· aquellos alumnos que durante 10 años han transitado por el curso de bioeconomia pesquera ......
TRANSCRIPT




iii
PREPARACION DE ESTE DOCUMENTO En su origen, la evaluación y representación matemática de las tendencias dinámicas en poblaciones de recursos pesqueros fue considerada únicamente en términos de la interacción entre la intensidad de pesca y los factores biológicos que determinan la productividad de los ecosistemas marinos. Si bien este enfoque es aún válido, la dinámica de poblaciones explotadas es analizada cada vez con mayor frecuencia en base a un análisis conjunto de aquellas fuerzas económicas que regulan, adicionalmente a los factores biológicos y ecológicos, la dinámica de una pesquería, tales como los ingresos, las fuentes de empleo y la producción de alimento. Esta actividad combinada es definida como ‘Bioeconomia’, sobre la cual la FAO ha elaborado una serie de publicaciones (FAO, Serie Informática - Pesca) relacionadas con el desarrollo de programas especializados de computación en el campo pero no había producido un texto que incluyera la teoría bioeconómica conjuntamente con ejemplos prácticos. El presente texto pretende llenar este vacío, enfocándose principalmente en problemas concretos sobre el tópico a los cuales se enfrentan los países en vías de desarrollo.
Agradecimientos Deseamos expresar nuestro agradecimiento al grupo de trabajo de Bioeconomía Pesquera del CINVESTAV, en especial a Miguel Angel Cabrera por su valioso apoyo en la edición del manuscrito. Agradecemos a Rögnvaldur Hannesson por su detallada revisión del manuscrito. Nuestro reconocimiento a Claudia Leonce y Anita de Alava por su escrupulosa revisión de las referencias citadas. Agradecemos a Eduardo P. Pérez el apoyo brindado en la elaboración del indice de tópicos. Enzo Acuña, Jorge González, Minerva Arce y Eduardo Balart proporcionaron comentarios y sugerencias valiosas al texto. Deseamos expresar nuestro aprecio sincero a todos aquellos alumnos que durante 10 años han transitado por el curso de Bioeconomia Pesquera que se ha ofrecido en la Unidad Mérida del CINVESTAV y en diversas instituciones de América Latina. Se agradece al Sr. Héctor Mares la realización del dibujo de la portada de este libro.
Se expresa un sincero reconocimiento al programa de Ecologia, Pesquerías y Oceanografía del Golfo de México (EPOMEX) de la Universidad Autónoma de Campeche, institución en la que el primer autor realizó una estancia académica que tuvo como uno de sus propósitos finalizar la primera versión de este manuscrito.
Juan Carlos Seijo desea expresar su agradecimiento a Cecilia, Juan Carlos Jr. y Adrianita por ser fuente permanente de motivación y cariño. Omar Defeo dedica este trabajo a Anita, Diego, a sus padres y hermanos, y Silvia Salas a su hija Nayelli y a sus padres.
Distribución:
FAO Departamento de Pesca FAO Oficinas regionales y subregionales Instituciones de pesca

iv
Seijo, J.C.; Defeo, O.; Salas, S. Bioeconomía pesquera. Teoría, modelación y manejo FAO Documento Técnico de Pesca. No. 368. Rome, FAO. 1997. 176p.
RESUMEN La sobreexplotación de los recursos pesqueros, la sobrecapitalización de las pesquerías, la eliminación de las rentas que generan a la sociedad y la creciente inquietud por la conservación de la biodiversidad marina, han motivado el desarrollo de lineas de investigación pesquera con un enfoque multidisciplinario, dinámico y precautorio. En este contexto, el análisis conjunto de factores biológicos, ecológicos y económicos surge como un tópico necesario a la hora de evaluar aquellas fuerzas que regulan la dinámica de una pesquería. Este documento desarrolla aspectos de la teoría bioeconómica dinámica a efectos de lograr un manejo sostenible de la actividad pesquera. Se define al manejo de un recurso pesquero como un proceso complejo que requiere la integración de su biología y ecologia con los factores socio-económicos e institucionales que afectan al comportamiento de los usuarios (pescadores) y a los responsables de su administración. El libro es presentado en ocho Capitulos. El Capítulo 1 describe los supuestos básicos en la asignación óptima de los recursos naturales renovables, así como las características inherentes de los recursos pesqueros y las interdependencias humanas que determinan, en un régimen de acceso abierto, el fracaso en dicha asignación y en consecuencia su sobreexplotación. Se describe cómo el aprovechamiento de este tipo de recursos bajo acceso abierto enfrenta altos costos de exclusión, altos costos de transacción (costos de información y vigilancia), generación de externalidades negativas y positivas y presencia de usuarios no contribuyentes. En el Capitulo 2 se discuten los punmtos de referencia bioeconómicos obtenidos del análisis de pesquerias bajo supuestos de equilibrio. Para reflejar la naturaleza dinámica de las pesquerías se reconoce la necesidad de que las estimaciones de biomasa, captura y renta en el tiempo sean estimadas con las trayectorias dinámicas de la intensidad de pesca. Para realizar tales estimaciones se presentan modelos bioeconómicos dinámicos tanto globales como estructurales. En el Capitulo 3 se discuten y presentan las extensiones de estos modelos bioeconómicos mediante la incorporación de interdependencias ecológicas (i.e. competencia y depredación) y tecnológicas (i.e. competencia entre flotas con diferente poder de pesca, tecnologia y costos unitarios del esfuerzo, pesquerías secuenciales con flotas artesanales e industriales que afectan diferentes componentes de la estructura de la población y pesquerías interdependentes con diferente especie objetivo). El precio del tiempo es incorporado en análisis a partir del Capítulo 4, en el cual se presenta el análisis intertemporal de pesquerías y se estima el nivel de esfuerzo óptimo resultante de la utilización de diferentes tasas de descuento en la estimación del máximo valor presente neto. En el Capitulo 5 se introduce el enfoque de ciencia de sistemas para la construcción de modelos matemáticos aplicados al análisis bioeconómico que sustente los planes de ordenación de pesquerias. En el Capitulo 6 se discuten las alternativas de ordenación de pesquerías y las correspondientes ventajas y limitaciones en su aplicación al manejo pesquero de países en desarrollo. En el Capítulo 7 se introducen modelos bioeconómicos que relajan el supuesto de distribución homogénea de recursos pesqueros a través de la incorporación de la dimensión espacial en el análisis dinámico de pesquerías. Con un enfoque precautorio, en el Capítulo 8, se analizan el riesgo e incertidumbre presentes en las pesquerías marinas. Se introducen elementos básicos de teoría de decisiones y se presentan criterios de decisión que reflejan diferentes actitudes de aversión al riesgo en la ordenación de pesquerías. Se discuten métodos de análisis a través de tablas de decisión con y sin probabilidades matemáticas sobre la ocurrencia de posibles estados de la naturaleza que pudieran afectar el desempeño de las pesquerías.

v
Contenido Lista de Figuras .......................................................................................................... ix Lista de Tablas ........................................................................................................... xi Glosario de Símbolos..................................................................................................... xii 1. Recursos Pesqueros: Características Inherentes ............................................ 1
Optima asignación de recursos naturales: supuestos básicos .......................... 1
El fracaso en la óptima asignación de recursos naturales: supuestos básicos ... 1 Regímenes de propiedad ............................................................................ 2 Externalidades ............................................................................................ 2 Altos costos de exclusión ........................................................................... 5 Trampa social en pesquerías ...................................................................... 5 Usuarios no contribuyentes ........................................................................ 6 Altos costos de transacción ......................................................................... 6
2. Modelo Bioeconómico Básico ............................................................................. 8
El modelo logístico ............................................................................................. 8
Modelo de rendimiento excedente y extensiones ............................................ 10 Supuestos básicos del modelo ................................................................. 12 Limitaciones .............................................................................................. 13
Estimación del rendimiento excedente en base a datos de captura y esfuerzo .. .14
Extensiones del modelo ................................................................................... 15
El modelo bioeconómico Gordon-Schaefer ..................................................... 15
Rendimiento promedio y marginal ............................................................ 18
Esfuerzo en máximo rendimiento sostenible (MRS), máximo rendimiento económico (MRE) y en equilibrio bioeconómico (EBE) ............................ 19 Supuestos del modelo .............................................................................. 21
Dinámica del esfuerzo pesquero ..................................................................... 22
Modelos de producción excedente usando mortalidad .................................. 26
Teoría y aproximaciones al ajuste de los modelos .................................. 26 Modelo logístico .......................................................................... 27 Modelo exponencial .................................................................... 29 Ventajas y limitaciones de la aplicación de los modelos captura-mortalidad ...................................................................... 31 Aproximación bioeconómica ..................................................................... 33
Análisis bioeconómico con modelos de estructura por edades de una población ........................................................................................ 34
Modelo de Beverton y Holt (1957) ............................................. 34 Supuestos y limitaciones del modelo ......................................... 35
Modelo dinámico de estructura por edades ..................................................... 37

vi 3. Interdependencias Ecológicas y Tecnológicas en Pesquerías ..................... 41
Conceptos Generales ...................................................................................... 42
Análisis de interdependencias ......................................................................... 43 Esfuerzo pesquero heterogéneo .............................................................. 43 Pesquerías tecnológicamente interdependientes: una flota afecta stocks
ecológicamente interdependientes .................................................... 49 Interdependencias tecnológicas: una flota afecta dos o más stocks con cierta segregación en el espacio ....................................................... 50 Pesquerías tecnológicamente interdependientes: dos flotas ................... 51 Interdependencias tecnológicas: pesquerías secuenciales ...................... 53 Stocks ecológicamente interdependientes: depredación ......................... 56 Stocks ecológicamente interdependientes: competencia ......................... 57 Stocks ecológicamente interdependientes afectados por dos flotas sin
interdependencia tecnológica ............................................................ 60 Interdependencias tecno-ecológicas: stocks ecológicamente interdependientes afectados por dos flotas con interdependencia
tecnológica ....................................................................................... 62
Análisis ecológico-pesquero ............................................................................ 65 Perspectivas .................................................................................................... 67
4. Análisis Intertemporal de Pesquerías ............................................................... 69 Preferencias intertemporales ........................................................................... 69
Preferencias neutras, positivas y negativas ............................................. 70
Inversiones públicas en el manejo de recursos pesqueros ............................. 72 Evaluación de proyectos y el criterio de Pareto-eficiencia ...................... 72 Valor presente y tasa de descuento ......................................................... 73 Costo de oportunidad de la inversión pública ........................................... 74 Depreciación ............................................................................................. 74 Precios “sombra” ...................................................................................... 75
Criterios de decisión para la inversión pública pesquera ................................ 75
Valor presente neto (VPN) ........................................................................ 75 Cociente beneficio-costo (B/C) .................................................................. 76 Tasa interna de retorno (TIR) ................................................................... 77 ABC en la práctica: la elección de un criterio adecuado .......................... 78 Una aplicación práctica en pesquerías: análisis discreto ......................... 80
Evaluación de proyectos pesqueros: preferencias intertemporales e impactos
distribucionales ......................................................................................... 82 La importancia de los impactos distribucionales ...................................... 82 La tasa social de descuento y preferencia social del tiempo .................... 83 Riesgo e incertidumbre en proyectos pesqueros ..................................... 84
Manejo intertemporal de recursos pesqueros ................................................. 87
El modelo bioeconómico dinámico y el precio del tiempo ........................ 87

vii
El efecto de la tasa de descuento ( δ ) en pesquerías: una visión alternativa .......................................................................................... 89
5. Análisis Bioeconómico de Pesquerías: Enfoque de Sistemas ....................... 91
Planes de manejo para recursos pesqueros ................................................... 91
Enfoque de ciencia de sistemas ...................................................................... 92 Identificación de necesidades de información bioeconómica ................... 93 Caracterización de la pesquería ............................................................... 94 Modelación matemática de la pesquería .................................................. 96 Recolección de datos de fuentes primarias y secundarias ..................... 102 Análisis de estabilidad, de error de integración numérica y de sensibilidad .................................................................................. 105 Validación del modelo ............................................................................. 105 Análisis de impacto bioeconómico de diversas estrategias de manejo .. 106 Ejemplo ilustrativo ................................................................................... 106
6. Ordenación de Pesquerías ............................................................................... 109
Criterios para la intervención del Estado ....................................................... 109 Criterio de conservación ......................................................................... 110 Criterio económico .................................................................................. 110 Criterio de equidad en el uso de recursos pesqueros ............................ 111 Criterio de equidad intergeneracional ..................................................... 114 Otros criterios de manejo de pesquerías ................................................ 114
Estrategias de manejo de pesquerías marinas ............................................. 115
Asignación de derechos de propiedad ................................................... 115 Regulación de la composición de la captura .......................................... 116 Regulación de la cantidad capturada ..................................................... 117 Cuotas individuales transferibles ............................................................ 119 Programas de extensionismo y educación ambiental ............................ 120
Manejo de pesquerías con criterios múltiples ................................................ 120
Función objetivo con criterios múltiples .................................................. 121 El método “Complex” para optimizar funciones no-lineales con criterios múltiples
122 7. Análisis Espacial de Pesquerías ...................................................................... 124
Asignación espacial de la intensidad de pesca ............................................. 125 Dinámica espacial en el corto plazo: Modelo ALLOC .................................... 126
Asignación espacial de la intensidad de pesca ...................................... 126 Distancia de los diferentes puertos a los sitios de pesca alternativos .... 127 Quasi-renta de los costos variables por tipo de flota proveniente de diferentes puertos de origen ............................................................ 128 Renta económica obtenida de los sitios de pesca .................................. 129 Variaciones espaciales de CPUE ........................................................... 131 Dinámica de la flota ................................................................................ 131 Supuestos del modelo ALLOC .............................................................. 132
Dinámica bioeconómica geográfica de corto y largo plazo: Modelo CHART .... 133

viii
Especificación espacial de la biomasa máxima y del reclutamiento ...... 133 Distribución espacial del reclutamiento por parches .............................. 134 Distribución espacial de la biomasa del recurso ..................................... 134 Captura por unidad de esfuerzo espacialmente desagregada ............... 135 Dinámica de la flota en el corto plazo: asignación espacial de la intensidad de pesca ......................................................................... 136 Dinámica de la flota en el largo plazo: entrada y salida de embarcaciones a la pesquería .................................................................................. 137 Supuestos del modelo CHART ............................................................... 137
Modelo bioeconómico espacial para especies sedentarias: la almeja amarilla de Uruguay, un estudio de caso ............................................................. 138
El recurso ................................................................................................ 139 La pesquería ........................................................................................... 140
8. Riesgo e Incertidumbre en Pesquerías: Un Enfoque Bioeconómico Precautorio ........................................................................................................ 142
Enfoque precautorio en el manejo de pesquerías ......................................... 142
Conceptos asociados al manejo precautorio de pesquerías ......................... 143 Principales fuentes de incertidumbre en pesquerías marinas ................ 143 Decisiones de manejo con probabilidades matemáticas ........................ 146 Reglas de decisión sin probabilidades matemáticas .............................. 146 Ejemplo ilustrativo: manejo precautorio con y sin probabilidades matemáticas ................................................................................... 147 Enfoque precautorio sin probabilidades matemáticas ............................. 148
Estimación de variabilidad en modelos pesqueros ........................................ 149
Estimaciones “jackknife-bootstrap” para intervalos de confianza ........... 150 Análisis de riesgo en modelos pesqueros ..................................................... 152
Aproximación derivada de la aplicación del “bootstrap” ......................... 152 Bibliografía citada .................................................................................................... 154 Indice de autores ..................................................................................................... 169 Indice de tópicos ..................................................................................................... 172

ix
Lista de Figuras 2.1 Función de crecimiento logístico de la biomasa de un recurso para una
capacidad de carga de 3.5 millones de toneladas y una tasa intrínseca de crecimiento de 0.36 ................................................................................................ 9
2.2 Curvas de biomasa en equilibrio (a), rendimiento sostenible (b) y CPUE (c)
basados en el modelo de Schaefer ....................................................................... 12 2.3 Curvas de biomasa sostenible (a), captura sostenible (b) e ingresos totales
sostenibles (ITS) y costos totales (CT) (c) del modelo bioeconómico estático Gordon-Schaefer .................................................................................................. 18
2.4 Rendimientos promedio y marginales (a), e ingresos, costos promedio y
marginales (b) en función del esfuerzo bajo condiciones de acceso abierto ........ 19 2.5 Trayectoria estática (equilibrio) y dinámica de la biomasa (a), las capturas (b) y
los costos e ingresos (c) resultantes de la aplicación de diferentes niveles de esfuerzo ................................................................................................................ 24
2.6 Dinámica de la biomasa (a), la captura (b), la renta económica (c) y el esfuerzo
pesquero (d) ......................................................................................................... 25 2.7 Ejemplo ilustrativo del modelo captura-mortalidad de Csirke y Caddy (1983) ..... 27 2.8 Modelo bioeconómico de estructura por edades: efecto dinámico de diferentes
tallas mínimas de captura en la biomasa (a), la captura (b), la renta (c) y el esfuerzo de pesca (d) ........................................................................................... 40
3.1 Comportamiento dinámico de la biomasa del recurso (a, b), las capturas por
flota (c, d) y de la distribución de la renta económica (e, f) por tipo de flota, como resultado de las interdependencias tecnológicas entre ellas ................... 47
3.2 Trayectoria dinámica de la biomasa (a), el esfuerzo (b), la captura (c) y la renta
económica (d) obtenida por las flotas industrial y artesanal ................................. 48 3.3 Modelo bioeconómico dinámico de interdependencias tecnológicas entre dos
flotas: biomasa de las especies objetivo e incidental (a), esfuerzo pesquero (b), capturas de ambas especies (c) y renta económica generada (d) ...................... 52
3.4 Interdependencias tecnológicas. Niveles de biomasa (a), esfuerzo (b), captura
(c) y renta económica generada por las flotas industrial y artesanal (d) .............. 55 3.5 Variación temporal en la abundancia de los bivalvos suspensívoros
Mesodesma mactroides y Donax hanleyanus de la costa atlántica uruguaya (de Alava, 1993) ................................................................................................... 58

x 3.6 Trayectorias dinámicas de la biomasa (a, b), la captura por especie (c, d), así
como la renta económica (e, f) generada por una flota con arte de pesca no discriminatorio ..................................................................................................... 61
3.7 Trayectoria dinámica de las biomasas de especies con relación depredador-
presa (a), el esfuerzo de dos flotas que inciden diferencialmente sobre ellas (b), las capturas correspondientes (c) y la renta económica estimada para cada flota (d). ....................................................................................................... 62
3.8 Interacción tecno-ecológica: stocks con interacción depredador-presa
capturados por dos flotas tecnológicamente interdependientes .......................... 65 4.1 Preferencia intertemporal neutra (adaptada de Randall, 1981) ........................... 71 4.2 Preferencia intertemporal positiva (adaptada de Randal, 1981) .......................... 71 4.3 Preferencia intertemporal negativa (adaptada de Randall, 1981) ........................ 72 5.1 Enfoque de análisis y modelación bioeconómica de una pesquería (Seijo,
1989) .................................................................................................................... 93 5.2 Caracterización de una pesquería ....................................................................... 95 5.3 Dinámica en el corto y mediano plazo del reclutamiento (a), del esfuerzo (b),
de las capturas (c, d) y de la renta y costos (e, f) para dos tipos de flotas en una pesquería hipotética de camarón . ........................................................... .... 108
6.1 Diagrama de bloques conceptual para el problema de optimización de
pesquerías con criterios múltiples ...................................................................... 121 7.1 Mesodesma mactroides. Comparación de la abundancia observada y estimada
por el modelo de largo plazo para reclutas (a) y adultos (b) ............................... 140 7.2 Variaciones trimestrales en la intensidad de pesca simulada por zona en base
al modelo de largo plazo (1983-1992) para la pesquería de almeja amarilla Mesodesma mactroides de las costas de Uruguay ............................................. 140
7.3 Mesodesma mactroides. Comparación de la intensidad de pesca observada y
simulada por estación del año (a); variaciones espaciales de la captura por zona de pesca (verano 1985) observada y estimada por el modelo de corto plazo (b) ................................................................................................................ 141

xi
Lista de Tablas 2.1 Parámetros para el modelo bioeconómico dinámico (Gordon-Schaefer) ............. 23 2.2 Parámetros para el modelo bioeconómico dinámico de estructura por edades ... 39 3.1 Parámetros definidos al inicio de la corrida de simulación para evaluar el
comportamiento dinámico de la biomasa poblacional, capturas, esfuerzo y renta económica generada por dos flotas con diferente poder de pesca ............. 46
3.2 Parámetros definidos al inicio de la corrida de simulación para evaluar el
comportamiento dinámico de la biomasa poblacional de dos especies X e Y capturadas por flotas tecnológicamente interdependientes .................................. 51
3.3 Parámetros incluidos en un modelo bioeconómico multiespecífico de
estructura por edades con interdependencias tecnológicas secuenciales .......... 54 3.4 Interdependencia tecno-ecológica. Parámetros definidos al inicio de la corrida
de simulación para evaluar el comportamiento dinámico de la biomasa poblacional de las especies X (presa) e Y (depredador) capturadas por flotas tecnológicamente interdependientes .................................................................... 64
4.1 Costos, beneficios y reglas de decisión de dos proyectos con similar vida de
ejecución. Se asume una tasa de descuento d del 8% (Pearce & Nash, 1981) .... 79 4.2 Flujo de costos y beneficios para cada una de las alternativas de artes de
pesca A, B y C ...................................................................................................... 80 4.3 VPN y B/C para cada una de las alternativas de artes de pesca A, B, y C,
evaluadas con diferentes tasas de descuento ..................................................... 81 5.1 Parámetros bioeconómicos de la pesquería hipotética de camarón .................... 107 6.1 Criterios de bienestar aplicados a pesquerías marinas ........................................ 112 7.1 Propiedades del modelo SAE ............................................................................... 129 8.1 Valor presente neto (US$ ‘000) de tres estrategias alternativas de ordenación
(D1, D2, D3 ) bajo diferentes estados de la naturaleza (S1, S2) y sus correspondientes probabilidades de ocurrencia (P1/P2) ....................................... 148
8.2 Aplicación del criterio Maximin ............................................................................. 148 8.3 Aplicación del criterio Minimax: matriz de pérdida de oportunidades (US$´000) . 149 8.4 Aplicación del criterio Maximax ............................................................................ 149

xii
Glosario de Símbolos Símbolo Significado Unidad
de medida
a constante de la relación peso-longitud --- ad factor de compensación --- área área barrida por día km²/día α1 coeficiente de competencia --- α3 constante que considera el efecto de la especie X sobre la Y --- α4 término o constante de depredación: especie X --- Aj tasa finita de mortalidad total 1/t ALIM(t) contribución de la pesquería a la disponibilidad de alimento en la
zona costera ton/año
AREA área ocupada por el stock km² b exponente de la relación peso-longitud --- bβ valor de la corrida base para el criterio β --- β1 coeficiente de competencia --- β3 constante que considera el efecto de la especie Y sobre la X --- β4 constante de depredación: especie Y --- βp constante que relaciona el número de presas con la biomasa
máxima capaz de ser alcanzada por el depredador ---
Be biomasa de la población en equilibrio ton B0 biomasa inicial ton BEBE biomasa en equilibrio bioeconómico ton B(t) biomasa de la población en el tiempo t ton Bmax biomasa máxima ton BT(t) beneficios totales en el tiempo t $ B∞ población virgen; biomasa virgen ton Bxp biomasa en el nivel de esfuerzo p para la especie X Bxc biomasa crítica de X ton BOP nivel óptimo de biomasa ton c costo unitario del esfuerzo pesquero $/ue cp costos de procesamiento $/año Co capital inicial $ C (t) captura en el tiempo t ton/año Cj rendimiento por edad ton Ct rendimiento anual ton CMRE captura en máximo rendimiento económico ton Cinc captura promedio incidental por viaje de pesca ton C/Rj rendimiento por recluta g/ind CT(t) costos totales en el tiempo t $ CV(t) costos variables diarios del esfuerzo pesquero en el tiempo t $/día CPUE (t) captura por unidad de esfuerzo en el tiempo ton/ue CPUEmax captura por unidad de esfuerzo máxima observada ton/ue δ tasa de descuento continua anual --- d tasa de descuento discreta --- dy1, dy2,..,dyn cambio de los beneficios netos de cada miembro de la comunidad US$/t dW cambio neto en el bienestar social US$/t D1, D2,...Dn estrategias alternativas de ordenación --- DE desviación estándar ---

xiii Dkh distancia del puerto h al sitio de pesca k km DEL retraso en el tiempo de ocurrencia de un fenómeno t DELHS valor esperado del tiempo de maduración de huevos de una especie meses DELm valor esperado del tiempo de entrada de embarcaciones a la
pesquería meses
DP número promedio de días efectivos de pesca por mes días/mes ε proporción de la captura por especie --- EBE equilibrio bioeconómico : π (t) = 0 $ EMP (t) empleos directos generados por la pesquería # EXPm (t) generación de divisas, sector capturas $/año EXP p(t) generación de divisas, sector procesamiento $/año fa asignación espacial de la intensidad de pesca acumulada a lo largo
de la temporada ---
fkl (t) intensidad de pesca ejercida en el sitio con coordenadas geográficas kl en el tiempo t
días
fEBE esfuerzo en equilibrio bioeconómico ue fMRE esfuerzo en máximo rendimiento económico ue fMRS esfuerzo en máximo rendimiento sostenible ue f(t) esfuerzo de pesca en el tiempo t ue fop esfuerzo óptimo ue F tasa instantánea de mortalidad por pesca 1/t FC costo fijo diario por embarcación $/día FEC fecundidad promedio por hembra # huevos FMRE tasa instantánea de mortalidad por pesca en MRE 1/t FMRS tasa instantánea de mortalidad por pesca en MRS 1/t FR(t) tasa de explotación en el tiempo t --- g orden del proceso de retraso --- gβφ objetivo para el criterio β del conjunto de objetivos φ --- H proporción de hembras presentes en la estructura de la población --- Hmax número máximo anual de huevos generados por el stock desovante # huevos HS (t) número de huevos estimados del stock desovante # huevos i especie --- IT (t) ingresos totales en el tiempo $ ITD (t) ingresos totales por día en el tiempo t $/día ITS ingresos totales sostenibles $ j edad --- k coordenada de localidad: latitud; sitio de pesca --- kp parámetro de curvatura ecuación de crecimiento individual 1/t K capacidad de carga del ambiente ton KX capacidad de carga de la especie X ton KY capacidad de carga de la especie Y ton λ índice de conversión alimenticia X a Y --- l coordenada de localidad: longitud --- L longitud (talla) individual mm L∞ longitud asintótica mm L50% longitud al 50 % de retención del arte de pesca mm L75% longitud al 75 % de retención del arte de pesca mm LLi límite inferior constante o en función de variables independientes --- m tipo de embarcación o flota --- M tasa instantánea de mortalidad natural 1/t MAGE edad máxima t MN(t) tasa finita de mortalidad natural en el tiempo t --- MRE máximo rendimiento económico $ MRS máximo rendimiento sostenible ton µ opción de manejo --- N abundancia poblacional # ind

xiv Nj número de individuos de cada cohorte # ind NT número de reclutas a distribuirse en un parche # ind OCVm parámetro que agrupa costos variables (carnada, mantenimiento,
reemplazo de artes de pesca y avituallamiento) $/año
Ωβδφ desviación normalizada del valor del criterio β con respecto a la opción de manejo µ y al conjunto de objetivos φ
---
π(t) ingresos netos en el tiempo t $/año πM renta marginal $/ue p precio (promedio) de la especie capturada $/ton h puerto --- pcap probabilidad de captura del arte de pesca utilizado --- Pescm número promedio de tripulantes por embarcación tipo m # ind Pinc precio promedio ponderado pagado al pescador por especies
incidentales $/ton
Pk probabilidad de encontrar a la especie objetivo en los sitios alternativos de pesca k
---
Pn probabilidad de ocurrencia de un evento alternativo n --- P1 , P2 probabilidades de ocurrencia --- Ptar precio promedio pagado al pescador por especie objetivo $/ton Ψt consumo total en el tiempo t --- q coeficiente de capturabilidad --- qua coeficiente de capturabilidad por unidad de área --- quasiπ (t) quasi renta de los costos variables en el tiempo t $/año Qn unidades de determinado bien # r tasa intrínseca de crecimiento poblacional --- R número de peces vivos al tiempo t=tr (edad de reclutamiento) # ind Rj reclutamiento estimado por edad j # ind R’ número de individuos reclutados a la pesquería # ind Re rendimiento en equilibrio ton RET retención del arte de pesca utilizado % Rkl (t) número de reclutas asentándose a partir del centro del parche con
coordenadas kl en el tiempo t ind/t
Riskkl riesgo percibido por los pescadores de pescar en los sitios alternativos de pesca kl
---
Rm rendimiento marginal ton/ue Rm (t) variable aleatoria de la captura exponencialmente correlacionada ton/t Rmax reclutamiento máximo observado # ind RNi(t) (0, var) variable aleatoria normalmente distribuida en el tiempo t
para modelar reclutamiento estocástico ind/t
Rp rendimiento promedio ton/ue ROS rendimiento óptimo sostenible ton RVkpm(t) variable aleatoria generada con una función de densidad
probabilística exponencialmente autocorrelacionada ---
si edad de primera madurez sexual de la especie i t S1 , S2 estados de la naturaleza --- SAE (t) distribución (0, 1) espacial del esfuerzo pesquero en el tiempo t --- SEL selectividad del arte de pesca % SG número de filas de la cuadrícula geográfica --- Sj tasa finita de sobrevivencia --- SRij (t) tasa de supervivencia de individuos de la especie i, edad j en el
tiempo t. ---
σ2 desviación estándar de la densidad del parche (0< σ < 1) --- φ fricción de la distancia --- t tiempo varias tc edad de primera captura t tl edad de máxima longevidad t

xv to parámetro de ajuste de la ecuación de von Bertalanffy t tr edad de reclutamiento a la pesquería t TG número de columnas de la cuadrícula geográfica # TMPT tasa marginal de preferencia temporal %/t TSG largo de la celda estimada para una resolución espacial
especificada (grados, minutos y segundos) km
TTG ancho de la celda estimada para una resolución espacial especificada (grados, minutos y segundos)
km
θ costo promedio de combustible y lubricantes por km recorrido $/Km ρ tasa interna de retorno (TIR) % ϕ parámetro de dinámica de la flota --- υβ vector de parámetros de control óptimo de la pesquería --- υµ vector de parámetros de decisión correspondiente a la opción de
manejo µ ---
ue unidad de esfuerzo varias U captura por unidad de esfuerzo (notación generalizada) ton/ue Ue captura por unidad de esfuerzo en equilibrio (notación generalizada) ton/ue ULi límite superior constante o en función de variables independientes --- Umn utilidad marginal del ingreso $ U∞ tasa de captura del stock virgen ton/t VAR varianza del valor esperado de VPN $ VE valor esperado del VPN $ VME valor del producto marginal del esfuerzo pesquero $/ue VPB valor presente de los beneficios $ VPC valor presente de los costos $ VPE valor promedio del esfuerzo pesquero $/ue VPN valor presente neto $ VPπ valor presente de los ingresos netos $ V número de embarcaciones # Vt,d factor de descuento --- ϖ proporción de la captura asignada a la tripulación como pago por su
esfuerzo pesquero [0,1]
w1, w2,..., wn coeficientes de distribución unitarios --- wpi precio de planta $/ton W peso individual g W∞ peso asintótico g x distancia de la unidad de área kl al centro del parche km xpi precio de playa $/ton Y Ingreso percibido por un pescador $/t γg (t) salidas del proceso de retraso (e.g. número de embarcaciones que
entran a la pesquería a lo largo de la temporada) # / t
γ1 (t)... γg -1 (t) tasas intermedias del proceso de retraso # / t Yβ* vector de valores óptimos de la función de criterios múltiples --- Yβµ valor del criterio β de la estrategia de manejo µ --- Yµ (T) vector de valores de la función con criterios múltiples
correspondiente a la opción de manejo µ, evaluada en el horizonte de tiempo (0,T)
---
Yβµφ desviación normalizada del valor del criterio β con respecto a la opción de manejo µ y al conjunto de objetivos φ
---
Z tasa instantánea de mortalidad total 1/t ZMRS tasa instantánea de mortalidad total en MRS 1/t Zt mortalidad total en el año t 1/t

1
CAPITULO 1
Recursos Pesqueros: Características Inherentes El manejo de un recurso pesquero es un proceso complejo que requiere la integración de su biología y ecología con los factores socio-económicos e institucionales que afectan al comportamiento de los usuarios (pescadores) y a los responsables de su administración. Aún cuando los planes de manejo han mejorado ostensiblemente a través del tiempo, gracias a la obtención de series de tiempo prolongadas y a la elaboración de modelos sofisticados, muchos recursos pesqueros críticos han sido inevitablemente sobreexplotados, llegando incluso a niveles cercanos a su colapso (Ludwig et al., 1993). ¿Cuáles son los motivos por los que los recursos pesqueros tienden a ser sobreexplotados? ¿Cómo podría explicarse este síndrome de sobreexplotación, tanto en las pesquerías artesanales como industriales? Entre las múltiples razones que han sido invocadas para explicar este fenómeno, algunas surgen inevitablemente de las características inherentes que definen a un recurso y a la pesquería en éste sustentada. En este Capítulo describiremos las características inherentes de los recursos pesqueros y las interdependencias humanas que pudieran condicionar los niveles de esfuerzo pesquero que llevarían a su sobreexplotación.
Optima asignación de recursos naturales: supuestos básicos Para que pueda existir una óptima asignación de recursos naturales en una economía específica, se requiere contar con derechos de propiedad no atenuados. Esto implica que dichos derechos sean (Randall, 1981; Schmid, 1978): (i) Completamente especificados en términos de los derechos que acompañan la propiedad sobre el recurso, las restricciones sobre esos derechos y las penalizaciones correspondientes a su violación. (ii) Exclusivos, de tal forma que el individuo que posee dichos derechos reciba las retribuciones y las penalizaciones correspondientes al uso del recurso natural en cuestión. (iii) Transferibles, a efectos de que los derechos sobre el uso de los recursos naturales estén en manos de quienes tienen la capacidad de conducirlos a su más alto valor de uso. (iv) Efectivamente vigilables, ya que un derecho no vigilable es un derecho vacío.
El fracaso en la óptima asignación de recursos pesqueros
En el caso de recursos pesqueros se violan los supuestos básicos del modelo neoclásico de mercado mencionados anteriormente, lo cual ha conducido inevitablemente a la sobreexplotación

2
de muchos recursos pesqueros de importancia en el mundo. A continuación se mencionan los factores relevantes que originan dicho fracaso. Regímenes de propiedad Los recursos pesqueros pueden estar sujetos a cuatro diferentes regímenes de propiedad (Bromley, 1991): estatal, privada, común y acceso abierto. (i) Si los usuarios del recurso tienen el deber de observar y cumplir reglas y normas de uso y acceso, determinadas por una institución del gobierno que es responsable y tiene el derecho de su manejo, entonces el recurso pesquero es de propiedad estatal. (ii) Si los pescadores tienen el derecho de realizar un uso socialmente aceptable del recurso pesquero, y tienen el deber de abstenerse de hacer un uso inadecuado del mismo, entonces el régimen de explotación es de propiedad privada. (iii) Si el Estado ha asignado derechos de propiedad a grupos de pescadores específicos, (e.g. cooperativas pesqueras), y éstos tienen el derecho de excluir a otros pescadores, entonces el régimen de explotación es de propiedad común (res communis). Esta situación implica una condición necesaria pero no suficiente para que se produzca el fracaso en la óptima asignación de un recurso. (iv) En condiciones de acceso abierto (res nullius) no existe propiedad sobre el recurso, por lo que cualquier miembro de la sociedad (e.g. cualquier pescador) puede obtener el recurso por apropiación directa. El síndrome de sobreexplotación de recursos pesqueros indica que este régimen conduce al fracaso en su óptima asignación. En otras palabras, el libre acceso constituye una condición necesaria y suficiente para la sobreexplotación de un recurso pesquero. Se suscitan dos situaciones básicas: (1) acceso irrestricto al recurso a aquellos que deseen hacer uso de él; y (2) generación de externalidades (negativas y positivas) entre los usuarios del ecosistema relevante a dicho recurso. Externalidades Una externalidad es definida como todo efecto externo no contabilizado por el pescador que lo genera, pero que sí afecta a otros usuarios del recurso. En pesquerías las externalidades son generalmente negativas, y ocurren cuando los pescadores pueden entrar libremente a capturar un recurso pesquero y además no existe un acuerdo de cooperación voluntaria; en tales casos los usuarios del recurso no toman en consideración los efectos externos que se imponen entre sí. Agnello & Donnelley (1976) han identificado tres tipos de externalidades usualmente presentes en la mayoría de las pesquerías: del stock, de aglomeración y del arte de pesca. Además de estas pueden mencionarse tipos adicionales de externalidades originadas por interdependencias tecnológicas y ecológicas (ver además Seijo & Defeo, 1994a). (i) Externalidades del stock. Ocurren cuando la entrada de nuevas embarcaciones a la pesquería reduce la magnitud del recurso a los pescadores ya existentes, incrementando sus costos de extracción. Este costo no es tomado en cuenta por los pescadores, debido a que cada

3
uno contabiliza únicamente sus costos privados (internos) incurridos en el viaje de pesca, desestimando los costos externos que le impone a otros en términos de la reducción del recurso. (ii) Externalidades de aglomeración. El acceso irrestricto a un recurso ocasiona el aumento y la congestión de embarcaciones en un sitio de pesca restringido. Luego de cierto umbral, que es función de la extensión del sitio de pesca y de la magnitud del recurso, cada barco que se incorpora a la zona de pesca le impone externalidades de aglomeración a los demás barcos. Ello resulta en el incremento de los costos marginales de captura, ya que las unidades de pesca no asignarán óptimamente su esfuerzo pesquero ni en el espacio (e.g. sobre las mayores concentraciones del recurso) ni en el tiempo (e.g. deberán esperar su turno para acceder al sitio específico de captura). Este tipo de externalidad es común en recursos sésiles o sedentarios con distribución fuertemente agregada, donde la estrategia de explotación tiende a ser exterminadora de parches. También suele darse en casos de agregaciones de individuos por la presencia de condiciones naturales (e.g. arrecifes) o artificiales (e.g. hábitats artificiales, dispositivos de agregación). (iii) Externalidades tecnológicas. Se producen cuando el arte de pesca utilizado cambia la estructura dinámica de las poblaciones objetivo y de aquellas que constituyen la captura incidental asociada, imponiéndole efectos externos negativos a otros pescadores dentro de la misma pesquería y afectando la abundancia de las especies incidentales que constituyen especies objetivo en otras pesquerías de la región. Relacionadas con este tipo de externalidad se encuentran las externalidades secuenciales y las incidentales. (iii.1) Externalidades secuenciales. En pesquerías donde existen flotas artesanales o de pequeña escala y flotas industriales o mecanizadas que inciden sobre diferentes componentes de la estructura de la población de una misma especie objetivo, se generan interdependencias tecnológicas entre los dos tipos de flotas. Usualmente las embarcaciones artesanales, por su autonomía y características físicas, ejercen su esfuerzo pesquero en zonas cercanas a la costa donde habitan juveniles o adultos jóvenes de especies de peces, crustáceos y moluscos, mientras que la flota mecanizada por su mayor autonomía y tamaño opera generalmente en aguas profundas, a mayor distancia de la costa, en zonas de ocurrencia de adultos. A modo de ejemplo, incrementos sustantivos en el esfuerzo pesquero efectivo de la flota artesanal causarán una disminución en la abundancia de adultos en períodos subsiguientes, generando externalidades negativas a pescadores de la flota mecanizada. Análogamente, el aumento del esfuerzo pesquero por parte de la flota mecanizada tenderá a disminuir el stock desovante, pudiendo afectar en períodos subsiguientes el reclutamiento de juveniles y adultos jóvenes en zonas donde opera la flota artesanal. (iii.2) Externalidades incidentales. En pesquerías tecnológicamente interdependientes las flotas utilizan artes de pesca que no discriminan entre especies, de tal forma que existe una captura incidental en una pesquería A, que disminuye la abundancia del recurso en otra pesquería B, para la cual dicha captura incidental constituye su captura objetivo. El efecto negativo externo no contabilizado por pescadores de la pesquería A constituye una externalidad incidental. Esta es común en pesquerías interdependientes como las de camarón y de peces demersales, donde cada flota captura la respectiva especie objetivo en la misma zona de pesca. La pesquería de camarón genera capturas incidentales de especies demersales, efecto negativo no contabilizado por la flota de camarón, lo cual genera una externalidad.

4
(iv) Externalidades debidas a interdependencias ecológicas. Ocurren en pesquerías ecológicamente interdependientes, e.g. las respectivas especies objetivo tienen una relación ecológica de competencia o depredación. Cuando dos especies que compiten, constituyen el objetivo de diferentes pesquerías, los diferentes niveles de esfuerzo pesquero ejercidos por ambas flotas podrán modificar la magnitud y dirección de la interacción y en consecuencia alterar la abundancia relativa de las especies en una misma zona de pesca (ver Capítulo 3). A modo de ejemplo, considérese dos pesquerías A y B que capturan las especies objetivo S1 y S2 respectivamente, las cuales, si bien compiten por cierto recurso limitante, coexisten en ausencia de impacto humano. Un aumento del esfuerzo sobre la especie competitiva dominante S1 determinará incrementos en la abundancia de la especie S2, sobre la cual se mantiene un esfuerzo pesquero sostenido por parte de la pesquería B. En este caso puede decirse que la pesquería A le genera efectos externos positivos a la pesquería B, definidos como externalidades en coexistencia competitiva. Por otra parte, supóngase que en ausencia de impacto humano la especie competitiva inferior o subordinada S2 es excluida por la especie dominante S1. La aplicación de un esfuerzo pesquero sostenido por parte de los pescadores de la pesquería A sobre S1 tenderá a disminuir el stock, lo cual producirá a su vez el aumento de la especie competitiva inferior S2 capturada por la pesquería B, debido a un aumento en la disponibilidad del recurso limitante (e.g. espacio, alimento). En consecuencia, la pesquería A genera un efecto externo positivo a la B, el cual constituye una externalidad por liberación competitiva. Como se puede apreciar, en ambos casos ocurre una liberación de recurso limitante como consecuencia de la explotación, permitiendo mayor viabilidad a la especie dominada. La diferencia situacional radica en que en ausencia de esfuerzo pesquero la especie subordinada está o no presente. De manera análoga a las anteriores, la interdependencia ecológica depredador-presa orienta la dirección de las externalidades. Un incremento en el esfuerzo pesquero de la pesquería A que tiene como especie objetivo a la presa Sp, generará un decremento en la abundancia del depredador Sd (especie objetivo de la pesquería B que para efectos del ejemplo se considera depredador especialista, ver Begon et al., 1990), ocasionando una externalidad negativa a ésta. El caso inverso sucede cuando el incremento del esfuerzo ocurre en la pesquería B: al disminuir la abundancia del depredador se incrementa la abundancia de la presa, generándose en consecuencia una externalidad positiva para la pesquería A. Ambos efectos externos constituyen externalidades por interdependencia trófica. (v) Externalidades tecno-ecológicas. Suceden cuando el arte de pesca perturba el hábitat de la especie objetivo y de otras especies acompañantes que eventualmente constituyen el objetivo de otras pesquerías. La alteración del hábitat disminuye la probabilidad de recolonización y reclutamiento de las especies que lo habitan. Este tipo de externalidad es común en recursos bentónicos capturados por medio de artes de arrastre, los cuales en muchos casos alteran la calidad del sustrato en el cual habitan las especies objeto de explotación.

5
Adicionalmente a las externalidades antes mencionadas, existen otras características en pesquerías marinas que requieren ser analizadas para comprender mejor las interdependencias entre los usuarios de los recursos y los esfuerzos pesqueros resultantes. Altos costos de exclusión La naturaleza de los recursos pesqueros hace que se viole el supuesto de exclusividad en propiedad, generándose altos costos de exclusión. En efecto, el régimen de libre acceso, combinado con la alta variabilidad espacio-temporal en la magnitud de los recursos, hace que un pescador no se beneficie en posponer la captura con la esperanza de obtener peces mayores y más valiosos en el futuro, debido a que otro pescador puede capturarlos en ese mismo lapso. En otras palabras, un pescador, aunque lo deseara, no puede incrementar el tamaño del stock a través de la reducción de su tasa de captura, a menos que los restantes participantes de la pesquería acuerden restringir proporcionalmente su esfuerzo pesquero (Eckert, 1979). Lo anterior genera un aumento en la competencia de los pescadores por capturar la mayor cantidad en el menor tiempo posible, ocasionando altos costos de exclusión. Los esquemas tradicionales destinados a evitar estos altos costos de exclusión involucran al menos cuatro aproximaciones básicas: (i) privatización del recurso a través de la asignación de cuotas individuales; (ii) intervención estatal a través de la regulación del tamaño y composición de las capturas y de la intensidad del esfuerzo de pesca; y más recientemente (iii) la adopción de sistemas de manejo de recursos regulados por la comunidad (Berkes, 1989; Smith & Berkes, 1991) o bien (iv) la combinación de los anteriores (Seijo, 1993). Trampa social en pesquerías Sin un acuerdo para limitar las capturas, la reducción de la tasa de captura de un pescador disminuirá los costos de extracción de otros pescadores, sin necesariamente incrementar sus propios beneficios en el futuro. Consecuentemente, cada pescador tenderá a incrementar su tasa de captura y por tanto contribuirá al colapso de la pesquería, resultado de largo plazo no deseado por la mayoría de los pescadores involucrados. Esto indicaría la existencia de una trampa social en pesquerías ya que, utilizando la terminología de Schelling (1978), los micro-motivos de un pescador en el corto plazo son inconsistentes e incompatibles con los macro-resultados que él y los demás pescadores desean en el largo plazo. Los micro-motivos de corto plazo consisten en explotar la mayor cantidad de recurso posible a efectos de incrementar los beneficios marginales de los pescadores, mientras que los macro-resultados de largo plazo conllevan a lograr un rendimiento máximo sostenible en el tiempo. La incertidumbre sobre la disponibilidad futura del recurso determina que la consecución de mayores beneficios marginales (micro-motivo) como resultado de un incremento del esfuerzo pesquero, predomine sobre los macro-resultados de largo plazo. El rendimiento sostenible de una pesquería será una alternativa viable únicamente cuando el número de pescadores que asigne su esfuerzo pesquero esté limitado por regulaciones que restrinjan la entrada a una pesquería. Ello determinará la existencia de un nivel de biomasa sostenido en el tiempo, en base a la aplicación de una intensidad de esfuerzo pesquero considerada óptima por la comunidad, dadas las preferencias intertemporales de los

6
usuarios del recurso. La asignación intertemporal del esfuerzo pesquero será discutida en el Capítulo 4. Usuarios no contribuyentes Una vez establecidas las normas de explotación del recurso (legal o comunitariamente), se observa con frecuencia la presencia de usuarios no contribuyentes al logro de un rendimiento sostenible en una pesquería (Clark, 1982). Pueden distinguirse dos tipos de usuarios no contribuyentes (Schmid, 1987): involuntarios e intencionales. La cantidad de pescadores es un factor relevante en la ocurrencia de la trampa social mencionada anteriormente y tiene directa relación con la presencia de usuarios no contribuyentes en pesquerías. Cuando el grupo de pescadores es grande, un pescador puede constituirse en usuario no contribuyente involuntario, al no percibir como evitar el macro-resultado (destrucción de la pesquería) cuando no puede estar seguro que otros pescadores actuarán en concierto para sostener el rendimiento del recurso. La carencia de información en cuanto a la ejecución de actividades coordinadas con otros pescadores, así como en lo referente a la situación del recurso, es relevante para que se produzca la presencia de este tipo de usuario. Los usuarios no contribuyentes intencionales toman la decisión de infringir alguna norma legal existente sobre el régimen de explotación de un recurso, aún a pesar de percibir cierta probabilidad de ser sorprendidos. Cuando el grupo es pequeño, los costos de exclusión no son necesariamente menores, pero el usuario no contribuyente podría ser más fácilmente identificado (Schmid, 1987), por lo cual su presencia pudiera tender a disminuir. Dadas las implicaciones en vigilancia y cumplimiento de las normas de explotación, éstos son aspectos fundamentales en el posible comportamiento de los pescadores que requieren mayores esfuerzos de investigación. Altos costos de transacción Las pesquerías marinas involucran altos costos de transacción, lo cual genera otra fuente de atenuación de derechos de propiedad que impide la eficiente asignación temporal de los recursos pesqueros. Los costos de transacción involucran un grupo de costos discutidos en la literatura como: costos de información, costos de vigilancia y costos contractuales (Schmid, 1987, Randall, 1981). (i) Costos de información. El manejo de recursos pesqueros involucra altos costos de información resultantes de esfuerzos de investigación multidisciplinaria sobre aspectos biológicos, ecológicos, estadísticos y socioeconómicos. Dichas investigaciones están dirigidas a mantener actualizada la información sobre la magnitud, dinámica poblacional y distribución espacio-temporal del recurso y de aquellas variables físicas y químicas del ecosistema, conjuntamente con la evolución histórica y dinámica espacial de las capturas y del esfuerzo aplicado. La incertidumbre existente en muchos de estos tópicos incide a la hora de ejecutar planes de regulación, lo cual introduce una nueva fuente de inseguridad a los usuarios del recurso,

7
aumentando la probabilidad de ocurrencia de usuarios no contribuyentes y de disipación de renta económica. (ii) Costos de vigilancia. A diferencia de los recursos donde existen derechos de propiedad no atenuados, el manejo de recursos pesqueros supone altos costos de vigilancia que resultan de la implementación y puesta en práctica de esquemas regulatorios de manejo (e.g. vigilancia en áreas de veda), así como de la asignación de derechos de propiedad. En muchos casos las áreas de vigilancia son tan extensas (pesquerías oceánicas) o bien tan accesibles a terceros (pesquerías en litorales arenosos y rocosos someros) que los esfuerzos de vigilancia son costosos e inefectivos. Cuando esto sucede, el derecho no vigilable se convierte en un derecho vacío. (iii) Costos contractuales. Se dan en países que poseen una legislación tendiente a promover cierto tipo de organización (e.g. cooperativas), otorgándoles el derecho de propiedad sobre la explotación de un determinado recurso. En tal situación, los costos derivados de promover dicho tipo de organización se vuelven importantes, por lo cual se debe identificar a aquél (e.g. pescadores o Estado) que se hace cargo de los costos involucrados a tales fines. Algo similar sucede cuando el Estado está interesado en promover cierta estrategia de manejo, como pudiera ser la implementación de Cuotas Individuales Transferibles (CIT; ver Geen & Nayar, 1988) o bien Zonas Individuales Transferibles (ZIT) entre los miembros de una comunidad pesquera (Seijo, 1993) a efectos de maximizar la renta del recurso en el tiempo. Las características inherentes de los recursos pesqueros desarrolladas en este Capítulo los diferencian de otros recursos naturales sujetos a explotación. El acceso irrestricto a los mismos aumenta la probabilidad de fracaso en su óptima asignación, y lleva finalmente a la sobreexplotación y disipación de los beneficios económicos derivados de la actividad. La literatura referida a bioeconomía pesquera invoca generalmente la asignación de derechos privados de explotación como medio para atenuar los problemas de sobreexplotación. A tales efectos, se han desarrollado modelos bioeconómicos estáticos y dinámicos que constituyen el soporte teórico fundamental para la propuesta de métodos de regulación del esfuerzo pesquero y de las capturas. Lo anteriormente expuesto será objeto de análisis en los restantes Capítulos del presente libro.

8
CAPITULO 2
Modelo Bioeconómico Básico Como ha sido expuesto en el primer Capítulo, en condiciones de acceso abierto fracasa la óptima asignación de recursos pesqueros, debido entre otros factores, a la ocurrencia de altos costos de exclusión. El esfuerzo pesquero ejercido por una embarcación se ve influenciado por la forma en que otras hagan uso del recurso, de tal manera que la entrada de más embarcaciones incrementará el esfuerzo de pesca sobre éste, afectando los rendimientos de los diferentes usuarios de la pesquería. Para realizar estimaciones y predicciones de los posibles impactos bioeconómicos en pesquerías sujetas a diferentes regímenes de explotación, se hace necesario modelar matemáticamente estos sistemas dinámicos. Para ello se requiere conocer los fundamentos de la teoría bioeconómica de pesquerías, los cuales permitirán explorar e introducir las complejidades inherentes al manejo de recursos pesqueros en los siguientes Capítulos. El presente Capítulo se centrará en el desarrollo del modelo bioeconómico clásico de Gordon (1954), también llamado Gordon-Schaefer, porque al igual que el de este último, parte del modelo logístico de crecimiento poblacional de Verhulst (1838). En la primera sección se presenta el modelo logístico, exponiendo algunas de sus limitaciones en especies marinas. En la segunda sección se detalla el modelo de rendimiento excedente propuesto por Schaefer (1954) en términos de biomasa y de captura por unidad de esfuerzo; se discuten sus supuestos y se presentan algunas variantes propuestas por otros autores. En la tercera sección se presenta el modelo bioeconómico de Gordon (1954) en términos de captura y esfuerzo, y en función de oferta y demanda, tanto en su versión estática como dinámica. Se plantean sus supuestos y limitaciones, así como modificaciones desarrolladas por otros autores. En la cuarta sección se desarrolla el modelo dinámico propuesto por Smith (1969), el cual condiciona el comportamiento de largo plazo de una flota pesquera a variaciones en los ingresos netos. En la quinta sección se presentan los modelos de producción excedente basados en mortalidad, en sus versiones biológica y bioeconómica. Por último, en la sexta sección se presenta el análisis bioeconómico de modelos que consideran la estructura por edades de una población, tanto en la versión clásica estática (Beverton & Holt, 1957) como dinámica (Seijo & Defeo, 1994b).
El modelo logístico La ecuación logística propuesta por Verhulst (1838) describe el crecimiento de una población en base a la siguiente expresión matemática (Graham, 1938):
dBdt
= rB(t) 1 -B(t)K
⎛⎝⎜
⎞⎠⎟ (2.1)
donde:
9
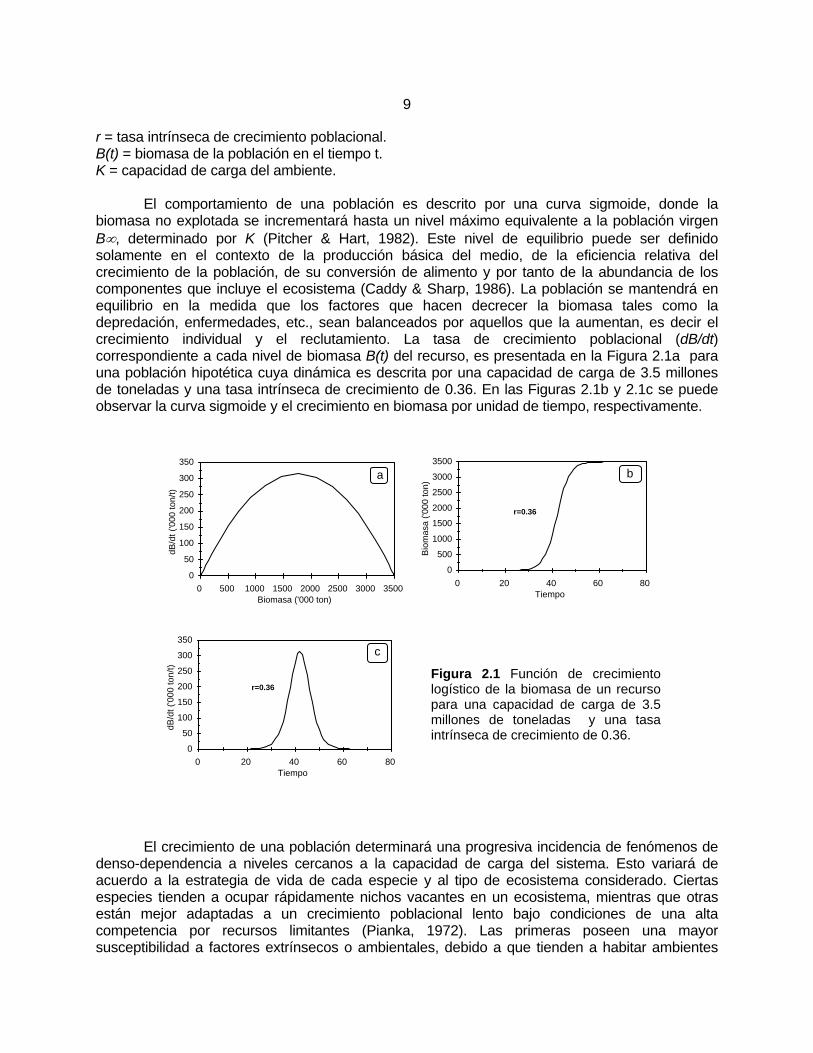
r = tasa intrínseca de crecimiento poblacional. B(t) = biomasa de la población en el tiempo t. K = capacidad de carga del ambiente. El comportamiento de una población es descrito por una curva sigmoide, donde la biomasa no explotada se incrementará hasta un nivel máximo equivalente a la población virgen B∞, determinado por K (Pitcher & Hart, 1982). Este nivel de equilibrio puede ser definido solamente en el contexto de la producción básica del medio, de la eficiencia relativa del crecimiento de la población, de su conversión de alimento y por tanto de la abundancia de los componentes que incluye el ecosistema (Caddy & Sharp, 1986). La población se mantendrá en equilibrio en la medida que los factores que hacen decrecer la biomasa tales como la depredación, enfermedades, etc., sean balanceados por aquellos que la aumentan, es decir el crecimiento individual y el reclutamiento. La tasa de crecimiento poblacional (dB/dt) correspondiente a cada nivel de biomasa B(t) del recurso, es presentada en la Figura 2.1a para una población hipotética cuya dinámica es descrita por una capacidad de carga de 3.5 millones de toneladas y una tasa intrínseca de crecimiento de 0.36. En las Figuras 2.1b y 2.1c se puede observar la curva sigmoide y el crecimiento en biomasa por unidad de tiempo, respectivamente.
0
50
100
150
200
250
300
350
dB/d
t ('0
00 to
n/t)
0 500 1000 1500 2000 2500 3000 3500 Biomasa ('000 ton)
a
0
500
1000
1500
2000
2500
3000
3500
Bio
mas
a ('0
00 to
n)
0 20 40 60 80 Tiempo
r=0.36
b
0
50
100
150
200
250
300
350
dB/d
t ('0
00 to
n/t)
0 20 40 60 80 Tiempo
r=0.36
c
El crecimiento de una población determinará una progresiva incidencia de fenómenos de denso-dependencia a niveles cercanos a la capacidad de carga del sistema. Esto variará de acuerdo a la estrategia de vida de cada especie y al tipo de ecosistema considerado. Ciertas especies tienden a ocupar rápidamente nichos vacantes en un ecosistema, mientras que otras están mejor adaptadas a un crecimiento poblacional lento bajo condiciones de una alta competencia por recursos limitantes (Pianka, 1972). Las primeras poseen una mayor susceptibilidad a factores extrínsecos o ambientales, debido a que tienden a habitar ambientes
Figura 2.1 Función de crecimiento logístico de la biomasa de un recurso para una capacidad de carga de 3.5 millones de toneladas y una tasa intrínseca de crecimiento de 0.36.

10
usualmente variables e impredecibles, y en consecuencia rara vez alcanzan tamaños poblacionales cercanos a la capacidad de carga del ambiente. Las segundas habitan sistemas estables o predecibles, el tamaño poblacional es poco variable en el tiempo y está limitado por procesos de competencia intra e interespecífica en comunidades usualmente saturadas (Pianka, 1972). En el caso de especies marinas, su ubicación dentro de un grupo u otro resulta difícil, dado que la complejidad de sus ciclos de vida parece ubicarlos en ocasiones dentro de un grupo y en otro período dentro de otro (Rothschild, 1986). Tal es el caso de especies con ciclos de vida complejos, que incluyen una fase dispersiva planctónica y una adulta bentónica; durante el estadío larval se encuentran expuestas de manera más marcada a efectos ambientales que en su etapa adulta, en la cual los efectos denso-dependientes pueden ocurrir en forma más intensa.
Modelo de rendimiento excedente y extensiones Bajo condiciones de explotación, Schaefer (1954) introduce el término de tasa de captura C(t), la cual se define como:
C( t )=q f ( t )B( t ) (2.2)
donde la captura depende del esfuerzo de pesca f(t) y de la biomasa poblacional B(t) en el tiempo t; y q representa el coeficiente de capturabilidad, definido como la fracción de la población que es extraída por una unidad de esfuerzo (Gulland, 1983). La ecuación 2.2 implica que la misma captura puede obtenerse a diferentes niveles de esfuerzo y biomasa. La limitante en este sentido es que q, considerado constante para un período de tiempo determinado, no puede obtenerse sin una estimación independiente de la biomasa. En el caso de pesquerías uniespecíficas donde se emplea un mismo tipo de arte, el concepto de q ha sido aplicado de manera más común, debido a que el esfuerzo puede ser estandarizado. En regiones tropicales, donde existe una alta diversidad de artes y especies, es difícil englobar en este coeficiente las complejidades que se presentan en el esfuerzo (Arceo & Seijo, 1991a). En estos casos se ha propuesto el uso de funciones de producción que den mejores aproximaciones (e.g. Prochaska & Cato, 1980; Taylor & Prochaska, 1985), dado que existen una serie de variables asociadas al esfuerzo que pueden intervenir en la producción (Staniford, 1987; Arceo, 1991; Arceo & Seijo, 1991b). Agnello & Anderson (1983) abordan la relación captura - esfuerzo en función de la eficiencia de artes de pesca, utilizando una función tipo Cobb-Douglas. A efectos del desarrollo del modelo de Schaefer, se seguirá manejando el concepto de q, no obstante las limitaciones antes mencionadas. Un análisis detallado de las variaciones de esta variable en función de las características de los recursos y de los artes de pesca empleados, es detallado en Caddy (1979a), Arreguín-Sánchez (1992) y Hilborn & Walters (1992). Considerando una población explotada, los cambios en biomasa se representarán de acuerdo a la siguiente expresión:

11
dBdt
= rB 1-BK
- C⎛⎝⎜
⎞⎠⎟ (2.3)
Cuando la población se encuentra en condiciones de equilibrio, la tasa de incremento poblacional repone las pérdidas debidas a la mortalidad, y por tanto no hay cambios de la biomasa en el tiempo (e.g. dB/dt =0); el rendimiento en equilibrio puede definirse como:
C(t) = rB 1-BK
⎛⎝⎜
⎞⎠⎟ (2.4)
Despejando B(t) se tiene que:
CrB
+ BK
= 1 (2.5)
y sustituyendo (2.2) en (2.5):
qfBrB
+ BK
= 1 (2.6)
Entonces, la biomasa de la población en equilibrio (Be) en función del esfuerzo de pesca se define como:
Be = 1 -qfr
K⎛⎝⎜
⎞⎠⎟ (2.7)
Para un nivel dado de esfuerzo, la población en equilibrio (Be) alcanzará un nivel donde la tasa de captura es igual a la de crecimiento. Por tanto, para cada nivel de esfuerzo existirá un nivel de población en equilibrio (Anderson, 1986). Como resultado se tiene una relación inversa entre la población en equilibrio y el esfuerzo pesquero (Fig. 2.2a). Este comportamiento se puede observar a través de la captura por unidad de esfuerzo (CPUE), considerándose en este caso como un índice relativo de la abundancia de la biomasa de la población en el tiempo (Fig. 2.2c). Para obtener la función que relaciona la captura con el esfuerzo bajo condiciones de equilibrio, se sustituye la ecuación (2.7) en (2.2):
C = qf K 1-qfr
⎛⎝⎜
⎞⎠⎟ (2.8)
La función (2.8) se representa mediante una parábola, donde la captura (C) para un esfuerzo (f) dado en una población en equilibrio es denominado rendimiento sostenible, dado que las pérdidas por mortalidad natural y por pesca son compensadas por un incremento poblacional debido a los procesos de crecimiento individual y reclutamiento; en consecuencia el rendimiento de la pesquería se puede mantener en el tiempo. Esta curva puede ser considerada como la

12
función de producción de la pesquería en el largo plazo (Fig. 2.2b), donde para un nivel dado de esfuerzo f1, se obtiene un rendimiento C1, el cual será mayor a medida que el primero incremente hasta un nivel de máximo rendimiento (MRS) correspondiente a un nivel de esfuerzo f2, después del cual decrece si el esfuerzo aumenta. En este punto, la población ya no podrá reponer las pérdidas por mortalidad, y a medida que la presión de pesca aumente, la captura tenderá a disminuir debido al decremento en la biomasa de la población (C3) a un nivel de esfuerzo f3.
500
1000
1500
2000
2500
3000
3500
Biom
asa
Sost
enib
le ('
000
ton)
0 200 400 600 800 Esfuerzo (barcos)
a
0
50
100
150
200
250
300
350
400
Cap
tura
('00
0 to
n)
0 200 400 600 800 Esfuerzo (barcos)
bMRS
f3f1
C2
C3
C1
f2
200
400
600
800
1000
1200
1400
CP
UE
(ton
/bar
co)
0 200 400 600 800 Esfuerzo (barcos)
c
Supuestos básicos del modelo a) Existen condiciones de equilibrio poblacional, mediante el cual se compensan los procesos de mortalidad con los de crecimiento y reclutamiento. b) Para obtener el rendimiento de la población sujeta a explotación asumiendo condiciones de equilibrio, se parte de que la mortalidad por pesca F es directamente proporcional al esfuerzo f, siendo q la constante de proporcionalidad, tal que:
F = qf (2.9)
Figura 2.2 Curvas de biomasa en equilibrio (a) rendimiento sostenible (b) y CPUE (c) basados en el modelo de Schaefer.

13
c) La CPUE es directamente proporcional a la abundancia de la población, o sea:
CPUE= Cf
= qB (2.10)
d) El recurso se mantiene en un ambiente físico estable determinado por una capacidad de carga constante. e) El recurso lo compone una sola especie, y por tanto el esfuerzo es aplicado exclusivamente a ésta. f) La tecnología utilizada en la extracción se mantiene constante. Limitaciones (i) El inciso (a) asume equilibrio, lo cual constituye un serio inconveniente a la hora de administrar los recursos (Caddy, 1984). Ello implica que se dejan de lado todos los procesos que gobiernan la productividad de un stock (e.g. reclutamiento), muchos de los cuales poseen un alto grado de estocasticidad inclusive en una escala de tiempo reducida. (ii) El coeficiente de capturabilidad q no siempre puede ser considerado constante. Diferentes comportamientos de agregación en cardúmenes de recursos pelágicos o en recursos bentónicos, así como factores relacionados con la diferente selectividad y accesibilidad por clases de talla al arte de pesca, violan dicho supuesto. (iii) La CPUE no siempre es un índice insesgado de abundancia relativa de una población como se asume en el inciso (c), en especial en recursos sedentarios donde la distribución espacial no es homogénea. La limitada movilidad de estos organismos implica que no puedan redistribuirse en el área de pesca una vez aplicado el esfuerzo pesquero. La estrategia dirigida de exterminación de parches determina un comportamiento agregado de los usuarios del recurso, lo cual limita la aplicabilidad del modelo (ver Caddy, 1975, 1989a, b; Conan, 1984; Orensanz et al., 1991 para revisiones detalladas acerca del tópico). (iv) El asumir que se conoce el tamaño de la población resulta riesgoso, debido a la dificultad en la obtención de estimadores insesgados de tamaño poblacional, así como a las fuentes de variabilidad que pudieran producir intensas fluctuaciones en su magnitud. (v) En el inciso (d) se ignora la dinámica de la distribución espacial del recurso, los procesos biológicos que generan la biomasa, así como las interacciones intra e interespecíficas y los efectos de las variaciones estocásticas en el medio ambiente y en la abundancia de la población. Al suponer que el recurso se mantiene en un ambiente físico estable que determina una capacidad de carga constante, se limita la aplicabilidad de este modelo determinístico (Pitcher & Hart, 1982; Sissenwine, 1984a, b). (vi) El supuesto (e) ignora la ocurrencia de interdependencias biológicas (e.g. competencia, depredación) y tecnológicas: ver Capítulo 3. Además, no considera la asignación diferencial del esfuerzo en el corto plazo, esto es, cómo el pescador decide entre la captura de un recurso u

14
otro, cuando el método y el arte de pesca le permiten tener más de una especie objetivo, así como definir cuándo y dónde salir a pescar (ver Hilborn, 1985; Eales & Wilen, 1986; Sampson, 1993). (vii) Por otro lado, incrementos en el poder de pesca (tecnificación de la captura) por parte de la flota determinan variaciones en q que restringen la aplicabilidad del modelo y limitan el supuesto (f). Estimación del rendimiento excedente en base a datos de captura y esfuerzo El modelo de Schaefer requiere información sobre la biomasa, tasa de crecimiento poblacional y capacidad de carga del ambiente, cuya obtención en la práctica es sumamente difícil. A efectos de mitigar este problema, una estimación alternativa del rendimiento sostenible es efectuada en términos de CPUE. Aún cuando esta información es más fácil de adquirir, en el caso de pesquerías multiespecíficas (sobre todo en áreas tropicales), se hace difícil el discernimiento del esfuerzo efectivo aplicado a cada especie. Por otra parte, las estadísticas de esfuerzo en la mayoría de los casos corresponden más al esfuerzo nominal que al efectivo. Sin embargo, esta información es más accesible que aquella específica de la población (e.g. composición por edades), razón por la cual los modelos globales se han utilizado ampliamente. La ecuación (2.10) asume proporcionalidad entre la CPUE y la biomasa en condiciones de equilibrio. Si se define CPUE como U (notación generalizada en los modelos globales) se tiene que:
B( t ) Uq
= (2.11)
Este nuevo valor de la biomasa es sustituido en el modelo para obtener el rendimiento en equilibrio (Re):
R r(U / q )[1-(U / q )]U / qe =
∞
(2.12)
o sea:
e2R = r(U / q )(U -U )∞
Reordenando para resolver en términos de CPUE en equilibrio (Ue):
( )U U U q r fe = −∞ ∞ / (2.13)
donde (U∞ q/r) es la pendiente (b) de la recta y U∞ es la ordenada al origen. En términos de esfuerzo, la función puede ser definida como sigue:

15
U = U f -bf ´e ∞ − (2.14)
y derivando la ecuación anterior con respecto al esfuerzo e igualándola a cero, se obtiene el esfuerzo para el MRS:
f U2bMRS = ∞ (2.15)
Como el nivel dado para el rendimiento sostenible ocurre a la mitad de la máxima biomasa (UMRS =U∞/2), el MRS se obtiene de la siguiente forma:
MRS U4b
= ∞ (2.16)
Extensiones del modelo Se han desarrollado diversas alternativas al modelo de rendimiento de Schaefer. Una de las variaciones comúnmente usadas es el modelo exponencial propuesto por Fox (1970), quien usa la curva de crecimiento de Gompertz (Pitcher & Hart, 1982), la cual muestra una asimetría que es más característica de crecimiento en peso. En este caso la población en equilibrio disminuye más lentamente con respecto al esfuerzo aplicado, obteniéndose una función exponencial a diferencia de la lineal propuesta por Schaefer en términos de Ue. Los cambios en la biomasa en función del peso bajo condiciones de equilibrio, serán definidos por la ecuación (2.17).
dB(t)dt
= rB(t) -B(t)K
- C(t)ln⎛⎝⎜
⎞⎠⎟ (2.17)
Mientras que en la curva logística del modelo de Schaefer el MRS se alcanza a un nivel de K/2, en el de Fox corresponde a rK/q. En este caso sólo se presenta la derivación del modelo de Schaefer (1954), el cual es utilizado como base para el modelo bioeconómico de Gordon. Para las variantes del modelo ver Fox (1970), Pella & Tomlinson (1969), Pope (1979) y Pitcher & Hart (1982).
El modelo bioeconómico Gordon-Schaefer La curva de rendimiento sostenible en función del esfuerzo es la base del modelo económico de Gordon (1954), quien deriva la función de rendimiento incorporando el componente económico. Aborda el problema de los recursos de propiedad común y desarrolla la teoría de acceso abierto. Su principal mérito ha sido clarificar en términos económicos el concepto de "sobrepesca" en pesquerías no reguladas.

16
El modelo establece que los ingresos netos π derivados de la actividad pesquera están en función de los ingresos totales sostenibles (ITS) y los costos totales (CT):
π = ITS - CT (2.18)
los cuales pueden expresarse de la siguiente forma:
π = pC - cf (2.19)
siendo p el precio (constante) de la especie capturada y c los costos (constantes) por unidad de esfuerzo. Estos últimos incluyen costos fijos, variables y de oportunidad, tanto de capital como de mano de obra. Los costos fijos son independientes de que la flota opere o no (depreciación, intereses, gastos administrativos, etc.), mientras que los variables sólo se producen cuando el pescador sale a pescar (combustible, carnada, alimentos, etc.). Los costos de oportunidad se definen en base a la siguiente mejor alternativa de inversión del capital o bien de ocupación de la mano de obra. Esto implica que los ingresos que hubieran podido ser generados en la siguiente mejor actividad alternativa deben ser incluídos dentro de los costos. El dueño de una embarcación desea obtener el mayor beneficio como resultado de cualquier tipo de inversión, y por tanto, puede decidir entre invertir en la pesca, en el banco o en otro proyecto que sea la siguiente mejor alternativa. Lo mismo sucede con respecto a su mano de obra, ya que podría dedicarla a otra actividad productiva. Así, esperará que al invertir en la pesca ésta le reditúe al menos lo que pudo haber ganado en su siguiente mejor opción de inversión de capital o de mano de obra. Ahora bien, si se sustituye la función correspondiente a la tasa de captura (2.2) en (2.19), los ingresos netos en función del esfuerzo serán definidos como:
( )π = pqB - c f (2.20)
Al igual que en el modelo biológico, Gordon (1954) supone condiciones de equilibrio en la población para obtener un rendimiento sostenible en el largo plazo. Considera asimismo que en equilibrio económico los ITS igualan a los CT y por tanto π(t) es igual a cero, no existiendo estímulo para entrar o salir de la pesquería. Si adicionalmente la biomasa de la expresión (2.20) se asume en equilibrio biológico para el nivel de esfuerzo correspondiente (la derivada de la biomasa con respecto al tiempo es igual a cero), entonces se presentan condiciones de equilibrio biológico y económico simultáneamente, razón por la cual se le denomina equilibrio bioeconómico. La biomasa en equilibrio bioeconómico (BEBE) se define igualando a cero la ecuación (2.20) y despejando B:
B cqpEBE = (2.21)
Bajo estas condiciones la biomasa será siempre mayor a cero dado que el esfuerzo se reducirá o cesará cuando los costos totales superen a los ingresos totales. En consecuencia, el

17
modelo pronostica: (1) sobreexplotación si la curva de costos totales intersecta a la de ingresos totales sostenibles con niveles de esfuerzo pesquero superiores a los requeridos para operar en MRS; y (2) no extinción del recurso, ya que antes que éste se agote deja de ser rentable ejercer esfuerzo sobre la especie en cuestión. La condición de no extinción dependerá del tipo de crecimiento asumido para el recurso y la forma que adquiera la función de captura definida en 2.2 (ver Clark, 1985; Anderson, 1986). Asimismo, debe precisarse que la predicción de no extinción será correcta si y sólo si la biomasa resultante en equilibrio bioeconómico excede el nivel de biomasa umbral requerido para darle viabilidad de existencia a la población. Los ingresos totales sostenibles (bajo condiciones de equilibrio) se obtienen al multiplicar la función de biomasa (2.4) por el precio p:
ITS = prB 1 -BK
⎛⎝⎜
⎞⎠⎟ (2.22)
La curva de ingresos totales sostenibles en función del esfuerzo tendrá la misma forma que la de rendimiento sostenible de Schaefer (Fig. 2.1) pero en términos monetarios. Los costos totales se obtienen partiendo de la misma función que en el caso anterior, pero en términos del esfuerzo pesquero (f):
qfB = rB 1-BK
⎛⎝⎜
⎞⎠⎟ (2.23)
Despejando f y multiplicando por el costo unitario (c) se definen los costos totales en función del esfuerzo en el largo plazo:
[ ]CT =
cr B Kq
1−⎛
⎝⎜
⎞
⎠⎟
( / ) (2.24)
En las Figuras 2.3a y b se presentan los niveles de biomasa en equilibrio y rendimiento sostenible, que resultarían de los niveles de esfuerzo en máximo rendimiento económico (fMRE), en máximo rendimiento sostenible (fMRS) y equilibrio bioeconómico (fEBE). La expresión gráfica del modelo de Gordon en términos de ITS y CT se presenta en la Figura 2.3c. En ausencia de regulación, el aumento del esfuerzo ocasiona un incremento en los ingresos hasta alcanzar el nivel de equilibrio bioeconómico, que corresponde a un nivel de esfuerzo (fEBE) donde los costos totales igualan a los ingresos totales sostenibles, en este caso es igual el esfuerzo requerido para alcanzar el MRS. El equilibrio económico indica que no existe incentivo para entrar o salir de la pesquería, y el biológico, que la población se mantiene en el tiempo. El máximo rendimiento económico (MRE) se alcanza en fMRE donde las diferencias entre los CT y ITS se maximizan. El área bajo la curva de ITS por encima de CT corresponde a los beneficios netos o renta económica, los cuales siempre serán mayores hacia la izquierda del MRS. En este sentido, la posición de la curva de costos es muy importante, ya que si éstos se reducen existirán ganancias (aunque con menores rendimientos) y por tanto habrá estímulos

18
para incrementar el esfuerzo hasta alcanzar un nuevo nivel de equilibrio, ateniéndose también un nuevo nivel de esfuerzo para obtener el máximo rendimiento económico. Se asume que el esfuerzo extra es producido por la operación de botes nuevos que operan de una manera eficiente, más que por la expansión del esfuerzo de los ya existentes (Anderson, 1986). Gordon (1954) pronostica como se demostrará posteriormente, que el esfuerzo en el equilibrio bioeconómico será el doble del ejercido al nivel de máximo rendimiento económico.
500
1000
1500
2000
2500
3000
3500
Biom
asa
Sost
enib
le ('
000
ton)
0 200 400 600 800 Esfuerzo (barcos)
a
B
B
B
ff
MRE
MRS
EBE
EBEf MRE MRS
0 50
100 150 200 250 300 350 400
Cap
tura
sos
teni
ble
('000
ton)
0 200 400 600 800 Esfuerzo (barcos)
f f f
MRSC CMRE EBE
MRE MRS EBE
b
0
5000
10000
15000
20000
25000
Cos
tos
e in
gres
os to
tale
s ('0
00 $
)
0 200 400 600 800 Esfuerzo (barcos)
CT
MRE
MRS
ITS
f MRE f fMRS EBE
c
Rendimiento promedio y marginal El análisis presentado anteriormente muestra a la pesquería como un todo. Tanto los rendimientos promedio (Rp) como los rendimientos marginales (Rm), que corresponden a la aplicación de una unidad adicional de esfuerzo, serán sostenibles por unidad de tiempo. Los primeros caerán continuamente conforme aumenta el esfuerzo hasta llegar a cero (Fig. 2.4a), es decir, cuando se agota el recurso. El Rm decrece igualmente hasta llegar a cero en el nivel de MRS, después del cual se hace negativo (Fig. 2.4a). Esto es, el rendimiento obtenido por cada unidad adicional de esfuerzo será progresivamente inferior al anterior. Esto resulta como consecuencia de que el pescador individual considera exclusivamente sus rendimientos, sin tomar en cuenta el efecto que el aumento en su esfuerzo pesquero tiene sobre otros usuarios, y por tanto opera siempre en términos de sus rendimientos promedio.
Figura 2.3 Curvas de biomasa sostenible (a), captura sostenible (b), e ingresos totales sostenibles (ITS) y costos totales (CT) (c) del modelo bioeconómico estático de Gordon-Schaefer.
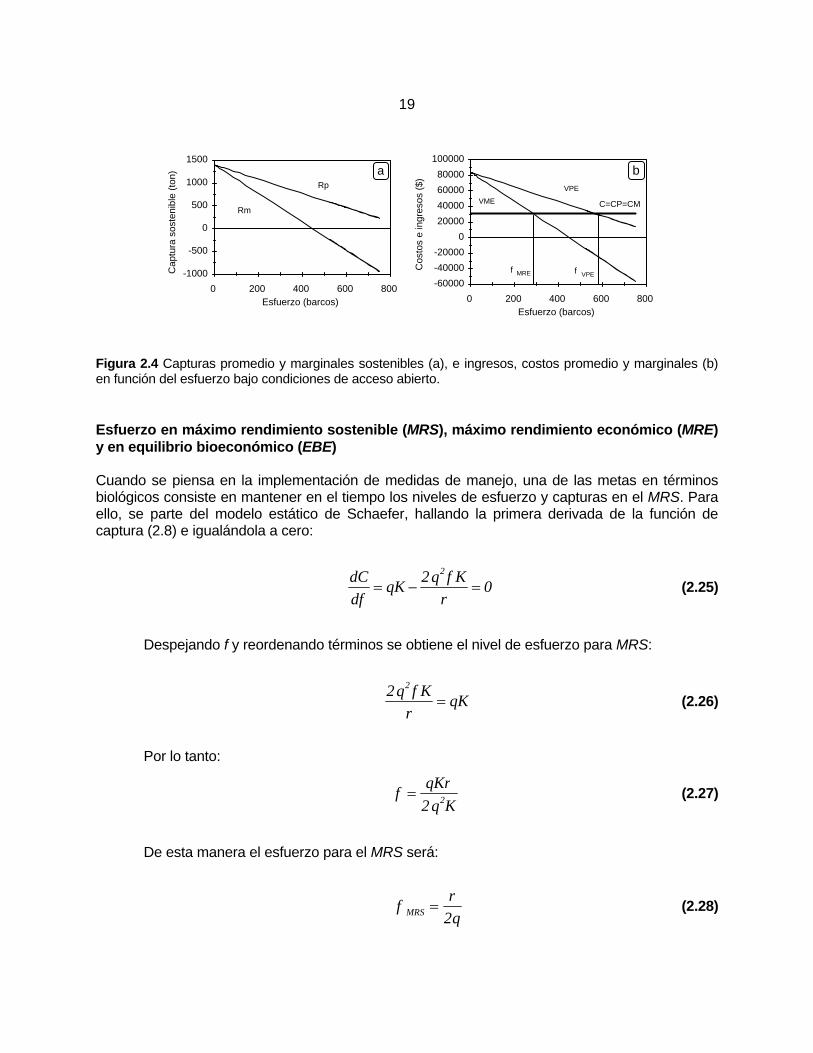
19
-1000
-500
0
500
1000
1500 C
aptu
ra s
oste
nibl
e (to
n)
0 200 400 600 800 Esfuerzo (barcos)
Rm
Rpa
-60000 -40000 -20000
0 20000 40000 60000 80000
100000
Cos
tos
e in
gres
os ($
)
0 200 400 600 800 Esfuerzo (barcos)
VME
f
C=CP=CM
b
f VPEMRE
VPE
Figura 2.4 Capturas promedio y marginales sostenibles (a), e ingresos, costos promedio y marginales (b) en función del esfuerzo bajo condiciones de acceso abierto. Esfuerzo en máximo rendimiento sostenible (MRS), máximo rendimiento económico (MRE) y en equilibrio bioeconómico (EBE) Cuando se piensa en la implementación de medidas de manejo, una de las metas en términos biológicos consiste en mantener en el tiempo los niveles de esfuerzo y capturas en el MRS. Para ello, se parte del modelo estático de Schaefer, hallando la primera derivada de la función de captura (2.8) e igualándola a cero:
dCdf
qK 2q f Kr
02
= − = (2.25)
Despejando f y reordenando términos se obtiene el nivel de esfuerzo para MRS:
2q f Kr
qK2
= (2.26)
Por lo tanto:
f qKr2q K2= (2.27)
De esta manera el esfuerzo para el MRS será:
MRSf r2q
= (2.28)

20
El MRS se obtiene al sustituir (2.28) en (2.8):
C qKr2q
q Kr4q r
2 2
2= − (2.29)
de tal modo que:
C = Kr
2- Kr
4 y en consecuencia la captura en MRS será:
MRSCKr4
= (2.30)
Para maximizar la renta económica que el recurso le podría generar a la sociedad, es necesario que la pesquería opere con un nivel de esfuerzo pesquero que maximice el rendimiento económico (e.g. esfuerzo en MRE). Este criterio se obtendrá al maximizarse las diferencias entre los ingresos totales sostenibles y los costos totales del esfuerzo pesquero. En este caso, se obtiene cuando el valor del producto marginal del esfuerzo pesquero (VME) es igual a los costos por unidad de esfuerzo, e.g. VME=c. Considerando la tasa de captura expresada en la ecuación (2.2), se sustituye la expresión que determina la biomasa en función del esfuerzo pesquero:
1
Kr
qf-1=B(t) ⎟⎠⎞
⎜⎝⎛
por lo tanto:
C(t) =qfK 1-qfr
⎛⎝⎜
⎞⎠⎟ (2.31)
Multiplicando (2.31) por el precio promedio de la especie objetivo y dividiendo entre el esfuerzo, se obtiene el valor promedio del esfuerzo pesquero (VPE):
VPE =qKp 1 -qfr
⎛⎝⎜
⎞⎠⎟ (2.32)

21
Derivando la ecuación de captura de Schaefer (2.8) con respecto al esfuerzo pesquero se tiene que:
dCdf
= qK 1-qfr
⎛⎝⎜
⎞⎠⎟
Multiplicando la ecuación anterior por el precio promedio de la especie (p), se obtiene el valor del producto marginal del esfuerzo pesquero (VME):
VME = pqK 1 -2qf
r⎛⎝⎜
⎞⎠⎟ (2.33)
Igualando la expresión (2.33) al costo unitario del esfuerzo pesquero, c, y despejando f, se obtiene el esfuerzo en máximo rendimiento económico (fMRE):
MREf =r
2q1
cpqK
−⎛
⎝⎜
⎞
⎠⎟ (2.34)
El equilibrio bioeconómico se alcanzará cuando el valor promedio del esfuerzo pesquero sea igual a los costos por unidad de esfuerzo (VPE = c). Este último se estima igualando la función (2.32) a los costos (c) y despejando f. Al hacer lo anterior se podrá observar que fEBE = 2fMRE , es decir que el esfuerzo en equilibrio bioeconómico es dos veces mayor que el esfuerzo en máximo rendimiento económico (Fig. 2.4b; ver también Fig. 2.3.c). Supuestos del modelo El modelo tiene implícitos varios supuestos, tales como: a) El precio es considerado constante. Asimismo, los costos marginales y promedio se asumen constantes para cualquier nivel de esfuerzo pesquero. b) Los cambios en el esfuerzo se darán de manera inmediata como respuesta a las variaciones en los ingresos netos. c) El recurso responderá inmediatamente a la reducción en el esfuerzo. d) Los costos totales crecen de manera proporcional al esfuerzo de tal manera que un cambio de pendiente en la curva de costos determinará un desplazamiento en el nivel en que se alcanza el equilibio bioeconómico y el MRE. e) Como resultado de su derivación con respecto al modelo de Schaefer, no considera cambios en la población dependientes de las características intrínsecas a su ciclo de vida, tales como longevidad, temporalidad del ciclo reproductivo y variabilidad en el reclutamiento (Lawson, 1984).

22
Entre las limitaciones de tipo logístico cabe mencionar que puede llevar años recopilar la información de datos biológicos requeridos y la colecta de las estadísticas de esfuerzo y rendimiento, y más aún los económicos. Además, en la mayoría de los casos la precisión de las estadísticas utilizadas es poco confiable, sobre todo en países en desarrollo donde los costos de información son muy altos. Clark (1985) menciona que es difícil distinguir si el efecto de disminución de una población se debe a presiones de pesca o a procesos naturales. En la práctica parece ser que la capacidad del esfuerzo para ciertas pesquerías es mucho más que dos veces el nivel óptimo; Clark (1985) concluye que el modelo sirve para explicar el problema de sobrepesca, pero parece inadecuado para proponer medidas de manejo.
Dinámica del esfuerzo pesquero
Smith (1969) propone un modelo donde asume que el esfuerzo pesquero cambia proporcionalmente a los ingresos netos:
dfdt
= ( t )ϕπ (2.35)
donde ϕ es una constante positiva que describe la dinámica de la flota en el largo plazo. Smith emplea esta ecuación para modelar los cambios en la flota de una pesquería, los cuales se obtienen de sustituir (2.20) en (2.35):
dfdt
[ pqB( t ) c ] f ( t )= −ϕ (2.36)
Los cambios en la biomasa serán dados por:
dBdt
rB(t) 1B(t)K
qB(t)f= −⎛⎝⎜
⎞⎠⎟− (2.37)
Si π(t) es mayor a cero, habrá un aumento en el esfuerzo resultante de la entrada de barcos a la pesquería, mientras que se producirá una disminución de éste en caso contrario. El parámetro ϕ puede estimarse empíricamente de acuerdo al comportamiento de la flota (entradas y salidas) en función de variaciones en los ingresos netos, que tendrán relación con los costos ejercidos para los correspondientes niveles de esfuerzo. El modelo de Smith aunque robusto, puede complementarse para incorporar un retraso entre la toma de decisión y la entrada o salida de embarcaciones. El capital tiene larga vida de trabajo y los factores institucionales, tales como sistema de pago compartido de grupos de pescadores, tienden a amortiguar el retorno del capital, de tal forma que las consecuencias del

23
período de ajuste del esfuerzo no se reflejan inmediatamente. El ingreso de nuevas embarcaciones a la pesquería tendrá un retraso si se considera el tiempo de obtención de permisos de pesca y de créditos y el tiempo requerido para la construcción de la embarcación. Los beneficios pueden obtenerse por varios períodos sin tener un cambio significativo sobre el esfuerzo. Seijo (1987) propone la adición de un término de retraso distribuido representado por la función de densidad probabilística Erlang (Manetsch, 1976), que incorpora el tiempo requerido para la salida o entrada de barcos a la pesquería, una vez que se manifiesta el efecto de cambios en el esfuerzo dados los ingresos netos anuales. El modelo incluye un parámetro que sirve para retrasar el tiempo de ocurrencia de un fenómeno (DEL), en este caso entrada y salida de barcos, el cual se define como tiempo de tránsito promedio y variará de acuerdo a la categoría de las embarcaciones. En el Capítulo 5 se describe la aplicación del modelo de retraso distribuido a pesquerías marinas. Las decisiones de dinámica del esfuerzo pesquero en el corto plazo no son expresadas en el modelo de Smith, por lo que serán introducidas, discutidas y modeladas en capítulos posteriores. Ejemplo 2.1 Modelo bioeconómico dinámico Dada una pesquería de una especie pelágica cuyos parámetros aparecen en la Tabla 2.1, se analiza el comportamiento dinámico de la biomasa de la población, los rendimientos, la flota, los ingresos y costos. Tabla 2.1 Parámetros para el modelo bioeconómico dinámico (Gordon-Schaefer).
Parámetro/Variable Valor Unidad de medida Tasa intrínseca de crecimiento 0.36 -- Coeficiente de capturabilidad 0.0004 -- Capacidad de carga del ecosistema 3500000 ton Precio de la especie 60 US$/año Costo unitario del esfuerzo 30000 US$/año Biomasa inicial de la población 3500000 ton Parámetro de dinámica de la flota 0.000005 --
Aplicando el modelo dinámico Gordon-Schaefer con los parámetros bioeconómicos de la Tabla 2.1 se puede observar la trayectoria dinámica de la biomasa, el rendimiento y los ingresos y costos totales (Fig. 2.5).

24
500
1000
1500
2000
2500
3000
3500
Biom
asa
('000
ton)
0 200 400 600 800 Esfuerzo (barcos)
aBiomasaDinámica
BiomasaEstática
0
100
200
300
400
500
Cap
tura
('00
0 to
n)
0 200 400 600 800 Esfuerzo (barcos)
CapturaDinámica
CapturaSostenida
b
0
5000
10000
15000
20000
25000
30000
Cos
tos
e in
gres
os to
tale
s ('0
00 $
)
0 200 400 600 800 Esfuerzo (barcos)
IngresoDinámico Ingreso
Sostenible
c
CostosTotales
Figura 2.5 Trayectoria estática (equilibrio) y dinámica de la biomasa (a), las capturas (b) y los costos e ingresos (c) resultantes de la aplicación de diferentes niveles de esfuerzo. En esta figura se incluyen también las curvas en equilibrio para las variables de desempeño antes mencionadas para observar la diferencia cuantitativa de considerar el efecto de diferentes niveles de esfuerzo utilizando el modelo estático y el dinámico. El modelo muestra el comportamiento dinámico de las diferentes variables donde se tiende al equilibrio en todos los casos, el cual se alcanza con 578 barcos. El esfuerzo en máximo rendimiento económico (fMRE) es de 289 embarcaciones. En la Figura 2.6 se presenta la evolución en el tiempo de las variables de desempeño bioeconómico en el modelo dinámico. En la Figura 2.6a se puede observar la dinámica de la biomasa como respuesta a la dinámica del esfuerzo. En la Figura 2.6b se presenta el rendimiento en el tiempo que obtiene una flota que va modificando su esfuerzo pesquero en la medida que la renta obtenida en el tiempo varía. En la Figura 2.6c pueden observarse las tendencias crecientes, decrecientes y estabilizantes del esfuerzo de pesca en la medida que la renta crece, decrece, se hace negativa y posteriormente tiende a cero, convergiendo a nivel de esfuerzo en el equilibrio bioeconómico.

25
500
1000
1500
2000
2500
3000
3500 Bi
omas
a (to
n)
0 20 40 60 80 Tiempo (años)
a
0
100
200
300
400
500
Cap
tura
(ton
)
0 20 40 60 80 Tiempo (años)
b
-5000
0
5000
10000
15000
Ren
ta e
conó
mic
a (U
S$)
0 20 40 60 80 Tiempo (años)
c
0
200
400
600
800
Esf
uerz
o (b
arco
s)
0 20 40 60 80 Tiempo (años)
d
Figura 2.6 Dinámica de la biomasa (a), la captura (b), la renta económica (c) y el esfuerzo pesquero (d).
Como se puede observar, los incrementos en el esfuerzo dan lugar a aumentos en las capturas y por tanto en los ingresos generados por la pesquería. Como consecuencia, la CPUE decrece de manera paulatina si el esfuerzo continúa creciendo, lo que indicaría disminución de la biomasa de la población. Si el esfuerzo pesquero sigue aumentando a un nivel mayor de 630 embarcaciones, se observa que las capturas y los ingresos netos disminuyen. Posteriormente se entra a una dinámica de entrada y salida de embarcaciones a la pesquería en la medida que la renta se hace positiva o negativa, hasta un nivel de esfuerzo en el que la renta tiende a cero, en condiciones de equilibrio bioeconómico. Analizando la información en el tiempo se observa que a los 33 años se alcanzan los mayores niveles de esfuerzo pesquero, lo que da lugar a una caída brusca de la biomasa en los años siguientes, repercutiendo en los ingresos netos. Al no obtener ganancias, no hay ingreso de embarcaciones, lo que permite a la población recuperarse y tiende a estabilizarse cerca de las 1200 ton después de 50 años de operación de la pesquería, al igual que el esfuerzo pesquero. En este punto los ingresos netos son iguales a cero, ya que los costos totales son iguales a los ingresos totales.

26
Modelos de producción excedente usando mortalidad Los modelos no lineales "captura-mortalidad" se basan en el análisis conjunto de series de datos correspondientes a la captura comercial (variable dependiente) en función de la tasa instantánea de mortalidad total Z (variable independiente) (Csirke & Caddy, 1983). La creciente adquisición de series de tiempo prolongadas, tanto de capturas como de valores de Z, provee un atrayente campo para la aplicación de estos modelos "output-output" (tanto la captura como la mortalidad total son salidas de los subsistemas biológico y económico), lo cual ha suscitado recientes publicaciones destinadas a realizar una revisión de la veracidad de diferentes tipos de ajuste a los modelos captura-mortalidad (Caddy & Defeo, 1996). Otro motivo fundamental es que dichas técnicas son adecuadas para formular estrategias de manejo aversas al riesgo y en consecuencia para adoptar medidas precautorias de administración de recursos pesqueros (ver Capítulo 8 y también FAO, 1993). En esta sección se describen los modelos "captura-mortalidad", tanto en base a las aproximaciones clásicas de Schaefer (1954) como de Fox (1970). La mayor parte está tomada de los clásicos artículos de Caddy & Csirke (1983) y Csirke & Caddy (1983), así como del trabajo realizado por Caddy & Defeo (1996), quienes extendieron la teoría desarrollada en los anteriores trabajos al uso del modelo exponencial, adicionando asimismo un análisis de la variabilidad, riesgo e incertidumbre en la estimación de parámetros y en la determinación de medidas precautorias de manejo (ver Capítulo 8 al respecto). Teoría y aproximaciones al ajuste de los modelos
Obtención de valores de mortalidad. Los valores anuales de Zt (mortalidad total en el año t) pueden ser obtenidos de la fracción descendente de una curva de captura basada en edades (Robson & Chapman, 1961) o longitudes (Sparre et al., 1989). También puede ser obtenida de tallas medias (e.g. Beverton & Holt, 1956) o bien usando métodos tales como el de Heinke ó Robson & Chapman (ver Ricker, 1975). Los análisis convencionales del Análisis de la Población Virtual/Análisis de Cohortes (Pope, 1972; Jones, 1981) también pueden ser utilizados para estimar valores de mortalidad por pesca F por edad, los cuales podrían ser transformados en Z al adicionar una estimación de mortalidad natural M. Es importante considerar que el análisis de cohortes opera en base a las "diagonales" que siguen la historia de una cohorte dada a través del tiempo; es posible sin embargo considerar las series horizontales de capturas (e.g. la composición por edades de las capturas en un año dado) en forma independiente a fin de generar curvas de captura que provean de valores de Z a ser usados como entradas a los modelos "captura-mortalidad". Un simple promedio de estimaciones independientes de Zt basadas en el cociente de los logaritmos de las abundancias de sucesivas clases de edad que forman una importante proporción de la captura anual, puede ser obtenido desde la edad de primera captura en adelante. Una discusión de las diferentes aproximaciones para estimar mortalidad puede ser encontrada en los textos clásicos de Ricker (1958), Pauly (1980), Jones (1981), Caddy (1986) y Sparre et al. (1989).

27
Modelo logístico a) Ajuste directo a través de regresión múltiple. Tal como fue expresado en la ecuación (2.4), el modelo logístico de crecimiento poblacional propuesto por Graham (1935) en condiciones de explotación pesquera (ver Ricker, 1975) está dado por:
C rB( K -B)K
= (2.38)
Csirke & Caddy (1983) expresaron este rendimiento en equilibrio en términos del valor en equilibrio de la tasa de mortalidad anual (ver p. 45 y también Caddy, 1986), y redujeron la ecuación (2.38) a una forma cuadrática:
t t2
tC = aZ +bZ + c (2.39)
donde: Ct es la captura correspondiente al año t.
Zt es el coeficiente promedio de mortalidad total para el año t.
Bajo los supuestos del modelo logístico, la ecuación (2.39) da una parábola que intercepta la abscisa a la derecha del origen (Fig. 2.7). De esta forma, usando regresión múltiple, donde Zt y Zt² son las dos variables independientes de este polinomio de segundo orden, podemos estimar la curva que relaciona los valores anuales de captura con la mortalidad total. Dicho ajuste provee una estimación del coeficiente de mortalidad natural M a través de la solución de esta ecuación para Z = M (Ct y F = 0) de la siguiente manera (Csirke & Caddy, 1983):
M =-b+ ( b - 4ac)
2a
2
(2.40)
0
8000
16000
24000
32000
40000
Cap
tura
(ton
)
0 0.2 0.4 0.6 0.8 1 1.2 1.4 Zi (1/año)
70 69
71
68
64
56
57
60
58
67
66
76
74 73 72
75
LOGISTICOFOX
5955
6265
Figura 2.7 Ejemplo ilustrativo del modelo captura-mortalidad de Csirke & Caddy (1983).

28
Asimismo, los parámetros del modelo logístico pueden obtenerse para aquellos valores de Z correspondientes a MRS, el cual ocurre cuando (Csirke & Caddy, 1983):
dCdZ
= ddZ
( aZ +bZ + c ) (2.41)
Entonces:
dCdZ
= b+ 2aZ = 0 (2.42)
por lo cual:
MRSZ = - b2a
(2.43)
FMRS, definido como el nivel de mortalidad por pesca en MRS, está dado por la substracción de
M a ZMRS :
MRS MRSF = Z - M (2.44)
Los otros parámetros del modelo logístico están dados por:
r FMRS= 2 (2.45)
MRS c ba
=2
4 (2.46)
K MRSr
=4
(2.47)
b) Aproximación lineal. Csirke & Caddy (1983) han sugerido una aproximación alternativa para el ajuste del modelo logístico, basada en el índice de abundancia:
U = C( Z - M )t
t
t (2.48)
el cual es analizado en relación a (Zt - M), de tal manera que:
t
tt
CZ - M
= a -b( Z - M ) (2.49)

29
La ecuación (2.49) ha sido preferida al ajuste anterior debido a objeciones teóricas al ajuste directo del modelo (Hoenig & Hoenig, 1986), el cual en algunos casos tiende a subestimar M. Este modelo requiere de una aproximación especial, ya que se emplean en forma secuencial diferentes valores semilla de M en la ecuación (2.49), de los cuales se selecciona aquél que provea el valor más alto de bondad de ajuste (e.g. coeficiente de determinación R²). Dicho valor puede ser usado para transformar las tasas anuales de mortalidad total Zt en tasas de mortalidad por pesca Ft. Es muy importante hacer notar que en este caso no se requieren de valores de esfuerzo o CPUE. Los parámetros de esta versión lineal del modelo logístico pueden ser obtenidos como
sigue:
∞B = a (2.50) 2
r = Bb∞ (2.51)
3
MRSF = r2
(2.52)
4
MRS MRSZ = F + M (2.53) 5 (
FB
=MRS MRS2∞ (2.54)
Modelo exponencial Caddy & Defeo (1996) extendieron la teoría de los modelos de producción excedente basados en mortalidad para dar cabida a la versión exponencial del modelo logístico. a) Aproximación no lineal. La versión exponencial del modelo logístico para datos de captura y esfuerzo (Fox, 1970) puede ser expresada como:
t t-bfC = f B e t
∞ (2.55)
donde el rendimiento anual Ct y el esfuerzo ft están relacionados en una forma exponencial, y U∞ es la tasa de captura correspondiente al stock virgen. A efectos de transformar el esfuerzo en tasas de mortalidad analíticas, si consideramos que q y M son constantes, se tiene que:
tt b( Z M )
qC =( Z M )q
U et−
∞−
− (2.56)

30
Definiendo b' =b/q y considerando que U∞/q = B∞, entonces:
t tb ( Z M )C ( Z M )B e t= − ∞
− ′ − (2.57)
Esto puede ser expresado como:
CZ - M
= B et
t
-b ( Z - M )t∞
′ (2.58)
donde B∞ y b' pueden ser estimados por técnicas de regresión no lineal, usando procedimientos de minimización de errores (e.g. quasi-Newton, Marquardt) para la estimación de parámetros. Como en el caso de la aproximación logística alternativa, el modelo es ajustado para diferentes valores semilla de M, seleccionando aquél que maximice cierto criterio de bondad de ajuste; una vez obtenido el mejor valor de M, los parámetros de manejo restantes pueden ser calculados diferenciando la ecuación (2.60) con respecto a F y llevándola a cero al nivel de MRS:
tt
-b F b FdCdF
= -b F B e + B e = 0MRS MRS′ ∞′
∞′ (2.59)
por tanto:
MRSF = - 1b′
(2.60)
De la ecuación (2.58) tenemos que:
U = U et-b ( Z - M )t
∞′ (2.61)
Sustituyendo esos valores en la ecuación (2.57) obtenemos:
MRS = F B e = Be
MRSb F
bMRS
∞′ ∞
′ (2.62)
y ZMRS se logra en función de la suma de FMRS a M.
b) Aproximación lineal. Una aproximación alternativa a la anterior puede lograrse al tomar los logaritmos naturales de ambos miembros de la igualdad y ajustando una regresión lineal de la forma Yi = a - b Xi:

31
ln (ln )t
tt
CZ -M
= B +b M -b Z⎛⎝⎜
⎞⎠⎟ ′ ′∞ (2.63)
usando a ln(Ct /Zt - M) y Zt como variables dependiente e independiente, respectivamente. La estimación de parámetros sigue el mismo razonamiento que en la aproximación previa, usando diferentes valores semilla de M, y difiriendo en la estimación de la biomasa virgen, la cual puede ser deducida de la ecuación (2.63) como:
B = e ( a -b M )∞
′ (2.64)
Ventajas y limitaciones de la aplicación de los modelos captura-mortalidad
Ventajas. Algunas de las conclusiones emanadas de la aplicación de estos modelos de "captura-mortalidad" sugieren lo siguiente (ver también Caddy & Defeo, 1996): 1) La pobre calibración de los errores en la estimación del esfuerzo y los sesgos que esto conlleva en el uso de esta variable en los modelos captura-esfuerzo tradicionales, proporciona ventajas comparativas a los modelos captura-mortalidad. 2) Los cambios en el coeficiente de capturabilidad q con la intensidad de pesca y biomasa pueden ser reducidos empleando esta aproximación. 3) Caddy & Defeo (1996) demostraron la utilidad de extender la teoría desarrollada por Csirke & Caddy (1983) al uso del modelo exponencial. Una de las principales críticas de los modelos logísticos basados en el uso del esfuerzo como variable de entrada ha sido que, particularmente luego de alguna aproximación al equilibrio de algún tipo, la relación tasa de captura vs esfuerzo no es siempre lineal, sino que tiende a caer a una tasa progresivamente decreciente, haciendo la aplicación de los modelos exponenciales más adecuada que aquella basada en el logístico. Esta crítica es menos válida cuando se usan tasas de mortalidad para ajustar modelos de producción (Csirke & Caddy, 1983), debido a que la experiencia sugiere que se obtienen curvas de rendimiento más simétricas al graficar las capturas contra la tasa de mortalidad total. La conclusión a la cual arriban Caddy & Defeo (1996) es que ambos tipos de modelo sean considerados con precaución, y a efectos de manejo, sea utilizado en mayor medida aquél que provea la aproximación más cauta, ésta evaluada en función del simple análisis de riesgo propuesto por dichos autores. En el Capítulo 8 se analizará con mayor detalle aspectos relacionados con la estimación en la variabilidad de los parámetros y del análisis de riesgo a efectos de manejo. 4) Estos modelos, en su versión estática, asumen equilibrio. Los valores medios de tasas de mortalidad Zt obtenidos de métodos basados en curvas de captura u otros (ver Caddy & Defeo, 1996) representan el impacto de la pesca en el presente y pasado reciente en forma más cercana que las aproximaciones que emplean valores anuales de esfuerzo de pesca (o bien de Zt basados en el cociente de la abundancia de una clase de edad simple en dos sucesivos períodos de tiempo). Por tanto, es posible compensar desviaciones de mortalidad anual con respecto al equilibrio, tal como el análogo a los promedios móviles (“past averaging”) de esfuerzo de pesca

32
para dar cuenta de los efectos de mortalidad en años previos (Gulland, 1961; Fox, 1975). Esta aproximación puede resultar bastante conservadora en caso de usar una prolongada serie de datos en el número de datos consecutivos a promediar. El problema se agrava en caso de usar modelos basados en esfuerzo (no en el caso de mortalidad), en los cuales es difícil decidir cuantos años deberían ser incluidos; en este caso variaría dependiendo de los cambios en la intensidad de pesca con el tiempo. En caso de considerar que el promedio móvil de estimaciones de mortalidad es apropiado, entonces los modelos "captura-mortalidad" tienen una ventaja definida (Caddy, 1986): el recíproco de la mortalidad promedio para las clases de edad plenamente reclutadas es en sí un estimador simple del número de clases de edad anuales que constituyen la captura. Entonces, es una medida del tiempo en el cual la aproximación de promedios móviles hacia atrás (“past mortality averaging'“) debería ser aplicado. Por tanto, 1/Zt podría ser considerado como un índice del número de grupos de edad plenamente reclutados que sobreviven de la explotación en años anteriores y que de una u otra forma constituyen la captura actual. Lo importante de este razonamiento radica en que cuando existen estimaciones de mortalidad para varios grupos de edad, hay un cierto grado de ajuste semiautomático de la mortalidad pasada en cada estimación individual anual de mortalidad (Caddy, 1986). 5) Las estimaciones de parámetros sin dar cuenta de ajustes al equilibrio son de todas maneras valiosas. Idealmente, a menos que existan variaciones anuales considerables en el esfuerzo de pesca, las series de captura-mortalidad tienden a mostrar cierto grado de autocorrelación serial sin las fuertes oscilaciones desde la izquierda a la derecha de las curvas de rendimiento características de muchos modelos de producción captura-esfuerzo, y por tanto con amplias desviaciones del equilibrio. Desventajas. Algunas de las principales limitaciones a la aplicación de este tipo de modelos consisten en: 1) La estructura de la base de datos debe abarcar un amplio rango de valores de tasas de mortalidad. Las estimaciones de los parámetros son en fuerte grado dependientes de los valores extremos, en particular de dicha variable independiente. La interpretación y evaluación de los efectos de estos puntos con criterios de manejo debe ser cautelosamente analizada, ya que los valores de Z situados en los extremos de las curvas de producción influyen en el comportamiento de los modelos y pueden ser responsables de amplios intervalos de confianza (Caddy & Defeo, 1996). Paradójicamente, dichos puntos proveen la información más importante desde un punto de vista biológico-pesquero, y contribuyen significativamente a la explicación de la variabilidad observada en las bases de datos. Caddy & Defeo (1996) mostraron que, al omitirse estos datos por un procedimiento de remuestreo, se obtuvieron valores de los parámetros que influenciaron fuertemente la estimación de sus medias e intervalos de confianza (ver también Capítulo 8). 2) Tal como se mencionó anteriormente, la versión analizada de estos modelos es estática y asume equilibrio. Uno de los tópicos que ha sido objeto de reciente debate (Hilborn & Walters, 1992, Polacheck et al., 1993, Caddy & Defeo, 1996) es la dicotomía entre modelos dinámicos vs estáticos, los cuales asumen equilibrio para la estimación de parámetros. Si bien la teoría de modelos de producción con tasas de mortalidad está mucho menos desarrollada que aquella basada en el esfuerzo de pesca, existe la posibilidad de obtener una versión dinámica, en la medida en que permita disipar dudas sobre los desajustes producidos por la ausencia de equilibrio.

33
Aproximación bioeconómica La aproximación anterior constituye un enfoque netamente biológico. Con el propósito de obtener un punto de referencia bioeconómico en el proceso de manejo precautorio de pesquerías, a continuación se presenta una expresión que estima la renta generada por el recurso (π) a partir de la ecuación captura-mortalidad. Se toma como ejemplo de desarrollo el modelo linearizado de producción excedente de Fox (1970) adaptado a criterios captura-mortalidad:
π = −∞− ′pF B e
cFq
( b F ) (2.65)
donde: p es el precio promedio de la especie capturada. c es el costo unitario del esfuerzo pesquero. Diferenciando la ecuación anterior, se obtiene una expresión de la renta marginal (πM) con respecto a cambios en el coeficiente de mortalidad por pesca:
πM(F)d
dFpFB e c
Fq
b F= −⎛
⎝⎜
⎞
⎠⎟∞
′( ) (2.66)
Ejecutando la derivada anterior se obtiene que:
πM( F )= pB e pFB ecq
(b F ) (b F )∞
′∞
′− − (2.67)
Ahora bien, igualando a cero la renta marginal y resolviendo la ecuación con respecto a F, se obtiene una expresión que representa el valor de mortalidad por pesca que genera el máximo rendimiento económico (FMRE):
pB e pFB b ecq
( -b F ) ( b F )∞
′∞
− ′− ′ − = 0 (2.68)
MRE
1
F =-W
epqB
c +1
b∞
⎛
⎝⎜
⎞
⎠⎟
⎡
⎣⎢
⎤
⎦⎥
′ (2.69)
haciendo ( )ae
pB qc=
∞
1
, se tiene que W [ a ] es una función especial de MathCad ®1 que puede
1 Mathcad 5.0 for Windows. 1994. Mathsoft, Inc.

34
expresarse de la forma siguiente:
( )a W W= exp (2.70)
En base a lo anterior se obtiene la expresión para FMRE :
MREF = -W +1b′
(2.71)
La captura en máximo rendimiento económico CMRE se obtiene como sigue:
( )C F B eMRE MRE
b FMRE= ∞− ' (2.72)
Finalmente, el esfuerzo en máximo rendimiento económico fMRE puede ser expresado como:
MREMREf = Fq
(2.73)
Ejemplo ilustrativo. Dados los parámetros del modelo linearizado exponencial captura-mortalidad: p = $60 ; q = 0.001; b' = .85; c = $ 200000; y B∞ = 15.0×106 ton y después de estimar numéricamente el valor de W = 0.404, la mortalidad por pesca que genera un máximo rendimiento económico FMRE se obtendrá al sustituir estos valores en la ecuación 2.53 teniéndose que FMRE = 0.701/año y una captura en máximo rendimiento económico CMRE = 5.795×106 ton correspondiente a un esfuerzo en fMRE igual a 701 embarcaciones.
Análisis bioeconómico con modelos de estructura por edades de una población
Modelo de Beverton & Holt (1957) Los modelos de estructura por edades consideran los factores de crecimiento y mortalidad que afectan la biomasa de una población distribuída de manera homogénea en el espacio y en el tiempo. A diferencia de los modelos globales, los cuales consideran la biomasa total de la población, las aproximaciones derivadas de estos modelos se caracterizan por incluir de manera explícita la estructura por edades de la población y los procesos que la alteran, donde la edad está asociada a la talla, crecimiento individual, reclutamiento, fecundidad y vulnerabilidad a la pesca (Hilborn & Walters, 1992).

35
Estos modelos parten de los trabajos iniciales de Beverton & Holt (1957) quienes definen a la captura (C) como el producto de la mortalidad por pesca (F) por la biomasa de la población, la cual resulta del producto del peso individual (W) por el número de individuos presentes en ese momento de tiempo (N). La captura en número y peso resulta de integrar la abundancia de las clases de edad de aquellos individuos reclutados a la pesquería (tr) hasta la de máxima longevidad (tl):
C = F( )N( )W( )dr
l
t
t
∫ τ τ τ τ (2.74)
Considerando la extrema variabilidad en los parámetros poblacionales y en la mortalidad por pesca (ésta ocasionada por una distribución agregada del esfuerzo de pesca) en especies sedentarias, el clásico análisis de rendimiento por recluta ( Beverton & Holt, 1957) debería ser realizado en forma separada para cada edad identificada (j edades), cada una sujeta a diferente intensidad de pesca y talla de media captura (Caddy, 1975):
j j jC = (C / R ) ( R )∑ (2.75)
donde Cj es el rendimiento por edad, C/Rj es el rendimiento por recluta y Rj es el reclutamiento estimado por edad j. De esta manera los supuestos clásicos de los modelos de agrupamiento dinámico se harían efectivos. La extensión del modelo clásico a efectos de un análisis multiespecífico es igualmente viable (Ursin, 1982), aunque en caso de especies sedentarias debería considerarse en una primera instancia la heterogeneidad espacial mencionada y luego integrar los resultados. Un análisis bioeconómico multiespecífico es posible con la inclusión de los costos unitarios del esfuerzo de pesca (c) y el precio de la especie capturada (p) en la ecuación tradicional (Arreguín-Sánchez, 1989). Supuestos y limitaciones del modelo El modelo supone las siguientes condiciones: a) Reclutamiento a la pesquería en forma de "filo de navaja", al igual que la selección de edades al arte de pesca. Se supone la incorporación de los reclutas a la pesquería en un mismo momento. b) Todos los peces de una cohorte son capturados al mismo tiempo. c) Distribución homogénea de la población, y por tanto los individuos que la componen tienen la misma probabilidad de ser capturados. d) La tasa de explotación se asume constante durante toda la vida de los organismos. Los rendimientos son estables en el tiempo si el patrón de explotación se mantiene, lo que implica reclutamiento constante.

36
e) La mortalidad natural es constante independientemente de la edad, inclusive a partir de aquella que es reclutada a la pesquería; la mortalidad por pesca es igual a cero para edades menores a la de primera captura, por lo que las pérdidas en N(t) antes de la edad de primera captura (tc) serán:
N( t )=R[ -M( t - tr )] (2.76)
donde R es el número de peces vivos al tiempo t=tr (edad de reclutamiento a la población). Ahora bien, N(t) para todas las edades capturadas se define como:
N( t )= R e[ -( F+ M )( t -t ) ]c' (2.77) donde R’ indica el número de individuos reclutados a la pesquería.
f) Otra suposición implícita es que el crecimiento individual se ajusta a la función propuesta por von Bertalanffy (1938):
W t W [1 - e ]bkp t -t )0( ) (= ∞ − (2.78)
donde W∞ es el peso asintótico que alcanzan los individuos, kp es el parámetro de curvatura, t0 es el parámetro de ajuste de la ecuación y b es el exponente de la relación peso-longitud. g) El modelo tradicional asume que el reclutamiento es independiente del tamaño del stock, y que tampoco se ve afectado por variaciones en la intensidad de pesca, supuesto que ha sido considerado como el menos satisfactorio e irreal. Sin embargo, se ha tratado de incorporar al modelo las clásicas curvas de reclutamiento de Beverton & Holt (1957) y la de Ricker (1975). Considerando el modelo de reclutamiento de los primeros e incorporando los elementos anteriores en el modelo, la ecuación que define el modelo estático de rendimiento por recluta queda expresada de la siguiente forma:
CR Fe W U e
F + M + nkpM ( tc tr ) n
[ -nkp ( t - t ) ]c 0
= ∑− −∞ (2.79)
Muchas de las limitaciones del modelo han sido mencionadas en los propios supuestos que lo sustentan (e.g. mortalidad natural y por pesca constantes, independencia del reclutamiento con respecto al tamaño del stock parental). El considerar la mortalidad por pesca constante puede tener un serio impacto en el manejo de pesquerías, debido a que en general hay diferencias marcadas con la edad producidas por ejemplo por diferencias en su accesibilidad al arte de pesca. En tal sentido, se omiten los diferentes tipos de tecnología empleados por diferentes grupos de pescadores en casos de esfuerzo pesquero heterogéneo. La no incorporación de elementos económicos dentro del modelo constituye una limitante adicional a la hora de evaluar diferentes estrategias de manejo.

37
Modelo dinámico de estructura por edades A diferencia del modelo estático propuesto por Beverton & Holt (1957), el modelo dinámico toma en cuenta los influjos y eflujos para cada cohorte de la estructura de la población, aplicando los principales conceptos de tablas de vida (Begon et al., 1990). La incorporación de los reclutas es dinámica, permitiendo un manejo estacional del reclutamiento y su distribución (Seijo, 1986; Seijo & Defeo, 1994b). Los cambios en el número de individuos en la población de acuerdo al modelo dinámico se definen como:
jj j j j j j
dNdt
S t N t A t N t S t N t= − −− −1 1( ) ( ) ( ) ( ) ( ) ( ) (2.80)
donde Sj es la sobrevivencia de los organismos de clase de edad j y Aj corresponde a la tasa finita de mortalidad total (Gulland, 1983). Dicha tasa de sobrevivencia es expresada como sigue:
j-1 j 1 j-1S (t) 1 [ MN (t) FR (t)]= − +− (2.81)
donde MN(t) es la tasa de mortalidad natural y FR(t) es la tasa de explotación, ambos derivados de los correspondientes coeficientes de mortalidad natural y por pesca. Factorizando y ordenando términos:
jj j j
dNdt
S t N t N t= −− −1 1( ) ( ) ( ) (2.82)
Así, el número de individuos de cada cohorte (Nj), se obtiene integrando en el intervalo de t+DT los individuos de la edad j-1 que sobreviven y crecen dentro de la cohorte en el tiempo t, considerando las pérdidas dadas por la tasa de la mortalidad total de los individuos de la cohorte (Aj), así como la tasa a la cual los organismos de la cohorte que sobreviven (Sj) son incorporados en la cohorte j+1 en el tiempo t (Seijo & Defeo, 1994b).
Integrando numéricamente la ecuación (2.84) con el método de Euler, se obtiene la dinámica de la estructura de la población:
N (t + Dt) N (t)+ [( S ( ) N ( ) - N ( )]dj jt
t+Dt
j - j - j= ∫ 1 1τ τ τ τ (2.83)
j j j-1 j-1 jN ( t + DT )=N ( t )+ DT[ S N ( t ) - N ( t )] (2.84)

38
En este caso el modelo también emplea la función de crecimiento de von Bertalanffy para la estimación del peso promedio de los individuos, por lo que la biomasa para cada clase de edad será:
j j jB ( t + Dt ) N ( t + Dt )tW= (2.85)
de donde se deriva la biomasa total B(t) como resultado de la sumatoria de la biomasa de todas las clases de edad:
B( t )= B ( t + Dt )j
j∑ (2.86)
La captura Cj (t) y la mortalidad por pesca (Fj ) por clase de edad se definen en base a las ecuaciones 2.2 y 2.9 respectivamente, pero en este caso tanto la biomasa como q son discriminados por clase de edad.
jj
jjF ( t ) C ( t )
B ( t )q f t= = ( ) (2.87)
j j jC ( t ) q B ( t )f ( t )= (2.88)
Los ingresos totales IT se obtendrán al multiplicar el precio por edad de la especie (pj ) por la captura derivada de cada clase de edad:
IT( t ) C ( t )p
jj j= ∑ (2.89)
El precio puede variar por tallas, y por tanto puede ser introducido en la ecuación. Los costos totales CT(t) e ingresos netos π(t) se obtienen de acuerdo a las ecuaciones desarrolladas en ocasión del modelo bioeconómico Gordon-Schaefer. Ejemplo 2.2 Modelo bioeconómico de estructura por edades dinámico Dada una pesquería de arrastre hipotética cuyos parámetros aparecen en la Tabla 2.2, se analiza el comportamiento dinámico de la biomasa de la población, los rendimientos, la flota y los ingresos, considerando variaciones en la edad de primera captura y el esfuerzo de pesca.

39
Tabla 2.2 Parámetros para el modelo bioeconómico dinámico de estructura por edades.
Parámetro/Variable
Valor
Unidad de medida
Edad máxima de la especie 10 años Edad de primera madurez sexual 2 años Fecundidad promedio 5000 huevos Edad de primera captura 2 años Proporción por sexos 0.5 -- Coeficiente de mortalidad natural 0.2 1/año Parámetro de curvatura ec. von Bertalanffy 0.5 1/año Parámetro de ajuste ec. von Bertalanffy (t0) 0.0 -- Longitud asintótica 100 mm Peso asintótico 200 g Parámetros para la ecuación de selectividad L50%=20
L75%=30 mm mm
Area barrida por día 0.1 Km2 Area total de distribución del recurso 10 Km2 Máximo reclutamiento observado (Rmax) 20000000 organismos Precio de la especie 10000 $/ton Costo unitario del esfuerzo 75000 US$/barco/año Parámetro de dinámica de la flota 0.00005 --
Como se puede observar en la Figura 2.8, a medida que se reduce la edad de primera captura (tc) la pendiente inicial de la curva de biomasa es mayor y la captura se incrementa, cayendo posteriormente a niveles menores a los de la edad inmediata superior, estabilizándose en ambos casos después de los 45 años (Fig. 2.8a, b y c). Sin embargo, un aumento indiscriminado en la edad de primera captura pudiera no justificar la salida de embarcaciones a la pesquería, ya que el componente de la estructura de la población autorizada para ser capturada posiblemente no cubra los costos del esfuerzo de pesca. En este ejemplo, con edades mínimas de captura mayores a 4 años este comportamiento podría esperarse. Debe observarse que, con los parámetros bioeconómicos proporcionados en este ejemplo ilustrativo, las tasas máximas de captura y de renta se obtienen al incrementar la edad mínima de captura de 1 a 2 años. Desde luego, la biomasa resultante de una mayor edad mínima de captura tiene un comportamiento dinámico que indica mayores niveles de biomasa y menor fluctuación de la misma. Respecto a la dinámica del esfuerzo de pesca, éste tiende a incrementarse de manera proporcional a las rentas generadas por la pesquería ante los diferentes escenarios de edades mínimas de primera captura (Fig. 2.8d). En el largo plazo, y bajo condiciones de acceso abierto, con una edad mínima de captura de 4 años la pesquería es capaz de soportar más del doble del número de embarcaciones que con la edad mínima de captura de 1 año.

40
1000
2000
3000
4000
5000
6000
Bio
mas
a (to
n)
0 10 20 30 40 50 Tiempo
tc=1tc=2
tc=3
tc=4
a
0
100
200
300
400
500
Cap
tura
(ton
)
0 10 20 30 40 50 Tiempo
tc=1
tc=2
tc=3
tc=4
b
-300
-200
-100
0
100
200
300
400
500
Ren
ta ('
000
US
$)
0 10 20 30 40 50 Tiempo
tc=1
tc=2
tc=3
tc=4
c
0
50
100
150
200
250
Esf
uerz
o (b
arco
s)
0 10 20 30 40 50 Tiempo
tc=1
tc=2
tc=3
tc=4
d
Figura 2.8 Modelo bioeconómico de estructura por edades: efecto dinámico de diferentes edades mínimas de captura en la biomasa (a), la captura (b), la renta (c) y el esfuerzo de pesca (d). Ahora bien, graficando las variables de desempeño bioeconómico de la pesquería (biomasa, rendimiento y renta) con respecto a cambios dinámicos en el esfuerzo de pesca, se pueden observar las trayectorias dinámicas ante diferentes edades mínimas de captura. Con una edad mínima de 4 años, la biomasa decrece ante diferentes niveles de esfuerzo a una tasa menor que las obtenidas con edades mínimas de captura de 1, 2 y 3 años (Fig. 2.8a). Asimismo puede observarse que, dados los parámetros bioeconómicos especificados, el esfuerzo de pesca máximo que llega a tener la pesquería es de cerca de 200 embarcaciones, cantidad que tiende a disminuir ya que a partir de ese nivel de esfuerzo la renta se hace negativa (Fig. 2.8c). El número de embarcaciones resultantes con las diferentes edades mínimas de captura, en condiciones de equilibrio bioeconómico, es de 67, 90, 115 y 142 (Fig. 2.8d). Es decir, el operar con una edad mínima de captura relativamente grande (4 años) permite soportar un mayor número de embarcaciones y por consiguiente generar un mayor número de empleos. La captura obtenible con el nivel de esfuerzo en equilibrio bioeconómico se incrementa de 158 ton con una edad mínima de 1 año a 272 ton con una edad mínima de 4 años (Fig. 2.8c). Como puede observarse de la discusión previa, el modelo bioeconómico de estructura por edades permite explorar el impacto de diferentes edades mínimas de captura. Esta importante variable de control en pesquerías donde la selectividad por tallas es viable, es un instrumento de ordenación que los modelos globales del tipo Gordon-Schaefer no tienen la capacidad matemática de analizar.

41
41
CAPITULO 3
Interdependencias Ecológicas y Tecnológicas en Pesquerías
El marco teórico-analítico dirigido a proponer pautas para el manejo de recursos pesqueros ha sido tradicionalmente desarrollado, tanto en el aspecto biológico como en el económico, en base a modelos que consideran una sola especie. Se ha argumentado que las interacciones entre los componentes de una comunidad carecen de relevancia al evaluar el estado de un recurso y sugerir estrategias para su explotación, en particular cuando el esfuerzo pesquero es moderado y dirigido a la especie ecológicamente dominante. Sin embargo, en pesquerías de varias regiones del mundo se han detectado cambios agudos en la abundancia y composición relativa de las especies que constituyen una comunidad, generalmente asociados con una creciente intensidad de pesca (ver ejemplos en Mercer, 1982; May, 1984; Pauly & Murphy, 1982). Se han observado fenómenos de sobrepesca, en términos de una progresiva disminución de las especies más valiosas en el mercado, que en general corresponden a los eslabones más altos de la cadena trófica (Pauly, 1982; Bustamante & Castilla, 1987). Esto ha llevado a reconocer que en la práctica la explotación pesquera opera sobre un recurso multiespecífico (May et al., 1979), obtenido incidental o intencionalmente por una flota variable en sus características básicas, tales como tamaño, poder de pesca y tipo de arte empleado (Gulland & Garcia, 1984). En las últimas dos décadas, el cuerpo teórico - práctico dirigido a manejar los recursos pesqueros ha intentado incluir las interdependencias ecológicas (e.g. competencia, relación depredador-presa) y la estabilidad de la comunidad asociada, es decir, su respuesta a perturbaciones diferenciales en cada uno de sus componentes, así como las interdependencias tecnológicas que resultan de la actividad extractiva de unidades pesqueras con características tecnológicas disímiles operando sobre diferentes componentes de una población o bien distintas especies objetivo (FAO, 1978; Huppert, 1979; May, 1984 y referencias contenidas en éstos). No obstante tales esfuerzos, la transición desde el análisis y manejo de recursos uni a multiespecíficos requiere mucho más que la extensión de los modelos tradicionales. Se ha documentado que la adición de una especie a un modelo uniespecífico incrementa su complejidad (e.g. número de interacciones en un modelo compuesto por subsistemas multicompartimentalizados: Silvert, 1982). El elevado número de parámetros requeridos como entrada para diseñar un modelo multiespecífico, así como las diversas fuentes de error y variabilidad natural en las estimaciones -que frecuentemente inducen a que el modelo no sea validado- ha sido la razón por la cual la aproximación multiespecífica es raramente aplicada en la práctica o sus resultados son considerados poco creíbles (Larkin, 1982; Majkowski, 1982; Sugihara et al., 1984). Si bien, a pesar de las dificultades, los conceptos anteriores denotan un creciente interés en el plano biológico de las implicaciones de manejo debidas a la explotación de recursos multiespecíficos, se ha dado poca importancia a la teoría económica aplicada a esta problemática (sin embargo, ver Mitchell, 1982; Clark, 1976; 1985; Heen, 1989; Anderson, 1986; May et al., 1979). Una de las razones principales de esto es la complacencia científica en cuanto a la aplicabilidad de los conceptos económicos desarrollados por Gordon (1953; 1954) para

42
42
pesquerías uniespecíficas (ver Cap. 2) en un contexto multiespecífico, lo cual no siempre es válido (Mitchell, 1982; Ströbele & Wacker, 1991; Flaaten, 1991). En este Capítulo se discute la extensión del modelo bioeconómico clásico para recursos multiespecíficos y se analizan las implicaciones bioeconómicas de las interdependencias ecológicas y tecnológicas que se suscitan en pesquerías. Se discute la aplicabilidad del modelo para diferentes tipos de recursos (e.g. pelágicos y bentónicos) en función de su biología, dinámica y estrategias de explotación. Finalmente, se sugieren futuros tópicos claves de investigación en el área.
Conceptos generales Una interdependencia tecnológica ocurre cuando flotas de diferentes características, ya sea en su poder de pesca (e.g. flota artesanal y mecanizada) y/o tipos de artes empleados, operan sobre diferentes componentes de un stock (e.g. juveniles y adultos) o bien sobre distintas especies objetivo de un stock múltiple ("mixed stock": Clark, 1985) y en consecuencia afectan su magnitud en forma diferencial (Fletcher et al., 1988). También ocurre en el caso de una flota que captura especies co-ocurrentes en el espacio, dependientes o no ecológicamente. El concepto de flota es aquí utilizado en un sentido amplio, incluyendo la recolección manual de organismos en pesquerías litorales artesanales de subsistencia. Una interdependencia ecológica es definida cuando existe cierta relación entre dos stocks explotados (Anderson, 1975a, b; Mitchell, 1982) o bien entre diferentes componentes poblacionales de un mismo stock (e.g. reclutas y adultos). En base a ello se definen dos tipos de interacciones: competencia y depredación. La competencia se define en base a los efectos negativos que un organismo ejerce sobre otro al consumir o interferir en el acceso a un recurso cuya disponibilidad (oferta) está limitada (Keddy, 1989). Esta interacción lleva a la reducción en la sobrevivencia, crecimiento y/o reproducción de los competidores, en especial el jerárquicamente subordinado (Giller, 1984; Begon et al., 1990). Es necesario discernir entre competencia interespecífica e intraespecífica, debido a que las consecuencias de estos procesos desde un punto de vista ecológico pueden ser muy diferentes. Los mecanismos por los cuales los competidores interactúan distinguen a la competencia por interferencia y por explotación (Schoener, 1983). Por medio de la dominancia competitiva una especie inhibe a otra en el uso de un recurso, ya sea a través de explotación o interferencia (Giller, 1984): el individuo competitivo dominante ó superior disminuye la abundancia del recurso limitante al competitivo subordinado ó inferior y en casos de recursos sedentarios tiende a establecer la monopolización en el uso del sustrato primario y en consecuencia un monocultivo (sensu Paine, 1984). La visión diferencial de la competencia desde la perspectiva del dominante o subordinado (ver Goldberg & Fleetwood, 1987; Goldberg, 1987) tendrá relevancia en un contexto ecológico-pesquero, al evaluar los efectos bioeconómicos derivados de esta interdependencia ecológica. La depredación es el consumo de un organismo (presa) por otro (depredador) a través de carnivoría, herbivoría u omnivoría (Begon et al., 1990). Un depredador puede ser clasificado de acuerdo a la amplitud de su dieta en monófago, oligófago y polífago, o también en especialista (monófagos y oligófagos) y generalistas (polífagos) según si reacciona o no a perfiles de

43
43
abundancia de presas. Exhibe preferencia trófica por alguna presa cuando la proporción de ésta en su dieta alimentaria es mayor que la proporción en el ambiente (Begon et al., 1990). Estos conceptos serán tomados en cuenta al evaluar el significado de dicha interdependencia ecológica en pesquerías. Análisis de interdependencias El marco teórico desarrollado en el Capítulo 2 suponía la explotación de un stock biológicamente independiente por parte de una flota homogénea. Las interdependencias ecológicas quedaban implícitamente consideradas en los parámetros K y r de la ecuación (2.1). En esta sección se expande el modelo a efecto de ilustrar las interdependencias ecológicas y tecnológicas relevantes en pesquerías multiespecíficas, manteniéndose los mismos supuestos que en el modelo uniespecífico. Esfuerzo pesquero heterogéneo En una pesquería se puede presentar heterogeneidad en el esfuerzo pesquero ejercido sobre una especie objetivo, generándose interdependencias de tipo tecnológico entre las flotas (o embarcaciones) con diferente poder de pesca y muchas ocasiones con costos unitarios heterogéneos del esfuerzo pesquero. La captura sostenible de dos flotas con diferente poder de pesca (e.g. artesanal y mecanizada) puede representarse de la siguiente forma:
dBdt
rB(t) 1B(t)K C (t) C (t)= −
⎛⎝⎜
⎞⎠⎟− −1 2 (3.1)
donde: C1(t) = captura de la flota 1 en el tiempo t. C2(t) = captura de la flota 2 en el tiempo t. Análisis estático. Bajo condiciones de equilibrio biológico dB/dt=0, por lo que:
rB 1BK
C C 0−⎛⎝⎜
⎞⎠⎟− − =1 2 (3.2)
por lo tanto la captura sostenible de la flota 1 puede expresarse como sigue:
1 2C rB 1BK
C= −⎛⎝⎜
⎞⎠⎟− (3.3)
Por otra parte se sabe que:

44
44
1 1 1C q f B= (3.4)
2 2 2C q f B= (3.5)
Despejando B en la ecuación (3.2) y sustituyendo C1 y C2 por las expresiones (3.4) y (3.5) se obtiene:
B 1-q f +q f
r K=
⎛⎝⎜
⎞⎠⎟1 1 2 2 (3.6)
Sustituyendo la ecuación (3.6) en las ecuaciones (3.4) y (3.5), la captura para cada una de las flotas será:
1 1 11 1 2 2C q f 1
q f q fr
K= −+⎛
⎝⎜
⎞⎠⎟ (3.7)
2 2 21 1 2 2C q f 1
q f q fr
K= −+⎛
⎝⎜
⎞⎠⎟ (3.8)
y la renta económica de cada flota:
1 1 1 1π = −pC c f (3.9)
2 2 2 2π = −pC c f (3.10)
Para obtener el equilibrio bioeconómico de las diferentes flotas, por ejemplo la flota 1, se hace π1 = 0 y se estima el correspondiente esfuerzo pesquero en equilibrio bioeconómico. En tal sentido, al sustituir la ecuación (3.7) en la (3.9) se obtiene:
π1 1 1
21
21 2 2 1 1
1 1 0= −⎛
⎝⎜
⎞
⎠⎟
⎛⎝⎜
⎞⎠⎟ − =Kpq f Kpq f
r+
q f q f Kr
c f (3.11)
Despejando f1 de la ecuación 3.11 se obtiene el esfuerzo pesquero en equilibrio bioeconómico para la flota 1.

45
45
f
1 cKpq r q f
qEBE1
1
12 2
1=
−⎛
⎝⎜
⎞
⎠⎟ −
⎡
⎣⎢⎢
⎤
⎦⎥⎥
(3.12)
El MRE de la flota 1 se obtiene al igualar a cero la primera derivada de los ingresos netos, lo cual puede expresarse de la siguiente manera:
c KpqKpq 2 f
rq f q pK
r1 1
21 1 2 2 1= −−⎛
⎝⎜
⎞
⎠⎟+
⎛
⎝⎜
⎞
⎠⎟ (3.13)
Factorizando se obtiene el costo unitario del esfuerzo pesquero:
c Kpq 12q f +q f
r1 11 1 2 2= − −
⎛
⎝⎜
⎞
⎠⎟ (3.14)
despejando f1 de la ecuación anterior se obtiene el esfuerzo de la flota 1 en máximo rendimiento económico:
f1
cKpq r q f
2qMRE1
1
12 2
1=
−⎛
⎝⎜
⎞
⎠⎟ −
⎡
⎣⎢
⎤
⎦⎥
(3.15)
Comparando las ecuaciones 3.11 y 3.14 se puede observar que dado el esfuerzo de la flota 2 el esfuerzo pesquero en máximo rendimiento económico fMRE de la flota 1 es igual a la mitad del esfuerzo en equilibrio bioeconómico fEBE de esa misma flota. De igual manera se obtiene el esfuerzo pesquero en MRE y en equilibrio bioeconómico para la flota 2. Análisis dinámico. Para observar la trayectoria hacia el equilibrio bioeconómico o bien el comportamiento de la dinámica de la estructura de la población explotada y de la intensidad de pesca, es necesario resolver numéricamente dicho modelo construyendo programas de computación. En el siguiente ejemplo se analiza la evolución dinámica de dos flotas con diferente poder de pesca que inciden sobre el mismo recurso, suscitándose una interdependencia tecnológica.

46
46
Ejemplo. 3.1 Flotas con diferente poder de pesca Dada una pesquería de una especie pelágica cuyos parámetros aparecen en la Tabla 3.1, se analiza el comportamiento dinámico de la biomasa de la población, de las capturas y esfuerzo pesquero desplegado por dos flotas con diferente poder de pesca (mecanizada = industrial y artesanal), así como los diferentes niveles de ingresos y costos generados por ambas flotas.
Tabla 3.1 Parámetros definidos al inicio de la corrida de simulación para evaluar el comportamiento dinámico de la biomasa poblacional, capturas, esfuerzo y renta económica generada por dos flotas con diferente poder de pesca.
Parámetro/Variable
Valor
Unidad de medida
Tasa intrínseca de crecimiento 0.36 -- Capacidad de soporte del ecosistema 4000000 ton Coef. capturabilidad flota artesanal 0.0002 -- Coef. capturabilidad flota mecanizada 0.00035 -- Precio de la especie 60 US$/ton Costo unitario esfuerzo artesanal 15000 US$viaje Costo unitario esfuerzo mecanizado 45000 US$/viaje Parámetro de dinámica flota artesanal 0.00001 -- Parámetro de dinámica flota mecanizada 0.00001 --
En la Figura 3.1 se pueden observar los resultados de la corrida de simulación a un horizonte de 50 años, con el correspondiente comportamiento dinámico de la biomasa del recurso, así como de las capturas de ambas flotas y las rentas económicas generadas por la actividad, todas en función del esfuerzo pesquero de ambas flotas. Los costos unitarios del esfuerzo y los coeficientes de capturabilidad relativos de ambas flotas determinan la expansión de la flota artesanal hasta llevar la biomasa a niveles inferiores de dos millones de toneladas con la consecuente eliminación de la renta de la flota industrial. Lo anterior motiva en consecuencia la salida de embarcaciones industriales liberando biomasa que es capturada posteriormente por la flota artesanal.

47
47
1000
1500
2000
2500
3000
3500
4000 Bi
omas
a ('0
00 to
n)
0 100 200 300 400 500 600 Esfuerzo industrial (barcos)
a
1000
1500
2000
2500
3000
3500
4000
Biom
asa
('000
ton)
0 200 400 600 800 1000 1200 1400 Esfuerzo artesanal (barcos)
b
0
100
200
300
400
500
Cap
tura
('00
0 to
n)
0 100 200 300 400 500 600 Esfuerzo industrial (barcos)
c
0
50
100
150
200
250
300
350
Cap
tura
('00
0 to
n)
0 200 400 600 800 1000 1200 1400 Esfuerzo artesanal (barcos)
d
-4000
-2000
0
2000
4000
6000
8000
Ren
ta ('
000
US$
)
0 100 200 300 400 500 600 Esfuerzo industrial (barcos)
e
0
1000
2000
3000
4000
5000
Ren
ta ('
000
US$
)
0 200 400 600 800 1000 1200 1400 Esfuerzo artesanal (barcos)
f
Figura 3.1 Comportamiento dinámico de la biomasa del recurso (a, b), las capturas por flota (c, d) así como la distribución de la renta económica (e, f) por tipo de flota, como resultado de las interdependencias tecnológicas entre ellas. La Figura 3.2. muestra el comportamiento dinámico de la biomasa y el esfuerzo pesquero aplicado por las flotas artesanal y mecanizada. Se observa el brusco descenso de la biomasa poblacional durante los 10 primeros años de la pesquería, como consecuencia de un fuerte incremento del esfuerzo pesquero en la fase de desarrollo o expansión de la actividad. Esto resulta evidente en el caso de la flota artesanal, que con sus menores costos operativos e inversión posibilita un crecimiento más acelerado del número de barcos. La tendencia en el largo plazo de ambas variables es a presentar ciclos levemente desfasados en el tiempo (e.g. 5 años) como producto del retraso en la respuesta de la decisión de inversión por parte de los pescadores con respecto a variaciones en la abundancia del recurso (Fig. 3.2a). La amplitud de las oscilaciones de ambas variables disminuye en magnitud, hasta llegar en el largo plazo a un

48
48
equilibrio en biomasa (1170 ton) y en el número de barcos de la flota mecanizada (7) y artesanal (475). En esa etapa los ingresos netos de ambas flotas tenderán a cero (ingresos totales = costos totales), y por tanto a un equilibrio bioeconómico donde no habrá incentivo para la entrada de nuevas unidades pesqueras. Es clara la tendencia de la flota artesanal a excluir a la mecanizada, debido a la ocurrencia de costos operativos mucho menores.
1000
1500
2000
2500
3000
3500
4000
Bio
mas
a ('0
00 to
n)
0 10 20 30 40 50 Tiempo (años)
a
0
200
400
600
800
1000
1200
1400
Esf
uerz
o (b
arco
s)0 10 20 30 40 50
Tiempo (años)
b
FlotaIndustrial
FlotaArtesanal
0
100
200
300
400
500
Cap
tura
('00
0 to
n)
0 10 20 30 40 50 Tiempo (años)
cFlotaIndustrial Flota
Artesanal
-4000
-2000
0
2000
4000
6000
8000
Ren
ta ('
000
US
$)
0 10 20 30 40 50 Tiempo (años)
dFlotaArtesanal
FlotaIndustrial
Figura 3.2 Trayectoria dinámica de la biomasa (a), el esfuerzo (b), la captura (c) y la renta económica (d) obtenida por las flotas industrial y artesanal.
Este caso pudiera ser considerado en el contexto de una pesquería secuencial (ver más abajo), sin embargo no considera la estructura por edades de la población y en consecuencia desconoce el efecto que cada flota pudiera tener en los diferentes componentes poblacionales. En efecto, es común encontrar que la pesquería artesanal tiende a afectar a organismos juveniles que se distribuyen en zonas costeras, mientras que la mecanizada tiende a hacer lo propio con los organismos adultos que se distribuyen hacia mayores profundidades (e.g. camarón: ver Willmann & Garcia, 1985). Este caso es desarrollado en la sección correspondiente a pesquerías secuenciales.

49
49
Pesquerías tecnológicamente interdependientes: una flota afecta stocks ecológicamente independientes En este caso dos stocks sin ninguna interdependencia ecológica (e.g. nichos disjuntos, sensu Pianka, 1978) aparecen representados en las capturas realizadas por una flota con artes de pesca no discriminatorios. Cada unidad de esfuerzo afecta por igual a ambos stocks, por lo cual su representación relativa en las capturas dependerá del tamaño poblacional y magnitud de esfuerzo utilizado. A continuación se desarrolla la teoría aportada por Clark (1985) como fundamento para el análisis de este tipo de interdependencias. Marco teórico (Clark, 1985). La ecuación básica del modelo de Schaefer para una población explotada (2.2) puede ser empleada para describir las tasas de cambio en biomasa de la especie X:
XX X
X
XX X
dB (t)dt
r B (t) 1B (t)K
q f(t)B (t)= −⎛⎝⎜
⎞⎠⎟ − (3.16)
y en forma análoga para la especie Y. En condiciones de equilibrio dBX(t)/dt=dBY(t)/dt=0, por lo tanto:
X
X
X
X
Y
Y
Y
Y
r (t)q 1
B (t)K
r (t)q 1
B (t)K
f(t)−⎛⎝⎜
⎞⎠⎟ = −
⎛⎝⎜
⎞⎠⎟ = (3.17)
A efectos ilustrativos asúmase que (Clark, 1985):
rq
rq
X
X
Y
Y
> (3.18)
En ausencia de esfuerzo pesquero, la curva de equilibrio resultante de (3.17) se origina en el nivel de la capacidad de carga de ambas especies KX,KY. Las especies decrecen en biomasa a medida que se intensifica el esfuerzo pesquero. La especie Y, con menor tasa de crecimiento según (3.18), se extinguirá en cierto nivel de esfuerzo P , mientras que X aún provee una captura sostenible. En el nivel de esfuerzo P, para la especie X se tendrá una biomasa (BXp ) que puede estimarse de la siguiente forma:
pX XY Y
X XB (t)= K 1 -
r / qr / q
⎛
⎝⎜
⎞
⎠⎟ (3.19)
La especie Y será eliminada si:
Y Y YF = q f ( t ) > r (3.20)

50
50
o sea, la mortalidad por pesca FY de Y excede su tasa intrínseca de crecimiento. En el caso de X, la condición (3.18) garantiza que FX < rX, por lo cual la pesquería persistirá como económicamente viable aún cuando Y no aparezca representada en las capturas. De acuerdo al análisis anterior, la eliminación de especies con menor tasa intrínseca de crecimiento poblacional es factible en condiciones de acceso abierto en un contexto multiespecífico. Considerando la extensión de (2.20), los ingresos netos generados por una pesquería serán:
π( t ) [ p q B ( t ) p q B ( t ) c ] f ( t )X X X Y Y Y= + − (3.21)
En una pesquería de libre acceso Y será eliminada si BX < BXp, o sea:
cp q K 1
r / qr / qX X
XY Y
X X= −
⎛
⎝⎜
⎞
⎠⎟ (3.22)
La sobreexplotación y probable extinción de una especie dependerá de su valor en el mercado, su posición y papel funcional en la cadena trófica y aspectos inherentes a sus características biológicas y poblacionales. Interdependencias tecnológicas: una flota afecta dos o más stocks con cierta segregación en el espacio En este tipo de pesquerías, el esfuerzo pesquero es dirigido a dos o más stocks. Si bien las actividades pesqueras son mutuamente excluyentes (Huppert, 1979; Anderson, 1981, 1982a), es factible obtener captura incidental. Esto es particularmente viable en el caso de stocks de especies sedentarias con cierto grado de segregación espacial, lo cual permite direccionar el esfuerzo pesquero. En este tipo de recursos los pescadores perciben a la costa como una colección de áreas discretas que son objeto de rotación en función de la magnitud de aquellas agregaciones de individuos que optimizen la combinación adecuada de tallas de interés comercial (Caddy, 1975; Hall, 1983). El "modelo de producción excedente en mosaico" (Munro, 1979; Polovina, 1989; Caddy & Garcia, 1983; Caddy, 1986) constituiría una buena aproximación uni ó multiespecífica al respecto: los datos colectados en diferentes zonas de pesca con cierta identidad geográfico-ambiental y diferentes intensidades de pesca, pueden ser empleados para definir estrategias óptimas de explotación y rendimientos sostenibles por unidad de área. Considerando k áreas con una extensión Ak en el mismo ó en diferentes años t, la ecuación 2.14 del modelo de Schaefer es modificada como:
kt
kt
kt
k
Cf
a b fA
= − (3.23)
Esta aproximación, que supone equilibrio en el sistema (aproximación estática), implica un buen mapeo de cada zona de pesca, así como una idea previa de la productividad y características poblacionales de los stocks. Dado que se asume en este caso que las actividades

51
51
pesqueras en cada stock son mutuamente excluyentes, es factible conjuntar los resultados de cada uno a posteriori y elaborar un "mapa de rendimiento bioeconómico", en base a los ingresos netos (ec. 3.21) generados por cada stock. Clark (1982) y Hilborn & Walters (1987) entre otros, han desarrollado modelos tendientes a simular la dinámica espacial de un recurso multiespecífico y del esfuerzo pesquero aplicado a éste. Parten de la ecuación (2.3) en una base multiespecífica:
ik ki ki iC B ( t ) f ( t )q= (3.24)
aplicada a i especies en k zonas de pesca, estimando los ingresos netos por unidad de esfuerzo, zona de pesca y especie. Pesquerías tecnológicamente interdependientes: dos flotas CASO 1: la flota 1 captura incidentalmente la especie objetivo de la flota 2. Considérense dos flotas: la flota 1, concentrada en la explotación de la especie X pero obteniendo incidentalmente cierto porcentaje de Y, la cual a su vez es objetivo de la flota 2. En la Tabla 3.2 se muestran los datos relevantes de ambas especies y flotas, a efectos de su introducción en el análisis bioeconómico dinámico de la corrida de simulación.
Tabla 3.2 Parámetros definidos al inicio de la corrida de simulación para evaluar el comportamiento dinámico de la biomasa poblacional de las especies X e Y capturadas por flotas tecnológicamente interdependientes.
Parámetro/Variable
Valor
Unidad de medida
r especie X 0.36 -- r especie Y 0.30 -- K especie X (ton) 4000000 ton K especie Y (ton) 2000000 ton q especie X flota 1 0.0004 -- q especie X flota 2 0 -- q especie Y flota 1 0.00015 -- q especie Y flota 2 0.0025 -- Precio especie X 60 US$/ton Precio especie Y 45 US$/ton Costo unitario flota 1 60000 US$/año Costo unitario flota 2 15000 US$/año Parámetro de dinámica flota 1 0.00001 -- Parámetro de dinámica flota 2 0.000015 --
La Figura 3.3 muestra los resultados del modelo bioeconómico dinámico que representa la interdependencia tecnológica. Las fluctuaciones en biomasa muestran un acentuado declive de

52
52
ambas especies como resultado del desarrollo de la pesquería (Fig. 3.3a). La flota 1, con mayor poder de pesca, realiza importantes capturas de X en los primeros 20 años, lo cual determina una reducción de la abundancia a la mitad de su capacidad de carga durante el mismo período. La flota 2, cuyo objetivo es la especie Y, alcanza un máximo de sus capturas y desarrollo de la pesquería a los 8 años de haberse iniciado la misma (Fig. 3.3b). Sin embargo, el aumento de la captura incidental a partir de dicho lapso determina una reducción de sus rendimientos y una renta económica negativa en los años subsiguientes, en concordancia con un aumento sostenido de capturas, esfuerzo y renta económica de la flota 1, la cual a su vez percibe ingresos adicionales derivados de la captura incidental de Y (Figs. 3.3c y 3.3d). En el largo plazo ambas flotas alcanzan el equilibrio bioeconómico a niveles de esfuerzo de 420 (flota 1) y 500 (flota 2) barcos. Sin embargo, la tendencia será a mostrar ciclos variables en amplitud de acuerdo a la presión pesquera ejercida por ambas flotas. En una situación ideal, el nivel de esfuerzo en 1 debería ser expandido hasta que el retorno marginal de la captura en ambas pesquerías iguale el costo marginal del esfuerzo pesquero en 1, mientras que para 2 es aplicable similar razonamiento.
500
1000
1500
2000
2500
3000
3500
4000
Bio
mas
a ('0
00 to
n)
0 10 20 30 40 50 Tiempo (años)
a
sp 1
sp 2
0
200
400
600
800
1000
1200 E
sfue
rzo
(bar
cos)
0 10 20 30 40 50 Tiempo (años)
b
Flota 2
Flota 1
0
200
400
600
800
1000
Cap
tura
('00
0 to
n)
0 10 20 30 40 50 Tiempo (años)
cFlota 1, sp1
Flota 2, sp2
Flota 1, sp2
-20000
-10000
0
10000
20000
30000
Ren
ta ('
000
US
$)
0 10 20 30 40 50 Tiempo (años)
d
Flota 2
Flota 1
Figura 3.3. Modelo bioeconómico dinámico de interdependencias tecnológicas entre dos flotas: biomasa de la especie objetivo e incidental (a), esfuerzo pesquero (b), capturas de ambas especies (c) y renta económica generada (d). CASO 2: las flotas 1 y 2 se afectan mutuamente. Esto se da cuando las dos flotas obtienen incidentalmente capturas del stock objetivo de la otra pesquería. El tamaño poblacional y rendimientos de los stocks X e Y son función del nivel de esfuerzo en ambas pesquerías. Sin

53
53
embargo, el retorno de una dependerá del nivel de esfuerzo generado por la otra. Para la pesquería 1 (especie objetivo X):
1 1 1 2 1 1 2 1 1π (t) p Y ( f , f ) p Y ( f , f ) c fX X Y Y= + − (3.25)
mientras que para la pesquería 2 (especie objetivo Y):
2 1 2 1 2 2 2π (t) p Y ( f , f ) p Y ( f , f ) c fY Y 2 X X 2= + − (3.26)
A medida que una flota incrementa su esfuerzo, el rendimiento en equilibrio por parte de la otra flota decaerá y en consecuencia su nivel de esfuerzo en acceso abierto. Interdependencias tecnológicas: pesquerías secuenciales Es el caso de dos o más flotas de diferentes características que explotan diferentes componentes poblacionales en el ciclo de vida de una especie (e.g. juveniles y adultos) generalmente segregados en el espacio. Es común en casos de pesca costera artesanal o deportiva y pesca mecanizada de altura, y generalmente incluye conflictos producidos por competencia por el recurso entre los diferentes usuarios. Al respecto se han desarrollado modelos bioeconómicos estáticos y dinámicos, a efectos de determinar la asignación óptima de capturas entre dos flotas que explotan un stock en forma secuencial (Geen & Nayar, 1988). En este tipo de pesquerías se observa el efecto de mutua interacción entre dos flotas segregadas en el espacio (e.g. artesanal costera y mecanizada de altura) que afectan diferencialmente a distintos componentes del ciclo vital de las especies involucradas. Por tanto, si bien la especie en cuestión posee una única curva de población en equilibrio, existirán dos trayectorias diferentes en las capturas, así como dos estructuras relativas precio-costo e ingresos generados por las flotas involucradas. La exclusión o coexistencia competitiva de las flotas ocurrirá en condiciones de libre acceso, mientras que un óptimo balance en la asignación de capturas dependerá especialmente de los cocientes entre precios y costos de ambas flotas, conjuntamente con aquellos parámetros biológicos que determinan la estructura por edades, reclutamiento y sobrevivencia del stock (Hoppensteadt & Sohn, 1981; Seijo et al., 1994b). Debe recalcarse la importancia de considerar la estructura por edades de las poblaciones explotadas, así como el efecto diferencial que cada flota pudiera tener en los diferentes componentes poblacionales. El siguiente ejemplo se basa en un modelo bioeconómico multiespecífico de estructura por edades con interdependencias tecnológicas secuenciales (Seijo et al., 1996). Los valores de entrada de los parámetros al modelo se muestran en la Tabla 3.3. En la Figura 3.4. se observan las fluctuaciones en el tiempo de los niveles de abundancia, captura, esfuerzo y renta económica para las flotas mecanizada y artesanal que componen la pesquería secuencial multiespecífica. En el caso de la biomasa (Fig. 3.4a), se produce un pronunciado decremento de ambas especies durante los primeros años de desarrollo de la pesquería, como resultado de una franca expansión de la flota mecanizada. Las capturas de dicha flota alcanzan su máximo a los 20 años (Fig. 3.4b), coincidiendo con el máximo número de barcos (Fig. 3.4c); a partir de ese período, la flota comienza a decrecer en forma paulatina hasta

54
54
el final del horizonte simulado (50 años). En contraste, la pesquería artesanal muestra una expansión no tan pronunciada pero constante de las capturas, como producto de un fuerte incremento del esfuerzo pesquero a partir del comienzo de la declinación de la flota mecanizada (20 años). Ejemplo 3.3 Pesquería multiespecífica con interdependencias tecnológicas secuenciales.
Tabla 3.3 Parámetros incluidos en un modelo bioeconómico multi-específico de estructura por edades con interdependencias tecnológicas secuenciales.
Parámetro/Variable
Valor
Unidad de medida
Edad máxima especie 1 20 años Edad máxima especie 2 12 años Edad primera madurez 1 3 años Edad primera madurez 2 2 años Proporción por sexos (ambas spp.) 0.50 -- Reclutamiento máximo observado 1 50000000 organismos Reclutamiento máximo observado 2 100000000 organismos Tasa mortalidad natural 1 0.4 1/año Tasa mortalidad natural 2 0.45 1/año Fecundidad promedio de 1 1200000 huevos Fecundidad promedio de 2 1500000 huevos Parámetro curvatura 1 0.12 1/año Parámetro curvatura 2 0.19 1/año Parámetro de ajuste 1 -0.55 anual Parámetro de ajuste 2 0.50 anual Longitud asintótica 1 920 mm Longitud asintótica 2 600 mm Peso asintótico 1 12500 g Peso asintótico 2 2400 g Precio mínimo 1 1500 US$/ton Precio mínimo 2 1000 US$/ton Pendiente curva de precios 1 50 -- Pendiente curva de precios 2 5 -- Costo unitario flota 1 55000 US$/barco/año Costo unitario flota 2 5000 US$/barco/año Parámetro de dinámica flota 1 0.00001 -- Parámetro de dinámica flota 2 0.0001 -- Area barrida flota 1 2.50 Km2 Area barrida flota 2 0.18 Km2 Long. retención 50% y 75% sp. 1 flota 1 450; 650 mm Long. retención 50% y 75% sp. 1 flota 2 300; 450 mm Long. retención 50% y 75% sp. 2 flota 1 300; 450 mm Long. retención 50% y 75% sp. 2 flota 2 200; 275 mm Area de distribución ambas spp. 5000 Km2

55
55
Los menores costos operativos de la flota artesanal tienden a inducir un aumento del número de barcos e indirectamente a excluir en el largo plazo a la flota mecanizada. Otro efecto del aumento del esfuerzo en la flota artesanal puede ser la sobrepesca en el reclutamiento, lo cual a su vez disminuye el potencial del stock a ser capturado por la flota mecanizada. En cuanto a la renta económica generada por la actividad pesquera, la sobreinversión como producto de amplios márgenes de ganancia en la flota mecanizada (entre los años 15 y 20) produce su sobredimensionamiento y posteriores pérdidas a la flota (entre años 20 y 25), para luego tender al equilibrio bioeconómico. La flota artesanal, con pequeños valores de renta y siempre cercana a este equilibrio, presenta una leve tendencia creciente de su renta económica en el largo plazo (Fig. 3.4d).
40
60
80
100
120
140
Bio
mas
a ('0
00 to
n)
0 10 20 30 40 50 Tiempo (años)
sp1
sp2
a
0
2000
4000
6000
8000
Cap
tura
(ton
)
0 10 20 30 40 50 Tiempo (años)
b
sp1. Flota artesanal
sp1. Flota industrial
sp2. Flota industrial
sp2. Flota artesanal
0
200
400
600
800
1000
1200
Num
ero
de b
arco
s
0 10 20 30 40 50Tiempo (años)
c
Flota industrial
Flota artesanal
-2000
-1000
0
1000
2000
3000
4000
5000
Ren
ta E
cono
mic
a ('0
00 U
S$)
0 10 20 30 40 50 Tiempo (años)
Flota industrial
Flota artesanal
d
Figura 3.4 Interdependencias tecnológicas. Niveles de biomasa (a), esfuerzo (b), captura (c) y renta económica generada por las flotas industrial y artesanal (d).
Análisis gráficos alternativos son desarrollados por Anderson (1977; 1986). En ambos casos se presenta a una pesquería secuencial en un contexto estático.

56
56
Stocks ecológicamente interdependientes: depredación El modelo básico empleado anteriormente asumía carencia de interacción entre las especies X e Y. En caso de especies ecológicamente interdependientes sujetas a explotación, un modelo general es descrito por el siguiente sistema de ecuaciones (Clark, 1985):
X X X X X 1 Y
XX X
dBdt
r B (t)[ K B (t) B (t)]K
q f(t) B (t)=− −
−α
(3.27)
Y X X X X 1 Y
XX X
dBdt
r B (t)[ K B (t) B (t)]K
q f(t) B (t)=− −
−β
(3.28)
donde los términos α1 y ß1 son coeficientes de competencia que miden el efecto per capita de una especie sobre otra (Begon et al., 1990); valores inferiores a 1 suponen un efecto inhibitorio interespecífico mayor que el intraespecífico. Un sistema simple depredador (especie Y) - presa (especie X) en el cual ambas especies son explotadas, puede expresarse en base a las ecuaciones 3.29 y 3.30 (Clark, 1985):
XX X X X Y X
dB (t)dt
R M B (t) B (t) B (t) C (t)= − − −α 2 (3.29)
donde: MX es la tasa de mortalidad de X. α2BX(t)BY(t) representa la tasa de consumo de X por Y. RX es la tasa de reclutamiento de X (denso-independiente). La ecuación (3.29) supone una distribución uniforme de la presa, lo cual impone limitaciones al modelo. Asimismo, el modelo supone que las capturas de cada especie son realizadas en forma independiente, e.g. no constituyen un stock múltiple. Formas alternativas de interacción depredador-presa, incluyendo agregación de presas y diferente comportamiento del depredador, han sido discutidas en el plano ecológico (Taylor, 1988; Comins & Hasell, 1979; Begon et al., 1990). La otra ecuación que conforma el sistema es:
YX Y Y Y Y
dB (t)dt
B (t) B (t) M B (t) C (t)= − −λα 2 (3.30)
El término BX(t)BY(t) representa la tasa natural de crecimiento poblacional del depredador. Y es depredador especialista de X. λ es un parámetro adimensional que puede interpretarse como un índice de conversión alimenticia de X a Y. En caso que el stock Y no sea explotado, existirá una biomasa crítica de X, BXc, dada por:

57
57
CXYB M=
λα (3.31)
de tal manera que habrá crecimiento poblacional en Y siempre que la biomasa de X supere a BXc. La ecuación (3.29) asume asimismo que X (presa) posee una tasa de reclutamiento R constante, e.g. denso-independiente. Se asume asimismo que BXc es menor a la capacidad de carga KX en ausencia de depredación y explotación, dada por:
XX
XK
RM
= (3.32)
El ingreso neto de la especie X será:
X XX X
X(t) p cq B (t)
C (t)π = −⎛
⎝⎜
⎞
⎠⎟ (3.33)
y para la especie Y:
Y YY Y
Y(t) p cq B (t)
C (t)π = −⎛
⎝⎜
⎞
⎠⎟ (3.34)
donde el término c/qB representa el costo de capturar una unidad de biomasa cuando la biomasa total es B (Clark, 1985), lo cual es fácilmente deducible a partir de la función de CT(t) definida en el Capítulo 2. El equilibrio bioeconómico simultáneo se dará cuando πX(t)=πY(t)=0. Un análisis completo sobre la construcción de curvas de rendimiento sostenible en pesquerías multiespecíficas es encontrado en Ströbele & Wacker (1991). Este estudio pierde relevancia si no se especifican las características trófico-conductuales del depredador (e.g. especialista ó generalista), así como la magnitud de las interacciones que unen a la presa X con otros competidores. Stocks ecológicamente interdependientes: competencia Exclusión competitiva. Este concepto es definido de acuerdo a la teoría de nicho (Hutchinson, 1958, 1959; Vandermeer, 1972). En ausencia de competidores, una especie ocupará su nicho fundamental, dado por la combinación óptima de condiciones y recursos que le

58
58
permiten mantener cierto nivel poblacional. La presencia de competidores lo restringe a su nicho realizado, definido como el conjunto observado de condiciones en las cuales la especie existe normalmente. Las dimensiones estarán determinadas por la magnitud de las interacciones, y en consecuencia el nicho realizado es menor o igual al fundamental (Giller, 1984; Begon et al., 1990). En este contexto, el Principio de Exclusión Competitiva (Gause, 1934) establece que si dos especies coexisten en un ambiente, lo harán como resultado de la diferenciación de sus nichos (e.g. coexistencia competitiva; nichos realizados). Si esta diferenciación de nichos no es viable, el competidor superior excluirá al subordinado cuando su nicho realizado se superponga totalmente con el nicho fundamental del subordinado. Pongamos por ejemplo dos recursos sedentarios, en los cuales la competencia interespecífica es generalmente muy intensa y se relaciona con la disponibilidad de espacio. En organismos filtradores de este tipo dicho proceso trae como consecuencia que una especie domine la comunidad a través de la exclusión del competidor subordinado (Branch, 1984). Un ejemplo típico lo constituye la interacción entre los bivalvos suspensívoros Mesodesma mactroides-Donax hanleyanus en el litoral Atlántico uruguayo (de Alava, 1993). En ausencia de impacto humano, M. mactroides es competidor dominante y reduce a mínimas densidades a D. hanleyanus. La explotación preferencial de la primera disminuye la magnitud de la interacción por un aumento del espacio disponible, en este caso el recurso limitante. De esta manera se suscita alternancia en la abundancia de dichos bivalvos, dependiendo de la magnitud del impacto humano (Fig. 3.5). Sin embargo, la característica variabilidad en el reclutamiento de estos recursos que suponen en su mayoría ciclos de vida complejos (Roughgarden et al., 1988), unida a la inestabilidad de las comunidades relacionadas (Warwick, 1986; Menge & Sutherland, 1987) y el grado variable de impacto humano (Castilla, 1988), alteran constantemente la magnitud de las interacciones. En este contexto, fuentes importantes de variación son ocasionadas por la competencia intraespecífica por espacio y/o alimento, lo cual es analizado en numerales siguientes.
0
20
40
60
80
100
120
140
Adu
ltos
Mes
odes
ma
(ind/
m2)
0
20
40
60
80
Adu
ltos
Don
ax (i
nd/m
2)
O I P V O I P V O I P V O I P V O I PTiempo (trimestres)
Mesodesma
Donax
1983 1987
Figura 3.5 Variación temporal en la abundancia de los bivalvos suspensívoros Mesodesma mactroides y Donax hanleyanus de la costa Atlántica uruguaya (de Alava, 1993). Coexistencia competitiva. En el contexto del modelo de Gause (1934), la competencia interespecífica puede expresarse en base a las ecuaciones (3.35) y (3.36) en ausencia de explotación, o sea:

59
59
XX X
X
X
Y
X
dB (t)dt
= r B (t) 1B (t)K
B (t)K
− −⎛⎝⎜
⎞⎠⎟
α (3.35)
para dominante X, mientras que para la subordinada Y:
YY Y
Y
Y
X
Y
dB (t)dt
= r B (t) 1B (t)K
ßB (t)K
− −⎛⎝⎜
⎞⎠⎟ (3.36)
las cuales responden al modelo Lotka-Volterra (Volterra, 1926; Lotka, 1932). Anderson (1986) desarrolla otra aproximación gráfica basada en un criterio estático, asumiendo equilibrio en las poblaciones: previo a la explotación pesquera, dos especies X e Y coexisten en forma estable (Keddy, 1989). X es competidor superior y posee una población mayor; Y es competidor inferior. Se asume que, si bien ambas especies son valoradas en el mercado, la explotación es dirigida a X en una primera instancia, y que hay cierta segregación espacial que permite una discriminación de las especies en la actividad extractiva. Asimismo, considérese que el espacio disponible para el asentamiento es el recurso limitante por el cual compiten las dos especies sedentarias. A medida que el esfuerzo f(t) aumenta sobre X, disminuye la magnitud de la interacción, hay liberación de espacio disponible para el asentamiento y en consecuencia aumenta el tamaño poblacional de la especie competitiva inferior. Esta tendencia ocurre hasta cierto nivel de esfuerzo f1, donde X virtualmente desaparece e Y alcanza su máximo tamaño poblacional. La disminución de la especie competitiva superior produce un direccionamiento del esfuerzo pesquero a la explotación a Y en niveles de esfuerzo mayores a f1 . La forma de las curvas de rendimiento sostenible y de ingresos sostenibles y costos totales dependerán de la magnitud de las interacciones biológicas, capacidad de crecimiento poblacional, el valor relativo de las especies en el mercado y los costos de extracción. Es importante hacer notar que tal como lo establece Anderson (1986), en un escenario de competencia entre especies que son capturadas por una misma flota es de esperarse que ocurran equilibrios bioeconómicos tanto estables como inestables en el tiempo. La relevancia de este análisis se ve minimizada por su naturaleza estática. Es común encontrar situaciones donde la especie competitiva subordinada tiene mucho mayor valor económico. Como resultado, la curva de ingresos totales es mayor que en el caso estático, al igual que su presión de explotación (Mitchell, 1979). Este análisis está limitado además desde un punto de vista ecológico. La sobreposición de nicho (Giller, 1984) entre dos especies con respecto a un recurso puede no ser un fiel indicador de la situación real, dado que el número potencial de competidores por otros recursos no tomados en consideración aumenta al examinar más ejes del nicho definidos por otras variables ambientales. Por tanto, si bien la sobreposición de nicho puede ser pequeña, el efecto acumulativo de esta competencia difusa (sensu Pianka, 1974) puede reducir severamente el tamaño del nicho realizado de una especie, lo cual a su vez repercutirá en el manejo del recurso en caso de ser explotado. Por ejemplo, los valores de MRS, MRE y EBE podrían suceder a menores niveles de esfuerzo pesquero. Una vez más, la jerarquía competitiva de las especies explotadas, así como la magnitud y número de los ligamientos

60
60
tróficos horizontales (competencia) y verticales (depredación) determinarán los efectos bioeconómicos derivados de esta interdependencia. Competencia intraespecífica. Si bien el Capítulo ha sido encarado bajo un marco multiespecífico, se considera de importancia mencionar a la competencia intraespecífica como una interdependencia ecológica relevante en poblaciones marinas. La competencia intraespecífica se manifiesta de diversas formas (e.g. en organismos bentónicos se produce por limitación del espacio disponible para el asentamiento larval: Branch, 1984; Holm, 1990; Raimondi, 1990). La abundancia de un stock inducirá a fenómenos de denso-dependencia de intensidad variable en el crecimiento, mortalidad y reclutamiento, dependiendo de la estrategia de vida de la especie (Caddy & Sharp, 1986) y de la estructura por edades del stock. Continuando con el ejemplo de Mesodesma mactroides, se ha observado una relación stock-reclutamiento sujeta a procesos de sobrecompensación y que por tanto se explica por el modelo stock-reclutamiento de Ricker (1975) (Defeo, 1993a). Dicha relación repercute directamente en el crecimiento del recurso (en longitud y peso individual) y por consiguiente en su biomasa disponible para explotación y en los ingresos netos derivados de la actividad pesquera. Esta interdependencia debe ser considerada en el manejo de un recurso pesquero, tomando en cuenta la variabilidad espacio-temporal de la magnitud del reclutamiento a diferentes niveles de stock y variables ambientales. La predicción de la magnitud del reclutamiento en función de la magnitud del stock tendrá un efecto bioeconómico sustancial en el manejo de un recurso explotado. Así por ejemplo, en el recurso sedentario de referencia, la adopción de medidas regulatorias podrían basarse en maximizar cierto nivel de stock que permita liberar el necesario espacio disponible para un asentamiento exitoso. Stocks ecológicamente interdependientes afectados por dos flotas sin interdependencia tecnológica En este caso, dos stocks que constituyen pesquerías independientes tienen una relación de competencia o depredación, por lo cual se suscitarán efectos directos e indirectos que beneficiarán o perjudicarán a una u otra pesquería. La magnitud de las interacciones entre especies en el caso de competencia, y el grado de especificidad del depredador en su dieta alimentaria, determinarán la estrategia de uso óptimo de ambos stocks. Dependiendo de ello, el equilibrio bioeconómico en libre acceso de cada pesquería estará supeditado al monto de esfuerzo producido en la otra. CASO 1: stocks sujetos a coexistencia competitiva. La Figura 3.6 muestra las trayectorias dinámicas de biomasa y capturas por especie y la renta económica generada por estos recursos a una flota que ejerce su esfuerzo con un arte de pesca no discriminatorio. Como puede observarse, en ausencia de esfuerzo de pesca la abundancia de la especie 1 (competidor dominante) es mayor que la de la especie 2. Al irse incrementando el esfuerzo de pesca, la abundancia de la especie dominante comienza a decrecer liberando el recurso por el que se compite (hábitat y/o alimento) en el ecosistema en cuestión. El incremento en la disponibilidad del recurso permite que la especie dominada incremente su abundancia en términos absolutos y relativos con respecto a la especie dominante, hasta un nivel de esfuerzo después del cual su abundancia comienza también a decrecer. Bajo condiciones de acceso abierto, la pesquería

61
61
alcanza su nivel de equilibrio bioeconómico con un nivel de esfuerzo pesquero que resulta en niveles bajos de abundancia de la especie 1, existiendo la posibilidad de que si el precio de la especie 2 fuera lo suficientemente alto, la biomasa de la especie 1 podría alcanzar niveles cercanos al agotamiento. Las biomasas relativas y las capturas resultantes de ejercer un esfuerzo de pesca que tiende a maximizar la renta económica que genera la pesquería se obtienen con un nivel de esfuerzo en el orden de 192 barcos (Fig. 3.6).
2000
2500
3000
3500
4000
Bio
mas
a ('0
00 to
n)
1000
1100
1200
1300
1400
1500
1600
1700
Bio
mas
a ('0
00 to
n)
0 10 20 30 40 50 Tiempo (años)
Especiedominante
Especiedominada
a
2000
2500
3000
3500
4000
Bio
mas
a ('0
00 to
n)
1000
1100
1200
1300
1400
1500
1600
1700
Bio
mas
a ('0
00 to
n)
0 100 200 300 400 500 Esfuerzo (barcos)
ffMRE EBE
b
Especiedominante
Especiedominada
0
100
200
300
400
500
Cap
tura
('00
0 to
n)
0
20
40
60
80
100
120
Cap
tura
('00
0 to
n)
0 10 20 30 40 50 Tiempo (años)
Especiedominante
Especiedominada c
0
100
200
300
400
500
Cap
tura
('00
0 to
n)
0
20
40
60
80
100
120
Cap
tura
('00
0 to
n)
0 100 200 300 400 500 Esfuerzo (barcos)
fMRE
fEBE
d
-2000
0
2000
4000
6000
Ren
ta E
conó
mic
a ('0
00 U
S$)
0 10 20 30 40 50 Tiempo (años)
e
-2000
0
2000
4000
6000
Ren
ta e
conó
mic
a ('0
00 U
S$)
0 100 200 300 400 500 Esfuerzo (barcos)
f fEBEMRE
f
Figura 3.6 Trayectorias dinámicas de la biomasa (a, b), la captura por especie (c, d), así como la renta económica (e, f) generada por una flota con arte de pesca no discriminatorio.
CASO 2: stocks con relación depredador-presa. En este caso las curvas de equilibrio bioeconómico y de rendimiento óptimo en equilibrio para la pesquería 1, dirigida sobre el depredador, tendrán pendiente negativa relativa (Fig. 3.7), dado que un incremento en el esfuerzo pesquero dirigido sobre la presa disminuirá su abundancia y en consecuencia la

62
62
magnitud de recurso disponible para el depredador en caso de ser éste especialista o bien oligófago. La tendencia inversa ocurrirá en el caso de la presa y las características de sus curvas de equilibrio bioeconómico y rendimiento máximo dependerán de la magnitud del esfuerzo ejercido en la pesquería del depredador, así como en la conducta trófica de éste.
2000
2500
3000
3500
4000
Bio
mas
a ('
000
ton)
50
100
150
200
250
300
Bio
mas
a ('0
00 to
n)0 10 20 30 40 50
Tiempo
presa
depredador
a
0
100
200
300
400
500
600
Esf
uerz
o (#
de
barc
os)
0
20
40
60
80
100
120
Esf
uerz
o (#
de
barc
os)
0 10 20 30 40 50 Tiempo
presadepredador
b
0
100
200
300
400
500
600
Cap
tura
('0
00 to
n)
0
1
2
3
4
5
Cap
tura
('00
0 to
n)
0 10 20 30 40 50 Tiempo
Flota 1:presa
Flota 1: depredador
Flota 2:depredador c
-4000
-2000
0
2000
4000
6000
8000
10000
Ren
ta E
conó
mic
a ('0
00 U
S$)
0 10 20 30 40 50 Tiempo
Flota 1
Flota 2
d
Figura 3.7 Trayectoria dinámica de las biomasas de especies con relación depredador-presa (a), el esfuerzo de dos flotas que inciden diferencialmente sobre ellas (b), las capturas correspondientes (c) y la renta económica estimada para cada flota (d). Interdependencias tecno-ecológicas: stocks ecológicamente interdependientes afectados por dos flotas con interdependencia tecnológica Una variante del caso anterior sería la explotación de stocks ecológicamente interdependientes por parte de dos flotas sujetas a interdependencias tecnológicas, lo cual constituye una interdependencia tecno-ecológica. El análisis en este caso se dificulta por la naturaleza múltiple de la interacción. Sin embargo, se aportan aquí dos ejemplos basados en interdependencias de competencia y depredación.

63
63
CASO 1: stocks sujetos a interacción competitiva Los modelos que analizan la relación de competencia entre dos especies están generalmente basados en el modelo Lotka-Volterra anteriormente desarrollado, así como en el modelo aportado por Gause & Witt (1935). En este caso, las ecuaciones diferenciales básicas involucradas son:
XX X
X X
X
X X Y
X
dNdt
r Nr N
Kr N N
K= −
′−
α 3 (3.37)
YY Y
Y Y
Y
Y Y X
X
dNdt
r Nr N
Kr N N
K= −
′−
β 3 (3.38)
donde NX y NY es la abundancia poblacional de las especies X (dominante) e Y (subordinada) respectivamente, rX y rY son sus tasas intrínsecas de crecimiento, KX y KY son las capacidades de carga del ambiente cuando ambas especies crecen solas, α3 es una constante que considera el efecto de la especie X sobre la Y, y ß3 es una constante que considera el efecto de la especie Y sobre la X. Básicamente, estas ecuaciones diferenciales expresan que el incremento en la densidad poblacional de una especie está limitado por efectos intra e interespecíficos, éstos dados por la naturaleza y magnitud de la interacción competitiva. CASO 2: stocks con relación depredador-presa En este caso el modelo depredador-presa utilizado para generar la interdependencia ecológica está basado en el sistema de ecuaciones logísticas aportadas por Leslie & Gower (1960):
dBdt
r B 1BK
BXX X
XY= − −⎛
⎝⎜⎞⎠⎟
α 4 (3.39)
dBdt
r B 1B
ß BY
Y YY
X
= −⎛⎝⎜
⎞⎠⎟
4
(3.40)
En estas ecuaciones, el crecimiento de la presa está limitado tanto por la capacidad de carga del sistema, K, como por el término de depredación, α.4 El crecimiento de los depredadores es función de su tasa intrínseca de crecimiento rY , y del número de presas. La constante ß4 relaciona el número de presas con la biomasa máxima capaz de ser alcanzada por el depredador, en función de la capacidad de carga del sistema.

64
64
A continuación se brinda un ejemplo numérico y gráfico basado en un programa de computación (Seijo et al., 1996) que considera una interdependencia tecno-ecológica en conjunto con una interdependencia ecológica de depredación.
Tabla 3.4 Interdependencia tecno-ecológica. Parámetros definidos al inicio de la corrida de simulación para evaluar el comportamiento dinámico de la biomasa poblacional de las especies X (presa) e Y (depredador) capturadas por flotas tecnológicamente interdependientes. Parámetro/Variable
Valor
Unidad de
Medida r especie X 0.36 -- r especie Y 0.15 -- K especie X 4000000 ton K especie Y 275000 ton q especie X flota 1 0.0004 -- q especie X flota 2 0.0 -- q especie Y flota 1 0.0002 -- q especie Y flota 2 0.0004 -- Precio especie X 60 US$/ton Precio especie Y 275 US$/ton Costo unitario flota 1 60000 US$ Costo unitario flota 2 9000 US$ Parámetro de dinámica flota 1 0.00001 -- Parámetro de dinámica flota 2 0.000015 -- α4 constante de depredación: especie X 0.00000001 -- ß4 constante de depredacion: especie Y 0.069 --
En este caso las variaciones en el tiempo de las curvas de biomasa de la presa y el depredador no serán las clásicas encontradas en la literatura ecológica (Begon et al., 1990), sino que se ven afectadas por la magnitud diferencial de la interdependencia tecnológica (Fig. 3.8a). La disminución en la abundancia de ambas especies resulta del aumento del esfuerzo pesquero durante los primeros 15 años de la pesquería. El efecto de la flota 1, cuyo objetivo es la presa, también repercute en la biomasa y capturas efectuadas sobre el depredador durante ese lapso de expansión, para luego decaer paulatinamente (Fig. 3.8b). Dicho efecto puede observarse en la Figura 3.8c, donde el aumento de la flota 1 aparece como respuesta a los rendimientos logrados tanto en base a la especie objetivo (presa) como incidental (depredador). Ese incremento repercute en la renta económica generada por esa flota, mientras la flota 2, cuyo único objetivo es el depredador, no sufre variaciones substanciales en la renta sino que se presenta siempre cerca del equilibrio bioeconómico (Fig. 3.8d).

65
65
2000
2500
3000
3500
4000 Bi
omas
a ('0
00 to
n)
50
100
150
200
250
300
Biom
asa
('000
ton)
0 10 20 30 40 50 Tiempo (años)
Depredador
Presa a
0.01
0.1
1
10
100
1000
Cap
tura
('00
0 to
n)
0 10 20 30 40 50 Tiempo (años)
b
Flota 2: Depredador
Flota 1Depredador
Flota 1Presa
0
100
200
300
400
500
600
Esf
uerz
o (#
bar
cos)
0 10 20 30 40 50 Tiempo (años)
cFlota 1
Flota 2
-4000
-2000
0
2000
4000
6000
8000
10000
Ren
ta E
conó
mic
a ('0
00 U
S$)
0 10 20 30 40 50 Tiempo (años)
d
Flota 2
Flota 1
Figura 3.8 Interacción tecno-ecológica: stocks con interacción depredador-presa capturados por dos flotas tecnológicamente interdependientes.
Es obvia la correspondencia entre las abundancias de ambas especies, dado que un incremento en el esfuerzo pesquero dirigido sobre la presa disminuirá su abundancia y en consecuencia la magnitud de recurso disponible para el depredador en caso de ser éste especialista o bien oligófago. La tendencia inversa ocurrirá en el caso de la presa y las características de sus curvas de equilibrio bioeconómico y rendimiento máximo dependerán de la magnitud del esfuerzo ejercido en la pesquería del depredador, así como en la conducta trófica de éste.
Análisis ecológico-pesquero La inclusión de la variabilidad espacio-temporal en la abundancia poblacional y en el ambiente, conjuntamente con la cuantificación de la magnitud de las interdependencias ecológicas a diferentes niveles de esfuerzo pesquero, debe ser el camino adecuado para el modelaje multiespecífico (Pauly, 1979; 1988; Pope, 1979; Sainsbury, 1988). Las propuestas deben contemplar asimismo la interacción larva-adulto como factor regulador de la abundancia, incluyendo mecanismos compensatorios y sobre-compensatorios (Sale, 1982; Menge & Sutherland, 1987, Fogarty et al., 1991). En un contexto pesquero, el conocimiento de estas

66
66
interdependencias posibilitará la predicción de biomasa y por tanto los ingresos netos derivados de la actividad. Teniendo en cuenta las observaciones realizadas en el numeral anterior, es importante discernir entre diferentes escalas espacio-temporales de análisis (Orensanz et al., 1991). Esto es relevante si se consideran las bruscas variaciones de abundancia observadas en las especies que poseen ciclos de vida complejos (e.g. fase larval planctónica y vida adulta bentónica: ver Roughgarden et al., 1985; 1988). La estrategia de estudio a implementarse dependerá de la complejidad (sensu Cohen, 1978) de cada comunidad. Estudios experimentales recientes han demostrado que la estructura de una comunidad explotada difiere drásticamente de una prístina debido a que la explotación induce una modificación de la magnitud de las interdependencias ecológicas. En tal sentido, comunidades de macrofauna bentónica intermareal, protegidas o con menor interferencia antropogénica, han sido comparadas con aquéllas donde el hombre ha ejercido una persistente influencia en el sistema (Castilla & Durán, 1985; Hockey & Bosman, 1986; Moreno, 1986; Bosman et al., 1987; Castilla & Paine, 1987; Bustamante & Castilla, 1990; Hockey et al., 1988). Se ha podido demostrar que la influencia del hombre en comunidades marinas costeras ha repercutido fundamentalmente en las especies depredadas, pero también en forma indirecta en otros organismos. Se han determinado importantes efectos de cascada (Paine, 1984; Castilla, 1988; Estes & Palmisano, 1974; Moreno et al., 1984; 1986) debidos a la explotación de un depredador tope ó especie clave ("keystone species", sensu Paine, 1980). Estos efectos indirectos son mayormente visibles cuando los ligamientos entre especies son muy fuertes (Moreno, 1986; Durán et al., 1987). Como consecuencia de lo anterior, la respuesta de una comunidad a la presión pesquera dependerá de la magnitud de las interacciones entre las especies, así como del papel funcional de la especie objeto de explotación. En efecto, características del depredador, tales como su espectro trófico, selectividad (especialista ó generalista) y eficiencia de depredación, así como de las presas (si compiten ó no y si la competencia es simétrica o no) incidirán en la respuesta de la comunidad a la explotación y por tanto sus efectos tendrán muy disímiles consecuencias a nivel pesquero y ecológico. A modo de ejemplo, la depredación sobre un competidor dominante tenderá a incrementar la diversidad en una comunidad (Paine, 1966; Durán & Castilla, 1989), mientras que la explotación de especies claves puede conducir a una simplificación de la comunidad y disminución de la diversidad. Ambas situaciones muestran la existencia de múltiples estados de equilibrio en una comunidad (Sutherland, 1974; Connell & Sousa, 1983). De acuerdo a los niveles de explotación, la magnitud de las interdependencias pueden ser modificadas y el esquema de manejo y regulación de los recursos alterado (de Alava, 1993). Teniendo en cuenta las características de estas interacciones, los ingresos netos derivados de la explotación pesquera podrán ser maximizados inclusive sobreexplotando deliberadamente algunas especies a efectos de producir el aumento de un recurso altamente valuado en el mercado, pero cuya disponibilidad es limitada como consecuencia de fuertes interdependencias ecológicas. La minimización de los impactos potenciales adversos derivados de la explotación multiespecífica puede alcanzarse asimismo durante períodos de máxima separación ó bien optimizando el nivel de esfuerzo en el momento de la co-ocurrencia espacial (Murawski & Finn, 1989). Es importante remarcar que investigaciones recientes no integran al hombre como depredador especialista eficiente sino que consideran una conducta generalista a medida que se van imponiendo vedas y cuotas sobre los recursos tradicionalmente explotados (Smith & McKelvey, 1986).

67
67
Se ha demostrado la dificultad de separar y cuantificar la importancia relativa de los diferentes agentes perturbadores de una comunidad (e.g. aspectos tecnológicos y ecológicos: ver Sainsbury, 1988; Deriso et al., 1986) lo cual a su vez dificulta el manejo de recursos desde una visión multiespecífica. Las aproximaciones holísticas (e.g. ECOPATH: Christensen & Pauly, 1990) pueden resultar útiles en casos en que la contribución de las especies (e.g. valor presente esperado, ver Sainsbury, 1988) a una pesquería es similar, sin embargo la dinámica de la comunidad ante diferentes grados de perturbación es difícilmente estimable (Sainsbury, 1987). La agregación de especies a efectos de un análisis más simple requiere de supuestos o del conocimiento de la dinámica de los componentes de dichas agregaciones (Sugihara et al., 1984), lo cual limita el poder predictivo de dichas aproximaciones. Por tanto, la verificación de la ocurrencia de competencia y depredación, así como la magnitud de las interacciones e incidencia de la pesca, requerirá necesariamente de manipulación experimental en terreno (Cobb & Caddy, 1989; Defeo, en prensa), dado que las comunidades marinas sujetas a explotación no constituyen sistemas estables en equilibrio espacialmente homogéneos, tal cual asume la teoría (Larkin, 1978; Caddy, 1983; Beddington, 1984; Chesson & Case, 1986). Perspectivas No obstante el importante desarrollo teórico (e.g. Pope, 1979; Clark, 1985; Saila & Erzini, 1987; Ströbele & Wacker, 1991), el estado actual del conocimiento permite afirmar que no existen a la fecha modelos multiespecíficos capaces de generar resultados adecuados para llevar a cabo un manejo a largo plazo. Tal como lo expresa claramente Clark (1985), la pregunta actual no debe ser si los modelos multiespecíficos pueden ser construidos, sino si éstos serán de algún valor a la hora de proponer pautas para el manejo de recursos. Esta inquietud surge como respuesta a la común observación que los modelos actuales han logrado un impacto relativo en el manejo de recursos pesqueros (Ursin, 1982; Larkin, 1982; Brander, 1988; Cobb & Caddy, 1989; Cobb et al., 1986). Como resultado de la complejidad de la estructura de una comunidad, la predicción de los efectos de la explotación de una especie en la abundancia de otra ecológicamente interdependiente es difícil de determinar (Miller, 1985; Akenhead et al., 1982). Esto puede resultar de extrema gravedad si se considera que una severa sobrepesca en una especie puede ocasionar impactos de diferente magnitud en sistemas cuya estructura trófica, así como la potencia relativa de las interrelaciones horizontales (e.g. competencia) y verticales (e.g. depredación) se desconoce (Paine, 1980; Sainsbury, 1982). Al igual que en los modelos tradicionales, la incertidumbre y estocasticidad en los procesos de estructuración comunitaria deben ser incorporados en modelos holísticos determinísticos simples y de uso masivo (Arreguín-Sánchez et al., 1993). Modelos tales como el ECOPATH (Christensen & Pauly, 1990, 1993), podrían ser modificados a efectos de adecuar los principios generales de la bioeconomía de recursos, los cuales por el momento emergen en una forma más clara y definida a partir de los modelos analíticos simples (Clark, 1985). La dificultad de análisis de la composición de la dieta en especies que constituyen ensambles o comunidades bentónicas, en contraposición a las nectónicas, limitan un análisis estructural epifenoménico de las mismas, tales como las que pudieran derivarse de la aplicación de los modelos mencionados. Por tal razón se ha desarrollado una rutina complementaria a ECOPATH denominada ECOSIM (Walters et al., en prensa), mediante la cual se amplía la capacidad de efectuar predicciones que incluyan el efecto de la explotación pesquera. En este contexto, las ecuaciones lineales que

68
68
describen los flujos tróficos bajo condiciones de equilibrio en ECOPATH se modifican para definir interacciones dinámicas que varían con cambios en la biomasa de los recursos y en el régimen de explotación. La incorporación de ECOSIM en ECOPATH permite conducir análisis de alternativas de manejo considerando explícitamente las interacciones tróficas dentro del sistema. Los estudios biológicos básicos y experimentales, en diferentes escalas de espacio y tiempo, pueden proveer la única alternativa válida para predecir los efectos bioeconómicos derivados de posibles interdependencias ecológicas y tecnológicas. Es necesario el desarrollo de modelos que incluyan una correcta identificación del sistema, análisis de sensibilidad a modo de índices de desempeño del modelo e intervalos de confianza para las predicciones, así como una cuidadosa estimación de los parámetros de entrada al mismo (Majkowski, 1982; Silvert, 1981; 1982; Walters, 1986; Seijo, 1986; Clark, 1989; Seijo et al., 1994a). Tanto la pesquería como las interacciones ecológicas, el ambiente físico y el desarrollo dinámico del sector pesquero tienen efectos en el recurso, con un retraso diferencial en su impacto. En el estado actual del conocimiento, la alternativa más viable parece ser el manejo adaptativo (Walters, 1984; 1986; Hilborn & Walters, 1987), esto es, manejar a través de experimentación, cambiando deliberadamente los niveles de esfuerzo a efectos de entender la dinámica de la comunidad y la pesquería. Esta estrategia permite la inclusión de mecanismos de control y regulación a corto plazo en respuesta a un flujo continuo de información sobre variaciones en el estado de un recurso. Tal como lo expresan Cobb & Caddy (1989): "nuestra habilidad para modelar la dinámica de poblaciones explotadas ha sobrepasado nuestra
habilidad para verificar el realismo de esos modelos... Es tiempo ya de elaborar hipótesis contrastables a través de experimentación en terreno y estudios comparativos".
Los efectos de diferentes tipos de regulación, incluyendo asignación de cuotas de pesca, cuotas individuales transferibles, rotación de áreas de pesca, así como la variabilidad en el descarte producida por la puesta en práctica de algún tipo de medida especie-dirigida, deberán ser considerados a través de modelos de optimización con criterios múltiples (Seijo, 1986; Charles, 1989). En caso contrario, " el manejo de recursos multiespecíficos permanecerá probablemente como un arte, más que una
ciencia, y la asignación de altos presupuestos dirigidos a resolver problemas científicos relacionados con el manejo de pesquerías multiespecíficas será injustificado en términos de los beneficios potenciales que se lograrían" (Clark, 1985).

69
CAPITULO 4
Análisis Intertemporal de Pesquerías En el Capítulo 2 se desarrolló el modelo bioeconómico clásico estático, del cual se derivaron fundamentalmente dos predicciones claves: 1) el libre acceso a los recursos pesqueros ocasionará la sobreexplotación del recurso y disipación de los excedentes generados por una pesquería; y 2) el máximo rendimiento económico (MRE) ocurrirá a una tasa de explotación menor a la del máximo rendimiento sostenible (MRS). Sin embargo, este modelo ignora la dimensión del tiempo en la estimación de los niveles óptimos de captura y esfuerzo. En este Capítulo se describe el modelo bioeconómico dinámico, a través del cual se demuestra que la tasa óptima de explotación de un recurso pesquero puede ser mayor o menor al MRE ó al MRS, dependiendo de las preferencias intertemporales en el uso del recurso por parte de la sociedad. A efectos de lograr una visión más profunda sobre la importancia del tiempo en la toma de decisiones, este Capítulo desarrolla en su primera sección conceptos clásicos relacionados con la preferencia intertemporal en el uso de un bien. La segunda sección considera conceptos claves en la evaluación de aquellos proyectos pesqueros, fundamentalmente financiados por el sector público, en los cuales el factor tiempo adquiere una dimensión relevante. La tercera analiza los criterios de decisión fundamentales para la inversión pública, en el contexto del análisis beneficio-costo. La cuarta sección comprende el estudio de aspectos relacionados con las preferencias intertemporales e impactos distribucionales en la sociedad derivados de la ejecución de un proyecto pesquero. La quinta sección se aboca exclusivamente al desarrollo del modelo bioeconómico dinámico en pesquerías, integrando el cuerpo teórico de conocimiento de las secciones precedentes. En el transcurso del Capítulo se intercalan ejemplos relacionados a pesquerías, a fin de discutir aplicaciones particulares de los principios básicos del análisis beneficio-costo y del modelo bioeconómico dinámico.
Preferencias intertemporales Las decisiones sobre inversión en esfuerzo pesquero están relacionadas con la promesa que éste asegure ingresos netos positivos durante la vida útil de una unidad de pesca (e.g. embarcación, artes y equipos que la componen). Una aproximación al problema de la dimensión e importancia del tiempo como factor clave en la inversión y desarrollo de actividades pesqueras, es considerar las preferencias en el consumo de un cierto bien en diferentes períodos. A modo de ejemplo, considérese a un individuo en un cierto momento que tiene la alternativa de consumir bienes en el presente año y en los subsecuentes. La persona no necesariamente será indiferente a la elección de gastar ahora o en el futuro, aún si los precios de los bienes permanecen constantes. En efecto, el consumo en un período constituye un bien diferente a consumir en otro período. Cada individuo de la sociedad tiene preferencias temporales respecto al consumo de un bien en diferentes períodos de tiempo. Esto es medido por su tasa marginal de preferencia en el tiempo (TMPT). Si un individuo es indiferente entre consumir en

70
forma extra 1 US$ en un año o bien, 1.10 US$ en el siguiente, se dice que posee una TMPT de 10 % por año. La palabra marginal es usada debido a que una TMPT mide la preferencia individual entre pequeños incrementos de consumo en el tiempo. Esto presupone que el individuo tiene disímiles expectativas acerca del monto que consumirá en diferentes períodos. Los conceptos anteriormente vertidos remarcan un punto verdaderamente importante: la preferencia en el tiempo considera preferencias en el consumo de un bien. Esto lleva a profundizar en el concepto de preferencias intertemporales, que en el caso de recursos renovables como los pesqueros adquiere una dimensión fundamental. En efecto, por dos grandes razones el análisis de las preferencias en el uso de este tipo de recursos no puede ser estático: 1) su naturaleza renovable implica variabilidad en el monto de recurso disponible, así como un cierto grado de incertidumbre en cuanto a su magnitud en diferentes períodos de tiempo; 2) según el tipo de pesquería analizada, existirá una distinta preferencia marginal del uso de los recursos en el tiempo. Así por ejemplo, en una pesquería con sistema de libre acceso existirá probablemente una alta TMPT, dado que por lo general hay un mayor incentivo para incrementar el monto de ingresos en la actualidad con respecto al futuro debido a la naturaleza inherente a los recursos pesqueros (e.g. ocurrencia de externalidades; ver Capítulo 1). Considerando pesquerías con cierto grado de mecanización (e.g. pesquerías de mediana y gran escala), la inversión realizada en planificar y efectuar actividades de pesca no se paga de inmediato, sino que existe una vida útil de la unidad de pesca (buque, equipos y artes de pesca), que debe ser tomada en consideración para evaluar la magnitud de la inversión, costos presentes y futuros, así como los probables ingresos que se deriven de dichas actividades. En estos casos es probable que bajo marcos conservativos de manejo y administración pesquera (e.g. entrada limitada de buques) ocurra una baja TMPT, a efectos de favorecer inversiones y al mismo tiempo explotar el recurso durante un lapso prolongado en el tiempo. Preferencias neutras, positivas y negativas Siguiendo el análisis intertemporal reportado por Randall (1981), considere un pescador A que debe decidir cómo distribuir sus actividades de consumo en dos períodos de tiempo t1 y t2. Se asume que el consumo total en cada período (Ψt) puede ser definido por su presupuesto en cada uno, esto es, Ψt1 en t1 y Ψt2 en t2. Asúmase que el pescador recibe un ingreso de Y1 en t1 e Y2 en t2, siendo Y1 = Y2. La Figura 4.1 muestra la curva de indiferencia Ia que define las preferencias (e.g. necesidades) del pescador A en el uso de los recursos en dos períodos de tiempo sucesivos. WW es la línea intertemporal de presupuesto, donde W = Y1 + Y2. WW tiene una pendiente negativa igual a -1, por lo cual se asume que el ingreso podría ser transferido de un período a otro en una base uno a uno. La curva de indiferencia pasa a través de un punto común representado por Y1 e Y2. Se dice que el individuo A tiene una preferencia neutra del tiempo si prefiere el mismo consumo en t1 que en t2 (con Ψt1 = Ψt2 e Y1 = Y2).
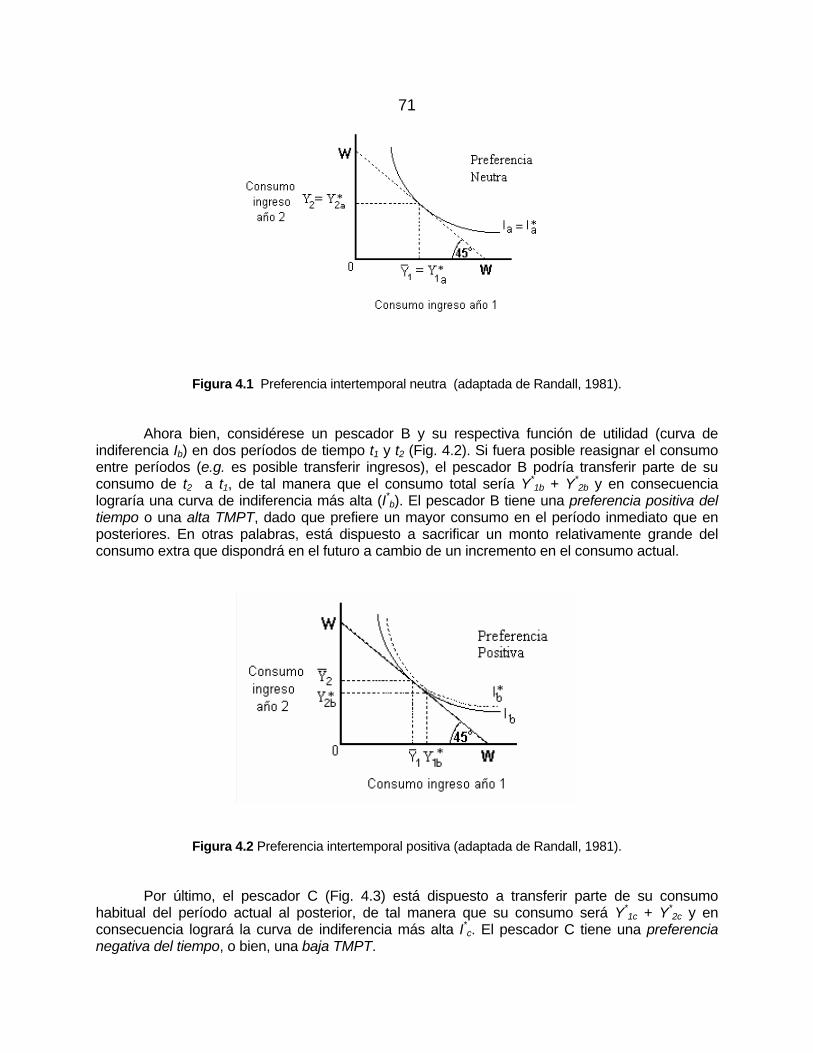
71
Figura 4.1 Preferencia intertemporal neutra (adaptada de Randall, 1981).
Ahora bien, considérese un pescador B y su respectiva función de utilidad (curva de indiferencia Ib) en dos períodos de tiempo t1 y t2 (Fig. 4.2). Si fuera posible reasignar el consumo entre períodos (e.g. es posible transferir ingresos), el pescador B podría transferir parte de su consumo de t2 a t1, de tal manera que el consumo total sería Y*
1b + Y*2b y en consecuencia
lograría una curva de indiferencia más alta (I*b). El pescador B tiene una preferencia positiva del tiempo o una alta TMPT, dado que prefiere un mayor consumo en el período inmediato que en posteriores. En otras palabras, está dispuesto a sacrificar un monto relativamente grande del consumo extra que dispondrá en el futuro a cambio de un incremento en el consumo actual.
Figura 4.2 Preferencia intertemporal positiva (adaptada de Randall, 1981).
Por último, el pescador C (Fig. 4.3) está dispuesto a transferir parte de su consumo habitual del período actual al posterior, de tal manera que su consumo será Y*
1c + Y*2c y en
consecuencia logrará la curva de indiferencia más alta I*c. El pescador C tiene una preferencia negativa del tiempo, o bien, una baja TMPT.

72
Figura 4.3 Preferencia intertemporal negativa (adaptada de Randall, 1981).
Inversiones públicas en el manejo de recursos pesqueros Evaluación de proyectos y el criterio Pareto-eficiencia El sector público está capacitado para llevar a cabo inversiones en el manejo, desarrollo y conservación de recursos naturales renovables como los pesqueros. En este contexto, el economista pesquero debe evaluar la viabilidad y factibilidad de aquellas inversiones en el sector que minimicen los requerimientos de subsidio. En la mayoría de los casos, los proyectos o programas dirigidos al manejo y administración de recursos pesqueros son de larga vida, por lo cual no sólo se debe realizar un análisis estático de su eficiencia (e.g. Pareto eficiencia, ver Capítulo 6), sino incluir aspectos que supongan diferentes preferencias intertemporales en el uso de los recursos por parte de distintos sectores de la sociedad. Un proyecto, definido en un sentido amplio, es una forma de usar recursos, por lo cual la decisión concerniente a tomarlo o desecharlo es una elección entre diferentes alternativas de uso de los recursos en el tiempo. En tal sentido, la ejecución del proyecto por parte de la agencia pública pesquera tendrá como objetivo clave lograr una mejora en el bienestar social. La idea de cuantificar y evaluar un objetivo social es a veces problemática, particularmente cuando los bienes en consideración no tienen o son inadecuadamente reflejados en el precio de mercado debido a sus características peculiares (e.g. arrecifes coralinos, manglares, reservas de la biósfera). Sin embargo, tales bienes reflejan las preferencias de la sociedad en la asignación de recursos de propiedad común por parte del Estado. Las herramientas más comunes en la evaluación de proyectos se basan en el análisis beneficio-costo (ABC) y métodos analíticos relacionados (ver Crooper y Oates, 1992). El ABC de un proyecto requiere la identificación de todos sus efectos en el bienestar individual de los miembros de una comunidad pesquera. Esos efectos son cuantificados a través de la comparación de los beneficios y costos derivados de la consecución del proyecto. El criterio más simple para evaluar diferentes alternativas, estrategias o proyectos es la Pareto-eficiencia intertemporal. Una estrategia o patrón del uso de un recurso en el tiempo es Pareto eficiente si y sólo si no es posible incrementar la utilidad de las partes afectadas en un período de tiempo sin disminuir las utilidades en otro período. Por medio de este mecanismo se identifican, miden y comparan cambios en el bienestar de una sociedad. En el lenguaje de la Teoría de Bienestar (ver

73
Capítulo 6), una mejora Pareto se producirá cuando una reorganización de la economía mejora el ingreso de un individuo sin perjudicar al resto de los miembros de la comunidad. En este contexto, un proyecto pesquero implicará una potencial mejora Pareto cuando la suma de dinero que los beneficiarios estarían preparados a pagar para garantizar que el proyecto sea llevado a cabo excede la suma total de dinero que los potenciales perjudicados aceptarían como compensación por su ejecución. En esencia, el criterio Pareto-mejora potencial ó Pareto eficiencia requiere que los cambios en el bienestar de una comunidad pesquera sean medidos por su disponibilidad a pagar, o sea, el monto de dinero que los individuos están dispuestos a erogar por los beneficios de un proyecto pesquero, así como por el monto que están dispuestos a aceptar como compensación por los daños impuestos a los grupos potencialmente perjudicados. De esto se desprende claramente que el criterio inherente en el ABC es el Valor Máximo del Producto Social y por tanto el daño económico es permisible si y sólo si la suma algebraica de los beneficios netos recibidos por los ganadores menos las pérdidas netas que enfrenten los perdedores sea mayor que cero y máxima. Valor presente y tasa de descuento El valor presente de un flujo de futuros ingresos es un concepto económico importante que permite realizar comparaciones monetarias en diferentes períodos de tiempo. En efecto, un igual monto de dinero a ser recibido ahora o en el futuro no significan lo mismo. En este contexto, la tasa de descuento (d) puede ser usada como un mecanismo para comparar el valor del dinero en el tiempo. Por ejemplo, supóngase que un individuo tiene una TMPT de d por período. En consecuencia es indiferente entre una propuesta de consumo extra de 1 unidad en el período 0 y otra de consumo 1 + d unidades en el período 1. En forma análoga, será indiferente a la elección de las propuestas anteriores con respecto a un consumo de (1 + d)² unidades extra de consumo en el período 2. Este razonamiento es similar a establecer que una unidad extra consumida en el período 1 tiene un valor presente de 1/(1 + d) unidades en el período 0; del mismo modo, una unidad extra consumida en el período 2 tiene un valor presente de 1/(1 + d)² unidades en el período 0, y así sucesivamente para t períodos (1/(1+d)t). La tasa d a la cual los ingresos futuros son descontados a valores presentes es denominada tasa de descuento. Es importante remarcar que este término, a diferencia del término TMPT, no implica una interpretación de la tasa a la cual hace referencia; una tasa de descuento es simplemente un número que es usado en una manipulación aritmética. En general, d es tratado como una constante. Conceptos relacionados, tales como el factor de descuento y factor de compensación, pueden ser empleados para cálculos de este tipo. El factor de descuento vt,d es el valor presente en el período 0 de una suma de una unidad acumulada en el período t, si la tasa de descuento es d. Aritméticamente el factor de descuento es descrito por un patrón geométrico regular de la forma:
t ,d tv1
(1+ d )= (4.1)
El factor de compensación ad es el valor presente en el período 0 de un flujo de ingresos consistente en la acumulación de una unidad en cada uno de los t períodos 1 a n, a una tasa de descuento d. Aritméticamente,

74
d ta1
(1+ d )= ∑ (4.2)
donde: t = 1, 2,...., n. En términos de factores de descuento:
d ( t ,d )a v= ∑ (4.3)
donde: t = 1, 2,...., n. Es importante remarcar que el factor de compensación es útil cuando el flujo de costos y beneficios a lo largo del tiempo es el mismo (ver Sugden & Williams, 1990). Este no es el caso de pesquerías. Costo de oportunidad de la inversión pública Dado que la evaluación de un proyecto involucra elecciones entre distintos cursos de acción, la utilidad de una estrategia puede ser determinada únicamente en relación a las alternativas posibles. En consecuencia, el costo de oportunidad de un proyecto es lo que se dejó de ganar en la siguiente mejor alternativa por haber adoptado el proyecto pesquero en cuestión (Shim & Siegel, 1995). De manera análoga, el retorno de un proyecto es el monto extra que se hubiera ahorrado si es puesto en marcha, en relación al generado si no es llevado a cabo. Es importante remarcar que en la evaluación de proyectos, los costos y ingresos son relacionados con las estrategias o cursos de acción alternativos, lo cual implica una concepción diferente en la estimación de costos con respecto a la aproximación financiera, donde se estiman en base a cosas concretas (e.g. infraestructura, horas - hombre de trabajo). Varias razones explican esta diferente aproximación metodológica, las cuales serán desarrolladas a continuación: Depreciación El ABC no incluye costos de depreciación como costos reales, en razón de que no hay un costo de oportunidad involucrado. En efecto, éste se relaciona con el valor de los bienes de capital en la siguiente mejor alternativa. A modo de ejemplo, supóngase que una unidad de pesca desea concluir sus actividades de pesca. Dicha unidad habrá sido pagada en el pasado, considerando un precio de depreciación por unidad/año calculado por métodos convencionales. Sin embargo, esta estimación carece de directa relevancia a la hora de decidir entre concluir o no las actividades pesqueras. El costo de oportunidad de usar la unidad de pesca será el valor que tiene para el pescador en su siguiente mejor alternativa. Tal como se expondrá posteriormente, los criterios de decisión que sustentan el uso del ABC muestran: 1) el valor actual de los excedentes generados por encima de los que se hubiera logrado si se invirtiese a una tasa marginal de interés; y 2) que el procedimiento de descuento automáticamente da cuenta de la recuperación del capital inicial. En razón de estas consideraciones, cuando se incluyen en el tiempo cero (al

75
inicio del proyecto) los costos de inversión, el flujo de costos de cualquier ABC no debe incluir ajustes por depreciación. Precios "sombra" Los precios que el mercado otorga a cierto bien representan las preferencias y la habilidad de los individuos de comprar y vender bienes libremente en el mercado. Esto es, los precios que establece el mercado son apropiados en la medida en que éstos denoten las preferencias de la sociedad y la libertad de los individuos en consumir determinada cantidad de cierto bien a un precio fijo establecido. Este estará determinado por el nivel en el cual la demanda y la oferta se igualan; se asume que el precio de un bien es igual al valor de su producto marginal. Cuando estos supuestos no son cumplidos, el precio que el mercado otorga a cierto bien no será necesariamente una buena aproximación de los costos sociales marginales del bien o servicio en cuestión. En este caso, en el ABC deben emplearse valores a efectos de tasar un bien que no son tomados directamente de precios reflejados por el mercado, los cuales son denominados precios sombra. El precio sombra es un valor impuesto a un bien o servicio que carece de precio en el mercado (Pearce, 1981) o que es incorrectamente representado en éste. Los precios sombra son utilizados en ABC para representar el costo de oportunidad de producir o consumir un bien que generalmente no es comercializado en el mercado. Ejemplos típicos de esto lo constituyen especies en peligro de extinción, parques o reservas marinas, cierto nivel de calidad ambiental, o funciones bióticas y abióticas de un ecosistema específico. Los precios sombra pueden ser estimados a partir de los costos marginales de producción o valor marginal de su uso como entradas al ABC.
Criterios de decisión para la inversión pública pesquera A continuación serán desarrolladas tres alternativas diferentes factibles de ser seleccionadas como criterios de decisión: valor presente neto (VPN), cociente beneficio-costo (B/C) y tasa interna de retorno (TIR). Valor presente neto (VPN)
Como se ha explicado anteriormente, un cambio en determinada regulación gubernamental, al igual que la puesta en marcha de un proyecto pesquero de interés social, genera un flujo de efectos positivos (beneficios) y negativos (costos) a través del tiempo. La diferencia entre las tasas de beneficios y costos en un momento dado es denominada tasa de beneficio neto. Visto en términos de intervalos de tiempo discretos (e.g. años), el concepto de valor presente neto (VPN) puede expresarse de la forma siguiente:

76
VPNBT t CT t
(1+ d )t= ∑−( ) ( )
(4.4)
donde BT(t) y CT(t) son los beneficios y costos totales en el tiempo t. Este mecanismo permite la evaluación de diferentes alternativas o proyectos a través de un valor único, independientemente de la forma en que los costos y beneficios están distribuidos en el tiempo, lo cual posibilita la comparación de flujos de beneficios netos en el tiempo y facilita la decisión a ser tomada entre aquellos proyectos que produzcan diferentes flujos de beneficios netos. Es importante remarcar que la decisión dependerá esencialmente de la elección de la tasa de descuento d. De (4.4) se desprende que d tiene una influencia sustancial en las magnitudes calculadas de VPN y relativa en los ingresos netos logrados en períodos distantes de tiempo. A mayor valor de d, menor será el VPN. Un bajo valor de d tenderá a favorecer aquellos proyectos que generan mayores beneficios netos positivos en el largo plazo, pues son menos penalizados al ser traídos a valor presente que en el caso de haberse utilizado una d mayor. Considerando al sector público como agente de inversión, es apropiado emplear estimaciones de d menores a aquéllas generadas por el mercado, aunque en general se utiliza la tasa de interés bancaria pues refleja de un modo u otro las preferencias de la sociedad. Tal como ya se ha mencionado, el objetivo de una agencia pública consiste en minimizar el valor presente de los requerimientos de subsidios netos en el tiempo, descontados a una tasa de descuento predeterminada por el Gobierno, ó "tasa de descuento testigo". A este nivel, la analogía entre las agencias públicas y privadas se rompe, dado que es muy difícil infundir un criterio comercial a la agencia pública, de tal forma que elija una correcta tasa de descuento. Por ello es preferible emplear diferentes tasas de descuento que reflejen distintas preferencias en el uso de los recursos por parte de la sociedad. Como regla de decisión, se considera que un proyecto o alternativa es ejecutable si el VPN es mayor a cero, y en consecuencia la inversión es aconsejable. Esta regla es mayormente viable en caso de no haber restricciones financieras. Ahora bien, dado que en la mayoría de los países, en especial en aquéllos en vías de desarrollo, se suscitan restricciones financieras, la regla de decisión será seleccionar aquella alternativa que maximice el bienestar social y que al mismo tiempo tenga el máximo VPN (mayor que cero). Este criterio puede darse también en presencia de proyectos mutuamente excluyentes. Dado que, tal como se ha expresado anteriormente, el criterio inherente a esta selección es el de valor máximo del producto social, supone implícitamente daño económico, por lo cual es recomendable identificar a los beneficiarios y perjudicados de la ejecución del proyecto, mostrando los impactos distribucionales en la sociedad. Cociente beneficio-costo (B/C) El cociente beneficio-costo (B/C) está dado por la ecuación:

77
VPBVPC
BT t(1+ d )CT t
(1+ d )
t
t
=∑
∑
( )
( ) (4.5)
donde: VPB y VPC son el valor presente de beneficios y costos respectivamente. El criterio de decisión en este caso será llevar a cabo el proyecto si el cociente es mayor a uno, mientras que si son varias alternativas se escogerá aquélla con mayor valor de dicho cociente. Tasa interna de retorno (TIR) La tasa interna de retorno (TIR) de un proyecto es definida como la tasa de descuento que hace el VPN igual a cero. O sea:
∑ =BT t - CT t
(1+ )0t
( ) ( )ρ
(4.6)
siendo ρ la TIR. Análogamente, ρ puede ser calculada llevando el valor descontado del flujo de beneficios netos igual al desembolso de capital inicial y resolviendo la ecuación resultante para el valor de la tasa de descuento. Esto es, una ecuación de la forma
[ ]∑ − =BT t CT t1
(1+ d )Ct 0( ) ( ) (4.7)
es resuelta para d, siendo C0 el capital inicial. La ecuación (4.7) puede ser expresada en la forma:
[ ] [ ]∑ = ∑BT t1
(1+ d )CT t
1(1+ d )t t( ) ( ) (4.8)
De (4.7) y (4.8) se obtendrá el mismo resultado, ya sea si se considera el valor presente del flujo de beneficios netos con respecto al capital inicial desembolsado, o bien el flujo de beneficios totales con respecto al flujo de costos totales. En ambas ecuaciones d es desconocido. El valor crítico de d que hace cero las ecuaciones (4.6) a (4.8) es la TIR (ρ). No existe un mecanismo simple para calcularla; sin embargo, una probable aproximación consiste en proponer una solución iterativa en base a la estimación de los valores presentes de costos y beneficios. Si el valor presente de éstos excede al de los costos, entonces la tasa escogida es demasiado pequeña y debe ser aumentada, y así sucesivamente. Un proyecto es aceptado cuando ρ es mayor o igual a un predeterminado valor de d que en general es la tasa de interés bancaria. En caso de proyectos mutuamente excluyentes o en

78
presencia de restricciones financieras, se seleccionará el proyecto con máximo valor de ρ. Estas pueden ser consideradas como reglas de decisión. ABC en la práctica: la elección del criterio adecuado El criterio de decisión para la inversión pública pesquera está basado en lograr la eficiencia económica en la asignación intertemporal de recursos con respecto a aquélla que hubiese logrado el sector privado. El argumento clave en este sentido ha sido considerar que una asignación eficiente de recursos en el tiempo será lograda si las tasas marginales de retorno en el uso de un recurso son iguales en los dos sectores. Cuando el sector público financia sus programas a través de préstamos ó impuestos provenientes del sector privado, las tasas de retorno derivadas de las inversiones no realizadas por estas causas, así como aquéllas inferidas de la preferencia del tiempo en el consumo no efectuado, pueden ser consideradas como el costo de oportunidad de los recursos canalizados a la inversión de dichos programas. El argumento de eficiencia económica en este caso es que se ahorraría una tasa de retorno en los servicios aportados por el sector público al menos igual a su tasa de retorno en los servicios generados por el sector privado. Esto puede ser afirmado en base a la siguiente regla de inversión en el sector público: aceptar sólo aquellos programas que exhiben un VPN positivo, usando una tasa ponderada del costo de oportunidad del sector privado como tasa de descuento. Esto aseguraría una eficiente asignación de recursos entre los sectores público y privado. Ahora bien, el VPN no provee información directa acerca de los requerimientos de capital de un proyecto en particular. Por ejemplo, si el proyecto A tiene un VPN apenas mayor que otro B, pero mucho mayor requerimiento de capital inicial, no es claro que el proyecto A tenga mayor prioridad que B en presencia de restricciones financieras. Tampoco una priorización por magnitudes en el caso del cociente beneficio-costo resulta estrictamente válido, dado que B/C es un cociente entre beneficios y costos promedio y no marginales, los cuales constituyen la única alternativa posible de optimización. En el caso de la TIR, tampoco la magnitud de ρ es un criterio válido para seleccionar proyectos en presencia de restricciones financieras. En efecto, dicho criterio es aplicable sólo en proyectos de inversión, donde los costos ocurren en forma temprana en el tiempo, mientras los ingresos son logrados a posteriori. En cambio no es aplicable a proyectos de desinversión, donde se producen beneficios a corto plazo y aumento de costos en el tiempo. Dado que altas tasas de descuento tenderán a favorecer aquellas alternativas con ocurrencia tardía de costos, cuanto más alta sea la tasa de interés, mejor considerado será el proyecto de desinversión. En consecuencia el valor crítico de ρ será el menor valor de la tasa de interés que justifique su ejecución. En otras palabras, un proyecto de desinversión sería ejecutable sólo si el verdadero valor de la tasa de interés es mayor que ρ, lo cual contradice el concepto de TIR. Una de las críticas de mayor peso efectuadas a la TIR es que su aplicación es problemática cuando se comparan proyectos cuyos flujos de costos y beneficios son muy diferentes en el tiempo, o sea, se intercalan períodos donde los beneficios son mayores a los costos con otros donde ocurre el caso inverso. Dado que la TIR es resultado de la solución de una ecuación polinomial, es factible en el caso indicado obtener más de un valor de ρ posible. En efecto, si el polinomio es de grado n, habrán n raíces, e.g. n valores de ρ probables. Por ejemplo, si la TIR de un proyecto tiene dos soluciones, 10% y 15%, y la tasa social de descuento es 12%, no hay un criterio evidente de

79
aceptación ó rechazo. Por otra parte, la determinación de una TIR óptima cuando hay restricciones financieras no es muy sencilla, y no equivale a ρ = d, sino a aquél incremento de ρ que iguala a los incrementos de ρ para otros proyectos. La TIR pondera en mayor grado a proyectos que acumulan sus beneficios en las primeras etapas de su ejecución con respecto a otros que generan beneficios en forma tardía en el tiempo. En efecto, 1 US$ invertido ahora tendrá una TIR de 100% si se acumulan 2 US$ al final del año. Una inversión de 10 US$ que acumule 15 US$ al final de año tendrá una TIR de 50%. El empleo de la TIR como criterio de decisión favorecerá la primera alternativa, aún cuando a una tasa de descuento del 10% el VPN será respectivamente 0.8182 y 3.6364. El uso de la TIR es complicado cuando se comparan proyectos mutuamente excluyentes, dado que no necesariamente el mejor proyecto es aquél que posee TIR más elevado. Considere los dos proyectos X e Y que se muestran en la Tabla 4.1 (Pearce & Nash, 1981), ambos con una vida de 10 años. Basándose en el TIR como regla de decisión, X es preferible a Y; sin embargo, de acuerdo a la regla de VPN, Y es preferible a X.
Tabla 4.1 Costos, beneficios y reglas de decisión de dos proyectos con similar vida de ejecución. Se asume una tasa de descuento d del 8 % (Pearce & Nash, 1981).
Proyecto Costo Beneficio TIR VPN X 1 0.20 15 0.34 Y 2 0.36 12 0.42
“Y-X” 1 0.16 9 -- En este caso la TIR como regla de decisión es falsa, dado que discrimina en contra de Y debido a la magnitud de capital desembolsado. Para evitar este problema, es necesario calcular la tasa de retorno en un proyecto hipotético "Y - X", o sea, basado en la diferencia entre capitales. Dado que la TIR en "Y - X" es mayor que el valor de d usado en el ejemplo, Y es preferible a X. En consecuencia, el contexto de exclusión mutua requiere de una regla de dos partes a efectos de que el proyecto Y sea aceptado: 1) TIRY > d y además: 2) TIR(Y-X) > d donde d es un valor predeterminado. Esta regla ha sido usualmente descrita como la "tasa de retorno sobre costo" de Fisher (Pearce & Nash, 1981). Del desarrollo de este Capítulo, resulta evidente que la TIR es un criterio de decisión más controvertido, ambiguo y con menor aplicabilidad que VPN y B/C. Sin embargo, las limitaciones que en común poseen los tres criterios surgen a raíz de su propia simplicidad, de su intento de

80
resumir una gran cantidad y variedad de información en un simple número. Como respuesta a esto, ninguna de las tres aproximaciones provee un “ranking” estricto en la eventualidad de existir varios proyectos. Dificultades similares surgen a la hora de seleccionar el tamaño óptimo de un proyecto, dado que dicha estimación requiere del cálculo del VPN, B/C y TIR para cada incremento en el tamaño de cada proyecto en consideración (e.g. desarrollo de análisis marginales). Los autores sugieren por tanto el uso de VPN o B/C como criterios de decisión. La elección entre éstos provendrá de una decisión alternativa: saber cuando B/C >0 contiene más o menos información que B/C >1. Una aplicación práctica en pesquerías: análisis discreto Un responsable de administrar recursos pesqueros está analizando tres alternativas de artes de pesca A, B y C, factibles de ser empleadas para desarrollar una pesquería de una especie de langosta en el Estado de Yucatán (México). Se le ha solicitado al especialista en bioeconomía pesquera que estime el VPN y el B/C de dichas alternativas, explicar sus resultados al administrador pesquero y efectuar recomendaciones con respecto a la mejor alternativa de arte de pesca. El flujo temporal de costos y beneficios (en millones de dólares) se presenta a continuación: Tabla 4.2 Flujo de costos y beneficios para cada una de las alternativas de artes de pesca A, B y C.
Año Costos Beneficios A B C A B C 0 4.0 2.0 1.0 0.0 0.0 0.0 1 8.0 4.0 2.0 7.0 4.0 6.0 2 6.0 4.0 4.0 7.0 5.0 7.0 3 4.0 4.0 6.0 8.0 6.0 8.0 4 3.0 4.0 8.0 9.0 7.0 9.0 5 2.0 4.0 8.0 8.0 9.0 8.0
Resultados En el ABC se ha optado por incluir 3 valores distintos de la tasa de descuento d a fin de reflejar diferentes preferencias intertemporales en el uso del recurso. El VPN fue estimado para cada alternativa de arte de pesca en particular, en base a la maximización del valor presente de los beneficios netos dada por la ecuación (4.4):
VPN = BTo - CTo + BT - CT(1+ )
+ BT - CT(1+ )
+...+ BT - CT(1+ )
1 1
1
2 2
2
5 5
5d d d (4.9)
donde: BT(t) son los beneficios totales en los años 1 a 5. CT(t) son los costos totales en los años 1 a 5.

81
d es la tasa de descuento (5%, 10% y 15%). Un criterio análogo se siguió con B/C, en este caso estimado en base a (4.5), tomando en cuenta el flujo de beneficios y costos en los 5 años. Los resultados derivados del análisis se muestran a continuación: Tabla 4.3 VPN y B/C para cada una de las alternativas de artes de pesca A, B y C, evaluadas con diferentes tasas de descuento.
ALTERNATIVA 5% 10% 15% VPN B/C VPN B/C VPN B/C
A 9.06 1.369 6.76 1.299 4.93 1.237 B 7.03 1.363 5.48 1.319 4.29 1.358 C 8.08 1.329 7.30 1.347 6.64 1.363
Tal como se ha argumentado anteriormente, el criterio de VPN maximiza la diferencia entre costos y beneficios, por lo cual si es mayor que cero, las alternativas son factibles de ser desarrolladas con éxito. Sin embargo, asumiendo restricciones financieras y/o que los proyectos son mutuamente excluyentes, debe hacerse un análisis comparativo de las distintas alternativas. Dicho análisis muestra que la alternativa C maximiza la relación beneficio-costo en base al empleo de tasas de descuento del 10% y 15%, y la oportunidad de llevar a cabo el proyecto de desarrollo es de 7.3 y 6.6 millones de US$ al valor presente (e.g. año cero) respectivamente. Para el caso del empleo de una tasa de descuento del 5%, el VPN se maximiza en la alternativa A, dado que otorga mayores beneficios netos al presente (9.06 millones US$). Por su parte, el análisis del cociente beneficio-costo muestra que todos los valores obtenidos de B/C son mayores a 1, por lo cual las alternativas son factibles de ser desarrolladas con éxito. Asumiendo proyectos mutuamente excluyentes, el análisis comparativo muestra que la alternativa C maximiza el cociente beneficio-costo en base al empleo de tasas de descuento del 10% y 15%. El empleo de una tasa de descuento del 5% maximiza la alternativa A, dado que otorga mayores beneficios netos al presente. De acuerdo a los resultados y análisis comparativo anteriormente expuesto, la selección del arte de pesca es sensible a la tasa de descuento empleada. La alternativa C sería la adecuada en caso de existir una alta preferencia intertemporal por el recurso, dado que optimiza el VPN y B/C para tasas de descuento mayores (10% y 15%). Estas a su vez tenderán a favorecer aquellas alternativas con ocurrencia de costos relativamente tardía y beneficios lográndose en el corto plazo. En el caso hipotético que la especie de langosta fuera de ocurrencia ocasional e impredecible, se le recomendaría al administrador del recurso utilizar el arte de pesca C para desarrollar la pesquería. Si por el contrario, los estudios de la biología y dinámica poblacional de la especie muestran que es de larga vida, con cierta constancia en la disponibilidad de biomasa a ser extraída, y que por otra parte se desea favorecer inversiones y explotarlo a largo plazo, el arte de pesca A junto al empleo de una tasa de descuento del 5% sería el curso de acción adecuado, dado que dicha estrategia otorga mayores beneficios al presente con una baja preferencia intertemporal. Por tanto beneficia a aquellos usuarios, presentes o futuros, que desean conservar el recurso a fin de favorecer su óptima explotación.

82
Evaluación de proyectos pesqueros: preferencias intertemporales e impactos distribucionales
La importancia de los impactos distribucionales Tal como se ha expresado anteriormente, el ABC es aplicado a problemas asociados con las preferencias intertemporales de la sociedad como un todo, para lo cual se aplica el criterio Pareto-eficiencia. Dicho criterio provee las herramientas adecuadas para hacer afirmaciones acerca de la contribución de un proyecto a la eficiencia económica y bienestar social, pero no provee ninguna guía sobre la diversidad de efectos del proyecto en la distribución del ingreso. En este entorno juega una decisión clave el administrador ó tomador de decisiones, quien debe escoger entre las distintas alternativas y en consecuencia hacer juicios de valor acerca de las diferentes formas en las que se plantea la distribución del ingreso. En este contexto, un ABC puede ser considerado como compuesto de dos etapas distintas: a) la primera consiste en evaluar, en términos monetarios, las ganancias y pérdidas generadas en cada uno de los sectores de la sociedad afectados por el proyecto. b) La segunda etapa consiste en combinar dichos flujos costo-beneficio en una cuantificación simple que represente de un modo u otro los cambios en el bienestar social. Es en esta etapa en la cual se integran al análisis juicios de valor sobre la distribución del ingreso derivados de la consecución del proyecto. El criterio de Pareto-eficiencia, que implica que los beneficios y costos son ponderados en una simple operación algebraica, es sólo un juicio. Sin embargo, la forma más conveniente de expresar dichos juicios es a través del uso de coeficientes de distribución. Supóngase que el efecto de un proyecto produce cambios en los beneficios netos de dy1, dy2 ,...., dyn a cada uno de los miembros de la comunidad 1, 2, .... , n. El cambio neto en el bienestar social dW es una suma ponderada de las pérdidas y beneficios individuales:
dW = w dy + w dy +...+ w dy1 1 2 2 n n (4.10)
En este caso, cada uno de los valores de w denotan los coeficientes de distribución, expresando los valores relativos sociales de incremento en el ingreso de los diferentes individuos. Un beneficio de 1 US$ en el individuo 1 tiene la misma significación en el bienestar social que uno de w1/w2 US$ para el individuo 2. En este contexto, el criterio de Pareto-eficiencia implica que ocurra un conjunto particular de valores de w:
1 2 nw = w =...= w = 1 (4.11)
Estos valores son denominados coeficientes de distribución unitarios.
dU =UM dy +UM dy +...+UM dy1 1 2 2 n n (4.12)
Los coeficientes de ponderación expresan de manera algebraica juicios de los administradores acerca de la redistribución del ingreso en los distintos sectores sociales como consecuencia de la puesta en marcha de un proyecto. En estos términos, el objetivo propio de un

83
gobierno es maximizar la suma de las utilidades correspondientes a cada uno de los miembros de la comunidad. Suponiendo que un proyecto produce beneficios netos (en dinero) dy1, dy2, .... , dyn a cada uno de los miembros de la comunidad 1, 2, .... , n, se generará una utilidad marginal del ingreso (UMn), esto es, la utilidad extra conferida a un aumento en el ingreso de 1 US$. El cambio neto en la utilidad total, o bienestar social, es entonces una suma ponderada de los cambios en el ingreso de los individuos; la ponderación en cada caso estará dada por la respectiva utilidad marginal en el ingreso. Dado que un incremento en una misma cantidad de dinero significa una utilidad marginal mucho mayor para un individuo con bajos ingresos, cuanto más bajo sea el ingreso promedio de determinado núcleo social, mayor será su coeficiente de distribución. En la práctica, la adopción de distintos coeficientes de distribución (e.g. no unitarios) dificulta en grado sumo al ABC, debido a que la información tradicionalmente recolectada (e.g. precios del mercado, cantidades de bienes consumidos y vendidos) permite la determinación del excedente del consumidor y productor en forma agregada, o sea, mide beneficios o pérdidas totales de los consumidores en general y asume implícitamente coeficientes de distribución unitarios. Es muy difícil obtener información para aplicar coeficientes de distribución ponderados por núcleo social, inclusive por la dificultad inherente a la propia identificación de dichos grupos. Otra dificultad radica en que los beneficios y costos sociales netos a partir de un cambio en el precio de un bien pueden ser calculados únicamente a partir de su curva de demanda, y este concepto está íntimamente ligado al concepto de Pareto-eficiencia, e.g. coeficientes unitarios de distribución. La tasa social de descuento y preferencia social del tiempo El ABC es aplicado a problemas que incluyen la dimensión del tiempo como factor clave. Debe escogerse entre consumir en la actualidad o en períodos subsiguientes, a través de la aplicación de la tasa de descuento "apropiada", que en este caso es la tasa social de descuento. Esta debe ser determinada a partir de las preferencias individuales de cada uno de los miembros de la sociedad. Estos podrían tener la misma TMPT, la cual reflejaría la tasa de interés en el mercado. En la práctica no existe ninguna tasa de interés patrón, sino que reproducen de una u otra manera la coyuntura económica, niveles de riesgo e incertidumbre asociados a la estabilidad política y desarrollo económico de un país. Por tanto, es más razonable asumir que los individuos que componen una sociedad no poseen la misma preferencia del tiempo, hecho que se agudiza si se consideran niveles distintos de ingreso e inclusive la propia expectativa de vida de cada individuo. Como resultado de estas disimilitudes, se hace necesario arribar a una tasa que de una u otra forma refleje las preferencias de la sociedad en su conjunto. Así habrán beneficiados y perjudicados como resultado de la ejecución de un proyecto (en el entendido que una evaluación personal del proyecto puede derivarse en un VPN negativo si se tiene en cuenta su propia TMPT). En este contexto, el Gobierno, representado por los administradores y tomadores de decisión, debe escoger dicho valor, para lo cual es factible seguir dos aproximaciones diferentes: la primera se basa en lograr cierta concordancia entre las tasas de descuento generadas en los sectores público y privado; la segunda se basa en el concepto de preferencia social del tiempo.

84
En el primer caso, se ha enfatizado que es posible interpretar las decisiones tomadas por firmas privadas en la evaluación de un proyecto como si proviniesen de un ABC. Esto es, en vez de considerar a las reglas de decisión de las empresas como provenientes de un análisis financiero (considerando una tasa de interés del mercado), tomar dichos resultados como si fuesen costos y beneficios sociales en función de una tasa social de descuento implícita. La toma de decisiones en el ámbito privado podrá ser considerada consistente para las agencias públicas siempre que la tasa social de descuento usada por éstas para un ABC sea igual a la tasa social de descuento implícita usada en el sector privado. Dicha tasa es también denominada costo de oportunidad social del capital, en razón de que mide el costo de oportunidad social de derivar recursos invertidos por el sector privado. La segunda alternativa supone el concepto de preferencia social del tiempo, la cual considera las preferencias temporales de la sociedad en el consumo, o sea, la cuantificación de efectos relativos en el bienestar de la sociedad a partir de cambios en el consumo en diferentes años. La selección de una tasa de descuento social (única) involucra un juicio de valor por parte de quien la selecciona. Este es en general el tomador de decisiones como puede ser el administrador pesquero, y su elección dependerá en gran medida de la coyuntura socio-económica, volumen de empleo complementario y disponibilidad de alimento entre otros factores. Sin embargo, la elección de un valor adecuado surgirá de un compromiso entre factores biológicos y socio-económicos (ver Sudgen y Williams, 1978). Es común la tendencia a identificar valores altos de d como sinónimo de "anticonservación", así como bajos valores a inversión y explotación sostenible en el largo plazo (Clark, 1985). La importancia de este principio como herramienta para un adecuado manejo de recursos renovables ha sido ampliamente analizada en la literatura (e.g. Clark, 1976; Dasgupta y Heal, 1979). Se ha argumentado que en el caso de recursos renovables en los cuales la sociedad como un todo es responsable de su conservación (no sólo los usuarios actuales sino también las futuras generaciones), el empleo de una tasa de descuento cero (Page, 1976) o muy pequeñas, sería una adecuada aproximación como tasa social de descuento. Dadas las características inherentes de los recursos pesqueros y su libre acceso a la explotación (ver Capítulo 1), así como su variabilidad en diferentes fases del sistema (e.g. recurso + impacto humano), la divergencia "micro-motivos macro-resultados" también se refleja a la hora de definir la tasa social de descuento adecuada: para los pescadores, a quienes concierne en mayor grado el corto plazo y que están sujetos a un fuerte componente de incertidumbre en su actividad, la elección de altos valores de d es preferible. Por el contrario, los biólogos pesqueros y administradores del recurso tenderán a suponer bajos valores de d con criterios de largo plazo (e.g. conservación). Estos conceptos realzan la necesidad de emplear análisis dinámicos que tomen en cuenta la preferencia del tiempo para el manejo de recursos pesqueros. Riesgo e incertidumbre en proyectos pesqueros A efectos de abordar este tópico, es importante distinguir los conceptos de riesgo e incertidumbre (Pearce & Nash, 1981). La existencia de riesgo en la ocurrencia de un evento implica que hay cierto conocimiento de las probabilidades de llevarlo a cabo, mientras que la incertidumbre supone variabilidad que no puede ser cuantificada debido al desconocimiento de dichas probabilidades de ocurrencia. A continuación se analizan brevemente ambos conceptos en el

85
contexto del ABC en pesquerías, ya que en el Capítulo 8 se discuten más a fondo en el marco del manejo precautorio de pesquerías. Riesgo La ejecución de un proyecto pesquero lleva asociado en general un componente de riesgo, más si se están considerando recursos naturales renovables como los pesqueros. Los criterios de decisión se verán afectados por dicho componente, en el sentido que los resultados de un ABC derivados del análisis de distintas alternativas podrán ser diferentes a los teóricamente esperados de acuerdo al clásico criterio determinístico presente en las reglas de decisión del ABC. En efecto, es probable que ocurra más de un valor de VPN en el caso de analizar posibles estrategias de desarrollo de un plan pesquero. En este caso, será en mayor grado apropiado considerar los resultados de un proyecto en términos de distribución de probabilidades de ocurrencia de las alternativas y seleccionar los cursos de acción adecuados en base a la media ponderada derivada de dicha distribución probabilística. Las decisiones alternativas pueden ser el resultado de la comparación de diferentes proyectos o bien de diferentes opciones de manejo o desarrollo para un mismo proyecto. En todos los casos deberá especificarse claramente la preferencia del tiempo (TMPT) asociada a la probabilidad de ocurrencia de cada alternativa (Schmid, 1989). A modo de ejemplo, supóngase que la magnitud de capturas de un recurso (y por tanto el flujo de ingresos totales asociados a un ABC) está asociada a la ocurrencia de diferentes condiciones ambientales, tales como magnitud de precipitaciones. La probabilidad de éxito del proyecto estará asociada a la ocurrencia de n alternativas posibles pero mutuamente excluyentes para desarrollarlo. La probabilidad de ocurrencia de cada alternativa es P1, P2 ,..., Pn. Si se considera que el resultado de cada una genera un beneficio neto de x1, x2 ,..., xn, el valor esperado de dichos beneficios (VE) será:
VE = P x + P x +...+ P x1 1 2 2 n n (4.13)
Esta evaluación ponderada de la utilidad de distintas alternativas se denomina criterio de valor esperado (VE), y puede resultar útil a efectos de atenuar el riesgo derivado del determinismo inherente al ABC. El VE puede constituir un criterio de decisión aceptable si, dada una serie de observaciones realizadas sobre el comportamiento del recurso en función de ciertas variables (e.g. climáticas) y estrategias de manejo, se asume como probable que los beneficios netos respondan al promedio de las expectativas generadas en dichos casos. El uso del VE como regla de decisión supone implícitamente que la variabilidad derivada de una serie estadísticamente consistente de observaciones sea despreciable. En caso de que no existan muchas alternativas o que la serie de observaciones no sea numerosa, es adecuado considerar la variabilidad de cada estrategia asociada al criterio de VE. Incertidumbre Se ha enfatizado anteriormente que existen situaciones donde el riesgo inherente a llevar a cabo ciertas alternativas existe, sin embargo se conoce la distribución de probabilidades de su

86
ocurrencia. En el manejo de pesquerías este caso ocurre sólo eventualmente. La situación que prevalece en mayor grado es un importante grado de incertidumbre acerca de la ocurrencia de dichas probabilidades, así como en lo referido a características del stock, magnitud del esfuerzo pesquero, proyecciones de costos y precios. La evaluación de un proyecto pesquero entonces llevará asociado en general un componente de incertidumbre. En efecto, en el ABC dirigido a evaluar la factibilidad de desarrollo de actividades pesqueras debieran tomarse en consideración dos componentes importantes de incertidumbre: a) Incertidumbre inherente al ABC. El desarrollo de las Secciones anteriores muestra que los criterios de decisión del ABC son determinísticos. Esto es, la viabilidad o factibilidad de un proyecto es definida a través de un valor único, sin especificar el grado de variabilidad asociado a su ejecución. Esto se debe a que los resultados obtenidos de un ABC son esencialmente proyecciones de flujos de costos y beneficios estimados de la planificación de actividades pesqueras y por tanto sujetas a un grado variable de incertidumbre. Por otra parte, el proyecto afectará en mayor o menor grado a los diferentes sectores de la sociedad, debido al criterio inherente de decisión que lo sustenta (e.g. valor máximo del producto social). Puede considerarse entonces que se dará incertidumbre también en esta etapa, para lo cual será adecuado incluir de un modo u otro una proyección del impacto que el proyecto generará en cada sector de la sociedad. A la hora de evaluar proyectos relacionados con pesquerías será necesario comparar los resultados provenientes del criterio de decisión empleado (sea preferiblemente VPN ó B/C) generados de distintas alternativas. Para ello, es conveniente emplear un amplio rango de valores de d que reflejen diferentes preferencias intertemporales en el uso de los recursos. Eventualmente podrá considerarse como patrón la tasa de interés existente en el mercado. Por ejemplo, si ésta es de 10%, podrían emplearse valores que oscilen entre 5 y 20%, representando respectivamente baja y alta TMPT. El empleo de tasas de descuento bajas beneficiará a aquellos usuarios (presentes ó futuros) de los recursos que desean conservarlo y en consecuencia explotarlo en un lapso prolongado en el tiempo. Este sería un ejemplo de un marco conservativo de manejo y al mismo tiempo una forma de estimular la inversión. La selección de altos valores de d (e.g. alta TMPT por el recurso) tenderá a favorecer aquellas alternativas con ocurrencia de costos relativamente tardía y beneficios produciéndose a corto plazo, ya que a mayores valores de d, mayor será la tasa de explotación del recurso. Es importante resaltar que una sola solución estaría mostrando las preferencias de sólo un sector de la sociedad, por lo que en cada evaluación realizada con determinado valor de d deberá especificarse el impacto distribucional que cada alternativa genera en los diferentes usuarios del recurso. b) Incertidumbre inherente a los recursos pesqueros. La mayoría de los modelos desarrollados en ciencias pesqueras han sido de carácter determinístico (ver Capítulo 2). Sin embargo, el cuerpo de conocimiento teórico-práctico generado en las últimas dos décadas, dirigido a adoptar decisiones inteligentes para el manejo de los recursos pesqueros, permite afirmar que tanto las pesquerías como los recursos que las sustentan no se comportan en base a tal aproximación, y brindan en general resultados incorrectos sobre la elección de la captura óptima (Hannesson, 1987). En efecto, los recursos naturales renovables, en especial los pesqueros, poseen un importante grado de variabilidad temporal en su magnitud. Ello impide predecir con certeza las fluctuaciones de su biomasa y abundancia a través del tiempo, y por ende la factibilidad de desarrollo de una pesquería, los costos de explotación (en caso de que éstos sean dependientes de la magnitud del stock) y dimensionamiento del fenómeno extractivo.

87
Dicho de otra manera, existe un fuerte componente de incertidumbre en las proyecciones de captura, y en consecuencia en los beneficios y costos incluidos como entrada en un ABC. Este hecho se ve agravado si se considera que los proyectos de pesca son en general a largo plazo, e incluyen componentes de inversión (e.g. barco de pesca) que tienen una vida útil que en general supera la década y presentan un bajo o nulo costo de oportunidad debido a la especialización de la actividad. Tres tipos de incertidumbre referidas a los recursos pesqueros deben ser tomadas en consideración a la hora de realizar un ABC (Lewis, 1982; Seijo, 1986): b1) incertidumbre en el tamaño del stock, el cual dependerá a su vez del nivel de esfuerzo efectuado en años anteriores y el grado en que éste afectó al stock; b2) fluctuaciones impredecibles en las variables ambientales, que a su vez inducen a variabilidad en el tamaño del stock y en consecuencia en la magnitud de la captura; b3) incertidumbre en la predicción de la demanda y precio del producto, así como en los costos de captura, en general debida a altibajos en la situación económica local o regional y aspectos coyunturales del mercado internacional. La incertidumbre asociada a la evaluación de un proyecto pesquero indicaría de una u otra manera que la decisión de llevarlo a cabo deba contener en forma explícita alguna corrección que considere dicha variabilidad. En este contexto, la generación de intervalos de confianza a través de enfoques tales como el Método de Monte Carlo puede ser empleada en aquellas variables críticas (e.g. proyecciones de captura, biomasa y diferentes niveles de esfuerzo) y considerar el límite inferior del intervalo a efectos de brindar resultados en un marco de acción precautorio. Ahora bien, qué criterio emplear como regla de decisión para escoger dentro de ese rango de alternativas aportadas por la aproximación estocástica? La decisión será dependiente de la visión e intereses sociales del administrador, y ésta a su vez del punto de vista de la sociedad frente a la adopción o no de nuevos cursos de acción. La toma de decisiones en la ejecución de proyectos se agrava aún más en aquéllos con una larga vida útil, donde las futuras generaciones puedan verse perjudicadas por su puesta en marcha, sin haber tomado parte en dicho proceso de decisión. Al respecto, resulta de significación la elección de una adecuada tasa de descuento, fundamentalmente en caso de recursos de larga vida e importancia en un contexto socio-económico (e.g. recursos demersales) donde las estrategias de manejo repercutirán en su disponibilidad para futuras generaciones.
Manejo intertemporal de recursos pesqueros El modelo bioeconómico dinámico y el precio del tiempo En el Capítulo 2 se desarrolló el modelo bioeconómico clásico en sus versiones estática y dinámica. Se presentaron los conceptos de máximo rendimiento sostenible (MRS), máximo rendimiento económico (MRE) y rendimiento en equilibrio bioeconómico (EBE), es decir, estimaciones bioeconómicas bajo condiciones de equilibrio. Asimismo, se presentaron las ecuaciones dinámicas que muestran la trayectoria de la biomasa, el rendimiento, la renta y el esfuerzo de pesca en el tiempo. Si bien los primeros constituyen conceptos claves en bioeconomía pesquera, el hecho de que provengan de un análisis estático les disminuye su potencialidad como predictores y en consecuencia como adecuadas herramientas de manejo.

88
Las trayectorias en el tiempo de las variables mencionadas anteriormente, es decir, la versión dinámica del modelo Gordon-Schaefer, permite contar con los flujos intertemporales de los costos y beneficios resultantes de diferentes niveles de esfuerzo de pesca. La influencia que la dinámica del esfuerzo tiene en la magnitud del stock, en las capturas, y en consecuencia en los flujos de costos y beneficios derivados de la pesquería, hacen que ésta deba ser manejada en base a la maximización de un patrón dinámico de explotación más que por un específico (estático) nivel de rendimiento sostenible (MRE; MRS). En efecto, dado que una pesquería será económicamente eficiente si maximiza el valor presente neto de las capturas, y que éstas están autocorrelacionadas en el tiempo (e.g. el nivel de explotación en un año repercutirá en forma directa en la magnitud del stock y en consecuencia en las capturas de años subsiguientes), el análisis bioeconómico basado en curvas de rendimiento sostenible pierde significación a efectos del manejo de un recurso pesquero.
Desde la perspectiva del administrador de pesquerías se busca identificar el nivel de esfuerzo pesquero f(t) que permita maximizar el valor presente del flujo de ingresos netos generados en el tiempo. Esto es:
PIT t CT t
(1+ d)t
(1+ d)t tππ
=−
=( ) ( ) ( )
(4.14)
donde VPπ es el valor presente de los ingresos netos π(t) en el tiempo. Clark (1985) desarrolla el modelo económico dinámico basado en el concepto de un solo propietario de la pesquería (que por ejemplo podría ser el Estado). Este concepto no significa un monopolio, ya que en éste se posee el control del mercado y en consecuencia se fija el precio del producto. Por el contrario, el modelo dinámico Gordon-Schaefer desarrollado por Clark (1985) supone que la industria es tomadora de precios que se asumen fijos y constantes. El modelo considera la tasa de descuento d en su forma continua. Esto es, el factor de descuento definido en (4.1) es expresado en su forma exponencial como:
d ,t t-t - tv
1(1+d )
(1+d ) e= = = δ (4.15)
En esta expresión δ es denominada tasa de descuento continua anual. De acuerdo a (4.15) δ se relaciona con d de la siguiente forma:
δ = ln(1+d ) (4.16)
El valor presente del flujo de los ingresos netos π(t) en un intervalo de tiempo dt será:
VP (t) (t)e dt0
T- tπ π δ= ∫ (4.17)

89
Asumiendo largo plazo (e.g. T=+∞) y sustituyendo en (4.17) a π(t), el objetivo de un solo propietario será maximizar el valor presente de la renta económica que genera el recurso:
Max [pqB(t)- c] f(t)e dt0
- t∞
∫ δ (4.18)
Sujeto a:
dB(t)dt
rB(t) 1 -B(t)K
qf(t)B(t)=⎛⎝⎜
⎞⎠⎟ − (4.19)
Debe considerarse asimismo la restricción de no negatividad en el monto del esfuerzo pesquero, de tal forma que f(t)≥0. Por otra parte se asume que B(t) ≥ 0 y que la biomasa inicial B0 es conocida. El nivel óptimo de biomasa BOP será función decreciente de δ y en consecuencia tenderá a acercarse a la biomasa en equilibrio bioeconómico (BEBE) en caso que δ -->+∞ (Clark, 1985). Resolviendo para f(t) en (4.19), sustituyendo en (4.18) e integrando por partes se obtiene una expresión para la biomasa óptima dada la tasa de descuento especificada:
BOP B kr
B kr
k BrEBE EBEEBE= + −
⎛⎝⎜
⎞⎠⎟ + + −
⎛⎝⎜
⎞⎠⎟
⎡
⎣⎢⎤
⎦⎥+
⋅ ⋅⎧⎨⎪
⎩⎪
⎫⎬⎪
⎭⎪
14
1 182δ δ δ
(4.20)
donde BEBE es definida como c/pq. Ahora bien, el rendimiento óptimo sostenible (ROS) y el esfuerzo óptimo (fop) para el precio del tiempo especificado con el parámetro δ se obtiene con la siguiente expresión:
ROS r BOPBOP
k= ⋅ −
⎛⎝⎜
⎞⎠⎟1 (4.21)
f ROSq BOP
op =⋅
(4.22)
El efecto de la tasa de descuento (δ ) en pesquerías: una visión alternativa La propuesta de Clark (1973; 1985) anteriormente desarrollada, ha sido discutida en la literatura por Hannesson (1986; 1987). El concepto clásico, que implica que altos valores de δ aumentan la probabilidad de sobreexplotación, es rechazado por Hannesson (1986) al expresar que tal propuesta ignora las implicaciones derivadas de un alto valor de la tasa de interés en el costo del capital, efecto previamente establecido por Farzin (1984) para recursos no renovables. Según

90
ambos autores, un alto valor de d aumentará el costo de extracción del recurso, dado que su explotación súbita requerirá inversiones a corto plazo en artes, equipos, etc. Hannesson (1986) discute la ambigüedad de δ: por un lado expresa la tasa de retorno requerida para un beneficio creciente obtenido en el corto plazo, de tal manera que mayores valores de d implicarán una mayor tasa de explotación y una disminución en el tiempo del stock explotable. Por otro, δ expresa el costo de oportunidad del capital a ser invertido en la unidad de pesca (e.g. buque, artes, equipos), por lo cual un alto valor de δ implicará mayores costos de operación y por tanto una explotación óptima menos intensa que en el caso anterior y un stock disponible de mayores magnitudes. El éxito en el desarrollo y sustentabilidad de una pesquería requerirá que la suma del valor presente de los ingresos netos totales sea maximizada. Esto significa que los niveles adecuados de captura en el tiempo dependerán de los valores esperados de costos y precios, fluctuaciones temporales en la tasa de crecimiento poblacional, y la situación económica y de mercado imperante en la región, probablemente reflejada en el valor de la tasa de interés. Las expectativas de cambios en los costos, precios y magnitud del stock deben ser incluidas en el estudio a través de un análisis ponderado de la distribución de probabilidades de diversas alternativas o cursos de acción. Estas razones suponen la necesidad de emplear aproximaciones dinámicas estocásticas para el manejo de un recurso pesquero. En este contexto, la tasa de descuento juega un papel fundamental para la adopción de un adecuado curso de acción a efectos de la explotación de un recurso pesquero. La elección de un valor de d será un paso crucial en la estrategia de explotación, y dependerá de la variabilidad esperada en variables intrínsecamente relacionadas al stock o bien al contexto socio-económico de los usuarios del recurso. Así pues, valores de δ que tiendan a +∞, debidos por ejemplo a una caída en el precio del producto o predicción de una drástica disminución del stock, supondrán estrategias de explotación que maximizarán los ingresos a corto plazo, no importando el efecto adverso en el largo plazo. En estos casos el MRE dinámico tenderá a un valor cercano al EBE. Cuando por el contrario, las características del recurso favorecen una explotación a largo plazo, hay cierta estabilidad en los precios y costos, y se desea favorecer la inversión, δ va a tender a 0 y en consecuencia el MRE dinámico coincidirá con el MRE estático. En general, el MRE dinámico fluctuará entre estas dos situaciones extremas (Anderson, 1986).