29/11/2016 x-chromosomal markers and f l xfamilias.name/sagf2016/theusefulnessofxinkinship.pdf ·...
TRANSCRIPT

29/11/2016
1
X-CHROMOSOMAL MARKERS AND FAMLINKX
THE USEFULNESS OF X CHROMOSOME
MARKERS IN KINSHIP ANALYSIS
Andrea ColussiLeonor Gusmão
TIPOS DE MARCADORES GENÉTICOS QUANTO A PROPRIEDADES DE TRANSMISSÃO
Marcadores com transmissão biparentallocalizados
Autossomas
Cromossoma X
Marcadores com transmissão uniparentallocalizados
Cromossoma Y DNA mitocondrial
Recombinantes
Não- recombinantes
Universidade do Estado do Rio de Janeiro

29/11/2016
2
EVOLUÇÃO DOS CROMOSSOMAS SEXUAIS
autossomas X Y X Y
SRY
Em muitos aspectos , o cromossoma X é semelhante a um autossoma, o que se entende dada a sua história evolutiva, como
propõe pela primeira vez Ohno, em 1967
Universidade do Estado do Rio de Janeiro
COMPARAÇÃO DOS CROMOSSOMAS X E YO tamanho de cromossoma X é 155M, cerca de 3 vezes
superior ao do Y (51Mb), e a sua região eucromática é cerca de 6 vezes mais longa que a do Y.
Na extremidade dos braços curtos do X e Y existe uma região pseudoautossómica - PAR1- que compreende 2.7 Mb.
A taxa de recombinação em PAR1 é cerca de 20 vezes superior à média registada no genoma!
Homologia entre X e Y em PAR1 é mantida pelo ocorrência de uma recombinação obrigatória na meiose masculina.
Nesta região, os genes estão presentes em 2 cópias em homens e mulheres e não estão sujeitos a inactivação.
Na extremidade do braço longo do X e Y existe uma 2ª região pseudoautossómica, PAR2, com 330 kb, que foi criada por duplicação de material do X para o Y depois da divergência
homem-chimpanzé, há cerca de 6 milhões de anos.
Fora das PARs, homologias entre X e Y localizam-se em regiões não-recombinantes, predominantemente no interior
de XAR, e ainda numa grande região XTR (X-transposedregion). A XTR surgiu por duplicação de material do X para Y
após divergência homem-chimpanzé. A região duplicada estende-se por 3.91 Mb no X, mas a região
correspondente no Y tem apenas 3.38 Mb.
Universidade do Estado do Rio de Janeiro

29/11/2016
3
PADRÃO DE TRANSMISSÃO
Y
X X X X
X X
X
Y
Y
X X
Y
X
X X
• Uma mulher transmite a cada descendente um dos 2 cromossomas Xs com 1/2 de probabilidade (tal como acontece para os autossomas)
• Um homem só possui um cromossoma X que transmite apenas às filhas.
EXCEPÇÃO: PAR1 e PAR2
Universidade do Estado do Rio de Janeiro
APLICAÇÕES FORENSES: IDENTIDADE ENTRE DUAS AMOSTRAS BIOLÓGICAS
Com poucas excepções, marcadores docromossoma X não são tão eficientescomo os autossómicos na investigaçãode identidade entre 2 amostrasbiológicas
O poder de discriminação demarcadores do X varia consoante osexo
Amostra biológica Suspeito
?
Universidade do Estado do Rio de Janeiro

29/11/2016
4
APLICAÇÕES FORENSES: IDENTIDADE ENTRE DUAS AMOSTRAS BIOLÓGICAS
Marcadores do cromossoma X têm o mesmo Poder de Discriminação (PD) que os
autossómicos
Amostra biológica e suspeito
Po
der
de
Dis
crim
inaç
ão
1112
22
12
2211
12
11
22
PD = 1 - (p4 + 4 p2q2 + q4)p2 (2pq+q2)
+
+A
mo
stra
Exc
luíd
os
Locus com 2 alelos p =q = 0.5
PD = 0.75
2pq (p2+q2)
q2 (p2+2pq)
SE:
APLICAÇÕES FORENSES: IDENTIDADE ENTRE DUAS AMOSTRAS BIOLÓGICAS
Marcadores do cromossoma X têm menorPD que os autossómicos
Amostra biológica e suspeito
SE:
Po
der
de
Dis
crim
inaç
ão
1 2
2 1
pq
pq
+
Am
ost
ra
Exc
luíd
os
Locus do X com 2 alelos p =q = 0.5
PD = 0.5
PD = 1 - (p2 + q2)

29/11/2016
5
APLICAÇÕES FORENSES: IDENTIDADE ENTRE DUAS AMOSTRAS BIOLÓGICAS
A probabilidade de ter todos os alelosmasculinos incluídos no perfil da mulher, émaior para marcadores do X queautossómicos.
Desaconselhável usar marcadores do Xpara identificar vestígios masculinos numfundo femininoEx: sémen em vagina
+
Alelos femininos só estarão totalmenteincluídos no perfil masculino se, por acaso,a mulher for homozigótica para todos osloci.
Aconselhável usar marcadores do X paraidentificar vestígios femininos num fundomasculinoEx: restos de pele feminina em unhas de indivíduomasculino; células vaginais em pénis.
SeAmostra biológica e suspeito
SeAmostra biológica e suspeito
+
APLICAÇÕES FORENSES: INVESTIGAÇÃO DE PARENTESCOS BIOLÓGICOS
Marcadores do cromossoma X são informativos para inferir
relações pais-filhos que envolvam pelo menos uma mulher.
Por exemplo:
duos
ou um trio clássico em que se disputa a paternidade de
uma filha
? ? ?
?
Universidade do Estado do Rio de Janeiro
Na maioria dos casos em que se investiga parentescos biológicos, a análise de
marcadores autossómicos é suficiente para responder à pergunta em causa.

29/11/2016
6
PROBABILIDADE DE EXCLUSÃO A PRIORI (MEAN EXCLUSION CHANCE –MEC)
1111
12
22
11
12
11
22
22
11
2212
22
22
11
12
Mãe
s
Fil
has
Pai
s ex
clu
ído
s
p3 q2
p2q p2
q3 p2
p2q q2
pq2 p2
q2p q2
p = q = 0.5
+
+
+
+
+
MEC = 0.25
1111
12
2
1
12
11
22
2
1
2212
22
2
1
12
Mãe
s
Fil
has
Pai
s ex
clu
ído
s
p3 q
p2q p
q3 p
p2q q
pq2 p
q2p q
p = q = 0.5
+
+
+
+
+
MEC = 0.375
Marcador autossómico Marcador cromossoma X
PROBABILIDADE DE EXCLUSÃO A PRIORI (MEAN EXCLUSION CHANCE –MEC)
A eficiência relativa de um marcador do cromossoma X é sobretudo maior comparativamente a um autossómico quando o nível de polimorfismo de um locus é baixo, p.ex, SNPs; o aumento
de eficiência diminui para loci muito polimórficos, p.ex, STRs.
Marcador autossómico
Marcador cromossoma X

29/11/2016
7
A GRANDE VANTAGEM DO CROMOSSOMA X: INVESTIGAÇÕES DE PARENTESCO INCOMPLETAS
Em averiguações de parentesco, nemsempre se dispõe de material biológico detodos os indivíduos-chave; por vezes tem-seacesso a amostras de familiares.
CASOS INCOMPLETOS
?
?
?
PP é o pai biológico?
São irmãs?
São meias-irmãs?
Universidade do Estado do Rio de Janeiro
EXEMPLO DA UTILIDADE DO CROMOSSOMA X NUM POSSÍVEL CASO DE INCESTO
Efectuou-se diagnóstico pré-natal
A grávida é portadora do haplótipode X que o seu pai lhe transmitiu.Logo, se este tivesse sido o pai dofeto (de sexo feminino) que está agerar, não se deveriam encontrarneste alelos adicionais aos da mãe
?
?
PP1
PP2
11-11
8-9
20-22
14-15
11-12
11-15
9-9
20-24
14-15
12-12
PP1 não é o pai do feto
Universidade do Estado do Rio de Janeiro

29/11/2016
8
PATERNIDADE ENVOLVENDO PRESUMÍVEIS PAIS APARENTADOS
Se pai e filho forem presumíveis pais, PP1 e PP2 não compartilham alelos do cromossomaX idênticos por descendência pois recebem o X de mães diferentes.
Neste caso, o poder de exclusão de marcadores autossómicos é substancialmente inferiorao dos X.
H0 – PP is the father of S H1 – PP is the grandfather of S
(For X-markers is the same as PP
and S are two unrelated persons)
INTERPRETAÇÃO ESTATÍSTICA DE EVIDÊNCIAS GENÉTICAS
Female Male
2 copies 1 copy
* Images from
http://members.cox.net/chromosome3
Universidade do Estado do Rio de Janeiro
Tamanho populacional efectivo relativamente a autossomas:
- mtDNA e cromossoma Y: ¼
- cromossoma X: ¾

29/11/2016
9
MARCADORES DO CROMOSSOMA X: ANÁLISE POPULACIONAL
Outra consequência: nível de subestruturação populacional superior no X que em Aut
Devido ao menor efectivo populacional do X relativamente a autossomas, os efeitos de deriva são mais acentuados no X
SUBESTRUTURAÇÃO POPULACIONAL MAIS MARCADA PARA O X, OU SEJA, MAIS DIFERENÇAS INTERPOPULACIONAIS PARA O X QUE PARA
AUTOSSOMAS
FST é uma medida da distância genética entre populações. Varia entre 0 e 1
Para loci autossómicos, a distância entre africanos e não-africanos varia entre valores de FST de 0.1 a 0.15
GENÉTICA POPULACIONAL
Mendelian Theory of HeredityPopulation extension, 1908
Godfrey Harold Hardy(1877 - 1947)
Wilhelm Weinberg
(1862-1937)
Frequência genótipica = produto das frequências alélicas
Exemplo, LR = 1 / 2f4 f7
Universidade do Estado do Rio de Janeiro

29/11/2016
10
EQUILÍBRIO DE HARDY-WEINBERG
AA
AA
AB
AB
BB
BB
1 locusA=B
f(AA) = 0.25f(AB) = 0.50f(BB) = 0.25
FREQUÊNCIAS GENOTÍPICAS
p = f(AA) + ½ f(AB) = 0.5
q = f(BB) + ½ f(AB) = 0.5
FREQUÊNCIAS GÉNICAS
What are the gene and genotype proportions after 1, 2, 3… generations of random mating?
Pressupostos do equilíbrio de HARDY-WEINBERG: a população é
infinitamente grande e panmítica; não ocorre seleção, migração e mutação
EQUILÍBRIO DE HARDY-WEINBERG
LR final = LR1 x LR2 x LR3 x ....
O resultado final é apresentado através de um índice calculado com base nas probabilidades obtidas individualmente para cada marcador.
O valor final obtido é a probabilidade de todos aqueles acontecimentos (conjunto de vários marcadores) ocorrerem simultaneamente.
INDEPENDÊNCIA
Universidade do Estado do Rio de Janeiro

29/11/2016
11
FREQUÊNCIAS HAPLOTÍPICAS = PRODUTO DAS FREQUÊNCIAS ALÉLICAS
Extensão dos princípios do equilíbrio de Hardy-Weinberg na análise simultânea de dois loci
1924, Felix Bernstein
(1878-1956)
Pressupostos: panmíxia; ausência de seleção, migração, mutação e
AUSÊNCIA DE ASSOCIAÇÃO GAMÉTICA
Universidade do Estado do Rio de Janeiro
ASSOCIAÇÃO GAMÉTICA
Frequências haplotípicas ≠ Produto das frequências alélicas
Associação gamética
ou
LD – Linkage disequilibrium ou desequilibrio de ligamento
Causas:
LINKAGE
SUB-ESTRUTURAÇÃO OU
ESTRATIFICAÇÃO POPULACIONAL
Universidade do Estado do Rio de Janeiro

29/11/2016
12
LINKAGE
CSF1PO
D5S818
D21S11
TH01
TPOX
D13S317
D7S820
D16S539 D18S51
D8S1179
D3S1358
FGA
VWA
AMEL
AMEL
SNPs – Single Nucleotide Polymophisms
Marcadores de cromossoma X

29/11/2016
13
Indels – Insertion/Deletion Polymophisms
Marcadores de cromossoma X
Indels SNPs STRs
Marcadores de cromossoma X

29/11/2016
14
MARCADORES DO CROMOSSOMA X: ANÁLISE POPULACIONAL
Como os homens só têm um cromossoma X, a sua genotipagem faculta o acesso imediato a haplótipos, o que não acontece nas mulheres. É impossível inferir haplótipos para
autossomas com base na informação genotípica de um indivíduo
A A C GT T G T A CA A GGT A G T AG
AA
GT
TG
C
T
AC
AA
GT
TG
G
A
AG
AA
GT
TGT
AC
AA
GT
TG
G
A
AG
C
AA
GT
TG
AC
AA
GT
TG
G
A
AG
C
T
AA
GT
TG
A
G
A
G C
AA
GT
TG
A
C
T
T
AT
GT
TG
A
C
T
A T C GT T G T A T
Outro
? ? ? ? ?
Haplótipos ? Haplótipos
X X X X X X X X X Y
MARCADORES DO CROMOSSOMA X: ANÁLISE POPULACIONAL
Como os homens só têm uma cópia de cromossoma X, qualquer cromossoma X actual passou 2/3 dasua história nas mulheres.Consequentemente, mutações ocorrem com menos frequência no cromossoma X que numautossoma porque a taxa de mutação nucleotídica nas mulheres é várias vezes inferior à doshomens (o número de divisões na linha germinativa masculina é muito superior à feminina).
MENOR DIVERSIDADE GENÉTICA NO XMENOR DIVERGÊNCIA INTER-ESPECÍFICA NO X ( entre homem e
chimpanzé, o grau de divergência quanto ao X é apenas cerca de 83% do observado em média para autossomas)
Tamanho populacional efectivo do cromossoma X relativamente a autossomas: ¾

29/11/2016
15
MARCADORES DO CROMOSSOMA X: ANÁLISE POPULACIONAL
Outra consequência: nível de desequilíbrio de ligação superior no X que em Aut
LD é geralmente baixo nostelómeros, elevado perto doscentrómeros e estácorrelacionado com otamanho dos cromossomas(os cromossomas maispequenos tendem a ter taxasde recombinação maiselevadas, Kong et al, 2002,Nat Genetics 31:241-247)
Os cromossomas Xs só recombinam nas mulheres porque os homens apenas têm uma cópia.Portanto, apenas 2/3 dos Xs recombinam em cada geração. Em consequência, é de prever que LDseja superior no X e que neste cromossoma seja maior o tamanho de regiões com a mesma históriagenética
LD
forte
fraco
The International HapMap Consortium (2005) Nature 437, 1299-1320
LD no genoma humano
A taxa de recombinação no X ( cM/Mb) é cerca de 2/3 da média do genoma
LINKAGE – TAXA DE RECOMBINAÇÃO
Regra 1 cM igual a 1 Mb?
Universidade do Estado do Rio de Janeiro
Marker Location Genetic distance ARGUS DECAPLEX
DXS6807 4753382 14.7608
DXS9895 7387107 17.0891
DXS10135 9266321 20.0256 0.1855
DXS8378 9330226 20.2111 70.539 12.1075
DXS9902 15233537 32.3186 58.4315
DXS6810 42803634 75.1188
DXS10076 48194253 85.0446
DXS10077 48201954 85.0611
DXS10078 48207132 85.0722
DXS7132 64572061 90.7501 0.0798 10.5403
DXS10079 66632579 90.8159
DXS10074 66893842 90.8299 58.8268
DXS10075 66914898 90.8311
DXS981 68114084 92.8093
DXS6800 78567066 97.4932
DXS6803 86317826 99.3954
DXS9898 87683075 101.2904 6.829
DXS6801 92397828 106.0777
DXS6809 94824809 108.1194 0.3539
DXS6789 95336070 108.4733 9.7114
DXS6799 97265570 110.7108
DXS7424 100505472 115.2536
DXS101 101299672 116.1513
DXS6797 107367721 117.7436
DXS7133 108928199 118.1847 6.1749
DXS6804 111999363 122.3232
GATA172D05113061249 124.3596 36.1791
DXS7130 118084196 130.2764
GATA165B12120705649 136.1791
HPRTB 133443071 149.6567 0.0908
DXS10101 133482143 149.7475 34.2139
GATA31E08140061921 160.5387 23.6527
DXS9908 142768992 169.8699
DXS8377 149317129 183.6568
DXS10146 149334927 183.7198
DXS10134 149400732 183.9614 0.23
DXS10147 149414073 184.0119
DXS7423 149461561 184.1914
DXS10011 150938682 188.6987

29/11/2016
16
LINKAGE – TAXA DE RECOMBINAÇÃO
Bases de dados de X-STR
É necessário ter em conta a frequência de recombinação entre cada par de
marcadores e respectivas distâncias genéticas
Dentro de cada um dos grupos de ligação, os marcadores tendem a apresentar associação,
sendo necessário estimar frequências haplotípicas na elaboração de bases de dados
populacionais.
Universidade do Estado do Rio de Janeiro
Haldane’s mapping function
r=0,4448
r=0,4137
r=2958
CÁLCULO DE TAXAS DE RECOMBINAÇÃO

29/11/2016
17
SUB-ESTRUTURAÇÃO OU ESTRATIFICAÇÃO POPULACIONAL
Subestruturação “Admixture”
Associação pode dever-se à existência de uma subestruturação, ou estratificação populacional
•Fenómenos de admixture recentes
• Estratificação populacional como consequência de factores socio-culturais (etnia/estrato social;
cultura; religião, etc.)
Universidade do Estado do Rio de Janeiro
ADMIXTURE E LINKAGE DESIQUILIBRIUM
Linkage desiquilibrium resulta de uma maior tendência para que 2 alelos de diferentes loci tendam a
ser herdados juntos a uma maior frequência que o esperado na ausência de associação.
Teoricamente, fenómenos de admixture irão criar LD entre todos os loci para os quais existam diferenças nas
frequências alélicas, nas 2 populações ancestrais.
LD entre loci não ligados tenderão a desaparecer em poucas gerações (HW)
Admixture e linkage desiquilibrium
LD entre loci ligados tenderão a desaparecer mais lentamente, dependendo da fracção de recombinação
entre eles.
Populações que sofreram fenómenos de admixture recente tendem a exibir LD num intervalo de distância genética mais alargado que populações não misturadas.
Admixture
Todos os loci em LD
Loci no mesmo cromossoma em
LD
Loci próximos em LD

29/11/2016
18
SUB-ESTRUTURAÇÃO OU ESTRATIFICAÇÃO POPULACIONAL
• Estratificação populacional como consequência de factores socio-culturais
(etnia/estrato social; cultura; religião, etc.)
•Numa população que sofreu admixture, determinado marcador pode apresentar-se associado a uma doença, devido a uma alta
frequência de ambos alelos
Admixture
Ex: diferentes grupos raciais dos EUA; diferentes castas na India; diferentes grupos religiosos que coexistem num mesmo país (ex.
judeos na Europa); Bascos na Península Ibérica, etc.
MARCADORES DE CROMOSSOMA X
Universidade do Estado do Rio de Janeiro
Recommendation #1
In paternity cases (duos or trios, with a daughter), the use of X-STR analysis is recommended as
supplementary testing when the information from standard autosomal markers are inconclusive (e.g. when
having paternity cases with few genetic inconsistencies).
Recommendation #2
The use of X chromosome markers is recommended in specific kinship cases where they have non-null
exclusion power in contrast to the autosomal markers. Important examples include full or paternal half sibling
duos involving two females, and paternal grandmother/granddaughter duos.
Recommendation #3
It is also recommended to use X chromosomal markers in situations where two alternative hypotheses with
the same likelihood for autosomal but not for X-chromosomal markers are expected. One should also
consider if X can help to distinguish possibly culprits in incest cases.
Recommendation #4
In many situations, the values of LR will be low for the usually available set of X-markers, and before the
analysis it is recommended to simulate the expected distribution of LRs to evaluate the performance of the
available markers in each case.
Recommendation #5
Linkage should be accounted for when calculating LRs given that the X-markers are linked and that linkage
will have an impact on the final LR. Before the use of a given X-chromosomal marker multiplex, it should be
evaluated if the markers are linked or not. A recombination rate lower than 0.5 indicates linkage. The
recombination rate should primarily be estimated from family studies or secondarily via mapping functions
based on genetic distances.

29/11/2016
19
MARCADORES DE CROMOSSOMA X
Universidade do Estado do Rio de Janeiro
Recommendation #6
A “cluster-approach” (assuming no recombination within a linkage group) is not valid for X-chromosomal
markers, unless…. and in LR calculations it is recommended to account for the possibility of recombination
events even within a linkage group.
Recommendation #7
For closely located markers on the X chromosome, not demonstrated to be in linkage equilibrium, it is
important to report haplotypes rather than single locus allele frequencies. Furthermore, the likelihood
calculations are more sensitive to using an appropriate haplotype frequency database compared to when
using individual allele frequencies.
Recommendation #8
Haplotype frequencies should be used for likelihood calculations when LD exists. It is important to note that
specific haplotypes may not always be unambiguously determined for the individuals in a pedigree and the
computations will therefore include an iteration over all possible haplotype setups.
Recommendation #9
When calculating LRs based on X-chromosomal markers in kinship analysis, the use of an appropriate
software is highly recommended to avoid hand-calculation errors. The software should rely on likelihood
calculations and can accommodate mutation, linkage and linkage disequilibrium between markers.
Recommendation #10
The final LR can be obtained by the product of individual autosomal or X chromosomal markers whenever
the same (and clearly defined) hypotheses/pedigrees are used for both autosomal and X-chromosomal data,
and if substructure or LD between autosomal and X chromosomal alleles does not play a role.
X-CHROMOSOMAL MARKERS AND FAMLINKX
LIKELIHOOD RATIOS WITH X EXAMPLES
Leonor Gusmão Andrea Colussi

29/11/2016
20
LIKELIHOOD RATIOS (LR)
fi LR
0,05 5
0,4 0,625
2
1
|
|
HGP
HGPLR
O
O
pares the genotipos observados:OG
Indivíduos associados por uma relação genética R1:1H
:2H Indivíduos associados por uma relação genética R2
Exemplo
F e C são Pai - Filho
F e C são 2 ind.não relacionados
:1H
:2H
kjiO fffHGP 5.02| 1
kijiO ffffHGP 22| 2 ifLR
4
1
F
i - j
i - k
C
Universidade do Estado do Rio de Janeiro
LIKELIHOOD RATIOS (LR)
Exemplo:
:1H
0| 1 HGP O
lkjiO ffffHGP 22| 2
0LR
F
i - j
k - l
C
2
1
|
|
HGP
HGPLR
O
O
pares the genotipos observados:OG
Indivíduos associados por uma relação genética R1:1H
:2H Indivíduos associados por uma relação genética R2 F e C são 2 ind.não relacionados:2H
F e C são Pai - Filho
Universidade do Estado do Rio de Janeiro

29/11/2016
21
LIKELIHOOD RATIOS (LR)
fi LR
0,05 5
0,4 0,625
2
1
|
|
HGP
HGPLR
O
O
pares the genotipos observados:OG
Indivíduos associados por uma relação genética R1:1H
:2H Indivíduos associados por uma relação genética R2
Exemplo
F e C são Pai - Filho
F e C são 2 ind.não relacionados
:1H
:2H
kjiO fffHGP 5.02| 1
kijiO ffffHGP 22| 2 ifLR
4
1
F
i - j
i - k
C
n
l
lfinal LRLR1
n: número de loci autossomicos analisados,
não ligados
likelihood ratio (razão de verosímilhança)
associado ao locus l
:lLR
R1 e R2 são hipóteses
mutuamente exclusivas
Na prática forense:
Universidade do Estado do Rio de Janeiro
ALELOS IBD
Dois alelos são ditos idêntico por
descendência (identical-by-descent ou
IBD) quando descendem de um
mesmo alelo ancestral
Identity-by-descent (IBD) ≠ Identity-by-state (IBS)
Exemplo:
IBS;
M
C
1 13 4
F
1 1
1 1and IBD
IB
S
1 and IBDF
1C
IBD
IBS IBD
IBS , excepto se ocorrer mutação
Nota: IBD é sempre medido a partir de
um mesmo grupo de ancestrais
“fundadores” que se assume não
estarem relacionados entre si.
Dois indivíduos dizem-se relacionados se
pelo menos um dos alelos de um poder ser
IBD em relação aos alelos do outro.
Universidade do Estado do Rio de Janeiro

29/11/2016
22
PADRÕES DE IBD PARA 2 INDIVÍDUOS
Δ4 I1:
I2:
Δ5 I1:
I2:
Δ6 I1:
I2:
Δ2 I1:
I2:
Δ3 I1:
I2:
Δ1 I1:
I2:
Δ7 I1:
I2:
Δ9 I1:
I2:
Δ8 I1:
I2:
Jacquard’s coefficients (1970)
Universidade do Estado do Rio de Janeiro
PADRÕES DE IBD PARA 2 INDIVÍDUOS
... ...
1/2 1/2
1/2 1/2
1/2 1/2
Indivíduo consanguíneo C
(Exemplo):
B
C
A
A e B Primos em primeiro
grauAlelos
:
42
16
P(C é inbred)
=
= %25.62
14
... ...
Universidade do Estado do Rio de Janeiro

29/11/2016
23
PADRÕES DE IBD PARA 2 INDIVÍDUOS, NA AUSÊNCIA DE CONSAGUINIDADE
k2 I1:
I2:
k0 I1:
I2:
k1 I1:
I2:
Um indivíduo diz-se consaguíneo se poder ser
portador de dois alelos IBD num locus.
Universidade do Estado do Rio de Janeiro
PADRÕES DE IBD PARA 2 INDIVÍDUOS, NA AUSÊNCIA DE CONSAGUINIDADE
k2 I1:
I2:
k0 I1:
I2:
k1 I1:
I2:
Não relacionados
k0 = 1
k1 = 0
k2 = 0
BA
Os exemplos mais
simples:
A
B
Pai- Filho
k0 = 0
k1 = 1
k2 = 0
Gémeos monozigóticos
BA
k0 = 0
k1 = 0
k2 = 1
Universidade do Estado do Rio de Janeiro

29/11/2016
24
PADRÕES DE IBD PARA 2 INDIVÍDUOS, NA AUSÊNCIA DE CONSAGUINIDADE
Full - siblings
k0 = 1/4
k1 = 1/2
k2 = 1/4
BA
Half - siblings
k0 = 1/2
k1 = 1/2
k2 = 0
BA
Avuncular
k0 = 1/2
k1 = 1/2
k2 = 0
B
A
k2 I1:
I2:
k0 I1:
I2:
k1 I1:
I2:
Exemplos:
Universidade do Estado do Rio de Janeiro
Universidade do Estado do Rio de Janeiro
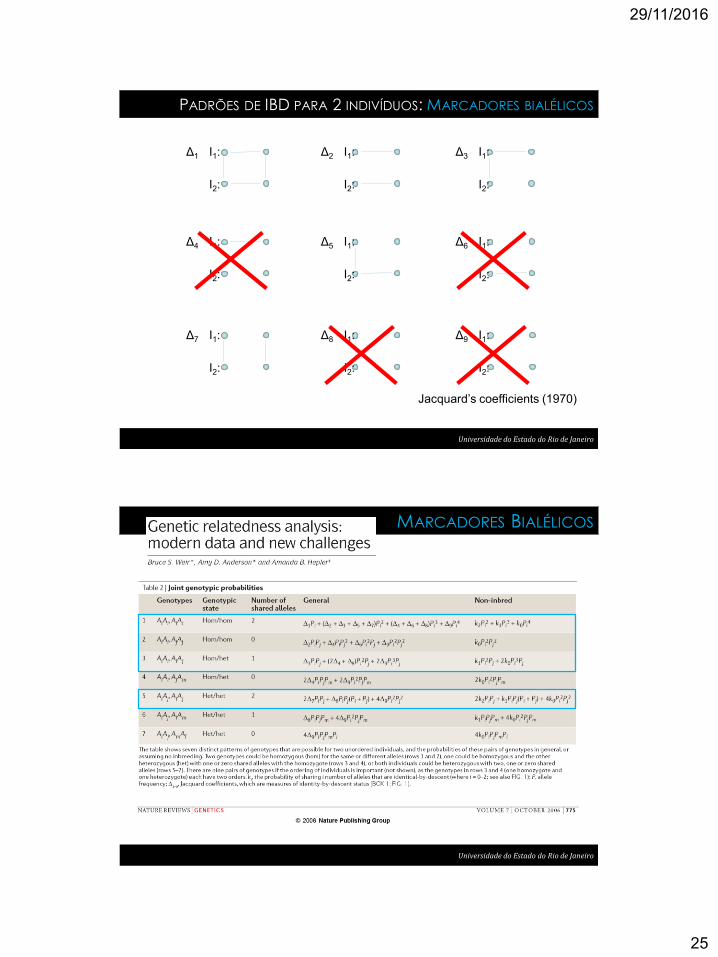
29/11/2016
25
PADRÕES DE IBD PARA 2 INDIVÍDUOS: MARCADORES BIALÉLICOS
Δ4 I1:
I2:
Δ5 I1:
I2:
Δ6 I1:
I2:
Δ2 I1:
I2:
Δ3 I1:
I2:
Δ1 I1:
I2:
Δ7 I1:
I2:
Δ9 I1:
I2:
Δ8 I1:
I2:
Jacquard’s coefficients (1970)
Universidade do Estado do Rio de Janeiro
MARCADORES BIALÉLICOS
Universidade do Estado do Rio de Janeiro

29/11/2016
26
PADRÕES DE IBD PARA MARCADORES DO CROMOSSOMA X
Para 2 mulheres:
F1:
F2:
F1:
F2:
F1:
F2:
F1:
F2:
F1:
F2:
ΛFF1 F1:
F2:
F1:
F2:
F1:
F2:
F1:
F2:
ΛFF2 ΛFF
3
ΛFF4 ΛFF
5 ΛFF6
ΛFF7 ΛFF
8 ΛFF9
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
PADRÕES DE IBD PARA MARCADORES DO CROMOSSOMA X
Para 2 mulheres na ausência de consaguinidade:
F1:
F2:
F1:
F2:
F1:
F2:
xFF2 xFF
1 xFF0
Examples:
paternal half-sisters
x2FF = 0
x1FF = 1
x0FF = 0
x2FF = 0
x1FF = 1/2
x0FF = 1/2
paternal aunt - niece unrelated
x2FF = 0
x1FF = 0
x0FF = 1
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32

29/11/2016
27
INTERPRETAÇÃO ESTATÍSTICA DE EVIDÊNCIAS GENÉTICAS
GenotypesJoint Genotypic Probabilities for X-markers and two females
General Non-inbred females
AiAi, AiAi
Λ1FFfi + (Λ2
FF+ Λ3FF+ Λ5
FF+ Λ7FF)fi
2+(Λ4FF+ Λ6
FF+
Λ8FF)fi
3+ Λ9FFfi
4 x2FFfi
2 + x1FFfi
3+ x0FFfi
4
AiAi, AjAjΛ2
FFfifj + Λ4FFfifj
2+ Λ6FFfi
2fj + Λ9FFfi
2fj2 x0
FFfi2fj
2
AiAi, AiAjΛ3
FFfifj +(2 Λ4FF + Λ8
FF)fi2fj+ 2Λ9
FFfi3fj x1
FFfi2fj+ 2x0
FFfi3fj
AiAj, AiAiΛ5
FFfifj +(2 Λ6FF + Λ8
FF)fi2fj+ 2Λ9
FFfi3fj x1
FFfi2fj+ 2x0
FFfi3fj
AiAi, AjAl 2Λ4FFfifjfl + 2Λ9
FFfi2fjfl 2x0
FFfi2fjfl
AjAl, AiAi 2Λ6FFfifjfl + 2Λ9
FFfi2fjfl 2x0
FFfi2fjfl
AiAj, AiAj2Λ7
FFfifj + Λ8FFfifj(fi + fj) + 4Λ9
FFfi2fj
2 2x2FFfifj + x1
FFfifj(fi + fj) + 4x0FFfi
2fj2
AiAj, AiAk Λ8FFfifjfk + 4Λ9
FFfi2fjfk x1
FFfifjfk + 4x0FFfi
2fjfk
AiAj, AkAl 4Λ9FFfifjfkfl 4x0
FFfifjfkfl
Marcadores de cromossoma X: Probabilidade genotípica conjugada para 2 mulheres
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
PADRÕES DE IBD PARA MARCADORES DO CROMOSSOMA X
Para um par mulher/homem:
ΛFM1 F:
M:
F:
M:
ΛFM2
ΛFM3 F:
M:
ΛFM4 F:
M:
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
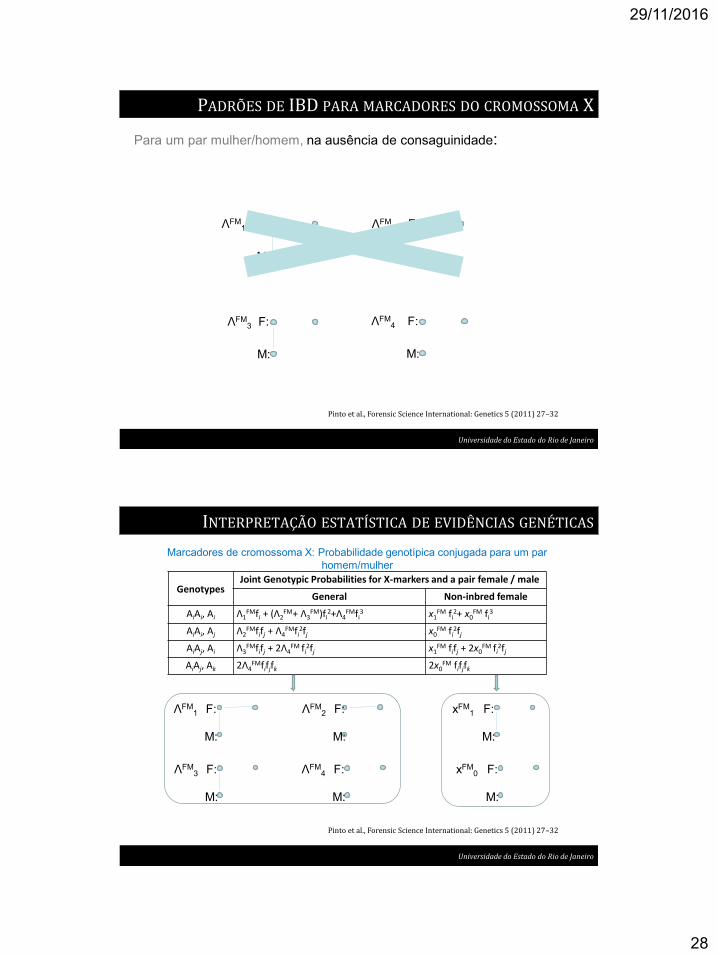
29/11/2016
28
PADRÕES DE IBD PARA MARCADORES DO CROMOSSOMA X
Para um par mulher/homem, na ausência de consaguinidade:
ΛFM1 F:
M:
F:
M:
ΛFM2
ΛFM3 F:
M:
ΛFM4 F:
M:
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
INTERPRETAÇÃO ESTATÍSTICA DE EVIDÊNCIAS GENÉTICAS
Marcadores de cromossoma X: Probabilidade genotípica conjugada para um par
homem/mulher
GenotypesJoint Genotypic Probabilities for X-markers and a pair female / male
General Non-inbred female
AiAi, Ai Λ1FMfi + (Λ2
FM+ Λ3FM)fi
2+Λ4FMfi
3 x1FM fi
2+ x0FM fi
3
AiAi, Aj Λ2FMfifj + Λ4
FMfi2fj x0
FM fi2fj
AiAj, Ai Λ3FMfifj + 2Λ4
FM fi2fj x1
FM fifj + 2x0FM fi
2fj
AiAj, Ak 2Λ4FMfifjfk 2x0
FM fifjfk
ΛFM1 F:
M:
F:
M:
ΛFM2
ΛFM3 F:
M:
ΛFM4 F:
M:
xFM1 F:
M:
xFM0 F:
M:
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro

29/11/2016
29
INTERPRETAÇÃO ESTATÍSTICA DE EVIDÊNCIAS GENÉTICAS
Marcadores de cromossoma X: Probabilidade genotípica conjugada para um par
homem/mulher
EXEMPLOS
paternal uncle - niece
x0FM = 1/2
x1FM = 1/2
unrelated
x0FM = 1
x1FM = 0
Genotypes
Joint Genotypic Probabilities for X-markers and a
female (non-inbred) / male pair
General paternal uncle - niece unrelated
AiAi, Ai x1FM fi
2+ x0FM fi
3 1/2fi2+ 1/2fi
3 fi3
AiAi, Aj x0FM fi
2fj 1/2fi2fj fi
2fj
AiAj, Ai x1FM fifj + 2x0
FM fi2fj 1/2fifj + fi
2fj 2 fi2fj
AiAj, Ak 2x0FM fifjfk fifjfk 2 fifjfk
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
PADRÕES DE IBD PARA MARCADORES DO CROMOSSOMA X
Para 2 homens:
ΛMM1 M1:
M2:
ΛMM2 M1:
M2:
GenotypesJoint Genotypic Probabilities for X-chromosome
markers and two males
Ai, Ai Λ1MMfi + Λ2
MMfi2
Ai, Aj Λ2MMfifj
Marcadores de cromossoma X: Probabilidade genotípica conjugada para um par de homens:
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro

29/11/2016
30
INTERPRETAÇÃO ESTATÍSTICA DE EVIDÊNCIAS GENÉTICAS
Marcadores de cromossoma X: Probabilidade genotípica conjugada para um par de homens:
Exemplo
s:
maternal uncle - nephew
x0MM = 3/4
x1MM = 1/4
unrelated
x0MM = 1
x1MM = 0
Genotypes
Joint Genotypic Probabilities for X-markers and two
males
Generalmaternal uncle -
nephewunrelated
Ai, Ai Λ1MMfi + Λ2
MMfi2 1/4fi + 3/4fi
2 fi2
Ai, Aj Λ2MMfifj 3/4fifj fifj
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Universidade do Estado do Rio de Janeiro
CASOS EM QUE MARCADORES DE CROMOSSOMA X SÃO NECESSÁRIOS
Pinto et al., Forensic Science International: Genetics 5 (2011) 27–32
Caso I (ausência de consaguinidade)
Universidade do Estado do Rio de Janeiro

29/11/2016
31
LRS PARA MARCADORES DE CROMOSSOMA X
Universidade do Estado do Rio de Janeiro
Mother
Marker 1: a/bMarker 2: c/d
SonMarker 1: aMarker 2: d
Caso mãe e filho (do sexo masculino). Os dados consistem em 2 marcadores em linkage (separados
por uma fração de recombinação r) localizados no cromossoma X.
𝐿𝑅 =Pr 𝐷𝑁𝐴 𝑚𝑜𝑡ℎ𝑒𝑟 𝑎𝑛𝑑 𝑐ℎ𝑖𝑙𝑑
Pr 𝐷𝑁𝐴 𝑢𝑛𝑟𝑒𝑙𝑎𝑡𝑒𝑑 =
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 0.5 ∙ 𝑟 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 0.5 ∙ 1 − 𝑟
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 𝑝𝑎𝑑
=𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 𝑟 + 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 1 − 𝑟
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 𝑝𝑎𝑑
Numa situação de LE (e.i. 𝑝𝑥𝑦 = 𝑝𝑥 ∙ 𝑝𝑦 ) o LR será:
𝐿𝑅 =2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 0.5 ∙ 𝑟 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 0.5 ∙ (1 − 𝑟)
(2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ) ∙ 𝑝𝑎𝑑
=𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑 ∙ 𝑟 + 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑∙(1 − 𝑟)
(2 ∙ 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑 + 2 ∙ 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑) ∙ 𝑝𝑎 ∙ 𝑝𝑑=
1
4 ∙ 𝑝𝑎 ∙ 𝑝𝑑
Neste caso, a recombinação não influencia o LR final. Não entanto, na presença de LD, devemos ter
em conta a fração de recombinação no cálculo do LR.
Caso mãe e filho (do sexo masculino). Os dados consistem em 2 marcadores em linkage (separados
por uma fração de recombinação r) localizados no cromossoma X.
𝐿𝑅 =Pr 𝐷𝑁𝐴 𝑚𝑜𝑡ℎ𝑒𝑟 𝑎𝑛𝑑 𝑐ℎ𝑖𝑙𝑑
Pr 𝐷𝑁𝐴 𝑢𝑛𝑟𝑒𝑙𝑎𝑡𝑒𝑑 =
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 0.5 ∙ 𝑟 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 0.5 ∙ 1 − 𝑟
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 𝑝𝑎𝑑
=𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 𝑟 + 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 1 − 𝑟
2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 𝑝𝑎𝑑
Numa situação de LE (e.i. 𝑝𝑥𝑦 = 𝑝𝑥 ∙ 𝑝𝑦 ) o LR será:
𝐿𝑅 =2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 ∙ 0.5 ∙ 𝑟 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ∙ 0.5 ∙ (1 − 𝑟)
(2 ∙ 𝑝𝑎𝑐 ∙ 𝑝𝑏𝑑 + 2 ∙ 𝑝𝑎𝑑 ∙ 𝑝𝑏𝑐 ) ∙ 𝑝𝑎𝑑
=𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑 ∙ 𝑟 + 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑∙(1 − 𝑟)
(2 ∙ 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑 + 2 ∙ 𝑝𝑎 ∙ 𝑝𝑐 ∙ 𝑝𝑏 ∙ 𝑝𝑑) ∙ 𝑝𝑎 ∙ 𝑝𝑑=
1
4 ∙ 𝑝𝑎 ∙ 𝑝𝑑
Neste caso, a recombinação não influencia o LR final. Não entanto, na presença de LD, devemos ter
em conta a fração de recombinação no cálculo do LR.