8 lecture mismatch repair 18 ug -...
TRANSCRIPT

1
Mismatch Repair
Mismatch-when conventional but noncomplementaryW-C bp occur in the double helix
8 classes:G-T, A-C pairs, cause transition mutationsG-G,A-A,T-T,C-C, C-T,G-A,cause transversion miut.
Insertion or deletion mutations are also recognized
Source- heteroduplexes formed during replication orrecombination
Transform with heteroduplex, found it was corrected
Wagner and Meselson-1976
Mismatches are subject to a repair system that dis-criminates newly synthesized from parental strands
Selective elimination of errors from newly synthesized DNA
Proposed the selectivity might be based on methylation
Methylation is a post-synthetic event, thus newly synthesized DNA is transiently hemi-methylated
N6-methyl adenine - dam methylase (GATC/CTAG)m5C-dcm methylase
dam mutants-hypermutable, overproducers also hypermutable
Miroslav Radman
Direct evidence - transform with heteroduplex DNA con-taining mismatched bases or small deletions and insertion
Mutants defective in correction in E. coli are:
mutH- binds hemi-methylated DNA, latent endonuclease
mutL-mediator, activates latent endo of mutH
mutS-recognize mismatch
xHemi-methylated Fully methylated Unmethylated

2
Mechanism
Paul Modrich, in vitro system- a cell free system that supportsmethyl directed reaction requiring mutH, mutL, and mutS
Strand specific process involving excision of long tracts, several kb
Adenine methylation d(GATC) modulates strandedness
Correction occurs on the unmethylated strand
Recognize mismatch, 97 kDa, AAA+ ATPase,promote loopBy sliding
Recognize GATC25 kDa
Latent endo
Cleaves unmethyla-ted strand-stimulatedby mismatch, mutL,and mutS
mutS
mutH
mutL
Architectural role70 kDa dimer
Excision reaction
Bidirectional - Either a 5’ to 3’ or a 3’ to 5’ nuclease, depending on which methyl group directs incision
Exonuclease. i.e. ssDNA end required
Therefore, a helicase involved, uvrD
Resynthesis reactionPol III

3
1993 Bert Vogelstein-two papers
1) Simple repeats are unstable in a set of sporadic colon cancer tumors
Deletions of up to 4 NT in poly(dA) tracts
Insertions and deletions in microsatellite sequences of di-or trinucleotide repeats
2) HNPCC-hereditary, nonpolyposis colorectal cancer- One of the most common cancer predisposition syndromesdevelop cancer before age 5020% of colon cancers, 2nd largest killer behind lung cancer
Role of heredity is established and 2 loci on chromosome 2had been linked to HNPCC in genetic studies
In studying the inheritance of the autosomal dominant marker,found that tumors developing in these individuals had alterations in their microsatellite sequences
They called this RER+, replication error phenotype
He dug in to clone this gene-positional cloning
One of the papers was published in the NYT
Paul Modrich noticed the similarity to mismatch repair in bacteria and to some yeast work (from Tom Petes)
In yeast, responsible for expansion and contraction of simple repeat sequences
Poly(dGT)10-30
Several human inherited disorders, fragile X syndrome,myotonic dystrophy, Huntington’s disease are associatedwith expansions of triplet repeats
Several other cancers are associated with alterations in dinucleotide repeat tracts

4
Collaborated to look for mut homologs in the region, which theypredicted should be affected by HNPCC mutations, and to show cell lines were defective in MMR
Collaborated to look for mut homologs in the region, which they predicted should be affected by HNPCC mutations, and to show cell lines were defective in MMR
hMSH2 (MutS Homolog) was found in the regionEvidence it was causative:
- in HNPCC patients, hMSH2 genes were mutated
-MSH2 segregates with HNPCC and is mutated in many sporadiccolon cancers
-Modrich showed that RER+ cells were defective in mismatch repair in vitro
- invented an assay similar to E. coli, strand specific MMR system dependent on MSH2- early step defective in RER+ cells
-3 additional genes, homologs of mutL (MLH) identified in otherHNPCC families
humanMutSα•heteroduplexcompleximagecourtesyofLorenaBeese
MechanismsinE.coli&humanmismatchrepair
PaulModrichHHMI&DeptBiochemistry
DukeUniversity
Modrich, P. J. Biol. Chem. 2006;281:30305-30309
Substrates and requirements for in vitro mismatch repair
5’ 3’ substrate
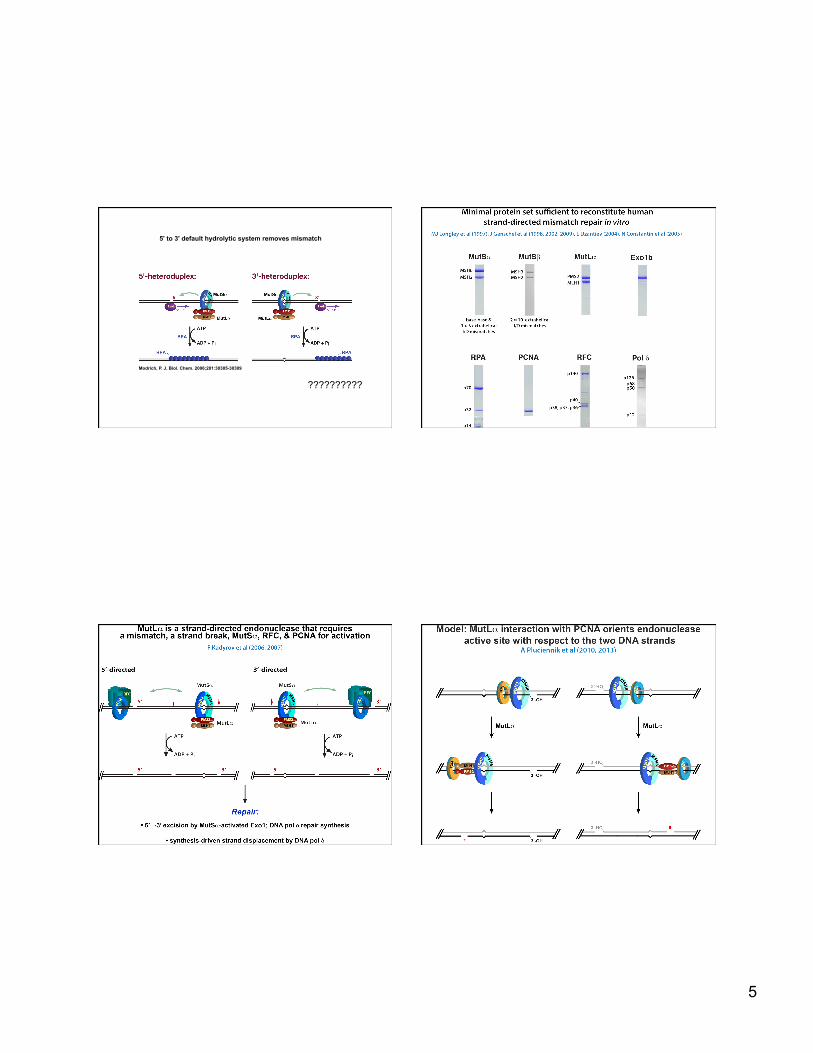
5
Modrich, P. J. Biol. Chem. 2006;281:30305-30309
5' to 3' default hydrolytic system removes mismatch
??????????

6
Conclusion:
Most theories of cancer attribute cancer to somatic andinherited genetic changes that lead to alterationsIn GROWTH CONTROL (tumor suppressor, oncogene)
Progressive somatic mutations are associated with several cancers
It has been postulated that an early step might confera mutator phenotype that leads to intrinsic genomicInstability (MMR mutants increase mutation rate 100-1000x)
Caveat: in yeast, mutations in mismatch repair genes led to increase in telomere maintenance by recombination - i.e. MMR tumors don’t need to reactivatetelomerase in order for tumor cells to proliferate
Gene rearrangement in cancer could be due to promotion ofillegitimate recombination between quasi-homologous sequences by MMR
Salmerichia - Salmonella and E. coli 5% apart but in mutmutants, they can recombine
Fischl et al. Cell 75, 10207-10038 (1993) KolodnerLeach et al. Cell 75, 1215-1225 (1993) VogelsteinParsons et al. Cell 75, 1227-1236 (1993) Modrich
DNA DAMAGE TOLERANCE Stalled Replication forks Upon replication fork stalling, PCNA is ubiquitinated to promote DNA damage toler
Chang and Cimprich, Nature Chem Bio 5,82 (2009)
Regulation of Post-replication Repair by PCNA Modification
Pathway choice is important: different modifications for different types of damage
PCNA
Chang and Cimprich, Nature Chem Bio 5,82 (2009)
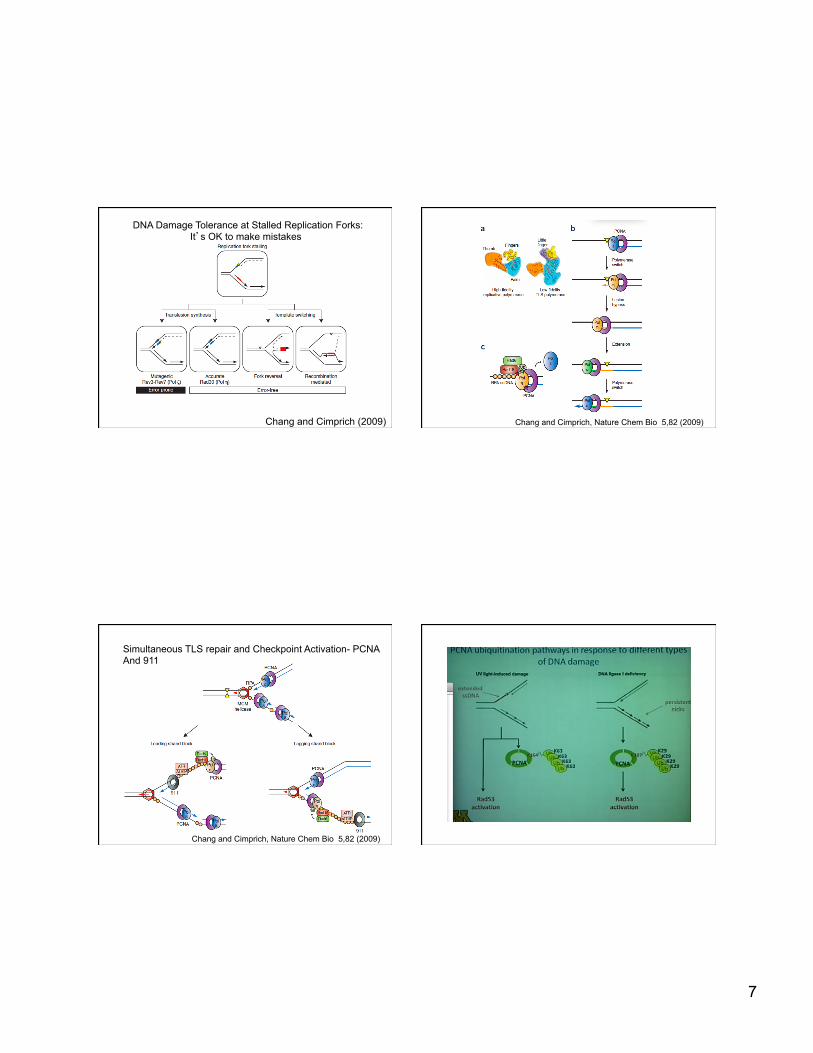
7
DNA Damage Tolerance at Stalled Replication Forks: It’s OK to make mistakes
Chang and Cimprich (2009)
Chang and Cimprich, Nature Chem Bio 5,82 (2009)
Simultaneous TLS repair and Checkpoint Activation- PCNA And 911
Chang and Cimprich, Nature Chem Bio 5,82 (2009)

8
d.
e.
f.
Translesion synthesis, Removal of unhooked X-link
DNA2 5’ to 3’ resection
RAD51 strand invasion
HR
a.
b.
c.
DNA replication
FancD2/FancI, Endonuclease,Unhooking of X-link
Interstrand crosslink Repair – The Fanconi anemia pathway
Long, Walter (2011)