apoptosis: death comes for the cell - hawaii.edulesaux/621/ewexternalfiles/jr lecture 3-2.pdf ·...
TRANSCRIPT

Apoptosis:Death Comes
for the Cell
Joe W. [email protected]
From Ingmar Bergman�s The Seventh Seal
1

2

Mutations in proteins that regulate cell proliferation, survival and death
can contribute to oncogenesis
3
The Chessboard

Ways to Die
4
From Okada and Mak, Nat. Rev. Cancer 4:592-603

The Morphology of Death
5

Apoptosis: Programmed Cell Death
A term used to describe the morphological changes associated with programmed cell death.
The term was originally used by Wyllie and his colleagues and is from the Greek meaning �dropping away� as the leaves from a tree.
6

Apoptosis
l Active cell deathl Requires energy and RNA and protein
synthesisl Characteristic morphological featuresl DNA cleaved, chromatin condensesl Cells shrinkl Formation of apoptotic bodyl Cleared by phagocytosisl No inflammation=no tissue damage 7

Necrosisl Passive cell deathl Cells swell upl Membrane breaks down and cellular contents
leak outl Nucleus disintegratesl Cell ghostsl Inflammatory=tissue damage
8

The function of Cell death
l Multicellular developmentl involved in deletion of entire structures, l sculpting of tissues, l and regulates the neuron number
l The immune responsel The body�s defense against
cancer
9

Death and the mouse�s paw
Fig 18-18
Dark Green fluorescence indicates apoptotic cells.
10

Apoptosis regulates nerve cell targeting
Fig 18-2011

Apoptosis in Lymphocyte development
12

How do we recognize Programmed Cell Death?
13
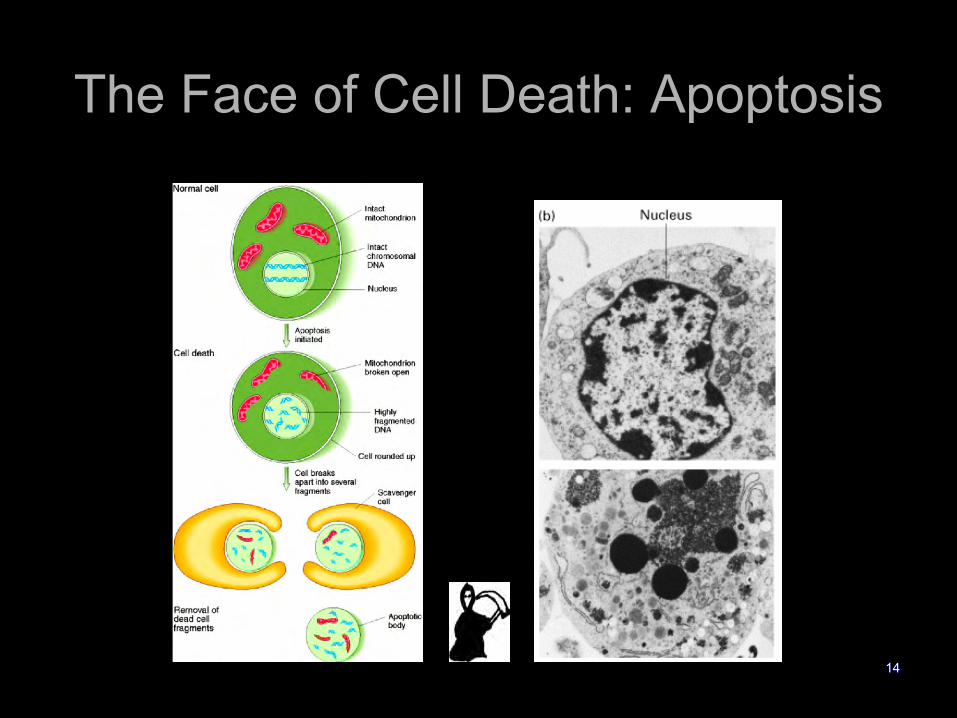
The Face of Cell Death: Apoptosis
14

Detection of apoptotic cells
l Microscopyl Cells have classic features (eg. small darkly stained nuclei)l Detection of free 3� ends of DNA by TUNEL assay (terminal
deoxytransferase-mediated dUTP-biotin nick end labeling)
l Gel electrophoresisl Detect DNA ladder of 180 bp intervals caused by
internucleosomal DNA cleavage
l Flow cytometryl Measure externalization of phosphatidylserine (PS) with
fluorescently labeled Annexin-Vl Measure DNA fragmentation with propidium iodide
fluorescence 15

Analysis of DNA content with a flow cytometer
Recall the fluorescence intensity of the DNA dye (amount of DNA) is measured for each cell.
16
Dead Cells
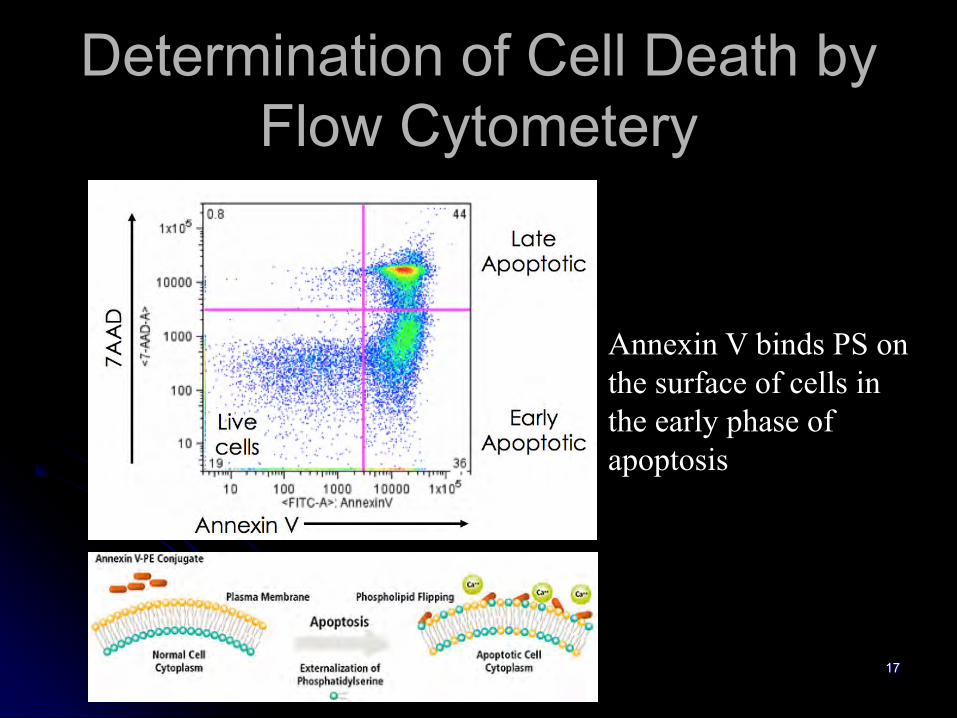
Determination of Cell Death by Flow Cytometery
Annexin V binds PS on the surface of cells in the early phase of apoptosis
17
18

Triggers of apoptosis
l Programmed cell death in which many more cells are produced than survive (e.g. development of lymphocytes)
l Toxic stimuli (viruses, chemicals, ionizing radiation)
l Extracellular signals (Fas, p75 NGF-R, TNF)
l DNA damage (p53)19

C. elegans has played a key role in our understanding of Apoptosis
ced-1 mutant(No engulfment)
ced-1/ced-3(No cells die)
1090 total cells131 die
Ced-3=no death
Ced-4=no death
Ced-9=all die
H.R Horvitz and colleagues responsible for much of this work, 2002 Nobel Prize in Medicine with Sulston and Brenner.20

C. elegans apoptosis
CED-9=Blocks apoptosisCED-4=linker molecule forms activating complex with CED-3CED-3=Protease that executes cell by chewing up proteinsEGL-1=Proapoptotic by blocking CED-9 function
21

Three classes of proteins function in the apoptotic pathway-conserved in
vertebrates
Mammalian Bcl-2 can substitute for Ced-9 in c. elegans
22

Death�s Methods: A protease cascade
Fig 18-22
These proteases are called caspases
23

Caspases
l Caspases are Cysteine directed proteases that cleave after ASPartate residues
l Ced-3 is the C. elegans homologuel At least 14 family membersl Synthesized as proenzymes with low levels of
caspase activity (~1-2 % of active form)l Activated upon after aggregation or cleavage to
mature forml Caspases –8 and –9 are �initiator� caspasesl Caspases –3 is the �effector� caspasel Caspase activation requires a stimulus l They proteolyze cellular proteins to carry out cell death program
24

The Caspase Family
25

Procaspase activation
26

Caspase cascade
27

Two Pathways that Initiate Apoptosis
l Intrinsic/ Mitochondrial Apoptosisl Regulated by Mitochondrial Cytochrome c release
l Extrinsic/ Death Receptor Apoptosisl Activated by ligation of Death Receptorsl Fas, TNF alpha
l These pathways intersect at the effector caspases
28

Activation of the Intrinsic Pathway
29

Intrinsic/Mitochondrial Pathway
CARD domain
30

Mitochondrial permeability
PT=Permeability transition, bursts outer membrane31

Intrinsic Pathway: Apaf-1 Induced Apoptosis
CARD domains
32
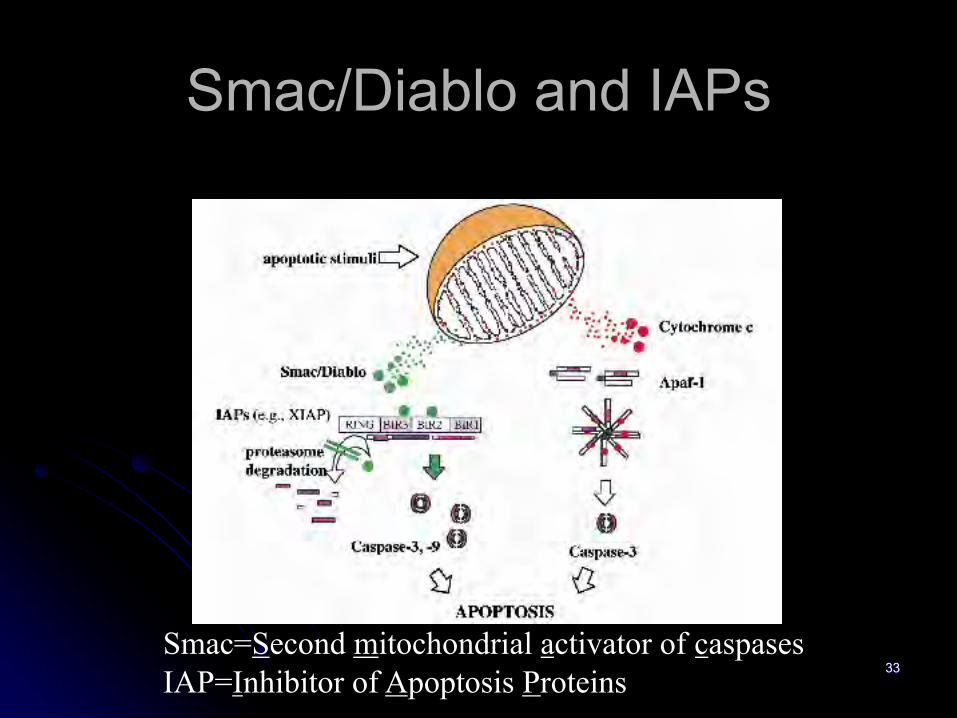
Smac/Diablo and IAPs
Smac=Second mitochondrial activator of caspasesIAP=Inhibitor of Apoptosis Proteins
33

Bcl-2 family membersl A very large family with 19 members identifiedl Bcl-2 (homologous to ced-9) is prototypel All have the BH3 domain (Bcl-2 Homology)
l BH-3 is the pro-apoptotic domain exposed on activation
l Act as dimers=either hetero or homodimersl Pro-apoptotic dimers (Bax) increase mitochondrial
permeabilityl Anti-apoptotic members (Bcl-2, Bcl-XL) form dimers
with pro-apoptotic members to inactivate them 34

The Bcl-2 FamilyBH domains=protein-protein interaction domains
35

Intrinsic Apoptosis
36

Some trophic factors prevent apoptosis by inducing inactivation of a pro-apoptotic regulator
Figure 23-5037

Two mechanisms for p53 activation of apoptosis
38

Extrinsic/Death Receptor Pathway
39

Death Receptors and Ligands
CD95=Fas40

TNF receptor family
Death Domains (DD)Bind DDs of other proteins (e.g. FADD)…
…Recruiting them to the plasma membrane.
Cysteine-Rich Domains (CRD)
41

Fas-FasL Apoptosis
l In response to antigenic stimulation, peripheral T cells expand
l The antigen specific T cells generated must be eliminated (except for the memory cells)
l Upon repeated antigenic stimulation via the T Cell receptor: T cells upregulate Fas and FasL
l Eliminate neighboring T Cells expressing Fas42

Activation of Apoptosis by Fas Ligand
43

Fas Induced Apoptosis
The Formation of the Death Initiating Signal Complex (DISC)
44

Adaptor Proteins contain conserved protein interaction domains
= inhibits apoptosis
-CARD domain of Apaf-1 binds CARD domain of procaspase-9.-DED domain of FADD binds DED domain of procaspase-8.-DED domains of FLIP can bind to the DED domain of FADD and block procaspase-8 recruitment.
45

Fas and the intrinsic pathway:Bid
/Bax
46

Proteolytic targets of effector caspases
l Cytoskeletal regulatory proteinsl Actin
l Nuclear Laminsl Poly(ADP-ribose) polymerase (PARP)
l PARP activity depletes ATP, thus cleavage of PARP may maintain store of ATP to drive apoptosis
l DNA-fragmentation factor (DFF)
47

Removal of apoptotic cell by phagocytosis
48

Removal of cell corpses
49
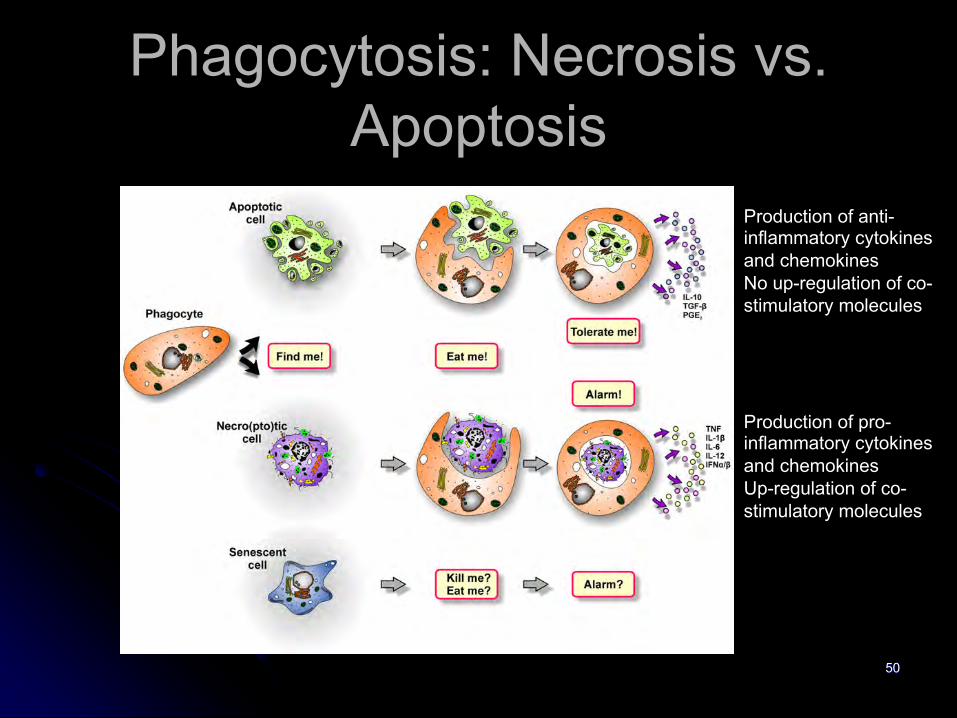
Phagocytosis: Necrosis vs. Apoptosis
50
Production of anti-inflammatory cytokines and chemokinesNo up-regulation of co-stimulatory molecules
Production of pro-inflammatory cytokines and chemokinesUp-regulation of co-stimulatory molecules

Phagocytosis tags and receptors
51

Two roads to activate apoptosisExtrinsic Intrinsic
52
Caspase 3, 6, 7
Apoptosis
Pro-apoptotic ligand
FADDFLIP
DR5
DR4
Cell-extrinsicpathway
Procaspase 8, 10
p53p53
Caspase 9
Caspase 8, 10
p53
BAX, BAK
Mitochondria
SMAC/DIABLO
ChemotherapyRadiotherapy
DNA damage
PUMA, NOXA
APAF1
Cytochrome c
DNA damage
BID
IAP
Cell-intrinsicpathway
BCL2, BCLXL, MCL1

TNFα Can also activate Necroptosis
53

Necroptosis: RIP3 Dependent programed necrosis
54

Autophagy
55
Two major functions:
o to eliminate damaged or superfluous proteins and organelles, commonly referred to as the protein and organelle quality control function of autophagy.
o to degrade and recycle intracellular components to sustain sustain metabolism and homeostasis in the absence of external nutrients, commonly referred to as the catabolic function of autophagy.
Autophagy, literally means ‘self-eating’.

The Autophagic Process
56

Autophagy under the microscope
57
Autophagic Vesicles LC3-II punctae

membrane recruitment of both PKB and its activatorPDK1 (phosphoinositide-dependent protein kinase 1),leading to the activation of PKB. PtdIns3K activity canbe opposed by PTEN, a 30-phosphoinositide phospha-tase, subsequently decreasing PKB activity, and inhibit-ing mTOR. PtdIns3K–PKB activation suppressesautophagy in mammalian cells. PKB further activatesmTORC1 through inhibiting a downstream protein com-plex, the tuberous sclerosis complex 1/2 (TSC1/TSC2).The TSC1/TSC2 heterodimer, which is a stable complex,senses the upstream inputs from various kinases, in-cluding PKB and ERK1/2 [35,36]. Phosphorylation ofTSC2 by PKB or ERK1/2 leads to the disruption of itscomplex with TSC1, and results in mTOR activation.
TSC1/TSC2 acts as the GTPase-activating protein forRheb, a small GTP-binding protein that binds to andactivates mTOR in its GTP-bound form. Ras has oppos-ing roles in autophagy regulation: it inhibits autophagy byactivating the PtdIns3K–PKB–mTORC1 pathway, andat the same time, it may induce autophagy via theRaf-1–MEK1/2–ERK1/2 pathway [37,38]. Finally, themTORC2 complex is also involved in autophagy regu-lation. Full activation of PKB requires mTORC2 [39],and inhibition of PKB, caused by mTORC2 depletion,reduces the phosphorylation of, and therefore activates,the forkhead box O (FoxO3) transcription factor, whichstimulates autophagy in muscle cells independent of theactivity of mTORC1 [40].
128 Cell regulation
Figure 2
Signaling cascades involved in the regulation of mammalian autophagy. Autophagy is regulated by a complex signaling network of various stimulatory(arrowheads) and inhibitory (bars) inputs. Activation of growth factor receptors stimulates the class I PtdIns3K complex and small GTPase Ras, whichleads to activation of the PtdIns3K–PKB–mTORC1 pathway and the Raf-1–MEK1/2–ERK1/2 pathway, respectively. PKB and ERK1/2 phosphorylateand inhibit the GTPase-activating protein complex TSC1/TSC2, leading to the stabilization of Rheb-GTPase, which, in turn, activates mTORC1,causing inhibition of autophagy. Activated ERK1/2 also stimulates autophagy. mTORC2 inhibits autophagy through the phosphorylation and activationof PKB. Metabolic stress, such as high AMP/ATP ratios resulting from energy depletion, or an increase in the cytosolic free Ca2+ concentration orcytokines, cause the AMP-activated protein kinase (AMPK) to be phosphorylated and activated by LKB1, CaMKKb, and TAK1, respectively. AMPKphosphorylates and activates TSC1/TSC2, leading to inactivation of mTORC1 and autophagy induction. Genotoxic and oncogenic stresses result innuclear p53 stabilization and activation, which stimulates autophagy through activation of AMPK or upregulation of DRAM. In contrast, cytosolic p53has an inhibitory effect on autophagy. Antiapoptotic proteins, Bcl-2 or Bcl-XL, associate with Beclin 1 and inhibit the Beclin 1-associated class IIIPtdIns3K complex, causing inhibition of autophagy. For additional details, see the main text.
Current Opinion in Cell Biology 2010, 22:124–131 www.sciencedirect.com
Signals Controlling Autophagy
Current Opinion in Cell Biology 2010, 22:124–131
58
Genotoxic and oncogenic stresses result in
nuclear p53 stabilization and activation, which
stimulates autophagy through activation of
AMPK
AMPK phosphorylates and activates the GAP
complex TSC1/TSC2, leading to inactivation of
mTORC1 and autophagy induction.
Rheb is a GTPase
AMPK->mTORC1 activates autophagy
PI3K-> AKT blocks autophagy

Autophagy in Cancer
59
Autophagy Can promote tumor formation or block it depending on context.
Clin Cancer Res; 21(3): 498

Questions?
60