activation of paneth cell -defensins in mouse small intestine
TRANSCRIPT

Activation of Paneth Cell -Defensins in Mouse Small Intestine
Tokiyoshi Ayabe‡*
1, Donald P. Satchell*
1, Patrizia Pesendorfer
3, Hiroki Tanabe
1, Carole L. Wil-
son4, Susan J. Hagen
5, and Andre J. Ouellette
1,2¶
Departments of 1Pathology and
2Microbiology & Molecular Genetics, University of California
College of Medicine, Irvine, CA 92697-4800, 3Department of Pediatric Surgery, Karl-Franzens-
Univerity Graz, Graz, Austria, 4Division of Allergy and Pulmonary Medicine, Department of
Pediatrics, Washington University School of Medicine, St. Louis, MO 63110, and 5Department
of Surgery, Beth Israel Deaconess Medical Center, 300 Brookline Avenue, Boston, MA 02115
*Equal contributors to the study.
‡Current address:
Third Department of Internal Medicine, Asahikawa Medical College, Asahikawa, Japan
Running title: Intracellular processing of mouse procryptdins
¶Corresponding author: Andre J. Ouellette, Ph.D., Department of Pathology,
College of Medicine, University of California, Irvine, CA 92697-4800.
Tel: (949)824-4647, Fax: (949)824-1098, E-mail: [email protected]
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 2
ABSTRACT
Paneth cells in small intestinal crypts secrete microbicidal -defensins, termed cryptdins,
as components of enteric innate immunity. The bactericidal activity of cryptdins requires prote-
olytic activation of precursors by the matrix metalloproteinase matrilysin (MMP-7a, Wilson et
al., Science: 286, 113-117, 1999). Here, we report on the intracellular processing of cryptdin
proforms in mouse Paneth cells. Peptide sequencing of MMP-7 digests of purified natural
procryptdins identified conserved cleavage sites in the proregion between Ser43 and Val44 as
well as at the cryptdin peptide N-terminus between Ser58 and Leu59. Immune staining co-
localized precursor prosegments and mature cryptdin peptides to Paneth cell granules, providing
evidence of their secretion. Extensive MMP-7-dependent procryptdin processing occurs in Pa-
neth cells as shown by western blot analyses of intestinal crypt proteins and proteins from gran-
ule-enriched subcellular fractions. The addition of soluble prosegment to in vitro antimicrobial
peptide assays inhibited the bactericidal activities of cryptdins 3 and 4 in trans, suggesting pos-
sible cytoprotective effects by prosegments prior to secretion. Levels of activated cryptdins are
normal in small bowel of germ-free mice and in sterile implants of fetal mouse small intestine
grown subcutaneously. Thus, the initiation of procryptdin processing by MMP-7 does not re-
quire direct bacterial exposure, and the basal MMP-7 content of germ-free Paneth cells is suffi-
cient to process and activate -defensin precursors. MMP-7-dependent procryptdin activation in
vivo provides mouse Paneth cells with functional peptides for apical secretion into the small in-
testinal lumen.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 3
INTRODUCTION
The release of endogenous antimicrobial peptides by mammalian epithelial cells contrib-
utes to innate mucosal immunity (1,2). The crypts of Lieberkühn in the small intestine of most
mammals contain Paneth cells that secrete -defensins (cryptdins), lysozyme, sPLA2, xanthine
oxidase, CD95 ligand, CD15, and TNF- as components of apically-oriented secretory granules
(3-10). Although certain Paneth cell -defensins have been detected in mouse skin and testis
(11,12) and in human oropharyngeal and urogenital mucosa (13,14), in the small intestine, -
defensins are specific to Paneth cells (9). Exposure of Paneth cells to cholinergic agonists or
bacterial stimuli elicits granule discharge into the crypt lumen (15), and carbamyl choline medi-
ates secretion via increased cytosolic Ca2+
(16). Regardless of how mouse Paneth cell secretion
is stimulated, cryptdins constitute ~70% of the released bactericidal activity, and the concentra-
tion of cryptdins is estimated to be 25 mM at the point of secretion in the crypt lumen (15).
-Defensins are processed from inactive proforms by specific proteolytic cleavage steps.
Both neutrophil and Paneth cell -defensins derive from ~10 kDa prepropeptides that contain
canonical signal sequences, acidic proregions, and a ~3.5 kDa mature -defensin peptide in the
C-terminal portion of the precursor. For example, maturation of myeloid pro -defensins ap-
pears to involve two primary cleavage steps, and most -defensins in mature phagocytic leuko-
cytes are completely processed (17-20). In a heterologously-expressed human neutrophil pro- -
defensin, deletions in the prosegment adjacent to the proregion-defensin junction impaired post-
translational processing in 32DCL3 cells (19).
In mouse Paneth cells, the matrix metalloproteinase matrilysin (MMP-7) mediates the
processing and activation of -defensins from 8.4 kDa proforms (21). MMP-7 gene disruption
ablates procryptdin processing, resulting in accumulation of cryptdin precursors and the absence
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 4
of activated, mature cryptdin peptides in the small intestine (21). Lacking functional cryptdin
peptides, MMP-7-null mice have a defect in clearance of intestinal infections, and they succumb
more rapidly and to lower doses of virulent Salmonella typhimurium compared to control mice
(21). Thus, the cryptdin deficiency resulting from defective procryptdin activation is associated
with a measurable deficit in mucosal immunity and increased risk of systemic disease.
In this study, cryptdin biosynthesis was investigated by characterizing details of intracel-
lular procryptdin processing in mouse Paneth cells. The products of in vitro cleavage of
procryptdin-1 and natural procryptdins by MMP-7, the localization of the cryptdin proregion in
the exocytotic pathway, and the extent of procryptdin activation in Paneth cells of adult mice
have now been characterized. Our results show that extensive intracellular procryptdin activa-
tion occurs in mouse Paneth cells and that exposure to bacterial antigens does not induce
procryptdin processing in mice.
EXPERIMENTAL PROCEDURES
Animals and Tissue Preparation -- All procedures on mice were performed in compli-
ance with the policies of the Institutional Animal Care and Use Committee of the University of
California, Irvine. Outbred Swiss mice [(Crl:CD-1)(ICR)BR], 45 day old males, 6 week-old
adult male BALB/cJ and C57/Bl6 mice, 6 week-old pregnant female BALB/cJ mice, and 6
week-old adult male, germ-free Swiss mice were purchased from Charles River Breeding Labo-
ratories, Inc. (North Wilmington, MA). Matrilysin (MMP-7) null mice were 6 to 8 week-old
males backcrossed for 10 generations onto the C57/BL6 background. Mice were housed under
12 h cycles of light and dark and had free access to standard rat chow and water.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 5
For preparation of fetal small intestinal implants, pregnant BALB/cJ mice were eutha-
nized at 15-17 days gestation by injection with 600 g/g body weight of Avertin (500 mg tri-
bromoethanol, 250 mg 2-methyl-2-butanol in 39.5 ml water). Segments (1-2 cm) of proximal
small intestine from each fetus were implanted aseptically under dorsal subcutaneous skin flaps
of individual 6 week old isogenic BALB/cJ male mice (22,23). Approximately 90% of implants
grew and were harvested for isolation of RNA or protein, or they were fixed by immersion in
phosphate-buffered formalin. Fixed tissue was processed into paraffin blocks, sectioned and
stained with hematoxylin-eosin by the Department of Pathology Histology Lab, UCI Medical
Center.
Preparation of small intestinal crypts -- Crypts were prepared by EDTA treatment of
everted small intestinal segments as described (15,24,25,26). Briefly, segments adult mouse
small bowel were agitated in buffered 30 mM EDTA (pH 7.4), and eluted crypts were deposited
by centrifugation and resuspended in ice-cold Ca++
, Mg++
-free buffer. Enteric -defensins derive
exclusively from Paneth cells in crypts (15,27,28). Certain experiments were conducted with
Protease Inhibitor Cocktail Set III from CalBiochem (Costa Mesa, CA) present in all buffers and
solutions to test for the possibility of proteolysis during sample preparation. After protease in-
hibitors were shown to have no effect on procryptdin recovery or on the state of cryptdin activa-
tion (Fig 4C), experiments were conducted in the absence of inhibitors.
Extraction of crypt proteins -- Peptides were prepared by extraction using 30% acetic
acid (28,29). For analysis of peptides from crypt-enriched fractions, crypts were resuspended in
30% acetic acid, sonicated, and extracted overnight at 4oC. Extracts were centrifuged 15 min at
10,000 rpm in the Sorvall SA-600 rotor, supernatants were clarified by centrifugation for 2 h at
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 6
28,000 rpm in the Beckman SW28.1 rotor, and high speed supernatants were diluted 10-fold and
lyophilized (21).
Preparation of Paneth cell secretory granules -- Subcellular fractions enriched for Pa-
neth cell secretory granules were prepared from duodenal and ileal crypts. Crypts deposited by
centrifugation at 700 rpm for 5 min in the Beckman GS-6R centrifuge were resuspended in ~10
ml ice-cold Ca++
, Mg++
-free phosphate-buffered saline (PBS), pH 7.5 (GIBCO, Gaithersburg,
MD), and placed under N2 at 750 psi for 15 min in a Model 1019HC nitrogen cavitation bomb
(Parr Instrument Company, Moline, IL). Cell lysates produced by equilibration to atmospheric
pressure were diluted two-fold with PBS containing 5 mM EDTA and centrifuged at 700 x g for
10 min at 4oC. Low-speed supernatants were reserved, and the deposited cell debris was washed
by resuspension in ice-cold Hanks-EDTA (GIBCO) and centrifugation at 700 x g for 10 min at
4oC. Granules in the combined supernatants were deposited by centrifugation at 27,000 x g for
40 min at 4oC in the Sorvall SA-600 rotor, and granules in the high-speed pellet were washed 2-3
times by resuspension and centrifugation in Hanks-EDTA under the same conditions. Granules
were stored frozen or dissolved immediately in 30% acetic acid and extracted as described
above.
Acid-Urea Polyacrylamide Gel Electrophoresis -- Lyophilized peptide samples were dis-
solved in 20 l of 5% acetic acid containing 3.0 M urea, and electrophoresed on 12.5% acid-urea
polyacrylamide gels (AU-PAGE) for 6 h at 150 V (29). Resolved proteins were visualized by
staining with Coomassie R-250 after fixation in formalin-containing acetic acid-methanol. -
Defensins were identified by their rapid co-migration with authentic mouse cryptdin peptides in
AU-PAGE (> 0.6 x Rf of methyl green dye) as described (30) and confirmed immunochemically
in western blots (15,31).
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 7
Cryptdin-1 prosegment antiserum -- The cryptdin-1 prosegment corresponds to residues
19-58 in preprocryptdin-1 as deduced from cryptdin-1 cDNA (32,33, Fig 1A). The prosegment
(DPIQNTDEET KTEEQPGEDD QAVSVSFGDP EGTSLQEES) was synthesized by Quality
Controlled Biochemicals, Inc., (QCB, Hopkinton, MA). The composition and concentration of
synthetic prosegment was determined by amino acid analysis on a Waters Model 2690 Alliance
Analyzer, and its mass was verified by matrix-assisted laser desorption ionization mode time-of-
flight mass spectrometry (MALDI-TOF MS) on a Voyager-DE instrument (PE-Biosystems, Fos-
ter City, CA) in the UCI Biomedical Protein and Mass Spectrometry Resource Facility. QCB
produced polyclonal anti-cryptdin-1 prosegment antiserum in a sheep by administering 4 dorsal
subcutaneous injections of prosegment conjugated to bovine serum albumin in complete Freund's
adjuvant. Injections were repeated twice, and the antiserum titer was evaluated by ELISA by
QCB. The primary structures of prosegments in all mouse defensin family precursors are highly
conserved (Fig 1A), and the antibody is likely to cross-react with all mouse defensin family pre-
cursors. Rabbit antisera to the CRS1C-1 prosegment (34,35, Fig 1A) react with mouse Paneth
cells specifically (36).
Immunolocalization of cryptdins and prosegments -- Immunoperoxidase staining was
performed by the Department of Pathology Histology Lab at the UCI Medical Center. Paraffin
sections of formalin-fixed mouse small bowel were deparaffinized with xylenes, treated for 30
min with 0.3% H2O2 and washed with water and PBS. Slides were incubated 3 times for 5 min
each in a microwave oven with antigen unmasking solution (Vector Laboratories, Inc., Burlin-
game, CA) then cooled in unmasking solution (Vector) for 30 min at room temperature. After
rinsing with PBS, sections were blocked by incubation with normal goat serum for 30 min, with
Avidin D blocking solution for 15 min, rinsed briefly with PBS, and then incubated with biotin
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 8
blocking solution (Vector) for 15 min. Slides were incubated with a 1:100 dilution of sheep
cryptdin-1 prosegment immune antiserum or with serum from the sheep prior to immunization.
After 30 min, slides were washed 3X with PBS, incubated 30 min with a 1:2000 (28) dilution of
biotinylated donkey anti-sheep IgG and washed as before. After 60 min incubation with
VECTASTAIN ABC peroxidase reagent (Vector), slides were washed, flooded with diamino-
benzidine, washed and counterstained before mounting.
For immunogold co-localization of mature peptide and cryptdin-1 prosegment, samples
of jejunum were fixed with 2% formaldehyde, 0.1% glutaraldehyde in 0.1 M phosphate buffer
(pH 7.4) and embedded in Unicryl (Ted Pella, Inc., Redding CA) at -20oC. Thin sections placed
on formvar and carbon-coated grids were stained with rabbit anti-cryptdin-1 antibody (28),
washed, and reacted with a 1:25 dilution of protein A labeled with 10 nm gold (Ted Pella) as de-
scribed (27). Next, sections were incubated with sheep anti-cryptdin-1 prosegment immune IgG
(1:200) overnight at 4oC. After washing, sections were incubated with donkey anti-sheep IgG
conjugated to 20 nm gold (Ted Pella). Sections were counterstained with uranyl acetate and lead
citrate, examined in a JEOL 100 CX electron microscope and photographed. Equivalent dilu-
tions of preimmune sera provided negative controls in all experiments.
Purification of mouse procryptdins – Recombinant procryptdin-1 was prepared as de-
scribed previously (21). Briefly, mouse procryptdin-1 cDNA cloned in pMalc2 (New England
Biolabs Inc. Beverly, MA) was expressed as an MBP-fusion protein in E. coli BL21(DE3)
CodonPlus cells (Stratagene, La Jolla, CA) that was purified by amylose resin affinity chroma-
tography. Procryptdin-1, released from the fusion protein by digestion with 1 g Factor Xa per
mg fusion protein at 30 C for 48 h (New England Biolabs, Inc., Beverly MA), was separated
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 9
from MBP by C-4 RP-HPLC (Vydac 214TP1010 column, Vydac, Hesperia, CA) and purified to
homogeneity by analytical C-18 RP-HPLC on a Vydac 218TP54 column (21).
For purification of mouse enteric procryptdins, small intestinal protein extracts were pre-
pared from MMP-7 null mice by extraction with 30% acetic acid as described above. Protein
samples were applied to analytical C-18 RP-HPLC columns (Vydac 218TP54) in aqueous 0.1%
trifluoroacetic acid (TFA) and eluted at ~35 min using a 10 to 45% acetonitrile gradient devel-
oped over 55 min. Protein fractions containing apparent procryptdins were analyzed by acid
urea polyacrylamide gel electrophoresis (AU-PAGE) as described (21,29). Procryptdins A-C
were purified to homogeneity by C-18 reverse phase HPLC using a 120 min, 10-40% acetonitrile
gradient, from which cryptdin precursors eluted between 18-30 % acetonitrile (data not shown).
The identification of the purified proteins as cryptdin precursors was achieved by N-
terminal sequencing and MALDI-TOF MS. Peptide concentrations were determined using the
Bradford assay (Bio-Rad Laboratories, Hercules, CA), and the molecular masses of purified pu-
tative procryptdins were determined by MALDI-TOF MS followed by sequencing in the UCI
Biomedical Protein and Mass Spectrometry Resource Facility.
MMP-7 cleavage of mouse procryptdins in vitro -- Recombinant procryptdin-1 and natu-
ral procryptdins were digested with MMP-7, analyzed by AU-PAGE and SDS-PAGE, and mix-
tures of proteolytic digests from MMP-7 cleavage were analyzed by N-terminal sequencing.
Samples (1 g) of recombinant procryptdin-1 (21) and of natural procryptdins A, B, and C puri-
fied from MMP-7-null mice were incubated with equimolar quantities of activated recombinant
human MMP-7 catalytic domain (Chemicon International, Inc., Temecula, CA) in buffer
containing 10 mM N-[2-hydroxyethyl]piperizine-N'-[2-ethanesulfonic acid] (HEPES, pH 7.4),
150 mM NaCl, 5 mM CaCl2 for 24 h at 37oC. Reactions were analyzed in 15% polyacrylamide
Tris-Tricine SDS-PAGE gels (BioRad Laboratories, Hercules, CA). Curiously, proregions or
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 10
Tricine SDS-PAGE gels (BioRad Laboratories, Hercules, CA). Curiously, proregions or frag-
ments of proregions are not seen by routine gel staining methods after digestion with MMP-7,
even though procryptdins and cryptdin peptides stain well (Fig 3A, D.P.S. & A.J.O., unpub-
lished). Samples (~ 200 ng) of complete digests were subjected to 8 cycles of N-terminal pep-
tide sequencing at The UCI Biomedical Protein and Mass Spectrometry Resource Facility.
Western blot analyses of Paneth cell -defensin precursors – Proteins extracted from
adult outbred Swiss mouse crypts were resolved by AU-PAGE, transferred to 0.2 m nitrocellu-
lose membranes, blocked, and incubated with sheep anti-cryptdin-1 prosegment immune IgG
diluted 1:2000 in TBST + 5% nonfat milk at room temperature with agitation (21). Washed
blots were incubated with peroxidase-conjugated donkey anti-sheep Ab diluted 1:5000 in TBST
for 30 min, washed and developed using SuperSignal chemiluminescent substrate (Pierce, Rock-
ford, IL) with a 10-15 min exposure (21). In western blots using rabbit anti-cryptdin-1 peptide
antiserum, goat anti-rabbit IgG was used as the secondary antibody at a 1:20,000 dilution (15).
Assays of Bactericidal Peptide Activity –To measure bactericidal activities, ~1 x 106 ex-
ponentially-growing Escherichia coli ML35 cells were incubated with 5 g/ml synthetic
cryptdin-3 or recombinant cryptdin-4 in 10 mM PIPES buffer (pH 7.4) with quantities of
prosegment, corresponding to preprocryptdin-1 residues 19 to 58 (Fig 1A). After 60 min at 37oC,
20 l of each incubation mixture was diluted 1:2000 with 10 mM PIPES, pH 7.4, and 50 l of
the diluted samples were plated on trypticase soy agar using a Spiral Biotech Autoplate 4000
(Spiral Biotech Inc., Bethesda, MD). Surviving bacteria were quantitated as colony forming
units (CFU/ml) on plates after incubation at 37 C for 12 h.
RESULTS
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 11
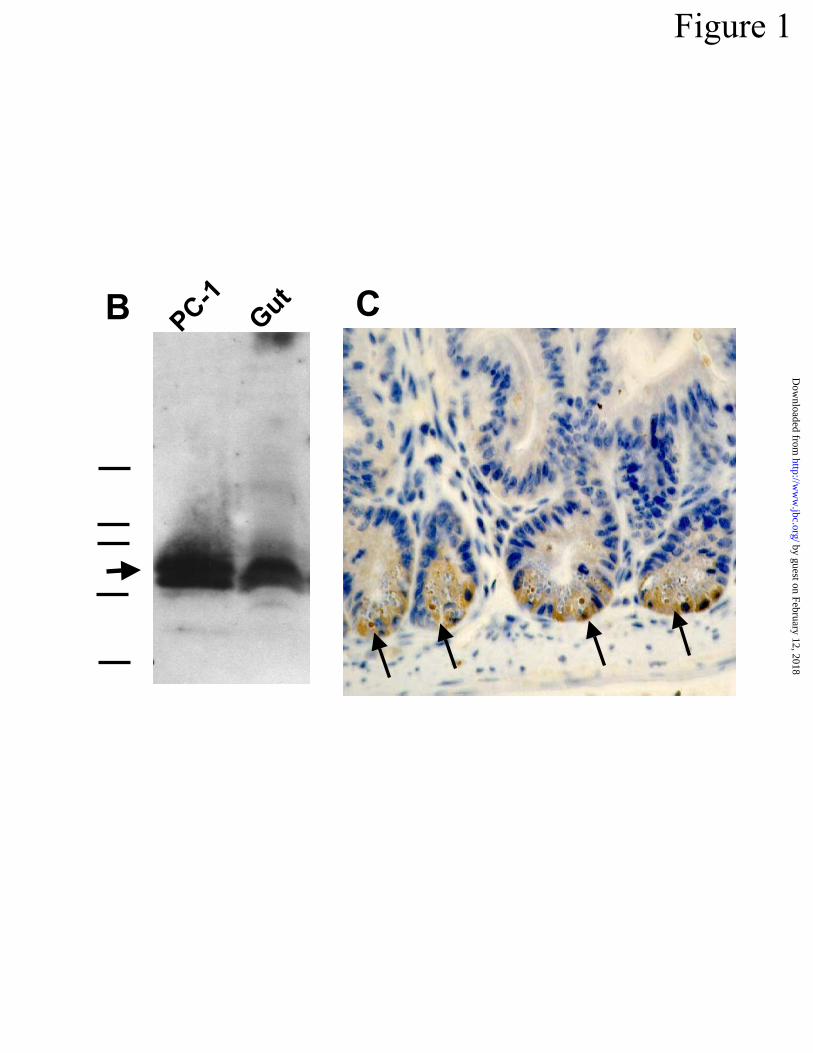
Cryptdin-1 prosegment in mouse Paneth cells – A polyclonal sheep antibody raised
against full-length synthetic cryptdin-1 prosegment (Fig 1A) reacts specifically with
procryptdin-1 and with procryptdins in extracts of mouse small intestinal proteins (Fig 1). West-
ern blot analysis (Experimental Procedures) showed that the antibody was specific for
procryptdins in intestinal protein extracts, which co-migrated with recombinant procryptdin-1
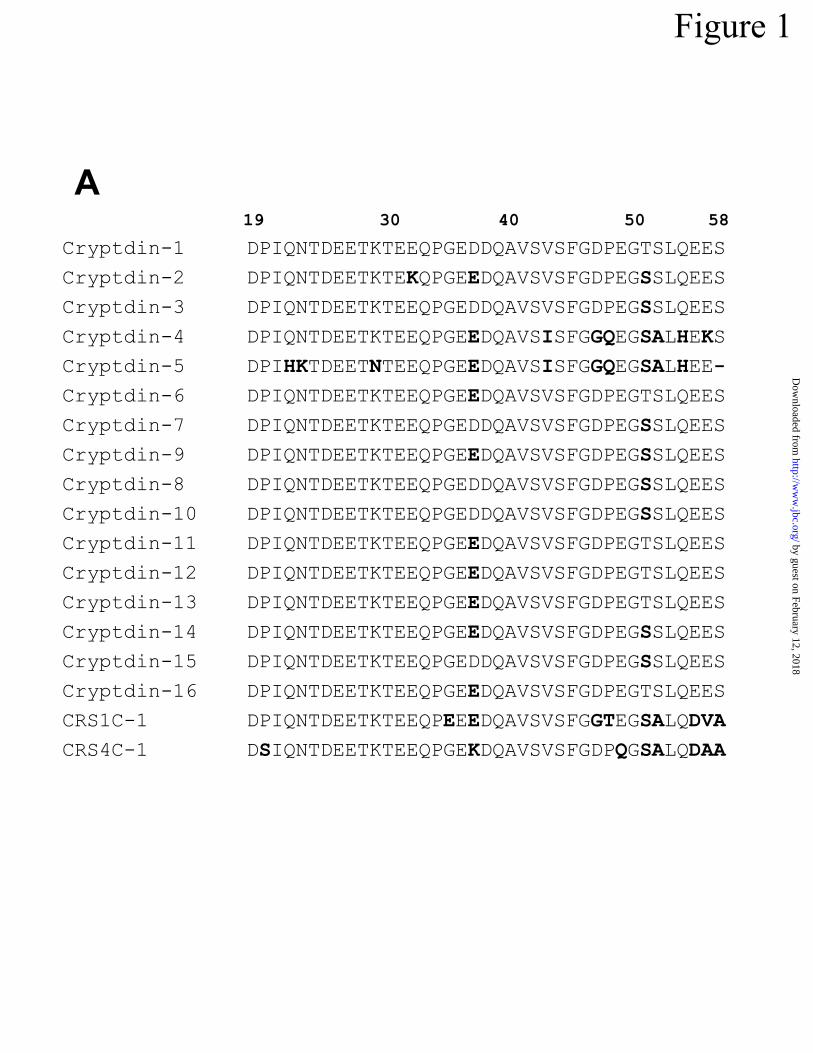
(21, Fig 1B). Because mouse defensin family proregions have extensive sequence similarity (Fig
1A), these data are probably a measure of immunoreactivity with procryptdin-1 and with the
many defensin and defensin-related precursors expressed by mouse Paneth cells (33-35,37).
In small intestine, cryptdin transcripts and peptides previously have been found only in
Paneth cells (9,15,27,28,38-42), and consistent with those findings, immunoperoxidase detection
of the cryptdin-1 prosegment showed that it also is Paneth cell-specific (Fig 1C). This finding is
in agreement with immunolocalization of the related mouse CRS1C prosegment (Fig 1A) using a
rabbit polyclonal antibody to the CRS1C-1 proregion (36, A.J. Ouellette, unpublished). The re-
active cryptdin prosegment antigen appeared to be associated with secretory granules, prompting
immunolocalization studies at the electron microscopic level.
Cryptdin precursors in Paneth cell secretory granules -- -Defensin prosegments as well
as cryptdin peptides are constituents of mouse Paneth cell secretory granules. The subcellular
location of cryptdin prosegments within Paneth cells of mouse mid small bowel was determined
using appropriate gold-conjugated protein A or second antibodies. As shown in Figure 2A-C,
both the prosegment and the cryptdin peptide antibodies reacted strongly and specifically with
Paneth cell granules. Preimmune negative control sera had very low background staining (Fig
2C, inset). With both prosegment and cryptdin peptide antisera, the respective antigens first
were detected in the trans-Golgi of the Paneth cell exocytotic pathway (Fig 2B, inset). Cyto-
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 12
plasmic staining was highly specific for the electron-dense region of secretory granules (Fig
2B,C). The electron-lucent haloes of Paneth cell granules, which contain high levels of GalNAc-
O-linked glycoconjugates (43), showed very little gold staining (Fig 2B,C). All granules were
immunoreactive, and staining was uniformly equivalent regardless of subcellular localization, as
shown by quantitation of gold particles over apical or supranuclear granules (Fig 2D). Despite
showing that the prosegment antibody reacted with Paneth cell granules, these findings did not
distinguish unprocessed procryptdins from soluble proregions generated by MMP-7 proteolysis
of cryptdin precursors. To resolve this question, the products of in vitro MMP-7 hydrolysis of
procryptdins in vitro and the status of procryptdin activation in Paneth cells in vivo were investi-
gated in detail.
Specificity of in vitro procryptdin cleavage by MMP-7 – Because Paneth cell -defensin
processing intermediates had not been characterized, mouse procryptdins were purified from
MMP-7 null mouse small intestine as substrates for analysis of the MMP-7 cleavage products.
MMP-7 null mice are an optimal source for cryptdin precursor purification, because cryptdin
gene expression occurs at wild-type levels, and procryptdins accumulate in MMP-7 deficient Pa-
neth cells (21). Also, studies of natural substrates avoid potential complications of analyzing
possibly misfolded recombinant cryptdin precursors.
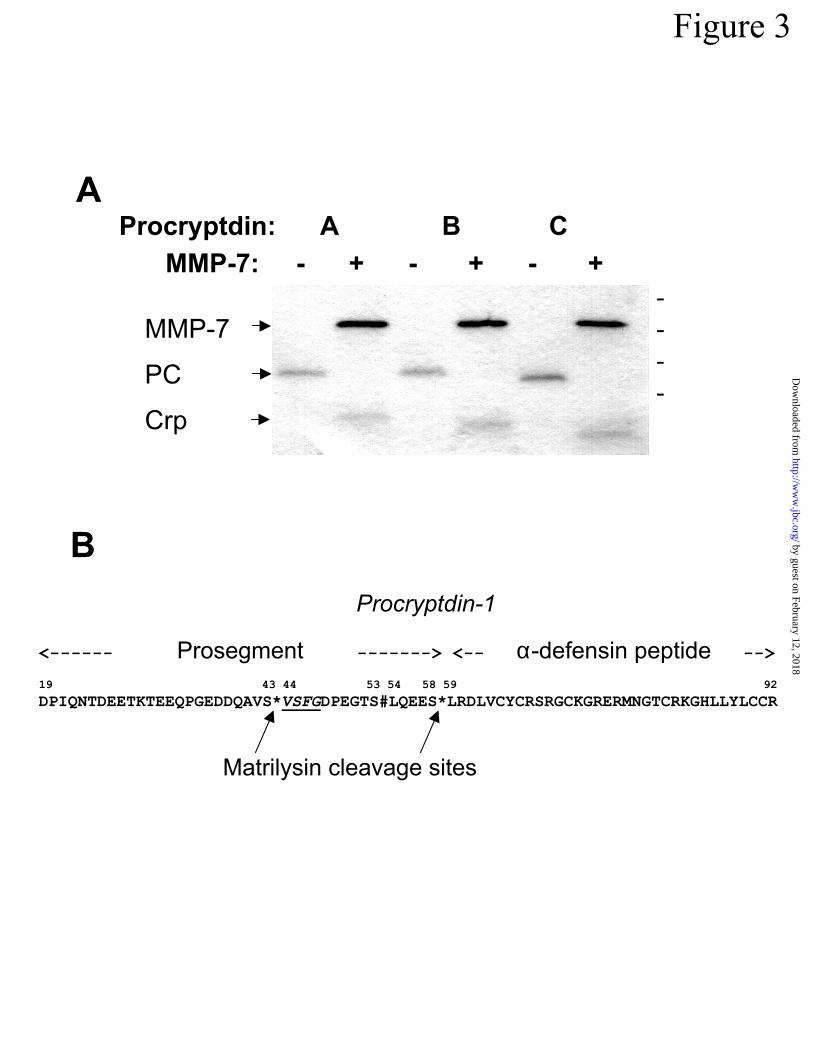
Putative mouse procryptdins A, B, and C were purified to homogeneity by combined C-4
and C-18 RP-HPLC (Fig 3A, Experimental Procedures). Candidate molecules were deduced to
be cryptdin precursors based on elution times from C18 RP-HPLC and co-migration with re-
combinant procryptdin-1 in SDS- (Figs 1B, 3A) and AU-PAGE (not shown). MALDI-TOF MS
of putative procryptdins A-C provided atomic masses of 8543, 8478, and 8277 a.m.u., respec-
tively, values that did not correspond to previously deduced procryptdin sequences (32,33,38).
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 13
Despite this apparent discrepancy, procryptdins A-C were shown to be -defensin precursors by
N-terminal sequencing, because they had N-termini identical to that of procryptdin-1, DPIQNTD
(Table), the consensus N-terminus of all known mouse procryptdins (32,33,38, Fig 1A). Analy-
sis of procryptdins A-C by SDS-PAGE following cleavage with MMP-7 in vitro produced only
one evident primary cleavage product of appropriate mobility for mature -defensin peptides
(Fig 3A).
The peptide bonds cleaved in procryptdins A-C by MMP-7 were determined by direct N-
terminal sequencing of MMP-7 digests of the precursors (Fig 3B). For each putative
procryptdin, only three N-termini were detected besides that of the activated MMP-7 enzyme
(Table, Fig 3B). The first N-terminal sequence was DPIQNTD..., the consensus procryptdin N-
terminus (Table). The second sequence was VSFGDPEG..., an internal cleavage site between
Ser43 and Val44 in the prosegment (Table, Fig 3B). The V-S-*-V-S-F-G sequence flanking the
cleavage site (“*”, Fig 3B) within the prosegment is conserved in all mouse defensin family pre-
cursors (32,33,37, Fig 1A). The third sequence was LRDLV_Y_..., where underscore characters
represent deduced cysteines, and that N-terminal sequence results from proteolysis between
Ser58 and Leu59 in all related procryptdins (21, Fig 3B). LRDLV is the consensus N-terminus
for all cryptdin peptides except cryptdins 4 and 5 (33). The masses determined for procryptdins
A-C were not in concordance with known mouse procryptdins, perhaps because the MMP-7
knockout is on the C57/Bl6 genetic background. Previous clones of procryptdin cDNAs and
genes were from inbred C3H/HeJ and 129/SvJ mouse strains or from outbred Swiss mice, and
these strains may have unreported proregion or cryptdin peptide amino acid substitutions that
differ from those in C57/Bl6 mice. Collectively, these results both confirmed procryptdins A-C
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 14
as cryptdin precursors and defined an MMP-7 catalyzed processing site within the proregions of
these precursors.
Activated -defensins in mouse Paneth cell secretory granules – The colocalization of
prosegments and cryptdins in secretory granules (Figs 1C,2) prompted an evaluation of the proc-
essing status of cryptdin precursors in Paneth cell granules. The distribution of cryptdins and
procryptdins in Paneth cell secretory granules was determined by AU-PAGE western blotting
(15,31). In AU-PAGE, activated -defensins are the most rapidly migrating intestinal peptides
(Experimental Procedures), and they are lacking in MMP-7 null mice (21, Fig 4A). Previously,
only low levels of procryptdins were detected in secretions elicited from Paneth cells by carba-
myl choline exposure (15). Partially purified Paneth cell secretory granules contained abundant
activated cryptdins at levels equivalent to those in intact crypts (Fig 4B, lane 2). Inclusion of a
complex of potent proteinase inhibitors in all solutions and buffers during crypt isolation, gran-
ule sedimentation, and protein extraction (Experimental Procedures) had no effect on the appar-
ent levels of activated cryptdins (Fig 4C), a fact taken as evidence that procryptdin processing
was not caused by experimental manipulation.
The relative distribution of cryptdins to procryptdins was evaluated by western blot
analysis of Paneth cell granule proteins from wild-type and MMP-7 null mice using a cryptdin-1
peptide antibody (15,28). As predicted from previous analyses of whole mouse small bowel pro-
teins (21), Paneth cell granules from MMP-7 null mice lacked rapidly-migrating, activated
cryptdins but contained high levels of procryptdins (Fig 4D, lane 2). In contrast, granule pro-
teins from C57/Bl6 wild-type mice gave strong immunoreactivity at the position of cryptdin mo-
bility, where the signal strength was approximately twice that of the procryptdin region (Fig 4D).
From these considerations and because cryptdins give weaker immune staining in westerns than
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 15
equimolar quantities of procryptdins (M.E. Selsted, personal communication), we estimate that
60-70% of the procryptdins in Paneth cells are processed to functional peptide before secretion
(see Discussion). Because extensive procryptdin activation is intracellular (Fig 4), prosegments
in granules (Fig 2) are subject to secretion, suggesting that proregions might inhibit the bacteri-
cidal activities of cryptdin peptides.
Soluble prosegment neutralizes cryptdin bactericidal activity in vitro – The cryptdin-1
prosegment lacks antimicrobial activity, but it inhibited the bactericidal activity of mature
cryptdin peptides in trans. The ability of soluble cryptdin-1 propeptide, corresponding to resi-
dues 19 to 58 in the cryptdin-1 precursor (Fig 1A), to inhibit the activity of cryptdins 3 and 4 was
tested in bactericidal assays against E. coli ML35. In agreement with the inhibition of myeloid
-defensins by human neutrophil proregions (20), prosegment to cryptdin molar ratios of 0.5 to 1
or greater inhibited both peptides to approximately the same extent (Fig 5). Perhaps as other au-
thors have suggested (17), the acidic proregions (pI ~ 3.4) may neutralize the activity of the cati-
onic defensins by charge neutralization. These in vitro inhibitory activities of secreted proseg-
ments in trans are consistent with a possible cytoprotective role.
Procryptdin activation in germ-free mice – Because MMP-7 is required for procryptdin
activation (21, Fig 4A), we evaluated the extent of cryptdin activation in Paneth cells of germ-
free mice to test whether their basal MMP-7 levels are adequate for cryptdin processing. Germ-
free mice contain less Paneth cell MMP-7 than conventional mice, and monocolonization of
germ-free mice with Bacteroides thetaiotaomicron induces expression of Paneth cell MMP-7 to
levels found in mice harboring conventional microflora (36). Whether raised conventionally or
germ-free, mouse intestinal extracts contained comparable levels of activated cryptdins (Fig 6),
and thus sufficient MMP-7 exists under germ-free conditions to activate cryptdins normally. Al-
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 16
though the mice were germ-free and consumed sterile chow, exposure to dietary bacterial anti-
gens may have been responsible for inducing elevated MMP-7 levels, even in the germ-free
state. For that reason, the level of cryptdin activation was examined in sterile implants of fetal
mouse small intestine grown subcutaneously (22,23).
Activated cryptdins in implants of fetal small intestine -- To test whether procryptdin ac-
tivation requires exposure to bacterial antigens, the state of cryptdin processing was investigated
during Paneth cell ontogeny in BALB/cJ isogenic implants. In mice, the ontogeny of the small
intestinal epithelium occurs during the first three weeks postpartum (45,46). Interestingly, sub-
cutaneous growth of fetal intestinal implants provides conditions that favor epithelial cell differ-
entiation in structures that develop to resemble the morphology of normal adult small intestine
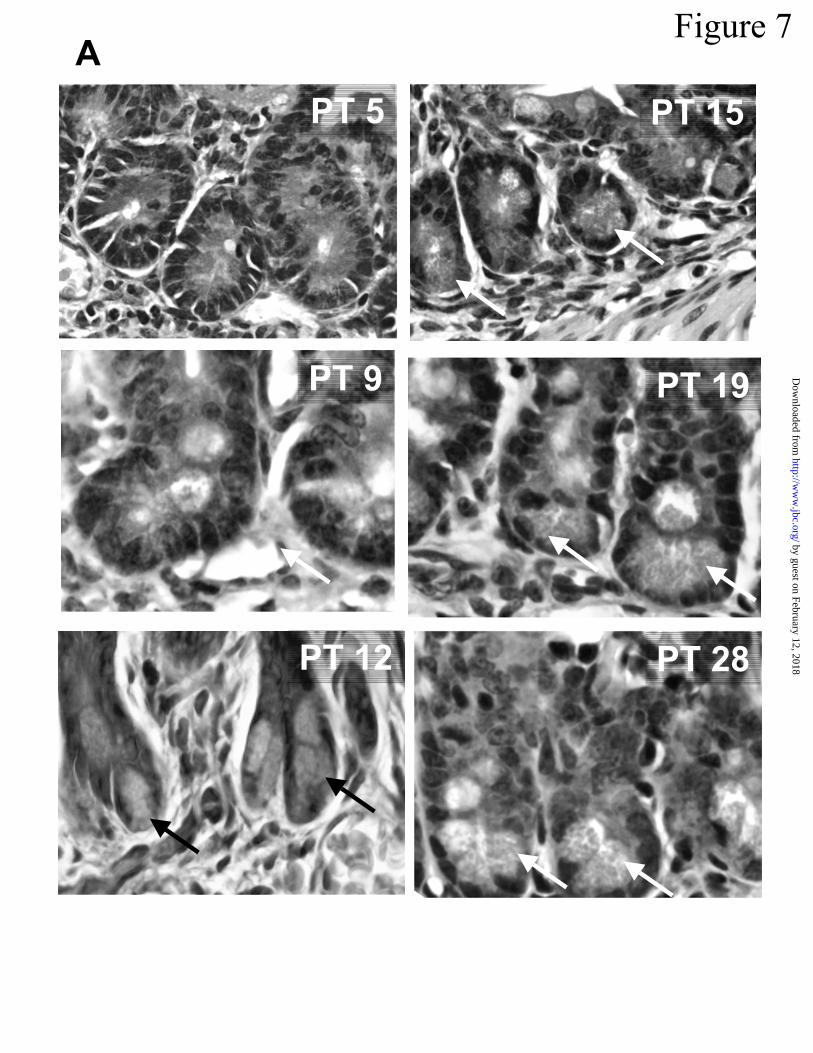
(22,23,47). In our experiments, ~ 90% of implants grew, and Paneth cells were evident at the
base of crypts by approximately 12 days after transplantation (PT 12) as judged by hematoxylin-
eosin staining. Paneth cell granules increased in number and size between PT 12 and PT 19 (Fig
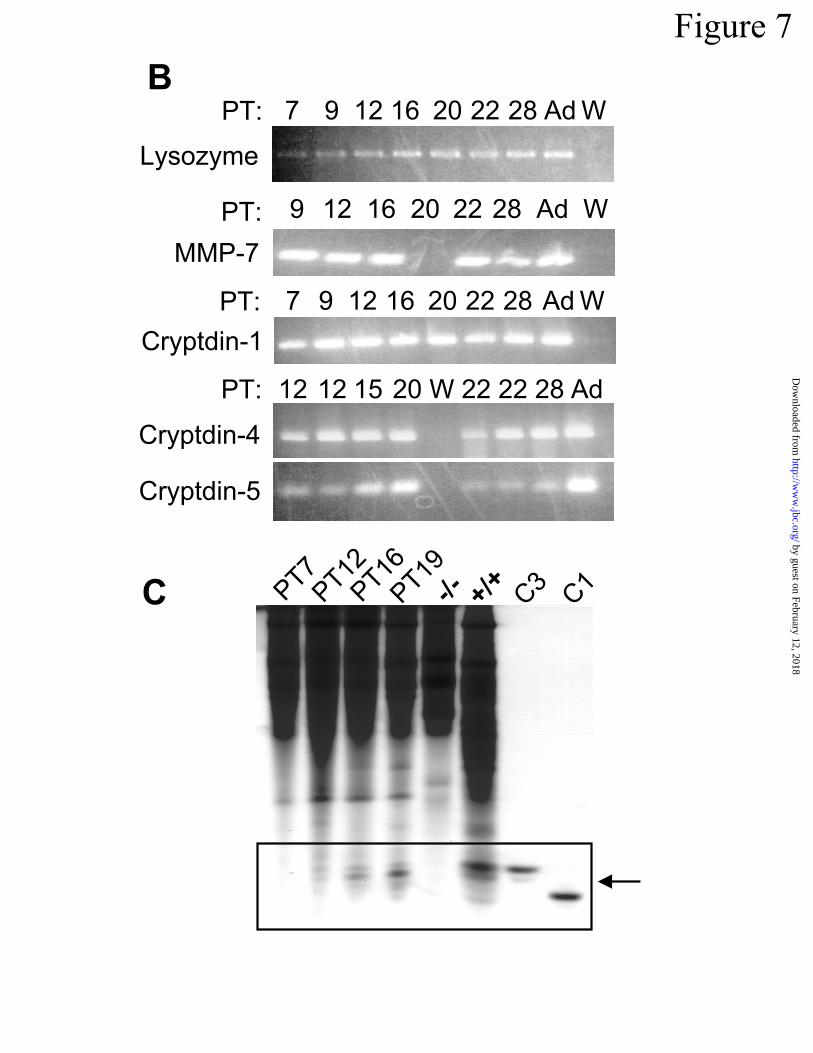
7A). RT-PCR amplification assays for Paneth cell specific mRNAs in PT 7 to PT 28 implant
RNAs detected lysozyme, MMP-7, and cryptdin-1, 4, and 5 mRNAs at all time points (Fig 7B).
AU-PAGE analysis of proteins extracted from implants removed on PT 7 to PT 19 showed that
activated cryptdins were evident from PT 12 onward, resembling adult levels by PT 19. Protein
extracts from PT 7 implants and MMP-7-null mice lacked activated cryptdins (Fig 7). Because
Paneth cells in implants PT 12 or older contain processed cryptdins, luminal exposure to bacte-
rial antigens cannot be required to initiate procryptdin processing. Furthermore, Paneth cells that
are naive to luminal bacterial antigen exposure contain MMP-7 in adequate quantities to provide
functional cryptdins for secretion.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 17
DISCUSSION
In mouse small intestine, a substantial fraction of procryptdin activation occurs in Paneth
cells and prior to secretion. This conclusion is supported by evidence from electrophoretic and
western blot analyses of proteins extracted from subcellular fractions enriched in secretory gran-
ules, where 60-70% of procryptdins exist already activated by MMP-7-dependent proteolytic
cleavage (Fig 3). This value for the fraction of processed precursors represents an overall aver-
age for all cryptdin precursors, with the exception of cryptdins 4 and 5, which do not react with
the cryptdin-1 antibody. Interpretation of these data is complicated, though, by the dynamics of
crypt cell biology and by the ongoing processes of Paneth cell differentiation and granule bio-
genesis in the regulated secretory pathway. For example, our findings do not distinguish be-
tween the procryptdin activation state in mature granules poised for vesicular fusion at the Pa-
neth cell apical membrane from that in nascent granules that are forming in the trans-Golgi. In
addition, we cannot discount the possibility of post-secretory activation of the procryptdin mole-
cules that are secreted. Also, Paneth cells differentiate in crypts over ~ 8 days as they emerge
from the stem cell zone and descend to the base of the crypt (25). The extent of procryptdin
processing in granules of maturing Paneth cells may differ relative to that in fully differentiated
cells at the crypt base.
Questions remain regarding the biology of cryptdin prosegments. For example, the inhi-
bition of cryptdin bactericidal activity by addition of complete prosegment in trans (Fig 5) is
consistent with comparable dose-dependent inhibition of HNP-1 activity by recombinant HNP-1
prosegment [proHNP-120-64] (20) but paradoxical in view of the bactericidal activity of Paneth
cell secretions (15). Perhaps, as suggested for myeloid -defensin prosegments (20), cryptdin
propeptides may interact with additional chaperones to neutralize the potential membrane disrup-
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 18
tive activities of mouse -defensins as they traverse the Paneth cell Golgi stack during granu-
logenesis. Because secretory granules containing activated cryptdins also react with prosegment
antibodies (36, Figs 1,2), the processed proregions or proregion fragments (Fig 3B) are likely to
be released along with activated cryptdins as Paneth cells degranulate. The high bactericidal
peptide activity in Paneth cell secretions (15), however, suggests that proregion inhibitory activ-
ity may be neutralized before or during secretion. Possibly, MMP-7 catalyzed proteolysis of
proregions between Ser43-Val44, Ser53-Leu54 (44), and Ser58-Leu59 during precursor activa-
tion may eliminate the inhibitory capabilities of the complete 39 amino acid proregion tested in
our studies. Also, MMP-7 cleaved procryptdins A-C reproducibly, but additional cleavage steps
may exist as suggested by the isolation of apparent procryptdin processing intermediates with
LQEESLRDLV N-termini from mouse small intestine (44). Those intermediates may be MMP-
7 cleavage products that our sequencing experiments did not detect, or they may be procryptdin
cleavage products of an MMP-7-dependent enteric protease(s) capable of cleaving the precursors
in vivo. Interestingly, preliminary studies of MMP-7 digests of recombinant procryptdin-4 have
not detected the proregion cleavage site between Ser42 and Ile43. Instead, an abundant LHEKS
N-terminal sequence was found, showing that MMP-7 cleaves procryptdin-4 between the Ala52
and Leu53 (Y. Shirafuji & A.J. Ouellette, unpublished), a site that corresponds to the intermedi-
ates purified by Putsep et al. (44). Thus, in vitro, MMP-7 appears to be capable of generating all
known cryptdin processing intermediates.
Paneth cells in germ-free mice have almost undetectable MMP-7 as previously deter-
mined immunohistochemically (36). Nevertheless, the base line level of MMP-7 suffices to in-
sure normal cryptdin activation (Figs 6, 7). Similarly, procryptdin processing in sterile intestinal
implants shows that enough MMP-7 exists (Fig 7B) to activate the pool of cryptdin precursors
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 19
without microbial stimuli in the lumen (Fig 7C). Although RT-PCR amplification of implant
RNAs detected lysozyme, MMP-7, and cryptdin mRNAs in all implants (Fig 7B), cryptdin
RNAs were not detected by northern blot hybridization before PT 12 (data not shown). Similar
findings have been obtained in fetal and newborn mouse intestine, which also lacks Paneth cells
prior to crypt ontogeny and where cryptdins accumulate in apparent secretory cells of the matur-
ing epithelial monolayer (48).
Paneth cell differentiation inherent to small bowel development includes programmed
mechanisms for procryptdin activation and the secretion of functional -defensins without envi-
ronmental cues from the lumen. The evidence in support of this conclusion does not exclude re-
sponses of Paneth cells or their progenitors to proinflammatory mediators, including TNF- or
IFN- that may be released by neighboring epithelial cells or by stromal cells. In fact, Trichi-
nella spiralis infection of mouse small intestine stimulates an increase in Paneth cell numbers as
well as recruitment of intermediate cells to accumulate cryptdins in dense granules, and both
outcomes are mediated by T lymphocytes (49,50). Similarly, Paneth cells increase rapidly in
number when T cells are activated by CD3 ligation, and those events are partly dependent on
TNF- (51). Perhaps proinflammatory cytokines influence the inherent plasticity of the g
testinal epithelium by redirecting lineage determination programs in the short term and modulat-
ing Paneth cell numbers during inflammatory episodes. The responsiveness of MMP-7 biosyn-
thesis and activation to proinflammatory cytokines (52,53) is consistent with this possibi
Regardless of the mechanisms regulating MMP-7 expression in Paneth cells, MMP-7-dependent
procryptdin processing insures the secretion of active -defensins to facilitate innate mucosal
immunity.
astroin-
lity.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 20
ACKNOWLEDGMENTS
Supported by NIH grant DK10184 (D.P.S), DE14040 (C.L.W.), DK15681 (S.J.H.), and
DK44632 (A.J.O.). The thank Drs. Michael E. Selsted, Charles L. Bevins, Dipankar Ghosh, and
William C. Parks for useful discussions and Dana M. Frederick, Khoa Nguyen, Hao Truong, and
Hong Yang for excellent technical assistance. We thank Ms. Tracey Kingsley and Dr. Philip M.
Carpenter, Histology Lab, Department of Pathology, UCI Medical Center for performing histo-
chemical and immunoperoxidase experiments and Dr. Agnes Henschen, UCI Biomedical Protein
and Mass Spectrometry Resource Facility, for peptide sequencing and analysis.
aAbbreviation footnote: MMP-7, matrilysin; MALDI-TOF MS, matrix-assisted laser desorption
ionization mode time-of-flight mass spectrometry; RP-HPLC, reverse-phase high performance
liquid chromatography, RT-PCR, reverse transcriptase polymerase chain reaction; AU-PAGE,
acid urea polyacrylamide gel electrophoresis.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 21
REFERENCES
1. Huttner, K. M., and Bevins, C. L. (1999) Pediatr Res 45, 785-794
2. Lehrer, R. I., and Ganz, T. (1999) Curr Opin Immunol 11, 23-27
3. Morita, Y., Sawada, M., Seno, H., Takaishi, S., Fukuzawa, H., Miyake, N., Hiai, H., and
Chiba, T. (2001) Biochim Biophys Acta 1540, 43-49
4. Nyman, K. M., Ojala, P., Laine, V. J., and Nevalainen, T. J. (2000) J Histochem Cytochem 48,
1469-1478
5. Alper, S. L., Rossmann, H., Wilhelm, S., Stuart-Tilley, A. K., Shmukler, B. E., and Seidler,
U. (1999) Am J Physiol 277, G321-332
6. Bevins, C. L., Martin-Porter, E., and Ganz, T. (1999) Gut 45, 911-915
7. Shimada, O., Ishikawa, H., Tosaka-Shimada, H., Yasuda, T., Kishi, K., and Suzuki, S. (1998)
J Histochem Cytochem 46, 833-840
8. Fernandes, P. R., Samuelson, D. A., Clark, W. R., and Cousins, R. J. (1997) Am J Physiol
272, G751-759
9. Porter, E. M., Liu, L., Oren, A., Anton, P. A., and Ganz, T. (1997) Infect Immun 65, 2389-
2395
10. Ariza, A., Lopez, D., Castella, E. M., Munoz, C., Zujar, M. J., and Mate, J. L. (1996) J Clin
Pathol 49, 474-477
11. Shirafuji, Y., Oono, T., Kanzaki, H., Hirakawa, S., and Arata, J. (1999) Clin Diagn Lab
Immunol 6, 336-340
12. Grandjean, V., Vincent, S., Martin, L., Rassoulzadegan, M., and Cuzin, F. (1997) Biol Reprod
57, 1115-1122
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 22
13. Frye, M., Bargon, J., Dauletbaev, N., Weber, A., Wagner, T. O., and Gropp, R. (2000) J Clin
Pathol 53, 770-773
14. Quayle, A. J., Porter, E. M., Nussbaum, A. A., Wang, Y. M., Brabec, C., Yip, K. P., and Mok,
S. C. (1998) Am J Pathol 152, 1247-1258
15. Ayabe, T., Satchell, D. P., Wilson, C. L., Parks, W. C., Selsted, M. E., and Ouellette, A. J.
(2000) Nat Immunol 1, 113-118
16. Satoh, Y., Habara, Y., Ono, K., and Kanno, T. (1995) Gastroenterology 108, 1345-1356
17. Michaelson, D., Rayner, J., Couto, M., and Ganz, T. (1992) J Leukoc Biol 51, 634-639
18. Valore, E. V., and Ganz, T. (1992) Blood 79, 1538-1544
19. Ganz, T., Liu, L., Valore, E. V., and Oren, A. (1993) Blood 82, 641-650
20. Valore, E. V., Martin, E., Harwig, S. S., and Ganz, T. (1996) J Clin Invest 97, 1624-1629
21. Wilson, C. L., Ouellette, A. J., Satchell, D. P., Ayabe, T., Lopez-Boado, Y. S., Stratman, J. L.,
Hultgren, S. J., Matrisian, L. M., and Parks, W. C. (1999) Science 286, 113-117
22. Rubin, D. C., Swietlicki, E., Roth, K. A., and Gordon, J. I. (1992) J Biol Chem 267, 15122-
15133
23. Rubin, D. C., Roth, K. A., Birkenmeier, E. H., and Gordon, J. I. (1991) J Cell Biol 113, 1183-
1192
24. Cheng, H., and Bjerknes, M. (1996) Anat Rec 244, 78-94
25. Cheng, H. (1974) Am J Anat 141, 521-535
26. Cano-Gauci, D. F., Lualdi, J. C., Ouellette, A. J., Brady, G., Iscove, N. N., and Buick, R. N.
(1993) Exp Cell Res 208, 344-349
27. Ouellette, A. J., Satchell, D. P., Hsieh, M. M., Hagen, S. J., and Selsted, M. E. (2000) J Biol
Chem 275, 33969-33973
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 23
28. Selsted, M. E., Miller, S. I., Henschen, A. H., and Ouellette, A. J. (1992) J Cell Biol 118, 929-
936
29. Selsted, M. E. (1993) Genet Eng (N Y) 15, 131-147
30. Selsted, M. E., and Becker, H. W., 3rd. (1986) Anal Biochem 155, 270-274
31. Wang, M. S., Pang, J. S., and Selsted, M. E. (1997) Anal Biochem 253, 225-230
32. Huttner, K. M., Selsted, M. E., and Ouellette, A. J. (1994) Genomics 19, 448-453
33. Ouellette, A. J., Hsieh, M. M., Nosek, M. T., Cano-Gauci, D. F., Huttner, K. M., Buick, R. N.,
and Selsted, M. E. (1994) Infect Immun 62, 5040-5047
34. Lin, M. Y., Munshi, I. A., and Ouellette, A. J. (1992) Genomics 14, 363-368
35. Ouellette, A. J., and Lualdi, J. C. (1990) J Biol Chem 265, 9831-9837, additions and errata J
Biol Chem 269, 18702 (1994)
36. Lopez-Boado, Y. S., Wilson, C. L., Hooper, L. V., Gordon, J. I., Hultgren, S. J., and Parks,
W. C. (2000) J Cell Biol 148, 1305-1315
37. Huttner, K. M., and Ouellette, A. J. (1994) Genomics 24, 99-109
38. Ouellette, A. J., Darmoul, D., Tran, D., Huttner, K. M., Yuan, J., and Selsted, M. E. (1999)
Infect Immun 67, 6643-6651
39. Ouellette, A. J., Greco, R. M., James, M., Frederick, D., Naftilan, J., and Fallon, J. T. (1989) J
Cell Biol 108, 1687-1695
40. Jones, D. E., and Bevins, C. L. (1993) FEBS Lett 315, 187-192
41. Jones, D. E., and Bevins, C. L. (1992) J Biol Chem 267, 23216-23225
42. Condon, M. R., Viera, A., D'Alessio, M., and Diamond, G. (1999) Infect Immun 67, 4787-
4793
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 24
43. Leis, O., Madrid, J. F., Ballesta, J., and Hernandez, F. (1997) J Histochem Cytochem 45, 285-
293
44. Putsep, K., Axelsson, L. G., Boman, A., Midtvedt, T., Normark, S., Boman, H. G., and
Andersson, M. (2000) J Biol Chem 275, 40478-40482
45. Gordon, J. I., Hooper, L. V., McNevin, M. S., Wong, M., and Bry, L. (1997) Am J Physiol
273, G565-570
46. Simon, T. C., and Gordon, J. I. (1995) Curr Opin Genet Devel 5, 577-586
47. Savidge, T. C., Morey, A. L., Ferguson, D. J., Fleming, K. A., Shmakov, A. N., and Phillips,
A. D. (1995) Differentiation 58, 361-371
48. Darmoul, D., Brown, D., Selsted, M. E., and Ouellette, A. J. (1997) Am J Physiol 272, G197-
206
49. Kamal, M., Wakelin, D., and Mahida, Y. (2001) Parasite 8, S110-113
50. Kamal, M., Wakelin, D., Ouellette, A. J., Smith, A. W., Podolsky, D. K., and Mahida, Y. R.
(2001) Clin Exper Immunol 126, 117-125
51. Alnadjim, Z., Cohn, S. M., Ayabe, T., Biafora, S., Ouellette, A. J., and Barrett, T. A. (2001)
Gastroenterology 120, A21 (Abstract)
52. Powell, W. C., Fingleton, B., Wilson, C. L., Boothby, M., and Matrisian, L. M. (1999) Curr
Biol 9, 1441-1447
53. Wilson, C. L., and Matrisian, L. M. (1996) Int J Biochem Cell Biol 28, 123-136
54. Darmoul, D., and Ouellette, A. J. (1996) Am J Physiol 271, G68-74
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 25
LEGENDS TO FIGURES
Figure 1. Immunochemical analysis of mouse Paneth cell prosegments and -defensin precur-
sors. In A, alignment of prosegment sequences from mouse cryptdin and defensin-related pre-
cursors deduced from intestinal cDNAs (33,37) illustrates their extensive sequence similarity.
Numerals identifying residue positions are based on the deduced preprocryptdin-1 sequence with
position #1 at the initiating Met residue. Amino acid differences with the cryptdin-1 proregion
are noted in bolded characters. In B, antibody to cryptdin-1 prosegment is shown to react with
procryptdin-1 and procryptdins by SDS-PAGE western blot analysis of adult mouse small bowel
protein extracts (Experimental Procedures). PC-1 denotes a lane containing 1 µg of recombinant
procryptdin-1 (21), and the lane labeled “Gut” contained ~500 µg extracted peptides from adult
mouse small intestine (Experimental Procedures). In descending order, the bars at left represent
28, 18, 15.6, 7.6, and 3.55 kDa protein markers. The arrow at left denotes the position of immu-
noreactive procryptdins. In C, the Paneth cell -defensin prosegment was immunolocalized to
Paneth cell secretory granules (Experimental Procedures). Arrows indicate the presence of reac-
tive prosegment antigen in apparent secretory granules of Paneth cells.
Figure 2. Immunostaining of mouse Paneth cells with cryptdin and procryptdin antibodies. In
A, electron micrograph of an adult mouse small intestinal crypt. Paneth cells (P) reside at the
base of the crypt and are surrounded by undifferentiated crypt epithelial cells (C). Paneth cells
contain granules (G), an extensive system of rough endoplasmic reticulum (rER), and
prominent Golgi (Gp). L, Lumen of the crypt. Original magnification = 5,375X; Bar = 5 µm. In
B, the boxed supranuclear region in A is shown at higher magnification to visualize gold-labeled
structures after staining with rabbit anti-cryptdin and protein A gold (10 nm) and sheep anti-
prosegment and anti-sheep IgG gold (20 nm). Note that the Golgi (Gp) and granules (G) in the
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 26
Paneth cell were labeled with both the 10 nm (short arrows) and 20 nm (long arrows) gold
probes, demonstrating that both peptides are present (inset). In contrast, the rough endoplasmic
reticulum (rER) is not labeled. Gold labeling of the Paneth cell Golgi (Gp) is much more exten-
sive than that of the Golgi (Gc) from adjacent undifferentiated crypt epithelial cells (C). Original
magnification = 22,306X. Bar = 1 µm. In C, a high magnification electron micrograph shows
co-localization of prosegment and cryptdin peptides in Paneth cell granules. This section was
labeled with rabbit anti-cryptdin-1/protein A gold (10 nm) and sheep anti-prosegment/anti-sheep
IgG gold (20 nm), and both 10 nm (short arrows) and 20 nm (long arrows) gold particles are pre-
sent in the electron dense (Gd), but not electron lucent (Gl), zones of Paneth cell granules. Inset
shows the relative lack of background staining in Paneth cell granules that were incubated with
preimmune sera prior to incubation with 10 and 20 nM gold-conjugates. Original magnification
= 53,750X. Bar = 0.5 µm. In D, sections labeled with rabbit anti-cryptdin-1/protein A gold (10
nm) were evaluated by counting gold particles in 38 apical and 40 supranuclear granules. The
labeling density of apical and supranuclear granules was identical. Data are presented as means
± S.E.M.
Figure 3. Recognition and cleavage of mouse procryptdins by MMP-7. In A, samples (1 µg) of
procryptdins A, B, and C, purified from MMP-7-null mice, were incubated overnight with (+) or
without (-) 2 µg MMP-7, and samples of digests were resolved by SDS-PAGE and stained with
Gel Code Blue (Pierce). Electrophoretic mobilities of individual components are noted at left in
descending order as follows: MMP-7, matrilysin; PC, purified procryptdins; Crp, MMP-7-
activated cryptdin peptides. Dashes at right denote, in descending order, the position of 28, 18,
15.6, and 7.6 kDa molecular weight markers. In B, the consensus cleavage sites disclosed by
protein sequencing of MMP-7 digests of procryptdins A-C (panel A) are noted by asterisks (*)
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 27
that interrupt the procryptdin-1 sequence, and the pound character (#) shows the N-terminus of
procryptdin intermediates purified from mouse small bowel by Putsep et al. (44) that were not
evident in these in vitro analyses. Numerals above the primary structure refer to residue posi-
tions, with the initiating Met residue in preprocryptdin-1 as residue #1.
Figure 4. Intracellular processing of mouse Paneth cell -defensin precursors. In A: samples
(250 µg) of protein extracts from adult mouse small intestine were resolved by AU-PAGE and
gels were stained with Coomassie Blue (Experimental Procedures). Lanes 1 & 3, extracts from
MMP-7-null mice lack activated defensins (boxed region); lanes 2 & 4, extracts from wild-type,
C57/BL6 mice. In B, activated cryptdins are shown in proteins extracted from combined duode-
nal and ileal Paneth cell granules (Experimental Procedures) after resolution in AU-PAGE, and
stained with Coomassie Blue R250. Lanes: 1, extract from intact crypts; 2, granule extract; C1,
1 µg cryptdin-1; C3, 1 µg cryptdin-3; and C4, 1 µg cryptdin-4. In C, proteins extracted from s
retory granules prepared from adult mouse crypts in the absence (lane 1) or in the presence (lan
2) of Protease Inhibitor Cocktail III (CalBiochem, Experimental Procedures) were subjec
AU-PAGE. Equivalent quantities of protein were electrophoresed and the gel was stained with
Coomassie Blue. Lanes marked C1, C3, C4, contain 1 µg of cryptdin-1, 3, and 4, respectively.
In D, proteins from Paneth cell granules purified from wild-type (lane 1) or MMP-7 null (lane 2)
adult mouse small intestine were run in AU-PAGE, western blotted and probed with anti-
cryptdin-1 antibody. Lanes C1 and C3 contain 1 µg cryptdin-1 and cryptdin-3, respective
all panels, boxed regions denote the positions at which cryptdin peptides migrate in the AU
system. The arrow at right indicates procryptdins.
ec-
e
ted to
ly. In
gel
Figure 5. Cryptdin-1 prosegment neutralizes cryptdin bactericidal activities in trans. Synthetic
prosegment, corresponding to residues 19-58 in preprocryptdin-1 (Fig 1A), was combined with 5
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 28
µg of cryptdin-3 (A) or cryptdin-4 (B) in the molar ratios shown, incubated with ~1 X 106 E. coli
ML35 cells for 60 min at 37oC, and surviving bacteria were determined by colony counts after
overnight growth on semi-solid media (Experimental Procedures). Bars labeled “C” denote bac-
terial survival in the absence of cryptdin peptides, and bars labeled Crp3 (in A) or Crp4 (in B)
show viability after exposure to 5 µg of cryptdin-3 or cryptdin-4, respectively, in the absence of
prosegment.
Figure 6. Activated cryptdins in germ-free mice. Proteins extracted from small intestines of
adult germ-free mice (lanes 1 & 2), mice conventionalized for 1d (lane 3) and 7 d (lanes 4 & 5),
and a conventionally-reared mouse (lane 6) were analyzed in an AU-PAGE gel and stained with
Coomassie Blue. The boxed region denotes the position of cryptdin peptides. Lanes C1, C3,
and C4 contain 1 µg of cryptdin-1, 3, and 4, respectively.
Figure 7. Activated cryptdins in fetal mouse intestinal implants grown subcutaneously. In A,
implanted tissue removed 5 to 28 days after implantation (PT 5 to PT 28) was fixed in buffered
formalin, processed, and stained with hematoxylin & eosin (Experimental Procedures). Arrows
indicate granule-containing Paneth cells in crypts of developed implants. In B, RNA from PT 7
to PT 28 implants was amplified by RT-PCR using primers specific for lysozyme, MMP-7,
cryptdin-1, cryptdin-4, and cryptdin-5 as reported previously (48,54). As in neonatal small
bowel (48), the Paneth cell marker mRNAs are present in the implanted tissues prior to the ap-
pearance of recognizable Paneth cells. Lanes marked “Ad” contain products amplified from to-
tal RNA from adult mouse small bowel, and lanes marked “W” contain equivalent samples of
amplification reactions in which water was substituted for template RNA. In C, samples (700
µg) of implant protein extracts were analyzed by AU-PAGE as in Fig 6. Lanes contain proteins
from implants taken 7 to 19 d after implantation (PT7 to PT19, respectively), intestinal protein
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Ayabe l et al.
Page 29
extracts from MMP-7 null (-/-) and control wild-type (+/+) mice, respectively, and 1 µg
cryptdin-1 (C1) and cryptdin-3 (C3) as noted. The boxed region of the gel shows the position of
activated cryptdin peptides.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
19 30 40 50 58
Cryptdin-1 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGTSLQEES
Cryptdin-2 DPIQNTDEETKTEKQPGEEDQAVSVSFGDPEGSSLQEES
Cryptdin-3 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGSSLQEES
Cryptdin-4 DPIQNTDEETKTEEQPGEEDQAVSISFGGQEGSALHEKS
Cryptdin-5 DPIHKTDEETNTEEQPGEEDQAVSISFGGQEGSALHEE-
Cryptdin-6 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGTSLQEES
Cryptdin-7 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGSSLQEES
Cryptdin-9 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGSSLQEES
Cryptdin-8 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGSSLQEES
Cryptdin-10 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGSSLQEES
Cryptdin-11 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGTSLQEES
Cryptdin-12 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGTSLQEES
Cryptdin-13 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGTSLQEES
Cryptdin-14 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGSSLQEES
Cryptdin-15 DPIQNTDEETKTEEQPGEDDQAVSVSFGDPEGSSLQEES
Cryptdin-16 DPIQNTDEETKTEEQPGEEDQAVSVSFGDPEGTSLQEES
CRS1C-1 DPIQNTDEETKTEEQPEEEDQAVSVSFGGTEGSALQDVA
CRS4C-1 DSIQNTDEETKTEEQPGEKDQAVSVSFGDPQGSALQDAA
Figure 1
A
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
CBPC-1
Gut
Figure 1 by guest on February 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
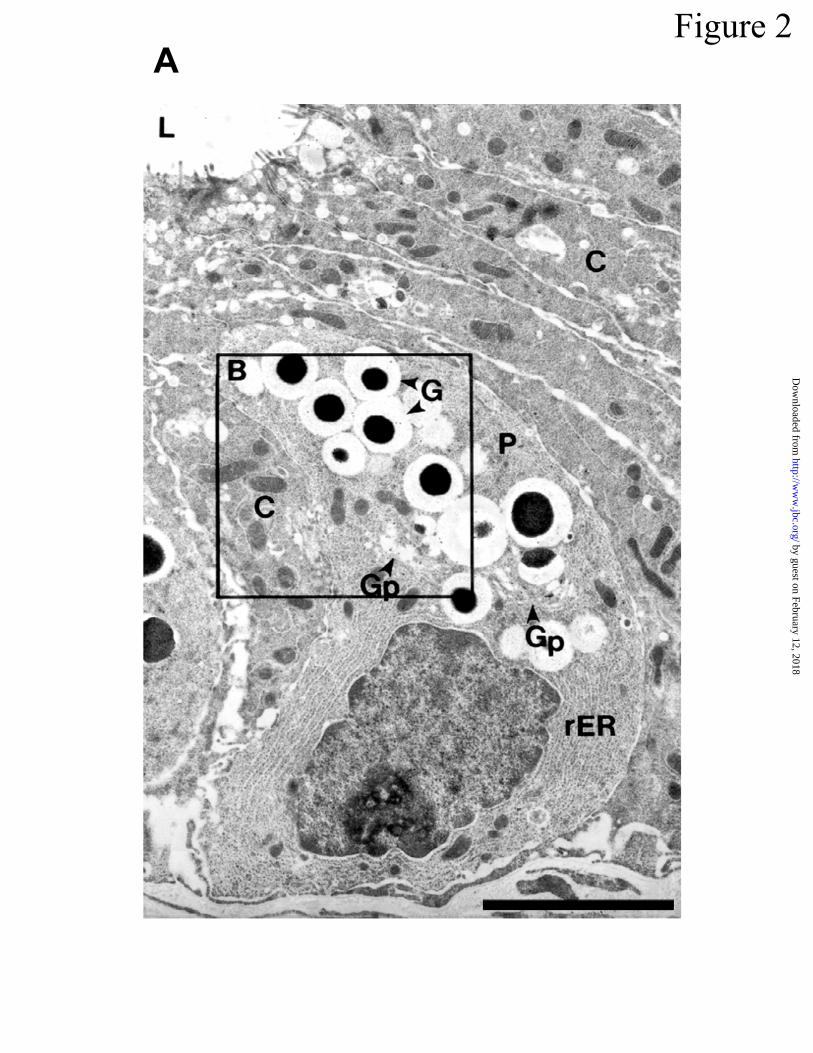
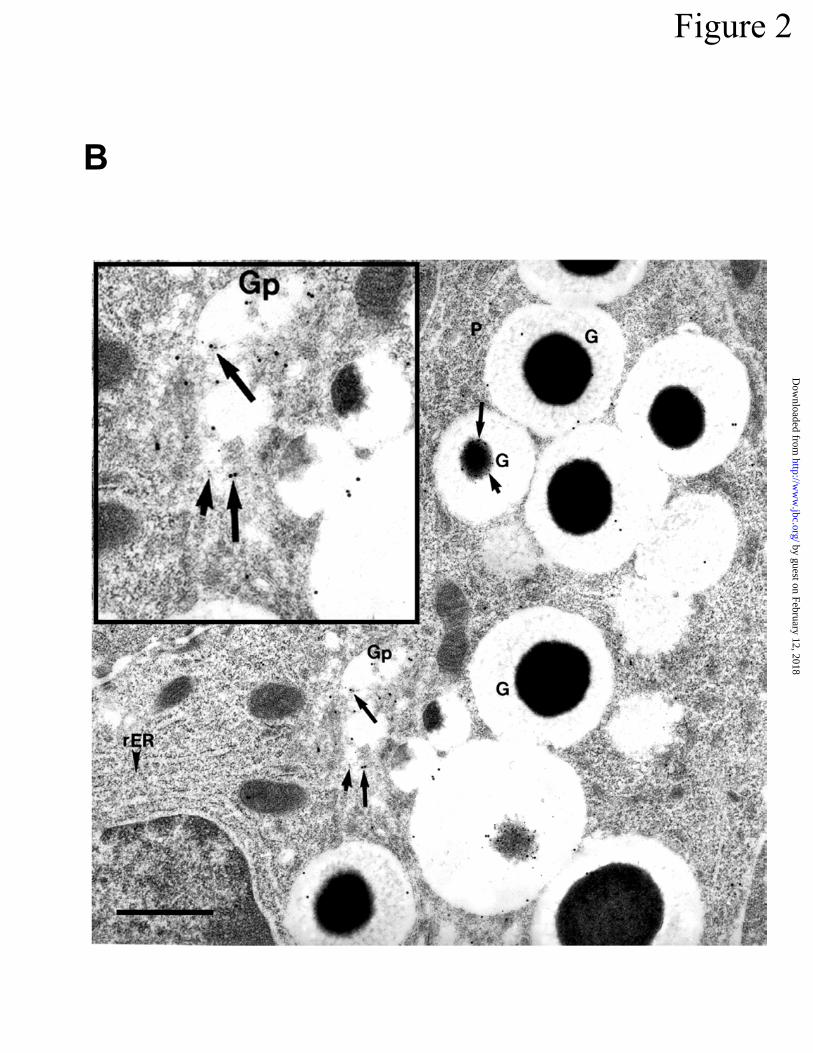
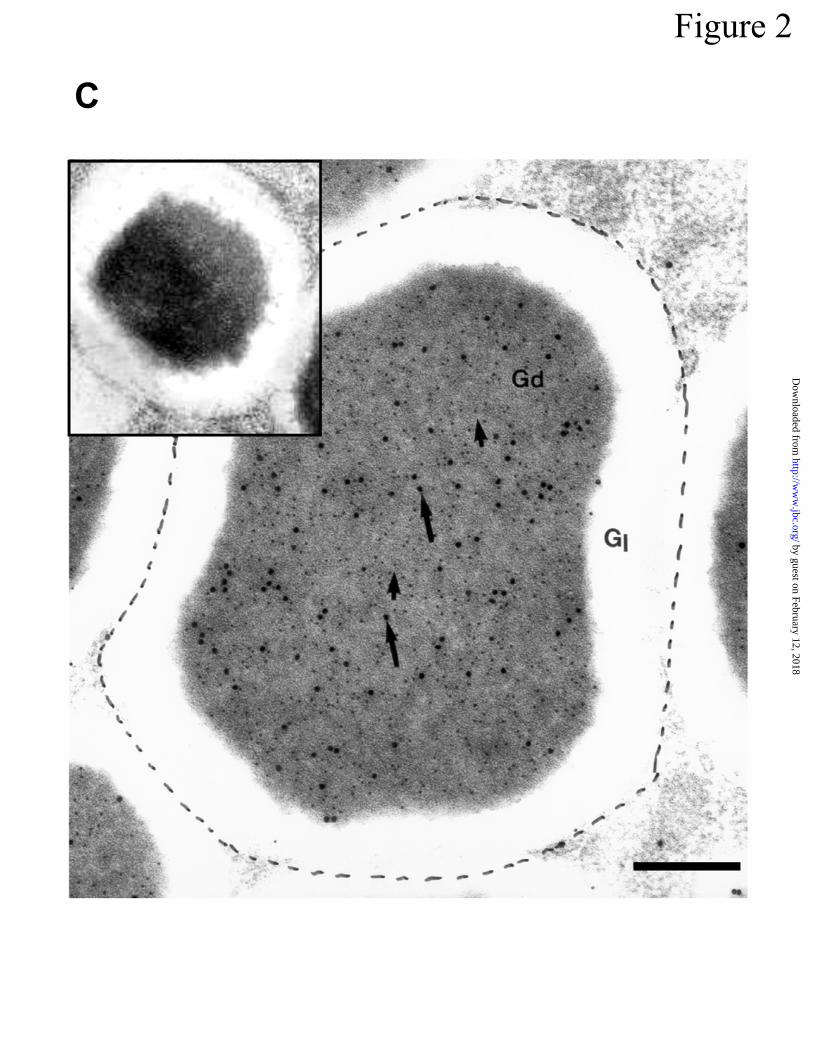
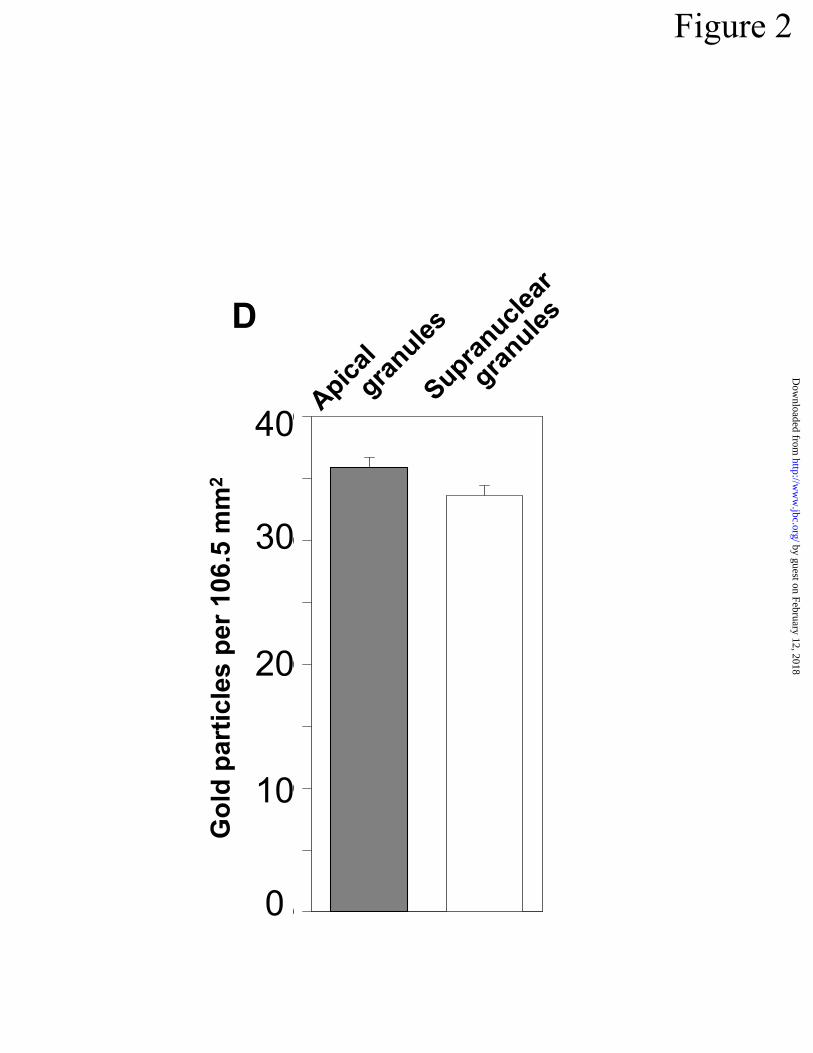
Figure 2
0
0
0
0
0
D
Go
ld p
art
icle
s p
er
106.5
mm
2
40
30
20
10
0
Apic
al
granule
s
Supra
nuclea
r
granule
s
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
Procryptdin-1
<------ Prosegment -------> <-- α-defensin peptide -->
19 43 44 53 54 58 59 92
DPIQNTDEETKTEEQPGEDDQAVS*VSFGDPEGTS#LQEES*LRDLVCYCRSRGCKGRERMNGTCRKGHLLYLCCR
Matrilysin cleavage sites
B
Figure 3
MMP-7
PC
Crp
MMP-7: - + - + - +
Procryptdin: A B C
A
-
-
-
-
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

1 2 3 4
A
B
Figure 4
C
1 2 C1C3C4
1 2 C1 C3
D
1 2 C1 C3C4
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
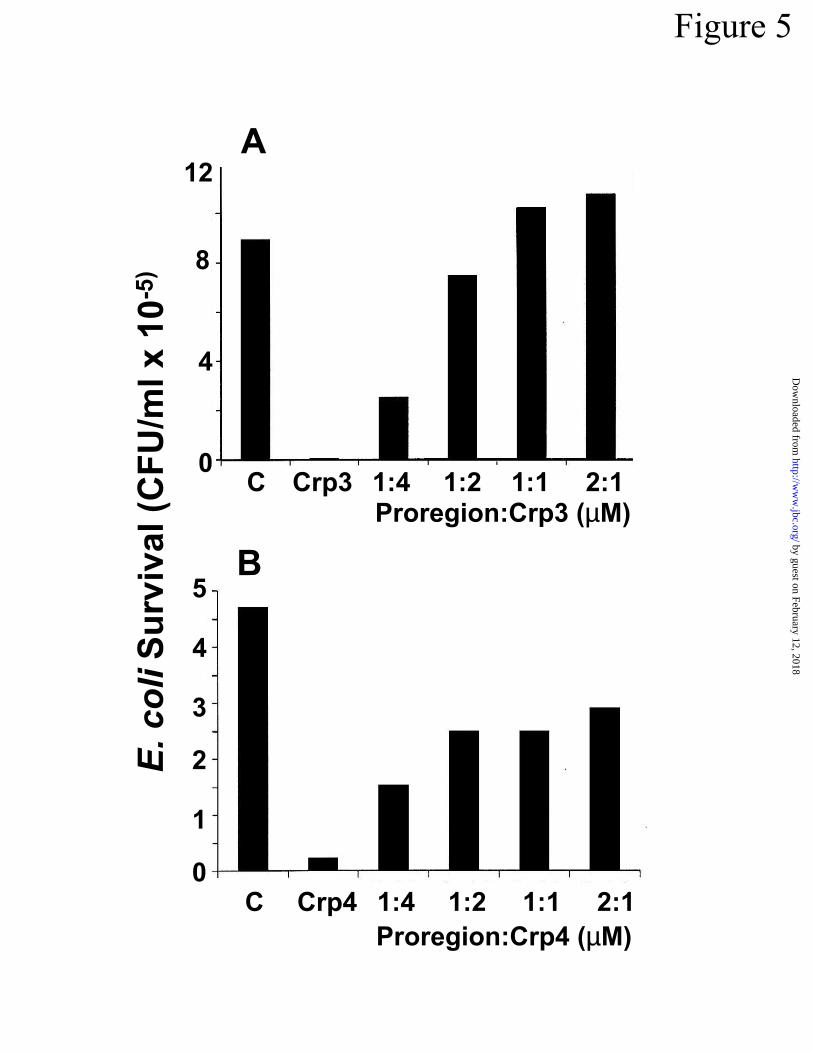
Figure 5
C Crp3 1:4 1:2 1:1 2:1
E. co
liS
urv
ival (C
FU
/ml x 1
0-5
)
12
8
4
0
A
C Crp4 1:4 1:2 1:1 2:1
B5
4
3
2
1
0
Proregion:Crp3 (µM)
Proregion:Crp4 (µM)
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
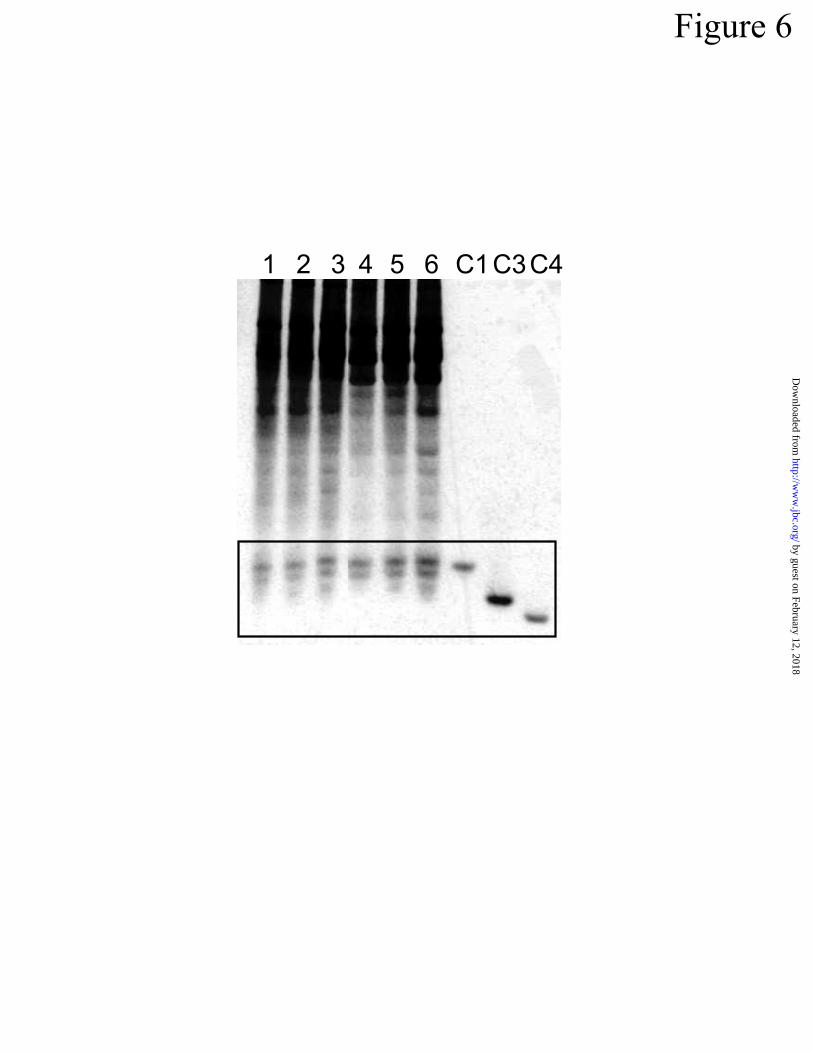
Figure 6
1 2 3 4 5 6 C1C3C4
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
AFigure 7
PT 12
PT 15
PT 19
PT 28
PT 5
PT 9
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
B
Lysozyme
MMP-7
Cryptdin-1
Cryptdin-4
Cryptdin-5
7 9 12 16 20 22 28 Ad W
12 12 15 20 W 22 22 28 Ad
7 9 12 16 20 22 28 Ad W
9 12 16 20 22 28 Ad WPT:
PT:
PT:
PT:
C
Figure 7
PT7
PT12
PT16
PT19
-/- +/+
C1
C3
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Table1
MMP-7 Cleavage Sites in Mouse Cryptdin Precursors Protein Mass Amino Terminal Sequences N-terminus (A.M.U.) Undigested MMP-7 Digests Procryptdin-1 N.D. DPIQNTD DPIQNTD Asp19 VSFGDPEG Val44 LRDLV_Y_ Leu59 Procryptdin A 8543 DPIQNTD DPIQNTD Asp19 VSFGDPEG Val44 LRDLV_Y_ Leu59 Procryptdin B 8478 DPIQNTD DPIQNTD Asp19 VSFGDPEG Val44 LRDLV_Y_ Leu59 Procryptdin C 8277 DPIQNTD DPIQNTD Asp19 VSFGDPEG Val44 LRDLV_Y_ Leu59 1The molecular masses of putative natural cryptdin precursors were determined by
MALDI-TOF mass spectrometry, and N-terminal sequences were determined by Edman deg-
radation of samples before and after digestion with MMP-7 (Experimental Proce-
dures). N.D. = not determined. N-terminal residue assignments are based on the
primary structure of preprocryptdin-1 (Fig 4B) with residue #1 at the initiating
Met.
by guest on February 12, 2018 http://www.jbc.org/ Downloaded from

Wilson, Susan J. Hagen and Andre J. OuelletteTokiyoshi Ayabe, Donald P. Satchell, Patrizia Pesendorfer, Hiroki Tanabe, Carole L.
Activation of Paneth cell alpha-defensins in mouse small intestine
published online December 3, 2001J. Biol. Chem.
10.1074/jbc.M109410200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from