accepted preprint first posted on 11 may 2012 as manuscript joe
TRANSCRIPT

1
Adiponectin inhibits KiSS-1 gene transcription through AMPK and specificity protein-1 in 1
the hypothalamic GT1-7 neurons 2
Jun-Ping Wen1,2, Chune Liu1, Wen-kai Bi1,Ya-Ting Hu1, Qingshi Chen1, Huibing Huang1, Ji-xing 3
Liang1, Lian-tao Li
1, Li-xiang Lin
1 and Gang Chen
1 4
1. Department of Endocrinology, Fujian Provincial Hospital, Fujian Medical University, 5
2. Department of Endocrinology, North Branch of Fujian Provincial Hospital, Fujian Provincial 6
Geriatric Hospital, Fuzhou 350001, China. 7
Corresponding author: Gang Chen, PhD,MD, 8
Department of Endocrinology, Fujian Provincial Hospital, Fujian Medical University, Fuzhou 9
350001, China. 10
Jun-Ping Wen, PhD, MD 11
Department of Endocrinology, Fujian Provincial Hospital, Fujian Medical University, 12
North Branch of Fujian Provincial Hospital, Fujian Provincial Geriatric Hospital, Fuzhou 350001, 13
China. 14
Tel: (86)591-87557768 ext. 3079; Fax: (86)591-87532356 15
Email: [email protected], [email protected] 16
The first four authors contributed to the study equally. 17
Reprint requests to corresponding author. 18
Short title: Adiponectin inhibits KiSS-1 gene transcription. 19
Keywords: Adiponectin; KiSS-1 gene; AMPK; SP1 20
21
22
Page 1 of 42 Accepted Preprint first posted on 11 May 2012 as Manuscript JOE-12-0054
Copyright © 2012 by the Society for Endocrinology.

2
Abstract 1
Adiponectin secreted from adipose tissues plays a role in regulation of energy homeostasis, food 2
intake, and reproduction in the hypothalamus. We have previously demonstrated that adiponectin 3
significantly inhibited GnRH secretion from GT1-7 hypothalamic GnRH neuron cells. In present 4
study, we further investigated the effect of adiponectin on the hypothalamic KiSS-1 gene 5
transcription, which is the upstream signal of GnRH. We found that globular adiponectin(gAd) or 6
AICAR, an artificial AMPK activator, decreased KiSS-1 mRNA transcription and promoter 7
activity. Conversely, inhibition of AMPK by Compound C or AMPKα1-SiRNA, augmented 8
KiSS-1 mRNA transcription and promoter activity. Additionally, gAd and AICAR decreased the 9
translocation of specificity protein-1(SP1) from cytoplasm to nucleus; however, Compound C 10
and AMPKα1-SiRNA played an inverse role. Our experiments in vivo demonstrated that the 11
expression of KiSS-1 mRNA was stimulated 2-fold in the Compound C treated rats and 12
decreased about 60-70% in gAd or AICAR treated rats compared with control group. The 13
numbers of kisspeptin immunopositive neurons in the arcuate nucleus (ARC) region of 14
Sprague-Dawley rats mimicked the same trend seen in KiSS-1 mRNA levels in animal 15
groups with different treatments. In conclusion, our results provide the first evidence that 16
adiponectin reduces KiSS-1 gene transcription in GT1-7 cells through activation of AMPK and 17
subsequently decreased translocation of SP1. 18
Keywords: Adiponectin; KiSS-1 gene; AMPK; SP1 19
20
Page 2 of 42

3
1. Introduction 1
Human puberty is a mystifying process involving a complex series of hormonal events. 2
From a general neuroendocrine perspective, puberty is defined by the full activation of the 3
hypothalamic -pituitary-gonadal (HPG) axis (Grumbach 2002; Ojeda, et al. 1986). The onset of 4
puberty is marked by an increase in the secretion of gonadotropin-releasing hormone (GnRH) 5
from the hypothalamus. A crucial role in puberty and reproduction has been recently assigned to 6
a new hypothalamic signaling pathway, KiSS-1/GPR54 system (Seminara, et al. 2003). KiSS-1 7
gene encodes a premature 145-amino-acid protein called kisspeptin, which is an active ligand of 8
GPR54 in human (Kotani, et al. 2001; Muir, et al. 2001; Ohtaki, et al. 2001). The master role of 9
the KiSS-1/GPR54 system in the central regulation of the HPG axis was first unraveled by the 10
finding that inactivating mutations of the GPR54 gene were associated with the lack of puberty 11
onset and hypogonadotropic hypogonadism both in human and in mice (de Roux, et al. 2003; 12
Seminara et al. 2003), which indicates that KiSS-1 and GPR54 are the new gatekeepers of 13
reproduction. 14
Interestingly, hypothalamic KiSS-1 has been proposed as key molecular conduit for 15
relaying a number of peripheral signals into the GnRH system in the HPG axis. These include not 16
only feedback actions of androgen and estrogen (Navarro, et al. 2004; Smith, et al. 2005) but also 17
metabolic cues (Castellano, et al. 2006). Castellano et al. (Castellano, et al. 2005) showed an 18
interaction between energy status and the hypothalamic KiSS-1 system, which may constitute a 19
target for disruption of pubertal development in conditions of negative energy balance. Several 20
neuropeptides and hormones, including leptin, insulin and kisspeptin, have been reviewed 21
involved in communicating energy status to the reproductive axis (Budak, et al. 2006; Hill, et al. 22
Page 3 of 42

4
2008). Besides, an increasing body of evidence indicates that adiponectin secreted from adipose 1
tissues could also directly regulate reproductive functions. For instance, our prior research had 2
demonstrated that adiponectin inhibited GnRH secretion in the hypothalamus (Wen, et al. 2008). 3
Adiponectin also inhibits both basal and GnRH-stimulated LH secretion in rat and mouse 4
pituitary cells (Lu, et al. 2008; Rodriguez-Pacheco, et al. 2007). Overexpression of adiponectin 5
impairs female fertility in mice (Campos, et al. 2008). And serum adiponectin levels in boys and 6
girls decrease throughout puberty (Bottner, et al. 2004). These findings draw immediately 7
attention to that the changes of the hypothalamic KiSS-1 gene may be mediated by adiponectin 8
according to metabolic status. 9
Adiponectin, also termed Acrp30 or AdipoQ, was identified in 1995-1996 as an adipocyte 10
specific secreted factor with an N-terminal collagenous domain and a C-terminal globular 11
domain (Maeda, et al. 1996). Adiponectin activates down-stream targets such as AMP-activated 12
protein kinase (AMPK), peroxisome proliferator activated receptor-a (PPAR-a), and mitogen 13
activated protein kinase (MAPK) through binding to its two receptors, AdipoR1 and AdipoR2 14
(Kadowaki and Yamauchi 2005; Yamauchi, et al. 2003). The roles of adiponectin plays in 15
antidiabetic (Yamauchi, et al. 2001), antiinflammatory, antiangiogenic, and antiatherosclerotic 16
effects in peripheral tissues (Brakenhielm, et al. 2004; Goldstein and Scalia 2004) have been 17
reported in the early studies. Recent studies showed adiponectin participates in the regulation of 18
energy homeostasis (Kubota, et al. 2007; Wen, et al. 2010) and GnRH secretion through 19
activation AMPK in the hypothalamus (Wen et al. 2008). However, it has not yet been studied 20
whether and how adiponectin participates in the KiSS-1 expression in the hypothalamus. 21
It was clear that KiSS-1 can be regulated at the transcriptional level by specificity protein-1 22
Page 4 of 42

5
(SP1) in different cells (Li, et al. 2007; Mitchell, et al. 2006; Mitchell, et al. 2007). SP1 is a 1
ubiquitously expressed, prototypic C2H2-type zincfinger-containing DNA binding protein that 2
can activate or repress transcription through GC-rich elements in response to physiologic and 3
pathological stimuli. SP1 can be phosphorylated by various kinases at different sites, and the 4
effects of these modifications can lead to its translocation from the cytoplasm to the nucleus, 5
followed by the regulation of gene expression (Majumdar, et al. 2006; Solomon, et al. 2008; Tan 6
and Khachigian 2009). Report(Chu and Ferro 2005) shows that AMPK downstream signaling 7
molecules, such as ERK, can mediate the phosphorytion of transcription factor SP1. 8
In the present study, we investigated the intracellular signaling mechanisms and AMPK 9
activation in adiponectin treated GT1-7 cells, an immortalized hypothalamic KiSS-1gene positive 10
neuron(Chu and Ferro 2005) (Li et al. 2007). We hypothesized that adiponectin dynamically 11
regulated KiSS-1 gene transcription through activation of AMPK and sequentially subcellular 12
compartmentalization of SP1 in GT1-7 cells. To test this hypothesis, we evaluated KiSS-1 gene 13
transcription levels in response to adiponectin, as well as AMPKα1-SiRNA, 14
5-Amino-imidazole-4-carboxamide riboside (AICAR), and Compound C which regulate the 15
activity of AMPK specifically or nonspecifically. In addition, we conducted mechanistic studies, 16
aiming to disclose the mechanism by which adiponectin regulates KiSS-1 gene transcription. 17
18
2. Materials and Methods 19
2.1. Culture of neurons 20
Immortalized hypothalamic neurons GT1-7 cells were provided gratis by Shanghai Clinical 21
Center for Endocrine and Metabolic Diseases, Shanghai Jiaotong University School of Medicine 22
Page 5 of 42

6
(Shanghai, China). GT1-7 cells were maintained in 4.5g/l glucose Dulbecco’s modified Eagle’s 1
medium (DMEM) supplemented with 10% fetal bovine serum, 100 IU/ml penicillin and 100 2
IU/ml streptomycin (at 37°C in a humidified atmosphere of 5% CO2). Cells were deprived of 3
serum overnight by changing incubation to serum-free medium prior to pharmaceutical exposure. 4
And cells were later exposed to globular adiponectin at concentrations of 0.1, 0.2, 1.0 and 5.0 ug 5
/ml, respectively, for 2, 6, 9, 18 and 24 h,, AICAR (0.1 and 1.0mM) for 2 or 24 h, compound C 6
(20 and 40uM) for 2 or 24 h, and mithramycin A (100 and 200nM) for 24 h. Mouse globular 7
adiponectin (Gingras, et al.) was purchased from R&D systems (Minneapolis, MN, USA), 8
5-amino-imidazole-4-carboxamide riboside (AICAR) and Compoud C from Sigma (Fluka 9
Chemie, USA) and mithramycin A from AppliChem (Germany). 10
11
2.2 Animals 12
Female Sprague-Dawley rats (5-6 months) were obtained from slac laboratory animal Inc, 13
shanghai, China. All animals were housed under an environment (12 h light/dark cycles; 20-24°C; 14
humidity 55±10%) with free access to food and water. The female rats having shown two 15
consecutive estrous cycles were bilaterally ovariectomized (OVX), and allowed to recover for 14 16
days. Animals were anaesthetized under intraperitoneal injection of pentobarbital anesthesia and 17
given an intracerebroventricular injection (-0.8 mm posterior, 4.0mm ventral to the bregma and 18
1.5mm lateral from the midline) of colchicines at 75µg/15µl using a protocol adapted from 19
Adachi(Adachi, et al. 2007). Two days later, the rats were randomly assigned into 4 groups (n=5) 20
and given intracerebro -ventricular injection of PBS/DMSO, gAd, AICAR or Compound C, 21
respectively. The method was described in Table 1. Three hours later, animals were perfused 22
Page 6 of 42

7
200ml 0.1M PBS and 200ml cold 4% paraformaldehyde in 0.1M PBS (pH7.4) through left 1
atrium in succession. Brains were immersed in paraformaldehyde for 12 h followed 25% 2
sucrose/0.1M PBS for 2 days. Fixed brain tissue was sectioned coronally at 30um using a sliding 3
microtome. All procedures were approved by the Animal Care Committee of Fujian Medical 4
University in accordance with the National Institutes of Health Guide for Care and Use of 5
Laboratory Animals. 6
7
2.3. Isolation of RNA and cDNA synthesis 8
GT1-7 cells, as well as hypothalamic tissues from female Sprague-Dawley rats were used in 9
this process. For Sprague-Dawley rats, their brain tissues were immediately removed on ice after 10
decapitation, the tissue of the ARC area was then dissected out of the brain with a micro-knife. 11
RNA extraction was conducted according to manufacturer’s instructions using TRIZOL reagent 12
(Invitrogen, Carlsbad CA). All RNA samples went through DNAse enzymatic digestion (Qiagen, 13
Carlsbad CA) to eliminate DNA contamination. In a subsequent analysis, cDNA was synthesized 14
based on 1ug total RNA from each sample, using the TaqMan Reverse Transcription kit 15
(Promega, CA) following manufacturer’s condition (60 min at 42°C, 5 min at 95°C, 5 min at 16
4°C). 17
18
2.4. Real-time quantitative PCR 19
Real-time quantitative PCR was carried out to evaluate KiSS-1 gene expression levels in the 20
GT1-7 cells and the hypothalamic tissues of Sprague-Dawley rats posterior to pharmaceutical 21
exposures (globular adiponectin, AICAR and Compound C). The extraction of RNA and 22
Page 7 of 42

8
synthesis of first-strand cDNA were as mentioned above. The conditions of this real-time PCR 1
were described as follows: 20ul reactions containing 4ul cDNA, 400nM primer pairs, and 10ul 2
SYBR green PCR master were mixed in 96 well plates on an ABI PRISM7300 sequence 3
detection system (Applied Biosystems, Foster City, CA). To normalize the quantity of cDNA in 4
the reaction, a housekeeping gene β-actin was run in all reactions separately under the same 5
experimental conditions. The primers for the PCR are KiSS-1 (forward 6
5’-GCTGCTGCTTCTCCTCTGTG-3’and reverse 5’-CTGCATACCGCGATTCCTT-3’ ) and 7
β-actin (forward 5’-GGCTGTATTCCCCTCCATCG-3’ and reverse 5’-CCAGTTGGTAA 8
CAATGCCATGT-3’). The PCR cycling conditions were 95°C for 10 min, followed by 40 cycles 9
at 95 °C for 5 sec and 60°C for 30 sec. Dissociation curve analysis was also carried out for each 10
gene at the end of the PCR. The raw data from each experiment was used to determine the 11
relative levels of expression for the gene by the -△△CT method. 12
13
2.5. Transient transfection and luciferase assay 14
Cells were cultured on 24-well plates in DMEM supplemented with 10% FBS for at least 24 15
h. When cells approximately reached 60% confluence, transient transfection process started. 16
PGL3-KiSS-1 promoter (provided gratis by Shanghai Clinical Center for Endocrine and 17
Metabolic Diseases, and the human KiSS-1 promoter was cloned from BAC clone RP11-203F10 18
using primers consisting of XhoI and KpnI sites for ligation into the pGL3-basic vector, as 19
previously described ) (Mitchell et al. 2007) was transfected with the aid of Lipofectamine 2000 20
transfection reagent (LF2000; Invitrogen Life Technologies) according to the manufacturer’s 21
protocol. To internally control the transfection efficiency, cells were cotransfected with pRL-TK 22
Page 8 of 42

9
(Promega), an expression vector for Renilla luciferase under the control of a thymidine kinase 1
promoter. This TK promoter would nullify any effect of adiponectin on the expression of 2
luciferase. Premixed DNA and Lipofectamine 2000 were applied to the cells for an 18 h term of 3
transfection. Cells were then incubated with globular adiponectin, AICAR, compound C, 4
AMPKα1-SiRNA, SP1SiRNA and mithramycin A for a further 24 h term. Cell lysates were 5
prepared using reporter lysates buffer (Promega) and other routine techniques such as 6
centrifugation (18,000 rpm for 5 min). Aliquots of cell lysates (20ul) were used to mix with 7
luciferase substrate for luminescent measurement (Berthold Detection Systems). 8
9
2.6. Western blot analysis 10
GT1-7 cells were harvested and lysed in radioimmunoprecipitation (RIPA) buffer 11
containing protease inhibitors for total cell protein extract. Nuclear and cytoplasmic proteins 12
were extracted using a kit (Beyotime Institute of iotechnology, Jiangsu, China). In brief, 13
after the cells were treated as indicated, cytoplasmic proteins were prepared by 14
repeating cycles of freezing and thawing in 0.2 ml of cold uffer A. Nuclear protein was extracted 15
by using ice-cold buffer C. Total cellular extracts (40ug of protein) were separated in 10% 16
SDS-PAGE gels, and then electrophoretically transferred onto polyvinylidene difluoride 17
membranes (Millipore Bedford, MA, USA). The membranes were incubated first with blocking 18
buffer (Tris-buffered saline containing 0.05% Tween 20 and 10% nonfat milk) for 3h at room 19
temperature. Primary polyclonal antibodies against SP1, β-actin, β-tubulin (Santa Cruz 20
Biotechnology, CA, USA) and Lamin B1(Bioworld Technology, USA) were applied overnight at 21
4°C. After three washes, the membranes were subsequently incubated with secondary antibody 22
Page 9 of 42

10
conjugated with HRP (invitrogen) for 2h at room temperature. The blots were visualized by ECL 1
using Kodak X-OMATLS film (Eastman Kodak). Quantitative data were obtained using a 2
computing densitometer and ImageQuant software. 3
4
2.7. Fluorescence microscopy 5
Cells were cultivated on coverslips, washed in PBS three times (5 min each), and fixed in 6
10% paraformaldehyde (10 min). Cells were washed three times in PBS (5 min each), and made 7
permeable in 0.3% Triton X-100 for 15 min, then washed three times in PBS (5 min each). To 8
block nonspecific antibody-binding sites, cells were incubated in 10% goat serum albumin in 9
PBS for 1 h at room temperature. The cells were then incubated overnight at 4°C with rabbit 10
polyclonal antibodies to SP1, washed with PBS for three times (5 min each), and incubated for 11
1h at 37°C with fluorescein isothiocyanate-conjugated goat antibodies to rabbit immunoglobulin 12
G (ZSGB-BIO, China)(green, 1:250) in the dark. The cells were washed three times with PBS 13
again and then were stained by 1-Aminopyridinium iodide for 30 min in the dark, followed by 14
three washes in PBS. Coverslips were air-dried and mounted with anti-fading glycerinum, then 15
mounted on glass slides, finally observed with a fluorescence microsc. 16
17
2.8. Immunohistochemistry 18
Brain tissues of the sacrificed rats were immersed in paraformaldehyde for 12 h followed 19
25% sucrose/0.1M PBS for 2 days. Fixed brain tissue was sectioned coronally at 30um using a 20
sliding microtome. Then the sections through ARC was preserved in ethylene glycol 21
cryoprotectant at -20°C until use. The free-floating sections were rinsed for three times (10 min 22
Page 10 of 42

11
each) in 0.01 M PBS and incubated in 3% H2O2/PBS for 10 min. Before incubation in the 1
primary antiserum, the sections were rinsed and treated in the blocking solution (PBS with 0.3% 2
Triton X-100, 5% sheep serum, and 0.25% bovine serum albumin) at room temperature for 1 h. 3
Then they were incubated at 4°C for 48 h in anti-KiSS-1 rabbit polyclonal primary antibody 4
(1:200,000; antiserum 566, a generous gift from Dr. A. Caraty, NOUZILLY, France). After 5
incubation in primary antiserum, the sections were washed and incubated in secondary 6
biotinylated goat anti-rabbit IgG (1:500; Vector Laboratories, Burlingame, USA) for 2 h. Rinsed 7
again for three times (10 min each), the sections were incubated for 1 h in ABC Elite Kit (Vector 8
Laboratories) at room temperature. Then the free-floating brain sections were stained with DAB/ 9
chromogen solution for 15 min, and sequentially rinsed in PBS for three times (2 min each). At 10
last sections were mounted on gelatinized glass slides, dried, dehydrated in ascending alcohol 11
concentrations, cleared by xylenes, and cover slipped with neutral gum. 12
13
2.9. Cell counting 14
To quantify kisspeptin immunopositive neurons in the ARC, a Nikon microscope attached to a 15
digital camera (NIKON, ECLIPSE-TS100, Japan) was utilized. Cell counting was performed by 16
two researchers. Bilateral cell counts for each section were averaged between researchers. Total 17
cell counts of the ARC (eight sections/rat) were summed and analyzed. 18
19
2.10. Statistical analysis 20
All experiments were repeated at least three times. All analyses were performed using the 21
Statistical Package for Social Sciences (SPSS for Windows, version 16.0; Chicago, IL). Data 22
Page 11 of 42

12
were log-transformed owing to skewed distributions, and were expressed as mean±SD for 1
quantitative values. Unless otherwise noted, data were analyzed using ANOVA followed by 2
Tukey post hoc tests. The significant differences of variate between two groups were assessed by 3
Student’s t tests. P<0.05 was considered statistically significant. 4
5
3. Results 6
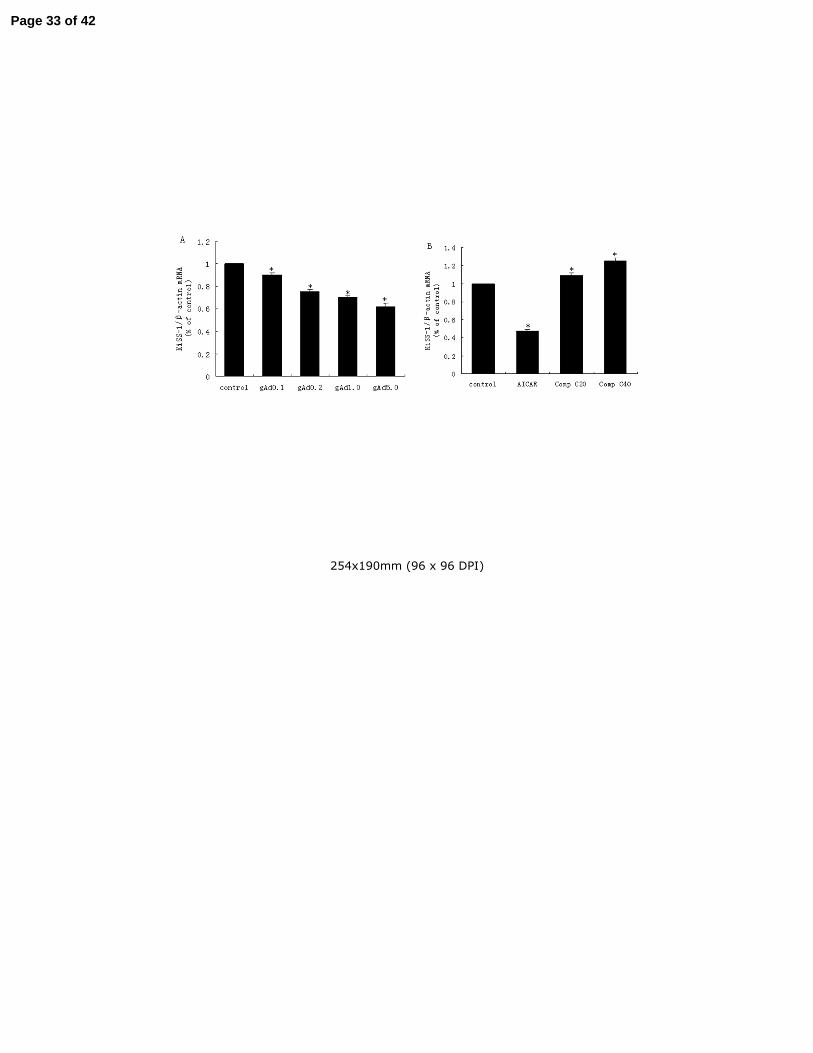
3.1. Globular adiponectin and AMPK inhibited KiSS-1 gene mRNA transcription in GT1-7 cells 7
GT1-7 cells exposed to 0.1, 0.2, 1.0 and 5.0ug/ml globular adiponectin for 2 h have lower 8
transcription of KiSS-1 mRNA determined by real-time PCR (shown in Fig.1A). The inhibition 9
of KiSS-1 mRNA by adiponectin is in a concentration-dependent manner with statistical 10
significance (P<0.05). 11
AMPK is believed to be a major cellular target of adiponectin and its activation has been 12
reported in GT1-7 cells. In order to discover whether AMPK plays a role in the modulation of 13
KiSS-1 mRNA transcription, we used artificial means to activate or inhibit AMPK on GT1-7 14
cells, and followed it with the quantification of mRNA transcription in our assay. As results 15
shown, AICAR (1mM), an AMPK activator, caused an outstanding decrease in KiSS-1 mRNA 16
transcription (Fig.1B) (P<0.001). Conversely, Compound C (20/40uM), a relatively specific 17
AMPK inhibitor, promoted the transcription of KiSS-1 mRNA with a statistical significance in a 18
concentration-dependent manner (P<0.05). 19
20
3.2. Globular adiponectin inhibited KiSS-1 gene promoter activity 21
To examine the transcriptional regulation of KiSS-1 gene in hypothalamus by adiponectin, a 22
Page 12 of 42

13
2-kb full-length KiSS-1 gene promoter (PGL3-KiSS-1 promoter) was transiently transfected into 1
GT1-7 cells. Cells were treated with adiponectin at the concentration of 1.0, 5.0, 10.0ug/ml for 24 2
h, or at 1.0ug/ml for 6, 9, 18, 24 h respectively. The promoter activity was significantly decreased 3
in a time-dependent (Fig.2A) (P<0.05) and concentration-dependent manner (Fig.2B) 4
(P<0.05).The promoter activity was significantly suppressed by the administration of globular 5
adiponectin at the lowest concentration of 1.0ug/ml for a 6 h term of incubation, and the activity 6
was further reduced as time extended. 7
8
3.3. AMPK contributed to KiSS-1 gene promoter activity 9
Following to our discovery of AMPK’s involvement in KiSS-1 gene transcription, it is 10
interesting to know whether AMPK functions as usual under some known pathways, or unusually 11
at the transcriptional level. With these questions unaddressed, we transiently transfected the 12
GT1-7 cells with 2-kb PGL3-KiSS-1 promoter. Then pharmacological regulators of AMPK, 13
AICAR and Compound C as well as the sequence specific interfering RNA of AMPKα1subunit, 14
were applied to the GT1-7 cell culture. The results showed that activation of AMPK by AICAR 15
decreased the promoter activity of KiSS-1gene (Fig.2C) (P<0.05). However, the suppression of 16
AMPK by Compound C or AMPKα1-SiRNA enhanced the promoter activity of KiSS-1gene 17
(Fig.2C,D) (P<0.001). We also pre-treated the GT1-7 cells for 30 min with AICAR (1uM) or 18
Compound C (20uM), and then incubated with globular adiponectin (5ug/ml) for 24 h. As shown 19
in Fig.2E, globular adiponectin (5ug/ml) and AICAR (1mM) decrease KiSS-1gene promoter 20
activity, while Compound C (20uM) augmented promoter activity of KiSS-1gene (P<0.05). 21
22
Page 13 of 42

14
3.4. Globular adiponectin regulated subcellular redistribution of SP1 in GT1-7 cells 1
SP1 functions as an important transcriptional factor by binding to GC-rich elements in 2
response to physiologic and pathological stimuli, and it can either activate or repress gene 3
transcription. Previous studies have shown that the mechanism of its gene regulation partially lies 4
in the translocation of SP1 triggered by its posttranslational modifications. And this translocation, 5
from the cytoplasm to the nucleus, is more commonly described as subcellular 6
compartmentalization. 7
This experiment is to investigate whether the sequential subcellular compartmentalization of 8
SP1 is involved in the regulation of KiSS-1 gene in GT1-7 cells after globular adiponectin 9
administration. Western blotting technique was used to observe the changes of SP1 content 10
before and after subcellular compartmentalization. As shown in Fig.3, the nuclear fraction of SP1 11
decreased by 50% compared with the control group; however, the cytoplasmic fraction increased 12
1.2 fold after global adiponectin incubation (10.0ug/ml) (P<0.05), and this increase is 13
corresponding in a dependent manner to the concentration of globular adiponectin administration. 14
Total SP1 content in whole cells remain constant in GT1-7 cells (P=0.55). 15
16
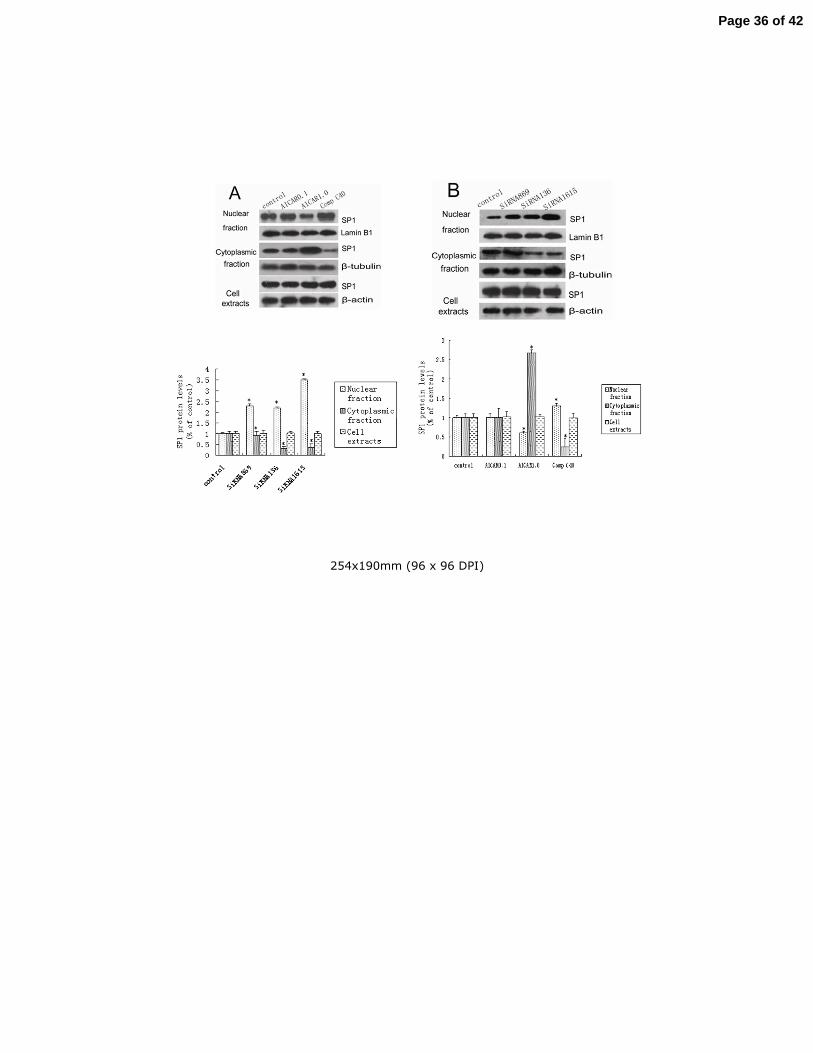
3.5. AMPK regulated subcellular redistribution of SP1 in GT1-7 cells 17
From the above mentioned experiments, it is clear that both AMPK and SP1 are taking roles in 18
regulating KiSS-1 gene transcription. Then our curiosity leads us to question whether AMPK 19
regulates the procedure of subcellular compartmentalization of SP1 or vice versa. We treated the 20
GT1-7 cells with AICAR, Compound C and AMPKα1-SiRNA, then we ran western blot to 21
evaluate the changes of SP1 content upon subcellular compartmentalization. The results showed 22
Page 14 of 42

15
that AICAR decreased the nuclear fraction and increased the cytoplasmic fraction of SP1 content 1
(Fig.4A) (P<0.05). Compound C decreased the cytoplasmic SP1 content and increased the 2
nuclear fraction, so did AMPKα1-SiRNA (P<0.05). As shown in Fig.4, pharmacological 3
regulators of AMPK, AICAR and Compound C, as well as AMPKα1-SiRNA had no effect on the 4
total SP1 content in whole GT1-7 cells (P=0.46). 5
6
3.6. Immunofluorescence analysis supported the regulation of subcellular redistribution of SP1 by 7
AMPK 8
Other than western blot analysis, we also performed immunofluorescence analysis to directly 9
visualize the effect of AMPK on the subcellular redistribution of SP1 protein. After incubation 10
with AICAR or Compound C for 24 h, changes in SP1 content were assessed by using highly 11
specific primary antibodies, which was later couple with fluorescent tagged secondary antibodies 12
(Witte, et al.). As Fig.5 fluorescence demonstrates, in response to AICAR treatment, SP1 in the 13
cytoplasm increased greatly, but the SP1 in the nucleus reduced compared with the control group. 14
To the contrary, Compound C demonstrated a reverse effect on the subcellular localization of SP1. 15
It increased the amount of SP1 in the nucleus and decreased the amount of SP1 in the cytoplasm. 16
These findings were in accordance with the results from western blot. 17
18
3.7. SP1 contributed to KiSS-1 promoter activity 19
We performed luciferase assay to demonstrate the important role of SP1 in the KiSS-1 20
promoter activity. The 2-kb full-length KiSS-1 gene promoter (PGL3-KiSS-1 promoter) was 21
transiently transfected into GT1-7 cells. After one passage, we treated the cells with SP1SiRNA 22
Page 15 of 42

16
and mithramycin A (known as a pharmacological suppressor of SP1). Fig.6 illustrated that 1
suppression of SP1 resulted in a greatly decrease in the activity of KiSS-1 promoter (P<0.05). 2
Our western blot results demonstrated that the SP1SiRNA we used in the luciferase assay 3
reduced the expression of SP1 dramatically in the GT1-7 cells (P<0.05). 4
5
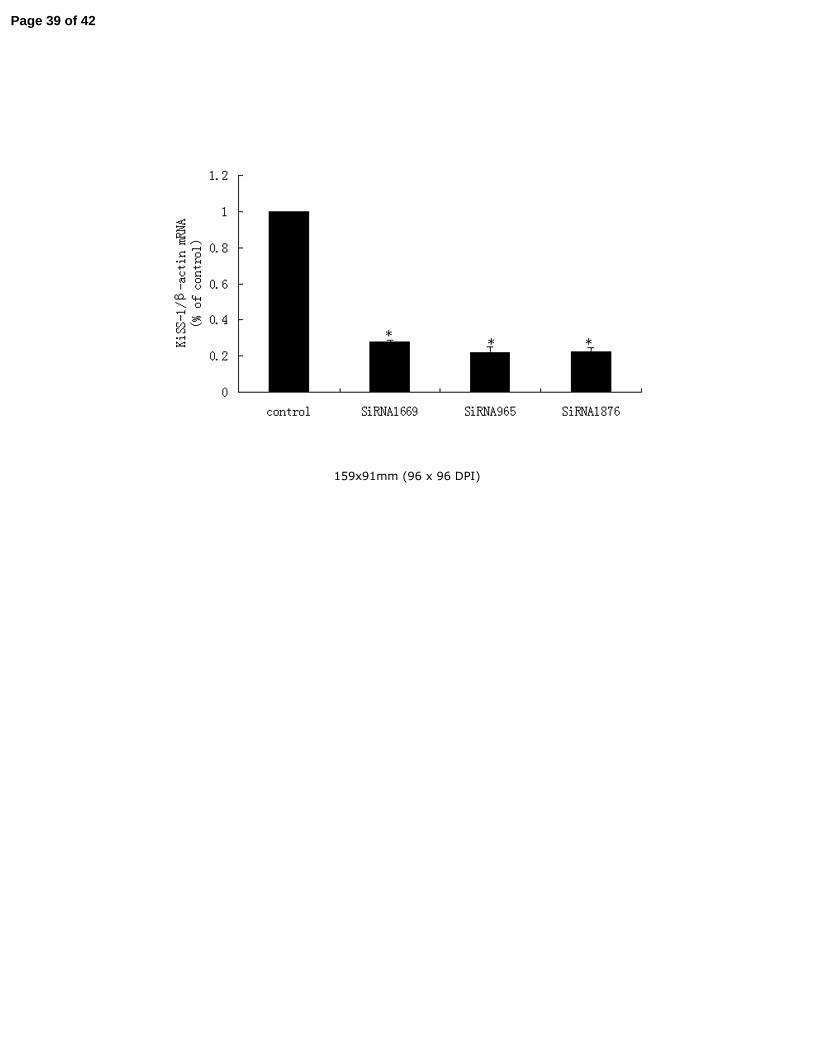
3.8. SP1 promoted KiSS-1 gene mRNA transcription in GT1-7 cells 6
In order to confirm SP1 has an definitely effect on the expression of KiSS-1 mRNA, real time 7
PCR was carried out to detect the KiSS-1 mRNA levels when SP1 SiRNA was transiently 8
transfected into in GT1-7 cells. SP1 protein was definitely knockdown by SP1SiRNA as shown 9
by western blot in Fig.6. As demonstrated in Fig.7, KiSS-1 mRNA transcription was decreased 10
obviously when the expression of transcription factor SP1 was knockdown by SP1SiRNA 11
(P<0.05). 12
13
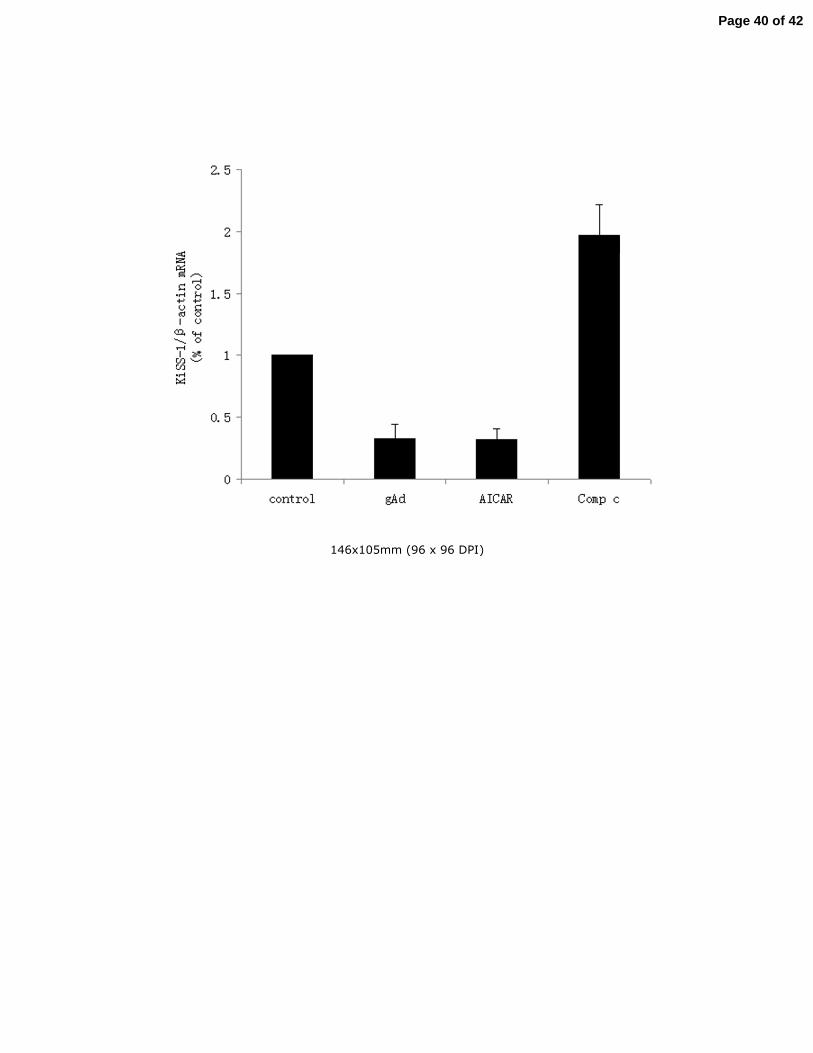
3.9. Globular adiponectin and AMPK contributed to KiSS-1 gene mRNA transcription in vivo 14
Except for the in vitro experiments, we used real-time PCR to detect the expression levels of 15
KiSS-1 mRNA in the hypothalamic tissues of Sprague-Dawley rats given intracerebro 16
-ventricular injection of PBS/DMSO, globular adiponectin, AICAR or Compound C. As 17
shown in Fig.8, the expression of KiSS-1 mRNA was stimulated almost 2-fold in the Compound 18
C treated rats (P<0.05). Correspondently, the KiSS-1 mRNA level decreased about 60-70% in 19
globular adiponectin or AICAR treated rats compared with control group (P<0.05). 20
21
3.10. Globular adiponectin and AMPK had an effect on the expression of kisspeptin in the ARC 22
Page 16 of 42

17
area of Sprague-Dawley rats 1
We performed immunohistochemistry and counted the cell number of kisspeptin 2
immunopositive neurons to demonstrate globular adiponectin and AMPK play an important role 3
in the expression of kisspeptin in the ARC area of Sprague-Dawley rats. As Fig.9 shown, 4
injection of Compound C increased the number of kisspeptin immunopositive neurons in ARC 5
area of Sprague-Dawley rats (Fig.9 D), while injection of globular adiponectin or AICAR 6
decreased the number of kisspeptin immunopositive neurons (Fig.9 B,C) compared with control 7
group treated with PBS/DMSO with significant difference (P<0.05). 8
9
4. Discussion 10
Puberty is a complicated process that has not been comprehensively understood by human 11
beings ourselves. Lots of factors are believed to play important regulatory roles in this process, 12
which include genetic heritages, environmental effects, neuroendocrinological encounters, 13
metabolic status and other growth related factors. Reproductive function is highly sensitive to 14
changes in the metabolic status and energy reserves of an organism, and adverse metabolic 15
conditions are commonly associated with defective reproductive capability involving the 16
alteration in the hypothalamic KiSS-1 system (Castellano et al. 2006). Kisspeptin, ligand of G 17
protein-coupled receptor 54 (GPR54), encoded by the KiSS-1gene, have recently emerged as 18
major gatekeepers of the gonadotropic axis. Shahab et al. (Shahab, et al. 2005) shows that KiSS-1 19
mRNA levels increases with puberty in both male and female monkeys. In intact females, GPR54 20
mRNA levels in the hypothalamus increased 3-fold from the juvenile to midpubertal stage. 21
Hybridization histochemistry indicated robust KiSS-1 and GPR54 mRNA transcription in the 22
Page 17 of 42

18
region of the arcuate nucleus. Dhillo (Dhillo, et al. 2005) reports that systemic administration of 1
kisspeptin to male volunteers results in a significant increase in plasma LH, FSH, and 2
testosterone. Converging evidence (Kauffman 2009; Messager, et al. 2005; Navarro et al. 2004) 3
demonstrates that a key action of KiSS-1/GPR54 system on the hypothalamo-pituitary-gonadal 4
axis occurs directly at the level of GnRH release. It is well established that GPR54-deficient mice 5
and human display phenocopy syndromes characterized as isolated hypogonadotropic 6
hypogonadism (de Roux et al. 2003; Seminara et al. 2003). So KiSS-1/GPR54 represents the 7
molecular ‘‘gatekeepers’’ underlying the onset of pubertal development. 8
Adiponectin is exclusively expressed and secreted by adipose tissue (Kadowaki and 9
Yamauchi 2005). Except for its regulation effect in energy metabolism and insulin sensitivity, 10
adiponectin has also been approved to be involved in antiinflammatory, antiangiogenic, and 11
antiatherosclerotic processes (Brakenhielm et al. 2004; Goldstein and Scalia 2004; Yamauchi et 12
al. 2001). An increasing body of evidence indicates that adiponectin could also directly regulate 13
reproductive functions. Adiponectin also inhibits both basal and GnRH-stimulated LH secretion 14
in rat and mouse pituitary cells (Lu et al. 2008; Rodriguez-Pacheco et al. 2007). Study by 15
Campos et al. sugests that overexpression of adiponectin impairs female fertility in mice 16
(Campos et al. 2008). Bottner’s research shows that serum adiponectin levels in boys and girls 17
decrease throughout puberty (Bottner et al. 2004). 18
In this study, we used globular domain of adiponectin (gAd) to treat GT1-7 cells and found 19
that both transcription levels and promoter activity of KiSS-1gene were significantly inhibited. 20
According to the literature, along with the initiation of puberty, adiponectin levels in men 21
decrease dramatically, while the KiSS-1 gene transcription levels increase rapidly, which also 22
Page 18 of 42

19
shows the intrinsic relationship may exist between adiponectin and KiSS-1 gene before puberty. 1
AMPK plays as an “energy sensor” in vivo as well as an important signaling molecule mediating 2
the physiological role of adiponectin. Adiponectin facilitates glucose uptake and fatty acid 3
oxidation through activation of AMPK in peripheral tissues (Ceddia, et al. 2005; Wu, et al. 2003; 4
Yamauchi, et al. 2002). Lu et al. (Lu et al. 2008) reported that adiponectin could reduce LH 5
secretion in the pituitary gonadotrophic cells in an AMPK-dependent manner. Additionally, our 6
prior research had demonstrated that adiponectin inhibited GnRH secretion via AMPK in GT1-7 7
cells (Wen et al. 2008). Therefore, AMPK may play as a conjunction between energy metabolism 8
and central reproduction, as well as a mediator for the regulation of metabolism and reproduction 9
by adiponectin. In order to investigate whether the AMPK signaling pathway is involved in the 10
regulation of KiSS-1 gene transcription by adiponectin, we used AMPK agonist AICAR as 11
control to observe their impact on the KiSS-1 gene transcription. The results showed that AICAR 12
also inhibited the KiSS-1 gene transcription and their promoter activity obviously, and the 13
AMPK inhibitor Compound C could increase the KiSS-1 gene transcription and enhance the 14
promoter activity. AMPKα1-SiRNA by RNA interference reduced AMPK α catalytic subunit 15
protein level, significantly enhanced the KiSS-1 gene transcription and promoter activity as 16
Compound C did. 17
Report (Chu and Ferro 2005) shows that AMPK downstream signaling molecules such as 18
ERK can mediate the phosphorytion of transcription factor SP1. SP1 has been proved to be a 19
vital regulator of gene transcription of KiSS-1gene (Li et al. 2007; Mitchell et al. 2006; Mitchell 20
et al. 2007). SP1 is a ubiquitously expressed, prototypic C2H2-type zincfinger-containing DNA 21
binding protein. The primary sequence and domain organization of SP1 protein is highly 22
Page 19 of 42

20
conserved. In addition to three zinc fingers, SP1 possesses two glutamine-rich modules that 1
together make up their transcriptional activation domains (AD). Adjacent to the two activation 2
domains is a serine/threonine (S/T)-rich module that may be targeted for posttranslational 3
modifications (e.g. phosphorylation or acetylation). Posttranslational modifications of SP1 not 4
only regulate its subcellular compartmentalization but also modulate its interactions with other 5
transcription factors to regulate gene expression. For example, phosphorylation of SP1 leads to 6
its enhanced nuclear migration and results in the decrease in SP1 DNA binding, and hence 7
suppression of transcription activation of EP4 gene in glioblastoma cells (Kambe, et al. 2008). 8
Therefore, we hypothesized that adiponectin regulates the gene transcription of KiSS-1 and the 9
onset of puberty through AMPK and the activity of its downstream signaling molecules. 10
Through the combination of GC/GT box and the interaction with other proteins, SP1 plays an 11
important role in the enhancement or inhibition of gene transcription and has been implicated in 12
the control of a diverse array of cellular processes exemplified by not limited to cell growth, 13
differentiation, apoptosis, angiogenesis, and immune response (Tan and Khachigian 2009). Study 14
by Mitchell et al. (Mitchell et al. 2007) indicate that co-expression of SP1 and DRIP-130, which 15
is encoded by human chromosome 6q16.3-q23 and has been proved to be SP1-coactivator protein, 16
not only rescues KiSS-1 expression, but also induces an inhibition of the invasive and migratory 17
behavior in highly metastatic melanoma cells. Mitchell et al. (Mitchell et al. 2006) has also 18
demonstrated that SP1 directly interacts with activator protein 2α(AP2)to form transcription 19
complexes at two tandem SP1-binding sites of the promoter of KiSS-1 gene which leads to 20
activation of KiSS-1 transcription in breast cancer cells. Dali Li (Li et al. 2007) has found that 21
there are four proximal GC-rich SP1 binding sites at -188 to -87 of the KiSS-1 promoter and the 22
Page 20 of 42

21
differential regulation of KiSS-1 expression by E2 in different brain regions can be achieved by 1
the different ratio of SP1 to SP3 in the protein complexes. It can be explained by the regulatory 2
mechanism of SP1: 1) SP1 combines to different sites of the KiSS-1 gene promoter, resulting in 3
changes in binding affinity; 2) The role of SP1 in regulation of gene expression depends on its 4
expression level which varies with cell type during different developmental stages; 3) Other 5
factors (as mentioned above involving in DRIP-130, AP-2α, SP3, etc.) can be involved in 6
regulating the transcriptional activity of SP1; 4) as a transcription factor, posttranslational 7
modifications of SP1 play a crucial role for its transcriptional regulatory function. Growing 8
evidence indicates that phosphorylation, acetylation, sumoylation, ubiquitylation, and 9
glycosylation are among the posttranslational modifications that influence the transcriptional 10
activity and stability of SP1. Phosphorylation of SP1 occurs in glutamate-rich region usually 11
enhances transcriptional activation of SP1 in the corresponding pathway, and phosphorylation 12
within the zinc finger always reduces affinity of SP1 to the DNA, thereby reducing 13
transcriptional activity. 14
In our study, the nuclear fraction of SP1 decreased in content while the cytoplasmic fraction 15
increased in a concentration-dependent manner after administration of globular adiponectin in 16
hypothalamic GT1-7 cells. Meanwhile, we didn’t find significant changes in the total cell extracts 17
of SP1 after globular adiponectin treatment in GT1-7 cells. Also in our study, we discovered that 18
AICAR mimicked globular adiponectin in enhancing the cytoplasmic fraction, while Compound 19
C and AMPKα1-SiRNA functioned inversely in the subcellular compartmentalization of SP1 in 20
hypothalamic GT1-7 cells. So we conclude that the changes of subcellular compartmentalization 21
of SP1, instead of the changes of total cell extracts of SP1, take part in the regulations of KiSS-1 22
Page 21 of 42

22
gene transcription by globular adiponectin and AMPK. Our results showed knockdown of SP1 1
expression by SP1SiRNA induced the inhibitory expression of KiSS-1 mRNA in GT1-7 cells. 2
Besides, the luciferase assay demonstrated that SP1 contributed to promote the promoter activity 3
of KiSS-1 gene, since both SP1 SiRNA and mithramycin A, the pharmacological suppressor of 4
SP1, dramatically suppressed the promoter activity of KiSS-1gene. Collectively, these results 5
demonstrated SP1 could activate KiSS-1 promoter activity and promote KiSS-1 mRNA 6
transcription, knockdown of SP1 induced lower promoter activity of KiSS-1 and reduced 7
expression of KiSS-1 mRNA. The suppressive effect of globular adiponectin and AMPK activitor 8
on KiSS-1 promoter activity and the mRNA transcription could be partly explained by the 9
detained transcriptional factor SP1 in the cytoplasm induced by globular adiponectin and AMPK 10
activitor which mimicked the effect on expression of KiSS-1 mRNA and promoter activity when 11
expression of SP1 was knockdown. While, Compound C or AMPKα1-SiRNA, which inhibited 12
the activity of AMPK nonspecifically or specifically, played an inverse role as globular 13
adiponectin and AICAR did. 14
Except for the in vitro studies, our results from in vivo experiments demonstrated injection 15
of globular adiponectin and AICAR could inhibit the expression of KiSS-1 mRNA and decrease 16
the number of kisspeptin immunopositive neurons in the ARC area of Sprague-Dawley rats, 17
while Compound C performed an inverse role as globular adiponectin and AICAR did, the 18
transcription of KiSS mRNA almost doubled and the number of kisspeptin immunopositive 19
neurons in the ARC area also increased significantly in Sprague-Dawley rats treated with 20
Compound C compared with control group. These results further confirmed the results 21
found in vitro studies. 22
Page 22 of 42

23
In summary, our study demonstrated that adiponectin caused the activation of AMPK in the 1
hypothalamic GT1-7 neurons and whereby decreased the translocation of SP1 from cytoplasm to 2
nucleus; as a result, the promoter activity and transcription levels of KiSS-1gene are repressed. 3
This may further influence the hypothalamic release of GnRH and the onset of puberty. To our 4
best knowledge, the data presented here is the first experimental evidence for the characterization 5
of reproductive regulation by adiponectin, as well as the involvement of AMPK and SP1 6
pathways in such a process. However, the limitation of the present study is that only one kind of 7
cell line was utilized, and hence the conclusions of the paper seem a little bit thin. Although we have 8
performed some in vivo studies in Sprague-Dawley rats, further investigations about the 9
molecular mechanism in vivo will be conducted more completely in our future studies. 10
Declaration of interest 11
The authors of this manuscript have nothing to declare. 12
Funding 13
This study was supported by the grants from Natural Science Foundation of China (No.30900510 14
and No.81170774) and Natural Science Foundation of Fujian Province (No. 2011J01127 and 15
No.2011J06012). 16
Acknowledgments 17
Immortalized Gonadotropin-Releasing Hormone Neurons (GT1-7 Cells), the 2-kb 18
full-length KiSS-1 gene promoter (PGL3-KiSS-1 promoter) were provided gratis by Shanghai 19
Clinical Center for Endocrine and Metabolic Diseases, Shanghai Jiaotong University School of 20
Medicine. This manuscript was reviewed for grammatical accuracies by G. Alexander Wen at 21
Lowell General Hospital, Lowell MA. We appreciate all the supports. 22
Page 23 of 42

24
References 1
Adachi S, Yamada S, Takatsu Y, Matsui H, Kinoshita M, Takase K, Sugiura H, Ohtaki T, 2
Matsumoto H, Uenoyama Y, et al. 2007 Involvement of anteroventral periventricular 3
metastin/kisspeptin neurons in estrogen positive feedback action on luteinizing hormone release 4
in female rats. J Reprod Dev 53 367-378. 5
Bottner A, Kratzsch J, Muller G, Kapellen TM, Bluher S, Keller E, Bluher M & Kiess W 2004 6
Gender differences of adiponectin levels develop during the progression of puberty and are 7
related to serum androgen levels. J Clin Endocrinol Metab 89 4053-4061. 8
Brakenhielm E, Veitonmaki N, Cao R, Kihara S, Matsuzawa Y, Zhivotovsky B, Funahashi T & 9
Cao Y 2004 Adiponectin-induced antiangiogenesis and antitumor activity involve 10
caspase-mediated endothelial cell apoptosis. Proc Natl Acad Sci U S A 101 2476-2481. 11
Budak E, Fernandez Sanchez M, Bellver J, Cervero A, Simon C & Pellicer A 2006 Interactions of 12
the hormones leptin, ghrelin, adiponectin, resistin, and PYY3-36 with the reproductive system. 13
Fertil Steril 85 1563-1581. 14
Campos DB, Palin MF, Bordignon V & Murphy BD 2008 The 'beneficial' adipokines in 15
reproduction and fertility. Int J Obes (Lond) 32 223-231. 16
Castellano JM, Navarro VM, Fernandez-Fernandez R, Nogueiras R, Tovar S, Roa J, Vazquez MJ, 17
Vigo E, Casanueva FF, Aguilar E, et al. 2005 Changes in hypothalamic KiSS-1 system and 18
restoration of pubertal activation of the reproductive axis by kisspeptin in undernutrition. 19
Endocrinology 146 3917-3925. 20
Castellano JM, Navarro VM, Fernandez-Fernandez R, Roa J, Vigo E, Pineda R, Dieguez C, 21
Aguilar E, Pinilla L & Tena-Sempere M 2006 Expression of hypothalamic KiSS-1 system and 22
rescue of defective gonadotropic responses by kisspeptin in streptozotocin-induced diabetic male 23
rats. Diabetes 55 2602-2610. 24
Ceddia RB, Somwar R, Maida A, Fang X, Bikopoulos G & Sweeney G 2005 Globular 25
adiponectin increases GLUT4 translocation and glucose uptake but reduces glycogen synthesis in 26
rat skeletal muscle cells. Diabetologia 48 132-139. 27
Chu S & Ferro TJ 2005 Sp1: regulation of gene expression by phosphorylation. Gene 348 1-11. 28
de Roux N, Genin E, Carel JC, Matsuda F, Chaussain JL & Milgrom E 2003 Hypogonadotropic 29
hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc Natl 30
Page 24 of 42

25
Acad Sci U S A 100 10972-10976. 1
Dhillo WS, Chaudhri OB, Patterson M, Thompson EL, Murphy KG, Badman MK, McGowan 2
BM, Amber V, Patel S, Ghatei MA, et al. 2005 Kisspeptin-54 stimulates the 3
hypothalamic-pituitary gonadal axis in human males. J Clin Endocrinol Metab 90 6609-6615. 4
Gingras ME, Masson-Gadais B, Zaniolo K, Leclerc S, Drouin R, Germain L & Guerin SL 2009 5
Differential binding of the transcription factors Sp1, AP-1, and NFI to the promoter of the human 6
alpha5 integrin gene dictates its transcriptional activity. Invest Ophthalmol Vis Sci 50 57-67. 7
Goldstein BJ & Scalia R 2004 Adiponectin: A novel adipokine linking adipocytes and vascular 8
function. J Clin Endocrinol Metab 89 2563-2568. 9
Grumbach MM 2002 The neuroendocrinology of human puberty revisited. Horm Res 57 Suppl 2 10
2-14. 11
Hill JW, Elmquist JK & Elias CF 2008 Hypothalamic pathways linking energy balance and 12
reproduction. Am J Physiol Endocrinol Metab 294 E827-832. 13
Kadowaki T & Yamauchi T 2005 Adiponectin and adiponectin receptors. Endocr Rev 26 14
439-451. 15
Kambe A, Iguchi G, Moon Y, Kamitani H, Watanabe T & Eling TE 2008 Regulation of EP4 16
expression via the Sp-1 transcription factor: inhibition of expression by anti-cancer agents. 17
Biochim Biophys Acta 1783 1211-1219. 18
Kauffman AS 2009 Sexual differentiation and the Kiss1 system: hormonal and developmental 19
considerations. Peptides 30 83-93. 20
Kotani M, Detheux M, Vandenbogaerde A, Communi D, Vanderwinden JM, Le Poul E, Brezillon 21
S, Tyldesley R, Suarez-Huerta N, Vandeput F, et al. 2001 The metastasis suppressor gene KiSS-1 22
encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J Biol 23
Chem 276 34631-34636. 24
Kubota N, Yano W, Kubota T, Yamauchi T, Itoh S, Kumagai H, Kozono H, Takamoto I, Okamoto 25
S, Shiuchi T, et al. 2007 Adiponectin stimulates AMP-activated protein kinase in the 26
hypothalamus and increases food intake. Cell Metab 6 55-68. 27
Li D, Mitchell D, Luo J, Yi Z, Cho SG, Guo J, Li X, Ning G, Wu X & Liu M 2007 Estrogen 28
regulates KiSS1 gene expression through estrogen receptor alpha and SP protein complexes. 29
Endocrinology 148 4821-4828. 30
Page 25 of 42

26
Lu M, Tang Q, Olefsky JM, Mellon PL & Webster NJ 2008 Adiponectin activates adenosine 1
monophosphate-activated protein kinase and decreases luteinizing hormone secretion in LbetaT2 2
gonadotropes. Mol Endocrinol 22 760-771. 3
Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y & Matsubara K 1996 cDNA 4
cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most 5
abundant Gene transcript 1). Biochem Biophys Res Commun 221 286-289. 6
Majumdar G, Harrington A, Hungerford J, Martinez-Hernandez A, Gerling IC, Raghow R & 7
Solomon S 2006 Insulin dynamically regulates calmodulin gene expression by sequential 8
o-glycosylation and phosphorylation of sp1 and its subcellular compartmentalization in liver cells. 9
J Biol Chem 281 3642-3650. 10
Messager S, Chatzidaki EE, Ma D, Hendrick AG, Zahn D, Dixon J, Thresher RR, Malinge I, 11
Lomet D, Carlton MB, et al. 2005 Kisspeptin directly stimulates gonadotropin-releasing hormone 12
release via G protein-coupled receptor 54. Proc Natl Acad Sci U S A 102 1761-1766. 13
Mitchell DC, Abdelrahim M, Weng J, Stafford LJ, Safe S, Bar-Eli M & Liu M 2006 Regulation 14
of KiSS-1 metastasis suppressor gene expression in breast cancer cells by direct interaction of 15
transcription factors activator protein-2alpha and specificity protein-1. J Biol Chem 281 51-58. 16
Mitchell DC, Stafford LJ, Li D, Bar-Eli M & Liu M 2007 Transcriptional regulation of KiSS-1 17
gene expression in metastatic melanoma by specificity protein-1 and its coactivator DRIP-130. 18
Oncogene 26 1739-1747. 19
Muir AI, Chamberlain L, Elshourbagy NA, Michalovich D, Moore DJ, Calamari A, Szekeres PG, 20
Sarau HM, Chambers JK, Murdock P, et al. 2001 AXOR12, a novel human G protein-coupled 21
receptor, activated by the peptide KiSS-1. J Biol Chem 276 28969-28975. 22
Navarro VM, Castellano JM, Fernandez-Fernandez R, Barreiro ML, Roa J, Sanchez-Criado JE, 23
Aguilar E, Dieguez C, Pinilla L & Tena-Sempere M 2004 Developmental and hormonally 24
regulated messenger ribonucleic acid expression of KiSS-1 and its putative receptor, GPR54, in 25
rat hypothalamus and potent luteinizing hormone-releasing activity of KiSS-1 peptide. 26
Endocrinology 145 4565-4574. 27
Ohtaki T, Shintani Y, Honda S, Matsumoto H, Hori A, Kanehashi K, Terao Y, Kumano S, Takatsu 28
Y, Masuda Y, et al. 2001 Metastasis suppressor gene KiSS-1 encodes peptide ligand of a 29
G-protein-coupled receptor. Nature 411 613-617. 30
Page 26 of 42

27
Ojeda SR, Urbanski HF & Ahmed CE 1986 The onset of female puberty: studies in the rat. 1
Recent Prog Horm Res 42 385-442. 2
Rodriguez-Pacheco F, Martinez-Fuentes AJ, Tovar S, Pinilla L, Tena-Sempere M, Dieguez C, 3
Castano JP & Malagon MM 2007 Regulation of pituitary cell function by adiponectin. 4
Endocrinology 148 401-410. 5
Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Jr., Shagoury JK, Bo-Abbas 6
Y, Kuohung W, Schwinof KM, Hendrick AG, et al. 2003 The GPR54 gene as a regulator of 7
puberty. N Engl J Med 349 1614-1627. 8
Shahab M, Mastronardi C, Seminara SB, Crowley WF, Ojeda SR & Plant TM 2005 Increased 9
hypothalamic GPR54 signaling: a potential mechanism for initiation of puberty in primates. Proc 10
Natl Acad Sci U S A 102 2129-2134. 11
Smith JT, Dungan HM, Stoll EA, Gottsch ML, Braun RE, Eacker SM, Clifton DK & Steiner RA 12
2005 Differential regulation of KiSS-1 mRNA expression by sex steroids in the brain of the male 13
mouse. Endocrinology 146 2976-2984. 14
Solomon SS, Majumdar G, Martinez-Hernandez A & Raghow R 2008 A critical role of Sp1 15
transcription factor in regulating gene expression in response to insulin and other hormones. Life 16
Sci 83 305-312. 17
Tan NY & Khachigian LM 2009 Sp1 phosphorylation and its regulation of gene transcription. 18
Mol Cell Biol 29 2483-2488. 19
Wen JP, Liu CE, Hu YT, Chen G & Lin LX 2010 Globular adiponectin regulates energy 20
homeostasis through AMP-activated protein kinase-acetyl-CoA carboxylase (AMPK/ACC) 21
pathway in the hypothalamus. Mol Cell Biochem 344 109-115. 22
Wen JP, Lv WS, Yang J, Nie AF, Cheng XB, Yang Y, Ge Y, Li XY & Ning G 2008 Globular 23
adiponectin inhibits GnRH secretion from GT1-7 hypothalamic GnRH neurons by induction of 24
hyperpolarization of membrane potential. Biochem Biophys Res Commun 371 756-761. 25
Witte JS, Greenland S, Haile RW & Bird CL 1994 Hierarchical regression analysis applied to a 26
study of multiple dietary exposures and breast cancer. Epidemiology 5 612-621. 27
Wu X, Motoshima H, Mahadev K, Stalker TJ, Scalia R & Goldstein BJ 2003 Involvement of 28
AMP-activated protein kinase in glucose uptake stimulated by the globular domain of 29
adiponectin in primary rat adipocytes. Diabetes 52 1355-1363. 30
Page 27 of 42

28
Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, 1
Tsunoda M, et al. 2003 Cloning of adiponectin receptors that mediate antidiabetic metabolic 2
effects. Nature 423 762-769. 3
Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, Yamashita S, Noda M, Kita S, 4
Ueki K, et al. 2002 Adiponectin stimulates glucose utilization and fatty-acid oxidation by 5
activating AMP-activated protein kinase. Nat Med 8 1288-1295. 6
Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y, Ide T, Murakami K, 7
Tsuboyama-Kasaoka N, et al. 2001 The fat-derived hormone adiponectin reverses insulin 8
resistance associated with both lipoatrophy and obesity. Nat Med 7 941-946. 9
10
11
12
Page 28 of 42

29
Figure legends 1
Fig.1. Regulation of KiSS-1 mRNA expression by different concentrations of gAd or AMPK. 2
GT1-7 cells were serum starved, and then treated with gAd for 2 h in different concentrations 3
(from 0-5 ug/ml) (A) or treated with 1mM AICAR or 20/40uM Compound C for 2 h (B). KiSS-1 4
mRNA levels were determined by realtime-PCR analysis. Results were representative of at least 5
three replicate determinations for each treatment group. *P<0.05, compared with the control 6
group. 7
8
Fig.2. Regulation of KiSS-1 promoter activity by gAd or AMPK. GT1-7 cells were serum starved, 9
and then transiently transfected with 2-kb full-length KiSS-1 pormoter (PGL3-2k), along with a 10
Renilla luciferase to control for transcription efficiency. After 18 h of transfection, cells were 11
treated with gAd (1.0 ug/ml) for different times (A) or at different concentrations for 24 h (B), or 12
treated with 1mM AICAR or 20uM Compound C (C) or AMPKα1-SiRNA (D), or pretreated with 13
AICAR (1mM), Compound C (20uM) for 30 min before incubation with gAd (5.0 ug/ml) for 24 14
h (E). Luciferase expression level driven by KiSS-1 promoter was measured and normalized to 15
Renilla luciferase activity. Results were representative of at least three replicate determinations 16
for each treatment group. *P<0.05, compared with the control group. 17
18
Fig.3 gAd induces subcellular redistribution of SP1 protein. GT1-7 cells were treated with gAd in 19
different concentrations (from1-10ug/ml), after which cell lysates were separated into nuclear and 20
cytoplasmic fractions. The nuclear and cytoplasmic fractions as well as total cell extracts were 21
subjected to western blot analysis with antibodies to SP1 protein , Lamin B1 (nuclear marker), 22
Page 29 of 42

30
β-tubulin (cytoplasmic marker) or β-actin (total protein marker), as indicated. Results are 1
representative of at least three replicate determinations for each treatment group. *P<0.05, 2
compared with the control group. 3
4
Fig.4 AMPK induces subcellular redistribution of SP1 protein. GT1-7 cells were treated with 5
AICAR (0.1, 1.0mM), Compound C (40uM) (A) or AMPK SiRNA (B), after which cell lysates 6
were separated into nuclear and cytoplasmic fractions. The nuclear and cytoplasmic fractions as 7
well as total protein extracts were subjected to western blot analysis with antibodies to SP1 8
protein, Lamin B1 (nuclear marker), β-tubulin (cytoplasmic marker) or β-actin (total protein 9
marker), as indicated. Results were representative of at least three replicate determinations for 10
each treatment group. *P<0.05, compared with the control group. 11
12
Fig.5. AMPK induces subcellular redistribution of SP1 protein by immunofluorescence analysis 13
(200×). GT1-7 cells were treated with AICAR (1mM) or Compound C (20uM) for 24 h, fixed, 14
immunostained with antibodies to SP1 which was visualized by a secondary antibody coupled to 15
FITC. Nucleus was stained by 1-Aminopyridinium iodide (red), and then examined with a 16
fluorescence microscope. SP1 is distributed in cytoplasm and nucleus of untreated GT1-7 cells 17
(Witte et al.) (a). In response to AICAR treatment, SP1 in the cytoplasm increased greatly (Witte 18
et al.) (b). SP1 in the nucleus increased greatly after treated with Compound C (Witte et al.) (c). 19
Nucleus was stained by 1-Aminopyridinium iodide (red) (d, e, and f). Merged figures with graph 20
a and d, b and e, c and f were respectively shown as graph g, h, and i. Color of SP1 localized in 21
nucleus merged with nucleus stain shows yellow. The yellow color was more deep in Compound 22
Page 30 of 42

31
C group compared with the control group and AICAR group demonstrated the subcellular 1
localization of SP1(g, h, and i). 2
3
Fig.6. Regulation of KiSS-1 promoter activity by SP1 SiRNA and mithramycin A. Cells were 4
transfected with full-length KiSS-1 promoter (PGL3-2k), along with a Renilla luciferase to 5
control for transcription efficiency, then passaged and transiently transfected with SP1 SiRNA or 6
treated with mithramycin A (100nM, 200nM). Luciferase activity was then assayed. Regulation 7
of SP1 protein levels by SP1 SiRNA from western blot analysis. Results were representative of at 8
least three independent experiments. *P<0.05, compared with the control group. 9
10
11
Fig.7. Regulation of KiSS-1 mRNA expression by SP1 SiRNA. GT1-7 cells were transfected 12
with SP1 SiRNA (SiRNA 1669, SiRNA 965, SiRNA 1867). KiSS-1 mRNA levels were 13
determined by realtime-PCR analysis. Results were representative of at least three replicate 14
determinations for each treatment group. *P<0.05, compared with the control group. 15
16
17
Fig.8. Regulation of KiSS-1 mRNA expression by gAd, AICAR or Compound C in 18
Sprague-Dawley rats. The rats were given intracerebroventricular injection of 19
PBS/DMSO, gAd, AICAR or Compound C, their heads were immediately removed on the 20
ice, then the ARC area was dissected out of the brains, followed by gene expression 21
assays as described in method part. KiSS-1 mRNA levels were determined by realtime-PCR 22
Page 31 of 42

32
analysis. Results were representative of at least three replicate determinations for each treatment 1
group. *P<0.05, compared with the control group. 2
3
4
Fig.9. Regulation of kisspeptin expression in the ARC area of Sprague-Dawley rats by gAd, 5
AICAR or Compound C. The Sprague-Dawley rats were anaesthetized under intraperitoneal 6
injection of pentobarbital anesthesia and given an intracerebroventricular injection of colchicines at 7
75µg/15µl and followed by intracerebroventricular injection of PBS/DMSO, gAd, AICAR and 8
Compound C, then kisspeptin levels were determined by immunohistochemistry as described in 9
method part. Graph A, B, C and D stands for treatment of PBS/DMSO, gAd, AICAR and 10
Compound C respectively. To quantify kisspeptin immunopositive neurons in the ARC, cell counting 11
was performed by two researchers. Bilateral cell counts for each section were averaged between 12
researchers. Total cell counts of the ARC (eight sections/rat) were summed and analyzed. Results 13
were representative of at least three replicate determinations for each treatment group. *P<0.05, 14
compared with the control group. 15
Page 32 of 42
254x190mm (96 x 96 DPI)
Page 33 of 42

254x190mm (96 x 96 DPI)
Page 34 of 42

254x190mm (96 x 96 DPI)
Page 35 of 42
254x190mm (96 x 96 DPI)
Page 36 of 42

529x533mm (96 x 96 DPI)
Page 37 of 42

254x190mm (96 x 96 DPI)
Page 38 of 42
159x91mm (96 x 96 DPI)
Page 39 of 42
146x105mm (96 x 96 DPI)
Page 40 of 42

254x190mm (96 x 96 DPI)
Page 41 of 42

Table1. The methods of i.c.v for the four groups
i.c.v
Control Adiponectin
(gAd)
AICAR CompoundC
(Comp C)
Drug(ug)
SolutionⅠ(ul)
----
PBS(10)
gAd (7)
PBS (10)
AICAR(10)
PBS(10)
Comp C (40)
DMSO (10)
SolutionⅡ(ul) DMSO(10) DMSO(10) DMSO(10) PBS(10)
Page 42 of 42