976 reports invest. ophthalmol. visual sci. void...
TRANSCRIPT
976 Reports Invest. Ophthalmol. Visual Sci.October 1977
Q_O
600
500
400
300
200
100
0
VOIDVOLUME
•
A
-\ 7V
I iIK
1;
!•
1
10 15MILLILITERS
20
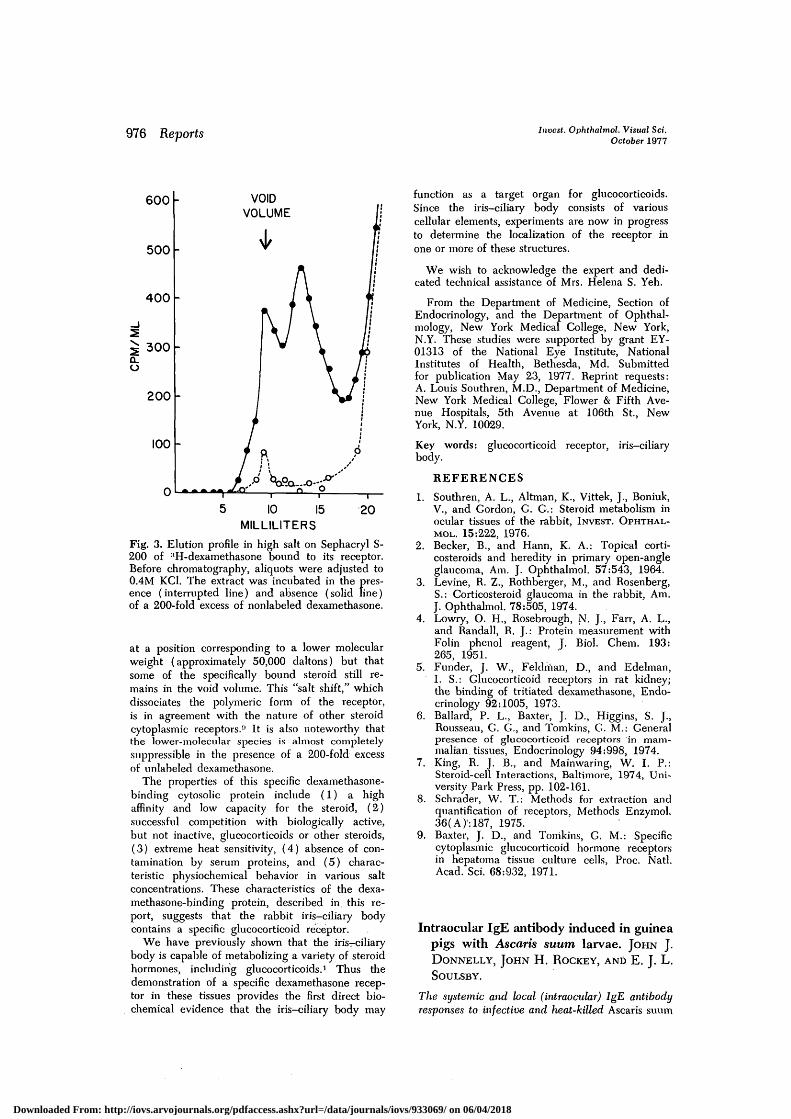
Fig. 3. Elution profile in high salt on Sephacryl S-200 of 8H-dexamethasone bound to its receptor.Before chromatography, aliquots were adjusted to0.4M KC1. The extract was incubated in the pres-ence (interrupted line) and absence (solid line)of a 200-fold excess of nonlabeled dexamethasone.
at a position corresponding to a lower molecularweight (approximately 50,000 daltons) but thatsome of the specifically bound steroid still re-mains in the void volume. This "salt shift," whichdissociates the polymeric form of the receptor,is in agreement with the nature of other steroidcytoplasmic receptors." It is also noteworthy thatthe lower-molecular species is almost completelysuppressible in the presence of a 200-fold excessof unlabeled dexamethasone.
The properties of this specific dexamethasone-binding cytosolic protein include (1) a highaffinity and low capacity for the steroid, (2)successful competition with biologically active,but not inactive, glucocorticoids or other steroids,(3) extreme heat sensitivity, (4) absence of con-tamination by serum proteins, and (5) charac-teristic physiochemical behavior in various saltconcentrations. These characteristics of the dexa-methasone-binding protein, described in. this re-port, suggests that the rabbit iris-ciliary bodycontains a specific glucocorticoid receptor.
We have previously shown that the iris-ciliarybody is capable of metabolizing a variety of steroidhormones, including glucocorticoids.1 Thus thedemonstration of a specific dexamethasone recep-tor in these tissues provides the first direct bio-chemical evidence that the iris-ciliary body may
function as a target organ for glucocorticoids.Since the iris-ciliary body consists of variouscellular elements, experiments are now in progressto determine the localization of the receptor inone or more of these structures.
We wish to acknowledge the expert and dedi-cated technical assistance of Mrs. Helena S. Yeh.
From the Department of Medicine, Section ofEndocrinology, and the Department of Ophthal-mology, New York Medical College, New York,N.Y. These studies were supported by grant EY-01313 of the National Eye Institute, NationalInstitutes of Health, Bethesda, Md. Submittedfor publication May 23, 1977. Reprint requests:A. Louis Southren, M.D., Department of Medicine,New York Medical College, Flower & Fifth Ave-nue Hospitals, 5th Avenue at 106th St., NewYork, N.Y. 10029.
Key words: glucocorticoid receptor, iris-ciliarybody.
REFERENCES
1. Southren, A. L., Altaian, K., Vittek, J., Boniuk,V., and Gordon, G. G.: Steroid metabolism inocular tissues of the rabbit, INVEST. OPHTHAL-MOL. 15:222, 1976.
2. Becker, B., and Hann, K. A.: Topical corti-costeroids and heredity in primary open-angleglaucoma, Am. J. Ophthalmol. 57:543, 1964.
3. Levine, R. Z., Rothberger, M., and Rosenberg,S.: Corticosteroid glaucoma in the rabbit, Am.J. Ophthalmol. 78:505, 1974.
4. Lowry, O. H., Rosebrough, N. J., Farr, A. L.,and Randall, R. J.: Protein measurement withFolin phenol reagent, J. Biol. Chem. 193:265, 1951.
5. Funder, J. W., Feldnian, D., and Edelman,I. S.: Glucocorticoid receptors in rat kidney;the binding of tritiated dexamethasone, Endo-crinology 92:1005, 1973.
6. Ballard, P. L., Baxter, J. D., Higgins, S. J.,Rousseau, G. G., and Tomkins, G. M.: Generalpresence of glucocorticoid receptors in mam-malian tissues, Endocrinology 94:998, 1974.
7. King, R. J. B., and Mainwaring, W. I. P.:Steroid-cell Interactions, Baltimore, 1974, Uni-versity Park Press, pp. 102-161.
8. Schrader, W. T.: Methods for extraction andquantification of receptors, Methods Enzymol.36(A)I:187) 1975.
9. Baxter, J. D., and Tomkins, C. M.: Specificcytoplasmic glucocorticoid hormone receptorsin hepatoma tissue culture cells, Proc. Natl.Acad. Sci. 68:932, 1971.
Intraocular IgE antibody induced in guineapigs with Ascaris suum larvae. JOHN J.DONNELLY, JOHN H. ROCKEY, AND E. J. L.
SOULSBY.
The systemic and local (intraocular) IgE antibodyresponses to infective and heat-killed Ascaris suum
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018

Volume 16Number 10
Reports 977
larvae were examined in guinea pigs. IgE anti-body, demonstrated by the 6-day P-K test, wasfound in anterior chamber fluid following primaryintravitreal injection of live second-stage A. suumlarvae and in some instances occurred when noserum P-K activity was demonstrable. No intra-ocular IgE antibody was induced by primaryintravitreal injection of heat-killed larvae. A briskintraocular and systemic IgE antibody responsefollowed the secondary intravitreal injection ofeither live or heat-killed larvae into animals sys-temically infected with A. suum. The guinea pigsystem promises to be a useful model for studyingthe immunopathology of human ocular parasiteinfections.
The intestinal ascarid of dogs, Toxocara canis,is the major cause of the human disease viscerallarva migrans, in which visual complications mayresult from the intraocular localization of theparasite.1 There is evidence that allergic mech-anisms play a role in the ocular reactions toparasites,2 but the individual roles played by thedifferent immunopathologic mechanisms are incom-pletely understood.
The present studies were undertaken to de-velop a model system to separate and define theroles of the different immunopathologic mech-anisms in ocular inflammatory reactions to para-sites. Ascaris suum infection of guinea pigs wasselected as a model for human infection withthe larval stages of T. canis because of the fol-lowing considerations. A. suum and T. canis areclosely related phylogenetically and antigenically:thus of 10 of the major diffusible antigens ofT. canis, six cross-react strongly with antigens ofA. suum.3 Detailed information is available aboutthe allergic reactions of the guinea pig, and theimmune response to A. suum infection in theguinea pig has been extensively characterized.4
IgE antibodies play a prominent role in immuneresponses to parasitic infections, and the IgEresponse to A. suum infection in the guinea pighas been studied in detail.5- ° The ready avail-ability of A. suum worms, the ability to growselected developmental stages in defined mediain vitro, and the similarity of the migrationpathways of the larval stages were further con-siderations in the selection. In the present report,we examine the induction of intraocular IgEantibodies with live and heat-killed A. . suumlarvae. .
Materials and methods. Female outbred (Hart-ley) guinea pigs weighing approximately 500gm. were obtained from Skippack Farms, Skip-pack, Pa.
Second-stage A. suum larvae were hatched invitro" from eggs by the method of Jas-koski arid Colucci,7 as modified by Leventhal andSoulsby,8 and suspended in phosphate-bufferedsaline, pH"7.4, at a concentration of 10,000 to
50,000 larvae/ml. The viability of the larvae wasverified by microscopic examination for larvalmotility. Only preparations containing more than80% viable larvae were used for "live larvae"experiments. Larvae were heat-killed when neces-sary by incubation at 56° C. for 20 min. Thelack of viability of the larvae was verified bymicroscopic examination.
Animals were systemically sensitized to larvaeby three subcutaneous injections of 10,000 A.suum infective eggs at 10-day intervals. Thisimmunization procedure results in a high degreeof resistance to reinfection and high levels ofIgE antibodies. Second-stage larvae hatch fromthe subcutaneously deposited eggs, some migratinglocally and others migrating to the lungs.9 Animalswere first bled by cardiac puncture, and anteriorchamber fluid was obtained by paracentesis, 10days after the last sensitizing injection of eggs.
A. suum antigen extract was prepared fromadult female worms according to the method ofWilliams and Soulsby.10
Intravitreal injection of 0.1 ml. of larvae sus-pension, containing 1,000 to 5,000 larvae, wascarried out under halothane anesthesia. Motilelarvae could be observed in the vitreous by di-rect ophthalmoscopy. Anterior chamber paracente-sis was performed under halothane anesthesia,and the 0.01 to 0.05 ml. of fluid obtained wasdiluted immediately in 0.1 ml. of sterile saline.This served to prevent clotting of anterior cham-ber fluid samples from inflamed eyes.
Prausnitz-Kustner (P-K) passive cutaneousanaphylactic tests were performed immediatelywith samples of aqueous humor. Each sample,diluted in 0.1 ml. of sterile saline, was injectedin toto into a single intradermal site on theshaved flank of an outbred female guinea pig.Saline and positive serum controls were includedon each test animal. P-K tests with samples ofserum obtained at the same time as paracentesiswere routinely performed on undiluted and 1:5diluted serum, and in some cases dilutions ofserum up to 1:1,000 were made. Recipient animalswere challenged 6 days later by intravenousinjection of 0.5 ml. of A. suum antigen extract(protein concentration, 0.5 mg./ml.) and 0.5 ml.of 10% Evans blue dye. Reactions were assessed30 min. later; the diameter of the area of bluingwas measured, and the degree of bluing wasassessed on an arbitrary scale of 1 to 4.
The 6-day P-K reaction has been shown to bespecific for guinea pig IgE antibodies, whichshare the following characteristics with IgE ofman and other species: fixation to mast cells inhomologous skin for 6 days or more, loss of ac-tivity after reduction of disulfides or heating, asedimentation coefficient of 8S, and elution froma Sephadex G-200 column before IgG immuno-globulins.6 The 6-day P-K test does not demon-strate guinea pig anaphylactic antibodies of the
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018

978 Reports Invest. Ophthalmol. Visual Set.October 1977
Fig. 1. Histopathology of an eye which had received a single intravitreal injection of liveA. suum larvae and was enucleated 12 days later. A, Parasites have penetrated into the choroid,producing a diffuse inflammatory reaction, with destruction of trie overlying retina. Serialsections of this eye were examined, but distinct retinal structures were not identified. Theinner layer of scarred, inflamed "choroid" has a distinctly different structure in some areas,suggesting that what is being seen here is both scarred choroid and retina. Examination ofother eyes where clearly identifiable retina merges into a similar inner, scarred, inflamedlayer on the choroid supports this suggestion. Some eyes show scarring and inflammation inretina detached by the subretinal collection of proteinaceous fluid, but such separation is theexception rather than the rule. Large irregular cystic changes throughout the retina are seenin other regions. B, Parasites in the anterior chamber, surrounded by acute inflammatory cells(including eosinophils) and chronic inflammatory cells {including histiocytes and epithelioidcells). Intact nucleated cells are visible in the alimentary tracts of parasites cut longitudinally,indicating that the larvae are still viable.
IgGl subclass, which fix at skin sites for 48 hr.or less, are not inactivated by heat or by disulfidereagents, and have sedimentation coefficients of7S and a molecular weight of about 150,000."
Results. When a single intravitreal injection of3,000 to 5,000 live A, suum larvae was given intoboth eyes, gross signs of ocular inflammation ap-peared within 3 days. These included conjunctivalinjection, hyperemia of the iris, and peripheralcorneal neovascularization. Within 2 weeks, hypo-pyon and extensive corneal vascularization ap-peared; after 4 weeks some animals developedocclusio pupillae and vascularization of the an-terior chamber, and a few eyes became phthisical.The histopathology of an eye infected with A.suum larvae is shown in Fig. 1. Aqueous humor,obtained at 7- to 9-day intervals, was tested for
the presence of IgE antibody, but because ofthe small amount of fluid (0.01 to 0.05 ml.)obtained from a single paracentesis, titration ofthe aqueous IgE antibody was not possible.Consequently, data are expressed as the proportionof the total number of eyes sampled which gavepositive P-K reactions. Positive reactions weredefined as 2 by 2 mm. in size or larger andhaving a dye accumulation of at least 2+. Of 55positive reactions obtained with samples of aqueoushumor, two measured 2 by 2 mm., two measuredbetween 2 by 2 and 4 by 4 mm., 28 measuredbetween 5 by 5 and 10 by 10 mm., and 23 mea-sured greater than 10 by 10 mm., with threemeasuring 20 by 15 and the largest lesion measur-ing 20 by 20 mm.
Intraocular IgE antibody was observed follow-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018

Volume 16Number 10
Reports 979
Aqueousw Serum v/w Humor t
* P K T i t e r PKSerum
t
Days* P-KTi ter P-K P-KTiter
10
17
AqueousHumor»P-K
14
2
4
1I
4
I4
2
•
10 50 100 )IOOO
1
•
pos. tc
Live Larvae!t
• 1
•
If
t.8
4
8
4
8
8
4
8
10-
14
2
4
10 50 100
• •
>IOOO
1
pos. tot.
II8
4
8
rrHeat-Killed Larvae"
4
4
• 1
. 1
il l r
31
38
Fig. 2. Serum and aqueous IgE (P-K) responses to systemic infection with A. suum andsubsequent intraocular challenge with live or heat-killed A. suum larvae. "Time in days afterlast systemic immunization. (Animals challenged with live larvae on day 24 had not been bledon day 10.) * "Serum P-K titers are expressed as histograms, with titer on the horizontal axisand number of animals demonstrating a given titer on the vertical axis. fThe number ofaqueous humor samples giving positive P-K reactions are represented by solid bars; the totalnumber of aqueous humor samples taken on a given day are represented by open bars, ft Ani-mals were challenged with 3,000 either live or heat-killed larvae by intravitreal injection afterparacentesis on day 24.
ing primary intraocular injection of live second-stage A. suum larvae. No IgE was detected inaqueous humor samples taken 7 days after intra-vitreal injection of larvae. However, three eyesout of 20 sampled gave positive P-K tests onday 14, and the maximum number of positiveocular P-K tests was six out of 20, which occurred29 days after infection (Table I) . P-K tests ofserum obtained by cardiac puncture at the sametime as the anterior chamber paracenteses showedno detectable IgE antibody until 19 days afterinfection (Table I) . Thus serum and aqueoushumor IgE appeared at approximately the sametime. However, in three eyes, ocular IgE anti-body was observed in the absence of serumIgE, and this suggests that intraocular IgE anti-body may be produced locally. The relative pro-portions of animals having IgE antibody in serumand aqueous are shown in Table I.
Migration of live larvae from the eye intoother organs of the guinea pig was demonstratedby the finding of live larvae in the lungs ofanimals 16 days after infection. To determine the
effect of systemic migration of larvae on theproduction of IgE antibody, the ocular and sys-temic responses to intravitreal injection of heat-killed A. suum larvae were studied. The grossinflammatory response to dead larvae was marked-ly different from that to live larvae. Beginningat 30 days after injection a mild inflammation,characterized by conjunctival injection and mildiritis, occurred. No IgE antibody was detected inthe aqueous humor at any time over the periodof 7 to 42 days after injection of dead larvae(Table I) . Serum samples, taken simultaneouslywith aqueous humor, did contain IgE antibodyat low titers with the frequencies shown in TableI. Thus the serum IgE response to intraocularinjection of dead larvae was not substantiallydifferent from that observed following the intra-ocular injection of live larvae.
A systemic immune response to A. suum in-duced by three subcutaneous injections of viableeggs elicited serum P-K titers ranging from 10to 1,000 or greater (Fig. 2). No IgE was detectedin the anterior chamber fluid of such animals,
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018

980 Reports Invest. Ophthalmol. Visual Sci.October 1977
Table I. Frequency of positive P-K reactions withserum and aqueous humor following a singleintravitreal injection of A. suum larvae
Inlravitrealinjection
Live larvae
Heat-killedlarvae
Days
114192629
7142142
SerumP-K test
No.pos.*
00213
0012
Totalno.f
1010101010
5555
Aqueous humorP-K test
No.pos.*
03*316
0000
Totalno.f
2020202020
1010.1010
"No. pos. = number of samples of serum or aqueousgiving positive P-K reactions.fTotal no. = number of animals (serum) or individualeyes (aqueous) tested for specific IgE antibody.(Positive P-K reactions measuring 5 by 4, 12 by 12, and20 by 20 mm., respectively, were obtained with aqueousfluid at 14 days, in the absence of positive P-K reactionswith undiluted serum from the same animals.
even in the presence of serum P-K titers of 1,000or more. Furthermore, multiple anterior chamberparacenteses did not induce the passage of IgEantibody into the aqueous humor (Fig. 2). Infec-tion of guinea pigs with viable second-stage A.suum larvae via the mesenteric vein, which alsoinduces serum P-K titers in excess of 1,000,likewise failed to elicit detectable IgE antibodyin anterior chamber fluid. Hence, it is apparentthat under normal ocular conditions, IgE anti-body does not leak from the peripheral circulationinto the aqueous humor.
Intraocular challenge of systemically sensitizedanimals 24 days after the last immunization, witheither 3,000 live or heat-killed A. suum larvae,reversed a decline in serum P-K titers (Fig.2). Thus three of four animals given live larvaehad serum P-K titers of 50 or greater on thefourteenth day after challenge, and three of fiveanimals given heat-killed larvae had similar titers(Fig. 2).
Ocular responses to intravitreal challenge alsowere dramatic in terms of gross inflammationand aqueous humor IgE antibody. The severityof the inflammation was essentially equal in ani-mals given live larvae and in those given heat-killed larvae. Within 2 days after challenge, pro-nounced flare and hypopyon were observed, withextensive corneal.vascularization, occlusio pupillae,and phthisis appearing within 2 weeks. After 2weeks, gradual clearing of the anterior chamberflare was observed, but the neovasculature andpupillary occlusions persisted. IgE antibody wasdetected in 90% of aqueous humor samples at7 and 14 days after challenge (Fig. 2). It waspresent with equal frequency in those animalschallenged with live organisms and in those given
heat-killed larvae: hence the serum and ocularresponses of these two groups of animals werevirtually identical.
Discussion. Heat-killed second-stage A. suumlarvae are clearly less able than live larvae toprovoke primary intraocular IgE antibody andinflammatory responses. This difference may re-sult from several factors. The reaction to livelarvae may depend on their ability to migratefrom the vitreous into uveal tissues and from theeye into other organs. In order to resolve thispoint, studies are now being conducted withthird-stage A. suum larvae, which are antigenicallysimilar to second-stage larvae but are probablynot able to leave the eye because of their largersize (1 mm. in length). Live larvae may providea different antigenic stimulus than heat-killedlarvae. Although it is possible but unlikely that asignificant heat-labile antigen is responsible forthe difference in the primary ocular response, theresponses of animals systemically infected withA. suum to intravitreal injection of live andheat-killed larvae are virtually the same, arguingagainst a significant role for a heat-labile antigen.However, it may be that antigens secreted and/orexcreted by living larvae do play a role in thedifference in the primary response. In this respect,a soluble antigen capable of eliciting high-titeranti-A. suum IgE antibody in guinea pigs hasbeen isolated from cultures, with the use of de-fined culture media, of third-stage A. suum larvaeundergoing the metamorphosis to fourth stage(Soulsby and Stromberg, unpublished observation).The ability of this antigen to induce IgE antibodyand histopathologic changes in guinea pig eyesis now under investigation.
Both cell-mediated and humoral immuno-pathologic mechanisms are probably involved inthe ocular response to parasites. The roles ofindividual mechanisms are now being studied bypassive transfer experiments. In our initial studies,we have observed that the ocular effects ofsystemic anaphylaxis in guinea pigs passivelysensitized with anti-A. suum IgE antibody arelargely limited to periocular and episcleral tissues(Rockey, Laties, Soulsby, and Donnelly, unpub-lished observations). This is consistent with ourobservation that IgE antibody is not found inaqueous fluid from normal eyes even in thepresence of high serum IgE antibody titers. Resultsfrom passive transfer studies will be reported indetail elsewhere. The guinea pig-A. suum systempromises to be a useful model for separating anddefining the roles of the various immunopathologicmechanisms in ocular inflammatory disease.
We are grateful to Ms. Norma Molina and Mr.Derek W. Muncey for their assistance in thepreparation of the parasites and to Mr. Nestor G.Menocal and Mrs. Dolores B. Ventura for theirassistance in preparing the histologic specimens.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018

Volume 16Number 10
Reports 981
From the Department of Ophthalmology, Schoolof Medicine, and the Department of Pathobiology,School of Veterinary Medicine, University ofPennsylvania, Philadelphia. This research wassupported by United States Public Health Servicegrants EYO1193 and AIO6262 and by a Fightfor Sight Student Fellowship, financed by a grantfrom Burroughs Wellcome Co. (U.S.A.), Inc., toFight for Sight, Inc., New York, N. Y. Submittedfor publication May 23, 1977. Reprint requests:Dr. John H. Rockey, Scheie Eye Institute, MyrinCircle, 51 North 39th St., Philadelphia, Pa. 19104.
Key words: Ascaris suum, toxocariasis model,ocular parasites, IgE antibody, six-day P-K test,human ocular immunopathology model, guineapig eye.
REFERENCES
1. Ash ton, N.: Larval granulomatosis of theretina due to Toxocara, Br. J. Ophthalmol.44:129, 1960.
2. Duguid, I. M.: Features of ocular infestationby Toxocara, Br. J. Ophthalmol. 45:789, 1961.
3. Jeska, E. L.: Antigenic analysis of a metazoanparasite. I. Extraction and assay of antigens,Exp. Parasitol. 20:28, 1967.
4. Khoury, P. B., and Soulsby, E. J. L.: Ascarissuum: immune response in the guinea pig. I.Lymphoid cell responses during primary in-fections, Exp. Parasitol. 41:141, 1977.
5. Dobson, C, Morseth, D. J., and Soulsby,E. J. L.: Immunoglobulin E-type antibodiesinduced by Ascaris suum infections in guineapigs, J. Immunol. 106:128, 1971.
6. Dobson, C, Rockey, J. H., and Soulsby,E. J. L.: Immunoglobulin E antibodies inguinea pigs: characterization of monomericand polymeric components, J. Immunol. 107:1431, 1971.
7. Jaskoski, B. J., and Colucci, A. V.: In vitrohatching of Ascaris suum eggs, Trans. Amer.Microscop. Soc. 83:294, 1964.
8. Leventhal, R., and Soulsby, E. J. L.: Cuticularreactivity of the early larval stages of A. suum:binding of fractions of immune serum to thesurface of infective and parasitic stage larvaeas detected by the mixed antiglobulin test,Int. J. Parasitol. 5:499, 1975.
9. Soulsby, E. J. L.: Immunization againstAscaris lumbricoides in the guinea pig, Nature179:783, 1957.
10. Williams, J. F., and Soulsby, E. J. L.: Anti-genic analysis of developmental stages ofAscaris suum. I. Comparison of eggs, larvae,and adults, Exp. Parasitol. 27:150, 1970.
Reflection of light by small areas of theocular fundus. ROBERT W. FLOWER, D.SCOTT MCLEOD, AND SHARON M. PITTS.
Light reflected from 50 nm spots on the fundi oftwo rhesus monkeys and three human volunteerswas measured as a function of wavelength in 10nm. jumps from 400 to 900 nm. The areas mea-
sured were a retinal artery and vein, areas of thedisk, macula, and retina devoid of visible blood,vessels, and the fovea.
A number of studies aimed at determiningreflection characteristics of the ocular fundus havealready been conducted on fundus fields as smallas 1 degree, with the use of a variety of tech-niques.1'4 However, as technological advancesmake it possible to observe and operate on smallerareas of the fundus, it becomes clearly moreimportant to know the optical properties of smallerdiscrete areas of the fundus. Hochheimer5 de-vised a method for measurement of reflectanceof small areas of the monkey fundus from fundusphotographs. We have refined that method tomeasure light reflected from 50 jum diameterareas of monkey and human fundi as a functionof wavelength and to determine the variability ofthese measurements among eyes widely varyingin density of fundus pigmentation.
Methods and proceduresModified fundus camera and film. Fundus photo-
graphs were made at 10 nm. intervals from400 to 900 nm. wavelength with a Zeiss funduscamera modified to permit placing narrow band-pass (about 10 nm. wide at 50 percent trans-mission) and neutral density filters in the path-way of light from the camera. Since transmissioncharacteristics of these filters are sensitive to theangle at which incident light strikes them, theywere placed in near-parallel light with the filtersurface normal to the light path. (Typically, tiltingthese filters up to 10 degrees off normal resultsonly in a 0.2 percent to 0.4 percent drop incenter wavelength with no change in bandwidthand an average reduction of less than 3 percentin transmission.) Illumination produced in thecentral retinal field in which reflectance measure-ments were made was measured to assure uni-formity. The same xenon flash-lamp was used asa light source for all experiments reported here,and its output stability was monitored by aradiometer mounted in the fundus camera hous-ing. All fundus photographs were made withKodak black and white, high-speed infrared-sensi-tive film (type 2841) always having the sameemulsion number. Film was developed in Kodakliquid x-ray developer for 3.5 min. at 70° F.
Characteristic curves for the photographic film(film transmission density vs. reflection density)were determined for each wavelength at whichfundus photographs were made, by photographinga Kodak calibrated gray paper scale using thesame series of narrow band-pass filters and de-velopment regimen used for the fundus photo-graphs. These characteristic curves were recalcu-lated during the course of the experiments toassure that the gamma values determined foreach wavelength were constant throughout thefilm batch.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933069/ on 06/04/2018