antenatal phosphodiesterase 4 inhibition restores...
TRANSCRIPT

JPET#179085
1
Antenatal phosphodiesterase 4 inhibition restores
postnatal growth and pulmonary development in a model
of chorioamnionitis in rabbits
L. Homer , E Launay , N. Joram , C. Jacqueline, P.-H. Jarreau , J Caillon,T
Moyon, B Branger, G Potel, JC Roze, C. Méhats , and C. Gras-Le Guen
LH : CHU Brest, Service de Gynécologie Obstétrique et Médecine de la Reproduction,
Brest, F- 29200, France ;
CJ, JC, CGL : Université de Nantes, Nantes Atlantique Universités, Thérapeutiques
Cliniques et Expérimentales des Infections, EA 3826, Faculté de Médecine, Nantes F -
44000, France ;
TM : a
Institut National de la Recherche Agronomique and University of Nantes, UMR-1280
Physiologie des Adaptations Nutritionnelles, CHU Hôtel Dieu, Place Alexis Ricordeau, HNB1,
44093 NANTES CEDEX 1, France (EU).
BB: Perinatal network “Pays de la Loire”, Nantes University Hospital,Nantes, France
JCR : National Institute of Health and Medical Research CIC004, Nantes University
Hospital,Nantes, France
NJ, EL, CGL : CHU de Nantes, Service de Néonatalogie, Hôpital Mère Enfant, 38
boulevard Jean Monnet, France Nantes cedex F - 44093, France ;
CM : Institut National de la Santé et de la Recherche Médicale (INSERM) U767, Paris,
France ;
PHJ : Université Paris Descartes, Faculté de Médecine, Paris, France ;
PHJ : CHU Port-Royal, Service de Néonatologie, 123 Boulevard de Port-Royal, Paris F-
75679 Cedex 14, France ;
JPET Fast Forward. Published on December 7, 2011 as DOI:10.1124/jpet.111.179085
Copyright 2011 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
2
RUNNING TITLE PAGE
• Running title : antenatal PDEA inhibition improves pulmonary development
• Number of text page:20
• Number of tables: 3
• Number of figures: 4
• Number of references: 42
• Number of words:
• Abstract :242
• Introduction: 464
• Discussion: 1257
• Corresponding author:
Christèle GRAS – LE GUEN
Service de Néonatalogie - Hôpital Mère Enfant CHU de Nantes
38 boulevard Jean Monnet
Nantes cedex F - 44093, France
Tel : +33240083483 Fax : +33240083483
e-mail : [email protected]
• Liste of non standard abbreviations :
• PDE4: Phosphodiesterase 4
• BPD: Bronchopulmonary dysplasia
• BAL: Bronchoalveolar lavage fluid
• MIC : Minimum inhibitory concentration
• DMSO: Dimethyl sulfoxide • CFU : Colony forming units • HES: Hematoxylin-Eosin-Saffron
• Recommended section assignment: Gastrointestinal, Hepatic, Pulmonary and renal
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
3
ABSTRACT 242words
Chorioamnionitis is implicated in the pathophysiology of bronchopulmonary disease, and the
associated inflammatory response is responsible for adverse effects on alveolar development.
The aim of this work was to analyze the effects of a phosphodiesterase 4 (PDE4) selective
inhibitor, rolipram (a modulator of the inflammatory response), in an experimental model of
chorioamnionitis on pulmonary development and on the processes of infection and
inflammation. Rabbit mothers were assigned to four groups: i) saline serum inoculation
(controls), ii) E. coli intra-uterine inoculation (C+), iii) rolipram infusion (R+), and iv) E. coli
inoculation + rolipram infusion (C+R+). High rates of morbility and mortality were noticed in
mothers and pups (5/13 pregnant rabbits in groups with rolipram). Alveolar development,
inflammation and infection were analyzed in pups at day 0 and day 5. At day 0, in the context
of chorioamnionitis, rolipram significantly decreased birth weight (p<0.01) relative to
controls (p<0.05). At day 5, weight normalized in the C+R+ group but not in group C+
relative to controls (p<0.001); moreover, alveolar airspace volume was preserved in group
C+R+ but not in group C+ animals (p<0.05). Interstitial volume decreased in group C+
animals versus controls (p<0.05) but was preserved in group C+R+. Specific alveolar area
was not significantly modified by rolipram. No significant difference was found concerning
bronchoalveolar lavage cellularity, and all blood cultures remained sterile. In this model of
impaired alveologenesis, rolipram significantly preserved specific alveolar density. However,
PDE4 inhibition induced antenatal fetal demise and growth retardation.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
4
INTRODUCTION 464 words
Bronchopulmonary dysplasia (BPD) remains a major concern in very premature
infants despite recent advances in perinatal care. BPD is characterized by the arrest of
secondary septation, previously described as an alveolar number decrease and an alveolar
enlargement, and is associated with thinned septa and minimal capillary development
(Kinsella et al; 2006). Alveolarization occurs postnatally in humans and premature, in utero
exogenous disturbances of immature lungs are thought to induce BPD through inflammatory
processes of the airway (Coalson JJ et al; 1999). Main aetiologies are the prematurity itself,
and also perinatal care such as hyperoxia and mechanical ventilation (Kraybill EN et al;
1989). Relationships between BPD and inflammation were established by Ogden et al., who
found increased inflammatory cell counts in bronchoalveolar lavage fluid (BAL) of neonates
exposed to hyperoxia or mechanical ventilation who subsequently developed BPD (Ogden BE
et al; 1983). Chorioamnionitis and neonatal infections have been implicated in the
development of pulmonary defects. Actually, correlation between inflammation, premature
delivery, and BPD has been demonstrated in humans (Kallapur SG et al; 2001) and animals
(Kallapur SG et al; 2004). Yoon, et al. have shown that high levels of IL-6, IL-1ß, IL-8, and
TNF-α found in amniotic fluid as early as 5 days before delivery was predictive of BPD in
premature neonates (Yoon BH et al; 1997). This observation was confirmed by Gomez et al.,
who showed that fetal morbidity correlated with an inflammatory response as a result of high
concentrations of IL-6 in fetal plasma, even in the case of asymptomatic chorioamnionitis
(Gomez R et al;1998).
As modulators of the inflammatory response, phosphodiesterase (PDE) inhibitors have
been widely studied in chronic pulmonary inflammation deseases (Hatzelmann A et al; 2010;
Giembycz MA; 2006). Among the 11 families of PDEs, selective inhibition of PDE4
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
5
increases intracellular cAMP levels, particularly in inflammatory cells (T and B lymphocytes,
polymorphonuclear neutrophils), respiratory epithelial cells, and endothelial cells (Torphy TJ;
1998), thus modulating cellular trafficking and cytokine and chemokine responses (Bender
AT and Beavo JA; 2006; Sanz MJ et al; 2005). Moreover, inhibition of PDE4 limits
interstitial fibrosis and enhances muscular relaxation in airways (Kohyama T et al; 2002).
Recently, selective PDE4 inhibition was tested in a common murine postnatal hyperoxia
model of BPD. While de Visser, et al. found that PDE4 inhibition prolonged median survival
and reduced alveolar fibrin deposition and lung inflammation (de Visser YP et al; 2008),
Mehats et al. observed diminished weight gain without restoration of impaired alveolarization
induced by hyperoxia (Mehats C et al; 2008).
The aim of this study was to test the antenatal effects of the prototypical PDE4
inhibitor, rolipram, in a previously described model of chorioamnionitis-induced BPD in the
rabbit (Gras-Le Guen C et al; 2008). Pulmonary development was assessed by lung
morphometry. Inflammation and infection were determined by cellularity and culture of blood
and BAL.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
6
METHODS
Antenatal infection
Pregnant and newborn New Zealand rabbits were used in the study. The RS 218 (O18:K1:H7)
strain of Escherichia coli K1 was used and has been characterized for virulence factors and
invasion genes (Huang SH et al; 1995). The minimum inhibitory concentration (MIC) of this
strain for ceftriaxone was 0.06 mg/L. The intra-uterine inoculation protocol, as previously
described (Gras-Le Guen C et al; 2003), was approved by the Animal Care Committee of the
University of Nantes experimental therapy department (Nantes, France). A total of 25 female
New Zealand white rabbits weighing 3.7–4.5 kg (CEGAV, Saint Marc d’Egrenne, France)
were obtained 10 days before the end of their usual 31- to 32-day gestation period and housed
at the Nantes School of Medicine (Nantes, France) in appropriate cages with a special place
for nidation. The 25 female rabbits were provided water and food ad libitum consisting of
antibiotic-free granules. Animals were randomly assigned to four groups: i) controls (saline
serum inoculation), ii) C+ (E.coli intra uterine inoculation), iii) R+ (saline serum inoculation
+ rolipram infusion), iv) C+R+ (E.coli inoculation + rolipram infusion). Pregnant rabbits in
all groups (n = 25 for both groups) were treated with 200 mg/kg/day intravenous ceftriaxone
(Rocephine®; Roche, Meylan, France) via a catheter inserted into a marginal vein of the ear.
Treatment began 6 h after intrauterine inoculation and lasted 4 days, until spontaneous
delivery. Pregnant rabbits in all groups received dimethyl sulfoxide (DMSO), the rolipram
diluent. DMSO was injected alone (0.02 mg + 20 mL saline serum) or with rolipram (R6520,
Sigma-Aldrich, France), according to the experimental group. Posology of rolipram was 0.250
mg/kg in our study. Suspension of rolipram was done at 7.3 mg/mL in DMSO. The rolipram
treatment was started as the antibiotic treatment 6 h after intrauterine inoculation and lasted 4
days, until spontaneous delivery. For administration, rolipram was infused with 20mL of
0.9% saline serum during a 60 min continuous infusion. On day 28 of gestation, the animals
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
7
were anesthetized with PROPOFOL LIPURO® 1% (B BRAUN PHARMA) and ketamine (25
mg/kg). Laparotomy was then performed via a 2-cm vertical midline incision below the
gravid uterus. After exposing the uterus, a single injection of E. coli (1×103 colony forming
units (CFU) in 1 mL of 0.9% NaCl solution) or sterile vehicle was performed under visual
control into one of the uterine horns facing a fetoplacental unit. Slight aspiration was
performed before injection to verify correct intra-amniotic positioning of the needle. The
incision was then closed in layers and the animals were returned to their cages.
After spontaneous birth, rabbit pups were suckled until euthanasia. They were
exsanguinated at birth (day 0) or on day 5, under deep anesthesia with intraperitoneal
Pentothal. Blood was obtained by cardiac puncture after animals failed to blink an eyelid or
move a leg muscle following local stimulation. Five to ten rabbit pups per group were used
for either bronchoalveolar lavage or morphometric study.
Assessment of lung morphometry
Lungs were gently extracted and fixed with 4% paraformaldehyde through a polyethylene
tracheal cannula at a constant pressure of 30 cm water for 10 minutes. The trachea was then
ligated, and the lung was immersed in 4% paraformaldehyde for 24 h. Lung volumes were
measured by the displacement method in the fixative solution (Scherle W, 1970). Both lungs
of each rabbit were paraffin-embedded and 4-m frontal sections were cut in the medial part of
the lungs, and stained with Hematoxylin-Eosin-Saffron (HES). All lung lobes were used. All
morphometric evaluations were performed by one observer (L.H.) blinded to group
assignment. A microscope (Leitz, Weszlar, Germany) connected to a television screen by a
video camera (Sony, Tokyo, Japan) was used. Volume densities of pulmonary parenchymal
structures (alveolar airspace, airways, blood vessels larger than 20 m in diameter, and
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
8
interstitial tissues), and alveolar surface density were measured using the point counting and
mean linear intercept methods, as previously described (Weibel ER, 1997). Lightmicroscope
fields were quantified at an overall magnification of 400×, using a 42point, 21-line eyepiece
graticule placed on the television screen. Twenty fields (10 per lung) were evaluated per
animal by a systematic sampling method from a random starting point. To correct for
shrinkage associated with fixation and paraffin processing, area values were multiplied by
1.22, a factor calculated during a previous evaluation (Franco ML et al; 2002). All
morphometric data were expressed as relative and absolute values, as described by Burri and
coworkers (Burri PH et al; 1974). Relative values (volume density or surface density) were
those obtained directly from morphometric measurements of tissue sections. Absolute values
(total volume or surface area per lung) were determined by multiplying the relative values by
lung volume.
Evaluation of cell proliferation
Cell proliferation was assessed at day 0 by immunohistochemistry, using a monoclonal mouse
antibody against rat proliferating cell nuclear antigen as primary antibody, diluated 1:1500
(Mouse anti-rat PCNA (Clone PC-10), DakoCytomation, Ca# M0879). Peroxidase-linked
goat anti-mouse immunoglobulin (Santa Cruz Biotechnology, Cat# SC 2005) was used as
secondary antibody, diluted 1:1500 (Horseradish Peroxydase, HRP). The alveolar PCNA
index was determined by counting the immunostained nuclei per alveoli in 15 fields of
alveolar parenchyma per animal using a Quantimet Q550 (Leica Microsystem, Cambridge,
UK) (Namyslowski G et al; 2004) .
Evaluation of cell death
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
9
Cell death was assessed at day 0 using the DNA specific dye Hoechst 33258 (Calbiochem,
Merck Biosciences, VWR, Fontenay-sous-Bois, France), at 1 mg/mL in Hank’s balanced salt
solution without phenol red on formalin-fixed paraffin sections. Sections were mounted with
Prolong Antifade medium (Molecular Probes, Eugene, OR, USA). Fluorescence was observed
with a fluorescence microscope (Axiovert 200-M; Carl Zeiss, Göttingen, Germany) equipped
with an ApoTome slider, which eliminates image blurring. Cells were visualized with a
63/1.4× oil immersion lens. Image processing was performed using an AxioCam MR CCD
camera and AxioVision 4.0 software (Carl Zeiss, Göttingen, Germany).
Evaluation of lung elastic fiber content
To evaluate lung parenchyma elastic fiber length content at day 5, lungs sections were stained
by the Weigert technique. The standard morphometric methods described above were then
used [i.e., counting the number of points touching alveolar parenchyma (point counting) and
the number of intersections between the graticule lines and the elastic fibers (mean linear
intercept)]. (Weibel ER, 1997)
Assessment of vascularization
Vascularization was assessed at day 5 by immunohistochemistry using a monoclonal antibody
against human CD31 (Clone JC/70A, DakoCytomation, Ca#M0823) as primary antibody,
diluted 1:50. The vascular development index was determined by counting the immunostained
nuclei per alveoli in 25 fields of alveolar parenchyma per animal, using a Quantimet Q550
(Leica Microsystem).
Assessment of total and specific PDE activity in pup lungs at day 0
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
10
Whole lung tissues were homogenized in ice-cold hypotonic buffer, (100 mM Tris-HCl pH
7.4, 2 mM MgSO4, 2 mM EDTA, 50% glycerol, and 1 mM 2-β-mercaptoethanol
supplemented with P2714 protease inhibitor cocktail, Sigma) using an all-glass homogenizer.
Aliquots of homogenates were assayed for cAMP-PDE activity according to the method of
Thompson and Appleman (Thompson WJ et Appelman MM; 1971). PDE activities were
measured with 1 µM cAMP as substrate (GE Healthcare, Amersham, Buckinghamshire, UK).
PDE4 activity was defined as the fraction of cAMP PDE activity inhibited by 10 µM rolipram
as described previously (Schmitz T et al; 2007). Protein concentrations were determined using
the protein assay kit from Bio-Rad (Bio-Rad Laboratories, Hercules, CA, USA) with BSA as
standard.
Assessment of inflammation: Broncho-alveolar lavage (BAL)
BAL was performed as previously described (Franco ML et al; 2002). Rabbits were
exsanguinated and the thorax was opened to expose the lungs and trachea. A short length of
tubing was inserted into the trachea and ligated, and then lungs were immediately lavaged.
BAL was performed at birth and at day 5 using a total of 1 mL sterile saline. BAL fluid was
centrifuged and total and differential cell counts were performed. Cytodifferentiation was
performed on cytospin preparations stained with Diff-Quick (Scientific Products, McGraw
Park, IN, USA).
Bacteriological analysis
Blood cultures
At the time of sacrifice, blood was withdrawn from the hearts of mothers and pups for
quantitative cultures; 1-mL blood samples were mixed with 900 mL of heparinized (5,000
IU/L) physiological serum. After centrifugation, 100 mL from each pellet were plated on MH
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
11
agar (Difco; Becton Dickinson, Franklin Lakes, NJ, USA) and incubated at 37°C for 24 h
before performing bacterial counts. This method allows detection of an inoculum of 102
CFU/mL. (Gras-Le Guen C et al; 2008)
Pulmonary cultures
At the time of sacrifice, lung samples were homogenized in 100 µL of sterile saline solution
and diluted to 10-2 and 10-4 to prevent the possibility of carry-over. Next, 50 mL of each
dilution was spread on EMB plates using a spiral system plater (Interscience, BV, Louvain,
Belgium). After 48 h of incubation at 37°C, viable bacteria were counted.
Statistical analysis
Statistical analyses were performed with SPSS Software (version 15.0, SPSS Inc.
Chicago, IL USA). Multiple group comparisons were performed using a 2X2 factor ANOVA
analysis. Survival was evaluated by Kaplan-Meier survival function and the logrank test. A p-
value < 0.05 was considered to be statistically significant.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
12
RESULTS
Assessment of survival and growth at day 0 and day 5
Survival. Five pregnant rabbits died between inoculation (day 28 of gestation) and
delivery, three in group C+R+ (days 29, 29 and 31) and two in group R+ (both at day 31);
however, mortality rates did not significantly differ between the four groups. No mother died
after delivery.
Among pups, intrauterine death was significantly increased in groups C+R+ and R+
compared to controls (p<0.0001) and group C+ (p<0.03), and postnatal mortality was
significantly increased in groups C+ (p<0.03) and C+R+ (p<0.04) relative to Controls (Figure
1). There were significant differences in survival of live born pups at day 5 between controls
(97±2%) and groups C+, C+R+ and R+ (31±6, 22±8 and 40±5%, respectively, p<0.0001, by
Logrank)
Body-weight gain. (Table 1). Animals in Rolipram groups (R+ and C+R+) and
infection groups had significantly lower weight at birth. Five days after birth, infection and
Rolipram had no significant effect, but we observed a strong interaction between these two
factors (F1/22=44,6, p=0,000001). Weights were preserved between control animals and
C+R+ animals
All maternal and fetal blood and tissue cultures remained sterile. We previously demonstrated
in the same experimental model early fetal infection 8 h after bacterial challenge with 5 (29%)
out of 17 fetal blood positive cultures and 15 (88%) out of 17 placental positive cultures with
a mean quantitative value of 4.5±1.4 log10CFU.g-1 of tissue) (Gras-Le Guen C et al; 2008)
Lung morphometry at day 0 and day 5
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
13
Examination of postnatal lung sections under light microscopy at day 5 showed that infected
rabbit lungs exhibited a diffuse and simplified lung structure with enlarged air spaces and
fewer secondary septa, whereas infected rabbit treated antenatally with rolipram exhibited an
alveolar morphology similar to control animals . Quantification by morphometric analysis
highlighted the following features (Tables 1 and 2):
-lung volume: At day 0, no difference was found between the four groups for absolute
lung volumes but we noticed a significant effect of Rolipram on specific lung volume
(F1/22 = 7,1; p=0,01). At day 5, absolute lung volume was significantly reduced in
infection groups (F1/21 = 7,6; p=0,01) with a strong interaction between infection and
rolipram effects (F1/21 = 52,3; p=0,000001), but we did not onserve any effect of
infection or Rolipram on specific lung volume.
-alveolar airspace (Vva): At day 0, we observed a significant effect of Rolipram on all
alveolar airspace parameters (volume density, absolute volume, specific volume).
Moreover, we noticed a strong interaction effect on volume density (F1/22 = 16,0;
p=0,0005). At day 5, we reported some significant interaction effects on all alveolar
airspace parameters. Rolipram seemed to have the same deleterious effect than
infection on alveolar airspace, but preserved alveolar volume when used in infected
animals.
-alveolar surface (Sva): At day 0, we did not observe any significant effect of
infection nor Rolipram on alveolar surface. At day 5, we reported a significant effect
of Rolipram on all alveolar surface parameters (area density, absolute area and specific
area).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
14
-interstitium (Vvi): At day 0, specific volume was significantly increased in Rolipram
groups (C+R+ and R+). At day 5, we observed an interaction effect on relative volume
density (F1/21 = 6,1; p=0,02).
-airways: At day 0, specific volume was significantly increased in Rolipram group,
and at day 5, we observed an interaction effect on absolute volume (F1/21 = 10,9;
p=0,003).
Blood vessels: At birth, specific volume was significantly increased in Rolipram group, with
an interaction effect on the 3 parameters. At day 5, we observed the same interaction effects
on the blood vessels. Once again, morphometric parameters were decreased in group C+ but
maintained in group C+R+ (table 2). Moreover, microvascularization studied by
immunohistochemistry labeling was significantly altered in all groups as compared to controls
(Figure 2A).
Evaluation of lung elastic fiber content
Morphological analysis of elastic fibers at day 5 showed a significant decrease in the
number of alveolar fibers in the pup’ lungs of groups C+, C+R+ and R+ versus controls [8±4,
16±5, 4±2, vs. 40±25, respectively, (p<0.05)]. The number of alveolar fibers tended to be less
decreased in group C+R+ than in groups C+ and R+, but the difference was not significant
(Figure 2B).
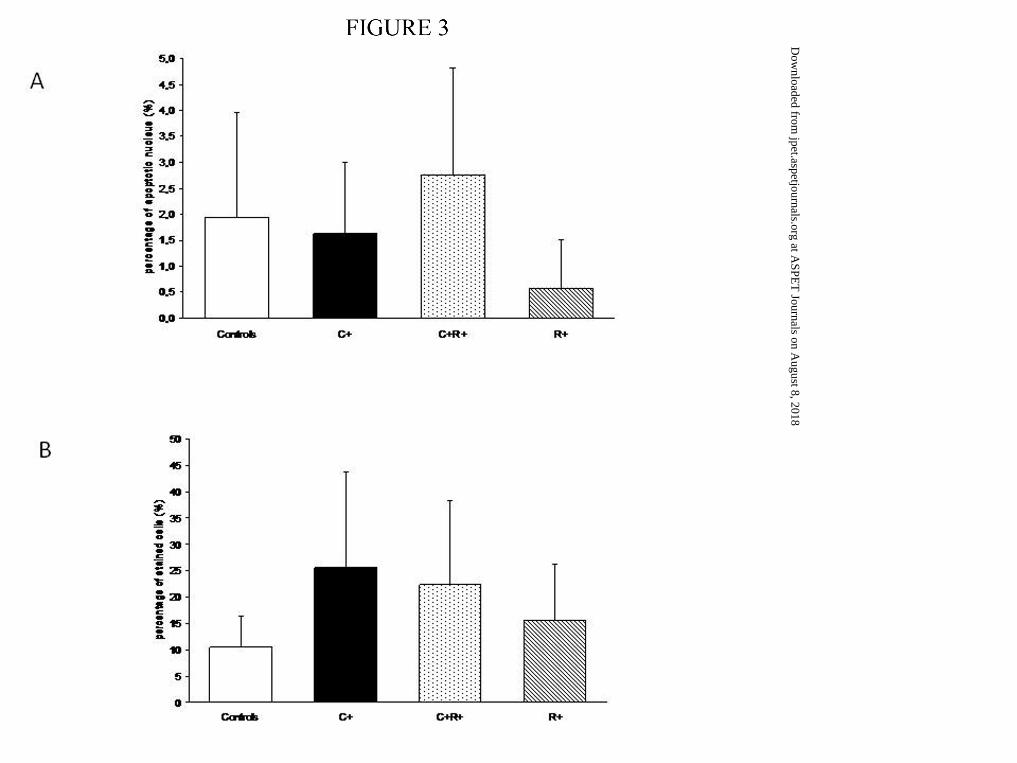
Assessment of apoptosis and cell proliferation
Apoptosis was present in the controls, and the group C+ parameters were not
associated with an increase in cell apoptosis (Figure 3A). A significant decrease was noticed
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
15
in group R+ compared to both controls and C+R+ (p<0.001). For cell proliferation, an
increase was noticed in the C+ group relative to controls (p<0.05; Figure 3B).
Assessment of inflammation in BAL fluid at day 0 and day 5 and PDE4 activity in lung
at day 0
Mononuclear cell count in BAL fluid. No difference was found in the inflammatory
cell count in BAL fluid, either at day 0 or at day 5. However, group C+R+ animals showed a
trend, although not significant, to an increase of mononuclear inflow (Table 3).
PDE4 activity in lung. A global and equivalent AMPc-phosphodiesterase activity was
noticed in all conditions at day 0. Mean PDE4 specific activity represented 48.7±5.1% of the
global AMPc-phosphodiesterase activity expressed in the pups’ lung for all conditions, and
did not change subsequent to the experimental condition (Figure 4).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
16
DISCUSSION 1257 words
In the present study, we report that inhibition of PDE4s by rolipram in rabbit pups exposed to
chorioamnionitis preserved ante and postnatal alveolarization, without modifying the
inflammatory response. However, we observed marked intrauterine growth retardation and a
very high incidence of stillbirth in animals treated with rolipram, results not yet reported in
this model. Rolipram is the prototypical PDE4 selective inhibitor. PDEA 4 enzyme is the
main cAMP metabolising enzyme in immune and inflammatory cells, airway smooth muscle,
and pulmonary nerves; its inhibition suppresses the recruitment and activation of several
inflammatory cells (neutrophils, CD8 T cells, and macrophages) known to have a crucial role
in the pathophysiological processes of bronchopulmonary dysplasia (Hayes D et al; 2010;
Sanz MJ et al; 2005). In this context, we chose to test this new treatment in a previously
described model of antenatal infection with subsequent impaired alveolarization in the rabbit
(Gras-Le Guen C et al; 2008). The detection of AMPc-phosphodiesterase activity in our study
confirmed that PDE4s are expressed in pups’ lungs. The very short half-life of the molecule (
1.9 ±0.6 hour in the rabbit when using the 0.2 mg/kg dose) can probably explain the absence
of significant changes in the level of expression between the different groups, because the
assays were possible only 6 to 12 hours after birth, several hours after the last injection
(Krause W et al, 1988). However, we can observe a tendency to a decreased expression in the
infected + rolipram group (C+R+) when compared to the infected group (C+). The small
posology that we used (0.25 mg/kg/d) is possibly responsible for this lack of difference, but
was imposed by the poor tolerance to higher doses observed in this model. However, these
results confirm that PDE4 is a potential target in fetal and neonatal lungs.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
17
Antenatal infusion of selective PDE4s inhibitor preserves alveologenesis in the
context of chorioamnionitis. We observed conservation of specific interstitial, vascular,
airways and alveolar airspace volumes at day 0, and persistent effects at day 5 with some
significant interaction effects on alveolar airspace, interstitium, airways and vascular
morphometric parameters. Rolipram effects seem clearly different in infected or in non-
infected fetuses and newborns. These observations are consistent with the results of Woyda et
al., using a different PDE4 inhibitor (cilomast) in a different experimental model of hyperoxia
alveologenesis impairment in the mice (Woyda K et al; 2009). However, the lack of a control
group and the quality of morphometric determinations made the interpretation of the exact
effects of PDE4 inhibition on disturbed alveolarization difficult (Mehats C et al; 2009).
Mehats et al., in the same model in rat indicated that PDE4 inhibition with rolipram did not
enhance alveolarization in rat pups exposed to hyperoxia, and may directly affect
alveolarization in rat pups exposed to room air (Méhats C et al; 2008). The authors suggested
that this last point could be explained by the profound effect of PDE4 inhibition on pups’
weight gain that could interfere with normal alveolarization as much as antenatal infection
interfere (Gras-Le Guen C et al, 2008). In this infectious and inflammatory context, benefic
effects of PDE4 inhibition observed on alveologenesis in infected pups is possibly mediated
by some specific anti-inflammatory mechanisms whereas poor weight gain and poor
alveolarization both could reflect poor animals nutrition.
Rolipram infusion was also concomitant with intrauterine growth restriction. In fact,
this experimental model of antenatal infection has previously been associated with neonatal
growth alteration, but only with a postnatal onset (Gras-Le Guen C et al; 2008). In this present
work, antenatal growth restriction is noticed at birth in the two groups exposed to rolipram
versus the infected group (C+) and controls. However, postnatal growth restriction occurred
as early as 5 days after birth in the infected group (C+) but not in group C+R+ animals, where
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
18
rolipram seemed to have preserved postnatal growth. However, de Visser et al. and Mehats et
al. observed a weight loss in rat pups exposed to rolipram in an experimental model of
hyperoxyia-altered alveolarization (de Visser YP et al; 2008; Méhats C et al; 2008). In
Humans, Fabbri et al. observed weight loss in the group of patients with chronic obstructive
pulmonary diseases treated with roflumilast compared to a control group (Fabbri LM et
al;2009 ). To explain the weight loss, one hypothesis was that pups treated with rolipram were
underfed because of side effects limiting the pregnant rabbits’ feeding. Rolipram is known to
have adverse effects on the central nervous system that accounts for nausea, vomiting, and
enhanced gastric acid secretions (Zeller E et al; 1984; Barnette MS et al; 1995). However,
mother rabbits did not lose weight during the delay from inoculation through delivery.
Secondly, rolipram on its own increases lipolysis as described in an in vitro model of rat pups
(Nakamura J et al; 2004; Snyder PB et al; 2005). In addition, toxicological reports during
preclinical studies demonstrated significant inflammation of the intestinal tract, suggesting
pathological absorption of food (Dagues N et al; 2007; Larson JL et al; 1996). This
mechanism would better explain the differences noticed between the C+ and control groups
and group C+ versus groups C+R and R+ at birth, because animals treated with rolipram
made up for the weight restriction as soon as rolipram infusion was stopped, and group C+R+
recovered normal weight at day 5 compared to controls.
Despite the fact that rolipram showed some protective effects on alveologenesis in the
chorioamnionitis model, high rates of morbidity and mortality were noticed in mothers and
pups. Such a mortality rate has not been reported with pregnant rabbits in this experimental
model –Gras-Le Guen C et al; 2008). Hemodynamic effets have been described on
anesthetized and ventilated animals (decrease in pulmonary arterial pressure and systemic
arterial pressure, increase in cardiac output ), without possibility to assess morbo-mortality
during these veru short-term experiences (Schermuly RT et al; 1999). Cardiac toxicity has
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
19
been described in rabbits by Shahid and Rodger, who showed an in vitro increase in the
intracellular concentration of cAMP that led to chronotropic effects on atrial muscle fibers
(Shahid M and Rodger IW; 1989). Nevertheless, this effect did not induce cardiac arrhythmias
(Vaughan Williams EM; 1987). PDE4 inhibitors have been used in humans with a well-
described adverse event profile, and such severe or fatal effects have not been reported
(Fabbri LM et al; 2009). The fatal adverse events in the rabbit are probably specific to this
experimental model and constitute a limitation, as previously observed with corticotherapy
(Pratt L et al; 1999). In utero death of pups in groups C+R+ (52.1%) and R+ (46.8%) differed
significantly from both controls (0.7%) and group C+ (18.9%), and the increase in mortality
in group C+R+ compared to group C+ was probably related to the cumulative deleterious
effects of chorioamnionitis and rolipram infusion. We hypothesized that the tocolytic effect of
rolipram described by Schmitz et al. may have played a role in this high rate of in utero
mortality (Schmitz T et al; 2007); however, inoculation-delivery delay did not differ between
the four groups. We also suggested that peripartum maternal behavior associated with
maternal neglect could be implicated in the mortality of the pups post-delivery as an indirect
consequence of rolipram. Interestingly, de Visser et al. and Mehats et al. observed a
prolonged median survival in rat pups treated with rolipram only after birth (de Visser et al;
2008; Mehats C et al; 2008). It seems clear that rolipram is implicated in the maternal and
neonatal deaths in these studies, but further studies are needed to understand the mechanism
involved in these severe adverse events.
PDE4 inhibitors appear as a promising new class of anti-inflammatory drugs that have
shown efficacy in the experimental model of chorioamnionitis-induced bronchopulmonary
dysplasia described here. However, the conflicting results and side effects observed in
different experimental models tempered the enthusiasm for this treatment. Further
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
20
investigations are needed in several different models before conclusions can be made
concerning the benefit/risk ratio of this treatment.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
21
ACKNOWLEDGEMENTS
The authors thank Mr. Arnaud LEGRAND, from his very helpful advice and suggestions
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
22
AUTHORSHIP CONTRIBUTION
Participated in research design: LH, JHJ, GP, CGL
Conducted experiments: LH, CJ, CM, CGL
Contributed analytic tools: LH, CM, JC, PHJ, CGL
Performed data analysis: TM, BB, JCR, LH, CJ, CGL
Wrote or contributed to the writing of the manuscript: LH, NJ, EL, CM, GP, CGL
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
23
REFERENCES
Barnette MS, Grous M, Cieslinski LB, Burman M, Christensen SB, Torphy TJ. J
(1995) Inhibitors of phosphodiesterase IV (PDE IV) increase acid secretion in rabbit
isolated gastric glands: correlation between function and interaction with a high-affinity
rolipram binding site. Pharmacol Exp Ther. 273:1396-402.
Bender AT, and Beavo JA (2006) Cyclic nucleotide phosphodiesterases: molecular
regulation to clinical use. Pharmacological reviews 58: 488-520.
Burri PH, Dbaly J, and Weibel ER (1974) The postnatal growth of the rat lung. I.
Morphometry. The Anatomical record 178: 711-730.
Coalson JJ, Winter VT, Siler-Khodr T, and Yoder BA (1999) Neonatal chronic lung
disease in extremely immature baboons. American journal of respiratory and critical care
medicine 160: 1333-1346.
Daguès N, Pawlowski V, Sobry C, Hanton G, Borde F, Soler S, Freslon JL, Chevalier
S (2007) Investigation of the molecular mechanisms preceding PDE4 inhibitor-induced
vasculopathy in rats: tissue inhibitor of metalloproteinase 1, a potential predictive
biomarker. Toxicol Sci 100 : 238-47.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
24
de Visser YP, Walther FJ, Laghmani EH, van Wijngaarden S, Nieuwland K, and
Wagenaar GT (2008) Phosphodiesterase-4 inhibition attenuates pulmonary inflammation
in neonatal lung injury. Eur Respir J 31: 633-644.
Fabbri LM, Calverley PM, Izquierdo-Alonso JL, Bundschuh DS, Brose M, Martinez
FJ, Rabe KF; M2-127 and M2-128 study groups (2009) Roflumilast in moderate-to-severe
chronic obstructive pulmonary disease treated with longacting bronchodilators: two
randomised clinical trials. Lancet 374 :695-703.
Franco ML, Waszak P, Banalec G, Levame M, Lafuma C, Harf A, and Delacourt C
(2002) LPS-induced lung injury in neonatal rats: changes in gelatinase activities and
consequences on lung growth. Am J Physiol Lung Cell Mol Physiol 282: L491-500.
Giembycz MA (2006) An update and appraisal of the cilomilast Phase III clinical
development programme for chronic obstructive pulmonary disease. British journal of
clinical pharmacology 62: 138-152.
Gomez R, Romero R, Ghezzi F, Yoon BH, Mazor M, and Berry SM (1998) The fetal
inflammatory response syndrome. Am J Obstet Gynecol 179: 194-202.
Gras-Le Guen C, Debillon T, Toquet C, Jarry A, Winer N, Jacqueline C, Kergueris
MF, Bingen E, Roze JC, Potel G, and Bugnon D (2003) Persistent bacteremia in rabbit
fetuses despite maternal antibiotic therapy in a novel intrauterine-infection model.
Antimicrobial agents and chemotherapy 47: 2125-2130.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
25
Gras-Le Guen C, Denis C, Franco-Montoya ML, Jarry A, Delacourt C, Potel G,
Bourbon J, Roze JC, and Jarreau PH (2008) Antenatal infection in the rabbit impairs post-
natal growth and lung alveolarisation. Eur Respir J 32: 1520-1528.
Hatzelmann A, Morcillo EJ, Lungarella G, Adnot S, Sanjar S, Beume R, Schudt C,
Tenor H (2010) The preclinical pharmacology of roflumilast--a selective, oral
phosphodiesterase 4 inhibitor in development for chronic obstructive pulmonary disease.
Pulm Pharmacol Ther 23 :235-56.
Hayes D Jr, Feola DJ, Murphy BS, Shook LA, Ballard HO (2010) Pathogenesis of
bronchopulmonary dysplasia. Respiration 79:425-36.
Huang SH, Wass C, Fu Q, Prasadarao NV, Stins M, Kim KS (1995) Escherichia coli
invasion of brain microvascular endothelial cells in vitro and in vivo: molecular cloning
and characterization of invasion gene ibe10. Infection and immunity 63: 4470-4475.
Kallapur SG, Bachurski CJ, Le Cras TD, Joshi SN, Ikegami M, and Jobe AH (2004)
Vascular changes after intra-amniotic endotoxin in preterm lamb lungs. Am J Physiol
Lung Cell Mol Physiol : 287: L1178-1185.
Kallapur SG, Willet KE, Jobe AH, Ikegami M, and Bachurski CJ (2001) Intra-
amniotic endotoxin: chorioamnionitis precedes lung maturation in preterm lambs. Am J
Physiol Lung Cell Mol Physiol 280: L527-536.
Kambayashi T, Jacob CO, Zhou D, Mazurek N, Fong M, and Strassmann G (1995)
Cyclic nucleotide phosphodiesterase type IV participates in the regulation of IL-10 and in
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
26
the subsequent inhibition of TNF-alpha and IL-6 release by endotoxin-stimulated
macrophages. J Immunol 155: 4909-4916.
Kinsella JP, Greenough A, and Abman SH (2006) Bronchopulmonary dysplasia.
Lancet 367: 1421-1431.
Kohyama T, Liu X, Wen FQ, Zhu YK, Wang H, Kim HJ, Takizawa H, Cieslinski LB,
Barnette MS, and Rennard SI (2002) PDE4 inhibitors attenuate fibroblast chemotaxis and
contraction of native collagen gels. American journal of respiratory cell and molecular
biology 26: 694-701.
Krause W, and Kühne G (1988) Pharmacokinetics of rolipram in the rhesus and
cynomolgus monkeys, the rat and the rabbit. Studies on species differences. Xenobiotica
18:561-71.
Kraybill EN, Runyan DK, Bose CL, and Khan JH (1989) Risk factors for chronic lung
disease in infants with birth weights of 751 to 1000 grams. J Pediatr 115: 115-120.
Larson JL, Pino MV, Geiger LE, Simeone CR (1996) The toxicity of repeated
exposures to rolipram, a type IV phosphodiesterase inhibitor, in rats. Pharmacol
Toxicol.78:44-9
Mehats C, Franco-Montoya ML, Boucherat O, Lopez E, Schmitz T, Zana E, Evain-
Brion D, Bourbon J, Delacourt C, and Jarreau PH (2008) Effects of phosphodiesterase 4
inhibition on alveolarization and hyperoxia toxicity in newborn rats. PloS one 3: e3445.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
27
Méhats C, Bourbon J, Jarreau PH (2009) Does PDE4 inhibition improve
alveolarisation in hyperoxia-exposed immature rodents? Eur Respir J 33 :1236;
Nakamura J, Okamura N, and Kawakami Y (2004) Augmentation of lipolysis in
adipocytes from fed rats, but not from starved rats, by inhibition of rolipram-sensitive
phosphodiesterase 4. Arch Biochem Biophys 425: 106-114.
Namyslowski G, Scierski W, Nozynski JK, and Zembala-Nozynska E (2004)
Morphometric characteristics of cell nuclei of the precancerous lesions and laryngeal
cancer. Med Sci Monit 10: CR241-245.
Ogden BE, Murphy S, Saunders GC, and Johnson JD (1983) Lung lavage of newborns
with respiratory distress syndrome. Prolonged neutrophil influx is associated with
bronchopulmonary dysplasia. Chest 83: 31S-33S.
Pratt L, Magnes RR, Phernetton T, and Bird IM (1999) Repeated use of
betamethasone in rabbits: Effects of treatment variation on adrenal suppression,
pulmonary maturation, and pregnancy outcome. Am J Obstet Gynecol;180:995-1005
Sanz MJ, Cortijo J, and Morcillo EJ (2005) PDE4 inhibitors as new anti-inflammatory
drugs: effects on cell trafficking and cell adhesion molecules expression. Pharmacol Ther
106: 269-297.
Scherle W (1970) A simple method for volumetry of organs in quantitative stereology.
Mikroskopie: 26: 57-60.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
28
Schermuly RT, Ghofrani HA, Enke B, Weissmann N, Grimminger F, Seeger W,
Schudt C; and Walmrath D (1999) Low-dose systemic phosphodiesterase inhibitors
amplify the pulmonary vasodilatory response to inhaled prostacyclin in experimental
pulmonary hypertension. Am J Respir Crit Care Med 160:1500-6.
Schmitz T, Souil E, Herve R, Nicco C, Batteux F, Germain G, Cabrol D, Evain-Brion
D, Leroy MJ, and Mehats C (2007) PDE4 inhibition prevents preterm delivery induced by
an intrauterine inflammation. J Immunol 178: 1115-1121.
Shahid M, and Rodger IW (1989) Chronotropic and inotropic actions of amrinone,
carbazeran and isobutylmethyl xanthine: role of phosphodiesterase inhibition. British
journal of pharmacology 98: 291-301.
Snyder PB, Esselstyn JM, Loughney K, Wolda SL, and Florio VA (2005) The role of
cyclic nucleotide phosphodiesterases in the regulation of adipocyte lipolysis. J Lipid Res
46: 494-503.
Thompson WJ, and Appleman MM (1971) Characterization of cyclic nucleotide
phosphodiesterases of rat tissues. The Journal of biological chemistry 246: 3145-3150.
Torphy TJ (1998) Phosphodiesterase isozymes: molecular targets for novel antiasthma
agents. American journal of respiratory and critical care medicine 157: 351-370.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
29
Vaughan Williams EM (1987) Is phosphodiesterase inhibition arrhythmogenic?
Electrophysiologic effects in pithed rats and in normoxic and hypoxic rabbit atria of
enoximone, a new cardiotonic agent. J Clin Pharmacol 27: 91-100.
Weibel ER, Cruz-Orive LM. Morphometric methods. In:Crystal RG, West JB, Weibel
ER, Barnes PJ, eds. The Lung: Scientific Foundations. Lippincott-Raven, Philadelphia,
PA, 1997; pp. 333–344.
Woyda K, Koebrich S, Reiss I, Rudloff S, Pullamsetti SS, Rühlmann A, Weissmann
N, Ghofrani HA, Günther A, Seeger W, Grimminger F, Morty RE, Schermuly RT (2009)
Inhibition of phosphodiesterase 4 enhances lung alveolarisation in neonatal mice exposed
to hyperoxia. Eur Respir J 33:861-70.
Yoon BH, Romero R, Jun JK, Park KH, Park JD, Ghezzi F, and Kim BI (1997)
Amniotic fluid cytokines (interleukin-6, tumor necrosis factor-alpha, interleukin-1 beta,
and interleukin-8) and the risk for the development of bronchopulmonary dysplasia. Am J
Obstet Gynecol 177: 825-830.
Zeller E, Stief HJ, Pflug B, and Sastre-y-Hernandez M (1984) Results of a phase II
study of the antidepressant effect of rolipram. Pharmacopsychiatry 17: 188-190.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
30
LEGENDS FOR FIGURES
Figure 1. Perinatal mortality from inoculation to birth of pups. Data are presented as
relative value according to the time of death. Intrauterine death was significantly increased in
groups C+R+ and R+ as compared with both Controls and C+; #: p<0.03 for intrauterine
demise compared to controls and group C+. ¶: p<0.05 for neonatal demise compared to
controls. Neonatal death : hached bar, intrauterine death : solid black bars and live pups: white
bars.
Figure 2. Microvascularization and elastogenesis at day 5.
A: Immunohistochemistry labeling for microvascularization. Monoclonal antibody against
human CD31 was used. Data are percentage of labeled cells out of total parenchymal cells.
Microvascularization was significantly decreased in groups C+ (solid black bars), C+R+
(pointed bars), and R+ (hatched bars) compared to controls (solid white bars). Data are
presented as mean ± SD. # : p<0.01 compared to controls.
B: Specific elastine fiber length assessed by morphometry (cm/100g). Fiber length was
significantly decreased in groups C+, C+R+ and R+ as compared with Controls. Data are
presented as mean ± SD. # : p<0.05 compared to Controls.
Figure 3. Cell Turnover at day 0. A. Relative value of cell apoptosis over total cells in
lungs. Apoptosis was significantly decreased in group R+ (hached bars) as compared to
controls (solid white bars) and group C+R+ (pointed bars). Group C+ is presented as solid
black bars, data are presented as mean ± SD. # : p<0.05 compared to controls and C+R+B
PCNA index in terminal alveolar epithelium. B. Alveolar PCNA index: ratio of PCNA-
positive cells to total epithelial cells, expressed as a percentage. Data are presented as mean ±
SD. # : p<0.05 compared to controls.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
31
Figure 4. PDE activity in whole lung homogenates of rabbit pups exposed to
chorioamnionitis with or without rolipram. No significant difference in PDE activity was
found between the groups at day 0.
Control group:solid white bars, groups C+ :solid black bars, group C+R+ : pointed bars, and
R+: hatched bars. Data are presented as mean ± SD.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
32
TABLES
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET#179085
33
Table 1. Morphometric measurements at day 0
Controls (n = 6)
C+ (n = 6)
C+ R+ (n = 6)
R+ (n = 8)
Body weight (g) 56.68 ± 9.3 47.18 ± 4.5 32.18 ± 6.1 39.90 ± 2.7
Lung volume (cm3) 2.89 ± 0.19 2.61 ± 0.26 2.32 ± 0.68 2.36 ± 0.33
Specific lung volume (cm3/100g)
5.23 ± 0.98 5.56 ± 0.62 7.09 ± 1.95 5.92 ± 0.73
Alveolar airspace
Volume density 0.66 ± 0.03 0.70 ± 0.02 0.64 ± 0.03 0.67 ± 0.01
Absolute volume (cm3) 1.93 ± 0.11 1.66 ± 0.48 1.49 ± 0.46 1.59 ± 0.22
Specific volume (cm3/100g) 3.50 ± 0.63 3.66 ± 0.78 4.54 ± 1.26 3.99 ± 0.47
Alveolar surface
Area density (cm2/cm3) 284 ± 34 225 ± 27 230 ± 84 223 ± 31
Absolute area(cm2) 822 ± 130 542 ± 193 520 ± 182 528 ± 108
Specific area (cm2/100g) 1485 ± 337 1187 ± 336 1660 ± 838 1320 ± 233
Interstitium
Volume density 0.28 ± 0.01 0.25 ± 0.02 0.28 ± 0.05 0.28 ± 0.02
Absolute volume (cm3) 0.82 ± 0.05 0.60 ± 0.19 0.64 ± 0.18 0.66 ± 0.11
Specific volume (cm3/100g) 1.48 ± 0.26 1.33 ± 0.31 2.00 ± 0.60 1.68 ± 0.26
Airways
Volume density 0.02 ± 0.01 0.02 ± 0.01 0.03 ± 0.01 0.02 ± 0.01
Absolute volume (cm3) 0.06 ± 0.04 0.04 ± 0.03 0.08 ± 0.05 0.05 ± 0.03
Specific volume (cm3/100g) 0.12 ± 0.09 0.10 ± 0.06 0.24 ± 0.15 0.13 ± 0.07
Blood vessels
Volume density 0.02 ± 0.01 0.01 ± 0.01 0.04 ± 0.02 0.01 ± 0.01
Absolute volume (cm3) 0.07 ± 0.02 0.04 ± 0.01 0.09 ± 0.06 0.04 ± 0.02
Specific volume (cm3/100g) 0.14 ± 0.05 0.09 ± 0.01 0.29 ± 0.19 0.11 ± 0.06
Data are expressed as mean ± SD, comparison results are expressed with the F1/22 value accompanied with the
corresponding p value.
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on D
ecember 7, 2011 as D
OI: 10.1124/jpet.111.179085
at ASPET Journals on August 8, 2018 jpet.aspetjournals.org Downloaded from

JPET#179085
34
Body weight; Infection effect F (1,22) =12.2, p=0.0002; Rolipram effect F (1,22) =27.7, p<0.0001. Interaction P>0.05
Alveolar airspace (volume density); Rolipram effect F(1,22) = 11.7, p=0.002; Interaction F (1,22)=16.0, p=0.0005
Alveolar airspace (specific volume), Rolipram effect F(1,22) =4.4, p=0,04
Interstitium (specific volume); Rolipram effect F(1,22) = 8.4, p=0.008;
Blood vessels (volume density); Interaction only F(1,22) = 6.8, p=0.05
Blood vessels (absolute volume ); Interaction only F(1,22) = 8.6, p=0.007
Blood vessels (specific volume );Rolipram effect F(1,22) = 4,7, p=0,04; Interaction F(1,22) = 7,8, p=0,01
All other comparisons not statistically significant
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on D
ecember 7, 2011 as D
OI: 10.1124/jpet.111.179085
at ASPET Journals on August 8, 2018 jpet.aspetjournals.org Downloaded from

JPET#179085
35
Table 2. Morphometric measurements at day 5
groups day 5
Controls (n = 10)
C+ (n = 8)
C+ R+ (n = 5)
R+ (n = 10)
Body weight (g) 112.9 ± 10.8 72.7 ± 15.8 102.20 ± 14.5 70.89 ± 20.5
Lung volume (cm3) 5.99 ± 0.41 3.10 ± 0.62 5.15 ± 0.71 3.85 ± 0.86
Specific lung volume (cm3/100g) 5.34 ± 0.65 5.09 ± 0.95 5.05 ± 0.69 5.63 ± 1.13
Alveolar airspace
Volume density 0.60 ± 0.05 0.69 ± 0.03 0.60 ± 0.05 0.66 ± 0.06
Absolute volume (cm3) 3.30 ± 0.54 2.75 ± 0.53 3.13 ± 0.62 2.55 ± 0.53
Specific volume (cm3/100g) 3.14 ± 0.39 3.80 ± 0.32 3.04 ± 0.41 3.79 ± 1.01
Alveolar surface
Area density (cm2/cm3) 253 ± 25 223 ± 34 205 ± 5 220 ± 32
Absolute area(cm2) 1380 ± 249 862 ± 107 1057 ± 158 857 ± 253
Specific area (cm2/100g) 1313 ± 207 1214 ± 191 1034 ± 134 1221 ± 218
Interstitium
Volume density 0.33 ± 0.04 0.26 ± 0.01 0.30 ± 0.02 0.28 ± 0.05
Absolute volume (cm3) 1.82 ± 0.29 1.05 ± 0.24 1.57 ± 0.17 1.11 ± 0.38
Specific volume (cm3/100g) 1.74 ± 0.29 1.45 ± 0.11 1.55 ± 0.29 1.56 ± 0.24
Airways
Volume density 0.02 ± 0.01 0.02 ± 0.01 0.04 ± 0.03 0.02 ± 0.01
Absolute volume (cm3) 0.14 ± 0.06 0.07 ± 0.04 0.22 ± 0.12 0.08 ± 0.04
Specific volume (cm3/100g) 0.13 ± 0.06 0.11 ± 0.06 0.23 ± 0.17 0.12 ± 0.04
Blood vessels Volume density 0.05 ± 0.05 0.02 ± 0.01 0.04 ± 0.01 0.02 ± 0.01
Absolute volume (cm3) 0.33 ± 0.35 0.08 ± 0.03 0.21 ± 0.07 0.10 ± 0.04
Specific volume (cm3/100g) 0.31 ± 0.36 0.14 ± 0.05 0.21 ± 0.06 0.15 ± 0.08
Data are expressed as mean ± SD, comparison results are expressed with the F(1,21) value accompanied with the
corresponding p value.
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on D
ecember 7, 2011 as D
OI: 10.1124/jpet.111.179085
at ASPET Journals on August 8, 2018 jpet.aspetjournals.org Downloaded from

JPET#179085
36
Body weight; Interaction effect only F (1,21) =44,6, p= 0,000001
Lung volume; infection effect F(1,22) = 7,6, p= 0,01; interaction effect F(1,21)=52,3, p=0,000001
Alveolar airspace (volume density); Interaction effect only F (1,21)= 10,4, p=0,003
Alveolar airspace (absolute volume), Interaction effect only F(1,21) =18,8, p=0,0002
Alveolar surface (area density); Rolipram effect F(1,21)= 11,7, p=0,002
Alveolar surface (absolute area ); Rolipram effect F(1,21)= 9 ,6, p=0,005
Alveolar surface (specific area ); Rolipram effect F(1,21)= 8,0, p=0,009
Interstitium ( volume density); interaction effect only F(1,21)= 6,1, p=0,02
Airways (absolute volume); interaction effect only F(1,21)= 10,9, p=0,003
Blood vessels (absolute volume ); Interaction only F(1,21) =5,5, p=0,02
All other comparisons not statistically significant
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on D
ecember 7, 2011 as D
OI: 10.1124/jpet.111.179085
at ASPET Journals on August 8, 2018 jpet.aspetjournals.org Downloaded from

JPET#179085
37
Table 3: Effects of antenatal infection, Rolipram or both on bronchoalveolar lavage cell count
from day 0 to day 5
BAL cell count data are expressed as mean ± SD
Day 0 Day 5
Controls C+ C+R+ R+ Controls C+ C+R+
BAL cell count (X 104/ml)
1,3±1,0 1,6±1,7 2,4±0,1 0,8±0,2 0,9±0,9 1,2±1,1 5,1±0,2
PMN, % 0,51±0,16 0,88±0,45 0,91±0,22 1,22±0,67 2,07±1,24 2,45±1,56 3,66±1,89
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 7, 2011 as DOI: 10.1124/jpet.111.179085
at ASPE
T Journals on A
ugust 8, 2018jpet.aspetjournals.org
Dow
nloaded from