EPA assessed the acute toxicology and other human health safety data for the Cry1A. 105 and Cry2Ab2 proteins and established an exemption from the requirement of a tolerance for both proteins (US EPA, 2008a, 2008b). In vitro digestibility Cry1A.105 (>95%) is rapidly digestible in simulated gastric fluid after 30 seconds incubation in simulated gastric fluid (SGF) and greater than 99% after 5 minutes in simluated intestinal fluid (SIF). More information on this study is provided below under the allergenicity paragraph of this section. Heat stability A study was conducted to assess the immunodetectability of the Cry1A.105 and Cry2Ab2 proteins in maize grain from MON 89034 following heat treatment. Test grain (MON 89034) and conventional control grain were ground and then heated in an oven to simulate the heating process used commercially to process maize grain. Heated and unheated samples of test and control grain were extracted with two buffers: 50 mM CAPS [3-(Cyclohexylamino)-1-propanesulfonic acid] and 50 mM CAPS containing 2% N-Lauroyl sarcosine (NLS). The extracts were analyzed using the western blot method to detect the presence of the Cry1A.105 and Cry2Ab2 proteins. Image analyses of immunoreactive bands on the blot films were used to semi-quantitatively estimate the amount of immunoreactive Cry1A. 105 and Cry2Ab2 proteins in the heated and unheated test and control grain extracts. Results from this study indicated that the amount of immunodetectable Cry1A. 105 protein after heating was below the lower LOD, or had decreased by greater than or equal to 94% and 78% relative to their original values, respectively. This study was provided in Attachment 12 of the application for General Release of MON 89034.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 68
Cry2Ab2 protein In vitro digestibility Cry2Ab2 (>99%) is rapidly digestible in simulated gastric fluid after 30 seconds incubation in SGF and greater than 97% after 15 minutes in SIF. More information is provided below under the allergenicity paragraph. Heat stability In the same heat stability study described for the Cry1A.105 protein, similar results were obtained for the Cry2Ab2 protein, with the amount of immunodetectable Cry2Ab2 protein after heating below the lower LOD, or had decreased by greater than or equal to 77% and 70% relative to their original values, respectively. Conclusion regarding Cry1A.105 and Cry2Ab2 The additional information provided in the paragraphs above support the fact that exposure to Cry1A.105 and Cry2Ab2 is very low, indicating a high margin of safety regarding exposure. In addition, rapid digestion of these proteins in simulated gastric fluid and lability of the protein to heat during food or feed processing provide additional
assurance to the safety of these proteins. Heating of ground maize grain from MON 89034, in a manner similar to the conditions employed for commercial processing, results in the loss of immunodetectable Cry1A. 105 and Cry2Ab2 proteins. This loss is likely due to protein degradation or aggregation into an insoluble complex as a result of heat treatment. MON 88017 Full details pertaining to the potential toxicity and allergenicity of the Cry3Bb1 and CP4 EPSPS proteins in MON 88017, was previously submitted in view of Monsanto’s applications for Commodity Clearance of MON 88017 (17/3/1-Monsanto-07/649 on 20 December 2006). The safety of the Cry3Bb1 protein The safety of this protein has been established based on the following data and information:
The Cry3Bb1 protein is functionally and structurally similar to Cry proteins that have a demonstrated history of safe use. The Cry3Bb1 protein produced in MON 88017 is nearly (99%) identical in amino acid sequence to the Cry3Bb1
protein in Raven10 Oil Flowable Bioinsecticide, a foliar-applied Bt microbial product that has been used commercially for many years.
The Cry3Bb1 protein has no structural similarity to known toxins or other biologically active proteins that could cause adverse effects in humans or animals, as demonstrated by detailed bioinformatics analyses. A detailed description of the structural similarity to known toxins or biologically active proteins of the Cry3Bb1 protein is given in the application for approval of MON 88017 Commodity Import.
10
Raven is a registered trademark of Certis USA, LLC.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 69
The Cry3Bb1 protein does not exhibit any acute toxic effects in mammals. Results from acute oral toxicity studies with mice demonstrated that the Cry3Bb1 protein is not acutely toxic and do not cause any adverse effects, even at high dose levels that are orders of magnitude greater than human exposure (Refer to respective application for MON 88017).
The safety of the CP4 EPSPS protein The safety of the CP4 EPSPS proteins has been established based on the following data and information:
CP4 EPSPS is structurally and functionally related to endogenous EPSPS enzymes. There is a history of safe consumption of related EPSPS enzymes found naturally in plant material of commonly consumed foods. There is also a history of more than 10 years of safe use/consumption of CP4 EPSPS present in herbicide tolerant soybeans and maize (Delaney et al., 2008).
The CP4 EPSPS protein has no structural similarity to known toxins or other biologically active proteins that could cause adverse effects in humans or animals, as demonstrated by detailed bioinformatics analyses. A detailed description of the structural similarity to known toxins or biologically active proteins of the CP4 EPSPS protein is given in the toxicology section of the application for approval of MON 88017 Commodity Clearance.
The CP4 EPSPS protein does not exhibit any acute toxic effects in mammals. Results from acute oral toxicity studies with mice demonstrated that the CP4 EPSPS protein is not acutely toxic and does not cause any adverse effects, even at high dose levels that are orders of magnitude greater than human exposure (Refer to respective application for MON 88017).
Mode of action of Cry and CP4 EPSPS proteins MON 87460 × MON 89034 × MON 88017 expresses three Cry proteins: the Cry1A.105 protein, which is closely related to Cry1Ab and Cry1Ac proteins derived from B. thuringiensis subsp. kurstaki and to Cry1F protein from subsp. aizawai, the Cry2Ab2 protein derived from B. thuringiensis subsp. kurstaki, and the Cry3Bb1 protein derived from B. thuringiensis subsp. kumamotoensis. The spectrum of activity of Cry1A.105 and Cry2Ab2 proteins remains within the Order Lepidoptera, while the spectrum of activity of Cry3Bb1 within the Order Coleoptera (corn rootworm). A review of the research characterizing the mechanism of action for Bt crystal proteins has been published by Schnepf et al. (1998) and De Maagd et al. (2001a). Based on the accumulated knowledge of Bt Cry proteins, a generalized mode of action has been proposed and includes the following steps: ingestion of crystals by the insect, solubilization of the crystals in the insect midgut, proteolytic processing of the released Cry protein by digestive enzymes to activate the toxin, binding of the toxin to receptors on the surface of midgut epithelial cells of target organisms, formation of membrane ion channels or pores, and consequent disruption of cellular homeostasis (English, 1992). Electrolyte imbalance and pH changes render the gut paralyzed, which causes the insect to stop eating and die (Sacchi et al., 1986). These high affinity Cry-specific receptors have not been detected in intestinal tissues of mammalian species, such as mouse, rat, monkey, or human (Hofmann et al., 1988a; Hofmann et al., 1988b). This

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 70
explains, in part, observations that mammals are not adversely affected by Cry proteins even when administered at oral doses millions of times higher than target insects would ever encounter (Betz et al., 2000). CP4 EPSPS provides tolerance to glyphosate herbicides. Glyphosate (N-phosphonomethyl glycine) inhibits EPSPS (5-enolpruvylshikimate-3-phosphate synthase), an enzyme in the shikimate pathway involved in the production of aromatic amino acids (Franz et al., 1997). Inhibition of the production of aromatic amino acids, which are necessary for plant growth and development, causes the plant to die. The shikimate pathway is found in all plants but is not present in mammals or other animal species. The modes and sites of biological activity are therefore different for the Cry and CP4 EPSPS proteins and there is no known or conceivable mechanism of interaction between these proteins, which could lead to adverse health effects in animals or humans. Taking into consideration the safety assessments already conducted for the single events MON 87460, MON 89034 and MON 88017 and that the individual proteins are not changed when combined in MON 87460 × MON 89034 × MON 88017, there is no reason to anticipate that the combination of the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1 and CP4 EPSPS proteins in MON 87460 × MON 89034 × MON 88017 would result in any toxic effects to humans or animals. Allergenicity Assessment The assessment of the allergenic potential of proteins compares the biochemical characteristics of these proteins to characteristics of known allergens. A protein is not likely to be an allergen if:
The protein is from a non-allergenic source.
The protein does not share structural similarities to known allergens based on the amino acid sequence.
The protein is rapidly digested in simulated gastric fluid. The above four characteristics have been discussed in detail for the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1 and CP4 EPSPS proteins produced in MON 87460 × MON 89034 × MON 88017, in the respective applications for approval of MON 87460, MON 89034 and MON 88017 in South Africa. Potential allergenicity of the inserted proteins in MON 87460 × MON 89034 × MON 88017 has therefore been submitted for evaluation to the Advisory Committee and Executive Council on previous occasions. The proteins are from non-allergenic sources, lack structural similarity to known allergens, are rapidly digested in simulated gastric fluid and constitute a very small portion of the total protein present in the grain of MON 87460 × MON 89034 × MON 88017 maize. Taken together the data lead to the conclusion that the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1 and CP4 EPSPS proteins are unlikely to have any allergenic potential and MON 87460 × MON 89034 × MON 88017 is as safe as conventional maize regarding the risk for allergenicity.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 71
Furthermore, maize is not considered a common allergenic food. Food allergies to maize are of low frequency. Rare cases of occupational allergy to maize dust have been reported. There is no reason to expect that the use of MON 87460 × MON 89034 × MON 88017 will significantly increase the intake and exposure to maize. A possible over expression of any endogenous protein which is not known to be allergenic would be unlikely to alter the overall allergenicity of the whole plant or the allergy risk for consumers. Nutritional Assessment The inherited traits in MON 87460 × MON 89034 × MON 88017 are of agronomic interest and not intended to change any nutritional aspects of this maize, nor is the presence of these traits expected to alter patterns or volumes of maize consumption. Several studies have been conducted to demonstrate equivalence of MON 87460 × MON 89034 × MON 88017 to conventional maize. These studies are summarised below. Compositional analysis Grain and forage tissues of MON 87460 × MON 89034 × MON 88017 and a conventional control with background genetics similar to the test substances were harvested from plants grown at each of the four field sites in Chile. The composition of forage and grain of the test substances from the three sites (CT, CL, and LUM) where both the well-watered and water-limited treatments were achieved were analyzed and statistically compared to the control. Tolerance intervals calculated from the conventional reference substances were also established for each compositional constituent. Seventy-seven different analytes were evaluated to determine the compositional profile of the T/C/R substances. The compositional analysis data for the T/C/R substances can be found in Attachment J DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. Each individual analyte for the test substances was compared to that of the control for both the well-watered and water-limited treatments at each of the three field sites and for the combination of all three sites. Of the 77 components analyzed, 14 fatty acids, sodium, and furfural had greater than 50% of the analytical values that were below the limit of quantitation. These components were not included in the statistical analyses (Harrigan et al., 2009) (Attachment J) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. The overall data set was examined for evidence of compositionally relevant changes. Based on this evaluation, analyte results were as expected from the respective assays and were similar to reference and published ranges for conventional maize. No compositionally relevant changes were obvious. A statistical summary was generated for each of the remaining 61 compositional analytes. Least square means, standard errors, and the range of observed values for the test and control substances are included in Appendix 2. Each test value that had a significant difference from the control (p<0.05) was compared to the 99% tolerance interval generated from the reference substances. A summary of the significant differences (p<0.05) for MON 87460 × MON 89034 × MON 88017 can be found in Table 13 and Table 14. Reported literature and ILSI

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 72
(International Life Sciences Institute) Crop Composition Database (ILSI, 2008)11 ranges for the analytical components present in forage and harvested grain are presented in Table 15. Composition of Forage and Harvested Grain from MON 87460 × MON 89034 × MON 88017 Compared to the Conventional Control under Well-watered Conditions Overall, the results of the analyses of both forage and grain samples showed that there were significant differences (p<0.05) between MON 87460 × MON 89034 × MON 88017 and the conventional control for six of 61 comparisons from the combined-site analysis and 14 of 183 comparisons from the individual site analyses. Mean component values observed to be significantly different between test and control fell within the 99% tolerance interval (Table 13) (Harrigan et al., 2009) (Attachment J) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. In the combined-site analysis of forage, significant differences were found between MON 87460 × MON 89034 × MON 88017 and the control in the values for carbohydrates by calculation, protein, and phosphorus. One individual site comparison for carbohydrates by calculation and phosphorus was observed to be significantly different. No individual site comparison for protein was observed to be significantly different. In the combined-site analysis of grain, three significant differences were observed. These included values for 16:0 palmitic acid, thiamine, and vitamin B6. All three individual site comparisons were observed to be significantly different for thiamine. However, all test values for thiamine fell within the 99% tolerance interval. Only one individual site comparison each for 16:0 palmitic acid and vitamin B6 was observed to be significantly different. For forage, individual site differences in components not recorded in the combined-site analysis included values for moisture and calcium. These were each observed only at a single individual site. For grain, individual site differences in components not recorded in the combined-site analysis included values for moisture, 18:3 linolenic acid, 20:1 eicosenoic acid, 22:0 behenic acid, and vitamin E (all one site). In summary, statistical analysis highlighted no consistent differences across sites in the levels of components from the test and control substances.
Composition of Forage and Harvested Grain from MON 87460 × MON 89034 × MON 88017 Compared to the Conventional Control under Water-limited Conditions
Overall, the results of the analyses of both forage and grain samples showed that there were significant differences (p<0.05) between MON 87460 × MON 89034 × MON 88017 and the conventional control for seven of 61 comparisons from the combined-site analyses and 36 of 183 comparisons from the individual site analyses. Mean component values observed to be significantly different between test and control fell within the 99% tolerance (Table 14) (Harrigan et al., 2009) (Attachment J) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act].
11
International Life Sciences Institute (ILSI) 2008. Crop Composition Database, Version 3.0,
http://www.cropcomposition.org.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 73
In the combined-site analysis of forage, two significant differences were found between MON 87460 × MON 89034 × MON 88017 and the control. These included values for protein and phosphorus. One individual site comparison for phosphorus was observed to be significantly different. No individual site comparisons were observed to be significantly different for protein. In the combined-site analysis of grain, five significant differences were observed. These included values for ADF, 16:0 palmitic acid, 18:3 linolenic acid, thiamine, and vitamin E. All three individual site comparisons were observed to be significantly different for thiamine. However, all test values for thiamine fell within the 99% tolerance interval. One individual site comparison each was observed to be significantly different for ADF, 16:0 palmitic acid, 18:3 linolenic acid, and vitamin E. For forage, individual site differences in components not recorded in the combined-site analysis included values for calcium observed at a single individual site. For grain, individual site differences in components not recorded in the combined-site analysis, included 18:0 stearic acid, 18:1 oleic acid, 18:2 linoleic acid (two sites), and moisture, total fat, NDF, TDF, copper, manganese, alanine, aspartic acid, glutamic acid, isoleucine, leucine, phenylalanine, proline, serine, threonine, tyrosine, valine, 20:0 arachidic acid, 22:0 behenic acid, vitamin B6, and raffinose (all one site). In summary, statistical analysis highlighted no consistent differences across sites in the levels of components from the test and control substances. Compositional analysis: Conclusions In 2006-2007, grain and forage tissues of MON 87460 × MON 89034 × MON 88017 and a conventional control with background genetics similar to that of the test substance were harvested from plants grown at each of four field sites in Chile. The composition of forage and grain of the test substances from the three sites (CT, CL, and LUM) where both the well-watered and water-limited treatments were achieved were analyzed and statistically compared to the control. For MON 87460 × MON 89034 × MON 88017 and the control grown under well-watered conditions, mean component values were not statistically significantly different for 224 (91.8%) of the 244 comparisons made as part of the individual and combined-site analyses. The statistical analysis further showed that for 201 (82.4%) of the 244 comparisons made between MON 87460 × MON 89034 × MON 88017 and the control grown under water-limited conditions, mean component values were not statistically significantly different. All mean component values observed to be statistically significantly different between test and control substances were within the 99% tolerance interval for each respective irrigation treatment. Thus, the grain and forage of MON 87460 × MON 89034 × MON 88017 were considered to be compositionally equivalent to that of conventional maize whether grown under optimal conditions or under a limited irrigation regimen.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 74
Table 13. Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Well-Watered Conditions
Tissue/Site/ Components (Units)
a
Mean MON 87460
× MON 89034 ×
MON 88017b
Mean Control
b
Mean Diff (% of Control)
b
Signif. (p-
value)b
MON 87460 × MON 89034 × MON 88017
Rangeb
99% Tolerance Interval
b,c
Forage Combination of all sites Phosphorus (% DW) 0.15 0.16 -8.85 0.022 0.12 - 0.17 [0.086, 0.22]
Carbohydrates (% DW) 87.94 87.11 0.95 0.040 86.67 - 89.49 [82.51, 92.09]
Protein (%DW) 6.25 6.71 -6.79 0.040 5.25 - 7.31 [4.12, 8.77] CT Moisture (% FW) 73.63 76.10 -3.24 0.001 72.60 - 75.50 [69.22, 81.25] Carbohydrates (% DW) 87.49 86.14 1.57 0.016 86.67 - 87.91 [82.51, 92.09]
Calcium (% DW) 0.25 0.32 -20.58 0.033 0.23 - 0.28 [0.043, 0.46]
LUM Phosphorus (% DW) 0.13 0.15 -13.71 0.039 0.12 - 0.15 [0.086, 0.22]
Grain Combination of all sites 16:0 Palmitic (% Total FA) 11.61 11.15 4.15 0.007 11.32 - 11.75 [7.43, 14.09] Thiamine HCl (mg/kg DW) 2.19 2.86 -23.47 <0.001 1.93 - 2.85 [1.01, 6.00] Vitamin B6 (mg/kg DW) 5.77 6.83 -15.46 0.005 4.67 - 7.75 [1.86, 8.29]
CL Thiamine HCl (mg/kg DW) 2.08 2.88 -27.70 <0.001 1.93 - 2.27 [1.01, 6.00]
Vitamin E (mg/kg DW) 12.63 10.50 20.33 0.001 12.49 - 12.71 [0, 19.32]
CT 16:0 Palmitic (% Total FA) 11.51 11.15 3.26 0.029 11.32 - 11.65 [7.43, 14.09]
Thiamine HCl (mg/kg DW) 2.39 2.91 -17.65 0.004 2.16 - 2.85 [1.01, 6.00]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 75
Table 13 (cont.). Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Well-Watered Conditions
Tissue/Site/ Components (Units)
a
Mean MON 87460
× MON 89034 ×
MON 88017b
Mean Control
b
Mean Diff (% of Control)
b
Signif. (p-
value)b
MON 87460 × MON 89034 × MON 88017
(Range)b
99% Tolerance Interval
b,c
LUM Moisture (% FW) 11.97 12.57 -4.77 0.001 11.80 - 12.10 [10.10, 13.35]
18:3 Linolenic (% Total FA) 1.19 1.23 -2.62 0.029 1.17 - 1.21 [0.72, 1.66]
20:1 Eicosenoic (% Total FA) 0.18 0.19 -7.79 0.018 0.18 - 0.18 [0.10, 0.36]
22:0 Behenic (% Total FA) 0.066 0.16 -58.60 0.011 0.063 - 0.067 [0, 0.48] Thiamine HCl (mg/kg DW) 2.08 2.78 -25.17 <0.001 2.05 - 2.15 [1.01, 6.00]
Vitamin B6 (mg/kg DW) 5.26 6.94 -24.18 0.014 4.67 - 6.43 [1.86, 8.29] aDW= dry weight; FW=fresh weight; FA=fatty acid.
bData obtained from Appendix 2.
cWith 95% confidence, the interval contains 99% of the values expressed in the population of commercial lines. Negative limits were set to
zero.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 76
Table 14. Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Water-limited Conditions
Tissue/Site/ Components (Units)
a
Mean MON 87460
× MON 89034 ×
MON 88017b
Mean Control
b
Mean Diff (% of Control)
b
Signif. (p-
value)b
MON 87460 × MON 89034 × MON 88017
Rangeb
99% Tolerance Interval
b,c
Forage Combination of all sites Protein (% DW) 7.12 7.67 -7.13 0.014 5.16 - 8.98 [2.94, 11.20]
Phosphorus (% DW) 0.15 0.17 -11.28 0.002 0.13 - 0.18 [0.077, 0.23]
CL Calcium (% DW) 0.27 0.35 -22.30 0.012 0.22 - 0.32 [0.085, 0.50]
Phosphorus (% DW) 0.14 0.17 -17.40 0.037 0.13 - 0.15 [0.077, 0.23]
Grain Combination of all sites Acid Detergent Fiber (% DW) 3.05 2.33 30.68 0.010 2.02 - 4.21 [0.88, 4.63] 16:0 Palmitic (% Total FA) 11.76 11.18 5.22 0.001 11.16 - 13.85 [7.71, 14.14]
18:3 Linolenic (% Total FA) 1.17 1.21 -3.50 0.004 1.03 - 1.23 [0.67, 1.76]
Thiamine HCl (mg/kg DW) 2.23 2.98 -25.25 <0.001 1.95 - 2.41 [1.55, 5.85]
Vitamin E (mg/kg DW) 13.76 12.16 13.13 0.009 10.82 - 15.17 [0, 22.61]
CL Total Fat (% DW) 4.09 3.77 8.48 0.017 3.88 - 4.21 [2.07, 5.10]
Acid Detergent Fiber (% DW) 3.61 2.36 52.83 0.031 3.01 - 4.21 [0.88, 4.63]
Neutral Detergent Fiber (% DW) 9.91 8.34 18.82 0.015 8.81 - 10.85 [2.87, 13.22]
Total Dietary Fiber (% DW) 14.75 12.69 16.28 0.011 13.73 - 16.16 [6.50, 17.54]
Tyrosine (% DW) 0.32 0.21 55.19 0.023 0.29 - 0.38 [0.12, 0.52]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 77
Table 14 (cont.). Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Water-limited Conditions
Tissue/Site/ Components (Units)
a
Mean MON 87460
× MON 89034 ×
MON 88017b
Mean Control
b
Mean Diff (% of Control)
b
Signif. (p-
value)b
MON 87460 × MON 89034 × MON 88017
(Range)b
99% Tolerance Interval
b,c
CL (cont.)
16:0 Palmitic (% Total FA) 12.44 11.28 10.37 0.002 11.69 - 13.85 [7.71, 14.14]
18:0 Stearic (% Total FA) 2.09 1.84 13.61 0.001 1.96 - 2.17 [0.71, 2.57]
18:1 Oleic (% Total FA) 21.91 20.74 5.63 0.015 21.63 - 22.38 [12.15, 35.55]
18:2 Linoleic (% Total FA) 61.69 64.30 -4.06 0.001 59.81 - 62.91 [50.63, 72.71]
18:3 Linolenic (% Total FA) 1.13 1.22 -7.40 0.010 1.03 - 1.23 [0.67, 1.76]
20:0 Arachidic (% Total FA) 0.35 0.32 10.77 0.010 0.32 - 0.39 [0.18, 0.52]
22:0 Behenic (% Total FA) 0.17 0.11 58.51 0.027 0.16 - 0.19 [0, 0.32]
Thiamine HCl (mg/kg DW) 2.12 2.91 -27.18 <0.001 1.95 - 2.36 [1.55, 5.85]
Vitamin E (mg/kg DW) 14.16 11.16 26.98 <0.001 12.91 - 15.17 [0, 22.61]
Raffinose (% DW) 0.12 0.14 -13.58 0.033 0.11 - 0.13 [0, 0.21]
CT
Manganese (mg/kg DW) 6.19 6.88 -9.99 0.036 5.68 - 6.46 [0.72, 11.82]
Alanine (% DW) 0.73 0.83 -12.21 0.022 0.60 - 0.83 [0.59, 1.09]
Aspartic Acid (% DW) 0.61 0.67 -9.64 0.018 0.52 - 0.67 [0.52, 0.88] Glutamic Acid (% DW) 1.89 2.14 -11.96 0.023 1.56 - 2.14 [1.51, 2.80]
Isoleucine (% DW) 0.34 0.39 -12.21 0.028 0.28 - 0.38 [0.27, 0.50]
Leucine (% DW) 1.28 1.46 -12.54 0.022 1.04 - 1.45 [0.98, 1.91] Phenylalanine (% DW) 0.50 0.57 -11.39 0.027 0.42 - 0.57 [0.41, 0.72] Proline (% DW) 0.92 1.03 -11.29 0.048 0.74 - 1.03 [0.64, 1.23]
Serine (% DW) 0.48 0.53 -9.81 0.045 0.41 - 0.54 [0.36, 0.71]
Threonine (% DW) 0.33 0.36 -7.97 0.035 0.29 - 0.36 [0.28, 0.44]
Valine (% DW) 0.47 0.52 -10.39 0.033 0.39 - 0.52 [0.39, 0.64]
Thiamine HCl (mg/kg DW) 2.28 3.08 -26.04 <0.001 2.27 - 2.28 [1.55, 5.85] Vitamin B6 (mg/kg DW) 6.93 5.73 20.95 0.024 6.58 - 7.41 [2.06, 9.98]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 78
Table 14 (cont.). Summary of Significant Differences (p<0.05) Comparing MON 87460 × MON 89034 × MON 88017 and the Conventional Control Produced in Chilean Field Trials Conducted in 2006-2007 under Water-limited Conditions
Tissue/Site/ Components (Units)
a
Mean MON 87460
× MON 89034 ×
MON 88017b
Mean Control
b
Mean Diff (% of Control)
b
Signif. (p-
value)b
MON 87460 × MON 89034 × MON 88017
(Range)b
99% Tolerance Interval
b,c
LUM Moisture (% FW) 12.57 11.93 5.31 <0.001 12.20 - 12.90 [10.50, 14.11]
Copper (mg/kg DW) 3.41 2.17 57.48 0.012 2.62 - 3.96 [0.22, 3.82]
18:0 Stearic (% Total FA) 1.78 1.89 -5.50 0.041 1.75 - 1.86 [0.71, 2.57]
18:1 Oleic (% Total FA) 19.86 20.89 -4.93 <0.001 19.66 - 20.18 [12.15, 35.55]
18:2 Linoleic (% Total FA) 64.98 64.17 1.26 0.035 64.35 - 65.35 [50.63, 72.71]
Thiamine HCl (mg/kg DW) 2.29 2.95 -22.51 <0.001 2.17 - 2.41 [1.55, 5.85] aDW= dry weight; FW=fresh weight; FA=fatty acid.
bData obtained from Appendix 2.
cWith 95% confidence, the interval contains 99% of the values expressed in the population of commercial lines. Negative limits were set to
zero.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 79
Table 15. Literature and ILSI Ranges for Components in Maize Forage and Grain
Tissue/Component1 Literature Values
2 ILSI Range
3
Forage Proximates (% dw)
Ash 2.43 - 9.64a; 2 - 6.6
b 1.527 - 9.638
Carbohydrates 83.2 - 91.6b; 76.5 - 87.3
a 76.4 - 92.1
Fat, total 0.35 - 3.62b; 1.42 - 4.57
a 0.296 - 4.570
Moisture (% fw) 56.5 - 80.4a; 55.3 - 75.3
b 49.1 - 81.3
Protein 4.98 - 11.56a 3.14 - 11.57
Fiber (% dw)
Acid detergent fiber (ADF) 18.3 - 41.0b; 17.5 - 38.3
a 16.13 - 47.39
Neutral detergent fiber (NDF) 26.4 - 54.5b; 27.9 - 54.8
a 20.29 - 63.71
Minerals (% dw) (mg/kg dw)
Calcium 0.0969 - 0.3184b 713.9 - 5767.9
Phosphorus 0.1367 - 0.2914b 936.2 - 3704.1
Grain Proximates (% dw)
Ash 1.1 - 3.9d; 0.89 - 6.28
b 0.616 - 6.282
Carbohydrates 77.4 - 87.2b; 82.2 - 88.1
a 77.4 - 89.5
Fat, total 3.1 - 5.7d; 2.48 - 4.81
b 1.742 - 5.823
Moisture (% fw) 7 - 23d; 8.18 - 26.2
b 6.1 - 40.5
Protein 6 - 12d; 9.7 - 16.1
c 6.15 - 17.26
Fiber (% dw)
Acid detergent fiber (ADF) 3.3 - 4.3d; 2.46 - 11.34
a,b 1.82 - 11.34
Neutral detergent fiber (NDF) 8.3 - 11.9d; 7.58 - 15.91
b 5.59 - 22.64
Total fiber 10.99 - 11.41h
8.85 - 35.31 Amino Acids (% dw) (mg/g dw)
Alanine 0.56 - 1.04i
4.39 - 13.93 Arginine 0.27 - 0.57
i 1.19 - 6.39
Aspartic acid 0.48 - 0.85i 3.35 - 12.08
Cystine/Cysteine 0.17 - 0.26i 1.25 - 5.14
Glutamic acid 1.26 – 2.58i 9.65 - 35.36
Glycine 0.29 - 0.45i 1.84 - 5.39
Histidine 0.21 - 0.38i 1.37 - 4.34
Isoleucine 0.22 - 0.50i 1.79 - 6.92
Leucine 0.84 - 1.84i 6.42 - 24.92
Lysine 0.21 - 0.38i 1.72 - 6.68

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 80
Table 15 (cont.). Literature and ILSI Ranges for Components in Maize Forage and Grain
Tissue/Component1 Literature Values
2 ILSI Range
3
Grain (cont.) Amino Acids
(mg/g dw)
Methionine 0.17 - 0.28i 1.24 - 4.68
Phenylalanine 0.32 - 0.64i 2.44 - 9.30
Proline 0.63 - 1.16i 4.62 - 16.32
Serine 0.37 - 0.91i 2.35 - 7.69
Threonine 0.27 - 0.49i 2.24 - 6.66
Tryptophan 0.05 - 0.08i 0.271 - 2.150
Tyrosine 0.12 - 0.48i 1.03 - 6.42
Valine 0.30 - 0.61i 2.66 - 8.55
Fatty Acids (% total FA)
16:0 Palmitic 7 - 19e
7.94 - 20.71 16:1 Palmitoleic 1
e 0.095 - 0.447
18:0 Stearic 1 - 3e
1.02 - 3.40 18:1 Oleic 20 - 46
e 17.4 - 40.2
18:2 Linoleic 35 - 70e
36.2 - 66.5 18:3 Linolenic 0.8 - 2
e 0.57 - 2.25
20:0 Arachidic 0.1 - 2e
0.279 - 0.965 20:1 Eicosenoic - 0.170 - 1.917 22:0 Behenic - 0.110 - 0.349 Vitamins (mg/kg dw) (mg/100g dw)
Folic acid 0.3d 0.0147 - 0.1464
Niacin 9.3 - 70d,e
Riboflavin (Vitamin B2) 0.25 - 5.6
d,e 0.050 - 0.236
Thiamine (Vitamin B1) 3 - 8.6d,e
0.126 - 4.00 Vitamin B6 5.3
d; 9.6
e 0.368 - 1.132
Vitamin E 3 - 12.1e; 17 - 47
d 0.0015 - 0.0687
(mg/g dw) Minerals (mg/kg dw)
Calcium (% dw) 0.01 - 0.1d,e
12.7 - 208.4 Copper (mg/kg dw) 0.9 - 10
d,e 0.73 - 18.50
Iron (mg/kg dw) 1 - 100d,e
10.42 - 49.07 Magnesium (% dw) 0.09 - 1
d,e 594.0 - 1940.0
Manganese (mg/kg dw) 0.7 - 54d,e
1.69 - 14.30 Phosphorus (% dw) 0.26 - 0.75
d,e 1470.0 - 5330.0
Potassium (% dw) 0.32 - 0.72d,e
1810.0 - 6030.0 Zinc (mg/kg dw) 12 - 30
d,e 6.5 - 37.2

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 81
Table 15 (cont.). Literature and ILSI Ranges for Components in Maize Forage and Grain
Tissue/Component1 Literature Values
2 ILSI Range
3
Anti-Nutrients (% dw)
Phytic Acid 0.48 - 1.12a
0.111 - 1.570 Raffinose 0.08 - 0.30
e 0.020 - 0.320
Secondary Metabolites (mg/kg dw)
Ferulic Acid 113 - 1194f; 3000
g 291.9 - 3885.8
p-Coumaric Acid 22 - 75f
53.4 - 576.2 1fw=fresh weight; dw=dry weight
2Literature range references:
a (Ridley et al., 2002), b (Sidhu et al.,
2000), c (Jugenheimer, 1976), d (Watson, 1982), e (Watson, 1987), f (Classen et al., 1990), g (Dowd and
Vega, 1996), h (Choi et al., 1999), I (OECD, 2002).ILSI Database, 200711
.
Conversions: % dw × 104 = mg/kg dw; % dw × 10 = mg/g dw; mg/g dw × 10
3 = mg/kg dw;
mg/100g dw × 10 = mg/kg dw
Agronomic analysis Based on centuries of experience with conventional, domesticated maize in the world, there is negligible potential for maize to be invasive of natural habitats or to persist in the agronomic environment without the aid of human intervention. The maize plant is known as a poor competitor, which outside cultivation has no meaningful impact on the environment.
Well-Watered Treatment (KS and NE sites) Results for the combined site and individual site phenotypic comparisons of MON 87460 × MON 89034 × MON 88017 to the control under well-watered conditions are shown in Table 16 and Table 17, respectively. Minimum and maximum mean values (reference range) observed among the commercially available reference corn hybrids provide benchmark values common to corn for each characteristic (Jones et al., 2007) (Attachment K) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. In the combined-site analysis for the well-watered treatment, no significant differences were detected for seedling vigor, early stand count, number of days after planting to 50% pollen shed and 50% silking, ear height, plant height, number of dropped ears, number of stalk and root lodged plants, final stand count, grain moisture, test weight, and yield (Table 16). A significant difference was detected between MON 87460 × MON 89034 × MON 88017 and the control for stay green. MON 87460 × MON 89034 × MON 88017 exhibited a significantly lower stay green rating (more green tissue) than the control. However, the mean value of MON 87460 × MON 89034 × MON 88017 for stay green was within the range of values of the reference substances; thus, the detected statistical difference is unlikely to be biologically meaningful in terms of increased pest potential (Jones et al., 2007) (Attachment K) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. In the individual site analyses for the well-watered treatment, no significant differences were detected for seedling vigor, early stand count, number of days after planting to 50%

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 82
pollen shed and 50% silking, stay green, ear height, plant height, number of dropped ears, number of stalk lodged plants, final stand count, test weight, and yield (Table 35) (Jones et al., 2007) (Attachment K) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. Two significant differences were detected between MON 87460 × MON 89034 × MON 88017 and the control in the well-watered treatment across two characteristics in the individual site analysis (Table 17). MON 87460 × MON 89034 × MON 88017 exhibited a significantly lower number of root lodged plants than the control (0.0 vs. 15.0 plants, respectively) and a significantly higher percent grain moisture than the control (13.6 vs. 12.4%, respectively) at the KS site (Table 17). However, the differences in root lodged plants and grain moisture were not detected in the combined-site analysis from plants in the well-watered treatment (Table 16). Thus the detected differences for root lodged plants and grain moisture at the KS site are unlikely to be biologically meaningful in terms of increased pest potential (Jones et al., 2007) (Attachment K) DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]. Ecological Interactions Ecological interaction evaluations were conducted as part of the plant characterization of MON 87460 × MON 89034 × MON 88017, and will be used as part of the environmental risk assessment. The ecological interaction evaluations included the collection and comparison of data on plant response to abiotic stressors, disease damage, and arthropod damage. Abiotic Stress Response In the well-watered treatment, no differences in plant response to abiotic stressors were observed between MON 87460 × MON 89034 × MON 88017 and the control from hail, heat, water logging, or wind evaluated using the observational severity scale at the KS and NE sites (Table 18). Disease Damage In the well-watered treatment, no differences in disease damage were observed between MON 87460 × MON 89034 × MON 88017 and the control from crazy top, ear rot, gray leaf spot, northern corn leaf blight, rust, seedling blight, smut, or stalk rot evaluated using the observational severity scale at KS and NE sites (Table 19). Arthropod Damage In an assessment of arthropod damage in the well-watered treatment, no differences in arthropod damage were observed between MON 87460 × MON 89034 × MON 88017 and the control for leafhoppers, grasshoppers, corn rootworm adults, wireworms, or leafrollers using the observational severity scale at the KS and NE sites (Table 20). The results of the ecological interactions evaluation for MON 87460 × MON 89034 × MON 88017 support the conclusion that the introduction of the drought tolerance trait did not unexpectedly alter MON 87460 × MON 89034 × MON 88017 compared to conventional corn in terms of increased pest potential.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 83
Conclusions for MON 87460 × MON 89034 × MON 88017 The phenotypic characteristics and ecological interactions evaluated in this study were used to characterize the plant and assess the pest or weed potential of MON 87460 × MON 89034 × MON 88017 compared a conventional maize control grown under well-watered conditions. Ultimately, the data will be considered in an ecological risk assessment. Based on the assessed characteristics, the results of this study support a conclusion that the introduction of the drought tolerance, lepidopteran-protected, corn rootworm-protected, or glyphosate tolerance traits did not unexpectedly alter the phenotype or ecological interactions of MON 87460 × MON 89034 × MON 88017 compared to conventional maize under well-watered conditions. Thus, the results support a conclusion of no increased pest or weed potential or altered environmental impact of MON 87460 × MON 89034 × MON 88017 compared to conventional maize.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 84
Table 16. Combined-Site Analysis of Phenotypic Characteristics of Test Substance MON 87460 × MON 89034 × MON 88017 Compared to the Control in the Well-Watered Treatment in 2007
Phenotypic Characteristic (units) Mean Reference Range
Test Control Min. Max.
Seedling vigor (0-9 scale) 2.5 2.2 1.3 2.7
Early stand count (#/plot) 59.2 59.2 52.0 68.7
Days to 50% pollen shed 64.5 63.3 61.0 69.0
Days to 50% silking 64.0 62.3 59.0 69.0
Stay green (0-9 scale) 5.3* 6.7 2.0 9.0
Ear height (in) 43.4 37.6 26.0 45.8
Plant height (in) 89.8 83.9 72.8 100.1
Dropped ears (#/plot) 0.5 0.3 0.0 1.0
Stalk lodged plants (#/plot) 0.2 1.9 0.0 14.3
Root lodged plants (#/plot) 0.0 5.1 0.0 9.7
Final stand count (#/plot) 58.7 56.1 52.7 63.7
Grain moisture (%) 14.9 13.7 12.3 17.3
Test weight (lbs/bu) 60.8 60.7 59.3 61.5
Yield (bu/a) 175.9 189.6 133.3 261.3 * Indicates a statistically significant difference between the test and control (α=0.05). Note: Reference Range is the minimum and maximum values of the reference substance means.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 85
Table 17. Individual Site Analysis of Phenotypic Characteristics of Test Substance MON 87460 × MON 89034 × MON 88017 Compared to the Control in the Well-Watered Treatment in 2007
Phenotypic Characteristic (units) Site Seedling vigor
(0-9 scale) Early stand count
(#/plot) Days to 50% pollen shed
Days to 50% silking
Stay green (0-9 scale)
Test Control Test Control Test Control Test Control Test Control
KS 2.7 3.0 67.7 65.3 62.7 62.3 62.0 62.0 6.7 8.0
NE 2.3 2.3 50.7 52.3 66.3 66.7 66.0 66.0 4.0 4.0
Phenotypic Characteristic (units) Site Ear height
(in) Plant height
(in) Dropped ears
(#/plot) Stalk lodged plants
(#/plot) Root lodged plants
(#/plot)
Test Control Test Control Test Control Test Control Test Control
KS 43.7 44.2 89.3 88.8 1.0 0.7 0.3 4.7 0.0* 15.0
NE 43.2 40.7 90.3 88.6 0.0 0.0 0.0 1.0 0.0 0.0
Phenotypic Characteristic (units) Site Final stand count
(#/plot) Grain moisture
(%) Test weight
(lbs/bu) Yield (bu/a)
Test Control Test Control Test Control Test Control
KS 63.0 61.3 13.6* 12.4 61.4 61.7 136.0 110.8
NE 54.3 50.3 16.1 15.7 60.1 60.3 215.9 202.8
* Indicates a statistically significant difference between the test and control (α=0.05). 1 The test substance MON 87460 × MON 89034 × MON 88017 was not planted at the TX site.
Note: No comparisons were made for Root lodged plants at the NE site due to a lack of variability.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 86
Table 18. Individual Site Analysis of Abiotic Stressors Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007
Abiotic Stressor
Site Observation 1 (V2-V4) Observation 2 (V10-V15) Observation 3 (VT-R3) Observation 4 (R6)
Test Control Reference Test Control Reference Test Control Reference Test Control Reference
Hail KS × × × × × × × × × no no no
NE no no no × × × no no no no no no
Heat NE × × × no no no no no no × × ×
Water logging
KS no no no-sl no-sl no no-sl no no no-sl × × ×
Wind KS sl sl sl no no no sl sl sl sl sl no-sl
NE no no no no no no no no no no no no
No = None; sl = Slight; × = abiotic stressors not evaluated at this observation and site. Note: No differences were detected between MON 87460 × MON 89034 × MON 88017 and the control.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 87
Table 19. Individual Site Analysis of Disease Damage Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007
Disease
Site Observation 1 (V2-V4) Observation 2 (V10-V15) Observation 3 (VT-R3) Observation 4 (R6)
Test Control Reference Test Control Reference Test Control Reference Test Control Reference
Crazy top
KS × × × × × × × × × no no no
Ear rot1 KS × × × × × × × × × no no-sl no-sl
NE × × × × × × × × × no no no-sl
Gray leaf spot
KS no no no no no no no-sl no-sl no-sl × × ×
NE no no no no no no no no no no no no
Northern corn leaf blight
NE no no no no no no no no no no no no
Rust2 KS no no no no no-sl no-sl no-sl sl no-sl sl sl sl
NE no no no no no no no no no no no no
Seedling blight
KS no no no-sl × × × × × × × × ×
Smut KS × × × no no no no no no no no no-sl
Stalk rot3 KS × × × × × × × × × no-sl no-sl no-mo
NE × × × × × × × × × no no-sl no-sl
no = None; sl = Slight; mo = Moderate; × = diseases not evaluated at this observation and site. Note: No differences were detected between MON 87460 × MON 89034 × MON 88017 and the control. 1 Ear rot assessed by pulling husk back from ears of 5 non-systematically selected plants and evaluating the disease.
2 Includes common rust and leaf rust.
3 Stalk rot assessed by splitting the stalks of 5 non-systematically selected plants and evaluating the disease.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 88
Table 20. Individual Site Analysis of Arthropod Damage Using an Observational Severity Scale for MON 87460 × MON 89034 × MON 88017, the Control, and References in the Well-Watered Treatment in 2007
Disease
Site Observation 1 (V2-V4) Observation 2 (V10-V15) Observation 3 (VT-R3) Observation 4 (R6)
Test Control Reference Test Control Reference Test Control Reference Test Control Reference
Leafhoppers NE × × × no no no × × × × × ×
Grasshoppers NE × × × sl sl no-sl sl sl no-sl no no no
Corn rootworm adults1
KS no-sl no-mo no-mo no no no no no-sl no-sl × × ×
NE × × × × × × no no no no no no
Wireworms NE no no no × × × × × × × × ×
Leafrollers NE no no no × × × × × × × × ×
no = None; sl = Slight; mo = Moderate; se = Severe; × = arthropods not evaluated at this observation and site. Note: No differences were detected MON 87460 × MON 89034 × MON 88017 and the control. 1 Includes western corn rootworm and southern corn rootworm.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 89
Consumption As indicated previously, the inherited traits in MON 87460 × MON 89034 × MON 88017 are of agronomic interest and are not intended to change any nutritional aspects of this maize. Furthermore the presence of MON 87460 × MON 89034 × MON 88017 is not expected to alter patterns or volumes of maize consumption. Extensive compositional analyses demonstrated the substantial equivalence of MON 87460 × MON 89034 × MON 88017 to conventional maize except for the inherited traits. Once compositional equivalence has been established in genetically modified feed that has been modified for agronomic input traits, nutritional equivalence can be assumed (Clark and Ipharraguerre, 2001). Thus, considering that results of the compositional analyses have established the compositional equivalence of MON 87460 × MON 89034 × MON 88017 grain to conventional maize grain, no further nutritional assessments of MON 87460 × MON 89034 × MON 88017 for use as or in food or feed are considered necessary. Human dietary assessment The human safety assessment for proteins introduced into biotechnology-derived crops takes the following into consideration: (a) the source organism from which they are derived, (b) their function and history of safe use, (c) a structural comparison to known allergens, toxins or other biologically active proteins known to have adverse effects in mammals, (d) their stability to heat treatment, (e) their digestibility in a simulated gastrointestinal system, and (f) their potential mammalian toxicity. A summary of the results of acute mammalian toxicity studies conducted with the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins and compares the results to the estimated levels of human exposure to these proteins from consumption of MON 87460 × MON 89034 × MON 88017-derived food products in South Africa. Potential health risks were evaluated by calculating a Margin of Exposure (MOE) between the No Observable Adverse Effect Level (NOAEL) from the acute toxicity studies and a conservative estimate of human dietary exposure. Mammalian Toxicity Most known protein toxins act through acute mechanisms to exert toxicity (Hammond and Fuchs, 1998; Pariza and Johnson, 2001; Sjoblad et al., 1992). The primary exceptions to this rule consist of certain anti-nutritional proteins such as lectins, which showed to manifest toxicity in a short-term (three week) rat feeding study (Liener, 1994). The amino acid sequences of the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins produced in MON 87460 × MON 89034 × MON 88017 are not similar to any of these anti-nutritional proteins or to any other known mammalian protein toxin (Burzio et al., 2008a; Kang and Silvanovich, 2012, 2013a, 2013b, 2014). Cry proteins act through acute mechanisms to control insect pests, and widely used Bt-pesticides (expressing Cry proteins) are not toxic to mammals (McClintock et al., 1995). Therefore, to assess the potential hazards of Bt Cry insect-control proteins that have an acute mode of action, the EPA has determined that the use of high (hazard) acute doses are generally adequate to address protein toxicity (US EPA, 2000). In practice, this is the only toxicity test required by the EPA for assessing safety of

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 90
insecticidal proteins. Furthermore, the results of in vitro digestibility assays conducted with the Cry1A.105, Cry2Ab2, Cry3Bb1, as well as the CspB, NPTII, and CP4 EPSPS proteins indicate that these proteins are rapidly digested in simulated gastric and intestinal fluid assays (Barlow et al., 2013; Fuchs et al., 1993b; Kapadia et al., 2008; Leach et al., 2002; Wang and Hernan, 2013), making systemic exposure to the intact proteins unlikely to occur via dietary consumption of foods derived from MON 87460 × MON 89034 × MON 88017. Based on limited exposure, the acute nature of toxicity for most toxic proteins, and the lack of similarity of the expressed proteins to known protein toxins, conduction of acute mouse oral toxicity studies was considered sufficient to evaluate the toxicity of the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins produced in MON 87460 × MON 89034 × MON 88017. The CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins used in the acute toxicity studies were purified from recombinant E. coli strains. These proteins were shown to be both biochemically and functionally equivalent to the proteins expressed in the MON 87460, MON 89034 and MON 88017 events (Bonner et al., 2003a; Bonner et al., 2003b; Burzio et al., 2008b; Goertz et al., 2005; Gu et al., 2008; Karunanandaa et al., 2006). Because MON 87460 × MON 89034 × MON 88017 is produced by conventional breeding, these protein equivalency studies are applicable to the proteins produced in the combined-trait product MON 87460 × MON 89034 × MON 88017. CspB Mammalian Toxicity CspB protein was administered by oral gavage to 10 male and 10 female CD-1 mice at a total dose level of 2160 mg/kg body weight (Smedley, 2012). There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for CspB was considered to be 2160 mg/kg, the highest dose tested. NPTII Mammalian Toxicity NPTII protein was administered by oral gavage to three groups of 10 male and 10 female CD-1 mice (Fuchs et al., 1993b; MSL 11940, 1992). The total dose levels administered were 100, 1000, and 5000 mg/kg body weight. There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for NPTII was considered to be 5000 mg/kg, the highest dose tested. Cry3Bb1 Mammalian Toxicity Cry3Bb1 protein was administered by oral gavage to 10 male and 10 female CD-1 mice (MSL 18711, 2003). The total dose administered was 1930 mg/kg body weight. There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for the Cry3Bb1 protein was considered to be 1930 mg/kg. Cry1A.105 Mammalian Toxicity Cry1A.105 protein was administered by oral gavage to 10 male and 10 female CD-1 mice at a total dose level of 2072 mg/kg body weight (MSL 20000, 2005). There were

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 91
no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for Cry1A.105 was considered to be 2072 mg/kg body weight, the highest dose tested. Cry2Ab2 Mammalian Toxicity Cry2Ab2 protein was administered by oral gavage to 10 male and 10 female CD-1 mice at a total dose level of 2198 mg/kg body weight (MSL 19901, 2006). There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for Cry2Ab2 was considered to be 2198 mg/kg body weight, the highest dose tested. CP4 EPSPS Mammalian Toxicity CP4 EPSPS protein was administered by oral gavage to three groups of 10 male and 10 female CD-1 mice at dose levels up to 572 mg/kg (Harrison et al., 1996; MSL13077, 1993). There were no treatment-related effects on survival, clinical observations, body weight gain, food consumption or gross pathology. Therefore, the NOAEL for CP4 EPSPS was considered to be 572 mg/kg, the highest dose tested.
ESTIMATED HUMAN EXPOSURE Maize Consumption A good model for estimating human consumption of maize and maize-derived products in South Africa is not readily available. Therefore, estimates of maize consumption were derived from the WHO Global Environmental Monitoring System - Food Contamination Monitoring and Assessment Programme (GEMS/Food,
www.who.int/foodsafety/chem/gems).
The GEMS/Food programme has developed 13 Cluster Diets which are considered to be representative of the major food consumption patterns exhibited by regional and cultural groups around the world. South Africa is represented by Cluster I. The estimates of chronic consumption for each cluster were derived from FAO Food Balance Sheets from 1997 to 2001, weighted by the population of each country. The Food Balance Sheets are based on the total amounts of food produced, imported, exported, and utilized for various purposes, not on food consumption surveys, and are generally considered to overestimate actual dietary consumption. However, they do provide conservative estimates of the average daily per capita consumption across large populations and are intended for use in assessing potential chronic or long-term health risks for the overall population when country-specific food consumption data are not available. The GEMS/Food programme also provides a list of the highest 97.5th percentile single-day “eater-only” consumption values reported by any of the countries providing actual food consumption survey data. These values represent a reasonable estimate of the maximum amount of a food that might be consumed in a single day in any world area, and are intended for use in assessing potential acute or short-term health risks. Separate 97.5th percentile acute consumption values are reported for the general population and for children aged six and under.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 92
Estimates for consumption of maize flour, sweet corn and popcorn from the GEMS/Food Programme are shown in Table 21. Based on the GEMS estimates, the total estimated chronic consumption of these maize products in Cluster I is approximately 211.3 g/person/day (mean per capita consumption). Table 21 also lists the GEMS estimates for acute consumption (97.5th percentile). The acute consumption of maize flour in the general population and in children aged six years and younger was derived from food consumption surveys in France and Australia, respectively. The values for sweet corn and popcorn were derived from food consumption surveys in Thailand and Japan, respectively. Because it is not appropriate to add 97.5th percentile consumption values and because these values are from different world areas, they cannot be summed up to estimate total consumption. Instead, acute consumption of each of these commodities must be evaluated separately. Estimated Consumption of MON 87460 × MON 89034 × MON 88017 Foods derived from MON 87460 × MON 89034 × MON 88017 will represent only a portion of the total maize consumed by South Africa. Regulatory approval is currently being requested only for importation of MON 87460 × MON 89034 × MON 88017 into South Africa, not for cultivation. In addition, most MON 87460 × MON 89034 × MON 88017 imported into South Africa will be field maize. Field maize is a blended commodity that is predominantly used for animal feed and is usually processed before being consumed by humans. Popcorn and some forms of sweet corn (all except corn-on-the cob) are also blended commodities. Thus, except for corn-on-the-cob, most MON 87460 × MON 89034 × MON 88017 grain entering the human food supply would likely be blended with other grain before being processed and/or consumed. However, estimating the percentage of consumed maize products that would likely be derived from MON 87460 × MON 89034 × MON 88017 is difficult. Therefore, for the purposes of this dietary risk assessment, the conservative assumption was made that 100% of all maize products consumed in South Africa will be derived from MON 87460 × MON 89034 × MON 88017. Actual consumption of MON 87460 × MON 89034 × MON 88017 is likely to be far lower.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 93
Table 21. Estimates of Maize Consumption by the WHO GEMS/Food Programme1
Commodity
Chronic Consumption (g/person/day)
Mean per capita in Cluster I,
General Population2
Acute Consumption (g/kg/day) Highest 97.5th Percentile, “Eater-
Only” Worldwide3
General Population
Children ≤ 6 yrs
Maize flour 206.7 2.04 3.16
Popcorn 0.1 3.33 3.33
Sweet corn 4.5 7.16 11.52
Total4 211.3 N/A N/A
N/A = not available. Note: GEMS/Food reports chronic consumption values as g/person/day while acute consumption data are reported as g/kg/day. 1 http://www.who.int/foodsafety/chem/gems/en/. Verified current as of 4/29/2014
2 Data obtained from http://www.who.int/entity/foodsafety/chem/ClusterDietsAug06.xls. Verified current as of 4/29/2014.
3 Data obtained from http://www.who.int/foodsafety/chem/en/acute_hazard_db1.pdf. Verified current as of 4/29/2014.
4 Totals for chronic consumption were calculated by summing the mean values for maize flour, sweet corn, and popcorn. Totals do not include consumption of maize oil or other highly refined fractions that do not contain significant amounts of protein. Total acute consumption cannot be calculated from these data since it is not appropriate to add 97.5
th percentile values
and since values for flour, sweet corn, and popcorn were derived from surveys in different countries.
Estimated Dietary Intake of CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS Proteins from MON 87460 × MON 89034 × MON 88017 The potential dietary intake of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins from the consumption of food derived from MON 87460 × MON 89034 × MON 88017 can be estimated by multiplying the consumption of each commodity by the level of these proteins in each commodity. Only maize flour, sweet corn and popcorn were included in this calculation. Maize oil was excluded from the assessment because this purified fraction (e.g. refined oil) is essentially devoid of protein (Martín-Hernández et al., 2008) and therefore would not be expected to contain significant amounts of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins. There are a number of processes used in maize processing, including high temperature treatments, hydrolyses, soaking in slightly acidic water, and drying that might lower intact protein levels in the final food product. Changes in temperature, pH, and physical disruptions associated with food processing and cooking/preparation generally lead to loss of protein structure and functionality (Hammond and Jez, 2011b). Like other proteins, the CspB, Cry3Bb1, NPTII, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins expressed in MON 87460 × MON 89034 × MON 88017 are labile to high temperatures as evidenced by loss of their functional activity (Goley and Thorp, 2005; Hernan and Bell, 2013; Hernan et al., 2011; Hernan et

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 94
al., 2010a; Hernan et al., 2010b) and are expected to be similarly susceptible to denaturation conditions during processing and cooking of foods containing MON 87460 × MON 89034 × MON 88017. As noted by Hammond and Jez (2011a), “If there is evidence that the introduced protein is heat labile when tested in vitro and is, therefore, unlikely to survive functionally intact during normal food processing, there would be negligible, if any, human dietary exposure to the functionally active protein in typically consumed foods”. Therefore, there is likely to be significantly lower exposures to the functionally active forms of these proteins through consumption of foods containing MON 87460 × MON 89034 × MON 88017 than the levels estimated below. For the purposes of this assessment, the concentration of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins in maize flour, sweet corn, and popcorn were conservatively assumed to be the mean expression values for these proteins in grain from MON 87460 × MON 89034 × MON 88017 grown under well-watered conditions in a 2006-2007 Chilean field trial (Shi, 2010). These values (Table 22) are considered to be representative of the average level of these proteins in MON 87460 × MON 89034 × MON 88017 maize, however, actual expression levels will vary slightly with the year of cultivation, location, and environmental conditions. In the case of NPTII,
protein expression was below the limit of quantification, 0.0047 g/g fresh weight and for the purposes of this dietary risk assessment, the conservative assumption was made that NPTII protein in maize and maize-derived commodities was expressed at a level equal to the limit of quantification (Table 22). The reported protein concentration estimates in this dietary risk assessment are conservative overestimates because they assume no loss of the proteins during storage, processing and/or cooking of the grain or food. Based on the above assumptions, estimated acute dietary intake of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins in South Africa from consumption of MON 87460 × MON 89034 × MON 88017 have been calculated and can be found in Table 23.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 95
Table 22. Mean Expression of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS Proteins in Grain from MON 87460 × MON 89034 × MON 88017 Grown Under Well-Watered Conditions in a 2006-2007 Chilean Field Trial1
Protein Mean Expression (μg/g fresh weight)
CspB 0.043
NPTII2 0.0047
Cry1A.105 4.7
Cry2Ab2 2.6
Cry3Bb1 6.6
CP4 EPSPS 4.2 1 Data derived from protein expression report (Shi, 2010).
2 NPTII expression was below the limit of quantification, 0.0047 μg/g fresh weight (Shi, 2010), but for the purposes of this assessment, expression was assumed to be equal to the limit of quantification.
Table 23. Intake of the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4
EPSPS Proteins from Consumption of Foods Derived from MON 87460 × MON 89034 × MON 88017 in South Africa
1 Acute consumption values fromTable 21.
2 Protein expression values fromTable 22. Values are multiplied by consumption estimates to obtain protein intake estimates.
Commodity
Consumption of Food
Item (g/kg/day)
1
Protein Intake 2 (µg/kg/day)
CspB NPTII Cry1A.
105 Cry2Ab2 Cry3Bb1 CP4
EPSPS
Maize Flour General population 2.04 0.088 0.010 9.6 5.3 13.5 8.6
Children ≤ 6 yrs 3.16 0.136 0.015 14.9 8.2 20.9 13.3
Popcorn
General population 3.33 0.143 0.016 15.7 8.7 22.0 14.0
Children ≤ 6 yrs 3.33 0.143 0.016 15.7 8.7 22.0 14.0
Sweet corn
General population 7.16 0.308 0.034 33.7 18.6 47.3 30.1
Children ≤ 6 yrs 11.52 0.495 0.054 54.1 30.0 76.0 48.4

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 96
MARGINS OF EXPOSURE A common approach used to assess potential health risks from chemicals or other potentially toxic products is to calculate a margin of exposure (MOE) between the NOAEL from an appropriate animal toxicity study and an estimate of human exposure. Since no evidence of mammalian toxicity has been reported for CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS, a dietary risk assessment would normally not be considered necessary. Nevertheless, to provide further assurances of safety, a dietary risk assessment was still conducted. Ideally, high-end (95th or 97.5th percentile) estimates of acute consumption should be used to calculate an MOE relative to the NOAEL from an acute toxicity study. However, high-end estimates of acute consumption were not readily available for the South African population. Therefore, potential health risks from dietary intake of these proteins from consumption of foods derived from MON 87460 × MON 89034 × MON 88017 were evaluated by calculating MOEs based on these dose levels and the 97.5th percentile estimates of acute dietary exposure from GEMS/Food. As indicated in Table 1, the MOEs for CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS were equal to, or several orders of magnitude greater than, 1.2 × 104. These very large MOEs indicate that there are no meaningful risks to human health from dietary exposure to Error! Reference source not found. MON 87460 × MON 89034 × MON 88017 that ay be present in foods derived MON 87460 × MON 89034 × MON 88017.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 97
Table 24. Margins of Exposure for the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS Proteins from Consumption of Foods Derived from MON 87460 × MON 89034 × MON 88017 in South Africa
1 Calculated by dividing NOAELs from acute mouse gavage studies with the CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins by the protein intake estimates (Table 23). MOEs rounded to two significant figures.
Conclusions
No adverse effects were noted in acute mouse oral gavage studies with the CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS proteins. In addition, very large MOEs have been demonstrated for consumption of these proteins in foods derived from MON 87460 × MON 89034 × MON 88017 for the general population and children aged six years and younger. Therefore, there is a reasonable certainty that consumption of MON 87460 × MON 89034 × MON 88017 will not adversely affect the health of adults or children in South Africa.
Commodity
Margin of Exposure1
CspB NPTII Cry1A.105 Cry2Ab2 Cry3Bb1 CP4
EPSPS
Maize Flour
General population 2.5× 10
7 5.2× 10
8 2.2× 10
5 4.1× 10
5 1.4× 10
5 6.7× 10
4
Children ≤ 6 yrs 1.6× 10
7 3.4× 10
8 1.4× 10
5 2.7× 10
5 9.3× 10
4 4.3× 10
4
Popcorn
General population 1.5× 10
7 3.2× 10
8 1.3× 10
5 2.5× 10
5 8.8× 10
4 4.1× 10
4
Children ≤ 6 yrs 1.5× 10
7 3.2× 10
8 1.3× 10
5 2.5× 10
5 8.8× 10
4 4.1× 10
4
Sweet corn
General population 7.0× 10
6 1.5× 10
8 6.2× 10
4 1.2× 10
5 4.1× 10
4 1.9× 10
4
Children ≤ 6 yrs 4.4× 10
6 9.2× 10
7 3.8× 10
4 7.3× 10
4 2.5× 10
4 1.2× 10
4

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 98
Estimated animal maize consumption Maize is fed to poultry and livestock animals such as broiler, swine, and dairy cow. The daily consumption of maize for several animals was estimated using the animal consumption and body weight values for European Union as provided by OECD (OECD, 2009) (Table 25). Estimated animal consumption of MON 87460 × MON 89034 × MON 88017 The exposure of poultry and livestock to MON 87460 × MON 89034 × MON 88017 can result from feeding MON 87460 × MON 89034 × MON 88017 derived maize grain, maize gluten feed, maize gluten meal, and/or, in the case of ruminants, maize silage. Maize bought for animal feed would be expected to have gone through a series of commingling steps with non-MON 87460 × MON 89034 × MON 88017 maize products as the maize makes its way through commerce. MON 87460 × MON 89034 × MON 88017 grown by a poultry or livestock feeder would be expected to be used on the farm, and therefore, livestock could be exposed to the highest anticipated levels of CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins. For this assessment, it will be assumed that maize from MON 87460 × MON 89034 × MON 88017 is the only maize in the diet. Estimated dietary intake of CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins from MON 87460 × MON 89034 × MON 88017 Animals can be exposed to the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins through dietary intake of ingredients derived from MON 87460 × MON 89034 × MON 88017. The quantity of maize consumed on a daily basis by poultry and livestock, as well as the levels of CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in MON 87460 × MON 89034 × MON 88017 are necessary to derive an estimate of daily dietary intake (DDI) of CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins. For example DDI of CSPB protein is computed as follows:
DDI of CSPB protein = Daily consumption of maize grain, maize gluten feed, maize gluten meal, and/or silage (g/kg body weight)
CSPB protein concentration (µg/g) For the purpose of calculating the dietary exposure of animals to CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins expressed in MON 87460 × MON 89034 × MON 88017, the mean and highest expressions of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins for MON 87460 × MON 89034 × MON 88017 were used. The mean and high end range values of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in grain and forage used in this assessment were from MON 87460 × MON 89034 × MON 88017 grown in Chile (Shi, 2010). To calculate the animal exposure, the following assumptions were made.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 99
First, the intake calculations make the conservative assumption that there is no loss of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins during the processing of maize grain or forage into animal feed. Second, it was assumed that 100% of the maize grain or forage ending up in animal feed is derived from MON 87460 × MON 89034 × MON 88017, which could be the case for farmers that grew the maize that was fed to their livestock. However, larger livestock operations purchases commodity maize that is a blend of many different maize including non-MON 87460 × MON 89034 × MON 88017. Third, it was assumed that MON 87460 × MON 89034 × MON 88017 derived maize grain, maize gluten feed, maize gluten meal, and maize silage (for dairy cow) are all provided as a diet which result in an animal diet that is 90-100% maize feedstuffs. This diet would be nutritionally imbalanced, significantly decrease the performance of the animal, and be economically disadvantageous. Thus, the feed ration used in this animal exposure analysis would not be used in actual practice and instead is used in this analysis to calculate the exposure in a worst-case scenario. Fourth, MON 87460 × MON 89034 × MON 88017 was produced in well-watered and water-limited conditions so it was assumed for this assessment that a conservative approach would be to use growing condition with the higher mean and maximum levels of protein expression. For example, CSPB protein expression level for forage was higher in water-limited condition. But the CSPB protein expression level for grain was higher in well-watered condition. Thus, for the calculation used in this assessment, water-limited forage values (mean and highest) were used and well-water grain values (mean and highest) were used. The mean and maximum levels of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in tissues and processed ingredients of MON 87460 × MON 89034 × MON 88017 are shown in Table 26. The estimated mean and maximum DDI of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins by poultry and livestock are shown in Table 3. The highest percentage of CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins per total protein intake was in the lactating dairy cow 0.0002%, 0.00007%, 0.03%, 0.06%, 0.05%, and 0.07%, respectively (Table 27). The percentage of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein intakes per total protein intake is much lower for broiler and pig than lactating dairy cow. Conclusion In the worst-case scenario, per kg bwt, poultry, pig, and lactating dairy cow would be consuming 0. 0.0002%, 0.00007%, 0.03%, 0.06%, 0.05%, and 0.07% or less of their total protein as CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins from MON 87460 × MON 89034 × MON 88017 maize, respectively.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 100
Table 25. The estimated daily consumption of maize in poultry and livestock.1
Species
Maize derived ingredient in
feed
Total diet intake
2
(kg/d)
Body weight
2
(kg)
Amount of ingredient in
diet2
(%)
Daily consumption
of maize1
(g/kg bwt/d)
Broiler
Grain 0.12 1.7 70% 49.4
Gluten Feed 0.12 1.7 10% 7.1
Gluten Meal 0.12 1.7 10% 7.1
Finishing pig
Grain 3.0 100 70% 21.0
Gluten Feed 3.0 100 20% 6.0
Gluten Meal 3.0 100 10% 3.0
Lactating dairy cow
Grain 25.0 650 0% 0.0
Gluten Feed 25.0 650 20% 7.7
Gluten Meal 25.0 650 20% 7.7
Silage 25.0 650 60% 23.1 1
Daily consumption of maize (on dry weight basis) = (total diet intake / bwt) x (amount of ingredient in diet) x 1000 2
Values for the total diet intake, body weight, and amount of ingredient in diet were obtained from OECD (OECD, 2009). For lactating dairy cow, the diet included maize silage, maize gluten meal, and maize gluten feed. The allocation of ingredient is based on first using the ingredient with the higher protein expression until 100% of daily intake is achieved. Thus, in this worst-case scenario, 100% of the diet was achieved without maize grain. Abbreviation: bwt = body weight, d = day.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 101
Table 26. The mean and highest (max) levels of CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein levels in the maize tissues collected from MON 87460 × MON 89034 × MON 880171
Maize tissue CSPB Protein Level (µg/g dwt)
2
NPTII Protein Level
(µg/g dwt)2,5
Cry1A.105 Protein Level
(µg/g dwt)2
Cry2Ab2 Protein Level
(µg/g dwt)2
Cry3Bb1 Protein Level
(µg/g dwt)2
CP4 EPSPS Protein Level
(µg/g dwt)2
Mean Max Mean Max Mean Max Mean Max Mean Max Mean Max
Grain 0.05 0.086 0.0028 0.0028 5.6 6.6 3 3.9 7.6 9.8 4.8 5.7
Gluten Feed3 0.13 0.22 0.01 0.01 14.6 17.2 7.8 10.1 19.8 25.5 12.5 14.8
Gluten Meal3 0.36 0.61 0.02 0.02 39.8 46.9 21.3 27.7 54.0 69.6 34.1 40.5
Forage/Silage4 0.17 0.23 0.14 0.17 30 48 94 140 58 96 72 150
1 (Shi, 2010)
2 dwt = dry weight. fwt = fresh weight.
3 The concentration of expressed protein in maize gluten feed and gluten meal was calculated to be 2.6-fold and 7.1-fold higher,
respectively, than the concentration in maize grain. This is based on an assumption that there is no loss of protein during the processing of maize grain to gluten feed and gluten meal, and the 2.6-fold and 7.1-fold higher protein content (as % of dry matter) of gluten feed and gluten meal than maize grain, respectively (OECD, 2002). 4 The expression levels of CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein levels in forage was used as the
silage values based on a conservative assumption that there is no protein loss.
5 The NPTII protein level in grain was < limits of quantitation (LOQ). When the protein level is <LOQ, the limits of detection (LOD) level
is used to estimate the exposure. The LOD level of 0.0024 (ug/g fwt) convert to 0.0028 (ug/g dwt) and used as the mean and maximum levels in this analysis.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 102
Table 27. Mean and highest (Max) daily dietary intakes (DDIs) of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in poultry and livestock (g/kg bwt/d)1
Total Consumption
of Maize2
(g/kg bwt/d)
CSPB Protein Intake, µg/kg bwt/d
(% of total protein intake)3
NPTII Protein Intake, µg/kg bwt/d
(% of total protein intake)3
Cry1A.105 Protein Intake, µg/kg bwt/d
(% of total protein intake)3
Species Mean Max Mean Max Mean Max
Broiler
63.5 5.9 10.1 0.3 0.3 660 778
(0.00004%) (0.00008%) (0.000002%) (0.000002%) (0.005%) (0.006%)
Finishing pig 30.0 2.9 5.0 0.2 0.2 324 382
(0.00007%) (0.0001%) (0.000004%) (0.000004%) (0.008%) (0.01%)
Lactating dairy cow
38.5 7.7 11.7 3.4 4.1 1110 1600
(0.0001%) (0.0002%) (0.00006%) (0.00007%) (0.02%) (0.03%) 1 The DDI of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein intake = (daily consumption of maize grain and /or silage)
×CspB, NPTII, CRY3Bb1, Cry1A.105, Cry2Ab2, and CP4 EPSPS (CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein concentration). 2
Total consumption of maize is the sum of maize derived ingredients (maize grain, maize gluten feed, maize gluten meal, and/or maize silage) in Table 1. 3 % of total protein intake (per bwt) = (CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein intake) ÷ (daily total protein intake
per bwt). The daily total protein intake per kg bwt is 13.4 g for broiler, 4.0 g for finishing pig, and 5.7 g for lactating dairy cow. The daily total
protein intake per bwt = (daily consumption) × (% of protein in daily consumption) ÷ bwt × 1000. The percent of protein in broiler diet is 19%
(NRC, 1998), finishing pig diet is 13.2% (NRC, 1994), and lactating dairy cow diet is 14.7% (NRC, 2001). bwt = body weight, d = day.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 103
Table 27. (Continued) Mean and highest (Max) daily dietary intakes (DDIs) of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS proteins in poultry and livestock (g/kg bwt/d) 1
Total Consumption
of Maize2
(g/kg bwt/d)
Cry2Ab2 Protein Intake, µg/kg bwt/d
(% of total protein intake)3
Cry3Bb1 Protein Intake, µg/kg bwt/d
(% of total protein intake)3
CP4 EPSPS Protein Intake, µg/kg bwt/d
(% of total protein intake)3
Species Mean Max Mean Max Mean Max
Broiler
63.5 354 460 896 1155 566 672
(0.003%) (0.003%) (0.007%) (0.009%) (0.004%) (0.005%)
Finishing pig
30.0 174 226 440 567 278 330
(0.004%) (0.006%) (0.01%) (0.01%) (0.007%) (0.008%)
Lactating dairy cow
38.5 2393 3522 1906 2947 2020 3887
(0.04%) (0.06%) (0.03%) (0.05%) (0.04%) (0.07%) 1 The DDI of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein intake = (daily consumption of maize grain and /or silage)
× (CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein concentration). 2
Total consumption of maize is the sum of maize derived ingredients (maize grain, maize gluten feed, maize gluten meal, and/or maize silage) in Table 1. 3 % of total protein intake (per bwt) = (CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1, and CP4 EPSPS protein intake) ÷ (daily total protein intake
per bwt). The daily total protein intake per kg bwt is 13.4 g for broiler, 4.0 g for finishing pig, and 5.7 g for lactating dairy cow. The daily total
protein intake per bwt = (daily consumption) × (% of protein in daily consumption) ÷ bwt × 1000. The percent of protein in broiler diet is 19% (NRC, 1998), finishing pig diet is 13.2% (NRC, 1994), and lactating dairy cow diet is 14.7% (NRC, 2001). bwt = body weight, d = day.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 104
6.3 If the foreign gene products are toxic or allergenic in any way, detail how the commodity clearance will be managed to prevent contact with animals or humans that will lead to discomfort or toxicity.
Studies to support a conclusion that the proteins expressed in MON 87460 × MON 89034 × MON 88017, viz CspB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1 and CP4 EPSPS, do not present toxic or allergenic risks to humans or animals have been provided in previous applications submitted for the single events MON 87460, MON 89034 and MON 88017. Maize and maize products derived from MON 87460 × MON 89034 × MON 88017 are therefore substantially equivalent to conventional maize and no special management of these products is required. 6.4 What are the common/major allergens present in the recipient organism
before modification? Evaluation of endogenous allergens are typically performed for crops that fall into the top eight “commonly” allergenic food groups (Chapman et al., 2006). Maize is not one of the top eight important allergenic foods since it only rarely produces clinically relevant allergenic responses (Chapman et al., 2006; Moneret-Vautrin et al., 1998; OECD, 2002). Maize does not contain any known allergens or produce significant quantities of toxins or anti-nutritional factors warranting analytical or toxicological tests (Watson, 1982; White and Pollak, 1995). 6.5 What evidence is there that the genetic modification described in this
application did not result in over-expression of the possible allergens indicated in 6.4 i.e. is the expression of the possible allergens in the non-GM counterpart substantially equivalent to that in the GM organism?
As indicated above, maize is not one of the top eight allergenic foods. There is no a priori reason to expect that the introduction of the CSPB, NPTII, Cry1A.105, Cry2Ab2, Cry3Bb1 and CP4 EPSPS proteins in MON 87460 × MON 89034 × MON 88017 would affect the general protein expression or the expression of specific allergenic proteins in the plant. 6.6 What are the implications of the proposed activity with regard to the health
and safety of the workers, cleaning personnel and any other person that will be directly or indirectly involved in the activity? Please take into consideration the provisions of the Occupational Health and Safety Act, 1993 (Act No. 181 of 1993) and accompanied regulations.
The grain produced from MON 87460 × MON 89034 × MON 88017 is substantially equivalent to conventional maize grains. All activities regarding the health and safety of individuals associated either directly or indirectly with grain/food products derived from MON 87460 × MON 89034 × MON 88017 are the same as conventional maize. 6.7 Indicate the proposed health and safety measures that would be applied to
safeguard employees during the proposed activity. Considering that MON 87460 × MON 89034 × MON 88017 is equivalent to conventional maize grain, no measures are required.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 105
7. ENVIRONMENTAL IMPACT AND PROTECTION 7.1 Detail any long-term effect the commodity clearance of the genetically
modified organism is likely to have on the biotic and abiotic components of the environment.
This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017 and is not intended for use in the environment of South Africa. 7.2 Provide data and information on ecosystems that could be affected by use
of the plant or its products. As indicated above, this is not an application for release of MON 87460 × MON 89034 × MON 88017 into the environment of South Africa. 7.3 Specify what effect the general release of the genetically modified plant will
have on biodiversity. As indicated above, this is not an application for release of MON 87460 × MON 89034 × MON 88017 into the environment of South Africa. 7.4 Specify the measures to be taken in the event of the plant or product being
misused or escaping into an environment for which it is not intended. This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017, i.e. use as food, feed or in processing. As indicated previously, grain imports are made by international grain traders, with the time of importation dependent on the local or regional need for grain. The grain traders would, as per the requirements in terms of the Genetically Modified Organisms Act, 1997 (Act No. 15 of 1997) obtain the necessary permits from the Registrar prior to importation. Importation activities would thus be subjected to the conditions (such as milling) prescribed in the relevant import permits. Information and data provided in this application supports the conclusion that except for the specifically introduced insect protection and herbicide tolerance traits, MON 87460 × MON 89034 × MON 88017 is equivalent to conventional maize. In the unlikely event that some grain may end up in the environment of South Africa before being milled, the impact would be negligible as maize plants cannot survive without human intervention. Furthermore, South Africa is not the centre of origin for Zea mays and there are no wild relatives in South Africa with which maize can outcross and if the grain was to be used as seed for planting, it would represent a F2 generation of which the growth, development and yield is extremely variable and predominantly weak. 7.5 If the foreign genes give rise to crops resistant to agrochemicals, provide
information on the registration of the agrochemicals to be used on the crop.
As indicated above, this is not an application for release of MON MON 87460 × MON 89034 × MON 88017 into the environment of South Africa.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 106
8. SOCIO-ECONOMIC IMPACTS 8.1 Specify what, if any, positive or negative socio-economic impacts the
genetically modified plant will have on communities in the proposed region of release.
As indicated above, this is not an application for release of MON 87460 × MON 89034 × MON 88017 into the environment of South Africa. 9. WASTE DISPOSAL 9.1 Where only a portion of the genetically modified plant will be used for the
product, how will the unused plant parts be disposed of? This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017. MON 87460 × MON 89034 × MON 88017, like any other maize grain contained in an imported consignment, would be used as food, feed or in processing. 10. MONITORING AND ACCIDENTS 10.1 Indicate the methods and plans for monitoring of the GMO (also refer to
Environmental Risk Assessment Framework for genetically modified organisms) in a comprehensive compliance plan
This is an application for commodity clearance approval of MON 87460 × MON 89034 × MON 88017, i.e. use as food, feed or in processing. Intentional release of MON 87460 × MON 89034 × MON 88017 into the environment of South Africa is not intended. As indicated previously, grain imports are made by international grain traders, with the time of importation dependent on the local or regional need for grain. The grain traders would, as per the requirements in terms of the Genetically Modified Organisms Act, 1997 (Act No. 15 of 1997) obtain the necessary permits from the Registrar prior to importation. Importation as well as use of the grain would thus be subjected to the conditions prescribed in the relevant import and use-as-commodity permits. Adherence to these permit conditions would be monitored by the Registrar’s office. 10.2 Indicate any emergency procedures that will be applied in the event of an
accident in a comprehensive contingency plan Information and data provided in this application supports the conclusion that except for the specifically introduced insect protection and herbicide tolerance traits, MON 87460 × MON 89034 × MON 88017 is equivalent to conventional maize. In the unlikely event that some grain may end up in the environment of South Africa before being milled, the impact would be negligible as maize plants cannot survive without human intervention. Furthermore, South Africa is not the centre of origin for Zea mays and there are no wild relatives in South Africa with which maize can outcross and if the grain was to be used as seed for planting, it would represent a F2 generation of which the growth, development and yield is extremely variable and predominantly weak.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 107
11. PATHOGENIC AND ECOLOGICAL IMPACTS
11.1 Submit an evaluation of the foreseeable impacts, in particular any pathogenic and ecologically disruptive impacts.
As indicated before, except for the specifically introduced insect protection and herbicide tolerance traits, MON 87460 × MON 89034 × MON 88017 is equivalent to conventional maize. With this application, MON 87460 × MON 89034 × MON 88017 is destined for use as food, feed and in processing. No environmental release is proposed. In the unlikely event that some grain should end up in the environment no differences in ecological impact are anticipated. 12. RISK MANAGEMENT
12.1 Please indicate any risk management measures that would be required for
commodity clearance. With this application, a comprehensive data package is provided to demonstrate that MON 87460 × MON 89034 × MON 88017 is, except for the specifically introduced traits, equivalent to conventional maize and have no negative impacts on human and animal health. This commodity clearance approval is to support the potential shortage of maize in South Africa and the subsequent need to import grain from exporting countries that may be commercially growing MON 87460 × MON 89034 × MON 88017 and other genetically modified maize. Grain imports are done by grain traders. Grain imported into South Africa for food and feed use would contain different GM events, depending on the events commercially grown in the exporting country. The measures pertaining to monitoring of the imported grain and emergency measures to be taken in an event of accidental spillage would be prescribed by the Executive Council and contained in the permits issued by the Registrar to the importers and the users (i.e. millers), as these are the parties that will handle and use the grain. The risk of accidental release of MON 87460 × MON 89034 × MON 88017 would be negligible. Maize plants are unable to survive without human intervention; they are dependent on cultivation and care to be able to grow, mature and produce seed. Therefore, no feral populations exist in South Africa even after 150 years of cultivation. Thus, even in case of any grain spillage from an imported consignment, possibly containing MON 87460 × MON 89034 × MON 88017, the grain would still require the typical agronomic practices required to ensure a sustainable crop. Furthermore, South Africa is not the centre of origin for maize and there are no close relatives with which maize can outcross. Therefore, should there be an accidental release into the environment, it is extremely unlikely that the traits contained in the consignment of grain would move into existing crop populations and survive.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 108
13. COMPLETE THE AFFIDAVIT The affidavit is an inseparable part of the application form.
COMMON FORMAT FOR Risk Assessment
(In accordance with Annex III of the Cartagena Protocol on Biosafety)
Risk assessment details
1. Country Taking Decision:
South Africa
2. Title: Application for Commodity Clearance of Genetically Modified Organisms (GMO) - MON 87460 × MON 89034 × MON 88017
3. Contact details: Monsanto Company, represented by Monsanto S.A.(Pty) Ltd Monsanto Company 800 N. Lindbergh Boulevard . St. Louis, Missouri 63167 U.S.A Monsanto House, Building No. 4 Fourways Office Park Maizeer Fourways Boulevard and Roos Streets Fourways South Africa
LMO information
4. Name and identity of the living modified organism:
Multi-event stack MON 87460 × MON 89034 × MON 88017
5. Unique identification of the living modified organism:
MON-8746Ø-4 × MON-89Ø34-3 × MON-88Ø17-3
6. Transformation event: MON 87460 × MON 89034 × MON 88017
7. Introduced or Modified Traits:
Choose the trait from the following list: Abiotic environmental tolerance - Drought or water tolerance Altered growth, development and product quality Chemical tolerance - Herbicide tolerance Pest resistance - Insect resistance
8. Techniques used for modification:
Agrobacterium-mediated transformation of embryonic maize cells.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 109
9. Description of gene modification:
The multi event stack contains the cspB gene from Bacillus subtilis, the cry1A.105, cry2Ab2 and cry3Bb1 genes isolated from Bacillus thuringiensis, and the cp4 epsps gene from Agrobacterium tumefaciens sp. strain CP4.
Characteristics of modification
10. Vector characteristics (Annex III.9(c)):
Not applicable, MON 87460 × MON 89034 × MON 88017 was obtained through conventional breeding methods.
11. Insert or inserts (Annex III.9(d)):
MON 87460 × MON 89034 × MON 88017 maize was obtained by conventional breeding of three single maize event products: MON 87460, MON 89034 and MON 88017.
MON 87460 × MON 89034 × MON 88017 expresses the CspB protein, providing against abiotic stress.
MON 87460 × MON 89034 × MON 88017 expresses the Cry1A.105 and Cry2Ab2 insecticidal proteins, providing tolerance to inter alia maize stalk borers in South Africa.
MON 87460 × MON 89034 × MON 88017 expresses the Cry3Bb1 insecticidal protein that provides protection against certain coleopteran insect pests belonging to the Chrysomelidae family (corn rootworm, Diabrotica spp.) and the CP4 5-enolpyruvyl shikimate-3-phosphate synthase (CP4 EPSPS) protein that confers tolerance to glyphosate.
Recipient organism or parental organisms (Annex III.9(a)):
12. Taxonomic name/status of recipient organism or parental organisms:
Common name: Maize
Family name: Gramineae
Genus: Zea
Species: mays (2n+20)
13. Common name of recipient organism or parental organisms:
Maize or Corn
14. Point of collection or acquisition of recipient or parental organisms:
MON 87460 × MON 89034 × MON 88017 was produced using elite parent lines into which the MON 87460, MON 89034 and MON 88017 events have been introgressed independently, using conventional breeding techniques. The original transformations that produced the individual events used privately owned germplasm acquired for this purpose.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 110
15. Characteristics of recipient organism or parental organisms related to biosafety:
Maize is the world’s third leading cereal, following rice and wheat, in terms of production and area harvested. It has a long history of safe use as a raw material for processed products, and direct uses as a human food or animal feed. Today, maize is produced on every continent except Antarctica, and is exported and imported as viable grain for use as foods or feeds, or directly in processing, without risk to the environment.
According to the OECD [Consensus Document on the Biology of Zea mays subsp. mays (Maize), 2003], “Maize has lost the ability to survive in the wild due to its long process of domestication, and needs human intervention to disseminate its seed”. In addition, “maize is incapable of sustained reproduction outside of domestic cultivation”, and “maize plants are non-invasive in natural habitats.” Despite the fact that maize frequently appears as a volunteer plant in a subsequent rotation, it has no inherent ability to persist or propagate. In all regions of the world, volunteer plants are managed with herbicides, tillage, or manual removal of plants. As such, maize is not considered to be a pest anywhere in the world. When it occurs outside of cultivation, it has no impact on the conservation and sustainable use of biological diversity.
Gene flow from maize occurs through dispersal of seed and pollen-mediated exchange of genes to sexually compatible plants. Since maize has no biological mechanism to scatter seed, low-level, incidental dispersal of viable grain occurs as a result of human-based activities such as transport and harvesting operations. As was noted by the OECD, the few plants that might result from incidental release will not persist or meaningfully reproduce without human intervention. Gene flow via pollen is only possible to other maize plants throughout the world except in Mexico and Guatemala where wild relatives occur. Maize reproduces sexually, is a wind-pollinated, monoecious species with separate staminate (tassels) and pistillate (silk) flowers, which encourages natural cross-pollination between maize plants. The distance that viable pollen can travel depends on prevailing wind patterns, humidity, and temperature. Generally, the pollen dissemination period lasts three to seven days. Because incidental release of maize during importation occurs at very low levels, and because maize in not competitive, pollen-mediated gene flow between local maize and rare volunteers has had no effect on the conservation and sustainable use of biological diversity.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 111
16. Centre(s) of origin of recipient organism or parental organisms:
Maize is thought to have its origin in Mexico, from where it spread northward to Canada and southward to Argentina. Although secondary centres of origin in South America are possible, the oldest archaeological evidence of domesticated maize (5000 B.C.) was discovered in the valley of Tehuacan in Mexico (Benson and Pearce, 1987). Several theories on the origin of maize have been proposed; the two theories most adhered to being that either teosinte (a wild relative of maize that is endemic to parts of Mexico and Guatemala) or a wild pod maize that is now extinct was the wild ancestor of maize (Benson and Pearce, 1987; Brown et al., 1984).
Maize is a member of the genus Zea, which is broken into 2 sections: ZEA and LUXURIENTES. The section ZEA includes one species (mays), which includes three subspecies: ssp. mays, ssp. mexicana (formerly Euchlaena mexicana), and ssp. parviglumis. The former subspecies is known as maize while the latter comprise a portion of the complex known as teosinte. Furthermore, ssp. mexicana and ssp. parviglumis are further separated into several races (OECD, 2003). Section LUXURIENTES encompasses 3 species: an annual Z. luxurians, and perennials Z. diploperennis and Z. perennis. While the classification of Zea continues to be modified, teosintes are the only know wild relatives of maize capable of forming hybrids in nature. Outcrossing and gene exchange between teosinte and maize has been reported with annual teosinte (Zea mays ssp. mexicana) (2n = 20) and maize (Zea mays L.) (2n = 20). A frequency of one F1 hybrid (maize × teosinte) for every 500 maize plants or 20 to 50 teosinte plants in the Chalco region of the Valley of Mexico was reported. However, newer information shows that annual teosintes may be a separate species because of the level of genetic isolation and that hybrids that do form are highly unsuccessful in introgressing genetic material (OECD, 2003). Regardless, Mexico and parts of Central America are regarded as the center of genetic diversity for maize. The natural distribution of teosinte is limited to the seasonally dry, suropical zone with summer rain along the western escarpment of Mexico and Guatemala and the Central Plateau of Mexico.
The belief that Central America and southern Mexico are both the center of origin and a center of diversity for maize was supported by Vavilov (1992).
17. Centres of genetic diversity, if known, of recipient organism or parental organisms:
Refer to 16 above

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 112
18. Habitats where the recipient organism or parental organisms may persist or proliferate:
As noted by OECD (2003), maize is not invasive of natural habitats, does not persist or disperse anywhere in the world without the human intervention. Early domestication and diversification through selection occurred in Meso-America. Maize is grown across a wide range of ecological conditions including soil types, altitude and rainfall. Currently, maize is grown over a wide range of conditions because of its many divergent types that have been bred for this purpose. The bulk of the maize is produced between latitudes 30°and 55°, with relatively little grown at latitudes higher than 47°latitude anywhere in the world. The greatest maize production occurs where the warmest month isotherms range between 21°C and 27°C and the frost-free season lasts 120 to 180 days. A summer rainfall of 15 cm is approximately the lower limit for maize production without irrigation.
Experience with maize imported for use as foods or feeds, or directly in processing, has demonstrated that stable populations do not establish, persist or proliferate as a result of this practice.
Donor organism or organisms (Annex III.9(b)):
19. Taxonomic name/status of donor organism(s)
MON 87460 × MON 89034 × MON 88017 was obtained from conventional breeding of the single events MON 87460, MON 89034 and MON 88017.
The donor organisms used in development of the single events MON 87460, MON 89034 and MON 88017 were Bacillus subtilis, Bacillus thuringiensis and Agrobacterium tumefaciens strain CP4.
20. Common name of donor organism(s):
Bacteria
21. Point of collection or acquisition of donor organism(s):
The organisms are ubiquitous in nature
22. Characteristics of donor organism(s) related to biosafety:
Not applicable, since the donor organisms are ubiquitous in nature and therefore do not pose a threat to biodiversity.
Intended use and receiving environment
23. Intended use of the LMO (Annex III 9(g)):
The intent of this application is for the commodity import of maize containing MON 87460 × MON 89034 × MON 88017
24. Receiving environment (Annex III.9(h)):
The receiving environments are varied and represent the maize producing area in South Africa. There are no reported and scientifically documented cases of any impact on biodiversity or animal and human safety concerns.
Risk assessment summary
NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 113
25. Detection/Identification method of the LMO (Annex III.9(f)):
Standard molecular biology techniques may be used for detection including, but not limited to Southern or PCR, for terminators and promoters in this construct.
A detection method for MON 87460 was provided to the Registrar in March 2009. Detection methods for MON 89034 and MON 88017 have been provided similarly.
26. Evaluation of the likelihood of adverse effects (Annex III.8(b)):
Based on the nature of the recipient species (unable to proliferate) and the lack of related and wild species with which
MON 87460 × MON 89034 × MON 88017 can outcross, the likelihood of adverse effects from out-crossing to other related species is negligible.
Transgenic maize hybrids with similar genes have been grown around the world for several years and in South Africa for 17 years without any recorded impact on the environment other than those created by conventional maize production.
Any volunteers could, like conventional maize, be removed by current agricultural practices such as ploughing and the use of herbicides.
27. Evaluation of the consequences (Annex III.8(c)):
Studies conducted with MON 87460 ×
MON 89034 × MON 88017 confirmed that this event is
agronomically and compositionally equivalent to conventional maize and has no increased tendency towards weediness or an increased susceptibility of tolerance to insects normally associated with maize. Thus, should any of the potential risks materialize, the consequences would be negligible.
No potentially adverse effects were detected based on extensive
characterization of MON 87460 × MON 89034 × MON 88017, which included molecular analysis, expression analysis, compositional analyses and phenotypic evaluation.
Testing of MON 87460 × MON 89034 × MON 88017 demonstrated no changes in its ability to persist in the environment without human intervention or to become invasive compared to conventional maize. As such, the potential
consequences to biodiversity resulting from MON 87460 × MON
89034 × MON 88017 for commercial use, including food, feed or processing, are the same as with conventional maize.
Any volunteer seed germinating in subsequent growing seasons would be detected in the fields that were planted and destroyed using chemical or mechanical means.
28. Overall risk (Annex III.8(d)):
Considering the potential risks and the consequences should the potential risks materialize, the overall risk of importing, cultivating or field testing MON 87460 × MON 89034 × MON 88017 is extremely low.
29. Recommendation (Annex III.8(e)):
No risks have been identified and therefore other than the containment parameters that might apply through the permit conditions, no additional actions need to be taken.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 114
30. Actions to address uncertainty regarding the level of risk (Annex III.8(f)):
The potential risks for the specific product is negligible; hence no additional actions are required except compliance with the conditions contained in the permit.
Additional information
31. Availability of detailed risk assessment information:
More information regarding the safety of MON 87460 × MON 89034 × MON 88017 is contained in the application preceding this section. Additional information can be obtained from the individual event dossiers submitted for MON 87460, MON 89034 and MON 88017.
32. Any other relevant information:
None
33. Attach document: Not applicable to applicant
34. Notes: None

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 115
14. AFFIDAVIT/VERKLARING/STATEMENT
DELETED: Section 68(a), (b) and (c) ii of the Promotion of Access to Information Act]

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 116
15. References
Adu-Tutu K, 2008. Amended Report for MSL0021095: Field Production of Tissues and Grain from Drought Tolerant Corn MON 87460, MON 87460 x NK603, MON 87460 x MON 89034 x NK603, and MON 87460 x MON 89034 x MON 88017 in Chile during 2006-2007. Monsanto Technical Report, MSL0021759, 1-67.
Arfin SM and Bradshaw RA, 1988. Cotranslational processing and protein turnover in eukaryotic cells. Biochem., 27, 7979-7984.
Ashikaga S, Nanamiya H, Ohashi Y and Kawamura F, 2000. Natural genetic competence in Bacillus subtilis natto OK2. Journal of Bacteriology, 182, 2411-2415.
Barker RF, Idler KB, Thompson DV and Kemp JD, 1983. Nucleotide sequence of the T-DNA region from the Agrobacterium tumefaciens octapine Ti Plasmid pTi15955. Plant. Mol. Biol., 2, 335-350.
Barlow R, Wang R and Hernan R, 2013. Assessment of the in vitro digestibility of Cry1A.105 protein in simulated gastric and simulated intestinal fluids. Monsanto Technical Report, MSL0024977, 1-30.
Baum JA, 1998. Transgenic Bacillus thuringiensis. Phytoproction, 79, 127-130. Baum JA, Johnson TB and Carlton BC, 1999. Bacillus thuringiensis; natural and
recombinant bioinsecticide products. Methods in biotechnology, 5: Biopesticides: use and delivery, 189-209.
Beck E, Ludwig G, Auerswald EA, Reiss B and Schaller H, 1982. Nucleotide sequence and exact localization of the neomycin phosphotransferase gene from transposon Tn5. Gene, 19, 327-336.
Benson GO and Pearce RB, 1987. Corn perspective and culture. Corn: Chemistry and Technology, Chapter 1,
Betz FS, Hammond BG and Fuchs RL, 2000. Safety and advantages of Bacillus thuringiensis-protected plants to control insect pests. Reg. Toxicology and Pharmacology, 32, 156-173.
Bevan M, Barnes WM and Chilton MD, 1983. Structure and transcription of the nopaline synthase gene region of T-DNA. Nucleic Acids Res., 11, 369-385.
Bienert R, Zeeb M, Dostal L, Feske A, Magg C, Max K, Welfle H, Balbach J and Heinemann U, 2004. Single-stranded DNA bound to bacterial cold-shock proteins: preliminary crystallographic and Raman analysis. Acta Crystallographica Section D-Biological Crystallography, 60, 755-757.
Blattner FR, Plunkett G, Bloch CA, Perna NT, Burland V, Riley M, ColladoVides J, Glasner JD, Rode CK, Mayhew GF, Gregor J, Davis NW, Kirkpatrick HA, Goeden MA, Rose DJ, Mau B and Shao Y, 1997. The complete genome sequence of Escherichia coli K-12. Science, 277, 1453-1462.
Bogosian G and Kane JF, 1991. Fate of recombinant Escherichia coli K-12 strains in the environment. Adv Appl Microbiol, 36, 87-131.
Bonner HKS, Ganguly T, Vaughn AP and Hileman RE, 2003a. Assessment of the physicochemical and functional equivalence of the Cry3Bb1.pvzmir39 protein produced in grain of MON 88017 corn to the E.coli- produced Cry3Bb1.pvzmir39 protein. Monsanto Technical Report, MSL 18816,
Bonner HKS, Ganguly T, Vaughn AP, Lee JL and Hileman RE, 2003b. Assessment of the physicochemical and functional equivalence of the CP4 EPSPS protein produced in grain of MON 88017 corn to the E.coli- produced CP4 EPSPS protein. Monsanto Technical Report, MSL 18815,
Bravo A, 1997. Phylogenetic relationships of Bacillus thuringiensis delta-endotoxin family proteins and their functional domains. J. Bacteriol., 179, 2793-2801.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 117
Brown SM and Santino CG, 1995. Enhanced expression in plants. United States Patent, Patent Number 5,424,412, 1-45.
Brown WL, Zuber MS, Darrah LL and Glover DY, 1984. Origin, adaptation, and types of corn. Corn: Chemistry and Technology,
Burzio LA, McClain JS and Silvanovich A, 2008a. Bioinformatics evaluation of the CSPB protein utilizing the AD8, TOXIN6, and PROTEIN databases. Monsanto Technical Report, MSL0021261, 1-751.
Burzio LA, Wang C and Rice EA, 2008b. Characterization of the Bacillus subtilis Cold Shock Protein B (CSPB) purified from the grain of drought tolerant corn MON 87460 and comparison of the physicochemical and functional properties of the MON 87460-produced and E. coli-produced CSPB proteins. Monsanto Technical Report, MSL0021140, 1-40.
Caplice E and Fitzgerald GF, 1999. Food fermentations: role of microorganisms in food production and preservation. International Journal of Food Microbiology, 50, 131-149.
Chaikam V and Karlson D, 2008. Functional characterization of two cold shock domain proteins from Oryza sativa. Plant Cell and Environment, 31, 995-1006.
Chapman JA, Bernstein IL, Lee RE, Oppenheimer J, Nicklas RA, Portnoy JM, Sicherer SH, Schuller DE, Spector SL, Khan D, Lang D, Simon RA, Tilles SA, Blessing-Moore J, Wallace D and Teuber SS, 2006. Food allergy: a practice parameter. Annals of Allergy Asthma & Immunology, 96, S1-S68.
Choi IJ, Son JH and Nahm KH, 1999. Dietary fiber fraction for grains containing high levels of water-soluble non-starch polysaccharides. Jpn. Poult. Sci, 36, 269-274.
Clark JH and Ipharraguerre IR, 2001. Livestock Performance: Feeding Biotech Crops. J. Dairy Sci., 84, 9-18.
Classen D, Arnason JT, Serratos JA, Lambert JDH, Nozzolillo C and Philogene BJR, 1990. Correlation of phenolic acid content of maize to resistance to Sitophilus zeamais, the maize weevil, in CIMMYT's collections. J. Chem. Ecol., 16, 301-315.
Codex Alimentarius Commission, 2003. Guideline for the conduct of food safety assessment of foods derived from recombinant-DNA plants. Codex, CAC/GL 45-2003, 1-13.
Codex Standard, 2003. Codex standard for fermented milks. Codex Stan 243-2003, Crickmore N, Zeigler DR, Feitelson J, Schnepf E, Van Rie J, Lereclus D, Baum J and
Dean DH, 1998. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiology and Molecular Biology Reviews, 62, 807-+.
CRO-2007-182, 2008. An acute toxicity study of Cold Shock Protein B administered by the oral (gavage) route to mice. Monsanto Technical Report, CRO-2007-182, 1-95.
De Block M, Herrera-Estrella L, Van Montagu M, Schell J and Zambryski P, 1984. Expression of foreign genes in regenerated plants and in their progeny. EMBO J., 3, 1681-1689.
de Boer AS and Diderichsen B, 1991. On the safety of Bacillus subtilis and B. amyloliquefaciens: a review. Appl Microbiol Biotechnol, 36, 1-4.
De Maagd RA, Bravo A, Berry C, Crickmore N and Schnepf HE, 2003. Structure, diversity, and evolution of protein toxins from spore-forming entomopathogenic bacteria. Annu. Rev. Genet., 37, 409-433.
De Maagd RA, Bravo A and Crickmore N, 2001a. How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Trends Genetics, 17, 193-199.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 118
de Maagd RA, Bravo A and Crickmore N, 2001b. How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Trends in Genetics, 17, 193-199.
Delaney B, Astwood J, Cunny H, Conn R, Herouet-Guicheney C, MacIntosh S, Meyer L, Privalle L, Gao Y, Mattsson J, Levine M and ILSI, 2008. Evaluation of protein safety in the context of agricultural biotechnology. Food and Chemical Toxicology, 46, S71-S97.
Depicker A, Stachel S, Dhaese P, Zambryski P and Goodman HM, 1982. Nopaline synthase: transcript mapping and DNA sequence. J. Mol. Appl. Genet., 1, 561-573.
Derzelle S, Hallet B, Francis KP, Ferain T, Delcour J and Hols P, 2000. Changes in cspL, cspP, and cspC mRNA abundance as a function of cold shock and growth phase in Lactobacillus plantarum. Journal of Bacteriology, 182, 5105-5113.
Dhaese P, De Greve H, Gielen J, Seurinck J, Van Montague M and Schell J, 1983. Identification of sequences involved in polyadenylation of higher plant nuclear transcripts using Agrobacterium T-DNA genes as models. The EMBO Journal, 2, 419-426.
Donovan WP, 1991. CryIIB crystal protein gene from Bacillus thuringiensis. United States Patent, Patent Number 5,073,632, 1-30.
Dowd PF and Vega FE, 1996. Enzymatic oxidation products of allelochemicals as a basis for resistance against insects: effects on the corn leafhopper Dalbulus maidis. Natural Toxins, 4, 85-91.
English LH, 1992. Recovery of Bt endotoxin protein from lysed cell mixtures. United States Patent, Patent Number 5,173,409,
Etchegaray JP and Inouye M, 1999. CspA, CspB, and CspG, major cold shock proteins of Escherichia coli, are induced at low temperature under conditions that completely block protein synthesis. Journal of Bacteriology, 181, 1827-1830.
FAO/WHO, 2000. Safety aspects of genetically modified foods of plant origin, a joint FAO/WHO expert consultation on foods derived from biotechnology. Geneva, Switzerland, 29 May - 2 June 2000,
FAO/WHO, 2001. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria, a joint FAO/WHO expert consultation on evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Córdoba, Argentina, 1 - 4 October 2001,
FDA, 1994. Secondary direct food additives permitted in food for human consumption; food additives permitted in feed and drinking water of animals; aminoglycoside 3'-phosphotransferase II; final rule. Federal Register, 59, 26700-26711.
FDA, 1998. Guidance for industry: use of antibiotic resistance marker genes in transgenic plants. Draft guidance. Editor.
FDA, 1999. Carbohydrase and protease enzyme preparations derived from Bacillus subtilis or Bacillus amyloliquefaciens: affirmation of GRAS status as direct food ingredients; final rule. Federal Register, 64, 19887-19895.
FDA, 2003. Advance notice of public rulemaking. Milk and cream products and yogurt products; petition to revoke standards for lowfat yogurt and nonfat yogurt and to amend standards for yogurt and cultured milk. Federal Register, 68, 39873-39877.
Federici B, 2002. Case study: Bt crops - a novel mode of insect resistance. Genetically Modified Crops: Assessing Safety, 164-200.
Flamm EL, 1991. How FDA approved chymosin: a case history. Bio/Technology, 9, 349-351.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 119
Flavell RB, Dart E, Fuchs RL and Fraley RT, 1992. Selectable marker genes: safe for plants? Bio/Technology, 10, 141-144.
Fraley RT, Rogers SG, Horsch RB, Sanders PR, Flick JS, Adams SP, Bittner ML, Brand LA, Fink CL, Fry JS, Galluppi GR, Goldberg SB, Hoffmann NL and Woo SC, 1983. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA, 80, 4803-4807.
Franz J, Mao MK and Sikorski JA, 1997. Glyphosate: a unique global herbicide. ACS Monograph, 189, 27-65.
Fuchs RL, Heeren RA, Gustafson ME, Rogan GJ, Bartnicki DE, Leimgruber RM, Finn RF, Hershman A and Berberich SA, 1993a. Purification and characterisation of microbially neomycin phosphotransferase II (NPTII) protein and its equivalence to the plant expressed protein. Bio/Technology, 11, 1537-1542.
Fuchs RL, Ream JE, Hammond BG, Naylor MW, Leimgruber RM and Berberich SA, 1993b. Safety assessment of the Neomycin Phosphotransferase II (NPTII) protein. Bio/Technology, 11, 1543-1547.
Goertz B, Ganguly T, Lee J and Rice EA, 2005. Characterization of the Cry1A.105 protein purified from the corn grain of MON 89034 and comparison of the physicochemicam and functional properties of the plant-produced and E.coli-produced Cry1A.105 proteins. Monsanto Technical Report, MSL 19960, 1-51.
Goley ME and Thorp JJ, 2005. Immunodetection of Cry2Ab2 and Cry1A.105 proteins in corn grain from MON 89034 following heat treatment. Monsanto Technical Report, MSL 19899, 1-38.
Graumann P, Wendrich TM, Weber MHW, Schroder K and Marahiel MA, 1997. A family of cold shock proteins in Bacillus subtilis is essential for cellular growth and for efficient protein synthesis at optimal and low temperatures. Molecular Microbiology, 25, 741-756.
Graumann PL and Marahiel MA, 1998. A superfamily of proteins that contain the cold-shock domain. Trends in Biochemical Sciences, 23, 286-290.
Graumann PL and Marahiel MA, 1999. Cold shock proteins CspB and CspC are major stationary-phase-induced proteins in Bacillus subtilis. Archives of Microbiology, 171, 135-138.
Gray CH and Tatum EL, 1944. X-ray induced growth factor requirements in bacteria. Proc Natl Acad Sci USA, 30, 404-410.
Groat JR, Wolff BJ, Rice JF and Masucci JD, 2006. Confirmation of the integrity of corn MON 89034 x MON 88017 by Southern blot analysis. Monsanto Technical Report, MSL 20145,
Gu X, Burzio LA and Rice EA, 2008. Assessment of the physicochemical equivalence of the plant-produced NPTII protein from the leaf of MON 87460 to the E. coli-produced NPTII protein. Monsanto Technical Report, MSL0021145, 1-18.
Hammond B and Fuchs RL, 1998. Safety evaluation of new varieties of food crops developed through biotechnology. Biotechnology and Safety Assessment., 61-79.
Hammond BG and Jez JM, 2011a. Impact of food processing on the safety assessment for proteins introduced into biotechnology. Food and Chemical Toxicology, 49, 711-721.
Hammond BG and Jez JM, 2011b. Impact of food processing on the safety assessment for proteins introduced into biotechnology-derived soybean and corn crops. Food and Chemical Toxicology, 49, 711-721.
Harrigan GG, Miller K and Sorbet R, 2009. Compositional analyses of forage and grain collected from drought tolerant corn MON 87460 × MON 89034 × MON 88017

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 120
grown in a 2006/2007 Chile field production Monsanto Technical Report, MSL 0021182, 1.
Harrison LA, Bailey MR, Naylor MW, Ream JE, Hammond BG, Nida DL, Burnette BL, Nickson TE, Mitsky TA, Taylor ML, Fuchs RL and Padgette SR, 1996. The expressed protein in glyphosate-tolerant soybean, 5- enolypyruvylshikimate-3-phosphate synthase from Agrobacterium sp. strain CP4, is rapidly digested in vitro and is not toxic to acutely gavaged mice. Journal of Nutrition, 126, 728-740.
Henderson L, Gregory J and Swan G, 2002. The national diet and nutrition survey: adults aged 19 to 64 years (Volume 1: Types and quantities of foods consumed). In: HMSO, Norwich,UK, 29-31.
Hernan R and Bell E, 2013. Amended report for MSL0022764: immunodetection of CP4 EPSPS following heat treatment. . Monsanto Techical Report, MSL0024759,
Hernan R, Heeren R, Uffman J and Finnessy J, 2011. The effect of heat treatment on Cry1A.105 functional activity. Monsanto Techical Report, MSL 0023289, 1-24.
Hernan R, Lee J, Deffenbaugh A and Finnessy J, 2010a. The effect of heat treatment on Cry2Ab2 immunodetection. Monsanto Technical Report, MSl 0022958, 1.
Hernan R, Roychowdhury S, Mueller G, Uffman J and Bell E, 2010b. Amended report for MSL 0022564: effect of temperature treatment on the functional activity of Cry2Ab2. Monsanto Techical Report, MSL 0023723, 1-30.
Herschlag D, 1995. RNA chaperones and the RNA folding problem. Journal of Biological Chemistry, 270, 20871-20874.
Hofmann C, Luthy P, Hutter R and Pliska V, 1988a. Binding of the delta endotoxin from Bacillus thuringiensis to brush- border membrane vesicles of the cabbage butterfly (Pieris brassicae). Eur J Biochem, 173, 85-91.
Hofmann C, Vanderbruggen H, Hoefte H, Van Rie J, Jansens S and Van Mellaert H, 1988b. Specificity of Bacillus thuringiensis delta-endotoxins is correlated with the presence of high-affinity binding sites in the brush border membrane of target insect midguts. Proc. Natl. Acad. Sci. USA, 85, 7844-7848.
Höfte H and Whiteley HR, 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev., 53, 242-255.
Hong HA, Duc LH and Cutting SM, 2005. The use of bacterial spore formers as probiotics. FEMS Microbiology Reviews, 29, 813-835.
Horsch RB, Fraley RT, Rogers SG, Sanders PR, Lloyd A and Hoffmann N, 1984. Inheritance of functional foreign genes in plants. Science, 223, 496-498.
Hosoi T, Hirose R, Saegusa S, Ametani A, Kiuchi K and Kaminogawa S, 2003. Cytokine responses of human intestinal epithelial-like Caco-2 cells to the nonpathogenic bacterium Bacillus subtilis (natto). International Journal of Food Microbiology, 82, 255-264.
Huppatz JL and Fitzgerald PA, 2000. Genetically modified food: to grow or not to grow? Medical Journal of Australia, 172, 170-173.
Inatsu Y, Nakamura N, Yuriko Y, Fushimi T, Watanasiritum L and Kawamoto S, 2006. Characterization of Bacillus subtilis strains in Thua nao, a traditional fermented soybean food in northern Thailand. Letters in Applied Microbiology, 43, 237-242.
Jefferson RA, Burgess SM and Hirsh D, 1986. Beta-Glucuronidase from Escherichia coli as a gene-fusion marker. Proc. Natl. Acad. Sci. USA, 83, 8447-8451.
Jensen OL, Podtelejnikov AV and Mann M, 1997. Identification of the components of simple protein mixtures by high-accuracy peptide mass mapping and database searching. Anal. Chem., 69, 4741-4750.
Jones S, Adu-Tutu K, Ahmad A and Sammons B, 2007. Phenotypic evaluations and ecological observations of drought tolerant corn MON 87460, MON 87460 × NK603, MON 87460 × MON 89034 × NK603 and MON 87460 × MON 89034 ×

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 121
MON 88017 under well-watered and water-limited conditions in U.S. field trials during 2007 Monsanto Technical Report, MSL0021700,
Jugenheimer RW, 1976. Corn for special purposes and uses. In: Corn, improvement, seed production and uses. John Wiley & Sons, Inc., New York, 215-258.
Kang H and Silvanovich A, 2012. Updated bioinformatics evaluation of the NPTII protein utilizing the AD_2012, TOX_2012 and PRT_2012 databases. Monsanto Technical Report, MSL0024370, 1.
Kang H and Silvanovich A, 2013a. Updated bioinformatics evaluation of MON 89034 utilizing the AD_2013, TOX_2013, PRT_2013, EST_2013, NT_2013, and NR_2013 databases. Monsanto Technical Report, RAR-2013-0422, 1-385.
Kang H and Silvanovich A, 2013b. Updated bioinformatics evaluation of the Cry3Bb1 protein utilizing the AD_2013, TOX_2013 and PRT_2013 databases. Monsanto Technical Report, MSL0024870, 1-74.
Kang H and Silvanovich A, 2014. Updated bioinformatics evaluation of the CP4 EPSPS protein utilizing the AD_2014, TOX_2014 and PRT_2014 databases. Monsanto Technical Report, MSL0025521, 1-49.
Kapadia SA, Goertz BE, Burzio LA and Rice EA, 2008. Assessment of the in vitro digestibility of the Cold Shock Protein B (CSPB) in simulated gastric and simulated intestinal fluids. Monsanto Technical Report, MSL0021138, 1-35.
Karlson D and Imai R, 2003. Conservation of the cold shock domain protein family in plants. Plant Physiology, 131, 12-15.
Karlson D, Nakaminami K, Toyomasu T and Imai R, 2002. A cold-regulated nucleic acid-binding protein of winter wheat shares a domain with bacterial cold shock proteins. Journal of Biological Chemistry, 277, 35248-35256.
Karunanandaa K, Thorp JJ, Goley ME, Levine SL and Silvanovich A, 2006. Characterization of the Cry2Ab2 protein purified from the corn grain of MON 89034 and comparison of the physicochemical and functional properties of the plant-produced and E. Coli-produced Cry2Ab2 proteins. Monsanto Technical Report, MSL 20071,
Kay R, Chan A, Daly M and McPherson J, 1987. Duplication of CaMV 35S promoter sequences creates a strong enhancer for plant genes. Science, 236, 1299-1302.
Kim JS, Park SJ, Kwak KJ, Kim YO, Kim JY, Song J, Jang B, Jung CH and Kang H, 2007. Cold shock domain proteins and glycine-rich RNA-binding proteins from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli. Nucleic Acids Research, 35, 506-516.
Klee HJ, Muskopf YM and Gasser CS, 1987. Cloning of an Arabidopsis thaliana gene encoding 5-enolpyruvylshikimate- 3-phosphate synthase: sequence analysis and manipulation to obtain glyphosate-tolerant plants. Mol. Gen. Genet., 210, 437-442.
König A, Cockburn A, Crevel RWR, Debruyne E, Grafstroem R, Hammerling U, Kimber I, Knudsen I, Kuiper HA, Peijnenburg AACM, Penninks AH, Poulsen M, Schauzu M and Wal JM, 2004. Assessment of the safety of foods derived from genetically modified (GM) crops. Food and Chemical Toxicology, 42, 1047-1088.
Lamppa GK, Morelli G and Chua NH, 1985. Structure and developmental regulation of a wheat gene encoding the major chlorophyll a/b-binding polypeptide. Mol. Cell. Biol., 5, 1370-1378.
Leach JN, Hileman RE, Thorp JJ, George C and Astwood JD, 2002. Assessment of the in vitro digestibility of purified E. coli-produced CP4 EPSPS protein in simulated gastric fluid. Monsanto Technical Report, MSL 17566, 1-46.
Leroy F, Verluyten J and De Vuyst L, 2006. Functional meat starter cultures for improved sausage fermentation. International Journal of Food Microbiology, 106, 270-285.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 122
Liener I, 1994. Implications of antinutritional components in soybean foods. Critical Reviews in Food Science and Nutrition, 34, 31-67.
Lopitz-Otsoa F, Rementeria A, Elguezabal N and Garaizar J, 2006. Kefir: a symbiotic yeasts-bacteria community with alleged healthy capabilities. Rev Iberoam Micol. , 23, 67-74.
Martín-Hernández C, Bénet S and Obert L, 2008. Determination of proteins in refined and nonrefined oils. Journal of Agricultural and Food Chemistry, 56, 4348-4351.
Masson L, Tabashnik BE, Mazza A, Prefontaine G, Potvin L, Brousseau R and Schwartz JL, 2002. Mutagenic analysis of a conserved region of domain III in the Cry1Ac toxin of Bacillus thuringiensis. Appl. Env. Microbiol., 68, 194-200.
Matsuoka M, Ka Y, Tanaka Y, Ozeki Y and Moto N, 1987. Nucleotide sequence of cDNA encoding the small subunit of ribulose-1,5-bisphosphate carboxylase from maize. J. Biochem, 102, 673-676.
Max KEA, Zeeb M, Bienert R, Balbach J and Heinemann U, 2006. T-rich DNA single strands bind to a preformed site on the bacterial cold shock protein Bs-CspB. Journal of Molecular Biology, 360, 702-714.
Max KEA, Zeeb M, Bienert R, Balbach J and Heinemann U, 2007. Common mode of DNA binding to cold shock domains - Crystal structure of hexathymidine bound to the domain-swapped form of a major cold shock protein from Bacillus caldolyticus. FEBS Journal, 274, 1265-1279.
May JB, 1987. Wet milling: process and products. Corn: Chemistry and Technology, Chapter 12, 377-397.
Mazza P, Zani F and Martelli P, 1992. Studies on the antibiotic resistance of Bacillus subtilis strains used in oral bacteriotherapy. Boll. Chim. Farm., 131, 401-408.
McClintock JT, Schaffer CR and Sjobald RD, 1995. A comparative review of the mammalian toxicity of Bacillus thuringiensis-based pesticides. Pestic. Sci., 45, 95-105.
McElroy D, Blowers AD, Jenes B and Wu R, 1991. Construction of expression vectors based on the rice actin 1 (Act1) 5' region for use in monocot transformation. Mol Gen Genet, 231, 150-160.
McElroy D, Zhang W, Cao J and Wu R, 1990. Isolation of an efficient actin promoter for use in rice transformation. Plant Cell, 2, 163-171.
McElwain EF and Spiker S, 1989. A wheat cDNA clone which is homologous to the 17 kd heat-shock protein gene family of soybean. Nucleic Acids Res., 17, 1764.
Miki B and McHugh S, 2004. Selectable marker genes in transgenic plants: applications, alternatives and biosafety. Journal of Biotechnology, 107, 193-232.
Milan-Carrillo J, Gutierrez-Dorado R, Cuevas-Rodriguez EO, Garzon-Tiznado JA and Reyes-Moreno C, 2004. Nixtamalized flour from quality protein maize (Zea mays L). optimization of alkaline processing. Plant Foods for Human Nutrition, 59, 35-44.
Moneret-Vautrin DA, Kanny G and Beaudouin E, 1998. L'allergie alimentaire au maïs existe-t-elle? Allerg Immunol (Paris), 30, 230.
Morea M, Baruzzi F and Cocconcelli PS, 1999. Molecular and physiological characterization of dominant bacterial populations in traditional Mozzarella cheese processing. Journal of Applied Microbiology, 87, 574-582.
Mozaffar S and Silvanovich A, 2008. Assessment of the CSPB and NPTII protein levels in tissues of drought tolerant corn MON 87460 produced in 2006 U.S. field trials Monsanto Technical Report, MSL0021033, 1-29.
MSL13077, 1993. Acute oral toxicity study of CP4 EPSPS protein in albino mice. Monsanto Technical Report, MSL 13077, 1-174.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 123
MSL 11940, 1992. Acute oral toxicity of Neomycin phosphotransferase (NPT II) in albino mice. Monsanto Technical Report, MSL 11940,
MSL 18711, 2003. An acute oral toxicity study in mice with E.coli- produced Cry3Bb1.pvzmir39 protein. Monsanto Technical Report, MSL 18711,
MSL 19901, 2006. An acute oral toxicity study in mice with Cry2Ab2 protein. Monsanto Technical Report, CRO-2005-049, MSL19901.
MSL 20000, 2005. An acute oral toxicity study in mice with Cry1A.105 protein. Monsanto Technical Report, CRO-2005-050, MSL20000.
Mühldorfer I and Hacker J, 1994. Genetic aspects of Escherichia coli virulence. Microb Pathog, 16, 171-181.
Nakaminami K, Karlson DT and Imai R, 2006. Functional conservation of cold shock domains in bacteria and higher plants. Proceedings of the National Academy of Sciences of the United States of America, 103, 10122-10127.
Nap JP, Bijvoet J and Stiekema WJ, 1992. Biosafety of kanamycin-resistant transgenic plants. Transgenic Research, 1, 239-249.
Newkirk K, Feng W, Jiang W, Tejero R, Emerson SD, Iouye M and Montelione GT, 1994. Solution NMR structure of the major cold shock protein (CspA) from Escherichia coli: identification of a binding epitope for DNA. Proceedings of the National Academy of Sciences, 91, 5114-5118.
NRC, 1998. Nutrient requirements of swine (Tenth revised edition). Editor. National Academy Press, Washington, D.C., 1-2.
NRC, 2001. Nutrient requirements of dairy cattle (Seventh revised edition). Editor. National Academy Press, Washington, D.C., 1-12.
Odell JT, Nagy F and Chua NH, 1985. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature, 313, 810-812.
OECD, 2002. Consensus document on compositional considerations for new varieties of maize (Zea mays): key food and feed nutrients, anti-nutrients and secondary plant metabolites. Organization of European Cooperation and Development, Series on the Safety of Novel Foods and Feeds, OECD ENV/JM/MONO (2002)25, 1-42.
OECD, 2003. Consensus Document on the Biology of Zea Mays Subsp. Mays (Maize). http://www.oecd.org/, 1-49.
OECD, 2009. Guidance document on overview of residue chemistry studies (as revised in 2009). ENV/JM/MONO(2009)31, 1-93.
Ogier JC, Son O, Gruss A, Tailliez P and Delacroix-Buchet A, 2002. Identification of the bacterial microflora in dairy products by temporal temperature gradient gel electrophoresis. Applied and Environmental Microbiology, 68, 3691-3701.
Oguntoyinbo FA, Sanni AI, Franz C and Holzapfel WH, 2007. In vitro fermentation studies for selection and evaluation of Bacillus strains as starter cultures for the production of okpehe, a traditional African fermented condiment. International Journal of Food Microbiology, 113, 208-218.
Olempska-Beer ZS, Merker RI, Ditto MD and DiNovi MJ, 2006. Food-processing enzymes from recombinant microorganisms - a review. Regulatory Toxicology and Pharmacology, 45, 144-158.
Padgette SR, Re DB, Barry GF, Eichholtz DE, Delannay X, Fuchs RL, Kishore GM and Fraley RT, 1996. New weed control opportunities: development of soybeans with a Roundup Ready gene. CRC Handbook, 4, 53-84.
Pariza MW and Johnson EA, 2001. Evaluating the safety of microbial enzyme preparations used in food processing: update for a new century. Regul Toxicol Pharmacol, 33, 173-186.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 124
Pedersen PB, Bjornvad ME, Rasmussen MD and Petersen JN, 2002. Cytotoxic potential of industrial strains of Bacillus sp. Regulatory Toxicology and Pharmacology, 36, 155-161.
Phadtare S, 2004. Recent developments in bacterial cold-shock response. Current Issues in Molecular Biology, 6, 125-136.
Phadtare S and Severinov K, 2005. Nucleic acid melting by Escherichia coli CspE. Nucleic Acids Research, 33, 5583-5590.
Phadtare S, Tyagi S, Inouye M and Severinov K, 2002. Three amino acids in Escherichia coli CspE surface-exposed aromatic patch are critical for nucleic acid melting activity leading to transcription antitermination and cold acclimation of cells. Journal of Biological Chemistry, 277, 46706-46711.
Polevoda B and Sherman F, 2000. N alpha-terminal acetylation of eukaryotic proteins. J. Biol. Chem., 275, 36479-36482.
Rademacher TW, Parekh RB and Dwek RA, 1988. Glycobiology. Annu Rev Biochem, 57, 785-838.
Rice JF, Wolff BJ, Groat JR, Scanlon NK, Jennings JC and Masucci JD, 2006. Molecular analysis of corn MON 89034. Monsanto Technical Report, MSL 20072,
Ridley WP, Sidhu RS, Pyla PD, Nemeth MA, Breeze ML and Astwood JD, 2002. Comparison of the nutritional profile of glyphosate-tolerant corn event NK603 with that of conventional corn (Zea mays L.). Journal of Agricultural and Food Chemistry, 50, 7235-7243.
Ritchie SW, Hanway JJ and Benson GO, 1997. How a corn plant develops: special report #48. Iowa State University of Science and Technology,
Rogers SG, 2000. Promoter for transgenic plants. United States Patent, Patent Number 6,018,100, 1-27.
Rolfe RD, 2000. The role of probiotic cultures in the control of gastrointestinal health. Journal of Nutrition, 130, 396S-402S.
Romano CP, 2002. Expression of Cry3B insecticidal protein in plants. United States Patent, Patent Number 6,501,009,
Russell SH, Hoopes JL and Odell JT, 1992. Directed excision of a transgene from the plant genome. Mol Gen Genet, 1, 49-59.
Sacchi VF, Parenti P, Hanozet GM, Giordana B, Lüthy P and Wolfersberger MG, 1986. Bacillus thuringiensis toxin inhibits K+-gradient-dependent amino acid transport across the brush border membrane of Pieris brassicae midgut cells. Febs Letters, 204, 213-218.
Sambrook J and Russell D, 2001. Protocol 1: Agarose gel electrophoresis. Molecular cloning (Third Edition), Chapter 5, 1-24.
Sanders ME, Morelli L and Tompkins TA, 2003. Sporeformers as human probiotics: Bacillus, Sporolactobacillus, and Brevibacillus. Comprehensive Reviews in Food Science and Food Safety, 2, 101-110.
Sauvageot N, Beaufils S, Maze A, Deutscher J and Hartke A, 2006. Cloning and characterization of a gene encoding a cold-shock protein in Lactobacillus casei. FEMS Microbiology Letters, 254, 55-62.
Schaffer P, Thoma R and Castiglioni P, 2007. Quantitation of CSPB protein from Japanese soy food, natto. Monsanto Technical Report, MSL-0020775,
Schindelin H, Jiang W, Inouye M and Heinemann U, 1994. Crystal structure of CspA, the major cold shock protein of Escherichia coli. Proceedings of the National Academy of Sciences, 91, 5119-5123.
Schindelin H, Marahiel MA and Heinemann U, 1993. Universal nucleic acid-binding domain revealed by crystal structure of the B. subtilis major cold-shock protein. Nature, 364, 164-168.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 125
Schindler T, Graumann PL, Perl D, Ma SF, Schmid FX and Marahiel MA, 1999. The family of cold shock proteins of Bacillus subtilis - Stability and dynamics in vitro and in vivo. Journal of Biological Chemistry, 274, 3407-3413.
Schnepf E, Crickmore N, Van Rie J, Lereclus D, Baum J, Feitelson J, Zeigler DR and Dean DH, 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62, 775-806.
Schröder K, Graumann P, Schnuchel A, Holak TA and Marahiel MA, 1995. Mutational analysis of the putative nucleic acid-binding surface of the cold-shock domain, CspB, revealed an essential role of aromatic and basic residues in binding of single stranded-DNA containing the Y-box motif. Molecular Microbiology, 16, 699-708.
Shi L, 2010. Amended report for MSL0021979: protein expression analyses of tissues collected from drought tolerant corn MON 87460 × MON 89034 × MON 88017 produced under well-watered and water-limited conditions in Chilean field trials during 2006-2007. Monsanto Technical Report, MSL 0022992, 1.
Sidhu RS, Hammond BG, Fuchs RL, Mutz JN, Holden LR, George B and Olson T, 2000. Glyphosate-tolerant corn: the composition and feeding value of grain from glyphosate-tolerant corn is equivalent to that of conventional corn (Zea mays L.). J. Agric. Food Chem., 48, 2305-2312.
Silvanovich A, 2009. The assembly of AD_2009, TOX_2009 and PRT_2009. Monsanto Technical Report, MSL0021840, 1-309.
Sjoblad RD, McClintock JT and Engler R, 1992. Toxicological considerations for protein components of biological pesticide products. Regul. Toxicol. Pharmacol., 15, 3-9.
Skipwith A, Feng D, Groat JR, Tian Q and Masucci JD, 2007. Molecular analysis of corn MON 87460. Monsanto Technical Report, MSL0020487, 1-58.
Smedley J, 2012. An acute toxicity study of cold shock protein B administered by the oral gavage route to mice. Monsanto Technical Report, MSL 0024004, 1.
Sorokulova IB, Pinchuk IV, Denayrolles M, Osipova IG, Huang JM, Cutting SM and Urdaci MC, 2008. The safety of two Bacillus probiotic strains for human use. Digestive Diseases and Sciences, 53, 954-963.
Tanasupawat S, Thongsanit J, Okada S and Komagata K, 2002. Lactic acid bacteria isolated from soy sauce mash in Thailand. Journal of General and Applied Microbiology, 48, 201-209.
Terlabie NN, Sakyi-Dawson E and Amoa-Awu WK, 2006. The comparative ability of four isolates of Bacillus subtilis to ferment soybeans into dawadawa. International Journal of Food Microbiology, 106, 145-152.
Thieringer HA, Jones PG and Inouye M, 1998. Cold shock and adaptation. Bioessays, 20, 49-57.
Tu H, 2009. Updated bioinformatics evaluation of the NPTII protein utilizing the AD_2009 and TOX_2009 databases. Monsanto Technical Report, MSL0021965, 1-13.
Tu H and Silvanovich A, 2009a. Updated bioinformatics evaluation of the CspB protein utilizing the AD_2009 and TOX_2009 databases. Monsanto Technical Report, RAR-09-346, 1-8.
Tu H and Silvanovich A, 2009b. Updated bioinformatics evaluation of the CSPB protein utilizing the PRT_2009 database. Monsanto Technical Report, RAR-09-465, 1-59.
US EPA, 1988. Guidance for the reregistration of pesticide products containing Bacillus thuringiensis as the active ingredient. U.S. Environmental Protection Agency, NTIS PB 89, 164198.

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 126
US EPA, 1995. Plant pesticide Bacillus thuringiensis Cry1A(c) delta-endotoxin and the genetic material necessary for its production in cotton; tolerance exemption; final rule. U.S. Environmental Protection Agency; Fed. Reg., 60, 47871.
US EPA, 1996. Bacillus thuringiensis CryIA (b) delta-endotoxin and the genetic material necessary for its production in all plants: exemption from the requirement of a tolerance; final rule. Fed. Reg. 61:40340,
US EPA, 1997. Bacillus thuringiensis subspecies kurstaki Cry1A(c) and the genetic material necessary for its production in all plants; exemption from the requirement of a tolerance on all raw agricultural commodities; final rule. U.S. Environmental Protection Agency Fed. Reg., 62, 17720.
US EPA, 2000. FIFRA Scientific Advisory Panel on sets of scientific issues being considered by the Environmental Protection Agency regarding: Bt plant-insecticides risk and benefit assessments. U.S. Environmental Protection Agency, October 18 - 2000.
US EPA, 2001. Bacillus thuringiensis Cry3Bb1 and Cry2Ab2 protein and the genetic material necessary for its production in corn and cotton; exemption from the requirement of a tolerance, final rule. Fed. Reg., 66, 24061-24066.
US EPA, 2002. Biopesticide registration action document: Bacillus thuringiensisCry2Ab2 protein and its genetic material necessary for its production in cotton. Amended. U.S. Environmental Protection Agency, http://www.epa.gov/oppbppd1/biopesticides/ingredients/tech_docs/brad_006487.pdf,
US EPA, 2008a. Bacillus thuringiensis Cry1A.105 protein; exemption from the requirement of a tolerance. Federal Register, 73, 40756-40760.
US EPA, 2008b. Bacillus thuringiensis Cry2Ab2 protein; exemption from requirement of a tolerance. Federal Register, 73, 37846-37850.
Vavilov NI, 1992. Origin and geography of cultivated plants. Wang R and Hernan R, 2013. Assessment of the in vitro digestibility of Cry2Ab2 protein
in simulated gastric and simulated intestinal fluids. Monsanto Technical Report, MSL0025099, 1-26.
Watson SA, 1982. Corn: amazing maize, general properties. In: CRC Handbook of Processing and Utilization in Agriculture Plant Products. IA Wolff. CRC Press, Boca Raton, FL, 3-29.
Watson SA, 1987. Structure and composition. In: Corn: chemistry and technology. SA Watson, PE Ramstad. American Association of Cereal Chemists, St. Paul, Minnesota, 53-82.
Weber MHW, Fricke I, Doll N and Marahiel MA, 2002. CSDBase: an interactive database for cold shock domain-containing proteins and the bacterial cold shock response. Nucleic Acids Research, 30, 375-378.
White PJ and Pollak LM, 1995. Corn as a food source in the United States: Part II. Processes, products, composition, and nutritive values. Cereal Foods World, 40, 756-762.
Whitsel JE and Sammons B, 2008. Pollen morphology and viability evaluation of drought tolerant corn MON 87460 under well-watered and water-limited conditions in a U.S. field trial during 2007. Monsanto Technical Report, MSL0021232, 1-26.
WHO 1993. Health aspects of marker genes in genetically modified plants. WHO/FNU/FOS/93,6 report of a WHO workshop. 1-32.
WI-2007-064, 2008. A 90-day feeding study in rats with drought tolerant corn: MON 87460. Monsanto Technical Report, WI-2007-064,

NON-CONFIDENTIAL BUSINESS INFORMATION
Commodity Clearance Application – MON 87460 × MON 89034 × MON 88017 Page 127
Widner WR and Whiteley HR, 1989. Two highly related insecticidal crystal proteins of Bacillus thuringiensis subsp. kurstaki possess different host range specificities. J Bacteriol, 171, 965-974.
Willimsky G, Bang H, Fischer G and Marahiel MA, 1992. Characterization of cspB, a Bacillus subtilis inducible cold shock gene affecting cell viability at low temperatures. J Bacteriol, 174, 6326-6335.