72952511 el-mundo-electrico
TRANSCRIPT

1
El mundo eléctrico,una invitación a pensar
ÁNGEL A. CAPUTI
DEPARTAMENTO DE NEUROCIENCIAS INTEGRATIVAS Y COMPUTACIONALES
INSTITUTO DE INVESTIGACIONES BIOLÓGICAS CLEMENTE ESTABLE
Estudiar es pensar.
Sol W. Caputi1
IntroducciónAlgunos peces poseen órganos especializados para emitir electricidad
(llamados órganos electrogénicos, OE) y por lo tanto se los denominacomúnmente peces eléctricos. Algunas especies utilizan la descarga delórgano electrogénico (DOE) como arma para atacar una presa o repelerla agresión de un predador. Sin embargo, en la mayoría de los casos, lamagnitud de la descarga no ejerce efectos nocivos sino que es portadorade señales para el propio sistema nervioso. En nuestro país hay variasespecies de peces eléctricos de débil descarga. En el desarrollo de estetrabajo nos referiremos a la especie Gymnotus carapo. Esta especiehabita en casi todos los ríos y arroyos de agua dulce y ocasionalmentese la encuentra en el Río de la Plata en época de creciente de los ríosParaná y Uruguay.
Como la luz de la linterna que escudriña en la oscuridad, la DOE deestos peces nocturnos explora la presencia de objetos que difieren enconductancia con el agua. Al ocurrir la DOE, todo el cuerpo del pez setransforma en una fuente electrogénica distribuida. Dicha fuente generacorrientes eléctricas que atraviesan la piel. La presencia de objetosgenera una “imagen eléctrica” al modificar el perfil de corrientes, de lamisma manera que una diapositiva interpuesta en un rayo de luz arrojaimágenes sobre una pantalla. Receptores, ubicados en la piel soncapaces de transducir y codificar dicha imagen en forma de trenes depotenciales de acción que viajan a lo largo de los aferentes primarioshacia el sistema nervioso central. De esta forma los peces generan unarepresentación de la realidad inimaginable por la intuición humana.
Este artículo está dedicado a los Profesores de Enseñanza Secundariaparticipantes en el curso-taller organizado por PEDECIBA y UNESCO enla primavera del 2003. Expone algunas características de un sistemasensorial activo, expresado por una especie de peces autóctonos, y la
1. Sol W. Caputi, además de ser mi padre fue mi profesor de Historia Natural ensegundo ciclo de Educacion Secundaria. Comenzaba cada año sus clases inaugu-rales con esta frase. Siento la obligación de transferir dicha herencia cultural a lasnuevas generaciones de docentes.

2
potencialidad de dicho sistema como modelo didáctico en la enseñanzade la neurobiología.
La existencia de múltiples grupos de investigación científica queutilizan los peces eléctricos como modelo experimental facilita la consul-ta y la integración de docentes de enseñanza media con investigadoresprofesionales y puede permitir su entrenamiento en el diseño y desarro-llo de actividades docentes.
Por obvias razones, se eligieron algunos ejemplos, renunciando a laconsideración de otros, pendientes para próximos artículos. Propone-mos utilizar dichos ejemplos como excusa tanto para la elaboraciónteórica como para el diseño de demostraciones experimentales permi-tiendo integrar distintas visiones cruzadas desde diversas disciplinas.Pensamos que la consideración de los ejemplos elegidos desde el puntode vista de distintas asignaturas puede ayudar a los estudiantes acumplir la misión más importante que tienen al cursar enseñanzasecundaria: aprender a pensar.
El cuerpo del pez como fuente eléctricaAsí como la iluminación es necesaria para generar imágenes visuales
también lo es la electrogénesis para generar imágenes eléctricas. Por estarazón, entender los mecanismos de electrogénesis es un eslabón esencialpara comprender cómo los peces crean una imagen eléctrica del ambien-te que los rodea.
Los peces eléctricos, inspiradores de la famosa pila de Volta, nospermiten analizar fácilmente un acto efector y comprobar que la accióndel sistema nervioso es crear o coordinar una potencialidad, unapropuesta, cuya realización se concreta en cantidad y forma en tanto larealidad circunstancial se lo permite.
Un corolario del teorema de Thevenin-Norton es que dada una redeléctrica lineal y compuesta de elementos resistivos puede sustituirseparte de la misma por una fuente de voltaje (o fuerza electromotriz) y unaresistencia en serie (Teorema de Thevenin) o por una fuente de corrienteconstante y una resistencia en paralelo (Teorema de Norton). La resisten-cia en paralelo o en serie es similar, y su valor es igual al cociente entrela fuerza electromotriz correspondiente a la fuente de Thevenin y lacorriente entregada por la fuente de Norton.
Utilizando estas nociones puede establecerse la ecuación caracterís-tica de una fuente electrogénica. La capacidad electrogénica de un pezpuede evaluarse de la misma forma.
En el caso de una fuente eléctrica, por ejemplo una pila, se puedeaplicar una carga entre sus bornes y medir la potencia disipada enfunción de la carga. Supongamos el circuito de la Figura 1 en el cual unapila está conectada a una resistencia de carga cuyo valor puede cambiar.De acuerdo a la ley de Ohm la corriente (I) que circula a través de la

3
resistencia (Rc) es proporcional a la caída de voltaje (∆V) entre sus
extremos.
∆V= Rc.I (1)
Por otra parte, dado que se trata de un circuito en serie la corrienteI es igual al cociente entre la fuerza electromotriz (FEM) y la suma de lasresistencias en serie (R
s) y de carga (R
c).
I = FEM / (Rs + R
c ) (2)
Por lo tanto, la corriente I es una función monótonamente decrecientede la resistencia de carga, con asíntota igual a cero para latendiendo a infinito y la caída de voltaje ∆V es una función monótona-mente creciente de la resistencia de carga, con asíntota igual a la FEM
∆V = FEM.Rc / (R
s + R
c ) (3)
De ambas relaciones surge que la caída de voltaje es una funciónlineal y decreciente de la corriente
∆V = FEM – I. Rs (4)
Un análisis similar puede hacerse considerando una fuente decorriente que entrega una intensidad I
0. Dicha corriente se reparte a
través de Rs y R
c de modo que
(I0 – I).R
s=I.R
c (5)
y por lo tanto
∆V =I0 R
s – I.R
s (6)
Figura 1

4
Cualquiera que sea la carga aplicada entre los bornes de una pila lagráfica corriente-voltaje a través de la carga es una recta que interceptala ordenada en el valor de la fuerza electromotriz y la abscisa en el valorde la corriente máxima entregada por la fuente. La pendiente de dicharecta es el opuesto de la resistencia interna de la pila. Para un valorconcreto de resistencia, la pareja de valores (∆V, I) puede ser determina-da gráficamente como la intersección de la recta descripta por laecuación 1 y la recta descripta por las ecuaciones 4 y 6.
Se puede mostrar experimentalmente que la fuente equivalente alcuerpo de un pez eléctrico de la especie Gymnotus carapo puederepresentarse como una fuente eléctrica caracterizada por una fuerzaelectromotriz de valor variable en el tiempo y una resistencia en serierelativamente constante. Para ello basta con colocar el pez con su cuerpofuera del agua y su cabeza en un pequeño recipiente lleno de agua delestanque (para permitirle respirar normalmente). La punta del extremocaudal se sumerge en un segundo recipiente conteniendo agua (Figura2A). Ambos recipientes se conectan con una resistencia de carga de valorconocido y la caída de voltaje a través de la resistencia de carga se registraen un osciloscopio.
La gráfica del voltaje obtenido utilizando una resistencia de carga de1 MΩ se muestra en el inserto de la Figura 2B. Se trata de un pulso conuna fase principal cabeza positiva llamada V
3 cercada por deflexiones
negativas. La deflexión que antecede a la positiva tiene dos componentescuya separación se señala con la flecha. Las gráficas del voltaje obtenidoscon otros valores de resistencia de carga son similares en forma yescalables en amplitud de acuerdo a la ecuación 4.
De acuerdo a la ecuación l, el curso temporal de la corriente a travésde la resistencia de carga tiene una morfología similar al curso temporalde la caída de voltaje. Por lo tanto para cada valor de tiempo y deresistencia de carga puede medirse una pareja (∆V, I) y calcular paracada momento de la descarga la ecuación característica de la fuenteequivalente.
Figura 2

5
Las medidas obtenidas utilizando este procedimiento en el momentodel pico positivo se muestran en la Figura 2C donde se comparan lascurvas características obtenidas en dos peces, uno de 8 cm de largo y otrode 20 cm de largo. Como se puede observar, si bien ambas curvaspresentan la misma fuerza electromotriz, el pez grande tiene menorresistencia que el pequeño.
La razón de esta diferencia puede considerarse una cuestión deescala: supongamos que el cuerpo del pez se compone de dos elementos,a) una fuente de corriente, el órgano eléctrico y b) resistencia en paralelorepresentada por los tejidos no electrogénicos que lo rodean. Dado queexiste una similaridad geométrica (homotecia) entre peces de distintotamaño, la resistencia longitudinal del pez, que es proporcional al largoe inversamente proporcional a la sección (es decir al cuadrado del largo),disminuye con la longitud del pez.
Adicionalmente, al incrementarse el diámetro del pez en la mismaproporción que el diámetro del órgano eléctrico, el área electrogénicaaumenta y por lo tanto aumenta la corriente máxima entregada por elmismo. Sin embargo, el número de unidades electrogénicas es constantey por lo tanto la fuerza electromotriz no cambia. Estas observacionespueden verificarse midiendo el volumen de varios peces de tamañossignificativamente diferentes. Para ello se puede sumergir el pez cabezaabajo en un cilindro graduado y medir el desplazamiento de líquido.Posteriormente se puede correlacionar el volumen con el cubo de lalongitud para verificar la homotecia.
También pueden medirse las áreas de sección transversal obteniendosecciones transversales de 2 mm de espesor de un cadáver de pez fijadopor inmersión en paraformaldehído al 4%.2 Dichas secciones, quepueden realizarse fácilmente utilizando una hoja de afeitar nueva o unanavaja afilada, pueden observarse con una lupa o proyectarse utilizandoun sistema óptico sobre una hoja de papel para obtener dibujosrepresentativos. En la sección se puede reconocer una zona de aspectogelatinoso ubicada en la región ventral del pez, el órgano eléctrico que seextiende desde la región abdominal a la punta de la cola.
En otros peces igualmente fijados pueden realizarse corteslongitudinales (sagitales u horizontales) de la mitad ventral del cuerpodel pez. En estas preparaciones puede comprobarse que el número decélulas electrogénicas (llamadas electrocitos) en una porción similar depez (por ejemplo entre el centro del cuerpo y la unión del tercio medio yel tercio caudal) es constante e independiente del largo del pez.
Las células electrogénicas se disponen como las pilas en una linternade mano. Sin embargo, dichas células están encerradas en una vainaconductora que permite el drenaje lateral de corrientes. Cuando el pezestá en el agua las corrientes circulan además a través de la piel del pez.
2. Idealmente por perfusión transaórtica.

6
De esta forma se comporta como un generador longitudinalmentedistribuido y representable como una serie de fuentes. Podemos concluirentonces que la medida de la fuente equivalente entre el extremo rostraly el extremo caudal del pez es global pero no necesariamente apropiadapara describir el cuerpo del pez como fuente distribuida.
La corriente que cada electrocito aporta es un porcentaje de lacorriente total que el OE genera. Debido a que los electrocitos sonpequeños y estan inmersos en tejidos conductivos, la caída de potenciala lo largo del pez aislado en el aire (fuerza electromotriz) presenta unaforma suave y el número de fuentes que caracterizan al cuerpo del pezpuede considerarse infinito. No obstante, una representación con ele-mentos discretos es suficiente para predecir con bastante precisión elcampo eléctrico generado (véase más abajo).
La fuerza electromotriz de dichas fuentes se puede medir utilizandoun arreglo experimental idéntico al utilizado para medir la fuerzaelectromotriz global pero registrando la caída de voltaje entre dos puntosdel cuerpo del pez mantenido en el aire. De esta forma se puede evaluarpor registro directo la fuerza electromotriz de trozos del cuerpo del pezcorrespondientes a un cierto porcentaje (por ejemplo 25%). Como se veen la Figura 3, el curso temporal de la fuerza electromotriz es diferentepara las distintas porciones de pez indicando que la estructura y funciónde los distintos trozos de órgano eléctrico son diferentes.
Figura 3
Hemos comprobado que la fuerza electromotriz que genera la activi-dad del órgano eléctrico es variable en el tiempo, y más aún, estágenerada por distintas fuentes ubicadas a lo largo del cuerpo del pez.Dichas fuentes generan fuerza electromotriz de distinta amplitud yforma de onda.
Por otra parte, la consideración de la forma del pez a la luz de losargumentos arriba expuestos sugiere que la resistencia interna tampoco

7
está homogéneamente distribuida. Esta hipótesis puede comprobarseexperimentalmente midiendo la caída de potencial entre el polo rostraly una serie de puntos ubicados a lo largo del cuerpo de un espécimenfijado mientras se aplica una corriente longitudinal de amplitud cons-tante. Los resultados obtenidos pueden modelarse utilizando una fun-ción polinomial de tercer grado.
Realizando estos experimentos en clase puede comprobarse que elcuerpo del pez actúa como una fuente distribuida. La corriente generadapor esta fuente distribuida atraviesa la piel (cuya resistencia puedeconsiderarse despreciable con respecto a la resistencia del agua) dondeestán ubicados electrorreceptores cutáneos que miden dicha corriente.La circulación de dichas corrientes genera un estímulo basal, pulso-a-pulso, de los electrorreceptores.
¿Es la electrogénesis particular, sólo observable en algunos peces? O,¿se trata de un ejemplo particular de un fenómeno general, la generaciónde actos efectores?
Un acto efector (motor o electrogénico) puede definirse como larealización de un trabajo sobre el mundo exterior. Potencia y trabajo sonmagnitudes que sintetizan un aspecto de determinado acto. Estasmagnitudes permiten comparar distintos actos y distintos tipos deenergía utilizando una única dimensión. No obstante, no lo describen.
Describir un acto implica describir el curso temporal de las variablescuya integración a lo largo de una trayectoria o tiempo resulta en trabajo.
En el caso de la fuente equivalente de la pila o del pez, la potencia esel producto de la corriente que circula a través de la resistencia de cargapor la caída de potencial que dicha corriente genera. Es decir, en laspeculiares circunstancias de nuestro experimento el acto electromotortiene dos componentes: corriente y voltaje. Esto no es una rareza, enefecto en otros sistemas como por ejemplo el control motor de unmúsculo o una articulación, también intervienen dos variables: fuerza yvelocidad en el primer caso, o torque y velocidad angular en el segundo.
Es importante destacar que la magnitud del trabajo realizado es sólomedible a posteriori de su realización. En efecto, el trabajo se calculaintegrando a lo largo del tiempo la potencia disipada. Dado que lapotencia disipada es función de elementos externos cuya magnitud es apriori impredecible, el trabajo lo es también.
No obstante, una de las características del sistema nervioso es la deimplementar sistemas cognitivos predictivos. Por esta razón, los sereshumanos hemos desarrollado la idea de potencial. El potencial es lacapacidad de actuar (de realizar trabajo) de un sistema enfrentado a unacarga dada. Dicho potencial se ha definido de distintas formas para losdistintos tipos de energía que los humanos generalmente concebimos:surgen así la fuerza mecánica, el potencial eléctrico el potencial químico,etcétera.

8
En nuestro experimento, dicha potencialidad es medible directamen-te. Cuando la carga opuesta impide la realización del acto electromotor(resistencia de carga infinita) la corriente por el circuito es nula. En estascircunstancias podemos medir uno de los parámetros que describen lacapacidad del pez para hacer circular corriente a través de la resistenciade carga. Sin embargo, potencialidad para realizar un determinado actono es el acto en sí. Un acto se realiza de la forma que las circunstancias(en nuestro caso la resistencia de carga) lo permiten. En el ejemplo de lapila, la potencia disipada por la resistencia de carga es máxima cuandoésta es igual a la resistencia en serie. El ejemplo nos enseña quecualquier sistema que ejerce un trabajo tiene dos tipos de acciones: unassobre objetos externos y otras sobre sí mismo. Podemos concluir queparte del trabajo siempre se realiza internamente. Por lo tanto la medidade la potencialidad de realizar trabajo incluye no sólo la fuerza electromotrizsino además la resistencia interna de la fuente.
En la realidad, la ejecución del acto electromotor, siendo la expresiónde una potencialidad estereotipada y genéticamente determinada, de-pende solamente de las circunstancias: sus cambios indican cambios enla escena que rodea al animal.
La medida de dichos cambios por un sistema sensorial adaptado pararesponder a las características temporo-espaciales de la descarga delórgano eléctrico permite la elaboración de una representación del mundobasado en la presencia de inhomogeneidades en la distribución deimpedancia en el agua que rodea al pez.
Adicionalmente, la corriente generada por esta fuente distribuidaatraviesa la piel donde están ubicados electrorreceptores cutáneos quemiden dicha corriente. La circulación de dichas corrientes estimula,pulso-a-pulso, a los electrorreceptores. Dicho estímulo depende de ladistribución de impedancia del medio, y provee al animal de informaciónsobre el mismo. La presencia de un determinado objeto imprime cambiossobre esta distribución. A dichos cambios del estímulo se los denomina“imagen eléctrica del objeto”.
La formación de imágenes por modulación de una portadoraautogenerada, tampoco es una caracteristica especial de los peceseléctricos. Pensemos por ejemplo las plantas de los pies, las palmas delas manos, o el cuerpo todo cuando nos zambullimos en el agua. En estoscasos la fuerza se distribuye en la superficie generando un patrón depresión sobre el cual objetos (por ejemplo una piedra en el zapato)generan una imagen mecánica.
El mundo eléctrico de los pecesEn el caso del sentido activo de los peces eléctricos, la descarga del
órgano eléctrico actúa como portadora de señales sobre el mundoexterno de la misma manera que la corriente que circula por una línea

9
telefónica actúa como portadora del lenguaje. La capacidad de respuestade los electrorreceptores ajusta a las características de la descarga delórgano eléctrico como las de un sintonizador a las de las ondas de unaradioemisora. Si se coloca un objeto en un determinado sitio de un fluidoconductor y en presencia de un campo eléctrico, dicho campo se modificaen función de tres variables principales: a) la intensidad y dirección delcampo basal, previo a la colocación del objeto en las cercanías en dichositio; b) la conductividad del objeto y c) la forma de dicho objeto. Modificarel campo tal como lo hace el objeto es equivalente a crear un nuevo campoeléctrico perturbador del campo basal. A la fuente eléctrica que alubicarse en el lugar del objeto genera un campo idéntico al campoperturbador se lo ha denominado impronta eléctrica del objeto.3
La presencia de objetos en la cercanía del cuerpo del pez causacambios en el campo y estos cambios se “proyectan” sobre la piel comouna imagen eléctrica física. Seguidamente describiremos algunos aspec-tos de los mecanismos de generación de dichas imágenes.
Consideremos este ejemplo: supongamos que el pez se encuentra en uncilindro cerrado de diámetro ligeramente mayor al diámetro de su sección.Podemos postular el modelo eléctrico que se observa en la Figura 4.
Dicho modelo supone que el espacio puede subdividirse en elementosdiscretos. En el caso representado el cuerpo del pez se representa portres elementos, la piel por cuatro y el medio exterior por otros tres. Esimportante destacar que los valores de estos elementos dependen de lasección del cilindro y de la longitud y sección del pez (todas estos valorespueden ser experimentalmente medidos).
En una red eléctrica de este tipo, la carga sobre la cual actúa cadafuente consiste en un arreglo compuesto por las resistencias internas detodas las otras fuentes, la resistencia de la piel y las resistenciasexternas. Extendiendo la observación que hacíamos previamente pode-
Figura 4
3. Recomendamos la lectura de los trabajos iniciales de Lissmann, H. W. (1958). “Onthe function and evolution of electric organs in fish”, J. Exp. Biol. 35, 156-191 y deLissmann H.W., Machin K.E., “The mechanism of object location in Gymnarchusniloticus and similar fish”, J. Exp. Biol. 35 (1958) 451-486.
1 2
3 4

10
mos decir que cada fuente es un lastre, para sí misma y también para lasotras fuentes. Podría razonarse que la corriente generada está enfrenta-da a circular a través de las resistencias internas de las otras fuentes oa través de las resistencias de la piel hacia el exterior. La proporción decorriente en cada rama puede ser calculada estableciendo un sistema deecuaciones basado en tres principios: 1) la regla de los nodos de Kirchoff:para cada nodo (es decir para cada contacto de tres o más elementos) lasumatoria de las corrientes que circulan por cada una de las ramasconvergentes al nodo es nula; 2) la ley de Ohm: la corriente que circulaentre los extremos de un elemento es igual al cociente de la caída depotencial y la resistencia del mismo (es útil sustituir la variable resisten-cia por su inverso, conductancia); 3) cuando entre dos nodos hay unafuente la caída de potencial entre ambos es igual a la caída de potencialcausada por la circulación de corriente más el valor de la fuente.
En nuestro modelo, el sistema tiene entonces 4 ecuaciones y 4incógnitas correspondientes al potencial en cada nodo. Obviamente estesistema es indeterminado dado que existen múltiples cuartetas devalores que satisfacen su solución. Puede entenderse fácilmente elfundamento físico de esta indeterminación: Si un punto cualquiera dedicha red es conectado a tierra (potencial nulo) a través de una bateríade valor controlable, puede observarse que podemos adicionar infinitosvalores de potencial basal a la solución particular que considera elpotencial nulo en dicho punto. Se levanta la indeterminación conside-rando que uno de los nodos tiene potencial nulo.
Este ejemplo nos muestra tres hechos 1) la corriente neta a través dela piel es nula (comprobación del teorema de Gauss); 2) si cambia laresistencia de uno de los elementos (como ocurriría por la presencia deun objeto) la contribución de cada una de las fuentes a la circulación decorriente en cada uno de los elementos se modifica; 3) los cambiosmayores ocurren en las cercanías de la resistencia que se modificó.
La impronta resultante al cambiar un valor de resistencia en elmodelo simple previamente expuesto, es la diferencia causada por elcambio de resistencia en los potenciales de cada uno de los nodos. Estadiferencia en la distribución de potenciales en cada uno de los nodospertenecientes al circuito corresponde a otro circuito cuyos elementospasivos (resistencias) son idénticos pero cuyo elemento activo consisteen una fuente en el sitio en el cual estaba la resistencia modificada.
Para representar una situación más realista pueden considerarsemodelos más complejos.4 Esto permite calcular la corriente transcutáneacuando el pez nada en un cierto espacio. Las conclusiones generalesobservables utilizando estos modelos son las mismas. Utilizando mode-los físicos de este tipo hemos demostrado que la imágenes eléctricas deobjetos compactos tienen forma de sombrero con la copa enfrentada al
4. Véase para este punto Rother D. 2002, tesis de Maestría, PEDECIBA, Uruguay.

11
objeto y orientada en sentido opuesto al ala. La copa representa unaumento (cuando el objeto es más conductivo que el agua) o unadisminución (cuando el objeto es menos conductivo que el agua) de lacorriente transcutánea. La Figura 5 representa el campo eléctricogenerado por un pez eléctrico en un instante, la deformación que unobjeto imprime sobre el mismo. Para simplificar la imagen se haconsiderado exclusivamente un plano horizontal y un instante corres-pondiente a una onda (V
3). Por esta razón el campo basal y el causado por
la impronta del objeto se representan en 2 dimensiones (en lugar de 3).
La imagen eléctrica se parece más a una sombra que a la imagenproyectada por una cámara de cine en una pantalla. Esto se debe a queen el caso de las imágenes eléctricas el sistema equivalente a la lente deproyección no existe. El diámetro de la copa es prácticamente idéntico aldel objeto cuando el objeto toca la piel y se ensancha al alejarlo. La alturade la copa es máxima cuando el objeto está en contacto con el pez y caerápidamente con la distancia. Por esta razón la pendiente relativa de la
Figura 5

12
transición entre copa y ala cae con la distancia y es utilizada por algunospeces como clave para medirla.5
En la Figura 6 se representa la imagen eléctrica normalizada pico apico y calculada a lo largo de una de sus dimensiones.
Distancia, forma, y tamaño son sólo algunos atributos que percibi-mos de los objetos que nos rodean. Dichos atributos se denominanprimarios porque pueden ser evaluados por un conjunto de imágenesobtenidas con un único subtipo receptorial en presencia de la mismaportadora.
Existen otros atributos que para ser detectados requieren la combi-nación o el procesamiento neural conjunto de varias imágenes prima-rias. Por ejemplo el color visto por algunos humanos, resulta de lainteracción de las imágenes físicas detectadas por tres tipos defotorreceptores llamados conos y un tipo llamado bastones. Este tipo decolor se basa entonces en el procesamiento conjunto de cuatro imágenessensoriales primarias. Variantes genéticas (llamadas daltonismo) deter-minan que otros hombres posean dos o sólo un tipo de conos y por lotanto perciban en forma diferente la misma realidad. Dichas variantesgenéticas son capaces de procesar menos imágenes sensoriales prima-rias y tienen, por lo tanto, una visión del color diferente entre sí y con loshumanos portadores de tres tipos de conos.
Análogamente al color pueden definirse cualidades secundarias paraotras modalidades sensoriales. Los peces eléctricos tienen distintos tiposde electrorreceptores cutáneos que analizan en forma diferente laimagen física, generando para cada grupo de receptores una imagensensorial. Existen entonces varias imágenes eléctricas sensoriales defi-nidas por la capacidad de los receptores de responder a la portadora ya sus modificaciones por inhomogeneidades de la impedancia en elentorno del pez. De la misma manera que existe color, olor o sabor, laspropiedades físicas de los objetos y las características de forma de ondade la descarga determinan la posibilidad de discriminar un atributo de
Figura 6
5. Véase von der Emde, G., Schwartz, S., Gómez, L., Budelli, R., Grant, K. (1998)“Electric fish measure distance in the dark”. Nature 395: 890-394.

13
los objetos relacionado con su impedancia. Por ejemplo en el circuito quehemos analizado podría sustituirse una resistencia por una impedanciaque tuviese elementos capacitivos (por ejemplo un condensador, Figura7).
Dado que se trata de un pulso, la respuesta del condensador esdistinta que la de una resistencia. Por lo tanto, la imagen difiere en el pesorelativo de cada una de las ondas de la misma manera que en una imagencoloreada cada “tinta” tiene distinto peso. Hemos llamado “color eléctri-co” a dicha cualidad secundaria.6
Como hemos visto en los modelos, la amplitud de la imagen de unobjeto cilíndrico con una de sus bases enfrentada a la superficie de la pieltiene forma de sombrero mexicano de modo que la copa es funcióncreciente de la conductancia longitudinal del objeto.
Dicha predicción puede verificarse en el laboratorio. Para ello seutiliza un pez de aproximadamente 12 a 15 cm de longitud y se loenvuelve en “papel higiénico” y se lo coloca en un corral de tul dentro deun estanque conteniendo una solución de cloruro de sodio al 0,5 por diezmil en agua destilada. De esta forma el pez esta impedido de navegar y
Figura 7
6. John Locke (1632-1704), introduce esta terminologías, postulando que cuandorepresentamos objetos combinamos tres clases de ideas: cualidades primarias,cualidades secundarias y potencialidad: “...Lo amarillo no se encuentra en el oro,sino que es un poder del oro para producirnos esa idea a través de la vista cuandoestá iluminado de manera adecuada; y el calor que no podemos eliminar de nuestraidea del Sol, realmente no está más en el Sol que el color blanco que produce enla cera”. Calor, color, olor, sabor y sonido son apariencias de los objetos a nuestrossentidos. Potencialidades que sólo se realizan en la mente del observador. Lockellama a estas cualidades de los objetos cualidades secundarias para distinguirlasde las llamadas cualidades primarias que son aquellas que no sólo se perciben através de los sentidos (por ejemplo: extensión, forma, movimiento o reposo ynúmero) sino cuya existencia implica necesariamente un estar, es decir ocupar unlugar en el espacio y en el tiempo. Sin embargo, las mismas nociones de espacioy tiempo son relativas al observador y, además, pueden ser solo una creación dela mente. Así, llevado a uno de sus límites, el empirismo iniciado por Lockedesemboca en el idealismo absoluto ya que el límite entre las cualidades primariasy secundarias puede considerarse absolutamente artificial.

14
la conductividad del agua es aproximadamente 100 microS/cm. Laamplitud de la imagen puede estimarse como la diferencia de camposeléctricos perpendiculares a la piel. Para ello se colocan muy cerca delpez un par de alambres finos (de aproximadamente 100 micras dediámetro, de transformador) aislados excepto en la superficie de corte ycuyos extremos estén ubicados sobre una recta perpendicular a la piel,separados 2 mm. Se mide la caída de potencial utilizando un amplifica-dor diferencial cuya salida se monitoriza en un osciloscopio.7
Si se acercan distintos objetos se pueden observar los cambios en laamplitud de la descarga. Si se utiliza un objeto cilíndrico cuyo contenidoy cara lateral no sean conductoras y sus bases sí sean conductoras (comoel que se muestra en la Figura 8A) se puede verificar la relación existenteentre resistencia del objeto y la amplitud de la imagen física. Para ello secoloca el cilindro-estímulo con una de sus bases enfrentadas a la región
Figura 8
A
B
7. Si se carece de este instrumento puede utilizarse un pequeño radiorreceptor. Paraello se abre la caja del mismo y se identifica el reóstato que controla el volumen.Se conecta uno de los cables al punto medio y otro a masa. Al mover la perilla deldial a un extremo se oirá la descarga del pez. Si se mide con un téster colocado enposición AC la caída de potencial en el parlante dicha medida es un índice de laamplitud de la descarga local.

15
donde se está midiendo la imagen eléctrica. Al conectar sucesivamenteentre sus bases resistencias de valor conocido se pueden medir lascorrespondientes amplitudes de la señal registrada en el osciloscopio
Análogamente se puede verificar, si se dispone de un osciloscopio,cómo las capacidades modifican no sólo la amplitud sino también laforma de onda de la señal registrada. Para ello se coloca un condensadorentre las bases del cilindro, y se registra en el osciloscopio el campo localenfrente del objeto.
En la Figura 8B se observan distintas formas de onda correspondien-tes a circuito abierto (izquierda), carga resistiva (22 kΩ centro arriba),carga capacitiva (10 nF, centro abajo) y cortocircuito. Las flechas indicanque el camino entre circuito abierto y cortocircuito puede recorrerse condiversidad de cargas y que una capacidad muy alta es equivalente a unaresistencia muy pequeña. La flecha vertical en gris indica que es posiblemodificar forma de onda sin cambiar amplitud de la señal. Los peces soncapaces de discriminar forma de onda. Dicha capacidad sugiere que lospeces pueden evaluar una cualidad de la señal asociada a la impedanciade los objetos. Es decir los peces podrían percibir una forma de “color
eléctrico”.
Memoria y discriminación de las imágenes eléctricas¿Cuán relevantes son para el pez las imágenes físicas que acabamos
de describir? Para responder a esta pregunta debemos primero subrayar5 conceptos básicos de fisiología sensorial: 1) detectar es contestar lapregunta ¿hay algo ahí?; 2) discriminar es contestar la pregunta ¿es estodistinto de esto otro?; 3) describir es contestar a la pregunta ¿cómo esesto? 4) localizar es contestar a la pregunta ¿dónde está esto?; y5) reconocer es contestar la pregunta ¿qué es esto? Cada una de estasfunciones requiere un procesamiento sensorial de grado cada vez máselevado.
Una de las herramientas adecuadas para evaluar algunas de estasfunciones fue diseñada por el físico y fisiólogo alemán Fechner en 1864a punto de partida de las observaciones de Weber, quien descubre querelación entre el peso de un objeto y su mínimo incremento detectable esconstante para un amplio rango de objetos, y que existe un peso mínimo,umbral, por debajo del cual no existe sensación de peso.
Fechner razona que la sensación o efecto psíquico causado por undeterminado estímulo puede cuantificarse suponiendo que la mismamínima sensación ∆S es causada por el mismo incremento relativo delestímulo ∆E/E (con E notando al estímulo y ∆E a su incremento). Estoimplica que el cociente incremental ∆E / ∆S para los mínimos incremen-tos medibles es proporcional al estímulo E. Fechner supone que laderivada de la sensación con respecto al estímulo es inversamenteproporcional al estímulo:

16
δS / δE = K/E (7)
de modo que la sensación es una función medible del estímulo:
S = K log (E) + C (8)
y como en el umbral absoluto Eo la sensación tiende a cero
C= -K log (E0) (9)
Y entonces
S = K log (E/E0) (10)
Finalmente, Fechner concluye que la sensación, una característicade la psiquis del sujeto, puede ser objetivamente medida utilizando lasreacciones del sujeto frente a distintos estímulos.
Las observaciones de Weber distan de ser generalizables para todaslas modalidades sensoriales en todo su rango dinámico y el razonamien-to de Fechner se basa en supuestos razonables pero no necesariamenteválidos desde el punto de vista cuantitativo. Sin embargo, el experimentoclásico de Weber y la aproximación teórica de Fechner dio origen a unanueva disciplina por medio de la cual se pueden evaluar en formaexperimental y comparable variables representativas de los procesospsíquicos: la psicofísica.
Utilizando técnicas psicofísicas hemos podido verificar que los pecesson capaces de detectar, discriminar y hasta cierto punto cuantificar lasdiferencias entre dos objetos.8 Sin embargo, la regla logarítmica deFechner no es válida en este caso. Como índice de detección hemosutilizado un tipo de conducta conocido como respuesta de novedadconsistente en la ejecución de un acto motor exploratorio disparado porun cambio en el entorno sensorial del animal. Son respuestas denovedad mirar o girar las orejas en la dirección desde donde proviene unruido, mirar hacia un objeto que ingresa en el campo visual. En el casode los peces eléctricos, ya los primeros trabajos de Lissmann, se reportanrespuestas de novedad caracterizadas por el incremento súbito de lafrecuencia de repetición de la descarga. Dichas respuestas se provocanpor cambios en las características eléctricas de los objetos cercanos.
La técnica utilizada consistió en estudiar la probabilidad y la ampli-tud (es decir la magnitud del aumento de la tasa de repetición de ladescarga del órgano eléctrico) en función del cambio en la imageneléctrica a partir de distintas imágenes basales. Dicha técnica se puedeimplementar en un salón de clases prácticas de enseñanza secundariasiempre que los grupos de estudiantes sean lo suficientemente reducidos
8. Véase P. Aguilera, tesis de Doctorado PEDECIBA.
17
para evitar estímulos vibratorios generados por la discusión en voz altay la repercusión a través del piso de los movimientos de distintosintegrantes del grupo. Ya hemos explicado cómo medir la imageneléctrica. En este caso el cambio de resistencia debe realizarse utilizandoun relé para poder registrar en forma precisa el momento en que dichocambio ocurre. La amplitud de la respuesta de novedad se mide por lareducción porcentual del intervalo entre pulsos inmediatamente alcambio de resistencia del objeto.
Figura 8
A
B

18
Para medir el tiempo de ocurrencia del cambio de resistencia y lostiempos de descarga del órgano eléctrico se requiere un programa decomputación que permite adquirir dichos sucesos a través del puertoserie o del puerto paralelo de un computador digital.9
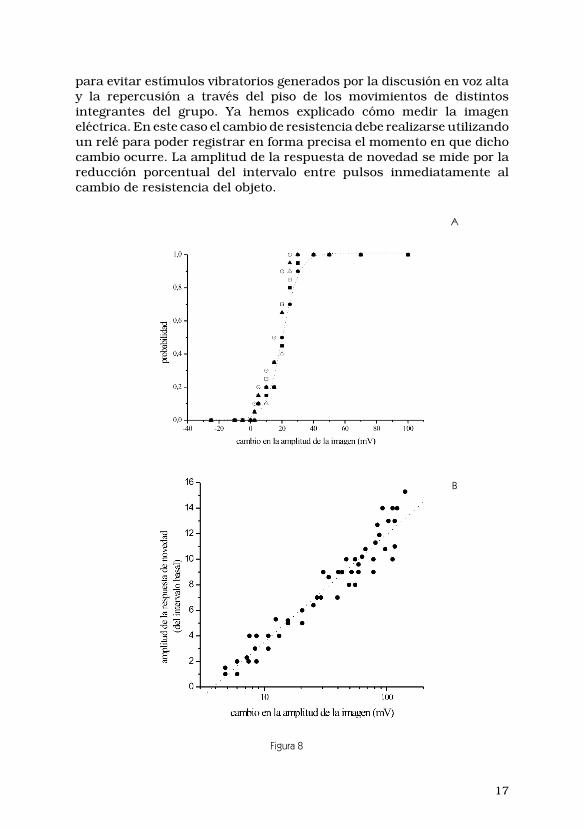
La probabilidad de obtener respuestas de novedad en función delcambio en la amplitud de la descarga del órgano eléctrico es una funciónsigmoide como se observa en la Figura 9A. Dicha función es independien-te de la amplitud de la imagen basal. Estos resultados indican que existeun umbral constante, positivo, e independiente del valor inicial de laimagen eléctrica, por encima del cual un cambio de amplitud de laimagen provoca respuestas de novedad. Es decir, la reducción del perfilde la imagen de un objeto menos conductivo que el agua es igualmentedetectable que el incremento de la imagen de un objeto cuya conductividades mayor a la del agua. Contrariamente, la reducción del perfil de laimagen de un objeto más conductivo que el agua y el incremento de laimagen de un objeto cuya conductividad es mayor a la del agua nocausan respuestas de novedad.
Este estudio nos permitió observar además otro fenómeno: la ampli-tud de la respuesta de novedad es función lineal del logaritmo delincremento de amplitud (∆amplitud, Figura 9B) con constante deproporcionalidad k:
RN = k. log (∆amplitud/∆umbral) (11)
Nuestros experimentos sugieren que el sistema nervioso central deestos peces es capaz de almacenar y comparar sucesivas imágenesdiscretas. A diferencia de otros sistemas en los cuales el flujo sensoriales continuo, en estos peces cada descarga del órgano eléctrico da origena una imagen individual. Dado un tren de descargas homogéneo, bastauna única imagen discordante en más de una diferencia umbral con elresto del tren para provocar una respuesta de novedad.
Surge entonces la pregunta ¿contra qué se compara la imagendiscordante? Una hipótesis es que cada imagen se compara con unaimagen fija genéticamente determinada. Dicha hipótesis no es plausibledado que si así fuese debería responderse a los valores absolutos y no acambios en la amplitud de las imágenes. La hipótesis alternativa queparece más probable es que el sistema compara cada nueva imagen conuna imagen neural resultante de almacenar imágenes recientes. Esdecir, existe una memoria de corto plazo implementada por el sistemasensorial. ¿Cuáles son las reglas que rigen dicha memoria? ¿Se comparaimagen a imagen o existe un efecto remanente?
9. Programa disponible a pedido. Corre en modelos 386 a pentium I siempre quetengan instalado DOS, (Windows 311, 95 o 98).

19
Para responder estas preguntas se diseñó el siguiente experimento:se aplicaron cambios máximos de amplitud del estímulo de modoperiódico. Cada 30 segundos se incrementó la amplitud del estímulocortocircuitando las bases del cilindro que oficiaba de objeto-estímulo yse mantuvo en esta condición durante una determinada fracción delciclo luego de la cual el circuito se volvió a abrir. En estas condiciones semodificó la fracción del ciclo en la cual se aplicaba el cortocircuito. Lapredicción antes del experimento fue que si cada imagen se comparasecon la inmediatamente anterior la respuesta no debería modificarsedado que el cambio fue siempre igual. Por el contrario, una memoriaremanente causaría un cambio gradual de la respuesta dependiente dela duración de dicha remanencia. La experiencia confirmó esta segundapredicción (Figura 10).
La gráfica de la Figura 10 fue obtenida con una intensidad de estímulomáxima dicho cambio en la respuesta. Dicha gráfica podría explicarsepor una disminución de la constante de proporcionalidad k, de unaumento del umbral o de ambos (véase ecuación 7).
Para dilucidar si solo uno o ambos parámetros intervenían en dichoproceso de memoria se realizó otro grupo de experimentos en los cualesse obtuvieron respuestas de novedad para los mismos 4 cambios deamplitud en tres condiciones. Se aplicaron cambios cíclicos de laimpedancia del objeto entre circuito abierto y circuito cerrado por una decuatro resistencias. Se compararon secuencias con distintas fraccionesde tiempo en circuito abierto (1/15 de ciclo, círculo claro; 1/3 de ciclocírculo lleno; y 14/15 de ciclo, cuadrado claro). Los resultados que semuestran en la Figura 11 indican que sólo la pendiente k es modificaday que ésta crece con el número de pulsos de amplitud basal.
Figura 10

20
Puede concluirse entonces que el sistema nervioso genera unamemoria renovable. Dicho proceso parece ocurrir, imagen a imagen, demodo que cada imagen compara la representación almacenada en elpasado. Simultáneamente se actualiza la representación de la escenaelectrosensorial integrando la información proveniente de la nuevaimagen. La exploración es realizada en el caso de los peces eléctricosaplicando una potencialidad idéntica (la activación del órgano eléctrico)a una realidad cambiante y evaluando la respuesta de dicha realidad(imagen eléctrica). Los experimentos muestran que el sistema es predictivo.La creación de una memoria de corto plazo y la comparación de dichamemoria con la imagen presente del mundo permite al pez estableceruna medida de la información contenida en cada imagen y reaccionarcon una conducta exploratoria tanto mayor cuanto más inesperada esla imagen presente.
Reflexiones finalesEl estudio de animales cuyo comportamiento está determinado en un
grado importante por un sentido intuitivamente inaccesible por los sereshumanos, como la representación eléctrica del mundo, nos provee de unexcelente ejemplo para entender la relación realidad y su representaciónpor el cerebro sin preconceptos derivados de las particularidades denuestros propios sistemas sensoriales.
La peculiaridad de que dichos sistemas sean activos, introduce unelemento adicional a favor de su utilización como modelo. La llamada“visión pura”, derivada de la aplicación de conceptos de inteligenciaartificial, intenta establecer formas de decodificación de la imagencomputacional que permitan inferir la realidad que la generó.
Figura 11

21
Este no es el caso de la percepción animal. La realidad no sólo serepresenta para su contemplación sino que se representa para la accióny toda acción condiciona una representación del mundo posterior. Esdecir no existe acción sin percepción y percepción sin acción.10 Se conoceentonces en tanto se explora y se explora en tanto se gana información,es decir cuando ocurren fenómenos inesperados. Se aprende a conocertanto como se aprende a explorar.
El problema de la existencia de una realidad externa a la mente delobservador y su correspondencia con su representación consciente hasido materia de estudio filosófico desde la más remota antigüedad hastanuestros días.
La coherencia de los hallazgos de la ciencia experimental sugiere lapresencia de una realidad externa coherente, independiente de lacapacidad perceptual de cada individuo. Sin embargo, la sensatez delarte11 muestra que la realidad percibida por un actor consciente nonecesariamente corresponde a la realidad objetiva.
Se sustituye entonces el problema de la existencia de una realidadexterna al individuo por el problema de cómo es dicha realidad y cómoel individuo representa dicha realidad.
De cualquier forma que sea el mundo externo al individuo, esnecesario convenir que la representación de la realidad externa por cadaindividuo es dependiente de sus órganos sensoriales, de la estructura yfunciones de su cerebro y de la capacidad exploratoria de dicho indivi-duo, es decir de la dinámica del ciclo acción-percepción.
La realidad externa es común pero su representación compartida conotros individuos es fragmentaria. Conciben el mismo mundo sóloaquellos individuos que han desarrollado sistemas de representacióncompatibles y viven en circunstancias similares. Surge entonces lanoción de mundo perceptual o Merkwelt. Nikolaas Tinbergen hace notarque “cada animal tiene su propio Merkwelt y este mundo difiere de su
ambiente tal como lo percibimos nosotros, es decir de nuestro propio
Merkwelt’’.12
Las propiedades básicas de los sistemas sensoriales están distribui-das ampliamente en la escala zoológica y derivan en general de laexperiencia individual azarosa y la estructura potencialmente presenteen su capital genético.
La capacidad de percibir y representar el mundo en el que operamoslos seres humanos, no sólo deriva de estos dos factores sino que se
10. “Cualquier pieza de conocimiento está conectada con una acción… conocer un objetoes usarlo asimilándolo dentro de un esquema de acción, es decir lo que tienen encomún varias repeticiones o superposiciones de la misma acción”. Biology andknowledge, J. Piaget, 1971.
11. “Hay, Horacio, en el cielo y en la tierra, algo más de lo que puede soñar tu filosofía”.Hamlet, W. Shakespeare.
12. Tinbergen N., The study of instinct, Oxford University Press, New York, 1951.

22
adquieren lentamente en el desarrollo y la maduración del cerebro.Dicho desarrollo y maduración requiere que cada cerebro individualelabore razonamientos, juicios, y estrategias coherentes de exploraciónde la realidad a partir del capital cognitivo acumulado por la humanidad.Los educadores debemos ser conscientes de que, independientemente dela asignatura, lograr dicha utilización, motivar el pensamiento de losestudiantes, es nuestra principal tarea.
Esperamos que el lector comparta con nosotros que plantear elproblema de cómo es el mundo eléctrico de los peces es una excelenteexcusa para ello.
Lecturas sugeridasLas siguientes son revisiones sobre estos temas que pueden aportar
detalles y extensa bibliografía que escapan a las características de estetrabajo. Sugerimos además la lectura de los originales mencionados enlas notas al pie.
Trujillo-Cenóz, O. (1991) “El enigma de los peces eléctricos”. Ciencia Hoy.Macadar, O. (1993) “Motor control of waveform generation” in Gymnotus carapo. J.
Comp. Physiol A. 173: 728-729.Caputi A.A., (1999) “The electric organ discharge of pulse gymnotiforms: the
transformation of a simple impulse into a complex temporal electromotor pattern”,J. Exp. Biol. 202 1229-1241.
Caputi, A. A. (1999) “Aprendiendo neurobiología con los peces eléctricos”. Actas deFisiología. 5: 109-157.
Lorenzo D, Silva A., Caputi A.A. , Borde M., Macadar O. (2001) “Electrogeneration inweakly electric fish” in Sensory biology of jawed fishes. New insights. Oxford & IBHPublishing Co. Pvt. Ltd.
Caputi A.A., Castelló M.E., Aguilera P.A., Trujillo-Cenóz O. (2002) “Electrolocation andelectrocommunication in pulse gymnotids: signal carriers, pre-receptor mechanismsand the electrosensory mosaic”. J Physiol. Paris 96: 493–505.
R. Budelli, A.A. Caputi, L. Gomez, D. Rother and K. Grant, (2002) “The electric image”in G.Petersii J. Physiol. (Paris) 96: 421–29
Caputi A. A., “Contribution of electric fish to the understanding of reafferent sensorysystems”. J. Physiol. Paris 98 (2004) 81–97
Caputi A.A., Budelli R. (2005) “Peripheral electric imaging by freshwater fish”. J. Comp.Physiol. A (in press).
Caputi A.A., Carlson, B., Macadar O., (2005) Electric organs and their control.Electroreception Ed. By T.H.Bullock, C. Hopkins, A.N. Popper and R.R. Fay.Springer-Verlag.