a study of the reciprocal connections between the septum and the entorhinal area using anterograde...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 225~327-343 (1984)
A Study of the Reciprocal Connections Between the Septum and the Entorhinal Area Using Anterograde and Retrograde
Axonal Transport Methods in the Rat Brain
ANGEL ALONSO rn CHRISTER KOHLER Astra Research Laboratories, Department of Pharmacology, Sodertalje, Sweden
ABSTRACT The reciprocal connections between the septum and the entorhinal area
(EA) was studied in the rat brain using antero- and retrograde axonal transport methods. After injections of large volumes (2 x 100 nl) of horserad- ish peroxidase (HRP) conjugated to wheat-germ agglutinin (WGA) into the medial septum (MS) and the diagonal band of Broca (dbB), anterogradely transported HRP-WGA was found primarily in layers I1 and IV of the medial and lateral EA. Injections of HRP-WGA (50-100 nl) or fluorescent dyes (50- 100 nl) into different parts of the retrohippocampal region resulted in label- ing, by retrograde axonal transport, of cells in the MS and dbB, both ipsi- and contralateral to the injected hemisphere. The labeled cells were either small (long axis of soma: 10-15 pm), round, and oval, or medium (15-25 pm) to large (25-35 pm) of fusiform or multipolar shape. By using the method of retrograde fluorescent double labeling, the septal afferents to the EA were found to give off collaterals to other parts of the hippocampal region. A much smaller number of septal cells appeared to send bilateral projections to the EA of both hemispheres. Studies employing retrograde transport of HRP in combination with acetylcholinesterase (AChE) histochemistry on the same tissue section showed that, while a large number of cells projecting to the EA contain AChE, many projecting cells are devoid of AChE reaction prod- ucts. These findings suggest that the septo-entorhinal projection consists of a cholinergic as well as a noncholinergic component.
The entorhinal efferents to the septum were studied after injections of HRP-WGA into different parts of the retrohippocampal region. Labeled fibers could be traced through the fimbria to their terminal fields in the intermediate parts of the lateral septal nucleus and to the most lateral aspect of the vertical limb of the dbB. The cells giving rise to this projection were situated in layer IV of the medial and layers I1 through V of the lateral EA. Taken together, the present findings demonstrate a close anatomical relationship between the septum and the entorhinal area, in addition to the better known connections between the septum and the Ammon’s horn.
Key words: septo-entorhinal connections, rat
‘The hippocampal region is used as a collective name for the Amman's horn, area dentata, subiculum, Pre- and Parasubiculum, and the entorhinal area (area 28 a, b; see Blackstad, ’56). The retrohippocampal region consists of the subicular complex and the entorhinal area. For more detailed descriptions of the different hippocampal subfields see Blackstad, ’56; Swanson and Cowan, ’79.
Accepted December 2, 1983. Angel Alonso’s permanent address is Research Department,
Address reprint requests to Christer KGhler, Department of Center “Ramon y Cajal,” Madrid, Spain.
Pharmacology, Astra Research Laboratories, SGde,-tglje, Sweden,
0 1984 ALAN R. LISS, INC.

328
The hippocampal region' of the rat is innervated by neu- rons located in the medial parts of the septal area (Meibach and Siegel, '77; Raisman, '66; Siegel and Tassoni, '71; Swanson and Cowan, '77). Several neuroanatomical studies have shown that the septal projections to the hippocampal formation originate in the medial septal nucleus (MS) and the nucleus of the diagonal band of Broca (dbB) (Segal and Landis, '74; Wyss et al., '79). The major terminal fields of these hippocampal afferents are found in the hilus and the molecular layer of the area dentata as well as in the stra- tum oriens and stratum pyramidalae of the regio inferior and regio superior (Mellgren and Srebro, '73; Rose et al., '76, Mosko et al., '73). While most earlier studies have focused on the organization of the septal projections to the hippocampal formation, far less is known about the septal innervation of the retrohippocampal structures, and, in par- ticular the entorhinal area. Nevertheless, studies using degeneration (Mellgren and Srebro, '73) and axonal trans- port (Segal, '77; Beckstead, '78; Swanson and Cowan, '77) methods have pointed out the possible existence of a projec- tion from the septum to the entorhinal area in the rat brain, although the exact origin, termination, and neurochemical identity of this projection have remained unknown.
In the rat, the EA is the major route by which cortical input reaches the hippocampal formation. The perforant path, which originates in layers I1 and I11 of the EA (Stew- ard, '76; Steward and Scoville, '761, provides a potent exci- tatory input to the granule cells of the area dentata (Andersen et al., '66). Thus, the neurons of the EA occupy a key position within the intrinsic neuronal circuit of the hippocampal region, and the identification of their afferent and efferent connections remains an important step in un- derstanding hippocampal function.
Recent studies have shown that the EA is innervated by neurons situated in a large number of different cortical and subcortical brain regions, including the septum (Srebro et al., '79; Wyss et al., 79). Furthermore, it has been suggested that the septo-entorhinal projection is part of a larger cho- linergic system connecting the septum with the hippocam- pal region (Mellgren and Srebro, '73; Shute and Lewis, '67). In the present study, we have examined the septo-ento- rhinal connections in more detail, using anterograde and retrograde axonal transport methods and acetylcholinester- ase (AChE) histochemistry in the rat brain.
MATERIALS AND METHODS Subjects
The present study was based on a n analysis of brains from 50 male albino rats of the Sprague-Dawley strain (Anticimex, Sweden; weight at surgery: 150-250 g). During the experiments the rats were housed under regular labo- ratory conditions.
HRP histochemistry The rats were anesthetized with pentobarbital (Mebumal;
60 mgkg) and positioned in a Kopf stereotaxic instrument with the head of the animal kept in a horizontal position. Injections of horseradish peroxidase (HRP, Sigma Chemical Co., St. Louis, USA; 30-40% (w/v) dissolved in saline con- taining 1% dimethylsulfoxide) or HRP coupled to wheat- germ agglutinin (HRP-WGA, Sigma; 2% (w/v) dissolved in saline) were made into the medial (MEA) and lateral (LEA) entorhinal area, respectively, or into the septal region, of individual rats. The injections were made either by pres- sure (50-100 nl) through a glass pipette (outer diameter 70- 100 pm) glued to a Hamilton microsyringe (1 pl), or by iontophoresis through a fine glass capillary. The injections
A. ALONSO AND C. KOHLER
into the entorhinal area were made with the pipette angled 10" (MEA) or 12" (LEA) to the vertical plane. Two to seven days after surgery the rats were deeply anesthetized, tra- cheotomized, provided with artificial respiration, and per- fused through the ascending aorta with warm (20°C) saline (50 ml) followed by 500 ml of either 3% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) or a mixture of paraformal- dehyde and glutaraldehyde (2% and 1%, respectively) in 0.1 M sucrose-phosphate buffer (pH 7.4). The fixation was fol- lowed by a rinse with 500 ml of 5% (w/v) sucrose-phosphate buffer (pH 7.4). Some of the brains injected with HRP were processed for the histochemical visualization of AChE (Ge- neser-Jensen and Blackstad, '71). These rats were perfused with saline (50 ml) followed by 500 ml of 4% paraformalde- hyde in 0.1 M phosphate buffer (pH 7.4). The brains were removed and stored in 20% (w/v) sucrose-phosphate buffer (pH 7.4) at 4°C until they were cut in a cryostat.
The HRP-injected brains were reacted according to the method of Mesulam ('76a) and Mesulam and Mufson ('80) using tetramethylbenzidine (TMB) as a chromagen. Briefly, the cut sections (30-40 pm thick) were collected onto gela- tine-albumin-coated glass slides and dried before they were washed and preincubated with TMB for 20 min a t room temperature. The reaction was initiated by adding 2-3 ml of 0.01% H202 to the TMB preincubation medium, and stopped by transfer of the sections to acetic buffer (pH 3.3) in distilled water. The reacted sections were either stained with neutral red or immediately coverslipped with Pertex (Histolab, Gothenburg, Sweden).
AChE histochemistry Horizontal or frontal sections were stained for the visual-
ization of AChE using the Koelle copper-thiocholine method described by Geneser-Jensen and Blackstad ('71) and by Mellgren and Srebro ('73). Briefly, glass-mounted sections were incubated for 60-90 min at 37°C in a solution (pH 5.01, containing acetylthiocholineiodide (4 mM), copper sul- phate (2 mM), and glycine (10 mM) in acetate buffer (50 mM). The incubation was terminated by several quick washes in distilled water and the sections subsequently immersed in 1.25% sodium sulphite (pH 7.5) for 45-60 sec. After several washes in distilled water the sections were developed for 60 sec in 1% silver nitrate. Nonspecific ester- ases were deactivated by the addition of etopropazine (N- diethylaminoethylphenothiazine, 0.2 mM) to the incuba- tion medium.
AChE histochemistry in combination with HRP on the same tissue section
The sections were reacted with the copper-thiocholine method as described above or a method based on the above protocol, but with exposure of the sections to 10% potassium ferricyanide for 2 min after the HRP reaction in order to produce a reddish-brown precipitate in the AChE-positive cells (Mesulam and Van Hoesen, '76).
Retrograde fluorescent tracing In a separate series of experiments, the fluorescent dye
Granular blue (GB; 5%1, dissolved in distilled water) was injected into different parts of the entorhinal area or into the septum. In studies of the branching projections of septal neurons GB and Propidium iodide (PI; Sigma Chemical Co., St. Louis, USA; 3%, dissolved in distilled water) were in- jected into different parts of the hippocampal region in the same rat.
The fluorochrome-injected brains were fixed by transcar- dial perfusion with a rinsing solution (50 ml) containing (w/

SEPTO-ENTORHINAL INTERCONNECTIONS
v) sucrose (0.8'70), glucose (0.4%), and NaCl (0.8%) in dis- tilled water, followed by 500 ml of a fixative containing (w/ v) paraformaldehyde (4%), tannic acid (l%), and magnesium sulphate (4%). For a detailed description of this method see DeOlmos and Heimer ('80). The brains were kept in a 20% (w/v) sucrose-phosphate buffer (pH 7.4) solution until they were cut in a cryostat at - 15°C. The sections (30 pm thick) were collected onto gelatine-albumin-coated glass slides and allowed to dry before they were coverslipped with Entellan (Histolab, Gothenburg) and examined under a Leitz Orto- plan fluorescent microscope equipped with epi-illumination.
RESULTS Several excellent reviews exist on the subdivisions of the
septal area in the rat (Harkmark et al., '75; Srebro et al., '75; Swanson and Cowan, '79), cat (Andy and Stephan, '64), and man (Harkmark et al., '75). In the present study we have followed the nomenclature proposed by Swanson and Cowan ('79) and by Harkmark et al. ('75). The major com- ponents of the septal area as revealed by AChE histochem- istry are shown in Figure 1. The nomenclature used in the present study for the EA is based primarily on the work of Blackstad ('56) and Haug ('76). The subdivisions of major areas and layers of the retrohippocampal region, including the EA, are shown in Figure 2.
General comments about the injection sites HRP injections The use of the TMB histochemical tech-
nique (Mesulam, '76) precludes any definitive statements about the actual spread of HRP around the site of infusion. The injection site is generally found to be larger when the tissue is reacted with the TMB as compared to diamino- benzidine, at least when the HRP is delivered by pressure (Morrell et al., '81). The issue of the effective injection site is further complicated by our lack of detailed knowledge about the active uptake and axonal transport of HRP from different sectors of the areak) containing the diffuse, blueish reaction product around the track of the injection pipette.
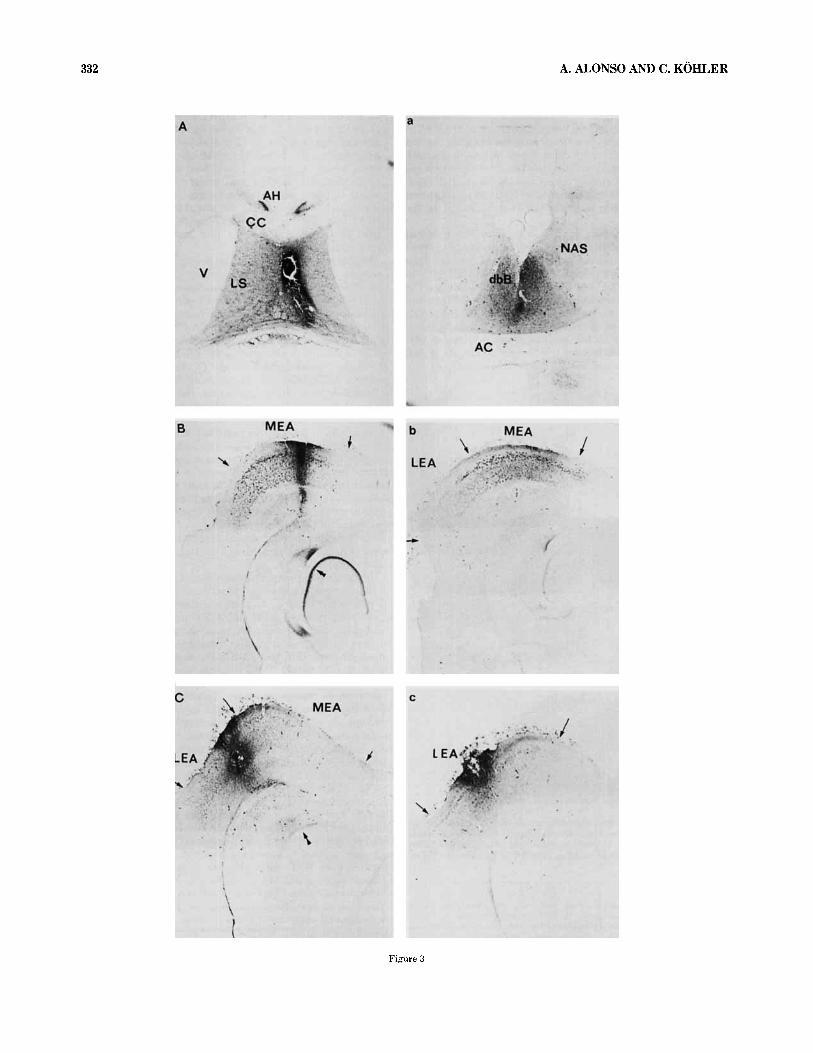
In the studies of anterograde transport of HRP-WGA from the septum to the entorhinal area, large (2 x 100 nl) intra- septal injections of HRP-WGA were used (Fig. 3a). These injections were aimed at the medial septum and diagonal band of Broca, but the area containing the HRP reaction product was never completely restricted to these subdivi- sions but included also the lateral septum, the medial part of the nucleus accumbens, and the cingulate cortex overly- ing the septal nuclei. In fact, injections of smaller volumes (50 nl) into either the medial or the lateral septum, respec- tively, also resulted in some spread of the HRP into the adjacent septal nuclei. In spite of the spread of HRP within the septum, after each of these smaller injections a separate population of neurons were labeled in the hippocampal formation (Alonso and Kohler, '82). Thus, it is possible that the area of uptake and transport of HRP is actually more restricted than would be predicted from the area containing the blue color around the injection site.
The present material includes relatively large injections (100-200 nl) of HRP-WGA into the EA with the spread of reaction product throughout the entire retrohippocampal region. Thus, it is likely that active uptake and retrograde transport of the conjugate occurred from structures other than the EA (such as the pre- and parasubiculum). In sev- eral cases, however, the HRP injections were restricted to either the medial or the lateral subdivisions of the EA, and, the presence of anterogradely transported HRP was noted only in that portion of the molecular layer of the area dentata which is known (Blackstad, '58; Hjort-Simonsen
329
and Jeune, '72; Steward, '76) to be occupied by the termi- nals of the medial and lateral perforant path, respectively (Fig. 3). This distribution of HRP in the molecular layer of the area dentata was used as a guide when determining if the injections were restricted to either part of the EA. Two representative HRP-WGA injections in the medial and lat- eral EA, respectively, are shown in Figure 3b. Comparisons between the retrograde transport found in brains with large or small injections showed a good correspondance with re- gard to the general distribution of labeled cells in different brain areas.
Fluomchrome injections Granular blue and Propidium iodide produced completely different patterns of labeling around the site of infusion. Injections of small quantities ( - 50 nl) of GB resulted in a well-restricted area of diffusion which extended spherically approximately 0.25-0.50 mm from the track of the injection pipette. In contrast to HRP and PI, the size of the GB injection site was well restricted and increased over time, which suggests that the injection site seen at 5-7 days after surgery represents the largest area contaminated by GB. It should be pointed out, how- ever, that in most cases severe damage of the nervous tissue was found along the track of the injection pipette. This is a common problem associated with GB injections in many brain regions, and may result in unwanted filling of dam- aged axons passing through the area of GB infusion.
Injections of PI did not result in the same nonspecific damage as GB. However, the exact area of the PI injection site was found to be extremely difficult to determine, since no clear border appears to exist between PI-labeled and nonlabeled tissue. It seems, however, that PI can spread for longer distances in the brain than GB. In the case of injec- tions of PI into the EA, we have assumed that little, if any, of the dye spread into the hippocampus, since no labeling of intrinsic hippocampal or commissural pathways could be found.
Septa1 projections to the entorhinal area Antemgrade transport studies The case described in de-
tail here is a large (2 x 100 nl) injection of HRP-WGA into the medial septum and the diagonal band of Broca (see Fig. 3a). Blue reaction product is present throughout the entire septal area including the MS, the vertical limb of dbB, the lateral septal nuclei, the anterior hippocampal rudiment, and the medial parts of the nucleus accumbens. Little or no
ab ad para prsb retrs e dbB,
ah AC cc F
dbBh
LSd LSj LS" LEA MEA MS NAS OT SF TA
A bbrevratioris Angular bundle Area dentata Parasubiculum Presubiculum Retrosplenial area e Diagonal band of Broca, vertical limb Diagonal band of Broca, horizontal limb Anterior hippocampal rudiment Anterior commissure Corpus callosum Fornix Lateral septum, dorsal part Lateral septum, intermediate part Lateral septum, ventral part Lateral entorhinal area (area 28b) Medial entorhinal area (area 28a) Medial septal nucleus Nucleus accumbens septi ,
Olfactory tubercle Septofimbria nucleus Transitional area

330 A. ALONSO Ah9 C. KOHLER
Figure 1

SEPTO-ENTORHINAL INTERCONNECTIONS 331
The distribution of septal efferents to the Ammon’s horn has been described in detail by Crutcher et al. (’81) and will not be dealt with further here, except for the drawings shown in Figure 4. As the HRP-WGA-filled axons leave the fornix-fimbria they assemble in the alveus and the stratum oriens, where they course in a caudal direction toward the angular bundle. At this level many of the fibers radiate toward the pre- and parasubiculum as well as the EA. Numerous fibers run along the angular bundle, and HRP- WGA-filled profiles, indicating labeled axons, can be fol- lowed as they run from the deep and toward the more superficial layers of the EA.
Within the EA, the dustlike precipitate of HRP-WGA reaction product is heterogeneously distributed between the different layers. Thus, what appears to be a dense terminal field is present in layer IV of both the medial and the lateral EA. A distinct, though sparser terminal field is present in layer I1 of the entire EA (Fig. 4). At the most ventral levels of the EA, anterogradely transported HRP- WGA is present in all layers, although most of the reaction product is present in layers I1 and IV. Analysis of horizontal sections cut through the hippocampal region clearly show HRP-WGA-filled fibers that enter the EA via a ventral route by way of the ansa lenticularis and through the amygdala and the piriform cortex. These fibers distribute to all layers of the ventral parts of the lateral EA and to a minor extent to the MEA. Fibers containing the HRP-WGA reaction product are present in the angular bundle of these ventral levels, indicating that some of the septal efferents ascend in the angular bundle to more dorsal parts of the hippocampus (Fig. 4). In addition to the projection from the septum to the EA, anterogradely transported HRP-WGA was found in both the parasubiculum and the deep layers of the presubiculum at dorsal levels, indicating the exis- tence of an efferent projection from the septum to the subi- cular complex.
The dense innervation of layers I1 and IV of the EA occurs in register with the laminar distribution of AChE staining in this area. Similar overlapping distribution of antero- gradely transported HRP and AChE is found in the para- subiculum and the deep layers of the presubiculum, and support earlier lesion studies (Mellgren and Srebro, ’73), which suggests that the EA like the hippocampus is inner- vated by cholinergic fibers from the medial septum (Mell- gren and Srebro, ’73).
Retrogrude transport studies Injections of HRP or HRP- WGA into either the medial or the lateral entorhinal area resulted in retrograde axonal transport of the protein to the parent cell bodies in the dbB and the MS, in addition to a large number of other brain areas. A majority of the retrogradely labeled cells are situated within the vertical and the horizontal limbs of the dbB and the horizontal nucleus of the dbB. Similarly, injections of either GB or PI into the EA resulted in retrograde labeling of cell bodies in the dbB and MS. The general position and morphology of the fluorochrome-labeled cells correspond well to those la- beled by HRP, although the fluorescent dyes labeled more cells per section. The superior sensitivity of the fluoro- chromes over HRP as retrograde tracers is particularly evident with regard to the labeling of septal cells located contralateral to the injected hemisphere.
Analysis of serial frontal sections cut through the septum showed that a majority of the cells projecting to the EA are located in the middle to caudal parts of the structure (Fig. 5). In all cases examined, a small number of HRP-positive
deposit of HRP-WGA is present in the horizontal compo- nent of the dbB or the olfactory tubercle. Our material also includes several smaller (< 100 nl) injections of HRP-WGA into the septum showing anterograde transport to the EA but the labeling found in these cases is less prominent, although the general distribution of fibers and preterminal processes in the EA closely resembles the large HRP injec- tion where anterogradely transported HRP-WGA is present throughout the entire hippocampal region, including the entorhinal area.
HRP-WGA-filled fibers are found to leave the injection site and to reach the hippocampal region by route of three main pathways: the dorsal fornix, the fimbria, and the ansa lenticularis and the piriform cortex (Fig. 4). The HRP-WGA- filled axons leave the septum and course caudally in the dorsal fornix and the lateral part of the fimbria. While the fibers in the dorsal fornix appear to be destined primarily for the dorsal parts of the subicular complex and the retro- splenial area, those in the fimbria show a more general distribution to all dorsal-ventral levels of the Ammon’s horn and the entorhinal area.
Fig. 2. Photomicrographs of horizontal sections cut through the retrohip- pocampal region at middle (a) and ventral cb) levels stained for the visual- ization of AChE activity. Arrows indicate borders between different areas of the region. The lamina dessicans is indicated by an asterisk. Original magnification 50 x.
Fig. 1. Photomicroflaphs of coronal sections through the septum stained for the histochemical visualization of AChE activity. The most rostra1 and caudal sections are shown in (a) andcd), respectively. The lateral and medial components of the horizontal diagonal band have been labeled 1 and rn, respectively. Magnification, 50 x .
332 A. ALONSO ANI) C. KOHLER
Figure 3

SEPTO-ENTORHINAL INTERCONNECTIONS 333
Fig. 4. Camera lucida drawings of horizontal sections showing the distri- bution of HRP-WGA-filled axons and apparent terminal deposit of the
cells are present in the dbB and MS contralateral to the injected hemisphere. Few, if any, retrogradely labeled cells are situated in the most rostra1 parts of the dbB.
The use of relatively long survival times (up to 5 days) in combination with a strong fixation of the tissue resulted in the visualization of HRP reaction product in the somata and proximal parts of the dendrites of the labeled cells. In some brains, extensive staining of the cell bodies and den- drites allowed a preliminary analysis of their morphology. A closer inspection of these cells suggests that they consti- tute a morphologically heterogeneous population (Fig. 6). Thus, the cells projecting to the EA are either of small (long axis of the soma: 10-15 pm) size and oval shape or medium (15-25 pm) to large (25-35pm) size and fusiform or multi- polar in shape. The caudal part of the MS is rich in medium to large multipolar and fusiform cells with long dendrites that cross the midline and invade the MS of the opposite side.
Neurons projecting to the medial and lateral part of the EA, respectively, occur intermingled in the horizontal limb of the dbB. In the MS and vertical limb of the dbB, however, the position of these cells shows a certain degree of geo- graphical segregation: cells projecting to the lateral EA tend to be located more lateral than those projecting to the
~
Fig. 3. Photomicrographs of horizontal sections showing examples of the apparent injection sites after injections of HRP-WGA into the septum IA,a), the medial (B,b) and the lateral (C,c) entorhinal area. Each injection is represented a t two levels. Double arrowheads indicate anterograde trans. port in the perforant path system after HRP-WGA injections into the ento- rhinal area. V, lateral ventricle. Original magnification 2 5 ~ .
protein throughout the longitudinal axis of the hippocampal region, includ- ing the EA. Compass gives medial (m) and caudal Ic) orientations.
medial EA (Fig. 5). This medial to lateral arrangement is also seen after small injections of GB into the lateral and PI into the medial EA. Analysis of the retrograde axonal transport of the dyes to the septum showed a separation between the neurons that give rise to these two projections: a majority of the cells retrogradely labeled from the medial part of the EA occupy a more medial position in the septum as compared to those labeled by GB after lateral EA injections.
Retrograde transport of HRP in combination with AChE In a separate series of experiments, retrograde ax- onal tracing of HRP from the EA to the septum was com- bined with AChE histochemistry on the same tissue section (for discussion of this method see Hardy et al., '76; Mesu- lam, '76; Mesulam and Van Hoesen, '76). Taken together, these experiments show that a large number of cells in the dbB and the MS contain both the blue HRP reaction prod- uct and the brown (or reddish) color which indicates the presence of cytoplasmatic AChE. The AChE-positive cells projecting to the entorhinal area do not belong to any mor- phologically distinct group of neurons, since small ovoid as well as large multipolar AChE-positive cells contain the retrogradely transported HRP (Figs. 7, 8). However, a rela- tively large number of cells that lack histochemically de- tectable AChE are retrogradely labeled by HRP after large injections of the protein into the hippocampal region, in- cluding the EA (Figs. 9, 10). This finding suggests that there may exist a prominent, noncholinergic projection from the septum to the entorhinal area (and the hippocampal formation), in addition to the one presumed to be cholin- ergic. The number and the relative positions of AChE-

334 A. ALONSO AND C. KOHLER
Fig. 5. Schematic drawings of coronal sections through the septum illus- trating the number and the relative position of HRP filled neurons after restricted injections of the protein into the medial (dots) and the lateral (triangles) entorhinal area, respectively, in two separate rats. The symbols
represent the actual numbers of HRP-filled neurons found in a 30-pm-thick section a t a rostrocaudal level corresponding to those shown in (a) through (d).
positive and AChE-negative cells containing HRP are shown schematically in Figure 7. Although many of the HRP-IabeledAChE-negative and HRP-labeledAChE-posi- tive cells in large part occur intermingled within the sep- tum, a closer inspection reveals that HRP-labeled neurons lacking AChE reactivity tend to be situated primarily in parts of the septum that contain relatively few AChE-posi- tive neurons in normal material (Fig. 8).
Branching of septa1 projections Injections of either GB or PI into the EA resulted in similar distribution of retro- gradely labeled neurons within the septum. Analysis of brains where GB (or PI) had been injected into the EA, and PI (or GB) into the dorsal or ventral hippocampal formation of the same side, showed a picture of retrograde labeling qualitatively similar to that seen with HRP. In one repre- sentative experiment, GB was injected into the dorsal hip-
pocampus and PI into the medial entorhinal area. The GB injection was situated at the border between the subiculum and CA1, with the tip of the pipette in the molecular layer of the area dentata. Spread of the dye extended approxi- mately 0.5 mm around the track of the pipette as well as along the pipette into the overlying neocortex. With excep- tion of the neocortex, diffusion of GB did not occur to any extrahippocampal region. The spread of PI after ento- rhinal injections is more difficult to determine, but a zone of bright red fluorescence extended spherically at least 0.5 Gm from the center of the injection. The neurons projecting to the hippocampal formation and the EA, respectively, occur intermingled along the rostrocaudal axis of the sep- tum, although the cells labeled from the dorsal hippocam- pus are present far more rostra1 in the diagonal band than are the EA projecting ones. In addition to these single

SEPTO-ENTORHINAL INTERCONNECTIONS 335
Fig. 6. Photomicrographs of HRP-filled neurons in the medial septal nucleus and in the diagonal band of Broca (a) after a large (100 nlj HRP injection into the MEA, that involved also parts of the lateral entorhinal area as well as the medial part of the parasubiculum. Cells in (b) and (cj are from the vertical limb of dbB; the cells in (d) and (el are situated in the caudal part of the medial septal nucleus. Arrows in (a) point a t weakly
labeled cells in the lateral part of dbB. Double arrowhead marks the mid- line. Arrowheads in (ej point a t the distal part of a labeled dendrite crossing the midline. Small midline cell marked by arrow in (el. Arrow in (f) indicates HRP deposits in efferents from the entorhinal area to the septum. Nomarski optics. Original magnification 130x (a); 540x (b-D.
labeled cells, a relatively large number of neurons in both double-labeled cells did not differ significantly when com- the dbB and the MS contained both fluorochromes within parisons were made between the dorsal and ventral hippo- their somata (Fig. 9). Cell counts performed in several rats campal injections vis-a-vis those made in the EA. In a showed that approximately 20% of all retrogradely labeled separate series of experiments, the fluorescent dyes were cells in the septum are double-labeled after these injections. injected into various parts of the right and left hippocampal In several instances, double-labeled cells are present also region, respectively. These experiments showed that, in contralateral to the injected hemipshere. The proportion of contrast to the relatively large proportion ( - 20%) of septal

336 A. ALONSO AND C. KOHLER
/ - - , /
Fig. 7. Camera lucida drawings of coronal sections through the septum a t three rostrocaudal levels (left to right) to illustrate the number and position of HRP-filled. AChE-positive (black dots) or AChE-negative (black
dots with slashes) cells containing HRP retrogradely transported fro,,, the entorhinal area. Small dots show cells containing only AChE i n their cyto. plasm or on their somatic membrane.

SEFTO-ENTORHINAL INTERCONNECTIONS 337
Fig. 8. Photomicrographs of coronal sections through the septum show- ing AChE-positive (large arrow) and AChE-negative (small arrow) cells containing HRP after injections into the MEA (a and b). Double arrowhead in (c) indicates HRP-positive cell lacking AChE. Single arrow shows HRP
cell with cytoplasmatic AChE reactivity, and double small arrows show AChE-positive cells lacking HRP granules. The position of cells (d) is indi- cated by arrow in (b). 300 X (a, b); 540 X (c, d).
neurons projecting to both the HPC and EA ipsilateral to the injection, few cells (< 5%) showed branching projections to the EA or HPC of both hemispheres.
onal transport of the protein to the septum, in addition to a large number of other subcortical structures. After HRP- WGA iniections into the MEA, labeled axons can be fol-
Entorhinal projections to the septum lowed from the injection site toward the angular bundle which thev either Denetrate on their way to the alveus or
Anterogmde tmnsport studies Injections of HRP-WGA into different parts of the EA resulted in anterograde ax-
ascend within to more dorsal levels. A- large number of HRP-WGA-labeled fibers run rostrally in the alveus to en-

338 A. ALONSO AW C. KOHLER
Figure 9

SEPTO-ENTORHINAL INTERCONNECTIONS
a
339
Fig, 10. Camera lucida drawings of the distribution of terminals and fibers in the septum in sagittal (a) and frontal (b) sections after an injection of HRP-WGA into the medial entorhinal area. The frontal section in (b) represents the level shown by the dotted line in (a). Crossed arrows mark
supracallosal fibers. Small arrow indicates HRP in the subcallosal stria. Double arrowhead indicates terminal field in the intermediate part of the LS.
ter the fimbria, where they occupy its most lateral portion. On their route to the terminal field in the LS, the labeled fibers traverse the dorsolateral part of the septum. Further rostrally they turn ventral as they enter the intermediate parts of the lateral septal nucleus. Throughout the rostro-
Fig. 9. Photomicrographs of cells in the diagonal band of Broca that were double-labeled (arrows) after injections of granular blue (a, c) and propidium iodide (b, d) into the hippocampus and medial entorhinal area, respectively, of the same rat. Double arrow in (b) indicates a cell labeled only by propi- dium iodide. 160x(a, b); 2 5 0 ~ (c, d).
caudal axis of the septum, the medial septal nucleus and the intermediate part of the lateral septum are the domi- nant terminal fields of the EA projections. At the most rostra1 levels the EA efferents occupy the entire lateral septal nucleus. The terminal fields of the efferents from the MEA and LEA show a high degree of overlap in the septum. The only apparent difference is a slightly less prominent input to the MS and dbB from the lateral than from the medial EA.
Retrograde transport studies Injections of either HRP- WGA or GB into the intermediate part of the lateral sep-

340 A. ALONSO AND C. KOHLER
tum resulted in labeling, by retrograde axonal transport, of cells in both the MEA and the LEA (Figs. 11, 12). Analysis of the laminar distribution of the retrogradely labeled neu- rons revealed that a majority of these cells are located in layer IV of both the MEA and the LEA. After long survival times (up to 3 days), retrogradely labeled cells are found also in layers I and I11 (Fig. 11). A majority of these cells, however, are found in the most lateral part of the LEA. Close to the rhinal fissure, scattered cells containing HRP- WGA reaction products or GB fluorescence are present in all cortical layers, including layers V and VI (Fig. 11). Analysis of serial horizontal sections cut through the EA showed an increasing number of retrogradely labeled cells at successively more ventral levels (see Fig. 11). A large number of the retrogradely labeled cells appears to be py- ramidal or stellate in shape.
DISCUSSION Using the methods of anterograde and retrograde axonal
tracing, we have analyzed the connections between the septum and the entorhinal area in the rat brain. The find- ings of reciprocal connections between the septum and the EA extend previous investigations showing prominent con- nections between the septum and the hippocampal forma- tion (Powell and Cowan, '55; Raisman, '66; Raisman et al., '66; Segal and Landis, '74; Swanson and Cowan, '79; Cragg and Hamlyn, '56; Siegel and Tassoni, '77; Meibach and Siegel, '77) in several animal species, including the rat. The findings of the present study indicate that the reciprocal
n
connections between the septum and the retrohippocampal structures are more extensive than previously recognized. The observation that the EA is innervated by neurons in the septum suggests that septal modulation of hippocampal activity can occur at different levels within the hippocam- pal region, including the entorhinal area.
The septo-entorhinal projection The existence of a path- way from the septum to the EA was suggested in early degeneration studies by Mellgren and Srebro ('73) and in autoradiographic experiments by Swanson and Cowan ('79) and Hunt and Schmidt ('79). In the two latter studies, however, the septo-entorhinal projection was mentioned only incidentally and no detailed analysis of the terminal distribution of this pathway was reported. It is clear from the present study, however, that the septal innervation of the EA is far more extensive than has been recognized in previous studies. The extensive labeling of this pathway in our experiment may have been due to the injections of relatively large volumes of tracer, in combination with rel- tively long survival periods (up to 5 days). The need for
Fig. 11. Schematic drawings of horizontal sections through the retrohip- pocampal region showing the distribution of neurons (dots) in the EA retro- gradely labeled by HRP after a large (100 nl) injection into the septal area. Each level is separated by approximately 1200 pm and summarizes the number of labeled cells found in eight consecutive 30-pm-thick sections.
Fig. 12. Photomicrographs showing cells in layer IV of the medial ento- rhinal area retrogradely labeled by granular blue after injection of the fluorochrome into the septal area. Original magnification 16Ox (a); 2 5 0 ~ 03). Caudal (c ) and medial (m) orientations are given by the compass.

SEPTO-ENTORHINAL INTERCONNECTIONS 341
large injections of tracer to demonstrate this pathway is probably due to the elongated and slightly curved shape of the dbB, which harbors a majority of the EA projecting cells, since restricted injections of HRP-WGA (Alonso and Kohler, unpublished observations) or 3H-amino-acids (Swanson and Cowan, '79) into small parts of the dbB re- sulted in modest labeling of the septal efferents to the entorhinal area.
A majority of the septal projections to the EA appear to terminate in layers I1 and IV. This pattern of distribution is in register with the laminar distribution of AChE. The overlap of HRP and AChE reaction products suggests that the AChE activity in the EA may derive from septal effer- ents to layers I1 and IV. That this may be the case was indicated previously by lesion experiments, showing a loss of AChE activity and the presence of terminal degeneration in these layers after electrolytic destruction of the medial septum (Harkmark et al., '75). The question remaining to be settled, however, is to what extent AChE in the EA indicates the existence of a cholinergic septo-entorhinal projection in the rat. The usefulness of AChE as a histo- chemical marker for cholinergic neurons has been chal- lenged in several recent studies (see Fibiger, '82, for review) and many studies have shown that in some brain regions AChE-positive cells do not contain choline acetyltransfer- ase immunoreactivity (CAT) (see Levey et al., '83). There exists good experimental evidence that AChE activity in the hippocampal formation resides partly in cholinergic afferents from the medial septum and the diagonal band of Broca, although far less is known about the identity of entorhinal AChE. Numerous CAT-immunoreactive fibers have been demonstrated in the hippocampal region (Ki- mura et al., '80), which suggests that this region does re- ceive a cholinergic innervation. However, the possibility that entorhinal AChE derives also from noncholinergic af- ferents cannot be ruled out at present. Interesting in this regard is the observation that in the spinal cord and sub- stantia nigra AChE hydrolyzes substance P (SP) (see Chubb et al., '80). Since SP-immunoreactive nerve terminals are present in the EA (Vincent et al., ,811, such a role for AChE cannot be excluded. Although further studies are needed to settle this issue, there is indirect evidence indicating that part of the septo-entorhinal projection indeed belongs to a general cholinergic pathway connecting the medial septum and the diagonal band with the hippocampal region (see Shute and Lewis, '67; Mellgren et al., '73). For example, the distribution of AChE- and CAT-positive cells appears to overlap in the medial septum (Kimura et al., '80), although this correlation is not perfect (see Levey et al., '83). The septal neurons retrogradely labeled from the EA contain AChE (present study), and lesions of the septum result in a depletion of AChE staining in both the hippocampus and EA with a similar time course (Mellgren and Srebro, '73). Furthermore, biochemical studies have shown loss of CAT and acetylcholine activity after destruction of the medial septum (Oderfeld-Nowak et al., '74; Pepeu et al., '71; Sethy et al., '73).
In the present study many of the cells in the septum, retrogradely labeled from the EA, were found to contain AChE as judged from experiments combining HRP and AChE histochemistry on the same tissue section (Mesulam, '76; Mesulam and Van Hoesen, '76; Hardy et al., '76). In addition, HRP-labeled cells lacking detectable amounts of AChE in their cytoplasm were found after entorhinal or hippocampal injections of HRP. This observation suggests
that there may exist a noncholinergic component of the pathway from the septum to the hippocampal region, in- cluding the EA. Recent studies using antibody to CAT (Kimura et al., '81) support the impression created from AChE-stained sections, namely that not all cells in the diagonal band of Broca are cholinergic. Furthermore, mas- sive injections of HRP (or fluorochromes) into the hippocam- pal region, including the EA, resulted in retrograde labeling of more cells per section than could be accounted for by the number of AChE-stained cells found in adjacent sections or in sections cut from fresh tissue (present study). The pres- ence of HRP-labeled neurons lacking AChE activity was established using two histochemical methods for the visu- alization of AChE that offer different optical resultion of AChE vis-a-vis HRP reaction products (see Materials and Methods). Both methods gave the same results. It seems unlikely that fixation procedures or optical factors reduced or abolished the visability of AChE in HRP-labeled neu- rons. First, the number of AChE-positive cells or the quality of staining did not differ significantly in sections of the septum from normal unfixed or formaldehyde-fixed brains. Second, in several brains the sections were "overincubated" for AChE. This procedure did not alter the number of HRP- AChE-positive to HRP-AChE-negative cells. Third, the sec- tions were analyzed using Nomarski optics, which en- hances the contrast in the section and greatly facilitates the identification of HRP-stained neurons lacking AChE (see Levey et al., '83). Under these conditions only cells with blue reaction product in their somata and dendrites, but with a white nucleus and a soma lacking brown precip- itate, were accepted as AChE-negative. This stringent cri- terion might in fact have resulted in a slight underestimation of the actual number of HRP-positive- AChE-negative cells, since many septal cells are cholino- ceptive (Kimura et al., '81; Fibiger, '82), with precipitate of AChE activity covering the outer surface of their soma membranes.
The neurochemical identity of the "noncholinergic" com- ponent of the septo-entorhinal projection remains unknown at present. However, the MS and dbB have recently been shown to contain a large number of cells immunoreactive for glutamic acid decarboxylase (GAD) (Kohler and Chan- Palay, '83), the enzyme responsible for the synthesis of y- aminobutyric acid. I t remains to be established if any of these GAD-positive neurons project to the hippocampal re- gion, including the entorhinal area.
Many of the septal cells with projections to the EA have axon collaterals to the hippocampal formation of the same hemisphere. However, in spite of the fact that many of the cells in septum have collaterals to the HPC and the EA, a very small number of cells showed bilateral projections to both hemispheres in the double-labeling experiments, al- though a single neuron may have an axon that crosses to the contralateral EA. The same type of organization has been observed in another midline structure, the dorsal raphe nucleus, where few cells project bilaterally to the EA (Kohler and Steinbusch, '82) or the striatum (Van der Kooy and Hattori, '80) as shown by the method of fluorescent retrograde tracing.
The enforhinul-septul projection Until recently, the per- forant path was regarded as the only efferent pathway of the entorhinal area in the rat. It is now well established that the EA has extensive extrahippocampal projections to a number of cortical and subcortical structures in the rat (Srebro et al., '79; Wyss, '81). One important target of these

342 A. ALONSO AND c. KOHLER
EA efferents is the septum as shown earlier using retro grade fluorescent tracing (Kohler and Eriksson, '81), and ii the present study using both anterograde and retrogradt tracing methods. Within the septum, the terminal distri- bution of the EA projection is restricted to the intermediate parts of the lateral septum, parts of the medial septal nu- cleus, and the vertical limb of the dbB. This distribution is different from the terminal field occupied by projections from the dorsal hippocampus, but overlaps to a certain degree with the projections originating from cells situated at ventral levels of the hippocampal formation (Swanson and Cowan, '79). Although the lateral septal area has been shown to project to several hypothalamic areas (see Swan- son and Cowan, '79), little is yet known about the efferent projection(s) of its intermediate parts. Until this informa- tion becomes available it is difficult to relate the EA inner- vation of the intermediate part of the lateral septum to a more general neuronal network connecting this particular septal nucleus with other parts of the brain. It is interest- ing, however, that a small component of the projections from the EA enters the MS-dbB complex, since this part of the septum contains neurons that project back upon the hippocampal region, including the entorhinal area.
A majority of the cells in the EA which contribute to the septal projection are situated in layer IV and to a minor extent in layer I1 of both the medial and lateral EA. These are the same layers that receive afferents from the medial septum and diagonal band of Broca. It remains to be shown if the same neurons of layer IV that project to the septum are innervated by the septal efferents to this layer. The projections from layer IV of the EA to the septum have axon collaterals to the nucleus accumbens septi as well as to the frontal cortex, as demonstrated in retrograde fluores- cence tracing experiments (Kohler and Eriksson, '81). Layer I1 of the EA contains neurons that give rise to the perforant path (Steward and Scoville, '76) as well as some of the septal projecting cells, but it is not known if these cells have axon collaterals to both the septum and the hippocam- pus. The chemical identity of the projection to the septum is not known at present. It is possible that at least some of these fibers may use glutamate or aspartate since it has been shown (Zaczek et al., '79; Walaas, '81) that their tran- section results in reductions of the high-affinity uptake of 3H-glutamate in the septal area.
ACKNOWLEDGMENTS We thank L.G. Eriksson for providing expert technical
assistance. A.A. was supported by the European Training Program in Brain and Behavioural Research.
LITERATURE CITED Alonso, A., and C. Kohler (1982) Evidence for separate projections of hippo-
campal pyramidal and non-pyramidal neurons to different parts of the septum in the rat brain. Neurosci. Lett. 31:209-214.
Andersen, P., B. Holmquist, and P.E. Voorhoeve 11966) Entorhinal activa- tion of dentate granule cells. Acta Physiol. Scand. 66:448-460.
Andy, O.J., and H. Stephan (1964) The Septum of the Cat. Springfield, Ill.: Thomas.
Biscoe, T.J., and D.W. Straughan (1966) Microelectrode studies of neurons in the cat hippocampus. J. Physiol. (Lond.) 183:341-359.
Bjorklund, A., and G. Skagerberg (1979) Simultaneous use of retrograde fluorescent tracers and fluorescent histochemistry for convenient and precise mapping of monoaminergic projection and collateral arrange- ments in the CNS. J. Neurosci. Meth. 1:261:277.
Blackstad, T.W. (1956) Commissural connections of the hippocampal region in the rat with special reference to their mode of termination. J. Comp. Neurol. 105:417-537.
Blackstad, T.W. (1958) On the termination of some afferents to the hippo- campus and fascia dentata. An experimental study in the rat . Acta Anat. 35:202-214.
Reckstead. R.M. 11978) Afferent connections of the entorhinal area in the rat as demonstrated by retrograde cell labeling with horseradish perox- idase. Brain Res. 152:249-264.
Chubb. I.W., A.J. Hodgson, and G.H. White (1980) Acetylcholinesterase hydrolyses substance P. Neuroscience. 52065-2072.
Cragg, B.G., and H. Hamlyn (1956) A histological study of some commis- sural and septal connections of the hippocampus in the rabbit. J. Anat. bond.) 90591-596.
Crutcher, K.A., R. Madison, and J.N. Davis 11981) A study of the rat septo- hippocampal pathway using anterograde transport of horseradish per- oxidase. Neuroscience 6:1961-1973.
DeOlmos, J., and L. Heimer (1980) Double and triple labeling of neurons with fluorescent substances: The study of collateral pathways in the ascending raphe system. Neursci. Lett. I9:7-12.
Fibiger, H.C. (1982) The organization and some projections of cholinergic neurons of the mammalian forebrain. Brain Res. Rev. 4:327-388.
Geneser-Jensen, F.A., and T. Blackstad (1971) Distribution of acetylcholin- esterase in the hippocampal region of the guinea pig. 1. Entorhinal area, parasubiculum and presubiculum. Z. Zellforsch. 114:460-481.
Hardy, H., L. Heimer, R. Switzer, and D. Watkins (1976) Simultaneous demonstration of horseradish peroxidase and acetylcholinesterase. Neu- rosci. Lett. 3:l-5.
Harkmark, W., S.I. Mellgren, and B. Srebro 11975) Acetylcholinesterase histochemistry of the septal region in rat and human: Distribution of enzyme activity. Brain Res. 95.281-289.
Haug. F.M.S. 11976) Sulphide silver pattern and cytoarchitectonics of para- hippocampal areas in the rat. Special reference to the subdivision of area entorhinalis (area 28) and its demarcation from the piriform cortex. Adv. Anat. Embryol. Cell. Biol. 525-73.
Hjort-Simonsen, A. 11971) Hippocampal efferents to the ipsilateral ento- rhinal area: An experimental study in the rat . J. Comp. Neurol. I42:417- 438.
Hjort-Simonsen, A., and B. Jeune (1972) Origin and termination of the hippocampal perforant path in the rat studied by silver impregnation. J. Comp. Neurol. I44:215-232.
Hunt, S., and J. Schmidt (1979) The relationship of a-bungarotoxin-binding activity and cholinergic termination within the rat hippocampus. Neu- rosci. 4.385-592.
Kimura, M., P.L. McGeer, F. Peng, and E.G. McGeer (1980) Choline acetyl- transferase-containing neurons in rodent brain demonstrated by immu- nohistochemistry. Science 208:1057-1059.
Kimura, M., P.L. McGeer, J.H. Peng, and E.G. McGeer (1981) The central cholinergic system studied by choline acetyltransferase immunohisto- chemistry in the cat. J. Comp. Neurol. 200:151-201.
Kohler. C., and L. Eriksson (1981) Efferent projections from the entorhinal area to the basal forebrain and frontal cortex originate in layer IV. Soc. Neurosci. 7:420 (abstract),
Kohler, C.. and V. Chan-Palay (1983) Distribution of gamma-aminobutyric acid containing neurons and terminals in the septal area. An immuno- histocheniical study using antibodies to glutamic acid decarboxylase in the rat brain. Anat. Embryol. 167153-65.
Kohler, C.. and H. Steinbusch (1982) Identification of serotonin and non- serotonin-containing neurons of the midbrain raphe projecting to the entorhinal area and the hippocampal formation. A combined immunoh- istochemical and fluorescent retrograde tracing study in the rat brain. Neuroscience 7:951-975.
Levey, A.I., B.H. Wainer, E.J. Mufson, and M.M. Mesulam (1983) Co-locali- zation of acetylcholinesterase and choline acetyltransferase in the rat cerebrum. Neurosci. 9:9-22.
Lorente de No, R. (1934) Studies on the structure of the cerebral cortex. 11. Continuation of the study of the ammonic system. J.Psychol. Neurol. 46:113-177.
Mellgren, S.I., and B. Srebro (1973) Changes in acetylcholinesterase and distribution of degenerating fibers in the hippocampal region after s e p tal lesions in the rat . Brain Res. 52:19-36.
Meibach, R.C., and A. Siege1 (1977) Efferent connections of the septal area in the rat: An analysis utilizing retrograde and anterograde transport methods. Brain Res. 119:1-20.
Mesulam, M.M. (1976aJ The blue reaction product in horseradish peroxidase histochemistry: Incubation parameters and visibility. J. Histochem. Cy- tochem. 24:1273-1280.

SEPTO-EhTORHINAL INTERCONNECTIONS 343
Mesulam, M.M. (1976b) A horseradish peroxidase method for the identifi- cation of the efferents of acetylcholinesterase containing neurons. J. Histochem. Cytochem. 24:1281-1286.
Mesulam M.M., and G.W. Van Hoesen (1976) Acetylcholinesterase-rich pro. jections from the basal fore-brain of the rhesus monkey to the neocortex. Brain Res. 109:152-157.
Mesulam, M.M., and E.J. Mufson (1980) The rapid anterograde transport of horseradish peroxidase. Neuroscience 5: 1277-1286.
Morell. J.I., L.M. Greenberger, and D.W. Pfaff (1981) Comparison of horse. radish peroxidase visualization methods: Quantitative results and fur. ther technical specifics. J . Histochem. Cytochem. 29:903-916.
Mosko, S., G. Lynch, and C.W. Cotman (1973) The distribution of septal projections to the hippocampal formation of the rat. J. Camp. Neural. 152163-174.
Oderfeld-Nowak, B., 0. Narkiewicz, J . Bialowas, J . Dabrowska, A. Wier- aszko, and M. Gradkowska (1974) The influence of septal nuclei lesion? on activity of acetylcholinesterase and choline-acetyltransferase in the hippocampus of the rat. Acta Neurobiol. Exp. 34:787-789.
Pepeu, G., A. Mulas, A. Rufi , and P. Satgui (1971) Brain acetylcholine levels in rats with septal lesions. Life Sci. IO:l81-184.
Powell, T.B.S., and W.M. Cowan (1955) An experimental study of the effer- ent connections of the hippocampus. Brain 78:115-135.
Raisman, G. (1966) The connections of the septum. Brain 89r317-348. Raisman, G., M.W. Cowan, and T.P.S. Powell (1966) An experimental anal-
ysis of the efferent projection of the hippocampus. Brain 89:83-108. Ropert, N., Y. Ben-Ari, K. Krnjevic, and P Pole (1980) Septohippocampal
pathway and cholinergic disinhibition of pyramidal cells. Can. J. Phys- iol. 11:118.
Rose, A.M., T. Hattori, and H.C. Fibiger (1976) Analysis of the septo-hippo- campal pathway by light and electron-microscopic autoradiography. Brain Res. 108r150-174.
Segal. M. 11977) Afferents to the entorhinal cortex of the rat studied by the method of retrograde transport of horseradish peroxidase. Exp. Neurol. 57.750-765,
Segal, M., and S.C. Landis (1974) Afferents to the hippocampus of the rat as studied with the method of retrograde transport of horseradish peroxi- dase. Brain Res. 78:l-15.
Sethy, V.H., M.J. Kuhar, R.H. Roth, M.H. Van Woert. and G.K. Aghajanian (1973) Cholinergic neurons: Effect of acute septal lesions on acetylcho- line and choline content of ra t hippocampus. Brain Res. 55:481-484.
Shute, C.C.D., and P.R. Lewis (1967) The ascending cholinergic reticular systems: Newortical, olfactory and subcortical projections. Brain 90:497- 521.
Siege], A,, and J.P. Tassoni (1971) Differential efferent projections of the lateral and medial septal nuclei to the hippocampus in the cat. Brain Behav. Evol. 4:201-219.
Srebro. B.. W. Harkmark, and C. Kohler (19791 Afferent and efferent projec- tions of the entorhinal cortex in the rat . Neurosci. Lett. Suppl. 3:143 (abstract).
Srebro, B., S.I. Mellgren, and W. Harkmark (1975) Distribution of acetylcho- linesterase in the septal region of the rat . In. J.F. De France (ed): First International Symposium on Septal Nuclei. Detroit: Wayne State Uni- versity Press.
Steward, 0. (1976) Topographic organization of the projections from the entorhinal area to the hippocampal formation of the rat. J. Camp. Neural. 167:285-314.
Steward, O., and S.A. Scoville (19761 Cells of origin of entorhinal cortical afferents to the hippocampus and fascia dentata of the rat . J. Camp. Neurol. 169r347-370.
S..anson, L.W.. and W.M. Cowan (1977) An autoradiogaphic study of the organization of the efferent connections at' the hippiicainpal formation in the rat. J. Camp. Neural. 17249-84.
Van der Kooy. D., and T. Hattori (1980) Dorsal raphe cells with collateral projections to the caudate-putamen and substantia nigra: A fluorescent retrograde double-labeling study in rat . Brain Res. 186:l-7.
Vincent. S.R., H. Kimura. and E.G. McGeer (19811 Organization of sub- stance P fibers within the hippocampal fbrmation demonstrated with a biotin-avidin immunoperoxidase techniquv. J. Comp. Neurol. 199:133- 143.
Vincent, S.R.. and E.C. McGeer (1982, A substance P prqiertion to the hippocampus. Brain Res. 215,349-351.
Walaas, I. (1981) Biochemical evidence for overlapping neocortical and allo- cortical glutamate projections to the nucleus accumbens and rostra1 caudatoputamen in the rat brain. Neuroscience 6:399-407.
Wyss. J.M., L.W. Swanson, and M.W. Cowan (1979) A study of subcortical afferents to the hippocampal formation in the rat. Neuroscience 4r463- 477.
Wyss, J.M. 119811 An autoradiographic study of the efferent connections of the entorhinal cortex in the rat. J . Camp. Neural. 199:495-512.
Zaczek, R., J.C. Hedreen, and J.T. Coyle (1979) Evidence for a hippocampal- septal glutamatergic pathway in the rat . Exp. Neural. 65r145-156.
NOTE ADDED IN PHOOF It has recently been shown that septal neurons in the MS
and dbB contain GAD immunoreactivity and have projec- tions to the hippocampal region, including the entorhinal area (Kohler C., V. Chan-Palay and J.Y. Wu (1984). Septal neurons containing glutamic acid decarboxylase immuno- reactivity project to the hippocampal region in the rat brain. Anat. Embryol. 169:41-44.