analysis of sex-mismatched human corneal transplants by fluorescence in situ hybridization of the...
TRANSCRIPT

Exp. Eye Res. (1999) 68, 341–346Article No. exer.1998.0611, available online at http:}}www.idealibrary.com on
Analysis of Sex-mismatched Human Corneal Transplants by
Flourescence in situ Hybridization of the Sex-chromosomes
GREGOR WOLLENSAK* WILLIAM RICHARD GREEN
Eye Pathology Lab, Wilmer Eye Institute, Johns Hopkins Hospital, Baltimore, MD 21287-9248, U.S.A.
(Received Lund 11 June 1998 and accepted in revised form 10 November 1998)
The fate of the cells of corneal transplants has been controversial from the early days of keratoplasty.Various methods such as histological evaluation, radiolabeling of donor cells or Barr-body analysis havebeen applied to clarify the issue. However, the question whether the transplanted cells are replaced orsurvive, remains unsolved.
In this study, we applied fluorescence in situ hybridization (FISH) of the X- and Y-chromosomes inparaffin sections of explanted sex-mismatched corneal transplants to distinguish between host and donorcells. Fourteen sex-mismatched cases with various reasons for explantation and different postoperativetime intervals ranging from 11 months to 30 years were analysed. We found that all cell types, includingepithelium, keratocytes and endothelial donor cells were replaced in most cases as early as 1 year aftertransplantation. In three cases, however, up to 26% of donor keratocytes were still detected up to 4±5years after transplantation, demonstrating a certain individual variability in the process of replacement.Further studies must show if the extent and timing of donor cell replacement in clinically successful,totally clear transplants is different.
Our results are in keeping with the phenomenon of recurrences of corneal dystrophies in the graft, thesignificant postoperative decline of the endothelial cell density, the fact that typical graft rejections usuallytake place within 1–2 years postoperatively and that relatively late rejections can occur in rare casesprobably due to some surviving stromal keratocytes.
Donor cell replacement is a special feature of corneal transplants when compared with other kinds oforgan transplants and might be due to the presence of the same tissue type in the immediateneighbourhood of the graft. # 1999 Academic Press
Key words : FISH; sex chromosomes; sex-mismatch; corneal transplantation; transplant replacement;transplant survival.
1. Introduction
The first successful human corneal transplantation
was performed by Zirm in 1905. From those early
beginnings of keratoplasty until today the fate of the
cells of the corneal transplant has been a controversial
issue. Salzer, Bonnefon, Lacoste, and Loehlein were
strong advocates of the hypothesis that the corneal
transplant is completely replaced by the recipient cells
while Fuchs, Ascher, Elschnig, Castroviejo and Filatov
favored the hypothesis that the corneal transplant
retains its identity and the transplanted cells survive
(Maumenee and Kornblueth, 1948; Salzer, 1990).
These opinions, however, were based mainly on the
histopathological analysis of cases with no reliable
possibility of distinguishing between donor and host
cells. Especially, the obvious survival of Bowman’s and
Descemet’s membrane was a strong argument in favor
of the survival theory while the successful trans-
plantation of non-viable, formalinized corneal trans-
plants backed up the replacement theory (Salzer,
1919; Scherschewskaya, 1940).
Later this question was addressed using the Barr-
body identification in sex-mismatched corneal trans-
plants (Basu and Carre! , 1973; Bourne, 1974; Espiritu,
* Corresponding author: Gregor Wollensak, Wildentensteig 4, D-14195 Berlin, Germany.
Kara and Tabowitz, 1961; Klen and Hradil, 1963;
Kinoshita, Friend and Thoft, 1981). However, the
Barr-body method only allows determination of the
statistical probability of the gender of a piece of tissue
but not of single individual cells. In addition, dis-
appearance of the Barr body has been described with
aging of the cells and the Barr body of the stromal
keratocytes, and mostly also of the endothelium can
only be detected in tangential sections of the cornea or
in cultural outgrowths (Basu and Carre! , 1973; Basu,
Sarkar and Carre! , 1964; Pedler and Ashton, 1955).
Other methods, which cannot be used in humans
for ethical reasons, were: radiolabeling of donor cells
with tritiated thymidine (Khodadoust and Silverstein,
1969; Polack, Smelser and Rose, 1964), labeling of
host cells with trypan blue (Maumenee and
Kornblueth, 1948), and karyotyping of host and
donor cells in heterografts with different species (Basu,
Sarkar and Carre! , 1964).
However, most of these studies yielded controversial
results, partly due to the method or animal model
used, but also, we believe because of the relatively
short postoperative time interval after the trans-
plantation.
Recently, we have applied fluorescence in situ
hybridization (FISH) of the X- and Y-chromosome to
ocular tissue. In situ hybridization allows the local-
0014–4835}99}03034106 $30.00}0 # 1999 Academic Press

342 G. WOLLENSAK AND W. R. GREEN
ization of specific DNA-sequences by using a labeled
complementary DNA-probe. This method is very
specific and has the advantage that it can identify the
gender of a cell to a high percentage in all cell types
present in the eye, without destroying the ocular
texture (Wollensak, Perlman and Green, 1998). In
situ hybridization of the sex-chromosomes has also
already been successfully used in the analysis of sex-
mismatched skin (Brain et al., 1989; Burt et al.,
1989), lung (Yousem and Sonmez-Alpan, 1991) and
cardiac transplantation (Hruban et al., 1993).
In the present study we have applied this approach
to analyse human eyes with sex-mismatched corneal
transplantations. In contrast to earlier studies, we
were able to use specimens with a postoperative time
interval of up to 30 years.
2. Materials and Methods
Corneal Explants
All corneal buttons were retrieved retrospectively
from the archives of the Eye Pathology Laboratory at
the Wilmer Eye Institute in Baltimore, U.S.A.
We selected 14 sex-mismatched corneal buttons
with a wide range of different postoperative time
intervals ranging from 11 months to 30 years (Table
I). Three of the recipients were male (cases 9, 11 and
13) and 12 were female.
The corneal transplants had been explanted for
various reasons as can be seen in Table I. The gender
of the donor of the corneal graft was established either
from the clinical charts if available or by application of
in situ hybridization of the sex-chromosomes on
paraffin sections of the donor corneoscleral rims which
routinely undergo histologic examination at the
Wilmer Eye Institute.
In addition, two control cases with no sex-mismatch
and metaphase spreads of male peripheral lympho-
cytes were tested (Fig. 1). The two control cases were
T I
Results of the analysis of the 14 sex-mismatched cases
Case number Donor keratocytes Time interval Reason for removal
1 none 11 mos pseudopemphigoid2 none 14 mos endothelial failure3 15% (63 of 420 cells) 2 y, 5 mos graft rejection4 20% (90 of 450 cells) 2 y, 6 mos endothelial failure5 none 3 y, 2 mos endothelial failure6 none 4 y endothelial failure7 none 4 y graft rejection8 26% (94 of 360 cells) 4±5 y endothelial failure9 none 9 y recurrent lattice dystrophy
10 none 11 y recurrent keratoconus11 none 12 y recurrent lattice dystrophy12 none 16 y endothelial failure13 none 20 y endothelial failure14 none 30 y endothelial failure
In no case, were there surviving epithelial or endothelial donor cells.
F. 1. Fluorescence photomicrograph of a peripherallymphocyte (right) and a metaphase spread of a maleperipheral lymphocyte. Note the green signal for the Y-(lower arrow) and the red signal for the X-chromosome(upper arrow). (¬416, oil immerson, DAPI counterstain).
cases of female recipients with female donor buttons
with a postoperative time interval of 1±5 years and 4±5years.
Fixation and Hybridization Conditions
All the specimens used in this study had been fixed
in 10% neutral buffered formalin for about 1 day and
embedded in paraffin. Six micrometer thick paraffin
sections were mounted on silanised slides. Three slides
with two sections were tested in each case.
The DNA-probes for the X (α-satellite)- and Y-
chromosome (classical satellite) were from Oncor,
Gaithersburg, MD, U.S.A. The details of the technical
procedure for the fluorescence in situ hybridization
have been described previously (Wollensak, Perlman
and Green, 1999).
In short, the procedure involved pretreatment with
hot 50%-glycerol for 10 min, and digestion with
proteinase K (0±1 µg ml−") for 45 min at 37°C. Dena-
turation was performed by applying a formamide

SEX-MISMATCHED CORNEAL TRANSPLANTS 343
solution for 10 min at 72°C. Incubation with the
denatured probes was carried out overnight. Dual
detection and dual amplification with a rhodamine-
labeled antibody for the detection of the X- and a
fluorescein-labeled antibody for the detection of the Y-
chromosome was used. The slides were counterstained
with the bluish fluorescent dye 4«-6-diamidino-2-
phenylindole (DAPI).
For the corneoscleral donor rims and the two
control cases the FISH-procedure was identical. The
metaphase spreads of the male lymphocytes were not
digested and were denatured for only 2 minutes.
Fluorescence Microscopy
The specimens were examined under a Zeiss-
axiomat microscope using a triple band pass filter for
fluorescein (460 nm), rhodamine (570 nm) and DAPI
(365 nm). Most of the micrographs were taken with
an attached Zeiss-camera. A few images were captured
by a CCD camera (Photometrics, Tucson, AZ, U.S.A.)
and processed using the Oncor Image analyzing
system.
3. Results
Two red signals for the two X-chromosomes in
female cells, and a green and a red signal in male cells
(Fig. 1) were visualized in about 98% of epithelial (Figs
2 and 3), in about 70% of the stromal keratocytes (Figs
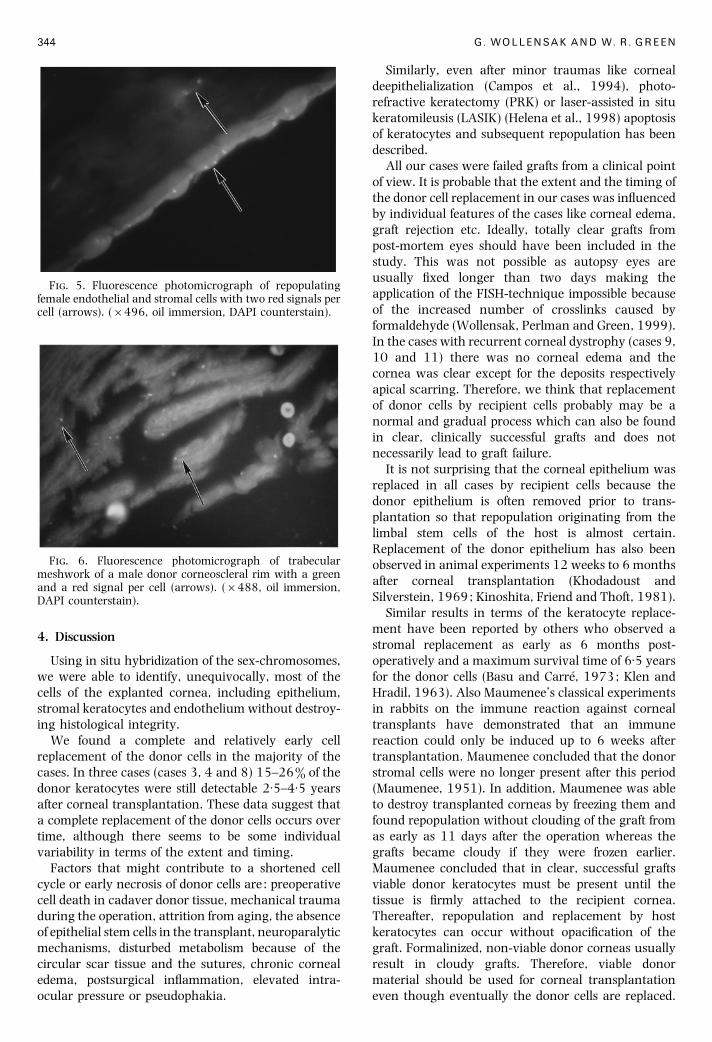
4 and 5) and 85% of endothelial cells (Fig. 5). The
signals had often to be observed at different levels of
focus. Sometimes, distortion of the stromal collagen
fibers occurred but was minimized by placing the slides
on ice at specific steps during the FISH-procedure
(Wollensak, Perlman and Green, 1998).
The FISH-procedure was also successful in the sex
determination of the donor corneoscleral rims (Fig. 6).
In the two control cases with identical gender of the
donor and recipient, only cells of one gender were
found.
In all the 14 sex-mismatched cases, the epithelium
and the endothelium were invariably replaced by cells
of the recipient as early as one year postoperatively
(Table I). The stromal keratocytes were completely
replaced by host cells in 11 cases while a cellular
mosaic (Fig. 4) with 15–26% of donor keratocytes was
present in three cases at 2±5, 2±6 and 4±5 years
postoperatively (cases 3, 4, 8). The surviving stromal
keratocytes were present in small groups of 2–4 cells
which were scattered throughout the corneal stroma
without a preferential localization.
The reason for transplant removal was recurrence
of the underlying dystrophy (3 cases), graft rejection
(2 cases), surface problems (1 case) or endothelial
failure (8 cases) with endothelial counts of up to only
4 nuclei per high power field (¬40). In 2 of the 8 cases
with endothelial failure (cases 5 and 6), pseudophakia
contributed to the endothelial atrophy.
F. 2. Fluorescence photomicrograph of repopulatingfemale epithelium with two red signals per cells (arrows)identifying them as female like the female recipient. (¬416,oil immersion, DAPI counterstain).
F. 3. Fluorescence photomicrograph of repopulatingmale epithelium with a green and red signal per cell (arrows)in the transplanted cornea of a male recipient. (¬488, oilimmersion, DAPI counterstain).
F. 4. Fluorescence photomicrograph of stromal mosaicwith two male keratocytes (green signal for the Y-chromosomes, lower arrows) and one repopulating femalekeratocyte (with two red signals for the X-chromosomes,upper arrow) in a male transplant that was grafted to afemale recipient. The red signals in the male cells are notvisible because they are at another level of focus. (¬496, oilimmersion, DAPI counterstain).
In three cases (cases 1, 2 and 4) a retrocorneal
membrane was present and shown to be of the
recipient gender.
344 G. WOLLENSAK AND W. R. GREEN
F. 5. Fluorescence photomicrograph of repopulatingfemale endothelial and stromal cells with two red signals percell (arrows). (¬496, oil immersion, DAPI counterstain).
F. 6. Fluorescence photomicrograph of trabecularmeshwork of a male donor corneoscleral rim with a greenand a red signal per cell (arrows). (¬488, oil immersion,DAPI counterstain).
4. Discussion
Using in situ hybridization of the sex-chromosomes,
we were able to identify, unequivocally, most of the
cells of the explanted cornea, including epithelium,
stromal keratocytes and endothelium without destroy-
ing histological integrity.
We found a complete and relatively early cell
replacement of the donor cells in the majority of the
cases. In three cases (cases 3, 4 and 8) 15–26% of the
donor keratocytes were still detectable 2±5–4±5 years
after corneal transplantation. These data suggest that
a complete replacement of the donor cells occurs over
time, although there seems to be some individual
variability in terms of the extent and timing.
Factors that might contribute to a shortened cell
cycle or early necrosis of donor cells are : preoperative
cell death in cadaver donor tissue, mechanical trauma
during the operation, attrition from aging, the absence
of epithelial stem cells in the transplant, neuroparalytic
mechanisms, disturbed metabolism because of the
circular scar tissue and the sutures, chronic corneal
edema, postsurgical inflammation, elevated intra-
ocular pressure or pseudophakia.
Similarly, even after minor traumas like corneal
deepithelialization (Campos et al., 1994), photo-
refractive keratectomy (PRK) or laser-assisted in situ
keratomileusis (LASIK) (Helena et al., 1998) apoptosis
of keratocytes and subsequent repopulation has been
described.
All our cases were failed grafts from a clinical point
of view. It is probable that the extent and the timing of
the donor cell replacement in our cases was influenced
by individual features of the cases like corneal edema,
graft rejection etc. Ideally, totally clear grafts from
post-mortem eyes should have been included in the
study. This was not possible as autopsy eyes are
usually fixed longer than two days making the
application of the FISH-technique impossible because
of the increased number of crosslinks caused by
formaldehyde (Wollensak, Perlman and Green, 1999).
In the cases with recurrent corneal dystrophy (cases 9,
10 and 11) there was no corneal edema and the
cornea was clear except for the deposits respectively
apical scarring. Therefore, we think that replacement
of donor cells by recipient cells probably may be a
normal and gradual process which can also be found
in clear, clinically successful grafts and does not
necessarily lead to graft failure.
It is not surprising that the corneal epithelium was
replaced in all cases by recipient cells because the
donor epithelium is often removed prior to trans-
plantation so that repopulation originating from the
limbal stem cells of the host is almost certain.
Replacement of the donor epithelium has also been
observed in animal experiments 12 weeks to 6 months
after corneal transplantation (Khodadoust and
Silverstein, 1969; Kinoshita, Friend and Thoft, 1981).
Similar results in terms of the keratocyte replace-
ment have been reported by others who observed a
stromal replacement as early as 6 months post-
operatively and a maximum survival time of 6±5 years
for the donor cells (Basu and Carre! , 1973; Klen and
Hradil, 1963). Also Maumenee’s classical experiments
in rabbits on the immune reaction against corneal
transplants have demonstrated that an immune
reaction could only be induced up to 6 weeks after
transplantation. Maumenee concluded that the donor
stromal cells were no longer present after this period
(Maumenee, 1951). In addition, Maumenee was able
to destroy transplanted corneas by freezing them and
found repopulation without clouding of the graft from
as early as 11 days after the operation whereas the
grafts became cloudy if they were frozen earlier.
Maumenee concluded that in clear, successful grafts
viable donor keratocytes must be present until the
tissue is firmly attached to the recipient cornea.
Thereafter, repopulation and replacement by host
keratocytes can occur without opacification of the
graft. Formalinized, non-viable donor corneas usually
result in cloudy grafts. Therefore, viable donor
material should be used for corneal transplantation
even though eventually the donor cells are replaced.

SEX-MISMATCHED CORNEAL TRANSPLANTS 345
During the time of the gradual replacement, viable
donor cells seem to be necessary to avoid irreversible
scarring of the cornea (Maumenee and Kornblueth,
1948).
The observed complete cell replacement of the donor
endothelium in the early postoperative years is in good
accordance with clinical studies that have shown that
the endothelial cell count in corneal transplants
decreases up to 65% during the first two postoperative
years which is much more than expected from normal
aging or due to pseudophakia (Waring et al., 1982).
Probably, the repopulating host endothelial cells
migrate towards the center of the tranplanted cornea
with flattening and enlargement of the cells and only
little actual mitotic proliferation (Waring et al., 1982).
It has also been found that the recipient age has an
unexpected significance as a predictor of endothelial
cell survival in the respect that endothelial cell counts
decrease with increasing recipient age regardless of
the donor age (Linn et al., 1981; Williams et al.,
1997). In animal experiments, survival of endothelial
cells has been observed from 5 weeks to 12 months
postoperatively. (Bourne, 1974; Polack et al., 1964)
and replacement by host endothelial cells between 4 to
7 months postoperatively (Espiritu, Kara and Tabo-
witz, 1961).
The time scheme which emerges from our results, is
in good accordance with the clinical observation of
graft rejection being mostly in the first two years but
also happening later in a few cases. In addition, these
results are in keeping with numerous reports about
recurrences of various corneal dystrophies like
Meesmann’s epithelial (Chiou et al., 1998), Reis-
Bu$ ckler’s (Olson and Kaufman, 1978), keratoconus
(Bechrakis et al., 1994), granular (Lyons et al., 1994),
macular (Klintworth et al., 1983), lattice (Klintworth
et al., 1982) or posterior polymorphus dystrophy
(Boruchoff et al., 1990) as early as 2–3 years after
corneal transplantation or later in some cases.
Moreover, recurrences of corneal dystrophies are a
clear clinical proof of the phenomenon of repopulation
of the graft by host cells.
The fact that the donor cornea seems to serve
primarily as a matrix or framework with a complete
replacement of the donor cells by recipient cells in the
long term makes corneal transplantation unique in
comparison with other organ transplants like lung
(Yousem and Sonmez-Alpan, 1991), kidney (Sedmak
et al., 1988; Williams et al., 1969), liver (Gouw et al.,
1987), small bowel (Iwaki et al., 1991) or cardiac
transplantats (O’Connell et al., 1991) where usually
only the tissue macrophages, lymphocytes and the
endothelium of the blood vessels are replaced by the
recipient cells. In addition, in other organ transplants,
no cadaver donor organs are used and vascular
anastomoses are necessary.
The nature of corneal transplantation can perhaps
be compared to blood transfusion (Salzer, 1900) in
contrast to a bone marrow transplantation. Interest-
ingly, also skin grafts (Burt et al., 1989; Plenat et al.,
1992), epidermal (Brain et al., 1989; Gielen et al.,
1987) and fascial allografts (West, Crawford and
Basu, 1969) are completely replaced by the recipient
cells.
In summary, our study has shown that all cell types
of corneal transplants tend to be replaced by recipient
cells in the long term. Epithelium and endothelium are
replaced relatively early within the first postoperative
years. Small proportions of stromal keratocytes can
survive for several years but are probably replaced
eventually. Individual variability in the process of
replacement exists. Further studies must determine if
there is a higher rate of surviving donor cells in totally
clear, clinically successful grafts. Our findings can help
to explain and understand better many features of
corneal transplantation like recurrences of corneal
dystrophies, postoperative decrease of endothelial cell
density and timing of graft rejections.
Acknowledgements
This work was supported by the Deutsche Forschungs-gemeinschaft and The International Order of Odd Fellows,Winston-Salem, North Carolina, U.S.A.
References
Basu, P. K. and Carre! , F. (1973). A study of cells in humancorneal grafts. Can. J. Ophthalmol. 8, 1–7.
Basu, P. K., Sarkar, P. and Carre! , F. (1964). Use of thekaryotype as biologic cell marker. Am. J. Ophthalmol.58, 569–72.
Bechrakis, N., Blom, M. L., Stark, W. J. and Green, W. R.(1994). Recurrent keratoconus. Cornea 13, 73–7.
Boruchoff, S. A., Weiner, M. J. and Albert, D. M. (1990).Recurrence of posterior polymorphous corneal dys-trophy after penetrating keratoplasty. Am. J. Ophthalmol.109, 323–8.
Bourne, W. M. (1974). In vivo survival of cryopreservedendothelial cells in primates. Arch. Ophthalmol. 92,146–8.
Brain, A., Purkis, P., Coats, P., Hackett, M., Navsaria, H. andLeigh, I. (1989). Survival of culturedallogeneic keratino-cytes transplanted to deep dermal bed assessed withprobe specific for Y-chromosome. Brit. Med. J. 298,917–19.
Burt, A. M., Pallett, C. D., Sloane, J. P., O’Hare, M. J.,Schafler, K. F., Yardeni, P., Eldad, A., Clarke, J. A. andGusterson, B. A. (1989). Survival of cultured allograftsin patients with burns assessed with probe specific for Y-chromosome. Brit. Med. J. 298, 915–17.
Campos, M., Szerenyi, K., McDonnell, J. M., Lopez, P. F. andMcDonnell, P. J. (1994). Keratocyte loss after cornealdeepithelialization in primates and rabbits. Arch.Ophthalmol. 112, 254–60.
Chiou, A. G.-Y., Florakis, G. J., Copeland, R. L., Williams,V. A., McCormick, S. A. and Chiesa, R. (1998). Re-current Meesmann’s corneal epithelial dystrophy afterpenetrating keratoplasty. Cornea 17, 566–70.
Espiritu, R. B., Kara, G. B. and Tabowitz, D. (1961). Studieson the healing of corneal grafts. II. The fate of theendothelial cells of the graft as determined by sexchromatin studies. Am. J. Ophthalmol. 52, 91–5.

346 G. WOLLENSAK AND W. R. GREEN
Gielen, V., Faure, M., Mauduit, G. and Thivolet, J. (1987).Progressive replacement of human cultured epithelialallografts by recipient cells as evidenced by HLA class Iantigen expression. Dermatologica 175, 166–70.
Gouw, A. S. H., Houthoff, H. J., Huitema, S., Beelen, J. M.,Gips, C. H. and Poppema, S. (1987). Expression of majorhistocompatibility complex antigens and replacement ofdonor cells by recipient ones in human liver grafts.Transplantation 43, 291–6.
Helena, M. C., Baerveldt, F., Kim, W.-J. and Wilson, S. E.(1998). Keratocyte apoptosis after corneal surgery.Invest. Ophthalmol. Vis. Sci. 39, 276–83.
Hruban, R. H., Long, P. P., Perlman, E. J., Hutchins, G. M.,Baumgartner, W. A., Baughman, K. L. and Griffin,C. A. (1993). Fluorescence in situ hybridization for theY-chromosome can be used to detect cells of recipientorigin in allografted hearts following cardiac trans-plantation. Am. J. Pathology 142, 975–80.
Iwaki, Y., Starl, T. E., Yagihashi, A., Taniwaki, S., Abu-Elmagd, K., Tzakis, A., Fung, J. and Todo, S. (1991).Replacement of donor lymphoid tissue in small-boweltransplants. Lancet 337, 818–19.
Khodadoust, A. A. and Silverstein, A. M. (1969). Thesurvival and rejection of epithelium in experimentalcorneal transplants. Invest. Ophthalmol. Vis. Sci. 8,169–79.
Kinoshita, S., Friend, J. and Thoft, R. A. (1981). Sexchromatin of donor corneal epithelium in rabbits.Invest. Ophthalmol. Vis. Sci. 21, 434–41.
Klen, R. and Hradil, I. (1963). Beitrag zur Biologie desHornhautpfropfes. Sex-chromatin-Charakter der im-plantierten Scheibe. Graefes Arch. Clin. Exp. Ophthalmol.166, 180–7.
Klintworth, G. K., Ferry, A. P., Sugar, A. and Reed, J.(1982). Recurrence of lattice corneal dystrophy type 1in the corneal grafts of two siblings. Am. J. Ophthalmol.94, 540–6.
Klintworth, G. K., Reed, J., Stainer, G. A. and Binder, P. S.(1983). Recurrence of macular corneal dystrophywithin grafts. Am. J. Ophthalmol. 95, 60–72.
Linn, J. G., Stuart, J. C., Warnicki, J. W., Sinclair, R. A. andMarsh, G. M. (1981). Endothelial morphology in long-term keratoconus corneal transplants. Ophthalmology88, 761–70.
Lyons, C. J., McCartney, A. C., Kirkness, C. M., Ficker, L. A.,Steele, A. D. M. and Rice, N. S. C. (1994). Granularcorneal dystrophy. Visual results and pattern of re-currence after lamellar or penetrating keratoplasty.Ophthalmology 101, 1812–17.
Maumenee, A. E. and Kornblueth, W. (1948). Symposium:Corneal transplantation. VI. Physiopathology. Am. J.Ophthalmol. 31, 1384–93.
Maumenee, A. E. (1951). The influence of donor recipientsensitization on corneal grafts. Am. J. Ophthalmol. 34,142–52.
O’Connell, J. B., Renlund, D. G., Bristow, M. R. andHammond, E. H. (1991). Detection of allograft en-dothelial cells of recipient origin following AB0-com-patible, non-identical cardiac transplantation. Trans-plantation 51, 438–42.
Olson, R. J. and Kaufman, H. E. (1978). Recurrence of Reis-Bu$ ckler’s corneal dystrophy in a graft. Am. J. Oph-thalmol. 85, 349–51.
Pedler, C. and Ashton, N. (1955). Sex of nuclei in oculartissues. Brit. J. Ophthalmol. 39, 362–7.
Plenat, F., Vignaud, J.-M., Guerret-Stocker, S., Hartmann, D.and Duprez, A. (1992). Host-donor interactions inhealing of human split-thickness skin grafts onto nudemice: In situ hybridization, immunohistochemical, andhistochemical studies. Transplantation 53, 1002–10.
Polack, F. M., Smelser, G. K. and Rose, J. (1964). Long-termsurvival of isotopically labeled stromal and endothelialcells in corneal homografts. Am. J. Ophthalmol. 57,67–78.
Salzer, F. (1910). Beitra$ ge zur Keratoplastik II. UeberImplantationvonisolirtenSchichtenconservirterPferde-hornhaut in die Cornea des Kaninchens. Arch. f.Augenheilk. 65, 214–28.
Salzer, F. (1900). Kritische und litterarische Studien u$ berTransplantation in Hinblick auf die Frage der Kerato-plastik. Zeitschr. f. Augenheilk. 4, 124–45.
Scherschewskaya, O. J. (1940). Keratoplastische Versuchemit Anwendung von Formollappen. Ophthalmologica99, 4–15.
Sedmark, D. D., Sharma, H. M., Czajka, C. M. and Ferguson,R. M. (1988). Recipient endothelialization of renalallografts. Transplantation 46, 907–10.
Waring, G. O., Bourne, W. M., Edelhauser, H. F. and Kenyon,K. R. (1982). The corneal endothelium. Normal andpathologic structure and function. Ophthalmology 89,531–90.
West, R. D., Crawford, J. S. and Basu, P. K. (1969). The fateof the cellular elements in fascial grafts. Can. J.Ophthalmol. 4, 64–71.
Williams, G. M., Krajewski, C. A., Dagher, F. J., TerHaar,A. M., Roth, J. A. and Santos, G. W. (1969). Hostrepopulation of the endothelium in allografts of kidneyand aorta. Surg. forum. 20, 293–4.
Williams, K. A., Muehlberg, S. M., Lewis, R. F. and Coster,D. J. (1997). Influence of advanced recipient and donorage on the outcome of corneal transplantation. Brit. J.Ophthalmol. 81, 835–9.
Wollensak, G., Perlman, E. and Green, W. R. (1999).Interphase fluorescence in situ hybridization analysis ofthe X- and Y-chromosome in human eyes. Cytogenet.Cell Res. forthcoming.
Yousem, S. A. and Sonmez-Alpan, E. (1991). Use of abiotinylated DNA probe specific for the human Ychromosome in the evaluation of the allograft lung.Chest 99, 275–9.