wildlife exposure factors handbook - risk assessment
TRANSCRIPT

United States Offi ce of Research EPA/600/R-93/187 Environment al Protect ion and Development December 1993 Agency (8603)
Wildlife Exposur eFactor s Handbook
Volume I of II

EPA/600/R-93/187 Decemb er 1993
WILDLIFE EXPOSURE FACTORS HANDBOOK
Volum e I of II
Offi ce of Health and Environmental Assessmen tOffi ce of Research and Development
U.S. Environmental Protect ion A gencyWashingt on, DC 20460
Addit iona l major funding f or this Handbook was p rovided by th e Offi ce of Emer gency an d Remedial Respons e,
Offi ce of Solid W aste an d Emergency Respons eand by th e
Offi ce of Sci ence and Technology , Offi ce of WaterU.S. Environmental Protect ion A gency
Washingt on, DC 20460

DISCLAIMER
This document has been reviewed in acco rdance wi th U.S. Environmental
Protect ion A gency polic y and appr oved for publi cation. Ment ion of tr ade names o r
commer cial products d oes not co nsti tute endor semen t or recommen dation f or use.
ii

CONTENTS
Foreword . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
Preface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi i
Author s, Managers, Cont ribut ors, and Re viewers . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi ii
1. INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-1
1.1. PURPOSE AND SCOPE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-11.2. ORGANIZATION OF THE HANDBOOK . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-51.3. LIST OF SELECTED SPECIES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-61.4. LIST OF EXPOSURE FACTORS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-11
1.4.1. Normalizing F acto rs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-11
1.4.1.1. Body W eight . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-131.4.1.2. Growth Rate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-131.4.1.3. Metabolic Rate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-14
1.4.2. Contact R ate Facto rs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-14
1.4.2.1. Oral Rout e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-141.4.2.2. Inhalation Rout e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-161.4.2.3. Dermal Rout e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-16
1.4.3. Popula tion Dy namics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-17
1.4.3.1. Social Organization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-171.4.3.2. Home Range/Territory Size/Foraging Ra dius . . . . . . . . 1-171.4.3.3. Popula tion De nsity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-191.4.3.4. Annua l Fecundit y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-191.4.3.5. Annua l Mor talit y and Longe vi ty . . . . . . . . . . . . . . . . . . . 1-19
1.4.4. Seasonal Acti vi ties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-20
1.5. DATA PRESENTATION FORMAT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-20
1.5.1. Normalizing a nd Con tact R ate Facto rs . . . . . . . . . . . . . . . . . . . . . 1-21
1.5.1.1. All Animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-211.5.1.2. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-221.5.1.3. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-231.5.1.4. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . 1-23
iii

CONTENTS (cont inued)
1.5.2. Dietary Composi tion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-24
1.5.2.1. All Animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-24
1.5.3. Popula tion Dy namics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-25
1.5.3.1. All Animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-251.5.3.2. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-261.5.3.3. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-271.5.3.4. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . 1-27
1.5.4. Seasonal Acti vi ties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-28
1.5.4.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-291.5.4.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-291.5.4.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . 1-29
1.5.5. Abbr eviations Used in T ables . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-29
1.6. LITERATURE SEARCH STRATEGY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-301.7. REFERENCES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-32
2. EXPOSURE FACTORS AND DESCRIPTIONS OF SELECTED SPECIES . . . . . . . 2-1
2.1. BIRDS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1
2.1.1. Great Blue Heron . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-32.1.2. Canada Goose . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-192.1.3. Mallard . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-392.1.4. Lesser Scaup . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-532.1.5. Osprey . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-652.1.6. Red-Tailed Hawk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-792.1.7. Bald Eagle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-912.1.8. Amer ican Kestr el . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1092.1.9. Nor thern Bobw hit e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1212.1.10. Amer ican Woodc ock . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1372.1.11. Spotted Sandpipe r . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1492.1.12. Herring G ull . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1572.1.13. Belted Kingf isher . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1732.1.14. Marsh Wren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1832.1.15. Amer ican Robin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-193
iv

CONTENTS (cont inued)
2.2. MAMMALS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-207
2.2.1. Short-Tailed Shrew . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2092.2.2. Red Fox . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2212.2.3. Racco on . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2332.2.4. Mink . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2472.2.5. River Otter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2612.2.6. Harbor Seal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2752.2.7. Deer Mous e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2912.2.8. Prairie Vole . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3112.2.9. Meadow Vole . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3232.2.10. Muskr at . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3372.2.11. Easter n Cotto ntai l . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-351
2.3. REPTILES AND AMPHIBIANS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-365
2.3.1. Snapping T urtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3672.3.2. Painted Turtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3812.3.3. Easter n Box Turtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3972.3.4. Racer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4072.3.5. Northern Water Snake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4192.3.6. Easter n Newt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4292.3.7. Green Frog . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4432.3.8. Bullf rog . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-453
3. ALLOMETRIC EQUATIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-1
3.1. FOOD INGESTION RATES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-3
3.1.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-43.1.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-53.1.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-6
3.2. WATER INTAKE RATES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-7
3.2.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-83.2.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-103.2.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-10
3.3. INHALATION RATES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-11
3.3.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-113.3.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-123.3.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-12
v

CONTENTS (cont inued)
3.4. SURFACE AREAS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-13
3.4.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-133.4.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-143.4.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-14
3.5. ALLOMETRIC EQUATIONS FOR METABOLIC RATE . . . . . . . . . . . . . . . . 3-15
3.5.1. Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-18
3.5.1.1 Basal Metabolic Rate . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-193.5.1.2. Existen ce Metabolic R ates . . . . . . . . . . . . . . . . . . . . . . . 3-203.5.1.3. Free-Liv ing M etabolic Rate . . . . . . . . . . . . . . . . . . . . . . . 3-223.5.1.4. Temperature and Metabolic Rate . . . . . . . . . . . . . . . . . . 3-24
3.5.2. Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-26
3.5.2.1. Basal Metablic Rate . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-263.5.2.2. Resting M etabolis m . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-273.5.2.3. Field Metabolic Rate . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-273.5.2.4. Temperature and Metabolic Rate . . . . . . . . . . . . . . . . . . 3-28
3.5.3. Rept iles and Amphibia ns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-29
3.5.3.1. Basal and Rest ing Metabolic R ates . . . . . . . . . . . . . . . . 3-293.5.3.2. Free-Liv ing M etabolic R ates . . . . . . . . . . . . . . . . . . . . . . 3-30
3.6. MATH PRIMER AND UNIT CONVERSIONS . . . . . . . . . . . . . . . . . . . . . . . . . 3-32
3.6.1. Summar y of Operations Involv ing Loga rithms . . . . . . . . . . . . . . 3-323.6.2. Summar y of Operations Involv ing P owers . . . . . . . . . . . . . . . . . . 3-323.6.3. Unit Conv ersions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-33
3.6.3.1. Approximate Facto rs fo r Metabolic Equa tions . . . . . . . 3-333.6.3.2. Exact Conv ersions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-33
3.7. ESTIMATING CONFIDENCE INTERVALS . . . . . . . . . . . . . . . . . . . . . . . . . . 3-34
3.8. REFERENCES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-38
4. EXPOSURE ESTIMATES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-1
4.1. GENERAL DOSE EQUATIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-1
4.1.1. Drink ing W ater . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-3
vi

CONTENTS (cont inued)
4.1.2. Diet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-6
4.1.2.1. Dose Equations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-64.1.2.2. Energy Cont ent and Assi mila tion Effi ciencies . . . . . . . . 4-10
4.1.3. Soil a nd Sedim ent Ing esti on . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-16
4.1.3.1. Backg round . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-184.1.3.2. Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-184.1.3.3. Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-194.1.3.4. Dose Equations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-21
4.1.4. Air . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-214.1.5. Dermal Exposure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-23
4.2. ANALYSIS OF UNCERTAINTY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-23
4.2.1. Natural Variation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-244.2.2. Sampling Unc ertaint y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-254.2.3. Model Unc ertaint y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-26
4.3. REFERENCES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-26
APPENDIX: LITERATURE REVIEW DATABASE . . . . . . . . . . . . . . . . . . . . . . . See Volum e II
vi i
2

LIST OF TABLES
Table 1-1. Character isti cs o f Selected Birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-8
Table 1-2. Character isti cs o f Selected Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-9
Table 1-3. Character isti cs o f Selected Repti les and Amphibia ns . . . . . . . . . . . . . . . 1-10
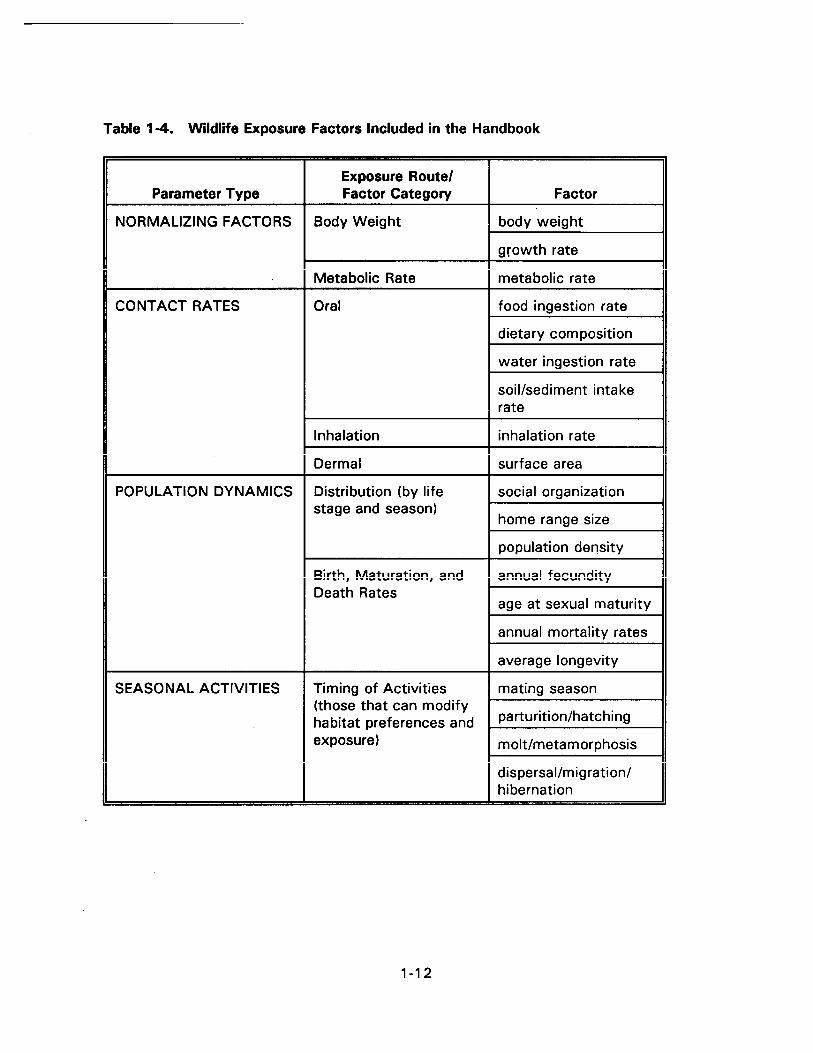
Table 1-4. Wildlif e Expos ure Facto rs Include d in t he Handbook . . . . . . . . . . . . . . . . 1-12
Table 1-5. Wildlif e Contact R ate Exp osure Facto rs . . . . . . . . . . . . . . . . . . . . . . . . . . 1-15
Table 1-6. Colum n Headers for Tables of Normalizing a nd Con tactRate Facto rs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1-21
Table 1-7. Colum n Headers for Tables on Di etary Composi tion . . . . . . . . . . . . . . . . 1-24
Table 1-8. Colum n Headers for Tables of F acto rs fo r Popula tion Dy namics . . . . . . 1-26
Table 1-9. Colum n Headers for Tables on Seasonal Acti vi ties . . . . . . . . . . . . . . . . . 1-28
Table 2-1. Birds Include d in t he Handbook . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2
Tables fo r 2.1.1. Great Blue Heron . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-82.1.2. Canada Goose . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-232.1.3. Mallard . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-432.1.4. Lesser Scaup . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-562.1.5. Osprey . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-682.1.6. Red-Tailed Hawk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-822.1.7. Bald Eagle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-952.1.8. Amer ican Kestr el . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1122.1.9. Nor thern Bobw hit e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1262.1.10. Amer ican Woodc ock . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1402.1.11. Spotted Sandpipe r . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1522.1.12. Herring G ull . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1622.1.13. Belted Kingf isher . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1762.1.14. Marsh Wren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-1862.1.15. Amer ican Robin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-197
Table 2-2. Mammal s Include d in t he Handbook . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-208
Tables for 2.2.1. Short-Tailed Shrew . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2132.2.2. Red Fox . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2242.2.3. Racco on . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2362.2.4. Mink . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2512.2.5. River Otter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-264
viii

LIST OF TABLES (cont inued)
2.2.6. Harbor Seal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2802.2.7. Deer Mous e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-2952.2.8. Prairie Vole . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3142.2.9. Meadow Vole . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3272.2.10. Muskr at . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3402.2.11. Easter n Cotto ntai l . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-355
Table 2-3. Repti les and Amphibia ns Inc lude d in t he Handbook . . . . . . . . . . . . . . . . 2-366
Tables for 2.3.1. Snapping T urtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3702.3.2. Painted Turtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-3862.3.3. Easter n Box Turtle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4002.3.4. Racer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4112.3.5. Northern Water Snake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4232.3.6. Easter n Newt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4332.3.7. Green Frog . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-4462.3.8. Bullf rog . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2-456
Table 3-1. Metaboliza ble En ergy (ME) of Var ious Diets fo r Birdsand Mammal s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-5
Table 3-2. Allometr ic Equations fo r Basal Metabolic R ate (BMR) in Bir ds . . . . . . . . 3-21
Table 3-3. Regress ion Stati sti cs fo r Nagy's (1987) Allometr ic Equationsfor Food Ingestion R ates fo r Free-Living A nim als . . . . . . . . . . . . . . . . . . 3-36
Table 3-4. Regress ion Stati sti cs fo r Nagy's (1987) Allometr ic Equationsfor Free-Liv ing ( Field) Metabolic R ates . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-37
Table 4-1. Gross En ergy and Water Composi tion of W ildlif e Foods :Animal Prey . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-13
Table 4-2. Energy and Water Composi tion of W ildlif e Foods :Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-14
Table 4-3. General Assi mi lation Effi ciency (AE) Values . . . . . . . . . . . . . . . . . . . . . . . 4-15
Table 4-4. Percent Soil or Sedim ent in Die t Esti mated FromAcid- Insoluble A sh of Scat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-20
Table 4-5. Other Esti mates o f Percentage of Soil or Sedim ent in Die t . . . . . . . . . . . 4-21
ix

LIST OF FIGURES
Figur e 3-1. Mont hly Var iation in E nergy Budge t Esti mated for aHouse Sparrow . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3-25
Figur e 4-1. Wildlif e Dose Equations fo r Drink ing W ater Exposures . . . . . . . . . . . . . 4-4
Figur e 4-2. Wildlif e Dose Equations fo r Dietary Exp osures . . . . . . . . . . . . . . . . . . . 4-6
Figur e 4-3. Esti mat ing NIR k When Dietary Composi tion Is Know n on aWet-Weight Basis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-8
Figur e 4-4. Esti mat ing NIR k Based on Differ ent ME Values WhenDietary Composi tion Is Exp ressed as Percentage ofTotal Prey Captured . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-9
Figur e 4-5. Utiliza tion of Food E nergy by Anim als . . . . . . . . . . . . . . . . . . . . . . . . . . 4-11
Figur e 4-6. Metaboliza ble En ergy (ME) Equation . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-12
Figur e 4-7. Examp le of Esti mat ing Food Ing estion R ates fo rWildlif e Species From Free-Liv ing M etabolic R ate andDietary Composi tion: Male Mink . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-17
Figur e 4-8. Wildlif e Oral Dose Equation f or Soil or Sedim entIngestion E xpos ures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-22
x

FOREWORD
The Exposure Assessmen t Group ( EAG) of EPA 's Offi ce of Research and
Developm ent has three main f unc tions : (1) to conduc t hum an health and ecologic al
exposure and risk assessmen ts, (2) to review exp osure and risk assessmen ts an d related
documents, and (3) to develop guide line s and ha ndbook s for use in t hese assessmen ts.
The acti vi ties unde r each of th ese fu nct ions ar e suppor ted by and r espond t o the needs of
the var ious progr am of fices, regiona l of fices, and the technical commu nity.
The Wildlif e Expos ure Facto rs Handbook was p roduced in response to the
increased interest i n assess ing r isks to ecologic al systems. Its p urpose is to improve
exposure assessmen ts fo r wildlif e and s uppor t the quant if ication of r isk esti mates. It is a
companion doc ument to the Expos ure Facto rs Handbook , whic h cont ains inf ormati on
useful f or quant ifying e xpos ure to hum ans. Because informat ion a nd methods fo r
esti mat ing e xpos ure are cont inua lly improv ing, we will r evise these handbook s as
necessar y in the future.
Michael A. CallahanDirecto rExposure Assessmen t Group
xi

PREFACE
The Exposure Assessmen t Group of th e Offi ce of Health and Environmental
Assessmen t (OHEA) has prepared the Wildlif e Expos ure Handbook in s uppor t of th e Offi ce
of Solid W aste an d Emergency Response and the Offi ce of Water . The Handbook prov ides
inf ormation on v arious facto rs used to assess exp osure to wildlif e. The goals of th e
project ar e (1) to promote th e appli cation of r isk assessmen t met hods to wildlif e species,
(2) to foster a consisten t approach to wildlif e expos ure and r isk assessmen ts, and (3) to
increase th e access ibilit y of th e lit erature applic able to these assessmen ts.
The bulk of th e doc ument summar izes l iterature values fo r exposure facto rs fo r 34
species of birds, mammal s, amphibia ns, and r ept iles. In addit ion, we inc lude a chapter on
allometr ic equations th at can be used to esti mate so me of th e exposure facto rs wh en data
are lacking. Fina lly , we descr ibe some co mmo n equations used to esti mate exp osure. The
basic l iterature search was co mpleted in May 1990 and was s upplemen ted by tar geted
searches conducted in 1992.
We anticipate upda ting t his Handbook and w ould a ppr eciate any assi stan ce in
ident ifying a ddit iona l sour ces o f informat ion t hat fill d ata gaps or otherwise improve th e
Handbook . Commen ts can be sent to :
Exposure Assessmen t Group Wildlif e Expos ure Facto rs Handbook Pr oject USEPA (8603) 401 M Street, SW Washingt on, DC 20460
xi i

AUTHORS, MANAGERS, CONTRIBUTORS, AND REVIEWERS
The Exposure Assessmen t Group ( EAG) wi thin E PA's Offi ce of Health and
Environmental Assessmen t (OHEA) was r espons ible fo r preparing t his doc ument.
Addit iona l major cont ract funding was p rovided by th e Offi ce of Emer gency an d Remedial
Respons e wi thin t he Offi ce of Solid W aste an d Emergency Response (OSWER) and by th e
Offi ce of Sci ence and Technology wi thin t he Offi ce of Water . The document was p repared
by ICF, Incorpor ated unde r cont racts 68-C 8-0003, 68-W8-0098, 68-DO-0101, and 68-C2-
0107. Susan Braen Norton served as th e EAG project man ager and provided overall
dir ection a nd c oor dina tion of th e project. Thanks to Doug Nor ton f or th e cover illus tration.
AUTHORS
Dr. Margaret McVeyKimberly Hall, Peter Trenham, Alexander Soast, Lesl ie Frymi er, and Ansara HirstICF, Incorpor ated
PROJECT MANAGER
Susan Braen NortonExposure Assessmen t Group, Offi ce of Health and Environmental Assessmen tOffi ce of Research and DevelopmentU.S. Environmental Protect ion A gency
TASK MANAGERS
Cynthia NoltOffi ce of Sci ence and TechnologyOffi ce of WaterU.S. Environmental Protect ion A gency
Ron Presto nOffi ce of Emer gency an d Remedial Respons eOffi ce of Solid W aste an d Emergency Respons eU.S. Environmental Protect ion A gency
CONTRIBUTORS
Dr. Sarah Gerould Offi ce of Water Quality U.S. Geologic al Survey
xiii

Dr. Ronald Kir byOffi ce of Informat ion T ransferU.S. Fish and Wildlife Service (USFWS)Departmen t of th e Inter ior
REVIEWERS
Dr. Cynthia Annett, USFWS, Arkansas Cooper ative Fish and Wildlife ResearchUnit, AR
Dr. Rich ard J. Auler ich, Michigan State Universi ty, MIDr. Richard Bennett, USEPA, Environmental Research Laboratory (ERL),
Corvallis, ORDr. Chr istine A. Bishop, Canadian Wildlife Service, Ontar io, CanadaDr. Brad Bortner, USFWS, Offi ce of Migr atory Bir d Management, MDDr. Steven Bradbur y, USEPA, ERL-Duluth, MNDr. Rober t Burr, U SFWS, VAMs. Janet Burris, ABB Environmental Services, Washington, DCDr. Joseph Chapman, Utah State Universi ty, UTDr. Lewis Cowar din, USFWS, Nor thern Prairie Wildlife Research Center , NDMs. Deanna Dawso n, USFWS, Patuxent Wildlife Research Center , MDDr. Donald L. DeAngelis, Oak Ridge National Labor atory, TNDr. Bruce Duncan, USEPA, Region 10, Seattl e, WADr. Rober t Foley, U SFWS, Annapolis Field Offi ce, MDDr. Glen Fox, Canadian Wildlife Service, Ontar io, CanadaDr. J.H.B. Garner, USEPA, OHEA, Environmental Criter ia and Assessmen t Offi ce
(ECAO), Research Triangle Park, NCDr. J. Whitfi eld Gibbons, Univer si ty of Georgia, Savannah River Ecology
Laboratory, GAMr. Keith A. Grasman , Virginia Polytec hnic Insti tute and State Universi ty, VADr. Fred Guthery, Caesar Kleberg Wildlife Research Insti tute, Texas A&I
Universi ty, TXDr. Richard S. Halbrook, Oak Ridge National Labor atory, TNDr. Michael J. Hamas, Central Michigan Univer si ty, MIDr. Charles J. Henny, USFWS, Paci fic Northwest R esearch Stat ion, ORDr. Paula F. P. Henry, USFWS, Patuxent Wildlife Research Center , MDDr. Gary Heinz, USFWS, Patuxent Wildlife Research Center , MDDr. Doug James, Universi ty o f Arkansas, ARMr. K. Bruce Jo nes, USEPA, Environmental Monito ring Systems L aboratory,
EMAP Terrestr ial Ecosystems, CADr. Dennis G. Jorde, USFWS, Patuxent Wildlife Research Center , MDDr. Norm Kowal, USEPA, OHEA, ECAO, Cincinnati , OHDr. Gary L. Krapu, USFWS, Nor thern Prairie Wildlife Research Center , NDDr. David Krementz, USFWS, Patuxent Wildlife Research Center , MDDr. James K ushlan, Univer si ty of Missi ssippi, MSDr. John T . Lokemo en, USFWS, Northern Prairie Wildlife Research Center , NDDr. Dave Ludwig, EA Engineer ing, Inc. , MDDr. Drew Major , USFWS, Conco rd, NHDr. Karen McBee, Oklahoma State Universi ty, OK
xiv

REVIEWERS (cont inue d)
Dr. Linda M eyers-Schöne , Internationa l Technology Corp., NMMr. Jer ry Moor e, USEPA, OHEA, Washingt on, DCDr. Lewis Oring, Univ ersi ty of N evada at Reno, NVDr. Raymond P ierot ti , Univ ersi ty of A rkansas, ARDr. Richard M. Poché, Genesis Laboratories, WIDr. Dougla s Reagan, Woodw ard-Clyde Cons ult ants, CODr. Lor enz Rhom berg, USEPA, OHEA, Human Health Assessmen t Group,
Washingt on, DCDr. Robert Ringe r, Mount Dora, FLDr. John L. Roseberry, Sout hern Illinois Univ ersi ty at Carbonda le, ILDr. Alan Sargeant, USFWS, Nor thern Prairie Wildlif e Research Center , NDDr. Edward Schafer , USDA, Denver Wildlif e Research Center , CODr. Laird Schut t, Canadian Wildlif e Service, Quebec, CanadaMs. Anne Sergeant, USEPA, OHEA, EAG, Washingt on, DCDr. Kim Smi th, Universi ty o f Arkansas, ARDr. Mark Sprenger, USEPA, OSWER, Environmental Response Team, Edison, NJDr. George A. Swanson, USFWS, Northern Prairie Wildlif e Research Center , NDDr. Sylvia Talmage, Oak Ridge Nationa l Labor atory, TNDr. Bob T rost, USFWS, Offi ce of Migr atory Bir d Management, MDDr. D.V. (Chip) W eseloh, Canadian Wildlif e Service, Ontar io, CanadaDr. Nathanie l Wheelwr ight , Bow doin Colle ge, MEDr. Donald H. Whit e, USFWS, Sout heast Research Stat ion, GADr. Stanley N. Wiemeyer , USFWS, Patuxent Wildlif e Research Center , MDDr. Bill W illi ams, Environmental Planning a nd T oxicology , Corvallis , ORDr. Dean M. Wilkinson, U.S. Departmen t of Commer ce, Nationa l Oceanic and
Atmo sphe ric Adminis tration, Nationa l Marine Fisheries ServiceDr. Jer ry O. Wolff, USEPA, ERL, Corvallis , ORDr. John P . Wolf in, USFWS, Annapolis Field Station, MD
xv

1. INTRODUCTION
The Wildlif e Expos ure Facto rs Handbook (h ereafter refer red to as th e Handbook )
provides data, references, and guida nce for conduc ting e xpos ure assessmen ts fo r wildlif e
species expos ed to toxic chemicals in t heir environm ent. It is th e produc t of a joint effo rt
by EPA's Offi ce of Research and Development (ORD), Offi ce of Solid W aste an d
Emergency Response (OSWER), and Offi ce of Water (OW). The goals of th is Handbook ar e
(1) to promote th e appli cation of r isk assessmen t met hods to wildlif e species, (2) to foster
a consisten t approach to wildlif e expos ure and r isk assessmen ts, and (3) to increase th e
access ibilit y of th e lit erature applic able to these assessmen ts.
1.1. PURPOSE AND SCOPE
The pur pos e of th e Handbook is to prov ide a conv enient sour ce of inf ormation a nd
an analyti c fr amewo rk fo r scr eening- level risk assessmen ts fo r commo n wildlif e species.
These scr eening- level risk assessmen ts may b e used for sever al purposes, including: to
assess p otential effects o f environmental contami nation on w ildlif e species and t o suppor t
si te-sp eci fic decisions (e. g., for hazardous waste si tes); to suppor t the development of
water -quality o r other media-speci fic cr iter ia for limi ting e nvironm ental levels of to xic
substan ces to protect w ildlif e species; or to focus research and monit or ing effo rts. The
Handbook prov ides data (analogous to EPA's Exposure Facto rs Handbook for humans,
USEPA, 1989c) and methods fo r esti mat ing w ildlif e intakes o r doses o f environmental
cont amina nts. Although t he data presented in the Handbook can be used for scr eening
analyses, we recommen d that anyone establis hing a cl eanup goa l or cr iterion on t he basis
of value s cont ained herein obtain the original literature on which the values are based to
conf irm that the study quality is suffi cient to suppor t the cr iter ion. This Handbook does
not inc lude d ata or extr apola tion methods r equir ed to assess th e toxici ty o f su bstan ces to
wildlif e species, nor does it inc lude an y chemical-speci fic data (e.g., bioa vaila bilit y facto rs).
For the Offi ce of Water , data gathered for the Handbook wer e used to ident ify
wildlif e species that are lik ely to be at gr eater risk fr om bioaccu mulative pollut ants in
surface water s and to esti mate l ikely exp osures fo r these sp ecies. Data on diets an d on
1-1

food a nd water inge stion r ates can be used wi th chemical-speci fic informat ion, such as
bioaccu mulation pot ential and w ildlif e toxici ty, to calcul ate si te- or region- speci fic
conc entrations of a chemical in water (or soil or sedim ent) that are unlik ely to cause
adverse effects.
For the Superfund pr ogr am, this Handbook suppl ements th e exist ing e nvironm ental
evalua tion guida nce. EPA's Risk A ssessmen t Guidance fo r Superfund: Volume II--
Envi ronmental Evaluation Manual (U.S. EPA, 1989a) provides an overview of ecologic al
assessmen t in the Superfund pr ocess. It includes a descr iption of th e statu tory and
regulatory bases fo r ecologic al assessmen ts i n Superfund a nd f und amental concepts fo r
unde rstanding e cologic al effects o f environmental contami nants. The Envi ronmental
Evaluation Manual also reviews el ements o f planning a n ecologic al assessmen t and how to
organize and pr esent th e results o f th e assessmen t. EPA's Ecological Assessmen t of
Hazardous Waste Si tes: A Field and Laboratory Reference (U.S. EPA, 1989b) and
Evaluation of Terrestr ial Indicato rs for Use in Ecological Assessmen ts at H azardous Waste
Sites (U.S. EPA, 1992) are companion doc uments that descr ibe biologic al assessmen t
str ategie s, fi eld s ampling de signs , toxici ty tests, biom arkers, biologic al field assessmen ts,
and data interpretat ion. The ECO Update intermitten t bulle tin s eries (publis hed by EPA 's
Offi ce of Solid W aste an d Emergency Respons e, publi cation no. 9345.0-05I, avai lable from
the Nationa l Technic al Information S ervice, Springf ield, Virginia ) prov ides suppl emental
guida nce for Supe rfund on s elected issu es. Although t hese documents h ave identi fied
decreases i n wildlif e popula tions as pot ential endpoint s for ecologic al assessmen ts, they
do not prov ide guida nce on how to conduc t a wildlif e expos ure assessmen t th at is
comparable to the guida nce prov ided by th e Superfund pr ogr am for hum an health
exposure assessmen ts. This Handbook prov ides bot h guida nce and d ata to fac ilitate
esti mat ing w ildlif e expos ure to con tami nants i n the environment.
Exposure assessmen ts fo r wildlif e and hum ans di ffer in sever al important ways.
One key d isti nct ion is th at many di ffer ent wildlif e species may b e exposed, as co mpared
wi th a single sp ecies of conc ern for a hum an health assessmen t. Exposure var ies between
differ ent sp ecies and even between differ ent popula tions of th e same sp ecies; behavioral
attr ibutes an d diet and habitat p references i nfluence th is var iation. Second, whereas i t is
1-2

�
seldom pos sible to conf irm esti mated levels of human exposure wi thout invasi ve sam pling
of human tissu es, confi rmato ry sam pling f or man y chemicals can be done in w ildlif e
species (protected species excep ted). However , the tissu e sampling r equir ed to quant ify
actu al exposure levels can be costl y, and interpretat ion of ti ssu e conc entrations can be
complex.
For bot h hum an health and w ildlif e expos ure assessmen ts, the most co st-effecti ve
approach is often to fi rst scr een for potentially s ignif icant expos ures us ing measu res (or
esti mates) o f environmental contami nation ( e.g., in s oils , water , prey sp ecies) to esti mate
contami nant intakes o r doses b y s ignif icant routes o f exp osure. If esti mated doses fal l far
below th e toxici ty val ues asso ciated wi th adverse effects, especially fr om ch ronic
exposures, further assessmen t may b e unnecessar y. If esti mated doses far exceed
reference to xici ty val ues, it may b e possible to determine appropriate act ions on t he basis
of th ese esti mates a lone . When a scr eening- level exposure assessmen t indi cates th at
adverse effects ar e likely, addit iona l conf irmato ry data may b e needed in the decision-
making pr ocess. For humans, it is usually not practi cable to obtain addit iona l types of data
(e.g., tissu e conc entrations , biom arkers), and hum an expos ure esti mates ar e often refined
by us ing m ore si te-sp eci fic data fo r exposure parameter s. For wildlif e, conf irmato ry data
may b e obtained from ch emical analyses o f ti ssu e samp les fr om potentially exp osed
wildlif e or their prey and f rom obs erved inc idence of disease, reproduc tive failur e, or death
in expos ed wildlif e. These are reviewed in EPA's fi eld and laboratory reference and
ter restr ial indi cato rs documents d escr ibed above (EPA, 1989b, 1992). If th is mo re direct
appr oach is not pos sible , the expos ure analysi s can be refine d on t he basis of mo re si te-
speci fic data fo r the species of co ncern.
Wildlif e can be expos ed to environm ental con tamina nts through inha lation, dermal
contact wi th contami nated water or soil, or ing estion of con tami nated food, water , or soil.
Exposure assessmen t seeks to answer sever al quest ions , inc luding:
� What organisms ar e actu ally or potentially exp osed to contami nants?
1-3

�
�
�
�
�
�
�
� Which organisms o r life stag es might be mo st vulne rable to environm ental
contami nants (e.g., ingest th e largest q uanti ties of co ntami nated media
relative to body size )?
� What are the signif icant routes o f exp osure?
� To what amo unts o f each contami nant are organisms actu ally or potentially
exposed?
� How long is each exposure?
� How often does or will e xpos ure to the environm ental con tami nants take
place?
� What seaso nal and cl imati c var iations in c ondit ions ar e lik ely to affect
expos ure?
� What are the si te-sp eci fic geophy sical , phy sical , and c hemical condit ions
affecting e xpos ure?
The parameter s fo r which data are presented in the Handbook ar e int ended to help a r isk
assesso r answer these quest ions . The popula tion pa rameter data (e.g., birth and death
rates) may b e usefu l for placing esti mates o f r isks to wildlif e popula tions in a broader
ecologic al con text an d for planning m onit or ing acti vi ties.
This Handbook fo cuses o n selected groups of mammal s, birds, amphibia ns, and
repti les. Fish and aquatic or ter restr ial inver tebrates wer e not include d in t his effo rt. The
prof iles on a mphibia ns and r ept iles are, in ge neral, less d evelope d than thos e for bir ds an d
mammal s. We emphasized birds and mammal s because met hods fo r assess ing t heir
expos ure are more common a nd w ell de velope d. As more assessmen ts ar e done fo r
amphibia ns and r ept iles, we anticip ate th at addit iona l methods and s uppor ting facto rs wi ll
be necessar y. Unti l then, we hope th e inf ormation pr esented here will e ncour age
assesso rs to begin c ons idering a nd qua nt ifying t heir expos ure.
For all exposure parameter s and species in the Handbook , we try to present data
indi cati ve of th e range of val ues th at differ ent popula tions of a species may assu me acr oss
North Amer ica. For si te-sp eci fic ecologic al risk assessmen ts, it is important to note th at
the values fo r exposure facto rs presented in th is Handbook may n ot accu rately represent
1-4

speci fic loc al popula tions . The species inc lude d in t he Handbook h ave broad geographic
ranges, and they may ex hibit di ffer ent val ues fo r many of th e exposure facto rs in differ ent
por tions of th eir range. Some species exhibit geogr aphic var iation in body size , surviv al,
and r eproduc tion. Breeding a nd m igr ation a lso inf luence expos ure. Site-sp eci fic val ues
for these parameter s can be determined more accu rately using publis hed s tudie s of loc al
popula tions and assi stan ce fr om th e U.S. Fish and Wildlif e Service, state d epartmen ts o f
fish and game, and organizations su ch as loc al Audubon S ociety ch apters. In addit ion, The
Nature Cons ervancy de velops and m aint ains wildlif e databases (i ncluding e ndangered
species) in coope ration w ith all 50 states. Local informat ion inc reases th e cer tainty o f a
risk assessmen t. Thus, for si te-sp eci fic assessmen ts, we str ongly r ecommen d contacti ng
loc al wildlif e experts to determine the presence and character isti cs o f sp ecies of co ncern.
Fina lly , we do not int end t o imply th at risk assessmen ts fo r wildlif e should be
restr icted to the species descr ibed in th is Handbook , or should a lways b e conducted for
these sp ecies. We emphasize that local ly important or rare species not include d in t his
Handbook may st ill be ver y im por tant for si te-sp eci fic r isk assessmen ts. To assi st u sers
who wish to evaluate other species, we l ist g eneral references fo r birds, mammal s,
rept iles, and a mphibia ns in Nor th Amer ica. The Handbook al so prov ides allo metr ic
equations to assi st i n extr apola ting e xpos ure facto rs (e.g., water inge stion r ate, surface
area) to closely related species on the basis of body size .
1.2. ORGANIZATION OF THE HANDBOOK
The Handbook is organized int o four chapters. The remainde r of this ch apter
provides an overview of th e species and exposure facto rs include d in t he Handbook an d
discu sses th e literature search str ategy used to identi fy facto rs. Chapter 2 presents
exposure profi les fo r the selected species (descr ibed in greater detai l below). Chapter 3
provides al lometr ic mo dels th at may b e used to esti mate f ood a nd water inge stion r ates,
inha lation r ates, surface ar eas, and metabolic r ates fo r wildlif e species on t he basis of
body size . Chapter 4 descr ibes co mmo n equations used to esti mate w ildlif e expos ure to
environm ental cont amina nts. Inc lude d are methods fo r esti mat ing di et-sp eci fic food
1-5

�
�
inge stion r ates o n the basis of meta bolic r ate and for esti mat ing e xpos ure to chemicals in
soil a nd s edim ent.
Chapter 2 is th e core of th e Handbook ; it presents exp osure profi les fo r selected
birds (Sect ion 2 .1), mammal s (Sect ion 2 .2), and r ept iles and amphibia ns (Sect ion 2 .3),
along w ith br ief descr iptions of th eir natural his tory. Each species profi le includes an
int roduc tion t o the species' general taxonom ic group, quali tati ve descr iption of th e
species, list o f si mi lar species, table of exp osure facto rs, and reference l ist (wh ich also
covers th at sp ecies' sect ion in V olum e II, the Appendix ). The value s inc lude d in t he
exposure facto rs tab les are a subset o f th ose we f ound in t he lit erature and a lso inc lude
values th at we esti mated using t he allo metr ic equations presented in Chapter 3. We
selected values fo r the tables in Chapter 2 based on a var iety o f facto rs including samp le
size, quant if ication of var iabilit y (e.g., standa rd deviations , stan dard errors, ranges),
relevance of th e measu rement tec hnique fo r expos ure assessmen t, and coverage of
habitats, subspecies, and the var iabilit y seen in the literature. A complete l ist ing of th e
parameter values identi fied in our literature survey i s provided in the Appendix . The
Appendix al so inc lude s more details conc erning sample size , methods , and qua lif ying
inf ormation t han the species pr of iles. Users are encour aged to cons ult th e Appendix to
select th e most ap propriate val ues fo r their particular assessmen t.
The remainde r of this int roduc tory chapter d escr ibes th e species and exposure
facto rs co vered in the Handbook in gr eater detai l. The literature search str ategy is
discu ssed in Sect ion 1 .6.
1.3. LIST OF SELECTED SPECIES
Wildlif e species were selected for the Handbook to prov ide sever al typ es of
coverage:
� Major taxonom ic groups (major ver tebrate groups , orders, and fami lies);
� A range of diets (e.g., pisci vore, probing in secti vore) likely to result in
contact wi th contami nated environmental media;
1-6


Table 1-1. Character isti cs o f Selected Birds
Diet General Foraging Ha bit at Body Size Selected Bird Species
Insecti vorea
probing/ soil- dwelling inv ertebratesgleaning/ insects
Herbiv ore gleaning/ seeds grazing/ shoot s
Omniv ore
Carnivoreb
Carnivore/Pisci vore/Scaven ger smal l birds & mammal s/fish/dead fish fish/inver tebrates/smal l birds/garbage
cPisci vore
Aquatic Insecti vored
probing/ soil- dwelling inv ertebrates div ing/ aquatic inv ertebrates
Aquatic Herbivore/Insecti vore
woodla nds , marshes marshes
woodla nds , fields and br ush open f ields
open woodla nd, subur bs
open fields, forest ed ge most ope n areas
open water bodie s Great Lakes an d coastal
most str eams, rivers, smal l lakes most fr eshwater and sal twater bodie s large water bodie s
most r ivers and str eams oceans and coastal areas
most wetl ands, ponds
medium smal l
medium large
smal l
medium medium
large medium
medium
large
large
smal l medium
medium
Amer ican woodc ock marsh wren
nor thern bobw hit e Canada goos e
Amer ican robin
Amer ican kestr el red-taile d hawk
bald eagle herring gull
belted kingf isher
great blue heron
osprey
spotted sandpipe r lesser scau p
malla rd
1-8
aIncludes consum pt ion of insect s, other arthropods, wor ms, and ot her terrestrial invertebrates.bIncludes consum ption of terrestrial vertebrates and l arge invertebrates. cIncludes consum ption of fish, amphi bians, crust aceans, and ot her larger aquat ic ani mals.dIncludes consum pt ion of aquat ic invertebrates and am phi bian larvae by gleaning or probi ng.

Table 1-2. Character isti cs o f Selected Mammal s
Diet General Foraging Ha bit at Body Size Mammal Species Selected
Insecti vorea
gleaning/ surface-d welling inv ertebrates most h abitat typ es
most d ry-land habitats grassy fi elds, marshes, bogs prairie grass co mmunit ies most h abitat typ es
woodla nds , subur bs mixed woodla nds and ope n areas
most ar eas near water
rivers coastal , estu aries, lakes
smal l
Herbiv ore gleaning/ seeds smal l deer mouse grazing or browsing/ shoot s, root s, or smal l meadow vo le leaves smal l prairie vole
medium easter n cotto ntai l
Omniv ore medium racco on medium red fox
Carnivore medium minkb
Pisci vore medium river o tterc
medium harbor seal
Aquatic Herbivore most aq uatic habitats medium muskr at
short-tai led shrew
1-9
aIncludes consum pt ion of insect s, other arthropods, wor ms, and ot her terrestrial invertebrates.bIncludes consum ption of aquat ic and t errestrial vertebrates and l arge invertebrates. cIncludes consum ption of fish, amphi bians, crust aceans, molluscs, and ot her large aquat ic ani mals.

Table 1-3. Character isti cs o f Selected Repti les and Amphibia ns
Adult Diet for Adult s Body Size Amphibia n Species General Foraging Ha bit at Selected Repti le or
REPTILES
Terrestr ial Carnivorea
Aquatic Pisci voreb
Omnivore
Aquatic Herbivore
AMPHIBIANS
Insecti vorec
Aquatic Pisci vore/Insecti vored
open woods , fields and br ush
most typ es of water bodie s
open fields, forest ed ge, marshes most fr eshwater bodie s
most wetl ands, ponds
shallow fr eshwater bodie s
lakes, ponds , bogs , str eams smal l lakes, ponds , str eams
medium
medium
medium
large
medium
smal l
medium smal l
racer
nor thern water snake
easter n box tu rtle
snapping t urtle
paint ed turtle
green frog
bullf rog easter n newt
1-10
aIncludes consum pt ion of terrestrial vertebrates and i nvertebrates, insect s, other arthropods, wor ms, and ot her terrestrial invertebrates.bIncludes consum pt ion of fish, amphi bians, and cr ust aceans. cIncludes consum pt ion of insect s, other arthropods, wor ms, and ot her terrestrial invertebrates.dIncludes consum pt ion of fish, amphi bians, crust aceans, molluscs, other aquat ic ani mals, and t errestrial insect s and ot her invertebrates.

�
�
� Species of so cietal signif icance or conc ern (e.g., game sp ecies, familia r
species); and
� Species th at have been shown to be particularly sen si tive to the str esso r
being a ddr essed .
When species of co ncern for a risk assessmen t include sp ecies for whic h data are
presented in th is Handbook , it can serve as a readily avai lable source of data fo r
scr eening- level exposure analyses.
1.4. LIST OF EXPOSURE FACTORS
Three routes o f exp osure may b e of co ncern for wildlif e in t he vicinit y of
contami nated surface water s and ter restr ial habitats: oral, inha lation, and de rmal. Oral
expos ures might occur vi a ing estion of con tami nated food ( e.g., aquatic pr ey) or water or
inc idental inge stion of con tami nated media (e.g., soils , sedim ents) d uring f oraging or other
acti vi ties. Inhalation of g ases o r particulates m ight be a signif icant rout e of expos ure for
some anim als. Dermal expos ures are lik ely to be mo st signif icant for bur rowing mammal s
(i.e., via contact wi th contami nated soils ) and anim als that spend c ons iderable amount s of
time su bmerged in surface water s. This Handbook tabul ates sel ected data fo r all three
routes o f exp osure (Table 1-4), emphasizing or al expos ures. It also prov ides qua nt itati ve
inf ormation on popula tion pa rameter s and qualitati ve informat ion r elated to seaso nal
acti vi ties, geographic r anges, habitats, and other life-histo ry ch aracter isti cs.
The exposure facto rs presented in the Handbook ar e conc eptually sep arated into
four typ es: nor malizing facto rs (Sect ion 1 .4.1), con tact r ates (Sect ion 1 .4.2), popula tion
dynamics (Sect ion 1 .4.3), and seaso nal acti vi ties (Sect ion 1 .4.4). Sect ion 1 .5 descr ibes th e
format i n which values fo r these exp osure facto rs are presented in Chapter 2.
1.4.1. Norm alizing Factors
Normalizing facto rs include body weight , growth rate, and metabolic r ate, which are
discu ssed in turn below.
1-11

1.4.1.1. Body Weight
Body weight s (in unit s of mass) ar e reported as fr esh weight as might be obt ained
by weighing a liv e anim al in t he field. Several of th e contact r ate parameter s are
nor malize d to body weight . For examp le, both food a nd water inge stion r ates ar e reported
on a per body weight basis (e.g., gram of fr esh food or water per gram of fr esh body weight
per day). Using e mpir ical mode ls , body weight d ata also were used to esti mate co ntact
rate parameter s fo r which we could not find measu red values.
Adult body weight s are listed for all species. For birds, we also provide egg weight ,
weight at hatching, nestling or chic k weight s, and w eight at fledging, when availa ble , to
assi st r isk assesso rs co ncerned wi th esti mat ing e xpos ures of emb ryos and y oung bir ds.
For mammal s, we also provide gestat ing femal e weight , bir th weight , pup w eight s at
var ious ag es, weight at weaning, and w eight at sexu al matu rity, when avai lable, for a
simila r pur pos e. Fina lly , for rept iles and amphibia ns, we also prov ide egg w eight , larval or
juv enile weight s wi th age, and w eight at metamo rphos is , if availa ble and a pplic able. Body
size fo r rept iles and amphibia ns is o ften reported as body lengt h in stead of body weight ,
so we also provide data on body lengt h and t he relations hip b etween body lengt h and body
weight , when availa ble .
1.4.1.2. Growth Rate
Young a nim als generally cons ume more food ( per unit body weight ) than adult s
because th ey grow and develop r apidly . Growth rates ch ange as animals matu re, whether
expressed as absolute (g/day) or relative (percent body weight ) terms. Weight gain is r apid
after birth, but sl ows o ver time. Differ ent typ es of animals exhibit di ffer ent patter ns of
growth over ti me. Plot s of body weight ver sus age for so me anim al groups ar e sigm oida l
whereas others may ap proximate logis tic func tions or other sh apes. As a result ,
investi gators often report growth rates as var ious con stan ts asso ciated wi th particular
math emati cal models (e.g., Gompertz equation, von Be rtalanffy eq uation; see Peter s, 1983)
that fi t th e growth patter n for a given species. Instead of presenting a var iety o f growth
constan ts an d models, however , we report growth rates fo r young a nim als, wh en availa ble ,
1-13

in grams p er day fo r speci fic age groups . Growth rates al so can be infer red from a ser ies
of juv enile body weight s wi th age. These measu res are include d unde r body weight (see
Sect ion 1 .4.1.1).
1.4.1.3. Metabolic Rate
Metabolic r ate is reported on the basis of k iloc alor ies per day nor malize d to body
weight (e. g., kcal /kg-day). If meta bolic r ate was measu red and reported on the basis of
oxygen cons umpt ion only , we pr ov ide thos e value s as lit ers O2/kg-day. Normalize d
metabolic r ates b ased on kilocalories can be used to esti mate n ormalized food ing esti on
rates (see Sect ion 4 .1.2). Metabolic r ates b ased on oxyg en consumption c an be used to
esti mate meta bolic r ates b ased on kilocalories fo r subsequent use in esti mat ing f ood
inge stion r ates (see Sect ion 3 .6.3.1).
1.4.2. Contact Rate Factors
Table 1-5 summar izes th e six co ntact r ate facto rs include d for th e oral, inha lation,
and dermal routes o f exp osure.
1.4.2.1. Oral Route
Three environm ental media ar e the pr imary cont ribut ors to wildlif e expos ure by th e
oral rout e: food, water , and soils and s edim ents. Four con tact r ate exp osure parameter s
related to these th ree exposure media are discu ssed below.
1.4.2.1.1. Food i ngesti on rates. Food ing estion r ates ar e expressed in th is
Handbook as gr ams o f food (wet we ight ) per gram of body weight (wet we ight ) per day
(g/g-day). Food ing estion r ates can vary by age, size, and sex an d by seaso nal changes in
ambient temp erature, acti vi ty l evels, reproducti ve acti vi ties, and the typ e of diet co nsumed.
Food ing estion r ates h ave not been measu red for many wildlif e species. Methods fo r
esti mat ing f ood ing estion r ates o n the basis of fr ee-living ( or fi eld) metabolic r ate, energy
content of th e diet, and assi mi lation effi ciency ar e discu ssed in Sect ion 4 .1.2.
1-14

Table 1-5. Wildlif e Contact R ate Exp osure Facto rs
Expos ure Rout e Medium Facto r Expressi on Unit s
ORAL Food ing estion r ate fraction body weight
6 g/g-day
dietary compos it ion
fraction of to tal intake represented by each food t ype
Water inge stion r ate fraction body weight
g/g-day
Soil/ Sedim ent intake r ate fract ion of to tal food intake
g/g-day
INHALATION Vapor or Particulates
inha lation r ate daily volum e m /day3
DERMAL Water or Soil/ Sedim ent
surface ar ea total area potentially exposeda
cm 2
aTotal unpr otected or permeable sur face area that might be exposed under som e ci rcumstances ( e.g., dust bathing), even t hough i t woul d not be exposed under other condi tions ( e.g., swi mming wi th a trapped ai r layer between the feathers or fur and ski n).
1.4.2.1.2. Dietary co mposi tion. Dietary co mposi tion v aries seaso nally and by age,
size, reproducti ve statu s, and habitat. Dietary co mposi tion ( e.g., propor tion of die t
cons is ting of var ious plant or anim al mater ials), often measu red by sto mach -content
analyses, is exp ressed whenever possible as pe rcentage of to tal intake o n a wet-we ight
basis. This conv ention faci litates co mparison wi th contami nant co ncentrations in di etary
items r eported on a wet-we ight basis. Methods fo r conv erting ot her measu res of dietary
compos it ion ( e.g., percentage of to tal prey items cap tured, propor tion of intake o n a dry-
weight basis) to esti mates o f dietary intake o n a wet-we ight basis are prov ided in Secti on
4.1.2.
1.4.2.1.3. Water ingesti on rates. For drinking-water exposures, inge stion r ates ar e
expressed in th is Handbook as gr ams o f water per gram of wet body weight per day (g/g
day). Water consumption r ates d epend on body weight , phy siologic al adaptations ,
1-15

diet, temp erature, and acti vi ty l evels. It is important to rememb er that, unde r so me
condit ions , some species can meet th eir water requir ements wi th only th e water contained
in the diet and metabolic water produc tion (see Sect ion 3 .2).
1.4.2.1.4. Incidental soil and sediment intakes. Wildlif e can inc identally inge st
soils or sedim ents while fo raging or dur ing ot her acti vi ties su ch as dust b athing a nd
preening or groom ing. Data quanti fy ing s oil a nd s edim ent ing estion a re lim ited; we
present avai lable values fo r selected species in Sect ion 4 .1.3.
1.4.2.2. Inhalation Route
Average daily inha lation r ates ar e reported in the Handbook in unit s of m 3/day.
Inhalation r ates var y wi th size, seaso nal acti vi ty l evels, ambient temp erature, and daily
acti vi ties. EPA's cu rrent approach to calculating inha lation e xpos ures requir es addit iona l
inf ormation on s pecies' respir atory phy siology to fully esti mate inha lation e xpos ures (see
Sect ion 4 .1.4).
1.4.2.3. Dermal Route
Dermal contact wi th contami nated soil, sedim ent, or water is l ikely to be an
exposure pathway fo r some wildlif e species. An anim al's surface ar ea could be used to
esti mate th e potential for uptake o f co ntami nants th rough it s skin. For some expos ures
(e.g., dus t bathing) , the enti re surface ar ea of th e animal might be impor tant. For other
typ es (e.g., swi mming) , only th e unins ulated portions (e. g., no f ur or feath ers th at cr eate a
trapped air layer ) of th e animal might con tact th e contami nated medium . In the Handbook ,
we provide measu res or esti mates o f th e enti re potentially exp osed surface ar ea of an
anim al, when pos sible . We have not attemp ted to determine what portions would be
exposed and protected for swi mming a nim als.
1-16

1.4.3. Population Dy namics
Several parameter s can be used to descr ibe the spatial distr ibut ion a nd abunda nce
of a popula tion of anim als in r elation t o the spatial exten t of co ntami nation. Three
parameter s related to spatial distr ibut ion a re social organization, home-range size, and
popula tion de nsity. These are important fo r esti mat ing t he num ber of indiv idua ls or
propor tion of a popula tion t hat might be expos ed to a con tami nated area. Parameter s
related to popula tion s ize and pe rsisten ce include ag e at sexu al matu rity an d matu ration,
mortalit y, and a nnua l fecundit y rates. These parameter s may b e usefu l to assesso rs
planning or evalua ting f ield s tudie s or monit or ing pr ogr ams.
1.4.3.1. Social Organization
The Handbook inc lude s a quali tati ve descr iption of each species' social
organization, whic h inf luences h ow animals of var ious ag es and s izes are dis tr ibut ed in
space. In some sp ecies, indiv idua l hom e ranges do not overlap. In ot hers, all indiv idua ls
use th e same h ome range. In between these extr emes, home ranges can be shared wi th
mates, offsp ring, or exten ded fami ly groups .
Social organization c an vary sub stan tially among s pecies that appear otherwis e
simila r; therefore, it is not pos sible to extr apolate th e social organization of si mila r sp ecies
from th e selected species in th is Handbook . Cons ult th e general bibliogr aphie s for
inf ormation s our ces to determine the social organization of sp ecies not co vered in t he
Handbook .
1.4.3.2. Home Range/Territ ory Size/Foraging Radius
Home range size can be used to determine the propor tion of ti me that an indiv idua l
animal is exp ected to contact co ntami nated environmental media. Home range is defined
as the geogr aphic ar ea encompassed by an animal 's acti vi ties (excep t mi gration) over a
speci fied time. While hom e range value s often are expressed in units o f area, for species
dependent on riparian or coastal habitats, a more meaningf ul measu re can be foraging
1-17

radius , or the distan ces th e animals are willing t o travel to potential food s our ces. Although
home ranges may b e roughly ci rcula r in hom ogeneous habitats, it is important to
rememb er that depending on ha bitat n eeds and condit ions , hom e ranges may b e irregula r
in shape. The size and spatial attr ibutes o f a home range often are defined by fo raging
acti vi ties, but also might depend on t he location of sp eci fic resour ces su ch as dens or nest
si tes i n other areas. An animal might not vi si t all areas of i ts h ome range every day or
even every week, but over longe r ti me periods , it can be exp ected to visi t mo st o f th e areas
wi thin t he hom e range th at cont ain ne eded resour ces su ch as fo rage, prey, or protected
resting a reas.
Home range size fo r indiv idua ls wi thin a popula tion c an vary wi th seaso n, lati tude,
or alti tude as a cons equence of ch anges in t he dis tr ibut ion a nd abunda nce of food or other
resources. It generally var ies wi th animal body size and a ge because of differ ences i n the
dis tr ibut ion of prefer red forage or prey. It can also depend on habitat q uality, increasing a s
habitat q uality d ecreases to a condit ion be yond w hic h the habitat d oes not su stai n even
sparse popula tions . Fina lly , hom e ranges can vary by sex an d seaso n. For examp le, if a
femal e is respons ible fo r mo st or all of th e feeding of young, her fo raging r ange might be
restr icted to an area close to her nest o r den when she has dependent young, whereas th e
foraging r ange of mal es would not be so restr icted .
Nonter ritorial species may a llow signif icant ov erlap of acti vi ty ar eas amo ng
neighbor ing indiv idua ls or groups . For examp le, sever al indiv idua ls or mated pairs may
share the same ar ea, although s igna lling be havior s may en sure temp oral segregation. For
these sp ecies, we report a home range size or foraging r adius . Other sp ecies are str ongly
ter ritorial and defend mutually excl usive areas: indiv idua ls , breeding pa irs, or family unit s
acti vely adver tise identi fiable bounda ries and exclude neighbor ing indiv idua ls or groups .
Foraging acti vi ties are usually restr icted to the defended ter ritories. For these sp ecies, we
repor t the size of th e defende d territory and not e whether fo raging oc curs out side of th e
terri tory .
1-18

1.4.3.3. Popul ation Density
Popula tion de nsity (the num ber of anim als per unit ar ea) influences h ow man y
indiv idua ls (o r wh at pr opor tion of a loc al popula tion) might be expos ed within a
contami nated area. For str ongly ter ritorial species, popula tion de nsity can be inf erred
from ter ritory si ze in many cases. For species wi th over lapping hom e ranges, partic ularly
colonia lly breeding a nim als (e.g., most seabir ds), popula tion de nsity cannot be inf erred
from hom e range size .
1.4.3.4. Annua l Fecundi ty
Attr ibutes r elated to the number of offsp ring pr oduc ed each year that reach sexu al
matu rity (annua l fecundi ty) ar e measu red in differ ent ways d epending on t he lif e his tory of
the species. For birds, data are generally avai lable for clutch size, number of cl utch es per
year , nest su ccess (g enerally reflect ing pr edation pr essu re), number of young f ledged per
successfu l nest (g enerally reflect ing f ood availa bili ty), and number of young f ledged per
acti ve nest (r eflect ing a ll c auses o f mo rtal ity). For mammal s, litter size in wild popula tions
often is determined by placental scar s or embryo count s, and t he num ber of young
surviv ing t o weaning is seldom know n. For rept iles that lay eggs , clut ch size and pe rcent
hatching c an be measu red in the field. For viviparous repti les, we report th e number born
in a lit ter . For amphibia ns, egg masses may i nclude thous ands of eggs , but th ese are
seldom count ed.
1.4.3.5. Annua l Mor tal ity and Longe vi ty
Longe vi ty can inf luence the pot ential for cu mula tive de leter ious effects an d the
appropriate aver aging t imes fo r chronic expos ures. For bir ds, annua l adult mo rtalit y tends
to be constan t. For large mammal ian species, however , annua l adult mo rtalit y tends to be
constan t fo r sever al years, and then increases r apidly wi th age. For rept iles and
amphibia ns, annua l adult mo rtalit y can decrease wi th age for some ti me as th e animals
cont inue to grow larger and be come less su scep tible to predation. In the Handbook , we
1-19

�
�
�
�
repor t annua l mortalit y rates b y age categ ory and typ ical or mean and maxi mum
longe vi ties, wh en pos sible .
1.4.4. Seasonal A ctiv ities
Many l ife-cycl e attr ibutes affect an animal 's acti vi ty an d foraging p atter ns in time
and space. For examp le, many sp ecies of birds are present in the northern hemisphere
only dur ing t he warmer mont hs or move seaso nally between the northern and southern
parts o f North Amer ica. Some sp ecies of mammal s, repti les, and amphibia ns hibe rnate or
spend a dor mant period in a bur row or den dur ing t he wint er mont hs. The species pr of iles
descr ibe these and other seaso nal acti vi ty p atter ns th at can influence exp osure frequency
and dur ation.
For each species, we su mmar ize informat ion on t he seaso nal occu rrence of sever al
acti vi ties including br eeding, molt ing, migr ation, dis persal , and oc currence of
dor mancy/denning ( if applic able). Depos it ion a nd ut iliza tion of fat r eserves ar e discu ssed
where inf ormation is avai lable. Trends in these facto rs wi th lati tude are identi fied.
1.5. DATA PRESENTATION FORMAT
Species-sp eci fic val ues fo r the exposure facto rs are presented in Chapter 2.
Quanti tati ve data fo r each species are presented in tables ar ranged in four main sect ions :
� Normalizing a nd Con tact R ate Facto rs;
� Dietary Composi tion;
� Popula tion Dy namics; and
� Seasonal Acti vi ties.
The parameter values and units u sed for each exposure facto r are descr ibed in the
remainde r of this sect ion. In the species pr of iles and in t he Appendix , all v alues are
identi fied as measu red or esti mated , and references ar e provided.
1-20

1.5.1. Norm alizing and Contact Rate Factors
Normalizing a nd c ontact r ate facto rs are presented unde r the heading "F acto rs" in
Chapter 2. Several of th em apply to all anim als inc lude d in t he Handbook , whereas so me
apply only to speci fic gr oups , as descr ibed in Sect ions 1. 5.1.1 through 1 .5.1.4. The colum n
headers fo r these facto rs are explained in Table 1-6.
Table 1-6. Colum n Headers for Tables of Normalizing a nd Con tact R ate Facto rs
Age/Sex/ Age (e.g., A for adult , J for juv enile ) Cond. /Seas. Sex (e.g., M for male, F for femal e)
Condit ion ( e.g., I for inc ubating, NB for nonbr eeding) Season (e.g., SP for spring, SU for su mmer ).
[Not e: Only inf ormation ne eded to correctl y interpret th e value is inc lude d.]
Mean Mean value fo r popula tion sample d ± standa rd deviation ( SD), if reported. If SD is not reported, mean value for popula tion sample d ± stan dard error (SE) of th e mean, if reported. For some st udies, a range of typ ical values may b e presented instead of a mean value (check th e notes).
Range or Range of value s repor ted for th e popula tion sample d, or (95% CI of (95th percent co nfidence interval of th e mean value). Mean)
Locati on State(s) o r province(s) i n which the stu dy was c onducted (subs pecies) (subs pecies studie d, if r epor ted).
Reference Reference fo r stu dy.
Note No. Foot not e num ber.
1.5.1.1. All An imals
Body weight (g rams o r Measured values only . Although w e use the term kilogr ams) weight , all d ata are presented in units o f mass. The age
and sex o f th e animal are speci fied as appropriate, and
1-21

Metabolic r ate (l iters O2/kg-day)
Metabolic r ate (kcal /kg-day)
Food ing estion r ate (g/g-day)
Water inge stion r ate (g/g-day)
Sedim ent/soil ing estion r ate
Inhalation r ate (m3/day)
Surface ar ea (cm2)
1.5.1.2. Bird s
Egg w eight (g rams)
Weight at hatching ( grams)
weights may i nclude ag e-weight ser ies for young animals.
Include d only if measu red values wer e avai lable. These data can be used to esti mate meta bolic r ate on a kcal basis.
Measured or esti mated basal and free-living ( or fi eld) metabolic r ates. Most o f th e free-living v alues were esti mated from body weight us ing a n appr opr iate allometr ic equation.
Measured on a wet-we ight basis. For bir ds an d mammal s, values measu red in captivi ty ar e generally lower than for free-ranging a nim als. For rept iles and amphibia ns, food ing estion r ates can be highe r in captivi ty than in t he field. Food ing estion r ates can also be differ ent in captivi ty th an in the wild if th e die t di ffer s substan tially fr om th at co nsumed in the wild ( e.g., dry laboratory ch ow has a su bstan tially lower water content than most n atural diets).
Most o f th ese val ues wer e esti mated from body weight us ing a n allo metr ic equation.
These val ues are not presented in the indiv idua l species profi les in Chapter 2; instead , the limi ted data avai lable for soil/ sedim ent ing estion r ates (as p ercent soil or sedim ent in die t on a dry weight basis) for sel ected species are presented in Sect ion 4 .1.3.
Note th at th is val ue is not normalized to body weight , but is th e total volum e inha led each day. Most val ues were esti mated from body weight us ing a n appr opr iate allometr ic equation.
Most val ues wer e esti mated from body weight us ing a n appropriate allometr ic equation.
Include d only if measu red values wer e avai lable.
Include d only if measu red values wer e avai lable.
1-22

Chic k or nestling gr owth rate (g/day)
Weight at fledging ( grams)
1.5.1.3. Mammal s
Neonate weight (g rams)
Pup growth rate (g/day)
Weight at weaning ( grams)
1.5.1.4. Repti les and Amphibians
Body lengt h (mm)
Egg w eight (g rams)
Weight at hatching ( grams)
Juvenile growth rate (g/day)
Tadpole weight (g rams)
Larval or eft we ight (g rams)
Inc lude d only if measu red values wer e avai lable. The ages to which the growth rate applie s are indi cated .
Include d only if measu red values wer e avai lable.
Include d only if measu red values wer e avai lable.
Include d only if measu red values wer e avai lable. The ages to which the growth rate applie s are indi cated .
Include d only if measu red values wer e avai lable.
Length is th e most co mmo n measu re of si ze and growth rate reported for repti les and amphibia ns. Body lengt h-weight r elations hips ar e repor ted whenever poss ible . Data fo r snakes i nclude snou t-to -vent lengths (SVL) and total lengt hs; for fr ogs , SVLs only ; and f or tu rtles, carapace (dorsal shell) and plastr on (ventral shell) lengt hs.
Include d only if measu red values wer e avai lable.
Include d only if measu red values wer e avai lable.
Include d only if measu red values wer e avai lable. The ages to which the growth rate applie s are indi cated .
For frogs only ; inc lude d only if measu red values wer e availa ble .
For newts only ; inc lude d only if measu red values wer e availa ble .
1-23

1.5.2. Dietary Composition
1.5.2.1. All An imals
The diet of al l animals is sep arated by seaso n whenever possible . Up to three
months of data wer e combine d for each of th e four seaso ns, provided the animals wer e in
the same l ocat ion a nd ha bitat d uring t he 3-mont h period ( Table 1-7). The diet co mpone nts
are listed in the fi rst c olum n shaded in gr ey. The measu re of dietary co mposi tion is
enclosed in parentheses unde r the "Lo cation ( subs pecies)/Habitat (measu re)" colum n
header.
Table 1-7. Colum n Headers for Tables on Di etary Composi tion
Dietary Composi tion List o f food t ypes.
Spring Dietary co mposi tion dur ing s pring ( March, April, May). Summer Dietary co mposi tion dur ing s ummer (June, July, August). Fall Dietary co mposi tion dur ing f all (Septemb er, Octo ber,
Novemb er). Winter Dietary co mposi tion dur ing w int er (Decemb er, January,
February).
Locati on State(s) o r Canadian province(s) i n which stu dy was (subspecies)/ conducted (subspecies st udie d, if r epor ted). Habitat Type of habitat asso ciated wi th the reported values (measu re) (measu re used to quanti fy d ietary co mposi tion) .
Reference Reference fo r stu dy.
Note No. Foot not e num ber.
Dietary co mposi tion c an be exp ressed in many ways. In the Appendix , we have
presented all measu res of dietary co mposi tion e ncount ered in t he lit erature review. In the
species profi les in Chapter 2, we have emp hasized dietary co mposi tion measu red as th e
percentage of th e total food in take o f each food t ype on a wet-we ight basis. These data
1-24

are usually determined by analysi s of sto mach or other digesti ve tr act co ntents. For
entr ies based on these measu res, the total of th e values l isted unde r each seaso nal
colum n should a ppr ox imate 100 p ercent. As Chapter 4 indi cates, it is relatively si mple to
esti mate co ntami nant intakes wh en dietary co mposi tion is measu red on a wet-we ight
basis. Dietary co mposi tion may al so be measu red on a dry-weight basis; inf ormation on
the relative water content of th e differ ent dietary i tems p rovided in Chapter 4 can be used
to conver t dry-weight co mpos it ion t o wet-we ight co mpos it ion if needed. Dietary
compos it ion is o ften reported as fr equency o f occu rrence in digesti ve tr act co ntents,
scats, or regurgitated pellets. For these measu res, the total of th e values in the seaso nal
colum ns can exceed 100 (e.g., fish occu rred in 90 percent of scats, amphibia in 75 percent
of scats, and mollu scs i n 15 percent of scats). We do not provide guida nce on how to
esti mate co ntami nant intakes b ased on these measu res; however , studie s us ing t hese
measu res can indi cate seaso nal and geographic di ffer ences i n diet.
1.5.3. Population Dy namics
Dis tr ibut ion a nd m ortalit y parameter s can be defined simi larly fo r birds, mammal s,
rept iles, and a mphibia ns (Sect ion 1 .5.3.1). Reproduc tive parameter s, however , differ
among t hese groups (Sect ions 1. 5.3.2 through 1 .5.3.5). The colum n headers for popula tion
dynamics ar e descr ibed in Table 1-8.
1.5.3.1. All An imals
Home range size (ha)/ Area usually l isted in hectar es, radius in k ilo meter s. The home Territory size (h a)/ range fo r sp ecies such as mink or kingf ishers, whic h spend Foraging r adius (m) most of th eir ti me along s hor eline ar eas, is so meti mes
descr ibed as k ilometer s of sh oreline . For some species wi th extr emely smal l breeding t erritories, we us ed m2 instead of hectar es. For colonia lly n esting bir ds, foraging r adii a re listed in ki lometer s. For frogs, we found inf ormation only on m ale breeding t erritory size, whic h doe s not inc lude th e foraging range of ei ther sex.
Popula tion de nsity Usually listed as number (N) of indiv idua ls per hectar e, (N/ha) although num bers of breeding pa irs or nests p er hectar e are
used for some sp ecies.
1-25

Table 1-8. Colum n Headers for Tables of F acto rs fo r Popula tion Dy namics
Age/Sex/ Age (e.g., A for adult , J for juv enile ) Cond. /Seas. Sex (e.g., M for male, F for femal e)
Condit ion ( e.g., I for inc ubating, NB for nonbr eeding) Season (e.g., SP for spring, SU for su mmer ).
[Not e: Only inf ormation ne eded to correctl y interpret th e value is inc lude d.]
Mean Mean value fo r popula tion sample d ± standa rd deviation ( SD), if reported. If SD is not reported, mean value for popula tion sample d ± stan dard error (SE) of th e mean, if reported. For some st udies, a range of typ ical values may b e presented instead of a mean .
Range Range of value s repor ted for th e popula tion sample d.
Locati on State(s) o r province(s) i n which the stu dy was c onducted (subspecies)/ (subspecies st udie d, if r epor ted). Habitat Type of habitat asso ciated wi th the reported values.
Reference Reference fo r stu dy.
Note No. Foot not e num ber.
Age at sexu al matu rity Age at wh ich fi rst su ccessfu l reproduc tion oc curs. In many long- liv ed species, only a por tion of th e popula tion br eeds at th is age.
Annua l mortalit y rates Usually l isted as percent per year . Can vary wi th age and sex of th e animal.
Longe vi ty Mean longe vi ty of adult memb ers of th e popula tion ( does not inc lude juv enile mo rtali ty). When avai lable, an esti mate o f maxi mum longe vi ty is al so prov ided (usually fr om studie s of captive indiv idua ls).
1.5.3.2. Bird s
Clutch size Number of eggs laid per acti ve nest (u sually th e number laid per femal e, but in some sp ecies, more than one femal e may l ay in a single n est).
1-26

Clutch es per year Number of su ccessfu l clutch es laid per year . Addit iona l clutch es may b e laid if a cl utch is lost ear ly in incubation.
Days i ncubation Measured from day incubation star ts (o ften after laying of last egg) to hatching.
Age at fledging Age at whic h young c an maint ain s ustaine d f light . Parents usually cont inue to feed or to acco mpany young f or so me time after fledging.
Number fledged per Number fledged for each nest fo r which incubation w as acti ve nest init iated.
Percent nests Percent of acti ve nests h atching e ggs . successfu l
Number fledged per Number fledged for each nest fo r which at least o ne young successfu l nest hatch ed.
1.5.3.3. Mammal s
Litter size Based on embryo counts wh enever possible . Use of placental scar s can result in overesti mat ion of li tter size and count s of liv e pups in de ns can result in unde resti mat ion of li tter size.
Litter s per year Number of l itter s born each year .
Days g estati on Days o f acti ve gestat ion. For species wi th delayed implantation, this period c an be sub stan tially sh orter than the period f rom mating t o bir th.
Pup growth rate Usually reported as grams p er day during a sp eci fied age interval . May be reported instead as a ser ies of weights fo r pups of sp eci fied ages.
Age at weaning Age when the pups begin t o leave th e nest o r den to acti vely feed for most o f th eir food.
1.5.3.4. Repti les and Amphibians
Clutch or litter size Number of eggs laid per femal e for egg-laying s pecies; num ber of l ive offsp ring bor n for sp ecies bearing liv e young ( e.g., water snake). Reported by age and size of th e femal e when appropriate.
1-27

Clutch es or litter s Number of cl utch es or litter s produced each year . Not l imi ted per year to successfu l clutch es because th ere is no parental care in
most temp erate sp ecies.
Days i ncubation Measured from laying of l ast eg g to hatching. The dur ation of inc ubation de pends on t he temp erature of th e substr ate into whic h eggs ar e laid.
Juvenile growth rate Usually r epor ted as gr ams p er day during a sp eci fied age (or size) interval . May be reported instead as a ser ies of weight s for juv enile s of sp eci fied sizes if thos e are the only data availa ble .
Length at sexu al matu rity Length at wh ich the fi rst su ccessfu l reproduc tion us ually occu rs (see ab ove). More commonly r epor ted than weight or age at sexu al matu rity.
1.5.4. Seasonal A ctiv ities
The meaning of mo st of th e facto rs include d unde r seaso nal acti vi ties are sel f-
evide nt . Thos e requir ing a ddit iona l expla nation a re descr ibed in Sect ions 1. 5.4.1 through
1.5.4.3. The colum n headers for this sect ion of th e table ar e show n in T able 1-9.
Table 1-9. Colum n Headers for Tables on Seasonal Acti vi ties
Begin Mont h that the acti vi ty u sually begins .
Peak Mont h(s) th at th e acti vi ty p eaks (mo st o f th e popula tion is inv olv ed).
End Mont h that the acti vi ty u sually ends.
Locati on State(s) o r province(s) i n which the stu dy was c onducted (subs pecies) (subs pecies studie d, if r epor ted).
Reference Reference fo r stu dy.
Note No. Foot not e num ber.
1-28

1.5.4.1. Bird s
Mating/ laying These two facto rs are combine d because birds lay eggs wi thin a day or two of mating ( they begin mating a day or two pr ior to laying t he fi rst egg) .
1.5.4.2. Mammal s
Mating Although f or mo st mammal s th e mat ing seaso n corresponds to conc eption a nd is follow ed immed iately by gestat ion, some species exhibit delayed implantat ion.
Parturition Bir th of th e pups (al so know n as whelping f or canids ).
1.5.4.3. Repti les and Amphibians
Mating Because fer tiliza tion is exter nal for many amphibia ns (i .e., most toads and f rogs and s ome salamande rs), mating oc curs at the same ti me as eg g-laying f or th ese sp ecies. For repti les, fer tiliza tion is int ernal, and f or so me species, sp erm may b e sto red for months or years follow ing mating.
Nesti ng Because man y femal e repti les can sto re sperm, nest ing ( i.e., egg- laying) o ften occu rs weeks o r months after mat ing.
1.5.5. Abbrev iations Used in T ables
Age (life stag e)
A adult (fo r all gr oups )B both adults an d juveniles/year lings (fo r all gr oups )C chic k (for bir ds)E eft (fo r newts)F fledgling ( for bir ds)H hatchling ( for bir ds, rept iles, and a mphibia ns)J juv enile (fo r all gr oups )N nestling ( for bir ds)
or neonate (fo r mammal s, water snakes)
P pup ( for mammal s) T tadpole (fo r fr ogs ) Y year ling ( for all gr oups )
1-29

�
Sex
B both sexes F femal eM male
Unit s
time: d day wk week yr year
mass: g gram kg kilogr am
lengt h: mm milli meter cm centimeter m meter km ki lometer
temp erature: �C degrees Centigrade
Other
NS not stated
energy: cal kcal
area: ha
2m
volum e: ml l
calorie kiloc alor ie
hectar e squa re meter
millilit er liter
1.6. LITERATURE SEARCH STRATEGY
The prof iles in t his Handbook ar e int ended to prov ide a readily availa ble
compendium of r epresentati ve data fo r each selected species to assi st i n conduc ting
scr eening- level exposure assessmen ts. They are not intended to provide complete
reviews o f al l avai lable publis hed and unpublis hed inf ormation or inde pth biologic al
summar ies. Moreover , the Handbook is not int ended to replace fi eld guide s or natural
his tory or anim al phy siology texts. We have attemp ted to balance generalities, accu racy,
and coverage of each species relative to the avai lable literature to meet o ur stated
purposes. We descr ibe the process b y wh ich we identi fied literature for the Handbook
below .
1-30

The U.S. Fish and Wildlif e Service (USFWS) Offi ce of Informat ion T ransfer
condu cted the primary l iterature search for species-sp eci fic informat ion us ing t heir Wildlif e
Review/Fisheries Review database. The database is co mpile d by USFWS personne l f rom a
review of over 1,130 publi cation s our ces (l argely jour nals , but al so USFWS publi cations )
from th e United States an d other count ries, mo st da ting b ack to 1971. The search was
condu cted in May 1990 using c ommon a nd s cienti fic species names, but no further
restr ict ions on search terms wer e applie d. All t itles ide nt if ied for each species wer e
reviewed to determine potential utilit y for th e Handbook , and pr omis ing r eferences wer e
reviewed in full. Recent review ar ticles, handbook s, and na tural his tory texts wer e used to
identi fy o ther relevant l iterature and literature from before 1971. Commer cial databases
were not sear ched init ially . Follow ing pe er review of th e Handbook in 1991 and 1992, all
references su bmitted or identi fied by peer reviewers wer e evaluated, and addit iona l
relevant ci tat ions wer e obt ained for review. Lim ited (1970 forward) literature searches fo r
some species were condu cted using c ommer cial databases i n 1992.
For inf ormation c onc erning phy siology , allo metr ic equations , energetics, and ot her
general topics, literature was i denti fied on the basis of recent review ar ticles or book s in
the field suggested by exp erts i n the field and by peer reviewers.
Because of resource l imi tat ions , we have include d some value s from seconda ry
ci tations . In these cases, our intent was to carefully r ecord the or igina l sour ce and t o
clearly indi cate fr om wh ich seconda ry sour ce it was o btained. Users are encouraged to
obtain the primary so urces to ver ify th ese val ues.
We used cer tain f ield guide s cons isten tly th roughout each taxonom ic categ ory to
provide greater comparabilit y of general species character isti cs. The use of a sp eci fic fi eld
guide does not cons ti tute endor semen t.
Because our literature search str ategy may n ot have include d all jour nals of int erest
and did not cons isten tly co ver other sources o f informat ion ( e.g., book s, th eses,
disser tat ions , state w ildlif e repor ts, conf erence pr oceedings ), we would a ppr eciate any
assi stan ce th at users might prov ide in ide nt ifying a ddit iona l sour ces o f informat ion t hat
1-31

would he lp t o fill d ata gaps or to improve th e informat ion in t he Handbook . In partic ular,
Ph.D. disser tat ions and master 's th eses o ften contain relevant but unpublis hed
inf ormation.
1.7. REFERENCES
Peters, R.H. (1983) The ecologic al impli cations of body size . Cambridge , Engla nd: Cambridge Univ ersi ty Press.
U.S. Environmental Protect ion A gency. (1989a) Risk assessmen t guida nce for Supe rfund: volume II--environmental evaluation m anual, int erim fina l. Washingt on, DC: Offi ce of Solid W aste, Offi ce of Emer gency an d Remedial Respons e; EPA repor t no. EPA/540/1-89/001A.
U.S. Environmental Protect ion A gency. (1989b) Ecologic al assessmen t of hazardous waste si tes: a field and laboratory reference. Corvallis , OR: Environm ental Research Laboratory; EPA report no. EPA/600/3-89/013.
U.S. Environmental Protect ion A gency. (1989c) Exp osure facto rs handbook . Washingt on, DC: Offi ce of Health and Environmental Assessmen t; EPA report no. EPA/600/8-89/043.
U.S. Environmental Protect ion A gency. (1992) Evaluation of ter restr ial indi cato rs fo r use in ecologic al assessmen ts at h azardous waste si tes. Washingt on, DC: Offi ce of Research and Development; EPA report no. EPA/600/R-92/183.
1-32

2. EXPOSURE FACTORS AND DESCRIPTIONS
OF SELECTED SPECIES
Chapter 2 includes exp osure profi les fo r the selected species in three subsect ions :
birds (Sect ion 2 .1), mammal s (Sect ion 2 .2), and r ept iles and amphibia ns (Sect ion 2 .3).
Each species profi le follow s the same fo rmat, beginning w ith an int roduc tion t o the
taxonom ic group t o whic h the species be longs and a quali tati ve descr iption of r elevant
aspects o f th e species' natural histo ry. Next, a list o f si mi lar species is provided to help
ident ify species that might sh are cer tain e xpos ure character isti cs, although t hey may h ave
differ ent geographic r anges, diets, and habitat p references. Each species profi le then
presents a ser ies of tab les presenting v alues for nor malizing a nd c ontact r ate facto rs,
dietary co mposi tion, popula tion dy namics, and seaso nal acti vi ty p atter ns th at represent
the range of val ues th at we i denti fied in our literature review. Table format i s descr ibed in
Sect ion 1 .5. Data on soil a nd s edim ent ing estion a re lim ited; we pr esent th ese data in a
separate sect ion ( 4.1.3) for easy co mparison among s pecies. Fina lly , each profi le include s
the references ci ted in the species profi le and in the corresponding A ppendix table s.
2.1. BIRDS
Table 2-1 lists th e bird species descr ibed in th is sect ion. For range map s, refer to
the general references i denti fied in indiv idua l species pr of iles. The remainde r of this
sect ion is organized by sp ecies in t he order presented in Table 2-1. The avai labilit y of
publis hed inf ormation v aries sub stan tially among s pecies, as is r eflected in the profi les.
Some species inc lude two or more subs pecies; these are indi cated in the profi les wh en
reported by th e investi gators. For many st udie s, th e subs pecies, al though not ident if ied,
can be inf erred f rom th e study loc ation a nd ge ogr aphic r ange of th e subs pecies. Average
lengths of birds are reported from mu seum stu dy ski ns measu red from bill t ip t o tail t ip.
Body weight is r epor ted as fresh wet we ight wi th plum age, unl ess o therwise noted.
2-1

Table 2-1. Birds Include d in t he Handbook
Order Family Commo n name Scienti fic name Secti on
Ciconif ormes Ardeidae great blue heron Ardea herodi as 2.1.1
Anser iformes Anatida e Canada goos e Branta can adensis 2.1.2
malla rd Anas platyr hynchos 2.1.3 lesser scau p Ayth ya affi nis 2.1.4
Falconif ormes Accipit ridae osprey Pandion haliaetus 2.1.5
red-taile d hawk Buteo jamaicensis 2.1.6 bald eagle Haliaeetus leucocephalus 2.1.7
Falconida e Amer ican kestr el Falco sparver ius 2.1.8
Gallif ormes Phasianida e nor thern bobw hit e Colin us virg inianus 2.1.9
Charadr iif ormes Scolopa cida e Amer ican woodc ock Scolopax mi nor 2.1.10
spotted sandpipe r Acti ti s macu laria 2.1.11
Laridae herring gull Larus argentatu s 2.1.12
Coraciif ormes Alcedinida e belted kingf isher Cery le alcyo n 2.1.13
Passer iformes Troglody tida e marsh wren Cis tothorus palustris 2.1.14 Muscicapida e Amer ican robin Turdus migratorius 2.1.15
2-2

2.1.1. Great Blue Heron (herons)
Order Ciconiif ormes, Family Ard eidae. Herons, egrets, and bitter ns are medium to large wading bir ds wi th long n ecks an d spear-like bills . Nearly all s pecies feed primar ily on aquatic animal life (e.g., fish, frogs, crayfi sh, insects) an d are commo n along t he margins of mo st fr eshwater and sal twater bodie s and wetl ands (Kushlan, 1978). Their long le gs, necks, and bills ar e adapted for wading in s hallow water and stabbing pr ey. Most species build t heir nests i n trees near their foraging ha bi tat, and many nest c olonia lly . Members of this group r ange in s ize fr om th e least b itter n (28 to 36 cm bill t ip t o tail t ip) to the great blue heron (106 to 132 cm tal l). The sexes ar e simi lar in size and appearance.
Selected species
The great blue heron (Ardea herodi as) is th e largest memb er of th e group in Nor th Amer ica and feeds primar ily on aquatic animals. It is wi dely distr ibuted in both sal twater and freshwater environments. There are four subspecies in the United States an d Canada: A. h. wardi (Kansas an d Oklahoma acr oss th e Missi ss ippi Riv er to Flor ida), A. h. herodi as (remainde r of th e Nor th and Central Amer ican range), A. h. fanni ni (Paci fic co ast o f North Amer ica fr om Alaska to Washingt on) , and A. h. occi dentalis (extr eme so uth of Flor ida) (Bancroft, 1969, ci ted in Hancock an d Kushlan, 1984). A. h. occi dentalis (the great wh ite heron) is an all w hit e color morph t hat was fo rmer ly co nsidered a separate sp ecies (Nationa l Geogr aphic So ciety, 1987).
Body size. Males aver age slight ly h eavier in weight th an femal es (Hartman , 1961; Palmer , 1962). Northern continental herons are somewh at smal ler than those found in t he south (Palmer , 1962). Quinne y (1982) determined a relations hip b etween age and body weight fo r nestling gr eat blue herons (r = 0.996, N = 16 nest lings , and 274 measu rements):
BW = 55.6 × A - 47.4
where BW equals body weight in gr ams an d A equals age in days.
Habitat . Great blue herons inha bit a var iety o f fr eshwater and mar ine areas, inc luding f reshwater lakes an d rivers, bracki sh marshes, lagoons , mangr oves, and coastal wetl ands, particularly wh ere smal l fish are plenti fu l in shallow ar eas (Spendelow and Patto n, 1988; Short and Coope r, 1985). They are often seen on tidal flats an d sandba rs and occas iona lly fo rage in wet mead ows, pastu res, and other ter restr ial habitats (Pal mer , 1962). Great blue herons tend t o nest i n dense colonie s, or heronr ies. The location of th e heronr y is generally clos e to foraging gr ounds , and t all t rees are prefer red over shorter trees or bushes fo r nest si tes (B ent, 1926; Palmer , 1962; Gibbs et al., 1987). They also may nest o n the ground, on r ock le dges, or on sea cl iffs (Pal mer , 1962).
Food ha bi ts . Fish are the prefer red prey, but great blues also eat amphibia ns, repti les, crustacean s, insects, birds, and mammal s (Alexander, 1977; Bent, 1926; Hoffman , 1978; Kirkpatr ick, 1940; Peifer , 1979). When fishing, they mainly use two foraging technique s: standing s till a nd w aiting f or fi sh to swi m within s tr ik ing di stan ce or
2-3 Great Blue Heron

slow wading t o catch more sedentary prey (su ch as f lounde r and s culpin) (B ent, 1926; Willa rd, 1977). To fish, they requir e shallow water s (up to 0.5 m) wi th a fi rm su bstr ate (Shor t and Coope r, 1985). Fish up to about 20 cm in le ngt h were dom inant in t he die t of herons fo raging in s out hwester n Lake Er ie (Hoffman , 1978), and 95 percent of fi sh consumed by great blue s in a W iscons in popula tion w ere less th an 25 cm i n length (Kirkpatr ick, 1940). Great blues so meti mes fo rage in wet mead ows an d pastu res in pursuit of lizards, smal l mammal s, and large insects (Pal mer , 1962; Peifer , 1979). In northern areas, smal l mammal s su ch as mead ow vo les may b e an important part of th e diet ear ly in the breeding seaso n, poss ibly b ecause so me aquatic fo raging a reas may st ill be partia lly frozen when the herons ar rive (Colla zo, 1985). Cons umpt ion of larger prey (fish, frogs , rodents) i s often follow ed by dr ink s of water (Hedeen, 1967); ter restr ial prey su ch as vo les are usually dunked in water before they are swallowed (Peifer , 1979). Adult herons tend t o deliver the same typ e and size of food t o their nestlings th at they cons ume themsel ves, but they deliver it wel l digested for young nestlings and l ess wel l digested as th e nest lings grow (Kushlan, 1978). Adults ten d to feed solit arily , although t hey may feed in single or mixed species fl ocks wh ere there are large concentrations of prey (Bayer , 1978; Krebs, 1974; Kushlan, 1978; Willa rd, 1977); fledglings ar e frequent ly seen foraging t ogether (Dow d and Flake, 1985). Kushlan (1978) develope d a regression e quation r elating t he amount of food ingested per day to body weight fo r wading bir ds (N = seven species):
log( FI) = 0.966 log( BW) - 0.640
where FI equals food ing estion in gr ams p er day and BW equals body weight in gr ams.
Mol t. Adult s unde rgo a co mpl ete mo lt in the late su mmer and fal l and a partial molt of th e contour feath ers in the late wi nter and early sp ring ( Bent, 1926). Young he rons attai n full a dult plum age in t he summer /fall m olt at the end of th eir second year (Bent, 1926).
Migration. In the northern part of i ts r ange, most g reat blues are migratory, some moving t o the sout hern Atlant ic and G ulf States to overwinter wi th the resident popula tions of herons (Bent, 1926; Palmer , 1962), others co ntinuing on t o Cuba and Ce ntral and Sout h Amer ica (Hancock an d Kushlan, 1984). Most mi grating he rons l eave th eir breeding grounds by Octo ber or Novemb er and return between February and April (Bent, 1926).
Breeding acti vi ties and social organization. The male great blue heron s elects th e si te for th e breeding t erritory, and n ests g enerally co nsist o f a sti ck p latfo rm over 1 m in diameter (Palmer , 1962). Great blues often use a nest fo r more than 1 year , expanding it wi th each use (Palmer , 1962). Mean clutch sizes range from th ree to five (see tab le); in general, clutch size tends to increase wi th lati tude (Pratt, 1972). Only one brood is r aised per year ; however , if a cl utch is destr oyed, great blues may l ay a replacemen t cl utch , usually wi th fewer eggs th an the init ial c lut ch (Palmer , 1962; Pratt an d Winkler, 1985). Both parents i ncubate and feed the young (Palmer , 1962; Hancock an d Kushlan, 1984). During the breeding seaso n, great blue s are monoga mous and c olonia l, wi th from a few to hundr eds of pairs nesting in t he same ar ea or heronry (Gibbs et al., 1987). Colonie s may
2-4 Great Blue Heron

�
inc lude other sp ecies, su ch as gr eat egrets o r double-cr ested cormorants (Pr att an d Winkler, 1985; Mock et al ., 1987).
Home range and r esour ces. Breeding c olonie s are generally clos e to foraging grounds (B ent, 1926; Palmer , 1962; Gibbs et al., 1987). Mathisen and Richards (1978) found t he distan ce between heronries and possible feeding a reas in Minne sota lakes to range from 0 to 4.2 km, averaging 1 .8 km. Anot her study found t hat most he ronr ies along the Nor th Carolina co ast wer e located near inlets wi th large concentrations of fi sh, an average of 7 to 8 km away (Par nell and Soots, 1978, ci ted in Short and Coope r, 1985). Fifteen to 20 km i s th e far thest g reat blue herons r egula rly travel between foraging a reas and c olonie s (Gibbs et al., 1987; Gibbs , 1991; Peifer , 1979). In the northern portion of th eir range, great blue herons o ften build n ests i n tal l trees over dry land, whereas in the southern part of th eir range, they usually nest i n swamp trees, including m angr oves (Palmer , 1962). Each breeding pa ir defends a small t erritory around t he nest, the size of which depends on local habitat an d the birds' stag e of reproduc tion ( Hancock and Kus hlan, 1984). Herons in some areas also defend feeding t erritories (Peifer , 1979). In other areas, great blue s appear to be oppor tunis tic foragers, lacking s tr ic t fide lit y to partic ular feeding si tes (Dowd and Flake, 1985). A stu dy in North Dakota found t hat herons o ften returned to the same g eneral areas, but differ ent indiv idua ls o ften used the same ar eas at d iffer ent times (Dowd and Flake, 1985).
Popul ation de nsity . Because great blues nest c olonia lly , loc al popula tion de nsity (i.e., colony density, colony size , and num ber of coloni es) var ies wi th the avai labilit y of suit able nesting ha bitat as wel l as fo raging ha bi tat. On islands in coastal Maine, Gibbs an d others (1987) found a signif icant correlation b etween colony size and t he area of tida l and inter tidal wetl ands wi thin 2 0 km of th e colonie s, whic h was th e longest d istan ce herons in the study colonie s traveled on foraging t rips . In wester n Oregon, the size of heronr ies was found t o range fr om 32 to 161 acti ve nests; the area enclosed by periphe ral nest tr ees wi thin t he colonie s ranged f rom 0.08 to 1.21 ha (Wersch kul e t al., 1977).
Popul ation dy namics. Most nestling los s is a result of star vat ion, although s ome losses to predation do oc cur (Colla zo, 1981; Hancock an d Kushlan, 1984). In a stu dy of 243 nests i n a coastal California colony , 65 percent of th e chi cks fl edged, 20 percent star ved, 7 percent wer e taken by predators, and 7 percent wer e lost to other causes (Pr att an d Winkler, 1985). Esti mates o f th e number of young f ledged each year by breeding pa irs range from 0.85 to 3.1 (Pratt, 1970; Pratt, 1972; McAloney, 1973; Pratt an d Winkler, 1985; Quinne y, 1982). Based on banding s tudie s, about two -thir ds of th e fledglings do not surviv e more than 1 year , although t hey may su rvive better in protected wildlif e refuge s (Bayer , 1981a). Values fo r later years indi cate th at about one-th ird to one-fi fth of th e 2-year -old a nd olde r bir ds ar e los t each year (Bayer , 1981a; Henny, 1972; Owen, 1959).
Simi lar species (fr om general references)
� The great egret (Casmer odius albus) is almost th e same si ze (96 cm l ength) as th e great blue heron and is found ov er a lim ited range in t he breeding seaso n, including a reas in the central and easter n United States an d the east and west co asts. It wi nters in coastal areas of th e United States an d in
2-5 Great Blue Heron

�
�
�
�
�
�
�
�
Mexico and far ther south. The great egret's habitat p references ar e simi lar to those of th e great blue heron.
� The snowy eg ret (Egretta th ula), one of th e medium-si zed herons (51 to 69 cm), shuffl es i ts feet to sti r up benthic aquatic prey. It is found m ostl y in freshwater and sal twater marshes but also someti mes f ollow s cattl e and other livesto ck as d oes th e cattl e egret. It breeds in parts o f th e wester n, southeaster n, and east co asts o f th e United States an d winters along bot h coasts o f th e southern United States an d far ther south.
� The cattl e egret (Bubul cus ibis) is seen in agricultural pastu res and fields, where it follow s li vesto ck to pick u p insects d istu rbed by grazing. An Old World species, it was i ntroduced into South Amer ica and reached Flor ida in the 1950's. It reached California by th e 1960's and has been continuing t o expand its r ange.
� The green-backed heron (Butorides str iatus), one of th e smalle r herons (41 to 56 cm), breeds over most o f th e United States excep t fo r the northwest and southern midwest. It has a wi nter range simi lar to that of th e snowy egret and seems to prefer water bodie s wi th woodla nd c over .
� The tr icolor ed heron (Egretta tr icolor) (former ly know n as the Louis iana heron) is co mmo n in sal t mar shes and mangrove swamp s of th e east an d gulf co asts, but i t is rare inla nd.
� The lit tle blue heron (Egretta caer ulea) is co mmo n in freshwater ponds , lakes, and marshes and coastal sal twater wetl ands of th e Gulf Coast States. Juvenile s are easily conf used wi th juv enile snow y egrets. This sp ecies hunt s by wal king s low ly in s hallow water s, and its d iet typ ical ly includes fi sh, amphibia ns, crayfi sh, and insects.
� The black-cr owned night heron (Nycti corax nycti corax), character ized by a heavy body , shor t thic k neck, and short legs (64 cm), is a co mmo n heron of freshwater swamp s and tidal marshes, roost ing by day in t rees. It typ ical ly feeds by night , predom inant ly on aquatic species, fi sh, amphibia ns, and insects. This heron is extr emely wi despread, occu rring in Nor th and Sout h Amer ica, Eurasia, and Afr ica. It breeds over much of th e United States an d parts of cen tral Canada and w int ers along bot h coasts o f th e United States and far ther south.
� The yel low-cr owned night heron (Nyctan assa vi olacea) (61 cm) i s si mi lar to the black-cr owned but is mo re restr icted in its r ange to the southeaster n United States. It roosts i n trees in wet w oods , swamp s, and low co astal shrubs .
� The Amer ican bitter n (Botaurus lentig inosus), anot her of th e medium -sized herons (58 to 70 cm), is a relatively co mmo n but elusive inha bit ant of freshwater and bracki sh marshes and reedy lakes. It is a solitary feed er,
2-6 Great Blue Heron

�
cons uming f ish, crayfi sh, repti les, amphibia ns, insects, and even smal l mammal s. Its b reeding r ange inc lude s most of Canada and t he Unit ed States, although m uch of th e sout hern Unit ed States i s inha bit ed only dur ing the winter .
� The least b itter n (Ixobrych us exilis ), the smal lest o f th e North Amer ican herons (33 cm), also is an elusive inha bit ant of r eedy areas. Its b reeding range is restr icted largely to the easter n half of th e United States.
General references
Hancock an d Kushlan (1984); Robbins et al. (1983); Nationa l Geogr aphic So ciety (1987); Palmer (1962); Short and Coope r (1985).
2-7 Great Blue Heron
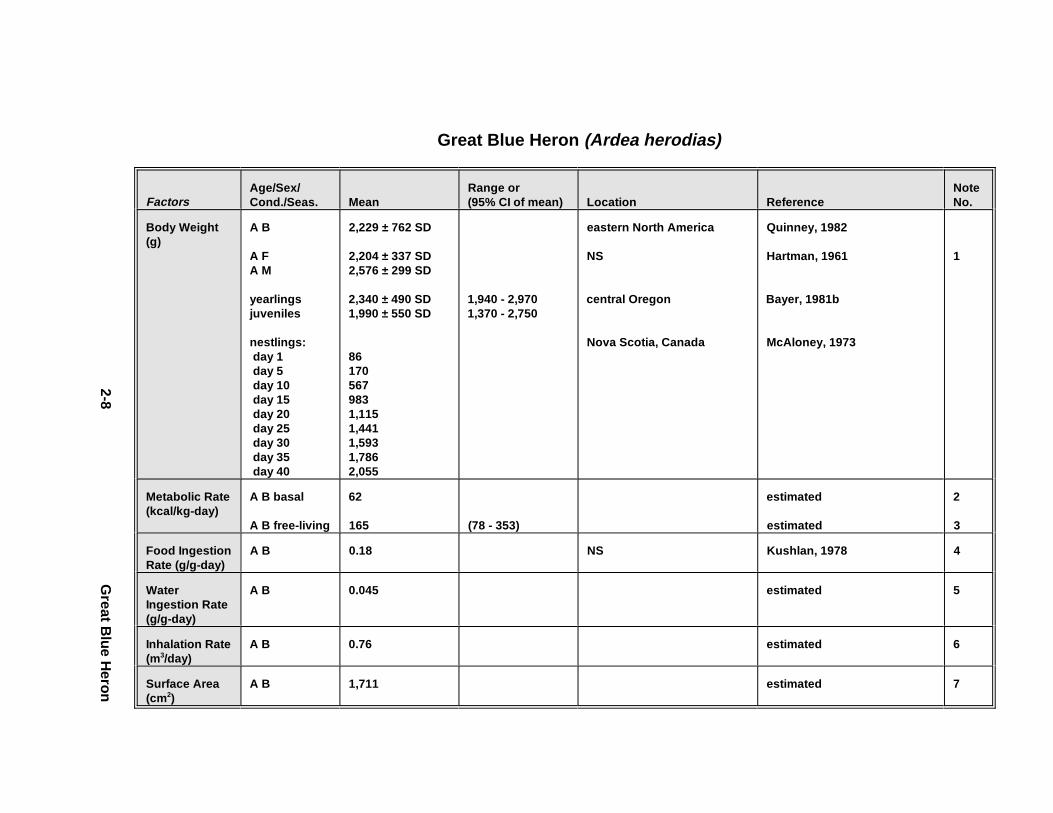
Great Blue Heron (Ardea herodias )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A B 2,229 ± 762 SD eastern Nor th Am erica Quinney , 1982 (g)
A F 2,204 ± 337 SD NS Hartman, 1961 1 A M 2,576 ± 299 SD
yearlings 2,340 ± 490 SD 1,940 - 2,970 cent ral Oregon Bayer, 1981b juveni les 1,990 ± 550 SD 1,370 - 2,750
nest lings: Nova Scot ia, Canada McAloney , 1973 day 1 86 day 5 170 day 10 567 day 15 983 day 20 1,115 day 25 1,441 day 30 1,593 day 35 1,786 day 40 2,055
Metabol ic Rate A B basal 62 est imated 2 (kcal /kg-day)
A B f ree-living 165 (78 - 353) est imated 3
Food Ingest ion A B 0.18 NS Kushl an, 1978 4 Rate (g/g-day)
Water A B 0.045 est imated 5 Ingest ion Rat e (g/g-day)
Inhal ation Rat e A B 0.76 est imated 6 (m /day)3
Surface Ar ea A B 1,711 est imated 7 (cm )2
2-8 G
reat Blue
Hero
n

2-9 G
reat Blue
Hero
n
Dietary Compositio n Spr ing
trout non- trout fish crust aceans/ amphi bian s
trout non- trout fish crust aceans amphi bians bi rds and m ammals
At lant ic si lverside mummi chog American eel Gaspereaux pol lack yello w perch
staghor n scul pin small medium large
starry flounder small medium large
other small medium
Great Blue Heron (Ardea herodias )
Summer Fall Winter (measur e) Locat ion/Habi tat
59 lower Michi gan/lake 39
2 (% wet weight ; stomach cont ents)
89 lower Michi gan/river 5 1 (% wet weight ; stomach 4 cont ents) 1
3.6 Nova Scot ia/Boot Island 2.4
52.6 (% wet weight ; items 29.9 regur gi tated by nest lings)
8.9 2.6
Vancouver , BC/coast al 27.8 island
7.6 2.2 (% of fish obser ved caught ;
small = less t han 1/3 beak 15.0 lengt h; medium = about 1/2
8.1 beak lengt h; large = longer 5.2 than beak; other includes
shi ner sea per ch and 30.6 penpoi nt gunnel s)
3.5
Reference No. Note
Alexander , 1977
Alexander , 1977
Quinney , 1982
Krebs, 1974

Great Blue Heron (Ardea herodias )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range ion/Habi tat Reference No.
Size Feeding A B fall 0.6 ± 0.1 SD ha Oregon/ freshwat er marsh Bayer, 1978 Territory A B wi nter 8.4 ± 5.4 SD ha Oregon/ estuary
Foraging A B sum mer 3.1 km up t o 24.4 km Sout h Dakot a/river & Dowd & Fl ake, 1985 Distance f rom streams Colony A B sum mer 7 to 8 km North Carolina/coast al Parnell & Soot s, 1978 8
Popul ation sum mer North Dakot a/rivers & Dowd & Fl ake, 1985 Densi ty along st ream 2.3 bi rds/km streams
along r iver 3.6 bi rds/km
sum mer 149 ± 53 SD Maine/coast al islands Gibbs et al., 1987
sum mer 461 nest s/ha 447 - 475 Oregon/ coast al island Werschkul et al., 1977 nest s/ha
Clutch Si ze 3.16 ± 0.04 SE 1 - 5 California/coast al cany on Pratt & Winkl er, 1985
4.17 ± 0.85 SD 3 - 6 Nova Scot ia/island McAloney , 1973
4.37 3 - 6 Pennsylvania/NS Mille r, 1943 9
Clutches/ Year 1 lvani a; Oregon/ NS Mille r, 1943; Engl ish, 1978 10
Days 27.1 25 - 30 Nova Scot ia/island McAloney , 1973 Incubat ion
28 Uni ted States/NS Bent , 1926
Age at Fledgi ng 45 Nova Scot ia/island McAloney , 1973 11 (days) 60 NS/NS Hancock & Kushl an, 1984
49 to 56 Nova Scot ia/island Quinney , 1982
Number Fledge 1.7 cent ral California/coast al Pratt, 1972 per Pair 1.96 nor thwest Oregon/ river Engl ish, 1978
2.8 Nova Scot ia/island Quinney , 1982
Locat
Pennsy
2-10 G
reat Blue
Hero
n

123456
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range ion/Habi tat Reference No.
Number 2.19 ± 0.25 SD 2 - 3 cent ral California/coast al Pratt & Winkl er, 1985 Fledge per 2.43 nor thwest Oregon/ river Engl ish, 1978 Successf ul 3.09 Nova Scot ia/island McAloney , 1973 Nest
Age at Sexual B 2 years NS Bent, 1926 Maturity
Annual dur ing 1st yr 64 Uni ted States and Henny , 1972 Mortality Rates dur ing 2nd y r 36 Canada/NS (percent ) dur ing 3r d yr 22
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Matin g/Laying Nov. to Dec. April Flo rid a Howell, 1932 9 mid-February mid-March June cent ral California Pratt & Winkl er, 1985 mid-March nor thwest Oregon Engl ish, 1978 late March early April Pennsylvania Mille r, 1943 9 mid-Apr il early May late May Nova Scot ia McAloney , 1973
Hatching early May nor thwest Oregon Engl ish, 1978 mid-Apr il Idaho Col lazo, 1981 mid-May mid-July Ohio Hoffman & Cur now , 1979
Migration (fall) mid-Sept. late October nor thern US Palmer, 1962
(spring arriv al) mid -February mid-March western Oregon Werschkul et al., 1977 mid-March Wisconsi n; Minnesot a Bent , 1926 late March Nova Scot ia Bent , 1926
Locat
Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Quinney (1982).Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Quinney (1982).Est imated f rom Kushl an's (1978) allometric equat ion f or wadi ng bi rds, assum ing a body weight of 2,230 g.Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Quinney (1982).Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Quinney (1982).
2-11 G
reat Blue
Hero
n
As ci ted in Dunni ng, 1984.
Great Blue Heron (Ardea herodias )

7 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Quinney (1982).
8 Cited in Shor t and Cooper (1985). 9 Cited in Palmer (1962).
10 May replace cl utch i f eggs ar e lost , but onl y rear one br ood ( Henny , 1972). 11 Young f ed around col ony for 10 days after leaving nest at 45 days of age.
2-12 G
reat Blue
Hero
n
Great Blue Heron (Ardea herodias )

References (including A ppendix)
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Altman , P. L.; Dittmer , D. S., eds. (1968) Biology d ata book . 2nd ed., 3v. Bethesda, MD: Federation of A mer ican Societies for Exp erimental Biology .
Baird, S. F.; Brewer , T. M.; Ridg eway, R. (1884) Water birds of North Amer ica. Bosto n, MA: Lit tle, Brown & Co.
Bancroft, G. (1969) (as ci ted in Hancock an d Kushlan, 1984). Auk 86: 141-142.
Bayer , R. D. (1978) Aspects o f an Oregon estu arine great blue heron popula tion. In: Sprunt , A.; Ogden, J.; Winc kler, S., eds. Wading bir ds. Natl . Audubon S oc. Res. Rep. 7; pp. 213-217.
Bayer , R. D. (1981a) Regiona l variation of great blue heron Ardea herodi as longe vi ty. J. Field Ornit hol. 52: 210-213.
Bayer , R. D. (1981b) Weights o f great blue herons ( Ardea herodi as) at the Yaquina Estu ary, Oregon. Mur relet 62: 18-19.
Benedict, F. G.; Fox, E. L. (1927) (ci ted in Altman and Dittmer , 1968). Proc. Am. Phil. Soc. 66: 411.
Bent, A. C. (1926) Life histo ries of North Amer ican marsh birds. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 135.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Colla zo, J. A. (1981) Some asp ects o f th e breeding e cology of th e great blue heron at Heyburn State Par k. Northwest Sci . 55: 293-297.
Colla zo, J. A. (1985) Food ha bit s of n estling gr eat blue herons ( Ardea herodi as) at Heyburn State Par k, Idaho. Northwest Sci . 59: 144-146.
Cottam, C. A.; Uhler, F. M. (1945) Birds in relation t o fishes. U.S. Fish Wildl. Serv. Leaflet 272.
Cottam, C. A.; Willi ams, J. (1939) (as ci ted in Palmer , 1962). Wilson Bull. 51: 150-155.
Dowd, E.; Flake, L. D. (1985) Foraging ha bi tats an d movemen ts o f nest ing gr eat blue herons in a prairie river ecosystem, South Dakota. J. Field Ornithol. 56: 379-387.
2-13 Great Blue Heron

Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Englis h, S. M. (1978) Distr ibut ion a nd ecology of great blue heron c olonie s on t he Willamette R iver , Oregon. In: Sprunt , A.; Ogden, J.; Winc kler, S., eds. Wading bir ds. Natl . Audubon S oc. Res. Rep. 7; pp. 235-244.
Forbes, L. S.; Simpson, K.; Kelsal l, J. P., et al . (1985) Reproducti ve su ccess o f great blue herons in Br it ish Colum bia. Can. J. Zool. 63: 1110-1113.
Gibbs , J. P. (1991) Spatial relations hips b etween nest ing c olonie s and f oraging a reas of great blue herons . Auk 108: 764-770.
Gibbs , J. P.; Woodw ard, S.; Hunt er, M. L., et al. (1987) Determinants o f great blue heron colony dis tr ibut ion in c oastal Maine. Auk 104: 38-47.
Hancock, J.; Kushlan, J. (1984) The herons handbook . New York, NY: Harper & Row .
Hartman , F. A. (1961) Locomotor mech anisms i n birds. Washingt on, DC: Smithsonia n Mis c. Coll. 143.
Hedeen, S. (1967) Feeding be havior of th e great blue heron in I tasca State Par k, Minne sota. Loon 3 9: 116-120.
Henny, C. J. (1972) An analysi s of th e popula tion dy namics of sel ected avian species wi th special reference to changes during t he mode rn pesti cide era. Washingt on, DC: Bur . Spor t. Fish. Wildl. ; Wildl. Res. Rep. 1.
Henny, C. J.; Bethers, M. R. (1971) Ecology of th e great blue heron w ith special reference to wester n Oregon. Can. Field-Nat. 85: 205-209.
Hoffman , R. D. (1978) The diets o f herons and egrets i n southwester n Lake Er ie. In: Sprunt, A.; Ogden, J.; Winc kler, S., eds. Wading bir ds. Natl . Audubon S oc. Res. Rep. 7: 365-369.
Hoffman , R. D.; Curnow, R. D. (1979) Mercury in herons, egrets, and their foods . J. Wildl. Manage 43: 85-93.
Howel l, A. H. (1932) Flor ida bir d lif e. Flor ida Dept . Game and Freshwater Fish and Bur. Biol. Survey, USDA.
Kelsal l, J. P.; Simpson, K. (1979) A three year stu dy of th e great blue heron in southwester n British Colum bia. Proc. Col. Waterbird Group 3 : 69-74.
Kirkpatr ick, C. M. (1940) Some foods of young gr eat blue herons . Am. Midl. Nat. 24: 594-601.
2-14 Great Blue Heron

Krebs, J. R. (1974) Colonia l nesting a nd s ocial feeding a s str ategie s for exploit ing f ood resources i n the great blue heron (Ardea herodi as). Behaviour 51: 93-134.
Kushlan, J. A. (1978) Feeding e cology of wading bir ds. In: Sprunt , A.; Ogden, J.; Winc kler, S., eds. Wading bir ds. Natl . Audubon S oc. Res. Rep. 7; pp. 249-296.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Mathisen, J.; Richards, A. (1978) Statu s of great blue herons on t he Chipp ewa Nationa l Forest. Loon 5 0: 104-106.
McAloney, K. (1973) The breeding biology of th e great blue heron on T obacco Island, Nova Scotia. Can. Field-Nat. 87: 137-140.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Mille r, R. F. (1943) The great blue herons : the breeding bir ds of th e Phila delphia r egion (Part II). Cass inia 33: 1-23.
Mitch ell, C. A. (1981) Reproducti ve su ccess o f great blue herons at Nu eces B ay, Corpus Christi , Texas. Bull. Texas Or nithol. Soc. 14: 18-21.
Mock, D. W.; Lamey, T. C.; Willi ams, C. F.; et al . (1987) Flexibilit y in t he development of heron s ibling a ggr ession: an int erspeci fic test o f th e prey-si ze hypothesis. Anim. Behav. 35: 1386-1393.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Owen, D. F. (1959) Mortal ity o f th e great blue heron as show n by banding r ecover ies. Auk 76: 464-470.
Page, P. J. (1970) Appendix . San Joaquin Riv er rook ery study 1970. Sacramento, CA: Calif. Dept. Fish and Game; statewi de heron rookery su rvey p rogress r eport.
Palmer , R. S. (1949) Marine birds. Bull. Mus. Comp. Zool. Harvard 102.
Palmer , R. S. (1962) Handbook of Nor th Amer ican bir ds: v. 1. New Haven, CT: Yale Universi ty Pr ess.
2-15 Great Blue Heron

Parnell, J. F.; Soots, R. F. (1978) The use of dredge islands by wading bir ds. Wading bir ds. Nat. Audubon S oc. Res. Rep. 7: 105-111.
Peifer , R. W. (1979) Great blue herons fo raging f or smal l mammal s. Wilson Bull. 91: 630-631.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Powel l, G. V.; Powel l, A. H. (1986) Reproduc tion by great wh ite herons Ardea herodi as in Flor ida Bay as an indi cato r of habitat q uality. Biol. Cons erv. 36: 101-113.
Pratt, H. M. (1970) Breeding biology of great blue herons and c ommon e grets i n central Calif ornia . Condor 72: 407-416.
Pratt, H. M. (1972) Nest ing s uccess o f co mmo n egrets an d great blue herons in t he San Francisco Bay region. Condor 74: 447-453.
Pratt, H. M.; Winkler, D. W. (1985) Clut ch size ti ming of laying a nd r eproduc tive success i n a colony of great blue herons ( Ardea herodi as) and great egrets (Casmer odius albus ). Auk 102: 49-63.
Quinne y, T. E. (1982) Growth , diet, and mortal ity o f nest ling gr eat blue herons . Wils on Bull. 94: 571-577.
Robbins , C. S.; Bruun, B.; Zim, H. S. (1983) A guide to field ide nt if ication: bir ds of Nor th Amer ica. New York, NY: Golde n Press.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Shor t, H. L.; Coope r, R. J. (1985) Habitat su itabilit y inde x mode ls : great blue heron. U.S. Fish Wildl. Serv. Biol. Rep. No. 82(10.99); 23 pp.
Spendelow, J. A.; Patto n, S. R. (1988) Nationa l atlas of co astal water bird colonie s: 1976-1982. U.S. Fish Wildl. Serv. Biol. Rep. No. 88(5).
Thompson, D. H. (1978) Feeding a reas of great blue herons and gr eat egrets n est ing w ithin the floodpla in of th e uppe r Mississippi Riv er. Colonia l Waterbirds 2: 202-213.
Vermeer , K. (1969) Great blue heron c olonie s in A lberta. Can. Field-Nat. 83: 237-242.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Wersch kul, D. F.; McMahon, E.; Leitschuh, M.; et al. (1977) Observat ions on t he reproduc tive ecology of th e great blue heron (Ardea herodi as) in wester n Oregon. Murrelet 58: 7-12.
2-16 Great Blue Heron

Willa rd, D. E. (1977) The feeding e cology and be havior of fi ve species of herons in southeaster n New Jer sey. Condor 79: 462-470.
Wood (1951) (ci ted in Palmer , 1962). Univ . Mich. Mus. Misc. Publ. No. 75.
2-17 Great Blue Heron

2.1.2. Canada Goose (geese)
Order Anserif ormes, Family Anatidae. Geese are large herbivorous water fowl that feed on grains, grass sp routs, and some aquatic veg etat ion. Although a dapted for lif e on the water , they fo rage primar ily in open fields. They breed in open forested areas near lake shores and coastal marshes fr om th e arcti c tundr a through temp erate cl imates. These birds mi grate in noisy fl ocks i n the familia r V-formation, stopping in c ult ivated fields, wetl ands, and grassl ands to feed. Geese sh ow a wi de var iation in s ize even wi thin a species; the sexes look alik e.
Selected species
The Canada goos e (Branta can adensis) is th e most wide spread and abunda nt goos e in Nor th Amer ica. It is a popula r game sp ecies and is co mmonly en count ered on c ult ivated fields , golf cour ses, other parklands, and wetl and refuge areas. Depending on s ubs pecies, these geese can range in size from 64 to 114 cm (bill t ip t o tail t ip) , the larger geese breeding in m ore sout herly lo cations th an the smal ler subspecies. The reverse is tr ue in wint er, wi th the larger subs pecies wint ering in t he more nor therly parts of th e range (Palmer , 1962). The number of exi st ing r ecognize d subs pecies var ies, but mo st ornit hologi sts ag ree that th ere are 11: canadensis (Atlantic Canada goose), fu lva (Vancouver Canada goose), hutch insii (Richardson's Canada goose), interio r (int erior Canada goose), leucopareia (Aleutian Canada goose), maxi ma (giant Canada goose), minima (cack ling Ca nada goo se), moffi tti (Great Basin or wester n Canada goose), occi dental is (dusky C anada goose), parv ipes (lesser Canada goose), and taver neri (Taverner's Canada goose) (Bellrose, 1976; Johns on et al., 1979; Palmer , 1962). Several subs pecies us ually mingle dur ing m igr ation a nd in w int ering a reas, but th ey breed in geographical ly disti nct r anges. Six of th e subspecies breed in Alaska ( fulva, leucopareia, minima, occi dental is, parvipes, and taver ni) (Johns on et al., 1979). The leucopareia subs pecies, found in O regon, Washingt on, Calif ornia , and A laska, currently is a Unit ed States fed erally designated threatened species (50 CFR 17.11, 1992). It is only know n to breed on one of th e wester n Aleutian islands off A laska (B yrd and Woolingt on, 1983). See Bellrose (1976) fo r ranges, migration c orridor s, and w int ering a reas of sp eci fic su bspecies and popula tions .
Body size. Canada geese su bspecies var y greatly in size, but mal es are on average larger than femal es (see tab le). Body weight r eaches i ts maxi mum just p rior to or during the spring m igr ation a nd t hen decline s dur ing e gg- laying a nd inc ubation, someti mes b y as much as 20 percent (Mainguy and T homas, 1985; McLandress an d Raveling, 1981). Most o f the weight los t dur ing inc ubation r eflects l oss o f fat, which can provide over 80 percent of the energy r equir ements fo r the incubating femal es (Mainguy and T homas, 1985; Murphy and Boag, 1989). Young a re simila r to parents in s ize by 2 mont hs of ag e (Palmer , 1962).
Habitat . Breeding ha bitat i nclude s tundr a, forest mu skeg in the far north, tal l- and shortgrass p rairie, marshes, ponds , and l akes. Most n est ing s ites ar e close to open water wi th high v isibilit y in a ll dir ections (Palmer , 1962; Steel et al ., 1957). In many areas, Canada geese nest p redominantly on islands in ponds or lakes (Gei s, 1956). Former
2-19 Canada Goose

muskr at houses o ften serve as n est si tes i n marshes (Steel et al ., 1957). Brood- rearing habitats, on the other hand, requir e adequate co ver , and riparian areas are used more frequently th an open water (Eberhardt et al ., 1989a). During t he fall a nd w int er in Maryland, Harvey et al . (1988) found Ca nada geese to spend 57 percent of th eir time in farmlands (mostl y co rn, soybeans, and winter wheat fi elds) and 24 percent in forested areas.
Food ha bi ts . Canada geese are almost excl usively veg etar ian, and feeding acti vi ty is co ncentrated in areas wh ere food is plent iful (e.g., standing c rops , scatter ed whole grain) (Palmer , 1962). They are primar ily grazers, but mu st co nsume grit at so me point to assu re proper dige stion (Palmer , 1962). They prefer cer tain foods , but will c hange th eir die t depending on t he availa bilit y of a food t ype (Coleman and Boag, 1987). For examp le, when water levels are low in the south Yukon (Canada) river delta, Canada geese fo rage on rhizo mes o f Potamoge ton r ichardsoni i even though ot her fo rage is availa ble ; at highe r water levels wh en the Potamo geton is unreachable, the geese will feed on other plants (Coleman and Boag, 1987). During f all, geese often consume green crops (e.g., winter wheat). During w int er, however , they co nsume mo re energy-r ich foods su ch as corn (Harvey et al ., 1988; McLandress an d Raveling, 1981). In late wi nter and early sp ring, green crops th at are high in nit rogen and ot her impor tant nut rients again c ons ti tute an impor tant part of th e diet (McLandress an d Raveling, 1981). Canada geese often feed preferentially on t he blade tips of man y pla nts, whic h are highe r in nit rogen than other parts of th e plant (Buchsbaum et al ., 1981). In Minne sota, Canada geese begin c ons uming gr een grasses as soon a s they are expos ed by th e melting s now (M cLandr ess an d Raveling, 1981). In Maryland, on the other hand, Harvey et al . (1988) found t hat Canada geese did not begin cons uming gr een crops before migr ation t o the breeding gr ounds , indi cating t hat this popula tion may r ely on green forage avai lable at sta ging a reas to obtain the protein and lipids r equir ed for reproduc tion. In the spring in Fa lmout h Harbor , Massach usetts, Canada geese init ially cons ume predom inant ly th e marsh grasses Spartina spp. and rushes Junc us gerardi, whic h are high in pr otein (Buc hsbaum and V alie la, 1987). As th e summer progresses, however , they feed increasingly on s ubm erged eelgr ass, Zoster a mar ina, whic h pr ov ides more carbohy drates (B uchsbaum and Valiela, 1987).
Mol t. Nonbr eeders and year lings migr ate to a separate mo lting gr ound s oon after arrival at the breeding gr ounds , while breeding bir ds molt on t he brood- rearing gr ounds (Bellrose, 1976). Molt ing oc curs earlier in nonbr eeders, at least a mo nth earlier in the larger subspecies (Palmer , 1962). Molt ing pa rents do not r egain f light feath ers unti l just pr ior to the time when their young f irst attai n flight (Palmer , 1962). The flight less p eriod of B. c . interio r is esti mated to be 32 days. For B. c. maxi ma and B. c. moffi tti , the flight less period l asts fr om 39 to 40 days (B alham, 1954; Hanson, 1965, as ci ted in Palmer , 1962).
Migration. Migr atory Canada geese leave th eir breeding gr ounds dur ing la te summer and early autumn; they return in the spring a round t he time the fi rst water is opening ( i.e., ice melting) but well be fore snow co ver has dis appeared (Bellr ose, 1976). Spring m igr ation be gins l ater for northerly popula tions , wi th geese th at wi nter in mi ld cl imates d eparting a s early as mid -Janua ry, while thos e wint ering in t he cold est ar eas do not mo ve nor thward unt il t he beginning of March (Bellr ose, 1976). The bulk of th e migr ants typ ical ly ar rive on the summer breeding gr ounds 3 weeks after the fi rst b irds
2-20 Canada Goose

(Bellrose, 1976). Some popula tions h ave become resident year -round, for examp le, B. c. maxi ma in Misso uri (Brakhage, 1965) and in southeast Geo rgia and southwest
Alabama (Combs et al ., 1984). During bot h the spring a nd f all m igr ations , geese ten d to gather in large flocks an d feed for sever al weeks i n "staging" a reas along m ajor water fowl flyways (B ellrose, 1976).
Breeding acti vi ties and social organization. Canada geese ar rive on the breeding grounds in f locks, and soon after , the male becomes ter ritorial and aggressi ve to ward other birds (Palmer , 1962). Lifelong m onog amy f ollow ing t heir fi rst br eeding is th e general rule wi th these geese (Palmer , 1962). Nests ar e built on t he ground in a pos it ion w ith good visibilit y (Palmer , 1962). During inc ubation t he male stands guard, while th e femal e incubates th e eggs, which she normal ly leaves two or three times d aily to feed, bath, drink, and preen (Murphy and Boag, 1989). Both parents acco mpany th e young t hrough t he brood pe riod ( Bellr ose, 1976; Brakhage, 1965). Canada geese return to the breeding grounds as fami ly units, but th e year lings l eave th eir parents s oon after arrival (Bellrose, 1976).
Home range and r esour ces. The foraging hom e range of Canada geese var ies wi th seaso n, lati tude, and breeding c ondit ion. Soon after hatching, goos e fami lies mo ve away from th e nesting s ites to other areas wi th adequate co ver and forage to rear their broods (Byrd and Woolingt on, 1983). Newly hatch ed families may h ave to travel 10 to 20 km fr om the nest si te to reach areas wi th adequate aquatic veg etat ion or pastu re grasses (Gei s, 1956). Although t he fami lies stay p redominantly on land, often in riparian areas, they usually are close to water . Eberhardt et al . (1989a) found gos lings wi thin 5 m of water most of th e time; only 7 pe rcent of sight ings wer e far ther th an 50 m away. During t he spring a nd f all m igr ations and in w int er, Canada geese can be found on ope n water or refuges near grain fields or coastal estu aries (Leopold e t al., 1981).
Popul ation de nsity . Breeding popula tion de nsit ies of Canada geese var y wi dely. Low nesting de nsit ies (i.e., less th an 0.005 per hectar e) are commo n in the Northwest Territories of Canada (Smith and Sutto n, 1953, 1954) and intermediate densi ties (i .e., 0.02 to 0.7 per hectar e) have been reported for Alaska (Cornley et al ., 1985). In some mo re sout herly loc ations (e. g., Calif ornia), colonia l nesting s ituations h ave been reported, wi th as man y as 32 n ests l ocated on half an acre (Naylor, 1953, as ci ted in Palmer , 1962).
Popul ation dy namics. The earliest Canada geese begin br eeding is ar ound 2 to 3 years of age (MacInnes and Dunn, 1988; Brakhage, 1965). In the larger subspecies, only a small pr opor tion of th e bir ds unde r 4 years may attemp t to breed. For examp le, in Manitoba, Moser and Rusch (1989) found t hat only 7 pe rcent of 2-year -old a nd 15 percent of 3-year -old B. c . interio r laid eggs. Canada geese only attemp t to rear one brood pe r year . In the more southerly lati tudes, Canada geese will r enest i f a cl utch is lost p rior to inc ubation ( Brakhage, 1965; Geis, 1956). In general, both clutch size and success at rearing gos lings inc rease wi th the age of th e breeder (Brakhage, 1965). Raveling (1981) found t hat olde r B. c. maxi ma (4 plus years) raised more than twi ce as man y goslings to fledging a s did y ounge r (2 to 3 years) birds. Popula tion a ge str ucture and annua l mortalit y vary wi th hunt ing pr essu re as wel l as natural facto rs.
2-21 Canada Goose

�
�
�
�
Simi lar species (fr om general references)
� The Brant goose (Branta bernic la) is approximatel y th e size of th e smal ler Canada geese su bspecies (length 25 cm). It is primar ily a sea goos e and is rare inla nd. It wint ers along bot h the east an d west co asts o f th e Unit ed States, where it feed s on aquatic plants i n shallow b ays an d estu aries. It breeds in t he high a rcti c.
� The greater white-fr onted goose (Anser albifrons) is l imi ted to cer tain areas west o f th e Missi ss ippi Riv er and averages 71 cm i n length. Its h abits ar e simi lar to those of other geese.
� The snow goose (Chen caerulescen s) breeds in the Arcti c and winters in selected coastal areas acr oss th e United States. However , th is aver age-sized goose (71 cm) i s a mi gratory vi si tor to much of th e central Unit ed States.
� The Ross' goose (Chen rosi i) breeds in t he high a rcti c tundr a and w int ers in some areas of th e southwest United States. This relatively smal l (58 cm) goose is a rare visi tor to the mid-Atlantic States an d is always seen wi th snow geese.
General references
Bellrose (1976); Kortr ight (1955); Nationa l Geogr aphic So ciety (1987); Palmer (1962).
2-22 Canada Goose

Canada Goose ( Branta canadensis )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion (subspeci es) Reference No.
Range or
Body Weight (g) A M late sum . 1,443 ± 32 SE 1,260 - 1,605 Alaska (minima) Ravel ing, 1979 A F late sum . 1,362 ± 54 SE 1,195 - 1,590
A M winter 2,769 ± 30 SE Colorado (parvipes) Grieb, 1970 A F w inter 2,472 ± 23 SE
A M not spec. 3,992 NS (canadensi s) Webst er (unpubl ished) in A F not spec. 3,447 Bel lrose, 1976
A M fall 4,212 ± 35 SE 3,799 - 4,727 Illin ois (in terio r) Ravel ing, 1968 A F fall 3,550 ± 31 SE 3,147 - 3,856
A M late sum . 4,960 Missour i (maxima) Brakhage, 1965 A F late sum . 4,160
M at hatching 108.7 Alberta (moffitti ) LeBlanc, 1987b F at hatching 109.5
B day 10 150 Alaska (minima) Sedinger , 1986 1 B day 20 450 B day 30 755 B day 40 950 B day 47 1,050
B day 0 110 NS (moffitti ) Willia ms (unpubl ished) in B day 9 240 Palmer, 1976 B day 16 440 B day 30 1,400 B day 44 2,400 B day 51 2,600
M at fledgi ng 87% adul t wt Alaska (minima) Sedinger , 1986 F at fledgi ng 89% adul t wt
2-23 C
anad
a Go
ose

Canada Goose ( Branta canadensis )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion (subspeci es) Reference No.
Range or
Body Fat F fall migr. 182 ± 24 SE 117 - 264 Alaska i n winter (minima) Ravel ing, 1979 (g lipid) F winter 57 ± 6 SE 34 - 71 California in sum mer
F spr ing m igr. 172 ± 25 SE 68 - 362 F prelaying 171 (no SE; N=2) 136 - 205 F end i ncub. 33 ± 5 SE 14 - 51 F early molt 108 ± 13 SE 62 - 179
F prelaying 751 ± 45 SE Ontario, Canada (maxima) Thom as et al., 1983 F incubat ing 611 ± 40 SE F late incub. 166 ± 18 SE F molting 485 ± 37 SE
Egg Weight 96 NS (minima) Owen, 1980 2 (g) 127 NS (leucopa) Owen, 1980 2
163 Alberta, Canada (moffitti ) LeBlanc, 1987a
Metabolic Rate free-liv ing: (kcal /kg-day) A M winter 105 - 209 Illin ois in winter (in terio r) Willia ms & Kendeigh, 1982 3
A M spr ing 105 - 203 Ontario, Canada i n A M sum mer 115 - 253 sum mer A M fall 100 - 209
A F spr ing 130 - 220 Willia ms & Kendeigh, 1982 3 A F sum mer 143 - 274 (in terio r)
free-liv ing: A M 185 (87 - 391) est imated 4a A F 187 (88 - 397) (minima)
A M 141 (65 - 304) est imated 4b A F 147 (69 - 316) (in terio r)
A M 135 (63 - 292) est imated 4c A F 142 (66 - 305) (maxima)
2-24 C
anad
a Go
ose
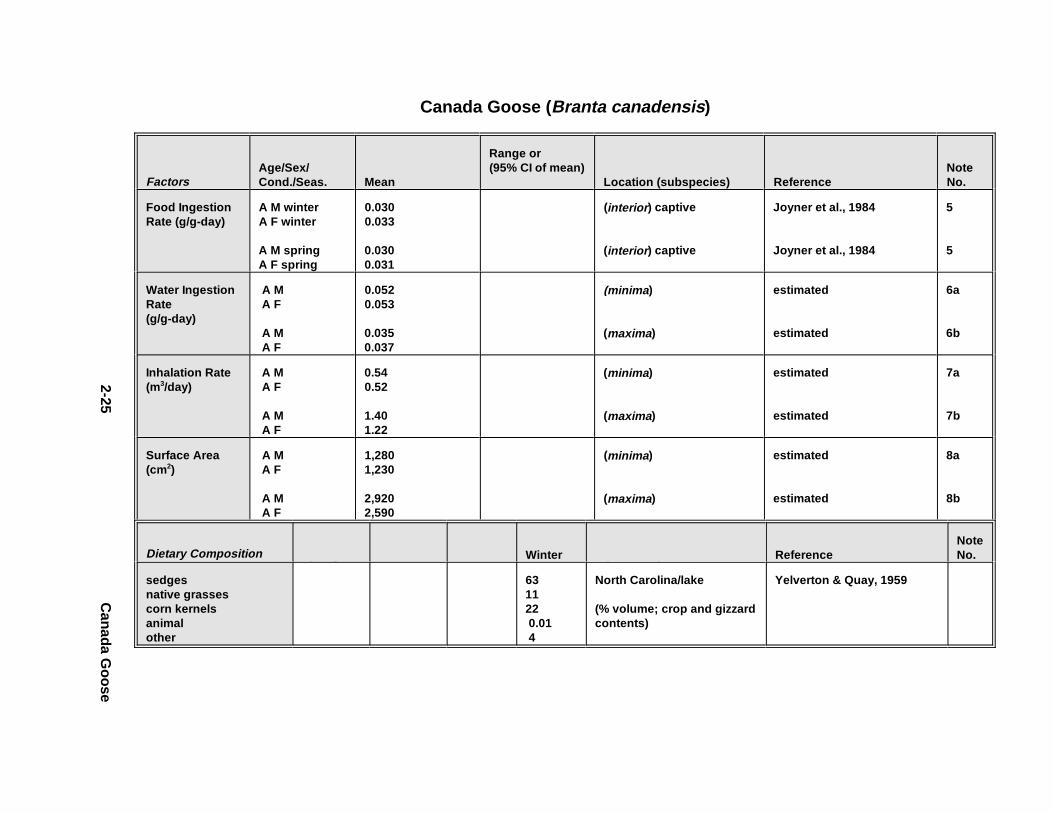
Canada Goose ( Branta canadensis )
2-25 C
anad
a Go
ose
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion (subspeci es) Reference No.
Range or
Food Ingest ion A M winter 0.030 (in terio r) captiv e Joyner et al., 1984 5 Rate (g/g-day) A F winter 0.033
A M spr ing 0.030 (in terio r) captiv e Joyner et al., 1984 5 A F spr ing 0.031
Water Ingest ion A M 0.052 (minima) est imated 6a Rate A F 0.053 (g/g-day)
A M 0.035 (maxima) est imated 6b A F 0.037
Inhal ation Rat e A M 0.54 (minima) est imated 7a (m /day) A F 0.523
A M 1.40 (maxima) est imated 7b A F 1.22
Surface Ar ea A M 1,280 (minima) est imated 8a (cm ) A F 1,2302
A M 2,920 (maxima) est imated 8b A F 2,590
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
sedges 63 North Carolina/lake Yelverton & Q uay, 1959 nat ive gr asses 11 cor n kernels 22 (% vol ume; crop and gi zzard animal 0.01 cont ents) other 4
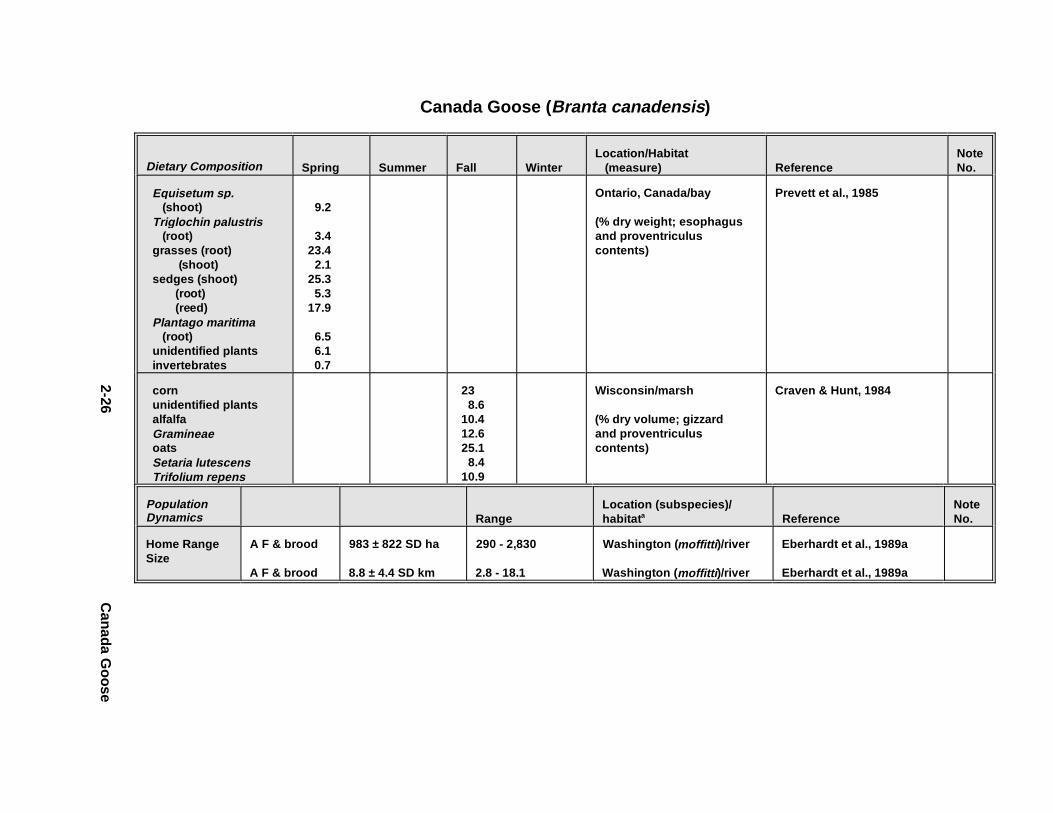
Canada Goose ( Branta canadensis )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
Equisetum sp. Ontario , Canada/bay Prevett e t al., 1985 (shoot ) 9.2
Triglochin palust ris (% dry weight ; esophagus (root ) 3.4 and pr ovent riculus
grasses ( root ) 23.4 cont ents) (shoot ) 2.1
sedges ( shoot ) 25.3 (root) 5.3 (reed) 17.9
Plantago maritima (root) 6.5
uni dent ified pl ants 6.1 invertebrates 0.7
cor n 23 Wisconsi n/marsh Craven & Hunt , 1984 uni dent ified pl ants 8.6 alfalfa 10.4 (% dry vol ume; gizzard Gramineae 12.6 and pr ovent riculus oats 25.1 cont ents) Setaria lutescens 8.4 Trifo liu m repens 10.9
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/ Note Cond. /Seas. Mean Range habi tat Reference No.a
Home Range A F & br ood 983 ± 822 SD ha 290 - 2,830 Washi ngt on (moffitti )/riv er Eberhard t et al., 1989a Size
A F & br ood 8.8 ± 4.4 SD km 2.8 - 18.1 Washi ngt on (moffitti )/riv er Eberhard t et al., 1989a
2-26 C
anad
a Go
ose

Canada Goose ( Branta canadensis )
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/ Note Cond. /Seas. Mean Range habi tat Reference No.a
Popul ation summer 0.02-12.4 nest s/ha var ious l ocat ions Cooper , 1978 9 Densi ty
fall 22 birds/ha Missouri/w ild life refuge Humburg et al., 1985
winter 4 birds/ha Missouri/w ild life refuge Humburg et al., 1985
16.6 nest s/ha Mont ana (moffitti )/ Geis, 1956
1.3 nest s/ha Mont ana (moffitti )/
0.35 nest s/ha Alaska ( leucopus)/ Byrd & Wool ingt on, 1983
on 0.2-0.8 ha island
on 8-121 ha island
island pr eferred habi tat
Clutch Si ze 4.7 Alaska (minima) Spencer et al., 1951 10 5.6 ± 0.1 SE 2 - 8 Alaska ( leucopa) Byrd & Wool ingt on, 1983 4.6 Ontario, Canada ( in terio r) Ravel ing & Lum sden, 1977 2 5.6 Alabama, Georgia (maxima) Combs et al., 1984
Clutches/ Year 1 issour i Brakhage, 1985
Days 25 NS (minima) Laidley, 1939 10 Incubat ion 28 Missour i (maxima) Brakhage, 1965
Age at 40-46 Alaska (minima) Mickel son, 1973 11 Fledgi ng 55 NS (leucopa) Lee (pers. com m.) in Byrd (days) & Wool ingt on, 1983
63 Ontario, Canada ( in terio r) Hanson, 1965 11 71-73 Michi gan (maxima) Sherwood, 1965 11
Percent Nests 91 89 - 93 Alaska/ island (leucopa) Byrd & Wool ingt on, 1983 Successf ul 44 27 - 64 Alabama, Georgia (maxima) Combs et al., 1984
Number 2.19 ± 2.42 SD 0 - 7 Washi ngt on (moffitti ) Eberhardt et al., 1989b Fledge per Act ive Nest
M
2-27 C
anad
a Go
ose
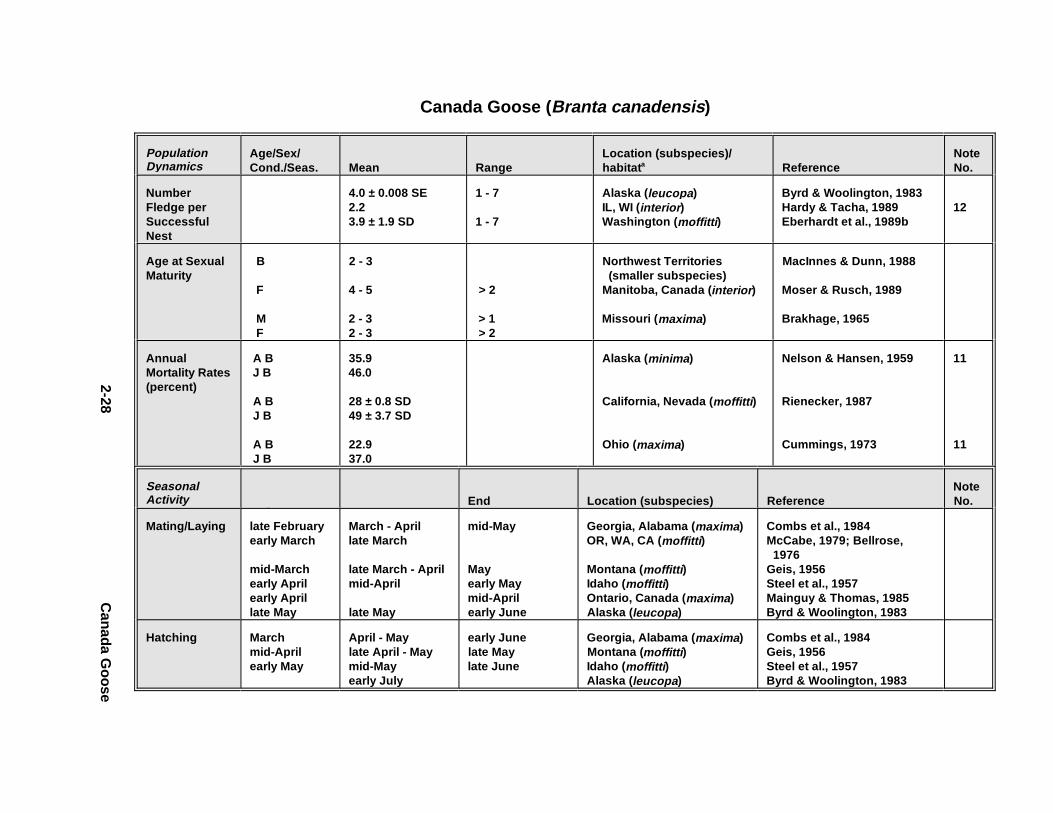
Canada Goose ( Branta canadensis )
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/ Note Cond. /Seas. Mean Range habi tat Reference No.a
Number 4.0 ± 0.008 SE 1 - 7 Alaska ( leucopa) Byrd & Wool ingt on, 1983 Fledge per 2.2 IL, WI (in terio r) Hardy & Tacha, 1989 12 Successf ul 3.9 ± 1.9 SD 1 - 7 Washi ngt on (moffitti ) Eberhard t et al., 1989b Nest
Age at Sexual B 2 - 3 Northwest Territories MacInnes & Dunn, 1988 Maturity (smaller subspeci es)
F 4 - 5 > 2 Mani toba, Canada (in terio r) Moser & Rusch, 1989
M 2 - 3 > 1 Missour i (maxima) Brakhage, 1965 F 2 - 3 > 2
Annual A B 35.9 Alaska (minima) Nelson & Hansen, 1959 11 Mortality Rates J B 46.0 (percent )
A B 28 ± 0.8 SD California, Nevada (moffitti ) Rienecker , 1987 J B 49 ± 3.7 SD
A B 22.9 Ohio (maxima) Cummings, 1973 11 J B 37.0
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating/Laying late February March - Apr il mid-May Georgia, Alabama (maxima) Combs et al., 1984 early March late March OR, WA, CA (moffitti ) McCabe, 1979; Bel lrose,
mid-March late March - Apr il May Mont ana (moffitti ) Geis, 1956 early April mid -April early May Idaho (moffitti ) Steel et al., 1957 early April mid -April Ontario , Canada (maxima) Mainguy & Thom as, 1985 late May late May early June Alaska (leucopa) Byrd & Wool ingt on, 1983
1976
Hatching March Apr il - May early June Georgia, Alabama (maxima) Combs et al., 1984 mid-Apr il late Apr il - May late May Mont ana (moffitti ) Geis, 1956 early May mid-May late June Idaho (moffitti ) Steel et al., 1957
early Jul y Alaska (leucopa) Byrd & Wool ingt on, 1983
2-28 C
anad
a Go
ose

�
Canada Goose ( Branta canadensis )
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Molt (fall) mid-June Idaho (moffitti ) Steel et al., 1957 mid-Jul y mid-August late August Alaska (leucopa) Byrd & Wool ingt on, 1983 late June late October Illin ois (in terio r) Willia ms & Kendeigh, 1982
Migration fall mid-Sept. Novem ber arrive sout h Illinoi s (in terio r) Bell & Kl imstra, 1970
spr ing February early March leave Illin ois (in terio r) Bell & Kl imstra, 1970
October early Novem ber mid-December arrive CO, TX (parvipes) Grieb, 1970
late March early Apr il leave Minnesot a (maxima) Ravel ing, 1978b
1 Weight s est imated from graph. 2 Cited in Dunn and M acInnes ( 1987).
2-29 C
anad
a Go
ose
3 Est imated range of existence t o maximum free-living m etabol ism at typical breeding gr ound ( Ontario, Canada i n spr ing and sum mer) and at typical wintering gr ound ( sout h Illinoi s in fall and w inter). Est imated usi ng r egressi on equat ions devel oped by the author s, measur es of metabol ic rates at temperatures f rom -40 to 41�C, and t emperatures typical for the season and l ocat ion.
4 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom (a) Ravel ing (1979); (b) Ravel ing (1968); and (c) Brakhage ( 1965). 5 Repor ted as gr ams dr y weight of feed; cor rected to grams wet weight of feed usi ng t he measur ed moisture cont ent of 11 percent (on aver age) of
the feed i tems (i.e., cor n, sunf lower seeds, wheat, and m ilo). 6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom (a) Ravel ing (1979) and (b) Brakhage ( 1965). 7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom (a) Ravel ing (1979) and (b) Brakhage ( 1965). 8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom (a) Raveling (1979)
and (b) Brakhage ( 1965). 9 Summarizing sever al studi es, ci ted in Byrd & Wool ingt on (1983).
10 Cited in Palmer (1976). 11 Cited in Bel lrose (1976). 12 For parents ol der than 5 years of age.

References (including A ppendix)
Akesso n, T. R.; Raveling, D. G. (1981) Endoc rine and body weight ch anges of n esting a nd non- nesting Ca nada geese. Biol. Reprod. 25: 792-804.
Balham, R. W. (1954) The behavior of th e Canada goose (Branta can adensis) in Manit oba [Ph.D. disser tat ion] . Colum bia, MO: Univ ersi ty of Missour i.
Bell, R. Q.; Klimstr a, W. D. (1970) Feeding acti vi ties of Canada geese (Branta can adensis interio r) in s out hern Illinois . Trans. Ill. State. Acad. Sci . 63: 295-304.
Bellrose, F. C. (1976) Ducks, geese, and swan s of North Amer ica. Harrisburg, PA: The Stack pole Co.
Best, R. G.; Fowler, R.; Hause, D., et al . (1982) Aerial thermal infrared census of Canada geese in South Dakota. Photogr. Eng. Remote Sens. 48: 1869-1877.
Brakhage, D. H. (1985) A second br ood by Canada geese. Wilson Bull. 97: 387-388.
Brakhage, D. H.; Baskett, T. S.; Graber, D. A., et al . (1987) Impacts o f a new reservoir on resident Canada geese. Wildl. Soc. Bull. 15: 192-196.
Brakhage, G. K. (1965) Biology and be havior of tub- nesting Ca nada geese. J. Wildl. Manage. 29: 751-771.
Buchsbaum, R.; Valiela, I. (1987) Variabilit y in t he chemistr y of estu arine plants an d its effects o n feeding by Canada geese. Oecologia (B erlin) 73: 146-153.
Buchsbaum, R.; Valiela, I.; Teal, J. M. (1981) Grazing by Canada geese and related aspects of th e chemistr y of sal t mar sh grasses. Colonia l Waterbirds 4: 126-131.
Buchsbaum, R.; Valiela, I.; Swain, T. (1984) The role of phenolic co mpounds and ot her plant cons ti tuents in f eeding by Canada geese in a coastal marsh. Oecologia (Berlin) 63: 343-349.
Bultsma, P. M.; Linde r, R. L.; Kuc k, T. L. (1979) Reproducti ve su ccess o f giant Canada geese in wester n South Dakota. Proc. SD Acad. Sci . 58: 35-38.
Byrd, G. V.; Woolingt on, D. W. (1983) Ecology of A leutian Canada geese at Buldir Island, Alaska. U.S. Fish Wildl. Serv. Spec. Sci . Rep. No. 253.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chapman, J. A. (1970) Weights an d measu rements o f dusky C anada geese wi nter ing in Oregon. Mur relet 51: 34-37.
2-30 Canada Goose

Chapman, J. A.; Henny , C. J.; Wight , H. M. (1969) The statu s, popula tion dy namics, an d harvest o f th e dusky C anada goos e. Wildl. Monogr . 18.
Coleman, T. S.; Boag, D. A. (1987) Foraging c haracter isti cs o f Canada geese on the Nisutl in River delta, Yukon. Can. J. Zool. 65: 2358-2361.
Collia s, N. E.; Jahn, L. R. (1959) Social behavior and breeding s uccess i n Canada geese (Branta can adensis) conf ined unde r semi -natural condit ions . Auk 76: 478-509.
Combs, D. L.; Ortego, B.; Kennamer , J. E. (1984) Nest ing biology of a reside nt floc k of Canada geese. Proc. Annu. Conf . Sout heast. Asso c. Fish Wildl. Agencies 38: 228-238.
Coope r, J. A. (1978) Canada geese at Marshy Point , Manit oba. Wildl. Monogr . 51: 1-87.
Cornley, J. E.; Campbell, B. H.; Jarvis, R. L. (1985) Produc tivi ty, mo rtalit y and popula tion statu s of dusky C anada geese. Trans. North Am. Wildl. Nat. Resour . Conf . 50: 540-548.
Craven, S. R. (1981) The Canada goose (Branta can adensis)--an annotated bibliogr aphy . U.S. Fish Wildl. Serv. Spec. Sci . Rep. No. 231.
Craven, S. R.; Hunt , R. A. (1984) Fall food ha bit s of Canada geese in Wisco nsin. J. Wildl. Manage. 48: 169-173.
Cummings , G. E. (1973) The Tennessee Val ley popula tion of Canada geese. U.S. Fish Wildl. Serv. Unpublis hed Repor t.
Dey, N. H. (1966) Canada goos e produc tion a nd popula tion stabilit y, Ogde n Bay water fowl management area, Utah. Utah State Dept. Fish and Game Publ. 66-7.
Dow, J. S. (1943) A stu dy of nest ing Ca nada geese in Honey Lake Val ley, California. Calif. Fish and Game 29: 3-18.
Dunn, E. H.; MacInne s, C. D. (1987) Geographic var iation in c lut ch size and body size of Canada geese. J. Field Ornithol. 58: 355-371.
Eberhardt, L. E.; Anthony , R. G.; Rickar d, W. H. (1989a) Movemen t and habitat u se by Great Basin Ca nada goos e broods . J. Wildl. Manage. 53: 740-748.
Eberhardt, L. E.; Anthony , R. G.; Rickar d, W. H. (1989b) Survival of juvenile Canada geese dur ing t he rearing pe riod. J. Wildl. Manage. 53: 372-377.
Eberhardt, L. E.; Book s, G. G.; Anthony , R. G.; et al. (1989c) Acti vi ty budgets o f Canada geese during br ood r earing. Auk 106: 218-224.
Estel , B. L. (1983) Winter weights o f Canada geese in southern Illinois dur ing 1982-83. Ill. Dep. Cons erv. Per . Rep. No. 38.
2-31 Canada Goose

Fitzner, R. E.; Rickar d, W. H. (1983) Canada goos e nesting pe rfor mance along t he Hanford Reach of th e Colum bia River, 1971-1981. Northwest Sci . 57: 267-272.
Geis, M. B. (1956) Producti vi ty o f Canada geese in the Flathead Valley, Mont ana. J. Wildl. Manage. 20: 409-419.
Geis, A. D.; Taber, R. D. (1963) Measuring hunt ing a nd ot her mo rtalit y. In: Mosby, H. S., ed. Wildlif e investi gationa l technique s. Washingt on, DC: The Wildlif e Society; pp. 284-298.
Grieb, J. R. (1970) The shortgrass p rairie Canada goos e popula tions . Wildl. Monogr . 22: 4-49.
Gulde n, N. A.; Johns on, L. L. (1968) Histo ry, behavior and management of a fl ock o f giant Canada geese in southeaster n Minne sota. In: Hine, R. L.; Schoe nfeld, C., eds. Canada goose man agement. 1st ed . Madison, WI: Dembar Educ. Res. Serv.; pp. 58-71.
Hanson, H. C. (1965) The giant Canada goos e. Carbonda le, IL: Sout hern Illinois Univ ersi ty Press.
Hanson, H. C.; Smith, R. H. (1950) Canada geese of th e Missi ssippi f lyway wi th special reference to an Illinois floc k. Ill. Nat. His t. Surv. Bull. 25: 67-210.
Hanson, W. C.; Eberhardt, L. L. (1971) A Colum bia River Canada goos e popula tion, 1950-1970. Wildl. Monogr . 28.
Hardy, J. D.; Tacha, T. C. (1989) Age-related recruitmen t of Canada geese fr om th e Mississippi V alley popula tion. J. Wildl. Manage. 53: 97-98.
Harvey, W. F., IV; Maleki , R. A.; Soutiere, E. C. (1988) Habitat u se by fo raging Ca nada geese in Ke nt Count y, Maryland. Trans Nor theast Sect. Wildl. Soc. 45: 1-7.
Hille y, J. D. (1976) Producti vi ty o f a resident giant Canada goose fl ock i n northwester n South Dakota [master 's th esis]. Brookings , SD: Sout h Dakota State Universi ty.
Humburg, D. D.; Graber, D. A.; Babcock, K. M. (1985) Facto rs affect ing a utumn and w int er dis tr ibut ion of Canada geese. Trans. North Am. Wildl. Nat. Resour . Conf . 50: 525-539.
Jensen, G. H.; Nelson, A. L. (1948) (ci ted in Palmer , 1962) U.S. Fish Wildl. Serv., Spec. Sci . Rept .-Wildlif e no. 60.
2-32 Canada Goose

Johns on, D. H.; Timm, D. E.; Springe r, P. F. (1979) Morphologic al character isti cs o f Canada geese in the Paci fic fl yway. In: Jarvis, R. L.; Bartonek, J. C., eds. Management and biology of fl yway g eese: a symp osium; February 16, 1979; Portland, OR. Corvallis , OR: OSU Book Sto res; pp. 56-68.
Joyner, D. E.; Arthur, R. D.; Jaco bson, B. N. (1984) Winter weight dynamics, grain cons umpt ion a nd r eproduc tive pot ential in Canada geese. Condor 86: 275-280.
Korsch gen, L. J. (1955) Fall foods of water fowl in Misso uri. Misso uri Dept. Conserv. P-R Ser. 14.
Kor tr ight , F. H. (1942) The ducks, geese, and swan s of North Amer ica. Harrisburg, PA: The Stack pole Co.
Kor tr ight , F. H. (1955) The ducks, geese, and swan s of North Amer ica. Harrisburg, PA: The Stack pole Co. and Washingt on, DC: Wildlif e Management Insti tute.
Laidley (1939) (ci ted in Palmer , 1962). Avicultural Mag. 5th Ser.: 102-103.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lebeda, C. S.; Ratti , J. T. (1983) Reproduc tive biology of Van couv er Canada geese on Admiralty Island, Alaska. J. Wildl. Manage 47: 297-306.
LeBlanc, Y. (1987a) Intraclutch var iation in e gg s ize of Canada geese. Can. J. Zool. 65: 3044-3047.
LeBlanc, Y. (1987b) Relations hips b etween sex o f gosling a nd pos it ion in t he laying sequence, egg mass, hatchling s ize, and f ledgling s ize. Auk 104: 73-76.
LeBlanc, Y. (1987c) Egg mass, posi tion in t he laying s equence, and br ood s ize in r elation t o Canada goose reproducti ve su ccess. Wilson Bull. 99: 663-672.
Leopold, A. S.; et al. (1981) North Amer ican game birds and mammal s. New York, NY: Charles Scr ibne r & Sons .
MacInnes, C. D. (1962) Nest ing of smal l Canada geese near Eski mo Point , Nor thwest Territories. J. Wildl. Manage. 26: 247-256.
MacInnes, C. D.; Davis, R. A.; Jones, R. N., et al . (1974) Reproducti ve effi ciency o f McConne ll Riv er smal l Canada geese. J. Wildl. Manage. 38: 686-707.
MacInne s, C. D.; Dunn, E. H. (1988) Esti mat ing pr opor tion of an age class n est ing in Canada geese. J. Wildl. Manage. 52: 421-423.
Mainguy , S. K.; Thomas, V. G. (1985) Comparisons of body r eserve buildup a nd us e in sever al groups of Canada geese. Can. J. Zool. 63: 1765-1772.
2-33 Canada Goose

Manning, T. H. (1978) Measurements an d weight s of eggs of th e Canada goos e, (Branta canadensis), analyzed and c ompared wi th thos e of other sp ecies. Can. J. Zool. 56: 676-687.
Martin, F. W. (1964) Behavior and survival of Canada geese in Utah. Utah State Dep. Fish and Game Inform. Bull. 64-7.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York, NY: McGraw-Hill Book Company, Inc.
McCabe, T. R. (1979) Produc tivi ty and ne sting ha bitat o f great basin Canada geese, Umatilla , Oregon. In: Jarvis, R. L.; Bartone k, J. C., eds. Management and biology of flyway g eese: a symp osium; February 16, 1979; Portland, OR. Corvallis , OR: OSU Book Sto res; pp. 117-129.
McLandress, M. R.; Raveling, D. G. (1981) Changes in diet and body co mpos it ion of Canada geese before spring m igr ation. Auk 98: 65-79.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Mickel son, P. G. (1973) Breeding biology of cack ling g eese (Branta can adensis mi nima Ridgeway) an d asso ciated species on the Yukon-Kusko kwi m Delta, Alaska [Ph .D. disser tat ion] . Ann A rbor , MI: Univ ersi ty of Michiga n.
Mille r, A. W.; Collins , B. D. (1953) A nest ing s tudy of Canada geese on Tule Lake and Lower Klamath Nationa l Wildlif e Refuge s, Siski you Count y, Calif ornia . Calif . Fish and Game 39: 385-396.
Moffi tt, J. (1931) The statu s of th e Canada goose in California. Calif. Fish and Game 17: 20-26.
Moser , T. J.; Rusch , D. H. (1989) Age-speci fic breeding r ates o f femal e interio r Canada geese. J. Wildl. Manage. 53: 734-740.
Murphy, A. J.; Boag, D. A. (1989) Body r eserve and food us e by inc ubating Ca nada geese. Auk 106: 439-446.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Naylor, A. E. (1953) Produc tion of th e Canada goos e on Hone y Lake Refuge, Lassen Count y, Calif ornia . Calif . Fish and Game 39: 83-94.
Nelson, A. L.; Martin, A. C. (1953) Gamebird weight s. J. Wildl. Manage. 17: 36-42.
2-34 Canada Goose

Nelson, U. C.; Hansen, H. A. (1959) The cack ling goo se--i ts mi gration a nd m anagement. Trans. Nor th Am. Wildl. Nat. Resour . Conf . 24: 174-187.
Owen, M. (1980) Wild g eese of th e world. Their life histo ry and ecology . London, UK: B. T. Batsfo rd Ltd.
Palmer , R. S. (1962) Handbook of Nor th Amer ican bir ds: v. 1. New Haven, CT: Yale Universi ty Pr ess.
Palmer , R. S. (1976) Handbook of Nor th Amer ican bir ds: v. 2. New Haven, CT: Yale Universi ty Pr ess.
Peach, H. C.; Thomas, V. G. (1986) Nutr ient co mposi tion of yolk in r elation t o early growth of Canada geese. Physiol. Zool. 59: 344-356.
Prevett, J. P.; Marshall, I. F.; Thomas, V. G. (1985) Spring f oods of snow and Ca nada geese at James B ay. J. Wildl. Manage. 49: 558-563.
Ratti , J. T.; Timm, D. E.; Robards, F. L. (1977) Weights an d measu rements o f Vancouver Canada geese. Bird Banding 4 8: 354-357.
Raveling, D. G. (1968) Weight s of Branta can adensis inter ior dur ing w int er. J. Wildl. Manage. 32: 412-414.
Raveling, D. G. (1978a) Morphology of th e cack ling Ca nada goos e. J. Wildl. Manage. 42: 897-900.
Raveling, D. G. (1978b) Dynamics o f distr ibut ion of Canada geese in winter . Trans. North Am. Wildl. Nat. Resour . Conf . 43: 206-225.
Raveling, D. G. (1979) The annua l cycl e of body co mpos it ion of Canada geese wi th special reference to control of reproduc tion. Auk 96: 234-252.
Raveling, D. G. (1981) Survival , experience, and age in relation t o breeding s uccess o f Canada geese. J. Wildl. Manage. 45: 817-829.
Raveling, D. G.; Lum sden, H. G. (1977) Nest ing e cology of Canada geese in the Huds on Ba y low lands of On tar io: evolut ion a nd popula tion r egula tion. U.S. Fish Wildl. Res. Rep. 98; pp. 1-77.
Rienecker , W. C. (1987) Popula tion t rends , dis tr ibut ion, and s urviv al of Canada geese in California and wester n Nevada. Calif. Fish and Game 73: 21-36.
Rienecker , W. C.; Anderson, W. (1960) A water fowl nest ing s tudy on T ule L ake and Lower Klamath Nationa l Wildlif e Refuge s, 1957. Calif. Fish and Game 46: 481-506.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
2-35 Canada Goose

Samuel, M. D.; Rusch , D. H.; Craven, S. (1990) Influence of neck b ands on recovery and survival rates o f Canada geese. J. Wildl. Manage. 54: 45-54.
Sedinge r, J. S. (1986) Growth and developm ent of Canada goos e gos lings . Condor 88: 169-180.
Sedinge r, J. S.; Raveling, D. G. (1984) Dietary sel ecti vi ty i n relation t o availa bilit y and qualit y of food f or gos lings of cack ling g eese. Auk 101: 295-306.
Sedinge r, J. S.; Raveling, D. G. (1986) Timing of n esting by Canada geese in relation t o the phenology and availa bilit y of th eir food pla nts. J. Anim . Ecol. 55: 1083-1102.
Sherwood, G. A. (1965) Canada geese of th e Seney Nationa l Wildlif e Refuge . Minne apolis , MN: U.S. Fish Wildl. Serv. Compl. Rep., Wildl. Manage. Stud. 1, 2.
Sherwood, G. A. (1966) Flexible plasti c colla rs compared to nasal discs fo r marking g eese. J. Wildl. Manage. 30: 853-855.
Smith, R. H.; Sutto n, E. L. (1953) Water fowl breeding gr ound s urvey i n northern Alberta, the Northwest T erritories, and the Yukon. In: Water fowl popula tion a nd br eeding condit ions . U.S. Fish Wildl. Serv. and Ca nadian Wildl. Serv.; Spec. Sci . Rep., Wildl. 25; pp. 7-15.
Smith, R. H.; Sutto n, E. L. (1954) Water fowl breeding gr ound s urvey i n northern Alberta, the Northwest T erritories, and the Yukon. In: Water fowl popula tion a nd br eeding condit ions . U.S. Fish Wildl. Serv. and Ca nadian Wildl. Serv.; Spec. Sci . Rep., Wildl. 27; pp. 11-20.
Spencer , D. L., et al . (1951) Amer ica's greatest B rant goos e nesting a rea. Trans. Nor th Am. Wildl. Nat. Resour . Conf . 16: 290-295.
Steel , P. E., et al . (1957) Canada goos e produc tion a t Gray's Lake, Idaho, 1949-1951. J. Wildl. Manage. 21: 38-41.
Szymcz ak, M. R. (1975) Canada goose resto ration a long t he foot hills of Color ado. Colo. Dept. Nat. Resources Wildl. Div . Tech. Publ. 31.
Thomas, V. G.; Peach Brown, H. C. (1988) Relations hips among e gg s ize, energy r eserves, growth rate, and fast ing r esistan ce of Canada goos e gos lings fr om sout hern Ontar io. Can. J. Zool. 66: 957-964.
Thomas, V. G.; Mainguy , S. K.; Prevett, J. P. (1983) Predict ing fat co ntent of geese fr om abdom inal fat we ight . J. Wildl. Manage. 47: 1115-1119.
Thor nbur g, D. D.; Tacha, T. C.; Estel , B. L., et al . (1988) Spatial and temp oral var iation in wint er weight s of Mississippi V alley Canada geese. In: Weller, M. W., ed. Water fowl in w int er. Minne apolis , MN: Univ ersi ty of Minne sota Press; pp. 271-275.
2-36 Canada Goose

Timm, D. (1974) Statu s of lesser Canada geese in Alaska. Juneau, AK: Alaska Dep. Fish and Game; Paci fic Flyway T ech. Com. Rep. 38-50.
Trainer, C. E. (1959) The 1959 wester n Canada goose (Branta can adensis occi dental is) study of th e Coppe r River Delta, Alaska. U.S. Bur. Sport Fish. Wildl. Annu. Water fowl Rep., Alaska (mi meo).
Vaught , R. W.; Kir sch , L. M. (1966) Canada geese of th e easter n prairie popula tion, wi th special reference to the Swan Lake fl ock. Misso uri Dept. Cons erv. Tech. Bull. 3.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
West, W. L. (1982) Annua l cycl e of th e giant Canada goos e floc k at the Trimble Wildlif e Area [master 's th esis]. Colum bia, MO: Univ ersi ty of Missour i.
Will, G. C. (1969) Producti vi ty o f Canada geese in Larimer Count y, Color ado, 1967-1968. [master 's th esis]. Fort Collins , CO: Color ado State Universi ty.
Willi ams, J. E.; Kendeigh, S. C. (1982) Energetics o f th e Canada goos e. J. Wildl. Manage. 46: 588-600.
Yelver ton, C. S.; Quay, T. L. (1959) Food ha bit s of th e Canada goos e at Lake Mattamu skeet, Nor th Carolina . Nor th Carolina W ildlif e Resour ces Commi ss ion.
Yocom, C. F. (1972) Weights an d measu rements o f Taverner's and Great Basin Canada geese. Murrelet 53: 33-34.
2-37 Canada Goose

2.1.3. Mallard (surface-feeding ducks)
Order Anserif ormes, Family Anatidae. Surface-fee ding du cks ar e the most fam ilia r ducks o f fr eshwater and sal twater wetl ands. They feed by dabbling a nd t ipping up in shallow water , often fi lter ing t hrough s oft mud f or food. They feed primar ily on seeds of aquatic plants an d cultivated grains, although t hey also cons ume aquatic inv ertebrates, partic ularly dur ing t he breeding seaso n (Jorde et al ., 1983; Swanson et al ., 1985). All species have a bright color ed patch of feath ers on the trailing e dge of each wing, and t he overall plum age of th e males is mo re color ful t han that of th e femal es. Dabbling duc ks range in s ize fr om th e green-winge d teal (average 37 cm bill t ip t o tail t ip) to the nor thern pintai l (average 66 cm).
Selected species
The mal lard (Anas platyr hynchos) feeds mo stl y on aquatic plants, seeds, and aquatic inver tebrates, depending on t he seaso n, and forages in ponds and wetl ands by dabbling a nd f ilt ering t hrough s edim ents. It is wide spread throughout mo st of th e Unit ed States an d is th e most abunda nt of th e Unit ed States d ucks (U SFWS, 1991). In the past decade, however , its n umbers have decline d markedly acr oss it s pr inc ipa l range in t he mid- cont inental region b ecause of habitat d egradation a nd dr ought (U SFWS, 1991). Mallards interbreed wi th domesti c ducks an d black d ucks ( Anas rubrip es).
Body size. Mallards aver age 58 cm fr om bill t ip t o tail t ip. Male malla rds are generally heavier than femal es (Delnicki and Reinecke, 1986; Whyte an d Bole n, 1984; see table). Female mal lards lose weight dur ing t he laying a nd inc ubation pe riods ; males los e weight fr om th eir sp ring a rrival through t he peak of th e breeding seaso n and then gain weight while th e femal es are incubating ( Lokemo en et al ., 1990a).
Habitat . Wint ering m allards pr efer natural botto mland wetl ands and rivers to reservoirs and farm ponds (H eitmeyer and Vohs, 1984); water depths of 20 to 40 cm ar e opt imum fo r fo raging ( Heitmeyer , 1985, ci ted in Allen, 1987). The primary habitat requir ement fo r nest ing a ppears to be dense grassy veg etat ion a t least a h alf meter high (Bellrose, 1976). Mallards prefer areas th at provide concealment fr om predators su ch as seeded cover (fields esta blis hed on f ormer cr opla nds ) (Klett et al ., 1988; Lokemo en et al ., 1990b), cool-seaso n introduced legumes an d grasses (D uebbert and Lokemo en, 1976), and idle grassl and wi th tal l, dense, rank co ver in the area (Duebbert and Kantrud, 1974). Nests usually are located wi thin a few k ilometer s of water , but i f choic e nesting ha bitat i s not avai lable nearby, femal es may n est fu rther away (B ellrose, 1976; Duebbert and Lokemo en, 1976).
Food ha bi ts . In winter , mal lards feed primar ily on seeds but also on inver tebrates asso ciated wi th leaf l itter and wetl ands, mast, agricultural grains, and to a limi ted exten t, leaves, buds , stems, rootlets, and tubers (Goodman and Fisher, 1962; Heitmeyer , 1985, ci ted in Allen, 1987). In spring, femal es sh ift fr om a largely herbivorous diet to a diet of mainly inv ertebrates to obtain protein for their prebasic mo lt and then for egg produc tion (Swanson and Meyer , 1973; Swanson et al ., 1979; Swanson et al ., 1985; Heitmeyer , 1988b). Laying femal es co nsume a highe r propor tion of anim al foods on t he breeding
2-39 Mallard

grounds th an do m ales or nonla ying femal es (Swan son et al ., 1985). The animal diet cont inue s throughout th e summer , as man y femal es lay cl utch es to replace destr oyed nests (Swan son et al ., 1979; Swanson et al ., 1985). Ducklings al so cons ume aquatic inver tebrates al most excl usively, particularly during t he period of r apid gr owth (Chur a, 1961). Mallards co ncentrate in wetl ands at night , apparent ly feeding on emerging in sects (Swanson and Meyer , 1973). Flocks may feed in unha rvested grain fields and stubble fi elds dur ing f all and w int er (Dillon, 1959). During pe riods of food s hor tage, fat r eserves ar e used as an energy sour ce. Dur ing br eeding, femal es co ntinue to feed but also use fat to meet the demands of egg pr oduc tion; femal es may l ose 25 percent of th eir body mass (i n fat) dur ing la ying a nd early inc ubation ( Krapu, 1981).
Mol t. Female mal lards mo lt into basic plumage in late wi nter or early sp ring, excep t for th e wing m olt , whic h is delayed unti l about th e time br oods ar e fledged. In males, head-body-tai l molt co mmen ces i n early su mmer and over laps or is follow ed by th e wing m olt . Mallards generally ar e flight less fo r about 25 days d uring t he wing m olt (Palmer , 1976).
Migration. Although t he malla rd wint ers in a ll f our water fowl flyways o f North Amer ica (i .e., Paci fic, Central, Missi ss ippi, and A tlant ic ), the Mississippi f lyway (a lluv ial valle y from Missour i to the Gulf of Mexico) cont ains th e high est n umbers (Bellrose, 1976). Human creation a nd a lteration of water bodie s and pla nt co mmunit ies have ch anged the migr ation a nd w int ering p atter ns of mal lards; in North Amer ica th e ducks wi nter far ther north than in the past (Jo rde et al ., 1983). Mallards ten d to arrive at th eir winter ing gr ounds in t he Mississippi V alley in m id-Septemb er through e arly Novemb er and depart fo r their nor therly breeding gr ounds ag ain in Ma rch (Fredr ickso n and Heitmeyer , 1988). Adult femal es th at reproduce su ccessf ully ar e lik ely to return to the same n est ing gr ound t he follow ing year (Lokemo en et al ., 1990a, 1990b).
Breeding acti vi ties and social organization. Older femal es ar rive at b reeding grounds ear lier th an year ling bir ds, whic h pr obably inc reases th eir chances o f reproducti ve su ccess b ecause th ey can select th e best n est si tes (L okemo en et al ., 1990b). First clut ches are generally finis hed by mid- April in t he sout hern part of th e breeding r ange and late April to May in the northern United States (Pal mer , 1976). High r ates o f nest fa ilur e requir e femal es to renest p ersisten tly to reproduce su ccessf ully (Swanson et al ., 1985). Average clutch size decreases as th e seaso n progresses b ecause th e clutch size of renesting femal es is smal ler than init ial c lut ches (Eldr idge and Kr apu, 1988; Lokemo en et al., 1990b). Older femal es produce larger clutch es th an do year lings (Lo kemo en et al ., 1990a). Mallards mate fo r one breeding seaso n, and males typ ical ly leave th e femal es at the onset o f incubation (Palmer , 1976). Females remain wi th the brood unt il f ledging. Mallards are ser ially monoga mous and t hus r emate annua lly (Palmer , 1976).
Home range and r esour ces. Each pair of mal lards uses a h ome range, and the drake co mmonly establis hes a ter ritory that he d efends against o ther mal lards (Bellrose, 1976). Home-range size depends on habitat, in particular the typ e and distr ibut ion of water habitats (e. g., prairie pothole s, r ivers), and popula tion de nsity (Bellr ose, 1976; Dwyer et al ., 1979; Kirby et al ., 1985).
2-40 Mallard

�
�
�
�
Popul ation de nsity . Mallard dens it ies dur ing t he breeding seaso n are posi tively correlated wi th avai labilit y of ter restr ial cover for nest ing a nd w ith availa bilit y of wetl ands and ponds th at pr ov ide th e aquatic die t of malla rds (Pospahala et al., 1974). Avai labilit y of suitable wetl and habitat fo r breeding a nd w int ering de pends on environm ental condit ions (e.g., rainfal l) (Heitmeyer and Vohs, 1984; Lokemo en et al ., 1990a). Average densi ties of breeding m allards in t he prairie pot hole r egion r ange fr om 0.006 to 0.67 pairs per hectar e (Duebbert and Kantrud, 1974; Duebbert and Lokemo en, 1976; Kantrud and Stewar t, 1977; Lokemo en et al ., 1990b). Mallards attai n their highest d ensi ties in prairie and parkland of the southern prairie provinces an d in the Coope r River and A thabasca River deltas o f Canada (Johns on and Grier, 1988).
Popul ation dy namics. Nest su ccess o r failur e is an impor tant facto r affecti ng malla rd popula tions . Mammal ian predation is th e main c ause of n est fa ilur e, follow ed by hum an dis turbance (e.g., farming ope rations ) and adverse weath er condit ions (Kl ett et al ., 1988; Lokemo en et al ., 1988). Mammal ian predators include fo x, badger, and s kunk ; crows also prey on mal lard nests (J ohns on et al., 1988). Mallards usually renest i f th e fi rst n est fai ls (Palmer , 1976). Juvenile su rviv al depends on f ood a nd pr efer red habitat avai labilit y, facto rs th at in turn are affected by environmental condit ions . For examp le, high r ainf all is related to increased wetl and area, which is posi tively co rrelated wi th duckling gr owth (Lokemo en et al ., 1990a). Annua l adult mo rtalit y rates var y wi th year , locat ion, hunt ing pressu re, age, and sex. Females su ffer greater natural mortal ity r ates (e.g., typ ical values of 40 to 50 percent) than do m ales (e.g., typic al value s of 30 to 40 percent) (Chu and Hestb eck, 1989). By fal l, there is a highe r propor tion of mal es than femal es in most popula tions (B ellr ose, 1976). Immatu re mortal ity r ates o f 70 percent have been recorded in many areas, although low er immatu re mortal ity r ates ar e more commo n (Bellrose, 1976; Chu and Hestb eck, 1989). Annua l mortalit y rates al so are greater in areas wi th highe r hunt ing pr essu re (Bellrose, 1976).
Simi lar species (fr om general references)
� The Amer ican black d uck (Anas rubrip es) is only present in the woode d parts o f northeaster n and north central United States. It nests n ear woodla nd l akes an d str eams o r in freshwater and tidal marshes. It is si mi lar in size (58 cm) to mal lards using t he same h abitats.
� The nor thern pint ail (Anas acu ta) is wide spread, occurring in m ost pa rts of Nor th Amer ica and br eeding t hroughout Canada and t he nor th central Unit ed States. Although f ormer ly far ily abunda nt , Nor th Amer ican pint ail popula tions h ave decline d dr amati cal ly during t he past d ecade (USFWS, 1991). It prefers mar shes and open areas wi th ponds and l akes. Pintai ls average slight ly longe r (66 cm) th an mal lards.
� The gadwall (Anas str epera) (51 cm) o ccu rs th roughout mo st of th e Unit ed States. In Canada, its b reeding r ange is lim ited to the sout h central pot hole s region. It is mo re common in t he west th an in the east.
� The Amer ican wigeon (Anas amer icana) (48 cm) b reeds th roughout mo st of Canada and in t he prairie pot hole r egions of th e Unit ed States. It wi nters
2-41 Mallard

�
�
�
�
along bot h the east an d west co asts o f th e United States as wel l as far ther sout h int o Mexico.
� Northern shovelers (Anas cl ypeata) (48 cm), inha bit ants of mar shes, ponds , and bays, breed throughout mid t o wester n Canada and the prairie pothole regions of th e Unit ed States. They wi nter along t he gulf co ast, southern Atlantic co ast, in Texas, and a few o ther southwester n states as wel l as throughout Mexico.
� Blue -winge d teal (Anas disco rs) (39 cm) ar e fai rly co mmo n in open count ry in m arshes and on ponds and l akes. Breeding popula tions occur th roughout the central United States an d Canada, but wi nter ing popula tions ar e restr icted to Atlantic and Paci fic co astal areas.
� The green-winge d teal (Anas cr ecca) (37 cm) i s th e smal lest o f th e dabbling ducks. A. c. carolinensis is th e most co mmo n subspecies in the United States. It breeds th roughout mo st of Canada and t he prairie pot hole r egion of th e United States. It overwinters in the southern half of th e United States and in Me xico.
� Cinna mon teal (Anas cyan optera) (41 cm) b reeding popula tions ar e restr icted to the wester n United States an d Mexico, wi th few r eaching s out hern Canada. Some popula tions in Calif ornia and Me xico are year -round residents.
General references
Allen (1987); Nationa l Geogr aphic So ciety (1987); Pospahala et al . (1974); Palmer (1976); Bellrose (1976).
2-42 Mallard
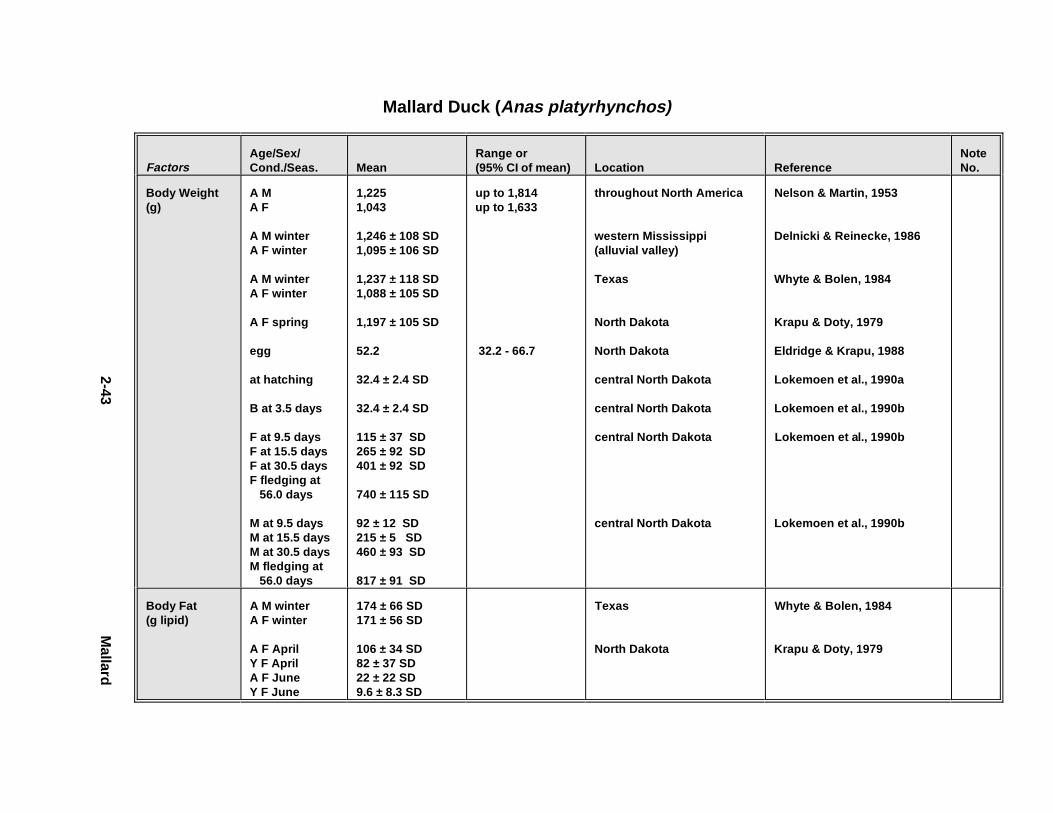
Mallard Duck ( Anas platy rhy nchos )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A M 1,225 up t o 1,814 throughout North Am erica Nelson & M artin, 1953 (g) A F 1,043 up t o 1,633
A M winter 1,246 ± 108 SD western Missi ssi ppi Delnicki & Reinecke, 1986 A F w inter 1,095 ± 106 SD (alluvial val ley)
A M winter 1,237 ± 118 SD Texas Whyte & Bol en, 1984 A F w inter 1,088 ± 105 SD
A F spr ing 1,197 ± 105 SD North Dakot a Krapu & Dot y, 1979
egg 52.2 32.2 - 66.7 North Dakot a Eldr idge & Kr apu, 1988
at hatching 32.4 ± 2.4 SD centra l North Dakota Lokemoen et al., 1990a
B at 3.5 days 32.4 ± 2.4 SD centra l North Dakota Lokemoen et al., 1990b
F at 9.5 days 115 ± 37 cent ral North Dakot a Lokem oen et al., 1990b F at 15.5 days 265 ± 92 F at 30.5 days 401 ± 92 F fledgi ng at
56.0 days 740 ± 115 SD
M at 9.5 days 92 ± 12 SD centra l North Dakota Lokemoen et al., 1990b M at 15.5 days 215 ± 5 M at 30.5 days 460 ± 93 M fledgi ng at
56.0 days 817 ± 91
Body Fat A M winter 174 ± 66 SD Texas Whyte & Bol en, 1984 (g lipid) A F winter 171 ± 56 SD
A F Apr il 106 ± 34 SD North Dakot a Krapu & Dot y, 1979 Y F Apr il 82 ± 37 SD A F June 22 ± 22 SD Y F June 9.6 ± 8.3 SD
SD SD SD
SD SD
SD
2-43 M
allard

Mallard Duck ( Anas platy rhy nchos )
2-44 M
allard
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Metabol ic Rate A F basal 77 est imated 1 (kcal /kg-day) A M basal 73
A F w inter 280 Texas Whyte & Bol en, 1984 2 A M winter 220
A F f ree-living 200 (94 - 424) est imated 3 A M free-living 192 (91 - 408)
Food Ingest ion 4 Rate (g/g-day)
Water A F 0.058 est imated 5 Ingest ion A M 0.055 Rate (g/g-day)
Inhal ation A F 0.42 est imated 6 Rate (m /day) A M 0.483
Surface Ar ea A F 1,030 est imated 7 (cm ) A M 1,1482
Dietary Compositio n
Locat ion/Habi tat Note (measur e) Reference No.
adul ts: Winter Loui siana/coast al marsh Dillon, 1959 rice 24 and pr airie jungl e rice 21 brownseed paspal um 19 (% vol ume; gul let cont ents) barnyard grass 8.0 red ric e 8.0 knot grass 6.5 signal grass 2.5 coast cockspur 1.9 Mamaica saw grass 1.3 snails 1.0 other 6.8

Mallard Duck ( Anas platy rhy nchos )
Dietary Compositio n
Locat ion/Habi tat Note (measur e) Reference No.
breeding female: April May June south centra l North Swanson et al., 1985 (total animal) (67.8) (66.8) (89.4) Dakot a/prairie pot hol es gast ropods trace 24.9 16.5 insect s 13.1 25.6 48.1 (% wet vol ume; esophagus crust acea 7.9 15.1 13.9 cont ents) annelid s 38.3 0.2 10.9 misc. animal 8.5 1.0 -
(total plant) (32.2) (33.2) (10.6) seeds 28.7 28.7 10.6 tubers 2.4 4.3 -stems 1.1 0.2 -
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Home Range spr ing: Size (ha) A F total 468 ± 159 SD 307 - 719 North Dakot a/prairie Dwyer et al., 1979
A F laying 111 ± 76 SD 38 - 240 pot hol es spr ing: A F 540 40 - 1,440 Minnesota/wetlands, riv er Kirby et al., 1985 A M 620 70 - 1,140
Popul ation A B spr ing 0.036 0.006 - 0.076 cent ral North Dakot a/range Lokem oen et al., 1990a Densi ty (area 1) of 6 years of data from (pairs/ha) A B spr ing 0.047 0.031 - 0.087 two di fferent pot hol e
(area 2) areas
Clutch yearling 9.3 ± 1.7 SE North Dakot a/prairie Krapu & Dot y, 1979 Size A 10.3 ± 1.1 SE pot hol es
9 1 - 18 NS/NS Bellrose, 1976
Clutches if lost up t o 4.5 North Dakot a/exper imental Swanson, unpubl ished i n /Year ponds ( nests pur posel y Swanson et al., 1985
if successf ul 1 North Am erica/NS Bel lrose, 1976 destroyed)
Days 26 23 - 29 NS/NS Bent, 1923 8 Incubat ion 25 North Dakot a/wetlands Klett & Johnson, 1982
2-45 M
allard

Mallard Duck ( Anas platy rhy nchos )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Age at 52 - 60 NS/NS Bellrose, 1976 Fledgi ng (days) 56 centra l North Dakota/ Lokemoen et al., 1990a
pot hol es
Percent Nests 51 - 61 Sout h Dakot a/prairie Duebber t & Lokem oen, 1976 Successf ul pot hol es and f ields
9 - 10 eastern Sout h Dakot a/ Klett et al., 1988 pot hol es
Number 4.9 NS/NS Cowar din & Johnson, 1979 9 Fledge per 8.4 Uni ted States/NS Bel lrose, 1976 Successf ul Nest
Age at 1 yr Uni ted States/NS Krapu & Dot y, 1979 Sexual Maturity
Annual A M 27.2 eastern-cent ral flyway/NS Bellrose, 1976 Mortality A F 38.2 Rates (percent ) A M fall 40.1 ± 3.1 SE 22 - 51 western mid-At lant ic/NS Chu & Hest beck, 1989
J M fall 41.1 ± 7.2 SE 31 - 59 1971 to 1985 A F all 49.9 ± 3.3 SE 20 - 72 J F all 48.8 ± 6.0 SE 15 - 68
A M fall 39.0 ± 2.3 SE 9 - 60 nor theastern Uni ted Chu & Hest beck, 1989 J M fall 48.1 ± 5.3 SE 7 - 69 States/NS A F all 51.5 ± 1.9 SE 33 - 64 1971 to 1985 J F all 56.8 ± 3.2 SE 38 - 68
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating May CA, UT, MT, SD, NY, VT Bel lrose, 1976 early Apr il early May mid-July sout h cent ral N Dakot a Krapu & Dot y, 1979
Hatching June NW Territo ry, Canada Toft e t al., 1984
ff
ff
2-46 M
allard

�
Mallard Duck ( Anas platy rhy nchos )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Molt spr ing December March Missi ssi ppi Valley Fredr ickson & Hei tmeyer, 1988 fall mid-Sept. Novem ber
Migration spr ing mid-March mid-May arrive nor th cent ral US Johnson et al., 1987 fall mid-October Novem ber leave nor thern US Palmer, 1976
1 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Nelson and M artin (1953).2 Est imated dai ly existence ener gy at 0�C.3 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Nelson and M artin (1953).4 See Chapt ers 3 and 4 f or methods of est imating f ood i ngest ion r ates f rom free-living m etabol ic rate and di etary com posi tion.5 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Nelson and M artin (1953).6 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Nelson and M artin (1953).7 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Nelson and Martin
(1953). 8 Cited in Palmer (1976). 9 Cited in Johnson et al. (1987).
2-47 M
allard

References (including A ppendix)
Allen, A. W. (1987) Habitat su itabilit y inde x mode ls : malla rd (wint er habitat, lower Mississippi V alley). U.S. Fish Wildl. Serv. Biol. Rep. No. 82(10.132).
Bellrose, F. C. (1976) Ducks, geese, and swan s of North Amer ica. Harrisburg, PA: The Stack pole Co.; pp. 229-243.
Bellrose, F. C.; Hawki ns, A. S. (1947) (ci ted in Palmer , 1976). Auk 64: 422-430.
Bent, A. C. (1923) Life histo ries of North Amer ican wild f owl. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 126.
Brownie, C.; Anderson, D. R.; Burnham, K. P.; et al . (1978) Stati sti cal inference fr om band recovery data--a handbook . U.S. Fish Wildl. Serv. Resour . Publ. 131.
Brownie, C.; Anderson, D. R.; Burnham, K. P.; et al . (1985) Stati sti cal inference fr om band recovery data--a handbook . U.S. Fish Wildl. Serv. Resour . Publ. 156.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chu, D. S.; Hestb eck, J. B. (1989) Temporal and geographic esti mates o f su rvival and recovery rates fo r the mal lard, 1950 through 1985. Washingt on, DC: U.S. Fish Wildl. Serv. Tech. Rep. No. 20.
Chur a, N. J. (1961) Food availa bilit y and pr eference of juvenile malla rds. Trans. N. Am. Wildl. Nat. Resour . Conf . 26: 121-134.
Coult er, M. W.; Mille r, W. R. (1968) Nest ing biology of black d ucks an d mal lards in northern New Engla nd. Vermont Fish and Game Dept. Bull. 68-2.
Cowardin, L. M.; Johns on, D. H. (1979) Mathemati cs an d mal lard management. J. Wildl. Manage. 43: 18-35.
Delnicki , D.; Reinecke, K. J. (1986) Mid-wint er food us e and body weight s of malla rds and wood ducks i n Missi ss ippi. J. Wildl. Manage. 50: 43-51.
Dillon, O. W. (1959) Food ha bit s of wild m allard ducks i n three Louis iana parishes. Trans. Nor th Am. Wildl. Nat. Resour . Conf . 24: 374-382.
Doty, H. A. (1975) Renest ing a nd second br oods of wild m allards. Wils on Bull. 87: 115.
Duebbert, H. F.; Kantrud, H. A. (1974) Upland duc k nesting r elated to land use and predator reduc tion. J. Wildl. Manage. 38: 257-265.
2-48 Mallard

Duebbert, H. F.; Lokemo en, J. T. (1976) Duck n est ing in f ields of undis turbed grass-l egume cover . J. Wildl. Manage. 40: 39-49.
Dwyer , T. J.; Krapu, G. L.; Janke, D. M. (1979) Use of prairie pothole habitat b y breeding malla rds. J. Wildl. Manage. 43: 526-531.
Dzubin, A. (1955) Some evi dences o f home range in water fowl. Trans. North Am. Wildl. Nat. Resour . Conf . 20: 278-298.
Eldr idge , J. L.; Krapu, G. L. (1988) The influence of diet quality o n clutch size and laying patter n in mal lards. Auk 105: 102-110.
Fredrickso n, L. H.; Heitmeyer , M. E. (1988) Water fowl use of fo rested wetl ands of th e southern United States: an overview. In: Weller, M. W., ed. Water fowl in winter . Minne apolis , MN: Univ ersi ty of Minne sota Press; pp. 307-323.
Fulle r, R. W. (1953) Studie s in t he lif e his tory and ecology of th e Amer ican pint ail, Anas acuta tz itzihoa (Vieillot ), in Ut ah [master 's th esis]. Loga n, UT: Utah State Agricult ural Colle ge.
Gilm er, D. S.; Ball, I. J.; Cowardin, L. M.; et al. (1975) Habitat u se and home range of malla rds br eeding in Minne sota. J. Wildl. Manage. 39: 781-789.
Girard, G. L. (1941) The mal lard: its man agement in wester n Mont ana. J. Wildl. Manage. 5: 233-259.
Gollop, J. B.; Marshall, W. H. (1954) A guide to aging duc k br oods in t he field (mimeo). MS: Mississippi Fl yway Counc il T ech. Sect.
Goodman, D. C.; Fisher, H. I. (1962) Func tiona l anatomy of th e feeding a pparatus in water fowl Aves: A natidae. Carbonda le, IL: Sout hern Illinois Univ ersi ty Press.
Heitmeyer , M. E. (1985) Winter ing s trategies of femal e mal lards related to dynamics o f low land ha rdwood wetl ands in the uppe r Mississippi De lta [Ph.D. disser tat ion] . Colum bia, MO: Univ ersi ty of Missour i.
Heitmeyer , M. E. (1988a) Body co mpos it ion of femal e mal lards in winter in relation t o annua l cycl e events. Condor 90: 669-680.
Heitmeyer , M. E. (1988b) Protein costs o f th e prebasic mo lt of femal e mal lards. Condor 90: 263-266.
Heitmeyer , M. E.; Vohs, P. A. (1984) Distr ibut ion a nd ha bitat u se of water fowl winter ing in Oklahom a. J. Wildl. Manage. 48: 51-62.
Johns on, D. H.; Grier, J. W. (1988) Determinants o f breeding dis tr ibut ions of ducks. Wildl. Monogr . 100: 1-37.
2-49 Mallard

Johns on, D. H.; Sparling, D. W.; Cowardin, L. M. (1987) A model of th e producti vi ty o f th e malla rd duc k. Ecol. Model. 38: 257-275.
Johns on, M. A.; Hinz, T. C.; Kuc k, T. L. (1988) Duck n est su ccess an d predators in North Dakota, South Dakota, and Montana: The Central Flyway Stu dy. In: Uresk, D. W.; Schenbeck, G. L.; Cefki n, R., tech . coords. Eight h Great Plains wildlif e damage cont rol w orkshop pr oceedings ; April 28-30, 1987; Rapid Cit y, Sout h Dakota. For t Collins , CO: U.S. Departmen t of Agriculture, Forest Ser vice, Rocky Mount ain For est and Range Experiment Stat ion; pp. 125-133.
Jorde, D. G.; Krapu, G. L.; Crawfo rd, R. D. (1983) Feeding e cology of malla rds wint ering in Nebraska. J. Wildl. Manage. 47: 1044-1053.
Kantrud, H. A.; Stewar t, R. E. (1977) Use of natural basin wetl ands by breeding water fowl in Nor th Dakota. J. Wildl. Manage. 41: 243-253.
Kir by, R. E.; Cowardin, L. M. (1986) Spring a nd s ummer survival of femal e mal lards fr om nor thcentral Minne sota. J. Wildl. Manage. 50: 38-43.
Kir by, R. E.; Riechmann, J. H.; Cowardin, L. M. (1985) Home range and habitat u se of forest-d welling m allards in Minne sota. Wils on Bull. 97: 215-219.
Klett, A. T.; Johns on, D. H. (1982) Variabilit y in n est su rvival rates an d implic ations to nesting s tudie s. Auk 99: 77-81.
Klett, A. T.; Shaffer , T. L.; Johns on, D. H. (1988) Duck n est su ccess i n the prairie pothole region. J. Wildl. Manage. 52: 431-440.
Krapu, G. L. (1981) The role of nutr ient reserves i n mal lard reproduc tion. Auk 98: 29-38.
Krapu, G. L.; Doty, H. A. (1979) Age-related aspects o f mal lard reproduc tion. Wildf owl 30: 35-39.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lee, F. B.; Jessen , R. L.; Ordal, N. J.; et al . (1964) In: Moyle, J. B., ed. Ducks an d land use in Minne sota. Minn. Dept . Cons erv. Tech. Bull. 8.
Lokemo en, J. T.; Schnaderbeck, R. W.; Woodward, R. O. (1988) Increasing water fowl produc tion on point s and is lands by r educ ing mammal ian predation. In: Uresk, D. W.; Schenbeck, G. L.; Cefki n, R., tech . coord. Eight h Great Plains wildlif e damage cont rol w orkshop pr oceedings ; April 28-30, 1987; Rapid Cit y, Sout h Dakota. For t Collins , CO: U.S. Departmen t of Agriculture, Forest Ser vice, Rocky Mount ain For est and Range Experiment Stat ion; pp. 146-148.
2-50 Mallard

Lokemo en, J. T.; Duebbert, H. F.; Sharp, D. E. (1990a) Homing a nd r eproduc tive habit s of malla rds, gadwalls , and blue -winge d teal . Wildl. Monogr . 106: 1-28.
Lokemo en, J. T.; Johns on, D. H.; Sharp, D. E. (1990b) Weight s of wild m allard Anas platyr hynchos, gadwall A. str epera, and blue -winge d teal A. disco rs dur ing t he breeding seaso n. Wildf owl 41: 122-130.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York, NY: McGraw-Hill Book Company, Inc.
McAtee, W. L. (1918) Food ha bit s of th e malla rd ducks o f th e United States. U.S. Dept. Agric. Bull. 720.
McEwan , E. H.; Koelink , A. F. (1973) The heat produc tion of oile d mallards and scaup. Can. J. Zool. 51: 27-31.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Nelson, A. L.; Martin, A. C. (1953) Gamebird weight s. J. Wildl. Manage. 17: 36-42.
Palmer , R. S. (1976) Handbook of Nor th Amer ican bir ds: v. 1. New Haven, CT: Yale Universi ty Pr ess.
Perret, N. G. (1962) The spring a nd s ummer foods of th e common m allard (Anas platyr hynchos platyr hynchos L.) in south central Manitoba [master 's th esis]. Vancouv er, BC: Univ ersi ty of Br it ish Colum bia.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Pospahala, R. S.; Anderson, D. R.; Henny, C. J. (1974) Breeding ha bi tat condit ions , size of the breeding popula tions , and pr oduc tion indi ces i n popula tion e cology of th e malla rd. Bur eau of Spor t Fis h. and Wildl. , Res. Publ. 115, U.S. GPO Stock No. 2410-00387.
Ringe lman, J. K.; Eddl eman, W. R.; Mille r, H. W. (1989) High pla ins r eservoirs and sloughs . In: Smith, L. M.; Pederson, R. L.; Kaminski , R. M., eds. Habitat man agement fo r wint ering water fowl in North Amer ica. Lubboc k, TX: Texas T ech Universi ty Pr ess; pp. 311-340.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
2-51 Mallard

Rutherford, W. H. (1966) Chronology of water fowl migration in Color ado. Colo. Div . Wildl. ; Game Inf. Leafl . No. 42.
Simpson, S. G. (1988) Duck n est su ccess o n South Dakota game produc tion a reas. In: Uresk, D. W.; Schenbeck, G. L.; Cefki n, R., tech . coords. Eight h Great Plains wildlif e damage control workshop pr oceedings ; April 28-30, 1987; Rapid Cit y, Sout h Dakota. For t Collins , CO: U.S. Departmen t of Agriculture, Forest Ser vice, Rocky Mount ain Forest an d Range Experiment Stat ion; pp. 140-145.
Stoudt , J. H. (1944) Food pr eferences o f mal lards on the Chippewa N ationa l For est, Minne sota. J. Wildl. Manage 8: 100-112.
Swanson, G. A.; Meyer , M. I. (1973) The role of inver tebrates i n the feeding e cology of Anatina e dur ing t he breeding seaso n. In: The Water fowl Habitat Management Symp osium at Moncto n, New Brunswi ck, Canada; July 30 - August 1, 1973; The Atlantic Water fowl Counc il; pp. 143-180.
Swanson, G. A.; Krapu, G. L.; Serie, J. R. (1979) Foods of laying femal e dabbling du cks o n the breeding gr ounds . In: Book hout , T. A., ed. Water fowl and wetl ands--an integrated review: proceedings of 1977 symp osium. Madison, WI: The Wildlif e Society, NC Sect. ; pp. 47-57.
Swanson, G. A.; Meyer , M. I.; Adomaitis, V.A. (1985) Foods cons umed by breeding m allards on wetl ands of so uth-central North Dakota. J. Wildl. Manage. 49: 197-203.
Toft, C. A.; Trauger, D. L.; Murdy, H. W. (1984) Seasonal decline in br ood s izes of symp atr ic water fowl (Anas and Athya, Anatidae) and a propos ed evolut iona ry expla nation. J. Anim. Ecol. 53: 75-92.
USFWS. (1991) 1991 Statu s of water fowl & fal l flight fo recast. Laurel, MD: U.S. Fish Wildl. Serv., Offi ce of Migr atory Bir d Management.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Whyte, R. J.; Bole n, E. G. (1984) Impact o f wi nter str ess o n mal lard body co mpos it ion. Condor 86: 477-482.
2-52 Mallard

2.1.4. Lesser Scaup (bay ducks)
Order Anserif ormes, Family Anatidae. Bay ducks ar e adapted for diving a nd character isti cal ly need a running star t to become airborne because th eir legs are located further back o n their body th an on ot her ducks. They breed at mi d to high la ti tude s and winter in flocks o n large water bodie s and in pr otected coastal bays an d river mouths. Bay ducks d ive fo r their food, and t heir die t is omniv orous (i .e., bot h pla nt and a nim al matter ) and depends on the seaso nal and regiona l abunda nce of food r esour ces. Because of th eir food ha bit s, bay du cks p refer deeper, more permanent ponds th an dabbling duc ks (Bellrose, 1976). The sexes var y in color ation, and di ffer ent bay duck sp ecies range in lengt h f rom 42 to 53 cm (bill t ip t o tail t ip) .
Selected species
The lesser scau p (Ayth ya affi nis) is one of th e most abunda nt Nor th Amer ican ducks (A llen, 1986). They breed principally th roughout wester n Canada and Alaska, although t heir breeding r ange exten ds into the wester n United States as far south as Color ado and Ohio. Lesser scau p winter in the United States i n the Missi ss ippi f lyway an d the Atlantic fl yway (B ellrose, 1976). They also winter along a ll c oastal areas in the southern states an d into Mexico (Nationa l Geogr aphic So ciety, 1987).
Body size. The lesser scau p averages 42 cm fr om bill t ip t o tail t ip. Males are larger and m ore color ful t han the brown femal es (Bellrose, 1976; see tab le). Follow ing t heir pos tbreeding m olt , scau ps increase th eir fat r eserves i n preparation f or migr ation ( Austin and Fredrickso n, 1987; see tab le).
Habitat . Lesser scau p are found on la rge lakes an d bays d uring t he fall a nd w int er and are common on s malle r bodie s of water (e.g., ponds ) dur ing t he spring. They br eed in the prairie pot hole s region, most often on permanent or semi permanent wetl ands of 0.85 to 2.0 ha wi th trees and shrubs bordering a t least h alf of th e shorelines (Bellrose, 1976; Smith, 1971, ci ted in Allen, 1986). Primary brood ha bi tat i s ch aracter ized by permanent wetl ands dominated by emer gent veg etat ion (Smith, 1971, ci ted in Allen, 1986). In a stu dy of ducks wi nter ing in S out h Carolina , Bergan and Smith (1989) found l esser scau p would forage primar ily in areas wi th submergent veg etat ion but al so in areas of emer gent vegetat ion, shallow open water , and floating- leaved vegetat ion. They found s ome differ ences i n foraging ha bi tat u se by seaso n and between males and femal es. In particular, femal es ten ded to use mo re shallow habitats th an males, and males prefer red open water in late fal l (Bergan and Smith, 1989).
Food ha bi ts . Most popula tions of l esser scau p consume primar ily aquatic inver tebrates, both from th e water colum n and f rom th e surfaces o f aquatic veg etat ion a nd other substr ates (T ome and Wrubleski , 1988; Bartonek and Hickey, 1969). Commo n prey inc lude sn ails , clams, scu ds (amphipods ), midge s, chir onom ids , and l eeches (see tab le). Scaup are omnivorous, however , and the percentage of plant mater ials (almost excl usively seeds) in the diet var ies seaso nally as th e avai labilit y of di ffer ent foods ch anges (Afton e t al., 1991; Dirsch l, 1969; Rogers and Kor schge n, 1966). When seeds are local ly abunda nt , they may b e consumed in large quanti ties (Dirsch l, 1969). Breeding femal es and ducklings
2-53 Lesser Scaup

eat mo stl y aquatic inver tebrates (Sugde n, 1973). Young duc klings feed primar ily on water -colum n inv ertebrates (e.g., phantom midges, clam sh rimps, water mites), whereas olde r duc klings fo rage mainly on bot tom-dwelling inv ertebrates (e.g., scu ds or amphipods , dragonf lies, caddis fli es) (Bartonek and Murdy, 1970). During t he wint er, there are no signif icant di ffer ences i n diet between juveniles and adults o r between males and femal es (Afto n et al ., 1991).
Mol t. Nonbr eeding a nd pos tbreeding m ales and nonbr eeding femal es generally leave th e breeding gr ounds in J une to molt on l akes. However , some mal es co mplete th eir molt on t he breeding gr ounds (T rauger, 1971, ci ted in Bellrose, 1976). Large flocks o f molt ing bir ds become flight less d uring t he wing m olt phase, which begins in J uly and is usually co mplete by late Augus t (McKnight and Bus s, 1962).
Migration. The axis of th e main m igr ation c orridor exten ds fr om th e breeding grounds on t he Yukon Fl ats, Alaska, to winter ing a reas in Flor ida (B ellr ose, 1976). Most scau p winter in the United States, wi th the greatest n umbers in the Missi ssippi f lyway an d the Atlantic fl yway. They star t to arrive at th eir winter ing a reas in mid-Octo ber (Bellrose, 1976). The timing of nor thward migr ation in t he spring v aries from February to May (Bellrose, 1976). Before migration, scaup ga in w eight by inc reasing t heir body fat co ntent (Austi n and Fredrickso n, 1987).
Breeding acti vi ties and social organization. Scaup build n ests o n the ground among t all gr asses, shrubs, or forbs wh ere plant heights r ange from 20 to 60 cm (Hines, 1977). Nests can be located along t he edge of shor eline s to upla nd areas (Bellrose, 1976). Cour tship a nd pa ir bonds star t to form on the winter ing gr ounds , and pa irs typic ally r emain toge ther fo r only one seaso n. Males do not remain long after incubation c ommen ces (Trauger, 1971, ci ted in Bellrose, 1976). The femal e and her brood l eave th e vicinity o f th e nest sh ortly after the ducklings h ave hatch ed. Most b roods ar e on t heir own by 4 to 5 weeks o f age (Gehrman, 1951, ci ted in Bellrose, 1976) and fledge between 7 and 9 weeks o f age (Bellrose, 1976; Light body and A nkney, 1984). Females of th is sp ecies often lay eggs in other lesser scau p nests (n est p arasi tism), which can result in large compound c lut ches of lesser scau p eggs in a single n est (Hines, 1977). Hines (1977) also found t hat mixing of broods was co mmo n in Saskatch ewan; by Augus t, groups of 15 to 40 duc klings led by two to three hens would be co mmon. Female lesser scau p also occas iona lly lay eggs in t he nests o f other ducks (e. g., gadwal l; Hines, 1977).
Home range and r esour ces. Relatively small ne sting t erritories and la rge highly overlapping f oraging r anges are character isti c of lesser scau p (Hammel , 1973, ci ted in Allen, 1986). Several pairs can nest i n close proximity wi thout aggr ession, each defending only a smal l area immed iately su rrounding t he nest (B ellrose, 1976; Vermeer , 1970). In Manitoba, Hammel (1973) esti mated the mean minim um fo raging hom e range to be 89 ± 6.5 ha. Init ial areas occupie d by pairs us ually cont ain s tumps, logs , boulde rs, or beaches as loafing s ites, but later lesser scau p rely so lely on open water (Gehrman, 1951, ci ted in Bellrose, 1976).
Popul ation de nsity . In wint er, loc al dens it ies of scaup c an be ver y high, as large flocks fl oat on favo red feeding a reas (Bellrose, 1976). In summer , the densi ty o f breeding
2-54 Lesser Scaup

�
�
�
�
pairs increases wi th the permanence and size of th e ponds (K antrud and Stewar t, 1977; see tab le).
Popul ation dy namics. In some popula tions , many year ling a nd s ome 2-year -olds do not breed; the propor tion br eeding t ends to inc rease wi th improving water and habitat condit ions (A fton, 1984; McKnight and Bus s, 1962). In a 4-year stu dy in Manitoba, Afto n (1984) found t hat, on aver age, 30 percent of 1-year -olds and 1 0 percent of 2-year -olds , did not breed. Clut ch size and r eproduc tive perfor mance of adult femal es generally increase wi th age (Afto n, 1984). Most n est fa ilur es are due to predation ( e.g., by mink , raccoons , red fox), and scau p often attemp t to renest i f th e init ial nest fai ls (Afto n, 1984; Bellrose, 1976). Annua l mortalit y for juv enile s is highe r th an that for adult s, and a dult femal e mortal ity exceed s adult mal e mortalit y (Smith, 1963; see tab le).
Simi lar species (fr om general references)
� The redhead (Ayth ya amer icana), a larger bay duck (48 cm), breeds on lakes and ponds in t he nor thwester n United States an d in midwester n Canada. They wi nter in coastal areas and the southern United States an d Mexico. In summer , adult femal e and juvenile r edheads cons ume predom inant ly anim al matter (e.g., caddis flie s, midge s, water fleas, snails), while mal es inc lude more plant mater ials in their diet.
� The canvasb ack (Ayth ya val isineria) is th e largest b ay duck (53 cm). They are commo n on lakes an d ponds in t he nor thern Unit ed States an d southern Canada dur ing t he breeding seaso n and along c oastal areas of th e Unit ed States d uring w int er. Studie s dur ing t he wint er in Nor th and Sout h Carolina have found v arying di ets fo r canvasb acks, consuming m ostl y anim al matter (e.g., clams); others eat only veg etation. In summer , adult femal e and juv enile can vasb acks eat p redominantly animal mater ial (e.g., caddis flie s, snails, mayfl ies, midges), whereas adult mal es may eat p redominantly vegetable mater ial, particularly tu bers of Potamo geton.
� The ring- necked duck (Ayth ya co llaris) is si mi lar in size (43 cm) to the lesser scau p and prefers fr eshwater wetl ands. They are commonly seen on woodla nd l akes an d ponds , but in w int er also use sout hern coastal marshes. Dur ing t he wint er, ring- necked ducks eat mo stl y plant mater ials (81 percent) and a var iety o f animal matter (19 percent).
� The greater scau p (Aythya marila ) (46 cm) i s co mmo n in coastal areas and the Great Lakes d uring w int er. They are omniv orous , eating 5 0 to 99 percent animal matter and the remainde r plant foods dur ing t he wint er.
General references
Allen (1986); Bartonek and Hickey (1969); Bellrose (1976); Nationa l Geogr aphic Society (1987); Perry and Uhler (1982).
2-55 Lesser Scaup

Lesser Scaup ( Aythya affinis )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight F preflight less 688 Mani toba, Canada Aust in & Fredr ickson, 1987 1 (g) F flight less 647
F post flight less 693 F migratory 842
F 770 up t o 950 Uni ted States Nelson & M artin, 1953 M 860 up t o 1,100
Adul t Body Fat F preflight less 50.7 7.4%) Mani toba, Canada Aust in & Fredr ickson, 1987 1 (grams lip id : F flig htless 37.2 (5.7%) % of total body F post flight less 46.5 6.7%) weight ) F migratory 188.1 (22.3%)
Duckl ing age in weeks growt h in g/day (final body weight ) Utah or Canada Sugden & Har ris, 1972 2 Growth Rate 0-3 6.9 (190 g)
3-6 (485 g) 6-9 5 (516 g) 9-12 2 (542 g)
Metabol ic Rate A F basal 83 est imated 3 (kcal /kg-day) A M basal 81
A B r est ing Canada McEwan & Koel ink, 1973 20 to 30��C
A F free-living 211 (102 - 457) est imated 4 A M free-living (99 - 445)
90
216
Food Ingest ion juveni les, dry matter intake/ Saskat chewan/ capt ive: Sugden & Har ris, 1972 5 Rate (g/g-day) bot h sexes: wet body weight reared in large brooder
1 - 5 weeks 0.162 and i n out door pens 6 - 12 weeks 0.077
Water A F 0.064 est imated 6 Ingest ion A M 0.062 Rate (g/g-day)
Inhal ation A F 0.34 est imated 7 Rate (m /day) A M 0.363
(
(
14 1.1.
2-56 L
esser Scau
p

Lesser Scaup ( Aythya affinis )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Surface Ar ea A F 842 est imated 8 (cm ) A M 9062
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
(animal) (60.9) Loui siana/lakes, marshes Afton et al., 1991 midges 45.9 snails 7.7 (% dry weight; grass shr imp 7.3 esophageal & pr ovent ricular
(plant - seeds) (36.1) cont ents) bul rush 36.0
(plant - veget ative) (3.0) green algae 2.3
juveni les onl y: Northwest Territories/lake Bartonek & M urdy, 1970 (animal) (100) (100) scuds 1 ± 1 57 ± 9 (% wet vol ume ± SE; phant om midges 54 ± 8 1 ± 1 esophageal cont ents) clam shr imps 30 ± 8 2 ± 2 dragon/ damsel flies - 17 ± 8 water bugs 4 ± 3 11 ± 7 water mites 8 ± 3 -caddis flie s - 6 ± 5 water beetles 1 ± 1 4 ± 3 mayflies 2 ± 1 -
(plants) (trace) (trace)
2-57 L
esser Scau
p
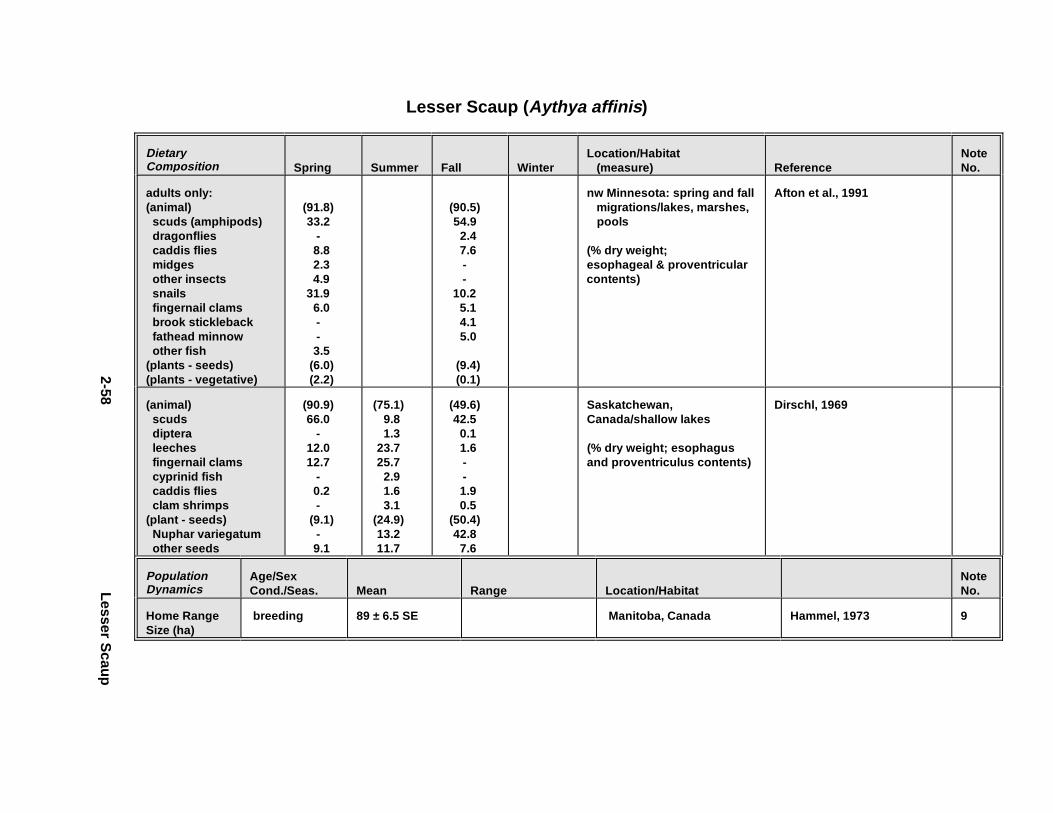
Lesser Scaup ( Aythya affinis )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
adults only: nw Minnesota: sprin g and fall Afto n et al., 1991 (animal) (91.8) (90.5) migrations/ lakes, marshes, scuds ( amphi pods) 33.2 54.9 pool s dragonf lies - 2.4 caddis flie s 8.8 7.6 (% dry weight; midges 2.3 - esophageal & pr ovent ricular other insect s 4.9 - cont ents) snai ls 31.9 10.2 fin gernail c lams 6.0 5.1 brook st ickl eback - 4.1 fathead minnow - 5.0 other fis h 3.5
(plants - seeds) (6.0) (9.4) (plants - veget ative) (2.2) (0.1)
(animal) (90.9) (75.1) (49.6) Saskat chew an, Dirschl , 1969 scuds 66.0 9.8 42.5 Canada/shal low lakes diptera - 1.3 0.1 leeches 12.0 23.7 1.6 (% dry weight ; esophagus finger nail clams 12.7 25.7 - and pr ovent riculus cont ents) cyprin id fis h - 2.9 -caddis flie s 0.2 1.6 1.9 clam shr imps - 3.1 0.5
(plant - seeds) (9.1) (24.9) (50.4) Nuphar var iegatum - 13.2 42.8 other seeds 9.1 11.7 7.6
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home Range breeding 89 ± 6.5 SE Mani toba, Canada Hammel, 1973 9 Size (ha)
Range
2-58 L
esser Scau
p

Lesser Scaup ( Aythya affinis )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Popul ation A B seasonal North Dakot a/ Kantrud & St ewart, 1977 Densi ty wetland 0.029 prairie pot hol es (pairs/ha) A B per manent
wetland 0.061
A B i sland i n 28.9 13.1 - 58.5 Alberta, Canada/ islands i n Vermeer, 1970 10 lake lakes of parklands and
boreal forest
Clutch 9.47 ± 0.18 SE 7 - 12 Saskat chew an/marsh i sland Hines, 1977 Size
2nd y r female 10.0 ± 0.2 SE 8 - 12 Mani toba/ lake Afton, 1984 4th yr female 12.1 ± 0.2 SE 11 - 14
Clutches 1, but of ten NS/NS Afton, 1984 /Year renest if lo st
Days 24.8 21 - 27 NS/NS Vermeer, 1968 10 Incubat ion
Age at B 65 ± 0.91 SE Mani toba/capt ive Light body & Ankney , 1984 Fledgi ng (days)
Percent Nests 1st yr female 26.3 Mani toba/ lake Afton, 1984 Hatching 2nd y r female 22.2
3rd yr female 45.5
76 Saskat chew an/marsh i slands Hines, 1977
Percent up t o 20 days 67.5 ± 4.9 SE Mani toba/ lake Afton, 1984 Broods of age Surv iv ing
Age at First M most in 2nd y r NS/NS Palmer, 1976 Breeding F 1 - 2 years Mani toba/ lake Palmer, 1976; Af ton, 1984
Annual juveni les 68 - 71 NS/NS Smith, 1963 Morta lity Rates A males 38 - 52 (percent ) A females 49 - 60
Range
2-59 L
esser Scau
p

�
Lesser Scaup ( Aythya affinis )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Laying early June Mani toba, Canada Afton, 1984 early May early June early July Montana Ellig , 1955 10
Hatching early Jul y mid-Jul y early August NW Territory and Toft et al., 1984; Hines, 1977 Saskat chew an, Canada
Molt (fall) Jul y September Mani toba, Canada Aust in & Fredr ickson, 1987
Migration spr ing early February March - Apr il May depar ting Uni ted States Bel lrose, 1976
mid-Apr il arriving Mani toba, Canada Afton, 1984
fall September mid-Novem ber Paci fic f lyway (s OR, n CA) Gammonl ey & Hei tmeyer, 1990
mid-October mid- December arriving Uni ted States Novem ber
Bel lrose, 1976
1 Four stages of feather molt evaluated.
2-60 L
esser Scau
p
2 Duckl ings st opped gr owi ng at rate typical of wi ld bi rds around 6 weeks of age. By 12 weeks, they weighed appr oxi mately 200 g less t han typical of wild scaup.
3 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Nelson and M artin (1953). 4 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Nelson and M artin (1953). 5 Young duckl ings m aintained in 18 to 27�C brooder , then in out door pens wi th same temperature range. Metabol izable energy of amphi pods
(est imated to be 3.11 kcal /g dry wt), a typical scaup f ood, is similar to the com mercial diet used i n the exper iment (3.09 kcal /g dry wt). Ducklin gs stopped gr owi ng as r apidly as woul d wi ld duckl ings at about 6 weeks of age. For methods of est imating f ood i ngest ion r ates for adul t scaup, see Chapt ers 3 and 4.
6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Nelson and M artin (1953).7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Nelson and M artin (1953).8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Nelson and Martin (1953).9 Relatively small, h ighly overlapping, home ranges. Cited in Allen (1986).
10 Cited in Bel lrose (1976).

References (including A ppendix)
Afto n, A. D. (1984) Influence of age and time on reproducti ve performance of femal e lesser scau p. Auk 101: 255-265.
Afton, A. D.; Hier, R. H.; Paulus , S. L. (1991) Lesser scau p diets d uring m igr ation a nd w int er in t he Mississippi f lyway. Can. J. Zool. 69: 328-333.
Allen, A. W. (1986) Habitat su itabilit y inde x mode ls : lesser scau p. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.117).
Austi n, J. E.; Fredrickso n, L. H. (1987) Body and or gan mass an d body co mpos it ion of pos tbreeding femal e lesser scau p. Auk 104: 694-699.
Bartonek, J. C.; Hickey, J. J. (1969) Food ha bit s of can vasb acks, redheads, and lesser scaup in Ma nit oba. Condor 71: 280-290.
Bartonek, J. C.; Murdy, H. W. (1970) Summer foods of l esser scau p in subarcti c tai ga. Arcti c 23: 35-44.
Bellrose, F. C. (1976) Ducks, geese, and swan s of North Amer ica. Harrisburg, PA: The Stack pole Co.
Bergan, J. F.; Smith, L. M. (1989) Differ ential habitat u se by diving du cks wi nter ing in S out h Carolina . J. Wildl. Manage. 53: 1117-1126.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chabreck, R. H.; Takagi, T. (1985) Foods of l esser scau p in crayfi sh impoundm ents in Louis iana. Proc. Annu. Conf . Sout heast. Asso c. Fish Wildl. Agencies 39: 465-470.
Chappel, W. A.; Titman , R. D. (1983) Esti mat ing r eserve lipids in gr eater scau p (Ayth ya marila ) and lesser scau p (A. affi nis). Can. J. Zool. 61: 35-38.
Dirsch l, H. J. (1969) Foods of l esser scau p and blue -winge d teal in the Saskatch ewan River Delta. J. Wildl. Manage. 33: 77-87.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Ellig, L. J. (1955) Water fowl relations hips to Greenfields L ake, Teton Count y, Mont ana. Mont . Fish. and Game Comm. Tech. Bull. 1.
Gammonley, J. H.; Heitmeyer , M. E. (1990) Behavior, body condit ion, and f oods of buffl eheads and lesser scau ps during s pring m igr ation t hrough t he Klamath Basin, Calif ornia . Wils on Bull. 102: 672-683.
2-61 Lesser Scaup

Gehrman, K. H. (1951) An ecologic al study of th e lesser scau p duck (Athaya affi nis Eyto n) at West Medical Lake, Spokane Count y, Washingt on [master 's th esis]. Pullman, WA: Washingt on State Colle ge.
Gollop, J. B.; Marshall, W. H. (1954) A guide fo r aging duc k br oods in t he field (mimeo). MS: Mississippi Fl yway Counc il T ech. Sect.
Hammel , G. S. (1973) The ecology of th e lesser scau p (Athaya affi nis Eyton) in southwester n Manitoba [master 's th esis]. Guelph, Ontar io: Univ ersi ty of Gu elph.
Hines, J. E. (1977) Nest ing a nd br ood e cology of l esser scau p at Waterhen Marsh, Saskatch ewan. Can. Field-Nat. 91: 248-255.
Hoppe , R. T.; Smith, L. M.; Wester , D. B. (1986) Foods of wint ering div ing du cks i n South Carolina . J. Field Ornit hol. 57: 126-134.
Hunt , E. G.; Anderson, W. (1966) Renest ing of ducks at Mount ain Meadow s, Lassen Count y, Calif ornia . Calif . Fish and Game 52: 17-27.
Kantrud, H. A.; Stewar t, R. E. (1977) Use of natural basin wetl ands by breeding water fowl in Nor th Dakota. J. Wildl. Manage. 41: 243-253.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Light body , J. P.; Ankney, C. D. (1984) Seasonal influence on the str ategies of growth and development of can vasb ack an d lesser scau p ducklings . Auk 101: 121-133.
McEwan , E. H.; Koelink , A. F. (1973) The heat produc tion of oile d mallards and scaup. Can. J. Zool. 51: 27-31.
McKnight , D. E.; Bus s, I. O. (1962) Evidence of breeding in year ling femal e lesser scau p. J. Wildl. Manage. 26: 328-329.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nasser , J. R. (1982) Management of impoundm ents for cr ayfi sh and water fowl [master 's thesis]. Baton Rouge , LA: Louis iana State Universi ty.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
2-62 Lesser Scaup

Nelson, A. L.; Martin, A. C. (1953) Gamebird weight s. J. Wildl. Manage. 17: 36-42.
Palmer , R. S. (1976) Handbook of Nor th Amer ican bir ds: v. 2, 3. New Haven, CT: Yale Universi ty Pr ess.
Perry, M. C.; Uhler, F. M. (1982) Food ha bit s of div ing du cks i n the Carolina s. Proc. Annu. Conf . Sout heast. Asso c. Fish Wildl. Agencies 36: 492-504.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Rienecker , W. C.; Anderson, W. (1960) A water fowl nest ing s tudy on T ule L ake and Lower Klamath Nationa l Wildlif e Refuge s, 1957. Calif. Fish and Game 46: 481-506.
Ringe lman, J. K.; Eddl eman, W. R.; Mille r, H. W. (1989) High pla ins r eservoirs and sloughs . In: Smith, L. M.; Pederson, R. L.; Kaminski , R. M., eds. Habitat man agement fo r wint ering water fowl in North Amer ica. Lubboc k, TX: Texas T ech Universi ty Pr ess; pp. 311-340.
Rogers, J. P. (1962) The ecologic al effects o f drought on r eproduc tion of th e lesser scau p, Ayth ya affi nis (Eyto n) [Ph.D. disser tat ion] . Colum bia, MO: Univ ersi ty of Missour i.
Rogers, J. P. ; Kor schge n, L. J. (1966) Foods of l esser scau ps on breeding, migr ation, and wint ering a reas. J. Wildl. Manage. 30: 258-264.
Rowinski , L. J. (1958) A review of water fowl investi gations and a co mparison of aer ial and ground c ensusing of water fowl at Mint o Flats, Alaska. Mimeogr. Rep.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Rutherford, W. H. (1966) Chronology of water fowl migration in Color ado. Colo. Div . Wildl. ; Game Inf. Leafl . No. 42.
Siegfr ied, W. R. (1974) Time budge t of behavior among l esser scau ps on Delta Marsh. J. Wildl. Manage. 38: 708-713.
Smith, A. G. (1971) Ecologic al facto rs affect ing water fowl produc tion in A lberta parklands . U.S. Fish Wildl. Serv. Resour . Publ. 92.
Smith, R. I. (1963) Lesser scau p and ring- necked duck s hoot ing pr essu re and mortal ity rates. Bur. Sport Fish & Wildl. Adm. Rep. 20.
Sugde n, L. G. (1973) Feeding e cology of pint ail, gadwall, Amer ican widge on and l esser scaup duc klings in Sout hern Alberta. Can. Wildl. Serv. Rep. Ser. No. 24.
Sugde n, L. G.; Harris, L. E. (1972) Energy requir ements an d growth of cap tive lesser scau p. Poult ry Sci . 51: 625-633.
2-63 Lesser Scaup

Swanson, G. A.; Krapu, G. L.; Bartonek, J. C.; et al . (1974) Advantages in math emati cal ly weight ing water fowl food ha bit s data. J. Wildl. Manage. 38: 302-307.
Toft, C. A.; Trauger, D. L.; Murdy, H. W. (1984) Seasonal decline in br ood s izes of symp atr ic water fowl (Anas and Athya, Anatidae) and a propos ed evolut iona ry expla nation. J. Anim. Ecol. 53: 75-92.
Tome, M. W.; Wrubleski , D. A. (1988) Underwater foraging be havior of can vasb acks, lesser scaups , and r uddy ducks. Condor 90: 168-172.
Townsend, G. H. (1966) A stu dy of water fowl nest ing on t he Saskatch ewan River delta. Can. Field. Nat. 80: 74-88.
Trauger, D. L. (1971) Popula tion e cology of l esser scau p (Athaya affi nis) in subarcti c tai ga [Ph.D. disser tat ion] . Ames, IA: Iowa State Universi ty.
Vermeer , K. (1968) Ecologic al aspects o f ducks n est ing in high de nsit ies among la rids . Wils on Bull. 80: 78-83.
Vermeer , K. (1970) Some asp ects o f th e nest ing of ducks o n islands in Lake Newel l, Alberta. J. Wildl. Manage. 34: 126-129.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
2-64 Lesser Scaup

2.1.5. Osprey (Pandion haliaetus )
Order Falconi formes, Family Acci pitr idae. The only Nor th Amer ican memb er of th e subf amily Pandionina e, these large birds of prey have long na rrow wings , a sharp hook ed bill, and pow erful t alons . Osprey are found ne ar fr eshwater or sal twater , and their diet is almost co mpletely restr icted to fish. They are adapted for hover ing ov er th e water and dive feet-fi rst, seizing f ish wi th their talons (Robbins et al., 1983). Once ver y rare owing t o DDT accu mulation in t heir food (1950's to early 1970's), osprey now are increasing in numbers. In the United States, there are five regiona l popula tions of osprey (in or der of abunda nce): Atlantic co ast, Flor ida and gulf co ast, Paci fic Northwest, wester n inter ior, and Great Lakes (Henny, 1983). In North Amer ica, osprey breed primar ily in a wide band from coast to coast acr oss Canada and the southern half of Alaska, where they are not restr icted to coastal and Great Lake areas as th ey are in the United States. However , osprey are reported from al l States d uring t he fall a nd s pring m igr ations (H enny , 1986).
Body size. The var ious subs pecies of osprey around t he world di ffer in size, and in general femal es are heavier than males (Poole , 1989a; see tab le). Osprey found in t he United States ar e considered to be of th e subspecies carolinenesis and average 56 cm from bill t ip t o tail t ip (Robbins et al., 1983) and weigh b etween 1.2 and 1.9 kg (see tab le).
Habitat . In the United States, the majority o f osprey popula tions ar e asso ciated wi th mar ine en vironm ents, but large inla nd r ivers, lakes, and reservoirs also may s uppor t osprey (Henny, 1986, 1988b). Good nesting s ites i n proximity to open, shallow water and a plent iful s upply of fi sh are the pr imary resour ces r equir ed for osprey success (P oole , 1989a). The tops of isolated and often dead trees and man-made str uctu res are prefer red nesting s ites. Osprey often nest i n colonie s (Poole , 1989a).
Food ha bi ts . Osprey are almost co mpletely pisci vorous, although t hey have been observed on occas ion taking ot her prey inc luding bir ds, frogs , and c rustacean s (Brown and Amadon, 1968). Their prey preferences ch ange seaso nally wi th the abunda nce of th e local fish (Edwards, 1988; Greene et al ., 1983). Osprey occas iona lly will pic k up de ad fish but only if fr esh (Bent, 1937). Osprey are most su ccessfu l catc hing s pecies of slow -moving f ish that eat benthic organisms i n shallow water s and fish that remain near the water 's su rface (Poole , 1989a). Osprey co nsume al l parts o f a fi sh excep t th e larger bone s; later , bone s and ot her undig estible parts are ejected in fecal pellets (B ent, 1937).
Mol t. Juvenile plum age is fully d evelope d by fl edging a t about 60 days o f age (Henny, 1988b). Juvenile s unde rgo a gradual molt to adult plum age at appr ox imatel y 18 mont hs of ag e (Brown and Amadon, 1968). For adults, the basic mo lt takes p lace in two phases; the fi rst p hase occu rs primar ily on the winter ing gr ounds pr ior to spring m igr ation. Completion of th e molt occurs in t he summer range prior to fal l migration ( Henny , 1988b).
Migration. Osprey are year -round r eside nts in t he most sout hern parts of th eir range (e.g., sout h Flor ida, Mexico) but ar e migr atory ov er th e rest o f th eir range in the United States an d Canada (Poole , 1989a). Studies of banded osprey have sh own that th e fall m igr ation be gins in l ate August i n the north temp erate zone, wi th adult s and juv enile s
2-65 Osprey

from th e easter n and central United States co mprising a broad f ront fl ying s out h and t hen directl y acr oss o pen ocean to their winter ing gr ounds in Central and Sout h Amer ica (Poole , 1989a). Spring m igr ation a ppears to follow th e same r outes wi th birds reaching, for examp le, the Chesapeake Bay area in mid-March (Reese, 1977) and Minne sota by th e fi rst half of April (Dunstan , 1973; Henny and Van Velzen, 1972). The majority o f mi grating osprey appear to follow th e coastline , perhaps because th ey co me fr om co astal colonie s or because th e coast o ffer s abunda nt food ( Poole , 1989a). After their fi rst mi gration s out h, juv enile s remain in t heir wint ering gr ounds fo r about a year and a half, returning nor th to the breeding gr ounds as 2-year -olds (H enny and V an Velzen, 1972).
Breeding acti vi ties and social organization. Nonm igr atory (i.e., year -round r eside nt) popula tions breed dur ing t he wint er; whereas mi gratory popula tions breed dur ing t he summer (Poole , 1989a). Monogamy i s th e general rule for osprey; breeding pa irs remain together and return to the same n est si te year after year (Fernandez and Fernandez, 1977; Henny, 1988b). Colonie s of osprey oc cur in areas su ch as islands, reservoirs, or lakes th at offer secu re nest ing s ites an d abunda nt food ( Henny , 1986), but mo st o sprey are solitary nester s, often separated from other nests b y ten s to hundr eds of kilo meter s (Poole , 1989a). The femal e performs mo st o f th e incubation a nd r elies compl etely on the male for food from just after mat ing unt il t he young h ave fl edged (Poole , 1989a). Van Daele and Van Daele (1982) found t hat os preys at su ccessfu l nests i ncubated 99.5 to 100 percent of th e daylight hour s; dis turbance of th e nest d uring t his ti me can kill t he eggs if th e adult s are kept from r eturning t o the nest fo r some ti me. After hatching, the femal e is in constan t atten dance at th e nest fo r the fi rst 35 d ays b ut may p erch nearby at i ntervals after that (Henny, 1988b). The femal e distr ibutes th e food de liv ered by th e male by bit ing of f pi eces to feed to the young ( Poole , 1989a). By 30 days, the nest lings h ave reached 70 to 80 percent of th eir adult weight and be gin t o be acti ve in the nest (Poole , 1989a). The young fledge by age 60 to 65 days i n nonm igr atory popula tions and by about 50 to 55 days i n migr atory popula tions (H enny et al., 1991). After fledging, the young r emain de pendent on bot h parents for food us ually fo r an addit iona l 2 to 3 weeks (P oole , 1989a), but dependency can cont inue up t o 6 weeks i n the more southern popula tions (H enny , 1986).
Home range and r esour ces. Osprey build la rge sti ck nests i n the tops of tal l trees or arti ficial str uctures such as buoy s and r adio t owers (Poole , 1989a). In the Chesapeake Bay area, less th an one third of th e 1,450 breeding pa irs built th eir nests i n trees, while over half nested on channel markers and duck blinds , and t he remainde r on m iscella neous man-made str uctu res (Henny et al ., 1974). Osprey build t heir nest at th e top of th e chosen si te, which can make i t vulne rable to destr uct ion f rom high w inds (H enny , 1986). If not lost, the same n est o ften is used year after year , and it can become quite large (e.g., over 2 m tal l and 1.5 m acr oss) (Dunstan , 1973; Henny, 1988a). On islands wh ere no predators are present, osprey will n est o n the ground ( Poole , 1989b). The distan ce osprey tr avel from their nests to forage (i.e., foraging r adius ) depends on t he availa bilit y of appr opr iate nest si tes n ear areas wi th suffi cient fi sh; osprey will t ravel up to 10 to 15 km to obtain food (Van Daele and Van Daele, 1982).
Popul ation de nsity . Popula tion de nsity depends on t he availa bilit y and dis tr ibut ion of resources an d can be highly var iable . Henny (1988a) reported as man y as 1.9 nests p er hectar e in one of th e largest o sprey colonie s in t he wester n United States i n 1899, wi th an
2-66 Osprey

esti mated 1.0 to 1.2 nests p er hectar e occupie d that year . Low er dens it ies on t he order of 0.005 to 0.1 nests p er hectar e are more commo n (see tab le).
Popul ation dy namics. Breeding d ata fr om man y locat ions in t he Unit ed States an d Canada dur ing t he years 1950 to 1976 show low producti vi ty (fewer than one chick fl edged per acti ve nest o n average). Evidence indi cates th e cause to be egg-shell thinning t hat resulted from th e ospreys' exposure to DDT that had bioaccu mulated in fish (Henny and Anthony , 1989; Henny et al ., 1977; Poole , 1989a). Thus, data fr om reproduc tive studie s condu cted during t his ti me can only be used wi th this in m ind ( Spit zer et al., 1978).a
Because of th eir terminal posi tion in t he aquatic food c hain, osprey can be a sensi tive indi cato r of to xic co ntami nants th at bioaccu mulate (Henny et al ., 1978; Henny, 1988b).
Osprey are only know n to star t a sec ond c lut ch if th e fi rst one is d estr oyed (Poole , 1989a). Juveniles do not return to their place of birth unti l 2 years of age, and they do not breed unti l their th ird seaso n (Henny and Van Velzen, 1972). Often , breeding is delayed unti l 4 to 7 years of age in areas su ch as th e Chesapeake Bay, where good nesting s ites are scar ce (Poole , 1989b).
General references
Poole (1989a); Brown and Amadon (1968); Henny (1986); Henny (1988b).
aIn the table beginni ng on t he next page, data on t he num ber f ledged per active nest and t he num ber f ledged per successf ul nest are provi ded onl y for st udi es of popul ations t hat appeared to be unaffected by DDT.
2-67 Osprey

Osprey (Pandion haliaetus )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight A F 1,568 1,250 - 1,900 NS Brown & Am adon, 1968 (g) A M 1,403 1,220 - 1,600
A F cour tship 1,880 ± 20 SE se Massachuset ts Pool e, 1984 1 A F incubat ion 1,925 ± 25 SE A F late nest l. 1,725 ± 25 SE A M cour tship 1,480 ± 15 SE A M late nest l. 1,420 ± 15 SE
F at fledgi ng 1,510 Maryland, Virginia McLean, 1986 M at fledgi ng 1,210
Egg Weight (g) 72.2 ± 5.35 SD 66.0 - 81.3 North Carolina Whittemore, 1984 (carol inensi s)
Metabol ic Rate A F basal 69 est imated 2 (kcal /kg-day) A M basal 71
A F f ree-living 181 (85 - 384) est imated 3 A M free-living 186 (87 - 395)
Food Ingest ion A F cour tship 0.21 se Massachuset ts Pool e, 1983 Rate (g/g-day) perio d
Water Ingest . A F 0.051 est imated 4 Rate (g/g-day) A M 0.053
Inhal ation A F 0.578 est imated 5 Rate (m /day) A M 0.5313
Surface Ar ea A F 1,353 est imated 6 (cm )2
2-68 O
sprey

Osprey (Pandion haliaetus )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
alewife 32 Nova Scotia, Canada/ Greene et al., 1983 7 smelt 5 harbor , bay pol lock 53 winter flounder 10 (% wet weight ; obser ved
capt ures)
starry flounder 95 se Al aska/NS Hughes, 1983 7 cutth roat tro ut 5
(% wet weight ; obser ved capt ures, not ing f ish l engt h)
carp 67 w Oregon/ NS Hughes, 1983 7 crappi e 33
(% wet weight ; obser ved capt ures, not ing f ish l engt h)
gizzard shad 63 Flor ida/lake Col lopy , 1984 sunf ish 29 largemout h bass 5 (% of prey caught ; ident ified gol den shi ner 3 at nest s)
brown bul lhead 37.7 Idaho/ reservoi r Van Daele & Van Dael e, 1982 salmoni ds 20.8 nor thern squawf ish 19.3 (% of fish caught ; obser ved yellow perch 11.6 capt ures) largescal e sucker 10.6
Size of fish caught : Idaho/ reservoi r Van Daele & Van Dael e, 1982 < 10 cm 3.3 11 - 20 cm 42.1 (% of fish i n each si ze class; 21 - 30 cm 46.7 determined f rom remains at 31 - 40 cm 6.6 nest) 41 + cm 1.3
2-69 O
sprey

Osprey (Pandion haliaetus )
Popul ation Dynamics
Age/Sex Note Cond/ Seas Mean Range Locat ion/Habi tat Reference No.
Foraging A M 1.7 0.7 - 2.7 Minnesot a/lakes Dunst an, 1973 Radius (km) A B s prin g 10 Nova Scotia/coastal Greene et al., 1983
A B 3 to 8 > 1 nw California/coast al, bay Kopl in, 1981
Popul ation A B sum mer 1.9 Oregon/ lake in 1899 onl y Henny , 1988a Densi ty A B spr ing 0.028 Flor ida/wet land Eichhol z, 1980 (nests/ha) A B spr ing 0.10 North Carolina/reservoi r Henny & Nol temeier, 1975
A B spr ing 0.005 North Carolina/lake Henny & Nol temeier, 1975
Clutch 3.23 ± 0.03 SE At lant ic Seaboar d/NS Judge, 1983 Size 2.84 ± 0.07 SE Georgia, Flor ida/NS Judge, 1983
2.67 ± 0.07 SE s Cal ifornia, n Mexico/NS Judge, 1983 3.23 ± 0.09 SE ne Uni ted States/NS Spi tzer, 1980 2.82 1 - 4 Idaho/ river, lakes Henny et al., 1991
Clutches/ Year 1 NS/NS Pool e, 1989a 8
Days 38.1 ± 3.2 SD 32 - 42 Baja California, Judge, 1983 Incubat ion Mexico/coast al islands
35 - 43 Massachuset ts/NS Pool e, 1989a
Age at non- migr. 62.5 ± 4.9 SD 52 - 76 Baja California, Judge, 1983 9 Fledgi ng pop. Mexico/coast al islands (days)
migratory pop.
54 3.0 SD 48 - 59 Maryland/Cheasapeake Bay Stotts & Henny , 1975 9
Number 1.16 0.79 - 1.47 (10 yrs) N. Carolina/lake Whittemore, 1984 Fledge per 1.34 S. Carolina/lake Henny & Nol temeier, 1975 Act ive Nest 1.58 1.17 - 1.89 (3 yrs) Idaho/reservoi r Van Daele & Van Dael e, 1982
1.92 e Uni ted States/coast al Pool e, 1984
±
2-70 O
sprey

Osprey (Pandion haliaetus )
Popul ation Dynamics
Age/Sex Note Cond/ Seas Mean Range Locat ion/Habi tat Reference No.
Number 1.7 Baja California, Judge, 1983 Fledge per Mexico/coast al islands Successf ul Nest 2.14 Idaho/riv er Henny et al., 1991
1.83 or ida/lake Col lopy , 1984 1.79 Delaware/coastal bay Henny et al., 1977 2.05 Montana/lake Henny et al., 1991
Age at B 3 - 5 yrs New York, Spi tzer, 1980 10 Sexual B 3 yrs Massachuset ts/NS Henny & Wight , 1969 Maturity North Amer ica/NS
Annual 1st year 57.3 New Yor k, New Jer sey/NS Henny & Wight , 1969 Mortality years 2 - 18 18.5 ± 1.8 Rates (percent ) J B 41 NS/NS Spi tzer, 1980
A B 15
Aver age if reach sex. 4.8 NS/NS Brown & Am adon, 1968 Longevi ty maturity
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating late Apr il May mid-June Delaware, New Jersey Bent , 1937
early Dec. late February Flor ida (nonm igratory) Pool e, 1989a early Januar y early March Baja California, Mexico Judge, 1983
May Minnesot a Dunst an, 1973
(nonm igratory)
Hatching mid-March early May late May Maryland, Virginia Bent , 1937 late Apr il mid-May mid-June New York/New Engl and Bent, 1937 February late Apr il Baja California, Mexico Judge, 1983
(nonm igratory)
Migration fall late August September Novem ber most of Uni ted States Henny , 1986 11
spr ing early Apr il Minnesot a Dunst an, 1973 early March North Carolina Parnel l & Walton, 1977
Fl
2-71 O
sprey

123456
Osprey (Pandion haliaetus )
Late nest l. indi cates late nest ling st age of the breeding season. Cited in Pool e (1989a).Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Brown and Am adon ( 1968).Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Brown and Am adon ( 1968).Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Brown and Am adon ( 1968).Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Brown and Am adon ( 1968).Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Brown and Am adon(1968).
7 Percent wet weight of food i ngest ed by free-flying ospr ey est imated by ident ifying speci es of fish capt ured (using bi nocul ars), est imating t he lengt h of each f ish capt ured by com parison w ith ospr ey, and usi ng l abor atory measur es of weight s and l engt hs of samples of these fish speci es.
8 Second cl utch pr oduced onl y if first is lost . 9 Nest lings i n migratory popul ations f ledge at an ear lier age than nest lings i n nonm igratory popul ations, such as t hose i n Mexico and sout h
Flor ida. 10 Cited in Henny (1988b). 11 Cited in Henny (1986).
2-72 O
sprey

References (including A ppendix)
Bent, A. C. (1937) Life histo ries of North Amer ican birds of prey. Part 1: Order Falconif ormes. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 167.
Brown, L.; Amadon, D. (1968) Eagles, hawks, and falcons of th e world, v. 1. New York, NY: McGraw-Hill.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Collopy , M. W. (1984) Parental care, produc tivi ty, and pr edator-prey relations hips of ospreys i n three North Flor ida L akes: preliminary report. In: Westal l, M. A., ed. Proc. southeaster n U.S. and Caribbean osprey symp osium; pp. 85-98.
Cramp, S., chief ed. (1980) Handbook of th e bir ds of Eu rope, the Middle East an d North Afr ica: v. 2. Oxfo rd, UK: Oxfo rd Universi ty Pr ess.
Dunstan , T. C. (1968) Breeding s uccess o f osprey in Minne sota from 1963-1968. Loon 4 0: 109-112.
Dunstan , T. C. (1973) The biology of ospreys i n Minne sota. Loon 4 5: 108-113.
Edwards, T. C., Jr . (1988) Temporal var iation in pr ey pr eference patter ns of adult ospreys. Auk 105: 244-251.
Eichholz, N. F. (1980) Osprey nest co ncentrations in nor thwest Flor ida. Fla. Field Nat. 8: 18-19.
Fernandez, G.; Fernandez, J. (1977) Some instan t benefi ts an d long r ange hope s of color banding os preys. Transact ions of th e Nor th Amer ican osprey research conference. U.S. Natl . Park Serv. Trans. Proc. Ser. 2.
French, J. M. (1972) Distr ibut ion, abunda nce, and br eeding statu s of ospreys i n northwester n California [master 's th esis]. Arcata, CA: Humboldt State Universi ty.
French, J. M.; Koplin, J. R. (1977) Distr ibut ion, abunda nce, and br eeding statu s of ospreys in northwester n California. Transact ions of th e Nor th Amer ican osprey research conference. U.S. Natl . Park Serv. Trans. Proc. Ser. 2: 223-240.
Garber, D. P. (1972) Osprey nest ing e cology in L assen and Plumas Count ies, Calif ornia [master 's th esis]. Arcata, CA: Humboldt State Universi ty.
2-73 Osprey

Greene, E. P.; Greene, A. E.; Freedman, B. (1983) Foraging be havior and pr ey selection by ospreys i n coastal habitats i n Nova Sco tia, Canada. In: Bird, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 257-267.
Grubb, T. C., Jr . (1977) Weather dependent fo raging in os preys. Auk 94: 146-149.
Hagan, J. M. (1984) North Carolina osprey popula tion: social group or breeding aggr egation? In: Westal l, M. A., ed. Proc. southeaster n U.S. and Caribbe an osprey sympos ium .
Henny, C. J. (1977) Research, management and statu s of th e osprey in North Amer ica. In: Chancellor , R. D., ed. Proc. world bir ds of prey conf . Internat. Counc il Bir d Preser v.; Vienna, Austr ia; pp. 199-222.
Henny, C. J. (1983) Distr ibut ion a nd abunda nce of n esting os preys i n the United States. In: Bird, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 175-186.
Henny, C. J. (1986) Osprey (Pandion haliaetus), Sect ion 4 .3.1, US Army Cor ps of Engine ers Wildlif e Resour ces Management Manual. Prepared by U.S. Fish Wildl. Serv., Patuxent Wildl. Res. Center, Corvallis , OR for US Army Engine er Waterways Experiment Stat ion, Vicksb urg, MS: Technical Report EL-86-5.
Henny, C. J. (1988a) Large osprey colony dis covered in Oregon. Mur relet 69: 33-36.
Henny, C. J. (1988b) Osprey [sect ions ]. In: Palmer , R. S., ed. Handbook of Nor th Amer ican birds: v. 4. New Haven, CT: Yale Universi ty Pr ess.
Henny , C. J.; Anthony , R. G. (1989) Bald eagle and osprey. Natl . Wildl. Fed. Sci . Tech. Ser. No. 12: 66-82.
Henny, C. J.; Noltemei er, A. P. (1975) Osprey nest ing popula tions in t he coastal Carolina s. Amer . Birds 29: 1073-1079.
Henny, C. J.; Van Velzen, W. T. (1972) Migr ation p atter ns and winter ing loc alit ies of Amer ican ospreys. J. Wildl. Manage. 36: 1133-1141.
Henny , C. J.; Wight , H. M. (1969) An endangered osprey popula tion: esti mates o f mo rtal ity and pr oduc tion. Auk 86: 288-198.
Henny, C. J.; Smith, M. M.; Stotts, V. D. (1974) The 1973 distr ibut ion a nd abunda nce of breeding os preys i n the Chesapeake Bay. Chesapeake Sci . 15: 125-133.
Henny, C. J.; Byrd, M. A.; Jaco bs, J. A.; et al . (1977) Mid-Atlant ic co ast o sprey popula tion: present numbers, produc tivi ty, pollut ant con tamina tion, and statu s. J. Wildl. Manage. 41: 254-265.
2-74 Osprey

Henny , C. J.; Collins , J. A.; Deibert, W. J. (1978) Osprey distr ibut ion, abunda nce, and statu s in wester n North Amer ica. Murrlet 59: 14-25.
Henny , C. J.; Blus , L. J.; Hoffman , D. J.; et al . (1991) Lead accu mulation a nd os prey produc tion ne ar a mining s ite on t he Coeur d'Alene River, Idaho. Arch. Environ. Contam. Toxicol. 21: 415-424.
Hughe s, J. (1983) On osprey habitat an d producti vi ty: a tale of two habitats. In: Bird, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 269-273.
Judge , D. S. (1983) Producti vi ty o f ospreys i n the gulf of Calif ornia . Wils on Bull. 95: 243-255.
Kennedy, R. S. (1973) Notes o n the migration of juv enile ospreys fr om Maryland and Virginia . Bir d-Banding 4 4: 180-186.
Koplin, J. R. (1981) Reproducti ve performance of ospreys (Pandion haliaetus) in northwester n California. Natl . Geogr. Soc. Res. Rep. 13: 337-344.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967) A re-exami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lind, G. S. (1976) Produc tion, nest si te sel ect ion, and f ood ha bit s of ospreys i n Desch utes Nationa l For est, Oregon [master 's th esis]. Corvallis , OR: Oregon State Universi ty.
MacCarter , D. S. (1972) Food ha bit s of osprey at Fla thead Lake, Montana [master 's th esis]. Arcata, CA: Humboldt State Universi ty.
MacNamara, M. (1977) Sexing t he Amer ican osprey us ing seconda ry sexu al character isti cs. Transact ions of th e Nor th Amer ican osprey research conference. U.S. Natl . Park Serv. Trans. Proc. Ser. 2: 43-45.
McLean, P. K. (1986) The feeding e cology of Chesapeake Bay ospreys an d the growth and behavior of th eir young [master 's th esis]. Willi amsbur g, VA: Colle ge of Willia m and Mary.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Melquis t, W. E.; Johns on, D. R.; Carrier, W. D. (1978) Migr ation p atter ns of northern Idaho and easter n Washingt on os preys. Bird-Banding 4 9: 234-236.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
2-75 Osprey

Nesbitt, S. (1974) Foods of th e osprey at Newnans Lake. Fla. Field Nat. 2: 45.
Ogden, J. C. (1977) Preliminary report on a stu dy of Flor ida ospreys. In: Ogden, J. C., ed. Transact ions of th e Nor th Amer ican osprey research conference; pp. 143-151.
Parnell, J. F.; Walton, R. (1977) Osprey reproducti ve su ccess i n southeaster n North Carolina . Transact ions Nor th Amer ican osprey research conference. U.S. Natl . Park Serv. Trans. Proc. Ser. 2: 139-142.
Peakal l, D. B. (1988) Know n effects o f pollut ants on f ish-eating bir ds in t he Great Lakes o f Nor th Amer ica. In: Toxic cont amina tion in la rge lakes; v. 1; pp. 39-54.
Poole , A. F. (1982) Brood r educ tion in temp erate and subtropical ospreys. Oecologia (B erl.) 53: 111-119.
Poole , A. F. (1984) Reproduc tive lim itation in c oastal ospreys: an ecologic al and evolut iona ry perspecti ve [Ph .D. disser tat ion] . Bos ton, MA: Bos ton Univ ersi ty.
Poole , A. F. (1983) Cour tship feeding, clut ch size, and egg s ize in os preys: a preliminary report. In: Bird, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 243-256.
Poole , A. F. (1989a) Ospreys: a natural and unna tural his tory. Cambridge , MA: Cambridge Universi ty Pr ess.
Poole , A. F. (1989b) Regula tion of osprey Pandion haliaetus popula tions : the role of nest side availa bilit y. In: Meybur g, B.-U.; Chancellor , R. D., eds. Raptors in t he mode rn world: proceedings of th e 3rd world c onf erence on bir ds of prey and ow ls ; 22-27 March 1987; Eilat, Israel. Berlin, London, Paris : World Working G roup on Bir ds of Prey and Owls; pp. 227-234.
Prevost, Y. A. (1977) Feeding e cology of ospreys i n Antigonis h Count y, Nova Scotia [master 's th esis]. Mont real, Quebec: MacDona ld Colle ge of McGill Univ ersi ty.
Prevost, Y. A. (1982) The winter ing e cology of ospreys i n Senegambia [Ph.D. disser tat ion] . Edinbur gh, Scotland: Univ ersi ty of Edinbur gh.
Prevost, Y. A.; Bancroft, R. P.; Seymo ur, N. R. (1978) Statu s of th e osprey in Antigonis h Count y, Nova Scotia. Can. Field-Nat. 92: 294-297.
Reese, J. G. (1977) Reproducti ve su ccess o f ospreys i n central Chesapeake Bay. Auk 94: 202-221.
Robbins , C. S.; Bruun, B.; Zim, H. S. (1983) A guide to field ide nt if ication: bir ds of Nor th Amer ica. New York, NY: Golde n Press.
2-76 Osprey

Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Spitzer, P. (1980) Dynamics o f a discr ete co astal breeding popula tion of ospreys i n the northeaster n U.S. during a period of decline and r ecovery, 1969-1979 [Ph.D. disser tat ion] . Ithaca, NY: Cornell Universi ty.
Spit zer, P. R.; Risebrough, R. W.; Walker, W. I.; et al. (1978) Producti vi ty o f ospreys i n Connecti cut - Long Is land inc reases as D DE residue s decline . Science 202: 333-335.
Stinson, C. H. (1977) Familia l longe vi ty in os preys. Bird-Banding 4 8: 72-73.
Stocek, R. F.; Pearce, P. A. (1983) Distr ibut ion a nd r eproduc tive success o f ospreys i n New Brunswi ck, 1974-1980. In: Bird, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and management of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 215-221.
Stotts, V. D.; Henny, C. J. (1975) The age at fi rst f light fo r young A mer ican ospreys. Wilson Bull. 87: 277-278.
Swenson, J. E. (1978) Prey and foraging be havior of ospreys o n Yellowsto ne Lake, Wyom ing. J. Wildl. Manage. 42: 87-90.
Swenson, J. E. (1979) Facto rs affect ing statu s and reproduc tion of ospreys i n Yellow stone Nationa l Park. J. Wildl. Manage. 43: 595-601.
Szaro, R. C. (1978) Reproducti ve su ccess an d foraging be havior of th e osprey at Seahorse Key, Flor ida. Wils on Bull. 90: 112-118.
Ueoka, M. L. (1974) Feeding be havior of ospreys at H umboldt Bay, Calif ornia [master 's thesis]. Arcata, CA: Humboldt State Universi ty.
Van Daele, L. J.; Van Daele, H. A. (1982) Facto rs affect ing t he produc tivi ty of ospreys nesting in west-cen tral Idaho. Condor 84: 292-299.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Whittemo re, R. E. (1984) Histo rical overview of osprey at th e Mattamu skeet N ationa l Wildlif e Refuge : result s from ten years of nest an d producti vi ty su rveys. In: Westal l, M. A., ed. Proceedings of th e Sout heaster n U.S. and Caribbean Osprey Symp osium; pp. 17-41.
Wilcox (1944) (ci ted in Henny, 1988b) Univ . of State o f N.Y. Bull. to the Schools 30: 262-264.
2-77 Osprey

2.1.6. Red-Tailed Haw k (buteo haw ks)
Order Falconi formes, Family Acci pitr idae. The family A ccipit ridae inc lude s most birds of prey excep t fal cons, owls, and Amer ican vultures. Buteo hawks ar e covered in th is sect ion. b But eo hawks ar e moderately large soaring h awks th at inha bit open or semi -open areas. They are the most co mmo n dayti me avi an predators on ground- dwelling ver tebrates, particularly rodents an d other smal l mammal s. They range in size from th e broad winge d hawk (41 cm bill t ip t o tail t ip) to the fer ruginous h awk (58 cm). Hawks eg est pellets th at co ntain undigestible parts of th eir prey, su ch as ha ir and feath ers, that can be useful in ide nt ifying t he typ es of prey eaten (bones usually are digested completely; Duke et al ., 1987).
Selected species
The red-tai led hawk (Buteo jamaicensis) is th e most co mmo n Buteo species in the United States (N ationa l Geogr aphic So ciety, 1987). Breeding popula tions ar e dis tr ibut ed throughout mo st woode d and semi woode d regions of th e Unit ed States an d Canada south of th e tundr a (Adamci k et al ., 1979), although s ome popula tions ar e found in d eser ts an d prairie habitats. Six su bspecies are recognize d (Brown and Amadon, 1968). Nesti ng pr imarily in w oodla nds , red-tai ls feed in open count ry on a wide var iety o f smal l- to medium -sized pr ey.
Body size. Males of th is med ium-si zed buteo (46 cm) we igh a bout 1 kg, and femal es are approximatel y 20 percent heavier than the males (see tab le). Otherwise, the sexes look alik e (Brown and Amadon, 1968).
Habitat . Red-tails ar e found in ha bitats r anging f rom woodla nds , wetl ands, pastu res, and prairies to deser ts (Bohm 1978b; Gates, 1972; MacLaren et al ., 1988; Mader, 1978). They appear to prefer a mixed landscap e containing old f ields , wetl ands, and pastu res fo r foraging int erspersed wi th groves o f woodla nds and blu ffs an d str eamsi de trees fo r perching a nd n esting ( Brown and Amadon, 1968; Presto n, 1990). Red-tai ls build their nests cl ose to the tops of tr ees in low-densi ty fo rests an d often in trees th at are on a slope (B ednarz and Dins more, 1982). In areas wh ere trees are scar ce, nests ar e built on other str uctu res, occas iona lly in cactu s (Mader, 1978), on rock pinna cles or ledges, or man-made str uctu res (Brown and Amadon, 1968; MacLaren et al ., 1988). In winter , night roosts u sually are in th ick c onifers i f avai lable and in other typ es of tr ees otherwise (Brown and Amadon, 1968).
Food ha bi ts . Red-tails hunt pr imarily fr om an elevated perch, often near woodla nd edges (Bohm , 1978a; Janes, 1984; Presto n, 1990). Smal l mammal s, including m ice, shrews, voles, rabbit s, and s quir rels , are impor tant pr ey, partic ularly dur ing w int er. Red-tai ls also eat a wi de var iety o f foods depending on availa bilit y, inc luding bir ds, lizards, snakes, and large insects (B ent, 1937; Craighead and Craighead, 1956; Fitch et al ., 1946). In general, red-tails ar e oppor tunis tic and w ill feed on whatever species are most abunda nt
bOther members o f the family Acci pitri dae, eagles and the osprey, are co vered in Secti ons 2.1.7 and 2.1.5, resp ecti vely.
2-79 Red-Tailed Hawk

(Brown and Amadon, 1968). Winter food c hoi ces var y wi th snow co ver ; when smal l mammal s su ch as vo les become unavai lable (unde r th e snow ), red-tai ls may co ncentrate on larger prey, such as pheasants (Gates, 1972).
Mol t. Juvenile s molt int o adult plum age in a gradual pr ocess fr om th e spring ( age about 14 mont hs) to summer or early fal l (Bent, 1937).
Migration. The more northerly red-tai led hawk popula tions ar e migr atory while th e more southerly are year -round r eside nts (Bent, 1937).
Breeding acti vi ties and social organization. Red-tai ls lay one clutch per year cons is ting of one to three eggs , although a r eplacemen t cl utch is possible if th e init ial clut ch is los t early in t he breeding seaso n (Bent, 1937). Their nests ar e large and built of twigs (Bohm , 1978b). Both sexes i ncubate, but th e male provides food f or th e femal e dur ing inc ubation a nd t he enti re family follow ing h atching ( Brown and Amadon, 1968). The parents cont inue to feed their young after fledging w hile th ey are learning t o hunt (B rown and Amadon, 1968).
Home range and r esour ces. Red-tai led hawks ar e ter ritorial throughout th e year , inc luding w int er (Brown and Amadon, 1968). Trees or other si tes fo r nest ing a nd pe rching are impor tant requir ements fo r breeding t erritories and c an determine which habitats ar e used in a particular area (Presto n, 1990; Rothfels and Lein, 1983). Home range size can vary fr om a few hundr ed hectar es to over 1,500 hectar es, depending on t he habit at (Andersen and Rong stad , 1989; Petersen, 1979). In a 10-year stu dy in Oregon, Janes (1984) found t hat the size of r ed-tail t erritories and t he loc ation of bounda ries between ter ritories var ied littl e from year to year , even though indiv idua l bir ds or pairs die d and were replaced.
Popul ation de nsity . Popula tion de nsit ies generally do not exceed 0.03 pairs per hectar e, and usually are lower than 0.005 pairs per hectar e (see Appendix ). Popula tions in southern areas su ch as Flor ida can inc rease su bstan tially in the winter wi th the influx of migr ants from th e more nor therly popula tions (Boha ll and Collopy , 1984).
Popul ation dy namics. Beginning a t 2 years of age, most r ed-tai led hawks attemp t to breed, although t he propor tion br eeding c an vary by popula tion a nd environm ental condit ions (H enny and W ight , 1970, 1972). Average clutch size var ies regiona lly , tending t o increase fr om east to west an d from so uth to north (Henny and Wight , 1970, 1972). In a 10-year stu dy of red-tai ls in Alberta, Canada, Adamci k et al . (1979) found t hat the breeding popula tion of adult s remaine d s table despit e str ong cyclic al fluc tuations in t he density of their main prey, the snowsh oe hare, over the years. The mean clutch size for the red-tai l popula tion, however , appeared to vary wi th prey densi ty, from 1.7 to 2.6 eggs/nest (Adamci k et al ., 1979). Over the course of th e stu dy, about 50 pe rcent of obs erved nestling losses o ccu rred wi thin 3 to 4 weeks after hatching due to star vat ion. Most of th e var iance in year ly mo rtal ity o f nest lings could be attr ibut ed to the amount of food s upplie d and t he frequency o f rain. Large raptors su ch as horned owls also can be important so urces o f mortalit y for red-tail ne stlings in s ome areas (Adamci k et al ., 1979).
2-80 Red-Tailed Hawk

�
�
�
�
�
�
�
�
Simi lar species (from general references)
� The fer ruginous h awk (Buteo regalis ), one of th e larger buteos (58 cm), inha bit s the dry ope n count ry of th e wester n United States.
� The red-shoulde red hawk (Buteo lineatus) is slight ly smal ler (53 cm) an d feeds on snakes, frogs, crayfi sh, mice, and some smal l birds. Its r ange is east o f th e Rocky Mount ains and in Ca lif ornia , wi th mois t mixed woodla nds prefer red.
� Swainson's hawk (Buteo swai nsoni) is restr icted to the open plains of th e wester n United States. Although it is as la rge (53 cm) as th e red-tai l, it preys mostl y on insects.
� The broad-winge d hawk (Buteo platyp terus) is one of th e smal ler buteos (41 cm) an d preys o n mice, frogs, snakes, and insects. It prefers woodla nds an d is found a lmost excl usively east o f th e Missi ss ippi Riv er.
� Harris' hawk (Parabuteo unicinctu s) is si mi lar in size (53 cm) to the red-tai led hawk b ut is restr icted to the semi arid wood a nd br ushlands of th e sout hwest. This bird nests i n saguaro, mesquite, and yucca an d preys o n rodents, lizards, and small bir ds.
� The rough- legged hawk (Buteo lagopus ) is one of th e larger buteos (56 cm). It wi nters th roughout mo st of th e Unit ed States i n open count ry but breeds only in t he high a rcti c of Nor th Amer ica.
� The zone-tai led hawk (Buteo albono tatu s) is slight ly smal ler (51 cm) than most b uteos and feeds on rodents, lizards, fi sh, frogs , and smal l bir ds. It can be found in mesa an d mount ain c ount ry wi thin it s lim ited range between the southwest United States an d Mexico.
� The short-tai led hawk (Buteo brachyurus) is th e smal lest b uteo (39 cm) an d can only be found in t he sout hern tip of Flor ida in m ixed woodla nd and gr assl and habitats.
General references
Brown and Amadon (1968); Craighead and Craighead (1956); Fitch et al . (1946); Nationa l Geogr aphic So ciety (1987).
2-81 Red-Tailed Hawk

Red-Tailed Haw k (Buteo jamaicensis )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A F 1,224 Michi gan, Pennsy lvani a Craighead & Cr aighead, 1956 (g) A M 1,028
A F 1,154 sw Idaho Steenhof , 1983 A M 957
A F 1,235 Ohio Spr inger & Osbor ne, 1983 1 A M 1,204 hatchl ing F 58 hatchl ing M 57 juvenile F 1,149 juvenile M 962
Metabolic Rate A B s tandard 17.7 ± 5.9 SD Michigan/metabolis m Pakpahan et al., 1989 (lO /kg-day) MR /spr ing chamber2
Metabol ic Rate A F basal 73 est imated 2 (kcal /kg-day) A M basal 77
A M breeding 109 California/mount ains Sol tz, 1984 3 A F br eeding 102
A F f ree-living 192 (91 - 408) est imated 4 A M free-living 201 (95 - 426)
Food Ingest ion A F winter 0.11 Michi gan/capt ive out door s Craighead & Cr aighead, 1956 5 Rate (g/g-day) A M winter 0.10
A M sum mer 0.086
Water A F 0.055 est imated 6 Ingest ion A M 0.059 Rate (g/g-day)
Inhal ation A F 0.48 est imated 7 Rate (m /day) A M 0.423
Surface Ar ea A F 1,147 est imated 8 (cm ) A M 1,0212
2-82 R
ed-T
ailed H
awk

Red-Tailed Haw k (Buteo jamaicensis )
2-83 R
ed-T
ailed H
awk
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
summary of 10 years: mean ± SD Alberta , Canada/ Adamcik et al., 1979 9 snowshoe har e 25.6 ± 19 farm & woodl ands Richar d's gr ound squi rrel 30.4 ± 10 (% wet weight of prey
Frankl in's ground brought to chi cks) squirre l 5.1 ± 2
vol es & m ice 4.8 ± 2 other mammals 7.8 ± 6 waterfowl 16.2 ± 10 ruffed gr ouse 2.0 ± 2 sharp-tailed grouse 1.2 ± 1 other grouse 0.9 ± 1 other bi rds 6.3 ± 3
(mammals) (78.5) nc Or egon/ Janes, 1984 9 Belding' s gr ound past ure and wheat fields squirre l 52.8
mtn cot tont ail 13.1 (% wet weight of prey pocket gopher 7.3 brought to nest s; March to Townsend' s gr ound June)
squirre l 2.9 (birds) (8.5) Alector is graeca 3.5 western meadow lark 1.8
(snakes) (13.1) gopher snake 6.1
ground squi rrel 60.8 c California/foot hills Fitch et al., 1946 9 rabbi t 26.5 pocket gopher 4.3 (% wet weight of prey other mammals 2.6 brought to nest s) gopher snake 3.8 whip tail liza rd 0.3 birds 1.3

Red-Tailed Haw k (Buteo jamaicensis )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Territo ry A B sprin g 60 - 160 c Califo rnia/foothills Fitch et al., 1946 Size (ha)
A B wi nter 697 ± 316 SD 381 - 989 s Michi gan/fields, woodl ots Craighead & Cr aighead, 1956
A B f all 1,770 957 - 2,465 Colorado/upl and pr airie, Ander sen & Rongst ad, 1989 10 pinyon- juni per woodl ands
Popul ation mer: pairs/ha: Densi ty A B 0.0017 - Colorado/open aspen McGovern & McNurney, 1986
A B ar ea a 0.0004 0.0002 - 0.0005 s Michi gan/fields, woodl ots Craighead & Cr aighead, 1956 A B ar ea b 0.0012 0.0010 - 0.0013
A B 0.0012 0.0010 - 0.0015 Alberta, Canada/farm, Adam cik et al., 1979
winter: N/ha: B B 0.014 Toront o, Canada/mixed ol d Baker & Br ooks, 1981
B B 0.0015 ± 0.0012 - 0.0018 s Michi gan/fields, woodl ots Craighead & Cr aighead, 1956
0.0050
0.0003 SD
woodl ands
fie lds
Clutch 2.0 ± 0.77 SD 1 - 3 c Califo rnia/foothills Fitch et al., 1946 Size 2.32 Ar izona/desert Mader, 1978
2.2 1.9 - 2.6 /10 yrs Alberta, Canada/farm, Adamcik et al., 1979 2.11 woodl ands Henny & Wight , 1972 2.96 Flor ida/NS Henny & Wight , 1970
Oregon, Washi ngt on/NS
Clutches/ Year 1 Bent , 1937
Days 32 Alberta , Canada Adamcik et al., 1979 Incubat ion
sum
2-84 R
ed-T
ailed H
awk
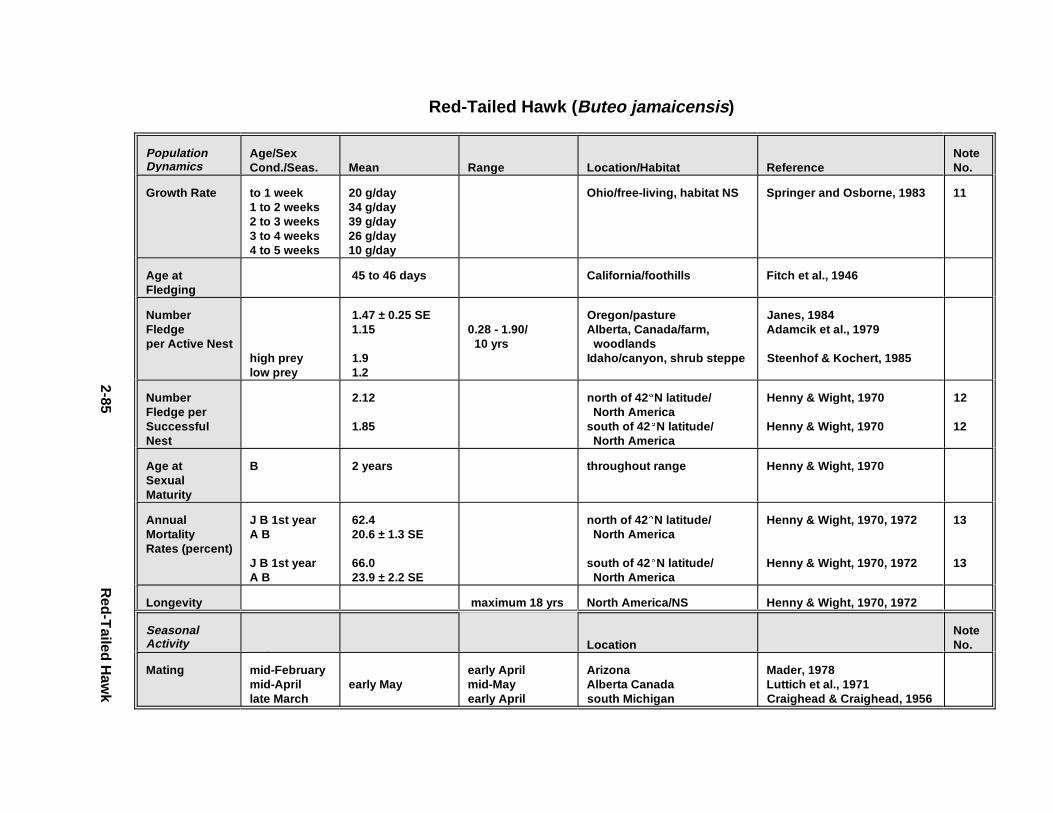
Red-Tailed Haw k (Buteo jamaicensis )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Growth Rate to 1 week 20 g/day Ohio/free-living, habi tat NS Spr inger and Osbor ne, 1983 11 1 to 2 weeks 34 g/day 2 to 3 weeks 39 g/day 3 to 4 weeks 26 g/day 4 to 5 weeks 10 g/day
Age at 45 to 46 days Califo rnia/foothills Fitch et al., 1946 Fledgi ng
Number 1.47 ± 0.25 SE Oregon/ past ure Janes, 1984 Fledge 1.15 0.28 - 1.90/ Alberta , Canada/farm, Adamcik et al., 1979 per Act ive Nest 10 yrs woodl ands
high pr ey 1.9 Idaho/ cany on, shr ub st eppe Steenhof & Kocher t, 1985 low prey 1.2
Number 2.12 nor th of 42��N latitude/ Henny & Wight , 1970 12 Fledge per North Amer ica Successf ul 1.85 sout h of 42��N latitude/ Henny & Wight , 1970 12 Nest North Amer ica
Age at B 2 years throughout range Henny & Wight , 1970 Sexual Maturity
Annual J B 1st year 62.4 nor th of 42��N latitude/ Henny & Wight , 1970, 1972 13 Mortality A B 20.6 ± 1.3 SE North Am erica Rates (percent)
J B 1st year 66.0 sout h of 42��N latitude/ Henny & Wight , 1970, 1972 13 A B 23.9 ± 2.2 SE North Am erica
Longevi ty maximum 18 yrs North America/NS Henny & Wight , 1970, 1972
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating mid-February early Apr il Arizona Mader, 1978 mid-April early May mid-May Alberta Canada Luttic h et al., 1971 late March early Apr il sout h Michi gan Craighead & Cr aighead, 1956
2-85 R
ed-T
ailed H
awk

� �
Red-Tailed Haw k (Buteo jamaicensis )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Hatching late March early May Arizona Mader, 1978 mid-May early June mid-June Alberta , Canada Luttic h et al., 1971 late Apr il early May sout h Michi gan Craighead & Cr aighead, 1956
Fall Migration mid-October Mont ana, Alberta, Canada Bent , 1937; Lut tich et al., 14
late October North Dakot a Bent , 1937 late Novem ber Minnesot a Bent , 1937
1971
Spr ing late February early March sout h Michi gan Craighead & Cr aighead, 1956 15 Migration mid-March Maine, Mont ana Bent, 1937
early April Alberta, Canada Luttic h et al., 1971
1 Est imated f rom data provi ded by author s. 2-86 R
ed-T
ailed H
awk
2 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Craighead and Cr aighead ( 1956).3 Est imated f rom time and ener gy budget s for breeding season onl y.4 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Craighead and Cr aighead ( 1956).5 Hawks m aintained out door s usi ng f alconer 's techni ques; fed lean raw beef suppl emented wi th natural prey. Overall act ivity levels not descr ibed.
Winter temperatures aver aged 3 t o 5�C and sum mer temperatures aver aged 15�C dur ing t rials. Females weighed 1, 218 g; males in winter weighed 1, 147 g; males in sum mer weighed 855 g.
6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Craighead and Cr aighead ( 1956).7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Craighead and Cr aighead ( 1956).8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Craighead and
Craighead ( 1956). 9 Percent biomass (wet weight ) est imated f rom obser vat ions of prey brought to the nest (ident ified to speci es) and r emains of prey found at the
nest s, usi ng st andar d wet weight s for each speci es of prey from other studi es or measur ed in the lab. 10 Home range determined by 95 percent ellip se method; radio-tagged hawks, two of each sex. 11 Estim ated fro m fig ure. 12 Summarizing dat a from sever al studi es. 13 Summarizing bandi ng r ecover ies pr ior to 1951. 14 Late depar ture dates. 15 Early arrival dates.

References (including A ppendix)
Adamci k, R. S.; Tood, A. W.; Keith, L. B. (1979) Demographic and di etary responses o f red-tai led hawks d uring a snow shoe hare fluc tuation. Can. Field-Nat. 93: 16-27.
Andersen, D. E.; Rong stad , O. J. (1989) Home-range esti mates o f red-tai led hawks b ased on r andom and systemati c relocat ions . J. Wildl. Manage. 53: 802-807.
Baker , J. A.; Brooks, R. J. (1981) Distr ibut ion p atter ns of raptors in relation t o dens ity of meadow vole s. Condor 83: 42-47.
Bednarz, J. C.; Dinsmo re, J. J. (1982) Nest-si tes an d habitat o f red-shoulde red and red-tai led hawks i n Iowa. Wilson Bull. 94: 31-45.
Bent, A. C. (1937) Life histo ries of North Amer ican birds of prey. Part 1: Order falconif ormes. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 167.
Boha ll, P. G.; Collopy , M. W. (1984) Seasonal abunda nce, habitat u se, and perch si tes o f four raptor species in north-central Flor ida. J. Field Ornit hol. 55: 181-189.
Bohm , R. T. (1978a) Observat ion of n est d ecoration a nd f ood ha bit s of r ed-tai led hawks. Loon 5 0: 6-8.
Bohm , R. T. (1978b) A stu dy of nest ing r ed-tai led hawks i n central Minne sota. Loon 5 0: 129-137.
Bosako wski , T.; Smith, D. G. (1992) Comparative diets o f symp atr ic nest ing r aptors in t he easter n deciduous fo rest biom e. Can. J. Zool 7 0: 984-992.
Brown, L.; Amadon, D. (1968) Eagles, hawks, and falcons of th e world: v. 1. New York, NY: McGraw-Hill.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Craighead, J. J.; Craighead, F. C. (1956) Hawks, owls and wildlif e. Harrisbur g, PA: The Stack pole Co. and Washingt on, DC: Wildl. Manage. Inst.
Duke, G. E.; Evanson, O. A.; Jegers, A. A. (1976) Meal to pellet intervals in 14 species of captive raptors. Comp. Bioc hem. Physiol. 53A: 1-6.
Duke, G. E.; Mauro, L.; Bird, D. M. (1987) Physiology . In: Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W.; et al., eds. Raptor management tec hnique s manua l. Washingt on, DC: Insti tute for Wildlif e Research, Nationa l Wildlif e Federation; Sci . Tech. Ser. No. 10; pp. 262-267.
2-87 Red-Tailed Hawk

Fitch , H. S.; Swenson, F.; Tillot son, D. F. (1946) Behavior and food ha bit s of th e red-tai led hawk. Condor 48: 205-237.
Gates, J. M. (1972) Red-tai led hawk popula tions and e cology in east-cen tral Wisco nsin. Wils on Bull. 84: 421-433.
Gatz, T. A.; Hegdal, P. L. (1987) Local winter movemen ts o f fo ur raptor species in central Color ado. West. Birds 17: 107-114.
Hagar, D. C., Jr . (1957) Nest ing popula tions of r ed-tai led hawks an d horned owls in central New York state. Wilson Bull. 69: 263-272.
Hardy, R. (1939) Nest ing ha bit s of th e wester n red-tai led hawk. Condor 41: 79-80.
Henny , C. J.; Wight , H. M. (1970) Popula tion e cology and e nvironm ental pollut ion: red-tai led and Coope r's hawks. Symp osium: Popula tion e cology of migr atory bir ds; Patuxent Wildlif e Research Center ; pp. 229-249.
Henny , C. J.; Wight , H. M. (1972) Popula tion e cology and e nvironm ental pollut ion: red-tai led and Coope r's hawks. U.S. Bur. Sport Fish. Wildl. , Wildl. Res. Rep. 2: 229-250.
Janes, S. W. (1984) Influences o f ter ritory co mposi tion a nd int erspeci fic competi tion on red-tai led hawk r eproducti ve su ccess. Ecology 65: 862-870.
Johns on, S. J. (1975) Producti vi ty o f th e red-tai led hawk i n southwester n Montana. Auk 92: 732-736.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lutti ch, S. N.; Keith, L. B.; Stephenson, J. D. (1971) Popula tion dy namics of th e red-tai led hawk (Buteo jamaicensis) at Rochester , Alberta. Auk 88: 75-87.
MacLaren, P. A.; Anderson, S. H.; Runde , D. E. (1988) Food ha bit s and n est ch aracter isti cs of breeding r aptors in s out hwester n Wyoming. Great Basin Nat. 48: 548-553.
Mader, W. J. (1978) A comparative nest ing s tudy of r ed-tai led hawks an d Harris' hawks i n sout hern Arizona . Auk 95: 327-337.
McGovern, M.; McNurney, J. M. (1986) Densi ties of red-tai led hawk n ests i n aspen stan ds in the Piceance Basin, Color ado. Raptor Res. 20: 43-45.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
2-88 Red-Tailed Hawk

Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Nice, M. M. (1954) Problems o f incubation pe riods in Nor th Amer ican bir ds. Condor 56: 173-197.
Orians, G.; Kuhlm an, J. (1956) The red-tai led hawk an d great horned owl popula tions in Wiscons in. Condor 58: 371-385.
Pakpahan, A. M.; Haufler, J. B.; Prince, H. H. (1989) Metabolic r ates o f red-tai led hawks an d great horned owls. Condor 91: 1000-1002.
Petersen, L. (1979) Ecology of great horned owls and red-tai led hawks i n southeaster n Wiscons in. Wisc. Dept . Nat. Resour . Tech. Bull. No. 111.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Presto n, C. R. (1990) Distr ibut ion of r aptor foraging in r elation t o prey bio mass an d habitat str ucture. Condor 92: 107-112.
Rothfels, M.; Lein, M. R. (1983) Territoriality i n symp atr ic popula tions of r ed-tai led and Swainson's hawks. Can. J. Zool. 61: 60-64.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Smith, D. G.; Murphy, J. R. (1973) Breeding e cology of r aptors in t he easter n Great Basin of Utah. Br igha m Young Univ . Sci . Bull. , Biol. Ser. 18: 1-76.
Soltz, R. L. (1984) Time and energy budgets o f th e red-tai led hawk (Buteo jamaicensis) in southern California. Southwest N at. 29: 149-156.
Springe r, M. A.; Kir kley, J. S. (1978) Inter - and intraspeci fic interact ions b etween red-tai led hawks an d great horned owls in central Ohio. Ohio J . Sci . 78: 323-328.
Springe r, M. A.; Osbor ne, D. R. (1983) Analysi s of growth of th e red-tai led hawk (Buteo jamaicensis). Ohio J . Sci 83: 13-19.
Steenhof , K. (1983) Prey weight s for co mput ing pe rcent bio mass i n raptor diets. Raptor Res. 17: 15-27.
Steenhof , K. (1987) Assess ing r aptor reproduc tive success an d producti vi ty. In: Giron Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W., et al., eds. Raptor management technique s manua l. Washingt on, DC: Nationa l Wildlif e Federation; pp. 157-170.
2-89 Red-Tailed Hawk

Steenhof , K.; Koc hert, M. N. (1985) Dietary sh ifts o f symp atr ic buteos during a prey decline . Oecologia 66: 6-16.
U. S. Departmen t of Inter ior. (1979) Snake River birds of prey sp ecial research report. Boise, ID: Bureau of Land Management.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
2-90 Red-Tailed Hawk

2.1.7. Bald Eagle (eagles)
Order Falconi formes, Family Acci pitr idae. Eagles have long r ounde d wings , large hook ed bills , sharp talons , and are the largest b irds of prey in the United States. They swoop dow n on t heir prey at high s peeds, and t heir die t var ies by sp ecies and considerably by habitat. In most sp ecies, the male is smal ler than the femal e, but otherwise th e sexes ar e simi lar in appearance. This fami ly also includes ki tes an d hawks.
Selected species
The bald eagle (Haliaeetus leucocephalus), our nationa l symbol, is a federally designated endangered species. Relatively co mmo n in Alaska, popula tions in t he low er 48 States h ave been ser ious ly dim inis hed, although t hey are recover ing in s ome areas. Bald eagles are most commonly sight ed in c oastal areas or near rivers or lakes. Bald eagles are pr imarily car rion feeders.
Body size. Females are signif icantly larger th an males, but otherwis e the sexes look alik e (Brown and Amadon, 1968). Body size inc reases wi th lati tude and is th e sole basis by wh ich the northern and southern subspecies are divided (Snow, 1973). Length from bill t ip t o tail t ip averages 81 cm i n the more northerly popula tions .
Habitat . Bald eagles generally are restr icted to coastal areas, lakes, and rivers (Brown and Amadon, 1968), although s ome may wi nter in areas not asso ciated wi th water (Platt, 1976). Prefer red breeding s ites i nclude prox imity to large bodie s of open water and large nest tr ees wi th stu rdy branches (often conifers) and areas of old- growth timber wi th an open and dis cont inuous canopy (A ndr ew and Mos her, 1982; Anthony et al., 1982; Grubb, 1980; Peterson, 1986). In an analysi s of mo re than 200 nests, Grubb (1980) found 55 percent wi thin 4 6 m of a shor eline and 9 2 percent wi thin 183 m of sh ore. During migr ation a nd in w int er, conif ers often are used for commu nal roost ing bot h dur ing t he day and at night , perhaps to minim ize heat loss (Anthony et al., 1982; Stalmaster , 1980). Mature trees wi th large open crowns and sto ut, horizont al perching lim bs ar e prefer red for roos ting in ge neral (Anthony et al., 1982; Chester et al ., 1990). Bald eagles reach maxi mum densi ties in areas of minimal human acti vi ty an d are almost n ever found in a reas of heavy human use (Peter son, 1986).
Food ha bi ts . Primarily car rion feeders, bald eagles eat d ead or dying f ish when availa ble but al so will catch live fi sh swi mming ne ar th e surface o r fish in shallow water s (Brown and Amadon, 1968). In general, bald eagles can be descr ibed as oppor tunis tic feeders, tak ing a dvantage of wh atever food s our ce is mo st ple nt iful and easy to scaven ge or to capture, inc luding bir ds and mammal s (Brown and Amadon, 1968; Green, 1985; Watso n et al ., 1991). In many areas, especially in winter , water fowl, kille d or injur ed by hunt ers, and s hor e bir ds ar e an impor tant food s our ce (Todd e t al., 1982). Eagles fo rage in upla nd areas in the winter when surface water s are frozen over , consuming c arrion inc luding r abbit s, squir rels , and de ad domesti c l ivesto ck su ch as pigs and c hicken s (Brown and Amadon, 1968; Harper et al ., 1988). Bald eagles also have been known to steal food f rom other memb ers of th eir own species as wel l as fr om hawks, osprey, gulls , and mergansers (Grubb, 1971; Jorde and Lingle , 1988; Sobkowiak and Titman , 1989). This
2-91 Bald Eagle

may o ccu r when there is a sh ortage of a primary food s our ce, su ch as fish, and an abunda nce of other prey such as water fowl being us ed by other predatory bir ds (Jo rde and Lingle , 1988). Some prey are important to a few popula tions ; for examp le, in the Chesapeake Bay region, turtles are cons umed dur ing t he breeding seaso n (Clark, 1982), and at Amch itka Island in Alaska, sea otter pups ar e found r egula rly in ba ld eagle n ests (Sherrod et al ., 1975). In the Paci fic Northwest d uring t he breeding seaso n, Watso n et al . (1991) found t hat ba ld eagles hunt ed liv e prey 57 percent of th e time, scaven ged for 24 percent of th eir prey, and pirated 19 percent (mo stl y fr om gulls or other eagl es). Because bald eagles scaven ge dead or dying pr ey, th ey are partic ularly vulne rable to environm ental contami nants an d pesti cides (e.g., from feeding on bir ds th at die d f rom p esti cides, cons uming le ad shot fr om water fowl kille d or dis abled by hunt ers) (Henny and A nthony , 1989; Harper et al ., 1988; Lingle and Kr apu, 1988). Bald eagles also are vulne rable to biom agnif ication of con tamina nts in f ood c hains . For examp le, near Lake Superior (WI), herring gulls , whic h were cons umed by over 20 percent of n esting ba ld eagle pairs, wer e found t o be a signif icant sour ce of DDE and PCB in take b y th e eagles (Kozie and A nderson, 1991). The gulls cont ained highe r con tami nant levels th an the local fish because of th eir highe r tr ophic l evel .
Mol t. Adult eagle s molt year ly. In northern popula tions , molt ing oc curs from late spring t o early fall; in s out hern popula tions , molt ing may b e init iated earlier (McCollough, 1989). It is l ikely th at th e molt is not co mplete, and that so me feath ers are retained for 2 years. Young ba ld eagles generally molt int o their adult plum age by th eir fi fth year (McCollough, 1989).
Migration. Bald eagles mi grate out of areas wh ere lakes ar e completely fr ozen over in w int er, but will r emain a s far nor th as the availa bilit y of open water and a reliable food supply allow (B rown and Amadon, 1968). Areas wi th ice-fr ee water ways, such as th e Colum bia River estu ary in Washingt on and Oregon, may s uppor t bot h reside nt an d migr atory popula tions in t he wint er (Watso n et al ., 1991). The far northern breeding popula tions migr ate so uth for the winter and often congr egate in areas wi th abunda nt food, partic ularly the Mississippi V alley and t he nor thwester n States (Sn ow, 1973). Some popula tions of eagle s that br eed in s out hern lati tude s (e.g., Arizona , Flor ida) show a reverse mi gration a nd m igr ate north in midsummer (follow ing br eeding) , returning s out h in early au tumn or wint er (Brown and Amadon, 1968; Grubb e t al., 1983).
Breeding acti vi ties and social organization. Bald eagles have been observed to nest su ccessf ully at 4 years of age, but mo st d o not breed unti l at least th eir fi fth year (Nye, 1983). Breeding pa irs remain t ogether as long a s bot h are aliv e (Brown and Amadon, 1968). Large sti ck n ests (ap proximatel y 1.5 m acr oss an d 0.6 m deep) are built near water and most o ften in a large tree, but so meti mes o n rocky o utcr ops or even on the ground on some is lands (B rown and Amadon, 1968; Grubb, 1980). In the absence of distu rbance, the same n est si te may b e used for many year s (Nash et al ., 1980). In Flor ida, eggs ar e laid in late autumn or winter , while over th e rest o f th e eagle's range, mat ing a nd egg la ying oc cur in s pring ( Brown and Amadon, 1968). Clutch sizes are larger in the north, and both sexes take r espons ibilit y for feeding t he young ( Brown and Amadon, 1968). Young f ledge at about 10 to 12 weeks o f age; after leaving t he nest, they are st ill de pendent on t heir parents for sever al weeks an d often return to the nest fo r food ( Sprunt et al., 1973). After nest ing, large groups will o ften gather at si tes wi th plenti fu l food
2-92 Bald Eagle

�
resources, such as along r ivers follow ing a sal mon s pawn (Fitzner and Ha nson, 1979; Keister et al ., 1987; McClelland, 1973).
Home range and r esour ces. Dur ing t he breeding seaso n, eagles requir e large areas in the vicinity o f open water , wi th an adequate supply of n esting t rees (Brown and Amadon, 1968). Distan ce fr om human distu rbance is an important facto r in nest si te sel ect ion, and nests h ave been reported to fai l as a result of d istu rbance (Andrew and Mosher, 1982). Dur ing inc ubation a nd br ooding, eagles show ter ritorial defense of an area around t he nest si te. Follow ing f ledging, there is lit tle need for nest d efense, and eagles are oppor tunis tic in their search for abunda nt sour ces o f prey (Mahaffy an d Frenzel, 1987). During w int er, eagles roos t communa lly in la rge aggr egations and s hare a foraging hom e range. For examp le, Opp (1980) descr ibed a popula tion of 150 eagles th at fed on meadow vo les in a 250-ha floode d f ield f or a 4-week p eriod. This group a lso establis hed a co mmuna l night roost i n the vicinity.
Popul ation de nsity . Because popula tion de nsity depends str ongly on t he conf igur ation of th e surface water bodie s us ed for fo raging, few i nvesti gators have publis hed explic it dens ity esti mates o n an area basis; most r eport breeding de nsit ies along a shor eline on a line ar basis. Dur ing t he breeding seaso n, 0.03 to 0.4 pairs have been recorded per km sh ore (see tab le). Eagles mi grating s out h f rom th eir su mmer ter ritories in Canada have aggregated in commu nal roosts o f up to 400 eagles in a 40-ha area (Crenshaw and McClelland, 1989). In the winter , commu nal roost si tes may al so contain large numbers of eagles. Opp (1980) descr ibed a group of 150 eagles th at roosted and foraged together in the Klamath Basin (OR/CA), and commu nal night r oosts o f up to 300 eagles in O regon in l ate wi nter .
Popul ation dy namics. Not all a dult s in a n area are part of th e breeding popula tion. Some pairs may esta blis h territories and not breed, while others may n ot even pair. The percentage of adult s br eeding a nd t he breeding s uccess o f th ose th at do vary wi th local food a bunda nce, weath er, and habitat c ondit ions (H ansen, 1987; Hansen and Hodge s, 1985; McAllister et al ., 1986). In past year s, bioaccu mulation of organoc hlor ine pollut ants reduced the reproducti ve su ccess o f bald eagles. Now, in many areas, these raptors are reproduc ing a t rates si mi lar to those prior to the widespread use of th ese pesti cides (Green, 1985). Eagles lay one clutch per year , although r eplacemen t cl utch es may b e laid upon los s of th e init ial one (Sh errod et al., 1987). Very l ittl e is kn own about mo rtalit y rates of bald eagles; Grier (1980) conclude d f rom popula tion m odels th at adult su rviv al is mo re important th an reproducti ve rate to the continue d success o f bald eagle popula tions . In captivi ty, bald eagles have l ived for up to 50 years (Snow, 1973), and one wild e agle, banded and recaptured in Alaska, was esti mated to be almost 22 year s old ( Cain, 1986). Upon los s of an init ial c lut ch, bald eagles may l ay replacemen t cl utch es i f su ffi cient ti me remains (Sherrod et al ., 1987).
Simi lar species (fr om general references)
� The golde n eagle (Aquila chrysaeto s) is si mi lar in size (81 cm) to the bald eagle, and its r ange encompasses al l but th e southeaster n United States. Smal l mammal s, snakes, birds, and car rion a re pr imary pr ey items, and golde n eagles pr efer mount ainous or hilly ter rain.
2-93 Bald Eagle

General references
Brown and Amadon (1968); Green (1985); Peterson (1986); Stalmaster and Gessaman (1982, 1984).
2-94 Bald Eagle
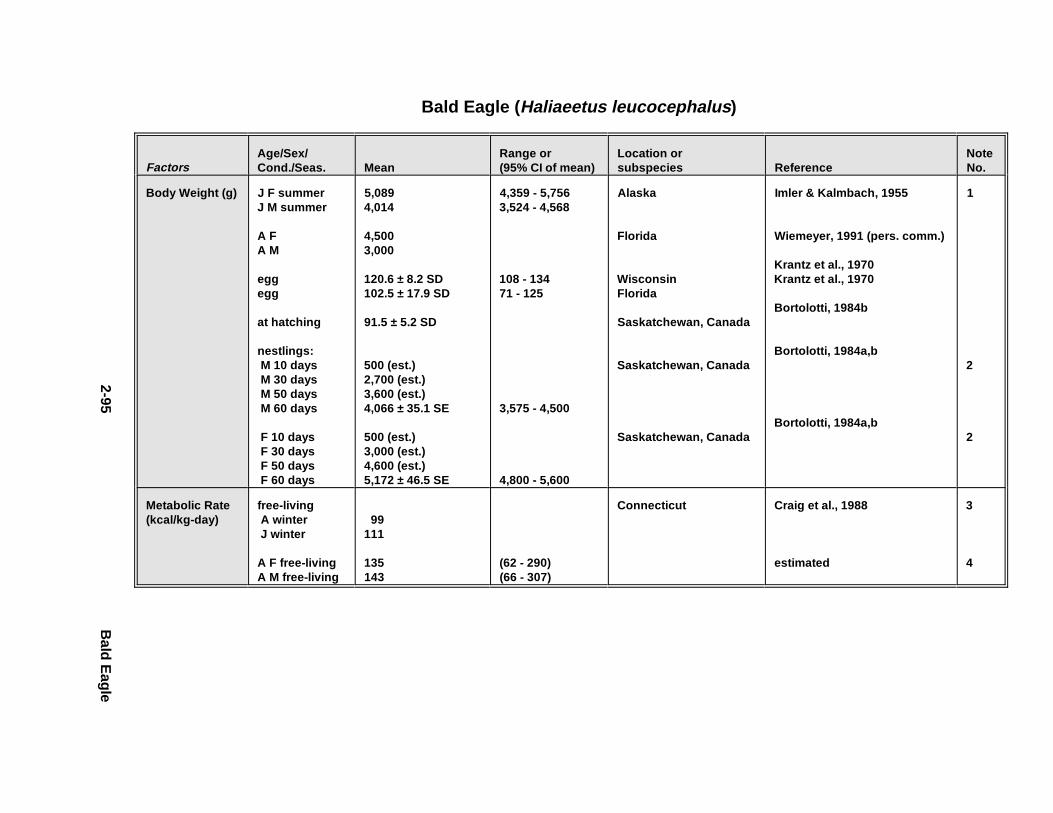
Bald Eagle ( Haliaeetus leucoce phalus )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight (g) J F sum mer 5,089 4,359 - 5,756 Alaska Imler & Kal mbach, 1955 1 J M sum mer 4,014 3,524 - 4,568
A F 4,500 Flor ida Wiemeyer, 1991 (pers. com m.) A M 3,000
egg 120.6 ± 8.2 SD 108 - 134 Wisconsi n Krantz et al., 1970 egg 102.5 ± 17.9 SD 71 - 125 Flor ida
at hatching 91.5 ± 5.2 SD Saskat chew an, Canada
nest lings: Bor tolot ti, 1984a,b M 10 days 500 (est .) Saskat chew an, Canada 2 M 30 days 2,700 (est .) M 50 days 3,600 (est .) M 60 days 4,066 ± 35.1 SE 3,575 - 4,500
F 10 days 500 (est .) Saskat chew an, Canada 2 F 30 days 3,000 (est .) F 50 days 4,600 (est .) F 60 days 5,172 ± 46.5 SE 4,800 - 5,600
Krantz et al., 1970
Bor tolot ti, 1984b
Bor tolot ti, 1984a,b
Metabolic Rate free-liv ing Connecticut Craig et al., 1988 3 (kcal /kg-day) A w inter 99
J winter 111
A F f ree-living 135 (62 - 290) est imated 4 A M free-living 143 (66 - 307)
2-95 B
ald E
agle

Bald Eagle ( Haliaeetus leucoce phalus )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Food Ingest ion winter: Rate (g/g-day) B B sal mon 0.092 ± 0.026 SD Utah (capt ive) Stalmaster & Gessam an, 1982
B B r abbi t 0.075 ± 0.013 SD B B duck 0.065 ± 0.012 SD
A B 0.12 Washi ngt on ( free-flying) Stalmaster & Gessam an, 1984 5 subadul t B 0.10 juvenile B 0.091
A B 0.12 Connecticut (free-fly ing) Craig et al., 1988 6 juvenile B 0.14
Water Ingest ion A F 0.035 est imated 7 Rate (g/g-day) A M 0.037
Inhal ation A F 1.43 est imated 8 Rate (m /day) A M 1.193
Surface Ar ea A F 2,970 est imated 9 (cm ) A M 2,5302
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
mallard 32 Washi ngt on/river Fitzner & Hanson, 1979 American w idgeon 9 American coot 9 (% biomass; prey remains other bi rds 3 found bel ow com munal Chinook sal mon 21 roost ) sucker 4 European car p 1 other fish 1 unaccount ed 20
2-96 B
ald E
agle

Bald Eagle ( Haliaeetus leucoce phalus )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
brown bul lhead 24.8 Maine/inland r iver Todd et al., 1982 white sucker 19.5 chai n pi cker el 20.1 (% occur rence i n pel lets) smallmout h bass 3.8 white perch 3.6 samples f rom all seasons other fish 4.9 except winter black duck 3.0 other bi rds 13.5 mammals 6.8
(fish) (57.6) cent ral Ar izona/desert Haywood & Ohm art, 1986 channel cat fish 21.8 scr ub, ripar ian Sonor a sucker 8.6 carp 17.3 (% biomass; prey obser ved other fish 8.5 brought to nest or found at
(birds) (14.1) nest s) American coot 8.1 great blue her on 4.4
(mammals) (28.1) deser t cot tont ail 8.1 jackr abbi t 14.9 rock squirre l 1.1
(rept iles) (0.2)
pink sal mon 15.5 Alaska/coast al Ofelt, 1975 herring 32.0 trout 4.5 (% frequency of occur rence; other fish 24.0 prey obser ved br ought to other animals 24.0 the nest )
2-97 B
ald E
agle

Bald Eagle ( Haliaeetus leucoce phalus )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Territory Area pair spr ing 3,494 ± 2,520 SD 1,821 - 6,392 Ar izona/desert, ripar ian r iver Haywood & Ohm hart, 1983 (ha) Territory pair 3.5 1.4 - 7.2 Washi ngt on/SJ Islands; Grubb, 1980 Lengt h (km) pair 15.8 11.1 - 26.6 Grays Harbor
Territory pair incubat . 0.56 ± 0.18 SE Minnesot a/lake, woods Mahaffy & Frenzel, 1987 Radius (km) pair broodi ng 0.72 ± 0.21 SE
Winter Home J B w inter 1,830 ± 1,460 SD Missour i/lake Griffin & Basket t, 1985 Range (ha) A B wi nter 1,880 ± 900 SD
Foraging B B w inter 3 to 7 Connecticut/riv er Craig et al., 1988 Distance (km)
Popul ation sum mer 0.38 se Al aska/river ine Hansen, 1987 Densi ty (pair/km summer WY, ID, MT/: Swenson et al., 1986 shor e) 0.035 Yellowstone
0.026 Cont inental 0.045 Snake
Clutch 2 1 - 3 NS/NS Brown & Am adon, 1968 Size 2.3 1 - 4 PA, DE, MD, NJ Schm id, 1966-67
Clutches/Yea 1 NS/NS Sherro d et al., 1987 r
Days 35 34 - 38 Maryland (capt ive) Maestrelli & Wiemeyer, 1975 Incubat ion
Age at M 79.9 ± 1.08 SE Saskat chew an/lake Bor tolot ti, 1989 Fledgi ng F 83.0 ± 0.94 SE (days)
Number 1.01 0.58 - 1.22/10 yr California/NS Henny & Ant hony , 1989 Fledge per 1.28 1.07 - 1.58/9 yr Mont ana/NS Henny & Ant hony , 1989 Act ive Nest 0.90 0.76 - 1.14/7 yr Washi ngt on/NS Henny & Ant hony , 1989
1.14 Flor ida/NS McEwan & Hi rth, 1979 1.00 ± 0.06 SE 0 - 3 Alaska/NS Sprunt et al., 1973
2-98 B
ald E
agle

Bald Eagle ( Haliaeetus leucoce phalus )
2-99 B
ald E
agle
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Number 1.65 ± 0.26 SD Arizona/desert scr ub, river Grubb et al., 1983 Fledge per 1.35 ± 0.11 SD 1.22 - 1.48/6 yr Washi ngt on/San Juan I sland Grubb et al., 1983 Successf ul 2.2 1 - 3 PA, DE, MD, NJ/NS Schm id, 1966-67 Nest 1.64 ID, MT, WY/riv er, lake Swenson et al., 1986
Age at B 4 3 - 5 Uni ted States/NS Nye, 1983 Sexual Maturity
Annual A B 5.4 Alaska/Amchi tka Island Sherrod et al., 1977 Mortality fledgi ng t o 1 yr 89.3 (percent )
Longevi ty A B up t o 50 yrs capt ivi ty Snow, 1973
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Layin late September Novem ber Florida, Texas Mager, 1977 10 g December late December late Januar y Ar izona Grubb et al., 1983
late October March se Uni ted States USFWS, 1989 February late March late Apr il MD, VA, DE LeFranc & Cl ine, 1983 early March early April WY, MT, ID Swenson et al., 1986 late March Vancouver BC Brown & Am adon, 1968
Fledgi ng Apr il May s Loui siana Harris et al., 1987 early Jul y late Jul y mid-August WY, MT, ID Swenson et al., 1986
late August se Al aska Hansen, 1987
Fall Migration June Ar izona Grubb et al., 1983 early October Novem ber mid-December Mont ana Crenshaw & McClelland, late October 1989
Novem ber December Januar y se Al aska Hodges et al., 1987 December/Januar y sc Or egon, n Cal ifornia Keister et al., 1987
Spr ing December Arizona Grubb et al., 1983 Migration Apr il sc Or egon, n Cal ifornia Keister et al., 1987
late March early April WY, MT, ID Swenson et al., 1986 early March Illinoi s Sabine, 1981

Bald Eagle ( Haliaeetus leucoce phalus )
1 Cited in Maestrelli a nd Wiemeyer (1975) and Bor tolot ti (1984a); juveni les up t o 3 years of age.2 Est imated f rom Figur e 4.3 Daily energy budget for free-living eagl es based on t ime-act ivity budget s and m etabol ic model s; assum ing 4.5 kg eagl e.4 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Imler and Kal mbach ( 1955).5 Est imated f rom obser ved capt ures of preweighed sal mon pr ovi ded at feeding st ations. Eagle body weight assum ed to be 4.5 kg. Some feeding
may have occur red elsewhere. 6 Est imate of food consum ed based on obser ved f eeding behavi ors and an eagl e body weight of 4.5 kg. 7 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Imler and Kal mbach ( 1955). 8 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Imler and Kal mbach ( 1955). 9 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Imler and Kal mbach
(1955). 10 Cited in Green, 1985.
2-100 B
ald E
agle

References (including A ppendix)
Andrew, J. M.; Mosher, J. A. (1982) Bald eagle nest si te sel ect ion a nd n esting ha bitat i n Maryland. J. Wildl. Manage. 46: 382-390.
Anthony , R. G.; Knight , R. L.; Allen, G. T.; et al. (1982) Habitat u se by nest ing a nd r oos ting bald eagles in the Paci fic Northwest. Trans. North Am. Wildl. Nat. Resour . Conf . 47: 332-342.
Bortolotti , G. R. (1984a) Sexual size dimorphism an d age-related size var iation in ba ld eagles. J. Wildl. Manage. 48: 72-81.
Bortolotti , G. R. (1984b) Physi cal developm ent of nestling ba ld eagles wi th empha sis on t he timing of growth events. Wilson Bull. 96: 524-542.
Bortolotti , G. R. (1989) Facto rs influencing t he growth of bald eagles in nor th central Saskatch ewan. Can. J. Zool. 67: 606-611.
Brown, L.; Amadon, D. (1968) Eagles, hawks, and falcons of th e world: v. 1. New York, NY: McGraw-Hill.
Cain, S. L. (1986) A new longe vi ty record for th e bald eagle. J. Field Ornit hol. 57: 173.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chester , D. N.; Stauffer , D. F.; Smith, T. J.; et al . (1990) Habitat u se by nonbr eeding ba ld eagles in Nor th Carolina . J. Wildl. Manage. 54: 223-234.
Chur a, N. J.; Stewar t, P. A. (1967) Care, food c ons umpt ion, and be havior of bald eagles used in DDT tests. Wilson Bull. 79: 441-458.
Clark, W. S. (1982) Turtles as a f ood s our ce of n esting ba ld eagles in t he Chesapeake Bay region. J. Field Ornit hol. 53: 49-51.
Craig, R. J.; Mitch ell, E. S.; Mitch ell, J. E. (1988) Time and energy budgets o f bald eagles wint ering a long t he Conn ecti cut River . J. Field Ornithol. 59: 22-32.
Crenshaw, J. G.; McClelland, B. R. (1989) Bald eagle use of a co mmu nal roost. Wilson Bull. 101: 626-633.
Dugoni, J. A.; Zwank, P. J. ; Furman, G. C. (1986) Food of n esting ba ld eagles in Louis iana. Raptor Res. 20: 124-127.
Duke, G. E.; Evanson, O. A.; Jegers, A. A. (1976) Meal to pellet intervals in 14 species of captive raptors. Comp. Bioc hem. Physiol. 53A: 1-6.
2-101 Bald Eagle

Duke, G. E.; Mauro, L.; Bird, D. M. (1987) Physiology . In: Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W.; et al., eds. Raptor management tec hnique s manua l. Washingt on, DC: Insti tute for Wildlif e Research, Nationa l Wildlif e Federation; Sci . Tech. Ser. No. 10; pp. 262-267.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Dunstan , T. C.; Harper, J. F. (1975) Food ha bit s of bald eagles in nor th-central Minne sota. J. Wildl. Manage. 39: 140-143.
Dzus, E. H.; Gerrard, J. M. (1989) Inter lake var iations of bald eagle, Haliaeetus leucocephalus, popula tions in nor th-central Saskatch ewan. Can. Field-Nat. 103: 29-33.
Fielder, P. C. (1982) Food ha bit s of bald eagles along t he mid- Colum bia River, Washingt on. Murrelet 63: 46-50.
Fielder, P. C.; Starkey, R. G. (1980) Winter ing ba ld eagle use along t he uppe r Colum bia River, Washingt on. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 177-193.
Fitzner, R. E.; Hanson, W. C. (1979) A congr egation of wint ering ba ld eagles. Condor 81: 311-313.
Fitzner, R. E.; Watso n, D. G.; Rickar d, W. (1980) Bald eagles of th e Hanford Nationa l Environmental Research Park. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 207-218.
Frenzel, R. W.; Anthony , R. G. (1989) Relations hip of diets an d environmental contami nants in w int ering ba ld eagles. J. Wildl. Manage. 53: 792-802.
Gessaman , J. A.; Fulle r, M. R.; Pekins, P. J.; et al . (1991) Rest ing metabolic r ate of golde n eagles, bald eagles, and ba rred ow ls wi th a tracking t ransmi tter or an equivalent load. Wils on Bull. 103: 261-265.
Green, N. (1985) The bald eagle. Audubon W ildl. Rep. 508-531.
Grier, J. W. (1977) Quadrant sam pling of a nesting popula tion of bald eagles. J. Wildl. Manage. 41: 438-443.
Grier, J. W. (1980) Modeling a ppr oaches to bald eagle popula tion dy namics. Wildl. Soc. Bull. 8: 316-322.
Grier, J. W. (1982) Ban of DDT and subsequent recovery of reproduc tion in ba ld eagles. Science 218: 1232-1235.
2-102 Bald Eagle

Griffi n, C. R.; Baskett, T. S. (1985) Food availa bilit y and w int er range size s of i mmatu re and adult bald eagles. J. Wildl. Manage. 49: 592-594.
Grubb, T. C. (1971) Bald eagles stea ling f ish from co mmon m ergansers. Auk 88: 928-292.
Grubb, T. G. (1976) A survey an d analysi s of nest ing ba ld eagles in wester n Washingt on [master 's th esis]. Seattl e, WA: Univ ersi ty of W ashingt on.
Grubb, T. G. (1980) An evaluation of bald eagle n esting in wester n Washingt on. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 87-103.
Grubb, T. G.; Hensel, R. J. (1978) Food ha bit s of n esting ba ld eagles on Kodia k Is land, Alaska. Murrelet 59: 70-72.
Grubb, T. G.; Knight , R. L.; Rubink , D. M.; et al. (1983) A five year comparison of bald eagle produc tivi ty in W ashingt on and A rizona . In: Bir d, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 35-45.
Hansen, A. J. (1987) Regula tion of bald eagle r eproduc tive rates i n southeast A laska. Ecology 68: 1387-1392.
Hansen, A. J.; Hodge s, J. I. (1985) High r ates o f non- breeding a dult bald eagles in southeaster n Alaska. J. Wildl. Manage. 49: 454-458.
Harper; R. G.; Hopk ins , D. S.; Dunstan , T. C. (1988) Nonf ish pr ey of wint ering ba ld eagles in Illinois . Wils on Bull. 100: 688-690.
Harris, J. O. ; Zwank, P. J. ; Dugoni, J. A. (1987) Habitat sel ect ion a nd be havior of n esti ng bald eagles in Louis iana. J. Raptor Res. 21: 27-31.
Haywood, D. D.; Ohmar t, R. D. (1983) Preliminary report on habitat u tiliza tion by two pairs of breeding ba ld eagles in A rizona . In: Bir d, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 87-94.
Haywood, D. D.; Ohmar t, R. D. (1986) Utiliza tion of benthic-feeding f ish by inla nd br eeding bald eagles. Condor 88: 35-42.
Henny , C. J.; Anthony , R. G. (1989) Bald eagle and osprey. Natl . Wildl. Fed. Sci . Tech. Ser. No. 12: 66-82.
Hensel , R. J.; Troyer , W. A. (1964) Nest ing s tudie s of th e bald eagle in A laska. Condor 66: 282-286.
Herrick, F. H. (1932) Daily l ife of th e Amer ican eagle: early phase. Auk 49: 307-323.
2-103 Bald Eagle

Hodge s, J. I.; King, J. G. (1979) Resurvey o f th e bald eagle breeding popula tion in southeast A laska. J. Wildl. Manage. 43: 219-221.
Hodge s, J. I.; Boeker , E. L.; Hansen, A. J. (1987) Movemen ts o f radio- tagge d bald eagles, Haliaeetus leucocephalus, in and from so utheaster n Alaska. Can. Field-Nat. 101: 136-140.
Howard, R. P.; van Daele, L. J. (1980) An overview of th e statu s of bald eagles in Idaho. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on bald eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 23-34.
Hulce, H. (1886) Eagles breeding in c aptivi ty. Forest an d Stream 27: 327.
Hulce, H. (1887) Eagles breeding in c aptivi ty. Forest an d Stream 28: 392.
Imler, R. H.; Kalmbach, E. R. (1955) The bald eagle and its ec onom ic statu s. U.S. Fish Wildl. Ser. Circ. 30.
Jorde, D. G.; Lingle , G. R. (1988) Kleptoparasi tism b y bald eagles wi nter ing in s out h-central Nebraska. J. Field Ornithol. 59: 183-188.
Keis ter , G. P., Jr .; Anthony , R. G.; Holbo, H. R. (1985) A model of energy co nsumption in bald eagles: an evalua tion of night co mmuna l roos ting. Wils on Bull. 97: 148-160.
Keis ter , G. P., Jr .; Anthony , R. G.; O'Neill, E. J. (1987) Use of co mmu nal roosts an d foraging areas by bald eagles wi nter ing in t he Klamath Basin. J. Wildl. Manage. 51: 415-420.
Kozie , K. D.; Anderson, R. K. (1991) Producti vi ty, diet, and environmental contami nants i n bald eagles nesting ne ar th e Wiscons in s hor eline of L ake Superior. Arch. Environ. Contam. Toxicol. 20: 41-48.
Krantz, W. C.; Mulhe rn, B. M.; Bagley, G. E.; et al. (1970) Organochlor ine and h eavy metal residues in bald eagle eggs. Pesti c. Monit . J. 4: 136-40.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
LeFranc, M. N., Jr .; Cline , K. W. (1983) The occu rrence of birds as p rey at acti ve bald eagle nests i n the Chesapeake Bay region. In: Bir d, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 79-86.
Lingle , G. R.; Krapu, G. L. (1988) Ingestion of lead shot and a lum inum bands by bald eagles dur ing w int er in Nebraska. Wilson Bull. 100: 326-327.
Maestr elli, J. R.; Wiemeyer , S. N. (1975) Breeding ba ld eagles in c aptivi ty. Wils on Bull. 87: 45-53.
2-104 Bald Eagle

Mager, D. (1977) The life and the future of th e southern bald eagle. In: Proceedings of th e bald eagle conference on eagle movemen ts. Apple River, IL: Eagle Valle y Environmental ists; pp. 115-117.
Mahaffy, M. S.; Frenzel, L. D. (1987) Territorial responses o f northern bald eagles near acti ve nests. J. Wildl. Manage. 51: 551-554.
McAllister , K. R.; Owens, T. E.; Lesch ner, L.; et al . (1986) Distr ibut ion a nd pr oduc tivi ty of nesting ba ld eagles in W ashingt on, 1981-1985. Murrelet 67: 45-50.
McClelland, B. R. (1973) Autumn concentrations of bald eagles in G lacier Nationa l Park. Condor 75: 121-123.
McCollough, M. A. (1989) Molt ing s equence and aging of bald eagles. Wils on Bull. 101: 1-10.
McEwan , L. C.; Hirth, D. H. (1979) Southern bald eagle producti vi ty an d nest si te sel ect ion. J. Wildl. Manage. 43: 585-594.
McEwan , L. C.; Hirth, D. H. (1980) Food ha bit s of th e bald eagle in nor th-central Flor ida. Condor 82: 229-231.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Murphy, J. R. (1965) Nest si te sel ect ion by th e bald eagle in Yello wstone Nationa l Park. Proc. Utah Acad. Sci . Arts an d Letter s 12: 261-264.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nash, C.; Pruett-Jo nes, M.; Allen, T. G. (1980) The San Juan Islands bald eagle nesti ng survey. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 105-176.
Nichols on, D. J. (1952) Littl e known facts a bout Flor ida bald eagles. Flor ida Nat. 25: 23-26.
Nye, P. E. (1983) A biologic al and econom ic r eview of th e hacking pr ocess fo r the resto ration of bald eagles. In: Bir d, D. M.; Seymo ur, N. R.; Gerrard, J. M., eds. Biology and m anagement of bald eagles and ospreys. St. Anne de Bellvue, Quebec: Harpell Press; pp. 127-135.
Ofel t, C. H. (1975) Food ha bit s of n esting ba ld eagles in s out heast A laska. Condor 77: 337-338.
Opp, R. R. (1980) Statu s of th e bald eagle in Oregon - 1980. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle sympos ium , June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 35-48.
2-105 Bald Eagle

Peterson, A. (1986) Habitat su itabilit y inde x mode ls : bald eagle (b reeding seaso n). U.S. Fish Wildl. Serv. Biol. Rep. 82(10.126); 25 pp.
Platt, J. B. (1976) Bald eagles wi nter ing in Ut ah deser t. Amer ican Birds 30: 783-788.
Rickl efs, R. E. (1973) Fecundit y, mo rtalit y and avian demogr aphy . In: Farner, D. S., ed. Breeding biology of bir ds. Washingt on, DC: Nationa l Academy o f Sci ences.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Sabine , N. (1981) Ecology of bald eagles wint ering in s out hern Illinois [master 's th esis] (abstr act). Carbonda le, IL: Sout hern Illinois Univ ersi ty.
Schmid, F. C. (1966-67) Numbers of eggs and young of bald eagles in f our middle A tlant ic states. Cassinia 50: 15-17.
Sherrod, S. K.; Estes, J. A.; White, C. M. (1975) Depredation of sea otter pups by bald eagles at Amch itka Island, Alaska. J. Mammal . 56: 701-703.
Sherrod, S. K.; Whit e, C. M.; Willi amso n, F. S. (1977) Biology of th e bald eagle on Amchit ka Island, Alaska. Living Bir d 15: 143-182.
Sherrod, S. K.; Jenkins, M. A.; McKee, G.; et al . (1987) Using w ild e ggs fo r produc tion of bald eagles for reint roduc tion int o the sout heaster n United States. In: Odom, R. R.; Riddle berger, K. A.; Ozier, J. C., eds. Proceedings of th e thir d sout heaster n nongame an d endangered wildlif e sympos ium ; Augus t; Athens, GA. Atlanta, GA: Georgia Dept. of Natural Resources; pp. 14-20.
Snow, C. (1973) Habitat man agement ser ies fo r endangered species report number 5: sout hern bald eagle Haliaeetus leucocephalus leucocephalus and nor thern bald eagle Haliaeetus leucocephalus alascan sus. Denver, CO: Bur eau of Land Management; BLM-YA-PT-81-019-6601.
Snyder, N. F.; Wiley, J. W. (1976) Sexual size dimorphism i n hawks an d owls of North Amer ica. Ornit hol. Monogr . 20.
Sobkowiak, S.; Titman , R. D. (1989) Bald eagles k illing A mer ican coot s and stea ling c oot carcasses fr om greater black-b acked gulls . Wils on Bull. 101: 494-496.
Sprunt , A., VI; Robertson, W. B., Jr .; Postupa lsky, S.; et al . (1973) Comparative producti vi ty of si x ba ld eagle popula tions . Trans. Nor th Am. Wildl. Nat. Resour . Conf . 38: 96-106.
Stalmaster , M. V. (1980) Management str ategies fo r winter ing ba ld eagles in t he Paci fic Northwest. In: Knight , R. L.; Allen, G. T.; Stalmaster , M. V.; et al ., eds. Proceedings of Washingt on ba ld eagle symp osium, June; Seattl e, WA. Seattl e, WA: The Nature Conservancy; pp. 49-67.
2-106 Bald Eagle

Stalmaster , M. V.; Gessaman , J. A. (1982) Food c ons umpt ion a nd energy r equir ements o f captive ba ld eagles. J. Wildl. Manage. 46: 646-654.
Stalmaster , M. V.; Gessaman , J. A. (1984) Ecologic al energetics and f oraging be havior of overwint ering ba ld eagles. Ecol. Monogr . 54: 407-428.
Stickel , L. F.; Chur a, N. J.; Stewar t, P. A.; et al . (1966) Bald eagle pesti cide relations . Trans. Nor th Am. Wildl. Natur. Resour . Conf . 21: 190-200.
Swenson, J. E. (1975) Ecology of th e bald eagle and os prey in Y ello wstone Nationa l Park [master 's th esis]. Bozeman, MT: Montana State Universi ty.
Swenson, J. E.; Alt, K. L.; Eng, R. L. (1986) Ecology of bald eagles in t he Greater Yellowsto ne ecosystem. Wildl. Monogr . 95: 1-46.
Todd, C. S.; Young, L. S.; Owen, R. B., Jr .; et al . (1982) Food ha bit s of bald eagles in Ma ine. J. Wildl. Manage. 46: 636-645.
United States F ish and Wildlif e Service (USFWS). (1989) Recovery plan: southeaster n states b ald eagle. Atlanta, GA: U.S. Fish and Wildlif e Service, Southeast Region.
Vermeer , K.; Morgan, K. H. (1989) Nest ing popula tion, nest si tes, and prey remains of bald eagles in Ba rkley Sound, Br it ish Colum bia. Nor thwest. Nat. 70: 21-26.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Watso n, J. W.; Garrett, M. G.; Anthony , R. G. (1991) Foraging e cology of bald eagles in t he Colum bia River estu ary. J. Wildl. Manage. 55: 492-499.
Weaver , J. (1980) Habitat man agement program: threatened and endangered plants an d animals. U.S. Forest Ser v., Bridge r-Teton Na t. Forest, Jackso n Hole, WY; 111 pp.
2-107 Bald Eagle

2.1.8. American Kestrel (falcons)
Order Falconi formes, Family Falconi dae. Falcons ar e the more str eamline d of th e raptor species, wi th long point ed wings bent b ack at th e wr ists an d large tai ls th at tap er at the tips . They cons ume many kinds of anim als inc luding in sects, repti les, smal l mammal s, and bir ds. Falcons ar e found in a var iety o f habitats, from ci ties to the most r emote areas. Strong f lie rs that achi eve high s peeds, fal cons r ange in s ize fr om th e Amer ican kestr el (27 cm bill t ip t o tail t ip) to the peregr ine fal con (41 to 51 cm).
Selected species
The Amer ican kestr el (Falco sparver ius), or sparrow hawk, is th e most co mmo n falcon in open and semi -open areas th roughout Nor th Amer ica. There are three recognize d subspecies: F. s. paulus (year -round r eside nt fr om Sout h Carolina to Flor ida and s out hern Alabama), F. s. peninsularis (year -round r eside nt of sout hern Baja Calif ornia), and F. s. sparver ius (wide spread and m igr atory) (Boha ll-Wood a nd Collopy , 1986). Predators of th e kestr el include large raptors such as gr eat horned owls, golde n eagles, and r ed-tai led hawks (M eyer and Balgooy en, 1987).
Body size. Weighing s light ly over one ten th of a kilogr am, th e kestr el is th e smal lest fal con native to the United States (B rown and Amadon, 1968). As fo r most raptors, femal es are 10 to 20 percent larger than males (Bloom , 1973; Craighead and Craighead, 1956). Kestr el body weight s vary seaso nally, wi th maxi mum weight (and f at deposi ts) b eing a chi eved in winter and minim um weight s in s ummer (Bloom , 1973; Gessaman and Haggas, 1987).
Habitat . Kestr els inha bit open deser ts, semi -open areas, the edges of groves (Brown and Amadon, 1968), and even ci ties (Nationa l Geogr aphic So ciety, 1987). In sever al areas, investi gators have found t hat male kestr els ten d to use woodla nd ope nings an d edges, while femal es ten d to utilize mo re open areas ch aracter ized by sh ort or sparse ground v egetation, partic ularly dur ing t he wint er (Koplin, 1973, ci ted in Mills , 1976; Meyer and Ba lgooy en, 1987; Mills , 1975, 1976; Smal lwood, 1987). In other areas, however , investi gators have found no s uch di ffer entiation ( Toland, 1987; Sfer ra, 1984). In Flor ida, kestr els appear to prefer sandhill c ommunit ies (particula rly pine /oak woodla nds ); these areas provide high- qualit y foraging ha bitat an d the majority o f avai lable nest si tes (Boha ll-Wood a nd Collopy , 1986). Kestr els are more likely to use habitats cl ose to centers of human acti vi ties th an are most o ther raptors (Fisch er et al ., 1984).
Food ha bi ts . Kestr els prey on a var iety o f smal l animals including inv ertebrates such as worms, spide rs, sco rpions , beetles, other large insects, amphibia ns and r ept iles such as frogs , lizards, and s nakes, and a wide var iety o f smal l- to medium -sized bir ds an d mammal s (Brown and Amadon, 1968; Mueller, 1987). Large insects, such as grasshoppe rs, are the kestr els' primary su mmer prey, although in t heir absence kestr els will s wi tch to smal l mammal s (Collopy , 1973) and birds (Brown and Amadon, 1968). In winter , smal l mammal s and birds co mprise mo st o f th e diet (Collopy and Koplin, 1983; Koplin e t al., 1980). Kestr els usually cach e their ver tebrate prey, often in clumps of grass o r in tree limbs and holes, to be retr ieved later (Collopy , 1977; Mueller, 1987; Rudolph, 1982; Toland,
2-109 Amer ican Kestr el

1984). Inver tebrate prey usually are eaten immed iately (Rudolph, 1982). In Flor ida, where smal l mammal s are scar ce and repti les are abunda nt , lizards are an impor tant compone nt of th e die t (Boha ll-Wood a nd Collopy , 1987). Kestr els fo rage by th ree differ ent tec hnique s: us ing ope n perches from whic h to spot and attack g round pr ey, hov ering in t he air to spot ground pr ey, and catc hing in sects o n the wing ( Rudolph, 1982, 1983).
Mol t. Females begin t heir molt dur ing inc ubation a nd c ompl ete i t by th e end of th e breeding seaso n. Males, who are respons ible fo r cap turing m ost of th e prey for th e fami ly, do not begin t heir molt unt il ne ar th e end of th e breeding seaso n (Smal lwood, 1988).
Migration. The Amer ican kestr el is a year -round r eside nt over mo st of th e Unit ed States, but is mi gratory over the northern-most p ortions of its range (Nationa l Geogr aphic Society, 1987). Because of th eir late mo lt, males mi grate and arrive at th e winter ing grounds l ater than femal es or immatu res (Smal lwood, 1988).
Breeding acti vi ties and social organization. Adult kestr els are solitary, excep t dur ing t he breeding seaso n, and maintain ter ritories even in winter (Brown and Amadon, 1968). Kestr els typ ical ly build t heir nests i n tree cavi ties, but have used hole s in t elephone pole s, buildings , or str eam banks wh en tree cavi ties are not avai lable (Brown and Amadon, 1968). Both parents p articipate in incubation, but th e femal e performs mo st o f th e inc ubation, while th e male prov ides her wi th food ( Brown and Amadon, 1968). Follow ing hatching, the male br ings th e major ity of th e prey to the nestlings (B rown and Amadon, 1968). After fledging, young kestr els remain dependent on their parents fo r food f or at least 2 to 4 addit iona l weeks (L ett an d Bird, 1987). Fledglings o ften perch and socialize wi th their siblings pr ior to dis persal (Lett an d Bird, 1987). In Flor ida, reside nt kestr els (paulus subspecies) mai ntain year -round pa ir bonds and joint ter ritories. The reside nt pairs have a co mpeti tive advantage over winter migrants (sparver ius subspecies) in their ter ritories (Boha ll-Wood a nd Collopy , 1986).
Home range and r esour ces. Although s ome in vesti gators have not noted ter ritorial defense (e.g., Craighead and Craighead, 1956), Mills (1975) demonstr ated that kestr els defend t erritories by int roduc ing c aptured bir ds int o ot her bir ds ' ter ritories. Wint er foraging t erritories range fr om a few h ectar es in producti ve areas (e.g., in California) (Meyer and Balgooy en, 1987) to hundr eds of h ectar es in less p roducti ve areas (e.g., Illinois , Michiga n) (Craighe ad and Cr aighe ad, 1956; Mills , 1975). Summer breeding t erritories probably follow th e same p atter n (Craighead and Craighead, 1956).
Popul ation de nsity . Although m uch smal ler than red-tai led hawks an d bald eagles, reported kestr el breeding popula tion de nsit ies can be si mila rly low (e. g., 0.0003 to 0.004 nests p er hectar e; see tab le).
Popul ation dy namics. Kestr els are sexu ally matu re in the fi rst b reeding seaso n after their birth (Carpenter et al ., 1987). Scarci ty o f su itable nest ing cavi ties probably l imi ts the size of kestr el popula tions in pa rts of th e Unit ed States (C ade, 1982). Three to four young may fl edge per nest p er year , but mo rtal ity o f juveniles in the fi rst year is high (60 to 90 percent) (Craighead and Craighead, 1956; Henny, 1972). Adult mo rtalit y can be low (e.g., 12 percent per year ) (Craighead and Craighead, 1956).
2-110 Amer ican Kestr el

�
�
�
Simi lar species (fr om general references)
� The peregr ine fal con ( Falco pe regr inus ), a rare reside nt of woods , mountains, and coasts, preys al most excl usively on birds. Though unc ommon, they can be found w int ering in m ost states, but rarely breeding. These large falcons (38 cm) h ave been reintroduced in some areas in the United States an d have nested in urban environments.
� The mer lin ( Falco columbariu s), larger (30 cm) th an the kestr el, can be found in a var iety o f habitats b ut nests i n open woods or woode d pr airies. Wint ering a long c oasts an d near ci ties of th e Great Plains, it primar ily eats bir ds.
� The prairie falcon (Falco mexi canus) also is larger (39 to 50 cm) th an the kestr el and inha bit s dr y, open count ry and pr airies. A year -round r eside nt of the wester n United States, prairie falcons prey ch iefly on birds and smal l mammal s.
General references
Cade (1982); Craighead and Craighead (1956); Nationa l Geogr aphic So ciety (1987); Brown and Amadon (1968).
2-111 Amer ican Kestr el
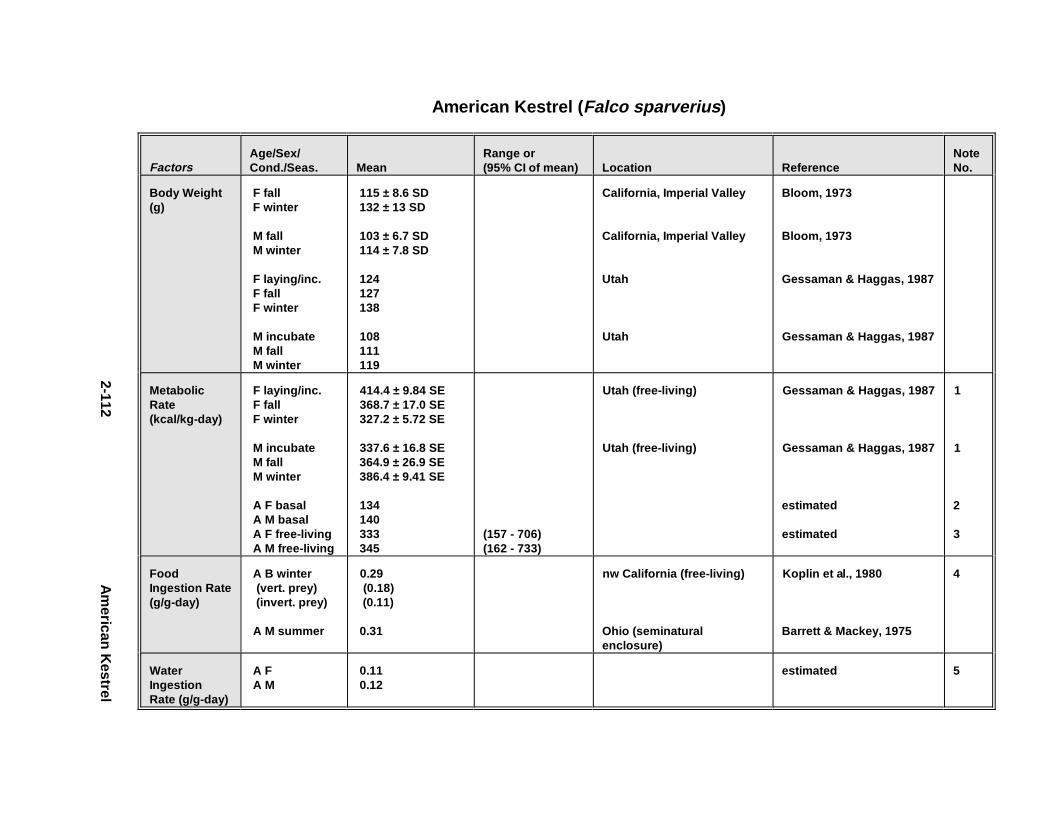
American Kestrel ( Falco s parverius )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight F fall 115 ± 8.6 SD California, Imperial Valley Bloom , 1973 (g) F winter 132 ± 13 SD
M fall 103 ± 6.7 SD California, Imperial Valley Bloom , 1973 M winter 114 ± 7.8 SD
F laying/ inc. 124 Utah Gessam an & Haggas, 1987 F fall 127 F winter 138
M incubat e 108 Utah Gessam an & Haggas, 1987 M fall 111 M winter 119
Metabol ic F laying/ inc. 414.4 ± 9.84 SE Utah (free-living) Gessam an & Haggas, 1987 1 Rate F fall 368.7 ± 17.0 SE (kcal /kg-day) F winter 327.2 ± 5.72 SE
M incubat e 337.6 ± 16.8 SE Utah (free-living) Gessam an & Haggas, 1987 1 M fall 364.9 ± 26.9 SE M winter 386.4 ± 9.41 SE
A F basal 134 est imated 2 A M basal 140 A F free-living 333 (157 - 706) est imated 3 A M free-living 345 (162 - 733)
Food A B wi nter 0.29 nw Cal ifornia (free-living) Kopl in et al., 1980 4 Ingest ion Rat e (vert. prey) (0.18) (g/g-day) (invert. p rey) (0.11)
A M sum mer 0.31 Ohio (seminatural Barrett & Mackey , 1975 encl osur e)
Water A F 0.11 est imated 5 Ingest ion A M 0.12 Rate (g/g-day)
2-112 A
merican
Kestrel

American Kestrel ( Falco s parverius )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Inhal ation A F 0.089 est imated 6 Rate (m /day) A M 0.0793
Surface Ar ea A F 267 est imated 7 (cm ) A M 2422
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
invertebrates 32.6 California/open ar eas, woods Meyer & Bal gooy en, 1987 mammals 31.7 bi rds 30.3 (% wet weight of prey rept iles 1.9 obser ved capt ured) other 3.5
vertebrates 49 Flor ida/dry pine-oak Bohal l-Wood & Col lopy , (primarily lizards) woodl ands ( sandh ill) 1987
invertebrates (% wet weight of prey obser ved capt ured)
Coleopt era 10.8 California/hayfields, past ure Col lopy & Kopl in, 1983 other invertebrates 14.2 frogs ( Rana aurora) 8.0 (% wet weight of prey other herpetofauna 12.2 obser ved capt ured) Microtus cal ifornicus Sorex vagrans other mammals
30.2 9.4
11.5
51
2-113 A
merican
Kestrel

American Kestrel ( Falco s parverius )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Territory A F wi nter 31.6 ± 10.7 SD 18.7 - 42.0 California/open ar eas, Meyer & Bal gooy en, Size (ha) A M winter 13.1 ± 2.0 SD 9.7 - 14.8 woods 1987
A B w inter 154 < 452
A B sum mer 202 ± 131 SD 41 - 500 Wyoming/grassl ands, Craighead & Cr aighead,
A B sum mer 131 ± 100 SD 21 - 215 Michi gan/woodl ots, fields Craighead & Cr aighead,
Illin ois/agric ultu ral area Mills , 1975
forests
1956
Popul ation pairs 0.0026 nest s/ha 0.0023 - 0.0031 Missour i/urban Toland & El der, 1987 Densi ty summer 0.0004 nest s/ha 0.0003 - 0.0006 Missour i/rural Toland & El der, 1987
pairs sum mer 0.0035 pairs/ha Wyoming/grassl ands, forest Craighead & Cr aighead,
pairs bird s/ha: sum mer 0.0007 ± 0.00004 SD 0.0005 - 0.0012 s Michi gan/fields, woodl ots Craighead & Cr aighead,
B B f all A B wi nter A B spr ing
0.0005 ± 0.0001 0.0005 - 0.0006 1956 0.0010 ± 0.0002 0.0008 - 0.0011
1956
Clutch 4.3 California/juni per, Bloom & Hawks, 1983 Size sagebr ush
4 to 5 3 - 7 NS/NS Brown & Am adon, 1968
Clutches/Year 1 Quebec, Canada/captiv e Carpenter et al., 1987
Days 33.7 ± 0.33 SE 33 - 35 Maryland/capt ive Porter & Wiemeyer, 1972 Incubat ion Brown & Am adon, 1968
29 to 30 NS/NS
Age at 27.4 days 26 - 30 days Maryland/capt ive Porter & Wiemeyer, 1972 Fledgi ng
Range
1956
SD SD
2-114 A
merican
Kestrel
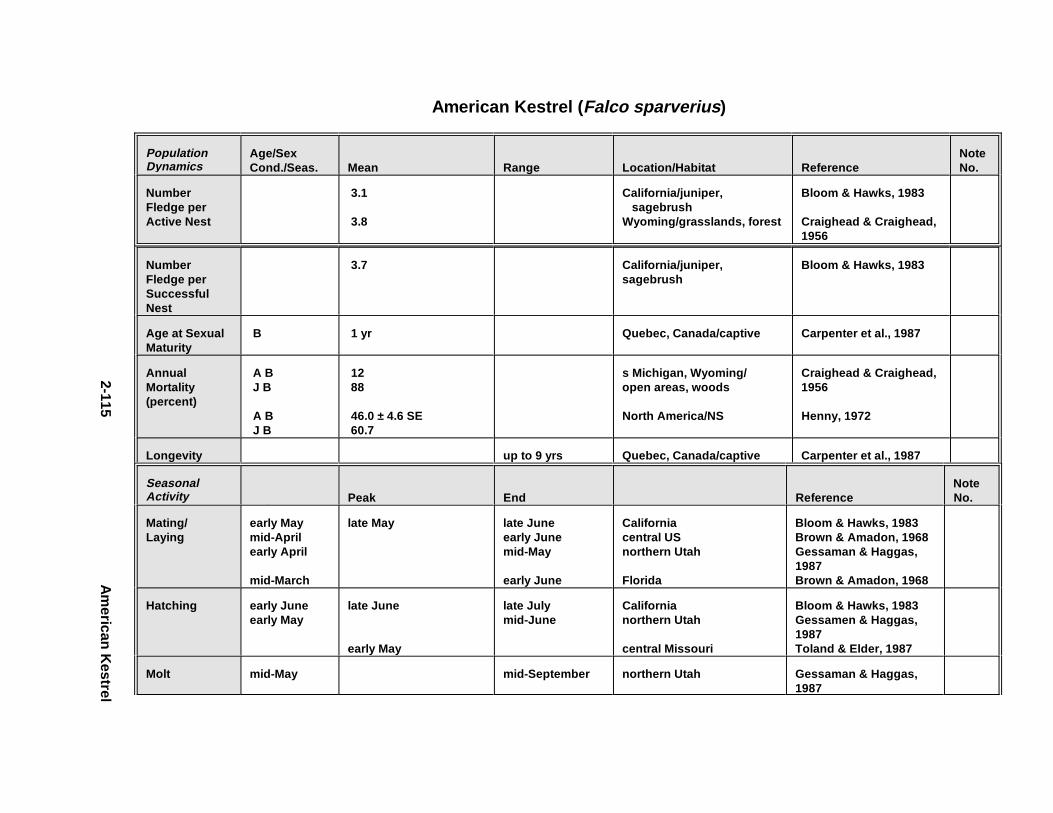
American Kestrel ( Falco s parverius )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Number 3.1 California/juni per, Bloom & Hawks, 1983 Fledge per sagebr ush Act ive Nest 3.8 Wyoming/grassl ands, forest Craighead & Cr aighead,
1956
Number 3.7 California/juni per, Bloom & Hawks, 1983 Fledge per sagebr ush Successf ul Nest
Age at Sexual B 1 yr Quebec, Canada/captiv e Carpenter et al., 1987 Maturity
Annual A B 12 s Michi gan, Wyoming/ Craighead & Cr aighead, Mortality J B 88 open ar eas, woods 1956 (percent )
A B 46.0 ± 4.6 SE North Am erica/NS Henny , 1972 J B 60.7
Longevi ty up t o 9 yrs Quebec, Canada/capt ive Carpenter et al., 1987
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/ early May late May late June California Bloom & Hawks, 1983 Laying mid-Apr il early June cent ral US Brown & Am adon, 1968
early Apr il mid-May nor thern Utah Gessam an & Haggas,
mid-March early June Flor ida Brown & Am adon, 1968 1987
Hatching early June late June late Jul y California Bloom & Hawks, 1983 early May mid-June nor thern Utah Gessam en & Haggas,
early May cent ral Missour i Toland & El der, 1987 1987
Molt mid-May mid-September nor thern Utah Gessam an & Haggas, 1987
Range
2-115 A
merican
Kestrel

American Kestrel ( Falco s parverius )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Migration fall early early Novem ber nor thern Utah Gessam an & Haggas,
spr ing early March sout h Michi gan Craighead & Cr aighead, September 1987
mid-Apr il Wyoming Craighead & Cr aighead, 1956
1956
1 Invest igators est imated val ues f rom time-act ivity budget studi es of kest rels in the field and r ates of energy expendi ture dur i ng di fferent act ivities measur ed in the labor atory.
2 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom winter measur ements by Gessam an and Haggas ( 1987). 3 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom winter measur ements by Gessam an and Haggas ( 1987). 4 Author s obser ved pr ey capt ured dai ly, and est imated total wet-weight prey intake usi ng m easur ed or repor ted weight s for ident ifiable prey and
est imated weight s for uni dent ifiable invertebrate prey (also, assum ed kest rel weighed 119 g) . Also, see Chapt ers 3 and 4 f or methods by est imating f ood i ngest ion r ates.
5 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom winter measur ements by Gessam an and Haggas ( 1987). 6 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom winter measur ements by Gessam an and Haggas ( 1987). 7 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, ci ted in Walsberg and Ki ng, 1978) and body weight s f rom winter measur ements by
Gessam an and Haggas ( 1987).
2-116 A
merican
Kestrel

References (including A ppendix)
Barrett, G. W.; Mackey, C. V. (1975) Prey sel ect ion a nd c alor ic ing estion r ate of cap tive Amer ican kestr els. Wilson Bull. 87: 514-519.
Bird, D. M.; Clark, R. G. (1983) Growth of body co mpone nts in pa rent- and ha nd-reared captive kestr els. Raptor Res. 17: 77-84.
Bloom , P. H. (1973) Seasonal var iation in body weight of sp arrow hawks i n California. Wester n Bird Bander 48: 17-19.
Bloom , P. H.; Hawks, S. J. (1983) Nest b ox use and reproduc tive biology of th e Amer ican kestr el in Lassen Count y, Calif ornia . Raptor Res. 17: 9-14.
Boha ll-Wood, P.; Collopy , M. W. (1986) Abunda nce and ha bitat sel ect ion of two Amer ican kestr el subspecies in north-central Flor ida. Auk 103: 557-563.
Boha ll-Wood, P. G.; Collopy , M. W. (1987) Foraging be havior of sout heaster n Amer ican kestr els in relation t o habi tat u se. Raptor Res. 6: 58-65.
Brown, L.; Amadon, D. (1968) Eagles, hawks, and falcons of th e world. New York, NY: McGraw Hill Book Co.
Cade, T. J. (1982) The falcons of th e world. Ithaca, NY: Cornell Universi ty Pr ess.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Carpenter , J. W.; Gabel, R. R.; Wiemeyer , S. N.; et al . (1987) Captive breeding. In: Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W.; et al., eds. Raptor management technique s manua l. Washingt on, DC: Nationa l Wildlif e Federation; pp. 350-355.
Collopy , M. W. (1973) Predatory effi ciency o f Amer ican kestr els wi ntering in nor thwester n Calif onia . Raptor Res. 7: 25-31.
Collopy , M. W. (1977) Food caching by femal e Amer ican kestr els in winter . Condor 79: 63-68.
Collopy , M. W.; Koplin, J. R. (1983) Diet, capture success, and mode of hunt ing by femal e Amer ican kestr els in winter . Condor 85: 369-371.
Craighead, J. J.; Craighead, F. C. (1956) Hawks, owls and wildlif e. Harrisbur g, PA: The Stack pole Co. and Washingt on, DC: Wildlif e Management Insti tute.
Duke, G. E.; Evanson, O. A.; Jegers, A. A. (1976) Meal to pellet intervals in 14 species of captive raptors. Comp. Bioc hem. Physiol. 53A: 1-6.
2-117 Amer ican Kestr el

Duke, G. E.; Mauro, L.; Bird, D. M. (1987) Physiology . In: Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W.; et al., eds. Raptor management tec hnique s manua l. Washingt on, DC: Insti tute for Wildlif e Research, Nationa l Wildlif e Federation; Sci . Tech. Ser. No. 10; pp. 262-267.
Enderson, J. H. (1960) A popula tion s tudy of th e sparrow hawk i n east-cen tral Illinois . Wils on Bull. 72: 222-231.
Fischer, D. L.; Ellis , K. L.; Meese, R. J. (1984) Winter habitat sel ect ion of diur nal raptors in central Utah. Raptor Res. 18: 98-102.
Gessaman , J. A. (1979) Premigratory fat i n the Amer ican kestr el. Wilson Bull. 91: 625-262.
Gessaman , J. A.; Haggas, L. (1987) Energetics o f th e Amer ican kestr el in northern Utah. Raptor Res. 6: 137-144.
Henny, C. J. (1972) An analysi s of th e popula tion dy namics of sel ected avian species wi th special reference to changes during t he mode rn pesti cide era. Washingt on, DC: Bur . Spor t. Fish. Wildl. ; Wildl. Res. Rep. 1.
King, J. R. (1974) Seasonal alloc ation of ti me and energy r esour ces i n birds. In: Paynter , R. A., Jr., ed. Avian energetics. Cambridge , MA: Nuttall O rnit hol. Club; pp. 4-70.
Koplin, J. R. (1973) Differ ential habitat u se by sexes o f Amer ican kestr els wi nter ing in northern California. Raptor Res. 7: 39-42.
Koplin, J. R.; Collopy , M. W.; Bamman n, A. R.; et al . (1980) Energetics o f two winter ing raptors. Auk 97: 795-806.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lett, D. W.; Bird, D. M. (1987) Postfl edging be havior of Amer ican kestr els in southwester n Quebec. Wils on Bull. 99: 77-82.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Meyer , R. L.; Balgooy en, T. G. (1987) A stu dy and implic ations of habitat sep aration by sex of wint ering A mer ican kestr els (Falco sparver ius L.). Raptor Res. 6: 107-123.
Mills , G. S. (1975) A winter popula tion s tudy of th e Amer ican kestr el in central Ohio. Wils on Bull. 87: 241-247.
Mills , G. S. (1976) Amer ican kestr el sex r atios and habitat sep aration. Auk 93: 740-748.
2-118 Amer ican Kestr el

Mueller, H. C. (1987) Prey sel ect ion by kestr els: a review. Raptor Res. 6: 83-106.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Porter , R. D.; Wiemeyer , S. N. (1972) Reproducti ve patter ns in captive Amer ican kestr els (sparrow hawks). Condor 74: 46-53.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Rudolph, S. G. (1982) Foraging s trategies of Amer ican kestr els during br eeding. Ecology 63: 1268-1276.
Rudolph, S. G. (1983) Aerial insect-catc hing by A mer ican kestr els. Condor 85: 368-369.
Sfer ra, N. J. (1984) Habitat sel ect ion by th e Amer ican kestr el (Falco sparver ius) and red-tai led hawk (Buteo jamaicensis) wint ering in Ma dis on Count y, Kentucky. Raptor Res. 18: 148-150.
Smallw ood, J. A. (1987) Sexual segregation by habitat i n Amer ican kestr els wi nter ing in sout hcentral Flor ida: vegetati ve str uctu re and responses to differ ential prey availa bilit y. Condor 89: 842-849.
Smallw ood, J. A. (1988) A mech anism o f sexu al segregation by habitat b y Amer ican kestr els (Falco sparver ius) wint ering in s out h-central Flor ida. Auk 105: 36-46.
Sparrowe, R. D. (1972) Prey-catc hing be havior in t he sparrow hawk. J. Wildl. Manage. 36: 279-308.
Toland, B. R. (1984) Unus ual pr edatory and caching be havior of Amer ican kestr els in central Misso uri. Raptor Res. 18: 107-110.
Toland, B. R. (1987) The effect o f veg etati ve co ver on foraging s trategie s, hunt ing s uccess and ne sting dis tr ibut ion of A mer ican kestr els in central Misso uri. Raptor Res. 21: 14-20.
Toland, B. R.; Elder, W. H. (1987) Influence of nest-b ox placemen t and densi ty o n abunda nce and pr oduc tivi ty of A mer ican kestr els in central Misso uri. Wilson Bull. 99: 712-717.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Wing, L.; Wing, A. H. (1939) Food c ons umpt ion of a sparrow hawk. Condor 41: 168-170.
2-119 Amer ican Kestr el

Zar, J. H. (1968) Standard metabolis m comparisons b etween orders of birds. Condor 70: 278.
Zar, J. H. (1969) The use of th e allometr ic mo del for avian stan dard metabolis m - body weight r elations hips . Comp. Bioc hem. Physiol. 29: 227-234.
2-120 Amer ican Kestr el

2.1.9. Northern Bobw hite (quail)
Order Gallif ormes, Fami ly Phasiadinae. Quail are ground- dwelling bir ds wi th shor t, heavy bills ad apted for fo raging on t he ground f or seeds and insects. Most sp ecies inha bit brush, abandone d f ields , and ope n woodla nds ; some inha bit parklands . Quail and m ost other gallin aceous bir ds ar e poor flyers that seldom l eave th e ground a nd do not migr ate. All species of th is fami ly gather in coveys (i .e., flocks o f var ying s ize) dur ing s ome part of the year . Quail range in size from Montezuma's quail (22 cm bill t ip t o tail t ip) to the mountain and Gambel's quail (28 cm); sexes ar e simi lar in size but differ in appearance.
Selected species
The nor thern bobw hit e (Colin us virg inianus) feeds mainly on seeds by gleaning on the ground a nd low veg etation. It ranges from sout heaster n Wyoming, east to southern Minne sota and across to sout hern Maine, sout h through t he central and easter n Unit ed States to easter n New Mexico in the west an d to Flor ida in t he east (Amer ican Ornit hologi sts' Union, 1983). It is th e most wi despread of th e North Amer ican quail and used to be very co mmo n, particularly east o f th e Rocky Mount ains . Over the past th ree decades, however , popula tions h ave been declining t hroughout its range (B rennan, 1991).
Body size. Nor thern bobw hit e are average-sized quail (25 cm). Wild bobw hit es typic ally weigh b etween 150 and 200 g depending on lo cation a nd seaso n (see tab le), while commer cially bred sto ck u sually exceed 200 g and may r each 300 g or more (Brenner and Reeder, 1985; Koerth and Guthery, 1991). Males and femal es are simi lar in size, and weights ten d to increase wi th lati tude and toward the west co ast o f th e United States (Hamilton, 1957; Rosene, 1969; Roseberry and Klimstr a, 1971). Females are heaviest i n the spring a nd s ummer when they are laying e ggs ; males are ligh test at th is ti me of year (Hamilton, 1957; Roseberry and Klimstr a, 1971). Juvenile s tend t o weigh s light ly l ess th an adult s through w int er (Hamilt on, 1957; Roseberry and Klimstr a, 1971). Koerth and Guthery (1987) found bot h males and femal es to maintain between 9 and 11 percent body fat (as a percentage of dry body weight ) throughout th e year in southern Texas; more northern popula tions may mai ntain highe r body fat r atios, particularly just p rior to breeding ( McRae and Dimmi ck, 1982).
Habitat . Dur ing t he breeding seaso n, grassl ands, idle fi elds , and p astu res are the prefer red nesting ha bitat, and bobw hit e often nest i n large clumps of grasses (R oseberry and Klimstr a, 1984). Shade, open herbaceous co ver , and green and growing v egetation a re requir ed for suit able n est si tes (L ehmann, 1984). Bobw hites fo rage in areas wi th open vegetat ion, some bare ground, and light li tter (Stodda rd, 1931). Nearby dry powdery soils are impor tant for dus t bathing ( Johns gard, 1988). Shrubby thi ckets u p to 2 m high a re used for co ver dur ing m idda y (Schroeder, 1985). Although t heir range is exten sive, nor thern bobw hit e reproduc e poor ly in t he arid wester n portions of th eir range and dur ing drought s elsewh ere (Schroeder, 1985). During t he wint er, they requir e woode d cover wi th unde rsto ry for dayti me co ver , preferably adjacent to open fields fo r foraging ( Yoho a nd Dimmi ck, 1972). They ten d to roost at night in m ore open habitats wi th short and sparse vegetat ion (Schroeder, 1985). In the more northern lati tudes, cover and food c an be lim ited dur ing t he wint er (Ros ene, 1969). Changes in land use, primar ily
2-121 Northern Bobw hit e

the dis tr ibut ion of far ms and f arming methods , have el iminated large areas of bobw hit e habitat i n the last th ree decades (Brennan, 1991).
Food ha bi ts . Bobw hites fo rage during t he day, pr imarily on t he ground or in a light litter layer less th an 5 cm d eep (Rosene, 1969). Seeds fr om weed s, woody plants, and grasses co mprise th e majority o f th e adult bobw hit e's die t throughout th e year (Handle y, 1931; Bent, 1932; Lehmann, 1984), although in w int er in t he sout h, green vegetation ha s been found t o dom inate th e plant mater ials in their diet (Campbell-Kisso ck et al ., 1985). Insects an d other inver tebrates can comprise up to 10 to 25 percent of th e adult s' die t dur ing t he spring a nd s ummer in more northerly areas and year -round in t he sout h (Campbell-Kisso ck et al ., 1985; Handley, 1931; Lehmann, 1984). Insects co mprise th e bulk of th e chicks' diet; up to 2 or 3 weeks o f age chicks may co nsume almost 85 p ercent insects, the remainde r of th e die t cons is ting of berries and seeds (Handley, 1931). Most insects co nsumed by bobw hit e chi cks ar e very smal l, less th an 8 mm i n length and 0.005 g (Hurst, 1972). Juvenile bobw hit e, on t he other hand, may co nsume only 25 pe rcent in sects, the remainde r of th eir die t be ing f ruit and seeds (Handley, 1931). Quail consume l ittl e grit. Korsch gen (1948) found gr it in only 3. 4 percent of over 5,000 crops exami ned, and agreed with Nestl er (1946) that hard seeds can replace grit as th e grinding a gent for nor thern bobw hit e.
In some areas, bobw hites ap parently can acquir e their daily water needs fr om dew, succu lent plants, and insects (St odda rd, 1931); in more arid areas or in times o f drought , however , northern bobw hit e need surface water for drinking ( Johns gard, 1988; Lehmann, 1984; Prasad and Guthery, 1986). Females need more water than males during t he breeding seaso n, and both sexes may r equir e more water in the winter than in the summer when their diet is mo re restr icted to seeds wi th low water content (Koerth and Guthery, 1990). Measurements o n captive quail have indi cated a daily water requir ement of up to 13 percent of th eir body mass (see tab le); however , water intake r equir ements fo r free-ranging birds may b e highe r, perhaps 14 to 21 percent of body mass p er day (Koerth and Guthery, 1990). In the absence of adequate water , femal es may fai l to reproduc e (Koerth and Guthery, 1991).
Dustb athing. Quail f requent ly dus tbathe, although t he reason for the behavior is cdebated. They scr atch in dry dirt or dust, toss th e dust u p into their feath ers, rub their
head and sides in the dust, and then shake th e dust fr om th eir plumage (Borchelt and Duncan, 1974). Experiments b y Driver et al . (1991) indi cate th at inge stion of mater ials preened from feath ers and direct d ermal uptake can be signif icant expos ure pathways fo r quail expos ed to aerial appli cation of p esti cides. Dust b athing m ight , therefore, prov ide a signif icant expos ure rout e for bobw hites u sing c ontami nated soils .
Mol t. Juveniles attai n adult plum age dur ing t heir fi rst fall m olt at about 3 to 5 months of age (Hamilton, 1957; Stodda rd, 1931). Adult s unde rgo a co mpl ete prebasic
cStoddard ( 1931) and ot hers have suggest ed that dust bathing hel ps to cont rol ectoparasi tes;Borchel t and Duncan ( 1974) suggest that dust bathing hel ps cont rol the amount of oi l on t he qua ils' feathers .
2-122 Northern Bobw hit e

molt in the late su mmer and fal l into winter plum age; in s pring, a lim ited renewal of feath ers around t he head and t hroat pr ov ides the breeding plum age (Bent, 1932).
Migration. The nor thern bobw hit e is a year -round r eside nt over its enti re range but may d isperse local ly to a differ ent co ver typ e or alti tude wi th the changing seaso n (Lehmann, 1984). Most wi nter in woode d or brushy ar eas, returning t o more open habitats in s pring f or th e breeding seaso n (Lehmann, 1984; Rosene, 1969). Popula tions n esting a t highe r elevat ions tend t o move to low er ground w here the wint ers are less sever e (Stodda rd, 1931). The more southerly popula tions may b e more sedentary; in a stu dy in Flor ida, nor thern bobw hit e were found no f urther th an 1 km from wh ere they were banded, and 86 percent were found w ithin 400 m fr om th eir banding s ite ov er a 1- to 5-year period (Smith et al ., 1982).
Breeding acti vi ties and social organization. Nor thern bobw hit e build n ests o n the ground in ope n woodla nds or in or around f ields used for fo raging. Most nests ar e constr ucted in grassy g rowth near open ground, often in areas wi th scatter ed shrubs and herbaceous growth (Klimstr a and Roseberry, 1975; Stodda rd, 1931). Both the male and femal e scr ape out a sau cer -shaped depression in t he ground 2 to 6 cm de ep and 10 to 12 cm across, lining it wi th dead grasses fr om th e previous year 's growth (Bent, 1932; Rosene, 1969). They lay large clutch es, 12 to 30 eggs, which one or both parents i ncubate for approximatel y 23 days (L ehmann, 1984; Simpson, 1976). As a general rule, clutch size and nest su ccess b oth decrease as th e seaso n progresses (R oseberry and Klimstr a, 1984). Family unit s, cons is ting of bot h the male and femal e as wel l as th e offsp ring, someti mes remain intact th rough t he summer , but mo re often , one or both parents ar e lost to predation ( some femal es leave th eir brood t o the male and be gin a not her), and ot her pairs or indiv idua l adult s may adopt chi cks fr om other broods (L ehmann, 1984). By fal l, northern bobw hites o f al l ages gather in larger coveys fo r the fal l and winter . The quail remain in coveys u nti l the next sp ring, when they dis perse as mating seaso n begins (L ehmann, 1984; Roseberry and Klimstr a, 1984). Coveys o f northern bobw hit e tend t o average 10 to 12 or 15 birds (up to 30) (Johns gard, 1988; Lehmann, 1984; Rosene, 1969). When roost ing in winter , the birds in a covey fo rm a smal l ci rcle on the ground unde r a tree or in t hic k br ush, wi th heads facing ou twar d and their bodie s clos ely packed to conserve heat.
Home range and r esour ces. In the breeding seaso n, the bobw hit e's hom e range inc lude s foraging a reas, cover , and the nest si te and may en compass sever al hectar es. Mated males and incubating femal es have th e smal lest sp ring a nd s ummer home ranges; bachelor males and post-n est ing m ales and femal es have mu ch larger foraging r anges (see table ). Bobw hit e tend t o use a por tion of th eir hom e range mo re int ensively than the remainde r of th e range (Urban, 1972). In the fal l and winter , the range of each bobw hit e covey mu st i nclude ad equate open foraging a reas and cover , typ ical ly sh rubby or woody thickets (R osene, 1969). Each covey may u tilize an area of sever al hectar es, although a s in summer , there tend to be acti vi ty cen ters wh ere the quail spend most o f th eir time (Yoho and Dimmi ck, 1972).
Popul ation de nsity . Bobw hit e dens ity depends on f ood a nd c over availa bilit y and var ies fr om year to year as wel l as fr om one locat ion t o anot her (Ros eberry and Kli mstr a, 1984). Densi ties are highest at th e end of th e breeding seaso n in the fal l. In the
2-123 Northern Bobw hit e

�
�
�
�
�
southeast, densi ties may r each values as high a s 7.5 bir ds (adult s and juv enil es) per hectar e, although average values of 2 to 3 may b e more commo n in these areas (Guthery, 1988; Lehmann, 1984; Smith et al ., 1982). Winter and spring de nsit ies between 0.1 and 0.8 birds per hectar e have been recorded in the spring f urther nor th (Roseberry et al., 1979).
Popul ation dy namics. Bobw hites attemp t to rear one or two broods per year (up to three in the south) (Bent, 1932; CKWRI, 1991; Stanford, 1972b). Bobw hit e clut ch sizes are generally smalle r in m ore sout herly popula tions (Ros eberry and Kli mstr a, 1984) and smalle r as the breeding seaso n progresses i n any given locale (Lehmann, 1984; Simpson, 1976). Predation is a major cause of n est l oss; once hatch ed, chicks l eave th e nest immed iately to follow bot h or one parent (L ehmann, 1984; Roseberry and Klimstr a, 1984). Juvenile s can surviv e wi thout parental care after about 6 weeks o f age (Lehmann, 1984). They reach matu rity b y 16 weeks o f age in the laboratory al though t hey cont inue to gain weight th rough a bout 20 weeks ( Moor e and Cain, 1975), and they may r equir e 8 to 9 months to matu re in the wild ( Johns gard, 1988; Jones and Hughe s, 1978). Adult mo rtalit y as well a s juv enile mo rtalit y is high, wi th 70 to 85 percent of bir ds su rviv ing l ess th an 1 year (Brownie et al ., 1985; Lehmann, 1984); thus, the bulk of th e popula tion t urns ov er each year .
Simi lar species (fr om general references)
� Calif ornia quail (Callip epla calif ornica), also known as val ley quail, are simi lar in size (25 cm) to the bobw hit e and a lso gather in c oveys d uring autumn and w int er. They are common in ope n woodla nds , brushy foot hills , str eam val leys, and subur bs, usually near permanent surface water s; however , their range is restr icted largely to the wester n coastal States an d Baja Calif ornia .
� Gambel's quail (Callip epla gambelii ) is larger (28 cm) th an the bobw hit e, and is a resident of th e southwester n deser t scr ubla nds , usually near permanent surface water s. It also gathers in coveys i n winter .
� The scal ed quail (Callip epla squamata), simi lar in size (25 cm) to the bobw hit e, is r estr icted to the mesas, plateaus, semi deser t scr ubla nds , and grassl ands mi xed wi th scr ub, primar ily of wester n Texas, New Mexico, and Mexico.
� Mount ain qua il (Oreortyx p ictu s) are found in t he chapparal, brushy r avine s, and mountain slopes of th e west u p to 3,000 m. These also are large quail (28 cm). During t he fall, they gather in c oveys an d descen d to lower alti tudes for the winter .
� The Mont ezuma quail (Cyr tonyx mo ntezumae), former ly kn own as th e harlequin qua il, is a smal l (22 cm), secr etive resident of th e southwest. This species is usually found in gr assy unde rgrowth of junipe r or oak-pine woodla nds .
2-124 Northern Bobw hit e

General references
Johns gard (1988); Lehmann (1984); Nationa l Geogr aphic So ciety (1987); Rosene (1969); Roseberry and Klimstr a (1984); Stodda rd (1931).
2-125 Northern Bobw hit e

Northern Bobw hite ( Colinus vir ginianus )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight A B f all 189.9 ± 3.28 SE Kansas Robel , 1969 (g) A B w inter 193.9 ± 4.56 SE
A B spr ing 190.0 ± 4.98 SE
A M winter 181 Illinoi s Roseber ry & Kl imstra, 1971 A M sum mer 163 A F w inter 183 A F sum mer 180
A M winter 161 west Rio Grande, Texas Guthery et al., 1988 A M sum mer 154 A F w inter 157 A F sum mer 157
at hatching 6.3 sout hwest Georgia/bot h Stoddar d, 1931 day 6 9 - 10 (0.5 - 0.75 g/day) capt ive and w ild bi rds day 10 10 - 13 living i n farms, woods, day 19 20 - 25 (1.5 g/day) and t hicket s day 32 35 - 45 day 43 55 - 65 (1.75 g/day) day 55 75 - 85 day 71 110 - 120 (1.75 - 2.0 g/day) day 88 125 - 150 day 106 140 - 160
J B f all 174.0 ± 3.49 SE Kansas Robel , 1969
(weight gain:)
Body Fat A M winter 15.5 ± 2.8 SD Tennessee McRae & Dimmick, 1982 (% dry weight ) A M spr ing 8.8 ± 3.2 SD
A F w inter 13.8 ± 2.7 SD A F spr ing 12.7 ± 2.4 SD
Body Fat A M winter 10.2 ± 0.6 SE 9.0 - 11.9 sout hern Texas/capt ive Koer th & Guthery, 1987 (% dry weight ) A M spr ing 7.9 ± 0.2 SE 6.5 - 10.0 (cont inued) A F w inter 10.6 ± 0.8 SE 8.3 - 19.9
A F spr ing 9.7 ± 0.3 SE 7.7 - 11.2
2-126 N
orth
ern Bobw
hite

Northern Bobw hite ( Colinus vir ginianus )
2-127 N
orth
ern Bobw
hite
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Egg Weight 9.3 ± 0.3 SE Texas Koer th & Guthery, 1991 (grams) 8.6 8.0 - 10.2 sout hwest Georgia Stoddar d, 1931
Metabol ic Rate A F nonbr eed 183.3 Nebraska/capt ive Case, 1982 1 (kcal /kg-day) A F laying 243.9
A M basal 129 est imated 2 A F basal 125
A M free-living 320 (151 - 677) est imated 3 A F f ree-living 311 (147 - 659)
Food Ingest ion A B wi nter 0.093 ± 0.0032 SE sout hern Texas/capt ive Koer th & Gut hery, 1990 4 Rate (g/g-day) A B spr ing 0.067 ± 0.0021 SE
(kcal /kg-day) A B w inter 587 Kansas Robel , 1969 5
A B sum mer 0.079 ± 0.0061 SE A B f all 0.072 ± 0.0017 SE
A B f all 657 A B spr ing 519
Water A M sum mer 0.10 ± 0.023 SD sout hern Texas/capt ive Koer th & Guthery, 1990 Ingest ion Rat e A F sum mer 0.13 ± 0.037 SD (g/g-day)
A M summer 0.11 est imated 6 A F summer 0.10
Inhal ation Rat e A M sum mer 0.10 est imated 7 (m /day) A F summer 0.113
Surface Ar ea A M sum mer 298 est imated 8 (cm ) A F sum mer 3202

Northern Bobw hite ( Colinus vir ginianus )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
adul ts: (total plant foods) (87.2) (78.7) (79.7) (96.8) sout heast ern Uni ted Handl ey, 1931 misc. seeds 21.1 6.0 11.1 2.6 States/NS other seeds: legum es 15.2 3.9 10.1 31.5 (% vol ume; crop and gi zzard senna 7.2 0.4 0.2 12.8 cont ents) cul tivated pl ants 2.1 2.1 5.3 2.6 grasses 3.1 11.3 26.0 2.3 sedges 1.1 1.2 2.4 1.1 mast 14.1 0.2 0.5 28.0 spurges 0.1 1.2 5.5 0.4
frui ts 11.1 45.8 11.3 9.5 forage pl ants 12 0.3 0.3 5.2
(total animal foods) (12.8) (19.6) (20.3) (3.2) grasshopper s 3.2 7.5 16.6 2.4 bugs 2.8 4.4 0.6 0.1 beetles 4.6 6.3 0.8 0.2
adul ts: seeds of weeds 43.64 33.7 30.0 34.3 sout h Texas/ semi-prairie, Lehm ann, 1984 seeds of woody 4.03 20.5 39.7 9.5 brushl and plants 13.2 24.8 0.7 7.2
seeds of grasses 3.7 1.9 8.3 15.4 (% dry vol ume; crop cul tivated gr ains, etc. 27.4 4.9 3.4 10.3 cont ents) greens 8.03 14.2 17.9 23.3 insect s
adul ts: seeds of forbs 3.5 19.0 12.0 sout hwest Campbel l-Kissock et al., 1985 seeds of grasses 51.7 42.9 4.9 Texas/grassl ands seeds/ frui ts of drought condi tions
woody plants 9.7 - 1.4 uni dent ified seeds 4.6 - 2.3 (% wet vol ume; crop green veget ation 4.8 1.8 72.4 cont ents) invertebrates 25.8 36.2 6.5
2-128 N
orth
ern Bobw
hite

Northern Bobw hite ( Colinus vir ginianus )
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Home Range summer: Size (ha/bi rd) A B 3.6 Iowa/State game area Crim & Sei tz, 1972
(ha/covey) win ter:
A M mated 7.6 ± 5.0 SD sout h Illinoi s/idle farms Urban, 1972 A M unm ated 16.7 ± 9.5 SD woods, brush, cor nfields A F nest ing 6.4 ± 4.0 SD A F post -nest 15.6 ± 9.1 SD
B B 6.8 ± 2.9 SD 4.0 - 11.7 Tennessee/ woods, old f ields Yoho & Di mmick, 1972
B B 15.4 12.1 - 18.6 sout h Illinoi s/NS Barthol omew, 1967 cul tivated f ields
Popul ation B B fall 0.21 ± 0.0031 SE sout h Texas/ upl and Guthery, 1988 Densi ty rangel and (N/ha) B B spr ing 0.10 ± 0.0003 SE
B B fall 0.63 ± 0.24 SD 0.28 - 0.92 south Illin ois/agric ultu ral Roseberry et al., 1979 B B spr ing 0.24 ± 0.05 SD 0.18 - 0.33
B B f all 5.0 ± 0.30 SE sout h Texas/ mixed br ush Guthery, 1988 B B spr ing 2.2 ± 0.21 SE rangel and
B B w inter 0.63 ± 0.18 SD 0.37 - 0.88 Sout h Carolina/farms, Rosene, 1969
B B w inter 2.25 ± 1.16 SD 0.6 - 3.9 Smith et al., 1982 B B wi nter 3.65 ± 2.22 SD 1.7 - 7.6 Florida/pine woods
woods
Clutch Si ze 12.9 4 - 33 sout h Texas/ prairie, brush Lehmann, 1984
March 25.0 sout hwest Georgia/pine Simpson, 1976 August 9.4 woods, farms
13.7 ± 3.28 SD 6 - 28 Illinoi s/agr icul tural Roseber ry & Kl imstra, 1984
Clutches/ Year 1 0 - 3 NS/NS CKWRI, 1991
Days 23 21 - 25 sout h Texas/ prairie, brush Lehmann, 1984 Incubat ion
2-129 N
orth
ern Bobw
hite

Northern Bobw hite ( Colinus vir ginianus )
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Percent Nests 17.5 15.4 - 19.0 sout hwest Georgia/pine Simpson, 1976 Successf ul woods, farms
32.6 ± 8.1 SD 21.0 - 52.8 sout h Illinoi s/agr icul tural Roseber ry & Kl imstra, 1984
Number Hatch spr ing/ 12.2 sout h Texas/ semiprairie, Lehmann, 1984 per summer brush Successf ul March 20.0 sout hwest Georgia/pine Simpson, 1976 Nest August 8.4 woods, farms
Age at Sexual B 8 - 9 mont hs NS/NS (wild) Johnsgar d, 1988 Maturity B 16 weeks Sout h Carolina/lab Jones & Hughes, 1978
Annual A M 78.8 ± 2.47 SE 64.7 - 94.8 Florida/open woods Browni e et al., 1985 Mortality Rates A F 85.3 ± 2.72 SE 68.4 - 98.6 (percent ) J M 81.8 ± 2.46 SE 73.0 - 93.7
J F 87.2 ± 1.68 SE 67.9 - 95.8
B B 81 Illinoi s/agr icul tural Roseber ry & Kl imstra, 1984
no hunt ing B M 52 Florida/pine woods Pol lock et al., 1989 B F 56
Longevi ty starting: 9 (mont hs) B Novem ber 10.6 Texas/semiprairie, brush Lehmann, 1984
B October 8.5 cent ral Missour i/NS Marsden & Basket t, 1958
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/ Laying March May - June August Florida Bent , 1932 mid-Apr il mid-August sout h Texas Lehm ann, 1984 Apr il mid-May - Jul y September sout h Illinoi s Roseber ry & Kl imstra, 1984
Hatching mid-March May - June mid-September sout h Texas Lehm ann, 1984 late Apr il May - August October sw Geor gia, nor thern Flor ida Stoddar d, 1931 early May mid-June October Missour i Stanford, 1972a mid-May June - August early October sout h Illinoi s Roseber ry & Kl imstra, 1984
2-130 N
orth
ern Bobw
hite

Northern Bobw hite ( Colinus vir ginianus )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Molt fall August September October NS Bent, 1932
spr ing early February March - Apr il early June sw Geor gia, nor thern Flor ida Stoddar d, 1931
1 Metabol ized ener gy requi rements of farm-raised bi rds in capt ivi ty: (1) 7 weeks pr ior to laying (mean weight of hens = 194 g) and (2) dur ing l aying (mean weight of hens = 215 g) .
2 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and sum mer body weight s f rom Roseber ry and Kl imstra (1971). 3 Est imated usi ng equat ion 3-37 (Nagy, 1987) and sum mer body weight s f rom Roseber ry and Kl imstra (1971). 4 Diet of com mercial game food wi th onl y 5 to 10 percent water cont ent; maintained at temperature, hum idity, and l ight cycle typical for Texas. 5 Gross ener gy intake cal cul ated from the average vol ume of crop cont ents in shot bi rds, assum ing a 1.5-hour retent ion per iod, 2.30 kcal /cm3 for
the cont ents, and const ant foraging t hroughout the daylight hour s, whi ch is likely to over est imate food i ntake. 6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Roseber ry and Kl imstra (1971). 7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Roseber ry and Kl imstra (1971). 8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Roseber ry and
Kl imstra (1971). 9 Expect ed remaining l ongevi ty for those j uveni le quai l that sur vived t o the mont h indi cated.
2-131 N
orth
ern Bobw
hite

References (including A ppendix)
Amer ican Ornit hologi sts' Union. (1983) Check-l ist o f North Amer ican birds. Lawrence, KS: Allen Press, Inc.
Andrews, T. L.; Harms, R. H.; Wilson, H. R. (1973) Protein requir ement of th e bobw hit e chic k. Poult . Sci . 52: 2199-2201.
Baldwin, W. P., Jr .; Handley, C. O. (1946) Winter food of th e bobw hit e quail in V irginia . J. Wildl. Manage. 10: 142-149.
Bartholo mew, R. M. (1967) A stu dy of th e winter acti vi ty o f th e bobw hit e through t he use of radiotelemetr y. Kalamazoo, MI: Occas. Pap. Adams Eco l. Center , Wester n Mich. Univ .; 25 pp.
Bent, A. C. (1932) Life histo ries of North Amer ican gallin aceous bir ds. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 162.
Blem, C. R.; Zara, J. (1980) The energetics o f young bobw hit e (Colin us virg inianus). Comp. Bioc hem. Physiol. A: Comp. Physiol. 67: 611-615.
Borchelt, P. L.; Duncan, L. (1974) Dustb athing a nd feath er lipid in bobw hit e (Colin us virg inianus). Condor 76: 471-472.
Brennan, L. A. (1991) How can we reverse th e northern bobw hit e decline ? Wildl. Soc. Bull. 19: 544-555.
Brenner, F. J.; Reeder, M. (1985) Effect o f temp erature on energy intake i n three str ains of bobw hit e quail (Colin us virg inianus). Proc. Penn. Acad. Sci . 59: 119-120.
Brownie, C.; Anderson, D. R.; Burnham, K. P.; et al . (1985) Stati sti cal inference fr om band recovery data - a handbook . Washingt on, DC: U.S. Fish Wildl. Serv., Resour . Publ. 156.
Buss, I. O.; Matti son, H.; Kozlik , F. M. (1947) The bobw hit e quail in Dunn Count y, Wiscons in. Wisc. Cons . Bull. 12: 6-13.
Caesar Kleberg Wildlif e Research Insti tute (CKWRI). (1991) Double broods r evisi ted. In: Quail news fr om CKWRI No. 14, March 1991. Kings ville , TX: Texas A&I Universi ty; p. 6.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Campbe ll-Kis sock, L.; Blankenship, L. H.; Stewar t, J. W. (1985) Plant and animal foods of bobw hit e and scal ed quail in southwest T exas. Southwest. Nat. 30: 543-553.
2-132 Northern Bobw hit e

Case, R. M. (1973) Bioe nergetics of a covey o f bobw hites. Wilson Bull. 85: 52-59.
Case, R. M. (1982) Adaptat ions of femal e bobw hites to energy demands of th e reproducti ve cycl e. Proc. Natl . Bobw hit e Quail Symp . 2: 74-78.
Case, R. M.; Robel, R. J. (1974) Bioe nergetics of th e bobw hit e. J. Wildl. Manage. 38: 638-652.
Craighead, J. J.; Craighead, F. C. (1956) Hawks, owls and wildlif e. Harrisbur g, PA: The Stack pole Co. and Washingt on, DC: Wildlif e Management Insti tute.
Crim, L. A.; Seitz, W. K. (1972) Summer range and habitat p references o f bobw hit e quail on a southern Iowa State Game A rea. Proc. Iowa Acad. Sci . 79: 85-89.
Driver, C. J.; Ligo tke, M. W.; Van Voris, P.; et al . (1991) Routes o f uptake an d their relative cont ribut ion t o the toxicologic r espons e of nor thern bobw hit e (Colin us virg inianus) to an organophos phate pesti cide. Environ. Toxicol. Chem. 10: 21-33.
Guthery, F. S. (1988) Line tr ansect sam pling of bobw hit e dens ity on r angeland: evaluation and r ecommenda tions . Wildl. Soc. Bull. 16: 193-203.
Guthery, F. S.; Koerth, N. E.; Smith, D. S. (1988) Reproduc tion of nor thern bobw hites i n semi arid environments. J. Wildl. Manage. 52: 144-149.
Hamilton, M. (1957) Weight s of wild bobw hites i n central Misso uri. Bird Banding 2 8: 222-228.
Handley, C. O. (1931) The food a nd feeding ha bit s of bobw hites. In: Stodda rd, H. L., ed. The bobw hit e quail: its habit s, preservat ion a nd inc rease. New York, NY: Charles Scr ibne r's Sons .
Heitmeyer , M. E. (1980) Foods of bobw hites i n northeaster n Misso uri related to land use. Trans. Misso uri Acad. Sci . 14: 51-60.
Hurst, G. A. (1972) Insects an d bobw hit e quail br ood ha bitat man agement. Proc. Natl . Bobw hit e Quail Symp . 1: 65-82.
Johns gard, P. A. (1988) The quails, partr idge s, and f rancolins of th e world. Oxfo rd, Engla nd: Oxfo rd Universi ty Pr ess; pp. 60-68.
Jone s, J. E. ; Hughe s, B. L. (1978) Comparison of growth rate, body weight , and feed conv ersion b etween Cortunix D1 quail and bobw hit e quail. Poult . Sci . 57: 1471-1472.
Judd, S. (1905) The bob-white and other quails of th e United States an d their econom ic relations . U.S. Biol. Survey Bull. 21: 1-66.
Kellogg, F. E.; Doster , G. L.; Willi amso n, L. L. (1970) A bobw hit e dens ity gr eater than one bir d per acr e. J. Wildl. Manage. 34: 464-466.
2-133 Northern Bobw hit e

Klimstr a, W. D.; Roseberry, J. L. (1975) Nest ing e cology of th e bobw hit e in s out hern Illinois . Wildl. Monogr . 41; 37 pp.
Koerth, N. E.; Guthery, F. S. (1987) Body fat l evels of northern bobw hites i n south Texas. J. Wildl. Manage. 51: 194-197.
Koerth, N. E.; Guthery, F. S. (1990) Water requir ements o f cap tive northern bobw hit es unde r subt ropic al seaso ns. J. Wildl. Manage. 54: 667-672.
Koerth, N. E.; Guthery, F. S. (1991) Water restr ict ion effects o n northern bobw hit e reproduc tion. J. Wildl. Manage. 55: 132-137.
Korsch gen, L. J. (1948) Late-fal l and early-wi nter food ha bit s of bobw hit e quail in Mis sour i. J. Wildl. Manage. 12: 46-57.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967) A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lay, D. W. (1954) Quail management fo r east T exas. Texas Par ks Wildl. Dept . Bull. No. 34.
Lehmann, V. W. (1984) Bobw hites i n the Rio Grande pla in of T exas. Colle ge Station, TX: Texas A&M Universi ty Pr ess.
Marsden, H. M., Baskett, T. S. (1958) Annua l mortalit y in a banded bobw hit e popula tion. J. Wildl. Manage. 22: 414-419.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York, NY: McGraw-Hill Book Company, Inc.
McRae, W. A.; Dimmi ck, R. W. (1982) Body fat an d blood-ser um val ues of breeding w ild bobw hites. J. Wildl. Manage. 46: 268-271.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Moor e, P. E.; Cain, J. R. (1975) Character ization of bobw hit e quail r eared for hunt ing. Poult . Sci . 54: 1798.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Nelson, A. L.; Martin, A. C. (1953) Gamebird weight s. J. Wildl. Manage. 17: 36-42.
2-134 Northern Bobw hit e

Nestl er, R. B. (1946) Mechanical value of grit fo r bobw hit e quail. J. Wildl. Manage. 10: 137-142.
Nice, M. (1910) Food of th e bobw hit e. J. Econom ic En tomology 3: 6-10.
Polloc k, K. H.; Moor e, C. T.; Davids on, W. R.; et al. (1989) Survival rates o f bobw hit e quail based on band recovery analyses. J. Wildl. Manage. 53: 1-6.
Prasad, N. L.; Guthery, F. S. (1986) Drinking by nor thern bobw hites i n Texas. Wilson Bull. 98: 485-486.
Reid, V. H.; Goodr um, P. D. (1960) Bobw hit e quail: a produc t of longl eaf pine fo rests. Trans. Nor th Am. Wildl. Conf . 25: 241-252.
Robel, R. J. (1969) Food ha bit s, weight dynamics and fat co ntent of bobw hites i n relation t o food pla nt ings in Ka nsas. J. Wildl. Manage. 38: 653-664.
Robel, R. J.; Bisset, A. R.; Dayto n, A. D.; et al . (1979a) Comparative energetics o f bobw hit es on six differ ent foods . J. Wildl. Manage. 43: 987-992.
Robel, R. J.; Bisset, A. R.; Clement, T. M., Jr .; et al . (1979b) Metaboliza ble en ergy of impor tant foods of bobw hites i n Kansas. J. Wild. Manage. 43: 982-987.
Robel, R. J.; Case, R. M.; Bisset, A. R.; et al . (1974) Energetics o f food plot s in bobw hit e management. J. Wildl. Manage. 38: 653-664.
Roseberry, J. L.; Klimstr a, W. D. (1971) Annua l weight cycl es in male and femal e bobw hit e quail. Auk 88: 116-123.
Roseberry, J. L.; Klimstr a, W. D. (1984) Popula tion e cology of th e bobw hit e. Carbonda le and Edwardsville , IL: Sout hern Illinois Univ ersi ty Press.
Roseberry, J. L.; Peter john, B. G.; Kli mstr a, W. D. (1979) Dynamics o f an unexploit ed bobw hit e popula tion in d eter iorating ha bi tat. J. Wildl. Manage. 43: 306-315.
Rosene, W. (1969) The bobw hit e quail, its lif e and m anagement. New Brunswi ck, NJ: Rutgers Press.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Schroeder, R. L. (1985) Habitat su itabilit y mode ls inde x: nor thern bobw hit e. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.104).
Sermons, W. O.; Speake, D. W. (1987) Produc tion of second br oods by nor thern bobw hites. Wils on Bull. 99: 285-286.
2-135 Northern Bobw hit e

Simpson, R. C. (1976) Certain aspects o f th e bobw hit e quails lif e his tory in s out hwest Georgia . Atlanta, GA: Georgia Dept . Nat. Resour .; Tech. Bull. WL1.
Smith, G. F.; Kellogg, F. I.; Doster , G. L.; et al . (1982) A 10-year stu dy of bobw hit e quail movemen t patter ns. Proc. Natl . Bobw hit e Quail Symp . 2: 35-44.
Stanford, J. A. (1972a) Bobw hit e quail popula tion dy namics: relations hips of weath er, nesting, produc tion p atter ns, fal l popula tion c haracter isti cs, and harvest i n Misso uri quail. Proc. Natl . Bobw hit e Quail Symp . 1: 115-139.
Stanford, J. A. (1972b) Second br oods in bobw hit e quail. Proc. Natl . Bobw hit e Quail Symp . 1: 21-27.
Stemp el, M. E. (1960) Quail hatching a nd pr imary feath er moult in adult s. Proc. Iowa Acad. Sci . 67: 616-621.
Stodda rd, H. L. (1931) The bobw hit e quail: its habit s, preservat ion a nd inc rease. New York, NY: Charles Scr ibne r's Sons .
Tomlins on, R. E. (1975) Weight s and w ing le ngt hs of wild S onor an masked bobw hit es dur ing f all and w int er. Wils on Bull. 87: 180-186.
Urban, D. (1972) Aspects o f bobw hit e quail m obilit y dur ing s pring t hrough f all m ont hs. Proc. Natl . Bobw hit e Quail Symp . 1: 194-199.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Wiseman , D. S.; Lewis, J. C. (1981) Bobw hit e use of habitat i n tallgr ass r angeland. Wildl. Soc. Bull. 9: 248-255.
Wood, K. N.; Guthery, F. S.; Koerth, N. E. (1986) Spring-summer nutr ition a nd c ondit ion of nor thern bobw hites i n south Texas. J. Wildl. Manage. 50: 84-88.
Yoho, N. S.; Dimmick, R. W. (1972) Habitat u tiliza tion by bobw hit e quail dur ing w int er. Proc. Natl . Bobw hit e Quail Symp . 1: 90-99.
2-136 Northern Bobw hit e

2.1.10. American Woodcock (w oodcock and snipe)
Order Charadriformes, Family Scolopacidae. These inla nd memb ers of th e sandpipe r fami ly have a sto cky build, long bill, and s hor t legs. However , their habitats an d die t are dis tinc t. Woodc ock inha bit pr imarily woodla nds and a bandone d f ields , whereas snipe ar e found in asso ciation w ith bogs and f reshwater wetl ands. Both species use th eir long bills to probe th e subs trate fo r inver tebrates. The woodc ock and s nipe ar e simila r in lengt h, although t he femal e woodc ock weighs al most twi ce as much as the femal e snipe .
Selected species
The Amer ican woodc ock (Scolopax mi nor) breeds from sout hern Canada to Louis iana th roughout fo rested regions of th e easter n half of North Amer ica. The highe st breeding de nsit ies are found in t he nor thern por tion of this r ange, especially in t he Great Lakes ar ea of th e United States, northern New Engla nd, and s out hern Canada (Gregg, 1984; Owen et al ., 1977). Woodc ock wint er pr imarily in t he sout heaster n United States an d are year -round r eside nts in s ome of th ese areas. Woodc ock are impor tant game animals over much of th eir range (Owen et al ., 1977).
Body size. Woodc ock are large for sandpipe rs (28 cm bill t ip t o tail t ip) , and femal es weigh m ore than males (Keppie and Re dmond, 1988). Most y oung a re full gr own by 5 to 6 weeks after hatching ( Gregg, 1984).
Habitat . Woodc ock inha bit bot h woodla nds and a bandone d f ields , partic ularly thos e wi th rich and moderately to poor ly drained lo amy s oils , whic h tend t o suppor t abunda nt earthworm popula tions (C ade, 1985; Owen and Galbraith, 1989; Rabe et al ., 1983a). In the spring, males us e early su ccess iona l ope n areas and woods openings , int erspersed wi th low brush and grassy veg etat ion, for singing dis plays at d awn and dusk (Cade, 1985; Keppie and Re dmond, 1985). Females nest i n brushy areas of sec onda ry gr owth woodla nds near th eir feeding a reas, often near the edge of th e woodla nd or near a br eak in the forest ca nopy (Gr egg, 1984). During t he summer , both sexes u se sec ond gr owth hardwood or early su ccess iona l mixed hardwood a nd c onif er woodla nds fo r diur nal cover (Cade, 1985). At night , they move int o ope n pastu res and early su ccess iona l abandone d agricult ural fields , inc luding f ormer mal e singing gr ounds , to roos t (Cade, 1985; Dunf ord and Owen, 1973; Krohn, 1970). During t he wint er, woodc ock us e bot tomland ha rdwood forests, hardwood t hickets, and upla nd m ixed hardwood a nd c onif er fo rests d uring t he day. At night , they us e open areas to some degree, but also forested habitats (C ade, 1985). Diur nal habitat an d noctu rnal roost ing f ields need to be in c los e prox imity to be usefu l for woodc ock (Owen et al ., 1977).
Food ha bi ts . Woodc ocks feed primar ily on inver tebrates f ound in m ois t upla nd soils by probing t he soil w ith their long pr ehensil e-tippe d bill (Owen et al ., 1977; Sperry, 1940). Earthworms ar e the prefer red diet, but wh en earthworms ar e not avai lable, other soil inv ertebrates ar e consumed (Mille r and Ca usey, 1985; Sperry, 1940; Str ibling a nd Doerr, 1985). Some seed s and other plant matter may al so be consumed (Sperry, 1940). Krohn (1970) found t hat dur ing s ummer most fee ding was done in w oode d areas prior to enter ing f ields at night , but other studie s have indi cated that a s ignif icant amount of food
2-137 Amer ican Woodc ock

is acquir ed dur ing noc turnal acti vi ties (Britt, 1971, as ci ted in Dunf ord and Owen , 1973). Dyer and Hamilton (1974) found t hat dur ing t he wint er in s out hern Louis iana, woodc ock exhibit ed three feeding pe riods : early mo rning (0100 to 0500 hour s) in t he noc turnal habitat, midday (1000 to 1300 hour s) in t he diur nal habitat, and at dusk (1700 to 2100 hour s) again in t he noc turnal fields ; earthworms and m illipe des were cons umed in bot h habitat typ es. Most o f th e woodc ocks' metabolic water needs are met b y th eir food (Mendall and A ldous , 1943, as ci ted in Cade, 1985), but cap tive birds have been observed to dr ink (Sh eldon, 1967). The chicks l eave th e nest s oon after hatching, but ar e dependent on the femal e for food f or th e fi rst week after hatching ( Gregg, 1984).
Mol t. Woodc ock molt twi ce annua lly . The prenupt ial molt inv olves body plum age, some wing c over ts, scapula rs, and t ertials and oc curs in l ate wi nter or early sp ring; the complete postnupt ial molt takes p lace in July or August (B ent, 1927).
Migration. Fall m igr ation be gins in l ate Septemb er and continue s through Decemb er, often follow ing t he fi rst heavy fr ost (Sh eldon, 1967). The migration may take 4 to 6 weeks (Sh eldon, 1967). Some woodc ock wint er in t he sout h Atlant ic r egion, while those th at breed west o f th e Appalachian Mount ains wint er in Louis iana and ot her Gulf States (M artin et al ., 1969, as ci ted in Owen et al ., 1977). Woodc ock are early sp ring migr ants, leaving t heir wint ering gr ounds in February and arriving on t heir nor thern breeding gr ounds in l ate March to early April (Gregg, 1984; Sheldon, 1967; Owen et al ., 1977). Dates o f woodc ock arrival at their breeding gr ounds var y from year to year depending on t he timing of sno wmel t (Gregg, 1984). Sheldon (1967) summar izes sp ring and f all m igr ation d ates b y States fr om numerous st udie s.
Breeding acti vi ties and social organization. From th eir ar rival in t he spring, male woodc ock perfor m daily cour tship f light s at dawn and at dus k, defending a si te on t he singing gr ounds in or der to attr act femal es fo r mat ing (Owen et al ., 1977; Gregg, 1984). Often sever al males display on a single singing gr ound, wi th each defending his own sect ion of th e area. Females co nstr uct th eir nests o n the ground, usually at the base of a tree or shrub located in a brushy area adjacent to an opening or male singing gr ound (Gregg and Hale, 1977; McAuley et al ., 1990; Owen et al ., 1977). Females are respons ible for all of th e inc ubation a nd c are of th eir brood ( Trippe nsee, 1948). The young l eave th e nest s oon after hatching a nd c an sustain f light by appr ox imatel y 18 days o f age (Gregg, 1984).
Home range and r esour ces. The hom e range of woodc ocks en compasses b oth diur nal cover ar eas and noctu rnal roost ing a reas and var ies in size depending on seaso n and t he dis tr ibut ion of feeding s ites an d suitable cover . During t he day, mo vemen ts ar e usually lim ited unt il dus k, wh en woodc ock fly to noc turnal roos t si tes. Hudgins et al. (1985) and Gregg (1984) found s pring a nd s ummer diur nal ranges to be only 1 to 10 percent of th e total home range. Movemen t on the noctu rnal roost si tes al so is l imi ted; however , during w int er, woodc ock are more lik ely to feed and move around a t night (Bor tner, pers. co mm.). Singing m ales generally r estr ict th eir movemen ts mo re than non-singing m ales, juv enile s, and femal es (Owen et al ., 1977).
Popul ation de nsity . The annua l s inging- ground s urvey c onducted by th e Unit ed States an d Canada provides informat ion on t he popula tion t rends of woodc ock in t he
2-138 Amer ican Woodc ock

�
northern states an d Canada during t he breeding seaso n (note fr om B. Bortner, U.S. Fish and Wildlif e Service, Offi ce of Migr ating Bir d Management, to Susan Norton, January 9, 1992). Gregg (1984) summar ized results o f sever al publis hed s inging- ground s urveys an d found esti mates to vary fr om 1.7 male singing gr ounds per 100 ha in Minne sota (Godf rey, 1974, ci ted in Gregg, 1984) to 10.4 male singing gr ounds per 100 ha in Maine (Mendall and Aldous , 1943, ci ted in Gregg, 1984). Although t his method is appr opr iate fo r assessi ng popula tion t rends , flus hing s urveys, telemetr y, and mark-recapture are better methods fo r esti mat ing w oodc ock densit ies because th ere are var iable numbers of femal es and nons inging m ales asso ciated wi th acti ve s inging gr ounds (Dilw or th, Krohn, Riffen berger, and Whitco mb pers. comm. , ci ted by Owen et al ., 1977). For examp le, Dwyer et al . (1988) found 2 .2 singing m ales per 100 ha in a wildlif e refuge in Maine, but wi th mark-recapture technique s, th ey found year ly su mmer densi ties of 19 to 25 birds per 100 ha in the same area.
Popul ation dy namics. Woodc ocks attemp t to raise only a single brood in a giv en year but may r enest i f th e init ial c lut ch is d estr oyed (McAuley et al ., 1990; Sheldon, 1967). In 12 years of stu dy in Wisco nsin, Gregg (1984) found 4 2 percent of all n ests to be lost to predators and another 11 percent lost to other causes. Survival of juveniles in their fi rst year ranges fr om 20 to 40 percent, and survival of adults r anges fr om 35 to 40 percent fo r males to approximatel y 40 to 50 percent fo r femal es (Dwyer and Nichols , 1982; Krohn e t al., 1974). Derleth and Sepik (1990) found high a dult su rviv al rates (0.88 to 0.90 for both sexes) between June and Octo ber in Maine, indi cating t hat adult mo rtalit y may o ccu r primar ily in the wint er and e arly sp ring. They found low er su mmer survival rates fo r young w oodc ock between fledging a nd m igr ation t han for adult s dur ing t he same mo nths, wi th most l osses of young attr ibut ed to predation.
Simi lar species (from general references)
� The common s nipe ( Gallin ago gallin ago) is si mi lar in length (27 cm) to the woodc ock, al though light er in w eight . Snipe ar e pr imarily found in asso ciation w ith bogs and f reshwater wetl ands and feed on the var ious inver tebrates asso ciated wi th wetl and soils . Snipe breed pr imarily in bor eal forest r egions and t hus ar e found s light ly nor th of th e woodc ock br eeding range, wi th some areas of over lap in the easter n half of th e continent. The breeding r ange of th e snipe , however , exten ds westwar d to the Paci fic co ast and t hroughout mo st of A laska, thus occu pying a mo re exten sive east-west range th an the woodc ock.
General references
Cade (1985); Dwyer et al . (1979); Dwyer and Storm (1982); Gregg (1984); Nationa l Geogr aphic So ciety (1987); Owen et al . (1977); Sheldon (1967); Trippe nsee (1948).
2-139 Amer ican Woodc ock

American Woodcock ( Scolo pax minor )
Factors Age/Sex/ (95% CI of Note Cond. /Seas. Mean mean) Locat ion erence No.
Range or
Body Weight A M 176 throughout range Nelson & M artin, 1953 (g) A F 218
A M Apr il 134.6 ± 2.9 SE Maine Dwyer et al., 1988 A M May 133.8 ± 5.8 SE A M June 151.2 ± 9.5 SE
A M sum mer 145.9 127 - 165 cent ral Massachuset ts Sheldon, 1967 J M sum mer 140.4 117 - 152 A F sum mer 182.9 162 - 216 J F sum mer 168.8 151 - 192
A M fall 169 Minnesot a Marshal l (unpubl .) 1 J M fall 164 A F fall 213 J F fall 212
at hatching 13.0 Wisconsi n Gregg, 1984 9 - 16
Egg Weight (g) at laying 18 - 19 Wisconsi n Gregg, 1984 near hatching 14 - 16
Chick Growth M 5.1 Maine Dwyer et al., 1982 Rate (g/day) F 6.2
Metabol ic Rate A F basal 115 s Michi gan Rabe et al., 1983b 2 (kcal /kg-day)
A M basal 126 est imated 3 A F basal 118
A F free-liv ing 315 s Michigan Rabe et al., 1983b 4 A F nest ing 553
A M free-living 313 (148 - 662) est imated 5 A F f ree-living 296 (140 - 627)
Ref
2-140 A
merican
Woodc
ock
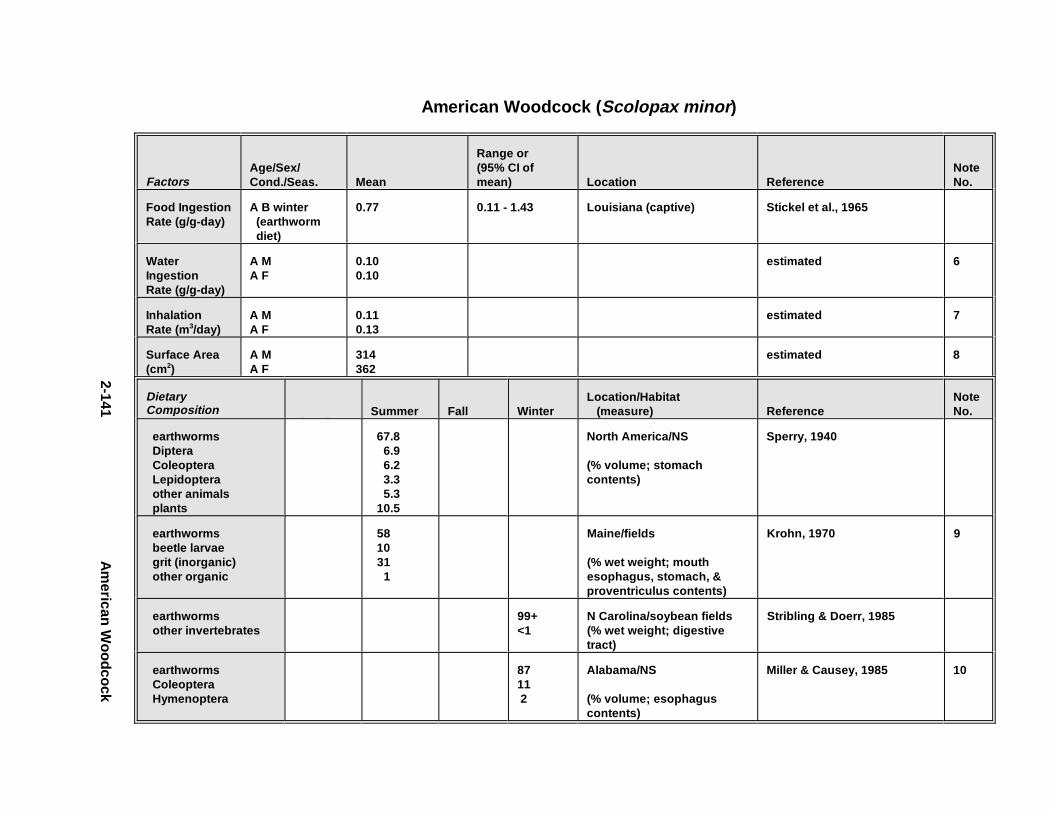
American Woodcock ( Scolo pax minor )
2-141 A
merican
Woodc
ock
Factors Age/Sex/ (95% CI of Note Cond. /Seas. Mean mean) Locat ion erence No.
Range or
Food Ingest ion A B wi nter 0.77 0.11 - 1.43 Loui siana (capt ive) Stickel et al., 1965 Rate (g/g-day) (earthworm
diet)
Water A M 0.10 est imated 6 Ingestion A F 0.10 Rate (g/g-day)
Inhal ation A M 0.11 est imated 7 Rate (m /day) A F 0.133
Surface Ar ea A M 314 est imated 8 (cm ) A F 3622
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
earthworms 67.8 North America/NS Sperry, 1940 Diptera 6.9 Coleoptera 6.2 (% volume; stomach Lepi dopt era 3.3 cont ents) other animals 5.3 plants 10.5
earthwor ms 58 Maine/fields Krohn, 1970 9 beetle larvae 10 grit (inor gani c) 31 (% wet weight ; mout h other organi c 1 esophagus, stomach, &
provent riculus cont ents)
earthworms 99+ N Carolina/soy bean f ields Stribl ing & Doer r, 1985 other invertebrates <1 (% wet weight ; digest ive
tract)
earthworms 87 Alabama/NS Mille r & Causey, 1985 10 Coleoptera 11 Hymenopt era 2 (% vol ume; esophagus
cont ents)
Ref

American Woodcock ( Scolo pax minor )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home Range A M inact ive 3.1 (median) 0.3 - 6.0 Pennsy lvani a/mixed f orests Hudgi ns et al., 1985 Size (ha) A M act ive 73.6 (median) 38.2 - 171.2 with shr ubs and f ields
A M singi ng 10.5 (median) 4.6 - 24.1
B B sum mer 32.4 ± 27.6 SD 7 - 98 Wisconsi n/woods, open Gregg, 1984 A F w ith 4.5 areas, brush brood
Popul ation North Carolina/agr icul tural: Connor s & Doer r, 1982 Density B B w inter 3.38 untille d soy stubbl e (bird s/ha) B B winter 0.20 untille d corn stubbl e
B B w inter 0.034 rebedded cor n fields
nest s in 0.21 (nests/ha) Pennsy lvani a/mixed pi ne Coon et al., 1982 spr ing and
A M sum mer 0.056 0.037 - 0.074 Maine/second gr owt h forest , A F sum mer 0.125 0.108 - 0.143 meadows, and ponds J B sum mer 0.223 0.190 - 0.250 B B summer
0.035 0.026 - 0.046 hardwoods, open f ields Dwyer et al., 1988
Clutch 4 3 - 5 throughout range and Bent, 1927 Size habi tats
1st clutch 3.8 ± 0.42 SD Maine/mixed fo rests, McAuley et al., 1990 2nd cl utch 3.0 ± 0.67 SD agr icul tural fields
Clutches/ 1 but renest if 1st throughout range and McAul ey et al., 1990 Year lost habi tats
Percent Nests about 50 Maine/mixed f orests, fields McAul ey et al., 1990 Hatching
Days 19 - 21 NS/NS Mendal l & Al dous, 1943; 11 Incubat ion Pettin gill, 1936
Age at 18 - 19 days Wisconsi n/woods, open Gregg, 1984 Fledgi ng areas, brush
Range
2-142 A
merican
Woodc
ock

American Woodcock ( Scolo pax minor )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Age at M < 1 year throughout range and Sheldon, 1967 Sexual F year habi tats Maturity
Annual A M east 65 ± 5.2 SD eastern and cent ral Uni ted Dwyer & Nichol s, 1982 Mortality A M cent ral 60 ± 15 SD States/NS Rates J M east 80 ± 4.8 SD
J M cent ral 64 ± 12 SD A F east 51 ± 7.3 SD A F cent ral 47 ± 9.6 SD J F east 64 ± 7.7 SD J F cent ral 69 ± 9.4 SD
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Laying early February mid-March Texas Whiting & Boggus, 1982 early April Maine Dwyer et al., 1982
Hatching early February Loui siana Pettingill, 1936 1 late February Virg in ia Pettin gill, 1936 1 late March Connecticut Pettin gill, 1936 1 mid-Apr il early May early June Massachuset ts Sheldon, 1967
mid-May Maine Dwyer et al., 1982
Molt August to NS/NS Owen & Kr ohn, 1973 12 early September
Migration spr ing mid-February early March leaving Nor th Carolina Connor s & Doer r, 1982
fall October December arriving Nor th Carolina Sheldon, 1967
March Apr il arriving i n nor thern range Gregg, 1984
late mid-December leaving Canada Owen et al., 1977 September
Range
1
2-143 A
merican
Woodc
ock

12345678
American Woodcock ( Scolo pax minor )
As ci ted in Sheldon ( 1967).Metabol ic rate est imated by author s f rom equat ion of Aschof f and Pohl (1970).Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and sum mer body weight s f rom Nelson and M artin (1953).Est imate of free-living m etabol ism based on ener gy budget model . Metabol ism dur ing nest ing est imated for peak needs dur ing eg g-laying.Est imated usi ng equat ion 3-37 (Nagy, 1987) and sum mer body weight s f rom Nelson and M artin (1953).Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and sum mer body weight s f rom Nelson and M artin (1953).Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and sum mer body weight s f rom Nelson and M artin (1953).Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and sum mer body weight s f rom Nelson andMartin (1953).
9 Grit com prised onl y 14 percent of total digest ive t ract cont ents vol ume. 10 Shoul d pr ovi de a more accur ate est imate of propor tion of so ft-bodi ed earthwor ms consum ed than woul d includi ng ot her por tions of the digest ive
tract . 11 Cited in Trippensee ( 1948). 12 Cited in Owen et al. (1977).
2-144 A
merican
Woodc
ock

References (including A ppendix)
Aldous , C. M. (1938) Woodc ock management st udies in Maine 1937. Trans. North Am. Wildl. Nat. Resour . Conf . 3: 839-846.
Aschof f, J. ; Pohl, H. (1970) Rhyth mic var iations in energy metabolis m. Fed. Proc. 29: 1541-1552.
Bent, A. C. (1927) Life histo ries of North Amer ican shore birds. Part 1. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 142.
Britt, T. L. (1971) Studie s of woodc ock on t he Louis iana wint ering gr ound [master 's th esis]. Shrevepor t, LA: Louis iana State Universi ty.
Cade, B. S. (1985) Habitat su itabilit y inde x mode ls : Amer ican woodc ock (wint ering) . U.S. Fish Wildl. Serv. Biol. Rep. 82(10.105).
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Connor s, J. I.; Doerr, P. D. (1982) Woodc ock us e of ag ricult ural fields in c oastal North Carolina . In: Dwyer , T. J.; Storm, G. W., tech . coords. Woodc ock ecology an d management. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 139-147.
Coon, R. A.; Willi ams, B. K.; Lindze y, J. S. ; et al. (1982) Exami nation of woodc ock nest si tes in c entral Penns ylvania . U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 55-62.
Derleth, E. L.; Sepik, G. F. (1990) Summer -fal l survival of th e Amer ican woodc ock in Ma ine. J. Wildl. Manage. 54: 97-106.
Dunf ord, R. D.; Owen, R. B. (1973) Summer behavior of immatu re radio- equippe d woodc ock in c entral Maine. J. Wildl. Manage. 37: 462-469.
Dwyer , T. J.; Nichols , J. D. (1982) Regiona l popula tion inf erences fo r the Amer ican woodc ock. In: Dwyer , T. J.; Storm, G. L., tech . coords. Woodc ock ecology an d management. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 12-21.
Dwyer , T. J.; Storm, G. L., eds. (1982) Woodc ock ecology and m anagement. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14.
Dwyer , T. J.; Coon, R. A.; Geissl er, P. H. (1979) The technical literature on the Amer ican woodc ock 1927-1978. Laurel, MD: U.S. Fish Wildl. Serv., Migr atory Bir d and Habit at Research Laboratory.
Dwyer , T. J.; Derleth, E. L.; McAuley, D. G. (1982) Woodc ock br ood e cology in Maine. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 63-70.
2-145 Amer ican Woodc ock

Dwyer , T. J.; Sepik, G. F.; Derleth, E. L.; et al . (1988) Demographic ch aracter isti cs o f Maine woodc ock popula tion a nd effects o f habitat man agement. Washingt on, DC: U.S. Fish Wildl. Serv. Res. Rep. 4.
Dyer , J. M.; Hamilton, R. B. (1974) An analysi s of feeding ha bit s of th e Amer ican woodc ock (Philohela minor ) in s out hern Louis iana. In: Fif th Amer ican woodc ock workshop proceedings ; Decemb er 3-5, 1974; Athens, GA. Athens, GA: Universi ty o f Georgia.
Godfrey, G. A. (1974) Behavior and ecology of Amer ican woodc ock on t he breeding r ange in Minne sota [Ph.D. disser tat ion] . Minne apolis , MN: Univ ersi ty of Minne sota.
Greeley, F. (1953) Sex and age studie s in f all-shot woodc ock (Philohela minor ) from sout hern Wiscons in. J. Wildl. Manage. 17: 29-32.
Gregg, L. (1984) Popula tion e cology of woodc ock in W iscons in. Wis. Dept . Nat. Resour . Tech. Bull. No. 144; 51 pp.
Gregg, L. E.; Hale, J. B. (1977) Woodc ock nesting ha bitat i n northern Wisco nsin. Auk 94: 489-493.
Hudgins , J. E.; Storm, G. L.; Wakeley, J. S. (1985) Local movemen ts an d diur nal-habit at selection by mal e woodc ock in P enns ylvania . J. Wildl. Manage. 49: 614-619.
Johns on, R.C.; Causey, M. K. (1982) Use of longl eaf pine stands by woodc ock in s out hern Alabama follow ing pr escr ibed burning. In: Dwyer , T. J.; Storm, G. W., tech . coords. Woodc ock ecology and m anagement. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 120-125.
Keppie , D. M.; Redmond, G. W. (1985) Body weight and t he possess ion of ter ritory for mal e Amer ican woodc ock. Condor 87: 287-290.
Keppie , D. M.; Redmond, G. W. (1988) A review of possible expla nations fo r reverse si ze dim orphis m of A mer ican woodc ock. Can. J. Zool. 66: 2390-2397.
Krohn, W. B. (1970) Woodc ock feeding ha bit s as related to summer field usage in central Maine. J. Wildl. Manage. 34: 769-775.
Krohn, W. B.; Martin, F. W.; Bur nham, K. P. (1974) Band-recovery distr ibtution a nd s urviv al esti mates o f Maine woodc ock. Proc. Am. Woodc ock Workshop 5 : 1-8.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
2-146 Amer ican Woodc ock

Martin, F. W.; Willi ams, S. O.; Newso m, J. D.; et al . (1969) Analysi s of records of Louis iana-banded woodc ock. Proc. Ann. Conf . Sout heast. Asso c. Game and Fish Comm. 23: 85-96.
McAuley, D. G.; Longc ore, J. R.; Sepik , G. F. (1990) Renest ing by A mer ican woodc ock (Scolopax mi nor) in Maine. Auk 107: 407-410.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Mendall, H. L.; Aldous , C. M. (1943) The ecology and m anagement of th e Amer ican woodc ock. Orono, ME: Maine Coop. Res. Unit , Univ ersi ty of Maine; 201 pp.
Mille r, D. L.; Causey, M. K. (1985) Food pr eferences o f Amer ican woodc ock wint ering in Alabama. J. Wildl. Manage. 49: 492-496.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Nelson, A. L.; Martin, A. C. (1953) Gamebird weight s. J. Wildl. Manage. 17: 36-42.
Nor ris, R. T.; Buele, J. D.; Studholm e, A. T. (1940) Banding w oodc ocks o n Pennsyl vania singing gr ounds . J. Wildl. Manage. 4: 8-14.
Owen, R. B.; Galbraith, W. J. (1989) Earthworm biomass i n relation t o forest typ es, soil, and land us e: implic ations fo r woodc ock management. Wildl. Soc. Bull. 17: 130-136.
Owen, R. B.; Krohn, W. B. (1973) Molt p atter ns and weight ch anges of th e Amer ican woodc ock. Wils on Bull. 85: 31-41.
Owen, R. B.; Morgan, J. W. (1975) Summer behavior of adult r adio- equippe d woodc ock in central Maine. J. Wildl. Manage. 39: 179-182.
Owen, R. B.; Anderson, J. M.; Artman n, J. W.; et al . (1977) Amer ican woodc ock. In: Sanderson, G. C., ed. Management of mi gratory sh ore and upla nd g ame birds in Nor th Amer ica. Washingt on, DC: Int . Asso c. Fish Wildl. Agencies; pp. 147-175.
Pettingill, O. S., Jr . (1936) The Amer ican woodc ock. Bos ton S oc. Nat. His t. Mem. 9(2).
Rabe, D. L.; Prince, H. H.; Beaver , D. L. (1983a) Feeding- si te selection a nd f oraging str ategie s of A mer ican woodc ock. Auk 100: 711-716.
Rabe, D. L.; Prince, H. H.; Goodman, E. D. (1983b) The effect o f weath er on bioe nergetics of breeding A mer ican woodc ock. J. Wildl. Manage. 47: 762-771.
2-147 Amer ican Woodc ock

Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Sheldon, W. G. (1967) The book of th e Amer ican woodc ock. Amherst, MA: Univ ersi ty of Massach usetts Pr ess.
Sperry, C. (1940) Food ha bit s of a gr oup of shor e bir ds; woodc ock, snipe , knot , and dow itcher. U.S. Dept . Int ., Bur . Biol. Survey, Wildl. Res. Bull. 1; 37 pp.
Stickel , W. H.; Hayne, D. W.; Stickel , L. F. (1965) Effects o f heptachlor -con tami nated earthworms on w oodc ocks. J. Wildl. Manage. 29: 132-146.
Str ibling, H. L.; Doerr, P. D. (1985) Noctu rnal use of fi elds by Amer ican woodc ock. J. Wildl. Manage. 49: 485-491.
Trippe nsee, R. E. (1948) Amer ican woodc ock. Wildlif e management. New York, NY: McGraw-Hill; pp. 323-332.
Tufts, R. W. (1940) Some st udie s in bill measu rements an d body weight s of A mer ican woodc ock (Philohela minor ). Can. Field-Nat. 54: 132-134.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Wetherbee, D. K.; Wetherbee, N. S. (1961) Arti ficial incubation of eggs of var ious bir d species and some attr ibutes o f neonates. Bird Banding 3 2: 141-159.
Whit ing, R. M.; Boggus , T. G. (1982) Breeding biology of Amer ican woodc ock in east T exas. In: Dwyer , T. J.; Storm, G. W., tech . coords. Woodc ock ecology and m anagement. U.S. Fish Wildl. Serv., Wildl. Res. Rep. 14; pp. 132-138.
2-148 Amer ican Woodc ock

2.1.11. Spotted Sandpiper (sandpipers)
Order Charadriif ormes, Family Scolopacidae. The fami ly Scolopacidae inc lude s num erous sp ecies of shor ebir ds, e.g., sandpipe rs, tattl ers, knots, godw its, cu rlews, yellow legs, will ets, and dowitch ers. Those kn own as sandpipe rs tend t o be smal l wi th moderately long le gs and bills . Most sandpipe rs forage on s andy b eaches and mudflats; a few u tilize upla nd areas. They feed almost excl usively on smal l inver tebrates, either by probing int o or gleaning f rom th e subs trate. Most sp ecies are highly migr atory, breeding in arcti c and s ubarcti c regions and e ither wint ering a long t he coasts o r in southern lati tudes and t he sout hern hemisphe re; therefore, many are only p assag e migrants th roughout mo st of th e United States. Scolapids r ange in s ize fr om th e least sa ndpipe r (11.5 cm bill t ip t o tail t ip) to the long- bille d curlew (48 cm).
Selected species
The spotted sandpipe r (Acti ti s macu laria) (19 cm) i s a ver y co mmo n summer resident of fr eshwater and sal twater bodie s throughout mo st of th e Unit ed States. These sandpipe rs are most often encount ered singly but may fo rm smal l flocks. Most wi nter in the neotropic s.
Body size. Females (approximatel y 50 g) are signif icantly larger th an males (approximatel y 40 g) (Oring a nd La nk, 1986).
Habitat . Spotted sandpipe rs br eed along t he edges of bodie s of water , usually in open habitats, from th e northern border of th e boreal forest acr oss North Amer ica, south to the central United States (Or ing a nd La nk, 1986). They requir e open water for bathing a nd dr ink ing, semi -open habitat fo r nest ing, and de nse vegetation f or breeding ( Bent, 1929; Oring e t al., 1983).
Food ha bi ts . In coastal areas, spotted sandpipe rs search the beach and muddy edges of inlets an d creeks, wading l ess fr equently th an most sa ndpipe rs; inla nd t hey feed along t he shor es of sandy ponds and a ll t ypes of str eams, someti mes str aying int o meadows, fields, and gardens in agricultural areas (Bent, 1929). Their diet is co mposed primar ily of ter restr ial and mar ine insects (B ent, 1929). While adult fl ying in sects co mprise the bulk of th e die t, cr ustacean s, leeches, mollu scs, smal l fish, and car rion a lso are eaten (Oring e t al., 1983). Young feed themsel ves i mmed iately after hatching, conc entrating on smal l inver tebrates (Or ing a nd La nk, 1986). During in sect o utbreaks, sandpipe rs will forage in w oode d areas near water , and they have been observed eating e ggs and f ish on occas ion ( Oring, pers. obs .).
Mol t. Partial prenupt ial molt of body plum age occurs in Ma rch and April, while th e pos tnupt ial molt begins by Augus t wi th the body feath ers and ends anywh ere from Octo ber to April w ith the los s of th e pr imary flight feath ers (Bent, 1929).
Migration. Spotted sandpipe rs generally migr ate in smal l flocks o r solit arily (Nationa l Geogr aphic So ciety, 1987). They wi nter from so uthern United States to northern Chile , Argent ina, and Ur aguay (Oring a nd La nk, 1986), and breed across North
2-149 Spotted Sandpipe r

�
�
Amer ica, nor th from Vi rginia and s out hern Calif ornia (Nationa l Geogr aphic So ciety, 1987). In the spring, femal es ar rive at th e breeding gr ounds ear lier th an males (in one study , by about 2 weeks; Oring a nd La nk, 1982).
Breeding acti vi ties and social organization. The pr imary cons ideration f or nesti ng si tes i s proximity to water , and spotted sandpipe rs have been known to build t heir ground nests i n such diverse condit ions as de pressions in v olc anic r ock and s trawberry patch es (Bent, 1929). Spotted sandpipe rs are poly andr ous (i .e., a single femal e lays eg gs fo r mult iple mal es), wi th males supply ing m ost of th e inc ubation a nd pa rental care (Oring, 1982). Thus reproduc tion is lim ited by th e num ber of mal es pr esent (Lank et al ., 1985). Spotted sandpipe rs lay a determinate cl utch of fo ur eggs. Females may l ay sever al clutch es in a year , often a dozen eggs per seaso n (Maxso n and Oring, 1980). Egg laying begins b etween late May and early Ju ne in Minne sota (Lank et al., 1985), and males incubate after the third egg is laid (Oring e t al., 1986). Females so meti mes i ncubate and brood w hen anot her mal e is not availa ble (Maxson a nd Oring, 1980). Parents b rood smal l chicks an d protect th em wi th warning c alls or by dis tracting or attack ing pr edators (Oring and Lank, 1986).
Home range and r esour ces. Although a var iety o f veg etat ion t ypes are used, nests usually are placed in semi -open vegetat ion ne ar th e edge of a lake, river , or ocean (Oring e t al., unpubl. , as ci ted in Oring e t al., 1983; McVey, pers. obs.). The suitabilit y of n esti ng habitat var ies fr om year to year in some locat ions due to levels of precipit ation a nd predators (Oring e t al., 1983).
Popul ation de nsity . Spotted sandpipe r nesting de nsit ies have been studie d well a t only one location, on Lit tle Pelic an Island, Leech Lake, Minne sota. At this location, densi ties ranged from 4 to 13 femal es per hectar e and 7 to 20 males per hectar e over a 10-year period, depending on weath er and other condit ions (Or ing e t al., 1983).
Popul ation dy namics. Females may l ay one to six cl utch es fo r differ ent mal es over one seaso n (Oring e t al., 1984), averaging 1 .3 to 2.7 mates p er year (Oring e t al., 1991b). Female mat ing a nd r eproduc tive success i ncrease wi th age, but mal e success d oes not (Oring e t al., 1991b). Lifeti me reproducti ve su ccess i s mo st affected by fl edging s uccess and longe vi ty for bot h males and femal es (Or ing e t al., 1991a).
Simi lar species (fr om general references)
� The solit ary sandpipe r (Trin ga solit aria) is usually seen singly in f reshwater swamp s or rivers. Present over much of th e United States d uring a nnua l migr ations , this average-sized sandpipe r (18 cm) wi nters along t he sout heast and Gulf co asts.
� The wester n sandpipe r (Calid ris mauri ) is a small s andpipe r (13 cm), commo n on mudflats an d sandba rs, th at wint ers on bot h the Atlant ic an d Paci fic sh ores of th e United States.
2-150 Spotted Sandpipe r

�
�
�
� The least sa ndpipe r (Calid ris minutilla ), the smal lest o f th is group (11 cm), is common in w int er on s alt marshes and m uddy shor es of r ivers and estu aries in coastal areas acr oss th e United States.
� The semi palmated sandpipe rs (Calid ris pusilla ) are smal l birds (13 cm) seen in the United States p rimar ily during m igr ation a nd r arely wint ering on Flor ida co asts.
� Most o ther memb ers of th e fami ly Scolopacidae forage by gleaning.
General references
Oring a nd La nk (1986); Lank et al . (1985); Nationa l Geogr aphic So ciety (1987); Oring et al . (1991a, 1991b).
2-151 Spotted Sandpipe r

Spotted Sandpiper ( Actitis macularia )
2-152 S
po
tted S
andpiper
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A F spr ing 47.1 43 - 50 Minnesot a island Maxson & O ring, 1980 (g) A M spr ing 37.9 34 - 41
Metabol ic Rate A F pr e-breed 404 - 787 Minnesot a Maxson & O ring, 1980 1 (kcal /kg-day) A F laying 383 - 745
A F 368 incubat ing 440 A M pre-breed 303 A M incubat ing 425 A M broodi ng
A F f ree-living 460 (213 - 994) A M free-liv ing
436 (202 - 937) est imated 2
Food Ingest ion 3 Rate (g/g-day)
Water A F 0.16 est imated 4 Ingest ion A M 0.17 Rate (g/g-day)
Inhal ation A F 0.039 est imated 5 Rate (m /day) A M 0.0333
Surface Ar ea A F 131 est imated 6 (cm ) A M 1132
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
mayflies �� Minnesot a/island i n lake Maxson & O ring, 1980 midges ��
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Territory appr ox. 0.25 NS/NS Maxson & O ring, 1980 Size (ha)
Range

Spotted Sandpiper ( Actitis macularia )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Popul ation A F sum mer 10 3.8 - 12.5 Minnesot a/island i n lake Oring et al., 1983 Densi ty (N/ha) A M sum mer 13.9 7.5 - 20.0
Clutch Si ze 4 3 - 5 NS/NS Bent , 1929; Oring et al., 1983 7
Clutches/Year 1 - 6 Minnesota/NS Orin g et al., 1983
Days 18 to 24 NS/NS Oring, unpubl ished Incubat ion
Age at approximately NS/NS Oring et al., 1991a Fledgi ng 18 days
Number 1.83 0.58 - 2.76 Minnesota/is land in lake Oring et al., 1984 Fledge per Nest That Hatches
Number 2.58 1.67 - 2.91 Minnesota/is land in lake Oring et al., 1984 Fledge per Successf ul Nest
Age at F 1 year Minnesota/is land in lake Oring et al., 1983 Sexual M 1 year Maturity
Annual F appr ox. 31 Minnesot a/island i n lake Oring et al., 1983; Oring & Mortality M appr ox. 30 Lank, 1982; Oring, Rates (percent) unpubl ished
Longevi ty A F 3.7 years Minnesot a/island i n lake Oring et al., 1983
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Matin g early May late May - early Minnesota Lank et al., 1985 June
Hatching early June late June Minnesota Lank et al., 1985
Range
2-153 S
po
tted S
andpiper
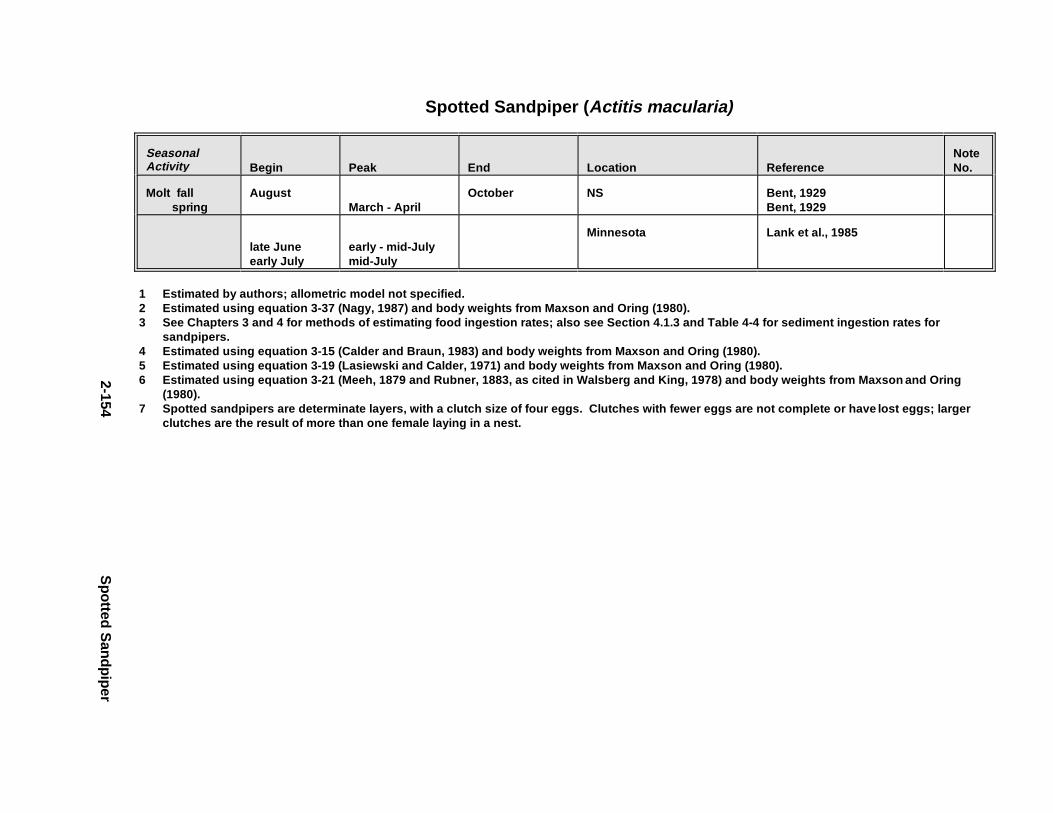
Spotted Sandpiper ( Actitis macularia )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Molt fall August October NS Bent, 1929 spr ing March - Apr il Bent, 1929
Migration Minnesota Lank et al., 1985 females late June early - mid-July males early July mid-July
1 Est imated by author s; allometric model not speci fied.2 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Maxson and Or ing (1980).3 See Chapt ers 3 and 4 f or methods of est imating f ood i ngest ion r ates; also see Sect ion 4.1.3 and Tabl e 4-4 for sedi ment ingest ion r ates for
sandpi pers. 4 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Maxson and Or ing (1980). 5 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Maxson and Or ing (1980). 6 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Maxson and Or ing
(1980). 7 Spot ted sandpi pers are determinate layers, wi th a clutch si ze of four eggs. Clutches wi th fewer eggs ar e not com plete or have lost eggs; larger
clutches ar e the resul t of more than one f emale laying i n a nest .
2-154 S
po
tted S
andpiper

References (including A ppendix)
Bent, A. C. (1929) Life histo ries of North Amer ican shore birds. Part 2. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 146.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Kuenzel, W. J.; Wiegert, R. G. (1973) Energetics o f a sp otted sandpipe r feeding on br ine fl y larvae (Paracoenia; Diptera: Ephydridae) in a thermal spring c ommunit y. Wils on Bull. 85: 473-476.
Lank, D. B.; Oring, L. W.; Maxso n, S. J. (1985) Mate and nutr ient l imi tat ion of egg- laying in a poly andr ous shor ebir d. Ecology 66: 1513-1524.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Maxson, S. J.; Oring, L. W. (1980) Breeding seaso n time and energy budgets o f th e polyandrous sp otted sandpipe r. Behaviour 74: 200-263.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Oring, L. W. (1982) Avian mat ing systems. In: Farner, D. S.; King, J. R., eds. Avian biology , v. 6. New York, NY: Academic Press; pp. 1-92.
Oring, L. W.; Lank, D. B. (1982) Sexual select ion, arrival times, philopa try and s ite fide lit y in the polyandrous sp otted sandpipe r. Behav. Ecol. Sociobiol. 10: 185-191.
Oring, L. W.; Lank, D. B. (1986) Polyandry in spotted sandpipe rs: the impact o f environment and experience. In: Rubenstein, D. I.; Wrangha m, R. W., eds. Ecologic al aspects o f social evolut ion - bir ds and mammal s; pp. 21-42.
Oring, L. W.; Lank, D. B.; Maxso n, S. J. (1983) Popula tion s tudie s of th e poly andr ous spotted sandpipe r. Auk 100: 272-285.
Oring, L. W.; Lank, D. B.; Maxso n, S. J. (1984) Mate and nutr ient l imi tat ion of breeding in the polyandrous sp otted sandpipe r (abstr act only ). Am. Zool. 24: 60A.
2-155 Spotted Sandpipe r

Oring, L. W.; Fiv izzani, A. J.; Halawani, M. E. (1986) Changes in plasma p rolacti n asso ciated wi th laying a nd h atch in the spotted sandpipe r. Auk 103: 820-822.
Oring, L.W.; Colw ell, M.A.; Reed, J.M. (1991a) Lifeti me reproducti ve su ccess i n the spotted sandpipe r (Acti ti s macu laria): sex d iffer ences an d var iance co mpone nts. Behav. Ecol. Sociobiol. 28: 425-432.
Oring, L.W.; Reed, J.M.; Colw ell, M.A.; et al. (1991b) Facto rs regula ting a nnua l mati ng success an d reproducti ve su ccess i n spotted sandpipe rs (Acti ti s macu laria). Behav. Ecol. Sociobiol. 28: 433-442.
Palmer , R. S. (1949) Maine birds. Bull. Mus. Comp. Zool. Harvard No. 102.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Walsberg, G. E.; King, J.R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Zar, J. H. (1968) Standard metabolis m comparisons b etween orders of birds. Condor 10: 278.
2-156 Spotted Sandpipe r

2.1.12. Herring Gull (gulls)
Order Charadriif ormes, Family Larid ae. Gulls ar e medium - to large-sized sea birds wi th long point ed wings , a stout , slight ly hook ed bill, and w ebbed feet. They are abunda nt in temp erate co astal areas and throughout th e Great Lakes. Although gulls may feed from garbage dum ps and la ndf ills , most take n atural prey. Gulls n est p rimar ily in colonie s, although s ome of th e larger sp ecies also nest s olit arily . Many popula tions migr ate annua lly b etween breeding a nd w int ering a reas. North Amer ican gull s pecies range in s ize from Bona parte's gull (33 cm bill t ip t o tail t ip) to the great black-b acked gull (76 cm).
Selected species
The herring gull ( Larus argentatu s) (64 cm) h as th e largest r ange of any North Amer ican gull, from Newfoundla nd s out h to the Chesapeake Bay along t he nor th Atlant ic and west th roughout th e Great Lakes i nto Alaska. Along t he Paci fic co ast, the simi lar-sized wester n gull ( L. occidentalis) is th e ecologic al equiv alent of th e herring gull. Bot h species take p rimar ily natural foods , especially fi sh, although s ome indiv idua ls of bot h species forage around f ishing ope rations and la ndf ills (Pi erot ti , 1981, 1987; Pierotti and Annett, 1987). The increase in number of herring gulls in t his cen tury has been attr ibuted to the inc reasing a bunda nce of year -round f ood s upplie s found in la ndf ills (D rury, 1965; Harris, 1970); however , birds sp ecializing on ga rbage have su ch low reproducti ve su ccess th at they cannot r eplace th emsel ves i n the popula tion ( Pierot ti and Annett, 1987, 1991). An alternative expla nation of th e species' expansion is th at cessat ion of taking of gulls by th e feath er indus try in t he late 1800's has allow ed gull num bers to return to pre-exploi tati on levels (Graham, 1975).
Body size. Adult femal es (800 to 1,000 g) are signif icantly smal ler than males (1,000 to 1,300 g) in both the herring gull ( Greig et al., 1985) and the wester n gull ( Pierot ti , 1981). Chicks g row fr om th eir hatching w eight of about 60 to 70 g to 800 to 900 g wi thin 3 0 to 40 days, after which time th eir weight stabilize s (Dunn a nd Br isbin, 1980; Norstr om et al ., 1986; Pierotti , 1982). Norstr om et al . (1986) fi tted chick g rowth rates to the Gompertz equation a s follow s:
BW = 997 e-e(-0.088(t - 14.8)) for femal es, and
BW = 1193 e-e(-0.075(t - 16.3)) for males,
where BW equals body weight in gr ams an d t equals days after hatching. Adult s show seaso nal var iation in body weight (Couls on et al., 1983; Norstr om et al ., 1986).
Habitat . Nesting c olonie s of herring gulls along t he nor theaster n coast o f th e Unit ed States ar e found pr imarily on s andy or rocky o ffsh ore or barrier beach islands (Kadlec and Drury, 1968). In the Great Lakes, they are found on t he more remote, seclude d, and protected islands and shorelines of th e lakes an d their conne cting r ivers (Weseloh, 1989). Smalle r colonie s or isolated pairs also can be found in c oastal marshes (Burger, 1980a), penins ulas, or cli ffs along seaco asts, lakes, and rivers (Weseloh, 1989), and occas iona lly in inla nd areas or on buildings or piers (Harris, 1964). Gulls ar e the most abunda nt seabir ds
2-157 Herring G ull

of fshor e from fall t hrough s pring, and are only found pr edom inant ly ins hor e dur ing t he breeding seaso n in late sp ring a nd s ummer (Powers, 1983; Pierotti , 1988). Gulls fo rage predom inant ly of fshor e, wi thin 1 to 5 km of th e coast (Pi erotti , 1988). In all seaso ns th e num ber of bir ds feeding a t sea outnumber those feeding ins hor e (data fr om Powers, 1983; Pierot ti , pers. co mm.). Inshor e, herring gulls fo rage pr imarily in int ertida l zone s but al so search for food in wet fi elds, around l akes, bays, and rock j etti es, and at landfills in s ome areas (Burger, 1988). In Flor ida, herring gull pr esence at l andfills is r estr icted to the winter months (Decemb er through A pril) and may co nsist p rimar ily of fi rst-year birds th at migrated from mo re northerly popula tions (e. g., from th e Great Lakes) (Patto n, 1988).
Food ha bi ts . Gulls feed on a var iety o f foods depending on availa bilit y, inc luding fish, squid, crustacea, mollu scs, worms, insects, smal l mammal s and birds, duck an d gull eggs and chicks, and garbage (Bour get, 1973; Burger, 1979a; Fox et al ., 1990; Pierotti and Annett, 1987). Gulls fo rage on ope n water by aer ial dipping a nd s hallow div ing a round conc entrations of prey. At sea, such concentrations o ften are asso ciated wi th whales or dolphins , other seabir ds, or fishing bo ats (McCleery and Sibly , 1986; Pierotti , 1988). In the Great Lakes, concentrations of sp ecies such as alewife oc cur seaso nally (e.g., when spawning) (Fox et al., 1990). Gulls al so forage by stea ling f ood f rom other bir ds and by scave nging a round hum an refuse si tes (e.g., garbage dumps, fish plants, docks, and seasi de parks) (B urger and Gochfeld, 1981; 1983; Chapman and Parker , 1985). Indiv idua l pairs of gulls may sp ecialize predom inant ly on a single typ e of food; for examp le, three quarters of a popula tion of herring gulls in Newfoundla nd w ere found t o specialize ei ther on blue mu ssel s, garbage, or adults o f Leach's sto rm-petrel, wi th 60 percent of th e specialists co ncentrating on m ussel s between 0.5 and 3 cm i n length (Pierotti and Annett, 1987; 1991). Diet choices may ch ange with the age and experience of adult bir ds as well a s wi th availa bilit y of prey (Pierot ti and Annett, 1987; 1991). Females take smal ler prey and feed less o n garbage than do males (Pierotti , 1981; Greig et al ., 1985). For examp le, Fox et al. (1990) found femal es to feed more on smel t (100 to 250 mm) an d males mo re on alewife (250 to 300 mm) i n the Great Lakes r egion. Adult gulls so meti mes attack an d eat ch icks o f neighbor ing gulls or other sp ecies of seabird (Brown, 1967; Schoen and Morris, 1984). Juveniles up to 3 years of age forage less effi ciently th an adults (Gr eig et al ., 1983; MacLean, 1986; Verbeek, 1977). In the Great Lakes, herring gulls ' high c ons umpt ion of alewife dur ing t heir sp awn may r esult in high e xpos ures of th e gulls to lipophilic contami nants th at biomagnify (Fox et al ., 1990).
Metabolism. Norstr om et al . (1986) have esti mated an annua l energy budge t for free-liv ing femal e herring gulls th at br eed in t he Great Lakes an d an annua l energy budge t for fr ee-liv ing juv enile herring gulls in t he Great Lakes i n their fi rst year . Between Septemb er and March, the nonbr eeding seaso n, they esti mate th at adult femal es requir e 250 to 260 kcal /day. Follow ing a dip in e nergy r equir ements to 210 kcal /day wh en the male feeds th e femal e during c our tship, the femal e's needs increase to peak at 280 kcal /day fo r egg pr oduc tion, then fall t o appr ox imatel y 210 kcal /day during inc ubation. The energy requir ed to forage fo r food f or th e chi cks i s su bstan tial, rising t hrough J uly to peak in August at 310 to 320 kcal /day, then declining a gain unt il Septemb er when feeding c hic ks has ceased . These esti mates co mpare wel l wi th those derived from Nagy's (1987) equation to esti mate fr ee-living metabolic r ates fo r seabirds, excep t th at th e energy peaks r equir ed to produce eggs and to feed chicks ar e not include d in Na gy 's mode l. Readers int erested in the metabolic r ates o f fi rst-year herring gulls ar e refer red to Norstr om et al . (1986). Ellis
2-158 Herring G ull

(1984) provides an overview of seab ird energetics an d addit iona l dis cussion of appr oaches and models fo r esti mat ing metabolic r ates o f fr ee-ranging seabir ds.
Mol t. Gull c hicks ar e downy gray wi th dark brown spott ing a nd m olt int o a dark-gray or brown mottl ed juv enile plum age. At the end of th e fi rst year , por tions of th e plumage have paled, and by th e second year , gray plum age develops along t he back an d top of wings . By th eir thir d year , young gulls r esemb le dirty adult s, and t hey acquir e their full a dult plum age by 4 years (Harrison, 1983; Kadlec and Drury, 1968). Adult gulls , at least in s ome popula tions , begin t heir pr imary feath er molt during inc ubation a nd c ompl ete th e molt by mi d- to late fal l (Couls on et al., 1983). They mo lt and replace th e large body feath ers fr om mi d-summer to early fal l (Couls on et al., 1983).
Migration. Herring gull popula tions along t he nor theast co ast o f North Amer ica ten d to be migr atory, while adult herring gulls of th e Great Lakes ar e year -round r eside nts. Along t he wester n North Atlantic, most h erring gulls ar rive on t heir breeding gr ounds between late February and late April. They remain unti l late August o r early Septemb er when they leave fo r their winter ing gr ounds along t he Atlant ic and G ulf co asts o r wel l offsh ore (Burger, 1982; Pierotti , 1988). Adult and olde r suba dult herring gulls in t he Great Lakes ar ea are essen tially nonm igr atory (Mineau et al., 1984; Weseloh e t al., 1990). Thus, in contrast to other fish-eating bir ds in t he Great Lakes system th at mi grate so uth in the wint er, herring gulls ar e expos ed to any con tami nants th at may b e in Great Lakes' fish throughout th e year (Mineau et al ., 1984). Postb reeding dis persal away fr om breeding colonie s begins in l ate Ju ly and ends in Augus t, wi th all ages traveling s hor t di stan ces. Great Lakes h erring gulls l ess th an a year old us ually migr ate to the Gulf or Atlantic co ast (Smith, 1959; Mineau et al ., 1984), traveling a long r iver systems an d the coast (Moor e, 1976).
Breeding acti vi ties and social organization. Gulls n est p rimar ily in colonie s on offsh ore islands, and nest d ensi ty i s str ongly affected by popula tion s ize (Pierot ti , 1981; 1982; 1987). Typical ly, males ar rive at th e breeding gr ounds fi rst and establis h territories. Both sexes build t he nest o f veg etat ion on t he ground in a reas th at are sheltered from wi nd but may b e exposed to the sun (Pierotti , 1981; 1982). Males feed femal es fo r 10 to 15 days prior to the star t of egg laying ( Pierot ti , 1981). From th e laying of th e fi rst egg unt il t he chicks ar e 3 to 4 weeks old, one or bot h parents will be present at al l times (T inbe rgen, 1960). Males perform mo st ter ritorial defense, femal es perform mo st i ncubation, and bot h parents feed the chicks u nti l they are at least 6 to 7 weeks old ( Bur ger, 1981; Pierotti , 1981; Tinbe rgen, 1960). All gulls ar e str ongly monoga mous ; pair bonds can persis t for 10 or more years and usually only ar e termin ated by th e death of a mate o r failur e to reproduc e successf ully (T inbe rgen, 1960). Males may b e promiscuous in popula tions wi th more femal es th an males (Pierotti , 1981). Herring gull c olonie s often are found in asso ciation wi th colonie s of other sp ecies, inc luding ot her gulls (Bour get, 1973; Brown, 1967). In some nesting c olonie s, gulls attack ch icks o f neighbor ing gulls and ot her sp ecies (Brown, 1967; Schoen and Morris, 1984).
Home range and r esour ces. Dur ing t he breeding seaso n, herring gulls d efend a ter ritory of sever al tens of sq uare meter s around t he immed iate vi cinity o f th e nest (B urger, 1980b). Their daily fo raging r ange depends on t he availa bilit y of prey and on t he foraging str ategy, age, and sex o f th e gull. Using r adiot elemetr y on gulls in t he Great Lakes, Morris
2-159 Herring G ull

�
�
�
and Black (1980) d emonstr ated that so me parents wi th chicks fo rage at sp eci fic locat ions wi thin 1 km of th e colony wh ereas other parents make exten ded flight s to destina tions across a l ake mo re than 30 km away. Simi larly, gulls th at feed at sea may r ange tens of ki lometer s fr om th eir nest wh ereas gulls fr om th e same c olony feeding in t he int ertida l zone may tr avel less th an 1 km (Pi erotti and Annett, 1987; 1991). Males typ ical ly range far ther than femal es and take l arger prey i tems (Pi erotti and Annett, 1987; 1991). At sea dur ing t he nonbr eeding seaso n, gulls may r ange hundr eds of kilo meter s during a day (Pierotti , pers. comm.).
Popul ation de nsity . As descr ibed above, popula tion de nsity is d etermined by availa ble nesting s pace, size of th e breeding popula tion, and qua lit y of habitat. Smal l is lands wi th good feeding a reas nearby can have sever al hundr ed nests p er hectar e (Kadlec, 1971; Parsons, 1976b; Pierotti , 1982). In poor qualit y habitat, some pairs nest solit arily wi thout anot her nest fo r sever al kilometer s (Weseloh, 1989).
Popul ation dy namics. Herring gulls and wester n gulls usually do not begin breeding unt il a t least 4 year s of age for males and 5 years of age for femal es (Burger, 1988; Pierotti , 1981; Pierotti , pers. comm.). Kadlec and Drury (1968) suggest th at in a given year , 15 to 30 percent of adult s of breeding a ge do not breed. Most br eeding femal es produce th ree-egg clutch es, but indiv idua ls in poor condit ion may l ay only one or two eggs (Parsons, 1976a; Pierotti , 1982; Pierotti and Annett, 1987; 1991). Herring gulls will la y replacemen t eggs i f al l or a portion of th eir or igina l c lut ch is d estr oyed (Parsons, 1976a). Hatching s uccess ap pears to be influenced by femal e diet, wi th garbage specialists hatching a smal ler percentage of eggs th an fish or inter tidal (mussel ) specialists (Pi erotti and Annett, 1987, 1990, 1991). Predation, often by gulls of th e same o r other species, also contr ibutes to egg losses (Payn ter , 1949; Harris, 1964; Davis, 1975). Many herring gull chicks th at hatch die before fledging, most wi thin t he fi rst 5 days after hatching ( Harris, 1964; Kadlec et al ., 1969; Brown, 1967). Adult mo rtalit y is low (ar ound 1 0 percent per year ), and some birds may l ive up to 20 years (Brown, 1967; Kadlec and Drury, 1968). Subadult bir ds exhibit highe r mo rtalit y (20 to 30 percent per year ) (Kadlec and Drury, 1968; Chabrzyk and Couls on, 1976).
Simi lar species (fr om general references)
� The wester n gull ( Larus occi dental is) (64 cm), found on t he Paci fic co ast o f the United States, is th e ecologic al equiv alent of th e herring gull a nd is simi lar in size (53 cm); males range from 1,000 to 1,300 g and femal es fr om 800 to 1,000 g (Pierotti , 1981).
� The glaucous gull ( Larus hyperbor eus) is larger (69 cm) th an the herring gull and is th e predom inant gull br eeding in t he high a rcti c. Bir ds fr om A laska are slight ly smal ler than birds fr om easter n Canada.
� The glaucous -winge d gull ( Larus glaucescen s) is si mila r in s ize to the herring gull (66 cm) an d is th e primary breeding s pecies nor th of th e Colum bia River. This sp ecies hy br idize s exten sively wi th the herring gull in Alaska.
2-160 Herring G ull

�
�
�
�
� The Calif ornia gull ( Larus cal ifornicus) is smal ler (53 cm) th an the herring gull. This sp ecies br eeds pr imarily in t he Great Basin Deser t and winters along t he Paci fic co ast.
� The great black-b acked gull ( Larus mar inus ) is th e largest sp ecies of gull ( 76 cm) i n North Amer ica and breeds fr om Labrador to Long Is land.
� The ring- bille d gull ( Larus delawarensis) is of aver age size (45 cm) an d is th e most common br eeding gull in t he Great Lakes an d northern prairies.
� Franklin' s gull ( Larus pipixcan ) is a smal l (37 cm), summer resident of th e Great Plains.
General references
For general inf ormation: Harrison (1983); Nationa l Geogr aphic So ciety (1987); Tinbe rgen (1960); Graham (1975). For discu ssion of die t: Bur ger (1988); Fox et al . (1990); Pierotti (1981); Pierotti and Annett (1987).
2-161 Herring G ull

Herring Gull ( Larus ar gentatus )
2-162 H
erring Gull
Factors Age/Sex Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A F spr ing 951 ± 88 SD Lake Hur on Norstrom et al., 1986 (g) A M spr ing 1,184 ± 116 SD
A F sum mer 999 ± 90 SD 832 - 1,274 Newfoundl and Threlfall & Jewer , 1978 A M sum mer 1,232 ± 107 SD 1,014 - 1,618
at hatching 65 50 - 80 Maine Dunn & Br isbin, 1980 10 days old 230 120 - 380 20 days old 590 420 - 800 30 days old 810 610 - 1,000
30 days old 964 ± 77 SD Newfoundl and/rocky island Pierot ti, 1982 1 30 days old 818 ± 99 SD Newfoundl and/grassy island
Chick Gr owt h < 5 days 8.8 - 13.1 Newfoundl and/ island Pierot ti, 1982 Rate (g/day) 5-30 days 26.3 ± 6.5 SD Newfoundl and/ island Pierot ti, 1982
5-30 days 33.4 ± 4.7 SD Newfoundl and/rocky island Pierot ti, 1982
5-25 days 30.2 ± 1.75 SD 26.7 - 31.4 Maine/coast al island Hunt , 1972
meadow
Egg Weight (g) 3 egg cl utch 87.2 New Brunsw ick Herbert & Bar clay, 1988 2 egg cl utch 85.7
in 1983 92.0 ± 5.9 SD Lake Super ior , Canada Meathrel et al., 1987 in 1984 98.0 ± 8.0 SD
Metabol ic Rate A M basal 86 est imated 2 (kcal /kg-day) A F basal 91
A standard 99 laboratory Lustic k et al., 1978
A M free- 233 (84 - 646) liv ing A F f ree- 248 (92 - 669) est imated 3 liv ing
Also see t ext for a discussi on of annual var iation i n free-living m etabol ic rate in herring gul ls.

2-163 H
erring Gull
Herring Gull ( Larus ar gentatus )
Factors Age/Sex Range or Cond. /Seas. Mean (95% CI of mean)
Food Ingest ion A M breeding 0.20 Rate (g/g-day) A F breeding 0.21
A M breeding 0.19 A F br eeding 0.18
Water A M 0.055 Ingest ion Rat e A F 0.059 (g/g-day)
Inhal ation Rat e A M 0.48 (m /day) A F 0.41 3
Surface Ar ea A M 1,150 (cm ) A F 1,001 2
Note Locat ion Reference No.
Newfoundl and - Pierotti & Annet t, 1991 4 diet of mussel s
Newfoundl and - Pierotti & Annet t, 1991 5 diet of garbage
est imated 6
est imated 7
est imated 8
Dietary Compositio n Summer Summer Summer Summer (measur e) Reference No.
Locat ion/Habi tat Note
mont hs: Mid-May/ Mid-June/ Mid-July/ Newfoundl and/ island Haycock & Thr elfall, 1975
Mytilus edul is 30.9 0.9 9.1 (% occur rence i n sea ur chi n 5.8 0.0 4.5 regur gi tations and fish 11.4 71.1 18.9 pellets) Oceanodr oma 22.4 7.0 15.9 leuchor hoa
Fratercul a arct ica 5.8 0.0 1.5 adults Fratercula, Uria 0.0 3.5 9.1 chi cks Larus sp. eggs 3.1 0.9 0.8 Vacci num - - 9.9 angust ifolium Gadus m orhua 12.4 1.7 14.4 offa l assor ted refuse 5.8 0.9 6.8
Mid-June Mid-Jul y Mid-Aug.

Herring Gull ( Larus ar gentatus )
Dietary Compositio n Summer Summer Summer Summer (measur e) Reference No.
Locat ion/Habi tat Note
year: 1978 1979 1980 1981 Lake Ontario Fox et al., 1990 American sm elt 46.1 18.4 61.2 57.8 alewife 23.1 73.7 16.7 23.4 (% occur rence i n other fish 20.5 0 3.4 3.1 regur gi tations f rom and birds 2.6 2.6 13.8 6.2 stomach cont ents of vol es 2.6 2.6 3.4 9.4 incubat ing adul ts) insect s & r efuse 12.8 0 3.4 0
lake: Ontario Erie Huron Super ior Great Lakes Fox et al., 1990 fish 91.8 94.1 75.8 38.6 insect s 5.5 5.9 5.6 42.1 (% occur rence i n bol i offal, garbage 0.5 2.9 13.6 21.0 regur gi tated by chi cks) gul l chi cks 2.2 0 1.0 0 adult b irds 1.6 0 1.0 3.5 amphi bians 0.5 0 0 0 earthworms 2.2 0 11.6 1.7 crayfish 0 0 0.5 0
snai ls 3 CA,FL,NY,NJ,TX/ Burger, 1988 crabs 14 coast al garbage 27 offal 5 (% of gul ls feeding on i tems) worms 23 other inverts. 28 offshor e feeding on f ish was fish unknown not included i n obser vat ions
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Foraging A M 10 to 15 3 - 50 NS/coast al Pierot ti, pers. com m. Radius (km) A F 5 to 10 3 - 25
Popul ation summer 227 138 - 350 Massachuset ts/coast al Kadl ec, 1971 Densi ty is lands (nests/ha)
sum mer 217 Newfoundl and/ island - rocky Pierot ti, 1982 75 Newfoundl and/ island - Pierotti, 1982
grassy slope
2-164 H
erring Gull

Herring Gull ( Larus ar gentatus )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Clutch 2.78 2.51 - 2.90 New Jersey/sal t marsh Burger, 1979b Size (over 8 si tes) islands
2.54 1 - 6 NE Uni ted States/coast al Nisbet & Drury, 1984
2.38 2.3 - 2.8 Maine/coast al islands Hunt , 1972
2.84 ± 0.44 SD Lake Super ior, Canada/ Meathrel et al., 1987
(per nest )
(over 11 years)
is lands
Clutches/ Year 1 1 - 2* (* if first eggs l ost) Burger, 1979a; Bour get, 1973
Days 30.5 28 - 33 Hol land/NS Tinber gen, 1960 Incubat ion 29 Newfoundl and/ island Pierot ti, 1982 9
Age at 51 35 - 44 to 56 - 61 Massachuset ts/coast al Kadl ec et al., 1969 Fledgi ng 43 31 to 52 island Paynter, 1949 (days) New Brunswick/is land
Number 3 col oni es 1.42 1.40 - 1.44 New Jersey/coast al Bur ger & Shi sler, 1980 Fledge per 6 col ony -yrs 1.65 1.40 - 2.13 Lake Ontario/lakeshor e Mineau et al., 1984 Act ive Nest 3 col ony -yrs 1.78 1.62 - 2.10 Lake Er ie/lakeshor e (minimum and m aximum are
6 col ony -yrs 2.19 2.16 - 2.25 Lake Hur on/ lakeshor e yearly means)
Number 3 col oni es 1.80 1.79 - 1.80 New Jersey/coast al Bur ger & Shi sler, 1980 Fledge per Successf ul Nest
Age at F 5 years throughout range/NS Greig et al., 1983; Pierot ti, Sexual M 4 - 5 years pers. com m. Maturity
B 4.3 to 5.8 3 - 8 Scotland/coastal Coulson et al., 1982
Annual A B 8 New Engl and/coast al Kadl ec & Dr ury, 1968 Mortality Rates J B 22 17 - 33 (percent ) A B 7.3 Scot land/coast al Chabr yzk & Coul son, 1976
2-165 H
erring Gull

Herring Gull ( Larus ar gentatus )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Longevi ty A B 10 up t o 30 years NS/NS Pierot ti, pers. com m.
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/ late Apr il early May early June ne shor e Lake Super ior Morris & Hay mes, 1977 Laying early May mid-May early June Maine Bour get, 1973
early May mid-May mid-June New Jersey Bur ger, 1977, 1979b early May late May end May Newfoundl and Pierotti, 1982
Hatching mid - late May Great Lakes Fox et al., 1990 May June Jul y Massachuset ts Kadl ec, 1971 early June mid-June end June Newfoundl and Pierotti, 1982, 1987 late June late June mid-Jul y New Brunsw ick Paynter, 1949
Migration spr ing February late Apr il nor thwestern At lant ic Bur ger, 1982
fall August September popul ations
Molt June Jul y August Newfoundl and Pierotti, pers. com m.
2-166 H
erring Gull
1 Weight of chi cks f rom first egg l aid in 1978 for the rocky island and i n 1977 for the grassy area. In som e years and som e loc ations, chi cks f rom the first egg w ere heavi er than the rest , and at other times and l ocat ions, the first chi ck w as l ight er.
2 Est imated usi ng equat ion 3-29 (Lasiewski and Dawson, 1967) and body weight s f rom Threlfall and Jewer (1978). 3 Est imated usi ng equat ion 3-38 (Nagy, 1987) and body weight s f rom Threlfall and Jewer (1978). 4 Est imated usi ng 11.2 meals of mussel consum ed per day per pair, weight of 80 g per mussel meal of which hal f is shel l and not included i n
ingest ion r ate, assum ing t hat the female account s for 46 percent of pair's energy requi rement and t he male account s for 54 perc ent, and usi ng the body weight s of Threlfall and Jewer (1978).
5 Est imated usi ng 4.2 meals of garbage consum ed per day per pair, weight of 100 g per garbage meal, assum ing t hat the female acc ount s for 46 percent of pair's energy requi rement and t he male account s for 54 percent , and usi ng t he body weight s of Threlfall and Jewer (1978).
6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Threlfall and Jewer (1978). 7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Threlfall and Jewer (1978). 8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Threlfall and Jew er
(1978). 9 Beginni ng wi th first egg.

References (including A ppendix)
Belopol' skii, L. O. (1957) (Ecology of sea colony bir ds of th e Barents Sea). Translated by: Israel Progr am for Sci enti fic Translations , Jerusalem (ci ted in Dunning, 1984).
Bour get, A. A. (1973) Relation of eide rs and gulls n esting in m ixed colonie s in P enobs cot Bay, Maine. Auk 90: 809-820.
Brown, R. G. (1967) Breeding s uccess an d popula tion gr owth in a colony of herring gulls and lesser black-b acked gulls Larus argentatu s and L. fuscu s. Ibis 109: 502-515.
Burger, J. (1977) Nest ing be havior of herring gulls : invasion int o Spartina sal t mar sh areas of New Jer sey. Condor 79: 162-169.
Burger, J. (1979a) Competi tion a nd pr edation: herring gulls ver sus laughing gulls . Condor 81: 269-277.
Burger, J. (1979b) Colony size : a test fo r breeding s ynchrony in he rring gull ( Larus argentatu s) colonie s. Auk 96: 694-703.
Burger, J. (1980a) Nest ing a daptation of herring gull ( Larus argentatu s) to sal t mar shes and sto rm tide s. Biol. Behav. 5: 147-162.
Burger, J. (1980b) Territory si ze differ ences i n relation t o reproduc tive stag e and typ e of int ruder in he rring gulls ( Larus argentatu s). Auk 97: 733-741.
Burger, J. (1981) On becoming inde pendent in he rring gulls : parent-young c onf lic t. Am. Nat. 117: 444-456.
Burger, J. (1982) Herring gull. In: Davis, D. E., ed. CRC handbook of cen sus methods fo r ter restr ial ver tebrates. Boca Raton, FL: CRC Press; pp. 76-79.
Burger, J. (1988) Foraging be havior in gulls : di ffer ences i n method, prey, and ha bitat. Colonia l Waterbirds 11: 9-23.
Burger, J.; Gochfeld, M. (1981) Age-related differ ences i n piracy b ehavior of fo ur species of gulls , Larus. Behaviour 77: 242-267.
Burger, J.; Gochfeld, M. (1983) Behavior of nine avi an species at a Flor ida garbage dum p. Colonia l Waterbirds 6: 54-63.
Burger, J.; Shisler, J. (1980) The process o f colony fo rmation a mong he rring gulls Larus argentatu s nesting in Ne w Jersey. Ibis 122: 15-16.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
2-167 Herring G ull

Chabrzyk, G.; Couls on, J. C. (1976) Survival and recruitmen t in the herring gull Larus argentatu s. J. Anim. Ecol. 45: 187-203.
Chapman, B.-A.; Parker , J. W. (1985) Foraging a reas, technique s, and s chedule s of wint ering gulls on s out heaster n Lake Er ie. Colonia l Waterbirds 8: 135-141.
Couls on, J. C.; Dunc an, N.; Thomas, C. (1982) Changes in the breeding biology of th e herring gull ( Larus argentus) induc ed by r educ tion in s ize and de nsity of th e colony . J. Anim. Ecol. 51: 739-756.
Couls on, J. C.; Monaghan, P.; Butter field, J.; et al . (1983) Seasonal changes in the herring gull in Br itain: weight , moult and m ortalit y. Ardea 71: 235-244.
Davis, J. W. (1975) Age, egg-size and breeding s uccess i n the herring gull Larus argentatu s. Ibis 117: 460-473.
Drury, W. H., Jr . (1965) Clash of co astal nester s. MA: Audubon.
Dunn, E. H. (1976) The developm ent of endot hermy and existen ce energy exp endit ure in herring gull c hicks. Condor 78: 493-498.
Dunn, E. H. (1980) On the var iabilit y in e nergy allo cation of n estling bir ds. Auk 1: 19-27.
Dunn, E. H.; Br isbin, I. L. (1980) Age-speci fic ch anges in the major body co mpone nts and calor ic value s of herring gull c hicks. Condor 82: 398-401.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Ellis , H. I. (1984) Energetics o f fr ee-ranging seabir ds. In: Whit tow, G. C.; Rhan, H., eds. Seabird energetics. New York, NY: Plenum Press; pp. 203-234.
Erwin, R. M. (1971) The breeding s uccess o f two species of symp atr ic gulls , the herring gull a nd t he great black-b acked gull. Wils on Bull. 83: 152-158.
Ewins , P. J.; Weseloh, D. V.; Groom , J. H.; et al. (unpublis hed 1991) The diet of herring gulls ( Larus argentatu s) dur ing w int er and e arly sp ring on t he low er Great Lakes (unpublis hed manus cr ipt , Canadian Wildlif e Service, Bur lingt on, Ontar io) .
Fox, G. A.; Allan, L. J.; Weseloh, D. V., et al. (1990) The diet of herring gulls dur ing t he nesting pe riod in Ca nadian water s of th e Great Lakes. Can. J. Zool. 68: 1075-1085.
Graham, F. (1975) Gulls : a social his tory. New York, NY: Random Hous e.
Greig, S. A.; Couls on, J. C.; Monaghan, P. (1983) Age-related differ ences i n foraging success i n the herring gull ( Larus argentatu s). Anim. Behav. 31: 1237-1243.
2-168 Herring G ull

Greig, S. A.; Couls on, J. C.; Monaghan, P. (1985) Feeding s trategies of mal e and femal e adult herring gulls Larus argentatu s. Behaviour 94: 41-59.
Gross, A. O. (1940) The migration of Kent Island he rring gulls . Bir d-Banding 1 1: 129-155.
Harris, M. P. (1964) Aspects o f th e breeding biology of th e gulls : Larus argentatu s, L. fuscu s, and L. mar inus . Ibis 106: 432-456.
Harris, M. P. (1970) Rates an d causes o f increases o f so me British gull popula tions . Bir d Study 17: 325-335.
Harrison, P. (1983) Seabirds: an identi ficat ion guide . Bos ton, MA: Hought on-Mif flin Co.
Hayco ck, K. A.; Threlfal l, W. (1975) The breeding biology of th e herring gull in Newfoundla nd. Auk 92: 678-697.
Hebert, P. N.; Barclay, R. M. (1986) Asyn chronous and s ynchronous h atching: effect o n early growth and s urviv orship of herring gull, Larus argentatu s, chicks. Can. J. Zool. 64: 2357-2362.
Hebert, P. N.; Barclay, R. M. (1988) Parental investmen t in herring gulls : clut ch appor tionm ent and c hic k surviv al. Condor 90: 332-338.
Holle y, A. J. (1982) Post-fl edging int eractions on t he ter ritory between parents an d young herring gulls Larus argentatu s. Ibis 124: 198-203.
Hunt , G. L. (1972) Influence of food dis tr ibut ion a nd hum an dis turbance on t he reproduc tive success o f herring gulls . Ecology 53: 1051-1061.
Kadlec, J. A. (1971) Effects o f introduc ing f oxes an d raccoons on he rring gull c olonie s. J. Wildl. Manage. 35: 625-636.
Kadlec, J. A. (1976) A re-evaluation of mo rtalit y rates i n adult herring gulls . Bir d-Banding 47: 8-12.
Kadlec, J. A.; Drury, W. H. (1968) Structu re of th e New Engla nd he rring gull popula tion. Ecology 49: 644-676.
Kadlec, J. A .; Drury, W. H.; Onion, D. K. (1969) Growth and mortal ity o f herring gull c hicks. Bir d-Banding 4 0: 222-233.
Keith, J. A. (1966) Reproduc tion in a popula tion of herring gulls ( Larus argentatu s) contami nated by DDT. J. Appl. Ecol. 3(suppl. ): 57-70.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
2-169 Herring G ull

Lasiewski , R. C.; Dawso n, W. R. (1967) A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lusti ck, S.; Batter sby, B.; Kelty, M. (1978) Behavioral thermoregula tion: or ientation t oward the sun in he rring gulls . Science 200: 81-83.
Lusti ck, S.; Batter sby, B.; Kelty, M. (1979) Effects o f insolation on juv enile herring gull energetics and be havior . Ecology 60: 673-678.
MacLean, A. A. (1986) Age-speci fic fo raging a bilit y and t he evolut ion of d efer red br eeding in t hree species of gulls . Wils on Bull. 98: 267-279.
McCleery, R. H.; Sibley, R. M. (1986) Feeding s pecialization a nd pr eference in herring gulls . J. Anim. Ecol. 55: 245-259.
Meathrel, C. E.; Ryder, J. P.; Termaat, B. M. (1987) Size and composi tion of herring gull eggs : relations hip t o pos it ion in t he laying s equence and t he body condit ion of femal es. Colonia l Waterbirds 10: 55-63.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Mendall, H. L. (1939) Food ha bit s of th e herring gull in r elation t o freshwater game fi shes in Maine. Wils on Bull. 41: 223-226.
Mineau, P.; Fox, G. A.; Norstr om, R. J.; et al . (1984) Using t he herring gull t o monit or levels and effects o f organochlor ine con tamina tion in t he Canadian Great Lakes. In: Nriagu, J. O.; Simmo ns, M. S., eds. Toxic co ntami nants i n the Great Lakes. New York, NY: John W iley & Sons ; pp. 425-452.
Moor e, F. R. (1976) The dynamics o f seaso nal distr ibut ion of Gr eat Lakes h erring gulls . Bir d-Banding 4 7: 141-159.
Morris, R. D.; Black, J. E. (1980) Radiotelemetr y and herring gull f oraging p atter ns. J. Field Ornit hol. 51: 110-118.
Morris, R. D.; Haymes, G. T. (1977) The breeding biology of two Lake Er ie herring gull colonie s. Can. J. Zool. 55: 796-805.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Niebuhr, V. (1983) Feeding s trategie s and inc ubation be havior of wild he rring gulls : an experiment us ing ope rant feeding bo xes. Anim. Behav. 31: 708-717.
2-170 Herring G ull

Nisbet, I. C.; Drury, W. H. (1984) Super-normal clutch es in herring gulls in New Engla nd. Condor 86: 87-89.
Norstr om, R. J.; Clark, T. P.; Kearney, J. P.; et al . (1986) Herring gull e nergy r equir ements and body cons ti tuents in t he Great Lakes. Ardea 74: 1-23.
Olsso n, V. (1958) Dispersal , migration, longe vi ty and d eath causes o f Str ix aluco, Buteo buteo, Ardea cinerea and Larus argentatu s. A stu dy based on recover ies of birds ringe d in Fe nno -Scandia . Acta Ver tebratica 1: 91-189.
Parsons, J. (1972) Egg size, laying d ate and incubation pe riod in t he herring gull. Ibis 114: 536-541.
Parsons, J. (1976a) Facto rs determining t he num ber and s ize of eggs laid by th e herring gull. Condor 78: 481-482.
Parsons, J. (1976b) Nest ing de nsity and br eeding s uccess i n the herring gull Larus argentatu s. Ibis 118: 537-546.
Patto n, S. R. (1988) Abunda nce of gulls at Tampa Bay landf ills . Wils on Bull. 100: 431-442.
Paynter , R. A. (1949) Clut ch size and t he egg and c hic k mortalit y of Kent Island he rring gulls . Ecology 30: 146-166.
Pierotti , R. (1981) Male and femal e parental roles in the wester n gull unde r di ffer ent environm ental condit ions . Auk 98: 532-549.
Pierotti , R. (1982) Habitat sel ect ion a nd it s effect o n reproduc tive out put in t he herring gull in Newfoundla nd. Ecology 63: 854-868.
Pierotti , R. (1987) Behavioral consequences o f habitat sel ect ion in t he herring gull. Studie s Avian Biol. 10: 119-128.
Pierotti , R. (1988) Asso ciations b etween mar ine birds and mammal s in the northwest Atlantic Ocean . In: Burger, J., ed. Seabirds and other mar ine ver tebrates. New York, NY: Colum bia Univ ersi ty Press; pp. 31-58.
Pierotti , R.; Annett, C. (1987) Reproducti ve co nsequences o f dietary sp ecialization a nd swi tching in a n ecologic al generalis t. In: Kamil, A. C.; Krebs, J. ; Pullia m, H. R., eds. Foraging be havior . New York, NY: Plenum Pr ess; pp. 417-442.
Pierotti , R.; Annett, C. A. (1990) Diet and reproduc tive out put in s eabir ds: food c hoi ces b y indiv idua l, free-liv ing a nim als can affect su rvival of offsp ring. BioS ci . 40: 568-574.
Pierotti , R.; Annett, C. A. (1991) Diet choic e in t he herring gull: cons traint s im pos ed by reproduc tive and ecologic al facto rs. Ecology 72: 319-328.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
2-171 Herring G ull

Powers, K. D. (1983) Pelagic distr ibut ions of mar ine bir ds of f the nor theaster n U.S. NOAA, Tech. Mem. NMFS-F/NED-27: 1-201.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Schoen, R. B.; Morris, R. D. (1984) Nest sp acing, colony location, and br eeding s uccess i n herring gulls . Wils on Bull. 96: 483-488.
Sibly , R. M.; McCleery, R. H. (1983) Increase in weight of herring gulls while feeding. J. Anim. Ecol. 52: 35-50.
Smith, W. J. (1959) Movemen ts o f Michiga n herring gulls . Bir d-Banding 3 0: 69-104.
Threlfal l, W.; Jewer , D. D. (1978) Notes o n the stan dard body measu rements o f two popula tions of herring gulls ( Larus argentatu s). Auk 95: 749-753.
Tinbe rgen, N. (1960) The herring gull' s world. New York, NY: Harper and Row , Publis hers.
Verbeek, N. A. (1977) Comparative feeding be havior of i mmatu re and adult herring gulls . Wils on Bull. 89: 415-421.
Vermeer , K. (1973) Food ha bit s and br eeding r ange of herring gulls in t he Canadian pr airie provinces. Condor 75: 478-480.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Weseloh, D. H. (1989) Herring gull. In: Cadman, M. D.; Eagles, P. F.; Helle iner, F. M., eds. Atlas of th e breeding bir ds of On tar io. Water loo, Univ ersi ty of W ater loo P ress; pp. 182-183.
Weseloh, D. V.; Mineau, P.; Struger, J. (1990) Geographic al dis tr ibut ion of con tamina nts and producti vi ty measu res of herring gulls in t he Great Lakes: Lake Er ie and conne cting c hannels 1978/79. Sci . Tot. Environ. 91: 141-159.
2-172 Herring G ull

2.1.13. Belted Kingfisher (kingfishers)
Order Coraciif ormes, Family Alc edinidae. Kingf ishers are sto cky, short-legged bir ds wi th large heads and bills . They exist on a die t mostl y of fi sh, whic h they catch by div ing, from a pe rch or the air, head fi rst int o the water . They nest i n burrows i n earthen banks that they dig us ing t heir bills and feet.
Selected species
The belted kingf isher (Cery le alcyo n, former ly Megaceryle alcyo n) is a medium -sized bird (33 cm bill t ip t o tail t ip) th at eats p rimar ily fi sh. It is one of th e few sp ecies of fish-eating bir ds found t hroughout inla nd areas as wel l as co astal areas. The belted kingf isher's range inc lude s most of th e Nor th Amer ican cont inent ; it breeds from nor thern Alaska an d central Labrador southward to the southern border of th e United States (B ent, 1940). Two subspecies so meti mes ar e recognize d: the easter n belted kingf isher (Cery le alcyo n alcyo n), which occu pies th e range east o f th e Rocky Mount ains and nor th to Quebec, and the wester n belted kingf isher (Cercyl e alcyo n caurina), which occu pies th e remaining r ange to the west (B ent, 1940).
Body size. The sexes ar e simi lar in size and appearance, although t he femal e tends to be slight ly larger (Salyer and Lagler, 1946). Bent (1940) reported that wester n popula tions ar e somewh at larger than easter n ones. Nest lings r each adult body weight by about 16 days after hatching, but th en may l ose so me weight before fledging ( Hamas, 1981).
Habitat . Belted kingf ishers are typic ally found a long r ivers and s treams an d along lake and pond e dges (Hamas, 1974). They are also commo n on seaco asts an d estu aries (Bent, 1940). They prefer water s th at are free of th ick veg etat ion t hat obs cures the view of the water and water that is not co mpletely overshadowed by tr ees (Bent, 1940; White, 1953). Kingf ishers also requir e relatively clear water in order to see th eir prey and are noticeably absent in areas wh en water s become tu rbid ( Bent, 1940; Davis, 1982; Salyer and Lagler, 1946). White (1953) suggested that water less th an 60 cm d eep is prefer red. They prefer str eam r iffl es fo r foraging s ites even when pools ar e more plent iful b ecause of th e conc entration of fi sh at riffl e edges (Davis, 1982). Belted kingf ishers nest i n burrows wi thin steep earthen banks d evoid of veg etation be side r ivers, str eams, ponds , and l akes; they also have been found t o nest i n slopes cr eated by human excavat ions su ch as roadcuts and la ndf ills (H amas, 1974). Sandy soil ba nks, whic h are easy to excavate an d provide good dr ainage, are prefer red (Brooks an d Davis, 1987; Cornwel l, 1963; White, 1953). In general, kingf ishers nest n ear suitable fishing a reas wh en possible but will n est away fr om water and feed in bodie s of water other than the one closest to home (Cornwel l, 1963).
Food ha bi ts . Belted kingf ishers generally feed on fish that swi m near the surface o r in s hallow water (Salyer and Lagler, 1946; White, 1953; Cornwel l, 1963). Davis (pers. comm. in Prose, 1985) believes th at th ese k ingf ishers generally catch fish only in t he uppe r 12 to 15 cm o f th e water colum n. Belted kingf ishers capture fish by div ing e ither fr om a perch ov erhanging t he water or after hover ing a bov e the water (Bent, 1940). Fish
2-173 Belted Kingf isher

are swallow ed whole , head fi rst, after being b eaten on a perch (Bent, 1940). The average lengt h of fi sh caught in a Michiga n s tudy was l ess th an 7.6 cm b ut ranged from 2.5 to 17.8 cm (Sal yer and Lagler, 1946); Davis (1982) found f ish caught in Ohio s treams to range from 4 to 14 cm i n length. Several studie s indi cate th at belted kingf ishers us ually catch the prey that are most avai lable (White, 1937, 1953; Salyer and Lagler, 1946; Davis, 1982). Diet therefore var ies cons iderably among di ffer ent water bodie s and w ith seaso n (see examp les in Appendix ). Although k ingf ishers feed predominantly on fish, they also someti mes consume large numbers of cr ayfi sh (Davis, 1982; Sayler and Lagler, 1946), and in shor tages of th eir prefer red foods , have been known to consume cr abs, mussel s, lizards, frogs, toads, smal l snakes, turtles, insects, salamanders, newts, young bir ds, mice, and berries (Bent, 1940). Parents b ring s urprisingly large fish to their young. Whit e (1953) found t hat nestlings only 7 to 10 days old w ere prov ided f ish up t o 10 cm long, and nestlings only 2 weeks old w ere prov ided with fish up t o 13 cm in le ngt h. After fledging, young be lted kingf ishers fed on f lying in sects fo r their fi rst 4 d ays after leaving t he nest, crayfi sh for the next week, and by th e 18th day post-fl edging, could catch fish (Salyer and Lagler, 1946).
Mol t. The juv enile plum age is maint ained through t he wint er, and y oung bir ds unde rgo t heir fi rst pr enupt ial molt in t he spring ( between February and April) involving most of th e body plum age (Bent, 1940). Adults h ave a co mplete postnupt ial molt in t he fal l (August to Octo ber) (Bent, 1940).
Migration. This kingf isher breeds ov er mo st of th e area of Nor th Amer ica and wint ers in m ost regions of th e cont inental Unit ed States (N ationa l Geogr aphic So ciety, 1987). Although m ost nor thern k ingf ishers migr ate to southern regions dur ing t he colde st months, some may stay i n areas th at remain ice-fr ee where fishing is pos sible (B ent, 1940).
Breeding acti vi ties and social organization. Dur ing t he breeding seaso n, pairs establis h territories for nesting a nd f ishing ( Davis, 1982); otherwise, belted kingf ishers are solit ary. They are not colonia l nester s and will d efend a n unus ed bank if it lies wi thin t heir ter ritory (Davis, 1982). In migrating popula tions , the males arrive before the femal es to find suit able nesting t erritories (Davis, 1982). Kingf ishers excavate th eir burrows i n earthen banks, fo rming a tunne l that averages 1 to 2 m in length, although s ome bur rows may b e as long a s 3 to 4 m (Hamas, 1981; Prose, 1985). The burrow entrance is usually 30 to 90 cm fr om th e top of th e bank (Bent, 1940; White, 1953) and at least 1.5 m fr om th e base (Cornwel l, 1963). Burrows cl oser to the top may c olla pse, and bur rows too low may f lood (Brooks an d Davis, 1987). Burrows may b e used for more than one seaso n (Bent, 1940). Five to seven eggs are laid on bare substr ate or on fish bone s wi thin t he bur row (Hamas, 1981; White, 1953). Only one adult , usually th e femal e, spends th e night in t he nest cavi ty; males usually roost i n nearby fo rested areas or heavy co ver (Cornwel l, 1963). Both parents i ncubate eggs and feed the young ( Bent, 1940). After fledging, the young r emain wi th their parents fo r 10 to 15 days (Sayl er and Lagler, 1946).
Home range and r esour ces. Dur ing t he breeding seaso n, belted kingf ishers requir e suit able nesting s ites wi th adequate nearby fi shing. Dur ing s pring a nd early su mmer , both male and femal e belted kingf ishers defend a ter ritory th at includes both their nest si te and their fo raging a rea (Davis, 1982). By autumn, each bird (including t he young of th e year )
2-174 Belted Kingf isher

�
�
defends an indiv idua l feeding t erritory only (D avis, 1982). The breeding t erritories (lengt h of water line protected ) can be more than twi ce as long a s the fall a nd w int er feeding ter ritories, and str eam ter ritories ten d to be longe r th an thos e on l akes (Davis, 1982; Salyer and Lagler, 1946). Foraging t erritory size is inv ersely related to prey abunda nce (Davis, 1982).
Popul ation de nsity . Breeding de nsit ies of b etween two and six pairs per 10 km o f river shor eline h ave been recorded, wi th densi ty i ncreasing w ith food availa bilit y (Brook s and Davis, 1987; White, 1936).
Popul ation dy namics. Kingf ishers are sensi tive to dis turbance and us ually do not nest i n areas near human acti vi ty (White, 1953; Cornwel l, 1963). Kingf ishers typic ally breed in the fi rst seaso n after they are born (Bent, 1940). Fledging s uccess d epends on food availa bilit y, sto rms, floods , predation, and t he int egrity of th e nest b urrow but can be as high a s 97 percent (M. J. Hamas, pers. comm.). Dispersal of young oc curs wi thin a mont h of fl edging ( Whit e, 1953). No data co ncerning a nnua l surviv orship r ates wer e found.
Simi lar species (fr om general references)
� The green kingf isher (Chloroceryle amer icana) is smal ler (22 cm) th an the belted kingf isher and is only co mmon in t he low er Rio Grande Valle y. It also is found in s out heaster n Arizona and a long t he Texas co ast, usually during fall a nd w int er.
� The ringe d k ingf isher (Cery le torquata) is larger (41 cm) an d resides in the low er Rio Grande Valle y in T exas an d Mexico.
General references
Bent (1940); Fry (1980); Nationa l Geogr aphic So ciety (1987); Prose (1985); White (1953).
2-175 Belted Kingf isher

Belted Kingfisher ( Cery le alc yon)
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight A B 148 ± 20.8 SD 125 - 215 Pennsy lvani a Powdermill N ature Center 1 (g) (unpubl .)
A B 136 ± 15.6 SE Pennsy lvani a Brooks & Davi s, 1987
A B 158 ± 11.5 SE Ohio Brooks & Davi s, 1987
at hatching 10 - 12 Minnesot a Hamas, 1981
at fledgi ng 148 ± 13.3 SE Pennsy lvani a Brooks & Davi s, 1987
at fledgi ng 169 ± 11.9 SE Ohio Brooks & Davi s, 1987
Nest ling 5 to 6 Pennsy lvani a, Ohio/streams Brooks & Davi s, 1987 2 Growth Rate (g/day)
Metabol ic Rate A B basal 132 est imated 3 (kcal /kg-day)
A B f ree-living 327 (154 - 693) est imated 4
Food Ingest ion A B 0.50 nor thcent ral lower Michi gan Alexander , 1977 5 Rate (g/g-day)
nest lings 1.0 - 1.75 Nova Scot ia White, 1936
Water A B 0.11 est imated 6 Ingest ion Rat e (g/g-day)
Inhal ation A B 0.094 est imated 7 Rate (m /day)3
Surface Ar ea A B 280 est imated 8 (cm )2
2-176 B
elted Kingfis
her

Belted Kingfisher ( Cery le alc yon)
Dietary Compositio n Spr ing Summer Fall Winter Locat ion/Habi tat (measur e) Reference No. Note
trout 17* lower Michi gan/lake Alexander , 1977 non- trout fish 29 crust acea 5 (% wet weight ; stomach insect s 19 cont ents) amphi bians 27 bi rds and m ammals 1 *data from spr ing and f all uni dent ified 2 also
trout 30 Michi gan/trout st reams Salyer & Lagl er, 1946 other game & pan f ish 13 (e.g., perch, (% wet volume; stomach cent rarchi ds) cont ents)
fo rage fis h (e.g., 15 minnow, st ickl eback, sculp ins)
uni dent ified f ish 1 crayfish 41 insect s < 1
salmon f ry 11 Nova Scot ia/riparian - White, 1936 salmon (1-yr-old) 42 streams salmon (2-yr-old) 1 (% of total num ber of prey; trout 15 fecal pel lets) st ickl ebacks 30 killifis h < 1 sucker s < 1
crayfish 13 sout hwest Ohio/creek Davis, 1982 cyprin ids 76 (minnows) (13) (% of total num ber of prey (stoner ollers) (38) brought to nest lings) (uni dent ified) (26)
other fish 10
2-177 B
elted Kingfis
her

Belted Kingfisher ( Cery le alc yon)
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Territo ry S ize early s ummer (km shor eline) breeding pai rs: 2.19 ± 0.56 SE Pennsy lvani a/streams Brooks & Davi s, 1987
late sum mer - 1.03 ± 0.22 SE sout hwest Ohio/streams Davis, 1980 nonbr eeding indi vidual s:
1.03 ± 0.28 SE Ohio/streams Brooks & Davi s, 1987
0.39 ± 0.093 SE sout hwest Ohio/streams Davis, 1980
Popul ation A B sum mer 0.11 - 0.19 Pennsy lvani a/streams Brooks & Davi s, 1987 Densi ty (pair/km shor e) A B sum mer 0.6 Nova Scot ia/streams White, 1936
Clutch Si ze 5.8 ± 0.7 SE Pennsy lvani a/streams Brooks & Davi s, 1987
6.8 ± 0.4 SE Ohio/streams Brooks & Davi s, 1987
Clutches/ Year 1 Pennsy lvani a, Ohio/streams Brooks & Davi s, 1987 9
1 Minnesot a/lake Hamas, 1975
Days 22 Minnesot a/lake Hamas, 1975 Incubat ion
Age at 28 days NS/NS Bent, 1940 Fledgi ng
Number 4.5 ± 1.9 SE Pennsy lvani a/streams Brooks & Davi s, 1987 Fledge per Act ive Nest 5.3 ± 2.2 SE Ohio/streams Brooks & Davi s, 1987
Age at Sexual 1 year throughout range Bent , 1940 Maturity
2-178 B
elted Kingfis
her

Belted Kingfisher ( Cery le alc yon)
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating Apr il Apr il to May early Jul y Minnesot a Hamas, 1975
Hatching May June late Jul y Minnesot a Hamas, 1975 early June Nova Scot ia White, 1936
Molt fall August October NS Bent, 1940 spr ing February Apr il NS Bent, 1940
Migration mid-October Maine Bent , 1940 fall depar tures mid-Novem ber NY, SD, WI, NE Bent , 1940
spr ing ar rivals late February PA, RI, MO Bent , 1940 mid-March NY, CT, IL, WI Bent , 1940 early Apr il Maine, Nova Scot ia Bent , 1940
mid-December Massachuset ts, New Jersey Bent , 1940
1 Cited in Dunni ng (1984).
2-179 B
elted Kingfis
her
2 Brooks and Davi s (1987) repor ted f ledgi ng wei ght s of 149 and 169 g f or two popul ations. Given a hat ching wei ght of about 10 g and 28 day s requi red to fledge, on aver age, chi cks m ust gain 5 to 6 g per day. Hamas (1981) found gai ns of appr oxi mately 8.5 g per day unt il day 18, and a loss of appr oxi mately 4.5 g per day unt il fledgi ng.
3 Est imated usi ng equat ion 3-28 (Lasiewski and Dawson, 1967) and body weight s f rom Powder mill N ature Center (unpubl .).4 Est imated usi ng equat ion 3-37 (Nagy, 1987) and body weight s f rom Powder mill N ature Center (unpubl .).5 Est imated by author .6 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Powder mill N ature Center (unpubl .).7 Est imated usi ng equat ion 3-19 (Lasiewski and Cal der, 1971) and body weight s f rom Powder mill N ature Center (unpubl .).8 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Powder mill N ature
Center (unpubl .). 9 They are know n to renest up t o three times i f clutches ar e lost early (Bent, 1940).

References (including A ppendix)
Alexander, G. R. (1974) The consumption of tr out by bir d and mammal predators on the Nor th Branch Au Sable River. Michiga n Dept . Nat. Resour ces, Dinge ll - Johns on Proj.; F-30-R, Fina l Repor t.
Alexander, G. R. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Bent, A. C. (1940) Life histo ries of North Amer ican cucko os, goat su cker s, hummingbir ds, and t heir allie s. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. US Nat. Mus., Bull. 176.
Brooks, R. P.; Davis, W. J. (1987) Habitat sel ect ion by breeding be lted kingf ishers (Cery le alcyo n). Am. Midl. Nat. 117: 63-70.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Cornwel l, G. W. (1963) Observat ions on t he breeding biology and be havior of a nesti ng popula tion of belted kingf ishers. Condor 65: 426-431.
Davis, W. J. (1980) The belted kingf isher, Megaceryle alcyo n: its ecology and t erritorialit y [master 's th esis]. Cinc inna ti , OH: Univ ersi ty of Cinc inna ti .
Davis, W. J. (1982) Territory si ze in Megaceryle alcyo n along a str eam habitat. Auk 99: 353-362.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Fry, C. (1980) The evolut iona ry biology of kingf ishers (Alc edinidae). In: The liv ing bir d, 1979-80. Ithaca, NY: The Laboratory of Ornithology , Cornell Univ ersi ty; pp. 113-160.
Hamas, M. J. (1974) Human incursion a nd n esting s ites o f th e belted kingf isher. Auk 91: 835-836.
Hamas, M. J. (1975) Ecologic al and phy siologic al adaptations fo r breeding in t he belted kingf isher (Megaceryle alcyo n) [Ph.D. disser tat ion] . Dulut h, MN: Univ ersi ty of Minne sota.
Hamas, M. J. (1981) Thermoregula tory developm ent in t he belted kingf isher. Comp. Bioc hem. Physiol. A: Comp. Physiol. 69: 149-152.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
2-180 Belted Kingf isher

Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Poole , E. L. (1938) Weight s and w ing a reas in North Amer ican birds. Auk 55: 511-517.
Prose, B. L. (1985) Habitat su itabilit y inde x mode ls : belted kingf isher. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.87).
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Salyer , J. C.; Lagler, K. F. (1946) The easter n belted kingf isher, Megaceryle alcyo n alcyo n (Linna eus), in r elation t o fish management. Trans. Am. Fish. Soc. 76: 97-117.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
White, H. C. (1936) The food of kingf ishers and m ergansers on t he Margaree River, Nova Scotia. J. Biol. Board Can. 2: 299-309.
White, H. C. (1937) Local feeding of kingf ishers and m ergansers. J. Biol. Board Can. 3: 323-338.
White, H. C. (1938) The feeding of kingf ishers: food of n estlings and effect o f water height . J. Biol. Board 4: 48-52.
White, H. C. (1953) The easter n belted kingf isher in t he mar it ime prov inces. Fish. Res. Board Can. Bull. 97.
2-181 Belted Kingf isher

2.1.14. Marsh Wren (w rens)
Order Passer iformes, Family Trogl ody tidae. Wrens are smal l insecti vorous birds that l ive in a var iety o f habitats th roughout th e Unit ed States. They have long, slender bills adapted for gleaning in sects fr om th e ground a nd v egetation. Most species are migr atory, although s ome popula tions ar e year -round r eside nts.
Selected species
The marsh wren (Cis tothorus palustris ) is a common bir d inha bit ing f reshwater cattai l marshes and sal t mar shes. Marsh wrens breed throughout mo st of th e nor thern half of th e United States an d in coastal areas as far south as Flor ida; they wint er in t he southern United States an d into Mexico, particularly in coastal areas. Marsh wrens eat mostl y insects, and occas iona lly sn ails , whic h they gle an from th e surface o f veg etat ion. This sp ecies was fo rmer ly kn own as th e long- bille d marsh wren (Telmato dytes p alustr is).
Body size. Although w rens are smal l (13 cm bill t ip t o tail t ip; about 10 g body weight ), males tend t o be about 10 pe rcent heavier than femal es (see tab le). Body weight var ies seaso nally; in Georgia, where marsh wrens are resident th roughout th e year , they tend to be heavier in the spring a nd s ummer than in the fal l and winter (Kale, 1965).
Habitat . Marsh w rens inha bit fr eshwater and sal twater marshes, usually nest ing in asso ciation w ith bulr ushes, cattai ls, and sedges or on occas ion in m angr oves (Welter , 1935; Bent, 1948; Kale, 1965; Verner, 1965). Standing water from sever al centimeter s to nearly a meter is typ ical of th e areas sel ected (Bent, 1948). Permanent water is necessar y to prov ide a food s upply of insects n ecessar y to maintain the birds and as a defense agains t pr edation (Verner and Engelsen, 1970). Deeper water and denser vegetat ion a re asso ciated wi th reduced predation r ates (L eonard and Picman , 1987).
Food ha bi ts . Marsh wrens co nsume aquatic inver tebrates, other insects, and spide rs, whic h they gle an from th e water surface, on stems an d leaves o f emer gent vegetat ion, and t he marsh floor (Kale, 1965; Welter , 1935). They so meti mes al so feed by flycatc hing ( Welter , 1935). The insect o rders mo st co mmonly taken include Coleopt era (both adults an d larvae), Diptera (adults an d larvae), Hemiptera (juveniles and adults), Lepidopt era (larvae mo st co mmonly fed to nestlings ); and Odon ata (newly emer ged) (Bent, 1948; Kale, 1964). When feeding t he young, at fi rst the parents br ing m osquit o adult s and larvae, midge s, larval tipulids , and ot her smal l insects (W elter , 1935). As th e young matu re, the parents b ring la rger insects su ch as ground beetl es, diving b eetles, long-horned beetles, cater pilla rs, dragonf lies, and sawfl ies to the nest lings (W elter , 1935). In a popula tion in G eorgia , spide rs (usually 1 to 3 mm in s ize, someti mes 12 to 15 mm), smal l crabs (5 to 7 mm), smal l snails (1 to 3 mm), and insect eg gs also were consumed and fed to nestlings (K ale, 1965). Thus, organisms th at are aquatic fo r all or part of th eir lives ar e an impor tant compone nt of th e die t of mar sh wren adult s and n estlings .
Migration. Marsh wrens are year -round r eside nts in s ome sout hern and c oastal mar it ime regions wh ere marshes do not fr eeze. Most migr atory wrens br eed throughout the northern half of th e United States th rough s out hern Canada and w int er in Mexico and
2-183 Marsh Wren

the southern half of th e United States (B ent, 1948; Verner, 1965; Amer ican Ornthologi sts' Union, 1983; Nationa l Geogr aphic So ciety, 1987).
Breeding acti vi ties and social organization. Many popula tions of mar sh wren are poly gynous , wi th some males mating w ith two , occas iona lly th ree, femal es in a seaso n, while th e remaining m ales have one mate o r remain bachelors. For examp le, Leonard and Picman (1987) found 5 to 11 percent bachelor males, 41 to 48 percent monoga mous mal es, 37 to 43 percent bigamous mal es, and 5 to 12 percent tr igamous mal es in two marshes in Manitoba, Canada. Simi larly, Verner and Engelsen (1970) found 1 6 percent bachelor s, 57 percent monoga mous , and 25 percent biga mous mal es in easter n Washingt on state. In contrast, Kale (1965) found m ost males to be monoga mous th rough 4 years of stu dy in Georgia.
Males arrive at the breeding m arshes before the femal es to establis h territories that inc lude bot h nest si tes an d foraging a reas (Kale, 1965; Verner, 1965; Welter , 1935). Males build sever al nests i n their ter ritories th roughout th e breeding seaso n (Kale, 1965; Verner, 1965). The femal e usually only adds lining mater ial to a nest o f her choic e, although s ome may h elp constr uct th e breeding n est (K ale, 1965). Breeding n ests ar e oblong in s hape, wi th a side opening, and are woven of cattai ls, reeds, and grasses an d lashed to standing vegetat ion, generally 30 cm to 1 m abov e standing water or high t ide (B ent, 1948; Verner, 1965). Incubation l asts ap proximatel y 2 weeks, as does th e nest ling pe riod ( Kale, 1965; Verner, 1965). After fledging, one or bot h parents cont inue to feed the young f or about 12 days (Ver ner, 1965). Many popula tions typic ally r ear two broods per year , although s ome may r ear three (Kale, 1965; Verner, 1965). In the more monoga mous popula tions , bot h parents regula rly feed young, but in t he more poly gynous ones, th e femal es may p rovide most of th e food, wi th males assi st ing only to ward the end of th e nestling pe riod ( Leonard and Picman , 1988; Verner, 1965).
Home range and r esour ces. Marshes smal ler than 0.40 ha usually are not used by breeding m arsh wrens (Bent, 1948). Average male ter ritory si ze for a given year and loc ation c an range fr om 0.006 to 0.17 ha, depending on t he habitat an d condit ions of th e year (see tab le). Also, there is a tr end in poly gynous popula tions fo r poly gynous mal es to defend la rger ter ritories than monoga mous mal es or males that end up a s bachelor s (Verner and Engelson, 1970; Verner, 1964; Kale, 1965).
Popul ation de nsity . Because th e species is poly gynous , there may b e more femal es than males inha bit ing br eeding m arshes. Popula tion de nsity var ies wi th the suit abilit y and patch iness o f th e habitat. Densi ties as high a s 120 adult bir ds per hectar e have been recorded (Kale, 1965).
Popul ation dy namics. Clutch size and number of cl utch es per year vary wi th lati tude and cl imate (see tab le). In some popula tions , marsh wrens commonly d estr oy eggs and k ill t he nestlings of other pairs of th eir own species and ot her mar sh-nesti ng passer ines (Or ians and Wilson, 1964; Picman , 1977; Welter , 1935). Fledging s uccess depends str ongly on n est l ocat ion; nests o ver deeper water are less v ulne rable to predation ( Leonard and Picman , 1987). Of nests l ost to all causes, Leonard and Picman (1987) found 4 4 percent due to mammal ian predators, 27 percent due to other wrens, 11 percent due to weath er, 8 percent due to nest ab andonm ent, and 1 3 percent unk now n. The
2-184 Marsh Wren

�
�
�
�
�
annua l mortalit y of adult s is low er th an that of fi rst-year birds. Both sexes o f th is sp ecies usually co mmen ce breeding in t he fi rst year follow ing h atching ( Kale, 1965).
Simi lar species
� The sedge wren (Cisto thorus platensis, former ly know n as the shor t-bille d marsh wren) nests l ocal ly in wet mead ows o r shallow sedge mar shes and hayfi elds in the northeaster n United States, winter ing pr imarily in t he southeaster n United States. It is s light ly smal ler (11 cm) th an the marsh wren.
Note: None of th e other wr en species inha bit mar shes, al though a ll f orage by gleaning in sects fr om veg etat ion a nd ot her su rfaces. Wrens th at inha bit mois t woodla nds and ope n areas are listed below.
� The house wr en (Troglodytes aed on) (12 cm) b reeds th roughout mo st of th e United States, into southern Canada. It inha bit s ope n habitats wi th brush and s hrubs and is found in or chards, farmyar ds, and urban gardens and parks.
� The winter wren (Troglodytes tr oglodytes ) (10 cm) b reeds in southern Canada, where it nests i n dense brush, especially along m ois t conif erous woodla nds . It wint ers pr imarily in t he sout heaster n United States, where it inha bit s many typ es of woodla nds .
� The Carolina wr en (Thryothor us ludov icianus ) (14 cm) i s nonm igr atory and can be found in bot h summer and winter in the easter n United States as far north as northern Delaware and as far west as Okl ahoma. It inha bit s mois t woodla nds and swamp s and woode d subur ban areas.
� Bewick' s wr en (Thryomanes bewicki i) (13 cm) i s mo re commo n in wester n States th an the hous e wren and is declining east o f th e Missi ss ippi. It is found in br ushland, str eam edges, and open woods .
General references
Kale (1965); Gutzwille r and A nderson (1987); Leonard and Picman (1987); Verner (1965), Nationa l Geogr aphic So ciety (1987).
2-185 Marsh Wren

Marsh Wren ( Cistothorus palustris )
2-186 M
arsh W
ren
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight F breeding 10.6 ± 0.99 SD 9.0 - 13.5 New Yor k Tint le (unpubl .) 1 (g) M breeding 11.9 ± 0.72 SD 10.5 - 13.5
A F 9.4 ± 1.1 SD Georgia Kale, 1965 2 A M 10.6 ± 0.7 SD J B 9.4 ± 1.6 SD
nest ling: New York, Minnesot a/fresh Welter, 1935 3 day 1 1.1 marshes day 3 2.1 day 5 4.7 day 7 6.8 day 9 10.0 day 11 10.6 day 13 11.3
at fledgi ng 8.84 ± 0.70 SD Georgia Kale, 1965
Egg Weight (g) 1.14 ± 0.10 SD Georgia Kale, 1965
Metabol ic Rate A B basal 91.2 Georgia (capt ive) Kale, 1965 4 (lO /kg-day) A B near 113 52
basal 6 A B l ight 169 activ ity
Metabol ic Rate A B basal 444 Georgia (capt ive) Kale, 1965 7 (kcal /kg-day) A B near 557 ± 115 SD Kale, 1965 8
basal A B l ight 788 ± 115 SD Kale, 1965 9
activity 880 ± 90 SD Kale, 1965 10 A B free-liv ing
A F f ree-living 1,174 (554 - 2,486) A M free-liv ing
1,209 (571 - 2,563) est imated 11

Marsh Wren ( Cistothorus palustris )
2-187 M
arsh W
ren
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Food Ingest ion A B f ree-living 1,155 ± 130 SD Georgia (capt ive) Kale, 1965 12 Rate kcal/kg-day
A B f ree-living 0.67 g/g-day Georgia (capt ive) est imated f rom Kale, 1965 13
A F f ree-living 0.99 g/g-day est imated 14 A M free- 0.96 g/g-day liv ing
Water A F 0.28 est imated 15 Ingest ion Rat e A M 0.26 (g/g-day)
Surface Ar ea A F 45 est imated 16 (cm ) A M 482
Dietary Compositio n Spr ing Summer Fall Winter Locat ion/Habi tat (measur e) Reference No. Note
Hymenopt era 17.3 12.4 Georgia/sal t marsh Kale, 1965 17 Homopt era 13.0 40.1 Coleopt era 11.6 12.6 (% wet vol ume; Lepi dopt era 14.6 2.9 stomach cont ents) Diptera 8.9 7.7 Hemiptera 5.4 10.0 Orthopt era 5.6 0.8 spi ders 15.1 6.2 other arthropods (crabs, amphi pods) 1.8 0.9 molluscs ( snai ls) 3.5 4.0 other (insect eggs, undet ermined, etc.) 4.5 3.3

Marsh Wren ( Cistothorus palustris )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range ion/Habi tat Reference No.
Territory A M spr ing 0.0060 ± 0.0014 SD Georgia/sal t marsh 1, 1958 Kale, 1965 Size (ha) 0.0156 ± 0.0050 SD Georgia/sal t marsh 2, 1958
A M spr ing 0.17 ± 0.021 SE 0.0242 - 0.360 west Washi ngt on/fresh Verner, 1965
A M spr ing 0.07 ± 0.06 SD Mani toba/fresh cat tail marsh Leonar d & Picman, 1986
0.0085 ± 0.0042 SD Georgia/sal t marsh 2, 1959
mixed-speci es marsh
Popul ation spr ing: Densi ty pairs/ha 48.3 ± 5.3 SD 45.1 - 56.2 Georgia/sal t marsh (4 years) Kale, 1965
males/ha 8.5 west Washi ngt on/fresh Verner, 1965
males/ha 3.7 ± 0.5 SD 3.4 - 4.3 Mani toba/fresh m ixed- Leonar d & Picman, 1987
16.9 mixed-speci es marsh (2 areas)
speci es marsh (3 years)
Clutch Si ze 4.5 3 - 5 Georgia/sal t marsh Kale, 1965
6.0 ± 0.19 SD 4 - 8 east Washi ngt on/fresh Verner, 1965
5.8 ± 0.8 SD Mani toba/fresh cat tail marsh Leonar d & Picman, 1987
pond- margin marsh
Clutches/ Year 1 - 2 0 - 3 Georgia/sal t marsh Kale, 1965
2 0 - 2 east Washi ngt on/fresh Verner, 1965
2 - 3 0 - 3 west Washi ngt on/fresh Verner, 1965
pond- margin marsh
mixed-speci es marsh
Days 13.1 12 - 14 Georgia/sal t marsh Kale, 1965 Incubat ion
15.1 13 - 16 west Washi ngt on/fresh Verner, 1965 marsh
Locat
2-188 M
arsh W
ren

Marsh Wren ( Cistothorus palustris )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range ion/Habi tat Reference No.
Age at B 12 - 13 10 - 15 Georgia/sal t marsh Kale, 1965 Fledgi ng B 14 11 - 16 Washi ngt on/ fresh m arshes Verner, 1965
Number 3.4 ± 3.4 SD Mani toba/fresh m ixed m arsh Leonar d & Picman, 1987 Fledge per Act ive Nest
Number 4.5 ± 1.3 SD Mani toba/fresh m ixed- Leonar d & Picman, 1987 Fledge per speci es marsh Successf ul 5.1 ± 1.2 SD Mani toba/fresh cat tail marsh Leonar d & Picman, 1987 Nest
Age at Sexual B 1 year Mani toba/fresh m arsh Leonar d & Picman, 1987 Maturity B 1 year Washi ngt on/ fresh m arsh Verner, 1971
Annual A B 32 Georgia/sal t marsh Kale, 1965 Morta lity J B 70 Rates (percent)
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Laying Apr il mid-August Georgia Kale, 1965 mid-Apr il May - June early Jul y eastern Washi ngt on Verner, 1965 late March Apr il - May mid-July (Turnbul l) Verner, 1965 late May early August western Washi ngt on (Seattle) Welter, 1935
New Yor k
Hatching early May mid-July eastern Washi ngt on Verner, 1965 mid-Apr il early August (Turnbul l) Verner, 1965
western Washi ngt on (Seattle)
Migration fall September late October New York, Minnesot a Welter, 1935
spr ing Apr il May June New York, Minnesot a Welter, 1935 mid-March eastern Washi ngt on Verner, 1965 (nonm igratory) (Turnbul l) Verner, 1965
western Washi ngt on (Seattle)
Locat
2-189 M
arsh W
ren

Marsh Wren ( Cistothorus palustris )
1 As ci ted in Dunni ng (1984).2 Col lect ion dat es not speci fied. Resident popul ation; presum ably averaged f rom birds capt ured throughout the year.3 Est imated from Welter's (1935) growth cur ve based on 50 nest lings.4 Measur ed by oxy gen respi rometry; lowest value of metabol ism of post absor pt ive wrens r est ing i n the dark (but not at night ) at temperatures
within the thermoneut ral zone. 5 Measur ed by oxy gen respi rometry; bi rds not post absor pt ive, but rest ing i n a dark box at temperatures w ithin the thermoneut ral zone. 6 Measur ed by oxy gen respi rometry; bi rds som ewhat act ive in their cage. 7 Est imated f rom oxy gen consum pt ion, for condi tions, see not e 3. 8 Est imated f rom oxy gen consum pt ion, for condi tions, see not e 4. 9 Est imated f rom oxy gen consum pt ion, for condi tions, see not e 5.
10 Est imated from measur ed dai ly food i ntake, excr etory losses, assi milation, and r espi ration f or act ive bi rds in small cages ( 173 weekly determinations t otal). Because of the birds' high act ivity levels, Kale (1965) consi dered the measur e represent ative of free-l iving bi rds.
11 Est imated usi ng al lometric equat ion 3-36 (Nagy, 1987) and body weight s f rom Kale (1965). 12 Measur ed dai ly food i ntake of bi rds in cages and m easur ed cal or ic cont ent of diet provi ded. Because of the bi rds' high act ivity levels, Kale (1965)
consi dered the measur e represent ative of free-living bi rds. 13 Est imated from Kale's (1965) measur ed dai ly food i ntake (see not e 11) assum ing 5.62 kcal /gram (dry weight ) insect s, a 70 perc ent assi milation
efficiency , and a 67 per cent water cont ent for insect s. 14 Est imated from free-living m etabol ic rate est imated from Nagy's (1987) equat ion 3-36 (see not e 10) assum ing t he same parameters descr ibed in
not e 12. These pr edicted food i ngest ion r ates (>0.95 g/g-day) for free-living bi rds exceed t he value est imated for Kale's (1965) caged bi rds (0.67 g/g-day); however , the latter does not include m etabol ic requi rements of searchi ng f or food, reproduct ion, or unusual thermoregulatory demands.
15 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Kale (1965).16 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Kale (1965).17 Summer col umn represent s com bination of spr ing and sum mer data; winter col umn represent s com bination of fall and w inter data.
2-190 M
arsh W
ren

References (including A ppendix)
Amer ican Ornit hologi sts' Union. (1983) Check-l ist o f North Amer ican birds. Lawrence, KS: Allen Press, Inc.
Bent, A. C. (1948) Life histo ries of North Amer ican nuthatch es, wrens, thrashers, and their allie s. Washingt on, DC: U.S. Government Print ing Offi ce; Smithsonia n Ins t. U.S. Nat. Mus., Bull. 195.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Gutzwille r, K. J.; Anderson, S. H. (1987) Habitat su itabilit y inde x mode ls : marsh wren. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.139).
Kale, H. W., II (1964) Food of th e long- bille d marsh wren, Telmato dytes p alustr is griseus, in the sal t marshes of Sap elo Is land, Georgia . Oriole 29: 47-61.
Kale, H. W., II. (1965) Ecology and bioe nergetics of th e long- bille d marsh wren Telmato idytes p alustr is griseus (Brewster ) in Georgia sal t mar shes. Publ. Nuttal l Ornit h. Club No. 5.
Leonard, M. L.; Picman , J. (1986) Why are nest ing m arsh wrens and y ellow -headed blackb irds sp atially seg regated? Auk 103: 135-140.
Leonard, M. L.; Picman , J. (1987) Nest ing m ortalit y and ha bitat sel ect ion by mar sh wrens. Auk 104: 491-495.
Leonard, M. L.; Picman , J. (1988) Mate choice by mar sh wrens: the influence of mal e and ter ritory quality. Anim. Behav. 36: 517-528.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Orians, G. H.; Wilson, M. F. (1964) Interspeci fic ter ritories of birds. Ecology 45: 736-745.
Picman , J. (1977) Intraspeci fic nest d estr uct ion in t he long- bille d marsh wren, Telmato dytes p alustr is plustr is. Can. J. Zool. 55: 1997-2003.
2-191 Marsh Wren

Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Verner, J. (1964) Evolut ion of poly gamy i n the long- bille d marsh wren. Evolut ion 1 8: 252-261.
Verner, J. (1965) Breeding biology of th e long- bille d marsh wren. Condor 67: 6-30.
Verner, J. (1971) Survival and dispersal of mal e long- bille d marsh wrens. Bir d-Banding 4 2: 92-98.
Verner, J.; Engelsen, G. H. (1970) Territories, multiple n est building, and poly gamy i n the long- bille d marsh wren. Auk 87: 557-567.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Welter , W. A. (1935) The natural histo ry of th e long- bille d marsh wren. Wils on Bull. 47: 3-34.
2-192 Marsh Wren

2.1.15. American Robin (thrushes)
Order Passer iformes, Family Musci capidae, Subf amily Turdinae. Thrushes are commo n, medium-si zed birds th at eat wo rms, insects, and fruit. They l ive in a var iety o f habitats, including w oodla nds , swamp s, subur bs, and pa rks. Most thrushes build n ests o f mud and vegetat ion on t he ground or in t he crotches of tr ees or shrubs; blue bir ds n est i n holes in trees and posts o r in nest b oxes. This group f orages pr imarily on t he ground a nd in low veg etat ion by probing a nd gle aning. Some thrushes are neotropic al migr ants while others reside year -round in Nor th Amer ica. Thrushes range in s ize fr om th e easter n and wester n blue bir ds (18 cm fr om bill t ip t o tail t ip) to the Amer ican robin (25 cm). Male and femal e plum ages are simila r in m ost thrushes, al though in s ome species, su ch as the blue bir ds, the males are more br ight ly color ed.
Selected species
The Amer ican robin ( Turdus migratorius) occurs throughout mo st of th e cont inental United States an d Canada during t he breeding seaso n and winters in the southern half of the United States an d in Mexico and Central Amer ica. The breeding r ange of th e robin ha s expanded in recent ti mes wi th the increasing a rea covered by lawns and ot her open habitats (H owel l, 1942; Martin et al ., 1951; James an d Shuga rt, 1974).
Body size. The sexes ar e simi lar in size and appearance. Their size var ies slight ly geographical ly; the smal lest r obins ar e found in t he easter n United States an d along t he Paci fic co ast, and the largest o nes occu r in the Rocky Mount ains , nor thern Great Plains, and northern deser ts (A ldrich and James, 1991).d The size of robins tends to inc rease wi th lati tude in easter n North Amer ica but does not in wester n North Amer ica (Aldrich and James, 1991). Fledglings attai n adult size at appr ox imatel y 6 weeks o f age (Howel l, 1942).
Habitat . Access to fresh water , protected nest ing s ites, and produc tive foraging areas are important requir ements fo r breeding r obins (Sp eirs, 1953). Breeding ha bi tats inc lude mois t forests, swamp s, open woodla nds , orchards, parks, and la wns. Robins forage on t he ground in ope n areas, along ha bi tat ed ges, or the edges of str eams; they also forage abov e ground in s hrubs and w ithin t he low er branches of tr ees (Paszkowski , 1982; Malmborg and Wills on, 1988). Nests i n woode d areas are usually near some typ e of opening s uch as the forest ed ge or a treefal l gap (Young, 1955; Knupp e t. al ., 1977). During the nonbr eeding seaso n, robins prefer moist w oods or fruit -bearing t rees and shrubs (Robbins et al., 1983). In the fal l, flocks o f mi gratory robins ar e often found a long f orest edges or clearings wh ere fruit s are most ple nt iful (Baird, 1980).
Food ha bi ts . Robins fo rage by hopping a long t he ground in search of ground-dwelling inv ertebrates an d by sear ching f or fr uit and f olia ge-dwelling in sects i n shrubs and low tr ee branches (Malmbor g and Wills on, 1988; Paszkowski , 1982). In the months preceding a nd dur ing t he breeding seaso n, robins feed mainly (g reater than 90 percent volume) on inver tebrates an d on some fr uits; during t he remainde r of th e year , their diet
dBased on linear measurements of museum stu dy ski ns.
2-193 Amer ican Robin

consists p rimar ily (over 80 to 99 percent by volume) of fr uits (Martin et al ., 1951; Gochfeld and Burger, 1984; Wheelwr ight , 1986). Robins eat a wi de var iety o f both plant and animal foods ; in a co mpila tion of die t records coll ected throughout th e Unit ed States an d southern Canada, Wheelwr ight (1986) found t hat robins cons umed fruit s from 51 ge nera and inver tebrates fr om 107 fam ilie s. Commonly eaten fruits i nclude plum s, dogw ood, summac, hackb erries, blackb erries, cherries, greenbriers, raspberries, and junipe r (Martin e t al., 1951; Wheelwr ight , 1986); commo n inver tebrates i nclude b eetles, cater pilla rs, mo ths, grasshoppe rs, spide rs, millipe des, and e arthworms (Martin e t al., 1951; Wheelwr ight , 1986; Paszkowski , 1982). Wheelwr ight (1986) has co mpile d seaso nal changes in the propor tion of plants an d inver tebrates co nsumed by robins in t hree di ffer ent sect ions of th e Unit ed States (see tab le). Wheelwr ight (1986) also has su mmar ized the average occu rrence of fruit s of var ious plant fami lies in the sto mach s of robins by mont h for th ese sect ions . Martin et al . (1951) have su mmar ized the occu rrence of fr uits o f various plant fami lies in more speci fic areas of th e United States (see A ppendix ).
Wheelwr ight (1986) found no di ffer ences b etween the sexes i n the propor tion or typ es of inver tebrates an d fruits eaten . Very young r obins (up t o at least 35 d ays o f age) feed almost en ti rely on insects an d other inver tebrates (Howel l, 1940). Older juveniles ten d to eat a highe r propor tion of fr uit and easy-to -capture prey th an adults (Go chfeld and Burger, 1984; Wheelwr ight , 1986). In a given area, robins o ften show food pr eferences: a popula tion in c entral New York seemed to prefer northern arrowwood a nd s pic e bus h f ruit s over mo st ot her plants (Wheelwr ight , 1988); in Illinois , a group ate predominantly fr ost grapes and Virginia cr eeper in t he late su mmer and fal l (Malmborg and Wills on, 1988).
Dur ing seaso ns wh en fruits d ominate th e diet, robins may n eed to consume quanti ties in excess o f th eir body weight to meet th eir metabolic needs each day (see table ). Robins as well a s ot her fr ui t-eating bir ds exhibit a low dig esti ve effi ciency fo r fruits; Karasov and Levey (1990) esti mated the metaboliza ble en ergy co effi cient (MEC) (i.e., the propor tion of food e nergy th at actu ally is assi mi lated) for robins eating a mi xed fruit die t to be only 55 pe rcent, perhaps because of th e low retention t ime of th e dig ested matter in the gut (Levey an d Karasov, 1992). The short retention t ime might actu ally be an adaptat ion t o eating f ruit b ecause large quanti ties of fr uit mu st b e processed to obtain an adequate protein intake. In contrast, when eating in sects, robins (as wel l as other bird species) exhibit a highe r dig esti ve effi ciency o f approximatel y 70 percent (Levey an d Karasov, 1989). Moreover , the energy co ntent of insects ten ds to be highe r th an that of mo st fruit s, particularly on a wet-we ight basis (see Chapter 4). Thus, during t he spring w hen robins are cons uming in sects, they should c ons ume a smalle r amount r elative to their body weight th an when eating f ruit s (Chapter 4 pr ov ides appr oaches that can be used to esti mate i nsect inge stion r ates fo r robins ).
Mol t. Postjuv enile and pos tbreeding ( prebasic) molt s oc cur fr om l ate Ju ly to Octobe r (Wheelwr ight , 1986; Sharp, 1990). During t his molt , robins ar e cons uming la rgely fruits an d other plant mater ials, which contain limi ted proteins. This may co ntr ibute to larger fr uit cons umpt ion r ates at th is ti me. During t he prebreeding ( prealternate) mo lt, robins ar e feeding pr imarily on in sects an d other inver tebrates (l etter
2-194 Amer ican Robin

from N.T. Wheelright , Departmen t of Biology , Bow doin Colle ge, Bruns wick, ME, to Sue Norton, March 18, 1992).
Migration. Most robins n esting in t he nor thern Unit ed States an d Canada winter in the Gulf Coast States an d the Carolina s (Speirs, 1953; Dorst, 1962, as ci ted in Henny, 1972). Winter ing r obins ar e most abunda nt b etween 30 and 35 degrees N lati tude (Speirs, 1953). Robin f locks mi grate during t he day (Robbins et al., 1983); most n orthern robins leave th eir breeding gr ounds fr om Sep temb er to Novemb er and return between February and April (Howel l, 1942; Young, 1951; Fulle r, 1977).
Breeding acti vi ties and social organization. The onset o f th e breeding seaso n is later at highe r lati tude s (appr ox imatel y 3 days fo r each addit iona l degree in t he east) an d alti tude s, but mating a nd egg la ying ge nerally occur in April or May (James an d Shuga rt, 1974; Knupp e t al., 1977). Males ar rive on the breeding gr ounds before femal es to establis h territories; femal es pair wi th establis hed males, usually fo r th e dur ation of th e breeding seaso n (Young, 1951). The femal e primar ily builds th e nest o ut of mu d, dried grass, weedy stems, and other mater ials, constr uct ing it on hor izont al lim bs, tree-branch crotches, wi thin s hrubs , or on any one of a num ber of man -made str uctu res wi th horizont al surfaces (H owel l, 1942; Klimstr a and Stieglit z, 1957). First cl utch es usually co ntain three or four eggs; later clutch es ten d to contain fewer eggs (Young, 1955). The femal e does al l of th e inc ubating, whic h cont inue s for 10 to 14 days f ollow ing t he laying of th e second e gg (Klimstr a and Stieglit z, 1957; Young, 1955). Both males and femal es feed the nest lings (Young, 1955). Follow ing f ledging, the brood o ften divides, wi th the male and femal e each feeding ha lf of th e fledglings fo r anot her 2 weeks (W eatherhead and McRae, 1990). Females may star t another brood be fore the current one is inde pendent , leaving t he male to feed all of th e fledglings (Young, 1955). After reaching inde pendence, juv enile s often form foraging f locks i n areas of high f ood availa bilit y (Hir th et al., 1969).
Early in t he breeding seaso n, robins o ften roost co mmu nally. Males can continue to use th ese roosts th roughout th e breeding seaso n, whereas femal es sto p once th ey begin inc ubating e ggs (How ell, 1940; Pitts, 1984). As fal l approaches and their diet tu rns mo re toward f ruit s, robins in m any areas begin t o roos t communa lly ag ain and may join ot her species, such as co mmo n grackl es and European star lings , in la rge roosts (M orrison and Caccami se, 1990).
Home range and r esour ces. Dur ing t he breeding seaso n, male robins establis h breeding t erritories, whic h the femal e helps to defend against o ther robins . Nonetheless, the ter ritories of differ ent pairs often over lap where neither pair can establis h dom inance (Young, 1951). Most fo raging dur ing t he breeding seaso n is co nfined to the ter ritory, but adults so meti mes l eave to forage in more producti ve areas th at are shared wi th other indiv idua ls (How ell, 1942; Young, 1951; Pitts, 1984). In some prime nest ing a reas (e.g., dense co niferous fo rest), where robin de ns it ies are high, ter ritories are small a nd t he bir ds might o ften forage elsewh ere (Howel l, 1942). Adult r obins o ften return to the same ter ritory in succeeding years (Young, 1951). During t he nonbr eeding r oos ting pe riod, robins ar e likely to return to the same fo raging s ites fo r many weeks an d to join r oosts wi thin 1 to 3 km o f th ese fo raging a reas (Morrison and Caccami se, 1990).
2-195 Amer ican Robin

�
�
�
�
Popul ation de nsity . Nesting popula tion de nsity var ies wi th habitat q uality. Densely forested areas th at provide wel l-protected nest si tes h ave been found t o suppor t high densit ies of nesting r obins ; however , the relatively smal l ter ritories found in t hese areas might not be used as much for fo raging a s thos e cont aining ope n areas (Howel l, 1942). In the nonbr eeding seaso n, robins o ften join s ingle - or mixed-species roosts th at can include tens of th ousands of birds (Morrison and Caccami se, 1990). Winter ing r obins ar e most common in pine or oak pine co mmunit ies of th e sout heaster n and southcentral Unit ed States, and decrease in abunda nce in dr ier, less fo rested areas westwar d (Speirs, 1953).
Popul ation dy namics. Robins fi rst attemp t to breed the year after they hatch (Henny, 1972) and will r aise mult iple broods in a seaso n (Howel l, 1942). Predation is o ften a major sour ce of mo rtalit y for bot h eggs and ne stlings (Knupp e t al., 1977; Klimstr a and Stieglit z, 1957). Approximatel y half of th e adult bir ds su rviv e from year to year (Farner, 1949; Henny, 1972); the average longe vi ty of a robin t hat survives to its fi rst Jan uary is from 1.3 to 1.4 years (Farner, 1949).
Simi lar species (fr om general references)
� The wood t hrush (Hylocichla mustelin a), whic h is smalle r th an the robin ( 18 cm), co-occu rs wi th the robin in s ome woodla nd ha bitats b ut is only present in the easter n United States. This sp ecies nests p rimar ily in the inter iors of matu re forests an d has been decreasing in a bunda nce over th e past d ecade as fo rested habitats i n North Amer ica become increasingly fr agmented (Robbins et al., 1989; Terborgh, 1989). This sp ecies is also primar ily a summer resident, winter ing in Flor ida and t he neotropic s.
� The hermit th rush (Catharus guttatu s) is found in c onif erous and m ixed woodla nds at nor therly lati tude s or high e levat ions and w int ers pr imarily in the southern half of th e United States. This sp ecies is also signif icantly smal ler (15 cm) th an the robin.
� Swainson's th rush (Catharus ustu latus) is present in the wester n and northeaster n United States d uring t he summer months, winter ing in t he neotropic s. It is al so smalle r th an the robin (16 cm).
� The var ied thrush (Ixoreus naevius) occu rs in moist co niferous fo rests o f th e Paci fic Northwest. This bird is si mi lar in size (21 cm) to the robin.
General references
Howel l (1942); Young (1955); Nationa l Geogr aphic So ciety (1987); Robbins et al. (1983); Sharp (1990).
2-196 Amer ican Robin

American Robin ( Turdus mi gratorius )
2-197 A
merican R
obin
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Body Weight A B al l seas. 77.3 ± 0.36 SE 63.5 - 103 Pennsy lvani a Clench & Leber man, 1978 1 (g)
A M nonbr eed. 86.2 ± 6.1 SD New Yor k Wheelwr ight , 1986 A F nonbr eed. 83.6 ± 6.4 SD
A M breeding 77.4 New York Wheelwright , 1986 A F br eeding 80.6
nest lings: at hatching 5.5 4.1 - 6.7 New York /fo rest Howell, 1942 day 2 12.6 8.4 - 17.5 day 4 24.3 17.9 - 32.3 day 6 39.4 32.5 - 45.9 day 8 50.9 42.0 - 59.3 day 10 55.2 49.0 - 63.2 day 14 55.0 51.8 - 58.2
Egg Weight (g) 6.26 4.6 - 8.4 New York Howell, 1942
Metabol ic Rate A B basal 259 est imated 2 (kcal /kg-day)
- B existence 344 Kansas Hazelto n et al., 1984 3
A B f ree-living 713 (336 - 1,513) est imated 4 (est imate)
Food Ingest ion A B f ree-living 1,070 ± 220 SD 760 - 1,330 Kansas Hazelton et al., 1984 5 Rate (kcal/kgday)
Food Ingest ion B B f ree-living 0.89 ± 0.73 SD California Skor upa & Hot hem, 1985 6 Rate (g/g-day)
- B free-liv ing 1.52 ± 0.25 SD 1.22 - 1.96 Kansas Hazelto n et al., 1984 7
Water A B 0.14 est imated 8 Ingest ion Rate (g/g-day)

American Robin ( Turdus mi gratorius )
Factors Age/Sex/ Range or Locat ion or Note Cond. /Seas. Mean (95% CI of mean) subspeci es Reference No.
Surface Ar ea A B 198 Walsberg & Ki ng, 1978 9 (cm )2
A B 182 est imated 10
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
nestlin gs/fledglin gs: south centra l New Howell, 1942 earthworms 15.0 York/forest sowbugs 1.7 spi ders 2.3 (% wet weight ; stomach millip edes 3.1 contents ) shor t-hor ned gr ass-
hopper s 4.9 (age of robi ns r anged f rom 3 beetles 11.6 to 35 day s after hatching; lepidopt eran larvae 24.7 presence of grass i s likely to ants 3.2 be acci dent al - carried along uni dent ified ani mal 5.2 with pr ey) grass (all parts) 19.5 mulberries 3.2 honey suckl e seeds 2.4 uni dent ified pl ants 4.2
adul ts: eastern Uni ted States Wheelwright , 1986 11 frui t 7 68 92 83 (% vol ume; invertebrates 93 32 8 17 stomach cont ents)
adul ts: cent ral Uni ted States Wheelwright , 1986 11 frui t 8 41 76 73 (% vol ume; invertebrates 92 59 24 27 stomach cont ents)
adul ts: western Uni ted States Wheelwright , 1986 11 fru it 17 29 63 70 (% volume; invertebrates 83 71 37 30 stomach cont ents)
2-198 A
merican R
obin
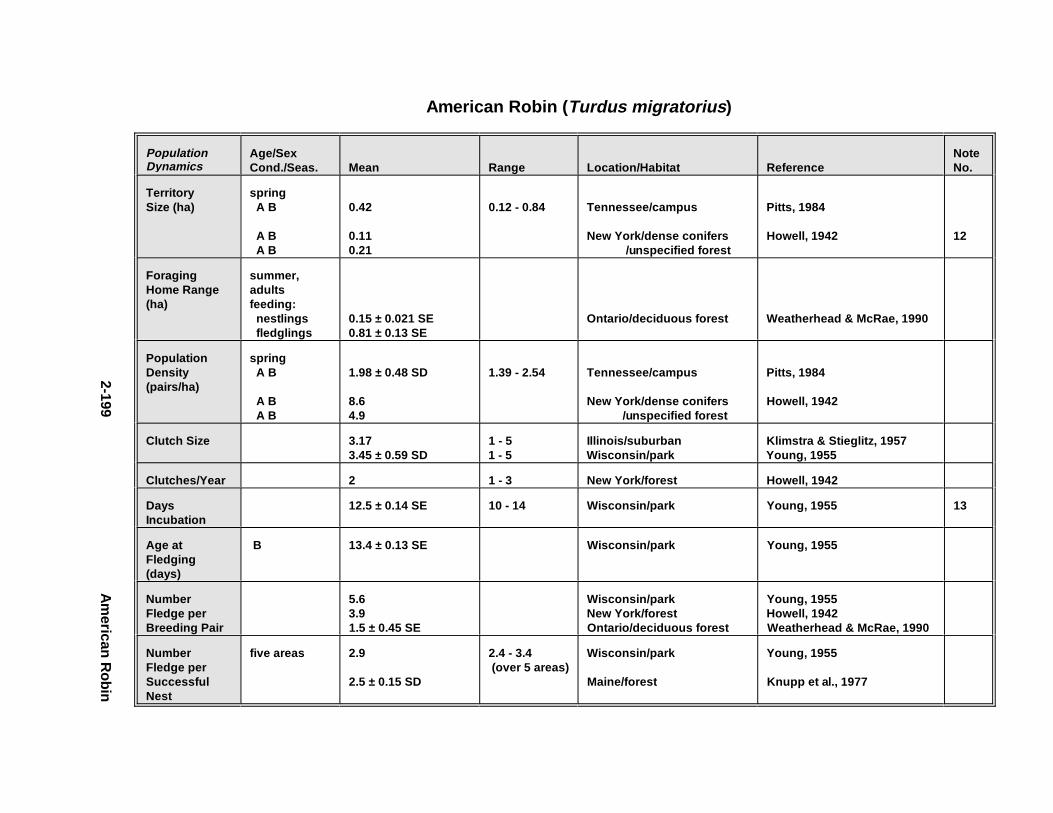
American Robin ( Turdus mi gratorius )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Territo ry sprin g Size (ha) A B 0.42 0.12 - 0.84 Tennessee/ campus Pitts, 1984
A B 0.11 New York /dense conifers Howell, 1942 12 A B 0.21 /unspecified fo rest
Foraging summer, Home Range adul ts (ha) feeding:
nest lings 0.15 ± 0.021 SE Ontario/deci duous f orest Weatherhead & McRae, 1990 fledgl ings 0.81 ± 0.13 SE
Popul ation spr ing Densi ty A B 1.98 ± 0.48 SD 1.39 - 2.54 Tennessee/ campus Pitts, 1984 (pairs/ha)
A B 8.6 New York /dense conifers Howell, 1942 A B 4.9 /unspecified fo rest
Clutch Si ze 3.17 1 - 5 Illinoi s/subur ban Klimstra & St iegl itz, 1957 3.45 ± 0.59 SD 1 - 5 Wisconsi n/park Young, 1955
Clutches/Year 2 1 - 3 New York /fo rest Howell, 1942
Days 12.5 ± 0.14 SE 10 - 14 Wisconsi n/park Young, 1955 13 Incubat ion
Age at B 13.4 ± 0.13 SE Wisconsi n/park Young, 1955 Fledgi ng (days)
Number 5.6 Wisconsi n/park Young, 1955 Fledge per 3.9 New York /fo rest Howell, 1942 Breeding Pai r 1.5 ± 0.45 SE Ontario/deci duous f orest Weatherhead & McRae, 1990
Number five areas 2.9 2.4 - 3.4 Wisconsi n/park Young, 1955 Fledge per (over 5 areas) Successf ul 2.5 ± 0.15 SD Maine/forest Knupp et al., 1977 Nest
2-199 A
merican R
obin

American Robin ( Turdus mi gratorius )
Population Dynamics
Age/Sex Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Age at B 1 year NS Henny , 1972 Sexual Maturity
Annual A B 51 ± 0.5 SE North Am erica Henny , 1972 Mortality Rates J B 78 - 82 (percent )
Longevi ty after Jan. 1 of 1.3 - 1.4 up t o 9 North America Farner, 1949 (years) first year
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Laying early Apr il mid-Apr il late Apr il Illinoi s Klimstra & St iegl itz, 1957 late April mid -July south centra l New York Howell, 1942 early May late May early Jul y n Maine Knupp et al., 1977
Hatching early May west : California, New James & Shugar t, 1974
early May east : VA, WV, DC, NY James & Shugar t, 1974 mid-May nor theast : VT, NH, CT James & Shugar t, 1974 mid-Apr il Kentucky James & Shugar t, 1974 early June Colorado James & Shugar t, 1974
Mexico
Molt fall Jul y & August North America Wheelwr ight , 1986
Migration fall mid-Sept. mid-October early Novem ber migrating t hrough Ful ler, 1977
sprin g February March Howell, 1942 mid-March mid-Apr il arriving New Yor k Young, 1951
early November Minnesota Howell, 1942 leaving New York
arriving Wisconsi n
1 As ci ted in Dunni ng (1984).
2-200 A
merican R
obin
2 Est imated usi ng equat ion 3-27 (Lasiewski and Dawson, 1967) and body weight s f rom Clench and Leber man (1978).3 Hazelton et al. (1984) est imate usi ng Kendei gh' s (1969) equat ions f or a 55-g bi rd.4 Est imated usi ng equat ion 3-36 (Nagy, 1987) and body weight s f rom Clench and Leber man (1978).

American Robin ( Turdus mi gratorius )
5 Est imated kcal consum ed in feeding t rials. Diet consi sted of paired of ferings of frui t (to test preferences) over a 2-day per iod, 12 tria ls per pairin g. Frui t included st rawberries (2.29 kcal /g), cher ries (4.34 kcal /g), green gr apes (2.59 kcal /g), and pur ple grapes (5.85 kcal /g). Mean weight of the bi rds = 55 g.
6 Based on gi zzard cont ents of robi ns caught foraging i n vineyards; diet 85 percent (wet weight ) grapes, 11.5 percent invertebrates, and 4.5 percent other plants. Mean wei ght of the bi rds = 82.3 g.
7 Based on sam e study descr ibed in not e 5 and est imated weight s of frui ts consum ed. 8 Est imated usi ng equat ion 3-15 (Calder and Br aun, 1983) and body weight s f rom Clench and Leber man (1978). 9 Beak sur face area 3.1 cm 2; leg sur face area 14.0 cm 2.
10 Est imated usi ng equat ion 3-21 (Meeh, 1879 and Rubner , 1883, as ci ted in Walsberg and Ki ng, 1978) and body weight s f rom Clench and Leber man (1978).
11 The U.S. Biologi cal Survey and U.S. Fish and W ildlife Service recor ds on w hich this study is based have sever al limitations: more birds were col lected in agr icul tural and subur ban t han nat ural areas; seasons and t ime of day of col lect ion wer e conveni ent to the col lectors; qui ckl y digest ed foods such as ear thwor ms and ot her so ft-bodi ed insect s are under represent ed.
12 Birds nest ing i n high densi ties in dense coni ferous f orest probabl y foraged el sewher e more of the time than di d bi rds wi th larger territo ries in less dense f orests.
13 Also i ncluded dat a from Howell (1942) (Ithaca, New York) in calcul ations.
2-201 A
merican R
obin

References (including A ppendix)
Aldrich, J. W.; James, F. C. (1991) Ecogeographic var iation in t he Amer ican robin ( Turdus migratorius). Auk 108: 230-249.
Armstr ong, J. T. (1965) Breeding hom e range in t he night hawk an d other birds: its evolut iona ry and ecologic al signif icance. Ecology 46: 619-629.
Baird, J. W. (1980) The select ion a nd us e of fr uit by bir ds in an easter n forest. Wilson Bull. 92: 63-73.
Bovi tz, P. (1990) Relations hips of fo raging s ubs trate sel ect ion a nd r oos ting in A mer ican robins and E uropean star lings [master 's th esis]. New Brunswi ck, NJ: Rutgers Universi ty.
Brackbill, H. (1952) Three-broode d Amer ican robin. Bir d-Banding 2 3: 29.
Butts, W. K. (1927) The feeding r ange of cer tain bir ds. Auk 44: 329-350.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Clench, M. H.; Leberman, R. C. (1978) Weights o f 151 sp ecies of Pennsyl vania birds analyzed by mo nth, age, and sex. Bull. Carnegie Mus. Nat. His t.
Dorst, J. (1962) The migration of bir ds. Bos ton: Hought on Mif flin Co.
Dunning, J. B., Jr . (1984) Body weight s of 686 species of North Amer ican birds. Wester n Bir d Banding A sso ciation, Monogr aph No. 1. Cave Creek, AZ: Eldon P ublis hing.
Farner, D. S. (1945) Age groups and longe vi ty in t he Amer ican robin. Wils on Bull. 57: 56-74.
Farner, D. S. (1949) Age groups and longe vi ty in t he Amer ican robin: commen ts, further dis cussion, and c ertain r evisions . Wils on Bull. 61: 68-81.
Fulle r, P. (1977) Fall robin m igr ation. Loon 4 9: 239-240.
Gochfeld, M.; Burger, J. (1984) Age differ ences i n foraging be havior of th e Amer ican robin (Turdus migratorius). Behaviour 88: 227-239.
Hamilton, W. J., Jr . (1940) Summer food of th e robin d etermined by fecal analyses. Wilson Bull. 52: 79-82.
Hamilton, W. J., Jr . (1943) Spring f ood of th e robin in c entral New York. Auk 60: 273.
2-202 Amer ican Robin

Hazelton, P. K., Robel, R. J.; Dayto n, A. D. (1984) Preferences an d influences o f paired food items o n energy intake o f Amer ican robins ( Turdus migratorius) and gr ay catbir ds (Dumetel la carolinensis). J. Wildl. Manage 48: 198-202.
Henny, C. J. (1972) An analysi s of th e popula tion dy namics of sel ected avian species wi th special reference to changes during t he mode rn pesti cide era. Washingt on, DC: Bur . Spor t. Fish. Wildl. , Wildl. Res. Rep. 1.
Hirth, D. H.; Hester , A. E.; Greeley, F. (1969) Dispersal and flocking of mar ked young r obins (Turdus m. migratorius) after fledging. Bir d-Banding 4 0: 208-215.
Howel l, J. C. (1940) Spring r oosts o f th e robin. Wils on Bull. 52: 19-23.
Howel l, J. C. (1942) Notes o n the nest ing ha bit s of th e Amer ican robin ( Turdus migratorius L.). Am. Midl. Nat. 28: 529-603.
James, F. C.; Shuga rt, H. H. (1974) The phenology of th e nesting seaso n of th e Amer ican robin ( Turdus migratorius) in the United States. Condor 76: 159-168.
Jung, R. E. (1992) Indiv idua l variation in f ruit choic e by Amer ican robins Turdus migratorius. Auk 109: 98-111.
Karasov, W. H.; Levey, D. J. (1990) Digesti ve system tr ade-offs an d adaptat ions of frugiv orous p asser ine birds. Physiol. Zool. 63: 1248-1270.
Kendeigh, S. C. (1969) Tolerance of cold a nd Be rgmann' s rule . Auk 86: 13-25.
Klimstr a, W. D.; Stieglit z, W. O. (1957) Notes o n reproducti ve acti vi ties of robins in Iow a and Illinois . Wils on Bull. 69: 333-337.
Knupp, D. M.; Owen, R. B.; Dimond, J. B. (1977) Reproduc tive biology of Amer ican robins in northern Maine. Auk 94: 80-85.
Lasiewski , R. C.; Dawso n, W. R. (1967). A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Levey, D. J.; Karasov, W. H. (1989) Digesti ve responses o f temp erate birds swi tch ed to fruit or insect d iets. Auk 106: 675-686.
Levey, D. J.; Karasov, W. H. (1992) Digesti ve modula tion in a seaso nal frugiv ore: the Amer ican robin Turdus migratorius. Am. J. Ph ysiol. 262: G711-G718.
Malmbor g, P. K.; Wills on, M. F. (1988) Foraging e cology of avi an frugiv ores and s ome consequences fo r seed dispersal in an Illinois woodlot . Condor 90: 173-186.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York, NY: McGraw-Hill Book Company, Inc.
2-203 Amer ican Robin

Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Morrison, D. W.; Caccami se, D. F. (1990) Comparison of roost u se by th ree species of commu nal roostmates. Condor 92: 405-412.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Geogr aphic So ciety. (1987) Field guide to the bir ds of Nor th Amer ica. Washingt on, DC: Nationa l Geogr aphic So ciety.
Paszkowski , C. A. (1982) Vegetat ion, ground, and f rugiv orous fo raging of th e Amer ican robin Turdus migratorius. Auk 99: 701-709.
Pitts, T. D. (1984) Descr iption of A mer ican robin t erritories in nor thwest T ennessee. Migr ant 55: 1-6.
Robbins , C. S.; Bruun, B.; Zim, H. S. (1983) A guide to field ide nt if ication: bir ds of Nor th Amer ica. New York, NY: Golde n Press.
Robbins , C. S.; Sauer, J. R.; Greenberg, R. S.; et al. (1989) Popula tion de cline s in Nor th Amer ican birds th at mi grate to the neotropics. Proc. Natl . Acad. Sci . USA 86: 7658-7662.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Sharp, M. H. (1990) Amer ica's songbir d--species profi le: Amer ican robin ( Turdus migratorius). Wild Bir d 4: 22-27.
Skorupa, J. P.; Hothem, R. L. (1985) Cons umpt ion of co mmer cially-grown grapes by Amer ican robins ( Turdus migratorius): a field evalua tion of labor atory esti mates. J. Field Ornit hol. 56: 369-378.
Speirs, J. M. (1953) Winter distr ibut ion of r obins east o f th e Rocky Mount ains . Wils on Bull. 65: 175-183.
Terborgh, J. (1989). Where have al l the birds gone ? Princeto n, NJ: Princeto n Universi ty Press.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Weatherhead, P. J.; McRae, S. B. (1990) Brood c are in Amer ican robins : impli cations fo r mixed reproducti ve str ategies by femal es. Anim. Behav. 39: 1179-1188.
Wheelwr ight , N. T. (1986) The diet of Amer ican robins : an analysi s of U.S. Biologic al Survey r ecords. Auk 103: 710-725.
2-204 Amer ican Robin

Wheelwr ight , N. T. (1988) Seasonal changes in food pr eferences o f Amer ican robins in captivi ty. Auk 105: 374-378.
Young, H. (1951) Territorial behavior of th e easter n robin. Proc. Linne an Soc. N.Y. 58-62: 1-37.
Young, H. (1955) Breeding be havior and n esting of th e easter n robin. Am. Midl. Nat. 53: 329-352.
2-205 Amer ican Robin

2.2. MAMMALS
Table 2-2 lists th e mammal ian species descr ibed in th is sect ion. For range map s,
refer to the general references i denti fied in the indiv idua l species pr of iles. The remainde r
of th is sect ion is organized by sp ecies in t he order presented in Table 2-2. The avai labilit y
of inf ormation in t he publis hed lit erature var ies sub stan tially among s pecies, as is r eflected
in the profi les. Some of th e selected species include two or more subs pecies; these are
indi cated in the profi les wh en reported by th e investi gators. Body lengt hs of th e mammal s
are reported for the length of th e outstr etch ed animal from th e tip of th e nose to the base
of th e tai l. The tai l measu rements d o not include th e hairs at the tip, but only th e tai l
ver tebrae. Body weight is r epor ted as fresh wet we ight wi th pelage, unl ess o therwise
noted.
2-207

l
Table 2-2. Mammal s Include d in t he Handbook
Order Subfamily Commo n name Scientif ic name Section
Soricidae short-tai led shrew Blarina brevicauda 2.2.1
Canidae red fox Vulpes vu lpes 2.2.2
Procyoni dae raccoon Procyon l otor 2.2.3
Mustel idae Mustelin ae min k Mustel a vison 2.2.4 Lut rinae river ot ter Lutra can adensis 2.2.5
Phocidae harbor seal Phoca vi tu ina 2.2.6
Criceti dae Sigmodont inae deer mous e Peromyscu s manicu latus 2.2.7 Arvico linae prairie vo le Microtus ochroga ster 2.2.8
meadow vole Microtus penns ylvanicus 2.2.9 muskrat Ondatra zibethicus 2.2.10
Leporidae eastern cotto ntail Sylv ilagus floridanus 2.2.11
2-208

�
�
2.2.1. Short-T ailed Shrew (shrew s)
Order Insecti vora, Family Soric idae. Shrews ar e smal l insecti vorous mammal s th at inha bit mo st regions of th e Unit ed States. They have high metabolic r ates an d can eat approximatel y th eir body weight in f ood each day. Most sp ecies are primar ily ver mivorous and insecti vorous, but so me also eat smal l birds and mammal s.
Selected species
The northern short-tai led shrew (Blarina brevicauda) ranges throughout th e nor th-central and northeaster n United States an d into southern Canada (George et al ., 1986). It eats i nsects, worms, snails, and other inver tebrates an d also may eat mi ce, voles, frogs, and other ver tebrates (Robins on and Br odie , 1982). Because th ey prey on other ver tebrates, shrews can concentrate DDT (and presumably other bioaccu mulative chemicals) to levels 10 ti mes highe r th an either Peromyscu s and Clethrionomys (Dimond and Sherburne, 1969). Shrews ar e an important co mpone nt of th e die t of man y ow ls (Palmer and Fowler, 1975; Burt and Grossen heider, 1980) and are also prey fo r other raptors, fox, weasel s, and other carnivorous mammal s (Buckn er, 1966).
Body size. Short-tai led shrews ar e 8 to 10 cm i n length wi th a 1.9 to 3.0 cm tai l (Burt and Grossen heider, 1980). The short-tai led shrew is th e largest memb er of th e genus , wi th some weighing ov er 22 g (George et al ., 1986; see tab le). Some st udies have found lit tle or no sexu al dimorphism i n size (Choate, 1972), while other repor ts show th at males are slight ly larger th an femal es (George et al ., 1986; Guilda y, 1957).
Metabolism. Short-tai led shrews ar e acti ve fo r about 16 pe rcent of each 24-hour period ( Martins en, 1969), in periods of ar ound 4 .5 minu tes at a ti me (Buckn er, 1964). The shrew's meta bolis m is inv ersely pr opor tiona l to the ambie nt temp erature, wi thin t he range of 0 to 25�C (Randolph, 1973). Sleeping metabolis m is half th at asso ciated wi th normal, explor ing acti vi ty (Randolph, 1973). Randolph (1973) develope d a regression e quation f or
emetabolis m (cc O 2/g-hour ) dur ing (1) i nter rupted sleep:
(Winter ) Y = 4.754 - 0.0869 (X - 16.4305) (Summer ) Y = 5.3448 - 0.1732 (X - 16.2310)
and (2) normal explor ing acti vi ty:
(Winter ) Y = 6.5425 - 0.0516 (X - 12.0600) (Summer ) Y = 7.949 - 0.2364 (X - 16.9554) where X= ambient temp erature
in �C.
Randolph (1973) also develope d a regression e quation f or overall metabolis m (cal /animalhour ) for sh rews sp ending e qual amount s of ti me sleeping a nd explor ing (cal /animal-hour ) as a func tion of ambie nt temp erature:
eRandol ph's ( 1973) equat ions coul d be si mplified to match that of Deavers and H udson ( 1981;next page) in form; how ever, we report the equat ions as R andol ph report ed them.
2-209 Short-Tailed Shrew

�
(Winter ) Y = 583.83 - 7.53 (X - 13.68)(Summer ) Y = 544.86 - 20.37 (X - 16.33), where X= ambient temp erature in �C.
Deavers and Huds on (1981) found a line ar inc rease in stan dard (near basal) metabolis m with decreasing temp erature that is si mi lar to that fo r inter rupted sleep descr ibed above (Y = stan dard metabolis m in c c O2/g-hour ):
Y = 8.84 - 0.22 (X) where X= ambient temp erature.
Deavers and Huds on (1981) found t hat wi thin t he thermone utral zone , the stan dard metabolic r ate of th e short-tai led shrew is approximatel y 190 percent th e metabolic r ate predicted from body weight .
Habitat . Short-tai led shrews inha bit a wide var iety o f habitats an d are commo n in areas wi th abunda nt veg etati ve co ver (Mille r and Getz, 1977). Short-tai led shrews n eed cool, mois t habitats b ecause of th eir high metabolic and water -loss r ates (Randolph, 1973).
Food ha bi ts . The short-tai led shrew is primar ily car nivorous. Stomach analyses indi cate th at insects, earthworms, slugs , and s nails can make u p most o f th e shrew's food, while plants, fungi, millipe des, cen tipe des, arachnids , and smal l mammal s also are consumed (Hamilton, 1941; Whitaker and Ferraro, 1963). Smal l mammal s are consumed more when inver tebrates ar e less avai lable (Allen, 1938; Platt an d Blakeley, 1973, ci ted in George et al ., 1986). Shrews ar e able to prey on smal l ver tebrates b ecause th ey produce a pois on secr etion in t heir saliv ary gla nds th at is tr ansmi tted during bit ing (Pearson, 1942, ci ted in Eadie, 1952). The short-tai led shrew sto res food, especially in t he autumn and winter (Hamilton, 1930; Martin, 1984). Robins on and Br odie (1982) found t hat shor t-tai led shrews cach ed most (86.6 percent) of th e prey cap tured; only 9. 4 percent was co nsumed immed iately. Short-tai led shrews co nsume approximatel y 40 percent mo re food in w int er than in summer (Randolph, 1973). The shrew mu st co nsume water to compensate fo r its high evaporative water loss, despite th e fact th at i t obtains water from both food a nd metabolic ox idation ( Chew, 1951). Deavers and Huds on (1981) indi cated that th e short-tai led shrew's evap orative water loss i ncreases wi th increasing a mbie nt temp erature even within it s thermone utral zone . Shor t-tai led shrews' digesti ve effi ciency i s about 90 pe rcent (Randolph, 1973).
Temperature regulation and molt. The short-tai led shrew does not unde rgo t orpor but uses nons hiv ering t hermoge nesis (NST) to compensate fo r heat loss d uring c old str ess i n winter (Zegers and Merritt, 1987). The short-tai led shrew exhibit s three molt s. Two are seaso nal molts, the fi rst i n Octo ber/Novemb er replaces su mmer wi th winter pelage and occu rs in fi rst- an d second-year shrews. The spring m olt can occur any time from February to Octo ber. The third molt occu rs in postj uveniles th at have reached adult size (Findle y and J ones, 1956).
Breeding acti vi ties and social organization. The short-tai led shrew probably breeds all y ear, inc luding lim ited breeding in w int er even in the northern portions of its range (Blus , 1971). In Illinois , males were found t o be mo st acti ve fr om Jan uary to July, femal es from March to Septemb er (Getz, 1989). There are two peak breeding pe riods , in
2-210 Short-Tailed Shrew

�
�
�
�
the spring a nd in l ate su mmer or early fal l (Blair, 1940). The home ranges of sh ort-tai led shrews i n summer over lap both wi thin a nd b etween sexes (B lair, 1940), although femal es wi th young do e xhibit so me ter ritorialit y (Platt, 1976). Nomadic sh rews ar e either young of the year or adult s moving t o areas wi th more abunda nt prey (Platt, 1976).
Home range and r esour ces. Short-tai led shrews inha bit r ound, unde rground n ests and m aint ain unde rground r unaways, usually in the top 10 cm o f soil, but so meti mes as deep as 50 cm (H amilton, 1931; and Jameso n, 1943, ci ted in George et al ., 1986). Winter , nonbr eeding hom e ranges can vary from 0.03 to 0.07 ha at high pr ey densit ies to 1 to 2.2 ha dur ing low prey densit ies wi th a minim um of ter ritory ov erlap. In the summer , ranges of oppos ite sex an imals over lap, but same sex indiv idua ls do not ; femal es wi th young exclude all others fr om th eir area (Platt, 1976).
Popul ation de nsity . Popula tion de nsit ies vary by habitat an d seaso n (Getz, 1989; Jackso n, 1961; Platt, 1968). In east-cen tral Illinois , popula tion de nsity was highe r in blue grass th an in tallgr ass o r alfal fa (Getz, 1989). In all three of th ese habitats, the short-taile d shrew exhibit ed annua l abunda nce cycl es, wi th peak densi ties ranging f rom 2.5 to 45 shrews p er hectar e, depending on t he habitat (Getz , 1989). The peaks o ccu rred from Ju ly to Octo ber (12.9/ha average for all three habitats), apparently just f ollow ing pe ak precipit ation l evels (Getz, 1989).
Popul ation dy namics. Winter mortal ity u p to 90 percent has been reported for the short-tai led shrew (Barbehenn, 1958; Gottsch ang, 1965; Jackso n, 1961, ci ted in George et al., 1986); however , Buckn er (1966) suggests th at mo rtal ity r ates i n winter may b e closer to 70 percent, which is si mi lar to the average monthly mo rtal ity r ate he found f or suba dult animals. Several litter s, averaging f our to five pups , are bor n each year (George et al ., 1986).
Simi lar species (fr om general references)
� The masked shrew (Sorex ci nereus) (lengt h 5.1 to 6.4 cm; weight 3 to 6 g) is smal ler than the short-tai led shrew and is th e most co mmo n shrew in moist forests, open count ry, and br ush of th e nor thern Unit ed States an d throughout Canada and A laska. It feed s primar ily on insects.
� Merriam's sh rew (Sorex mer riami ) (5.7 to 6.4 cm) i s found in a rid areas and sagebrush or bunc hgr ass o f th e wester n United States an d is smal ler than the short-tai led shrew.
� The smo key sh rew (Sorex fu meus) (6.4 to 7.6 cm; 6 to 9 g), smal ler than the short-tai led, prefers birch and hemlock fo rests wi th a thick l eaf mold on t he ground t o bur row in. It uses b urrows mad e by smal l mammal s or nests i n stu mps, logs , and among r ocks. Range is l imi ted to the northeast Unit ed States an d east o f th e Great Lakes i n Canada.
� The southeaster n shrew (Sorex longiro stris ) (5.1 to 6.4 cm; 3 to 6 g) prefers moist ar eas. Found m ostl y in ope n f ields and w oodlot s, its range is lim ited
2-211 Short-Tailed Shrew

�
�
�
�
�
�
to the southeaster n United States. It nests i n dry grass o r leaves i n a shallow depression.
� The long -tai led shrew (Sorex dispar) (7.0 cm; 5 to 6 g) inha bit s cool, mois t, rocky ar eas in deciduous or deciduou s-co niferous fo rests o f th e northeast, extending s out h to the Nor th Carolina and T ennessee b order.
� The vagrant sh rew (Sorex vag rans) (5.9 to 7.3 cm; 7 ± g) inha bit s marshy wetl ands and forest str eams. Its r ange is co nfined to the wester n Unit ed States, exc luding m ost of Calif ornia and N evada. In addit ion t o insects, it also eats p lant mater ial.
� The Paci fic sh rew (Sorex paci fi cus) (8.9 cm) i s s light ly larger th an the shor t-taile d shrew. It is lim ited to redwood a nd s pruce forests, marshes, and swamp s of th e northern California and southern Oregon c oasts.
� The dwar f sh rew (Sorex nanus) (6.4 cm) i s rare throughout its lim ited range in the wester n United States.
� The least sh rew (Cryptotis parva) (5.6 to 6.4 cm; 4 to 7 g) is easi ly dis tinguis hed f rom other sh rews b y i ts c inna mon c olor . It inha bit s gr assl and and marsh; its r ange is si mi lar to the short-tai led shrew but does not exten d as far north.
� The deser t sh rew (Notiosorex cr awfo rdi) (Gray sh rew) (5.1 to 6.6 cm) i s rarely seen and is found only in t he arid c ondit ions , chaparral s lope s, alluv ial fans, and a round low d eser t sh rubs of th e extr eme so uthwest. It nests beneath plants, boards, or debris.
General references
Burt and Grossen heider (1980); George et al . (1986).
2-212 Short-Tailed Shrew

Short-T ailed Shrew (Blarina brevicauda )
2-213 S
ho
rt-Tailed
Sh
rew
Factors Age/Sex/ Range Note Cond. /Seas. Mean or (95% CI) Locat ion Reference No.
Body Weight A B 15.0 ± 0.78 SD New Hampshi re Schl esinger & Pot ter, 1974 (g)
M sum mer 19.21 ± 0.42 SD 17.0 - 22.0 w Pennsy lvani a Guilday, 1957 F sum mer 17.40 ± 0.48 SD 14.0 - 21.0 M fall 16.87 ± 0.21 SD 13.0 - 22.0 M fall 15.58 ± 0.23 SD 12.5 - 22.5
neonat e 0.67 - 1.29 Maryland/ lab Blus, 1971
Metabol ic basal 82 80 - 84 Pennsy lvani a/lab Pearson, 1947 1 Rate average dai ly 125 106 - 150 (lO /kg-day)2
average dai ly 127 ± 15.3 SD 94 - 218 NS/lab Morrison, 1948 2
+ 20��C 126.5 Ontario, CAN/lab Randol ph, 1973 - 20��C 207.1
Metabol ic Rate basal 390 Pennsy lvani a/lab Pearson, 1947 3 (kcal /kg-day) average dai ly 600
average dai ly 680 Wisconsi n/lab Morrison et al., 1957 4
Food Ingest ion A B: 22 - 23��C 7.95 ±0.17 g/d SD Ohio/lab Barrett & Stuek, 1976 5 Rate 0.49 g/g-day
A B: 25��C Wisconsin /lab Morris on et al., 1957 6 0.62 g/g-day
Water A B 0.223 Illinoi s/lab Chew, 1951 Ingest ion Rate (g/g-day)
Inhal ation A B 0.026 est imated 7 Rate (m /day)3
Surface Ar ea A B 54 Pennsy lvani a/lab Pearson, 1947 8 (cm )2
A B 84 est imated 9

Short-T ailed Shrew (Blarina brevicauda )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
earthworms 31.4 New York/NS Whitaker & Ferraro, 1963 slugs & snai ls 27.1 misc. animals 8.1 (% vol ume; stomach Endegon ( fungi ) 7.7 cont ents) beetles 5.9 veget ation 5.4 (June t hrough Oct ober lepidopt eran 4.3 col lect ions com bined) larvae
chi lopoda 1.8 other 8.6
insect s 77.6 eastern Uni ted States Hamilton, 1941 annelid s 41.8 (primarily New York )/NS veget able matter 17.1 cent ipedes 7.4 (% frequency of arachni ds 6.1 occur rence; stomach snai ls 5.4 cont ents) small mammals 5.2 (all seasons com bined) crust acea 3.7 undet ermined 2.4
Popul ation Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home Range A F sum mer < 0.1 - 0.36 s Michi gan/bluegrass Blair, 1940 Size (ha) A M sum mer < 0.1 - 1.8
B B al l 0.39 ± 0.036 SD s Mani toba/tamarack bog Buckner , 1966
B B w inter (a) 0.03 - 0.07 c New York/old field Platt, 1976 10 B B w inter (b) 0.10 - 0.22
Range
2-214 S
ho
rt-Tailed
Sh
rew

Short-T ailed Shrew (Blarina brevicauda )
Popul ation Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Popul ation winter 2.3 ec Illinoi s/alfalfa Getz, 1989 Densi ty (N/ha) spr ing 5.9
sum mer 11.4 fall 10.0
B B 0.06 - 0.16 Mani toba, Canada/ Buckner , 1966
1.6 - 121 Wisconsi n/beech-maple Jackson, 1961; Willia ms, 11
tamarack bog
1936
Li tter 5.4 2 - 8 Indi ana/NS French, 1984 Size
4.7 ± 0.2 SE 1 - 8 Maryland/ lab Blus, 1971
Li tters sever al NS/NS George et al., 1986 /Year
Days 21 - 22 Maryland/ lab Blus, 1971 12 Gestation
Age at 25 - 30 Maryland/ lab Blus, 1971 Weaning (days)
Age at M > 65 days Maryland/ lab Blus, 1971 Sexual Maturity M > 83 days NS/NS Pearson, 1944 11
F < 1 year Indi ana/NS French, 1984
Annual B B 93 % MD, PA, NY, MA/NS Pearson, 1945 Morta lity
Longevi ty M 4.6 mont hs Maryland/ lab Blus, 1971 13 F 4.4 mont hs
B 20 mont hs c New Yor k/woods, field Dapson, 1968
Range
<
2-215 S
ho
rt-Tailed
Sh
rew
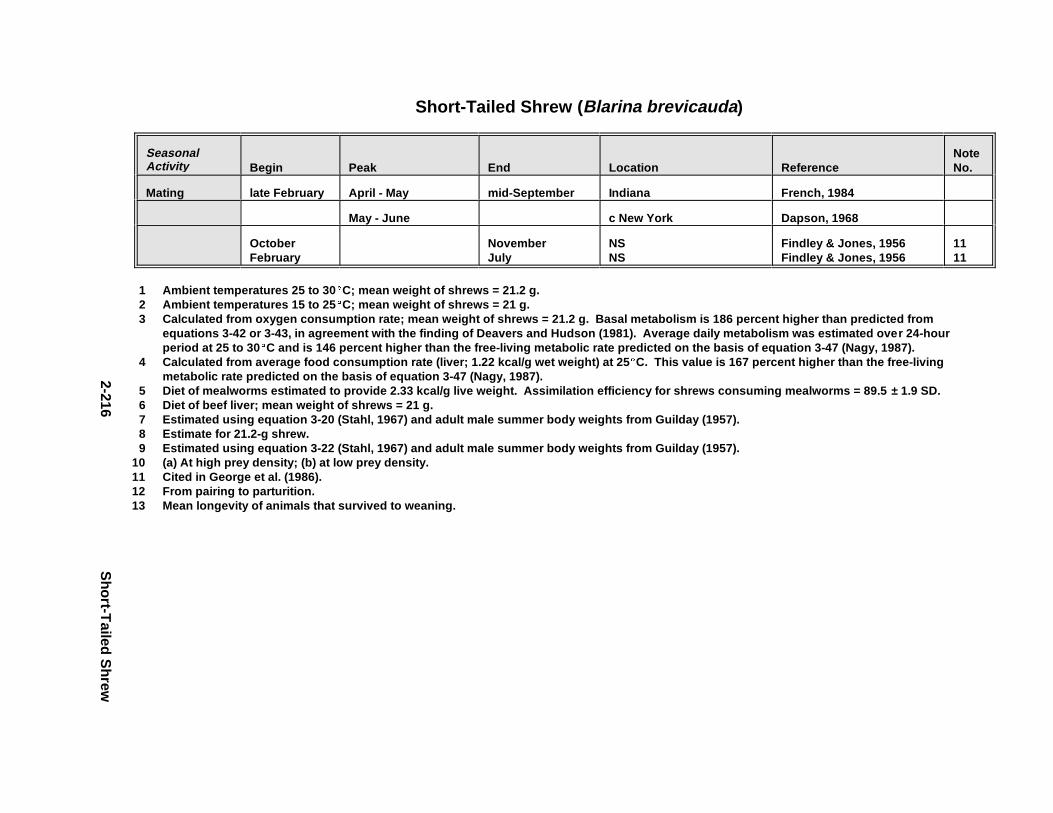
�
�
�
�
Short-T ailed Shrew (Blarina brevicauda )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating late February Apr il - May mid-September Indi ana French, 1984
Partur ition May - June c New York Dapson, 1968
Molt fall October Novem ber NS Findl ey & Jones, 1956 11 spr ing February Jul y NS Findl ey & Jones, 1956 11
1 Ambient temperatures 25 to 30�C; mean weight of shr ews = 21.2 g. 2 Ambient temperatures 15 to 25�C; mean wei ght of shr ews = 21 g.
2-216 S
ho
rt-Tailed
Sh
rew
3 Calcul ated f rom oxy gen consum pt ion r ate; mean weight of shr ews = 21.2 g. Basal metabol ism is 186 percent higher than pr edicted f rom equat ions 3- 42 or 3-43, in agreement with the findi ng of Deavers and Hudson ( 1981). Aver age dai ly metabol ism was est imated ove r 24-hour per iod at 25 to 30�C and i s 146 percent higher than the free-living m etabol ic rate predicted on t he basi s of equat ion 3-47 (Nagy, 1987).
4 Calcul ated f rom average food consum pt ion r ate (liver; 1.22 kcal /g wet weight ) at 25�C. This val ue is 167 percent higher than the free-living metabol ic rate predicted on t he basi s of equat ion 3-47 (Nagy, 1987).
5 Diet of mealworms est imated to provi de 2.33 kcal /g live weight . Assi milation ef ficiency for shr ews consum ing m ealworms = 89.5 ± 1.9 SD. 6 Diet of beef liver; mean weight of shr ews = 21 g. 7 Est imated usi ng equat ion 3-20 (Stahl , 1967) and adul t male sum mer body weight s f rom Gui lday (1957). 8 Est imate for 21.2-g shr ew. 9 Est imated usi ng equat ion 3-22 (Stahl , 1967) and adul t male sum mer body weight s f rom Gui lday (1957).
10 (a) At high pr ey densi ty; (b) at low pr ey densi ty.11 Cited in George et al. (1986).12 From pairing t o partur ition.13 Mean longevi ty of animals that sur vived t o weaning.

References (including A ppendix)
Allen, D. L. (1938) Ecologic al studie s on t he ver tebrate fau na of a 500-acr e farm in Kalamazoo Count y, Michiga n. Ecol. Monogr . 8: 347-436.
Barbehenn, K. R. (1958) Spatial and popula tion r elations hips b etween Microtus and Blarin a. Ecology 39: 293-304.
Barrett, G. W.; Stueck, K. L. (1976) Caloric inge stion r ate and assi mi lation effi ciency o f th e short-tai led shrew, Blarina brevicauda. Ohio J . Sci . 76: 25-26.
Blair, W. F. (1940) Notes o n home ranges and popula tions of th e shor t-tai led shrew. Ecology 21: 284-288.
Blair, W. F. (1941) Some data on the home ranges and general life histo ry of sh ort-tai led shrews, red-backed voles and woodla nd jum ping m ice in nor thern Mic higa n. Am. Midl. Nat. 25: 681-685.
Blus , L. J. (1971) Reproduc tion a nd s urviv al of shor t-tai led shrews (Blarina brevicauda) in captivi ty. Lab. Anim. Sci . 21: 884-891.
Buckn er, C. H. (1964) Metabolis m, food c apaci ty, and feeding be havior in f our species of shrews. Can. J. Zool. 42: 259-279.
Buckn er, C. H. (1966) Popula tions and e cologic al relations hips of sh rews i n tamar ack bogs of so utheaster n Manitoba. J. Mammal . 47: 181-194.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Chew, R. M. (1951) The water exch anges of so me smal l mammal s. Ecol. Monogr . 21: 215-225.
Choate, J. R. (1972) Variation w ithin a nd among popula tions of shor t-tai led shrews, Blarin a brevicauda. J. Mammal . 53: 116-128.
Dapson, R. W. (1968) Reproduc tion a nd age str ucture in a popula tion of shor t-tai led shrews, Blarina brevicauda. J. Mammal . 49: 205-214.
Deavers, D. R.; Huds on, J. W. (1981) Temperature regula tion in t wo rodents (Clethrionomys gapperi and Peromyscu s leucopus ) and a shrew (Blarina brevicauda) inha bit ing t he same en vironment. Physiol. Zool. 54: 94-108.
Dimond, J. B.; Sherbur ne, J. A. (1969) Persisten ce of DDT in wild popula tions of smal l mammal s. Nature 221: 486-487.
2-217 Short-Tailed Shrew

Eadie, R. W. (1952) Shrew predation a nd v ole popula tions on a loc alized area. J. Mammal . 33: 185-189.
Findle y, J. S. ; Jone s, J. K., Jr . (1956) Molt of th e shor t-tai led shrew, Blarina brevicauda. Am. Midl. Nat. 56: 246-249.
French, T. W. (1984) Reproduc tion a nd age str ucture of th ree India na sh rews. Proc. India na Acad. Sci . 94: 641-644.
George, S. B.; Choate, J. R.; Genoways, H. H. (1986) Blarina brevicauda. Amer ican Society of Mammalogi sts; Mammal ian Species 261.
Getz, L. L. (1989) A 14-year stu dy of Blarina brevicauda popula tions in east-cen tral Illinois . J. Mammal . 70: 58-66.
Gottsch ang, J. L. (1965) Winter popula tions of smal l mammal s in old f ields of sout hwester n Ohio. J. Mammal . 46: 44-52.
Guilda y, J. E. (1957) Indiv idua l and ge ogr aphic var iation in Blarina brevicauda from Pennsyl vania. Ann. Carnegie Mus. 35: 41-68.
Hamilton, W. J., Jr . (1929) Breeding ha bit s of th e shor t-tai led shrew, Blarina brevicauda. J. Mammal . 10: 125-134.
Hamilton, W. J., Jr . (1930) The food of th e Soricida e. J. Mammal . 11: 26-39.
Hamilton, W. J., Jr . (1931) Habits o f th e short-tai led shrew, Blarina brevicauda (Say). Ohio J. Sci . 31: 97-106.
Hamilton, W. J., Jr . (1941) The foods of smal l forest mammal s in easter n United States. J. Mammal . 22: 250-263.
Jackso n, H. H. T. (1961) Mammal s of Wisco nsin. Madison, WI: Universi ty o f Wisco nsin Press.
Jameso n, E. W., Jr . (1943) Notes o n the habits an d sipha napterous parasi tes o f th e mammal s of Welland Count y, Ontar io. J. Mammal . 24: 194-197.
Lom olino, M. V. (1984) Immi grant sel ect ion, predation, and t he dis tr ibut ion of Microtus pennsyl vanicus and Blarina brevicauda on islands. Am. Nat. 123: 468-483.
Martin, I. G. (1984) Facto rs affect ing f ood hoa rding in t he shor t-tai led shrew Blarin a brevicauda. Mammal ia 48: 65-71.
Martinsen, D. L. (1969) Energetics an d acti vi ty p atter ns of sh ort-tai led shrews (Blarin a) on restr icted diets. Ecology 50: 505-510.
2-218 Short-Tailed Shrew

Mille r, H.; Getz, L. L. (1977) Facto rs influencing loc al dis tr ibut ion a nd s pecies div ersi ty of forest smal l mammal s in new Engla nd. Can. J. Zool. 55: 806-814.
Morrison, P. R. (1948) Oxyg en consumption in sever al smal l wild mammal s. J. Cell. Comp. Physiol. 31: 69-96.
Morrison, P. R.; Pierce, M.; Ryser , F. A. (1957) Food c ons umpt ion a nd body weight in t he masked and short-tai led shrews (g enus Blarin a) in Kansas, Iowa, and Misso uri. Ann. Carnegie Mus. 51: 157-180.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Mono. 57: 111-128.
Neal, C. M.; Lusti ck, S. I. (1973) Energetics an d evaporative water loss i n the short-tai led shrew Blarina brevicauda. Physiol. Zool. 46: 180-185.
Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
Pearson, O. P. (1942) The cause and nature of a pois onous action pr oduc ed by th e bit e of a shrew (Blarina brevicauda). J. Mammal . 23: 159-166.
Pearson, O. P. (1944) Reproduc tion in t he shrew (Blarina brevicauda Say). Am. J. Anat. 75: 39-93.
Pearson, O. P. (1945) Longe vi ty of th e shor t-tai led shrew. Am. Midl. Nat. 34: 531-546.
Pearson, O. P. (1947) The rate of meta bolis m of so me smal l mammal s. Ecology 29: 127-145.
Platt, A. P. (1968) Differ ential trap mortal ity as a measu re of str ess d uring t imes o f popula tion inc rease and decrease. J. Mammal . 49: 331-335.
Platt, W. J. (1974) Metabolic r ates o f sh ort-tai led shrews. Physiol. Zool. 47: 75-90.
Platt, W. J. (1976) The social organization a nd t erritorialit y of shor t-tai led shrew (Blarin a brevicauda) popula tions in old- field ha bitats. Anim. Behav. 24: 305-318.
Platt, W. J.; Blakeley, N. R. (1973) Short-ter m effects o f sh rew predation upon inv ertebrate prey sets i n prairie ecosystems. Proc. Iowa Acad. Sci . 80: 60-66.
Randolph, J. C. (1973) Ecologic al energetics of a hom eothermic pr edator, the shor t-tai led shrew. Ecology 54: 1166-1187.
Richardson, J. H. (1973) Locomotory and feeding acti vi ty o f th e shrews, Blarina brevicauda and Sunc us murinus . Am. Midl. Nat. 90: 224-227.
2-219 Short-Tailed Shrew

Robins on, D. E.; Brodie , E. D. (1982) Food hoa rding be havior in t he shor t-tai led shrew Blarina brevicauda. Am. Midl. Nat. 108: 369-375.
Schle singe r, W. H.; Potter , G. L. (1974) Lead, coppe r, and c admium conc entrations in smal l mammal s in the Hubba rd Brook Exp erimental Forest. Oikos 25:148-152.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Whitaker , J. O., Jr .; Ferraro, M. G. (1963) Summer food of 220 short-tai led shrews fr om Ithaca, New York. J. Mammal . 44: 419.
Willi ams, A. B. (1936) The composi tion a nd dy namics of a beech-maple cl imax co mmunit y. Ecol. Monogr . 6: 317-408.
Zegers, D. A.; Merritt, J. F. (1987) Adaptat ions of Peromyscu s for winter survival in an Appalachian montane forest. J. Mammal . 69: 516-523.
2-220 Short-Tailed Shrew

2.2.2. Red Fox (foxes and coy otes)
Order Carnivora, Family Canidae. Unlik e the more social wol ves, foxes an d coyotes tend t o hunt alone , although c oyotes may hunt larger prey in pa irs. Foxes an d coyotes ar e pr imarily car niv orous , preying pr edom inant ly on smal l mammal s, but th ey also may eat insects, fruits, berries, seeds, and nuts. Foxes ar e found t hroughout mo st of th e Unit ed States an d Canada, including t he arcti c, as are coyotes wi th the excep tion of th e southeaster n United States. Foxes an d coyotes ar e acti ve primar ily at night .
Selected species
Red foxes (Vulpes vu lpes) are present th roughout th e Unit ed States an d Canada excep t in the southeast, extr eme so uthwest, and parts o f th e central states. Red fox prey exten sively on mice and voles but also feed on other smal l mammal s, insects, hares, game bir ds, poult ry, and o ccas iona lly seeds, berries, and fruits (Pal mer and Fowler, 1975). Twelve subs pecies are recognize d in Nor th Amer ica (Ables, 1974).
Body size. The dog- sized red fox has a body about 56 to 63 cm in le ngt h, wi th a 35 to 41 cm tai l (Burt and Grossen heider, 1980). They weigh f rom 3 to 7 kg, wi th the males usually out weighing t he femal es by about 1 kg (Voigt , 1987; see tab le).
Habitat. As th e most wi dely distr ibuted carnivore in the world, the red fox can live in habitats r anging f rom ar cti c areas to temp erate deser ts (Voigt , 1987). Red foxes u tilize many typ es of habitat--cr opla nd, rolling f armland, brush, pastu res, hardwood stan ds, and coniferous fo rests (M acGregor, 1942; Eadie, 1943; Cook and Ha milt on, 1944; Ables, 1974). They prefer areas wi th broken and diverse upla nd ha bitats su ch as occu r in most agricultural areas (Ables, 1974; Samuel and Nelson, 1982; Voigt , 1987). They are rare or absent from cont inuous stands of pine fo rests i n the southeast, moist c onifer forests a long the Paci fic co ast, and semi arid grassl ands and deser ts (Ables, 1974).
Food ha bi ts . The red fox feed s on both animal and plant mater ial, mostl y smal l mammal s, birds, insects, and fruit (Korsch gen, 1959; Samuel and Nelson, 1982). Meadow vole s are a major food in m ost areas of North Amer ica; other commo n prey include mi ce and rabbits (K orsch gen, 1959; Voigt , 1987). Game birds (e.g., ring- necked pheasant and ruffed grouse) and water fowl are seaso nally important prey in some areas (Pi ls and Martin, 1978; Sargeant, 1972; Voigt and Br oadfoot , 1983). Plant mater ial is mo st co mmo n in red fox diets i n summer and fal l when fruits, berries, and nuts b ecome avai lable (Johns on, 1970; Major and Sherburne, 1987). Red foxes o ften cach e food in a hole fo r fu ture use (Samuel and Nelson, 1982). They also are noted scaven gers on carcasses o r other refuse (Voigt , 1987). Most acti vi ty i s noctu rnal and at tw ilight (No wak an d Paradiso, 1983).
Temperature regulation and molt. In winter , foxes d o not unde rgo hibe rnation or torpor; instead , they are acti ve year -round. They unde rgo one molt per year , whic h us ually begins in April and is finis hed by June . The wint er co at is r egrown by Octobe r or Novemb er in northern lati tudes (Voigt , 1987).
2-221 Red Fox

Breeding acti vi ties and social organization. Breeding oc curs earlier in t he sout h than in the red fox's northern ranges (Samu el and Nelson, 1982) (see tab le). A mated pair maint ains a ter ritory throughout th e year , wi th the male cont ribut ing m ore to its defense than the femal e (Presto n, 1975). Pups are born and reared in an unde rground de n, and t he male assi sts th e femal e in rearing y oung, br inging f ood t o the den for th e pups (Samuel and Nelson, 1982). Pups fi rst emer ge from th e den when 4 to 5 weeks old (Samuel and Nelson, 1982). Once co nsidered solitary, red foxes n ow are reported to exhibit mo re comple x social habit s (MacDona ld and Voigt , 1985). A fox fami ly, the basic so cial unit , generally co nsists o f a mated pair or one male and sever al related femal es (MacDonald, 1980; Voigt , 1987). The addit iona l femal es are usually nonbr eeders that often help the breeding femal e (Voigt , 1987).
Home range and r esour ces. The hom e ranges of indiv idua ls fr om th e same fami ly overlap cons iderably , cons ti tuting a fami ly ter ritory (Sargeant, 1972; Voigt and Ma cDona ld, 1984). Territories of neighbor ing r ed fox familie s are largely nonov erlapping a nd cont iguous , usually r esult ing in a ll pa rts of a la ndscap e being oc cupie d by fo xes. Territory sizes range from less th an 50 to over 3,000 ha (see tab le). Territories in urban areas ten d to be smal ler than those in rural areas (Ables, 1969). Adults vi si t mo st p arts o f th eir ter ritory on a r egula r basis; however , they ten d to concentrate th eir acti vi ties near to their dens, prefer red hunt ing a reas, abunda nt food s upplie s, and r esting a reas (Ables, 1974; Keenan, 1981). Territory bounda ries often conform to physi cal landscap e featu res su ch as wel l-traveled roads and str eams (Ables, 1974). Territory defense is primar ily by nona ggr essi ve mech anisms i nvolving ur ine scen t-mar king a nd avoida nce behavior s. Scent mar king oc curs throughout th e ter ritory; there is lit tle patrolling of ter ritory bounda ries. Each fox or fami ly usually has a mai n unde rground de n and one or more other bur rows within t he hom e range (No wak an d Paradiso, 1983). Most d ens are abandone d bur rows of other sp ecies (e.g., woodc hucks, badgers) (Samu el and Nelson, 1982). Tunne ls are up to 10 m in length and lead to a chamber 1 to 3 m below th e surface (Nowak an d Paradiso, 1983). Pup-rearing de ns ar e the focal point of fo x acti vi ty d uring s pring a nd early summer . Foxes h ave so me rest si tes an d usually fo rage away fr om th e den (Voigt , 1987).
Popul ation de nsity. One red fox fami ly per 100 to 1,000 ha is typ ical (Voigt , 1987; see tab le). Red foxes h ave larger home ranges wh ere popula tion de nsit ies are low and in poor er habitats (Voigt , 1987). Most y oung f oxes, especially mal es, disperse before the age of 1 (Voigt , 1987), usually during S eptemb er to March, wi th peaks i n dispersal in Octo ber and Novemb er (Phillips et al., 1972; Storm et al ., 1976).
Popul ation dy namics. Foxes u sually produc e pups th eir fi rst year , excep t in extr emely high de nsity areas and in some year s in northern portions of th eir range wh ere they may d elay breeding unt il t he next seaso n (Allen, 1984; Harris, 1979; Storm et al ., 1976; Voigt and Ma cDona ld, 1984). Litter size generally aver ages fo ur to six pups (see tab le). The pups l eave th e den about 1 mont h after birth, and they are weaned by about 8 to 10 weeks o f age (Ables, 1974). Red foxes i ncur high m ortalit y rates as a r esult of shoot ing, trapping, disease, and acci dents (e.g., roadkills) (Sto rm et al ., 1976). Two facto rs th at ten d to lim it r ed fox abunda nce are competi tion w ith other canids , especially co yotes, and seaso nal limi ts o n food availa bilit y (Voigt , 1987). Fecundit y is highe r in areas of high mortalit y and low popula tion de nsit ies (Voigt , 1987).
2-222 Red Fox

�
�
�
�
�
Simi lar species (fr om general references)
� The arcti c fo x (Alopex lagopus ) is smalle r th an the red fox (body lengt h approximatel y 51 cm; weight 3. 2 to 6.7 kg) and is r estr icted in its d istr ibut ion to the arcti c, found in t he Unit ed States only in A laska. This sp ecies primar ily scaven ges fo r food but al so eats l emmings , hares, bir ds, and eggs as wel l as berries in seaso n.
� The swi ft fo x (Vulpes vel ox) is smalle r th an the red fox (body lengt h 38 to 51 cm; weight 1. 8 to 2.7 kg) and inha bit s the deser ts an d plains of th e southwest and central United States. It dens in ground bur rows and feeds on smal l mammal s and insects.
� The ki t fo x (Vulpes macr oti s) is si mi lar in size to the swi ft fo x and is considered by so me to be the same sp ecies, although it has not iceably larger ears. It inha bit s the sout hwester n United States an d prefers open, level , sandy areas and low deser t veg etat ion. It feeds on smal l mammal s and insects.
� The gray fo x (Urocyo n cinereoargenteus) is si mila r in s ize (body lengt h 53 to 74 cm; weight 3. 2 to 5.8 kg) to the red fox and r anges ov er mo st of th e Unit ed States excep t th e northwest an d northern prairies, inha bit ing c haparral, open forests, and rimrock r egions . Secretive and noctu rnal, gray fo xes w ill c lim b trees to evade enemies. They feed primar ily on smal l mammal s but also eat insects, fruits, acorns, birds, and eggs.
� The coyote (Canis latrans) is mu ch larger (body lengt h 81 to 94 cm; weight 9 to 22 kg) than the red fox and is found t hroughout mo st of th e Unit ed States (excep t poss ibly easter n), wester n Canada, and Alaska. It inha bit s pr airies, open woodla nds , brushy and boulde r-str ewn areas, and dens in the ground. Coyotes sh are some feeding ha bit s wi th the red fox but al so scaven ge and hunt larger prey in pa irs.
General references
Ables (1974); Burt and Grossen heider (1980); Palmer and Fowler (1975); Voigt (1987).
2-223 Red Fox

Red Fox ( Vulpes vul pes)
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A M sprin g 5.25 ± 0.18 SE 4.54 - 7.04 Illin ois Storm et al., 1976 (kg) A F spr ing 4.13 ± 0.11 SE 3.27 - 4.72
A M fall 4.82 ± 0.081 SE 4.13 - 5.68 Iowa Storm et al., 1976 A F fall 3.94 ± 0.079 SE 2.95 - 4.59
neonat e B 0.102 ± 0.12 SD 0.071 - 0.109 Wisconsi n Storm & Abl es, 1966 at weaning B 0.70 North Dakot a Sargeant, 1978
Pup Growth bi rth to 15.9 North Dakot a/lab Sargeant , 1978 Rate (g/day) weaning
Metabol ic Rate J sum mer 193 ± 56 SD Ohio/lab Vogt sber ger & Bar rett, 1973 (kcal /kg-day)
A M basal 47.9 est imated 1 A F basal 51.1
A M free-living 161 (68 - 383) est imated 2 A F free-liv ing
168 (71 - 400)
Food Ingest ion J 5-8 wks 0.16 North Dakot a/lab Sargeant , 1978 Rate (g/g-day) J 9-12 wks 0.12
J 13-24 wks 0.11
A bef ore 0.075 North Dakot a/capt ive Sargeant, 1978 3 whe lp F afte r whelp 0.14
A 0.069 North Dakot a/capt ive Sargeant, 1978 nonbr eeding
Water A M 0.084 est imated 4 Ingest ion A F 0.086 Rate (g/g-day)
Inhal ation A M 2.0 est imated 5 Rate (m /day) A F 1.73
2-224 R
ed F
ox
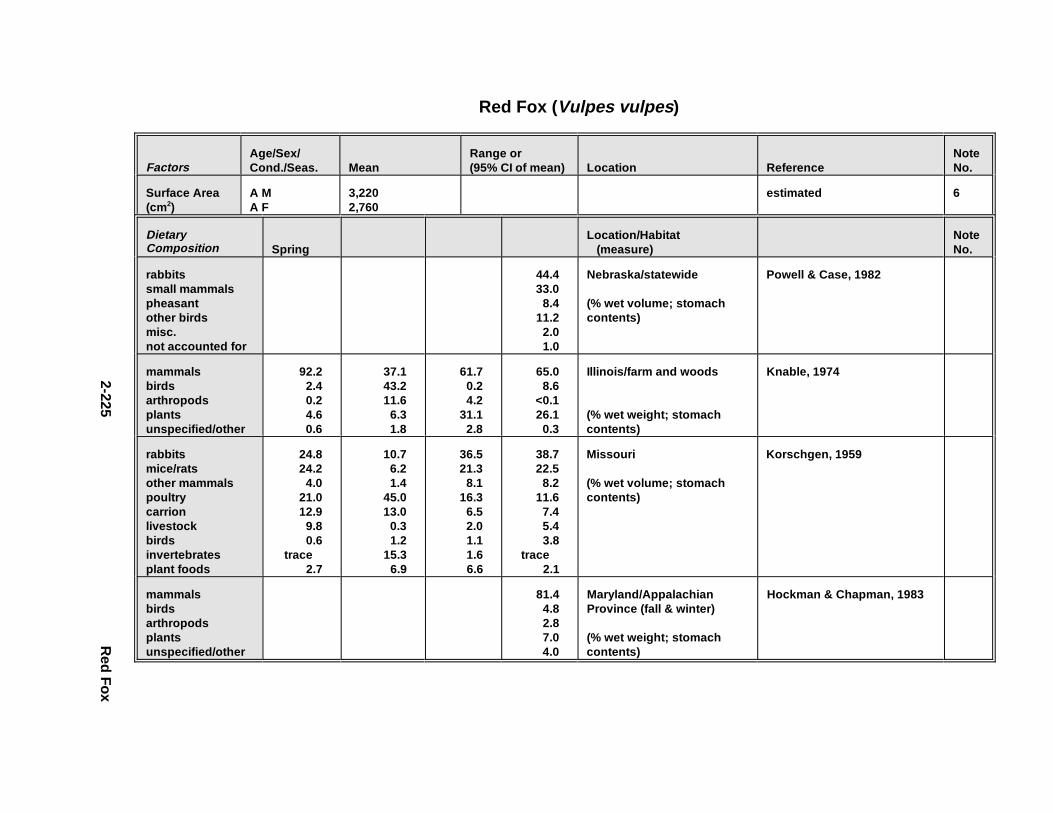
Red Fox ( Vulpes vul pes)
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Surface Ar ea A M 3,220 est imated 6 (cm ) A F 2,7602
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
rabbi ts 44.4 Nebraska/statewide Powell & Case, 1982 small mammals 33.0 pheasant 8.4 (% wet vol ume; stomach other bi rds 11.2 cont ents) misc. 2.0 not account ed for 1.0
mammals 92.2 37.1 61.7 65.0 Illinoi s/farm and woods Knabl e, 1974 birds 2.4 43.2 0.2 8.6 arthropods 0.2 11.6 4.2 <0.1 plants 4.6 6.3 31.1 26.1 (% wet weight ; stomach unspeci fied/other 0.6 1.8 2.8 0.3 cont ents)
rabbi ts 24.8 10.7 36.5 38.7 Missour i Korschgen, 1959 mice/rats 24.2 6.2 21.3 22.5 other mammals 4.0 1.4 8.1 8.2 (% wet vol ume; stomach poul try 21.0 45.0 16.3 11.6 cont ents) carrion 12.9 13.0 6.5 7.4 livest ock 9.8 0.3 2.0 5.4 birds 0.6 1.2 1.1 3.8 invertebrates trace 15.3 1.6 trace plant foods 2.7 6.9 6.6 2.1
mammals 81.4 Maryland/Appal achi an Hockm an & Chapm an, 1983 birds 4.8 Province (fall & winter) arthropods 2.8 plants 7.0 (% wet weight ; stomach unspeci fied/other 4.0 cont ents)
2-225 R
ed F
ox

Red Fox ( Vulpes vul pes)
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Territory size A B sum mer 1,611 277 - 3,420 nw Br itish Col umbia/ Jones & Theber ge, 1982 (ha) A M sum mer 1,967 514 - 3,420 alpine and subal pine
A F sum mer 1,137 277 - 1,870
A F spr ing 699 ± 137 SD 596 - 855 ec Minnesot a/woods, fields, Sargeant , 1972
A M all year 717 Wisconsi n/diverse Abl es, 1969 A F al l year 96 57 - 170
swamp
Popul ation B B spr ing 0.001 Canada/nor thern bor eal Voigt, 1987 Densi ty (N/ha) forests/arct ic tundr a
B B spr ing 0.01 s Ontario, Canada/sout hern Voigt, 1987
B B 0.046 - 0.077 "good f ox range" i n Abl es, 1974 habi tats
North America
Li tter 5.5 s Wisconsi n/farm, marsh, Pils & Martin, 1978 7 Size pasture
6.8 2 - 9 Illinoi s/farm and woods Storm et al., 1976 8 6.7 3 - 12 Iowa/farm and woods Storm et al., 1976 7 4.2 upper Michi gan/NS Switzenber g, 1950 8 4.1 North Dakot a/prairie Al len, 1984 7
pot hol es
Li tters/Year 1 NS/NS Samuel & Nelson, 1982
Days 51 - 54 New Yor k/NS Sheldon, 1949 9 Gestation
Age at 8 - 10 weeks NS/NS Ables, 1974 Weaning
Age at F 10 mont hs Illinoi s, Iowa/farm woods Storm et al., 1976 Sexual Maturity
Range
2-226 R
ed F
ox

1234567
Red Fox ( Vulpes vul pes)
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Annual B B 79.4 s Wisconsi n/var ious Pils & Martin, 1978 Morta lity Rates (percent) J M 83 Illin ois/Iowa/ Storm et al., 1976
J F 81 farms and woods A F 74 A B 77
Longevi ty < 1.5 yrs up t o 6 yrs NS/NS Storm et al., 1976
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Matin g early Dec. late January late February Iowa Storm et al., 1976 late December Jan. - Feb. March New York Layne & McKeon, 1956; 9
late Januar y early February sout hern Ontario, Canada Voigt, 1987 February March nor thern Ontario, Canada Voigt, 1987
Sheldon, 1949
Partur ition March sout hern CAN Voigt, 1987 late March, Apr il e North Dakot a Sargeant, 1972
Molt Apr il June NS/NS Voigt , 1987
Disperal late March Illin ois, Iowa Storm et al., 1976 September
Range
2-227 R
ed F
ox
Est imated usi ng ext rapol ation equat ion 3-45 (Boddi ngt on, 1978) and body weight s f rom Storm et al. (1976) (Illin ois). Est imated usi ng ext rapol ation equat ion 3-47 (Nagy, 1987) and body weight s f rom Storm et al. (1976) (Illin ois). Food consum pt ion of an adul t pair for 11 days pr ior to whel ping ( i.e., partur ition) and of the adul t female for the first 4 weeks af ter whelping.Est imated usi ng ext rapol ation equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Storm et al. (1976) (Illin ois).Est imated usi ng ext rapol ation equat ion 3-20 (Stahl , 1967) and body weight s f rom Storm et al. (1976) (Illin ois).Est imated usi ng ext rapol ation equat ion 3-22 (Stahl , 1967) and body weight s f rom Storm et al. (1976) (Illin ois).Litter size determined from embryo count . Using pl acent al scar s gener ally over est imates l itter size, and count ing l ive pups o ften under est imateslitter size (Allen, 1983; Lindst rom, 1981).
8 Method of determining l itter size not speci fied. 9 Cited in Samuel and Nel son (1982).

References (including A ppendix)
Ables, E. D. (1969) Home range studies of red foxes (Vulpes vu lpes). J. Mammal . 50: 108-120.
Ables, E. D. (1974) Ecology of th e red fox in Nor th Amer ica. In: Fox, M. W., ed. The wild canids . New York, NY: Van Nostrand Reinhold; pp. 148-163.
Allen, S. H. (1983) Comparison of red fox l itter sizes determined from count s of emb ryos and placental scar s. J. Wildl. Manage. 47: 860-863.
Allen, S. H. (1984) Some asp ects o f reproducti ve performance in the red fox in North Dakota. J. Mammal . 65: 246-255.
Allen, S. H.; Gulke, J. (1981) The effect o f age on adult r ed fox body weight s. Prairie Nat. 13: 97-98.
Asdell, S. A. (1946) Patter ns of mammal ian reproduc tion. Ithaca, NY: Comsto ck Publ. Co.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Cook , D. B.; Hamilt on, W. J., Jr . (1944) The ecologic al relations hip of r ed fox food in easter n New York. Ecology 24: 94-104.
Dalke, P. D.; Sime, P. R. (1938) Home and seaso nal ranges of th e easter n cotto ntai l in Connecti cut. Trans. N. Amer . Wildl. Conf . 3: 659-669.
Dekker , D. (1983) Denning a nd f oraging ha bit s of r ed foxes, Vulpes vu lpes, and their int eraction w ith coyotes, Canis latrans, in c entral Alberta. Can. Field-Nat. 97: 303-306.
Eadie, W. R. (1943) Food of th e red fox in s out hern New Hampshir e. J. Wildl. Manage. 7: 74-77.
Green, J. S.; Flinde rs, J. T . (1981) Diets o f symp atr ic red foxes an d coyotes i n southeaster n Idaho. Great Basin Nat. 41: 251-254.
Halpin, M. A.; Bis son ette, J. A. (1983) Winter resource use by red fox (Vulpes vu lpes) (abstr act only). Trans. Northeast Sect. Wildl. Soc. 40: 158.
2-228 Red Fox

Hamilton, W. J., Jr . (1935) Notes o n food of r ed foxes i n New York and New Engla nd. J. Mammal . 16: 16-21.
Harris, S. (1979) Age-related fer tility an d producti vi ty i n red foxes, Vulpes vu lpes, in subur ban London. J. Zool. (London) 187: 195-199.
Harris, S.; Smith, G. C. (1987) Demography of two urban fox (Vulpes vu lpes) popula tions . J. Appl. Ecol. 24: 75-86.
Hockman , J. G.; Chapman, J. A. (1983) Comparative feeding ha bit s of r ed foxes (Vulpes vulpes) and gray fo xes (Urocyo n cinereoargentus) in Maryland. Am. Midl. Nat. 110: 276-285.
Hoffman , R. A.; Kirkpatr ick, C. M. (1954) Red fox weight s and r eproduc tion in T ipp ecanoe Count y, India na. J. Mammal . 55: 504-509.
Johns on, W. J. (1970) Food ha bit s of th e red fox in Is le Royale Nationa l Park, Lake Superior . Am. Midl. Nat. 84: 568-572.
Johns on, D. H.; Sargeant, A. B. (1977) Impact o f red fox predation on t he sex r atio of prairie malla rds. Washingt on, DC: U.S. Fish Wildl. Serv.; Wildl. Res. Rep. 6.
Jones, D. M.; Theberge, J. B. (1982) Summer home range and habitat u tiliza tion of th e red fox (Vulpes vu lpes) and gray fo xes (Urocyo n cinereoargentus) in Maryland. Am. Midl. Nat. 110: 276-285.
Keenan, R. J. (1981) Spatial use of home range among r ed foxes (Vulpes vu lpes) in s out h-central Ontar io. In: Chapman, J. A.; Pursley, D., eds. Worldwide furbearer conf erence pr oceedings , Augus t, 1980; Frostb urg, Maryland; pp. 1041-1063.
Knable, A. E. (1970) Food ha bit s of th e red fox (Vulpes fu lva) in Union Count y, Illinois . Trans. Ill. State Acad. Sci . 63: 359-365.
Knable, A. E. (1974) Seasonal trends in the utiliza tion of major food gr oups by th e red fox (Vulpes fu lva) in Union Count y, Illinois . Trans. Ill. State Acad. Sci . 66: 113-115.
Korsch gen, L. J. (1959) Food ha bit s of th e red fox in Mis sour i. J. Wildl. Manage. 23: 168-176.
Kuehn, D. W.; Berg, W. E. (1981) Notes o n movemen ts, popula tion stati sti cs, and foods of the red fox in north-central Minne sota. Minn. Wildl. Res. Q. 41: 1-10.
Layne, J. N.; McKeon, W. H. (1956) Some asp ects o f red fox and gray fo x reproduc tion in New York. N.Y. Fish and Game J. 3: 44-74.
Linds trom, E. (1981) Reliabilit y of placental scar counts i n the red fox (Vulpes vu lpes L.) wi th special reference to fading of th e scar s. Mammal Rev. 11: 137-49.
2-229 Red Fox

Llewelly n, L. M.; Uhler, F. M. (1952) The foods of fu r anim als of th e Patuxent Research Refuge , Maryland. Am. Midl. Nat. 48: 193-203.
MacDona ld, D. W. (1980) Social facto rs affect ing r eproduc tion a mongs t red foxes (Vulpes vulpes L. 1758). In: Zimen, E., ed. The red fox, bioge ogr aphic a: v. 18. The Netherlands: W. Junk, The Hague; pp. 123-175.
MacDona ld, D. W.; Voigt , D. R. (1985) The biologic al basis of r abies mode ls . In: Bacon, P. J., ed. Popula tion dy namics of r abies in w ildlif e. London, UK: Academic Press; pp. 71-107.
MacGregor, A. E. (1942) Late fal l and winter foods of fo xes i n central Massach usetts. J. Wildl. Manage. 6: 221-224.
Major, J. T.; Sherburne, J. A. (1987) Interspeci fic relations hips of co yotes, bobcats, and red foxes i n wester n Maine. J. Wildl. Manage. 51: 606-616.
Maurel, D. (1980) Home range and acti vi ty r hyth m of adult mal e foxes d uring t he breeding seaso n. In: Amlaner, C. J.; MacDonald, D. W., eds. A handbook on biot elemetr y and radio t racking. Edmonds , WA: The Franklin P ress; pp. 697-702.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Mono. 57: 111-128.
Nowak, R. M.; Paradiso, J. L. (1983) Foxes. In: Walker 's mammal s of th e world. 4th ed. Balt imore, MD: Johns Hopk ins Univ ersi ty Press; pp. 932-980.
Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
Phillips , R. L.; Andr ews, R. D.; Storm, G. L.; et al . (1972) Dispersal and mortal ity o f red foxes. J. Wildl. Manage. 36: 237-248.
Pils, C. M.; Martin, M. A. (1978) Popula tion dy namics, predator-prey relations hips an d management of th e red foxes i n Wisco nsin. Madison, WI: Wisc. Dept. Nat. Resour. Tech. Bull. No. 105; 56 pp.
Pils, C. M.; Martin, M. A.; Lange, E. (1981) Harvest, age str uctu re, survivorship, produc tivi ty of red foxes i n Wisco nsin. Madison, WI: Wisc. Dept. Nat. Resour. Tech. Bull. No. 125; 19 pp.
Powel l, D. G.; Case, R. M. (1982) Food ha bit s of th e red fox in Ne braska. Trans. Nebr. Acad. Sci . and Affi l. Soc. 10: 13-16.
Presto n, E. M. (1975) Home range defense in the red fox, Vulpes vu lpes L. J. Mammal . 56: 645-652.
2-230 Red Fox

Richards, S. H.; Hine, R. L. (1953) Wisco nsin fox popula tions . Madis on, WI: Wisc. Cons . Dept . Tech. Wildl. Bull. No. 6; 78 pp.
Samuel, D. E.; Nelson, B. B. (1982) Foxes. In: Chapman, J. A.; Feldhammer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 475-490.
Sargeant, A. B. (1972) Red fox sp atial character isti cs i n relation t o water fowl predation. J. Wildl. Manage. 36: 225-236.
Sargeant, A. B. (1978) Red fox prey demands and implic ations to prairie duc k pr oduc tion. J. Wildl. Manage. 42: 520-527.
Sargeant, A. B.; Pfei fer , W. K.; Allen, S. H. (1975) A spring a erial census of r ed foxes i n Nor th Dakota. J. Wildl. Manage. 39: 30-39.
Sargeant, A. B.; Allen, S. H.; Fleskes, J. P. (1986) Commer cial sunflowers: food f or red foxes i n North Dakota. Prairie Nat. 18: 91-94.
Sargeant, A. B.; Allen, S. H.; Hastings , J. O. (1987) Spatial relations b etween symp atr ic coyotes an d red foxes i n North Dakota. J. Wildl. Manage. 51: 285-293.
Sargeant, A. B.; Allen, S. H.; Johns on, D. H. (1981) Determination of ag e and w helping d ates of liv e red fox pups . J. Wildl. Manage. 45: 760-765.
Schoon maker , W. J. (1938) Notes o n mat ing a nd br eeding ha bit s of fo xes i n New York state. J. Mammal . 19: 375-376.
Scott, T. G. (1943) Some food c oactions of th e nor thern pla ins r ed fox. Ecol. Monogr . 13: 427-480.
Sheldon, W. G. (1949) Reproducti ve behavior of fo xes i n New York state. J. Mammal . 30: 236-246.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stanley, W. C. (1963) Habits o f th e red fox in northeaster n Kansas. Univ . Kansas Mus. Nat. Hist. Misc. Pub. 34: 1-31.
Storm, G. L.; Ables, E. D. (1966) Notes o n newborn and fullt erm wild r ed foxes. J. Mammal . 47: 116-118.
Storm, G. L.; Andrews, R. D.; Phillips , R. L.; et al. (1976) Morphology , reproduc tion, dispersal and mortal ity o f mi dwester n red fox popula tions . Wildl. Monogr . 49: 1-82.
Switzenberg, D. F. (1950) Breeding pr oduc tivi ty in Mic higa n red foxes. J. Mammal . 31: 194-195.
2-231 Red Fox

Tulla r, B. J. (1983) An unus ually long- liv ed red fox. N.Y. Fish Game J. 30: 227.
Tulla r, B. J., Jr .; Berchie lli, L. T. (1980) Movemen t of th e red fox in central New York. N.Y. Fish Game J. 27: 197-204.
Vogtsb erger, L. M.; Barrett, G. W. (1973) Bioe nergetics of cap tive red foxes. J. Wildl. Manage. 37: 495-500.
Voigt , D. R.; MacDona ld, D. W. (1984) Variation in t he spatial and s ocial behaviour of th e red fox, Vulpes vu lpes. Acta Zool. Fenn. 171: 261-265.
Voigt , D. R.; Tinline , R. L. (1980) Strategies fo r analyzing r adio t racking d ata. In: Amlaner, C. J., Jr .; MacDona ld, D. W., eds. A handbook on biot elemetr y and radio t racking. Oxfo rd, Unit ed Kingdom : Pergamon P ress; pp. 387-404.
Voigt , D. (1987) Red fox. In: Novak, M.; Baker , J. A.; Obbarel, M. E.; et al ., eds. Wild furbearer management and conservat ion. Pittsb urgh, PA: Universi ty o f Pi ttsb urgh Press; pp. 379-392.
Voigt , D. R.; Broadfoot , J. (1983) Locat ing pup- rearing de ns of r ed foxes wi th radio- equippe d woodc hucks. J. Wildl. Manage. 47: 858-859.
2-232 Red Fox

2.2.3. Raccoon (raccoons, coatis, ringtails)
Order Carnivora, Family Procyoni dae. Procyonids ar e mediu m-size d om niv ores that range th roughout mu ch of Nor th Amer ica. Raccoons , coatis, and r ingt ails feed on insects, smal l mammal s, birds, lizards, and f ruit s. Ringt ails ar e much smalle r and m ore slender th an raccoons and c ons ume a highe r propor tion of anim al matter (Kaufman n, 1982). Coatis are slight ly smalle r th an racoons and a re lim ited in t heir dis tr ibut ion in t he United States to just n orth of th e Mexican border.
Selected species
The raccoon ( Procyo n lotor) is th e most abunda nt and w idespread mediu m-si zed omniv ore in t he Nor th Amer ica. They are found t hroughout Mexico, Central Amer ica, the United States, excep t at th e highe r elevat ions of th e Rocky Mount ains , and int o sout hern Canada (Kaufman n, 1982). During t he last 50 year s, raccoon popula tions in t he Unit ed States h ave increased greatly (Sanderson, 1987). In subur ban areas, they fr equently raid garbage cans and dum ps. Raccoons ar e preyed on by bobcats, coyotes, foxes, and great horned owls (Kaufman n, 1982). Twenty-fi ve su bspecies are recognize d in t he Unit ed States an d Canada; however , most r esearchers do not identi fy th e subspecies st udie d because differ ent su bspecies inha bit essen tially nonov erlapping ge ogr aphic r anges.
Body size. Raccoons measu re from 46 to 71 cm wi th a 20 to 30 cm tai l. Body weight s vary by loc ation, age, and sex fr om 3 to 9 kg (Kaufman n, 1982; Sanderson, 1987). The largest r accoons r ecorded are from Idaho and ne arby states, while th e smal lest r eside in t he Flor ida K eys (L otze and Anderson, 1979). Juveniles do not reach adult size unt il a t least th e end of th eir second year (Stuewer , 1943b). In the autumn, fat r eserves acc ount for 20 to 30 percent or more of th e raccoon' s weight (W hit ney and Unde rwood, 1952, ci ted in Kaufman n, 1982). In Minne sota, Mech et al. (1968) found t hat juv enile s ga ined weight almost l inearly unti l mid-Novemb er, after which they began to lose weight unt il April. Weight los s in a dult s and year lings can reach 50 percent during t he 4 mont hs of wint er dormancy (e.g., 4.3-kg loss fo r a 9.1-kg raccoon) (T hor kelson a nd Maxwel l, 1974; Mech et al., 1968). In Alabama, where raccoons ar e acti ve al l year , winter weight losses ar e less, 16 to 17 percent on average (Johns on, 1970).
Habitat . Raccoons ar e found ne ar vi rtua lly every aquatic habitat, particularly in hardwood swamp s, mangroves, floodpla in f orests, and freshwater and sal twater marshes (Kaufman n, 1982). They are also commo n in subur ban reside nt ial areas and cultivated and abandone d farmlands (K aufman n, 1982) and may fo rage in farmyar ds (Greenwood, 1982). Stuewer (1943a) stated that a permanent water supply , tree dens, and availa ble food a re essen tial. Raccoons use surface water s fo r both drinking a nd f oraging (Stuewer , 1943a).
Food ha bi ts . The raccoon is an omniv orous and oppor tunis tic feeder. Although primar ily acti ve fr om su nset to sunrise (Kaufman n, 1982; Stuewer , 1943a), raccoons will change their acti vi ty p eriod t o acco mmo date th e avai labilit y of food a nd water (Sanderson, 1987). For examp le, sal t mar sh raccoons may b ecome acti ve during t he day to take advantage of low ti de (Ivey, 1948, ci ted in Sanderson, 1987). Raccoons feed primar ily on fleshy fr uits, nuts, acorns, and corn (Kaufman n, 1982) but also eat grains, insects, frogs,
2-233 Racco on

crayfi sh, eggs, and vi rtually any animal and vegetable matter (Palmer and Fowler, 1975). The propor tion of di ffer ent foods in t heir die t depends on lo cation a nd seaso n, although plants are usually a more impor tant compone nt of th e die t. They may fo cus on a prefer red food, such as turtle eggs , when it is avai lable (Stuewer , 1943a). They also will feed on garbage and c arrion. Typic ally , it is only in t he spring a nd early su mmer that raccoons eat more animal than plant mater ial. Their late su mmer and fal l diets co nsist p rimar ily of fruit s. In wint er, acorns tend t o be th e most im por tant food, although r accoons will take any corn or fruit s that are still avai lable (Kaufman n, 1982; Stuewer , 1943a).
Temperature regulation and molt. From th e central United States i nto Canada, raccoons unde rgo a wint er dor mancy lasting up t o 4 mont hs (Stuewer , 1943a). It is not a true hibe rnation, however , and they can be easi ly awaken ed (Kaufman n, 1982). Animals in the south are acti ve year -round ( Goldm an, 1950). Snow co ver , more than low temp eratures, tr igge rs wint er dor mancy (Stuewer , 1943a; Mech et al ., 1966; Kaufman n, 1982). The raccoon' s annua l molt begins ear ly in s pring a nd l asts a bout 3 mont hs (Kaufman n, 1982).
Breeding acti vi ties and social organization. Although s olit ary, adult r accoons co me toge ther fo r a shor t time dur ing t he mating pe riod ( Kaufman n, 1982), which begins ear lier (Janua ry to March) in t heir nor thern range th an in t heir sout hern range (March to June ) (Johns on, 1970; Sanderson, 1987). Male and femal e home ranges over lap freely and each male may mate wi th sever al femal es during t he breeding seaso n (Mech et al ., 1966; Johns on, 1970; Kaufman n, 1982; Stuewer , 1943a). The most co mmo n group of r accoons is a mother and he r young of th at year . Fur ther nor th in t heir range, a family will de n together for the winter and break up the follow ing s pring ( Kaufman n, 1982). Males are ter ritorial toward one another but not to ward femal es; femal es are not ter ritorial (Fritzell, 1978).
Home range and r esour ces. The size of a raccoon' s hom e range depends on it s sex and age, habitat, food s our ces, and the seaso n (Sanderson, 1987). Values fr om a few hectar es to more than a few t housand hectar es have been reported, although hom e ranges of a few hundr ed hectar es appear to be most co mmo n (see Appendix ). In general, hom e ranges of mal es are larger than those of femal es, the home range of femal es wi th young is restr icted , and winter ranges are smal ler than ranges at o ther times o f th e year for both sexes (San derson, 1987). During t he wint er, raccoons co mmonly den in hollow tr ees; they also use th e burrows o f other animals su ch as fo xes, groundhogs , skunk s, and ba dgers. These si tes ar e used for sleeping dur ing w armer periods . After winter ing in one den, the femal e will c hoos e a new den in w hic h to bear her young ( Kaufman n, 1982). Schneider et al. (1971) found t hat onc e the cubs l eave th e den, the fami ly will not use it ag ain t hat year .
Popul ation de nsity . Popula tion de nsity depends on t he qualit y and qua nt ity of food resources an d den si tes. Values between 0.005 and 1.5 raccoons per hectar e have been repor ted, although 0 .1 to 0.2 per hectar e is mo re commo n (see Appendix ). Popula tions exceeding one r accoon pe r hectar e have been reported in residential areas (Hoffman and Gottsch ang, 1977). Although r accoons may p refer tree dens over ground de ns, partic ularly for rais ing y oung (Stuewer , 1943a), Butter field (1954) found high r accoon de nsit ies in a n area wi th few tr ee dens but numerous ground de ns.
2-234 Racco on

�
�
Popul ation dy namics. Males generally are not sexu ally matu re by th e time of th e fi rst regula r breeding seaso n follow ing t heir bir th, but th ey may matu re later that su mmer or fall ( Johns on, 1970; Sanderson, 1951). Females may b ecome pregnant in their fi rst year (Johns on, 1970). In a review of sever al studies, Kaufman n (1982) found t hat up t o 60 percent of bot h wild a nd c aptive femal es mate an d produce l itter s in their fi rst year . In Illinois and Mis sour i, Fritzell e t al. (1985) found pr egnancy rates o f year lings fr om 38 to 77 percent. After their fi rst year , almost al l femal es breed annua lly (F ritzell e t al., 1985). Females produc e only one li tter each year , and the femal e alone car es for th e young (Sanderson, 1987; Stuewer , 1943a, 1943b). With some excep tions (Bis sonn ette an d Csech , 1937), larger litter sizes usually occu r in the raccoon' s nor thern range (Lot ze and Anderson, 1979). Some juveniles of both sexes d isperse fr om th e areas wh ere they wer e bor n dur ing t he fall or wint er of th eir fi rst year , while others stay an d raise young w ithin their parents' home range (Stuewer , 1943a). The highest mo rtal ity r ates o ccu r wi thin t he fi rst 2 year s; the age str uctu re of popula tions in A labama suggests th at mo rtal ity i s highe r for suba dult s than for juv enile s (Johns on, 1970).
Simi lar species (fr om general references)
� The coati (Nasua nasua) is slight ly smalle r th an the raccoon ( 4 to 6 kg) but wi th a much longe r tail (51 to 64 cm). Ranging t hroughout Central Amer ica from Panama to Mexico (Kaufman n, 1982), the coati is rare in the Unit ed States wh ere it inha bit s ope n forests o f th e southwest, near the Mexican border. It fo rages primar ily fo r grubs and tubers but also feeds on fruits, nut s, bir d eggs , lizards, sco rpions , and t arantula s. Coatis roll a rthropods on the ground t o remove wings and scal es.
� The ringt ail (Bassar iscu s astu tus) is smal ler (36 to 41 cm; 0.9 to 1.13 kg) than the raccoon, wi th a tail e qual to its body lengt h. It ranges throughout th e southwester n United States i nto northern California and Oregon, inha bit ing chaparral, rocky r idges, and cl iffs n ear water . Ringt ails ar e omniv orous lik e the raccoon but cons ume a highe r propor tion of anim al matter , feeding mainly on smal l mammal s, insects, birds, and lizards as well a s fruit s. They den in caves o r crevices along c li ffs, hollow tr ees, unde r rocks, and in unus ed buildings . Although r ingt ails so meti mes l ive in colonie s, mated pairs are more common. More noc turnal than the raccoon, the ringt ail is only acti ve at d awn and dusk (K aufman n, 1982).
General references
Burt and Grossen heider (1980); Goldman (1950); Johns on (1970); Kaufman n (1982); Palmer and Fowler (1975); Sanderson (1987).
2-235 Racco on

Raccoon ( Proc yon lotor )
Factors Age/Sex/ Range Note Cond. /Seas. Mean or (95% CI) Locat ion Reference No.
Body Weight A M 7.6 7.0 - 8.3 wc Illinoi s Sander son, 1984 (kg) A F par ous 6.4 5.6 - 7.1
A F nullip arous 6.0 5.1 - 7.1 J M 5.1 4.6 - 5.7 J F 4.8 4.2 - 5.3
A M 6.76 Missour i Nagel , 1943 A F 5.74
A M 4.31 up t o 8.8 Alabama Johnson, 1970 A F 3.67 up t o 5.9
neonat e 0.075 w New York/capt ive Hamilton, 1936
Pup Growth bi rth to 7 days 17 w New York Hamilton, 1936 Rate (g/day) 8 to 19 days 21
20 to 30 days 11 31 to 40 days 12 41 to 50 days 23
bi rth to 6 wks 17.8 NS/lab Mont gom ery, 1969 6 to 9 wks 3.9 10 to 16 wks 29.5
Metabol ic Winter 9.36 ± 1.68 SD Washi ngt on, DC/National Mugaas et al., 1984 Rate 15-35��C Zoo (lO /kg-day)2
Metabol ic Rate J B 304 Ohio/lab Teubner & Bar rett, 1983 (kcal /kg-day)
A M basal 44.8 est imated 1 A F basal 46.8
A M free-living 183 (83 - 400) est imated 2 A F f ree-living 187 (85 - 408)
Food Ingest ion 3 Rate (g/g-day)
2-236 R
accoo
n

Raccoon ( Proc yon lotor )
Factors Age/Sex/ Range Note Cond. /Seas. Mean or (95% CI) Locat ion Reference No.
Water A M 0.082 est imated 4 Ingest ion Rat e A F 0.083 (g/g-day)
Inhal ation A M 2.47 est imated 5 Rate (m /day) A F 2.173
Surface Ar ea A M 3,796 est imated 6 (cm ) A F 3,4142
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
crayfish 37 8 aryland/forested Llewellyn & Uhler, 1952 snails 5 ottomland insect s 40 39 18 12 reptiles/amphibians 6 % wet volume; digestive fish 3 2 trace 2 tract) rodent s 7 2 trace 8 corn 0 9 Smilax 0 trace trace 6 acor ns 0 trace 5 17 pokeber ry 0 trace 17 2 wild cherry 0 17 2 0 blackber ries 0 16 trace 0 grapes 0 trace 23 8 persimmo n 0 0 11 7
M9 3 b6 3 5
(7 3 5
12 1
2-237 R
accoo
n

Raccoon ( Proc yon lotor )
2-238 R
accoo
n
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
frogs 8.1 trace 0 0 Tennessee/ NS Tabatabai & Kennedy , 1988 fish 1.2 0 0 0 bi rds trace 0 trace 8.4 (% wet vol ume; digest ive mammals 1.7 0 1.4 0 tract) other/unspeci fied 7.8 6.7 1.8 7.2 persimmon 0 35.8 57.3 27.4 cor n 57.6 0 10.0 25.9 grapes 0 trace 10.2 0 pokeber ry 0 20.5 4.5 0 acor ns 0 0 5.4 4.2 sugar hackber ry 0 0 5.5 18.4 cher ry 0 29.5 0 0 insect s 22.0 3.5 2.4 trace crayfish 1.6 4.0 1.5 1.4
Mollusca 44 sw Washi ngt on/tidewater Tyson, 1950 (mussel s and oy sters) mudf lats Crust acea (shr imp & 25 crabs) (% wet vol ume; stomach Pisces ( goby 9 cont ents) & cabez on) Annel ida 20 (marine worms) Echi ur ida (worm) 1
frui ts 37.9 New York/NS Hamilton, 1951 7 insect s 8.2 mammals 14.3 (% wet vol ume; stomach grains (e.g. cor n) 14.7 cont ents) earthworms 7.2 amphi bians 4.4 veget ation 6.1 reptiles 3.0 molluscs 1.9 bi rds 1.5 carrion 1.5 unspeci fied 0.2

Raccoon ( Proc yon lotor )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Home A M spr ./sum. 2,560 670 - 4,946 North Dakot a/prairie Fritzell, 1978 8 Range A F spr ./sum. 806 229 - 1,632 pot hol es Size (ha) A M May - Dec 204 18.2 - 814 Michi gan/riparian Stuewer, 1943a
A F May - Dec 108 5.3 - 376
A M all year 65 ± 18 SE Georgia/coast al island Lot ze, 1979 9 A F al l year 39 ± 16 SE
Popul ation NS 1.46 Ohio/resident ial woods Hoffman & Got tschang, 1977 Density (N/ha)
spr ing 0.17 Lake Er ie, Ohio/ Urban, 1970 10
spr ing 0.022 Wisconsi n/marsh Dorney, 1954 Sandusky Bay, marsh
Litte r 1 to 3 yrs 3.4 n Illin ois/NS Fritze ll e t al., 1985 Size 4 yrs + 3.8
2.43 Alabama/bot tomlands, Johnson, 1970 marsh
Li tters 1 most of range/NS Sander son, 1987 /Year
Days 63 North Am erica/NS Hamilton, 1936; Sander son, Gestation 1987; Stuewer, 1943b
Age at 84 63 - 112 NS/lab Mont gom ery, 1969 Weaning (days)
Age at M 15 mont hs Alabama/NS Johnson, 1970 Sexual Maturity F 1 year IL, MO/NS Fritze ll e t al., 1985
Annual A B 56 Missour i/NS Sander son, 1951 11 Morta lity Rates A B 38 sw Iowa/agricultu ral Clark et al., 1989 (percent ) J B 42
2-239 R
accoo
n

123456789
101112
Raccoon ( Proc yon lotor )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Longevi ty A B 3.1 years Alabama/NS Johnson, 1970 A B 1.8 years Missour i/NS Sander son, 1951 11
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating February March August sw Geor gia, nw Fl or ida McKeever , 1958 Januar y February March n Uni ted States Johnson, 1970
Partur ition Apr il early Apr il May Michi gan Stuewer, 1943b Apr il May October sw Georgia, nw Flor ida McKeever , 1958
Molt sum mer nor thern latitudes Goldman, 1950
Torpor late March/Apr il ec Minnesot a Whitney & Under wood, 1952 12 Novem ber
Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom Nagel (1943). Est imated usi ng equat ion 3-45 (Nagy, 1987) and body weight s f rom Nagel (1943).
2-240 R
accoo
n
See Chapt ers 3 and 4 f or methods f or calcul ating f ood i ngest ion r ates f rom free-living m etabol ic rate and di et.Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Nagel (1943).Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Nagel (1943).Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Nagel (1943).Col lect ions f rom Apr il through Oct ober .Measur ed f rom Apr il through Jul y.Based on r adiotracki ng.Aver age of three methods of est imating densi ty.Hunt ed popul ation.Cited in Schnei der et al. (1971).

References (including A ppendix)
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Arthur, S. C. (1928) The fur animals of Louis iana. Louis iana Dept . Cons erv.; Bull. No. 18.
Asdell, S. A. (1964) Patter ns of mammal ian reproduc tion. Ithaca, NY: Comsto ck Publ. Co.
Bailey, V. (1936) The mammal s and life zones of Or egon. U.S. Dept . Agr., Bur. Biol. Survey, North Am. Fauna 55: 1-416.
Bisso nnette, T. H.; Csech , A. G. (1937) Modif ication of mammal ian sexu al cycl es. Part 7, fer tile matings of r accoons in Decemb er instead of February induc ed by inc reasing daily periods of light . Proc. R. Soc. London, ser . B 827: 246-254.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Brown, C. E. (1936) Rearing w ild a nim als in c aptivi ty, and g estat ion pe riods . J. Mammal . 17: 10-13.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Butter field, R. T. (1944) Popula tions , hunt ing pr essu re, and movemen t of Ohio r accoons . Trans. Nor th Am. Wildl. Conf . 9: 337-344.
Butter field, R. T. (1954) Some raccoon a nd gr oundhog r elations hips . J. Wildl. Manage. 18: 433-437.
Cagle, F. R. (1949) Notes o n the raccoon, Procyon l otor megalodous Lowery. J. Mammal . 30: 45-47.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Cauley, D. L.; Schinne r, J. R. (1973) The Cinc inna ti raccoons . Nat. His t. 82: 58-60.
Clark, W. R.; Hasbrouc k, J. J. ; Kienzler, J. M.; et al. (1989) Vital stati sti cs an d harvest o f an Iowa raccoon popula tion. J. Wildl. Manage. 53: 982-990.
Cowan, W. F. (1973) Ecology and lif e his tory of th e raccoon ( Procyo n lotor hirt us Nelson and Goldman) in the northern part of i ts r ange [Ph.D. disser tat ion] . Grand For ks, ND: Universi ty o f North Dakota.
2-241 Racco on

Cunningha m, E. R. (1962) A stu dy of th e easter n raccoon, Procyo n lotor (L.), on t he Atom ic Energy Commission Savannah River Plant [master 's th esis]. Athens, GA: Universi ty of Geo rgia .
Dew, R. D. (1978) Biology of th e raccoon, Procyo n lotor; I. Genic var iation. II. Popula tion age str uctu re and average litter size [master 's th esis]. Memphis , TN: Memphis State Universi ty.
Dorney, R. S. (1954) Ecology of mar sh raccoons . J. Wildl. Manage. 18: 217-225.
Eisenberg, J. F. (1981) The mammal ian radiations ; an analysi s of tr ends in evolut ion, adaptation, and be havior . Chic ago, IL: Univ ersi ty of Chic ago Press.
Ewer , R. F. (1973) The carnivores. Ithaca, NY: Cornell Universi ty Pr ess.
Flower , S. S. (1931) Cont ribut ions to our know ledge of th e dur ation of lif e in v ertebrate animals. V. Mammal s. Zool. Soc. London. Proc. (part 1): 145-234.
Fritzell, E. K. (1978) Habitat u se by prairie raccoons dur ing t he water fowl breeding seaso n. J. Wildl. Manage. 42: 118-127.
Fritzell, E. K.; Hubert, G. F., Jr .; Meyen, B. E.; et al . (1985) Age-speci fic reproduc tion in Illinois and Mis sour i raccoons . J. Wildl. Manage 49: 901-905.
Goldman, E. A. (1950) Raccoons of Nor th and Middle A mer ica. Washingt on, DC: U.S. Fish Wildl. Serv.; Nor th Am. Fauna 60.
Greenwood, R. J. (1982) Noctu rnal acti vi ty an d foraging of prairie raccoons ( Procyo n lotor) in Nor th Dakota. Am. Midl. Nat. 107: 238-243.
Hamilton, W. J., Jr . (1936) The food a nd br eeding ha bit s of th e raccoon. Ohio J . Sci . 36: 131-140.
Hamilton, W. J., Jr . (1940) The summer food of mink s and r accoons on t he Mont ezuma Marsh, New York. J. Wildl. Manage. 4: 80-84.
Hamilton, W. J., Jr . (1951) Warm weath er foods of th e raccoon in Ne w York state. J. Mammal . 32: 341-344.
Hoffman , C. O.; Gottsch ang, J. L. (1977) Numbers, distr ibut ion, and m ovemen ts o f a raccoon popula tion in a subur ban reside nt ial communit y. J. Mammal . 58: 623-636.
Ivey, R. D. (1948) The raccoon in t he sal t marshes of nor theaster n Flor ida. J. Mammal . 29: 290-291.
Johns on, A. S. (1970) Biology of th e raccoon ( Procyo n lotor var ius Nelson and Goldm an) in Alabama. Alabama Coope rative Wildlilf e Research Unit ; Aubur n Univ . Agric. Exp . Stn. Bull. 402.
2-242 Racco on

Kaufman n, J. H. (1982) Raccoon a nd a llie s. In: Chapman, J. A.; Feldh amer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 567-585.
Llewelly n, L. M.; Uhler, F. M. (1952) The foods of fu r anim als of th e Patuxent Research Refuge , Maryland. Am. Midl. Nat. 48: 193-203.
Lotze, J.-H. (1979) The raccoon ( Procyo n lotor) on St. Catherines Island, Georgia. 4. Comparisons of home ranges determined by l ivetr apping a nd r adiot racking. New York, NY: Amer ican Museum of Natural Histo ry; Rep. No. 2664.
Lotze, J.-H.; Anderson, S. (1979) Procyo n lotor. Amer ican Society o f Mammalogi sts; Mammal ian Species No. 119.
Lowery, G. H., Jr . (1936) A preliminary report on the distr ibut ion of th e mammal s of Louis iana. Louis iana Acad. Sci . Proc. 3: 1-39.
McComb, W. C. (1981) Effects o f land use upon f ood ha bit s, produc tivi ty, an d gastr ointesti nal parasi tes o f raccoons . In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1, Augus t 1980; Frostb urg, MD; pp. 642-651.
McKeever , S. (1958) Reproduc tion in t he raccoon in t he sout heaster n United States. J. Wildl. Manage. 22: 211.
Mech, L. D.; Tester , J. R.; Warner, D. W. (1966) Fall dayti me rest ing ha bit s of r accoons as determined by tel emetr y. J. Mammal . 47: 450-466.
Mech, L. D.; Barnes, D. M.; Tester , J. R. (1968) Seasonal weight ch anges, mo rtalit y, and popula tion s tructure of r accoons in Minne sota. J. Mammal . 49: 63-73.
Mont gom ery, G. G. (1969) Weaning of cap tive raccoons . J. Wildl. Manage. 33: 154-159.
Moor e, D. W.; Kennedy, M. L. (1985) Weight ch anges and popula tion s tructure of r accoons in wester n Tennessee. J. Wildl. Manage. 49: 906-909.
Mugaas, J. N.; Mahlke, K. P.; Broudy , E.; et al. (1984) Metabolis m of r accoons , Procyo n lotor, in winter and summer (abstr act only ). Am. Zool. 24: 89A.
Nagel, W. O. (1943) How big is a 'coon. Missour i Cons ervationis t 6-7.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Mono. 57: 111-128.
Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
2-243 Racco on

Sanderson, G. C. (1951) Breeding ha bit s and a his tory of th e Missour i raccoon popula tion from 1941 to 1948. Trans. North Am. Wildl. Conf . 16: 445-461.
Sanderson, G. C. (1984) Coope rative raccoon c oll ections . Ill. Nat. His t. Survey Div.; Pittman -Robertson P roj. W-49-R-31.
Sanderson, G. C. (1987) Raccoon. In: Novak, M.; Baker , J. A.; Obbarel, M. E.; et al ., eds. Wild f urbearer man agement and conservat ion. Pittsb urgh, PA: Universi ty o f Pittsb urgh Press; pp. 487-499.
Sanderson, G. C.; Hubert, G. F. (1981) Selected demographic ch aracter isti cs o f Illinois (U.S.A.) raccoons ( Procyo n lotor). In: Chapman, J. A.; Pursley, D., eds. Worldwide furbearer conf erence pr oceedings : v. 1. Augus t 1980; Frostb urg, Maryland.
Sanderson, G. C.; Nalbandov , A. V. (1973) The reproducti ve cycl e of th e raccoon in Illinois . Illinois Nat. His t. Surv. Bull. 31: 29-85.
Schneider, D. G.; Mech, D. L.; Tester , J. R. (1971) Movemen ts o f femal e raccoons and t heir young a s determined by radio- tracking. Anim . Behav. Monogr . 4: 1-43.
Schoonov er, L. J.; Marshall, W. H. (1951) Food ha bit s of th e raccoon ( Procyo n lotor hirt us) in north-central Minne sota. J. Mammal . 32: 422-428.
Seton, E. T. (1929) Lives o f game animals. Garden City, NJ: Double day, Doran and Company.
Sherfy, F. C.; Chapman, J. A. (1980) Seasonal home range and habitat u tiliza tion of raccoons in Maryland. Carniv ore 3: 8-18.
Slate, D. (1980) A stu dy of New Jer sey r accoon popula tion s--determination of th e densit ies, dynamics an d incidence of disease i n raccoon popula tions in New Jersey. N.J. Div. Fish, Game, and Wildl. ; Pittman -Robertson P roj. W-52-R-8, Fina l Rep.
Sonenshine , D. E.; Wins low , E. L. (1972) Cont rasts i n distr ibut ion of r accoons in t wo Virginia loc alit ies. J. Wildl. Manage. 36: 838-847.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stains, H. J. (1956) The raccoon in Ka nsas: natural histo ry, management, and econom ic impor tance. Univ . Kansas Mus. Nat. Hist. , Misc. Publ. 10: 1-76.
Stuewer , F. W. (1943a) Raccoons : their habit s and m anagement in Michiga n. Ecol. Monogr . 13: 203-257.
Stuewer , F. W. (1943b) Reproduc tion of r accoons in Mic higa n. J. Wildl. Manage. 7: 60-73.
2-244 Racco on

Tabatabai, F. R.; Kennedy, M. L. (1988) Food ha bit s of th e raccoon ( Procyo n lotor) in Tennessee. J. Tenn. Acad. Sci . 63: 89-94.
Tester , J. R. (1953) Fall food ha bit s of th e raccoon in t he Sout h Platte Val ley of northeaster n Color ado. J. Mammal . 34: 500-502.
Teubne r, V. A.; Barrett, G. W. (1983) Bioe nergetics of cap tive raccoons . J. Wildl. Manage. 47: 272-274.
Thorkelson, J.; Maxwel l, R. K. (1974) Design a nd testing of a heat tr ansfer model of a raccoon (Procyo n lotor) in a clos ed tree den. Ecology 55: 29-39.
Tyso n, E. L. (1950) Summer food ha bit s of th e raccoon in s out hwest W ashingt on. J. Mammal . 31: 448-449.
Urban, D. (1970) Raccoon popula tions , movemen t patter ns, and predation on a man aged water fowl marsh. J. Wildl. Manage. 34: 372-382.
VanDruff, L. W. (1971) The ecology of th e raccoon a nd opos sum, wi th empha sis on t heir role as water fowl nest p redators [Ph .D. disser tat ion] . Ithaca, NY: Cornell Universi ty.
Whit ney, L. F.; Underwood, A. B. (1952) The raccoon. Orange, CT: Practi cal Science Publ.
Wood, J. E. (1954) Food ha bit s of fu rbearers of th e upla nd pos t oak region in T exas. J. Mammal . 35: 406-415.
Yeager, L. E.; Rennels, R. G. (1943) Fur yield and autumn foods of th e raccoon in Illinois river bot tom lands . J. Wildl. Manage. 7: 45-60.
2-245 Racco on

2.2.4. Mink (m ink, w easels, erm ine)
Order Carnivora, Family Mustel idae. Although v aried in s ize, most memb ers of th is fami ly have long, slender bodie s and s hor t legs. Throughout th e fami ly, the male is usually larger than the femal e. The more ter restr ial species feed primar ily on smal l mammal s and bir ds. Mustelids th at liv e around l akes an d str eams feed on aquatic prey su ch as fi sh, frogs, and inver tebrates (B urt and Grossen heider, 1980).
Selected species
The mink (Mustel a vison) is th e most abunda nt and w idespread carniv orous mammal in North Amer ica. Mink ar e dis tr ibut ed throughout Nor th Amer ica, excep t in the extr eme north of Canada, Mexico, and arid areas of th e southwester n United States. It is common t hroughout its range but o ften over looked because of i ts s olitary nature and noctu rnal acti vi ty. Mink ar e partic ularly sensi tive to PCBs and s imila r ch emicals, and ha ve been found t o accu mulate PCBs in subcutaneous fat to 38 to 200 times d ietary conc entrations , depending on t he PCB conge ner (Hor nshaw et al., 1983).
Body size. Body size var ies gr eatly th roughout th e species' range, wi th males weighing m arkedly mo re than femal es (in some popula tions , almost twi ce as much, see table). Males measu re from 33 to 43 cm wi th a 18 to 23 cm tai l. Females measu re from 30 to 36 cm wi th a 13 to 20 cm tai l (Burt and Grossen heider, 1980). Farm-raised mink ten d to be larger th an wild m ink (l etter from R.J. Aulerich, Departmen t of Animal Science, Michiga n State Universi ty, East L ansing, MI, to Susan Nor ton, Janua ry 7, 1992).
Metabolism. Harper et al . (1978) evaluated the energy requir ements o f growing farm-raised male mink dur ing a 21-day period w hen about 20 pe rcent of th eir to tal growth would oc cur. They expressed food in take o n the basis of meta bolic body size (MBS ) instead of body weight (B W) where MBS = BW(kg)0.73. Metaboliza ble en ergy (ME) requir ements wer e 147.8 ± 6.06 (kcal /kgMBS-day). Account ing f or assi mi lation effi ciency, this co rresponde d to a gross energy (GE) intake o f approximatel y 203 (kcal /kgMBS-day).
Iversen (1972) found t hat basal metabolic r ate fo r mink and other muste lids weighing 1 kg or more could be exp ressed by th e equation:
BMR = 84.6Wt0.78 (±0.15),
where BMR = basal metabolic r ate in kcal /day and Wt = body weight in k ilogr ams. This model reflects th e finding t hat the larger mu stelids h ave a s light ly (10 to 15 percent) highe r basal metabolic r ate th an expected for mammal s in general.f Free-living metabolic r ates would be exp ected to be three to five ti mes highe r (see tab le).
Habitat . Mink ar e found asso ciated wi th aquatic habitats o f al l kinds , inc luding water ways su ch as r ivers, str eams, lakes, and ditch es, as wel l as swamp s, marshes, and
fMustel id species much smaller th an 1 kg (i.e., the sto at and weasel) have much higher basal metabolic rates than predic ted for mammals in general.
2-247 Mink

backwater areas (Linsco mbe et al ., 1982). Mink prefer irregula r shor eline s to more open, exposed banks (A llen, 1986). They also tend to use brushy or woode d cover adj acent to the water , where cover for prey is abunda nt and w here dow nfall and de br is prov ide den si tes (A llen, 1986).
Food ha bi ts . Mink ar e predom inant ly noc turnal hunt ers, al though t hey are someti mes acti ve during t he day. Shor eline s and emergent veg etat ion a re the mink 's pr inc ipa l hunt ing a reas (Arnold, 1986, ci ted in Eagle and Whitman , 1987). Mink ar e oppor tunis tic feeders, tak ing w hatever prey is abunda nt (Hamilt on, 1936, 1940; Erringt on, 1954; Sargeant et al ., 1973). Mammal s are the mink's mo st i mportant prey year -round in many parts o f th eir range (Eagle and Whitman , 1987), but mi nk also hunt aqua tic pr ey such as fish, amphibia ns, and c rustacean s and other ter restr ial prey su ch as bird, repti les, and insects, depending on t he seaso n (Linsco mbe et al ., 1982). In marsh habitats i n summer , muskr ats can be an important food s our ce depending on t heir popula tion de nsity and vulne rabilit y (e.g., health) (Hamilt on, 1940; Sealander, 1943; Erringt on, 1954). Mink die t also can depend on marsh water level ; Proulx et al. (1987) found t hat wi th high water levels, mink cap tured predominantly cr ayfi sh and meadow vo les, but during pe riods of low water , the mink preyed on aquatic birds and muskr ats d eeper in the marsh. Simi larly, Erringt on (1939) found t hat mink predation on m uskr ats i n the prairie pothole r egion c an increase dramati cal ly in times o f drought as the muskr at bur rows be come more expos ed. Also in t his r egion, duc klings and m olt ing a dult ducks th at fr equent sh orelines are partic ularly vulne rable to mink predation ( Arnold a nd Fr itzell, 1987; Sargeant et al ., 1973). In winter , mink often supplemen t th eir diet wi th fish (Eagle and Whitman , 1987). Females tend t o be lim ited to smalle r prey than males, who a re able to hunt larger prey such as rabbits an d muskr ats mo re successf ully (Bir ks and Duns tone , 1985; Sealander, 1943).
Temperature regulation and molt. In wint er, mink do not unde rgo hibe rnation or torpor; instead , they are acti ve year -round. Mink r eplace th eir summer coat in mid to late fall w ith a darker mo re dense coat and m olt ag ain in t he spring (Eagle and Whitman , 1987; Linsco mbe et al ., 1982).
Breeding acti vi ties and social organization. Mating oc curs in l ate wi nter to early spring (Eagle and Whitman , 1987). Variation in t he lengt h of mating seaso n with differ ent subspecies reflects ad aptat ions to di ffer ent cl imates ( Linsco mbe et al ., 1982). Ovulation is induc ed by mating, and im plantation is delayed (Eagle and Whitman , 1987). Parturition generally occu rs in the late sp ring, and t he mink ki ts are altr ic ial (helpl ess) at b irth (Linsco mbe et al ., 1982). Mink ar e generally solit ary, wi th femal es only asso ciating w ith their young of th e year . Female home ranges generally do not over lap wi th the home ranges of other femal es, nor do the home ranges of mal es over lap wi th each other (Eagle and Whitman , 1987). The home range of a mal e may o ver lap the home range of sever al femal es, however , particularly during t he breeding seaso n (Eagle and Whitman , 1987).
Home range and r esour ces. The home range of mi nk encompasses b oth their foraging a reas around water ways an d their dens. When denning, mink use bank bur rows of other animals, particularly mu skr ats, as wel l as cavi ties in tree roots, rock o r brush pile s, logj ams, and beaver lodge s (Melquis t et al., 1981; Birks an d Linn, 1982; Eagle and Whitman , 1987). Indiv idua l mink may u se sever al differ ent dens wi thin t heir hom e range, males mo re so than femal es (Birks an d Linn, 1982). Melquis t et al. (1981) found t hat den
2-248 Mink

�
�
�
�
si tes i n Idaho were 5 to 100 m fr om th e water , and they never observed mink mo re than 200 m fr om water . The shape of mi nk home ranges depends on habitat typ e; river ine home ranges are basical ly l inear, whereas th ose in marsh habitats ten d to be more ci rcular (Birks and Linn, 1982; Eagle and Whitman , 1987). Home range size depends mo stl y on food abunda nce, but al so on t he age and sex o f th e mink, seaso n, and social stabilit y (Arnold, 1986; Birks an d Linn, 1982; Eagle and Whitman , 1987; Linn a nd Bir ks, 1981; Mitch ell, 1961). In wint er, mink sp end m ore time near dens and us e a smalle r por tion of th eir hom e range than in summer (Gerell, 1970, ci ted in Linsco mbe et al ., 1982). Adult mal e hom e ranges are generally larger th an adult femal e home ranges (Eagle and Whitman , 1987), particularly dur ing t he mating seaso n when males may r ange over 1,000 ha (Arnold, 1986).
Popul ation de nsity . Popula tion de nsity depends on avai lable cover and prey. Popula tion de nsit ies typic ally r ange fr om 0.01 to 0.10 mink per hectar e (see tab le). In river ine en vironm ents, it can be mo re meaningf ul t o measu re densi ties in terms o f number of mink per unit lengt h of shor eline co vered rather th an in t erms of num ber per hectar e.
Popul ation dy namics. Mink r each sexu al matu rity at 10 mo nths to a year and may reproduce fo r 7 years, possibly mo re (Enders, 1952; Ewer , 1973). Female mink can reproduce once per year and usually give birth to their fi rst l itter s at th e age of 1 year (Eagle and Whitman , 1987). Females often live to the age of 7 year s in captivi ty (En ders, 1952).
Simi lar species (fr om general references)
� The long -tai led weasel (Mustel a frenata) is smal ler (males 23 to 27 cm, 200 to 340 g; femal es 20 to 23 cm, 85 to 200 g) than the mink. It is co nsidered beneficial in agriculture because i t k ills small r odents, but it does not harm poult ry. Although it does not r ange as far nor th as the mink , the long -tai led weasel does inha bit parts of th e sout hwest.
� The least weasel (Mustel a nival is) is smal ler than the mink (mal es 15 to 17 cm, 39 to 63 g; femal es 14 to 15 cm, 38 to 40 g) and inha bit s meadows, fields, and w oode d areas. The least weasel feeds exten sively on mice and insects. Its h abitat i s l imi ted to the north central United States an d Canada.
� The ermine (Mustel a erminea), or shorttai l weasel , is smal ler (males 15 to 17 cm, 71 to 170 g; femal es 13 to 19 cm, 28 to 85 g) than the mink. The ermine inha bit s woody ar eas near water and feeds primar ily on smal l mammal s. The ermine's range is l imi ted to the northern and wester n United States an d Canada.
� The black-fo oted fer ret (Mustel a nigripes) is larger (36 to 46 cm; up to 1.1 kg) than the mink and inha bit s wester n prairies in the United States, although it now is an endangered species. It feed s on prairie dogs and ot her smal l animals.
2-249 Mink

General references
Burt and Grossen heider (1980); Eagle and Whitman (1987); Hall (1981); Linsco mbe et al . (1982); Palmer and Fowler (1975).
2-250 Mink

Mink ( Mustela vison )
2-251 M
ink
Range or
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion erence No.
Weight A M < 2,300 western races Harding, 1934 1 (g) A M < 1,400 eastern races Harding, 1934 1
A M spr ing 1,734 ± 350 SD Michi gan (farm-raised) Hornshaw et al., 1983 A F spr ing 974 ± 202 SD
A M summer 1,040 Montana Mitc hell, 1961 J M sum mer 777 A M fall 1,233 J M fall 952
A F summer 550 Montana Mitc hell, 1961 J F sum mer 533 A F fall 586 J F fall 582
neonat e 6 - 10 NS Eagle & Whitman, 1987 neonate 8.3 ± 1.54 SD Michigan (farm-raised) Hornshaw et al., 1983
Pup Gr owt h 0-30 days; M 7.0 NS/(farm-raised) Wehr et al. (unpubl ished) 2 Rate (g/day) 31-90 d; M 21
91-120 d; M 15 121-150 d; M 9.0 151-180 d; M 4.3
0-30 days; F 6.5 31-90 d; F 13 91-120 d; F 6.7 121-150 d; F 1.7 151-180 d; F 0.6
Ref
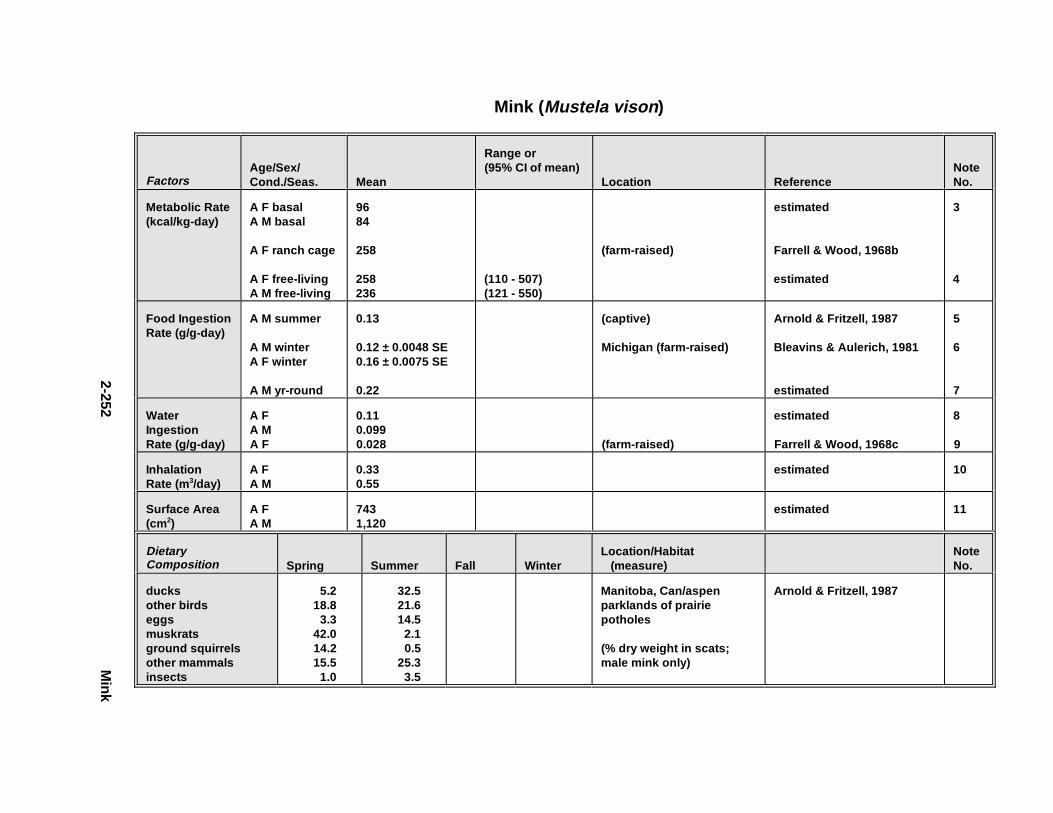
Mink ( Mustela vison )
Range or
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion erence No.
Metabol ic Rate A F basal 96 est imated 3 (kcal /kg-day) A M basal 84
A F ranch cage 258 (farm-raised) Farrell & Wood, 1968b
A F f ree-living 258 (110 - 507) est imated 4 A M free-living 236 (121 - 550)
Food Ingest ion A M sum mer 0.13 (capt ive) Arnol d & Fr itzell, 1987 5 Rate (g/g-day)
A M winter 0.12 ± 0.0048 SE Michi gan (farm-raised) Bleavins & Aul erich, 1981 6 A F w inter 0.16 ± 0.0075 SE
A M yr-round 0.22 est imated 7
Water A F 0.11 est imated 8 Ingest ion A M 0.099 Rate (g/g-day) A F 0.028 (farm-raised) Farrell & Wood, 1968c 9
Inhal ation A F 0.33 est imated 10 Rate (m /day) A M 0.553
Surface Ar ea A F 743 est imated 11 (cm ) A M 1,1202
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
ducks 5.2 32.5 Manito ba, Can/aspen Arnold & Fritze ll, 1987 other bi rds 18.8 21.6 parklands of prairie eggs 3.3 14.5 pot hol es muskr ats 42.0 2.1 ground squi rrels 14.2 0.5 (% dry weight in scat s; other mammals 15.5 25.3 male mink onl y) insect s 1.0 3.5
Ref
2-252 M
ink

Mink ( Mustela vison )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
(habi tat/season) (stream; year-round) (river; year-round) Michi gan/stream, river Alexander , 1977 trout 52 56 non- trout fish 6 26 (% wet weight ; stomach uni dent ified f ish 3 3 cont ents) crust aceans 11 4 amphi bians 2 3 birds/mammals 5 6 veget ation 17 1 uni dent ified 4 1
(sex of mink) (M) (F) Michigan/NS Sealander , 1943 12 muskr at 43 14 cot tont ail 16 12 (% vol ume; stomach small mammals 5 17 contents) large birds 18 11 small bi rds trace snakes 2 2 frogs 10 37 fis h 5 4 crayfish 1 3
frogs 24.9 Missour i/statewide Kor schgen, 1958 mice & r ats 23.9 fish 19.9 rabbi ts 10.2 (% dry vol ume; stomach crayfish 9.3 cont ents) bi rds 5.6 fox squi rrels 2.2 muskr ats 1.3 other 2.7
2-253 M
ink

Mink ( Mustela vison )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home Range 259 - 380 ha North Dakot a/prairie Eagle (unpubl ished) 13 Size pot hol es
A M 770 ha Mani toba, Canada/prairie Arnol d & Fr itzell, 1987
A F 7.8 ha heavy veget ation A F 20.4 ha spar se veget ation
A M 2.63 km 1.8 - 5.0 km Sweden/stream Gerell, 1970 1 J M 1.23 km 1.1 - 1.4 km A F 1.85 km 1.0 - 2.8 km
pot hol es Montana/riv erine: Mitchell, 1961
Popul ation 0.03 - 0.085 N/ha Mont ana/river Mitchell, 1961 Densi ty
A F w inter 0.006 N/ha Michi gan/river Marshall, 1936 14 A F w inter 0.6 N/km river
Litte r 4.2 Michigan/(farm-raised) Hornshaw et al., 1983 Size 4 2 - 8 Montana/riv er Mitchell, 1961
4 - 10 North Am erica/NS Hall & Kel son, 1959
Li tters 1 North America/NS Hall & Kel son, 1959 /Year
Days 39 - 76 North Am erica/NS Hall & Kel son, 1959 Gestation 51 40 - 75 Uni ted States/(farm-raised) Ender s, 1952
Age at eat meat 37 days Loui siana/NS Svi lha, 1931 14 Weaning fully
hom eothermic 7 weeks NS/NS Kost ron & Kukl a, 1970 14
Age at B 10 mont hs Uni ted States/(farm-raised) Ender s, 1952 Sexual B 1 year NS/NS Ewer, 1973 15 Maturity
Longevi ty maximum 10 years NS/zoo Eisenber g, 1981 F 7 maximum 11 years NS/(farm-raised) Ender s, 1952
Range
2-254 M
ink
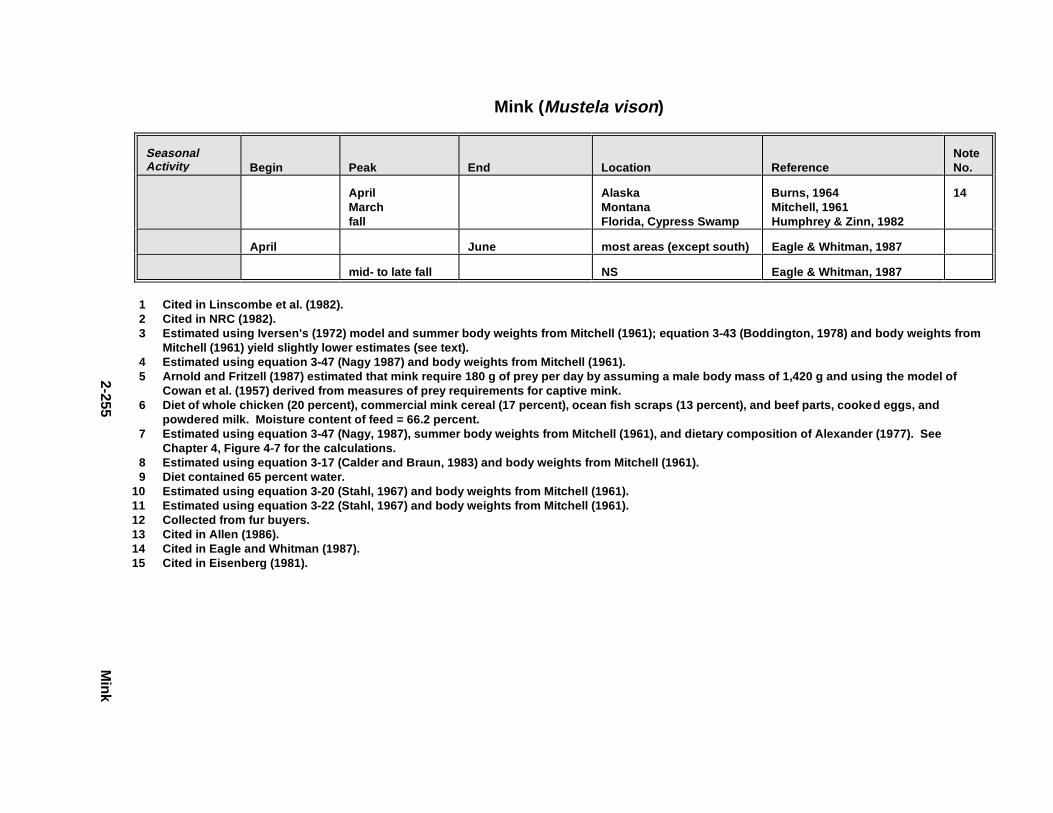
Mink ( Mustela vison )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating Apr il Alaska Bur ns, 1964 14 March Montana Mitc hell, 1961 fall Flor ida, Cypress Sw amp Humphr ey & Zinn, 1982
Partur ition Apr il June most areas (except sout h) Eagle & Whitman, 1987
Molt mid- to late fall NS Eagle & Whitman, 1987
1 Cited in Linscom be et al. (1982).2 Cited in NRC (1982).3 Est imated usi ng Iversen' s (1972) model and sum mer body weight s f rom Mitchel l (1961); equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom
Mitchel l (1961) yield sl ight ly lower est imates (see text). 4 Est imated usi ng equat ion 3-47 (Nagy 1987) and body weight s f rom Mitchel l (1961). 5 Arnol d and Fr itzell (1987) est imated that mink r equi re 180 g of prey per day by assum ing a m ale body mass of 1,420 g and usi ng the model of
Cowan et al. (1957) der ived f rom measur es of prey requi rements for capt ive mink. 6 Diet of whol e chi cken ( 20 percent ), com mercial mink cer eal (17 percent ), ocean f ish scr aps (13 percent ), and beef parts, cooke d eggs, and
powder ed milk. Moisture cont ent of feed = 66.2 percent . 7 Est imated usi ng equat ion 3-47 (Nagy, 1987), sum mer body weight s f rom Mitchel l (1961), and di etary com posi tion of Alexander (1977). See
Chapt er 4, Figur e 4-7 for the calcul ations. 8 Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Mitchel l (1961). 9 Diet cont ained 65 per cent water.
10 Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Mitchel l (1961).11 Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Mitchel l (1961).12 Col lected f rom fur buy ers.13 Cited in Al len (1986).14 Cited in Eagle and Whitman (1987).15 Cited in Eisenber g (1981).
2-255 M
ink

References (including A ppendix)
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Allen, A. W. (1986) Habitat su itabilit y inde x mode ls : mink . U.S. Fish Wildl. Serv. Biol. Rep. 82(10.127).
Arnold, T. W. (1986) The ecology of prairie mink dur ing t he water fowl breeding seaso n [master 's th esis]. Colum bia, MO: Univ ersi ty of Missour i.
Arnold, T. W.; Fritzell, E. K. (1987) Food ha bit s of prairie mink dur ing t he water fowl breeding seaso n. Can. J. Zool. 65: 2322-2324.
Birks, J. D.; Dunsto ne, N. (1985) Sex-related differ ences i n the diet of th e mink Mustel a vison. Holarcti c Ecol. 8: 245-252.
Bir ks, J. D.; Linn, I. J. (1982) Studies of home range of th e feral mink, Mustel a vison. Symp . Zool. Soc. Lond. 49: 231-257.
Bleavins, M. R.; Aulerich, R. J. (1981) Feed consumption a nd f ood p assag e in mink (Mustel a vison) and European fer rets (Mustel a putorius fu ro). Lab. Anim. Sci . 31: 268-269.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Burgess, S. A.; Bide r, J. R. (1980) Effects o f str eam habitat i mprovemen ts o n inver tebrates, trout popula tions , and m ink acti vi ty. J. Wildl. Manage. 44: 871-880.
Burns, J. J. (1964) The ecology , econom ics, and m anagement of mi nk in the Yukon-Kusko kwi m Delta, Alaska [master 's th esis]. Anchorage, AK: Universi ty o f Alaska.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chanin, P. R.; Linn, I. (1980) The diet of th e feral mink (Mustel a vison) in southwest B ritain. J. Zool. (London) 192: 205-223.
Cowan, I. M.; Wood, A. J.; Ki tts, W. D. (1957) Feed requir ements o f deer, beaver , bear, and mink fo r growth and m aint enance. Trans. Nor th Am. Wildl. Conf . 22: 179-188.
2-256 Mink

Cowan, W. F.; Reilly , J. R. (1973) Summer and fal l foods of mink on t he J. Clark Salyer Nationa l Wildlif e Refuge . Prairie Nat. 5: 20-24.
Eagle, T. C.; Whitman , J. S. (1987) Mink . In: Novak, M.; Baker , J. A.; Obbarel, M. E.; et al ., eds. Wild f urbearer man agement and conservat ion. Pittsb urgh, PA: Universi ty o f Pittsb urgh Press; pp. 615-624.
Eberhardt, L. E. (1974) Food ha bit s of prairie mink ( Mustel a vison) dur ing t he water fowl breeding seaso n [master 's th esis]. St. Paul, MN: Univ ersi ty of Minne sota.
Eisenberg, J. F. (1981) The mammal ian radiations . Chic ago, IL: Univ ersi ty of Chic ago Press.
Enders, R. K. (1952) Reproduc tion of th e mink ( Mustel a vison). Proc. Am. Philos . Soc. 96: 691-755.
Erringt on, P. L. (1939) React ions of mu skr at popula tions to drought . Ecology 20: 168-186.
Erringt on, P. L. (1954) The special respons iveness o f mi nks to epizoot ics in m uskr at popula tions . Ecol. Monogr . 24: 377-393.
Ewer , R. F. (1973) The carnivores. Ithaca, NY: Cornell Universi ty Pr ess.
Farrell, D. J.; Wood, A. J. (1968a) The nutr ition of th e femal e mink (Mustel a vison). I. The metabolic r ate of th e mink. Can. J. Zool. 46: 41-46.
Farrell, D. J.; Wood, A. J. (1968b) The nutr ition of th e femal e mink (Mustel a vison). II. The energy r equir ement fo r maintenance. Can. J. Zool. 46: 47-52.
Farrell, D. J.; Wood, A. J. (1968c) The nutr ition of th e femal e mink (Mustel a vison). III. The water requir ement fo r maintenance. Can. J. Zool. 46: 53-56.
Gerell, R. (1970) Home ranges and movemen ts o f th e mink Mustel a vison Schreber in southern Sweden. Oikos 20: 451-460.
Gilbe rt, F. F.; Nanceki vel l, E. G. (1982) Food ha bit s of mink ( Mustel a vison) and otter (Lut ra canadensis) in northeaster n Alberta. Can. J. Zool. 60: 1282-1288.
Guilda y, J. E. (1949) Winter food of Penns ylvania mink . Penns ylvania Game News 20: 12-32.
Hall, E. R. (1981) The mammal s of North Amer ica. 2nd ed. New York, NY: John W iley and Sons .
Hall, E. R.; Kelson, K. R. (1959) The mammal s of North Amer ica. 1st ed . New York, NY: The Ronald Press Co.
Hamilton, W. J., Jr . (1936) Food ha bit s of th e mink in New York. J. Mammal . 17: 169.
2-257 Mink

Hamilton, W. J., Jr . (1940) The summer food of mink s and r accoons on t he Mont ezuma Marsh, New York. J. Wildl. Manage. 4: 80-84.
Hamilton, W. J., Jr . (1959) Foods of mink in New York. N.Y. Fish and Game J. 6: 77-85.
Harding, A. R. (1934) Mink tr apping. Colum bus , OH: A. R. Harding.
Harper, R. H.; Travis, H. F.; Glinsky; M. S. (1978) Metaboliza ble en ergy r equir ement fo r maint enance and body co mpos it ion of growing f arm raised male mink ( Mustel a vison). J. Nutr . 108: 1937-1943.
Hornshaw, T. C.; Aulerich, R. J.; Johns on, H. E. (1983) Feeding G reat Lakes fi sh to mink: effects o n mink and accu mulation a nd e lim ination of PCBs by mink . J. Toxicol. Environ. Health 11: 933-946.
Humphr ey, S. R.; Zinn, T. L. (1982) Seasonal habitat u se by r iver otter s and everglades mink . J. Wildl. Manage. 46: 375-381.
Iversen, J. A. (1972) Basal energy meta bolis m of mu stelids . J. Comp. Physiol. 81: 341-344.
Korsch gen, L. J. (1958) Decemb er food ha bit s of mink in Mis sour i. J. Mammal . 39: 521-527.
Kostr on, K.; Kukla, F. (1970) Changes in thermoregula tion in m ink ki ts wi thin t he 45 days o f ontogenesis. Acta Univ . Agric., Facul tas Agronom ica, Sbor nik Vyso ke Sko ly Zemedelske (B runn) (r ada A) 18: 461-469.
Linn, I. J.; Bir ks, J. D. (1981) Observat ions on t he hom e ranges of fer al Amer ican mink (Mustel a vison) in Devon, Engla nd, as revealed by radio- tracking. In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. Augus t 1980; Frostb urg, MD; pp. 1088-1102.
Lins combe, G.; Kinle r, N.; Aulerich, R. J. (1982) Mink . In: Chapman, J. A.; Feldh ammer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 329-643.
Marshall, W. H. (1936) A stu dy of th e winter acti vi ties of th e mink. J. Mammal . 17: 382-392.
McCabe, R. A. (1949) Notes o n live-tr apping m ink . J. Mammal . 30: 416-423.
McDonne ll, J. A.; Gilbe rt, F. F. (1981) The responses o f mu skr ats (Ondatra zibethicus) to water level fluctu ations at Lut her Marsh, Ontar io. In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. Augus t 1980; Frostb urg, MD; pp. 1027-1040.
Melquis t, W. E.; Whitman , J. S.; Hornocker , M. G. (1981) Resource parti tioning a nd coexisten ce of symp atr ic mi nk and river otter popula tions . In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. Augus t 1980; Frostb urg, MD; pp. 187-220.
2-258 Mink

Mitch ell, J. L. (1961) Mink mo vemen ts an d popula tions on a Mont ana river . J. Wildl. Manage. 25: 48-54.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nationa l Research Counc il (NRC) (1982) Nutr ient requir ements o f mi nk and foxes. In: Nutrient requir ements o f domesti c animals ser ies, No. 7. Washingt on, DC: Nationa l Academy o f Sci ences, Nationa l Academy Pr ess.
Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
Pendleton, G. W. (1982) A selected annotated bibliogr aphy of mink behavior and ecology . Brook ings , SD: Sout h Dakota State Univ ersi ty; Tech. Bull. No. 3.
Perel'dik , N. S.; Milov anov , L. V.; Erin, A. T. (1972) Feeding f ur bearing a nim als. Washingt on, DC: Translated from Russi an by th e Agricultural Research Service, U.S. Departmen t of Agriculture and the Nationa l Science Founda tion.
Proulx , G.; McDonne ll, J. A.; Gilbe rt, F. F. (1987) The effect o f water level fluctu ations on muskr at, Ondatra zibethicus, predation by mink , Mustel a vison. Can. Field-Nat. 101: 89-92.
Sargeant, A. B.; Swanson, G. A.; Doty, H. (1973) Selecti ve predation by mink , Mustel a vison, on water fowl. Am. Midl. Nat. 89: 208-214.
Sealander, J. A. (1943) Winter food ha bit s of mink in s out hern Michiga n. J. Wildl. Manage. 7: 411-417.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Svilha , A. (1931) Habits o f th e Louis iana mink ( Mustel a vison vulgivagus). J. Mammal . 12: 366-368.
Willi ams, T. M. (1980) A comparison of running a nd s wimming e nergetics in t he mink (abstr act). Am. Zool. 20: 909.
Willi ams, T. M. (1983) Locomotion in t he Nor th Amer ican mink , a semi -aquatic mammal . I. Swimming e nergetics and body drag. J. Exp . Biol. 103: 155-168.
2-259 Mink

2.2.5. River Otter
Order Carnivora, Family Mustel idae. Mustelids h ave long, slender bodie s, shor t legs, and anal scen t glands. Throughout th e fami ly, the male is usually larger than the femal e. The more ter restr ial species of th is fami ly occu py var ious habitats an d feed primar ily on smal l mammal s and birds. Mustelids th at liv e around l akes an d str eams feed primar ily on aquatic sp ecies su ch as fi sh, frogs, and inver tebrates (Pal mer and Fowler, 1975; Burt and Grossen heider, 1980).
Selected species
The northern river otter (Lutra canadensis) histo rical ly l ived in or near lakes, marshes, str eams, and seash ores th roughout mu ch of th e Nor th Amer ican cont inent (Hall, 1981). Currently, many popula tions along t he coastal United States an d Canada are stab le or inc reasing, but this sp ecies is r are or exti rpated throughout mu ch of th e mid wester n United States (T oweill a nd T abor , 1982). The river otter dens in banks an d hollow logs . Indiv idua ls r ange over large areas daily, feeding pr imarily on f ish. Although o tter s have few n atural predators, while on la nd, they may b e taken by co yotes, fox, or dogs (M elquis t and Hornocker , 1983). Otter s cl ean themsel ves fr equently by rubbing a nd r olling in a ny dr y surface (T oweill a nd T abor , 1982). Otter s appear to unde rgo br adycar dia while subm erged and can stay unde rwater for up to 4 minutes (Melquis t and Dr onk ert, 1987). Because of i ts pis civorous die t and high t rophic l evel , the river otter is a notewo rthy indi cato r of bioaccu mulative pollut ion in a quatic ecosystems (M elquis t and Dr onk ert, 1987).
Body size. River otter s measu re 66 to 76 cm wi th a 30 to 43 cm tai l. Sexual dimorphism i n size is seen among a ll s ubs pecies (Harris, 1968; van Zyl l de Jong, 1972, ci ted in T oweill a nd T abor , 1982), and adult mal es (5 to 10 kg) outweigh femal es (4 to 7 kg) by approximatel y 17 percent (Melquis t and Hor nocker , 1983; see Table). Full a dult weight generally is not attai ned unti l sexu al matu rity after 2 years of age (Melquis t and Hor nocker , 1983). Along t he Paci fic Coast, there is so me evi dence th at si ze decreases fr om north to sout h (Toweill a nd T abor , 1982).
Metabolism. Iversen (1972) found t hat basal metabolic r ate of otter s and other mustelids weighing 1 kg or more could be exp ressed by th e equation:
BMR = 84.6Wt0.78 (±0.15),
where BMR = basal metabolic r ate in kcal /day and Wt = body weight in k ilogr ams. Free-liv ing metabolic r ates w ould be exp ected to be three to five ti mes highe r (see tab le).
Habitat. Almost excl usively aquatic, the river otter is found in f reshwater , estu arine, and some mar ine environments al l the way fr om co astal areas to mountain lakes (T owei ll and Tabor, 1982). They are found pr imarily in f ood- rich coastal areas, such as th e lower por tions of str eams an d rivers, estu aries, nonpollut ed water ways, the lakes an d tr ibut aries that feed rivers, and areas sh owing lit tle hum an impact (Mowbray et al ., 1979; Tabor and Wight , 1977).
2-261 River Otter

Food ha bi ts. Otter s usually are acti ve in the evening a nd f rom dawn to midm orning, although t hey can be acti ve any ti me of day (Melquis t and Hor nocker , 1983). The bulk of the river otter 's diet is fi sh; however , otter s are oppor tunis tic and w ill feed on a var iety o f prey depending on availa bilit y and ease of cap ture. River otter s take d iffer ent fi sh species acco rding t o their availa bilit y and how well t he fish can escap e capture (Loranger, 1981). Depending on availa bilit y, otter s also may co nsume cr ustacean s (esp ecially cr ayfi sh), aquatic insects (e. g., sto nefly nymp hs, aquatic beetles), amphibia ns, insects, birds (e.g., ducks), mammal s (e.g., young beaver s), and turtles (Burt and Grossen heider, 1980; Lagler and Osten son, 1942; Liers, 1951b; Melquis t and Hor nocker , 1983; Palmer and Fowler, 1975; Toweill a nd T abor , 1982). Gilbe rt and Nanceki vel l (1982) observed that otter s co nsume more water fowl in the northerly lati tudes th an in the south, probably because of th e ease of capturing t he water fowl during t heir molt in t he nor th. Otter s probe the botto ms o f ponds or str eams fo r inver tebrates an d may ingest mu d or other debris in the process (L iers, 1951b). Otter s in captivi ty r equir ed 700-900 g of food da ily (H arris, 1968, ci ted in Towei ll and Tabor, 1982).
Temperature regulation and molt. Seasonal patter ns in otter s are not wel l unde rstood. Otter s are acti ve th roughout th e year (Toweill a nd T abor , 1982), wi th the most intense acti vi ty l evels during t he wint er (Larsen, 1983; Melquis t and Hor nocker , 1983). They unde rgo a gradual molt in s pring a nd f all (Melquis t and Dr onk ert, 1987).
Breeding acti vi ties and social organization. Adult mal es are usually solit ary; an adult femal e and two or three pups make u p a typ ical fami ly group ( Melquis t and Dr onk ert, 1987). River otter s breed in late wi nter or early sp ring ov er a period of 3 mont hs or more. Birth of a l itter follow s mating by about 1 year ; however , implantat ion is delayed for approximatel y 10 mo nths, and acti ve gestat ion l asts only 2 mont hs (Pearson and Enders, 1944, ci ted in Toweill a nd T abor , 1982; Melquis t and Dr onk ert, 1987). Newborn otter s are bor n blind but fully fu rred and de pend on t heir mo ther fo r milk unt il 3 to 5 mont hs of ag e (Johnsto ne, 1978; Liers, 1951b). Fami ly groups dis perse about 3 mont hs after the pups ar e weaned (Melquis t and Hor nocker , 1983).
Home range and resources. The river otter 's home range encompasses th e area needed for fo raging a nd r eproduc tion ( Melquis t and Dr onk ert, 1987). The shape of th e home range var ies by habitat typ e; for examp le, near rivers or coastal areas, it may b e a long s tr ip a long t he shor eline (measu red in kilometer s), but in marshes or areas wi th many smal l str eams, the home range may r esemb le a poly gon (measu red in hectar es; Melquis t and Dronker t, 1987). All parts o f a home range are not used equally; instead , sever al acti vi ty cen ters may b e interconnected by a str eam or coast (Melquis t and Hor nocker , 1983). Food ha s the greatest i nfluence on habitat u se, but adequate sh elter in the form of tempor ary dens and r esting s ites al so plays a r ole (Anderson and Woolf , 1987a; Melquis t and Hornocker , 1983). River otter s use dens dug by other anim als or natural shelters such as hollow logs , logj ams, or drift pile s (Toweill a nd T abor , 1982; Melquis t and Dr onk ert, 1987). Beaver bank dens and lodge s account ed for 38 percent of r esting s ites u sed by radio-tracked otter s in Idaho (Melquis t and Hor nocker , 1983). River otter s appear to prefer flow ing water habitats (e. g., str eams) o ver more stat iona ry water (e.g., lakes, ponds ) (Idaho study ; Melquis t and Hor nocker , 1983).
2-262 River Otter

�
River otter s mai ntain disti nct ter ritories wi thin t heir hom e ranges: femal es mai ntain a feeding a rea for th eir familie s, mal es for breeding pur poses (T oweill a nd T abor , 1982). Indiv idua ls tend t o avoid c onf rontation t hrough m utual avoida nce (Melquis t and Hor nocker , 1983). Home ranges are most r estr icted for lactat ing femal es (Melquis t and Dr onk ert, 1987). Adult and s ubadult mal es have larger, more var iable home ranges th an femal es.
Popul ation de nsity. River otter popula tions show var iable sp acing in r elation t o prey densi ty an d habitat (Hornocker et al ., 1983). This ch aracter isti c, along w ith their secr etive habits an d use of sever al den si tes, makes i t d iffi cult to esti mate r iver otter popula tions (M elquis t and Dr onk ert, 1987). Popula tion de nsity of o tter s often is exp ressed in terms o f number per ki lometer of water way o r coast line b ecause of th eir dependence on aquatic habitats. Densi ties between one otter every k ilometer to one otter every 10 km o f river or shor eline ar e typic al (see tab le).
Popul ation pa rameter s. Otter s generally are not sexu ally matu re unti l 2 years of age (Liers, 1951b; Hamilton and Eadie, 1964; Tabor and Wight , 1977; Lauhachinda , 1978). Adult femal es appear to reproduce year ly in Oregon ( based on a pregnancy r ate of almost 100 percent; Tabor and Wight , 1977), but Lauhachinda (1978) conclude d that they br eed every other year in Alabama and Georgia. Litter s usually co nsist o f two to three pups , although li tter s as l arge as si x pups occur (see tab le). As adults, river otter mortal ity r ates are low, between 15 and 30 percent per year (Lauhachinda , 1978; Tabor and Wight , 1977).
Simi lar species (from general references)
� The sea otter (Enhy dra lutr is) (76 to 91 cm body and 2 8 to 33 cm tail; weight 13 to 38 kg) inha bit s kelp be ds and r ocky sh ores fr om th e Aleutian Islands to California. Its d iet includes fi sh, abalones, sea urchins , and ot her mar ine animals.
General references
Burt and Grossen heider (1980); Melquis t and Dr onk ert (1987); Palmer and Fowler (1975); Toweill a nd T abor (1982).
2-263 River Otter

River Otter ( Lutra canadensis )
Factors Age/Sex/ (95% CI of Note Cond. /Seas. Mean mean) Locat ion erence No.
Range or
Weight A B 5.0 - 15 throughout range Melqui st & Dronker t, 1987 1 (kg)
A M 8.13 ± 1.15 SD 5.84 - 10.4 Alabama, Georgia Lauhachi nda, 1978 A F 6.73 ± 1.00 SD 4.74 - 8.72 Y M 6.36 ± 0.98 SD 4.41 - 8.31 Y F 5.83 ± 1.82 SD 3.75 - 7.01
A M 9.20 ± 0.6 SE wc Idaho Melqui st & Hor nocker , 1983 A F 7.90 ± 0.2 SE Y M 7.90 ± 0.4 SE Y F 7.20 ± 0.1 SE
neonat e 0.132 New York Hamilton & Eadi e, 1964 neonat e 0.140 to 0.145 Alabama, Georgia Hill & Lauhachinda, 1981
Pup Growth 10 to 20 days 26.7 NS Liers, 1951a 2 Rate (g/day)
Metabol ic Rate A F basal 44.8 est imated 3 (kcal /kg-day) A M basal 42.6
A F f ree-living 183 (83 - 400) est imated 4 A M free-living 178 (81 - 391)
Food Ingest ion 5 Rate (g/g-day)
Water A F 0.082 est imated 6 Ingest ion A M 0.080 Rate (g/g-day)
Inhal ation A F 2.5 est imated 7 Rate (m /day) A M 2.93
Surface Ar ea A F 3,785 est imated 8 (cm ) A M 4,2802
Ref
2-264 R
iver Otter

---- --
River Otter ( Lutra canadensis )
2-265 R
iver Otter
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
fish 100 93 97 99 wc Idaho/mount ain st reams Melqui st & Hor nocker , 1983 (sucker ) (52) (47) (17) (30) and l akes (scul pins) (40) (31) (38) (42) (squawfish) ( 5) ( 4) ( 1) ( 6) (percent fre quency of (perch) (22) ( 3) ( 7) ( 9) occur rence i n scat s) (whi tefish) (21) (10) (24) (66)
invertebrates 2 7 10 12 (most of the fish wer e longer bi rds <1 12 1 <1 than 30 cm ) mammals 1 4 3 1 reptiles 0 1 0 0
invertebrates 41.6 44.2 33.3 26.3 nw Mont ana/ Greer, 1955 (aquat ic insect s) 19.6 19.2 10.7 4.0 lakes and st reams (fr water shr imp) 14.3 8.9 10.7 4.0
fishes 91.4 92.9 100 100 (percent frequency of (trout ) 23.7 9.8 33.3 29.3 occur rence i n scat s) (scul pin) 20.5 20.9 21.3 25.3 (sunf ish) 47.1 72.8 60.0 33.3
frog 19.6 19.2 10.7 9.1 salamander 0.3 0.7 1.3 snake 0.2 0.7 birds 6.7 4.1 1.3 1 mammals 8.1 5.3 2.7 4.0
fish 97 69 98 99 nw Illinoi s/Missi ssi ppi River Ander son & W ool f, 1987b (sunfish) (31) (31) (80) (52) (minnow/ carp) (52) ( 0) (17) (44) (percent frequency of (herring) (49) (38) (10) (40) occur rence i n scat s) (bass) (26) ( 0) ( 5) (14)
frogs 3 6 11 16 crayfish 12 50 8 7 dragonf ly nymphs 2 bi rds 4 13 3 1
2 6 0

River Otter ( Lutra canadensis )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
game & pan f ish 32 Michi gan/habi tat NS Lagl er & Ostenson, 1942 forage f ish 17.6 fish r emains 3.0 (% vol ume; stomach amphi bians 16.1 cont ents) other invertebrates 25.8
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Home Range A B 400 - 1,900 ha Missour i/marsh, st reams Erickson et al., 1984 9 Size (ha or km river) A B 2,900 - 5,700 ha Colorado ( fall-spr )/NS Mack, 1985 9
A M 400 ha se Texas/ coast al marsh Foy, 1984 9 A F 295 ha
yearling M 43 ± 20 SD km 10 - 78 km wc Idaho/river drainage Melqui st & Hor nocker , 1983 yearling F 32 ± 6.2 SD km 25 - 40 km (no t rends seen w ith adul t F 31 ± 9.2 SD km 23 - 50 km season) B B 28 ± 7.5 SD km 15 - 39 km
Popul ation B B 0.26/km 0.17 - 0.37/km wc Idaho/river drainage Melqui st & Hor nocker , 1983 Densi ty A F breeding 0.05/km (N/ha or N/km A M breeding 0.019/km shor eline) yearling B 0.071/km
B B 0.85/km se Al aska/coast al - island Wool ingt on, 1984 9
B B 0.0094 - 0.014 /ha se Texas/ coast al marsh Foy, 1984 9
A B 0.0025/ha Missour i/marsh, st reams Erickson et al., 1984 9
2-266 R
iver Otter

River Otter ( Lutra canadensis )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Litte r 2.73 ± 0.77 SD 1 - 4 Maryland/wetlands Mowbray et al., 1979 10 Size 2.68 ± 0.71 SD 1 - 4 Alabama, Georgia/NS Hill & Lauhachinda, 1981
1 yr old 0.53 ± 0.91 SD 0 - 3 Maine/NS Docktor et al., 1987 11 2 yr old 0.87 ± 0.96 SD 0 - 3 3 yr old 1.60 ± 1.42 SD 0 - 4 4 yr old 2.29 ± 1.25 SD 1 - 5 5 to 12 yrs ol d 2.67 ± 1.40 SD 0 - 6
2.1 ± 0.7 SD New York/NS Hamilton & Eadi e, 1964
Li tters 1 NS Trippensee, 1953 /Year
Days total 290 - 380 Wisconsi n/capt ive Liers, 1951b 12 Gestation
act ive 60-63 NS Lanci a & Hair, 1983 13
Age at > 90 days NS Harris, 1968 Weaning
Age at F 2 yrs New York/NS Hamilton & Eadi e, 1964 Sexual M 2 yrs Maturity
Annual birth - 1 yr 32 Oregon/ NS Tabor & Wight , 1977 Morta lity 1 - 2 yrs 54 Rates (percent ) 2 - 11 yrs 27
A M 17.8 Alabama, Georgia/river ine Lauhachi nda, 1978 A F 20.3
Longevi ty A B < 15 yrs Alabama, Georgia/river ine Lauhachi nda, 1978
2-267 R
iver Otter

123456789
10111213
River Otter ( Lutra canadensis )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating Januar y March to Apr il May Michi gan Hooper & Ost enson, 1949 14 March Apr il New York Hamilton & Eadi e, 1964 winter late winter spr ing AL, FL, GA Lauhachi nda, 1978
Partur ition mid-March mid-May Maryland, Chesapeake Bay Mowbray et al., 1979 late March early Apr il wc Idaho Melqui st & Hor nocker , 1983 late Januar y May Alabama Lauhachi nda, 1978
Disper sal Apr il to May wc Idaho Melqui st & Hor nocker , 1983 15
2-268 R
iver Otter
Summary of studi es di scussed by Hall (1981) and Wool ingt on (1984).Cited in Toweill a nd Tabor (1982).Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and adul t body weight s f rom Lauhachi nda (1978).Est imated usi ng equat ion 3-47 (Nagy, 1987) and adul t body weight s f rom Lauhachi nda (1978).See Chapt ers 3 and 4 f or methods of est imating f ood i ngest ion r ates.Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and adul t body weight s f rom Lauhachi nda (1978).Est imated usi ng equat ion 3-20 (Stahl , 1967) and adul t body weight s f rom Lauhachi nda (1978).Est imated usi ng equat ion 3-22 (Stahl , 1967) and adul t body weight s f rom Lauhachi nda (1978).Cited in Melqui st and Dr onker t (1987).Determined f rom implanted embryo count s.Determined f rom cor por a lutea count s.Total gest ation per iod ( incl udi ng pr eimplantation).Act ive gest ation per iod (post implantation), ci ted in Melqui st and Dr onker t (1987).
14 Cited in Toweill a nd Tabor (1982). 15 Disper sal at age 12 to 13 mont hs.

References (including A ppendix)
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Anderson, K. L. (1981) Popula tion a nd r eproduc tion c haracter isti cs o f th e river otter in Virginia and t issu e conc entrations of en vironm ental con tami nants [master 's th esis]. Blacksb urg, VA: Virginia Pol ytec hnic Insti tute.
Anderson, K. L.; Scanlon, P. F. (1981) Reproduc tion a nd popula tion c haracter isti cs o f r iver otter s in Virginia . Virginia J. Sci . 32: 87.
Anderson, E. A.; Woolf , A. (1987a) River otter habitat u se in northwester n Illinois . Trans. Illinois A cad. Sci . 80: 107-114.
Anderson, E. A.; Woolf , A. (1987b) River otter food ha bit s in nor thwester n Illinois . Trans. Illinois A cad. Sci . 80: 115-118.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chabreck, R. H.; Holc ombe, J. E.; Lins combe, R. G.; et al. (1982) Winter foods of r iver otter s fr om sa line and f resh environm ents in Louis iana. Proc. Annu. Conf . Southeast Asso c. Fish Wildl. Agencies 36: 473-483.
Dockto r, C. M.; Bowyer , T. R.; Clark, A. G. (1987) Number of co rpora lutea as r elated to age and dis tr ibut ion of r iver o tter in Maine. J. Mammal . 68: 182-185.
Eisenberg, J. F. (1981) The mammal ian radiations ; an analysi s of tr ends in evolut ion, adaptation, and be havior . Chic ago, IL: Univ ersi ty of Chic ago Press.
Erickson, D. W.; McCullough, C. R.; Porath, W. R. (1984) River otter investi gations in Misso uri. Misso uri Dept. Conserv.; Pittman -Robertson P roj. W-13-R-38, Fina l Repor t.
Foy, M. K. (1984) Seasonal movemen t, home range, and habitat u se of r iver otter in southeaster n Texas [master 's th esis]. Colle ge Station, TX: Texas A&M Universi ty.
Gilbe rt , F. F.; Nanceki vel l, E. G. (1982) Food ha bit s of mink ( Mustel a vison) and otter (Lut ra canadensis) in northeaster n Alberta. Can. J. Zool. 60: 1282-1288.
2-269 River Otter

Greer, K. R. (1955) Yearly food ha bit s of th e river o tter in the Thompson Lakes r egion, Northwester n Mont ana, as indi cated by scat an alyses. Am. Midl. Nat. 54: 299-313.
Greer, K. R. (1956) Fur resources an d investi gations : study of th e otter food ha bit s along a segment of th e Gallatin River . Montana Fish and Game Dept.; Job Comp. Rep. W-049-R-06:35-59.
Grenfel l, W. E., Jr . (1974) Food ha bit s of th e river o tter in Suisin Marsh, central California [master 's th esis]. Sacramento, CA: California State Universi ty.
Grinne ll, J.; Dixon, J. S.; Lins dale, J. M. (1937) Fur-bearing mammal s of California. Berkeley, CA: Universi ty o f California Press.
Hall, E. R. (1981) The mammal s of North Amer ica. 2nd ed. New York, NY: John W iley and Sons .
Hall, E. R.; Kelson, K. R. (1959) The mammal s of North Amer ica. 1st ed . New York, NY: The Ronald Press Co.
Hamilton, W. J., Jr . (1961) Late fal l, winter and early sp ring f oods of 141 otter s fr om New York. N. Y. Fish and Game J. 8: 106-109.
Hamilton, W. J., Jr .; Eadie, W. R. (1964) Reproduc tion in t he otter , Lutra canadensis. J. Mammal . 45: 242-252.
Harris, C. J. (1968) Otter s: a stu dy of th e recent Lutr inae. London, U.K.: Weidenf ield & Nicols on.
Harris, J. (1969) Breeding t he Canadian otter Lutra c. canadensis in a private c olle ction. Int . Zoo Year book 9: 90-91.
Hill, E. P.; Lauhachinda , V. (1981) Reproduc tion in r iver o tter s fr om Alabama and Georgia. In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. August 1980; Frostb urg, MD.
Hoope r, E. T.; Osten son, B. T. (1949) Age groups in Mic higa n otter . Ann A rbor , MI: Univ ersi ty of Michiga n; Mus. Zool. Occas. Pap. 518.
Hornocker , M. G.; Messi ck, J. P.; Melquis t, W. E. (1983) Spacial str ategies in three species of Mustel idae. Acta Zool. Fenn. 174: 185-188.
Humphr ey, S. R.; Zinn, T. L. (1982) Seasonal habitat u se by r iver otter s and Everglades mink . J. Wildl. Manage. 46: 375-381.
Iversen, J. A. (1972) Basal energy meta bolis m of Mustelids . J. Comp. Physiol. 81: 341-344.
Johnsto ne, P. (1978) Breeding a nd r earing t he Canadian otter (Lutra canadensis) at Mole Hall Wildlif e Park, 1966-1977. Int. Zoo Yearbook 18: 143-147.
2-270 River Otter

Knuds en, G. J.; Hale, J. B. (1968) Food ha bit s of o tter s in the Great Lakes r egion. J. Wildl. Manage. 32: 89-93.
Lagler, K. F.; Osten son, B. T. (1942) Early sp ring f ood of th e otter in Michiga n. J. Wildl. Manage. 6: 244-254.
Lancia, R. A.; Hair, J. D. (1983) Popula tion statu s of bobcat (Felis rufus) and river otter (Lutra canadensis) in Nor th Carolina . Raleigh, NC: Nor th Carolina State Univ ., Proj. E-1; 65 pp.
Larsen, D. N. (1983) Habitats, movemen ts, and foods of r iver o tter s in coastal southeaster n Alaska [master 's th esis]. Fairbanks, AL: Universi ty o f Alaska.
Larsen, D. (1984) Feeding ha bit s of r iver o tter s in coastal southeaster n Alaska. J. Wildl. Manage. 48: 1446-1452.
Lauhachinda , V. (1978) Life histo ry of th e river otter in Alabama wi th emphasis on food habits [Ph .D. disser tat ion] . Aubur n, AL: Univ ersi ty of A labama.
Liers, E. E. (1951a) My fr iends th e land otter s. Nat. Hist. 60: 320-326.
Liers, E. E. (1951b) Notes o n the river otter (Lutra canadensis). J. Mammal . 32: 1-9.
Liers, E. E. (1966) Notes o n breeding t he Canadian otter Lutra canadensis in captivi ty an d longe vi ty records of b eavers Casto r canadensis. Int . Zoo Yearbook 6: 171-172.
Loranger, A. J. (1981) Late fal l and early wi nter foods of th e river o tter (Lutra canadensis) in Massach usetts, 1976 - 1978. In: Chapman, J. A.; Pursley, D., eds. Worldwide furbearer conf erence pr oceedings , v 1; Augus t 3-11, 1980; Frostb urg, MD; pp. 599-605.
MacFarlane, R. (1905) Notes o n mammal s collected and observed in the northern Mackenzie River Dis tr ic t. Proc. U.S. Natl . Mus. 23: 716-717.
Mack, C. M. (1985) River otter resto ration in G rand Count y, Color ado [master 's th esis]. Fort Collins , CO: Color ado State Universi ty.
McDaniel, J. C. (1963) Otter popula tion s tudy . Proc. Annu. Conf . Sout heast. Asso c. Game and Fish Comm. 17: 163-168.
Melquis t, W. E.; Dronk ert, A. E. (1987) River otter . In: Novak, M.; Baker , J. A.; Obbarel, M. E.; et al., eds. Wild f urbearer man agement and conservat ion. Pittsbur gh, PA: Universi ty o f Pi ttsb urgh Press; pp. 627-641.
Melquis t, W. E.; Hornocker , M. G. (1983) Ecology of r iver o tter s in west cen tral Idaho. In: Kir kpatr ick, R. L., ed. Wildlif e monogr aphs : v. 83. Bethesda, MD: The Wildlif e Society; 60 pp.
2-271 River Otter

Melquis t, W. E.; Whitman , J. S.; Hornocker , M. G. (1981) Resource parti tioning a nd coexisten ce of symp atr ic mi nk and river otter popula tions . In: Chapman, J. A.; Pursley, D., eds. Worldw ide fu rbearer conf erence pr oceedings , v 1; Augus t 3-11, 1980; Frostb urg, MD; pp. 187-220.
Modaffer i, R.; Yocom, C. F. (1980) Summer food of r iver o tter in north coastal California lakes. Murrelet 61: 38-41.
Mowbray, E. E.; Pursley, D.; Chapman, J. A. (1979) The statu s, popula tion c haracter isti cs and harvest o f th e river otter in Maryland. Waver ly Press; Maryland Wildl. Admin. , Publ. Wildl. Ecol. 2; 16 pp.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Mono. 57: 111-128.
Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
Pearson, O. P.; Enders, R. K. (1944) Duration of pregnancy in c ertain Mu stelids . J. Exp . Zool. 95: 21-35.
Pierce, R. M. (1979) Seasonal feeding ha bit s of th e river o tter (Lutra canadensis) in ditch es of th e Great Dismal Swamp [master 's th esis]. Norfolk, VA: Old Dominion Univ ersi ty.
Reid, D. G. (1984) Ecologic al int eractions of r iver o tter s and beavers in a boreal ecosystem [master 's th esis]. Calgary, Canada: Univ ersi ty of Calgary.
Ryder, R. A. (1955) Fish predation by th e otter in Michiga n. J. Wildl. Manage. 19: 497-498.
Scheffer , V. B. (1958) Long lif e of a river o tter . J. Mammal . 39: 591.
Sheldon, W. G.; Toll, W. G. (1964) Feeding ha bit s of th e river o tter in a reservoir in central Massach usetts. J. Mammal . 45: 449-455.
Shirley, M. G. (1985) Spring f ood ha bit s of r iver o tter in southwester n Louis iana (abstr act only). Proc. La. Acad. Sci . 48: 138.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stenson, G. B.; Badgero, G. A.; Fisher, H. D. (1984) Food ha bit s of th e river o tter Lut ra canadensis in t he mar ine en vironm ent of Br it ish Colum bia. Can. J. Zool. 62: 88-91.
Tabor , J. E.; Wight , H. M. (1977) Popula tion statu s of r iver otter in wester n Oregon. J. Wildl. Manage. 41: 692-699.
2-272 River Otter

Toll, W. G. (1961) The ecology of th e river o tter (Lutra canadensis) in t he Quabbin Reservat ion of cen tral Massach usetts [master 's th esis]. Amherst, MA: Universi ty o f Massach usetts.
Toweill, D. E. (1974) Winter food ha bit s of r iver o tter s in wester n Oregon. J. Wildl. Manage. 38: 107-111.
Toweill, D. E.; Tabor , J. E. (1982) River otter . In: Chapman, J. A.; Feldhammer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 688-703.
Trippe nsee, R. E. (1953) Wildlif e management: fur bearers, water fowl, and fish. New York, NY: McGraw-Hill.
Tuml ison, R.; Shalaway, S. (1985) An annotated bibliogr aphy on t he Nor th Amer ican river otter Lutra canadensis. Sti llwater , OK: Okla. Fish Wildl. Res. Unit & Dept . Zool. , Oklahoma State Universi ty.
van Zyll de Jong, C. G. (1972) A systemati c review of th e nearcti c and neotropical river otter s (Genus Lut ra, Mustelida e, Carniv ora). Ontar io, Canada: R. Ontar io Mus ., Lif e Sci . Cont r. 80.
Wilson, K. A. (1959) The otter in North Carolina . Proc. Sout heast. Asso c. Fish and Game Comm. 13: 267-277.
Wilson, K. A. (1985) The role of mi nk and otter as mu skr at predators in northeaster n North Carolina . Proc. Annu. Conf . Sout heast. Asso c. Game Fish Comm. 18: 199-207.
Woolingt on, J. D. (1984) Habitat u se and movemen ts o f r iver otter s at K elp Bay, Baranof Island, Alaska [master 's th esis]. Fairbanks, AK: Universi ty o f Alaska.
2-273 River Otter

2.2.6. Harbor Seal (hair seals)
Order Carnivora, Family Phoc idae. Seals, sea lions , and w alruses ar e collecti vely refer red to as pinnipe ds (L atin f or wing- foot ed). Pinnipe ds ar e div ided int o three fami lies: otar ids (sea lions and f ur seals); phoc ids (h air seals, also cal led true seals or earless seals); and walruses. Most pinnipe ds feed on mar ine species su ch as fi sh, squid, and other inver tebrates (B urt and Grossen heider, 1980). Unlik e fur seals, which are protected from th e cold m arine en vironm ent by a de nse layer of unde rfur , phoc ids r ely only on a thic k blubbe r layer for insulation ( Pierot ti and Pierot ti , 1980). Phocids include bot h the smal lest (r ing seals) and the largest (el ephant seal s) of th e pinnipe ds. The geogr aphic range of mo st phoc id s pecies is fr om th e arcti c Atlant ic and Paci fic so uth to the coasts o f Canada and Alaska, although s ome do inha bit war mer water (Burt and Grossen heider, 1980). Most phoc ids , wi th the excep tion of th e elephant seal , do not exhibit th e large disparity i n size between the sexes, which is ch aracter isti c of otar ids (sea lions and f ur seals) (Burt and Grossen heider, 1980).
Selected species
In North Amer ica, harbor seals (Phoca vi tu lina) range from Alaska to Baja California, Mexico, along t he Paci fic co ast (su bspecies ric hardsi ; Hoover , 1988), and from Newfoundla nd t o easter n Long Is land a long t he Atlant ic co ast (su bspecies conc olor ; Payne and Selzer, 1989). They are one of th e most co mmonly seen pinnipe d species, in pa rt due to their tende ncy to inha bit co astal areas (Hoover , 1988). Harbor seals can be found a long the Paci fic co ast o n a year -round ba sis (excep t during s tormy pe riods in w int er), but Atlant ic popula tions wint er offsh ore when coastal ice has fo rmed in their usual haul-out areas (Boulv a and Mc Laren, 1979). The recent increases i n harbor seal popula tions in New Engla nd water s appear to be due to a southward dispersal of seal s fr om rooker ies in Maine follow ing t he termina tion of a Massach usetts bount y on ha rbor seals (1962) and the passag e of th e Marine Mammal Protect ion A ct (1972) (Payne and Schneider, 1984).
The spotted or largha seal (Phoc a largha) is a cl osely related species th at unti l recently was co nsidered a subspecies of th e harbor seal . It is si mi lar in size, appearance, and f eeding ha bit s to the Paci fic harbor seal , but i t ten ds to inha bit colde r water s along t he Paci fic co asts (A shwel l-Erickso n and Elsner, 1981). In North Amer ica, it seldom ven tures further south than the northern coast o f Alaska (Ashwel l-Erickso n and Elsner, 1981). The spotted seal requir es ic e for breeding ha ul-out s and gi ves b irth about 2 mont hs ear lier th an the Paci fic harbor seal (Ashwel l-Erickso n and Elsner, 1981; Boulv a and Mc Laren, 1979). The harbor seal , in contrast, breeds on land (Boulv a and Mc Laren, 1979).
Body size. The lengt h and w eight of harbor seals var y geographical ly, but sexu ally matu re adult s tend t o be about 1. 5 m in le ngt h and w eigh f rom 65 to 90 kg (Ashwell-Erickso n and Elsner, 1981; Pitch er and Calkins, 1979). Harbor seals exhibit so me sexu al dim orphis m, th e male being la rger (Pi tch er and Ca lk ins , 1979). Body lengt h us ually is used to measu re size because weight can vary sub stan tially wi th facto rs su ch as seaso n, food availa bilit y, and m olt ing ( Ronald et al., 1982). Newborn pups ar e around 8 0 cm long a nd weigh f rom 8.6 to almost 15 kg, wi th femal es often weighing l ess th an males (Newby, 1973; Pitch er and Calkins, 1979; Rosen, 1989). Harbor seal pups ar e highly precocial and are
2-275 Harbor Seal

able to swi m within hour s of bir th (Boulv a and Mc Laren, 1979; Lawso n and Renouf , 1987). Seal mi lk co nsists o f about half fat, and the pups mo re than double th eir weight before they are weaned at approximatel y 30 days (Bigg, 1969a, as ci ted in Pitch er and Calkins, 1979). Harbor seals co ntinue to grow wi th age for sever al years beyond t he age of sexu al matu rity (Boulv a and Mc Laren, 1979; Pitch er and Calkins, 1979). Body fat var ies seaso nally wi th food intake, while to tal body weight and le an body mass i ncrease wi th age (Ashwel l-Erickso n and Elsner, 1981). Harbor seals, unlik e many ot her pinnipe ds, do not fast fo r exten ded periods dur ing t he molt ing pe riod or breeding seaso n (Boulv a and Mc Laren, 1979; Pierotti and Pierotti , 1980).
Habitat . Harbor seals inha bit a var iety o f environments an d are able to to lerate a wide range of temp eratures and water salinit ies (Boulv a and Mc Laren, 1979; Hoover , 1988). In its easter n range, the harbor seal inha bit s inl ets, islets, reefs, and sandba rs (Boulv a and McLaren, 1979). In wester n North Amer ica, the harbor seal inha bit s tida l mud f lats, sand bars, shoals, river deltas, estu aries, bays, coastal rocks, and offsh ore islets (J ohns on and Jeffr ies, 1977), even ranging up r ivers int o f reshwater areas in search of food ( Roffe an d Mate, 1984). Harbor seals also inha bit so me freshwater lakes (Po wer and Gregoir e, 1978). Habitats u sed for haul-outs i nclude cobble and s and b eaches, tidal mud flats, offsh ore rocks an d reefs, glacial and sea ice, and man-made objects su ch as piers and log boom s (Hoover , 1988).
Food ha bi ts . Harbor seals' diet var ies seaso nally and include s bot tom-dwelling fishes (e.g., flounde r, sole , eelpout ), inv ertebrates (e.g., octopus), and species th at can be caught in pe riodic sp awning a ggr egations (e. g., herring, lance, squid) (Ever itt et al ., 1981; Lowry and Frost, 1981; Pitch er and Calkins, 1979; Roffe an d Mate, 1984).g Harbor seals are oppor tunis tic, cons uming di ffer ent prey in relation t o their availa bilit y and ease of cap ture (Pitch er and Calkins, 1979; Pitch er, 1980; Shaffer , 1989). They may mo ve into rivers on a seaso nal basis in pursuit of prey (e.g., eulachon in t he Colum bia River dur ing w int er; Brown et al ., 1989). They hunt alone or in small gr oups (Hoov er, 1988). Fish species consumed range between 40 and 280 mm, wi th mean values of between 60 and 180 mm (Brown and Mate, 1983). Recently wean ed pups tend t o feed on prey th at are more easi ly captured than fish, such as sh rimp or other crustacean s (Hoover , 1988; Pitch er and Calkins, 1979). During t he breeding a nd m olt ing seaso ns, when harbor seals sp end more time on la nd, adult s rely on t heir blubbe r layer as an addit iona l sour ce of en ergy (Ashwell-Erickso n and Elsner, 1981). During t his ti me, th ey may b e more suscep tible to lipophilic contami nants (e.g., PCBs) th at may h ave accu mulated in their blubbe r (Hoov er, 1988).
g Studi es of harbor seal diet of ten rel y on count s of fish sagi ttal otol iths f ound i n scat s or st omachcont ents. These ot ol iths can be i dent ified to the level of speci es, annul i on t he otol iths count ed to determ ine age, and fish weights and lengths esti mated fro m otolith dimensions. However, parti al or complete d igesti on of otoliths, parti cularly of small fish species, may resu lt in significant underest imates of the proport ion of these prey i n seal diets, part icularly from scat analysi s (da Silva and Neilson, 1985; Harvey, 1989). Studies of sto mach contents o f stran ded seals also may present a b iased pictu re of dietary co mposition due to exten ded periods of fasti ng prior to stranding (Selzer et al., 1986).
2-276 Harbor Seal

�
In general, food c ons umpt ion by adult seals is highest i n winter and lowest i n the summer (Ashwel l-Erickso n and Elsner, 1981; Ashwel l-Erickso n et al ., 1979). Innes et al . (1987) esti mated allometr ic equations fo r maint enance food ing estion r ates (IR; wet-we ight biomass) wi th body weight (B W, kg) for phoc ids :
IRmaint(kg/day) = 0.079 BW(kg)0.71 adult (N = 11; r2 = 0.84);
IRmaint(kg/day) = 0.032 BW(kg)1.00 juveniles (N = 19; r2 = 0.68); and
IRmaint(kg/day) = 0.068 BW(kg)0.78 both adults an d juveniles (N = 30; r2 = 0.68).
Allometr ic equations fo r food ing estion r ates o f growing a nim als (IR; wet-we ight biomass) wi th body weight (B W, kg) for phoc ids al so have been esti mated (Innes et al ., 1987):
IRgrow th(kg/day) = 0.0919 BW(kg)0.84 adult (N = 11; r2 = 0.84); and
IRgrow th(kg/day) = 0.0547 BW(kg)0.84 juveniles (N = 19; r2 = 0.68).
Innes et al . (1987) found t hat gr owing juv enile phoc id seals ingested 1.7 times mo re biomass p er day th an a simi lar-sized growing a dult and 1 .4 times mo re than juvenile phoc ids th at were not growing.
Boulv a and Mc Laren (1979) esti mated a relations hip b etween body weight and da ily food ingestion f or harbor seals fr om easter n Canada:
IRfree-livi ng(kg/day) = 0.089 BW(kg)0.76 adults (N = 26).
Perez (1990) esti mated the average energy val ue of th e harbor seal 's diet to be 1.4 kcal /g wet we ight . Ashw ell-Erickso n and Elsner (1981) provide age-speci fic esti mates o f fo od inge stion r ates fo r the closely related spotted seal (see Appendix) and summar ize studie s in w hic h food ing estion r ates fo r harbor and spotted seals have been esti mated .
Temperature regulation and molt. Harbor seals can maintain their heat balance while div ing in water as low as 13�C without inc reased muscl e acti vi ty o r metabolic r ate (Ronald et al., 1982). For seals in general, molting is si mply part of an ongoing pe lage cycl e that is influenced by th e seal 's environment, physiology , and be havior (Ling, 1974). Phocids get an enti rely new co at wi th each annua l molt (Ling, 1970), a process th at takes about 5 weeks (Sch effer and Slipp, 1944, as ci ted in Ashwel l-Erickso n and Elsner, 1981). Dur ing t heir molt , they spend m ore time hauled and exhibit a slow er metabolic r ate (e.g., 83 percent of premolt levels), which decreases th eir food r equir ements (Ashwel l-Erickso n and Elsner, 1981). After molting, harbor seals increase th eir fat r eserves (an d weight) fo r the wint er and e arly sp ring; metabolic r ates al so might be low ered dur ing t his ti me to cons erve energy (Renouf , 1989).
Breeding acti vi ties and social organization. The timing of r eproduc tion in ha rbor seals var ies wi th locat ion. Mating a nd pupping a re init iated earlier in the year in more
2-277 Harbor Seal

�
sout hern lati tude s, but wi thin popula tions , breeding is syn chronize d (Hoov er, 1988; Slater and Markowitz, 1983). Harbor seals may fo rm large breeding a ggr egations on la nd in a reas where food r esour ces ar e plenti fu l (Slater and Markowitz, 1983); however , pupping acti vi ties are not restr icted to large, discr ete rooker ies (Pi tch er and Calkins, 1979). Mating occurs soon after weaning, whic h is 3 to 6 weeks after birth (Ashwel l-Erickso n and Elsner, 1981). It is l ikely th at harbor seals are promiscuous (Pi erot ti and Pierot ti , 1980), although there is so me evide nce that they are mildly poly gynous , wi th males defending t erritories at the haul-out si tes (Boulv a and Mc Laren, 1979; Perry, 1989; Slater and Markowitz, 1983). Follow ing mating, implantation is delayed for 1.5 to 3 months, during w hic h t ime the femal e molt s (Bigg, 1969a; Hoover , 1988; Pitch er and Calkins, 1979). At other times o f th e year , harbor seals also can be found in gr oups of 30 to 80 in s ome haul-out ar eas (Hoover , 1988).
Home range and r esour ces. Harbor seals generally inha bit highly produc tive coastal areas, wi th upwelling o cean currents th at bring nut rients to the surface s uppor ting abunda nt mar ine lif e (e.g., the Calif ornia cu rrent system, the Gulf of Alaska, and the Gulf of Maine; Ronald et al., 1982). Harbor seals also requir e adequate places to haul out, and their dis tr ibut ion is inf luenced by th e availa bilit y of suit able si tes (Boulv a and Mc Laren, 1979). In general, seals stay n ear particular haul-out si tes wi th only loc al movemen ts (Brown and Mate, 1983; Pitch er and Calkins, 1979; Slater and Markowitz, 1983). Haul-out patter ns are determined by sever al facto rs, including weath er, tidal patter n, time of day, seaso n, and human proximity (Sl ater and Markowitz, 1983). Harbor seals are considered fairly sedentary, wi th indiv idua ls show ing year -round s ite fide lit y, al though s ome seaso nal movemen t asso ciated wi th pupping a nd long- distan ce mo vemen ts ar e recorded (Pitch er and Calkins, 1979; Slater and Markowitz, 1983). Data on likely daily or monthly fo raging distan ces ar e lack ing.
Popul ation de nsity . Harbor seals are found pr inc ipa lly in c oastal areas wi thin 2 0 km of sh ore; they ten d to concentrate in estu aries and protected water s (Hoover , 1988). Their dis tr ibut ion is highly p atch y, and local popula tion de nsit ies in ha ul-out ar eas wi th favo rable food r esour ces n earby can be quit e high ( Pitch er and Ca lk ins , 1979).
Popul ation dy namics. Females are sexu ally matu re by 3 to 5 years of age, whereas males are sexu ally matu re later , at 4 to 6 years of age (Boulv a and Mc Laren, 1979; Pitch er and Calkins, 1979). Females only produc e one pup pe r year (Hoover , 1988). Three major causes o f preweaning pup m ortalit y are stillbir th, deser tion by th e mother, and s hark kills (Boulv a and Mc Laren, 1979). Mortal ity fr om birth to 4 years of age was esti mated to be 74 percent fo r femal es and 79 percent fo r males in one stu dy, after which it remained at about 10 percent per year (Pitch er and Calkins, 1979). Life exp ectan cy fo r harbor seals is about 30 years (Newby, 1978).
Simi lar species (fr om general references)
� The ringe d seal (Phoca hispida) is smalle r (1.4 m lengt h; weight to 90 kg) than the harbor seal and inha bit s colde r water s. It feed s mainly on m arine inver tebrates.
2-278 Harbor Seal

�
�
�
� The harp seal (Phoc a groenlandi cus) (1.8 m; weight to 180 kg) inha bit s deep, icy water . It ranges fr om th e Arcti c Atlantic so uth to Huds on Ba y; it is only rarely found f urther sout h. It feeds on macr opla nkton a nd f ish.
� The largha or spotted seal (Phoc a largha) (1.5 m) is a cl osely related species that unti l recently was co nsidered a subspecies of th e harbor seal . Its character isti cs ar e compared wi th those of th e harbor seal unde r Selected species.
� The ribbon seal (Phoca fasci ata) (1.6 m; males to 90 kg, femal es to 76 kg) lives n ear pack i ce in the Bering Sea and feeds on botto m inver tebrates, fish, and oc topus and s quid.
General references
Ashwel l-Erickso n and Elsner (1981); Burt and Grossen heider (1980); Hoover (1988); Pitch er and Calkins (1979); Ronald et al. (1982).
2-279 Harbor Seal

Harbor Seal ( Phoca vitulina )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion Reference No.
Range or
Body Weight A M (> 7 yrs) 84.6 ± 11.3 SD Gulf of Alaska Pitcher & Calkins, 1979 (kg) A F (> 7 yrs) 76.5 ± 17.7 SD
J M 2 yrs 49 Aleut ian Ridge and Pr ibilof Ashw ell-Erickson & El sner , 1 J M 4 yrs 70 Islands, Ber ing Sea, Alaska 1981 J M 6 yrs 84 A M 8 yrs 95 A M 12 yrs 110 A M 16 yrs 120 A M 24 yrs 124
J F 2 yrs 40 J F 4 yrs 56 J F 6 yrs 67 A F 8 yrs 76 A F 12 yrs 90 A F 16 yrs 101 A F 24 yrs 112
neonat e M 12.0 ± 0.51 SE Alaska Pitcher & Calkins, 1979 neonat e F 11.5 ± 0.31 SE
at weaning B 24.0 British Col umbia, Canada Bigg, 1969a 2
Pup Growth bi rth to Gulf of St. Lawrence/ island Rosen, 1989 Rate (g/day) weaning marin e
M 520 F 790
Metabolic J B resting 7.3 Califo rnia/lab Davis et al., 1985 3 Rate A F rest ing 6.6 (lO /kg-day)2
2-280 H
arbo
r Seal

Harbor Seal ( Phoca vitulina )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean Locat ion Reference No.
Range or
Metabol ic Rate 1 to 4 yrs ol d/ 57.5 Bering Sea, Alaska Ashw ell-Erickson & El sner , (kcal /kg-day) basal 1981
A F basal 24.3 est imated 4 A M basal 22.4
A F f ree-living 131 (57 - 300) est imated 5 A M free-living 129 (56 - 296)
Food Ingest ion A B 0.05 e Canada/marine Boul va and M cLaren, 1979 Rate (g/g-day)
A B 0.06 to 0.08 review of sever al studi es Ashw ell-Erickson & El sner , A F lact ./gest . 0.10 1981
J B 1st year 0.13 Ber ing Sea (1 harbor & Ashw ell-Erickson & El sner , 1 spot ted seal ) 1981
Water A B 0.0048 0.0028 - 0.0091 seawater ingest ion (most Depocas et al., 1971 Ingest ion water obt ained f rom food) Rate (g/g-day) A B 0.064 est imated 6
Inhal ation A M 18.6 est imated 7 Rate (m /day) A F 17.23
Surface Ar ea A M 19,620 est imated 8 (cm ) A F 18,3802
2-281 H
arbo
r Seal

Harbor Seal ( Phoca vitulina )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
walleye pollo ck 3.7 27.3 32.2 1.3 Washington/ Everitt e t al., 1981 Engl ish sol e 37.0 0.0 27.0 0 coast al island shi ner perch 0.0 0.0 0.5 63.6 Paci fic her ring 0 54.6 3.9 28.6 (% of total otol iths Paci fic cod 0 0 10.1 0 recover ed f rom scat rex sol e 37 9.1 2.9 0 samples) Paci fic tomcod 3.7 0 4.7 0 rockf ish 3.7 0 4.7 0 Dover sol e 3.7 0 3.4 2.6 Petrale sol e 7.4 0 1.8 0 other fish 3.8 9.0 8.8 3.9
oct opus 17.6 17.7 30.4 Kodi ak Island, Alaska/ Pitcher & Calkins, 1979 salmon 5.4 0.0 0.0 coast al marine capel in 20.3 4.8 5.4 Paci fic cod 6.8 8.1 10.7 (% frequency of occur rence; walleye pol lock 12.2 9.7 14.3 stomach cont ents) Paci fic sandl ance 4.1 21.0 0.0
squi d & oct opus 20 Gul f of Alaska/ Pitcher, 1980 shr imp, crabs 3.7 coast al marine herring 6.4 salmoni ds 4.4 (% wet vol ume; stomach osm erids 22.5 cont ents) cod, tomcod, 26.0
walleye, pol lock all seasons com bined other 14.1
2-282 H
arbo
r Seal
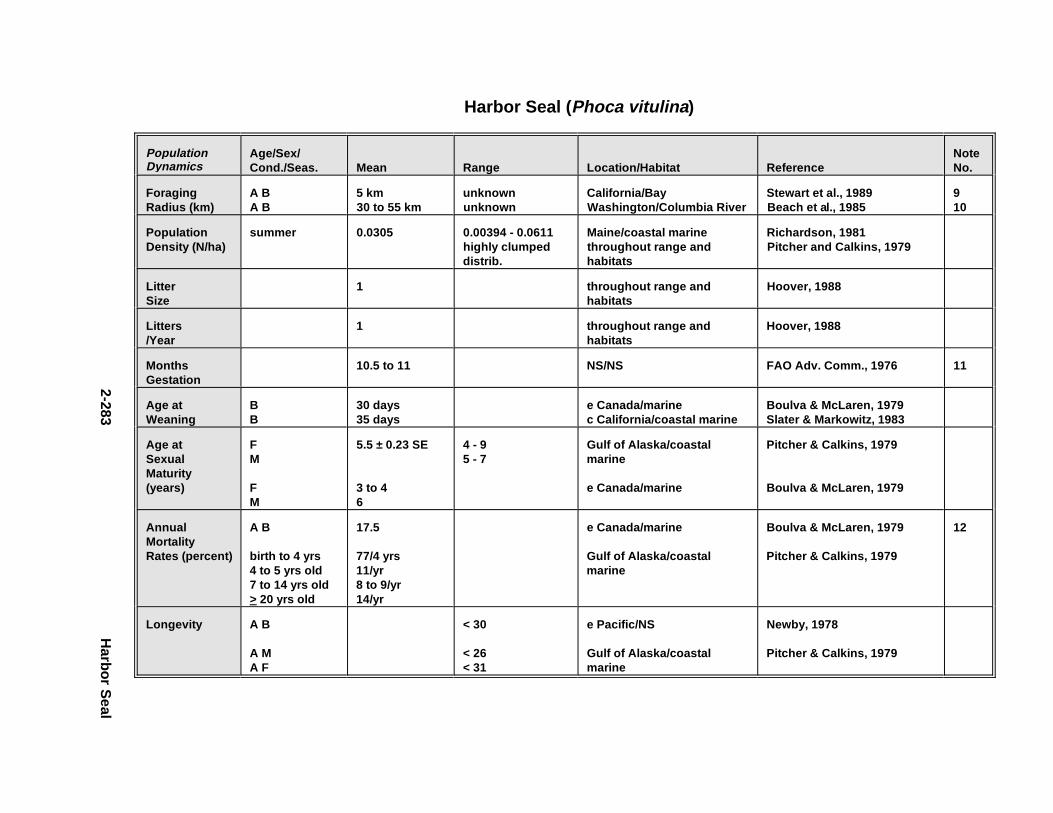
Harbor Seal ( Phoca vitulina )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Foraging A B 5 km unknown Califo rnia/Bay Stewart e t al., 1989 9 Radius (km) A B 30 to 55 km unknow n Washi ngt on/Columbia River Beach et al., 1985 10
Popul ation summer 0.0305 0.00394 - 0.0611 Maine/coast al marine Richar dson, 1981 Densi ty (N/ha) highl y clumped throughout range and Pitcher and Cal kins, 1979
dist rib. habi tats
Li tter 1 throughout range and Hoover , 1988 Size habi tats
Li tters 1 throughout range and Hoover , 1988 /Year habi tats
Mont hs 10.5 to 11 NS/NS FAO Adv. Comm., 1976 11 Gestation
Age at B 30 days e Canada/marine Boul va & McLaren, 1979 Weaning B 35 days c Cal ifornia/coast al marine Slater & Markowitz, 1983
Age at F 5.5 ± 0.23 SE 4 - 9 Gul f of Alaska/coast al Pitcher & Calkins, 1979 Sexual M 5 - 7 marine Maturity (years) F 3 to 4 e Canada/marine Boul va & McLaren, 1979
M
Annual A B 17.5 e Canada/marine Boul va & McLaren, 1979 12 Morta lity Rates (percent ) bi rth to 4 yrs 77/4 yrs Gul f of Alaska/coast al Pitcher & Calkins, 1979
4 to 5 yrs ol d 11/yr marine 7 to 14 yrs ol d 8 to 9/yr > 20 yrs ol d 14/yr
Longevi ty A B < 30 e Paci fic/NS Newby , 1978
A M < 26 Gulf of Alaska/coast al Pitcher & Calkins, 1979 A F < 31 marine
Range
6
2-283 H
arbo
r Seal
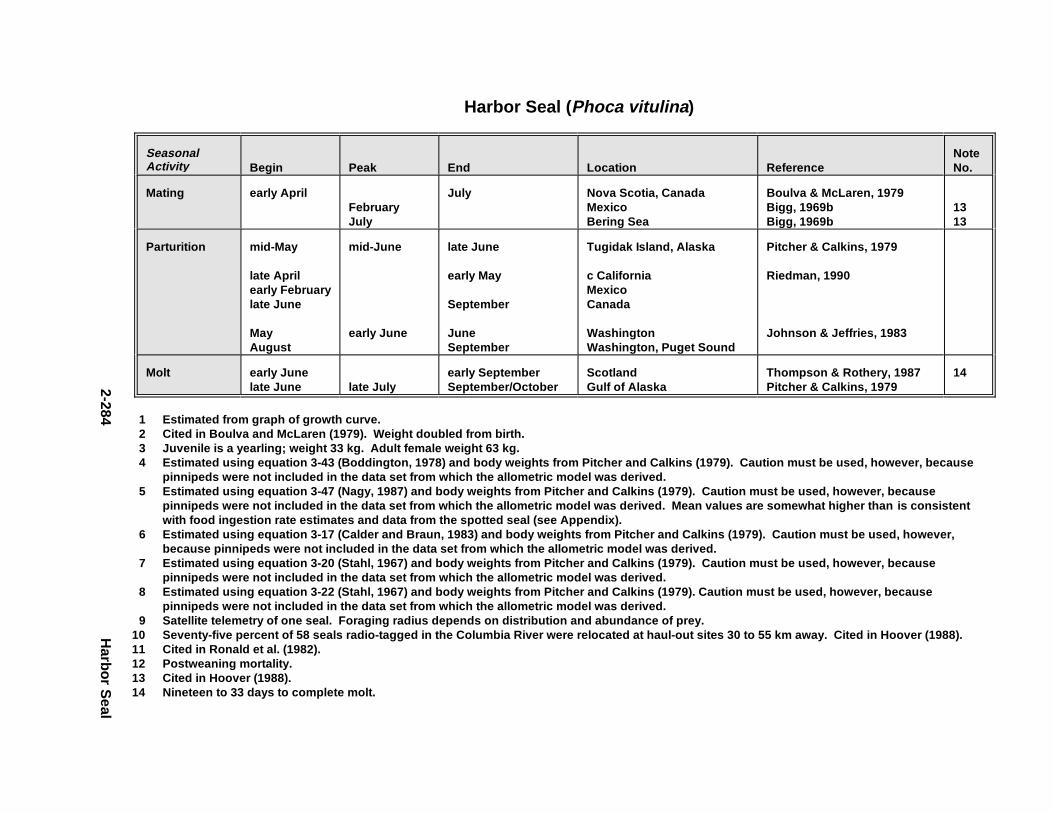
Harbor Seal ( Phoca vitulina )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating early Apr il July Nova Scot ia, Canada Boul va & McLaren, 1979 February Mexico Bigg, 1969b 13 Jul y Ber ing Sea Bigg, 1969b 13
Partur ition mid-May mid-June late June Tugi dak Island, Alaska Pitcher & Calkins, 1979
late Apr il early May c California Riedman, 1990 early February Mexico late June September Canada
May early June June Washi ngt on Johnson & Jef fries, 1983 August September Washi ngt on, Puget Sound
Molt early June early September Scot land Thom pson & Rot hery, 1987 14 late June late Jul y September/October Gul f of Alaska Pitcher & Calkins, 1979 2-284
Harb
or S
eal
1 Est imated f rom graph of growt h cur ve. 2 Cited in Boul va and M cLaren (1979). Weight doubl ed f rom birth. 3 Juvenile is a yearlin g; weight 33 kg. Adult female weight 63 kg. 4 Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom Pitcher and Cal kins (1979). Caution m ust be used, however , because
pinni peds wer e not included i n the data set from whi ch the allometric model was der ived. 5 Est imated usi ng equat ion 3-47 (Nagy, 1987) and body weight s f rom Pitcher and Cal kins (1979). Caution m ust be used, however , because
pinni peds w ere not included i n the data set from which the allometric model was der ived. Mean val ues ar e som ewhat higher than is consis tent wi th food i ngest ion r ate est imates and dat a from the spot ted seal (see Appendi x).
6 Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Pitcher and Cal kins (1979). Caution m ust be used , how ever, because pi nni peds w ere not included i n the data set from which the allometric model was der ived.
7 Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Pitcher and Cal kins (1979). Caution m ust be used, however , because pinni peds wer e not included i n the data set from whi ch the allometric model was der ived.
8 Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Pitcher and Cal kins (1979). Caut ion m ust be used, however , because pinni peds wer e not included i n the data set from whi ch the allometric model was der ived.
9 Satellite telemetry of one seal . Foraging r adius depends on di stribut ion and abundance of prey. 10 Sevent y-five per cent of 58 seals radio-tagged i n the Columbia River were relocated at haul -out si tes 30 to 55 km away. Cited in Hoover (1988). 11 Cited in Ronal d et al. (1982). 12 Post weaning m ortality. 13 Cited in Hoover (1988). 14 Nineteen to 33 days to com plete molt.

References (including A ppendix)
Allen, S. G.; Huber, H. R.; Ribic , C. A.; et al. (1989) Popula tion dy namics of harbor seals in the Gulf of th e Farallone s, Calif ornia . Calif . Fish Game 75: 224-232.
Angell-James, J. E.; Elsner, R.; de Burgh Daly, M. (1981) Lung inf lation: effects o n heat rate, respiration, and v agal affer ent acti vi ty i n seals. Am. J. Physiol. 240: H190-H198.
Ashwel l-Erickso n, S.; Elsner, R. (1981) The energy co st o f fr ee existen ce fo r Bering S ea harbor and spotted seals. In: Hood, D. W.; Calder, J. A., eds. The Easter n Bering S ea shelf: oceanogr aphy and r esour ces. v. 2. Washingt on, DC: Departmen t of Commer ce; pp. 869-899.
Ashw ell-Erickson, S.; Elsner, R.; Wartzol, D. (1979) Metabolis m and nut rition of Bering S ea harbor and spotted seals. Proc. Alaska Sci . Conf . 29: 651-665.
Beach, R.J.; Geiger, A.; Jeffr ies, S. J., et al . (1985) Marine mammal s and their interact ions wi th fisheries of th e Colum bia River and a djacent water s, 1980-1982. Seattl e, WA: Third Ann. Rept. to U.S. Dept. Commer ce, NOAA, Natl . Mar. Fish. Serv.
Bigg, M. A. (1969a) The harbour seal in British Colum bia. Fish. Res. Board Can.; Bull. 172.
Bigg, M. A. (1969b) Cline s in t he pupping seaso n of th e harbour seal , Phoca vi tu lina. J. Fish. Res. Board Can. 26: 449-455.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Boulv a, J.; McLaren, I. A. (1979) Biology of th e harbor seal , Phoca vi tu lina, in easter n Canada. Quebec, Canada: Fish. Res. Board Can. Bull. 200.
Brown, R. F.; Mate, B. R. (1983) Abunda nce, movemen ts, and feeding ha bit s of harbor seals, Phoca vi tu lina, at Nearts an d Tillamook Bays, Oregon. U.S. Natl . Mar. Fish. Serv. Fish. Bull. 81: 291-301.
Brown, R. F.; Jeffr ies, S. J.; Harvey, J. T. (1989) Seasonal abunda nce and w int er feeding ecology of harbor seals in the Colum bia River (abstr act). In: 8th Biennia l Conf erence on t he Biology of Marine Mammal s; Decemb er 7-11, 1989; Paci fic Grove, CA; p. 9.
Bryden, M. M. (1972) Growth and development of mar ine mammal s. In: Harrison, R. J., ed. Func tiona l anatomy of mar ine mammal s. New York, NY: Academic Press; pp. 2-79.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
2-285 Harbor Seal

Craig, A. B., Jr .; Pasch e, A. (1980) Respiratory physiology of fr eely div ing ha rbor seals (Phoca vi tu lina). Physiol. Zool. 53: 419-432.
da Silva, J.; Neilson, J. D. (1985) Limitat ions of us ing ot olit hs r ecovered in scats to esti mate p rey co nsumption in seals. Can. J. Fish. Aquat. Sci . 42: 1439-1442.
Davis, R. W.; Willi ams, T. M.; Kooyman , G. L. (1985) Swimming metabolis m of year ling a nd adult harbor seals (Phoca vi tu lina). Physiol. Zool. 58: 590-596.
Depocas, F.; Hart, J. S.; Fisher, H. D. (1971) Sea water drinking a nd water flux in star ved and in fed harbor seals, Phoca vi tu lina. Can. J. Physiol. Pharmaco l. 49: 53-62.
Ever itt, R. D.; Gearin, P. J.; Skidmore, J. S.; et al . (1981) Prey i tems o f harbor seals and California sea lions in Puget Sound, Washingt on. Mur relet 62: 83-86.
Food a nd Agricult ure Organization ( FAO) of th e Unit ed Nations , A. C. on Marine Resour ces Research. (1976) Mammal s in the seas. Ad Hoc Group III on seals and mar ine otter s, draft r eport. In: Symp osium: Scienti fic co nsultat ion on m arine mammmal s; Augus t 13 to Septemb er 9, 1976; Food a nd Agric. Advis. Comm. Mar. Resour . Res., Mar. Mammal , Sci . Cons ult . Organ. of U.N., Bergen, Norway.
Harkonen, T. J. (1988) Food- habitat r elations hip of harbour seals and black co rmorants i n Skagerrak and Katteg at. J. Zool. (London) 214: 673-681.
Harvey, J. T. (1989) Assessmen t of er rors asso ciated wi th harbour seal (Phoca vi tu lina) faecal sampling. J. Zool. (Lond. ) 219: 101-111.
Hoover , A. A. (1988) Harbor seal , Phoca vi tu lina. In: Lentfer , J. W., ed. Selected mar ine mammal s of Alaska: species acc ounts wi th research and management recommenda tions . Washingt on, DC: Marine Mammal Commi ss ion; pp. 125-157.
Innes, S.; Lavigne , D. M.; Earle, W. M.; et al. (1987) Feeding r ates o f seal s and whales. J. Anim. Ecol. 56: 115-130.
Irving, L. (1972) Arcti c l ife of b irds and mammal s including m an. New York, NY: Springe r-Verlag.
Johns on, M. L.; Jeffr ies, S. J. (1977) Popula tion evalua tion of th e harbour seal (Phoc a vi tu lin a ric hardii ) in the water s of th e state o f Washingt on. U.S. Mar. Mammal Comm. ; Rep. MMC-75/05.
Johns on, M. L.; Jeffr ies, S. J. (1983) Popula tion biology evalua tion of th e harbor seal (Phoca vi tu lina richardii) in the water s of th e State o f Washingt on: 1976-1977. Tacoma, WA: Univ ersi ty of Puge t Sound; MMC-76/25.
Jones, R. E. (1981) Food ha bit s of smal ler mar ine mammal s fr om northern California. Proc. Calif. Acad. Sci . 42: 409-433.
2-286 Harbor Seal

Klink hart, E. G. (1967) Birth of a harbor seal pup. J. Mammal . 48: 677.
Lawson, J. W.; Renouf , D. (1987) Bonding a nd weaning in ha rbor seals, Phoca vi tu lina. J. Mammal . 68: 445-449.
Ling, J. K. (1970) Pelage and molting in w ild a nim als wi th special reference to aquatic forms. Q. Rev. Bio. 45: 16-54.
Ling, J. K. (1974) The integument of mar ine mammal s. In: Harrison, R. J., ed. Func tiona l anatomy o f mar ine mammal s: v. 2. New York, NY: Academic Press; pp. 1-44.
Lowry, L. F.; Frost, K. J. (1981) Feeding a nd t rophic r elations hips of phoc id seals and walruses i n the Easter n Bering Sea. In: Hood, D. W.; Calder, J. A., eds. The Easter n Bering Sea shelf: oceanogr aphy and r esour ces: v. 2. Washingt on, DC: Departmen t of Commer ce; pp. 813-824.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Mono. 57: 111-128.
Newby, T. C. (1973) Observat ions on t he breeding be havior of th e harbor seal in the State of Washingt on. J. Mammal . 54: 540-543.
Newby, T. C. (1978) Paci fic harbor seal . In: Haley, D., ed. Marine mammal s of easter n north Paci fic and arcti c water s. Seattl e, WA: Paci fic Search Press; pp. 184-191.
Payne, P. M.; Schneider, D. C. (1984) Yearly ch anges in abunda nce of harbor seals, Phoc a vi tu lina, at a wi nter haul-out si te in Massach usetts. Fish. Bull. 82: 440-442.
Payne, P. M.; Selzer, L. A. (1989) The distr ibut ion, abunda nce and s elected prey of th e harbor seal , Phoca vi tu lina concolor, in s out hern New Engla nd. Mar. Mammal . Sci . 5: 173-192.
Perez, M. A. (1990) Review of mar ine mammal s popula tion a nd pr ey inf ormation f or Bering Sea ecosystem st udie s. Washingt on, DC: U.S. Dept . Commer ce, Nat. Oceanic Atm. Admin. , Nat. Mar. Fish. Serv.; NOAA Tech. Mem. NMFS F/NWC-186.
Perry, E. (1989) Evidence fo r poly gyny in ha rbour seals, Phoca vi tu lina. In: 8th Biennia l conf erence on t he biology of mar ine mammal s; Decemb er; Paci fic Grove, CA.
Pierotti , R.; Pierotti , D. (1980) Effects o f cold c limate o n the evolut ion of pinnipe d br eeding systems. Evolut ion 3 4: 494-507.
Pitch er, K. W. (1977) Popula tion pr oduc tivi ty and f ood ha bit s of harbor seals in the Prince Willia m Sound- Coppe r River Delta area, Alaska. Fina l repor t to U. S. Marine Mammal Commission No. MMC-75103. USDC NTIS. PB-226 935.
Pitch er, K. W. (1980) Food of th e harbor seal , Phoca vi tu lina richardsi , in t he Gulf of Alaska. U.S. Natl . Mar. Fish. Serv. Fish. Bull. 78: 544-549.
2-287 Harbor Seal

Pitch er, K. W.; Calkins, D. G. (1979) Biology of th e harbor seal , Phoca vi tu lina richardsi , in the Gulf of Alaska. Fina l repor t. Outer Cont inental Shelf Environm ental Assessmen t Program Research Unit 229, Cont ract No. 03-5-002-69.
Pitch er, K. W.; McAllister , D. C. (1981) Movemen ts an d haulout behavior of r adio -tagged harbor seals, Phoca vi tu lina. Can. Field-Nat. 95:292-297.
Power , G.; Gregoir e, J. (1978) Predation by fr eshwater seals on the fish communit y of lower Seal Lake, Quebec. J. Fish. Res. Board Can. 35: 844-850.
Renouf , D. (1989) Weight inc reases i n harbour seals in spite of reduced food in take an d heightened thermal demands: adjustab le metabolis m? In: 8th Biennia l Conf erence on t he Biology of Marine Mammal s; Decemb er; Paci fic Grove, CA.
Richardson, D. T. (1973) Distr ibut ion a nd abunda nce of harbor and gr ay seals in Acadia Nationa l Park. Fina l repor t to Nationa l Park Service and Maine Departmen t of Sea and Shore Fisheries. State o f Maine Cont ract No. MM4AC009.
Richardson, D. T. (1981) Feeding ha bit s and popula tion s tudie s of Maine 's harbor and gr ay seals. Natl . Geogr. Soc. Res. Rep. 13: 497-502.
Riedman, M. (1990) The pinnipe ds: seals, sea lions , and w alruses. Berkeley, CA: Universi ty of California Press.
Roffe, T. J.; Mate, B. R. (1984) Abunda nces an d feeding ha bit s of pinnipe ds in t he Rogue River, Oregon. J. Wildl. Manage. 48: 1262-1274.
Ronald, K.; Selley, J. ; Healey, P. (1982) Seals. In: Chapman, J. A.; Feldhammer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 769-827.
Rosen, D. A. (1989) Neonatal growth rates an d behaviour in t he Atlant ic harbour seal , Phoca vi tu lina (abstr act). In: 8th Biennia l Conf erence on t he Biology of Marine Mammal s; Decemb er 7-11, 1989; Paci fic Grove, CA; p. 57.
Scheffer , V. B.; Slipp, J. W. (1944) The harbor seal in Washingt on state. Am. Midl. Nat. 32: 373-416.
Schneider, D. C.; Payne, P. M. (1983) Facto rs affect ing ha ul-out of harbor seals at a si te in southeaster n Massach usetts. J. Mammal . 64: 518-520.
Selzer, L. A.; Early , G.; Fior elli, P. M.; et al. (1986) Stranded animals as indi cato rs of prey ut iliza tion by harbor seals, Phoca vi tu lina concolor, in s out hern New Engla nd. Fish. Bull. 84: 217-220.
Shaffer , K. E. (1989) Seasonal and size var iations in di ets o f harbor seals, Phoca vi tu lina (abstr act). In: 8th Biennia l Conf erence on t he Biology of Marine Mammal s; Decemb er 7-11, 1989; Paci fic Grove, CA; p. 62.
2-288 Harbor Seal

Slater , L. M.; Markowitz, H. (1983) Spring popula tion t rends in Phoca vi tu lina richardi in t wo central California coastal areas. Calif. Fish Game 69: 217-226.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stewar t, B. S.; Leatherwood, S.; Yochem, P. K.; et al. (1989) Harbor seal track ing a nd telemetr y by sate llite. Mar. Mamm. Sci . 5: 361-375.
Stutz, S. S. (1966) Moult and pe lage patter ns in the Paci fic harbor seal , Phoca vi tu lina [master 's th esis]. Vancouver , Canada: Univ ersi ty of Br it ish Colum bia.
Thompson, P; Rothery, P. (1987) Age and sex d iffer ences i n the timing of moult in t he commo n seal , Phoca vi tu lina. J. Zool. (London) 212: 597-603.
Wilson, S. C. (1978) Social organization a nd be havior of harbor seals Phoca vi tu lina conc olor in Maine. Fina l repor t to Marine Mammal Commi ss ion, Cont ract No. GPO PB 280-3188. NTIS PB 280 188.
2-289 Harbor Seal

2.2.7. Deer Mouse (deer and w hite-footed m ice)
Order Rodent ia, Family Murid ae (Genus Peromyscu s).h New wo rld mice (fami ly Muridae) are smal l, ground- dwelling r odents that liv e in a large var iety o f habitats i ncluding woodla nds , prairies, rocky h abitats, tundr a, and d eser ts. All are noctu rnal and are preyed on by owls, hawks, snakes, and carnivorous mammal s. Most sp ecies eat p rimar ily seed s, but so me also regula rly eat smal l inver tebrates. Many sp ecies sto re food. The genus Peromyscu s is th e most wi despread and geographical ly var iable of North Amer ican rodents (MacMille n and Garland, 1989).
Selected species
The deer mouse (Peromyscu s man iculatus) is pr imarily graniv orous and ha s the widest g eographic dis tr ibut ion of an y Peromyscu s species (Milla r, 1989; Brown and Zeng, 1989). It is resident and commo n in nearly ever y dry-land habitat wi thin it s range, inc luding alpine tundr a, conif erous and de ciduous fo rest, and grassl ands as wel l as deser ts. There are many recognize d subs pecies or races o f th e deer mouse asso ciated wi th differ ent loc ations or ins ular habitats, including artemi siae, auster us, bairdii, b alaclavae, blandus , borealis , carli, c ooledgei, gambelii, g racilis , labecula, maniculatus, oreas, nebrascen sis, nubi ter rae, rufinus , and sonoriensis (MacMille n and Garland, 1989; Milla r, 1982)
Body size. Deer mice range from 7.1 to 10.2 cm i n length, wi th a 5.1 to 13 cm tai l, and adult s weigh f rom 15 to 35 g (Bur t and Grossen heider, 1980; see tab le). Body size var ies so mewh at among popula tions and s ubs pecies throughout th e species' range. Body weight al so var ies seaso nally, being low er in autumn and w int er and a few g rams highe r in spring a nd s ummer (Zegers and Merritt, 1988). There may (Fleharty et al ., 1973) or may n ot (Milla r and S chi eck, 1986) be seaso nal differ ences i n fat co ntent.
Habitat. Deer mi ce inha bit nearly all t ypes of dry-land ha bitats wi thin t heir range: short-grass p rairies, grass-sag e communit ies, co astal sage scr ub, sand dunes, wet prairies, upla nd m ixed and c edar fo rests, deciduous fo rests, ponde rosa pine fo rests, other coniferous fo rests, mixed deciduous-ever green forests, junipe r/piñon f orests, and other habitats ( Holbr ook , 1979; Kaufman and Kaufman , 1989; Ribble and Samso n, 1987; Wolff and Hurlbutt, 1982). Few st udies have found m icrohabitat featu res th at dist inguis h the deer mouse, and some stu dies have co me to differ ent co nclusions r egarding ha bit at str uctu re preferences (Ribble and Samso n, 1987). For examp le, Vicker y (1981) found t hat deer mice appeared to prefer areas wi th moderate to heavy g round a nd m id-sto ry co ver to more open gr ound a reas, whereas others have found m ore deer mi ce in m ore open than in more vegetated areas (see K aufman and Kaufman , 1989).
Food ha bi ts . Deer mi ce are omniv orous and highly oppor tunis tic, whic h leads to substan tial regiona l and seaso nal var iation in t heir die t. They eat principally seed s, arthropods , some gr een vegetation, root s, fr uit s, and f ungi a s availa ble (Johns on, 1961; Menhusen, 1963; Whitaker , 1966). The nonseed plant mater ials provide a signif icant
hPeromyscu s is considered a member of the family Criceti dae by some mammalogists.
2-291 Deer Mous e

�
�
propor tion of th e deer mous e's da ily water requir ements (MacMille n and Garland, 1989). Food dig estibilit y and assi mi lation f or mo st of th eir die t have been esti mated to be as high as 88 percent (Mont gom ery, 1989). Deer mice may cach e food dur ing t he fall a nd w int er in the more northern parts o f th eir range (Barry, 1976; Wolff, 1989). They are noctu rnal and emerge shortly after dark to forage for sever al hour s (Marten, 1973).
Temperature regulation. The deer mouse has a meta bolic r ate about 1. 3 times highe r th an the other sp ecies in t he genus (M acMille n and Garland, 1989; Morris and Kendeigh, 1981; see tab le). Its meta bolic r ate is su bstan tially highe r in w int er th an in summer (Morris and Kendeigh, 1981; Stebbins , 1978; Zegers and Merritt, 1988). Outsi de the thermone utral zone (25 to 35�C), metabolic r ate var ies acco rding t o the follow ing equation:
VO2 = 0.116 - 0.003(Ta) + 0.0304 (V0.5)
where VO2 = volume oxyg en consumed (ml/g-min); Ta = ambient temp erature; and V = wind speed (Chappell and Holscl aw, 1984). Deer mice can enter torpor (body temp erature, 19 to 30�C) to reduce meta bolic d emands in the winter and also in response to brief fo od shortages (Tannenbaum and Pivorun, 1988, 1989). The deer mouse uses nons hiv ering thermogenesis (NST) to quickl y awaken from to rpor and to maintain body temp erature dur ing t he wint er (Zegers and Merritt, 1987). The deer mouse may b urrow in soils to assi st thermoregula tion; one study measu red the burrow dimensions to be 24 cm de ep (range 13 to 50 cm) an d 132 cm long ( range 30 to 470 cm) (Reynolds and W akki nen, 1987).
Breeding acti vi ties and social organization. The dur ation of th e reproduc tive seaso n var ies wi th lati tude and longit ude acco rding t o the regression e quation:
Y = -33.0 + 2.79 X + 0.0748 Z - 0.0370 X2
where Y = dur ation of th e breeding seaso n in weeks, X = lati tude, and Z = longit ude (r = 0.58; Milla r, 1989). Lactat ing femal es have longe r gestat ion pe riods th an nonl actati ng femal es. Newborn deer mice are highly al tr ic ial (Layne, 1968). Several studies have indi cated that daily food c ons umpt ion inc reases o ver 15 percent during e arly pregnancy and m ore than double s dur ing l actat ion ( Glazier, 1979; Milla r, 1975, 1978, 1979, 1982, 1985; Milla r and Inne s, 1983; Stebbins , 1977). Deer mice are promiscuous ; in one study , 19 to 43 percent of l itter s resulted from mu ltiple insemina tions (Bir dsall and Nash, 1973, as ci ted in Milla r, 1989).
Home range and r esour ces. Deer mice ten d to occu py mo re than one nest si te, most frequent ly in t ree hollow s up t o 8 m from th e ground ( Wolf f and Du rr, 1986) but also among t ree root s and unde r rocks an d logs (W olf f and Hur lbut t, 1982; Wolff, 1989). At low densit ies, hom e ranges are maint ained by mu tual avoida nce, but at highe r dens it ies, femal es may d efend a core area or ter ritory (Wolff, 1989). The home range of femal e deer mice encompasses b oth their foraging a reas and their nests. Male home ranges are larger and over lap the home ranges of man y femal es (Cranford, 1984; Taitt, 1981; Wolff, 1985a, 1986; Wolff et al ., 1983).
2-292 Deer Mous e

�
�
�
�
�
Popul ation de nsity . Popula tion de nsity var ies cons iderably over sp ace and time and is often posi tively co rrelated wi th food a bunda nce (Taitt, 1981; Wolff, 1989), moistu re content of p lants (B owers and Smith, 1979), and vegetati ve co ver (van Horne, 1982) as wel l as seaso n (Mont gom ery, 1989; Taitt, 1985). Interspeci fic co mpeti tion a lso can play a role in determining popula tion de nsit ies (Kaufman and Kaufman , 1989).
Popul ation dy namics. Although la bor atory and f ield s tudie s have demonstr ated that femal es can produce th eir fi rst l itter by 3 mo nths of age, femal es of th e more northern popula tions do not matu re unde r natural condit ions unt il t he spring after the year of th eir birth. First l itter s are consisten tly smal ler than subsequent l itter s (Milla r, 1989), and lati tude and elevat ion e xpla in a signif icant amount of th e var iation in li tter size among P. maniculatus popula tions (Smith and McGinnis , 1968, as ci ted in Milla r, 1989). Milla r (1989) esti mated the relations hip b etween litter size and lati tude and longit ude to be
Y = -1.62 + 0.0103X + 0.106Z + 0.0004X2 - 0.0005Z2
where Y is th e mean litter size; X, the lati tude; and Z, the longit ude. The largest l itter s are produced in northwester n North Amer ica. Pups wean within a bout 3 weeks, and femal es may h ave up to four litter s per year in the more southern parts o f th e species' range (Milla r, 1989). Mortal ity r ates ar e high, and m ost de er mi ce liv e for less th an 1 year (Milla r and Innes, 1983).
Simi lar species (fr om general references)
� The cactu s mo use (Peromyscu s eremicus), almost th e same si ze as th e deer mous e (8.1 to 9.1 cm; 17 to 40 g), is found only in low d eser ts o f th e extr eme southwest an d Mexico. It may feed on green vegetat ion, seeds, and berries and can cl imb trees fo r food.
� The Calif ornia mous e (Peromyscu s cal ifornicus) (9.6 to 11.7 cm; 42 to 50 g) is found in s out hwester n California and lives am ong oaks an d dense chaparral. It sto res aco rns in nests mad e of twi gs and sti cks.
� The canyon mouse (Peromyscu s cr initus) (7.6 to 8.6 cm) i s l imi ted to the wester n United States. It l ives i n rocky can yons and on lava-co vered slope s, nesting a mong r ocks.
� The oldf ield m ous e (Peromyscu s polionot us), smal ler than the deer mouse (4.1 to 6.1 cm), is l imi ted to the extr eme so utheaster n United States, where it inha bit s sandy b eaches and fields and feeds on seeds and berries. Females may b e ter ritorial during t he breeding seaso n.
� The white-fo oted mouse (Peromyscu s leucopus ) is approximatel y th e same size as th e deer mouse (9.1 to 10.7 cm; 14 to 31 g). Its r ange exten ds north into Canada and west to Arizona but does not exten d as far north and west as th e deer mouse's range. Like th e deer mouse, the white-fo oted mouse's diet co nsists ma inly of ar thropods , seeds, and other vegetat ion, and it usually nests o ff th e ground. It is mo st abunda nt in ha bitat th at includes a
2-293 Deer Mous e

�
�
canopy , such as br ushy fi elds and de ciduous woodlot s in nor thern regions and riparian areas and ravines in prairie and semi deser t regions .
� The cotto n mouse (Peromyscu s gossyp inus) (9.1 to 11.7 cm; 28 to 51 g) is found in t he sout heaster n United States wh ere it inha bit s woode d areas, swamp land, str eam banks, and field edges. This tr ee cl imber nests i n trees, unde r logs , and in buildings .
� The brush mous e (Peromyscu s boyl ii) (9.7 to 10.7 cm; 22 to 36 g) is l imi ted to chaparral and rocky ar eas of th e arid and semi arid west an d southwest United States. A good c lim ber, it lives unde r rocks an d debris and in crevices. It feed s on pine nut s, aco rns, seeds, and berries.
General references
Burt and Grossen heider (1980); Kirkland and Lane (1989); Milla r (1985, 1989); Wolff (1989).
2-294 Deer Mous e
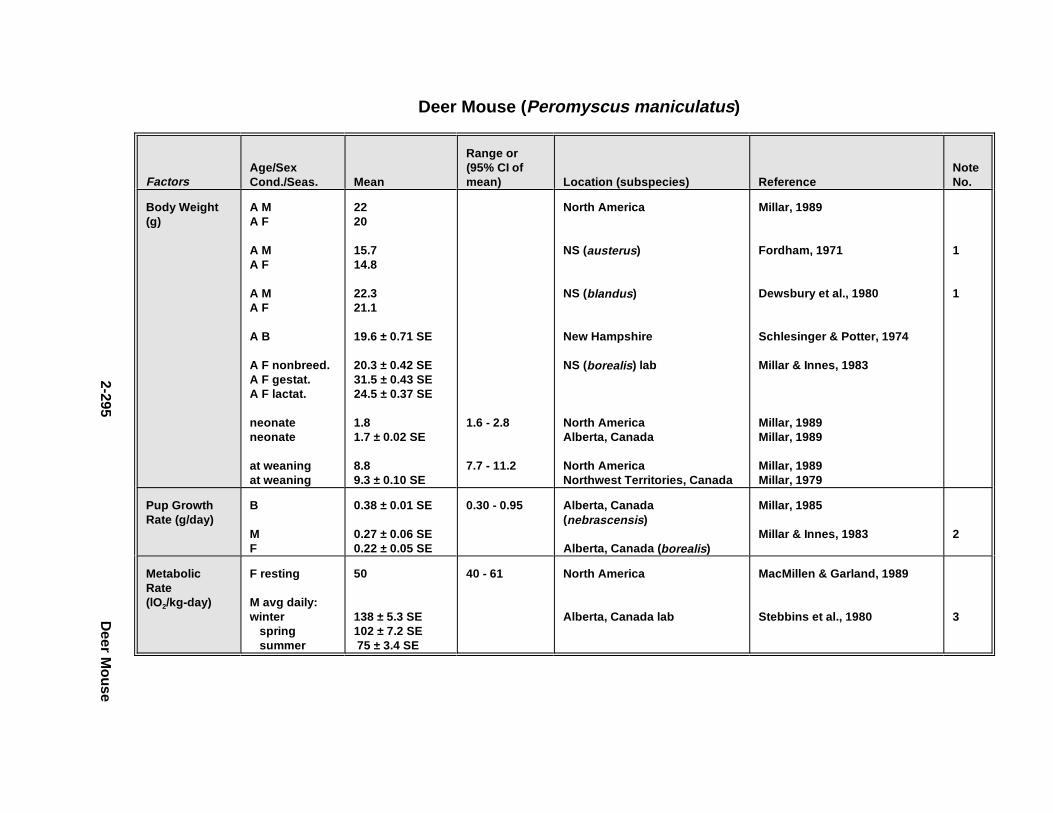
Deer Mouse ( Perom yscus maniculatus )
2-295 D
eer Mous
e
Factors Age/Sex (95% CI of Note Cond. /Seas. Mean mean) Locat ion (subspeci es) Reference No.
Range or
Body Weight A M 22 North America Milla r, 1989 (g) A F 20
A M 15.7 NS (aust erus) Fordham , 1971 1 A F 14.8
A M 22.3 NS (blandus ) Dewsbury et al., 1980 1 A F 21.1
A B 19.6 ± 0.71 SE New Hampshi re Schl esinger & Pot ter, 1974
A F nonbr eed. 20.3 ± 0.42 SE NS (bor ealis) lab Milla r & Innes, 1983 A F gest at. 31.5 ± 0.43 SE A F lactat. 24.5 ± 0.37 SE
neonate 1.8 1.6 - 2.8 North America Milla r, 1989 neonate 1.7 ± 0.02 SE Alberta , Canada Milla r, 1989
at weaning 8.8 7.7 - 11.2 North America Milla r, 1989 at weaning 9.3 ± 0.10 SE North west Territo ries, Canada Milla r, 1979
Pup Growth B 0.38 ± 0.01 SE 0.30 - 0.95 Alberta , Canada Milla r, 1985 Rate (g/day) (nebr ascensi s)
M 0.27 ± 0.06 SE Milla r & Innes, 1983 2 F 0.22 ± 0.05 SE Alberta, Canada (bor ealis)
Metabolic F resting 50 40 - 61 North America MacMille n & Garland, 1989 Rate (lO /kg-day) M avg daily :2
winter 138 ± 5.3 SE Alberta, Canada l ab Stebbi ns et al., 1980 3 spr ing 102 ± 7.2 SE sum mer 75 ± 3.4 SE

Deer Mouse ( Perom yscus maniculatus )
Factors Age/Sex (95% CI of Note Cond. /Seas. Mean mean) Locat ion (subspeci es) Reference No.
Range or
Metabolic Rate M avg daily : Alberta, Canada lab Stebbins et al., 1980 3 (kcal /kg-day) winter 668 ± 25 SE
spr ing 623 ± 35 SE sum mer 360 ± 17 SE
B f ree-living: Illinoi s lab Morris & Kendei gh, 1981 4 winter 790 sum mer 592
A M free-living 547 (259 - 1,153) est imated 5 A F f ree-living 574 (271 - 1,212)
Food A F nonbr eed. 0.19 Mani toba, Canada Milla r, 1979 6 Ingest ion (maniculatus) lab 7 Rate A F nonbr eed. 0.18 Alberta, Canada Milla r & Innes, 1983 (g/g-day) (bor ealis) lab 6
A F lactating 0.45 Manito ba, Canada Milla r, 1979
A F lactating 0.38 Alberta, Canada Milla r & Innes, 1983
A F nonbr eed. 0.19 Virginia lab Croni n & Br adley, 1988 A M nonbr eed. 0.22
J M 0.21 ± 0.01 SE Sout h Dakot a lab Nelson & Desj ardins, 1987
(maniculatus) lab 8
(bor ealis) lab
7
9
Water A B 0.19 (sonor iensi s) lab Ross, 1930 10 Ingest ion A B 0.19 0.123 - 0.287 Illin ois (bairdi i) lab Dice, 1922 11 Rate (g/g-day) J M 0.34 ± 0.02 SE Sout h Dakot a lab Nelson & Desj ardins, 1987 12
0.15 est imated 13
Inhal ation A M 0.025 est imated 14 Rate (m /day) A F 0.0233
Surface Ar ea A M 91 est imated 15 (cm ) A F 862
2-296 D
eer Mous
e

Deer Mouse ( Perom yscus maniculatus )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion (subspeci es)/Habi tat Note
nut s/seeds 0 24 23 Virginia (nubi terrae)/ Wolff e t al., 1985 arthropods 56 30 46 oak-maple-hickor y forest Lepi dopt . larvae 4 trace 2 Lepi dopt . adul ts 3 26 7 (% frequency of occur rence; green veg. 5 12 18 stomach cont ents) fungus 7 trace 1 fruit 25 unknown 1 4 3
Lepi dopt . larvae 20.6 34.5 16.7 4.8 Indi ana/sever al habi tats Whitaker, 1966 cor n 4.1 4.2 3.2 8.7 misc. veg. 15.8 3.1 8.0 13.4 wheat seeds 6.5 1.6 3.2 23.7 (% vol ume; stomach uni dent . seeds 5.4 5 8.8 8.3 cont ents) green veg. 7.6 0 4.3 3.7 Echi nochl oa
seeds Coleopt era soy beans Hemiptera
0 1.2 6.4 0 3.9 5.3 5.1 1.4
13.4 3.1 6.9 10.7 1.3 2.7 4.2 0.9
beet les 14.6 23.8 9.4 4.9 Colorado/shor t grass pr airie Flake, 1973 grasshopper s 6.4 4.2 6.4 2.5 leafhopper s 13.3 1.8 1.9 2.5 (% vol ume by a ranki ng Lepi dopt erans 21.7 12.7 1.5 1.8 method; stomach cont ents) spi ders 2.6 2.7 2.5 0.3 seeds 22.5 25.9 56.8 65.4 forbs 4.7 10.0 5.6 4.3 grasses & sedges 4.0 2.6 2.8 4.8
shr ubs 3.8 1.4 0.8 2.6
1 4
2-297 D
eer Mous
e
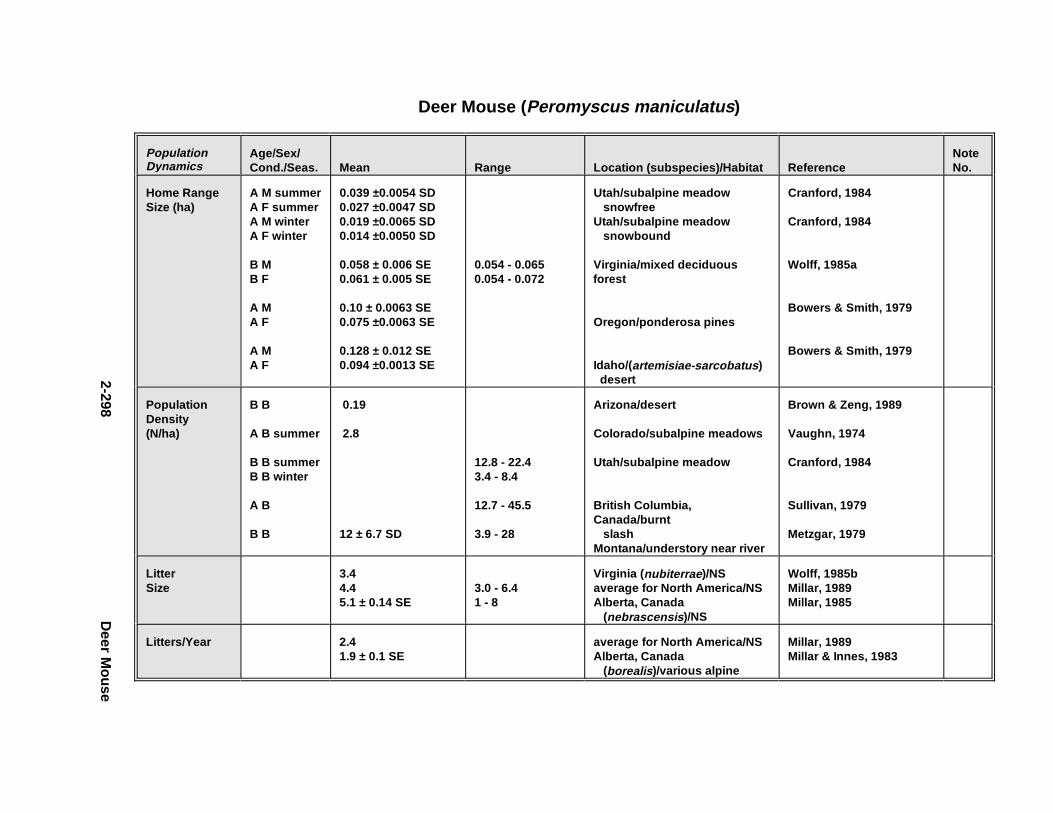
Deer Mouse ( Perom yscus maniculatus )
2-298 D
eer Mous
e
Popul ation Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion (subspeci es)/Habi tat Reference No.
Home Range A M sum mer 0.039 ±0.0054 SD Utah/subal pine meadow Cranford, 1984 Size (ha) A F sum mer 0.027 ±0.0047 SD snow free
A M winter 0.019 ±0.0065 SD Utah/subal pine meadow Cranford, 1984 A F wi nter 0.014 ±0.0050 SD snowbound
B M 0.058 ± 0.006 SE 0.054 - 0.065 Virginia/mixed deci duous Wolff, 1985a B F 0.061 ± 0.005 SE 0.054 - 0.072 forest
A M 0.10 ± 0.0063 SE Bow ers & Smith, 1979 A F 0.075 ±0.0063 SE Oregon/ ponder osa pi nes
A M 0.128 ± 0.012 SE Bow ers & Smith, 1979 A F 0.094 ±0.0013 SE Idaho/(artemisiaesarcobat us)
deser t
Popul ation B B 0.19 Ar izona/desert Brown & Zeng, 1989 Densi ty (N/ha) A B sum mer 2.8 Colorado/subal pine meadows Vaughn, 1974
B B sum mer 12.8 - 22.4 Utah/subal pine meadow Cranford, 1984 B B w inter 3.4 - 8.4
A B 12.7 - 45.5 British Col umbia, Sul livan, 1979
B B 12 ± 6.7 SD 3.9 - 28 slash Metzgar, 1979 Canada/bur nt
Mont ana/under story near river
Litte r 3.4 Virg in ia (nubi terrae)/NS Wolff, 1985b Size 4.4 3.0 - 6.4 average for North Am erica/NS Milla r, 1989
5.1 ± 0.14 SE 1 - 8 Alberta, Canada Milla r, 1985 (nebr ascensi s)/NS
Li tters/Year 2.4 average for North Am erica/NS Milla r, 1989 1.9 ± 0.1 SE Alberta , Canada Milla r & Innes, 1983
(bor ealis)/var ious al pine

Deer Mouse ( Perom yscus maniculatus )
Popul ation Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion (subspeci es)/Habi tat Reference No.
Days F non- lact . 23.6 average for Uni ted States/NS Milla r, 1989 Gestation F lactating 26.9
F non- lact . 22.4 ± 0.1 SE 22 - 23 Kansas/ NS Svendsen, 1964 16 F lactating 24.1 ± 0.3 SE 22 - 27
F non- lact . 25.5 ± 0.3 SE 23 - 26 Alberta, Canada Milla r, 1985 F lactating 29.5 ± 1.4 SE 24 - 35 (nebr ascensi s)/lab
Age at B 20.2 16 - 25 average for North Am erica/NS Milla r, 1989 Weaning B 24.9 Alberta, Canada Milla r & Innes, 1983 (days) (bor ealis)/var ious al pine
B 17.5 Colorado/NS Halfpenny , 1980 16
Age at M 35 days Alberta , Canada Milla r, 1985 Sexual F 60 days (nebr ascensi s)/lab Maturity
Mortality A F winter 100%/winter Alberta, Canada (bor ealis)/ Milla r & Innes, 1983 Rates A M winter 33%/winter various al pine
J F w inter 56%/winter J M winter 70%/winter
A B sum mer 20%/2 weeks Alberta, Canada (bor ealis)/ Milla r & Innes, 1983 J B sum mer 19%/2 weeks various al pine
Longevi ty B B < 1 yr Alberta, Canada (bor ealis)/ Mille r & Innes, 1983 various al pine
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating Apr il August Massachuset ts Drickam er, 1978 16 Novem ber Apr il Texas Blair, 1958 16 March October Virginia (nubi terrae) Wolff, 1985b 16 May August California Dunm ire, 1960 16
Disper sal spr ing Vancouver , Canada Fairbairn, 1977 (males)
2-299 D
eer Mous
e

� � � � � �
� � � �
�
�
� �
Deer Mouse ( Perom yscus maniculatus )
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Torpor winter nor thern par ts of range Tannenbaum & Pivor un, 1989
1 Cited in Mont gom ery (1989).2 Growth rate of "newl y emerged" pups, soon af ter leaving t he nest .3 Temperatures dur ing w inter averaged -17.7�C (-6 to -22�C); dur ing spr ing aver aged 14.5�C (8 to 22�C); dur ing sum mer 20.6�C (14 to 32�C).4 Est imated by author s f rom labor atory-der ived m odel assum ing no r eproduct ion, molt, or weight change and assum ing sum mer temperatures
averaged 17.5�C above gr ound and 20. 2�C in bur rows and w inter temperatures aver aged -3�C above gr ound and 10. 7�C in burro ws. 5 Est imated usi ng equat ion 3-48 (Nagy, 1987) and body weight s f rom Milla r (1989). 6 Diet of rat chow wi th 3 percent water cont ent and 4.5 kcal /g dry weight . 7 Diet of Pur ina lab chow no. 5001; com posi tion not speci fied. 8 Diet of lab chow; com posi tion not speci fied. 9 Diet of lab chow wi th 8 to 10 percent water cont ent .
10 Mean var ied by subspeci es; sonor iensi s, eremicus, gambelii , and fratercul us tested. Dry diet prepared in lab, probabl y less t han 10 per cent water cont ent; air temperature 21 to 24�C.
11 Dry air at 32 to 34�C; diet of wheat and peanut s, about 10 percent water cont ent. 12 Temperature 20�C ± 2�C; diet of lab chow wi th 8 to 10 percent water cont ent. 13 Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Milla r (1989). 14 Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Milla r (1989). 15 Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Milla r (1989). 16 Cited in Milla r (1989).
2-300 D
eer Mous
e

References (including A ppendix)
Abbott, K. D. (1974) Ecotyp ic and racial var iation in t he water and energy meta bolis m of Peromyscu s man iculatus from th e wester n United States an d Baja California, Mexico [Ph.D. disser tat ion] . Irvine , CA: Univ ersi ty of Calif ornia .
Agnew, W. J.; Uresk, D. W.; Hansen, R. M.; et al . (1988) Arthropod c ons umpt ion by smal l mammal s on prairie dog c olonie s and adjacent ungr azed mixed grass p rairie in wester n South Dakota. In: Uresk, D. W.; Schenbeck, G. L.; Cefki n, R., tech . coords. Eight h Great Plains wildlif e damage control workshop pr oceedings ; April 28-30, 1987; Rapid Cit y, Sout h Dakota. For t Collins , CO: U.S. Dept . Agr., Forest Ser v., Rocky Mount ain For est an d Range Experiment Stat ion; pp. 81-87.
Barry, W. J. (1976) Environmental effects o n food hoa rding in de ermice (Peromyscu s). J. Mammal . 57: 731-746.
Birdsal l, D. A.; Nash, D. (1973) Occu rrence of su ccessfu l multiple insemina tion of femal es in na tural popula tions of deer mi ce (Peromyscu s man iculatus). Evolut ion 2 7: 106-110.
Blair, W. F. (1940) A stu dy of prairie deer mouse popula tions in s out hern Michiga n. Am. Midl. Nat. 24: 273-305.
Blair, W. F. (1958) Effects o f x-i rradiation of a na tural popula tion of deer-mous e (Peromyscu s man iculatus). Ecology 39: 113-118.
Bowers, M. A.; Smith, H. D. (1979) Differ ential habitat u tiliza tion by sexes o f th e deermouse, Peromyscu s man iculatus. Ecology 60: 869-875.
Brower , J. E.; Cade, T. J. (1966) Ecology and phy siology of Napaeozapus insignis (Mille r) and ot her woodla nd m ice. Ecology 47: 46-63.
Brown, J. H.; Zeng, Z. (1989) Comparative popula tion e cology of el even species of rodents in t he Chihua huan Deser t. Ecology 70: 1507-1525.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chappell, M. A.; Hols claw, D. S., III (1984) Effects o f wind on t hermoregula tion a nd energy balance in deer mice (Peromyscu s man iculatus). J. Comp. Physiol. B Bioc hem. Syst. Environ. Physiol. 154: 619-625.
Cook , J. C.; Topping, M. S.; Stombaugh, T. A. (1982) Food ha bit s of Microtus ochrogaster and Peromyscu s man iculatus in symp atry. Trans. Misso uri Acad. Sci . 16: 17-23.
2-301 Deer Mous e

Cranford, J. A. (1984) Popula tion e cology and hom e range ut iliza tions of two suba lpine meadow r odents (Microtus longi caudus and Peromyscu s man iculatus). In: Merri t, J. F., ed. Wint er ecology of smal l mammal s: v. 10. Spec. Publ. Carnegie Mus. Nat. Hist. ; pp. 1-380.
Cronin, K. L.; Bradley, E. L. (1988) The relations hip b etween food in take, body fat an d reproduc tive inhibit ion in pr airie deermice (Peromyscu s man iculatus bairdii). Comp. Bioc hem. Physiol. A Comp. Physiol. 89: 669-673.
Deavers, D. R.; Huds on, J. W. (1981) Temperature regula tion in t wo rodents (Clethrionomys gapperi and Peromyscu s leucopus ) and a shrew (Blarina brevicauda) inha bit ing t he same en vironment. Physiol. Zool. 54: 94-108.
Dewsb ury, D. A.; Baumgardner, D. J.; Evans, R. L.; et al . (1980) Sexual dimorphism fo r body mass i n 13 taxa o f mu roid r odents unde r labor atory condit ions . J. Mammal . 61: 146-149.
Dice, L. R. (1922) Some facto rs affect ing t he dis tr ibut ion of th e prairie vole , forest d eer mous e, and pr airie deer mous e. Ecology 3: 29-47.
Drickamer , L. C. (1970) Seed preferences i n wild c aught Peromyscu s man iculatus bairdii and Peromyscu s leucopus nov ebor acensis. J. Mammal . 51: 191-194.
Drickamer , L. C. (1976) Hypothesis link ing f ood ha bit s and ha bitat sel ect ion in Peromyscu s. J. Mammal . 57: 763-766.
Drickamer , L. C. (1978) Annua l reproduc tion p atter ns in popula tions of two symp atr ic species of Peromyscu s. Behavior . Biol. 23: 405-408.
Drickamer , L. C.; Bernstei n, J. (1972) Growth in two subspecies of Peromyscu s maniculatus. J. Mammal . 53: 228-231.
Dunm ire, W. W. (1960) An alti tudina l survey o f reproduc tion in Peromyscu s man iculatus. Ecology 41: 174-182.
Eisenberg, J. F. (1981) The mammal ian radiations . Chic ago, IL: Univ ersi ty of Chic ago Press.
Fairbairn, D. J. (1977) The spring de cline in de er mi ce: death or dispersal? Can. J. Zool. 55: 84-92.
Fairbairn, D. J. (1978) Dispersal of deer mice, Peromyscu s man iculatus: proximal causes and effects o n fi tness. Oecologia 32: 171-193.
Flake, L. D. (1973) Food ha bit s of four species of r odents on a shor t-grass p rairie in Color ado. J. Mammal . 54: 636-647.
2-302 Deer Mous e

Fleharty, E. D.; Krause, M. E.; Stinnett, D. P. (1973) Body co mpos it ion, energy cont ent, and lipid cycl es of fo ur species of rodents. J. Mammal . 54: 426-438.
Fordham, R. A. (1971) Field popula tions of deermice wi th suppl emental food. Ecology 52: 138-146.
Glazier, D. S. (1979) An energetic and ecologic al basis for di ffer ent reproducti ve rates i n five sp ecies of Peromyscu s (mice) [Ph .D. disser tat ion] . Ithaca, NY: Cornell Universi ty.
Green, D. A.; Milla r, J. S. (1987) Changes in gut dimensions and c apaci ty of Peromyscu s maniculatus relative to die t qua lit y and energy needs. Can. J. Zool. 65: 2159-2162.
Gyug, L. W. (1979) Reproducti ve and developmental adjustmen ts to breeding seaso n lengt h in Peromyscu s [master 's th esis]. London, Ontar io: Univ ersi ty of W ester n Ontar io.
Gyug, L. W.; Milla r, J. S. (1980) Fat levels in a subarcti c popula tion of Peromyscu s maniculatus. Can. J. Zool. 58: 1341-1346.
Halford, D. K. (1987) Densi ty, movemen t, and transuranic ti ssu e inventory of smal l mammal s at a liquid r adio acti ve waste d isposal area. In: Pinde r, J. E., III; Alberts, J. J.; McLeod, K. W., et al ., eds. Environmental research on act inide el ements; Novemb er 7-11, 1983; Hilt on Head, Sout h Carolina . U.S. Departmen t of Energy, Offi ce of Sci enti fic and Technic al Information. Rep. No. CONF-841142 (DE86008713); pp. 147-156.
Halfpenny, J. C. (1980) Reproducti ve str ategies: intra and interspeci fic co mparison wi thin the genus Peromyscu s [Ph.D. disser tat ion] . For t Collins , CO: Univ ersi ty of Color ado.
Hamilton, W. J., Jr . (1941) The foods of smal l forest mammal s in easter n United States. J. Mammal . 22: 250-263.
Harris, J. H. (1986) Microhabitat seg regation in t wo deser t rodent sp ecies: the relation of prey availa bilit y to die t. Oecologia (B erl.) 68: 417-421.
Haywar d, J. S. (1965) Metabolic r ate and its temp erature-adaptive s ignif icance in s ix geogr aphic r aces o f Peromyscu s. Can. J. Zool. 43: 309-323.
Hock, R. J.; Roberts, J. C. (1966) Effect o f al ti tude on oxyg en consumption of deer mi ce: relation of temp erature and seaso n. Can. J. Zool. 44: 365-376.
Holbr ook , S. J. (1979) Habitat u tiliza tion, competi tive int eractions , and c oexisten ce of th ree species of cr iceti ne rodents i n east-cen tral Arizona . Ecology 60: 758-769.
Howard, W. E. (1949) Dispersal , amount of inbr eeding, and longe vi ty of a loc al popula tion of prairie deer mice on the George Reserve, southern Michiga n. Cont r. Lab. Vert. Biol. , Univ ersi ty of Michiga n 43:1-52.
2-303 Deer Mous e

Johns on, D. R. (1961) The food ha bit s of r odents in r ange lands of sout hern Idaho. Ecology 42: 407-410.
Kantak, G. E. (1983) Behavioral, seed preference, and habitat sel ect ion e xperiments wi th two symp atr ic Peromyscu s species. Am. Midl. Nat. 109: 246-252.
Kaufman , D. W.; Kaufman , G. A. (1989) Popula tion biology . In: Kir kland, G. L.; Lane, J. N., eds. Advances i n the stu dy of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech Universi ty Pr ess.
King, J. A.; Deshaies, J. C.; Webster , R. (1963) Age of weaning of two subs pecies of deer mice. Science 139: 483-484.
Kirkland, G. L.; Lane, J. N., eds. (1989) Advances i n the stu dy of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech Universi ty Pr ess.
Layne, J. N. (1968) Ontogeny. In: Biology of Peromyscu s (Rodent ia). Spec. Publ. , Amer . Soc. Mammal . 2: 1-593.
Linze y, A. V. (1970) Postn atal growth and development of Peromyscu s man iculatus nubi ter rae. J. Mammal . 51: 152-155.
MacMille n, R. E.; Garland, T. J. (1989) Adaptive physiology . In: Kir kland, G. L.; Lane, J. N., eds. Advances i n the stu dy of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech Universi ty Pr ess.
Marine lli, L.; Milla r, J. S. (1989) The ecology of b each-dwelling Peromyscu s man iculatus on the Paci fic co ast. Can. J. Zool. 67: 412-417.
Martel l, A. M.; MacAuley, A. L. (1981) Food ha bit s of deer mi ce (Peromyscu s man iculatus) in northern Ontar io. Can. Field Nat. 95: 319-324.
Marten, G. G. (1973) Time patter ns of Peromyscu s acti vi ty an d their correlations wi th weath er. J. Mammal . 54: 169-188.
May, J. D. (1979) Demographic adju stmen ts to breeding seaso n length in Peromyscu s [master 's th esis]. London, Ontar io: Univ ersi ty of W ester n Ontar io.
McCabe, T. T.; Blanchard, B. D. (1950) Three species of Peromyscu s. Santa Barbara, CA: Rood A sso ciates.
McLaren, S. B.; Kirkland, G. L., Jr . (1979) Geographic var iation in li tter size of smal l mammal s in the central Appalachian region. Proc. Penns ylvania A cad. Sci . 53: 123-126.
McNab, B. K.; Morrison, P. (1963) Body temp erature and metabolis m in s ubs pecies of Peromyscu s from ar id and mesi c environments. Ecol. Monogr . 33: 63-82.
2-304 Deer Mous e

Menhusen, B. R. (1963) An investi gation on t he food ha bit s of four species of r odents in captivi ty. Trans. Kansas Acad. Sci . 66: 107-112.
Metzgar, L. H. (1973a) Explor atory and feeding hom e ranges in Peromyscu s. J. Mammal . 54: 760-763.
Metzgar, L. H. (1973b) Home range shape and acti vi ty i n Peromyscu s leucopus . J. Mammal . 54: 383-390.
Metzgar, L. H. (1979) Dispersion p atter ns in a Peromyscu s popula tion. J. Mammal . 60: 129-145.
Metzgar, L. H. (1980) Dispersion a nd num bers in Peromyscu s popula tions . Am. Midl. Nat. 103: 26-31.
Meyers, P.; Master , L. L.; Garrett, R. A. (1985) Ambient temp erature and rainfal l: an effect on sex r atio and litter size. J. Mammal . 66: 289-298.
Milla r, J. S. (1975) Tacti cs o f energy parti tioning in br eeding Peromyscu s. Can. J. Zool. 53: 967-976.
Milla r, J. S. (1978) Energetics o f reproduc tion in Peromyscu s leucopus : the cost o f lactat ion. Ecology 59: 1055-1061.
Milla r, J. S. (1979) Energetics o f lactat ion in Peromyscu s man iculatus. Can. J. Zool. 57: 1015-1019.
Milla r, J. S. (1982) Life cycl e character isti cs o f northern Peromyscu s man iculatus borealis. Can. J. Zool. 60: 510-515.
Milla r, J. S. (1985) Life cycl e character isti cs o f Peromyscu s man iculatus nebrascen sis. Can. J. Zool. 63: 1280-1284.
Milla r, J. S. (1989) Reproduc tion a nd d evelopm ent. In: Kir kland, G. L.; Lane, J. N., eds. Advances i n the stu dy of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech Universi ty Pr ess; pp. 169-205.
Milla r, J. S.; Innes, D. G. (1983) Demographic and lif e cycl e character isti cs o f mo ntane deer mice. Can. J. Zool. 61: 574-585.
Milla r, J. S.; Schi eck, J. O. (1986) An annua l lipid cycl e in a montane popula tion of Peromyscu s man iculatus. Can. J. Zool. 64: 1981-1985.
Milla r, J. S.; Willie , F. B.; Iverson, S. L. (1979) Breeding by Peromyscu s in seaso nal environm ents. Can. J. Zool. 57: 719-727.
2-305 Deer Mous e

Mont gom ery, W. I. (1989) Peromyscu s and Apodemus: patter ns of si mi larity i n ecologic al equivalents. In: Kirkland, G. L.; Lane, J. N., eds. Advances i n the stu dy of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech Universi ty Pr ess; pp. 293-366.
Mor ris, J. G. ; Kendeigh, C. S. (1981) Energetics o f th e prairie deer mouse Peromyscu s maniculatus bairdii. Am. Midl. Nat. 105: 368-76.
Morrison, P. R. (1948) Oxyg en consumption in sever al smal l wild mammal s. J. Cell. Comp. Physiol. 31: 69-96.
Morrison, P.; Dieter ich, R.; Presto n, D. (1977) Body growth in s ixteen rodent sp ecies and subs pecies maint ained in la bor atory colonie s. Physiol. Zool. 50: 294-310.
Murie, M. (1961) Metabolic ch aracter isti cs o f mountain, deser t and coastal popula tions of Peromyscu s. Ecology 42: 723-740.
Myers, P.; Master , L. L. (1983) Reproduc tion by Peromyscu s man iculatus: size and compromise. J. Mammal . 64: 1-18.
Myers, P.; Master , L. L.; Garrett, R. A. (1985) Ambient temp erature and rainfal l: an effect o n sex r atio and litter size in deer mice. J. Mammal . 66: 289-298.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nelson, R. J.; Desjardins , C. (1987) Water avai labilit y affects r eproduc tion in de er mi ce. Biol. Reproduc . 37: 257-260.
Reynolds , T. D.; Wakki nen, W. L. (1987) Character isti cs o f th e burrows o f fo ur species of rodents in undis turbed soils in s out heaster n Idaho. Am. Midl. Nat. 118: 245-250.
Ribble , D. O.; Samso n, F. B. (1987) Microhabitat asso ciations of smal l mammal s in southeaster n Color ado, wi th special empha sis on Peromyscu s (Rodent ia). Southwest. Nat. 32: 291-303.
Rood, J. K. (1966) Observat ions on t he reproduc tion of Peromyscu s in c aptivi ty. Am. Midl. Nat. 76: 496-503.
Ross, L. G. (1930) A comparative stu dy of daily water -intake am ong c ertain taxonom ic an d geogr aphic groups wi thin t he genus Peromyscu s. Biol. Bull. 59: 326-338.
Sadleir, R. M. (1970) Popula tion dy namics and br eeding of th e deermous e (Peromyscu s maniculatus) on Bur naby Mount ain, Br it ish Colum bia. Syesi s 3: 67-74.
Schle singe r, W. H.; Potter , G. L. (1974) Lead, coppe r, and c admium conc entrations in smal l mammal s in the Hubba rd Brook Exp erimental Forest. Oikos 25: 148-152.
2-306 Deer Mous e

Sieg, C. H.; Uresk, D. W.; Hansen, R. M. (1986) Seasonal diets o f deer mice on bentonit e mine spoils and s agebrush grassl ands in southeaster n Montana. Northwest Sci . 60: 81-89.
Smith, M. H.; McGinnis , J. T. (1968) Relations hips of lati tude , alti tude and body size to li tter size and mean annua l pr oduc tion of o ffsp ring in Peromyscu s. Res. Pop. Biol. 10: 115-126.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stebbins , L. L. (1977) Energy requir ements d uring r eproduc tion of Peromyscu s maniculatus. Can. J. Zool. 55: 1701-1704.
Stebbins , L. L. (1978) Some asp ects o f overwinter ing in Peromyscu s man iculatus. Can. J. Zool. 56: 386-390.
Stebbins , L. L.; Orich, R.; Nagy, J. (1980) Metabolic r ates o f Peromsyscu s man iculatus in wint er, spring, and s ummer . Acta. Theriol. 25: 99-104.
Sulliv an, T. P. (1979) Repopula tion of cl ear-cut habitat an d conifer seed predation by deer mice. J. Wildl. Manage. 43: 861-871.
Svendsen, G. (1964) Comparative reproduc tion a nd d evelopment in two species of mi ce in the genus Peromyscu s. Trans. Kansas Acad. Sci . 67: 527-538.
Svihla , A. (1932) A comparative l ife histo ry stu dy of th e mice of th e genus Peromyscu s. Misc. Publ. Mus. Zool. , Univ . Michiga n 24: 1-39.
Svihla , A. (1934) Development and growth of deermice (Peromyscu s man iculatus artemi siae). J. Mammal . 15: 99-104.
Svihla , A. (1935) Development and growth of th e prairie deer mouse, Peromyscu s maniculatus bairdii. J. Mammal . 16: 109-115.
Taitt, M. J. (1981) The effect o f extr a food on small r odents popula tions : deer mi ce (Peromyscu s man iculatus). J. Anim. Ecol. 50: 111-124.
Taitt, M. J. (1985) Cycl es and annua l f luc tuations : Microtus townsendi i and Peromyscu s maniculatus. Acta. Zool. Fenn. 173: 41-42.
Tannenbaum, M. G.; Pivorun, E. B. (1988) Seasonal stu dy of daily to rpor in southeaster n Peromyscu s man iculatus and Peromyscu s leucopus from mount ains and f oot hills . Physiol. Zool. 61: 10-16.
Tannenbaum, M. G.; Pivorun, E. B. (1989) Summer torpor in montane Peromyscu s maniculatus. Am. Midl. Nat. 121: 194-197.
2-307 Deer Mous e

Thomas, B. (1971) Evolut iona ry relations hips among Peromyscu s from th e Georgia Strait, Gordon, Goletas, and Scott Islands of British Colum bia, Canada [Ph .D. disser tat ion] . Vancouv er, BC: Univ ersi ty of Br it ish Colum bia.
Tomasi , T. E. (1985) Basal metabolic r ates an d thermoregula tory abilit ies in f our smal l mammal s. Can. J. Zool. 63: 2534-2537.
van Horne, B. (1982) Niches of adult and juv enile deer mi ce (Peromyscu s man iculatus) in seral stag es of coniferous fo rest. Ecology 63: 992-1003.
Vaughn, T. A. (1974) Resource alloc ation in s ome symp atr ic su balpine r odents. J. Mammal . 55: 764-795.
Vicker y, W. L. (1981) Habitat u se by northeaster n forest r odents. Am. Midl. Nat. 106: 111-118.
Whitaker , J. O., Jr . (1966) Food of Mus muscu lus, Peromyscu s man iculatus bairdi, and Peromyscu s leucopus in Vigo Count y, India na. J. Mammal . 47: 473-486.
Wolff, J. O. (1985a) The effects o f densi ty, food, and int erspeci fic int erference on hom e range size in Peromyscu s leucopus and Peromyscu s man iculatus. Can. J. Zool. 63: 2657-2662.
Wolff, J. O. (1985b) Comparative popula tion e cology of Peromyscu s leucopus and Peromyscu s man iculatus. Can. J. Zool. 63: 1548-1555.
Wolff, J. O. (1986) The effects o f food on m ids ummer demography of wh ite-fo oted mice, Peromyscu s leucopus . Can. J. Zool. 64: 855-858.
Wolff, J. O. (1989) Social behavior. In: Kirkland, G. L.; Lane, J. N., eds. Advances i n the study of Peromyscu s (Rodent ia). Lubboc k, TX: Texas T ech. Universi ty Pr ess; pp. 271-291.
Wolff, J. O.; Durr, D. S. (1986) Winter nest ing be havior of Peromyscu s leucopus and Peromyscu s man iculatus. J. Mammal . 67: 409-412.
Wolff, J. O.; Hurlbutt, B. (1982) Day refuges of Peromyscu s leucopus and Peromyscu s maniculatus. J. Mammal . 63: 666-668.
Wolff, J. O.; Freeberg, H.; Dueser , R. D. (1983) Interspeci fic ter ritoriality i n two symp atr ic species of Peromyscu s (Rodent ia: Criceti dae). Behav. Ecol. Sociobiol. 12: 237-242.
Wolff, J. O.; Dueser , R. D.; Berry, D. S. (1985) Food ha bit s of symp atr ic Peromyscu s leucopus and Peromyscu s man iculatus. J. Mammal . 66: 795-798.
Zegers, D. A.; Merritt, J. F. (1987) Seasonal changes in non- shiv ering t hermoge nesis of three smal l mammal s (abstr act only ). Bull. Ecol. Soc. Am. 68: 455.
2-308 Deer Mous e

Zegers, D. A.; Merritt, J. F. (1988) Adaptat ions of Peromyscu s for winter survival in an Appalachian montane forest. J. Mammal . 69: 516-523.
2-309 Deer Mous e

2.2.8. Prairie Vole (v oles)
Order Rodent ia Family Murid ae (subfami ly Arv icolin ae). New wo rld voles are smal l, herbivorous rodents th at reside in all areas of th e United States wh ere good gr ass co ver exists. Their presence is ch aracter ized by narrow runways th rough matted grasses. Microtus species are adapted to unde rground, ter restr ial, and someti mes semi amphibious habitats (J ohns on and J ohns on, 1982). They are acti ve by day and night and feed mainly on shoots, grasses, and bark (Johns on and J ohns on, 1982). Voles are prey fo r snakes, raptors, and mammal ian predators su ch as sh ort-tai led shrews, badgers, raccoons , coyotes, and foxes (Ead ie, 1952; Johns on and J ohns on, 1982; Martin, 1956).
Selected species
The prairie vole (Microtus ochrogaster ) represents th e ground- bur rowing memb ers of this group. This vole is found in t he nor th and c entral pla ins of th e Unit ed States an d in southern Canada, usually in dry places su ch as prairies and along f encerows and railroads. Its r ange has exp anded eastwar d to West Vi rginia as a result of cl ear-cutting of forests (Jo nes et al ., 1983). Voles are acti ve by day or night (Johns on and J ohns on, 1982). Although pr airie and meadow vo les usually occu py differ ent habitats, where they co exist their popula tion de nsit ies tend t o be negatively correlated (Klatt, 1985; Krebs, 1977).
Body size. The prairie vole measu res fr om 8.9 to 13 cm i n length and has a 3.0- to 4.1-cm tai l (Burt and Grossen heider, 1980). After reaching sexu al matu rity, voles co ntinue to grow fo r sever al months (Johns on and J ohns on, 1982). Adult s weigh f rom 30 to 45 g (see tab le). Prairie voles mai ntain a relatively co nstan t propor tion of th eir body weight as fat (15 to 16 percent on a dry-weight basis) throughout th e year (Fleharty et al ., 1973).
Habitat . The prairie vole inha bit s a wide var iety o f prairie plant co mmunit ies and moistu re regimes, including r iparian, shor t-grass, or tal l-grass co mmunit ies (Kaufman and Fleharty, 1974). Prairie voles prefer areas of dense veg etat ion, such as gr ass, alfal fa, or clov er (Carroll a nd Getz, 1976); their presence in a habitat d epends on suitable cover for runways (K aufman and Fleharty, 1974). They will t olerate sp arser plant co ver than the meadow vo le because th e prairie vole usually nests i n burrows at l east 50 mm unde rground or in gr ass n ests unde r logs or boards (Klatt an d Getz, 1987).
Food H abi ts . Meadow vole s, as ot her vole s, are largely herbiv orous , cons uming primar ily green succu lent veg etat ion but al so root s, bark, seeds, fungi, arthropods , and animal matter (Johns on and J ohns on, 1982; Lomolino, 1984; Stalling, 1990). Voles have masti cato ry and digesti ve systems th at allow th em to dig est fi brous grasses su ch as cereals (Johns on and J ohns on, 1982). Diet var ies by seaso n and habitat acco rding t o plant availa bilit y, al though meadow and other voles sh ow a preference fo r young, tende r vegetat ion ( Johns on and J ohns on, 1982; Martin, 1956). Voles can damage pastu res, grassl ands, crops su ch as hay and grain, and fruit tr ees (by eat ing ba rk and r oot s) (Johns on and J ohns on, 1982).
2-311 Prairie Vole

�
�
Temperature regulation and molt. Unlik e some ot her mammal s, prairie voles do not hibe rnate or exhibit to rpor (Johns on and J ohns on, 1982). They overwinter wi thout us ing their lipid r eserves, finding f ood t o meet th eir metabolic r equir ements year -round ( Fleharty et al ., 1973). Prairie voles use burrows, runways, nests, and snow co ver to help maintain their body temp erature. They also modify wh en they are acti ve to avoid excessi vely hot or cold temp eratures (Johns on and J ohns on, 1982). Voles unde rgo t hree molt s (juv enile , suba dult , and adult ), and m olt ing may o ccu r at any ti me during t he year (Jameso n, 1947, as ci ted in Stalling, 1990). The subadult-to -adult molt occurs between 8 and 12 weeks o f age (Martin, 1956).
Breeding acti vi ties and social organization. Prairie vole s are monoga mous ; a mated pair occupies th e same h ome range (Thomas an d Birney, 1979). Reproduc tion occurs throughout th e year , and gestat ion l asts ap proximatel y 3 wk (M artin, 1956; Keller, 1985; Nadeau, 1985). Both sexes car e for the young; paternal acti vi ties include r unw ay cons truction, food caching, groom ing, retr ieving, and br ooding t he young ( Thomas an d Birney, 1979). The young a re weaned by about 3 weeks o f age (Thomas an d Birney, 1979). Reproducti ve acti vi ty p eaks fr om May to Octo ber, coinc iding w ith high m ois ture availa bilit y (Martin, 1956; Keller, 1985). Monoga mous fami ly units ap parently defend ter ritories against other family groups (Ostfel d et al ., 1988; Johns on and J ohns on, 1982; Thomas an d Birney, 1979).
Home range and r esour ces. Prairie voles excavate unde rground n ests th at are used as nur ser ies, resting a reas, and as sh elter from sever e weath er (Klatt an d Getz, 1987). They sp end very l ittl e time away fr om th is nest (B arbour, 1963). In th ick veg etat ion, prairie vole s move about in s urface r unways, and the number of runways i s propor tiona l to popula tion de nsity (Carroll a nd Getz, 1976). Female home range size decreases wi th inc reasing pr airie vole dens ity acco rding t o the follow ing r egression e quation (Gaines and Johns on, 1982):
Y= �0.23X + 20.16 where Y= home range length in meter s and X= minim um num ber aliv e per 0.8 ha gr id.
Abramsky an d Tracy (1980) f ound a si mila r co rrelation us ing bot h sexes acco rding t o the equation:
Y= �0.20X + 27.12 where Y= home range length in meter s and X=number of indiv idua ls per hectar e.
Popul ation dy namics. Female prairie voles can reach sexu al matu rity i n about 35 d, males in 42 to 45 d (Gier and Cooksey, 1967, as ci ted in Stalling, 1990). Martin (1956) found in Kansas th at femal es matu re wi thin a bout 6 wk in t he summer , but may r equir e 15 wk or more to matu re if born in the fal l. Male prairie voles ten d to disperse fr om th eir natal si te; approximatel y twi ce as man y femal es as mal es matu re near their birthplace (Boons tra et al., 1987). Popula tions tend t o fluc tuate wi th avai lable moistu re (Gier, 1967, as ci ted in Stalling, 1990). Mortal ity r ates i n prairie vole postn est ling juv enile s and y oung a dult s are simila r and highe r th an adult mo rtalit y rates; nest lings h ave th e lowest mo rtal ity r ate (Golley, 1961). Average life exp ectan cy i n the field is about 1 yr (Martin, 1956).
2-312 Prairie Vole

�
�
Simi lar species (fr om general references)
� The pine vole ( Microtus pinetorum) (7 to 11 cm), despite i ts n ame, usually inha bit s deciduous fo rest floor s, among a thic k layer of duff, where it tunne ls through loos e soil ne ar th e surface. It is found in t he easter n half of th e United States, excep t Flor ida; in t he sout h, it inha bit s pine fo rests. In addit ion t o feeding on ba rk, it bur rows for bulbs , tube rs, and c orms.
� See also simi lar species l isted for the meadow vo le in th is ch apter .
General references
Burt and Grossen heider (1980); Johns on and J ohns on (1982); Stalling (1990); Tamar in (1985).
2-313 Prairie Vole

Prairie Vole ( Microtus ochro gaster )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A B 41.6 ne Colorado Abr amsky & Tracy, 1980 (g)
A B sum mer 41.9 ne Colorado Abr amsky & Tracy, 1980 A B f all 44.2 A B w inter 39.0 A B spr ing 41.3
A M 31.3 ± 0.35 SE s Indi ana Myers & Kr ebs, 1971 A F 33.3 ± 0.30 SE
neonat e B 2.8 ± 0.4 SD ne Kansas Martin, 1956
Metabol ic Rate A wi nter 51.8 ± 8.2 SD NS/lab Wunder et al., 1977 (lO /kg-d)2
A sum mer 41.8 ± 4.8 SD
Metabol ic Rate A B basal 177 est imated 1 (kcal /kg-d)
A B f ree-living 399 (190 - 833) est imated 2
Food Ingest ion A B at 21��C 0.13 - 0.14 Illinoi s/lab Dice, 1922 3 Rate (g/g-d) A B at 28��C 0.09 - 0.10
Water A B 0.37 NS/lab Chew, 1951 4 Ingest ion Rat e (g/g-d) A B 0.29 ± 0.02 SE Kansas/ lab Dupr e, 1983 5
A B 0.21 0.15 to 0.26 Illinoi s/lab Dice, 1922 6
A B 0.14 est imated 7
Inhal ation A B 0.043 est imated 8 Rate (m /d)3
Surface Ar ea A B 139 est imated 9 (cm )2
2-314 P
rairie Vo
le

Prairie Vole ( Microtus ochro gaster )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
Sporobol us as per Kochi a sco paria Bout eloua gracilis Bromus japoni cus Rumex cr ispus Triticum aest ivum Carex sp. other (grasses) (fo rbs)
19.5 Kansas/ forb and gr ass f ield Fleharty & Olson, 1969 22.5
6.5 (% vol ume; stomach 8.5 cont ents) 9.2 3.4 (Items less t han 2% of 2.0 volume were com bined as
28.3 "other" ) (53.5) (46.5)
Festuca ar undi nacea Dactylis glomerata Phleum pratense Tridens f lavus Setaria vi ridis Taraxacum officinale Lamium amplexicaul e Bromus t ectorum Setaria faberi Capsel la bur sapast . Trifol ium stoloni fera arthropods animal material other
20.5 25.0 10.6 28.9 Missouri/o ld fie ld Cook et al., 1982 6.7 1.7 1.1 4.2 8.3 2.0 2.1 5.3 (mean num ber of food
17.1 11.1 1.9 11.0 items; stomach cont ents) 6.7 6.2 1.7 6.2 5.8 4.8 3.9 1.5 (Plant parts consum ed: leaf, 3.9 2.9 5.2 3.4 stem, and seeds of Festuca 2.8 4.7 2.5 4.8 and Bromus; leaf and st em 5.6 3.9 0.7 21.0 of Tridens and Setaria 2.7 1.2 0.5 0.6 faberi ; leaf and seeds of 2.4 0.8 0.5 1.4 Dactylis and Seteria vi ridis; 0.2 0.3 0.0 0.1 and l eaves onl y of all other 0 0.2 0.2 0.0 plant speci es) 3.9 1.4 1.5 0.9
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home A B al l yr 0.098 ± 0.012 SE Illin ois/bluegrass Jike et al., 1988 Range Size (ha) A M all yr 0.037 ± 0.0029 SE Kansas/ NS Swihart & Slade, 1989
A F al l yr 0.024 ± 0.0018 SE
A M 0.011 ne Col orado/shor t-grass Abr amsky & Tracy, 1980 A F 0.0073 prairie
Range
2-315 P
rairie Vo
le
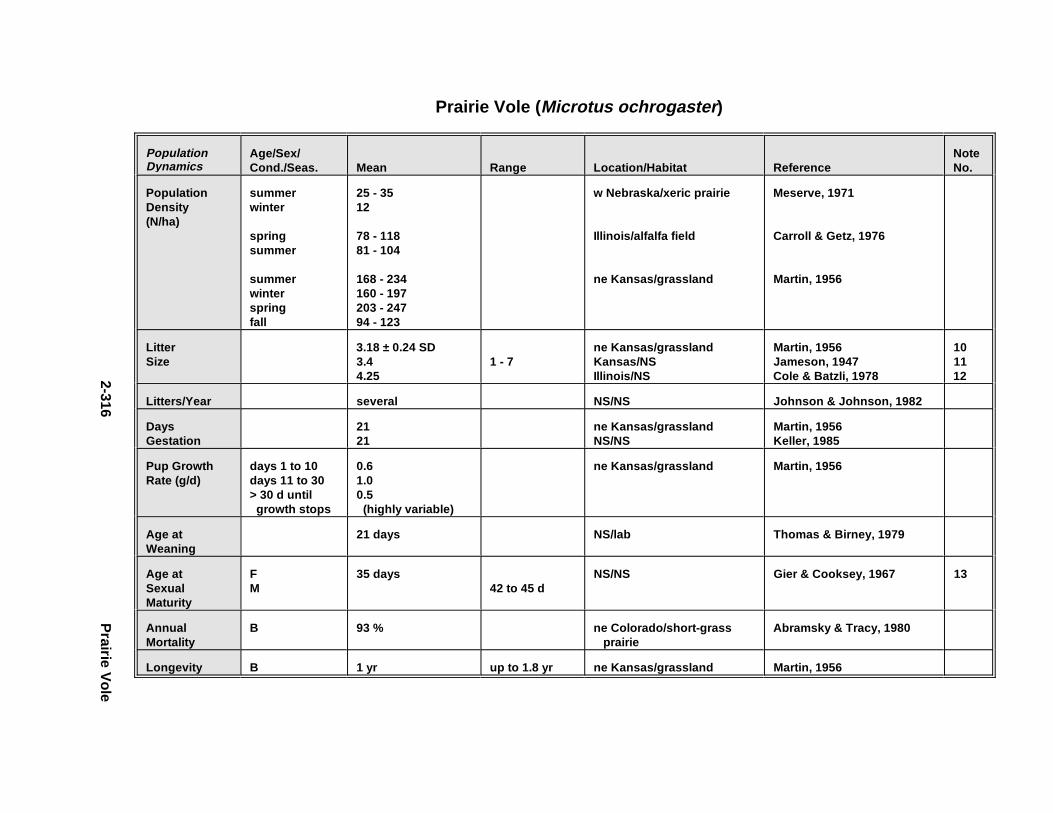
Prairie Vole ( Microtus ochro gaster )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Popul ation summer 25 - 35 w Nebr aska/xer ic prairie Meserve, 1971 Density win ter 12 (N/ha)
spr ing 78 - 118 Illinoi s/alfalfa field Carroll & Getz, 1976 sum mer 81 - 104
sum mer 168 - 234 ne Kansas/ grassl and Martin, 1956 winter 160 - 197 spr ing 203 - 247 fall 94 - 123
Li tter 3.18 ± 0.24 SD ne Kansas/ grassl and Martin, 1956 10 Size 3.4 1 - 7 Kansas/ NS Jameson, 1947 11
4.25 Illin ois/NS Cole & Batzli, 1978 12
Li tters/Year sever al NS/NS Johnson & Johnson, 1982
Days 21 ne Kansas/ grassl and Martin, 1956 Gestation 21 NS/NS Kel ler, 1985
Pup Growth days 1 to 10 0.6 ne Kansas/ grassl and Martin, 1956 Rate (g/d) days 11 to 30 1.0
> 30 d unt il 0.5 growt h stops (highl y var iable)
Age at 21 days NS/lab Thom as & Bi rney, 1979 Weaning
Age at F 35 days NS/NS Gier & Cooksey , 1967 13 Sexual M 42 to 45 d Maturity
Annual B 93 % ne Colorado/shor t-grass Abr amsky & Tracy, 1980 Morta lity prairie
Longevi ty B 1 yr up t o 1.8 yr ne Kansas/ grassl and Martin, 1956
Range
2-316 P
rairie Vo
le

�
�
Prairie Vole ( Microtus ochro gaster )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating May to Oct NS Keller, 1985; Martin, 1956
Partur ition May to Oct NS Keller, 1985; Martin, 1956
Molt any time NS Jameson, 1947 13
1 Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s (summer) from Abr amsky and Tr acy (1980). 2 Est imated usi ng equat ion 3-48 (Nagy, 1987) and body weight s (summer) from Abr amsky and Tr acy (1980).
2-317 P
rairie Vo
le
3 Est imated f rom ingest ion r ate for diet of oats (74 to 78 percent of total weight of diet) and dr y grass, assum ing 31 t o 34 g body weight. Diet was low in water (probabl y less t han 10 per cent ).
4 Measur ed water drunk f rom water bot tles; diet consi sted of rolled oat s with sunf lower seeds; temperature 28�C. 5 Measur ed wat er drunk; diet of dry food. 6 Temperature 21�C; dry air. 7 Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s (summer) from Abr amsky and Tr acy (1980). 8 Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s (summer) from Abr amsky and Tr acy (1980). 9 Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s (summer) from Abr amsky and Tr acy (1980).
10 Determined from pup count , whi ch may under est imate litter size at bi rth.11 Cited in Kel ler (1985); embryo or pup count .12 Cited in Kel ler (1985); embryo or placent al scar count .13 Cited in Stalling (1990).

References (including A ppendix)
Abramsky, Z.; Tracy, C. R. (1980) Relation b etween home range size and regula tion of popula tion s ize in Microtus ochrogaster . Oikos 34: 347-355.
Agnew, W. J.; Uresk, D. W.; Hansen; et al . (1988) Arthropod c ons umpt ion by smal l mammal s on prairie dog c olonie s and adjacent ungr azed mixed grass p rairie in wester n South Dakota. In: Uresk, D. W.; Schenbeck, G. L.; Cefki n, R., tech . coord. Eight h Great Plains wildlif e damage control workshop pr oceedings ; April 28-30, 1987; Rapid Cit y, Sout h Dakota. For t Collins , CO: U.S. Departmen t of Agriculture, Forest Ser vice, Rocky Mount ain For est an d Range Experiment Stat ion; pp. 81-87.
Barbour, R. W. (1963) Microtus: a simple method of r ecording t ime spent in t he nest. Science 141: 41.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Boons tra, R.; Krebs, C. J.; Gaines, M. S.; et al. (1987) Natal philopa try and br eeding systems i n voles (Microtus spp.). J. Anim. Ecol. 56: 655-673.
Bradley, S. R. (1976) Temperature regula tion a nd bioe nergetics of so me microt ine r odents [Ph.D. disser tat ion] . Ithaca, NY: Cornell Universi ty.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Carroll, D.; Getz, L. L. (1976) Runway u se and popula tion de nsity in Microtus ochrogaster . J. Mammal . 57: 772-776.
Chew, R. M. (1951) The water exch anges of so me smal l mammal s. Ecol. Monogr . 21: 215-225.
Cole, F. R.; Batzli, G. O. (1978) Influence of supplemen tal feeding on a vole popula tion. J. Mammal . 59: 809-819.
Colv in, M. A.; Colv in, D. V. (1970) Breeding a nd fecundit y of si x species of vole s (Microtus). J. Mammal . 51: 417-419.
Cook , J. C.; Topping, M. S.; Stombaugh, T. A. (1982) Food ha bit s of Microtus ochragaster and Peromyscu s man iculatus in symp atry. Trans. Misso uri Acad. Sci . 16: 17-23.
Corthum, D. W., Jr . (1967) Reproduc tion a nd dur ation of placental scar s in the prairie vole and the easter n vole. J. Mammal . 48: 287-292.
2-318 Prairie Vole

Dice, L. R. (1922) Some facto rs affect ing t he dis tr ibut ion of th e prairie vole , forest d eer mous e, and pr airie deer mous e. Ecology 3: 29-47.
Dupr e, R. K. (1983) A comparison of th e water relations of th e his pid c otton r at, Sigmodon hispidus , and the prairie vole, Michrotus ochrogaster . Comp. Bioc hem. Physiol. 75A: 659-663.
Eadie, R. W. (1952) Shrew predation a nd v ole popula tions on a loc alized area. J. Mammal . 33: 185-189.
Fitch , H. S. (1957) Aspects o f reproduc tion a nd d evelopment in the prairie vole (Microtus ochrogaster ). Univ . Kansas Publ., Mus. Nat. Hist. 10: 129-161.
Fleharty, E. D.; Olson, L. E. (1969) Summer food ha bit s of Michrotus ochrogaster and Sigmodon hi spidus . J. Mammal . 50: 475-486.
Fleharty, E. D.; Krause, M. E.; Stinnett, D. P. (1973) Body co mpos it ion, energy cont ent, and lipid cycl es of fo ur species of rodents. J. Mammal . 54: 426-438.
Gaines, M. S.; Johns on, M. L. (1982) Home range size and popula tion dy namics in t he prairie vole, Microtus ochrogaster . Oikos 39: 63-70.
Gaines, M. S.; Rose, R. K. (1976) Popula tion dy namics of Microtus ochrogaster in easter n Kansas. Ecology 57: 1145-1161.
Getz, L. L.; Hofman n, J. E.; Klatt, B. J.; et al . (1987) Four teen years of popula tion fluc tuations of Microtus ochrogaster and M. penns ylvanicus in east cen tral Illinois . Can. J. Zool. 65: 1317-1325.
Gier, H. T. (1967) The Kansas smal l mammal census: terminal report. Trans. Kansas Acad. Sci . 70: 505-518.
Gier, H. T.; Cooksey, B. F., Jr . (1967) Michrotus ochrogaster in the laboratory. Trans. Kansas Acad. Sci . 70: 256-265.
Golley, F. B. (1961) Interact ion of n atal ity, mortal ity an d movemen t during one annua l cycl e in a Microtus popula tion. Am. Midl. Nat. 66: 152-159.
Harvey, M. J.; Barbour, R. W. (1965) Home range of Microtus ochrogaster as determined by a modif ied minim um ar ea method. J. Mammal . 46: 398-402.
Jameso n, E. W., Jr . (1947) Natural histo ry of th e prairie vole (mammal ian genus Michrotus). Misc. Publ. Mus. Nat. His t. Univ . Kansas 1: 125-151.
Jike, L.; Batzli, G. O.; Getz, L. L. (1988) Home ranges of prairie voles as d etermined by radiot racking a nd by pow dertracking. J. Mammal . 69: 183-186.
2-319 Prairie Vole

Johns on, M. L.; Johns on, S. (1982) Voles (Microtus species). In: Chapman, J. A.; Feldha mer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 326-353.
Jones, J. K. Jr .; Armstr ong, R. S.; Hoffman , R. S.; et al . (1983) Mammal s of th e Great Nor thern Plains . Linc oln, NE: Univ ersi ty of Nebraska Pr ess.
Kaufman , D. W.; Fleharty, E. D. (1974) Habitat sel ect ion by nine sp ecies of r odents in north-central Kansas. Southwest. Nat. 18: 443-451.
Keller, B. L. (1985) Reproducti ve patter ns. In: Tamar in, R. H., ed. Biology of new world Microtus. Amer ican Society o f Mammalogi sts; Special Publi cation No. 8; pp. 725-778.
Keller, B. L.; Krebs, C. J. (1970) Microtus popula tion biology . III. Reproduc tive changes in fluc tuating popula tions of M. ochrogaster and M. penns ylvanicus in s out hern India na, 1965-1967. Ecol. Monogr . 40: 263-294.
Kenney, A. M.; Evans, R. L.; Dewsb ury, D. A. (1977) Posti mplantat ion pr egnancy dis rupt ion in Microtus orchogaster , Microtus penns ylvanicus , and Peromyscu s man iculati s. J. Reprod. Fert. 49: 365-367.
Klatt, B. J. (1985) The role of habitat p reference and interspeci fic co mpeti tion in determining t he loc al dis tr ibut ion of Microtus penns ylvanicus and M. ochrogaster in central Illinois (ab str act). Bull. Ecol. Soc. Am. 66: 209.
Klatt, B. J.; Getz, L. L. (1987) Vegetat ion c haracter isti cs o f Microtus ochragaster and M. pennsyl vanicus habitats i n east-cen tral Illinois . J. Mammal . 68: 569-577.
Krebs, C. J. (1977) Competi tion b etween Microtus penns ylvanicus and Microtus ochragaster . Am. Midl. Nat. 97: 42-49.
Kruckenbe rg, S. M.; Gier, H. T.; Dennis , S. M. (1973) Postn atal development of th e prairie vole , Microtus ochrogaster . Lab. Anim. Sci . 23: 53-55.
Lom olino, M. V. (1984) Immi grant sel ect ion, predation, and t he dis tr ibut ion of Microtus pennsyl vanicus and Blarina brevicauda on islands. Am. Nat. 123: 468-483.
Martin, E. P. (1956) A popula tion s tudy of th e prairie vole ( Michrotus ochragaster ) in northeaster n Kansas. Univ . Kansas Publ. Mus. Nat. His t. 8: 361-416.
Martin, E. P. (1960) Distr ibut ion of native mammal s among t he communit ies of th e mixed prairie. Fort Hays Stu d. N. S. Sci . Ser. 1: 1-26.
Meserve, P. L. (1971) Popula tion e cology of th e prairie vole , Microtus ochrogaster , in t he wester n mixed prairie of Nebraska. Am. Woodl. Nat. 86: 417-433.
2-320 Prairie Vole

Morrison, P. R.; Dieter ich, R.; Presto n, D. (1976) Breeding a nd r eproduc tion of fi fteen wi ld rodents maint ained as labor atory colonie s. Lab. Anim . Care 26: 237-243.
Myers, J. H.; Krebs, C. J. (1971) Genetic, behavioral, and reproducti ve attr ibutes o f dis persing f ield v oles Microtus penns ylvanicus and Michrotus ochrogaster . Ecol. Monogr . 41: 53-78.
Nadeau, J. H. (1985) Ontogeny. In: Tamar in, R. H., ed. Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 254-285.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Ostfel d, R. S.; Pugh, S. R.; Seamon, J. O.; et al . (1988) Space use and reproducti ve su ccess in a popula tion of meadow vo les. J. Anim. Ecol. 57: 385-394.
Quick, F. W. II (1970) Smal l mammal popula tions in an old f ield c ommunit y [Ph.D. disser tat ion] . Louis ville , KY: Univ ersi ty of Louis ville .
Richmond, M. E. (1967) Reproduc tion of th e vole , Microtus orchogaster [Ph.D. disser tat ion] . Colum bia, MO: Univ ersi ty of Missour i.
Richmond, M. E.; Conaway, C. H. (1969) Management, breeding a nd r eproduc tive performance of th e vole, Microtus ochrogaster in a labor atory colony . Lab. Anim . Care 19: 80-87.
Rolan, R. G.; Gier, H. T. (1967) Correlation of emb ryo and pl acental scar count s of Peromyscu s man iculati s and Microtus orchogaster . J. Mammal . 48: 317-319.
Rose, R. K.; Gaines, M. S. (1978) The reproducti ve cycl e of Microtus orchogaster in easter n Kansas. Ecol. Monogr . 48: 21-42.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Stalling, D. T. (1990) Microtus ochrogaster . Amer ican Society o f Mammalogi sts; Mammal ian Species No. 355; 9 pp.
Swihart, R. K.; Slade, N. A. (1989) Differ ences i n home-range size between sexes o f Microtus ochrogaster . J. Mammal . 70: 816-820.
Tamar in, R. H. (1985) Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8.
Thomas, J. A.; Birney, E. C. (1979) Parental care and mat ing system o f th e prairie vole, Microtus ochrogaster . Behav. Ecol. Sociobiol. 5: 171-186.
Wooster , L. D. (1939) An ecologic al evalua tion of predatees o n a mixed prairie area in wester n Kansas. Trans. Kans. Acad. Sci . 42: 515-517.
2-321 Prairie Vole

Wunde r, B. A. (1985) Energetics an d thermoregula tion. In: Tamar in, R. H., ed. Biology of new wo rld Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 812-844.
Wunde r, B. A.; Dobk in, D. S.; Gettinge r, R. D. (1977) Shifts o f th ermogenesis in the prairie vole ( Microtus ochrogaster ), str ategies fo r survival in a seaso nal environment. Oecologia (B erl.) 29: 11-26.
Zimmer man, E. G. (1965) A comparison of habitat an d food of two species of Microtus. J. Mammal . 46: 605-612.
2-322 Prairie Vole

2.2.9. Meadow Vole (v oles)
Order Rodent ia Family Murid ae (subfami ly Arv icolin ae). New wo rld voles are smal l, herbivorous rodents th at reside in all areas of Canada and the United States wh ere there is good gr ass co ver . Their presence is ch aracter ized by narrow runways th rough matted grasses. Microtus species are adapted to unde rground, ter restr ial, and someti mes semi amphibious habitats (J ohns on and J ohns on, 1982). They are acti ve by day and night , feeding m ainly on s hoot s, grasses, and bark. Voles are prey fo r hawks an d owls as wel l as sever al mammal ian predators su ch as sh ort-tai led shrews, badgers, and foxes (J ohns on and J ohns on, 1982; Eadie, 1952).
Selected species
The meadow vo le (Microtus penns ylvanicus ) makes i ts b urrows a long s urface runways i n grasses o r other herbaceous veg etat ion. It is th e most wide ly dis tr ibut ed smal l grazing he rbiv ore in Nor th Amer ica and is found ov er mo st of th e nor thern half of th e United States. Meadow vo les have been used in bioassays to indi cate th e presence of toxins in t heir foods (K endall and Sherwood, 1975, ci ted in Reich, 1981; Schillinge r and Elliot , 1966). Although pr imarily ter restr ial, the meadow vo le also is a str ong s wimmer (Johns on and J ohns on, 1982).
Body size. The meadow vo le measu res 8.9 to 13 cm i n length (head and body) and has a 3.6- to 6.6-cm tai l. They weigh b etween 20 and 40 g depending on a ge, sex, and loc ation (see tab le). Mature males are approximatel y 20 percent heavier than femal es (Boons tra and Rodd, 1983). Meadow vo les lose weight dur ing t he wint er, reaching a low around Fe bruary, th en regain w eight dur ing s pring a nd s ummer , reaching a high a round Augus t in m any popula tions (see tab le; Iverson and Turner, 1974).
Habitat . The meadow vole inha bit s gr assy fi elds, marshes, and bogs (Getz, 1961a). Compared wi th the prairie vole, the meadow vo le prefers fi elds wi th more grass, more cover , and fewer woody plants (Getz, 1985; Zimmer man, 1965). The meadow vo le also tends to inha bit mois t to wet h abitats, whereas th e prairie vole is relatively uncommo n in si tes wi th standing water (Getz, 1985).
Food ha bi ts . Meadow vo les co nsume green succu lent veg etat ion, sedge s, seeds, root s, bark, fungi, insects, and animal matter (see tab le). They are agricultural pests i n some areas, feeding on p astu re, hay, and grain (Johns on and J ohns on, 1982; Burt and Grossen heider, 1980). At high popula tion de nsit ies, th e meadow vole has been know n to girdle trees, which can damage orchards (Byers, 1979, ci ted in Reich, 1981). In seaso nal habitats, meadow vo les favo r green vegetat ion w hen it is availa ble and c ons ume other foods mo re when green vegetation is l ess avai lable (Johns on and J ohns on, 1982; Riewe, 1973; Getz, 1985). Although Zi mmer man (1965) found s ome evide nce of food s election, he found t hat meadow vo les generally ate th e most co mmo n plants i n their habitat. Meadow vole s liv ing on pr airies cons ume more seeds and fewer dicots an d monoc ots than vole s in a blue grass h abitat (Lindr oth and Ba tzli, 1984). The meadow vo le's large cecu m allow s it to have a high dig esti ve effi ciency o f 86 to 90 percent (Golley, 1960). Copr opha gy (eating of feces) h as been observed in th is sp ecies (Ouellete and Heisinge r, 1980).
2-323 Meadow Vole

�
Temperature regulation and molt. In wi nter, Microtus species do not unde rgo hibe rnation or torpor ; instead , they are acti ve year round ( Didow and H aywar d, 1969; Johns on and J ohns on, 1982). Behaviors th at help meadow vo les to maintain their body temp erature include th e use of bur rows, r unways, nests, and snow co ver for insulation. They also can change when they are acti ve; when temp eratures exceed 20�C, meadow voles are most acti ve at night (Getz, 1961b; Johns on and J ohns on, 1982). In winter , meadow vo les increase th eir brown fat co ntent (a maj or si te of th ermoregulatory heat produc tion) . Mature indiv idua ls average 0.5 percent brown fat i n summer , increasing t o 1.7 percent in early wi nter ; juveniles aver age 1.0 percent in the summer , increasing t o 2.3 percent in t he wint er (Didow and H aywar d, 1969). Voles unde rgo t hree molt s: juv enile , pos tjuv enile , and adult . The timing v aries by sp ecies (Johns on and J ohns on, 1982). Adult Arv icolin ae also unde rgo w int er and s ummer molts (J ohns on and J ohns on, 1982).
Breeding acti vi ties and social organization. Meadow vole s are poly gynous (McShea, 1989). Males fo rm a hierarchy in which the most d ominant mal e voles breed (Boons tra and Rodd, 1983). Voles produce l itter s th roughout th e breeding seaso n, the number of l itter s per seaso n increases wi th decreasing la ti tude (Johns on and J ohns on, 1982).
Home range and r esour ces. The area encompassed by a mead ow vo le's home range depends on seaso n, habitat, popula tion de nsity, and t he age and sex o f th e animal. Summer ranges ten d to be larger than winter ranges, and ranges in marshes ten d to be larger than ranges in meadows (Getz , 1961c; Reich, 1981). Home range size also decline s wi th inc reasing popula tion de nsity (Getz, 1961c; Tamar in, 1977a). Female meadow vo les defend ter ritories against o ther femal es, whereas mal e home ranges are larger and over lap wi th home ranges of both sexes (M adison, 1980; Ostfel d et al ., 1988; Wolff, 1985). Meadow vole s build r unways i n grasses an d vegetat ion a t the ground' s surface an d use th e runways fo r foraging a bout 45 pe rcent of th e time, depending on weath er and other facto rs (Gauthier and Bide r, 1987). The meadow vo le exhibit s dayti me acti vi ty wh ere dense co ver is avai lable and becomes mo re crepuscu lar wi th less co ver (Graham, 1968, ci ted in Reich, 1981). All Microtus species apparent ly do s ome bur rowing, excavat ing unde rground n ests that are used as nur ser ies, resting a reas, and as sh elter from sever e weath er (Johns on and Johns on, 1982). Nests ar e built wi th the use of dead grass i n patch es of dense, live grass; widened spaces, cal led forms, are used off mai n runways (A mbrose, 1973).
Popul ation de nsity . Meadow vole popula tion de nsit ies fluc tuate wi dely fr om seaso n to seaso n and year to year , someti mes cr ashing t o near zero before recover ing in a few years to densi ties of sever al hundr ed per hectar e (Boons tra and Rodd, 1983; Lindr oth and Batzli, 1984; Getz et al ., 1987; Myers and Krebs, 1971; Taitt an d Krebs, 1985). Krebs and Myers (1974) noted popula tion cycl es of 2 to 5 yr , whereas Tamar in (1977b) reported 3- to 4-year popula tion cycl es in southeaster n Massach usetts. However , Getz et al . (1987) found no indi cation of mult iannua l abunda nce cycl es in their three habitat stu dy (i .e., blue grass, tallgr ass p rairie, and alfal fa) in east cen tral Illinois . Meadow vole s avoid s hor t-tai led shrews (Fulk , 1972), and the vole popula tion de nsity decreases as th e number of sh ort-tai led shrews i n the area increases (Ead ie, 1952).
2-324 Meadow Vole

�
�
�
�
�
�
Popul ation dy namics. Voles reach sexu al matu rity u sually wi thin sever al weeks after birth, wi th femal es matu ring be fore males, but still c ont inue to grow fo r sever al mont hs (Johns on and J ohns on, 1982). Innes (1978) reported that l itter size is inde pendent of lati tude or elevat ion. However , summer litter s wer e, on average, 14 percent larger than litter s produc ed dur ing ot her seaso ns, and larger femal es produced larger litter s (Keller and Krebs, 1970). Young f rom th e spring a nd early su mmer litter s reached adult weight in about 12 wk (Brown, 1973). Mortal ity r ates ar e highest i n postn est ling juv enile s and y oung adults an d lowest i n nest lings (ag es 1 to 10 d) (Golle y, 1961). Dispersing meadow vole s (predom inant ly young m ales) ten d to weigh l ess th an resident mead ow vo les (Boons tra et al., 1987; Myers and Krebs, 1971; Boons tra and Rodd, 1983; Brochu et al ., 1988).
Simi lar species (fr om general references)
� The Calif ornia vole ( Microtus cal ifor nicus) is larger than the meadow vo le (12 to 14 cm he ad and body ) and is found t hroughout Calif ornia and s out hern Oregon. It inha bit s freshwater and sal twater marshy areas, wet mead ows, and grassy hills ides from th e seash ore to the mountains and feeds on green vegetat ion.
� Townsend's vo le (Microtus townsendi i) usually is found ne ar water in moist fields, sedges, tu les, and meadows (fr om ti dewater to alpine meadows). Its range is l imi ted to extr eme northwester n California, wester n Oregon a nd Washingt on, and s out hern Br it ish Colum bia (inha bit s sever al islands off th e coast o f Washingt on and Br it ish Colum bia). It is easily dis tinguis hed by its large size (12 to 16 cm) an d black-b rown color .
� The montane vole (Microtus mont anus ) (mount ain v ole) is slight ly larger (10 to 14 cm) th an the meadow vo le and is found in v alleys o f th e mount ainous Great Basin area of th e wester n and northwester n United States.
� The long -taile d vole (Microtus longi caudus ) (tai l 5 to 9 cm) i s s light ly larger (11 to 14 cm) th an the meadow vo le. It is found in t he wester n United States and Canada to Alaska an d lives a long s treambanks, in mount ain m eadow s, someti mes i n dry si tuations , and in br ushy ar eas during w int er. In addit ion to grasses an d bark, it feed s on bulbs . It nests ab ove ground in w int er and burrows i n summer .
� The creeping v ole (Microtus oregoni ) (Oregon v ole) (10 to 11 cm) i s an inha bit ant of wester n Oregon a nd Washingt on and extr eme northwest Calif ornia . Seldom abov e ground, it sp ends mo st of its time bur rowing through f orest floor duf f or grass r oots. It l ives i n forests, brush, and grassy areas.
� The sagebrush vole (Lagurus curtatu s) (9.7 to 11 cm) l ives i n loos e soil a nd arid c ondit ions and feeds on green vegetat ion, especially sag ebrush. It also bur rows around s agebrush; a vole found liv ing in s agebrush is al most cer tainly this sp ecies.
2-325 Meadow Vole

General references
Burt and Grossen heider (1980); Reich (1981); Johns on and J ohns on (1982); Tamar in (1985).
2-326 Meadow Vole

Meadow Vole (Microtus penns ylvanicus )
2-327 M
eado
w V
ole
Factors Age/Sex/ (95% CI of Note Cond. /Seas. Mean mean) Locat ion Reference No.
Range or
Body Weight A M summer 40.0 ± 8.3 SE Quebec, Canada Brochu et al., 1988 (g) A F sum mer 33.4 ± 8.2 SE
A M spr ing 52.4 Ontario, Canada Boonst ra & Rodd, 1983 A F spr ing 43.5 A M & F Manito ba, Canada Anderson et al., 1984 spr ing 26.0 sum mer 24.3 fall 17.0 winter 17.5
A M avg. all yr 35.5 ± 0.1 SE sout h Indi ana Myers & Kr ebs, 1971 A F avg. all yr 39.0 ± 0.3 SE
neonat e M & F 2.1 1.6 - 3.0 not speci fied Hamilton, 1941 1 neonate M & F 2.3 ± 0.1 SD Innes & Milla r, 1981 2
Pup Growth bi rth - 21 days 0.95 sout h Michi gan/old field Gol ley, 1961 Rate (g/d) 22 - 33 days 0.81
34 - 54 days 0.45 55 - 103 days 0.19
Body Fat (g ) summer: Alberta , Canada Milla r, 1987 J F 0.37 ± 0.04 SE A F gest ating 1.20 ± 0.15 SE A F lactating 0.60 ± 0.09 SE
Metabol ic basal 60.0 lab Wiegert, 1961 3 Rate average dai ly 82.8 ± 12 SD 43.2 - 146 lab Morrison, 1948 4 (lO /kg-d)2
Metabol ic Rate A M basal 166 est imated 5 (kcal /kg-d) A F basal 175
A B avg. dai ly 395 lab 25-30��C Pearson, 1947 A M free-living 357 (170 - 747) est imated 6 A F f ree-living 485 (231 - 1,020)

Meadow Vole (Microtus penns ylvanicus )
2-328 M
eado
w V
ole
Factors Age/Sex/ (95% CI of Note Cond. /Seas. Mean mean) Locat ion Reference No.
Range or
Food Ingest ion Rate (g/g-d) 0.30 - 0.35 Russi a Ognev, 1950 7
(cal /g-d) A M shor t-day 370 ± 20 SE NS Dark et al., 1983 8 A M long- day 410 ± 10 SE
Water A B 0.21 ± 0.02 SE NS Ernst , 1968 9 Ingest ion Rate (g/g-d) A B 0.14 est imated 10
Inhal ation A M 0.052 est imated 11 Rate (m /d) A F 0.0443
Surface Ar ea A M 161 est imated 12 (cm ) A F 1432
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
dicot shoot s 41 60 66 12 Illin ois/bluegrass Lindroth & Batzli, 1984 monocot 50 26 9 40 shoot s
seeds 1 9 1 13 (% vol ume; stomach root s 0 1 12 34 cont ents) fungi 6 4 10 0 insect s 2 0 2 1
dicot shoot s 53 65 41 41 Illinoi s/tallgrass pr airie Lindr oth & Bat zli, 1984 monocot 23 29 12 5 shoot s (% vol ume; stomach
seeds 7 1 16 36 cont ents) root s 4 0 6 17 fungi 12 1 20 0 insect s 1 4 5 1

Meadow Vole (Microtus penns ylvanicus )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Home Range A M sum mer 0.019 ± 0.011 SD Virginia/old field Madison, 1980 Size (ha) A F sum mer 0.0069 ±0.0039 SD
A B sum mer 0.014 Mont ana/alluvial bench Dougl ass, 1976 A B w inter 0.0002
A M sum mer 0.083 ± 0.037 SD Massachuset ts/grassy Ostfeld et al., 1988 A F sum mer 0.037 ± 0.020 SD meadow
Popul ation A B 96 - 549 Ontario, Canada/grassl and Boonst ra & Rodd, 1983 Density A B 2 - 28 Illin ois/bluegrass Lindroth & Batzli, 1984 (N/ha) A B 25 - 163 Indi ana/grassl and Myers & Kr ebs, 1971
fall 28 - 51 Michi gan/grass-sedge Getz, 1961a winter 20 - 51 marsh spr ing 22 - 53 sum mer 38 - 64
Li tter 3.82 1 - 11 Mani toba, Canada/NS Iverson & Tur ner, 1976 13 Size 4.46 1 - 9 Indi ana/NS Corthum , 1967 13
6.05 1 - 8 Pennsy lvani a/NS Goin, 1943 13
Li tters/Year sever al NS/NS Bailey, 1924 14
Days 21.0 ± 0.2 SD NS/NS Kenney et al., 1977 2 Gestation
Age at 21 s Michi gan/NS Gol ley, 1961 Weaning (d)
Age at F at least 3 wk NS/NS Johnson & Johnson, 1982 Sexual M at least 6-8 wk Maturity
Mortality nest lings (0-10 g) 50% sout h Michi gan/old field Gol ley, 1961 Rates juveni les (11-20 g) 61%
young adul ts (21-30 g) 58% adul ts (31-50 g) 53% old adul ts (>50 g) 100%
Range
2-329 M
eado
w V
ole

1234 �
56789
101112131415
Meadow Vole (Microtus penns ylvanicus )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Locat ion/Habi tat Reference No.
Longevi ty 2-3 mo NS Beer & MacLeod, 1961 9 < 24 mo NS Johnson & Johnson, 1982
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating early Apr il mid-October Mani toba, Canada Mihok, 1984 Oct. - Nov. Michi gan (fall-winter peak) Getz, 1960 15 Apr il - June Michi gan (spr ing-sum mer Getz, 1960 15
peak)
Dispersal fall/w in ter Indiana/grassl and Myers & Kr ebs, 1971
sum mer (females) Massachuset ts/coast al field Tamarin, 1977b winter (males)
Range
Cited in Reich (1981) and Johnson and Johnson ( 1982). Cited in Nadeau (1985).
2-330 M
eado
w V
ole
Body weight 35.6 g; temperature not speci fied; ci ted in Deavers and Hudson ( 1981).Temperature 15 to 25�C; weight 26.2 to 32 g.Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom Ander son et al. (1984).Est imated usi ng equat ion 3-48 (Nagy, 1987) and body weight s f rom Ander son et al. (1984).Cited in Johnson and Johnson ( 1982).Shor t-day phot oper iod = 10 h of light , 14 of dark; long- day phot oper iod = 14 h of light , 10 of dark.Cited in Reich (1981).Est imated usi ng equat ions 3- 17 (Calder and Br aun, 1983) and 3-18 and body weight s f rom Ander son et al. (1984).Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Ander son et al. (1984).Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Ander son et al. (1984).Cited in Kel ler (1985).Cited in Johnson and Johnson ( 1982).Cited in Getz (1961b).

References (including A ppendix)
Ambr ose, H. W., III. (1973) An experimental stu dy of so me facto rs affect ing t he spatial and temp oral acti vi ty o f Microtus penns ylvanicus . J. Mammal . 54: 79-100.
Anderson, M.; Prieto, J.; Rauch, J. (1984) Seasonal changes in white and brown adipos e tissu es in Clethrionomys g apperi (red-backed vole) and in Microtus penns ylvanicus (meadow vo le). Comp. Bioc hem. Physiol. A Comp. Physiol. 79: 305-310.
Bailey, V. (1924) Breeding, feeding a nd ot her lif e habit s of meadow mi ce (Microtus). J. Agric. Res. 27: 523-526.
Barbehenn, K. R. (1955) A field stu dy of growth in Microtus penns ylvanicus . J. Mammal . 36: 533-543.
Beer, J. R.; MacLeod, C. F. (1961) Seasonal reproduc tion in t he meadow vo le. J. Mammal . 42: 483-489.
Benton, A. H. (1955) Observat ions on t he lif e his tory of th e nor thern pine mous e. J. Mammal . 36: 52-62.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Boons tra, R.; Rodd, F. H. (1983) Regula tion of breeding de nsity in Microtus pennsyl vanicus. J. Anim. Ecol. 52: 757-780.
Boons tra, R.; Krebs, C. J.; Gaines, M. S.; et al. (1987) Natal philopa try and br eeding systems i n voles (Microtus spp.). J. Anim. Ecol. 56: 655-673.
Bradley, S. R. (1976) Temperature regula tion a nd bioe nergetics of so me microt ine r odents [Ph.D. disser tat ion] . Ithaca, NY: Cornell Universi ty.
Brochu, L.; Caron, L.; Bergeron, J.-M. (1988) Diet quality an d body condit ion of dis persing and r eside nt vole s (Microtus penns ylvanicus ). J. Mammal . 69: 704-710.
Brooks, R. J.; Webster , A. B. (1984) Relations hips of seaso nal change to changes in age str ucture and body size in Microtus pennsyl vanicus. Carnegie Mus. Nat. Hist. Spec. Publ. No. 10; pp. 275-284.
Brown, E. B., III. (1973) Changes in patter ns of seaso nal growth of Microtus pennsyl vanicus. Ecology 54: 1103-1110.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Byers, R. E. (1979) Meadow vo le control using a nticoagula nt baits. Hor t. Sci . 14: 44-45.
2-331 Meadow Vole

Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Corthum, D. W., Jr . (1967) Reproduc tion a nd dur ation of placental scar s in the prairie vole and the easter n vole. J. Mammal . 48: 287-292.
Dark, J.; Zucker , I. (1986) Photoperiodic r egula tion of body mass an d fat r eserves i n the meadow vole . Physiol. Behav. 38: 851-854.
Dark, J.; Zucker , I.; Wade, G. N. (1983) Photoperiodic r egula tion of body mass, food in take, and r eproduc tion in meadow vole s. Am. J. Ph ysiol. 245: R334-R338.
Deavers, D. R.; Huds on, J. W. (1981) Temperature regula tion in t wo rodents (Clethrionomys gapperi and Peromyscu s leucopus ) and a shrew (Blarina brevicauda) inha bit ing t he same en vironment. Physiol. Zool. 54: 94-108.
Didow , L. A.; Haywar d, J. S. (1969) Seasonal var iations in t he mass an d composi tion of brown adipos e tissu e in t he meadow vole , Microtus penns ylvanicus . Can. J. Zool. 47: 547-555.
Dieter ich, R. A.; Presto n, D. J. (1977) The meadow vo le (Microtus penns ylvanicus ) as a laboratory animal. Lab. Anim. Sci . 27: 494-499.
Douglass, R. J. (1976) Spatial interact ions and m icrohabitat sel ect ions of two loc ally symp atr ic vo les, Microtus mont anus and Microtus penns ylvanicus . Ecology 57: 346-352.
Dueser , R. D.; Wilson, M.; Rose, R. K. (1981) Attr ibutes o f dispersing meadow vo les in open-gr id popula tions . Acta T heriol. 26: 139-162.
Eadie, R. W. (1952) Shrew predation a nd v ole popula tions on a loc alized area. J. Mammal . 33: 185-189.
Ernst, C. H. (1968) Kidne y effi ciencies of th ree Pennsyl vania mice. Trans. Kentucky A cad. Sci . 29: 21-24.
Fulk , G. W. (1972) The effect o f sh rews o n the space utiliza tion of vole s. J. Mammal . 53: 461-478.
Gauthie r, R.; Bide r, J. R. (1987) The effects o f weath er on runway u se by rodents. Can. J. Zool. 65: 2035-2038.
Getz, L. L. (1960) A popula tion s tudy of th e vole , Microtus penns ylvanicus . Am. Midl. Nat. 64: 392-405.
Getz, L. L. (1961a) Facto rs influencing t he loc al dis tr ibut ion of Microtus and Synaptomys in sout hern Michiga n. Ecology 42: 110-119.
2-332 Meadow Vole

Getz, L. L. (1961b) Responses o f smal l mammal s to live-tr ap and weath er condit ions . Am. Midl. Nat. 66: 160-170.
Getz, L. L. (1961c) Home ranges, ter ritoriality, and movemen t of th e meadow vo le. J. Mammal . 42: 24-36.
Getz, L. L. (1985) Habitat. In: Tamar in, R. H., ed. Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 286-309.
Getz, L. L.; Hofman n, J. E.; Klatt, B. J.; et al . (1987) Four teen years of popula tion fluc tuations of Microtus ochrogaster and M. penns ylvanicus in east cen tral Illinois . Can. J. Zool. 65: 1317-1325.
Goin, O. B. (1943) A stu dy of indiv idua l variation in Microtus penns ylvanicus pennsyl vanicus. J. Mammal . 24: 212-223.
Golley, F. B. (1960) Anatomy o f th e digesti ve tr act o f Microtus. J. Mammal . 41: 89-99.
Golley, F. B. (1961) Interact ion of n atal ity, mortal ity an d movemen t during one annua l cycl e in a Microtus popula tion. Am. Midl. Nat. 66: 152-159.
Graham, W. J. (1968) Daily acti vi ty p atter ns in the meadow vo le, Microtus penns ylvanicus [Ph.D. disser tat ion] . Ann A rbor , MI: Univ ersi ty of Michiga n.
Hamilton, W. J., Jr . (1937) Growth and life sp an of th e field mouse. Am. Nat. 71: 500-507.
Hamilton, W. J., Jr . (1941) Reproduc tion of th e field m ous e (Microtus penns ylvanicus ). Cornell Univ . Agric. Exp . Sta. Mem. 237: 3-23.
Harris, V. T. (1953) Ecologic al relations hips of meadow vo les and rice rats i n tidal marshes. J. Mammal . 34: 479-487.
Innes, D. G. (1978) A reexami nation of li tter sizes in some North Amer ican microtines. Can. J. Zool. 56: 1488-1496.
Innes, D. G.; Milla r, J. S. (1979) Growth of Clethrionomys g apperi and Microtus pennsyl vanicus in captivi ty. Growth 43: 208-217.
Innes, D. G.; Milla r, J. S. (1981) Body weight , li tter size, and energetics o f reproduc tion in Clethrionomys g apperi and Microtus penns ylvanicus . Can. J. Zool. 59: 785-789.
Iverson, S. L.; Turner, B. N. (1974) Winter weight dynamics in Microtus penns ylvanicus . Ecology 55: 1030-1041.
Iverson, S. L.; Turner, B. N. (1976) Smal l mammal radioe cology : natural reproduc tive patter ns of seven species. Pinawa, Manitoba: Whitesh ell Nuclear Research Establis hment; AECL-5393; 53 pp.
2-333 Meadow Vole

Johns on, M. L.; Johns on, S. (1982) Voles (Microtus species). In: Chapman, J. A.; Feldha mer , G. A., eds. Wild mammal s of North Amer ica; pp. 326-353.
Keller, B. L. (1985) Reproducti ve patter ns. In: Tamar in, R. H., ed. Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 812-844.
Keller, B. L.; Krebs, C. J. (1970) Microtus popula tion biology . III. Reproduc tive changes in fluc tuating popula tions of M. ochrogaster and M. penns ylvanicus in s out hern India na, 1965-1967. Ecol. Monogr . 40: 263-294.
Kendall, W. A.; Sherwood, R. T. (1975) Palatabilit y of l eaves o f tal l fescu e and reed canary-grass an d some of th eir alkaloids to meadow vo les. Agron. J. 67: 667-671.
Kenney, A. M.; Evans, R. L.; Dewsb ury, D. A. (1977) Posti mplantat ion pr egnancy dis rupt ion in Microtus orchogaster , Microtus penns ylvanicus , and Peromyscu s man iculati s. J. Reprod. Fert. 49: 365-367.
Kot t, E.; Robins on, W. L. (1963) Seasonal var iation in li tter size of th e meadow vo le in southern Ontar io. J. Mammal . 44: 467-470.
Krebs, C. J. (1977) Competi tion b etween Microtus penns ylvanicus and Microtus ochragaster . Am. Midl. Nat. 97: 42-49.
Krebs, C. J.; Myers, J. H. (1974) Popula tion cycl es in smal l mammal s. Adv. Ecol. Res. 8: 267-399.
Lee, C.; Horvath , D. J. (1969) Management of th e meadow vo le (Microtus penns ylvanicus ). Lab. Anim. Care 19: 88-91.
Lindr oth, R. L.; Batzli, G. O. (1984) Food ha bit s of th e meadow vole ( Microtus pennsyl vanicus) in blue grass an d prairie habitats. J. Mammal . 65: 600-606.
Lom olino, M. V. (1984) Immi grant sel ect ion, predation, and t he dis tr ibut ion of Microtus pennsyl vanicus and Blarina brevicauda on islands. Am. Nat. 123: 468-483.
Madison, D. M. (1978) Movemen t indi cato rs of reproducti ve even ts am ong femal e meadow voles as r evealed by radio-telemetr y. J. Mammal . 59: 835-843.
Madison, D. M. (1980) Space use and social str uctu re in meadow vo les, Microtus pennsyl vanicus. Behav. Ecol. Sociobiol. 7: 65-71.
McShea, W. J. (1989) Reproducti ve syn chrony and home range size in a ter ritorial microtine. Oikos 56: 182-186.
McShea, W. J.; Madison, D. M. (1989) Measurements o f reproducti ve tr aints (l itter size, pup growth, and bir th int erval) in a fi eld popula tion of meadow vo les. J. Mammal . 70: 132-141.
2-334 Meadow Vole

Mihok , S. (1984) Life histo ry profi les of boreal meadow vo les (Microtus penns ylvanicus ). Carnegie Mus. Nat. His t. Spec. Publ. No. 10; pp. 91-102.
Mihok , S.; Br ian, T. N.; Iverson, S. L. (1985) The character ization of vole popula tion dynamics. Ecol. Monogr . 55: 399-420.
Milla r, J. S. (1987) Energy reserves i n breeding smal l rodents. Symp . Zool. Soc. Lond. 57: 231-240.
Morrison, P.; Dieter ich, R.; Presto n, D. (1977) Body growth in s ixteen rodent sp ecies and subs pecies maint ained in la bor atory colonie s. Physiol. Zool. 50: 294-310.
Morrison, P. R. (1948) Oxyg en consumption in sever al smal l wild mammal s. J. Cell. Comp. Physiol. 31: 69-96.
Myers, J. H.; Krebs, C. J. (1971) Genetic, behavioral, and reproducti ve attr ibutes o f dis persing f ield v oles Microtus penns ylvanicus and Michrotus ochrogaster . Ecol. Monogr . 41: 53-78.
Nadeau, J. H. (1985) Ontogeny. In: Tamar in, R. H., ed. Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 254-285.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Ognev, S. I. (1950) Mammal s of th e U.S.S.R. and adjacent count ries. Translated from Russian by: Israel Progr am for Sci enti fic Translations (1964), Jerusalem; 626 pp.
Ostfel d, R. S.; Pugh, S. R.; Seamon, J. O.; et al . (1988) Space use and reproducti ve su ccess in a popula tion of meadow vo les. J. Anim. Ecol. 57: 385-394.
Ouellette, D. E.; Heisinge r, J. F. (1980) Reinge stion of feces b y Microtus penns ylvanicus . J. Mammal . 61: 366-368.
Pearson, O. P. (1947) The rate of meta bolis m of so me smal l mammal s. Ecology 29: 127-145.
Reich, L. M. (1981) Microtus penns ylvanicus . Mammal ian species. The Amer ican Society o f Mammalogi sts; Species No. 159; 8 pp.
Riewe, R. R. (1973) Food ha bit s of ins ular meadow vo les, Microtus penns ylvanicus ter raenovae (Rodentia: Criceti dae), in Notre Dame Bay, Newfoundla nd. Can. Field-Nat. 87: 5-13.
Schillinge r, J. A., Jr .; Elliot t, F. C. (1966) Bioassays fo r nutr itive val ue of indiv idua l alfal fa plants. Q. Bull. Michiga n Agric. Exp . Sta. 48: 580-590.
2-335 Meadow Vole

Schwar tz, B.; Mihok , S. (1983) Body co mpos it ion of meadow vo les co ntrasted during annua l and m ult iannua l popula tion f luc tuations (ab str act). Bull. Ecol. Soc. Am. 64: 102.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Taitt, M. J.; Krebs, C. J. (1985) Popula tion dy namics and cycl es. In: Tamar in, R. H., ed. Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 567-620.
Tamar in, R. H. (1977a) Dispersal in island and mainla nd v oles. Ecology 58: 1044-1054.
Tamar in, R. H. (1977b) Demography of th e beach vole (Microtus brewer i) and the meadow vole ( M. penns ylvanicus ) in southern Massach usetts. Ecology 58: 1310-1321.
Tamar in, R. H. (1984) Body mass as a cr iter ion of dis persal in v oles: a cr it ique . J. Mammal . 65: 691-692.
Tamar in, R. H., ed. (1985) Biology of new world Microtus. Spec. Publ. Amer . Soc. Mammal . 8.
Townsend, M. T. (1935) Studies on some of th e smal l animals of cen tral New York. Roosevel t Wildl. Ann. 4: 1-20.
Van Vleck, D. B. (1969) Standardization of Microtus hom e range cal cula tion. J. Mammal . 50: 69-30.
Wiegert, R. (1961) Respiratory energy loss an d acti vi ty p atter ns in the meadow vo le, Microtus penns ylvanicus penns ylvanicus . Ecology 42: 245-253.
Wolff, J. O. (1985) Behavior. In: Tamar in, R. H., ed. Biology of New World Microtus. Spec. Publ. Am. Soc. Mammal . 8; pp. 340-372.
Wunde r, B. A. (1985) Energetics an d thermoregula tion. In: Tamar in, R. H., ed. Biology of new wo rld Microtus. Spec. Publ. Amer . Soc. Mammal . 8; pp. 812-844.
Zimmer man, E. G. (1965) A comparison of habitat an d food of two species of Microtus. J. Mammal . 46: 605-612.
2-336 Meadow Vole

2.2.10. Muskrat (w ater rats and m uskrats)
Order Rodent ia Family Murid ae. Water rats an d muskr ats ar e the most aq uatic of th is fami ly of rodents, wi th most o f th eir lives sp ent in or near bogs , marshes, lakes o r str eams. These two rodents feed mostl y on aquatic veg etat ion. Only one sp ecies exists i n each genus (Burt and Grossen heider, 1980).
Selected species
The muskr at (Ondatra zibethicus) is indige nous and c ommon t hroughout mo st of the United States (excep t in the extr eme so utheast, central Texas, and most o f California) and Canada (excep t in the extr eme north) (Burt and Grossen heider, 1980). Muskr ats feed primar ily on aquatic plants. They are prey fo r hawks, minks, otter s, raccoons , owls , red fox, dogs , snapping t urtles, and water snakes (B ednarik, 1956; Erringt on, 1939a; Wilson, 1985), and are more vulne rable to predation dur ing t imes o f drought wh en low water levels leave th eir dens or lodge s more expos ed (Erringt on, 1939a). Many ver tebrates u se muskr at homes fo r shelter or to find f ood ( Kiv iat, 1978). The muskr at is one of th e most valua ble fu r anim als in Nor th Amer ica (Dozier, 1953; Perry, 1982). Including t he Newfoundla nd m uskr at, fo rmer ly Ondatra obscu rus, 16 recognize d subs pecies of O. zibethicus exist i n North Amer ica (Per ry, 1982). Of th ese, O. z. zibethicus (easter n Unit ed States, southeaster n Canada), O. z. osoyoosensis (Rocky Mount ains , sout hwester n Canada), and O. z. riv alic ius (sout hern Louis iana, coasts o f Missi ss ippi, wester n Alabama, and easter n Texas) ar e most o ften studie d.
Body size. The muskr at measu res 25 to 36 cm (h ead and body) wi th a 20- to 25- cm tai l (Burt and Grossen heider, 1980), and adult weight s can range fr om 0.5 kg to over 2 kg (see Appendix ). Willne r et al. (1980) reported no sexu al dimorphism, whereas Dozier (1950), Parker and Maxwel l (1984), and others (see Appendix) reported that mal es are slight ly h eavier than femal es. Muskr ats ten d to be larger and heavier in northern lati tudes (Perry, 1982), although t he smal lest mu skr ats ar e found in Ida ho (Reeves an d Willi ams, 1956). Fat levels in adult mal es inc rease fr om sp ring t hrough f all, and s ubs equent ly decrease fr om wi nter to spring (Schacher and Pelton, 1975). In nonpr egnant femal es, fat levels decrease fr om wi nter through s ummer ; in pregnant femal es, body fat i ncreases fr om spring t o summer (Schacher and Pelton, 1975).
Habitat . Muskr ats inha bit sal twater and bracki sh marshes and freshwater creeks, str eams, lakes, marshes, and ponds (Dozie r, 1953; Johns on, 1925; Kiviat, 1978; O'Neil, 1949). Muskr ats th at l ive along t he banks or shor es of water ways g enerally excavate d ens in the banks, whereas mu skr ats l iving in ponds wi th ample plant mater ial constr uct lodge s (Johns on, 1925; Perry, 1982). When avai lable, bank dens seem p refer red over constr ucted lodge s (Johns on, 1925).
Food ha bi ts . Muskr ats ar e primar ily herbivorous, but so me popula tions ar e more omniv orous (Dozie r, 1953; Erringt on, 1939b). Muskr ats u sually feed at night , div ing t o gnaw on aquatic veg etat ion gr owing ne ar th eir hou ses (Dozier, 1953; Johns on, 1925; Perry, 1982). The roots an d basal portions of aqua tic pla nts make u p most o f th e muskr at's diet, although s hoot s, bulbs , tube rs, stems, and leaves al so are eaten (Dozier,
2-337 Muskr at

1950, 1953; Willne r et al., 1980; Svihla and S vihla , 1931). Marsh grasses an d sedges (Svihla and S vihla , 1931) and cattai ls (Johns on, 1925; Willne r et al., 1975) seem to be impor tant muskr at foods ; in Maryland, green algae is al so impor tant (Willne r et al., 1975). Although m uskr ats fo rage near their dens or lodge s, th ey show preferences fo r some plant species (e.g., cattai ls, bulr ushes) over others (Bellrose, 1950). Muskr ats ar e a major consumer of mar sh grasses (K iviat, 1978). They also dig f or food on l ake and pond botto ms (B ailey, 1937; Dozier, 1953; Hanson et al ., 1989). Among t he anim als that muskr ats co nsume are crayfi sh, fish, frogs, turtles, and young bir ds (Er ringt on, 1939b; Johns on, 1925; Willne r et al., 1980). Mollu scs ar e an important co mpone nt of th e die t of some popula tions (Con vey et al ., 1989; Neves an d Odom, 1989; Parmalee, 1989; Willne r et al., 1980). Young m uskr ats feed more on bank veg etat ion t han do adult s (Warwick, 1940, ci ted in Perry, 1982).
Temperature regulation and molt. Acti ve year -round ( Kiv iat, 1978), muskr ats usually begin t heir annua l molt in t he summer , wi th fur reaching it s minim um density dur ing A ugus t (Willne r et al., 1980). Muskr ats u se th eir dens or lodge s to ins ulate themsel ves fr om su mmer heat and winter cold ( O'Neil, 1949; Willne r et al., 1980). During extr eme cold, muskr ats may fr eeze to death if th ey are unable to plug t heir den entrances (Erringt on, 1939a).
Breeding acti vi ties and social organization. Muskr ats ar e solit ary or form br eeding pairs that remain in a hom e range exclus ive of other pairs (Erringt on, 1963; Proulx an d Gilbe rt, 1983). They are ter ritorial, particularly during p eak reproducti ve acti vi ty, wi th their houses u sually sp aced at least 8 m ap art (Johns on, 1925; Sather, 1958; Trippe nsee, 1953). In southern parts o f th eir range, muskr ats b reed throughout th e year , wi th late fal l and early sp ring p eaks (O' Neil, 1949; Svihla and S vihla , 1931; Wilson, 1955). In northern lati tude s, breeding oc curs only in t he spring a nd s ummer , wi th fi rst l itter s born in late April or early May (Mathiak, 1966; Beer, 1950; Erringt on, 1937b; Gashwi ler, 1950). Erringt on (1937b) found t hat pos tpartum estr us occu rs in the muskr at, and suggested that th e period between litter s is about 30 d. Neonates ar e almost h airless b ut by age 2 wk ar e covered wi th fur and a ble to swi m (Erringt on, 1963).
Home range and r esour ces. Muskr ats h ave relatively smal l home ranges th at var y in c onf igur ation de pending on t he aquatic habitat (Per ry, 1982; Willne r et al., 1980). They build t wo di ffer ent typ es of houses: a main dwelling a nd a feeding hous e (feeder) that is smalle r th an the main hous e (Dozier, 1953; Johns on, 1925; Sather, 1958). The feeder provides protect ion f rom th e elements an d predators wh en feeding in pr ime foraging a reas, as wel l as access to oxyg en during f rozen condit ions . The hous e prov ides a dry nest an d stab le temp eratures. Muskr ats u sually fo rage wi thin 5 to 10 m of a hous e (Willne r et al., 1980). Using r adiot elemetr y, MacArthur (1978) found m uskr ats wi thin 1 5 m of th eir pr imary dwelling 5 0 percent of th e time and only r arely mo re than 150 m. Mathiak (1966) reported other exp eriments show ing t hat muskr ats r emain close to their dwellings .
In the winter , muskr ats build pus hups , whic h are cavi ties fo rmed in 30 to 46 cm high piles of veg etat ion pus hed up t hrough hole s in t he ice of a marsh (Perry, 1982). Muskr ats use pus hups as resting pl aces d uring f rozen condit ions to minim ize th eir expos ure to cold water (Fulle r, 1951). In the summer , muskr ats o ften change the use of th eir home range in response to water levels; during dr ought s they will m ove if th e area
2-338 Muskr at

�
around t he hous e dr ies up, whic h can lead to int ense aggr ession in t he more favo rable habitat (Er ringt on, 1939a). Usually only a minor propor tion of drough t-evi cted muskr ats can find ne w ho mes (Er ringt on, 1939a). In the winter , drought s can result in sever e mortalit y (Erringt on, 1937a).
Popul ation de nsity . Bellrose and Brown (1941, ci ted in Perry, 1982) conclude d that cattai l communit ies suppor t more muskr at hou ses th an other plant typ es in the Illinois River val ley. Cattai l communit ies also suppor t high de nsit ies of mu skr ats i n other areas (Erringt on, 1963; Dozier, 1950). In pond a nd l ake habitats, shoreline lengt h is a more important facto r than overall habitat ar ea in determining m uskr at dens ity (Glass, 1952, ci ted in Perry, 1982). Many investi gators esti mate mu skr at densi ties by count ing t he number of houses o r push-ups and multiply ing by a facto r ranging f rom 2.8 (Lay, 1945, ci ted in Bout in and Bir kenholz, 1987) to 5.0 (Dozier et al., 1948), although t his method is questiona ble (Bout in and Bir kenholz, 1987).
Popul ation dy namics. The age at fi rst br eeding v aries but usually occurs dur ing t he fi rst spring after birth (Erringt on, 1963; Perry, 1982). Southern popula tions produc e more litter s but wi th fewer pups in each than do northern popula tions (Bo yce, 1977; Perry, 1982; see tab le). Muskr ats i n lower quality h abitats h ave both smal ler litter sizes and fewer litter s than muskr ats i n better quality ar eas (Neal, 1968). They disperse in the spring t o establis h breeding t erritories or to move int o uninha bit ed areas (Er ringt on, 1963). Muskr at popula tion cycl es of 5, 6, and 10 y have been reported (Butler, 1962; Willne r et al., 1980); Perry (1982) su mmar ized sever al studies th at reported cycl es ranging f rom 10 to 14 yr or more. Butler (1962) found t hat muskr ats follow a 10-yr cycl e in most p arts o f Canada.
Simi lar species (fr om general references)
� The Flor ida water rat (Neofib er alleni) is mu ch smal ler (20 to 22 cm) than the muskr at, wi th a rounde d tail (11 to 17 cm) to dist inguis h it fu rther. The Flor ida water rat inha bit s bogs , marshes, weedy lake borders, and savan na str eams, though it s range is lim ited to Flor ida. It feeds on aquatic plants an d crayfi sh.
General references
Bout in and Bir kenholz (1987); Burt and Grossen heider (1980); Perry (1982); Willne r et al . (1980).
2-339 Muskr at

Muskrat ( Ondatra zibethicus )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight B M winter 1,480 1,400 - 1,520 New York Dozier, 1950 (g) B F winter 1,350 1,300 - 1,400
B M winter 1,326 ± 45.9 SE e Tennessee Schacher & Pel ton, 1978 B F w inter 1,221 ± 54.2 SE
B M winter 1,180 730 - 1,550 Nebraska, nc Kansas Sather, 1958 B F w inter 1,090 770 - 1,450
A M spr ing 909 Idaho Reeves & W illia ms, 1956 A F spr ing 837
neonat e 21.3 16 - 28 Iowa Erringt on, 1939b neonat e 20 - 25 New York Dean, 1957
at weaning 200 112 - 184 Iowa Erringt on, 1939b at weaning New Brunswick, Canada Parker & Maxwell, 1984
Pup Growth 0 to 30 d 5.4 4.3 - 5.6 Iowa/marsh Erringt on, 1939b Rate (g/d) weaning to 1st
fall; M 7.5 New Brunswick, Canada/ Parker & Maxwell, 1980 F 7.1 marsh
Metabol ic float ing 21 ± 7.9 SE lab (water temperature 25 C) Fish, 1982 Rate swimming 38 (lO /kg-d)2
o
Metabol ic Rate float ing 101 lab (water temperature 25 C) Fish, 1982 (kcal /kg-d) swimming 182
A M basal 71.6 est imated 1 A F basal A M free-living 213 (90 - 505) est imated 2 A F f ree-living 216 (91 - 513)
o
2-340 M
uskrat

Muskrat ( Ondatra zibethicus )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Food greens 0.34 Loui siana, capt ive Svihla & Svi hla, 1931 3 Ingest ion greens & cor n 0.26 (riv alic ius) Rate (g/g-d)
Water A M 0.97 est imated 4 Ingestion A F 0.98 Rate (g/g-d)
Inhal ation A M 0.61 est imated 5 Rate (m /d) A F 0.573
Surface Ar ea A M 1,221 est imated 6 (cm ) A F 1,1592
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
catta il 25 - 50 ne United States/NS Martin et al., 1951 bul rush 10 - 25 bur reed 5 - 10 (rough appr oxi mation of % waters tarwort 2 - diet; stomach contents ) pondweed 2 - 5 arrowhead 2 - 5 cor n 2 - 5
cat tail 59 Somerset Co., MD/bracki sh Willn er et al., 1975 rush 17 marsh mille t 8 algae 5 (% of diet; stomach grass 4 cont ents) cor d grass 4 seeds 2 other 3
5
2-341 M
uskrat
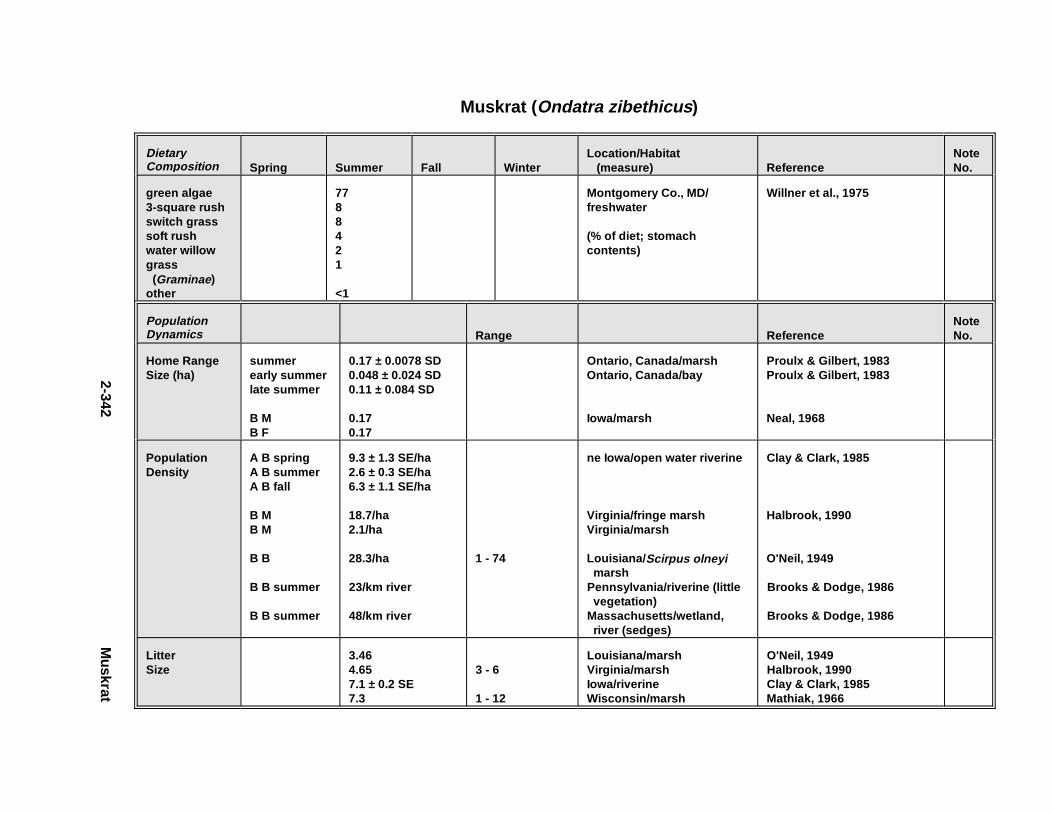
Muskrat ( Ondatra zibethicus )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
green algae 77 Montgomery Co., MD/ Willn er et al., 1975 3-squar e rush 8 freshw ater switch gr ass 8 sof t rush 4 (% of diet; stomach water w illo w 2 contents ) grass 1 (Graminae)
other <1
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Home Range sum mer 0.17 ± 0.0078 SD Ontario, Canada/marsh Proul x & Gilbert, 1983 Size (ha) early sum mer 0.048 ± 0.024 SD Ontario, Canada/bay Proul x & Gilbert, 1983
late sum mer 0.11 ± 0.084 SD
B M 0.17 Iowa/marsh Neal, 1968 B F 0.17
Popul ation A B spr ing 9.3 ± 1.3 SE/ha ne Iowa/open wat er r iver ine Clay & Clark, 1985 Density A B s ummer 2.6 ± 0.3 SE/ha
A B f all 6.3 ± 1.1 SE/ha
B M 18.7/ha Virginia/fringe m arsh Halbrook, 1990 B M 2.1/ha Virg inia/marsh
B B 28.3/ha 1 - 74 Loui siana/Scirpus ol neyi O'Neil, 1949
B B sum mer 23/km river Pennsy lvani a/river ine (little Brooks & Dodge, 1986
B B sum mer 48/km river Massachuset ts/wetland, Brooks & Dodge, 1986
marsh
veget ation)
river (sedges)
Li tter 3.46 Loui siana/marsh O'Neil, 1949 Size 4.65 3 - 6 Virginia/marsh Halbrook, 1990
7.1 ± 0.2 SE Iowa/river ine Clay & Clark, 1985 7.3 1 - 12 Wisconsi n/marsh Mathiak, 1966
2-342 M
uskrat
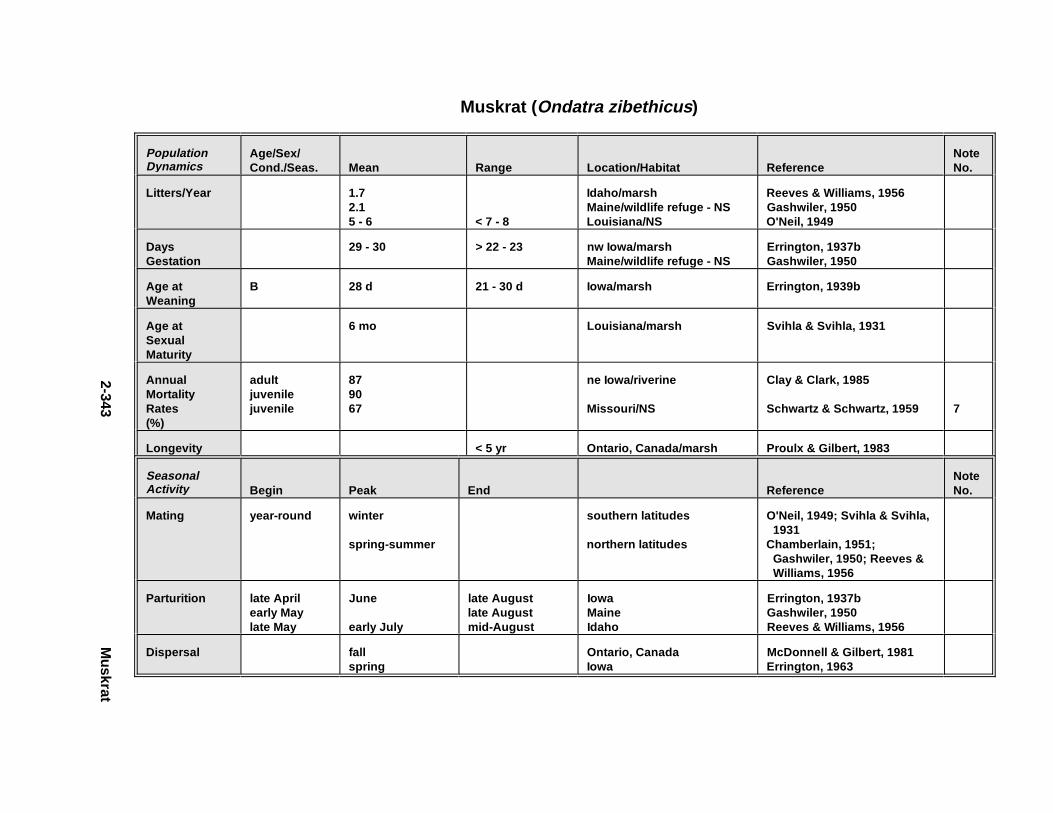
Muskrat ( Ondatra zibethicus )
Population Dynamics
Age/Sex/ Note Cond. /Seas. Mean Range Locat ion/Habi tat Reference No.
Li tters/Year 1.7 Idaho/ marsh Reeves & W illia ms, 1956 2.1 Maine/wildlife refuge - NS Gashw iler, 1950 5 - 6 < 7 - 8 Loui siana/NS O'Neil, 1949
Days 29 - 30 > 22 - 23 nw Iowa/marsh Erringt on, 1937b Gestation Maine/wildlife refuge - NS Gashw iler, 1950
Age at B 28 d 21 - 30 d Iowa/marsh Erringt on, 1939b Weaning
Age at 6 mo Loui siana/marsh Svihla & Svi hla, 1931 Sexual Maturity
Annual adul t 87 ne Iowa/river ine Clay & Clark, 1985 Morta lity juvenile 90 Rates juveni le 67 Missour i/NS Schw artz & Schw artz, 1959 7 (%)
Longevi ty < 5 yr Ontario, Canada/marsh Proul x & Gi lbert, 1983
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating year-round winter sout hern latitudes O'Neil, 1949; Svihla & Svi hla,
spr ing-sum mer nor thern latitudes Chamberlain, 1951; 1931
Gashw iler, 1950; Reeves & Willia ms, 1956
Partur ition late Apr il June late August Iowa Erringt on, 1937b early May late August Maine Gashwi ler, 1950 late May early Jul y mid-August Idaho Reeves & Willia ms, 1956
Disper sal fall Ontario, Canada McDonnel l & Gi lbert, 1981 spr ing Iowa Erringt on, 1963
2-343 M
uskrat

1234567
Muskrat ( Ondatra zibethicus )
Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom Sather (1958).Est imated usi ng equat ion 3-46 (Nagy, 1987) and body weight s f rom Sather (1958).Based on wet weight of food; greens i ncluded Panicum hemitomum, P. virgatum, and Spartina patens .Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Sather (1958).Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Sather (1958).Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Sather (1958).Cited in Perry (1982.)
2-344 M
uskrat

References (including A ppendix)
Arthur, S. C. (1931) The fur animals of Louis iana. Louis . Dept . Cons erv. Bull. 18 (revised).
Asdell, S. A. (1964) Patter ns of mammal ian reproduc tion. Compstock Publis hing Co.
Bailey, V. (1937) The Maryland muskr at mar shes. J. Mammal . 18: 350-354.
Bednarik, K. (1956) The muskr at in Ohio L ake Er ie marshes. Colum bus , OH: Ohio Div is ion of Wildlif e Departmen t of Natural Resources; 67 pp.
Beer, J. R. (1950) The reproducti ve cycl e of th e muskr at in Wisco nsin. J. Wildl. Manage. 14: 151-156.
Bellrose, F. C. (1950) The relations hip of mu skr at popula tions to var ious mar sh and aquatic plants. J. Wildl. Manage. 14: 299-315.
Bellrose, F. C.; Brown, L. G. (1941) The effect o f fl uctu ating water levels on the muskr at popula tion of th e Illinois River valle y. J. Wildl. Manage. 5: 206-212.
Beshears, W. W., Jr . (1951) Muskr ats i n relation t o farm ponds . Proc. Annu. Conf . Southeast. Asso c. Game and Fish Comm. 5: 1-8.
Beshears, W. W.; Haugen, A. O. (1953) Muskr ats i n farm ponds . J. Wildl. Manage. 17: 540-456.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Bout in, S.; Bir kenholz, D. E. (1987) Muskr at and round-tai led muskr at. In: Novak, M.; Baker , J. A.; Obbarel, M. E.; et al., eds. Wild f urbearer man agement and conservat ion; pp. 316-324.
Boyce, M. S. (1977) Life histo ries in var iable environments: appli cations to geogr aphic var iation in t he muskr at (Ondatra zibethicus) [Ph.D. disser tat ion] . New Haven, CT: Yale Universi ty.
Brook s, R. P.; Dodge , W. E. (1986) Esti mat ion of habi tat q uality an d summer popula tion densi ty fo r muskr ats o n a water shed basis. J. Wildl. Manage. 50: 269-273.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Butler, L. (1940) A quanti tati ve stu dy of mu skr at food. Can. Field-Nat. 54: 37-40.
Butler, L. (1962) Periodic it ies in t he annua l muskr at popula tion f igur es for th e prov inc e of Saskatch ewan. Can. J. Zool. 40: 1277-1286.
2-345 Muskr at

Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Chamberlain, J. L. (1951) The life histo ry and management of th e muskr at on Great Meadows Refuge [master 's th esis]. Amherst, MA: Universi ty o f Massach usetts.
Clay, R. T.; Clark, W. R. (1985) Demography of mu skr ats i n the uppe r Mississippi Riv er. J. Wildl. Manage. 49: 883-890.
Convey, L. E.; Hanson, J. M.; MacKay, W. C. (1989) Size-selecti ve predation on unionid clams b y mu skr ats. J. Wildl. Manage. 53: 654-657.
Dean, F. C. (1957) Age cr iter ia and ki t growth of cen tral New York mu skr ats [Ph .D. disser tat ion] . NY: State Univ ersi ty of New York, Colle ge of Forestr y.
Dibble e, R. L. (1971) Reproduc tion, produc tivi ty and f ood ha bit s of mu skr ats o n Prince Edward Island [master 's th esis]. Wolfv ille , Nova Scotia, Canada: Acadia Univ ersi ty.
Dilw or th, T. G. (1966) The life histo ry and ecology of th e muskr at, Ondatra zibethicus zibethicus, unde r sever e water level fluctu ations [master 's th esis]. Fredericto n, New Brunswi ck, Canada: Universi ty o f New Brunswi ck.
Donohoe , R. W. (1961) Muskr at produc tion in a reas of co ntrolle d and unc ont rolle d water -level units [Ph .D. disser tat ion] . Colum bus , OH: Ohio State Universi ty.
Dorney, R. S.; Rusch , A. J. (1953) Muskr at growth and litter produc tion. Wisc. Cons erv. Dept . Tech. Wildl. Bull. 8; 32 pp.
Dozier, H. L. (1948) Esti mat ing m uskr at popula tions by hous e count s. Trans. Nor th Amer . Wildl. Conf . 13: 372-392.
Dozier, H. L. (1950) Muskr at tr apping on t he Mont ezuma Nationa l Wildlif e Refuge , New York, 1943-1948. J. Wildl. Manage. 14: 403-412.
Dozier, H. L. (1953) Muskr at produc tion a nd m anagement. U.S. Fish Wildl. Serv. Circ. 18; 42 pp.
Dozier, H. L.; Markley, M. H.; Llewel lyn, L. M. (1948) Muskr at investi gations on t he Blackwater Nationa l Wildif e Refuge , Maryland, 1941-1945. J. Wildl. Manage. 12: 177-190.
Erickso n, H. R. (1963) Reproduc tion, growth, and m ovemen t of mu skr ats inha bit ing smal l water areas in New York state. N.Y. Fish Game J. 10: 90-117.
Erringt on, P. L. (1937a) Habitat r equir ements o f str eam-dwelling m uskr ats. Trans. North Amer . Wildl. Conf . 2: 411-416.
2-346 Muskr at

Erringt on, P. L. (1937b) The breeding seaso n of th e muskr at in northwest I owa. J. Mammal . 18: 333-337.
Erringt on, P. L. (1939a) React ions of mu skr at popula tions to drought . Ecology 20: 168-186.
Erringt on, P. L. (1939b) Observat ions on y oung m uskr ats i n Iowa. J. Mammal . 20: 465-478.
Erringt on, P. L. (1948) Environmental control for increasing m uskr at pr oduc tion. Trans. Nor th Amer . Wildl. Nat. Resour . Conf . 13: 596-609.
Erringt on, P. L. (1963) Muskr at popula tions . Ames, IA: Iowa State Universi ty Pr ess.
Fish, F. E. (1982) Aerobic en ergetics of su rface swi mming in t he muskr at Ondatra zibethicus. Physiol. Zool. 55: 180-189.
Fish, F. E. (1983) Metabolic effects o f swi mming v eloc ity and water temp erature in the muskr at (Ondatra zibethicus). Comp. Bioc hem. Physiol. A Comp. Physiol. 75: 397-400.
Fulle r, W. A. (1951) Measurements an d weights o f northern muskr ats. J. Mammal . 32: 360-362.
Gashwi ler, J. S. (1948) Maine muskr at investi gations . Maine Dept . Inland Fis h. Game Bull. ; 38 pp.
Gashwi ler, J. S. (1950) A stu dy of th e reproducti ve cap aci ty o f Maine muskr ats. J. Mammal . 31: 180-185.
Glass, B. P. (1952) Facto rs affect ing t he surviv al of th e pla ins mu skr at Ondatra zibethica cinnamomina in Oklahom a. J. Wildl. Manage. 16: 484-491.
Godin, A. J. (1977) Wild mammal s of New Engla nd. Balt imore, MD: Johns Hopk ins Universi ty Pr ess; 304 pp.
Halbrook, R. S. (1990) Muskr at popula tions in V irginia 's Elizabeth River: Inf luence of environmental contami nants [Ph .D. disser tat ion] . Blacksb urg, VA: Virginia Polytec hnic Insti tute and State Universi ty.
Hall, E. R. (1981) The mammal s of North Amer ica. 2nd ed. New York, NY: John W iley and Sons; 181 pp.
Hanson, J. M.; Mackay, W. C.; Prepas, E. E. (1989) Effect o f si ze-selecti ve predation by muskr ats (Ondatra zibethicus) on a popula tion of unionid c lams (Anodont a grandi s simpsoniana). J. Anim. Ecol. 58: 15-28.
Harris, V. T. (1952) Muskr ats o n tidal marshes of Dorchester Count y. Maryland Boa rd Nat. Resour., Chesapeake Bio. Lab., Dept . Res. Edu cation P ubl. 91; 36 pp.
2-347 Muskr at

Johns on, C. E. (1925) The muskr at in New York: its n atural histo ry and econom ics. Roosevel t Wildl. Bull. 3: 205-321.
Kiviat, E. (1978) The muskr at's role in the marsh ecosystem: a qualitati ve syn thesis (abstr act). Bull. Ecol. Soc. Am. 59: 124.
Lay, D. W. (1945) Muskr at investi gations in T exas. J. Wildl. Manage. 9: 56-76.
MacArthur, R. A. (1978) Winter movemen ts an d home range of th e muskr at. Can. Field-Nat. 92: 345-349.
MacArthur, R. A.; Krause, R. E. (1989) Energy requir ements o f fr eely diving m uskr ats (Ondatra zibethicus). Can. J. Zool. 67: 2194-2200.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York: McGraw-Hill.
Mathiak, H. A. (1966) Muskr at popula tion s tudie s at Hor icon Marsh. Tech. Bull. Wiscons in Conserv. Dept. 36: 1-56.
McDonne ll, J. A.; Gilbe rt, F. F. (1981) The responses o f mu skr ats (Ondatra zibethicus) to water level fluctu ations at Lut her Marsh, Ontar io. In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. Augus t 1980; Frostb urg, MD; pp. 1027-1040.
McLeod, J. A.; Bonda r, G. F. (1952) Studie s on t he biology of th e muskr at in Ma nit oba. Part I. Oestr us cycl e and breeding seaso n. Can. J. Zool. 30: 243-253.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Neal, T. J. (1968) A comparison of two muskr at popula tions . Iowa State J. Sci . 43: 193-210.
Neves, R. J.; Odom, M. C. (1989) Muskr at predation on e ndangered f reshwater mussel s in Virginia . J. Wildl. Manage. 53: 934-941.
O'Neil, T. (1949) The muskr at in the Louis iana co astal marshes (A stu dy of th e ecologic al, geologic al, biologic al, tida l, and c limati c facto rs governing t he produc tion a nd management of th e muskr at indus try in Louis iana). New Orleans, LA: Louis . Dept Wildl. Fish., Fed. Aid Sect. Fish and Game Div.; 152 pp.
O'Neil, T.; Linsco mbe, G. (1976) The fur animals, the alliga tor, and t he fur indus try in Louis iana. New Orleans, LA: Louis iana Wildl. and Fis h. Comm. Wildl. Education Bull. 106; 66 pp.
Olsen, P. F. (1959) Muskr at breeding biology at De lta, Manit oba. J. Wildl. Manage. 23: 40-53.
2-348 Muskr at

Parker , G. R.; Maxwel l, J. W. (1980) Character isti cs o f a popula tion of mu skr ats (Ondatra zibethicus zibethicus) in New Brunswi ck. Can. Field-Nat. 94: 1-8.
Parker , G. R.; Maxwel l, J. W. (1984) An evaluation of sp ring a nd autumn trapping seaso ns for muskr ats, Ondatra zibethicus, in easter n Canada. Can. Field-Nat. 98: 293-304.
Parmalee, P. W. (1989) Muskr at predation on s oftsh ell turtles. J. Tenn. Acad. Sci . 64: 225-227.
Perry, H. R., Jr . (1982) Muskr ats. In: Chapman, J. A.; Feldha mer , G. A., eds. Wild mammal s of Nor th Amer ica: biology , management and econom ics. Balt imore, MD: Johns Hopk ins Univ ersi ty Press; pp. 282-325.
Proulx , G.; Gilbe rt, F. F. (1983) The ecology of th e muskr at Ondatra zibethicus at Lut her Marsh, Ontar io. Can. Field-Nat. 97: 377-390.
Reeves, H. M.; Willi ams, R. M. (1956) Reproduc tion, size, and m ortalit y in Ro cky Mount ain muskr at. J. Mammal . 37: 494-500.
Sather, J. H. (1958) Biology of th e Great Plains mu skr at in Nebraska. Wildl. Monogr . 2. 35 pp.
Schacher, W. H.; Pelton, M. R. (1975) Producti vi ty o f mu skr ats i n east T ennessee. Proc. Annu. Conf . Sout heast. Asso c. Game and Fish Comm. 29: 594-608.
Schacher, W. H.; Pelton, M. R. (1978) Sex ratios, morphology and c ondit ion pa rameter s of muskr ats i n east T ennessee. Proc. Annu. Conf . Sout heast. Asso c. Fish and Wildl. Agencies 30: 660-666.
Schwar tz, C. W.; Schwar tz, E. R. (1959) The wild mammal s of Misso uri. Colum bia, MO: Universi ty o f Misso uri Press an d Misso uri Cons ervation Com mission.
Seamans, R. (1941) Lake Champlain fur survey. Vermont Fish Game Bull. 3-4.
Smith, F. R. (1938) Muskr at investi gations in Dor chester Count y, Maryland, 1930-34. U.S. Dept. Agr. Circ. 474; 24 pp.
Smith, H. R.; Jordan, P. A. (1976) An exploit ed popula tion of mu skr ats wi th unus ual biomass, produc tivi ty, and body size . Conn. Dept . Environ. Prot . Rep. Invest. No. 7; 16 pp.
Smith, H. R.; Sloan, R. J.; Walton, G. S. (1981) Some man agement implic ations b etween harvest r ate and popula tion r esilie ncy of th e muskr at (Ondatra zibethicus). In: Chapman, J. A.; Pursley, D., eds. Proceedings wo rldw ide fu rbearer conf erence: v. 1. August 1980; Frostb urg, MD; pp. 425-442.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
2-349 Muskr at

Steven s, W. E. (1953) The northwester n muskr at of th e Mackenzie Delta, Nor thwest Territories, 1947-48. Can. Wildl. Serv., Wildl. Manage. Bull. , Ser. 8; 55 pp.
Stewar t, R. W.; Bide r, J. R. (1974) Reproduc tion a nd s urviv al of dit ch-dwelling m uskr ats i n southern Quebec. Can. Field-Nat. 88: 420-436.
Svihla , A. (1931) The field biologis t's repor t. In: Arthur , S. C., compil. The fur anim als of Louis iana. New Orleans, LA: Louis iana Dept . Cons erv., Bull. 18 (rev.); 439 pp.
Svihla , A.; Svihla , R. D. (1931) The Louis iana mu skr at. J. Mammal . 12: 12-28.
Trippe nsee, R. E. (1953) Muskr ats. In: Wildlif e management: fur bearers, water fowl, and fish: v. 2. New York, NY: McGraw-Hill; pp. 126-139.
Walker , E. P.; et al . (1975) In: Paradiso, J. L., ed. Mammal s of th e world: v. 2. 3rd ed. Balt imore, MD: Johns Hopk ins Univ ersi ty Press; pp. 645-1500.
Warwick, T. (1940) A contr ibut ion t o the ecology of th e muskr at (Ondatra zibethica) in the Br it ish Is les. Proc. Zool. Soc. London, Ser. A 110: 165-201.
Willne r, G. R.; Chapman, J. A.; Golds berry, J. R. (1975) A stu dy and review of mu skr at fo od habits wi th special reference to Maryland. Maryland Wildl. Adm. Publ. Wildl. Ecol. 1; 25 pp.
Willne r, G. R.; Feldh amer , G. A.; Zucker , E. E.; et al . (1980) Ondatra zibethicus. Mammal ian species. No. 141. Amer . Soc. Mammal .; 8 pp.
Wilson, K. A. (1954) Litter produc tion of co astal North Carolina mu skr ats. Proc. Annu. Conf . Southeast. Asso c. Game and Fish Comm. 8: 13-19.
Wilson, K. A. (1955) A compendium of th e pr inc ipa l data on muskr at reproduc tion. Raleigh, NC: Nor th Carolina W ildl. Resour . Comm., Game Div.; Fed. Aid Wildl. Resto ration Proj. W-6-R.
Wilson, K. A. (1956) Color , sex r atios, and weight s of Nor th Carolina mu skr ats. Raleigh, NC: Nor th Carolina W ildl. Resour . Comm.; Fed. Aid in W ildl. Resto ration P roj. W-6-R-15; 20 pp.
Wilson, K. A. (1985) The role of mi nk and otter as mu skr at predators in northeaster n North Carolina . Proc. Annu. Conf . Sout heast. Asso c. Fish and Wildl. Agencies 18: 199-207.
2-350 Muskr at

2.2.11. Eastern Cottontail (rabbits)
Order Lagom orpha Family Lepor idae. Rabbit s and ha res are medium -sized gr azing herbiv ores found t hroughout Nor th Amer ica. Most species are noc turnal and c repus cula r. Many ar e social, travelling in small gr oups . Rabbit s are prey for large carniv orous bir ds and mammal s. Most sp ecies also are important game animals.
Selected species
The easter n cotto ntai l (Sylvila gus florid anus) is th e most wi dely distr ibuted of th e medium-si zed rabbits (Chapman et al ., 1982). It is found ov er mo st of th e easter n half of the United States an d southern Canada and has been widely introduced into the wester n United States (C hapman et al ., 1980). North of Mexico, 14 subspecies are recognize d (Chapman et al ., 1982). The easter n cotto ntai l feeds on green vegetat ion in s ummer and bark and twi gs in winter . The cotto ntai l is acti ve fr om ear ly evening t o late mo rning a nd is preyed on by owls, hawks, and carnivorous mammal s (Palmer and Fowler, 1975; Burt and Grossen heider, 1980).
Body size. The easter n cotto ntai l measu res 35 to 43 cm i n length and weighs 0. 7 to 1.8 kg (Lord, 1963; see tab le) wi th femal es s light ly larger th an the males (Nowak an d Paradiso, 1983; see tab le). Cotto ntai l body weight var ies seaso nally, increasing dur ing spring a nd s ummer and declining dur ing w int er in s ome areas; differ ent patter ns occu r in other areas (Chapman et al ., 1982; Pelton and Jenkins, 1970).
Habitat . The easter n cotto ntai l is unique to the genus b ecause of th e large var iety of habitats th at i t occupie s, inc luding gla des and w oodla nds , deser ts, swamp s, prairies, hardwood f orests, rain forests, and boreal forests (N owak an d Paradiso, 1983). Open grassy ar eas generally are used for foraging a t night , whereas dense, heavy co ver typ ical ly is used for sh elter dur ing t he day (Chapman et al., 1982). During w int er, cottont ails r ely more on w oody veg etation f or adequate co ver (Allen, 1984).
Food ha bi ts . Dur ing t he growing seaso n, cotto ntai ls eat h erbaceous plants (e.g., grasses, clover , timoth, alfal fa). During t he wint er in areas wh ere herbaceous plants ar e not availa ble , they cons ume woody vine s, sh rubs , and t rees (e.g., birch, maple, apple ) (Chapman et al ., 1982). In Ohio, blue grass an d other grasses mad e up a large portion of the easter n cotto ntai l's d iet, excep t during s now co ver (Cha pman et al., 1982). During t he wint er in Conn ecti cut, the principle die t of easter n and New Engla nd c ottont ails cons ists o f bark and t wigs , shrubs and v ines, berries, and w illow (D alke and Sime, 1941). In agricultural areas, corn, soybeans, wheat, and other crops may co mprise a large portion of their diet (Chapman et al ., 1982). Younge r rabbit s pr efer the more succu lent weed y fo rbs that cont ain m ore dige stible en ergy and pr otein (Chapman et al., 1982). Copr opha gy (inge stion of feces) h as been reported in S. floridanus (Kirkpatr ick, 1956).
Temperature regulation and molt. Easter n cotto ntai ls do not unde rgo hibe rnation or torpor; they are acti ve al l year , showing p eaks o f daily acti vi ty at d awn and dusk (Chapman et al ., 1980). Adult s molt gradually over about 9 mo of th e year , wi th two peak mo lting periods (Spinne r, 1940). In Connecti cut, the spring p eak occu rs in May and
2-351 Easter n Cotto ntai l

June and the fal l peak occu rs in Septemb er and Octo ber (Spinne r, 1940). In Texas, spring and fal l molts p eak in April and Octo ber, respecti vely (Bothma and Teer, 1982).
Breeding acti vi ties and social organization. Breeding acti vi ty b egins l ater at highe r elevat ions and a t highe r lati tude s (Conaway et al ., 1974), by Jan uary in Alabama and by late March in southern Wisco nsin (Chapman et al ., 1980). Several studies have sh own that cont inue d harsh w int er weath er may d elay th e onset o f th e breeding seaso n (Hamilton, 1940; Conaway an d Wight , 1962; Wight and Con away, 1961). Breeding seaso ns are longe r in the southern states (L ord, 1960). The onset o f breeding v aries between differ ent popula tions and w ithin t he same popula tion f rom year to year (Chapman et al ., 1980). Males may f ight to establis h dom inance hierarchie s for access to femal es (Chapman and Ceballos , 1990; Nowak an d Paradiso, 1983). Lagomorphs in general are induc ed ovulators, and cotto ntai ls in particular demonstr ate a syn chronize d br eeding seaso n, wi th conception immed iately after the birth of a l itter (Chapman et al ., 1982).
Home range and r esour ces. Cot tont ails ar e found in a var iety o f habitats th at contain weedy fo rbs and perennia l gr asses; they prefer th ick, short, woody perennia ls th at provide escap e si tes (Chapman and Ceballos , 1990). Cotto ntai ls usually do not defend ter ritories; the home ranges of differ ent age and sex g roups tend t o overlap, especially in fall a nd w int er wh en they look fo r areas offer ing a co mbina tion of food a nd c over (Chapman et al ., 1980, 1982). Home ranges are smal ler when thick veg etat ion pr ov ides abunda nt food a nd la rger in ha bitats wi th less f ood ( Chapman et al., 1982). Home ranges also are smalle r dur ing sever e winter weath er than at other times (Chapman et al ., 1982). Dur ing t he breeding seaso n, femal es build e labor ate nests wi thin s lanting hole s in t he ground w here they giv e bir th to their al tr ic ial (helpl ess) y oung. These burrows ar e vulne rable to flooding ( Chapman et al., 1982). The size of mal e home ranges during t he breeding seaso n can be more than double th at in w int er (Nowak an d Paradiso, 1983; Trent and Rong stad , 1974).
Popul ation de nsity . Popula tion de nsity depends on t he availa bilit y of r esour ces (e.g., food, cover ) in an area, and t ends to cycl e over a period of sever al years (Chapman and Ceballos , 1990). Usual densi ties range from 1 to 5 animals per hectar e, although value s as high a s 14 per hectar e have been reported (Chapman and Ceballos , 1990; Chapman et al ., 1982).
Popul ation dy namics. The easter n cotto ntai l exhibit s the high est fec undit y of th e genus; they often produc e 25 to 35 young pe r year (Chapman and Ceballos , 1990). Gestat ion l asts ap proximatel y 1 mo (Chapman et al ., 1982). Females may p roduce fi ve to seven litter s per year , and juvenile breeding ha s been repor ted (Chapman et al., 1982). The fi rst an d last l itter s of th e year are usually th e smal lest (Chapman et al ., 1977). Cotto ntai ls have mo re litter s wi th fewer young each in the southern states (L ord, 1960). Young l eave the nest wh en about ag e 14 to 16 d, although t hey may n ot be fully weaned unti l a few weeks l ater (Ecke, 1955). Female cotto ntai ls are capable of breeding by ag e 5 mo, and males as ear ly as 3 mo (Bothma and Teer, 1977). Adult mo rtalit y is high, from approximatel y 65 to 75 percent per year in some places (Eb erhardt et al ., 1963). Juvenile mortal ity i s even highe r, between 85 and 90 percent in the same ar eas (Eberhardt et al ., 1963).
2-352 Easter n Cotto ntai l

�
� �
�
�
�
�
�
�
�
Simi lar species (fr om general references)
� The mountain cotto ntai l (Sylvila gus nuttallii ) (Nuttal l's co tto ntai l) is smal ler (30 to 36 cm i n length and 0.7 to 1.3 kg) than the easter n cotto ntai l. The only cottont ail t hrough m ost of its range � the wester n United States � it l ives i n th ickets an d sagebrush, around loos e rocks, cl iffs, and mount ains . In the southwest, it l ives i n forests.
� The New Engla nd c ottont ail (Sylvi lagus transi tionalis) is si mila r in s ize to the easter n cotto ntai l and inha bit s br ushy ar eas, open forests, and mountain ter rain in Ne w Engla nd, extending dow n the Appalachia ns int o the sout hern United States. In recent year s, it has disappeared throughout mu ch of th e northeaster n United States, apparently because of co mpeti tion w ith S. flor idanus .
� The deser t co tto ntai l (Sylvi lagus auduboni i) (Audubon' s cottont ail) (30 to 38 cm i n length and 0.6 to 1.2 kg) is co mmo n in val leys i n the arid southwest, although it s range exten ds so uth to Mexico and north into the Rocky Mount ains . It inha bit s ope n pla ins , foot hills , and low vall eys an d also areas of grass, sagebrush, piny ons and junipe rs. It is mo st acti ve fr om late after noon t hroughout th e night .
� The brush rabbit (Sylvi lagus bachmani) (28 to 33 cm; 0.6 to 0.8 kg) is usually seen around t hic k cover and r arely u ses a b urrow. It feed s on green vegetat ion, inc luding la wns when in s ubur ban areas. The species is found along t he Paci fic co ast fr om th e Colum bia River in t he nor th to the tip of Baja Calif ornia in t he sout h.
� The marsh rabbit (Sylvila gus palustris ) is si mi lar in size to the easter n cotto ntai l and ranges fr om so utheaster n North Carolina to Flor ida. As the name implie s, it inha bit s swamp s and hummo cks, as wel l as wet botto mlands. Mostl y noctu rnal, it feed s on marsh vegetat ion, rhizo mes, and bulbs .
� The swamp rabbit (Sylvi lagus aquaticus) is si mi lar in size to the easter n cottont ail and is a good s wimmer found in swamp s, marshes, and wet botto mlands. It ranges primar ily in the south, from Texas eastwar d. It nests beneath logs or in t he bases o f stu mps, rarely using a bur row and may h arm crops near swamp s.
� The pygmy rabbit (Sylvi lagus idahoensis) is mar kedly smal ler (22 to 28 cm; 0.2 to 0.5 kg) than the easter n cotto ntai l, lacks a co nspicuous tail, and is cons idered by so me to be a dis tinc t genus ( Brachylagus). Its range is limi ted to sever al wester n states, where it inha bit s clum ps of tall s agebrush. It is mo stl y noctu rnal.
� The white-tai led jackr abbit (Lepus townsendi i), larger (46 to 56 cm; 2.2 to 4.5 kg) than the easter n cotto ntai l, is l imi ted to the northern United States
2-353 Easter n Cotto ntai l

�
�
west o f th e Great Lakes, into southern Canada. It inha bit s ope n, grassy, or sagebrush plains and may d amage hay cr ops and smal l trees.
� The black-tai led jackr abbit (Lepus cal ifor nicus) (43 to 53 cm; 1.3 to 3.1 kg) is the most co mmo n jackr abbit in t he grassl ands and open areas of th e wester n United States, where it inha bit s ope n pr airies and d eser ts wi th littl e vegetat ion. It is mo stl y noc turnal.
� The snow shoe hare (Lepus amer icanus ) (33 to 46 cm; 0.9 to 1.8 kg) inha bit s swamp s, forests, and thickets i n the northern United States an d Canada. Dur ing s ummer , it feed s on succu lent veg etat ion a nd dur ing w int er on t wigs , buds , and ba rk. Its hom e range is about 4 ha , but popula tions fluc tuate wide ly .
General references
Allen (1984); Burt and Grossen heider (1980); Chapman et al . (1980, 1982); Lord (1963); Nowak an d Paradiso (1983); and Palmer and Fowler (1975).
2-354 Easter n Cotto ntai l

Eastern Cottontail ( Sylvila gus floridanus )
2-355 E
astern C
otto
ntail
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight A M 1,134 ± 122 SD 801 - 1,411 w Maryland, West Virginia Chapm an & Morgan, 1973 (g) A F 1,244 ± 165 SD 842 - 1,533
A B inter 1,176 793 - 1,671 Georgia Pelton & Jenki ns, 1970 A B ing 1,286 898 - 1,630 all areas com bined A B mer 1,197 910 - 1,608 A B all 1,255 886 - 1,669
A B not breed. 1,229 ± 113 SD 1,093 - 1,461 mount ain Pelton & Jenki ns, 1970 A B not breed. 1,313 ± 141 SD 986 - 1,671 coast al A B not breed. 1,132 ± 136 SD 793 - 1,579 Piedmont
A B 1,231 ± 164 700 - 1,800 Illinoi s Lor d, 1963
neonate 42.2 36.0 - 49.0 Alabama Hill, 1972b
age: 10 d 58 Illinoi s Lor d, 1963 30 d 159 50 d 401
101 d 822 149 d 1,106
Georgia
Growth Rate day 0 - 30 3.2 Illinoi s Lor d, 1963 (g/d) day 11 - 30 3.7
day 31 - 50 8.8 day 51 - 100 11.3 day 101 - 150 6.4
Metabol ic Rate A B basal 71 est imated 1 (kcal /kg-d)
A B f ree-living 203 (77 - 535) est imated 2
Food Ingest ion 3 Rate (g/g-d)
Water A B 0.097 est imated 4 Ingest ion Rate (g/g-d)
wsprsumf

Eastern Cottontail ( Sylvila gus floridanus )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Inhal ation A B 0.63 est imated 5 Rate (m /d)3
Surface Ar ea A B 1,254 est imated 6 (cm )2
Dietary Compositio n Spr ing Summer Fall Winter Habi tat(measur e) Reference No.
Locat ion (subspeci es)/ Note
trees 13 2 7 39 Connect icut (malla rus)/ Dalke & Si me, 1941 shr ubs & vi nes 4 2 27 40 various (85% for malla rus herbs 44 23 34 5 subspeci es, grasses, sedges, (% frequence of occur rence; remainder for similar
rushes 26 56 30 6 obser vat ions of feeding on speci es S. transi tional is) crops 13 17 2 10 plants)
woody plants 17 23 20 100 Maryland/ forest Spencer & Chapm an, 1986 fo rbs 19 30 46 grasses 64 47 34 (% frequency of occur rence;
stomach cont ents)
bluegr ass 34 34 25 32 Ohio (mearnsi )/NS Dusi , 1952 orchar d gr ass 4 1 - 1 timothy grass 5 12 7 1 (% frequency of occur rence; Noddi ng wi ld rye 5 11 8 4 scat s) Canada gol denr od - - 3 -red cl over - - 6 - (in wi nter, woody tissues uni dent ified 52 42 51 62 predom inated in the
uni dent ified cat egor y)
2-356 E
astern C
otto
ntail

Eastern Cottontail ( Sylvila gus floridanus )
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/Habi tat Note Cond. /Seas. Mean Range Reference No.
Home Range A M winter 3.05 ± 0.72 SE Wisconsi n/woodl ot Dixon et al., 1981 Size (ha) A F w inter 2.99 ± 0.28 SE
A M winter 3.2 c Pennsy lvani a/mixed Al thof f and St orm, 1989 A M spr ing 7.2 A M sum mer 7.8 A M fall 3.1
A F w inter 2.1 c Pennsy lvani a/mixed Al thof f and St orm, 1989 A F spr ing 2.8 A F sum mer 2.4 A F fall 1.5
A M spr ing 2.8 sw Wisconsi n/woodl ot Trent & Rongst ad, 1974 A M early sum mer 4.0 late summer 1.5
A F spr ing 1.7 A F sum mer 0.8
Popul ation fall 1.1 ± 0.41 SD 0.41 to 2.08 c Michi gan/woods, Eberhardt et al., 1963 Densi ty marsh, fie lds (N/ha) fall 3.0 - 5.9 Illinoi s/old f ield Lor d & Cast eel, 1960
winter 0.67 - 1.5
sum mer 4.2 sw Wisconsi n/farm Trent & Rongst ad, 1974 fall 10.1 spr ing 3.7
Li tter 3.5 ± 0.042 SE Alabama/across si x habi tats Hill, 1972c Size 5.3 Illinoi s/NS Lor d, 1963
6.0 Missouri/w ild life area Conaway et al., 1963
Litte rs /Year 4.6 w Maryland/NS Chapman et al., 1977 5 - 7 sever al locat ions and Chapm an et al., 1980 7
habi tats
Days 28 25 - 35 sever al locat ions and Chapm an et al., 1982 Gestation habi tats
2-357 E
astern C
otto
ntail

Eastern Cottontail ( Sylvila gus floridanus )
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/Habi tat Note Cond. /Seas. Mean Range Reference No.
Age at 20 - 25 days Illinoi s/NS Ecke, 1955 Weaning
Age at F 3 - 6 mont hs s Texas/ grassl and Lor d, 1961, Negus, 1959b 8 Sexual M 3 - 6 mont hs Missour i/NS Conaw ay & Wight , 1963 Maturity
Annual B B 80 sw Wisconsi n/farm Trent & Rongst ad, 1974 Morta lity Rates (%) B B 65 ± 7 SD Illinoi s/sanct uary Lor d, 1963
Longevi ty B 1.25 Kentucky /NS Bruna, 1952 9
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating mid-March mid-September Connect icut Dalke, 1942 9 year-round Januar y - Apr il s Texas Bot hma & Teer, 1977
Partur ition Apr il May - Jul y August wc New Yor k Hamilton, 1940
Molt fall August October December s Texas Bot hma & Teer, 1982
spr ing February Apr il July s Texas Bot hma & Teer, 1982
September Sept. - Oct. Novem ber Connect icut Spinner , 1940
March May - June August Connect icut Spinner , 1940
2-358 E
astern C
otto
ntail
1 Est imated usi ng equat ion 3-43 (Boddi ngt on, 1978) and body weight s f rom Lor d (1963).2 Est imated usi ng equat ion 3-46 (Nagy, 1987) and body weight s f rom Lor d (1963).3 See Chapt ers 3 and 4 f or appr oaches t o est imating f ood i ngest ion r ates.4 Est imated usi ng equat ion 3-17 (Calder and Br aun, 1983) and body weight s f rom Lor d (1963).5 Est imated usi ng equat ion 3-20 (Stahl , 1967) and body weight s f rom Lor d (1963).6 Est imated usi ng equat ion 3-22 (Stahl , 1967) and body weight s f rom Lor d (1963).7 Summary of sever al studi es.8 Cited in Conaw ay and Wight (1963).9 Cited in Chapm an et al. (1980).
10 Cited in Chapm an et al. (1982).

References (including A ppendix)
Allen, D. L. (1938) Breeding of th e cottont ail r abbit in s out hern Michiga n. Am. Midl. Nat. 20: 464-469.
Allen, D. L. (1939) Michiga n cottont ails in w int er. J. Wildl. Manage. 3: 307-322.
Allen, A. W. (1984) Habitat su itabilit y inde x mode ls : Easter n cotto ntai l. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.66); 23 pp.
Althoff, D. P.; Storm, G. L. (1989) Dayti me sp atial character isti cs o f co tto ntai l rabbit s in central Pennsyl vania. J. Mammal . 70: 820-824.
Bailey, J. A .; Siglin, R. J. (1966) Some food pr eferences o f young c ottont ails . J. Mammal . 47: 129-130.
Barkalow, F. S., Jr . (1962) Lati tude related to reproduc tion in t he cottont ail r abbit . J. Wildl. Manage. 26: 32-37.
Beule, J. D. (1940) Cotto ntai l nest ing- study in Penns ylvania . Trans. Nor th Am. Wildl. Nat. Resour . Conf . 5: 320-328.
Bittn er, S. L.; Chapman, J. A. (1981) Reproduc tive and phy siologic al cycl es in an island popula tion of Sylvila gus florid anus. In: Myers, K. and MacInne s, C. D., eds. Proceedings wo rld la gom orph c onf erence; Augus t 1979; Guelph, Ontar io. Guelph, Ontar io, Canada: Univ ersi ty of Gu elph; pp. 182-203.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Bothma, J. P.; Teer, J. G. (1977) Reproduc tion a nd pr oduc tivi ty in s out h Texas co tto ntai l rabbits. Mammal ia 41: 253-281.
Bothma, J. P.; Teer, J. G. (1982) Moult ing in t he cottont ail r abbit in s out h Texas. Mammal ia 46: 241-245.
Bruna, J. F. (1952) Kentucky r abbit investi gations . Fed. Aid Proj. 26-R. Kentucky; 83 pp.
Burt, W. H.; Grossen heider, R. P. (1980) A field guide to the mammal s of North Amer ica nor th of Mexico. Bos ton, MA: Hought on Mif flin Co.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
2-359 Easter n Cotto ntai l

Chapman, J. A.; Ceballos , G. (1990) Chapter 5: the cotto ntai ls. In: Chapman, J. A.; Flux , J. E., eds. Rabbits, hares and pikas; statu s su rvey an d conservat ion action pla n. Internationa l Union f or Cons ervation of Nature and Natural Resour ces i n colla bor ation w ith World Wide Fund f or Nature. Oxfo rd, UK: Informat ion P ress.
Chapman, J. A.; Morgan, R. P., II. (1973) Systemati c statu s of th e cotto ntai l complex in wester n Maryland and nearby West Vi rginia . Wildl. Monogr . 36: 1-54.
Chapman, J. A.; Harman, A. L.; Samuel, D. E. (1977) Reproduc tive and phy siologic al cycl es in the cotto ntai l complex in wester n Maryland and nearby West Vi rginia . Wildl. Monogr . 56: 1-73.
Chapman, J. A.; Hockman , J. G.; Edwards, W. R. (1982) Cotto ntai ls. In: Chapman, J. A.; Feldhammer , G. A., eds. Wild mammal s of North Amer ica. Baltimore, MD: Johns Hopk ins Univ ersi ty Press; pp. 83-123.
Chapman, J. A.; Hockman , J. G.; Ojeda, M. M. (1980) Sylvila gus florid anus. Amer ican Society o f Mammalogi sts; Mammal ian Species No. 136; 8 pp.
Conaway, C. H.; Wight , H. M. (1962) Onset o f reproducti ve seaso n and fi rst p regnancy o f the seaso n in cotto ntai ls. J. Wildl. Manage. 26: 278-290.
Conaway, C. H.; Wight , H. M. (1963) Age at sexu al matu rity o f young m ale cottont ails . J. Mammal . 44: 426-427.
Conaway, C. H.; Wight , H. M.; Sadler, K. C. (1963) Annua l pr oduc tion by a cottont ail popula tion. J. Wildl. Manage. 27: 171-175.
Conaway, K.; Sadler, C.; Hazelwood, D. H. (1974) Geographic var iation in li tter size and onset o f breeding in c ottont ails . J. Wildl. Manage. 38: 473-481.
Crunde n, C. W.; Hendr ickso n, G. O. (1955) Evaluations of technique s esti mat ing a Mearns cottont ail popula tion. Proc. Iowa Acad. Sci . 62: 498-501.
Dalke, P. D. (1942) The cotto ntai l rabbit s in Conn ecti cut. Bull. Conn ecti cut Geol. Nat. Hist. Surv. 65: 1-97.
Dalke, P. D.; Sime, P. R. (1941) Food ha bit s of th e easter n and new Engla nd c ottont ails . J. Wildl. Manage. 5: 216-228.
de Poor ter , M.; van der Loo, W. (1981) Report on the breeding a nd be havior of th e volc ano rabbit at the Antwer p Zoo. In: Myers, K.; MacInne s, C. D., eds. Proceedings of th e world lagomorph conference; August 1979; Guelph, Ontar io. Guelph, Ontar io, Canada: Univ ersi ty of Gu elph; pp. 956-972.
2-360 Easter n Cotto ntai l

Dixon, K. R.; Chapman, J. A.; Rong stad , O. J.; et al . (1981) A comparison of home range size in Sylvila gus florid anus and S. bachmani. In: Myers, K.; MacInnes, C. D., eds. Proceedings of th e world la gom orph c onf erence; Augus t 1979; Guelph, Ontar io. Guelph, Ontar io, Canada: Univ ersi ty of Gu elph; pp. 541-548.
Dusi , J. L. (1952) The food ha bit s of sever al popula tions of co ttont ail r abbit s in O hio. J. Wildl. Manage. 16: 180-186.
Eberhardt, L.; Peter le, T. J.; Schofield, R. (1963) Problems i n a rabbit popula tion s tudy . Wildlif e Society Wildlif e Monogr aphs No. 10, Michiga n Departmen t of Conservati on and Ohio State Universi ty; 51 pp.
Ecke, D. H. (1955) The reproducti ve cycl e of th e Mearns co tto ntai l in Illinois . Am. Midl. Nat. 54: 294-311.
Edwards, W. R.; Havera, S. P.; Labisky, R. F.; et al . (1981) The abunda nce of co ttont ails in relation t o agricult ural land us e in Illinois (U .S.A.) 1956-1978, wi th commen ts o n mech anism o f regula tion. In: Myers, K.; MacInne s, C. D., eds. Proceedings of th e world lagomorph conference; August 1979; Guelph, Ontar io. Canada, Guelph, Ontar io: Univ ersi ty of Gu elph; pp. 761-789.
Eisenberg, J. F. (1981) The mammal ian radiations ; an analysi s of tr ends in evolut ion, adaptation, and be havior . Chic ago, IL: Univ ersi ty of Chic ago Press.
Evans, R. D.; et al . (1965) Regiona l comparisons of co ttont ail r eproduc tion in Mis sour i. Amer . Midla nd Nat. 74: 176-184.
Gerstel l, R. (1937) Management of th e cotto ntai l rabbit in Penns ylvania . Pa. Game News; May.
Hamilton, W. J., Jr . (1940) Breeding ha bit s of th e cottont ail r abbit in New York state. J. Mammal . 21: 8-11.
Haugen, A. O. (1942) Life histo ry st udie s of th e cottont ail r abbit in s out hwester n Michiga n. Am. Midl. Nat. 28: 204-244.
Heard, L. P. (1963) Notes o n cotto ntai l rabbit studie s in Mis sissippi. Proc. Annu. Conf . Southeast. Asso c. Game and Fish Comm. 17: 85-92.
Hendrickso n, G. O. (1943) Gestat ion pe riod in Me arns co ttont ail. J. Mammal . 24: 273.
Hill, E. P., III. (1972a) The cotto ntai l rabbit in A labama. Aubur n Univ ., Agric Exp. Stn. Bull. 440; 103 pp.
Hill, E. P., III. (1972b) An evaluation of sever al body measu rements fo r determining a ge in liv e juv enile co ttont ails . Proc. Annu. Conf . Sout heast. Asso c. Game and Fish Comm. 25: 269-281.
2-361 Easter n Cotto ntai l

Hill, E. P., III. (1972c) Litter size in Alabama co tto ntai ls as i nfluenced by soil f ertilit y. J. Wildl. Manage. 36: 1199-1209.
Hinds , D. S. (1973) Accl imat ion of th ermoregula tion in t he deser t co tto ntai l, Sylvi lagus auduboni i. J. Mammal . 54: 708-728.
Janes, D. W. (1959) Home range and movemen ts o f th e easter n cotto ntai l in Kansas. Univ . Kansas Publ., Mus. Nat. Hist. 10: 553-572.
Jurewicz, R. L.; Cary, J. R.; Rong stad , O. J. (1981) Spatial relations hips of breeding femal e cotto ntai l rabbits i n southwester n Wisco nsin. In: Myers, K.; MacInnes, C. D., eds. Proceedings of th e world la gom orph c onf erence; Augus t 1979; Guelph, Ontar io. Guelph, Ontar io, Canada: Univ ersi ty of Gu elph; pp. 295-309.
Kirkpatr ick, C. M. (1956) Copr opha gy in t he cottont ail. J. Mammal . 37: 300.
Leite, E. A. (1965) Relation of habi tat str uctu re to cotto ntai l rabbit produc tion, surviv al and harvest r ates. Job Prog. Rep., Ohio J .A.P.R. Proj. W-103-R-8, Job 12; 12 pp.
Lord, R. D., Jr . (1960) Litter size and lati tude in North Amer ican mammal s. Am. Midl. Nat. 64: 488-499.
Lord, R. D., Jr . (1961) Magnit udes of r eproduc tion in c ottont ail r abbit s. J. Wildl. Manage. 25: 28-33.
Lord, R. D., Jr . (1963) The cotto ntai l rabbit in Illinois . Tech. Bull. Illinois Dept . Cons erv. 3: 1-94.
Lord, R. D., Jr .; Casteel , D. A. (1960) Importance of food t o cottont ail w int er mo rtalit y. Trans. Nor th Am. Wildl. Conf . 25: 267-274.
Marsden, H. M.; Conaway, C. H. (1963) Behavior and the reproducti ve cycl e in the cottont ail. J. Wildl. Manage. 27: 161-170.
Martin, A. C.; Zim, H. S.; Nelson, A. L. (1951) Amer ican wildlif e and pla nts. New York, NY: McGraw-Hill Book Company, Inc.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Negus, N. C. (1959a) Pelage stag es in the cotto ntai l rabbit . J. Mammal . 39: 246-252.
Negus, N. C. (1959b) Breeding of suba dult co ttont ail r abbit s in O hio. J. Wildl. Manage. 23: 451-452.
Nowak, R. M.; Paradiso, J. L. (1983) Walker 's mammal s of th e world, v. 3, 4th ed. Baltimore, MD: Johns Hopk ins Univ ersi ty Press.
2-362 Easter n Cotto ntai l

Palmer , E. L.; Fowler, H. S. (1975) Fieldbook of natural his tory. New York, NY: McGraw-Hill Book Co.
Pelton, M. R.; Jenkins, J. H. (1970) Weights an d measu rements o f Georgia cotto ntai ls and an ecologic al pr inc iple . Proc. Annu. Conf . Sout heast. Asso c. Game and Fish Comm. 24: 268-277.
Pelton, M. R.; Jenkins, J. H. (1971) Produc tivi ty of Geo rgia co ttont ails . Proc. Annu. Conf . Southeast. Asso c. Game and Fish Comm. 25: 261-268.
Pelton, M. R.; Provost, E. E. (1972) Onset o f breeding a nd br eeding s ynchrony by Geo rgia cottont ails . J. Wildl. Manage. 36: 544-549.
Peterson, R. L. (1966) The mammal s of easter n Canada. Toronto, Canada: Oxfo rd Universi ty Pr ess.
Pils, C. M.; Martin, M. A. (1978) Popula tion dy namics, predator-prey relations hips an d management of th e red foxes i n Wisco nsin. Wisc. Dept. Nat. Resour.; Tech. Bull. 105; 56 pp.
Prouty, J. (1937) Cotto ntai ls of Massach usetts [master 's th esis]. MA: Massach usetts State Colle ge.
Rongstad , O. J. (1966) Biology of penned cottont ail r abbit s. J. Wildl. Manage. 30: 312-319.
Sandt, J. L.; McKee, R. M. (1978) Upland w ildlif e investi gations . Unpublis hed Federal Aid Performance Report (Maryland); W-47-7; 4 pp.
Schierbaum, D. (1967) Job completion r epor t, evalua tion of co ttont ail r abbit produc tivi ty. Albany, NY: Pittman -Robertson P roj. W-84-R-12; 21 pp.
Seton, E. T. (1929) Lives o f game animals. Garden City, NY: Double day, Doran and Co. , Inc .
Sheffer , D. E. (1957) Cotto ntai l rabbit propagation in small br eeding pe ns. J. Wildl. Manage. 21: 90.
Spencer , R. K.; Chapman, J. A. (1986) Seasonal feeding ha bit s of New Engla nd and easter n cotto ntai ls. Proc. Penn. Acad. Sci . 60: 157-160.
Spinne r, G. P. (1940) Molt ing c haracter isti cs i n the easter n cotto ntai l rabbits. J. Mammal . 21: 429-434.
Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
Trent, T. T.; Rongstad , O. S. (1974) Home range and survival of co tto ntai l rabbit s in southwester n Wisco nsin. J. Wildl. Manage. 38: 459-472.
2-363 Easter n Cotto ntai l

Trethewey, D. E.; Verts, B. J. (1971) Reproduc tion in easter n cotto ntai l rabbits i n wester n Oregon. Am. Midl. Nat. 86: 463-476.
Wainr ight , L. C. (1969) A literature review on cotto ntai l reproduc tion. Colo. Div . Game, Fish Parks, Spec. Rep. No. 19; 24 pp.
Wight , H. M.; Conaway, C. H. (1961) Weather influences o n the onset o f breeding in Missour i cottont ails . J. Wildl. Manage. 25: 87-89.
2-364 Easter n Cotto ntai l

2.3. REPTILES AND AMPHIBIANS
Table 2-3 summar izes th e species of repti les and amphibia ns inc lude d in t his
sect ion. For range map s, refer to the general references i denti fied in the indiv idua l species
prof iles. The remainde r of this sect ion is organized by sp ecies in t he order presented in
Table 2-3. The avai labilit y of inf ormation in t he publis hed lit erature var ies sub stan tially
among s pecies, whic h is r eflected in the profi les. The measu res used to descr ibe body
lengt h are inc lude d in each species profi le. Body weight is r epor ted as fresh wet we ight
(inc luding t he shell f or tu rtles), unless o therwise noted.
Unlik e bir ds and mammal s fo r which a single co mmon n ame usually co vers al l
subs pecies, man y rept ile and a mphibia n subs pecies are recognize d by differ ent co mmo n
names. For examp le, there are two subspecies of Rana clami tans: the green frog and the
bronze fr og (Sect ion 2 .3.7). There are four subs pecies of Terrapene carol ina: easter n box
turtle, three-toed box tu rtle, Flor ida box tu rtle , and Gulf Coast b ox tu rtle (Sect ion 2 .3.3). In
th is case, other species exi st th at are also known as box tu rtles: the ornate and deser t box
turtles be long t o the species T. ornata. For species that could be conf used wi th other
species unless a su bspecies co mmo n name is used, we sel ected the commo n name of th e
most wi despread subspecies to use in the tables and ti tles of th e species profi le. As wi th
the other sp ecies in t he Handbook , however , the profi le covers al l subspecies fo r the
selected species th at wer e represented in the literature reviewed.
In these profi les, we use th e word hibe rnation f or th e period of dor mancy that
rept iles and amphibia ns unde rgo dur ing w int er, when they change th eir metabolis m to
acco mmo date th e low (often near freezing) temp eratures and lack o f food ( and ox ygen).
Use of th e word for this group is cont roversial, however , because th e word was d evelope d
init ially to descr ibe mammal ian winter dormancy. Some investi gators argue that a differ ent
word, brumation, should be establis hed to descr ibe the overwinter ing dor mancy and
asso ciated metabolic ch anges for rept iles and amphibia ns (Hut chis on, 1979). Others
disagree, because s ignif icant phy siologic al changes also occur in r ept iles and amphibia ns
dur ing w int er dor mancy. They argue th at, al though t he phy siologic al changes are differ ent
from th ose in mammal s, the word hibe rnation is a ge neral term that doe s not sp eci fy what
2-365
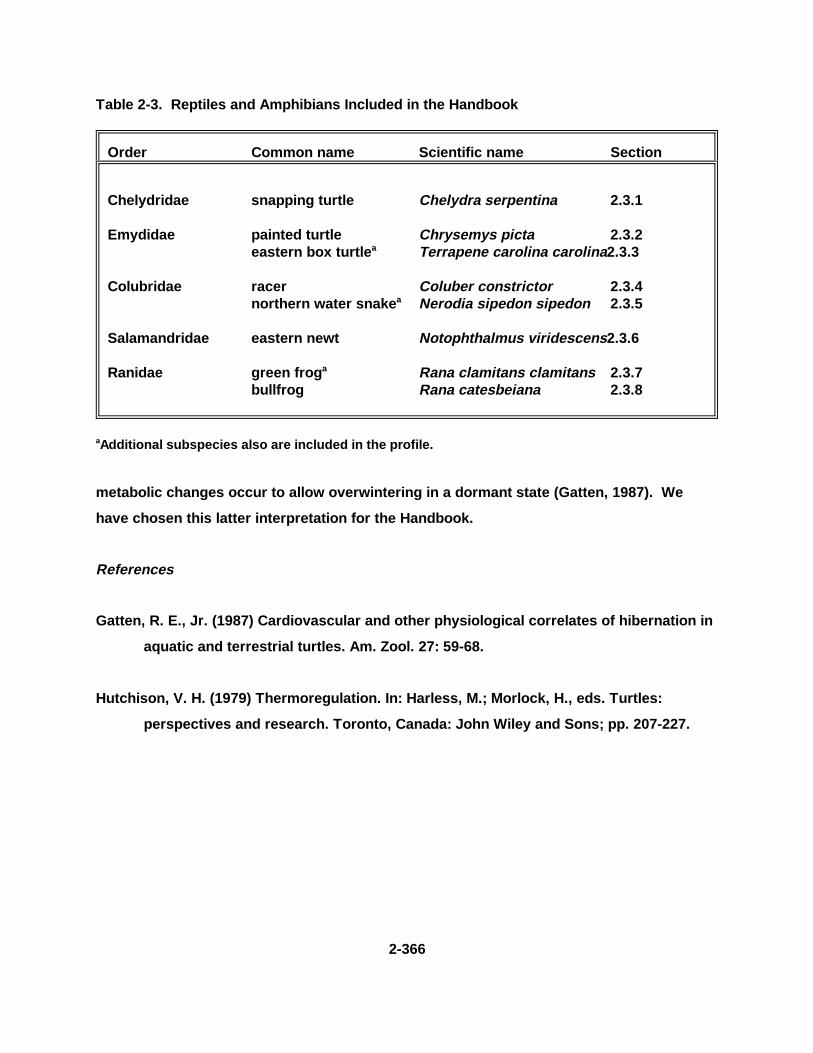
Table 2-3. Repti les and Amphibia ns Inc lude d in t he Handbook
Order Commo n name Scienti fic name Secti on
Chelydr idae snapping t urtle Chelydra serpentina 2.3.1
Emydida e paint ed turtle Chrysemys p icta 2.3.2 easter n box tu rtle Terrapene carolin a carolin a2.3.3a
Colubr idae racer Coluber constric tor 2.3.4 northern water snake Nerodi a sipedon s ipedon 2.3.5a
Salamandridae easter n newt Notopht halmus virid escen s2.3.6
Ranida e green frog Rana clami tans cl ami tans 2.3.7a
bullf rog Rana catesb eiana 2.3.8
aAdditional subspecies also are included in the profile.
metabolic ch anges oc cur to allow overwint ering in a dor mant state (Gatten , 1987). We
have ch osen this latter interpretat ion f or th e Handbook .
References
Gatten , R. E., Jr . (1987) Cardiovascu lar and other physiologic al correlates o f hibe rnation in
aquatic and ter restr ial turtles. Am. Zool. 27: 59-68.
Hutch ison, V. H. (1979) Thermoregula tion. In: Harless, M.; Morlock, H., eds. Turtles:
perspecti ves an d research. Toronto, Canada: John W iley and Sons ; pp. 207-227.
2-366

2.3.1. Snapping T urtle (snapping turtles)
Order Testudi nes, Family Chelydridae. Snapping t urtles are among t he largest o f the freshwater turtles. They are character ized by large heads wi th power ful hooked jaws. There are only two species of this family in Nor th Amer ica (the snapping t urtle , inc luding bot h the common a nd Flor ida sn apping t urtles, and t he alliga tor sn apping t urtle).
Selected species
The snapping t urtle (Chelydra serpentina) is pr imarily aqua tic, inha bit ing f reshwater and bracki sh environments, although t hey will t ravel over land (DeGraaf and Rudis , 1983; Ernst an d Barbour, 1972; Smith, 1961). There are two subspecies recognize d in Nor th Amer ica that are pr imarily dis tinguis hed by r ange: C. s. serpentin a (the common s napping turtle, which is th e largest su bspecies, primar ily occupies th e United States east o f th e Rocki es, excep t fo r the southern portions of T exas an d Flor ida), and C. s. osceo la (the Flor ida sn apping t urtle , found in t he Flor ida penins ula) (Conant and Collins , 1991). In th is profi le, stu dies refer to the serpent ina subspecies unless o therwise noted.
Body size. Adult sn apping t urtles are large, 20 to 37 cm in c arapace length, and males attai n larger sizes th an femal es (Congdon e t al., 1986; Ernst an d Barbour, 1972; Galbraith et al ., 1988). In a large oligot rophic l ake in Ontar io Canada, adult mal es averaged over 10 kg, whereas th e femal es aver aged 5.2 kg (Galbraith et al ., 1988). In other popula tions , the di ffer ence in size between males and femal es often is less (Congdon e t al., 1986; Galbraith et al ., 1988; Hammer , 1969). They reach sexu al matu rity at ap proximatel y 200 mm i n carapace length (Mosimann and Bide r, 1960). The cool, shor t acti vi ty seaso n in more northern areas results i n slower growth rates an d longe r ti mes to reach sexu al matu rity (B ury, 1979).
Habitat . In the east, snapping t urtles are found in a nd ne ar permanent ponds , lakes, and marshes. However , in the arid west, the species is primar ily found in la rger r ivers, because th ese are the only permanent water bodie s (Toner, 1960, ci ted in Graves an d Anderson, 1987). They are most o ften found in t urbid water s wi th a slow cu rrent (Graves and Anderson, 1987). They sp end most o f th eir time lying on t he bot tom of deep pools or bur ied in t he mud in s hallow water wi th only th eir eyes an d nostr ils exp osed. Froese (1978) observed that young s napping t urtles show a pr eference fo r areas wi th some obs tructions th at may p rovide cover or food.
Food ha bi ts . Snapping t urtles are omniv orous . In early sp ring, when lim ited aquatic veg etat ion e xists i n lakes an d ponds , they may eat p rimar ily animal matter ; however , when aquatic veg etat ion be comes abunda nt , they become more herbiv orous (Pell, 1941, ci ted in Graves an d Anderson, 1987). Young s napping t urtles are pr imarily carnivorous and prefer smal ler str eams wh ere aquatic veg etat ion is l ess abunda nt (L agler, 1943; Pell, 1941, ci ted in Graves an d Anderson, 1987). Snapping t urtles cons ume a wide var iety o f animal mater ial including in sects, crustacean s, clams, snails, earthworms, leeches, tubif ic id w orms, fr eshwater sponge s, fi sh (adult s, fr y, and e ggs ), frogs and t oads, salamanders, snakes, smal l turtles, birds, smal l mammal s, and car rion a nd pla nt mater ial inc luding v arious alga e (Alexander, 1943; Graves an d Anderson, 1987; Hammer , 1969;
2-367 Snapping T urtle

�
Punzo, 1975). Budha batti and Moll (1988) observed no differ ence between the diets o f males and femal es wh o fed at th e surface, midpe lagic , and be nthic l evels. Bramble (1973) suggested that th e pharyngeal mech anism o f feeding ( i.e., drawing water wi th food obj ects into the mouth) prevents sn apping t urtles from ing esting f ood a bov e the air-water inter face.
Temperature regulation and daily acti vi ties. Snappers are most acti ve at night . Dur ing t he day, th ey occas iona lly l eave th e water to bask o n shore, but basking is probably restr icted by intolerance to high temp eratures and by rapid los s of mois ture (Ernst and Barbour, 1972). In a stu dy in Ontar io, Canada, Obbard and Brooks (1981) f ound t hat the turtles wer e acti ve in the early mo rning a nd early evening a nd b asked in the after noon but were rarely acti ve at night . Acti ve turtles were found in de eper water s th an inacti ve snappers (Obbard and Brooks, 1981). Cloacal temp eratures of 18.7 to 32.6�C were repor ted for sn apping t urtles captured in t he water in Sarasota Count y, Flor ida, between May and Octobe r (Punzo, 1975).
Hibernation. Snapping t urtles us ually en ter hibe rnation by l ate Octo ber and emerge someti me between March and May, depending on la ti tude and temp erature. To hibe rnate, they bur row int o the debr is or mud bot tom of ponds or lakes, settl e beneath logs , or retreat int o muskr at bur rows or lodge s. Snapping t urtles have been seen moving on or below th e ice in m idw int er. Large congr egations so meti mes hibe rnate to gether (Budha batti and Moll, 1988; Ernst an d Barbour, 1972).
Breeding acti vi ties and social organization. Mating oc curs any time turtles are acti ve fr om sp ring t hrough f all, depending on la ti tude (Er nst and Ba rbour , 1972). Some investi gators believe th at mal e snapping t urtles are ter ritorial (Kiv iat, 1980; Pell, 1941, ci ted in Galbraith et al ., 1987), but Galbraith et al . (1987) doubt s that males defend their home ranges against o ther males. Sperm may r emain viable in the femal e for sever al years (Smith, 1956). Nest ing oc curs from l ate sp ring t o early fall, peaking in J une (Er nst and Barbour, 1972). Hammer (1969) observed that larger, olde r femal es nested earlier in the seaso n than did smalle r, younge r ones. Females often move up smal l str eams to lay eggs (Ewer t, 1976, ci ted in Graves an d Anderson, 1987). The nest si te may b e in the soil of banks o r in muskr at houses b ut mo re commonly is in t he open on s out h-facing s lope s and may b e sever al hundr ed meter s fr om water (DeGraaf and Rudis , 1983). The turtle digs a 4-to 7-in cavi ty o n dry land, preferably in sand, loam, or vegetable debris. The duration of inc ubation is inv ersely related to soil temp erature (Ernst an d Barbour, 1972; Yntema, 1978, ci ted in Graves an d Anderson, 1987). In more northerly popula tions , hatchlings may overwinter in the nest (DeGraaf and Rudis , 1983).
Home range and r esour ces. Most tu rtles stay p rimar ily wi thin t he same mar sh or in one general area from year to year ((Hammer , 1969; Obbard and Brooks, 1981). The summer home range include s a turtle 's aquatic foraging a reas, but femal es may n eed to travel some distan ce outsi de of th e foraging hom e range to find a suit able n est si te (DeGraaf and Rudis , 1983). Obbard and Brooks (1980) f ound t hat femal es tag ged at th eir nesting s ite moved an average of 5.5 km (± 1.8 SD) from th e nest si te after wards. Lonk e and Obbard (1977) observed that 91.9 percent of th e turtles in one popula tion r eturned to the same n est ing s ite a year after having be en tagge d there. Home ranges ov erlap bot h between and wi thin sexes (Ob bard and Brooks, 1981). Young s napping t urtles us e
2-368 Snapping T urtle

�
differ ent habitats th an adults; they ten d to remain in smal l str eams u nti l shortly before matu rity, when they mi grate to habitats p refer red by adult s (e.g., ponds , marshes, lakes) (Hammer , 1971; Mint on, 1972, ci ted in Graves an d Anderson, 1987).
Popul ation de nsity . The density of sn apping t urtles appears to be pos it ively correlated wi th the producti vi ty o f th e surface water body (e. g., dens ity in a eu trophic surface water body is highe r th an in an oligot rophic l ake) (Galbraith et al ., 1988). Speci fic habitat ch aracter isti cs an d intraspeci fic interact ions cont ribut e to the var iabilit y of obs erved popula tion de nsit ies in s napping t urtles (Froese and Burghardt, 1975).
Popul ation dy namics. Females do not begin la ying e ggs unt il age 6 to 19 yr depending on la ti tude and w hen they reach an appropriate si ze (approximatel y 200 mm carapace) (Galbraith et al . 1989; Mosimann and Bide r, 1960). Males matu re a few year s earlier than femal es (see tab le). Females may l ay one or two clutch es per seaso n (Mint on, 1972, ci ted in Graves an d Anderson, 1987). Clutch size increases wi th femal e body size ; Congdon e t al. (1987) calculated the relations hip b etween clutch size (CS) and plastr on length (PL in mm) fo r a popula tion in s out heaster n Michiga n:
CS = -21.227 + 0.242 PL, (r2 = 0.409, n = 65).
Clutch size has also been posi tively co rrelated wi th lati tude (Petokas an d Alexander, 1980). Hammer (1969) found t hat mammal ian predators destr oyed over 50 percent of th e turtle nests i n a South Dakota mar sh, and in undis turbed nests, hatchling s uccess was l ess th an 20 percent. Petokas an d Alexander (1980) observed a 94 percent predation r ate of nests unde r study in nor thern New York. Adult mo rtalit y is low , corresponding w ith the long lives ex hibit ed by th ese tu rtles (see tab le).
Simi lar species (fr om general references)
� The alliga tor sn apping t urtle (Macroclemys temmi ncki ) is mu ch larger (16 to 68 kg; 38 to 66 cm car apace) th an the commo n snapping t urtle and is one of the largest tu rtles in the world. Its r ange is fr om northern Flor ida to east-central Texas an d north in the Missi ss ippi V alley.
General references
Conant and Collins (1991); DeGraaf and Rudis (1983); Ernst an d Barbour (1972); Graves an d Anderson (1987).
2-369 Snapping T urtle

Snapping T urtle ( Chelydra ser pentina )
2-370 S
napping T
urtle
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight A M summer 10.5 ± 2.85 SD Ontario , Canada/large Galbraith et al., 1988 (kg) A F sum mer 5.24 ± 0.85 SD ol igot rophi c lake
J B sum mer 1.15 ± 0.80 SD
A M sum mer 5.52 ± 2.23 SD Ontario, Canada/eutrophi c Galbraith et al., 1988 A F sum mer 5.03 ± 1.12 SD pond J B sum mer 1.40 ± 0.20 SD
A M 4.16 ± 0.28 SE Michi gan Congdon et al., 1986 A F 3.16 ± 0.20 SE J B 0.80 ± 0.07 SE
at hatching 0.0057 NS Ernst & Bar bour , 1972 at hatching 0.0089 NS Ewert, 1979
mm carapace: 118 0.33 Massachuset ts Graham & Perkins, 1976 127 0.44 134 0.53 167 1.03 192 1.51 220 2,362
Egg Weight 7 - 15 NS Ernst & Bar bour , 1972 (g) 11.1 nor thern New York Petokas & Al exander , 1980
9.6 Sout h Carolina Congdon et al., 1986 9.3 5.7 - 13.8 New Jersey Hotalin g et al., 1985
Body Lengt h age in years Michigan Gibbons, 1968 (mm carapace) 1 62 ± 4.5 SD 54 - 66
2 102 ± 5.8 SD 83 - 108 3 137 ± 9.4 SD 124 - 145 4 168 ± 14.2 SD 146 - 184 5 198 ± 13.7 SD 177 - 211 6 222 ± 12.9 SD 204 - 238

Snapping T urtle ( Chelydra ser pentina )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Metabol ic Rate 7.18 kg, rest 2.54 Lynn & von Br and, 1945 1 (lO /kg-d) 25��C2
Metabol ic Rate A F basal 3.2 est imated 2 (kcal /kg-d) A M basal 3.0
Food Ingest ion B sum mer 0.01 - 0.016 New Yor k/capt ivi ty Kiviat, 1980 Rate (g/g-d)
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
adul ts & juveni les: locat ion not speci fied Smith, 1956 3 plants 35 - 70 animals 6 - 35 (% of diet; measur e NS)
adul ts: Tennessee/ embayment Meyers-Schoene & W alton, fish 83.7 1990 veget ation 13.6 (% wet vol ume; gast roclams 0.2 intest inal tract cont ents) mud & r ocks 2.5
adul ts & juveni les: Connect icut /lakes, ponds, Alexander , 1943 (plants) (36.5) streams, swamps algae 12.8
(animals) (54.1) (% wet vol ume; stomach crayfish 8.9 cont ents) fiddl er crab 2.7 sucker 3.2 bullh ead 6.3 sunfish 7.5 unknown f ish 12.4
(miscel laneous) (9.4)
2-371 S
napping T
urtle

Snapping T urtle ( Chelydra ser pentina )
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Home Range A M summer 0.7 ± 0.29 SD 0.24 - 1.3 Ontario , Canada/lake Galb raith et al., 1987 Size (ha)
A F sum mer 3.79 ± 1.46 SD 2.5 - 5.19 Ontario, Canada/ lake Obbar d & Br ooks, 1981 A M sum mer 3.21 ± 2.67 SD 0.95 - 8.38 A B sum mer 3.44 ± 2.18 SD
A M 8.9 New York/fresh t idal Kiviat, 1980 A F nonbr eed 7.2 wetland
Popul ation Ontario, Canada/ Densi ty A M sum mer 1.5 ol igot rophi c lake Galbraith et al., 1987 (N/ha) B B sum mer 2.3 ± 1.45 SD 1.0 - 4.9 ol igot rophi c wat ers Galbraith et al., 1988 4
B B sum mer 60.4 40.3 - 95.0 eutrophi c pond Galbraith et al., 1988 B B sum mer 29.3 ± 27.6 SD 4.4 - 65.9 eutrophi c ponds ( other Galbraith et al., 1988 5
A B sum mer 59 Tennessee/ pond Froese & Bur ghar dt, 1975 studies)
Clutch Si ze 49.0 31 - 87 Sout h Dakot a/marsh Hammer, 1969 27.9 ± 0.76 SE 12 - 41 se Michi gan/NS Congdon et al., 1987 16.6 ± 1.6 SD 14 - 20 Flor ida/NS Iverson, 1977 6
Clutches/ Yea 1 - 2 Indi ana/NS Minton, 1972 7 r > 1 NS/summarizing ot her Ernst & Bar bour , 1972
studi es
Days 105 90 - 119 Ontario, Canada/ lake Obbar d & Br ooks, 1981 Incubat ion 67 - 73 se Wisconsi n/NS Ewert, 1979
Age at Sexual F nesting 6 - 8 New York /NS Pell, 1941 8 Maturity (yr)
F nest ing 9 - 10 Iowa/NS Christiansen & Bur ken, 1979 M - 5
F nest ing 17 - 19 at least 14 to 15 Ontario, Canada/river ine, Galbraith et al., 1989 mixed f orest
4
2-372 S
napping T
urtle

�
Snapping T urtle ( Chelydra ser pentina )
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Lengt h at A B 200 mm carapace Quebec, Canada/NS Mosi mann & Bi der, 1960 Sexual Maturity A B 145 mm plastron Tennessee/ NS White & Murphy , 1973 9
Annual A B 3 - 7 NS/NS Galbraith & Br ooks, 1987 10 Morta lity Rates (%)
Longevi ty (yr) at least 24 Michi gan/marsh Gibbons, 1987
at least 19 Sout h Carolina/river Gibbons, 1987
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating Apr il June Novem ber depends on l atitude Ernst & Bar bour , 1972 early June mid-June end of June New York Kiviat, 1980 mid-June Flor ida Punzo, 1975
Nest ing May June September depends on l atitude Ernst & Bar bour , 1972 late May early to mid-June late June nor thern New York Petokas & Al exander , 1980 early June mid-June end of June Sout h Dakot a Hammer, 1969
Hatching August October depends on l atitude Ernst & Bar bour , 1972 late August September early October se Michi gan Congdon et al., 1987
Hibernation October March-May depends on l atitude Ernst & Bar bour , 1972 late September mid-March Iowa Chr ist iansen & Bur ken, 1979 mid-October early May Ontario, Canada Obbar d & Br ooks, 1981
2-373 S
napping T
urtle
1 Cited in Sievert et al. (1988). 2 Est imated assum ing t emperature of 20�C, using equatio n 3-50 (Robinson et al., 1983) and body weight s f rom Congdon et al. (1986), after
subt ract ing 30 per cent of body weight to eliminate the weight of the shel l (Hall, 1924). More information on est imating ener gy budget s for rept iles is provi ded in Congdon et al. (1982).
3 Method of est imating per cent diet not speci fied. 4 Summary of six f ield st udi es, includi ng t he author 's. 5 Summary of data from var ious aut hor s for eleven eut rophi c ponds. 6 Cited in Petokas and Al exander (1980). 7 Cited in Graves and Ander son (1987).

Snapping T urtle ( Chelydra ser pentina )
8 Cited in Galbraith et al. (1989). 9 Cited in Bur y (1979).
10 Cited in Frazer et al. (1991).
2-374 S
napping T
urtle

References (including A ppendix)
Alexander, M. M. (1943) Food ha bit s of th e snapping t urtle in Conn ecti cut. J. Wildl. Manage.7: 278-282.
Barbour, R. W. (1950) The repti les of Big Bl ack Mount ain, Harlan Count y, Kentucky. Copeia 1950: 100-107.
Bramble, D. M. (1973) Media dependent feeding in t urtles. Am. Zool. 13: 1342.
Brecken ridge , W. J. (1944) Repti les and amphibia ns of Minne sota. Minne apolis , MN: Univ ersi ty of Minne sota Press.
Budha bhatti , J.; Moll, E. O. (1988) Diet and acti vi ty p atter ns of th e commo n snapping t urtle , Chelydra serpentina (Linna eus) at Chain O'Lakes State Par k, Lake Count y, Illinois (abstr act only ). Bull. Ecol. Soc. Am. Suppl. 69: 86.
Bury, R. B. (1979) Popula tion e cology of fr eshwater turtles. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 571-602.
Bush, F. M. (1959) Foods of so me Kentucky h erpti les. Herpetologic a 15: 73-77.
Cahn, A. R. (1937) The turtles of Illinois . Illinois Biol. Monogr . 35: 1-218.
Carr, A . F. (1952) Handbook of tu rtles. Ithaca, NY: Comsto ck.
Christi ansen, J. L.; Burken, R. R. (1979) Growth and matu rity o f th e snapping t urtle (Chelydra serpentina) in Iow a. Herpetologic a 35: 261-266.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns - easter n and central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
Congdon, J. D.; Gibbons , J. W. (1985) Egg compone nts and r eproduc tive character isti cs o f turtles: relations hips to body size . Herpetologic a 41: 194-205.
Congdon, J. D.; Dunha m, A. E.; Tink le, D. W. (1982) Energy budgets an d life histo ries of rept iles. In: Gans, C., ed. Biology of th e rept ilia . v. 13. New York, NY: Academic Press; pp. 233-271.
Congdon, J. D.; Greene, J. L.; Gibbons , J. W. (1986) Biomass o f fr eshwater turtles: A geogr aphic co mparison. Am. Midl. Nat. 115: 165-173.
Congdon, J. D.; Tink le, D. W.; Rosen, P. C. (1983) Egg compone nts and ut iliza tion dur ing developm ent in a quatic turtles. Copeia 1983: 264-268.
2-375 Snapping T urtle

Congdon, J. D.; Breitenba ch, G. L.; van Lobe n Sels, R. C.; et al. (1987) Reproduc tion a nd nesting e cology of sn apping t urtles (Chelydra serpentina) in southeaster n Michiga n. Herpetologic a 43: 39-54.
DeGraaf, R. M.; Rudis , D. D. (1983) Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess; p. 42.
Ernst, C. H. (1968) A turtle's ter ritory. Int. Turtle Tortoise Soc. J. 2: 9-34.
Ernst, C. H. (1971) Popula tion dy namics and acti vi ty cycl es of Chrysemys p icta in southeaster n Pennsyl vania. J. Herpetol. 140: 191-200.
Ernst, C. H.; Barbour, R. W. (1972) Turtles of th e United States. Lexingt on, KY: Univ ersi ty Press o f Kentucky.
Ewer t, M. A. (1976) Nests, nest ing a nd aerial basking of Macroclemys unde r natural condit ions , and c omparisons wi th Chelydra (Testudine s: Chelydr idae). Herpetologic a 32: 150-156.
Ewer t, M. A. (1979) The embryo and its eg g: development and natural histo ry. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John Wiley and Sons, Inc.; pp. 333-413.
Frazer, N. B.; Gibbons , J. W.; Greene, J. L. (1991) Growth , survivorship a nd longe vi ty of painted turtles Chrysemys p icta in a southwester n Michiga n marsh. Am. Midl. Nat. 125: 245-258.
Froese, A. D. (1978) Habitat p references o f th e commo n snapping t urtle , Chelydra s. serpent ina (Reptilia , Testudine s, Chelydr idae). J. Herpetol. 12: 53-58.
Froese, A. D.; Burghardt, G. M. (1975) A dense natural popula tion of th e common s napping turt le (Chelydra s. serpentina). Herpetologic a 31: 204-208.
Galbraith, D. A.; Brooks, R. J. (1987) Survivorship of adult femal es in a northern popula tion of co mmon s napping t urtles, Chelydra serpentina. Can. J. Zool. 65: 1581-1586.
Galbr aith, D. A.; Chandle r, M. W.; Brook s, R. J. (1987) The fine str uctu re of home ranges of male Chelydra serpentina: are snapping t urtles ter ritorial? Can. J. Zool. 65: 2623-2629.
Galbr aith, D. A.; Bis hop, C. A.; Brook s, R. J.; et al. (1988) Facto rs affect ing t he density of popula tions of co mmon s napping t urtles (Chelydra serpentina serpentina). Can. J. Zool. 66: 1233-1240.
Galbraith, D. A.; Brooks, R. J.; Obbard, M. E. (1989) The influence of growth rate on age and body size at matu rity i n femal e snapping t urtles (Chelydra serpentina). Copeia 1989: 896-904.
2-376 Snapping T urtle

Gerholdt , J. E.; Oldf ield, B. (1987) Chelydra serpentina serpentina (common s napping turtle), size. Herpetol. Rev. 18: 73.
Gibbons , J. W. (1968) Growth rates o f th e commo n snapping t urtle , Chelydra serpentina, in a pollut ed river . Herpetologic a 24: 266-267.
Gibbons , J. W. (1987) Why do turtles l ive so long? BioS ci . 37: 262-269.
Graham, T. E.; Perkins, R. W. (1976) Growth of th e commo n snapping t urtle , Chelydra s. serpent ina, in a pollut ed marsh. Bull. Md. Herpetol. Soc. 12: 123-125.
Graves, B. M.; Anderson, S. H. (1987) Habitat su itabilit y inde x mode ls : snapping t urtle . U.S. Fish Wildl. Serv. Biol. Rep. 82(10.141); 18 pp.
Hall, F. G. (1924) The respiratory exch ange in turtles. J. Metab. Res. 6: 393-401.
Hammer , D. A. (1969) Parameter s of a mar sh snapping t urtle popula tion, La-creek Refuge, Sout h Dakota. J. Wildl. Manage. 33: 995-1005.
Hammer , D. A. (1971) The durable snapping t urtle . Nat. His t. 80: 59-65.
Hotaling, E. C.; Wilhof t, D. C.; McDowell, S. B. (1985) Egg posi tion a nd w eight of h atchling snapping t urtles, Chelydra serpentina, in natural nests. J. Herp. 19: 534-536.
Hutch ison, V. H. (1979) Thermoregula tion. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 207-227.
Iverson, J. B. (1977) Reproduc tion in f reshwater and ter restr ial turtles of north Flor ida. Herpetologic a 33: 205-212.
Kiviat, E. (1980) A Huds on Riv er tid e-marsh snapping t urtle popula tion. In: Trans. Northeast. Sec. Wildl. Soc. 37th Northeast, Fish and Wildl. Conf .; April 27-30, 1980; Ellenv ille , NY; pp. 158-168.
Lagler, K. F. (1943) Food ha bit s and econom ic r elations of th e turtles of Michiga n with special reference to game man agement. Am. Midl. Nat. 29: 257-312.
Lagler, K. F.; Apple gate, V. C. (1943) Relations hip b etween the length and the weight in t he snapping t urtle Chelydra serpentina Linna eus. Am. Nat. 77: 476-478.
Lonke, D. J.; Obbard, M. E. (1977) Tag success, dimensions , clut ch size and n esting s ite fide lit y for th e snapping t urtle , Chelydra serpentina (Reptilia , Testudine s, Chelydridae), in Algonquin P ark, Ontar io, Canada. J. Herpetol. 11: 243-244.
Lynn, W. G.; von Brand, T. (1945) Studie s on t he oxygen cons umpt ion a nd water metabolis m of tu rtle emb ryos . Biol. Bull. 88: 112-125.
2-377 Snapping T urtle

Macnamara, C. (1919) Notes o n turtles. Ottawa N atural. 32: 135.
Major, P. D. (1975) Densi ty o f sn apping t urtles, Chelydra serpentina in wester n West Virginia . Herpetologic a 31: 332-335.
Meyers-Schone , L.; Walton, B. T. (1990) Comparison of two freshwater turtle species as monit ors of en vironm ental con tamina tion. Oak Ridge , TN: Oak Ridge Nationa l Lab.; Environmental Sciences Division P ubli cation No. 3454. ORNL/TM-11460.
Mint on, S. A., Jr . (1972) Amphibia ns and r ept iles of India na. India napolis , IN: India na Academy o f Sci ence.
Moll, E. O. (1979) Reproducti ve cycl es and adaptat ions . In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 305-331.
Mosimann, J. E.; Bide r, J. R. (1960) Variation, sexu al dimorphism, and matu rity i n a Quebec popula tion of th e snapping t urtle , Chelydra serpentina. Can. J. Zool. 38: 19-38.
Obbard, M. E. (1983) Popula tion e cology of th e common s napping t urtle , Chelydra serpent ina, in north-central Ontar io [Ph.D. disser tat ion] . Guelph, Ontar io Canada: Univ ersi ty of Gu elph.
Obbard, M. E.; Brooks, R. J. (1980) Nest ing m igr ations of th e snapping t urtle (Chelydra serpent ina). Herpetologic a 36: 158-162.
Obbard, M. E.; Brooks, R. J. (1981) A radio-telemetr y and mark-recapture stu dy of acti vi ty in t he common s napping t urtle , Chelydra serpentina. Copeia 1981: 630-637.
Pearse, A. S. (1923) The abunda nce and m igr ation of tu rtles. Ecology 4: 24-28.
Pell, S. M. (1940) Notes o n the food ha bit s of th e common s napping t urtle . Copeia 2: 131.
Pell, S. M. (1941) Notes o n the habits o f th e commo n snapping t urtle , Chelydra serpentina (Linn. ) in c entral New York [master 's th esis]. Ithaca, NY: Cornell Universi ty.
Petokas, P. J.; Alexander, M. M. (1980) The nest ing of Chelydra serpentina in nor thern New York. J. Herpetol. 14: 239-244.
Punzo, F. (1975) Studie s on t he feeding be havior , die t, nesting ha bit s and temp erature relations hips of Chelydra serpentina osceo la (Chelonia : Chelydr idae). J. Herp. 9: 207-210.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
2-378 Snapping T urtle

Siever t, L. M.; Siever t, G. A.; Cupp, P. V., Jr . (1988) Metabolic r ate of feeding a nd fasti ng juv enile midla nd pa int ed turtles, Chrysemys p icta mar ginata. Comp. Bioc hem. Physiol. A Comp. Physiol. 90: 157-159.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9; pp. 134-136.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28: 118-120.
Toner, G. C. (1960) The snapping t urtle . Can. Audubon 2 2:97-99.
White, J. B.; Murphy, G. G. (1973) The reproducti ve cycl es and sexu al dimorphism o f th e common s napping t urtle , Chelydra serpentina serpentina. Herpetologic a 29: 240-246.
Wilhof t, D. C.; del Bagliv o, M. G.; del Bagliv o, M. D. (1979) Observat ions of mammal ian predation of sn apping t urtle n ests (R eptilia , Testudine s, Chelydr idae). J. Herpetol. 13: 435-438.
Yntema, C. L. (1968) A ser ies of stag es in the embryonic d evelopment of Chelydra serpent ina. J. Morphol. 125: 219-252.
Yntema, C. L. (1970) Observat ions on femal es and eggs of th e commo n snapping t urtle , Chelydra serpentina. Am. Midl. Nat. 84: 69-76.
Yntema, C. L. (1978) Incubation t imes fo r eggs of th e turtle Chelydra serpentina (Testudine s: Chelydr idae) at var ious temp eratures. Herpetologic a 34: 274-277.
2-379 Snapping T urtle


2.3.2. Painted T urtle (pond and m arsh turtles)
Order Testudi nes, Family Emyd idae. Pond a nd m arsh turtles (i.e., slide rs, coot ers, red-bellie d turtles, and pa int ed turtles) are smal l to medium-si zed semi aquatic tu rtles wel l know n for basking in t he sun. Paint ed turtles are the most wide spread of th ese in North Amer ica, ranging a cross the cont inent .
Selected species
The painted turtle (Chrysemys p icta) is largely aquatic, liv ing in s hallo w-water habitats, and is among t he most cons pic uous of th e basking t urtles. There are four subspecies in the United States ( only one r eaching s light ly int o Canada), dis tinguis hed by color var iations , body size , and r ange: C. p. picta (easter n painted turtle; 11.5 to 15.2 cm; range Nova Sco tia to Alabama), C. p. marginata (midla nd pa int ed turtle ; 11.5 to 14 cm; range southern Quebec and southern Ontar io to Tennessee), C. p. dorsalis (sout hern paint ed turtle ; 10 to 12.5 cm; range sout hern Illinois to the Gulf ), and C. p. bellii (wester n painted turtle; the largest o f th e subspecies, 9 to 18 cm; range southwest On tar io and Misso uri to the Paci fic Northwest) ( Conant and Collins , 1991). C. p. dorsalis is th e smal lest subspecies and also one of th e smal lest emy did t urtles in Nor th Amer ica (Moll, 1973). Hybr idiza tion oc curs between subspecies in areas wh ere their ranges over lap (e.g., bellii × marginata hybrids may o ccu r in areas of Michiga n) (Snow , 1980).
Body size. Painted turtles are medium-si zed turtles (10 to 18 cm). Males are smal ler than femal es; adult mal es average from 170 to 190 g, whereas adult femal es average from 260 to 330 g in some popula tions (Congdon e t al., 1986; Ernst 1971b ). In general, the shell comprises ap proximatel y 30 percent of th e total wet we ight of tu rtles of th is si ze (Hall, 1924). Frazer et al . (1991) esti mated a relations hip b etween plastr on length (PL in mm) an d age (t in years) fo r a popula tion in Mic higa n in t he 1980's using v on Bertalanffy g rowth equations :
PL = 111.8(1 - 0.792e-0.184t) for males, and
PL = 152.2(1 - 0.852e-0.128t) for femal es.
Congdon e t al. (1982) reported a relations hip b etween plastr on length (PL in mm) an d body weight (W t in gr ams) fo r painted turtles:
loge(Wt) = -6978 + 2.645 loge(PL).
Eggs weigh 4 to 6 g, and ne onates r etain a large yolk mass th at th ey draw on for the fi rst few mo nths of l ife (Cagle, 1954).
Habitat . Painted turtle habitat r equir ements i nclude so ft and m uddy bot toms, basking s ites, and aquatic veg etat ion (Sexto n, 1959). Painted turtles prefer slow-mo ving shallow water such as ponds , marshes, dit ches, prairie sloughs , spring r uns , canals, and occas iona lly bracki sh tidal marshes (Conant and Collins , 1991). They fr equent areas wi th floa ting s urface veg etat ion f or feeding a nd f or co ver (Sexto n, 1959). These areas ten d to
2-381 Painted Turtle

�
�
� �
be warmer than more open water , which is important in the early fal l as temp eratures begin to drop (Sexto n, 1959). For winter hibe rnation or dor mancy, paint ed turtles seek d eeper water (Sexto n, 1959). If outlying m arsh areas are dry during t he summer , the turtles may return to the more permanent bodie s of water soone r (McAuli ffe, 1978). Painted turtles someti mes inha bit stagna nt and pollut ed water (Smith, 1956).
Food ha bi ts. Paint ed turtles are omniv orous . Depending on ha bitat an d on age, painted turtles may co nsume predominantly veg etat ion or predom inant ly anim al matter . Marchand (1942, ci ted in Mahmoud a nd Kli cka, 1979) found in one popula tion t hat juv enile s consumed approximatel y 85 percent animal matter and 15 percent plant matter , whereas the adult s were pr imarily herbiv orous , cons uming 8 8 percent pla nt matter and 12 percent insects an d amphipods . Knight and G ibbons (1968) found oligoc haets, cladocera, dragonf ly nymphs , lepidopt eran larvae, and tendipe did la rvae and pupae to dominate th e anim al compone nt of th e die t and f ilamentous algae to dominate th e plant co mpone nt of the die t in a popula tion liv ing in a pollut ed river in Mic higa n. Adult paint ed turtles in a Penns ylvania popula tion w ere found t o cons ume only 40 pe rcent pla nt matter (Ernst an d Barbour, 1972), whereas in a Michiga n marsh and e lsewh ere, painted turtles of al l ages apparently co nsumed 95 to 100 percent plant matter (Cahn, 1937, ci ted in Smith, 1961; Gibbons , 1967). Some car rion a lso may b e consumed (Mount , 1975).
Temperature regulation and daily acti vi ties. Paint ed turtles are diur nal and us ually spend t heir night s sleeping s ubm erged (Ernst, 1971c). During t he day, th ey forage in t he late mo rning a nd l ate after noon a nd b ask d uring t he rest o f th e day (Ernst, 1971c). Acti ve feeding doe s not occur unt il water temp eratures approach 20�C, and these tu rtles are most acti ve around 2 0.7 to 22.4�C (Ernst, 1972; Ernst an d Barbour, 1972; Hutchins on, 1979). Basking is mo st frequent in t he spring, summer , and fal l, but occas iona lly paint ed turtles bask d uring w arm spells in t he wint er (Ernst and Ba rbour , 1972). Sexto n (1959) divided the annua l acti vi ty cycl e of painted turtles into five parts: (1) the prevernal, which begins wi th the fina l melting of wint er ice and l asts u nti l late March, or when the turtles begin t o move in mass o ut of th e hibe rnation ponds ; (2) the vernal, from late March to late May, when the submerged aquatic plants i mportant to the turtles grow to the surface o f th e water (the init iation of feeding a nd mating acti vi ties and the emergence of th e hatchling t urtles from the nests o f th e previous year also occu r during t his seaso n); (3) the aesti val , extending from June th rough A ugus t, wh en the turtles forage, grow, nest, and return to their winter hibe rnation ponds ; (4) the autumnal, including S eptemb er through Novemb er or when a permanent ice co ver forms; and (5) the winter seaso n, which lasts w hile th e water is permanently co vered wi th ice.
Hibernation. Most pa int ed turtles become dor mant dur ing t he colde r mont hs but will be come acti ve during w arm periods in t he wint er (Ernst and Ba rbour , 1972). C. picta usually hibe rnates i n muddy bot toms of ponds (D eGraaf and Rudis , 1983). Taylor and Nol (1989) found pa int ed turtles ov erwint ering in a n Ontar io pond in a reas wi th a mean water depth of 0.32 m (range 0.2 to 0.48 m), mean sediment depth of 0.79 m (0.5 to 0.95 m), and mean sediment temp erature of 4.1�C (3 to 6�C). Dur ing hibe rnation, paint ed turtles shif t toward more anaerobic metabolis m, suppor ted by gly coly sis of liv er and skel etal muscl e glyco gen (Seymo ur, 1982). After emerging f rom hibe rnation, the turtles conv ert the accu mulated lactate to gluc ose in t he liv er (using a erobic metaboli sm) (Seymo ur, 1982).
2-382 Painted Turtle

�
Breeding acti vi ties and social organization. Mating us ually occurs in s pring a nd summer but may co ntinue int o the fall ( Ernst, 1971c; Gibbons , 1968a; Gist et al ., 1990). Nesting oc curs somewh at later (Cagle, 1954; Ernst an d Barbour, 1972; Moll 1973). Eggs are often laid in high ba nks (DeGraaf and Rudis , 1983). The species does not appear to be ter ritorial and c an be found in la rge aggr egations , partic ularly at favo rite bask ing s ites (Ernst, 1971c).
Home range and resources. In spring, as the wint er ice melts, man y paint ed turtles move away fr om th e ponds in w hic h they hibe rnated to more shallow ponds and m arshes wi th surface veg etat ion (Sexto n, 1959). Movemen ts aver aging 6 0 to 140 meter s character ized one popula tion in Mic higa n (Sexto n, 1959). The summer home range inc lude s the paint ed turtle 's foraging a reas and basking s ites. Females f ind n esting s ites on dr y land out side of th e foraging r ange; Congdon a nd Gatten (1989) found nests to average 60 meter s fr om th e edge of a fo raging m arsh. Females init iate nest ing m igr ations dur ing da ylight hour s, and m ost finis h their nests b efore dark on the same d ay (Congdon and Gatten , 1989). In winter , painted turtles generally mo ve back to the deeper ponds fo r hibe rnation ( DeGraaf and Rudis , 1983).
Popul ation de nsity. Reported densi ties range from 11.1/ha in Saskatch ewan (MacCulloc h and Secoy, 1983) to 830/ha in Michiga n marshes (Frazer et al., 1991). Accu rate cen suses ar e diffi cult, however (Bayless, 1975), and the distr ibut ion of paint ed turtles in summer is highly clum ped, corresponding t o the patch es of fl oating a quatic vegetat ion (Sexto n, 1959).
Popul ation dy namics. Sexual matu rity i s attai ned in about 2 to 7 years, depending on the sex an d size of th e turtle and growing seaso n (Christi ansen and Moll, 1973; Ernst and Barbour, 1972). Males reach sexu al matu rity 1 to a few year s ear lier than femal es (Moll, 1973). Once sexu al matu rity i s reached, growth of painted turtles sl ows o r essen tially ceases (Er nst an d Barbour, 1972). Older, larger femal es ten d to produc e larger clut ch sizes and la rger eggs th an younge r, smal ler femal es (Mitch ell, 1985). In more sout herly popula tions , paint ed turtles pr oduc e more clut ches annua lly wi th fewer eggs each than in more northerly popula tions (Moll, 1973; Snow, 1980; Schwarzkopf and Brooks, 1986). Predation c auses mo st n est l osses, usually wi thin t he fi rst 2 days after laying (Tinkle et al ., 1981). The duration of th e inc ubation pe riod de pends on s oil temp erature, and ha tchlings may o verwinter in the nest i n more northerly popula tions (Gibbons an d Nelson, 1978).
Simi lar species (fr om general references)
Many sp ecies of pond a nd m arsh turtles can be found in s imila r habitats; however , there are important dietary differ ences am ong s pecies that can affect exp osure to environmental contami nants, as descr ibed below. Size is l isted acco rding t o carapace lengt h, whic h is longe r th an plastr on length.
cooters
� The Flor ida coot er (Pseudemys fl oridana) is larger (23 to 33 cm) th an the painted turtle. The floridana subspecies ranges fr om th e coastal plain of
2-383 Painted Turtle

�
�
�
�
�
�
�
Virginia to easter n Texas an d north in the Missi ss ippi V alley to sout hern Illinois , while th e peninsularis subspecies is restr icted to the Flor ida penins ula. The Flor ida coot er reside s in pe rmanent bodie s of water . In their fi rst year , young c oot ers feed on both aquatic plant and animal life; later they become totally herbiv orous .
� The river cooter (Pseudemys co ncinna), composed of fi ve su bspecies, also is larger (23 to 33 cm) th an the painted turtle. It inha bit s coastal plains ranging f rom sout heaster n Virginia to Georgia , sout heast i nto Flor ida, west into Texas an d New Mexico, and north in the Missi ss ippi V alley to sout hern Illinois . It is chie fly a reside nt of str eams an d relatively large lakes. In their fi rst year , young r iver coot ers are omniv orous ; the adult s are almost enti rely herbiv orous .
� The Texas r iver cooter (Pseudemys texan a) (18 to 25.5 cm) p refers r ivers but can be found in smal ler creeks an d ditch es. Its r ange is restr icted to most o f central and southeaster n Texas.
red-bellie d turtles
� The Flor ida r ed-bellie d turtle (Pseudemys n elsoni) is larger (20 to 31 cm) than the paint ed turtle and ha s a range in t he Flor ida penins ula an d panhandle . It can be found b asking on logs over fr esh to mode rately bracki sh water , and it prefers abunda nt subm erged aquatic vegetation, its pr inc ipa l food.
� The Alabama red-bellie d turtle (Pseudemys al abamensis) is larger (23 to 33 cm) th an the painted turtle and is found only in t he low er por tion of th e Mobile Bay dr ainage in A labama. It prefers fr esh to moderately bracki sh water wi th abunda nt aqua tic vegetation, its pr inc ipa l food.
� The red-bellie d turtle (Pseudemys r ubriventr is) is mu ch larger (25 to 32 cm) than the paint ed turtle and is found in t he mid- Atlant ic states an d easter n Massach usetts.
slide rs
� The pond s lide r (Trachemys scr ipta) is si mi lar in size or a littl e larger (12 to 20 cm) th an the painted turtle and has th ree subspecies ranging f rom southeaster n Virginia to nor thern Flor ida and west to New Mexico. During the fi rst year , pond s lide rs are pr inc ipa lly car niv orous , cons uming a quatic insects, crustacean s, mollu scs, and tadpole s. As they matu re, slide rs become herbiv orous , cons uming a wide var iety o f aquatic plants.
� The big be nd s lide r (Trachemys g aigeae) (12 to 20 cm) i s si mi lar to the pond slide r in s ize and ha bit s. It is abunda nt loc ally in it s lim ited range along t he uppe r Rio Grande and s ome of its tr ibut aries.
2-384 Painted Turtle

General references
Behler and King (1979); Conant and Collins (1991); Congdon e t al. (1986); Ernst an d Barbour (1972); Moll (1973); Sexto n (1959).
2-385 Painted Turtle

Painted T urtle ( Chrysem ys picta )
2-386 P
ainted
Tu
rtle
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight A F 266.5 ± 60.1 SD 83.5 - 450.3 Pennsy lvani a (pic ta x Ernst , 1971b (g) A M 189.1 ± 52.3 SD 102.0 - 274.5 marginata)
A F 326.7 ± 4.95 SE Michi gan Congdon et al., 1986 A M 176.9 ± 1.92 SE J B 64.2 ± 1.59 SE
at hatching 3.7 ± 0.2 3.5 - 3.9 cent ral Virginia (pic ta) Mitc hell, 1985
at hatching 4.1 ± 0.61 SD Iowa Ratterman & Acker man, 1989
Body Lengt h A F 157 ± 2.6 SE 136 - 185 Wisconsi n (bellii ) Moll, 1973 (mm plastron) A M 132 ± 2.9 SE 96 - 155
(mm plastron) A F 125.1 ± 0.64 SE Michi gan Congdon et al., 1986
(mm carapace) A F 134.2 ± 0.81 SE Michi gan Congdon et al., 1986
A M 99.9 ± 0.48 SE J B 65.0 ± 0.65 SE
A M 109.7 ± 0.54 SE J B 71.5 ± 0.69 SE
Egg Weight (g) ini tial mass 6.17 Georgia (dor sal is) Congdon & Gi bbons,
initial mass 6.65 ± 0.67 SD Iowa Ratterman & Acker man, final mass 8.62 ± 1.06 SD 1989
1985
Growth Rate J F - 1 yr 35 mm/yr Quebec, Canada (marginata) Chr istens & Bi der, 1986 J F - 2 to 3 yr 19 - 20 mm/yr J F - 4 to 5 yr 12 mm/yr (measur ed usi ng pl astron) J F - 6 to 7 yr 8 - 10 mm/yr A F - 8 to 12 yr 3 - 6 mm/yr A F - > 12 yr < 3 mm/yr
SD
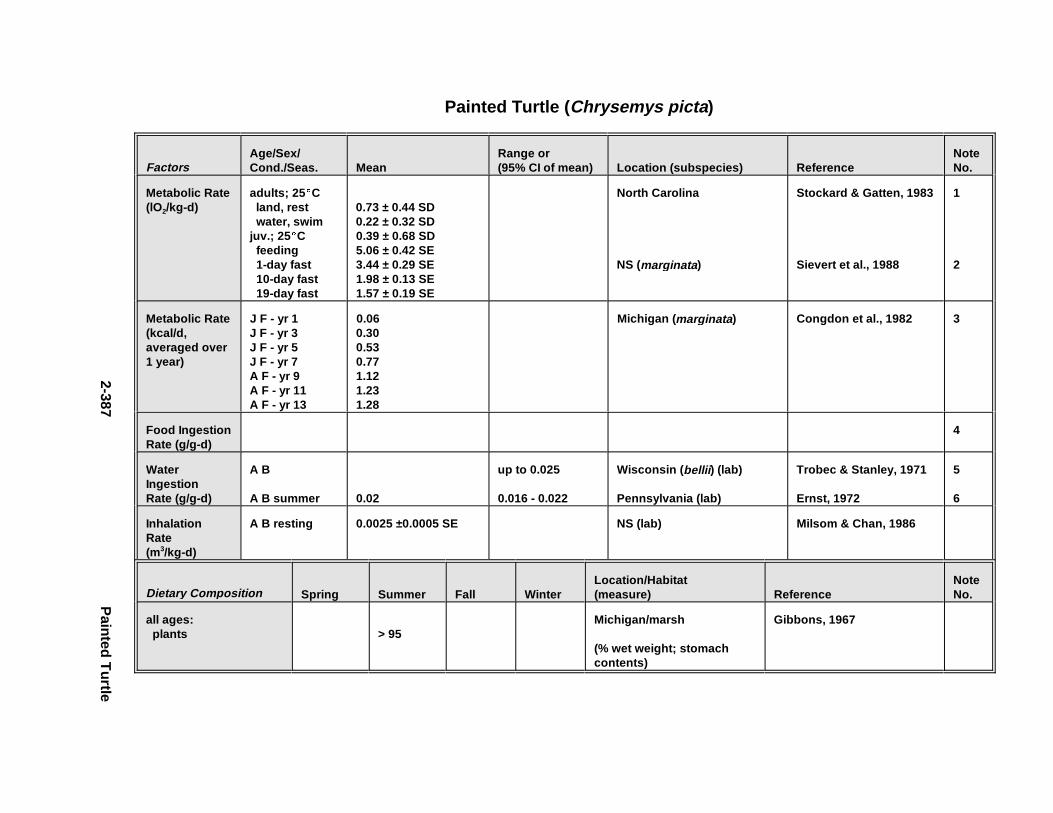
Painted T urtle ( Chrysem ys picta )
2-387 P
ainted
Tu
rtle
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Metabolic Rate adults ; 25��C North Carolina Stockar d & Gatten, 1983 1 (lO /kg-d) land, rest 0.73 ± 0.44 SD2
water, swim 0.22 ± 0.32 SD juv.; 25��C 0.39 ± 0.68 SD feeding 5.06 ± 0.42 SE 1-day fast 3.44 ± 0.29 SE NS (marginata) Sievert et al., 1988 2 10-day fast 1.98 ± 0.13 SE 19-day fast 1.57 ± 0.19 SE
Metabol ic Rate J F - yr 1 0.06 Michigan (marginata) Congdon et al., 1982 3 (kcal/d , J F - yr 3 0.30 averaged over J F - yr 5 0.53 1 year) J F - yr 7 0.77
A F - yr 9 1.12 A F - yr 11 1.23 A F - yr 13 1.28
Food Ingest ion 4 Rate (g/g-d)
Water A B up t o 0.025 Wisconsi n (bellii ) (lab) Trobec & St anley, 1971 5 Ingest ion Rate (g/g-d) A B sum mer 0.02 0.016 - 0.022 Pennsy lvani a (lab) Ernst , 1972 6
Inhal ation A B r est ing 0.0025 ±0.0005 SE NS (lab) Milsom & Chan, 1986 Rate (m /kg-d)3
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
all ages: Michi gan/marsh Gibbons, 1967 plants > 95
(% wet weight ; stomach cont ents)

Painted T urtle ( Chrysem ys picta )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
all ages: Michi gan/pol luted r iver Kni ght & Gibbons, 1968 plants 31.6 38.7 animals 77.3 72.3 (% wet weight ; stomach Oligochaet a - 30.0 cont ents) Cladocer a 1.5 48.5 Odonat a nymphs 60.0 38.3 Lepidopt era larvae 1.0 50.0 Tendipedi dae larva 30.8 7.7 Tendipedi dae pupae 36.7 10.0
detritu s 7.8 1.9
adul ts: snai ls 12.1 Pennsy lvani a (pic ta)/NS Ernst & Bar bour , 1972 amphi pods 3.0 crayfish 7.5 (% wet vol ume; stomach insect s 11.5 cont ents) fish 13.0 other animals 14.1 season not speci fied algae 14.7 vascul ar plants 24.1 other plants 0.8
Population Dynamics
Age/Sex Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Movements A B spr ing 63 - 144 up t o 301 Michi gan (marginata)/NS Sexton, 1959 7 (m) A B sum mer 86 - 91 up t o 300
A B f all 88 - 130 up t o 336
Popul ation B B sum mer 11.1 Saskat chewan, Canada MacCul loch & Secoy , 1983 Densi ty (bellii )/river (N/ha) B B 98 - 410 Michi gan (marginata)/ponds, Sexton, 1959
B B 590 240 - 941 Pennsy lvani a/pond, marsh Ernst , 1971c
B B 828 Michi gan/lake, marsh Frazer et al., 1991
marsh
2-388 P
ainted
Tu
rtle

Painted T urtle ( Chrysem ys picta )
Population Dynamics
Age/Sex Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Clutch Si ze 19.8 17 - 23 Saskat chew an, Canada MacCul loch & Secoy , 1983
10.7 4 - 16 Wisconsi n (bellii )/NS Moll, 1973
7.6 2 - 11 Michi gan (marginata)/NS Congdon & Ti nkl e, 1982
4.8 2 - 9 Tennessee ( dor sal is × Moll, 1973
(bellii )/creek
marginata)/NS
Clutches/ Year 1 - 2 2 Ontario, Canada/NS Schw arzkopf & Br ooks,
1 - 2 2 Michi gan (bellii × marginata) Snow , 1980
> 2 3 Illinoi s (bellii × marginata) Moll, 1973
> 3 5 Tennessee, Loui siana Moll, 1973
/NS
/ket tle ponds
(dor sal is and d. × marginata)/NS
1986
Days 65 - 80 se Pennsy lvani a/NS Ernst , 1971c Incubat ion 60 - 65 se Wisconsi n/NS (natural) Ewert, 1979
72 - 99 nw Minnesot a/NS (natural) Ewert, 1979
Age at Sexual F 5 - 6 New Mexico (bellii )/NS Chris tiansen & Moll, 1973 Maturity (yr) M 3
F 8 Wisconsin (bellii )/NS Chris tiansen & Moll, 1973 M 4
F 6 Pennsylvania (pic ta)/NS Ernst & Bar bour , 1972 M 5
F 4 - 5 Tennessee ( dor sal is x Moll, 1973 M 2 - 3 marginata)/NS
2-389 P
ainted
Tu
rtle

Painted T urtle ( Chrysem ys picta )
Population Dynamics
Age/Sex Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Lengt h at M 90 nor thern Michi gan Cagle, 1954 Sexual F 120 - 130 (marginata, dor sal is)/NS Maturity (mm plastron) M 70 sout hern Illinoi s (marginata, Cagle, 1954
F 120 - 125 dor sal is)/NS
M 123 88 - 170 New Mexico (bellii )/NS Chris tiansen & Moll, 1973 F 150 132 - 205
Annual A F 0 - 14 Saskat chewan, Canada, MI, Zwei fel, 1989 8 Mortality Rates A M 2 - 46 NY, NE/NS (%)
A B 4 - 6 Virg inia/NS Mitchell, 1988 8 J B 54
Longevi ty M up t o 31 yrs Michigan/marsh Frazer et al., 1991 F up t o 34 yrs
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating late Apr il mid-June se Pennsy lvani a Ernst , 1971c March Apr il - early May May Michi gan Gibbons, 1968a
October Ohio Gis t et al., 1990
Nest ing June Jul y se Pennsy lvani a Ernst , 1971c June Jul y Illinoi s, Kansas Smith, 1956, 1961 late May June late June se Michigan (marginata) Tinkle et al., 1981
Hatching late sum mer se Michi gan (marginata) Tinkle et al., 1981 September spr ing Illinoi s (marginata) Cahn, 1937 9 August September Kansas ( bellii ) Smith, 1956
Hibernation late October late March se Michi gan (marginata) Congdon et al., 1982 late October Apr il Kansas ( bellii ) Smith, 1956
2-390 P
ainted
Tu
rtle

Painted T urtle ( Chrysem ys picta )
1 Average mass of test animals rest ing on l and and i n water = 215 g (79 to 395 g) and of test animals sw imming and m easur ed for existence metabol ism = 143 g (79 to 297 g).
2 Average weight of juveni le turtles = 7.7 g. 3 Based on an annual energy budget est imated by the author s assum ing t hat females lay one cl utch of eggs per year after their sevent h year. 4 See Chapt ers 3 and 4 f or appr oaches t o est imating f ood i ngest ion r ates f rom metabol ic rate and di et. 5 Uptake of water by turtles hel d in tap water. 6 Measur ed as evapor ative water loss. 7 Spr ing: from hibernation t o other ponds; sum mer: back t o hibernation ponds; fall: to deep-water areas for hibernation. 8 Cited in Frazer et al., 1991. 9 Cited in Smith, 1961.
2-391 P
ainted
Tu
rtle

References (including A ppendix)
Bayless, L. E. (1975) Popula tion pa rameter s fo r Chrysemys p icta in a New York pond. Am. Midl. Nat. 93: 168-176.
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York, NY: Alfred A. Knopf , Inc .
Blanchard, F. N. (1923) The amphibia ns and r ept iles of Dickins on Count y, Iowa. Univ . Iowa Stud. Nat. Hist. 10: 19-26.
Brecken ridge , W. J. (1944) Repti les and amphibia ns of Minne sota. Minne apolis , MN: Univ ersi ty of Minne sota Press.
Breitenba ch, G. L.; Congdon, J. D.; van Lobe n Sels, R. C. (1984) Winter temp eratures of Chrysemys p icta nests i n Michiga n: effects o n hatchling s urviv al. Herpetologic a 40: 76-81.
Bury, R. B. (1979) Popula tion e cology of fr eshwater turtles. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 571-602.
Cagle, F. R. (1954) Observat ions on t he lif e cycl es of painted turtles (Genus Chrysemys ). Am. Midl. Nat. 52: 225-235.
Cahn, A. R. (1937) The turtles of Illinois . Illinois Biol. Monogr . 1-218.
Christen s, E.; Bide r, J. R. (1986) Reproduc tive ecology of th e paint ed turtle (Chrysemys picta mar ginata) in southwester n Quebec. Can. J. Zool. 64: 914-920.
Chr is tiansen, J. L.; Moll, E. O. (1973) Lati tudina l reproduc tive var iation w ithin a single subspecies of painted turtle, Chrysemys p icta b ellii . Herpetologic a 29: 152-163.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns - easter n and central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
Congdon, J. D.; Gatten , R. E., Jr . (1989) Movemen ts an d energetics o f nest ing Chrysemys picta . Herpetologic a 45: 94-100.
Congdon, J. D.; Gibbons , J. W. (1985) Egg compone nts and r eproduc tive character isti cs o f turtles: relations hips to body size . Herpetologic a 41: 194-205.
Congdon, J. D.; Tink le, D. W. (1982) Reproducti ve energetics o f painted turtle (Chrysemys picta). Herpetologic a 38: 228-237.
2-392 Painted Turtle

Congdon, J. D.; Dunha m, A. E.; Tink le, D. W. (1982) Energy budgets an d life histo ries of rept iles. In: Gans, C., ed. Biology of th e rept ilia : v. 13. New York, NY: Academic Press; pp. 233-271.
Congdon, J. D.; Greene, J. L.; Gibbons , J. W. (1986) Biomass o f fr eshwater turtles: a geogr aphic co mparison. Am. Midl. Nat. 115: 165-173.
DeGraaf, R. M.; Rudis , D. D. (1983) Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Ernst, C. H. (1971a) Sexual cycl es and matu rity o f th e turtle, Chrysemys p icta. Biol. Bull. 140: 191-200.
Ernst, C. H. (1971b) Growth of th e painted turtle, Chrysemys p icta, in southeaster n Penns ylvania . Herpetologic a 27: 135-141.
Ernst, C. J. (1971c) Popula tion dy namics and acti vi ty cycl es of Chrysemys p icta in southeaster n Pennsyl vania. J. Herpetol. 5: 151-160.
Ernst, C. H. (1972) Temperature-acti vi ty r elations hip in t he paint ed turtle , Chrysemys p icta. Copeia 1972: 217-222.
Ernst, C. H.; Barbour, R. W. (1972) Turtles of th e United States. Lexingt on, KY: Univ ersi ty Press o f Kentucky.
Ewer t, M. A. (1979) The embryo and its eg g: development and natural histo ry. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John Wiley and Sons, Inc.; pp. 333-413.
Frazer, N. B.; Gibbons , J. W.; Greene, J. L. (1991) Growth , survivorship a nd longe vi ty of painted turtles Chrysemys p icta in a southwester n Michiga n marsh. Am. Midl. Nat. 125: 245-258.
Gemmel l, D. J. (1970) Some observat ions on t he nesting of th e wester n painted turtle, Chrysemys p icta b elli, in nor thern Minne sota. Can. Field-Nat. 84: 308-309.
Gibbons , J. W. (1967) Variation in gr owth rates i n three popula tions of th e paint ed turtle , Chrysemys p icta. Herpetologic a 23: 296-303.
Gibbons , J. W. (1968a) Reproducti ve potential, acti vi ty an d cycl es in Chrysemys p icta. Ecology 49: 399-409.
Gibbons , J. W. (1968b) Popula tion s tructure and s urviv orship in t he paint ed turtle , Chrysemys p icta. Copeia 2: 260-268.
Gibbons , J. W. (1987) Why do turtles l ive so long? BioS ci . 37: 262-269.
2-393 Painted Turtle

Gibbons , J. W.; Nelson, D. H. (1978) The evolut iona ry signif icance of delayed emergence from th e nest b y hatchling t urtles. Evolut ion 3 2: 297-303.
Gist, D. H.; Michaelson, J. A.; Jones, J. M. (1990) Autumn mat ing in t he paint ed turtle , Chrysemys p icta. Herpetologic a 46: 331-336.
Hall, F. G. (1924) The respiratory exch ange in turtles. J. Metab. Res. 6: 393-401.
Hutchins on, V. H. (1979) Thermoregula tion. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 207-227.
Iverson, J. B. (1982) Biomass i n turtle popula tions : a neglected subject. Oecologia (B erl.) 55: 69-76.
Knight , A. W.; Gibbons , J. W. (1968) Food of th e paint ed turtle , Chrysemys p icta in a pollut ed river . Am. Midl. Nat. 80: 558-562.
Lagler, K. F. (1943) Food ha bit s and econom ic r elations of th e turtles of Michiga n with special reference to game man agement. Am. Midl. Nat. 29: 257-312.
Legler, J. M. (1954) Nest ing ha bit s of th e wester n painted turtle, Chrysemys p icta b elli Gray. Herpetologic a 10: 137-144.
Lynn, W. G.; von Brand, T. (1945) Studie s on t he oxygen cons umpt ion a nd water metabolis m of tu rtle emb ryos . Biol. Bull. 88: 112-125.
MacCulloc h, R. D.; Secoy, D. M. (1983) Demography, growth , and food of wester n painted turtles, Chrysemys p icta b ellii (Gray), from so uthern Saskatch ewan. Can. J. Zool. 61: 1499-1509.
Mahmoud, I. Y.; Kli cka, J. (1979) Feeding, dr ink ing, and excr etion. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 229-243.
Marchand, L. J. (1942) A contr ibut ion t o a know ledge of th e natural his tory of cer tain freshwater turtles [master 's th esis]. Gainesville , FL: Univ ersi ty of Flor ida.
McAuliffe, J. R. (1978) Seasonal migrationa l movemen ts o f a popula tion of th e wester n paint ed turtle (Chrysemys p icta b ellii ) (reptilia , testudine s, testudinid ae). J. Herpetol. 12: 143-149.
Mils om, W. K.; Chan, P. (1986) The relations hip b etween lung v olum e, respir atory dr ive and breathing p atter n in the turtle, Chrysemys p icta. J. Exp . Biol. 120: 233-247.
Mitch ell, J. C. (1985) Female reproducti ve cycl e and life histo ry attr ibutes i n a Virginia popula tion of paint ed turtles, Chrysemys p icta. J. Herpetol. 19: 218-226.
2-394 Painted Turtle

Mitch ell, J. C. (1988) Popula tion e cology and lif e his tories of th e freshwater turtles Chrysemys p icta and Sternotherus odor atus in an urban lake. Herpetol. Monogr . 2: 40-61.
Moll, E. O. (1973) Lati tudina l and int ersubs peci fic var iation in r eproduc tion of th e paint ed turt le, Chrysemys p icta. Herpetologic a 29: 307-318.
Mor loc k, H.; Herringt on, S.; Oldha m, M. (1972) Weight los s dur ing f ood de pr ivation in t he easter n painted turtle, Chrysemys p icta p icta. Copeia 1972: 392-394.
Mount , R. H. (1975) The repti les and amphibia ns of A labama. Aubur n, AL: Aubur n Universi ty Agricultural Experiment Stat ion.
Packar d, G. C.; Packar d, M. J.; Boardman, T. J.; et al . (1983) Influence of water exch anges by fl exible-shelled eggs of painted turtles Chrysemys p icta on metabolis m and growth of emb ryos . Physiol. Zool. 56: 217-230.
Pearse, A. S. (1923) The abunda nce and m igr ation of tu rtles. Ecology 4: 24-28.
Pope, C. H. (1939) Turtles of th e United States an d Canada. New York, NY: Alfred A. Knopf .
Powel l, C. B. (1967) Female sexu al cycl es of Chrysemys p icta and Clemmys i nscu lpta in Nova Sco tia. Can. Field-Nat. 81: 134-140.
Ratter man, R. J.; Acker man, R. A. (1989) The water exch ange and hydric mi crocl imate o f paint ed turtle (Chrysemys p ica) eggs inc ubating in f ield n ests. Physiol. Zool. 62: 1059-1079.
Ream, C. H. (1967) Some asp ects o f th e ecology of paint ed turtles, Lake Mendot a, Wisco nsin [Ph.D. disser tat ion] . Madis on, WI: Univ ersi ty of W iscons in.
Schwarzkopf, L.; Brooks, R. J. (1986) Annua l variations in r eproduc tive character isti cs o f painted turtles (Chrysemys p icta). Can. J. Zool. 64: 1148-1151.
Sexto n, O. J. (1959) Spatial and temp oral movemen ts o f a popula tion of th e paint ed turtle , Chrysemys p icta mar ginata (Agassiz) . Ecol. Monogr . 29: 113-140.
Seymo ur, R. S. (1982) Physiologic al adaptations to aquatic lif e. In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia , phy siology D; phy siologic al ecology : v. 13. New York, NY: Academic Press; pp. 1-51.
Siever t, L. M.; Siever t, G. A.; Cupp, P. V., Jr . (1988) Metabolic r ate of feeding a nd fasti ng juv enile midla nd pa int ed turtles, Chrysemys p icta mar ginata. Comp. Bioc hem. Physiol. A Comp. Physiol. 90: 157-159.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9; 365 pp.
2-395 Painted Turtle

Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28.
Snow, J. E. (1980) Second c lut ch laying by paint ed turtles. Copeia 1980: 534-536.
Snow, J. E. (1982) Predation on pa int ed turtle n ests: nest su rvival as a fu nct ion of n est ag e. Can. J. Zool. 60: 3290-3292.
Stockar d, M. E.; Gatten , R. E. (1983) Acti vi ty meta bolis m of paint ed turtles (Chrysemys picta). Copeia 1983: 214-221.
Taylor , G. M.; Nol, E. (1989) Movemen ts an d hibe rnation s ites o f overwinter ing pa int ed turtles in s out hern Ontar io. Can. J. Zool. 67: 1877-1881.
Tink le, D. W.; Congdon, J. D.; Rosen, P. C. (1981) Nest ing f requency and s uccess: Implic ations fo r th e demogr aphy of paint ed turtles. Ecology 62: 1426-1432.
Trobec, T. N.; Stanley, J. G. (1971) Uptake o f ions and water by th e painted turtle, Chrysemys p icta. Copeia 1971: 537-542.
Wade, S. E.; Giffo rd, C. E. (1965) A preliminary stu dy of th e turtle popula tion of a nor thern India na lake. Proc. India na Acad. Sci . 74: 371-374.
Wilbur , H. M. (1975a) The evolut iona ry and math emati cal demography of th e turtle Chrysemys p icta. Ecology 56: 64-77.
Wilbur , H. M. (1975b) A growth model for the turtle Chrysemys p icta. Copeia 1975: 337-343.
Zweifel , R. G. (1989) Long-ter m ecologic al studie s on a popula tion of paint ed turtles, Chrysemys p icta, on Long Is land, New York. Am. Mus. Novit . 2952: 1-55.
2-396 Painted Turtle

2.3.3. Eastern Box T urtle (box turtles)
Order Testudi nes, Family Emyd idae. Box tu rtles are the most ter restr ial of th e Emydid t urtles, having c lose-fi tting s hells th at have allowed them to adapt wel l to ter restr ial lif e. They are found t hroughout th e easter n and central United States an d into the southwest. They are omnivorous.
Selected species
The easter n box tu rtle (Terrapene carolin a carolin a) ranges fr om northeaster n Massach usetts to Georgia, west to Michiga n, Illinois , and T ennessee (Conant and Collins , 1991). There are four subspecies of T. carolin a, all f ound w ithin t he easter n United States: T. c. carolina (above), T. c. major (Gulf Coast b ox tu rtle; the largest su bspecies, restr icted to the Gulf Coast), T. c. tr iungui s (three-toed box tu rtle; Misso uri to south-central Alabama and Texas), and T. c. baur i (Flor ida box tu rtle ; restr icted to the Flor ida penins ula and keys) (Conant and Collins , 1991).
Body size. The easter n box tu rtle is smal l, wi th adult s ranging f rom 11.5 to 15.2 cm in length (plastr on) and approximatel y 300 to over 400 g. Hatchlings weigh a ppr ox imatel y 8 to 10 g. Turtles cont inue to grow th roughout th eir lives; however , their growth rate sl ows after reaching sexu al matu rity (Er nst an d Barbour, 1972), and growth rings ar e no longe r discer nable after 18 to 20 years (Sti ckel , 1978). Body fat r eserves i n a Georgia popula tion averaged 0.058 to 0.060 g of fat p er gram of lean dry weight fr om sp ring t hrough f all (Brisbin, 1972).
Habitat. Typical box tu rtle habitats i nclude open woodla nds , thi ckets, and wel l-drained but mo ist fo rested areas (Sti ckel , 1950), but occas iona lly p astu res and marshy meadow s are ut ilize d (Ernst and Ba rbour , 1972). In areas wi th mixed woodla nds an d grassl ands, box tu rtles use grassl and areas in times o f mo derate temp eratures and peak mois ture condit ions ; otherwis e, they tend t o use the more mois t forested habitats (R eagan, 1974). Many tu rtles are kille d attemp ting t o cross roads, and f ragmentation of habi tat b y roads can sever ely reduce popula tions (D eGraaf and Rudis , 1983; Stickel , 1978).
Food ha bi ts. Adult T. carolin a are omnivorous (Ernst an d Barbour, 1972). When young, they are pr imarily car niv orous , but th ey become more herbiv orous as they age and as growth slows (Er nst an d Barbour, 1972). They co nsume a wi de var iety o f animal mater ial, including e arthworms, slugs , snails , insects an d their larvae (particularly grasshoppe rs, mo ths, and b eetles), crayfi sh, frogs, toads, snakes, and car rion; they also consume veg etable matter , including l eaves, grass, berries, fruits, and fungi ( DeGraaf and Rudis , 1983). A high pr opor tion of sn ails and s lugs may co mprise th e animal matter in the diet (Barbour, 1950), and seeds can become an important co mpone nt of th e plant mater ials in the late su mmer and fal l (Klimstr a and Newso me, 1960).
Temperature regulation and daily acti vi ties. The species is diur nal and s pends th e night r esting in a scoope d depression or form that the turtle digs in t he soil w ith its front feet (Er nst an d Barbour, 1972; Stickel , 1950). T. carolin a are most acti ve in temp erate,
2-397 Easter n Box Turtle

�
�
�
humid weath er (Stickel , 1950). In the summer , they avoid high temp eratures during m idda y by r esting unde r logs or leaf l itter , in mammal burrows, or by congr egating in m udhole s (Smith, 1961; Stickel , 1950). In the hottest weath er, they may en ter shaded shallow pools for hour s or days (Er nst an d Barbour, 1972). In the coole r temp eratures, they may r estr ict their fo raging acti vi ties to midday (Sti ckel , 1950). In the laboratory, locomotion is maxi mal between 24 and 32�C (Adams et al ., 1989). In the field, their mean acti ve body temp erature is approximatel y 26�C (Brattstr om, 1965, ci ted in Hutchins on, 1979).
Hibernation. In the northern parts o f i ts r ange (northeaster n Massach usetts, Michiga n, Illinois ), the easter n box tu rtle enters hibe rnation in l ate Octo ber or Novemb er and emerges in April. In Louis iana, Penn and Potth arst (1940, ci ted in Ernst an d Barbour , 1972) found t hat T. c. major hibe rnated when temp eratures fel l below 65�F. To hibe rnate, the box tu rtle bur rows int o loos e soil a nd de br is or mud of ponds or str eam botto ms. Congdon e t al. (1989) found a Sout h Carolina popula tion of box tu rtles to occupy r elatively shallow bur rows (less th an 4 cm) co mpared wi th those occupie d by box tu rtles in c olde r regions (up t o 46 cm). Dolbe er (1971) found hibe rnacula of box tu rtles in T ennessee to be unde r 15.5 cm of l eaf l itter and 5.8 cm o f soil on average. In southern states, during r ainy and w arm periods , box tu rtles may b ecome acti ve again (Dolbe er, 1971). In Flor ida, the box tu rtle may b e acti ve al l year (Ernst an d Barbour, 1972).
Breeding acti vi ties and social organization. Box tu rtles are solitary excep t briefly dur ing t he mating seaso n. Indiv idua ls r estr ict th eir acti vi ties to a foraging hom e range, but home ranges of differ ent indiv idua ls can overlap substan tially (Sti ckel , 1950). Mating usually occurs in t he spring but may co ntinue int o fall, and eggs ar e laid in l ate sp ring a nd summer (Ernst an d Barbour, 1972). The femal e digs a 3- to 4-inc h cavi ty i n sandy or loamy soil in w hic h she depos its her eggs and t hen covers the nest wi th soil. Nests ten d to be constr ucted sever al hundr ed meter s fr om th e femal e's fo raging hom e range in t he warmer and dr ier upla nds (Stickel , 1989). The duration of inc ubation de pends on s oil temp eratures, and someti mes h atchlings overwint er in t he nest. The young a re semi aquatic but se ldom seen (Smith, 1956).
Home range and resources. Measures of th e foraging hom e range fo r box tu rtles range fr om .5 ha to jus t ov er 5 ha (Dolbe er, 1969; Schwar tz et al ., 1984). A femal e may need to search for suitable nest si te (e.g., slight ly el evated sandy soils) (Ernst an d Barbour, 1972) outsi de of her foraging hom e range (Stickel , 1950). Winter hibe rnacula ten d to be wi thin t he foraging hom e range (Stickel , 1989).
Popul ation de nsity. Popula tion de nsity var ies wi th habitat q uality, but st udie s link ing de nsity to partic ular habitat ch aracter isti cs ar e lacking. In some areas, popula tion densi ties have decline d steadily over th e past sever al decades (Sch war tz and Schwar tz, 1974; Stickel , 1978). Some investi gators attr ibut e the decline to inc reasing ha bit at fragmentation a nd ob stacl es (e.g., highways) th at prevent femal es fr om reaching or returning f rom appr opr iate nest ing a reas (Sti ckel , 1978; DeGraaf and Rudis , 1983).
Popul ation dy namics. Sexual matu rity i s attai ned at about 4 or 5 years (Ernst an d Barbour, 1972) to 5 to 10 years of age (Mint on, 1972, ci ted in DeGraaf and Rudis , 1983). One to four clutch es may b e laid per year , depending on la ti tude (Oliv er, 1955, ci ted in
2-398 Easter n Box Turtle

�
Moll, 1979; Smith, 1961). Clut ch size r anges from th ree to eight eggs , averaging t hree to four in some areas (Congdon a nd Gibbons , 1985; Ernst an d Barbour, 1972; Smith, 1956). Juvenile s generally co mprise a small pr opor tion of box tu rtle popula tions , for examp le, 18 to 25 percent in one popula tion in Mis sour i (Schwar tz and Schwar tz, 1974) and 10 percent in a stu dy in Maryland (Stickel , 1950). Some indiv idua l box tu rtles may l ive over 100 years (Graham and Hut chins on, 1969, ci ted in DeGraaf and Rudis , 1983; Oliver , 1955, ci ted in Auffen berg and Iverson, 1979).
Simi lar species (fr om general references)
� The ornate box tu rtle (Terrapene ornata ornata) and the deser t box tu rtle (Terrapene ornata luteola) are simi lar in size and habits to the easter n box turtle. They occu r in the wester n, midwester n, and southern midwester n states. Prefer red habitats i nclude open pr airies, pastu reland, open woodla nds , and water ways i n arid, sandy-soil t errains . The ornate box tu rtle and deser t box tu rtle forage primar ily on insects b ut also on berries and car rion.
General references
Behler and King (1979); Conant and Collins (1991); DeGraaf and Rudis (1983); Ernst and Ba rbour (1972); Stickel (1950).
2-399 Easter n Box Turtle

Eastern Box T urtle ( Terrapene carolina )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight A F fall 381 ± 29 SE Georgia (carol ina), capt ive Br isbin, 1972 (g) A M fall 398 ± 47 SE
A F spr ing 388 ± 29 SE Georgia (carol ina) Brisbi n, 1972 A M spr ing 369 ± 47 SE
A F 372 Sout h Carolina Congdon & Gi bbons, 1985
at hatching 8.8 Florida (major) Ewert, 1979
2 mont hs 21 Tennessee Al lard, 1948 1 1.3 years 40 3.3 years 54
8.4 Indi ana (carol ina) Ewert, 1979
Body Fat B fall 0.058 ± 0.014 SE Georgia (carol ina), capt ive Br isbin, 1972 (g/g lean dr y B spr ing 0.060 ± 0.016 SE weight ) B sum mer 0.059 ± 0.006 SE
Lengt h A F 129 mm plastron Sout h Carolina Congdon & Gi bbons, 1985 A up t o 198 mm NS/NS Oliver, 1955 2 at hatching 28 mm carapace carapace NS/NS Oliver, 1955 2
Egg Weight (g) 9.02 ± 0.17 SE Sout h Carolina Congdon & Gi bbons, 1985 6 - 11 NS/NS Ernst & Bar bour , 1972
Metabol ic Rate A F basal 5.4 est imated 3 (kcal /kg-d)
Food Ingest ion 4 Rate (g/g-d)
2-400 E
astern B
ox T
urtle

Eastern Box T urtle ( Terrapene carolina )
Dietary Compositio n Spr ing Summer Fall Winter Habi tat (measur e) Reference No. Locat ion (subspeci es)/ Note
snai ls 60 Kent ucky (carol ina)/ Barbour , 1950 crayfish 15 Cumberland Mount ains plants 12.5 cr icket s 7.5 (% vol ume; stomach uni dent ified seeds 5 cont ents)
plant matte r 35 39 20 Illin ois (carol ina)/forest , Klimstra & Newsome, 1960 insect s (adul ts) 18 12 12 prairie insect s (larvae) 4 5 9 seeds 8 16 33 (% wet vol ume; digest ive Gastropoda 18 6 8 tract) Isopoda <1 5 3 Diplopoda 3 2 5 Decapoda 2 2 0 Annel ida 1 1 4 mammals 2 <1 2 reptiles 1 3 1 bi rds 3 1 <1
Population Dynamics
Age/Sex/ Range or Locat ion Note Cond. /Seas. Mean (95% CI of mean) (subspeci es)/Habi tat Reference No.
Home Range sum mer 0.46 Tennessee ( carol ina)/ Dolbeer, 1969 5 Size (ha) woodl and
B M 1.2 Maryland (carol ina)/ Stickel , 1989 5 B F 1.1 botto mland fo rest
B M 5.2 Missouri (triu nguis)/mixed Schwartz et al., 1984 5 B F 5.1 woods, fields
Popul ation 2.8 - 3.6 Tennessee/ woodl and Dolbeer, 1969 Densi ty (N/ha) 17 - 35 Maryland ( triu nguis)/fo rest Schwartz et al., 1984
2-401 E
astern B
ox T
urtle

�
Eastern Box T urtle ( Terrapene carolina )
Population Dynamics
Age/Sex/ Range or Locat ion Note Cond. /Seas. Mean (95% CI of mean) (subspeci es)/Habi tat Reference No.
Clutch Si ze 3.4 ± 0.3 SE Sout h Carolina/NS Congdon & Gi bbons, 1985 4 2 - 7 Washi ngt on, DC/NS Smith, 1956
Clutches/ Year up t o 4 Florida/NS Oliver, 1955 6 1 Illinoi s/NS Smith, 1961
Days 78 - 102 nor thwest Ewert, 1979 Incubat ion 99 69 - 161 Minnesot a/(natural) Ewing, 1933 7
Washi ngt on, DC/(natural)
Age at Sexual B 4 - 5 NS/NS Ernst & Bar bour , 1972 Maturity (yr)
B 5 - 10 NS/NS Minton, 1972 8
Lengt h at B 100 - 130 NS/NS Oliver, 1955 2 Sexual Maturity (mm carapace)
Longevi ty (yr) 20 up t o 80 NS/NS Nichol s, 1939a 8 up t o 138 capt ivi ty Oliver, 1955 2
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating spr ing nor thern range Ernst & Bar bour , 1972 June Jul y ne Carolinas, Washi ngt on, DeGraaf & Rudi s, 1983;
DC Smith, 1956
Hatching September October nor thern range Ernst & Bar bour , 1972 August September ne Carolinas DeGraaf & Rudi s, 1983
Hibernation Novem ber Apr il nor thern range Ernst & Bar bour , 1972 October Apr il Missour i (triu nguis) Schw artz & Schw artz, 1974
2-402 E
astern B
ox T
urtle
1 Cited in Ernst and Bar bour (1972). 2 Cited in Auf fenber g and Iverson ( 1979). 3 Est imated assum ing t emperature of 20�C, using Equatio n 3-50 (Robinson et al., 1983) and body weight s of Br isbin (1972) after subt ract ing
30 percent of the body weight to eliminate the weight of the shel l (Hall, 1924). 4 See Chapt ers 3 and 4 f or methods of est imating i ngest ion r ates f rom metabol ic rate and di et.

Eastern Box T urtle ( Terrapene carolina )
5 Foraging hom e range; nest si tes can be sever al hundr ed meters away from the foraging hom e range.6 Cited in Moll (1979).7 Cited in Ewert (1979).8 Cited in DeGraaf and Rudi s (1983).
2-403 E
astern B
ox T
urtle

References (including A ppendix)
Adams, N. A.; Claussen , D. L.; Skillings , J. (1989) Effects o f temp erature on volunt ary loc omot ion of th e easter n box tu rtle, Terrapene carolin a carolin a. Copeia 4: 905-915.
Allard, H. A. (1935) The natural histo ry of th e box tu rtle. Sci . Mont hly 41: 325-338.
Allard, H. A. (1948) The easter n box-tu rtle and its b ehavior, part 1. J. Tenn. Acad. Sci . 23: 307-321.
Auffen berg, W.; Iverson, J. G. (1979) Demography of ter restr ial turtles. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons; pp. 541-569.
Barbour, R. W. (1950) The repti les of Big Bl ack Mount ain, Harlan Count y, Kentucky. Copeia 1950: 100-107.
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York, NY: Alfred A. Knopf , Inc .
Brattstr om, B. H. (1965) Body temp eratures of repti les. Am. Midl. Nat. 73: 376-422.
Breder, R. B. (1927) Turtle trailing: a new technique fo r study ing t he lif e habit s of cer tain testudinata. Zoologic a 9: 231-243.
Br isbin, I. L., Jr . (1972) Seasonal var iations in t he liv e weight s and m ajor body co mpone nts of cap tive box tu rtles. Herpetologic a 28: 70-75.
Bush, F. M. (1959) Foods of so me Kentucky h erpti les. Herpetologic a 15: 73-77.
Cahn, A. R. (1937) The turtles of Illinois . Illinois Biol. Monogr . 1-218.
Carr, A . F. (1952) Handbook of tu rtles. Ithaca, NY: Comsto ck.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns: easter n/central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
Congdon, J. D.; Gibbons , J. W. (1985) Egg compone nts and r eproduc tive character isti cs o f turtles: relations hips to body size . Herpetologic a 41: 194-205.
Congdon, J. D.; Gatten , R. E., Jr .; Morreale, S. J. (1989) Overwinter ing acti vi ty o f box tu rtles (Terrapene carol ina) in Sout h Carolina . J. Herpetol. 23: 179-181.
DeGraaf, R. M.; Rudis , D. D. (1983) Box tu rtle. Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Dickson, J. D., III (1953) The private l ife of th e box tu rtle. Everglades Nat. Hist. 1: 58-62.
2-404 Easter n Box Turtle

Dodge , C. H.; Dimond, M. T.; Wunde r, C. C. (1979) The influence of temp erature on the inc ubation of box tu rtle eggs (ab str act only ). Am. Zool. 19: 981.
Dolbe er, R. A. (1969) Popula tion de nsity and hom e range size of th e easter n box tu rtle (Terrapene c. carolin a) in easter n Tennessee. A.S.B. Bull. 16: 49.
Dolbe er, R. A. (1971) Winter behavior of th e easter n box tu rtle, Terrapene c. carolin a L., in easter n Tennessee. Copeia 1971: 758-760.
Ernst, C. H.; Barbour, R. W. (1972) Turtles of th e United States. Lexingt on, KY: Univ ersi ty Press o f Kentucky.
Ewer t, M. A. (1979) The embryo and its eg g: development and natural histo ry. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John Wiley and Sons, Inc.; pp. 333-413.
Ewing, H. E. (1933) Reproduc tion in t he easter n box-tu rtle Terrapene carolin a carolin a. Copeia 1933: 95-96.
Graham, T. E.; Hutchins on, V. H. (1969) Centenarian box tu rtles. Int. Turtle Tortoise Soc. J. 3: 24-29.
Hall, F. G. (1924) The respiratory exch ange in turtles. J. Metab. Res. 6: 393-401.
Hutchins on, V. H. (1979) Thermoregula tion. In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 207-227.
Klimstr a, W. D.; Newso me, F. (1960) Some observat ions on t he food c oactions of th e common box tu rtle , Terrapene c. carolin a. Ecology 41: 639-647.
Lynn, W. G.; von Brand, T. (1945) Studie s on t he oxygen cons umpt ion a nd water metabolis m of tu rtle emb ryos . Biol. Bull. 88: 112-125.
Mint on, S. A., Jr . (1972) Amphibia ns and r ept iles of India na. India napolis , IN: India na Academy o f Sci ence.
Moll, E. O. (1979) Reproducti ve cycl es and adaptat ions . In: Harless, M.; Morlock, H., eds. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 305-331.
Nichols , J. T. (1939a) Data on size, growth and age in the box tu rtle, Terrapene carol ina. Copeia 1939: 14-20.
Nichols , J. T. (1939b) Range and homing of indiv idua l box tu rtles. Copeia 1939: 125-127.
Oliver , J. A. (1955) The natural histo ry of North Amer ican amphibia ns and r ept iles. Princ eton, NJ: Van Nostrand Co.
2-405 Easter n Box Turtle

Penn, G. H.; Potth arst, K. E. (1940) The reproduc tion a nd dor mancy of Terrapene major in New Orleans. Herpetologic a 2: 25-29.
Reagan, D. P. (1974) Habitat sel ect ion in t he three-toed box tu rtle Terrapene carol ina tr iungui s. Copeia 2: 512-527.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Rosenberger, R. C. (1972) Interest ing facts a bout tu rtles. Int . Turtle T ortois e Soc. J. 6: 4-7.
Schwar tz, C. W.; Schwar tz, E. R. (1974) The three-toed box tu rtle in central Misso uri: its popula tion, hom e range, and m ovemen ts. Misso uri Dept. Conserv. Terr. Ser. No. 5; 28 pp.
Schwar tz, E. R.; Schwar tz, C. W.; Kiester , A. R. (1984) The three-toed box tu rtle in central Misso uri, part II: a nineteen-year stu dy of home range, movemen ts an d popula tion. Misso uri Dept. Conserv. Terr. Ser. No. 12; 29 pp.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28: 118-120.
Stickel , L. F. (1950) Popula tion a nd hom e range r elations hips of th e box tu rtle , Terrapene c. carolina (Linna eus). Ecol. Monogr . 20: 351-378.
Stickel , L. F. (1978) Changes in a box tu rtle popula tion dur ing t hree decades. Copeia 1978: 221-225.
Stickel , L. F. (1989) Home range behavior among box tu rtles (Terrapene c. carolin a) of a botto mland forest i n Maryland. J. Herpetol. 23: 40-44.
Stickel , L. F.; Bunck, C. M. (1989) Growth and morphometr ics o f th e box tu rtle, Terrapene c. carolina. J. Herpetol. 23: 216-223.
Strang, C. A. (1983) Spatial and temp oral acti vi ty p atter ns in two ter restr ial turtles. J. Herpetol. 17: 43-47.
2-406 Easter n Box Turtle

2.3.4. Racer (and w hipsnakes)
Order Squamata, Family Colubrid ae. All racer snakes (Coluber constric tor) and whips nakes (Masti cophi s) belong t o the fami ly Colubrid ae, along w ith 84 percent of th e snake sp ecies in North Amer ica. Colubr ids var y wide ly in f orm and s ize and c an be found in numerous ter restr ial and aquatic habitats. The more ter restr ial memb ers of th is fami ly also inc lude so me br own and ga rter sn akes; line d snakes; earth snakes; hognos e snakes; small w oodla nd s nakes; green snakes; speckl ed racer and indigo s nakes; rat sn akes; glossy sn akes; pine , bull, and gophe r sn akes; kings nakes an d mi lk sn akes; scar let, long-nosed, and short-tai led snakes; ground s nakes; rear-fanged snakes; and crowned and black-h eaded snakes (Conant and Collins , 1991).
Selected species
Racer snakes (Coluber constric tor) are slender and fast mo ving a nd are found in a wide var iety o f ter restr ial habitats. They are one of th e most co mmo n large snakes i n North Amer ica (Smi th, 1961). There are 11 subspecies in North Amer ica, limi ted to the United States an d Mexico: C. c. constr icto r (northern black r acer ; southern Maine to northeaster n Alabama), C. c. flaviventr is (easter n yellow belly r acer ; Montana, wester n North Dakota, and Iowa so uth to Texas), C. c. foxi i (blue r acer ; northwest O hio t o easter n Iowa and southeast Minne sota), C. c. anthicus (butter mi lk racer ; south Arkansas, Louis iana, and east T exas), C. c. etheridgei (tan racer ; west-cen tral Louis iana and a djacent Texas), C. c. helv igularis (brownchin r acer ; lower Chipola and A palachic ola River Vall eys i n Flor ida panhandle and a djacent Georgia), C. c. latrunc ulus (blackmask r acer ; southeast Louis iana along east si de of Missi ss ippi Riv er to nor thern Mis sissippi) , C. c. mormon (wester n yellow -bellie d racer ; south British Colom bia to Baja Calif ornia , east to southwest Montana, wester n Wyoming, and wester n Color ado) , C. c. oaxaca (Mexican racer ; south Texas an d Mexico), C. c. paludicola (Everglades racer ; southern Flor ida Everglades region and Cape Canaveral area), and C. c. priapus (southern black r acer ; southeaster n states and north and west i n Missi ss ippi V alley).
Body size. Adult r acer snakes ar e usually 76 to 152 cm i n total length (Conant an d Collins , 1991). Brown and Parker (1984) develope d an empir ical relations hip b etween snout-to -vent length (SVL)i and body weight fo r mal e and femal e racers of th e mormon subspecies in northern Utah:
weight (g ) = -100.80 + 2.93 SVL (cm) femal es,j and
weight (g ) = -82.65 + 2.57 SVL (cm) males.
The equations apply only over a lim ited range of body size s (40 to 70 cm) wh ere the relations hip is appr ox imatel y linear instead of expone nt ial. Kaufman and Gibbons (1975)
iMeasures o f SVL excl ude the tail. Fitch (1963) esti mated that th e tai l measures 28 p ercen t o f the SVL of young f emales and 31 percent of the SVL of young m ales.
jFemales collected when nonre produc tive.
2-407 Racer

�
determined a relations hip b etween length and weight fo r bot h sexes o f a South Carolina popula tion:
weight (g ) = 0.0003 SVL (cm)2.97 (± 0.15 2SE) both sexes. k
Racers fr om popula tions in t he nor theaster n United States ten d to be the largest, while those fr om th e far west an d south Texas ar e the smal lest (Fitch , 1963). Just p rior to egg-laying, the eggs can account fo r over 40 percent of a gr avid femal e's body weight (B rown and Parker , 1984). At hatching, racers weigh a bout 8 or 9 g. Weight gain dur ing t he fi rst year is rapid, wi th bot h sexes i ncreasing t heir weight after hatching by appr ox imatel y 3.2 times i n the fi rst year (Brown and Parker , 1984). One-year -old femal es nearly double th eir weight dur ing t heir second year (Brown and Parker , 1984). By th e time femal es are 3 years old ( when most reach sexu al matu rity), they are 1.3 times h eavier than the males (Brown and Parker , 1984).
Habitat . Racers can be found in m ois t or dry areas, abandone d f ields , open woodla nds , mount ain meadows, rocky w oode d hills ides, grassy-b ordered str eams, pine flatwoods , roadside s, and m arshes from sea level to 2,150 m in elevat ion ( Behler and King, 1979). Racers are partially arboreal (Behler and King, 1979; DeGraaf and Rudis , 1983). C. c. constr icto r seems to prefer forest ed ges and open grassy, shrubby ar eas (Fitch , 1963, 1982). In autumn, most C. constr icto r move int o woodla nds to find r ock crevices i n which to overwinter (Fitch , 1982).
Food ha bi ts . Racers are foraging ge neralists th at acti vely seek th eir prey. Their var ied diet includes smal l mammal s (e.g., mice, voles), insects, amphibia ns (especially frogs), smal l birds, birds' eggs, snakes, and lizards (Brown and Parker , 1982; Fitch , 1963; Klimstr a, 1959). In early sp ring, C.c. flaviventr is feeds primar ily on mammal s and from May to Octo ber feeds primar ily on insects ( Klimstr a, 1959). They often capture new prey before fully dig esting pr evious ly cap tured pr ey (Fitch, 1982). Females, which are larger than males, tend t o cons ume a highe r propor tion of ver tebrate prey th an do the males (Fitch , 1982). Males ten d to spend more time cl imbing a mong f olia ge in low sh rubs an d trees and consuming in sects (F itch , 1982).
Temperature regulation and daily acti vi ties. C. constr icto r is diur nal and s pends a good por tion of th e daylight hour s foraging (Vermersch and Kunt z, 1986). The species is fast mo ving a nd may b e encountered in almost an y ter restr ial si tuation ( Fitch, 1982). Hammer son (1987) observed California racers to bask i n the sun after emerging f rom th eir night bur rows or crevices u nti l their internal body temp erature reached almost 34�C, after whic h they would be gin acti vely fo raging. When temp eratures are moderate, racers wi ll spend m uch of th eir ti me dur ing t he day in t he open abov e ground; at high temp eratures, racers may r etreat unde rground ( Brown and Parker , 1982). Although r acers are good clim bers, th ey spend m ost of th eir ti me on t he ground ( Behler and King, 1979). When searching f or food or being pur sued, the racer snake will not hesi tate to cl imb or swi m (Smith, 1961).
k95 percen t co nfidence interval for co nstant (intercep t in log-tran sfo rm regressi on) = 0.00015 to 0.00058.
2-408 Racer

�
�
�
Hibernation. In fal l, racers mo ve to their hibe rnacula fai rly dir ectl y and begin hibe rnation s oon t hereafter (Brown and Parker , 1982; Fitch , 1963). Racers hibe rnate in congr egations of ten s to hundr eds of sn akes (B rown and Parker , 1984), someti mes wi th coppe rheads and r attl esnakes, often using de ep rock crevices o r abandone d woodc huc k holes (Parker and Brown, 1973). They are among t he earliest sn akes to emerge from hibe rnation ( DeGraaf and Rudis , 1983).
Breeding acti vi ties and social organization. The species br eeds in t he spring or early su mmer . Racers defend home ter ritories (DeGraaf and Rudis , 1983; Smith, 1956). Eggs are laid in the summer in rott ing w ood, stu mps, decaying v egetable matter , or loos e soil a nd h atch about 2 mont hs l ater (Behler and King, 1979; DeGraaf and Rudis , 1983). More than one male may mate wi th one femal e in a breeding seaso n. Eggs may double in size before hatching by ab sorbing water from th e surrounding s oil ( Fitch, 1963).
Home range and r esour ces. C. c. constr icto r appears to have a definit e hom e range (Smith, 1956) and requir es large tracts o f mi xed old f ields and w oodla nds (M . Klemens, pers. comm. , ci ted in DeGraaf and Rudis , 1983). Fitch (1963) descr ibed four typ es of movemen t depending on t he seaso n and acti vi ty: (1) those in areas wh ere hibe rnation occu rs (e.g., rocky l edges), (2) seaso nal migration b etween hibe rnation a nd s ummer ranges dur ing s pring a nd f all, (3) daily acti vi ties wi thin a hom e range dur ing t he acti ve seaso n, and (4) wandering m ovemen ts d uring w hic h the racer shifts i ts acti vi ties.
Popul ation de nsity . Popula tion de nsit ies of b etween 0.3 and 7 acti ve sn akes p er hectar e have been recorded in differ ent habitats an d areas (Fitch , 1963; Turner, 1977). Data on popula tion de nsit ies are lim ited due to the dif ficult y in accu rately cen susing snakes.
Popul ation dy namics. Male racers can reach sexu al matu rity b y 13 to 14 months, whereas femal es ten d not to matu re unti l 2 or 3 years of age (Behler and King, 1979; Brown and Parker , 1984). Adult femal es produc e at most a single clut ch each year (some may reproduc e only in a lternate year s) (Fitch , 1963). In general, the number of eggs in a clutch is propor tiona l to the size of th e femal e and ranges fr om 4 to 30 eggs (Fitch , 1963). Incubation l asts ap proximatel y 40 days to 2 months, depending on temp erature (Behler and King, 1979; Smith, 1956). Juvenile sn akes su ffer highe r mo rtalit y rates (e.g., 80 percent) than adult sn akes (e.g., 20 percent) (Brown and Parker , 1984).
Simi lar species (from general references)
� The easter n coachwhip ( Masti cophi s flagellum flagellum) (black p hase) is simila r in s ize and r anges from Nor th Carolina and s out h Flor ida to Texas, Oklahoma, and Kansas.
� The wester n coachwhip ( Masti cophi s flagellum testaceu s) is si mila r in s ize to the racer . It ranges fr om wester n Nebraska so uth to Mexico.
� The central Texas w hips nake (Masti cophi s taeniatus girardi), Schott' s whips nake (Masti cophi s taeniatus sch otti ), and Rut hven's whips nake
2-409 Racer

�
�
�
(Masti cophi s taeniatus ruthveni) are all simi lar in size to the racer and are restr icted to southern Texas an d northern Mexico.
� The Sonor a whips nake (Masti cophi s bilin eatus) can be slight ly larger (76 to 170 cm) th an the racer and is found f rom Arizona sout hwest to New Mexico and Mexico.
� The str iped racer (Masti cophi s lateralis) is also simi lar in size to the racer snake. It ranges fr om so uth-central Washingt on s out heast i n Great Basin to southern New Mexico and wester n and central Texas, south to west-cen tral Mexico.
� The deser t str iped whips nake (Masti cophi s taeniatus taen iatus) is si mi lar to the central Texas w hips nake. It ranges fr om northern Texas an d northern Calif ornia to Washingt on state.
General references
Behler and King (1979); Brown and Parker (1984); Conant and Collins (1991); DeGraaf and Rudis (1983); Fitch (1963).
2-410 Racer

Racer Snake ( Coluber constrictor )
2-411 R
acer
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight males: Utah (mormon) Brown & Parker, 1984 (g) yrs/mm SVL
<1 8.3 1 27.0 2 41.0 3 49.1 4 53.4 5 60.4 6 61.2
females: Utah (mormon) Brown & Parker, 1984 yrs/mm SVL <1 8.8
1 28.4 2 51.6 3 66.2 4 71.4 5 79.4 6 84.0
males: Kansas ( flavivent ris) Fitch, 1963 yrs/mm SVL 2 68.2 3 102.1 4 139.0 5 152.4 6 175.9 7 181.2 8 217.5
266 420 486 520 541 564 573
272 430 524 575 599 620 632
615 706 757 806 827 845 868

Racer Snake ( Coluber constrictor )
2-412 R
acer
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight females: Kansas ( flavivent ris) Fitch, 1963 (g) yrs/mm SVL (cont inued) 2 83.5
3 149.4 4 212.3 5 209.6 6 245.9 7 251.3
neonat e 4.16 2.4 - 5.8 Kansas ( flavivent ris) Fitch, 1963 215 mm SVL
Egg Weight (g) female size: Kansas ( flavivent ris) Fitch, 1963 892 mm SVL 5.5 4.4 - 6.0 773 mm SVL 4.9 4.4 - 5.2
size NS 7.8 ± 0.17 SE 5.9 - 10.8 Utah (mormon) Brown & Parker, 1984
Juveni le bot h sexes; 0.116 Kansas ( flavivent ris) Fitch, 1963 1 Growth Rate 0 to 10 wks (g/d)
Body A B sum mer 31.8 ± 0.20 SE 18.6 - 37.7 Utah (mormon) Brown, 1973 2 Temperature (��C) A B sum mer 26 - 27 (mode) 15.5 - 32.4 Kansas ( flavivent ris) Fitch, 1963
Metabol ic Rate M basal 6.78 est imated 3 (kcal /kg-d) F basal 6.19
Food Ingest ion B B: spr ing 0.02 Kansas ( flavivent ris) Fitch, 1982 4 Rate (g/g-d) through f all
644 810 866 914 965 974

Racer Snake ( Coluber constrictor )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No. Locat ion/Habi tat Note
insect s 20 40 64 s Illinoi s/past ures, meadow s Klimstra, 1959 5 small mammals 62 27 21 amphi bians 5 13 3 (% vol ume; digest ive t racts) reptiles 7 8 -birds 4 6 8 other 2 6 4
small mammals 65.7 Kansas ( flavivent ris)/ Fitch, 1963 orthopt erans 14.3 locat ions t hroughout state lizards 9.2 snakes 4.2 (% wet weight ; scat s and misc. insect s 1.9 stomach cont ents) birds 3.5 frogs 1.2
mice 15.4 Kansas ( flavivent ris)/ Fitch, 1963 orthopt erans 4.6 woodl and, grassl and lizards 61.5 frogs 12.6 (% wet weight ; stomach snakes 5.1 cont ents) cricket s 0.8
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Home Range A F sum mer 1.8 Kansas ( flavivent ris)/ Fitch, 1963 Size (ha) A M sum mer 3.0 woodl and, grassl and
Popul ation A B sum mer 7.0 Kansas ( flavivent ris)/ Fitch, 1963 Densi ty upl and pr airie, weeds, (N/ha) grasses
B B 0.32 Utah (mormon)/deser t shr ub Brown & Parker, 1984
2-413 R
acer
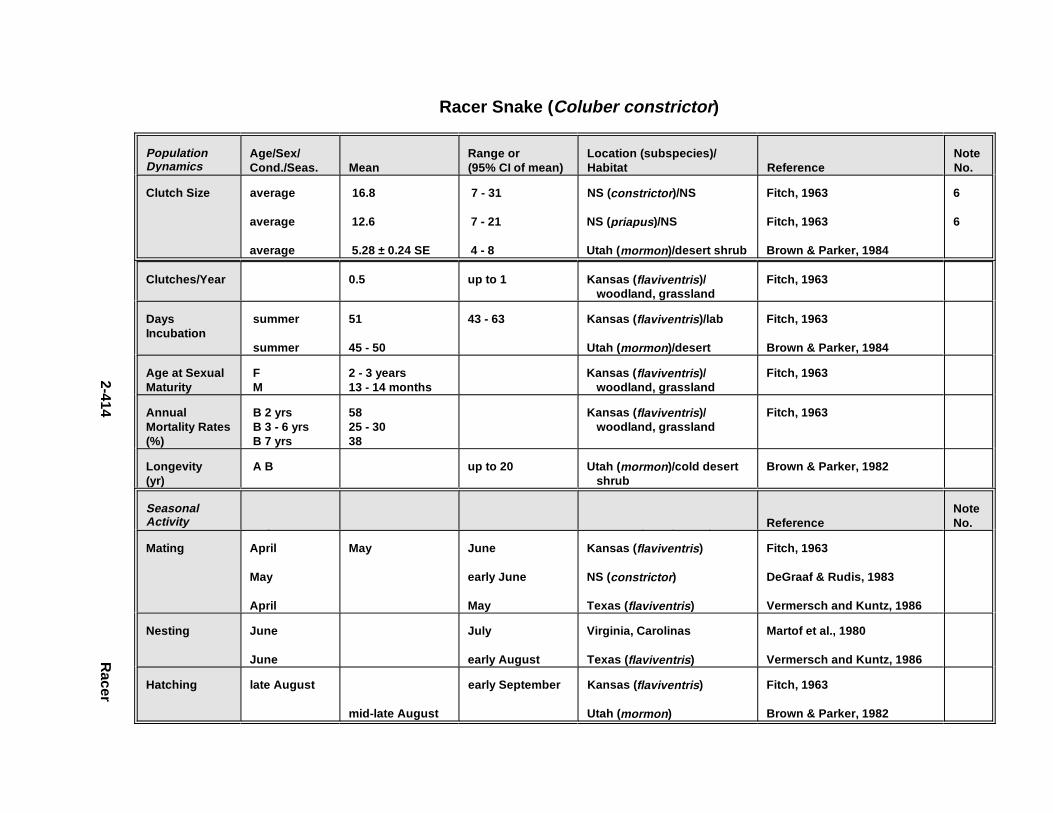
Racer Snake ( Coluber constrictor )
2-414 R
acer
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Clutch Si ze average 16.8 7 - 31 NS (constric to r)/NS Fitch, 1963 6
average 12.6 7 - 21 NS (priapus)/NS Fitch, 1963 6
average 5.28 ± 0.24 SE 4 - 8 Utah (mormon)/deser t shr ub Brown & Parker, 1984
Clutches/ Year 0.5 up t o 1 Kansas ( flavivent ris)/ Fitch, 1963 woodl and, grassl and
Days sum mer 51 43 - 63 Kansas ( flavivent ris)/lab Fitch, 1963 Incubat ion
sum mer 45 - 50 Utah (mormon)/deser t Brown & Parker, 1984
Age at Sexual F 2 - 3 years Kansas ( flavivent ris)/ Fitch, 1963 Maturity M 13 - 14 mont hs woodl and, grassl and
Annual B 2 yrs 58 Kansas ( flavivent ris)/ Fitch, 1963 Mortality Rates B 3 - 6 yrs 25 - 30 woodl and, grassl and (%) B 7 yrs 38
Longevi ty A B up t o 20 Utah (mormon)/col d deser t Brown & Parker, 1982 (yr) sh rub
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating Apr il May June Kansas ( flavivent ris) Fitch, 1963
May early June NS (constric to r) DeGraaf & Rudi s, 1983
Apr il May Texas ( flavivent ris) Vermersch and Kunt z, 1986
Nesting June July Virg in ia, Caro lin as Marto f et al., 1980
June early August Texas (flavivent ris) Vermersch and Kunt z, 1986
Hatching late August early September Kansas ( flavivent ris) Fitch, 1963
mid-late August Utah (mormon) Brown & Parker, 1982
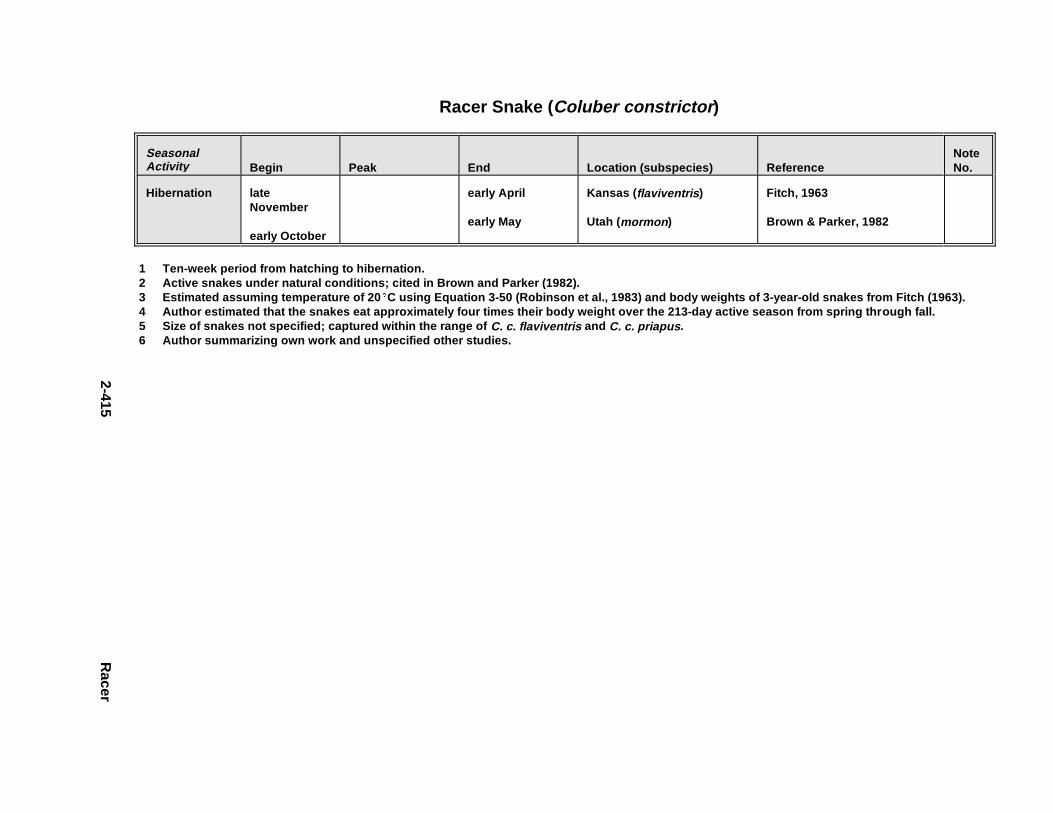
�
Racer Snake ( Coluber constrictor )
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Hibernation late early Apr il Kansas ( flavivent ris) Fitch, 1963 Novem ber
early October early May Utah (mormon) Brown & Parker, 1982
1 Ten-week per iod f rom hatching t o hibernation. 2 Act ive snakes under natural condi tions; ci ted in Brown and Par ker (1982). 3 Est imated assum ing t emperature of 20�C using Equatio n 3-50 (Robinson et al., 1983) and body weight s of 3-year-old snakes f rom Fitch (1963).4 Author est imated that the snakes eat appr oxi mately four times their body weight over the 213-day act ive season f rom spr ing t hrough f all.5 Size of snakes not speci fied; capt ured within the range of C. c. flavivent ris and C. c. priapus .6 Author sum marizing own wor k and unspeci fied ot her studi es.
2-415 R
acer

References (including A ppendix)
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York, NY: Alfred A. Knopf , Inc .
Brown, W. S. (1973) Ecology of th e racer , Coluber constric tor mormon (Serprentes, Colubr idae), in a cold temp erate deser t in northern Utah [Ph.D. disser tat ion] . Salt Lake City, UT: Universi ty o f Utah.
Brown, W. S.; Parker , W. S. (1982) Niche dimensions and r esour ce partit ioning in a great basin deser t sn ake co mmunit y. U.S. Fish Wildl. Serv. Wildl. Res. Rep. 13: 59-81.
Brown, W. S.; Parker , W. S. (1984) Growth , reproduc tion a nd de mogr aphy of th e racer , Coluber constric tor mormon, in nor thern Utah. In: Seige l, R. A.; Hunt , L. E.; Knight , J. L.; et al ., eds. Vertebrate ecology and systemati cs. Lawrence, KS: Museum of Natural Histo ry, The Universi ty o f Kansas; pp. 13-40.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns: easter n/central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
Corn, P. S.; Bury, R. B. (1986) Morphologic al var iation a nd zooge ogr aphy of r acers (Coluber constric tor) in the central Rocky m ount ains . Herpetologic a 42: 258-264.
DeGraaf, R. M.; Rudis , D. D. (1983) Racer snake. Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess; pp. 68.
Fitch , H. S. (1963) Natural histo ry of th e racer Coluber constric tor: v. 15. Lawrence, KS: Universi ty o f Kansas Publi cations , Museum of Natural His tory; pp. 351-468.
Fitch , H. S. (1982) Resources o f a sn ake co mmunit y in pr airie woodla nd ha bitat o f northeaster n Kansas. In: Scott, N. J., Jr., ed. Herpetologic al communit ies. U.S. Fish Wildl. Serv. Wildl. Res. Rep. 13; pp. 83-98.
Gibbons , J. W.; Semlitsch , R. D. (1991) Guide to the rept iles and amphibia ns of th e Savannah River si te. Athens, GA: The Universi ty o f Georgia Press.
Hammer son, G. A. (1987) Thermal behaviour of th e snake Coluber constric tor in west-cen tral California. J. Therm. Biol. 12: 195-197.
Kaufman , G. A.; Gibbons , J. W. (1975) Weight -lengt h relations hip in t hir teen species of snakes i n the southeaster n United States. Herpetologic a 31: 31-37.
Klimstr a, W. D. (1959) Foods of th e racer , Coluber constric tor, in s out hern Illinois . Copeia 1959: 210-214.
Martof, B. S.; Palmer , W. M.; Bailey, J. R.; et al . (1980) Amphibia ns and r ept iles of th e Carolina s and Virginia . Chapel Hill, NC: Univ ersi ty of Nor th Carolina Press.
2-416 Racer

Parker , W. S.; Brown, W. S. (1973) Species co mposi tion a nd popula tion c hanges in t wo complexes o f sn ake hibe rnacula in nor thern Utah. Herpetologic a 29: 319-326.
Pope, C. H. (1944) Amphibia ns and r ept iles of th e Chic ago area. Chic ago, IL: Chic ago Natural Histo ry Museum Press.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Ruben, J. A. (1976) Aerobic and a naerobic metabolis m dur ing acti vi ty i n snakes. J. Comp. Physiol. B: Metab. Transp. Funct. 109: 147-157.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9; 356 pp.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28.
Turner, F. B. (1977) The dynamics o f popula tions of squ amates, crocodilia ns an d rhynchoc ephalians. In: Gans, C.; Tink le, D. W., eds. Biology of th e rept ilia : v. 7. New York, NY: Academic Press; pp. 157-264.
Uhler, F. M.; Cot tom, C.; Clarke, T. E. (1939) Food of sn akes o f th e George Washingt on Nationa l For est, Virginia . Trans. Nor th Am. Nat. Resour . Wildl. Conf . No. 4; pp. 605-622.
Vermersch , T. G.; Kunt z, R. E. (1986) Snakes o f so uth central Texas. Austi n, TX: Eakin Press; pp. 18-19.
Wright , A. H.; Wright , A. A. (1957) Handbook of sn akes: v. 2. Ithaca, NY: Comsto ck.
2-417 Racer

2.3.5. Northern Water Snake (w ater snakes and salt m arsh snakes)
Order Squamata, Family Colubrid ae. Water snakes an d sal t mar sh snakes (g enus Nerodia) belong t o the fami ly Colubrid ae, along w ith 84 percent of th e snake sp ecies in Nor th Amer ica. Colubr ids var y wide ly in f orm and s ize and c an be found in num erous habitats, including t errestr ial, arboreal, aquatic, and burrowing. The more aquatic typ es of snakes i n th is fami ly include water snakes, sal t mar sh snakes, swamp snakes, brown snakes, and garter and ribbon s nakes (Conant and Collins , 1991).
Selected species
The northern water snake (Nerodi a sipedon s ipedon ) is largely aquatic and riparian. It ranges from Maine and s out hern Quebec to Nor th Carolina . It also inha bit s the upla nds of wester n North Carolina and a djacent portions of T ennessee an d Virginia , and it s range exten ds west to easter n Color ado (Conant and Collins , 1991). Three addit iona l subs pecies are recognize d, dis tinguis hed by r ange and ha bitat: N. s. pleuralis (midla nd water snake; ranges from India na to Oklahom a and t he Gulf of Mexico and s out h of th e mount ains to the Carolina s, prefer ring fast-mo ving s treams), N. s. insularum (Lake Er ie water snake; inha bit s is lands of Pu t-in- Bay, Lake Er ie), and N. s. willi amengelsi (Carolina sal t marsh water snake; inha bit s the Outer Bank is lands and m ainla nd c oast o f Paml ico and Core sounds , Nor th Carolina ) (Behler and King, 1979; Conant and Collins , 1991).
Body size. The northern water snake is typ ical ly 61 to 107 cm i n total length (Conant and Collins , 1991). Island popula tions of th e species tend t o be larger th an mainla nd one s (King, 1986). King (1986) esti mated the relations hip b etween snout-to -vent length (SVL)l and body weight fo r Lake Er ie water snakes (N. s. insularum):
weight (g ) = 0.0005 SVL (cm)3.07 all snakes;
weight (g ) = 0.0009 SVL (cm)2.88 femal es; and
weight (g ) = 0.0008 SVL (cm)2.98 males.
Kaufman and Gibbons (1975) determined a relations hip b etween length and weight fo r bot h sexes o f a South Carolina popula tion:
weight (g ) = 0.0004 SVL (cm)3.15 (± 0.12 SE) all snakes
(95% CI for intercept = 0.00015 to 0.0011). Immed iately after emergence fr om hibe rnation, femal es begin t o gain w eight and c ont inue gaining w eight unt il giv ing bir th in l ate su mmer . Weight los s asso ciated wi th parturition in one popula tion r anged f rom 28.2 to 45.5 percent of th e femal e's weight jus t pr ior to partur it ion ( King, 1986).
lMeasures o f SVL excl ude the tail. Kaufman and Gibbons (1975) esti mated that th e tai l represents 21.8 percen t (± 0.010 SE) of the to tal length of a female and 25.7 percen t (± 0.006 SE) of the total lengt h of a male.
2-419 Northern Water Snake

�
�
�
Habitat . The northern water snake prefers str eams b ut can be found in l akes an d ponds and ne arby r iparian areas (King, 1986; Smith, 1961). In the Carolina s and Virginia , they can be found f rom mount ain l akes an d str eams to large coastal estu aries (Martof et al., 1980). They are absent fr om water bodie s wi th soft muddy bot toms whic h may int erfere wi th foraging ( Lagler and S alyer , 1945). In Lake Er ie, N. s. insularum occu rs i n shor eline habitats wh ere rocks o r vegetat ion pr ov ide r efugia (King, 1986).
Food ha bi ts . Northern water snakes co nsume primar ily fi sh and amphibia ns and, to a lesser exten t, insects an d smal l mammal s (Raney and Roecker , 1947; Smith, 1961). Diet var ies acco rding t o the age (and s ize) of th e snake and food availa bilit y (DeGraaf and Rudis , 1983). Young s nakes fo rage in shallow r iffl es and c obble bars, pr imarily wai ting f or prey to move within r ange (l etter from K.B. Jones, U.S. Environmental Protect ion A gency Environm ental Monit or ing Systems L aboratory, to Susan B. Norton, January 6, 1992).
mTadpole s comprise a large propor tion of th e die t of young s nakes in some areas (Raneyand Roecker , 1947). Adult s are str ong s wimmer s and can swi m and dive fo r fish midstr eam, often capturing la rge specimens (e.g., 20 to 23 cm br own t rout ; 19 cm bullhe ad; 20+ cm l amprey) (Lagler and Salyer , 1945). They also tend to consume botto m-dwelling fish species (e.g., sucker s) (Raney and Roecker , 1947). In New York, Brown (1958) found that N. s. sipedon cons umed the most food b etween June and August; they co nsumed littl e dur ing t he remaining m ont hs pr ior to hibe rnation.
Temperature regulation and daily acti vi ties. The northern water snake is acti ve bot h day and night but is mo st acti ve between 21 and 27�C (Brown, 1958; Smith, 1961). Dur ing t he day, th ey are found in a reas th at provide basking s ites an d are not found in heavi ly sh aded areas (DeGraaf and Rudis , 1983; Lagler and Salyer , 1945). They may become inacti ve and seek sh elter , however , if temp eratures exceed 27�C (Brown, 1958; Lagler and Salyer , 1945). They become to rpid a t temp eratures less th an 10�C (Brown, 1958).
Hibernation. In autumn, the N. sipedon leaves th e aquatic habitats to overwinter in rock cr evices o r in banks n earby (DeGraaf and Rudis , 1983; Fitch , 1982).
Breeding acti vi ties and social organization. The northern water snake breeds pr imarily in early sp ring, and t he young a re bor n f rom l ate su mmer to fal l (i.e., viviparous) (DeGraaf and Rudis , 1983). The rate of developm ent before hatching is temp erature dependent (Bauman and Metter , 1977).
Home range and r esour ces. The northern water snake usually stays i n the same area of a str eam, in the same pond, or in an adjacent pond f or sever al years (Fraker , 1970). Snakes a long s treams h ave larger home ranges th an snakes i n ponds and l akes (Fraker , 1970). Fraker (1970) found t hat for large ponds (e. g., 1,500 to 2,000 m2), the home range of an indiv idua l snake is essen tially th e enti re pond. In f ish hatch eries wi th smal ler ponds , indiv idua l snakes fr equent mo re than one pond ( Fraker , 1970).
mSnakes l ess th an 36 cm in length for th is example.
2-420 Northern Water Snake

�
�
�
�
�
�
Popul ation de nsity . Popula tion de nsity esti mates fo r water snakes u sually are expressed relative to a length of sh oreline . Values from 34 to 380 snakes p er km o f shor eline h ave been reported for str eams an d Lake Er ie islands (see tab le).
Popul ation dy namics. Northern water snakes r each sexu al matu rity at 2 o r 3 years of age, wi th males generally matu ring e arlier and a t a smal ler size than femal es (Feaver , 1977, ci ted in King, 1986; King, 1986). Clut ch sizes vary from 5 or 10 to 50 or 60 depending on loc ation a nd on femal e size (see tab le). The propor tion of femal es breeding in a giv en year increases wi th increasing femal e size, as does cl utch size and offsp ring w eight (King, 1986). King deter mined the relations hip of li tter size to femal e SVL for Lake Er ie water snakes (N. s. insularum):
litter size = -12.45 + 0.41 SVL (cm).
Feaver (1977, ci ted in King, 1986) determined the relations hip f or a Mic higa n popula tion:
litter size = -23.55 + 0.55 SVL (cm).
Females produc e only one clut ch per year (Beatso n, 1976). Informat ion on a nnua l surviv orship of juv enile s or adult s was n ot identi fied in the literature reviewed.
Simi lar species (fr om general references)
� The Mississippi gr een water snake (Nerodia cycl opion) can be slight ly larger (76 to 114 cm) th an the northern water snake and is found in quie t water s of the Mississippi V alley.
� The blotch ed water snake (Nerodia eryth rogaster transver sa) is larger than the northern water snake (76 to 122 cm) an d is found in wester n Misso uri and Kansas to northeaster n Mexico.
� The nor thern coppe rbelly ( Nerodia eryth rogaster neglecta) is larger than the northern water snake (76 to 122 cm) an d ranges fr om wester n Kentucky to southeaster n Illinois and t o Michiga n.
� The redbelly water snake (Nerodia eryth rogaster eryth rogaster ) of th e midwester n United States i s cl ose in size to the water snake. It is best su ited to swamp y areas and sluggis h s treams.
� The yellow belly water snake (Nerodia eryth rogaster flavigaster ) is found in the low er Mississippi V alley and adjacent areas. Like th e redbelly, it is simi lar in size to the water snake and likely to be found in swamp y areas and sluggis h s treams.
� The banded water snake (Nerodia fasci ata fasci ata) is si mila r in s ize, and it s range includes th e coastal plain, North Carolina to Mississippi.
2-421 Northern Water Snake

�
�
�
�
�
�
�
� The broad banded water snake (Nerodia fasci ata co nfluens) (56 to 90 cm) occurs in t he Mississippi Riv er delta region in m arshes, swamp s, and shallow water s, including br acki sh water s along t he Gulf Coast.
� The Flor ida water snake (Nerodia fasci ata picti ventr is) is si mila r in s ize to the northern water snake and ranges fr om th e extr eme so utheast o f Georgia to the sout hern tip of Flor ida. It lives p rimar ily in shallow fr eshwater habitats.
� Harter 's water snake (Nerodia harter i) is relatively smal l (51 to 76 cm) an d is found in c entral Texas.
� The diamondb ack water snake (Nerodi rhom bi fer rhom bi fer ) can be slight ly longe r (76 to 122 cm) th an the northern water snake and is mo re thick-bodie d than most Nerodia. Its r ange exten ds so uth from th e Missi ss ippi V alley int o Mexico.
� The Gulf sal t mar sh snake (Nerodia clarki i clarki i) inha bit s the Gulf Coast from west-cen tral Flor ida to sout hern Texas. It is abunda nt in c oastal sal t meadows, swamp s, and marshes.
� The Atlantic sal t mar sh snake (Nerodia clarki i taeniata) is restr icted to Volus ia Count y along t he Atlant ic Coast o f north Flor ida.
� The mangrove sal t mar sh snake (Nerodia clarki i compressi cauda) is smal l (38 to 76 cm) an d inha bit s the mangr ove swamp s of Flor ida 's low er co asts.
Dietary differ ences ar e evident among t hese sp ecies. Mushinsky et al . (1982) found in Louis iana fo rested wetl ands th at N. eryth rogaster and N. fasci ata change from a diet of fish to one dominated by fr ogs wh en they exceed an SVL of 50 cm. N. rhom bi fer and N. cycl opion, on t he other hand, cons ume pr imarily fi sh throughout th eir lives, although t he species and s ize co mpos it ion of th eir die t changes as they gr ow larger (Mus hin sky et al ., 1982). As N. rhombifer exceed s 80 cm SVL , it begins to prey upon la rger fi sh that oc cupy deeper open-water habitats. N. cycl opion eats a l arger propor tion of cen trarchid f ish as it s body size inc reases. In a stu dy of th e diet of N. rhom bi fer , Plummer and Goy (1984) found a relations hip b etween the SVL of th e snakes an d the stan dard length (SL) of th e fish prey (defined as 80 percent of to tal length):
SLfish (cm) = -5.9 + 0.23 SVLsnake (cm) for males, and
SLfish (cm) = -3.6 + 0.17 SVLsnake (cm) for femal es.
The regression line s are not signif icantly di ffer ent, however .
General references
Behler and King (1979); Conant and Collins (1991); DeGraaf and Rudis (1983); King (1986).
2-422 Northern Water Snake

Water Snake ( Nerodia si pedon )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight A B 207 up t o 480 Kansas Fitch, 1982 (g)
J B 1 y r 7.0 ± 2.3 SD 5.3 - 10.4 New York (sipedon ) Brown, 1958 J B 2 y r 29.0 (N = 2) 25.2 - 32.7 J M 3 yr 53.2 (N = 1) A B 5 - 6 yr 210.0 ± 65 SD 114 - 255
neonat e B 4.8 3.6 - 6.6 Ohio, Ontario (insularum) King, 1986
Lengt h A M 620 SVL Ohio, Ontario (insularum) King, 1989 1 (mm) A F 745 SVL
J B 1 y r 285 total New York (sipedon ) Brown, 1958 2 J B 2 y r 496 total J M 3 yr 607 total A B 5 - 6 yr 868 total
neonat e 181 SVL 125 - 210 SVL Ohio, Ontario (insularum) King, 1986 1
Juveni le J 1 yr 0.18 ± 0.08 SD 0.13 - 0.27 New York (sipedon ) Brown, 1958 Growth Rate J 2 yr 0.42 0.40 - 0.45 (g/d) J 3 yr 0.80
Metabol ic B rest ing: Oklahom a, Nerodi a Gratz & Hut chinson, 1977 Rate 15��C 0.607 ± 0.035 SE 0.39 - 0.94 (lO /kg-d) 25��C 3.29 ± 0.10 SE 2.81 - 4.442
35��C 7.33 ± 0.23 SE 5.70 - 9.99
rhom bifera (simila r species)
Food J B 1 y r 0.088 New Yor k (sipedon ) Brown, 1958 3 Ingest ion Rat e J B 2 y r 0.043 (g/g-d) J M 3 yr 0.043
A B 5 - 6 yr 0.061
Surface Ar ea 155 mm SVL 131.16 Arkansas, Nerodi a Baeyens & Rount ree, 1983 (cm ) rhom bifera (simila r2
speci es)
2-423 N
orth
ern W
ater Sn
ake

Water Snake ( Nerodia si pedon )
Dietary Compositio n Spr ing Summer Fall Winter Habi tat (measur e) Reference No. Locat ion (subspeci es)/ Note
Esocidae 7.0 Georg ia/aquatic (NS) Camp et al., 1980 Catostomidae 22.5 Percidae 15.7 (% wet vol ume; Proteidae 51.9 stomach cont ents) Cyprin idae 1.5 Centrarchidae 0.3 season not speci fied crawfish 1.5
trout 64 n lower Michi gan/streams Alexander , 1977 4 non- trout fish 7 uni dent ified f ish 1 (% wet weight ; Crust acea 1 stomach cont ents) Amphi bia 14 birds & mammals 12 uni dent ified 1
minnows 9.1 n lower Michi gan/lakes Brown, 1958 5 darte rs 1.4 Amphi bia 52.8 (% vol ume; stomach scul pin (Cottidae) 2.2 cont ents) trout perch 2.8 (Percopsi s) 14.1 game fishes ( Perca) 17.4 bur bot (Lot a) 0.3 catfish ( Ictalur idae)
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/ Note Cond. /Seas. Mean Range Habi tat Reference No.
Popul ation A B 138 22 - 381 Ohio, Ontario (insularum)/ King, 1986 Densi ty Lake Er ie islands (N/km shor e) B B mer 34 - 41 Kansas ( sipedon )/stream Beatson, 1976 sum
2-424 N
orth
ern W
ater Sn
ake

Water Snake ( Nerodia si pedon )
Population Dynamics
Age/Sex/ Locat ion (subspeci es)/ Note Cond. /Seas. Mean Range Habi tat Reference No.
Li tter Size 11.8 4 - 24 Michi gan (sipedon )/ponds, Feaver, 1977 6
20.8 ± 8.2 SD 6 - 34 Ohio, Ontario (insularum)/ Camin & Ehr lich, 1958
22.9 9 - 50 Ohio, Ontario (insularum)/ King, 1986
33 13 - 52 Illinoi s (pleuralis )/NS Smith, 1961
marshes
Lake Er ie islands
Lake Er ie islands
Li tters/Year 1 cent ral Missour i Bauman & Metter, 1977
1 Kansas ( sipedon )/stream Beatson, 1976 (sipedon )/fis h hatchery
Days Gestation 58 cent ral Missour i Bauman & Metter, 1977 (sipedon )/fis h hatchery
Age at Sexual F 34 mo Michi gan (sipedon )/ponds, Feaver, 1977 6 Maturity (d) M 23 - 24 mo marshes
F 3 yrs Ohio, Ontario (insularum)/ King, 1986 M 2 yrs Lake Er ie islands
Lengt h at F 476 - 649 Michi gan (sipedon )/ponds, Feaver, 1977 6 Sexual M 375 - 425 marshes Maturity (mm SVL) F 590 Ohio , Ontario (insularum)/ King, 1986
M 430 Lake Er ie islands
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating Apr il - May Kansas ( sipedon ) Smith, 1956
mid-May mid-June cent ral Missour i (sipedon ) Bauman & Metter, 1977 May Michigan (sipedon ) Feaver, 1977 6
2-425 N
orth
ern W
ater Sn
ake

123 �
456
Water Snake ( Nerodia si pedon )
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Partur ition late August September Illin ois (sipedon ) Smith, 1961 mid-August late September Ohio, Ontario (insularum) King, 1986
late summer Virg inia, Caro lin as (sipedon ) Marto f et al., 1980
Hibernation mid-October mid-April Ohio , Ontario (insularum) King, 1986 Novem ber late March Michigan (sipedon ) Feaver, 1977 6
SVL = snout-to -vent length, which excl udes t he tail beyond t he vent .Total = total lengt h, from nose t o tip of tail.Snakes i n capt ivi ty; mean temperatures = 23�C. Snakes f ed f ish (one f ed f rogs) .Col lected whenever they were found; thought to be act ive in area from May to September.Mont hs of col lect ion and si ze of snakes not speci fied.Cited in King (1986).
2-426 N
orth
ern W
ater Sn
ake

References (including A ppendix)
Aldr idge , R. D. (1982) The ovar ian cycl e of th e water snake, Nerodi a sipedon , and effects o f hypoph ysecto my an d gona dot ropin a dminis tration. Herpetologic a 38: 71-79.
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Academician 10: 181-195.
Baeyens, D. A.; Rount ree, R. L. (1983) A comparative stu dy of evap orative water loss an d epide rmal permeabilit y in a n arbor eal snake, Opheodrys aesti vus, and a semi -aquatic sn ake, Nerodi a rhom bi fera. Comp. Bioc hem. Physiol. 76A: 301-304.
Barbour, R. W. (1950) The repti les of Big Bl ack Mount ain, Harlan Count y, Kentucky. Copeia 1950: 100-107.
Bauman, M. A.; Metter , D. E. (1977) Reproducti ve cycl e of th e northern water snake Natrix s. sipedon (Reptilia . Serpentes, Colubr idae). J. Herpetol. 11: 51-59.
Beatso n, R. R. (1976) Environmental and genetical correlates o f disruptive color ation in t he water snake, Natrix s. sipedon. Evolut ion 3 0: 241-252.
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York: Alfred A. Knopf , Inc .
Brown, E. E. (1958) Feeding ha bit s of th e nor thern water snake, Natr ix sipedon s ipedon Linna eus. Zoologic a (N.Y.) 43: 55-71.
Bush, F. M. (1959) Foods of so me Kentucky h erpti les. Herpetologic a 15: 73-77.
Camin, J. H.; Ehrlich, P. R. (1958) Natural select ion in water snakes (Natrix sipedon L.) on islands in Lake Er ie. Evolut ion 1 2: 504-511.
Camp, C. D.; Sprewel l, W. D.; Powders, V. N. (1980) Feeding ha bit s of Nerodia taxi spilota wi th comparative notes o n the foods of symp atr ic conge ners in G eorgia . J. Herpetol. 14: 301-304.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns: easter n/central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
DeGraaf, R. M.; Rudis , D. D. (1983) Water snake. Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Feaver , P. E. (1977) The demography of a Michiga n popula tion of Natrix sipedon with dis cussions of ophidia n gr owth and r eproduc tion [ Ph.D. disser tat ion] . Ann A rbor , MI: Univ ersi ty of Michiga n.
2-427 Northern Water Snake

Fitch , H. S. (1982) Resources o f a sn ake co mmunit y in pr airie woodla nd ha bitat o f northeaster n Kansas. In: Scott, N. J., Jr., ed. Herpetologic al communit ies. U.S. Fish Wildl. Serv. Wildl. Res. Rep. 13; pp 83-98.
Fraker , M. A. (1970) Home range and homing in t he water snake, Natrix s. sipedon. Copeia 1970: 665-673.
Gratz, R. K.; Hutchins on, V. H. (1977) Energetics fo r acti vi ty i n the diamondback water snake, Natrix Rhombifera. Physiol. Zool. 50: 99-114.
Justy, G. M.; Mallor y, F. F. (1985) Thermoregula tory behaviour in t he nor thern water snake, Nerodi a s. sipedon, and the easter n garter snake, Thamnophi s s. si rtal is. Can. Field-Nat. 99: 246-249.
Kaufman , G. A.; Gibbons , J. W. (1975) Weight -lengt h relations hip in t hir teen species of snakes i n the southeaster n United States. Herpetologic a 31: 31-37.
King, R. B. (1986) Popula tion e cology of th e Lake Er ie water snake, Nerodi a sipedon insularum. Copeia 1986: 757-772.
King, R. B. (1989) Body size var iation a mong is land and m ainla nd s nake popula tions . Herpetologic a 45: 84-88.
Lagler, K. F.; Salyer , J. C., II. (1945) Food a nd ha bit s of th e common water snake, Natrix sipedon , in Mic higa n. Pap. Michiga n Acad. Sci ., Arts and L etter s 31: 169-180.
Martof, B. S.; Palmer , W. M.; Bailey, J. R.; et al . (1980) Water snake. Amphibia ns an d rept iles of th e Carolina s and Virginia . Chapel Hill, NC: Univ ersi ty of Nor th Carolina Press.
Mushinsky, H. R.; Hebrard, J. J.; Vodopic h, D. S. (1982) Ontogeny of water snake fo raging ecology . Ecology 63: 1624-1629.
Plummer , M. V.; Goy, J. M. (1984) Ontogenetic dietary sh ift o f water snakes (Nerodia rhom bi fera) in a fish hatch ery. Copeia 1984: 550-552.
Raney, E. C.; Roecker , R. M. (1947) Food a nd gr owth of two species of water snakes fr om wester n New York. Copeia 1947: 171-174.
Smith, H. M. (1956) Water snake. Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. Hist. Misc. Publ. 9; 356 pp.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28.
Uhler, F. M.; Cot tom, C.; Clarke, T. E. (1939) Food of sn akes o f th e George Washingt on Nationa l For est, Virginia . Trans. Nor th Am. Wildl. Nat. Resour . Conf . 4: 605-622.
Wright , A. H.; Wright , A. A. (1957) Handbook of sn akes: v. 2. Ithaca, NY: Comsto ck.
2-428 Northern Water Snake

2.3.6. Eastern New t (salam anders)
Order Caudata, Family Salamandridae. Notopht halmus , the genus co mprising t he easter n newts, inha bit s easter n North Amer ica. A differ ent genus, Taricha, comprises th e wester n newts a long t he Paci fic co ast o f North Amer ica. Unlik e other sal amanders, the ski n of newts i s rough textu red, not sl imy. Easter n newts ar e primar ily aquatic; wester n newts ar e ter restr ial. The life cycl e of easter n newts i s co mplex. Females deposi t th eir eggs int o shallow su rface water s. After hatching, the larvae remain aquatic fo r 2 to sever al mont hs before transfo rming int o br ight ly color ed ter restr ial forms, cal led efts (H ealy, 1974). Postl arval migr ation of efts fr om ponds to land may take p lace fr om Ju ly th rough Novemb er, but ti ming v aries between popula tions (Hur lbert, 1970). Efts l ive on land (forest floor ) for 3 to 7 years (Healy, 1974). They th en return to the water and assu me adult character isti cs. In changing f rom an eft to an adult , the newt d evelops fins and t he ski n changes to permit aqua tic respir ation (Smith, 1961). Occas iona lly n ewts o mit th e ter restr ial eft stag e, especially in the species located in the southeast co astal plain (Conant and Collins , 1991) and along t he Massach usetts co ast (Healy, 1974). These aquatic juveniles have th e same ad aptat ions (i .e., smoot h skin a nd f latten ed tai l) as th e aquatic adults b ut are not sexu ally matu re (Healy, 1973). Under favo rable condit ions , adult s are permanently aquatic; however , adults may mi grate to land after breeding due to dry ponds , high water temp eratures, and low oxyg en tension ( Hur lbert, 1969). The life cycl e of wester n newts d oes not include th e eft stag e (Conant and Collins , 1991).
Selected species
The easter n newt (Notopht halmus virid escen s) has both aquatic and ter restr ial forms. The aquatic adult is usually yellow ish-brown or oli ve-green to dark brown above, yellow below . The land-dwelling eft i s orange-red to reddis h-brown, and it s skin c ont ains tetr odot ox in, a neurotox in and pow erful emeti c. There are four subspecies of easter n newts: N. v. virid escen s (red-spotted newt; ranges fr om Nova Sco tia west to Great Lakes and south to the Gulf states), N. v. dorsalis (broken-str iped newt; ranges along t he coastal pla in of th e Carolin as), N. v. louisianensis (central newt; ranges fr om wester n Michiga n to the Gulf), and N. v. piaropicola (penins ula n ewt; restr icted to penins ular Flor ida) (Conant
nand Collins , 1991). Neoteny occu rs co mmonly in t he penins ula and br oken-str iped newts.In the central newt, neoteny is fr equent in the southeaster n coastal plain. In the red-spotted newt, neoteny is rare (Conant and Collins , 1991).
Body size. Adult easter n newts u sually are 6.5 to 10.0 cm i n total length (Conant and Collins , 1991). In North Carolina , N. v. dorsalis efts r anged from 2.1 to 3.8 cm s nout-to -vent length (SVL), which exc ludes th e tai l, and adults r anged from 2.0 to 4.4 cm SVL (Harris, 1989; Harris et al ., 1988). Healy (1973) found a quatic juv enile s 1 year of age to range fr om 2.0 to 3.2 cm SVL. Adult easter n newts we igh a ppr ox imatel y 2 to 3 g (Gill, 1979; Gillis and Br euer, 1984), whereas th e efts g enerally weigh 1 to 1.5 g (Bur ton, 1977; Gillis and Breuer, 1984).
nNeotenic newts are mature and c apable of reproduc tion but retain the larval form, appearance, and habits (Conant and Collin s, 1991).
2-429 Easter n Newt

�
Habitat . Larval and adult easter n newts ar e found in ponds , especially thos e wi th abunda nt subm erged vegetation, and in weedy areas of lakes, marshes, ditch es, backwater s, and pools of sh allow slo w-mo ving s treams o r other unpollut ed shallow or semi permanent water . Terrestr ial efts inha bit mi xed and de ciduous fo rests (B ishop, 1941, ci ted in Sousa, 1985) and are found in m ois t areas, typ ical ly unde r damp leaves, brush pile s, logs , and s tumps, usually in w oode d habitats (D eGraaf and Rudis , 1983). Adequate surface l itter is important, especially during dr y periods , because efts se ldom bur row (Healy, 1981, ci ted in Sousa, 1985).
Food ha bi ts . Adult easter n newts ar e oppor tunis tic pr edators that pr ey unde rwater on worms, insects an d their larvae (e.g., mayfl y, caddis fly, midge , and m osquit o larvae), smal l crustacean s and mollu scs, spide rs, amphibia n eggs , and o ccas iona lly smal l fish. Newts cap ture prey at th e surface o f th e water and on the botto m of th e pond, as well a s in the water colum n (Ries and Be llis , 1966). The shed ski n (exuvia) is eaten and may comprise greater than 5 percent of th e total weight of food items o f both the adult and e ft diets (MacNamara, 1977). Snails are an important food s our ce for th e ter restr ial eft (Burton, 1976). Efts feed only dur ing r ainy su mmer periods (B ehler and King, 1979; Healy, 1973). Healy (1975) noted that in late August an d Septemb er, efts o ften were found cluster ed around decay ing m ushroom s feeding on a dult and la rval dipt erans. In a nor thern hardwood he mloc k forest i n New York, MacNamara (1977) found t hat most pr ey of adult migrants an d immatu re efts wer e from th e uppe r li tter layer , soil s urface, or low veg etat ion.
Temperature regulation and daily acti vi ties. Adult n ewts ar e often seen foraging in shallow water , and efts ar e often found in la rge num bers on t he forest floor after it rains (Behler and King, 1979). Efts may b e found on t he open forest floor even during da ylight hour s (Conant and Collins , 1991), but th ey rarely emer ge if th e air temp erature is below 10�C (Healy, 1975).
Hibernation. Most ad ults r emain acti ve al l winter unde rwater on pond bot toms or in str eams (DeGraaf and Rudis , 1983). Some adults o verwinter on land (Hurlbert, 1970) and migrate to ponds dur ing t he spring t o breed (Hur lbert, 1969). If th e water body fr eezes to the botto m, adults may b e forced to hibe rnate on land or to migrate to another pool (Smith, 1956). Efts hibe rnate on land, burrowing unde r logs and de br is . Hur lbert (1969) observed that efts mi grated to ponds fo r th e fi rst time in t he spring a nd f all.
Breeding acti vi ties and social organization. In south-central New York, breeding takes p lace in late wi nter or early sp ring, usually in l akes, ponds , and swamp s (Hurlbert, 1970). Ovulation a nd egg de pos it ion oc cur over an exten ded period ( McLaughlin a nd Humphries, 1978). Females overwinter ing on la nd c an sto re sperm for at least 10 mo nths (Massey, 1990). Spawning unde rwater , the femal e deposi ts eg gs s ingly on l eaves o f subm erged pla nts, hiding a nd w rapping each in vegetat ion ( Gibbons and Semlitsch , 1991; Smith 1956). The time to hatching de pends on temp erature (DeGraaf and Rudis , 1983). Smith (1961) found t ypic al inc ubation pe riods to be 14 to 21 days i n Illinois , whereas th e inc ubation pe riod obs erved by Behler and King (1979) was 21 to 56 days.
Growth and metamo rphos is. In late su mmer or early fal l, the larvae tr ansfo rm into either aquatic juveniles or ter restr ial efts (B ehler and King, 1979). Harris (1987) sh owed
2-430 Easter n Newt

�
�
that low larval densi ty sti mulated neoteny in larvae unde r exp erimental condit ions . Larval growth rates wer e highe r in ponds wi th low larval dens it ies (Harris, 1987; Morin et al ., 1983). Growth rates fo r aquatic juvenile s are high est i n the spring; however , maxi mum seaso nal growth for the ter restr ial efts o ccu rs between June and Septemb er when the temp erature is optimal for acti ve fo raging ( Healy, 1973).
Home range and r esour ces. For adult n ewts, Bellis (1968) found t he mean distan ce between capture and recapture si tes to be about 7 m, indi cating smal l home ranges. Harris (1981, ci ted in DeGraaf and Rudis , 1983) did not find a ny define d hom e range or any ter ritoriality fo r males. Most efts ar ound a pond in P enns ylvania r emaine d within 1 .5 m of the shor e (Bellis , 1968). Healy (1975) esti mated the home range for ter restr ial efts i n a Massach usetts w oodla nd t o be 270 m2 and located approximatel y 800 m fr om th e ponds where the adults an d larvae wer e located .
Popul ation de nsity . Popula tions of aqua tic adult s may r each high loc al dens it ies, whereas ter restr ial efts ex hibit low er popula tion de nsit ies. Recorded popula tion de nsit ies for ter restr ial efts r ange from 34 per hectar e (ranging f rom 20 to 50 efts p er hectar e) in a Nor th Carolina mi xed deciduous fo rest (Sh ure et al ., 1989) to 300 per hectar e in a Massach usetts w oodla nd (Healy, 1975). Harris et al . (1988) observed a densi ty o f 1.4 adult newts p er m2 (14,000 adult n ewts p er hectar e) in a shallow pond in Nor th Carolina in t he winter , whereas th e summer popula tion de nsity was only 0. 2 adult s per m2 (2,000 adult s per hectar e).
Popul ation dy namics. Many popula tions of th e easter n newt r each sexu al matu rity when the eft stag e returns to the water and changes to the adult fo rm (Healy, 1974). However , unde r cer tain c ondit ions su ch as low larval dens ity, mo st of th e larvae present have been shown to metamo rphose directl y into adults o r even into sexu ally matu re larvae (Harris, 1987). In experimental ponds , dens it ies of 22 la rvae per m2 resulted in metamo rphos is to eft b y th e majority, while a de ns ity of 5. 5 larvae per m2 resulted in metamo rphos is dir ectl y to the adult fo rm or sexu al matu ration w ithout metamo rphos is (Harris, 1987). Adult dens ity also inf luences r eproduc tion. Mor in e t al. (1983) found t hat doubling a dult dens ity result ed in a r educ tion of o ffsp ring pr oduc ed to one-quarter th at produced by adults at th e lower densi ty (i .e., from 36 offsp ring pe r femal e in tanks cont aining 1 .1 femal es per m2 to 9.7 offsp ring pe r femal e in tanks co ntaining 2 .2 femal es per m 2). The adult lif e expectan cy n oted by Gill (1978b) was 2.1 breeding seaso ns fo r males and 1.7 breeding seaso ns fo r femal es. Amphibia n blood l eeches (ecto parasi tes) ar e likely to be a primary so urce of mo rtal ity fo r adults; they also prey directl y on larvae (Gill, 1978a).
Simi lar species (from general references)
� The black-sp otted newt (Notopht halmus mer idionalis) is si mila r in s ize (7.5 to 11.0 cm) to the easter n newt. It has large black sp ots an d is found in south Texas i n ponds , lagoons , and swamp s. There is no eft stag e.
� The str iped newt (Notopht halmus perstr iatus) is smal ler (5.2 to 7.9 cm) th an the easter n newt an d ranges fr om so uthern Georgia to central Flor ida. It is found in a lmost any body of sh allow , standing water .
2-431 Easter n Newt

�
�
� The wester n newts ( Taricha) are found a long t he Paci fic co ast. They do not unde rgo t he eft stag e but rather transfo rm into land-dwelling a dult s that return to the water at breeding t ime.
� Other small s alamande rs are simila r but var y by having s lim y skin a nd cons pic uous co stal grooves. They differ in life histo ry, however ; in the fami ly Plethudont idae, all are lungl ess an d breathe through t hin, mois t skin. Many are completely ter restr ial.
General references
Behler and King (1979); Conant and Collins (1991); DeGraaf and Rudis (1983); Hurlbert (1969); Smith (1961).
2-432 Easter n Newt
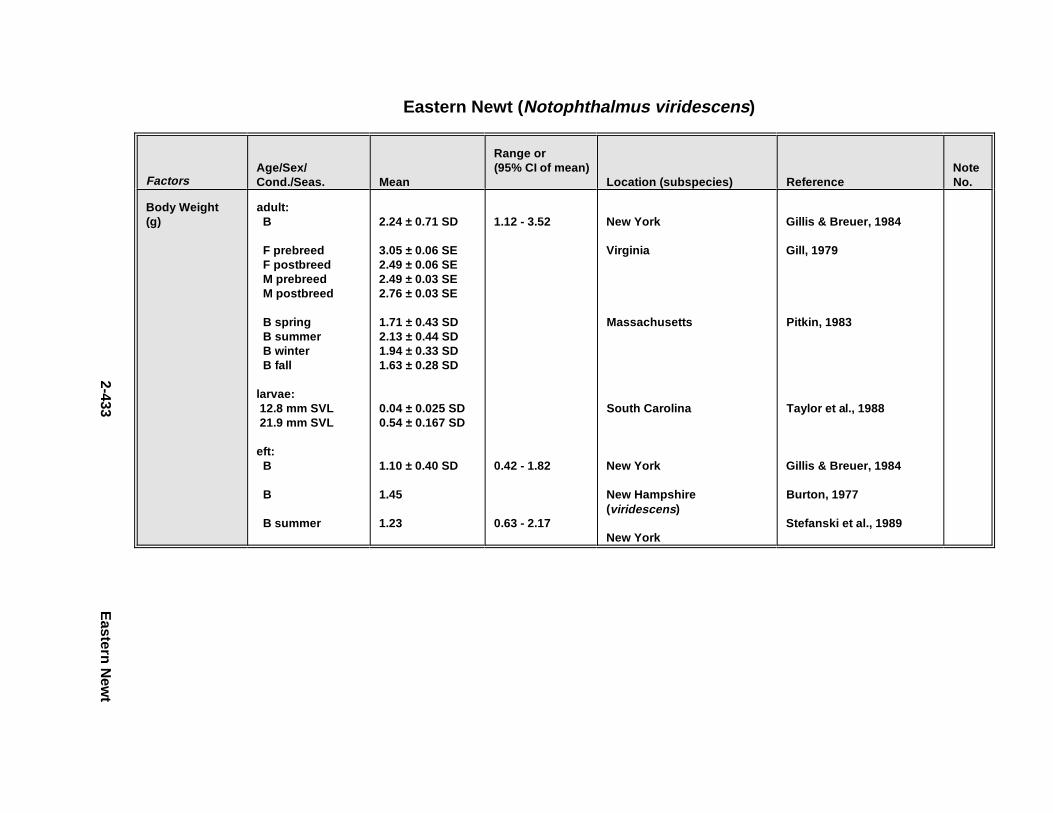
Eastern New t (Noto phthalmus viridescens )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean ion (subspeci es) Reference No.
Range or
Body Weight adul t: (g) B 2.24 ± 0.71 SD 1.12 - 3.52 New York Gillis & Breuer, 1984
F prebreed 3.05 ± 0.06 SE Virg in ia Gill, 1979 F post breed 2.49 ± 0.06 SE M prebreed 2.49 ± 0.03 SE M post breed 2.76 ± 0.03 SE
B spr ing 1.71 ± 0.43 SD Massachuset ts Pitkin, 1983 B sum mer 2.13 ± 0.44 SD B winter 1.94 ± 0.33 SD B fall 1.63 ± 0.28 SD
larvae: 12.8 mm SVL 0.04 ± 0.025 SD Sout h Carolina Taylor et al., 1988 21.9 mm SVL 0.54 ± 0.167 SD
eft: B 1.10 ± 0.40 SD 0.42 - 1.82 New York Gillis & Breuer, 1984
B 1.45 New Hampshi re Burton, 1977
B summer 1.23 0.63 - 2.17 Stefanski et al., 1989 (vi ridescens )
New Yor k
Locat
2-433 E
astern N
ewt

Eastern New t (Noto phthalmus viridescens )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean ion (subspeci es) Reference No.
Range or
Lengt h adul t: (mm SVL) M 35.0 24 - 44 North Carolina (dor sal is) Harris et al., 1988
F 35.0 20 - 42
B sum mer 38.9 33 - 48 New York MacNamara, 1977
juvenile : B spr ing 26.1 ± 0.35 SE 20 - 32 Massachuset ts (vi ridescens ) Healy, 1973
larvae: B spr ing 12.3 s Illinoi s Brophy , 1980 B fall 19.2
eft: B (mm total) 50.4 ± 0.5 SE North Carolina (dor sal is) Harris et al., 1988
B spr ing 20.5 Massachuset ts (vi ridescens ) Healy, 1973
B sum mer 32.7 18 - 41 New York MacNamara, 1977
Larval Growth high densi ty: North Carolina Harris, 1987 Rate (g/d) -> efts 0.00310 high densi ty: 55,000/ha
-> adul ts 0.00421 -> neonat es 0.00536
low densi ty: -> efts 0.00635 low densi ty: 220/ha -> adul ts 0.00685 -> neonat es 0.00676 1
1
Metabolic efts at 15��C: New York Stefanski et al., 1989 Rate resting 1.47 (lO /kg-d) act ive 4.272
Locat
2-434 E
astern N
ewt

Eastern New t (Noto phthalmus viridescens )
Factors Age/Sex/ (95% CI of mean) Note Cond. /Seas. Mean ion (subspeci es) Reference No.
Range or
Metabol ic Rate basal : (kcal /kg-d) A M post breed 16.2 est imated 2a
A F post breed 16.7 larvae (12.8 mm) 43.5 est imated 2b eft (71.0 mm) 20.1 est imated 2c
Food Ingest ion 3 Rate (g/g-d)
Surface Ar ea A M 17 est imated 4 (cm ) A F 152
Dietary Compositio n Spr ing Summer Fall Winter Habi tat (measur e) Reference No. Locat ion (subspeci es)/ Note
aquat ic adul ts: New Hampshi re Burton, 1977 Ephemeroptera 7.5 7.5 (vi ridescens )/small Odonat a 31.9 1.9 ol igot rophi c lake Lepi dopt era 13.7 0.9 Diptera 5.8 0.3 (% wet weight ; stomach other insect s 9.9 0.6 and gut cont ents) Cladocer ans 5.1 84.1 Amphi poda 5.6 3.1 Pelycepoda 6.2 1.5 N. viridescens
larvae 11.4 0.0 other 3.2 0.1
Locat
2-435 E
astern N
ewt

Eastern New t (Noto phthalmus viridescens )
Dietary Compositio n Spr ing Summer Fall Winter Habi tat (measur e) Reference No. Locat ion (subspeci es)/ Note
efts : New York/leaf litter MacNamara, 1977 Basom matophor a 5.5 sur face in forest Stylommatophor a 18.3 Acar i 13.8 (% dry weight ; stomach Collembola 10.4 contents ) Thysanopt era 3.4 Homoptera 4.7 Coleoptera adult 2.3 Coleopt era larvae 3.5 Lepi dopt era larvae 7.9 Diptera adul t 9.7 Diptera larvae 10.6 Hymenopt era adul t 5.8
larvae: New Hampshi re Burton, 1977 Zygopt era (Odonat a) 0.8 (vi ridescens )/small Chironom idae 16.2 ol igot rophi c lake (Dip tera)
Cladocer a 12.7 (% wet weight ; stomach Ostracoda 5.3 and gut cont ents) Hyalle la azteca 55.1 (Amphi poda)
Sphaerium sp. 9.4 (Pelycepoda)
Planor bidae 0.5 (Gastropoda)
Rhizopoda ( Protozoa) 0.01
2-436 E
astern N
ewt

Eastern New t (Noto phthalmus viridescens )
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Home Range eft: Size B 0.0087 ha 0.0028 - 0.0153 Massachuset ts Healy, 1975
adul t: summer 6.86 m Pennsylvania Bellis , 1968 5
(vi ridescens )/ oak/pine forest
(vi ridescens )/pond
Popul ation A B ent ire lake 130 - 173 New Hampshi re Burton, 1977 Densi ty A B f ringe onl y 50 - 2,600 (vi ridescens )/small (N/ha) ol igot rophi c lake
A winter 50,000 ± 9,000 SE North Carolina (dor sal is)/ Harris et al., 1988 A sum mer 3,000 ± 1,000 SE shal low pond
eft spr ing 300 Massachuset ts Healy, 1975
eft s ummer 34 20 - 50 North Caro lin a Shure et al., 1989
larvae spr ing 21,000 0 - 350,000 Sout h Carolina/pond, Taylor et al., 1988
larvae spr ing 65,000 ± 15,000 SE North Carolina (dor sal is)/ Harris et al., 1988 summer 25,000 ± 5,000 SE shal low pond fall 10,000 ± 3,000 SE
(vi ridescens )/ oak/pine forest
(vi ridescens )/mixed deci duous f orest
wetland
Clutch Si ze 200 - 400 NS/NS Behler & Ki ng, 1979 (eggs)
Days to Hatching 14 - 21 Illinoi s/NS Smith, 1961
21 - 56 NS/NS Behler & Ki ng, 1979
2-437 E
astern N
ewt

Eastern New t (Noto phthalmus viridescens )
2-438 E
astern N
ewt
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Age at larvae -> eft 2 - 3 mo Illinoi s Smith, 1961 Metamorphosi s (loui sianensi s)/NS
eft -> adult 1 - 3 yrs Gibbons & Sem litsch,
6 mo Healy, 1974 Massachuset ts
(vi ridescens )/in land ponds
Sout h Carolina/ponds 1991
Age at Sexual 3 - 7 years ef t 5 - 6 yrs 4 - 8 Massachuset ts Healy, 1974 Maturity (vi ridescens )/in land
no ef t stage 2 yrs coast al ponds
ponds
Annual A F 54.1 - 59.5 Virginia/mount ain ponds Gill, 1978a Mortality Rates A M 45.8 - 53.1 (%)
Longevi ty A F 1.7 Virginia/mount ain ponds Gill, 1978b (breeding A M 2.1 seasons)
Seasonal Act ivi ty Begi n Peak End Locat ion erence No.
Note
Mating/Laying February - Apr il - May Sout h Carolina Gibbons & Sem litsch, March 1991
April June North Caro lin a Harris et al., 1988
Hatching June Virg in ia Gill, 1978a
late April North Caro lin a Harris et al., 1988
spr ing NS Behl er and Ki ng, 1979
Ref

�
Eastern New t (Noto phthalmus viridescens )
Seasonal Act ivi ty Begi n Peak End Locat ion erence No.
Note
Metamorphosi s June September Sout h Carolina Gibbons & Sem litsch, to eft 1991
mid-August late Novem ber Virginia Gill, 1978a
mid-Jul y August - Sept. early Novem ber New Yor k Hur lbert, 1970
Fall Migration August - Sept. Novem ber Virginia Gill, 1978a (of adults to hibernacul ae)
Spr ing Migration late March late Apr il Virginia Massey , 1990 (of adults to breeding ponds)
Ref
1 "Neonat es" r efers to newts that becom e sexual ly mature in the larval form (i.e., neot eny).
2-439 E
astern N
ewt
2 Est imated assum ing t emperature of 20�C using Equatio n 3-50 (Robinson et al., 1983) and post breeding body weight s f rom (a) Gill ( 1979); (b) Taylor et al. (1988); and (c) Gillis and Breuer (1984). The val ues f or the larvae shoul d be used wi th caut ion because t hese ani mals are smaller than any used t o devel op t he allometric equat ions.
3 See Chapt ers 3 and 4 f or methods of est imating f ood i ngest ion r ates f rom metabol ic rate and di et. 4 Est imated usi ng Equat ion 3-26 (Whitford and Hut chinson, 1967) and post breeding body weight s f rom Gill, 1979. 5 Mean di stance bet ween capt ure and r ecapt ure si tes, suggest ing sm all hom e range si ze.

References (including A ppendix)
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York, NY: Alfred A. Knopf , Inc .
Bellis , E. D. (1968) Summer movemen t of red-spotted newts i n a smal l pond. J. Herpetol. 1: 86-91.
Bennett, S. M. (1970) Homing, dens ity and popula tion dy namics in t he adult n ewt, Notopht halmus virid escen s Rafinesque [Ph.D. disser tat ion] . Hanov er, NH: Dartmout h Colle ge.
Bis hop, S. C. (1941) The salamanders of New York. N. Y. State Mus. Bull. 324; 365 pp.
Brophy , T. E. (1980) Food ha bit s of symp atr ic larval Ambysto ma ti grinum and Notopthalmus virid escen s. J. Herpetol. 14: 1-6.
Burton, T. M. (1976) An analysi s of th e feeding e cology of th e salamande rs (Amphibia , Urodela) of th e Hubba rd Brook Exp erimental Forest, New Hampshire. J. Herpetol. 10: 187-204.
Burton, T. M. (1977) Popula tion esti mates, feeding ha bit s and nut rient and energy relations hips of Notopht halmus v. virid escen s, in Mirror Lake, New Hampshire. Copeia 1977: 139-143.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns: easter n/central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
DeGraaf, R. M.; Rudis , D. D. (1983) Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Gage, S. H. (1891) The life histo ry of th e vermillion- spo tted newt. Am. Nat. 25: 1084-1103.
Gibbons , J. W.; Semlitsch , R. D. (1991) Guide to the rept iles and amphibia ns of th e Savannah River Site. Athens, GA: Universi ty o f Georgia Press.
Gill, D. E. (1978a) The metapopula tion e cology of th e red-spo tted newt, Notopht halmus virid escen s (Rafine sque ). Ecol. Monogr . 48: 145-166.
Gill, D. E. (1978b) Effecti ve popula tion s ize and int erdemic migr ation r ates i n a metapopula tion of th e red-spo tted newt, Notopht halmus virid escen s. Evolut ion 3 2: 839-849.
Gill, D. E. (1979) Densi ty d ependence and homing be havior in adult r ed-spo tted newts Notopht halmus virid escen s (Rafine sque ). Ecology 60: 800-813.
2-440 Easter n Newt

Gillis , J. E.; Breuer, W. J. (1984) A comparison of rates o f evap orative water loss an d tole rance to dehydration in t he red-eft an d newt o f Notopht halmus virid escen s. J. Herpetol. 18: 81-82.
Harris, R. N. (1981) Intrapond hom ing be havior in Notopht halmus virid escen s. J. Herpetol. 15: 355-356.
Harris, R. N. (1987) Densi ty-d ependent paedomorphos is in t he salamander Notopht halmus virid escen s dorsal is. Ecology 68: 705-712.
Harris, R. N. (1989) Ontogenetic ch anges in size and shape of th e facu ltati vely paedom orphic sal amander Notopht halmus virid escen s dorsal is. Copeia 1989: 35-42.
Harris, R. N.; Alford, R. A.; Wilbur , H. M. (1988) Densi ty an d phenology of Notopht halmus virid escen s dorsal is in a natural pond. Herpetologic a 44: 234-242.
Healy, W. R. (1973) Life histo ry var iation a nd t he growth of th e juv enile Notopht halmus virid escen s from Massach usetts. Copeia 1973: 641-647.
Healy, W. R. (1974) Popula tion c ons equences o f al ternative l ife histo ries in Notopht halmus v. virid escen s. Copeia 1974: 221-229.
Healy, W. R. (1975) Terrestr ial acti vi ty an d home range in efts o f Notopht halmus virid escen s. Am. Midl. Nat. 93: 131-138.
Healy, W. R. (1981) Field test o f an HSI model for the red-spotted newt (Notopht halmus virid escen s) for the Habitat Eval uation P rocedur es Group [ unpublis hed mater ial]. For t Collins , CO: U.S. Fish Wildl. Serv.; 21 pp.
Hurlbert, S. H. (1969) The breeding m igr ations and int erhabitat wan dering of th e vermilion- spo tted newt, Notopht halmus virid escen s (Rafine sque ). Ecol. Monogr . 39: 465-488.
Hurlbert, S. H. (1970) The post-l arval migration of th e red-spo tted newt, Notopht halmus virid escen s (Rafine sque ). Copeia 1970: 515-528.
Logie r, E. B. (1952) The frogs, toads and salamanders of easter n Canada. Toronto, Canada: Universi ty o f Toronto Press.
MacNamara, M. C. (1977) Food ha bit s of ter restr ial adult migr ants and i mmatu re red efts o f the red-spotted newt, Notopht halmus virid escen s. Herpetologic a 33: 127-132.
Massey, A. (1990) Notes o n the reproduc tive ecology of r ed-spo tted newts ( Notopht halmus virid escen s). J. Herpetol. 24: 106-107.
McLaughlin, E. W.; Humphr ies, A. A., Jr . (1978) The jelly envelope s and f ertiliza tion of eggs of th e newt, Notopht halmus virid escen s. J. Morphol. 158: 73-90.
2-441 Easter n Newt

Morin, P. J. (1986) Interact ions b etween intraspeci fic co mpeti tion a nd pr edation in a n amphibia n pr edator-prey system. Ecology 67: 713-720.
Mor in, P. J.; Wilbur , H. M.; Harris, R. N. (1983) Salamander predation a nd t he str ucture of experimental communit ies: respon ses o f Notopht halmus and Microcrustacea . Ecology 64: 1430-1436.
Pitki n, R. B. (1983) Annua l cycl e of body size and blood pa rameter s of th e aquatic adult red-spotted newt, Notopht halmus virid escen s. J. Exp . Zool. 226: 372-377.
Ries, K. M.; Bellis , E. D. (1966) Spring f ood ha bit s of th e red-spo tted newt i n Pennsyl vania. Herpetologic a 22: 152-155.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Shure, D. J.; Wilson, L. A.; Hochwender, C. (1989) Predation on a pos omati c efts o f Notopht halmus virid escen s. J. Herpetol. 23: 437-439.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9; 356 pp.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28; pp. 118-120.
Sousa, P. J. (1985) Habitat su itabilit y inde x mode ls : Red-spo tted newt. U.S. Fish Wildl. Serv. Biol. Rep. 82(10.111); 18 pp.
Stefan ski , M.; Gatten , R. E.; Pough, F. H. (1989) Acti vi ty meta bolis m of sal amanders: tole rance to dehydration. J. Herpetol. 23: 45-50.
Taylor, B. E.; Estes, R. A.; Pechmann, J. H.; et al . (1988) Trophic r elations in a temp orary pond: larval salamanders and their microinver tebrate prey. Can. J. Zool. 66: 2191-2198.
Whit ford, W. G.; Hutchins on, V. H. (1967) Body size and metabolic r ate in salamanders. Physiol. Zool. 40: 127-133.
2-442 Easter n Newt

2.3.7. Green Frog (true frog fam ily )
Order Anur a, Family Ranidae. These are typ ical frogs wi th adult s be ing t ruly amphibious , liv ing a t the edge of water bodie s and enter ing t he water to catch prey, flee danger, and s pawn (Behler and King, 1979). This profi le covers medium -sized ranids . The next p rofi le (Sect ion 2 .3.8) covers large ranids .
Selected species
The green frog (Rana clami tans) is usually found ne ar sh allow fr esh water throughout mu ch of easter n North Amer ica. Two subspecies are recognize d: R. c. clami tans (the bronze fr og; ranges from th e Carolina s to nor thern Flor ida, west to easter n Texas, and north along t he Mississippi V alley to the mout h of th e Ohio Riv er) and R. c. melanota (the green frog; ranges fr om so utheaster n Canada to North Carolina , west to Minne sota and Oklahom a but r are in m uch of Illinois and India na, int roduc ed int o Br it ish Colum bia, Washingt on, and Ut ah) (Conant and Collins , 1991).
Body size. The green frog is a medium -sized ranid us ually b etween 5.7 and 8.9 cm snout-to -vent length (SVL) (Conant and Collins , 1991; Martof et al ., 1980). Its g rowing period is pr imarily conf ined to the period b etween mid May and mid Septemb er (Martof, 1956b). Females are usually larger than males (Smi th, 1961). Adult s typic ally weigh between 30 and 70 g (Wells, 1978). Hutchins on et al. (1968) develope d an allo metr ic equation r elating gr een frog s urface ar ea (SA in cm) to body weight (W t in gr ams):
SA = 0.997 Wt0.712.
This equa tion a lso is presented in Chapter 3 as Equation 3-25.
Habitat . Adult green frogs liv e at the margins of permanent or semi permanent shallow water , springs , swamp s, str eams, ponds , and l akes (Wells, 1977). Martof (1953b) found gr een frogs pr imarily to inha bitat th e banks o f str eams. They also can be found among r otting de br is of falle n t rees (Behler and King, 1979; Conant and Collins , 1991). Juveniles prefer shallower aquatic habitats wi th denser vegetat ion t han thos e prefer red by adults (Martof, 1953b). McAlpine and Dilw or th (1989) observed that green frogs inha bit ed aquatic habitats a bout two -thir ds of th e time and t errestr ial habitats th e remaining t ime. Simi larly, Martof (1953b) found t hat the green frog r elies on t errestr ial habitats fo r feeding and aquatic habitats fo r refuge from desiccat ion, temp erature extr emes, and enemies. Ponds used by green frogs ar e usually mo re permanent than thos e used by other anur an species (Pough a nd K amel, 1984).
Food ha bi ts . Adult R. clami tans are ter restr ial feeders among s hor eline veg etation. They co nsume insects, worms, smal l fish, crayfi sh, other crustacean s, newts, spide rs, small f rogs , and m ollu scs. Stewar t and Sandis on (1973) found t hat ter restr ial beetles often are their mo st im por tant food i tem b ut noted that any local ly abunda nt insect a long t he shor eline may b e consumed in large numbers. There is a pronounc ed reduc tion in f ood cons umpt ion dur ing t he breeding pe riod f or bot h males and femal es (Mele, 1980). During the breeding seaso n, males sp end most o f th eir energy defending br eeding t erritories, and
2-443 Green Frog

�
�
femal es exp end their energy produc ing e ggs (W ells , 1977). Fat reserves ac quir ed dur ing the prebreeding pe riod c ompensate fo r reduced food in take d uring t he breeding pe riod (Mele, 1980). Jenssen and Klimstr a (1966) found t hat gr een frogs cons ume most of th eir food in t he spring a nd eat l ittl e during t he wint er. Food eaten in the spring, summer , and fal l consists mo stl y of ter restr ial prey, whereas wi nter food is co mpos ed mostl y of aqua tic prey (Jen ssen and Klimstr a, 1966). Juveniles (sexu ally immatu re frogs) eat about half th e volum e of food a s do a dult s ov er th e cour se of a year (Jenssen and Klimstr a, 1966). Tadpole s are herbiv orous (D eGraaf and Rudis , 1983). Green frogs eat th eir cast ski ns follow ing m olt ing; the cast ing of skin is fr equent dur ing m ids ummer (Hamilton, 1948).
Temperature regulation and daily acti vi ties. Martof (1953b) found t hat the green frog's acti vi ty p eriod v aries by fr og s ize, wi th larger fr ogs being pr imarily noc turnal, smal l frogs being diur nal, and m iddl e-sized f rogs (5 to 7 cm SVL) being e qually acti ve during da y and night .
Hibernation. Adult green frogs overwint er by hibe rnating unde rground or unde rwater from fal l to spring ( Ryan, 1953). Martof (1956a) observed frogs hibe rnating in mud and debris at th e botto m of str eams ap proximatel y 1 m d eep. Jenssen and Klimstr a (1966) noted that adult s us ually hibe rnate in restr icted chambers wi thin r ock pile s or beneath plant debris, while juv enile s are more often found in lo cations wi th access to passing pr ey. The frogs begin emerging w hen the mean daily temp erature is about 4. 4�C and the maxi mum temp erature is about 15. 6�C for 3 to 4 days (Martof, 1953b). Juvenile frogs en ter and e xi t hibe rnation after adult fr ogs (M artof, 1956a).
Breeding acti vi ties and social organization. Green frogs breed from sp ring t hrough the summer , spawning a t night (Smith, 1961; Wells, 1976). Female green frogs stay i n nonbr eeding ha bitat u nti l it is ti me to spawn (Martof, 1956a). In preparation f or breeding, males esta blis h territories near shor e that serve as areas fo r sexu al display and as defende d ov ipos it ion s ites (Wells, 1977). Males esta blis h calling s ites wi thin t heir ter ritories wh ere they attemp t to attr act femal es (Wells, 1977). Females vi si t mal e ter ritories to mate an d lay th eir egg masses. The masses ar e contained in fi lms o f jelly and are deposi ted in emergent, floating, or subm erged vegetation; they hatch in about 3 to 6 days (B ehler and King, 1979; Martof, 1956a; Ryan, 1953). Adult s are solit ary dur ing non-breeding pe riods (Smith, 1956).
Tadpol e and metamo rphos is. In the sout hern part of th eir range, green frog tadpole s metamo rphose into frogs in the same seaso n in which they hatch ed, while in t he northern part, 1 or 2 years pass b efore metamo rphos is (Martof, 1956b). Tadpole s that hatch from egg masses l aid in the spring us ually metamo rphos e that fall, while thos e hatching f rom su mmer -laid eggs typ ical ly overwinter as larvae and metamo rphose th e follow ing s pring ( Pough a nd K amel, 1984). Ryan (1953) found t hat most tadpole s are 2.6 to 3.8 cm SVL at the time of tr ansfo rmation. Thos e that transfo rm in l ate Ju ne or early Ju ly grow r apidly , adding 1 .4 to 2.0 cm SVL in t he fi rst 2 mont hs and 0 .4 to 0.7 cm SVL more before hibe rnation. Tadpole s that transfo rm at appr ox imatel y 3.1 cm SVL may r each between 5.0 and 5.8 cm SVL before hibe rnation ( Ryan, 1953). Newly tr ansfo rmed frogs often move fr om lakes an d ponds wh ere they were tadpole s to shallow str eam banks, usually dur ing pe riods of r ain (Martof, 1953b).
2-444 Green Frog

�
�
�
�
Home range and r esour ces. The species' hom e range inc lude s it s foraging a nd refuge areas in and around a quatic environm ents. Dur ing t he breeding pe riod, the male's hom e range al so inc lude s it s br eeding t erritory (Wells , 1976). Martof (1953b) found t hat roughly 80 pe rcent of adult fr ogs cap tured in t he spring a nd again in t he fall oc cupie d the same h ome ranges.
Popul ation de nsity . Dur ing t he breeding seaso n, green frog densi ties at b reeding ponds can exceed sever al hundr ed indiv idua ls per hectar e (Wells, 1978). Adult mal e frogs space th eir breeding t erritories about 2 to 3 m apart (Martof, 1953a).
Popul ation dy namics. Sexual matu rity i s attai ned in 1 or 2 years after metamo rphos is ; indiv idua ls may r each matu rity at th e end of th e fi rst year but not attemp t to breed unti l the next year (Martof, 1956a,b). Most femal es lay one clutch per year , although s ome may l ay two clutch es, about 3 to 4 weeks ap art (Wells, 1976). In natural popula tions , green frogs can liv e to appr ox imatel y 5 year s of age (Martof, 1956b).
Simi lar species (from general references)
� The river frog (Rana hecksch eri) is slight ly larger th an the green frog (8.0 to 12.0 cm SVL) and is found in swamp s fr om so utheast North Carolina to central Flor ida and s out hern Mississippi.
� The leopard and picker el frogs (Rana pipiens and its r elatives, and Rana palustris ) are medium size d and s trongly spo tted . There are four leopard frogs wh ose ranges are mostl y excl usive fr om each other, but over lap wi th the green frog. The picker el frog has a si mi lar range with gaps in the uppe r midwest an d the southeast.
� The mink fr og (Rana septentrio nalis ) is only slight ly smal ler (4.0 to 7.0 cm) and is found on t he bor ders of ponds and l akes, especially near water lilie s. It ranges from Minne sota to New York, nor th to Labrador .
� The carpenter frog (Rana virg atipes) is about th e same si ze as th e green frog (4.1 to 6.7 cm) an d is cl osely asso ciated wi th sphagnum bogs an d grassl ands. It has a co astal plain range from New Jer sey to Georgia and Flor ida.
The bullf rog and pig f rog are much larger ranid s pecies and are covered in t he next p rofi le (Sect ion 2 .3.8).
General references
Behler and King (1979); Conant and Collins (1991); DeGraaf and Rudis (1983); Martof (1953a, b, 1956a, b); Smith (1956, 1961).
2-445 Green Frog

Green Frog ( Rana clamitans )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion (subspeci es) Reference No.
Body Weight B 49.1 ± 20.0 SD 25.5 - 103.5 New Brunsw ick, Canada McAlpine & Di lworth, 1989 (g)
A breeding 44.0 ± 10.0 SD 27.0 - 66.0 New York (melanot a) Wells, 1978
at 3 New Yor k Pough & Kam el, 1984 1 metamorphosi s
Lengt h A 54 - 102 NS Behl er and Ki ng, 1979 (mm SVL) A M 79.8 ± 8.5 SD 103 maximum s Michi gan Martof , 1956b
A 80.3 ± 8.9 SD 105 maximum J 32.6 28.4 - 36.3 s Michi gan Martof , 1956b
Metabol ic basal : Rate A 8.08 est imated 2 (kcal /kg-d) at 15.8 est imated 3
metamorphosi s
Food Ingest ion 4 Rate (g/g-d)
Surface Ar ea A 17 est imated 5 (cm ) at 2 est imated 62
metamorphosi s
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion (subspeci es)/Habi tat Note
adul ts: New York/lake Stewart & Sandi son, 1973 7 plant material 10.8 Araneae 12.1 (% total vol ume; Coleopt era 32.8 stomach cont ents) Hemiptera 12.9 Hymenopt era 14.4 Diptera 6.8 Ephem eropt era 5.6 Mollu sca 5.4 Lepi dopt era 2.5
B
M
F B
2-446 G
reen F
rog

Green Frog ( Rana clamitans )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion (subspeci es)/Habi tat Note
adul ts: s Illinoi s/swamp, stream Jenssen & Kl imstra, 1966 mineral - - - 2.6 plant 5.7 8.3 4.2 0.5 (% wet vol ume; stomach Pulmonat a 15.7 18.3 6.4 11.0 cont ents) Olig ochaeta 2.1 0.8 2.3 6.4 Amphi poda 1.2 0.1 - 4.6 Isopoda 5.6 1.4 - 4.6 Decapoda - - 4.1 -Ju lio fo rma 7.5 0.3 1.7 -Araneida 2.8 3.4 6.6 7.4 Odonat a 1.6 12.4 5.9 -Orthopt era 0.9 3.0 1.5 -Hemiptera 1.0 7.0 6.1 2.2 Coleopt era 9.6 19.6 15.9 9.1 Lepi dopt era 25.4 7.0 25.1 -Diptera 6.0 5.2 4.5 10.3 Hymenopt era 9.9 6.0 13.5 -Salientia - - 3.9 -
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Home Range A B 0.0065 ± 0.0036 SD 0.0020 - 0.020 ha s Michi gan (melanot a)/ Martof , 1953b 8 Size nonbr eeding ha shal low wat er
A M breeding 4.0 - 6.0 open near shor e areas
A M breeding 1.0 - 1.5 densel y veget ated
meters shorelin e: New York (melanot a)/ Wells, 1977
meters shorelin e: New York (melanot a)/ Wells, 1977
of ponds
nearshor e areas of ponds
Popul ation A M 476 New Yor k (melanot a)/ Wells, 1978 9 Densi ty A F 567 artificial pond (N/ha)
2-447 G
reen F
rog
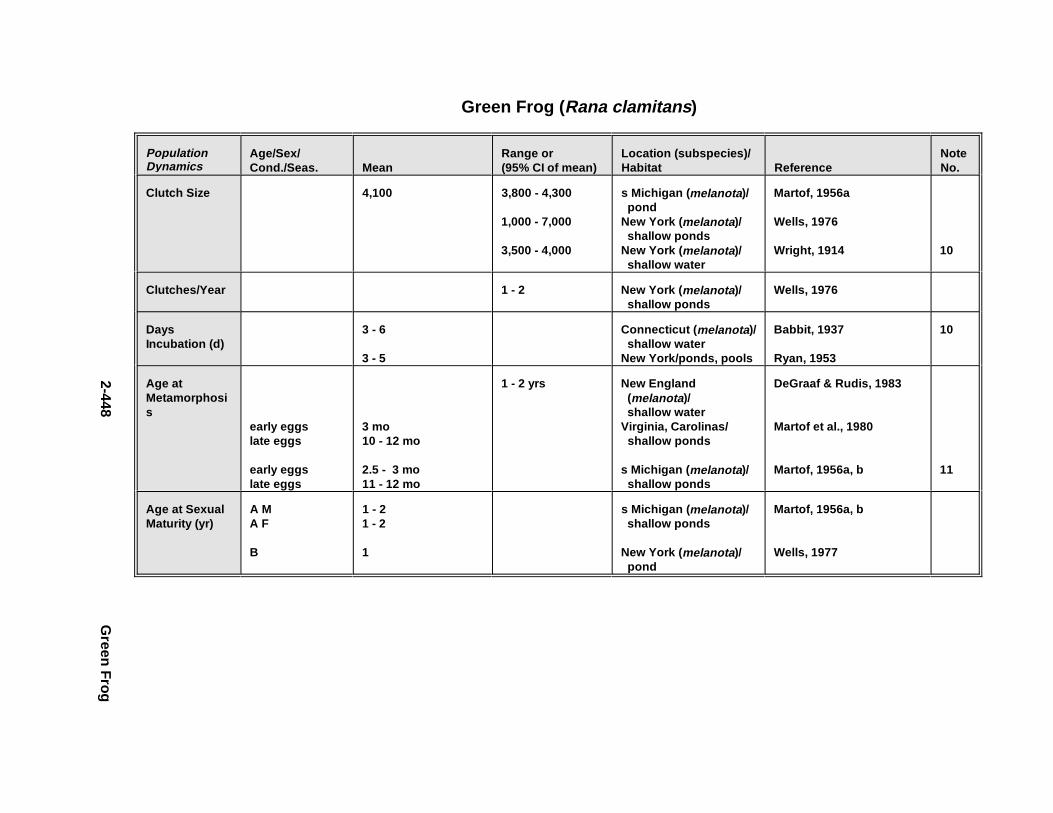
Green Frog ( Rana clamitans )
Population Dynamics
Age/Sex/ Range or Locat ion (subspeci es)/ Note Cond. /Seas. Mean (95% CI of mean) Habi tat Reference No.
Clutch Si ze 4,100 3,800 - 4,300 s Michi gan (melanot a)/ Martof , 1956a
1,000 - 7,000 New York (melanot a)/ Wells, 1976
3,500 - 4,000 New York (melanot a)/ Wright , 1914 10
pond
shal low ponds
shallo w water
Clutches/ Year 1 - 2 New York (melanot a)/ Wells, 1976 shal low ponds
Days 3 - 6 Connect icut (melanot a)/ Babbi t, 1937 10 Incubat ion (d) shallo w water
3 - 5 New Yor k/ponds, pool s Ryan, 1953
Age at 1 - 2 yrs New Engl and DeGraaf & Rudi s, 1983 Metamorphosi (melanot a)/ s shallo w water
early eggs 3 mo Virg in ia, Caro lin as/ Marto f et al., 1980 late eggs 10 - 12 mo shal low ponds
early eggs 2.5 - 3 mo s Michi gan (melanot a)/ Martof , 1956a, b 11 late eggs 11 - 12 mo shal low ponds
Age at Sexual A M 1 - 2 s Michi gan (melanot a)/ Martof , 1956a, b Maturity (yr) A F 1 - 2 shal low ponds
B New York (melanot a)/ Wells, 1977 pond
1
2-448 G
reen F
rog
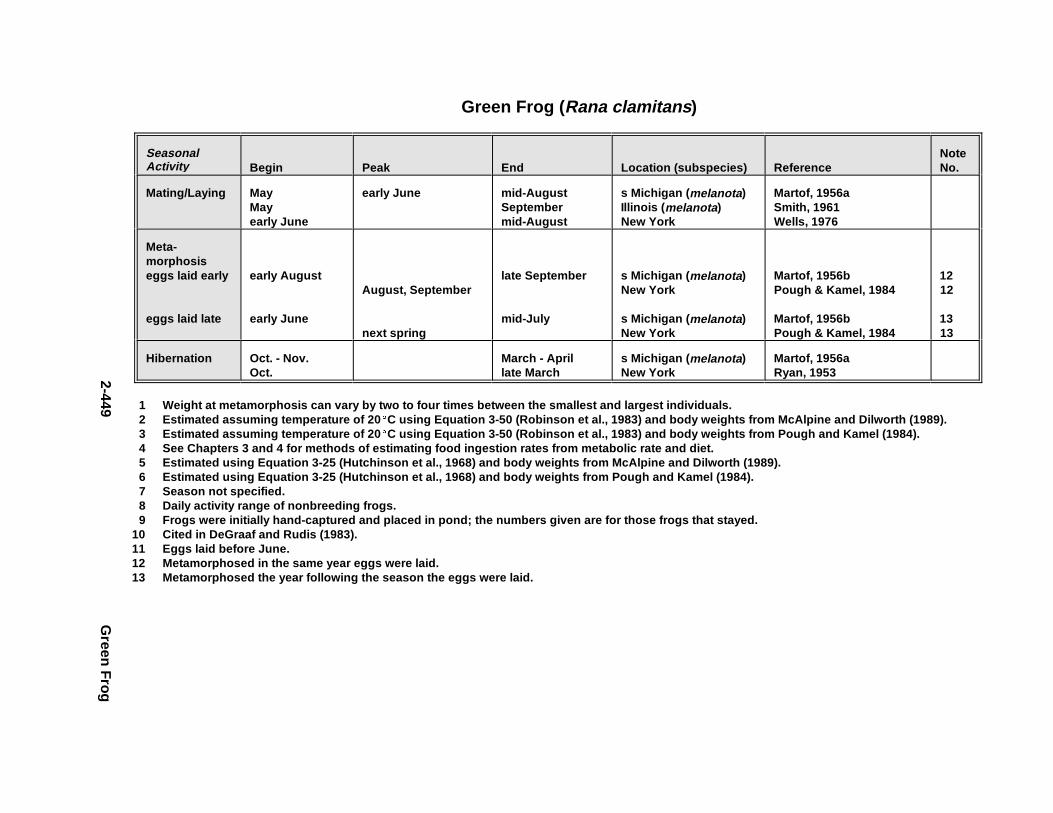
�
�
Green Frog ( Rana clamitans )
Seasonal Act ivi ty Begi n Peak End Locat ion (subspeci es) Reference No.
Note
Mating/Laying May early June mid-August s Michi gan (melanot a) Martof, 1956a May September Illinoi s (melanot a) Smith, 1961 early June mid-August New Yor k Wells, 1976
Metamorphosi s eggs l aid early early August late September s Michi gan (melanot a) Martof, 1956b 12
eggs laid late early June mid-July s Michigan (melanot a) Martof, 1956b 13
August , September New Yor k Pough & Kam el, 1984 12
next spr ing New Yor k Pough & Kam el, 1984 13
Hibernation Oct. - Nov. March - Apr il s Michi gan (melanot a) Martof, 1956a Oct. late March New York Ryan, 1953 2-449
Green
Fro
g
1 Weight at metamorphosi s can var y by two to four times bet ween the smallest and l argest indi vidual s. 2 Est imated assum ing t emperature of 20�C using Equatio n 3-50 (Robinson et al., 1983) and body weight s f rom McAlpine and Di lworth (1989).3 Est imated assum ing t emperature of 20�C using Equatio n 3-50 (Robinson et al., 1983) and body weight s f rom Pough and Kam el (1984).4 See Chapt ers 3 and 4 f or methods of est imating f ood i ngest ion r ates f rom metabol ic rate and di et.5 Estimated using Equatio n 3-25 (Hutchinson et al., 1968) and body weight s f rom McAlpine and Di lworth (1989).6 Estimated using Equatio n 3-25 (Hutchinson et al., 1968) and body weight s f rom Pough and Kam el (1984).7 Season not speci fied.8 Daily act ivity range of nonbr eeding f rogs.9 Frogs wer e initially hand- capt ured and pl aced i n pond; the num bers given ar e for those f rogs t hat stayed.
10 Cited in DeGraaf and Rudi s (1983).11 Eggs l aid before June.12 Metamorphosed i n the same year eggs w ere laid.13 Metamorphosed t he year following t he season t he eggs w ere laid.

References (including A ppendix)
Babbitt, L. H. (1937) The amphibia of Conn ecti cut. Hartfo rd, CT: State Geo l. and Nat. Hist. Surv.; Bull. No. 57. 9-50
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York: Alfred A. Knopf , Inc .
Bush, F. M. (1959) Foods of so me Kentucky h erpti les. Herpetologic a 15: 73-77.
Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns. Bos ton, MA: Hought on Mif flin Co.
DeGraaf, R. M.; Rudis , D.D. (1983) Green frog. Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Hamilton, W. J., Jr . (1948) The food a nd feeding be havior of th e green frog, Rana clami tans (Latreille), in New York state. Copeia 1948: 203-207.
Hutchins on, V. H.; Whit ford, W. G.; Kohl, M. (1968) Relation of body size and s urface ar ea to gas exch ange in anurans. Physiol. Zool. 41: 65-85.
Jenssen , T. A.; Klimstr a, W. D. (1966) Food ha bit s of th e green frog, Rana clami tans, in sout hern Illinois . Amer . Midl. Nat. 76: 169-182.
Martof, B. S. (1953a) Territoriality i n the green frog, Rana clami tans. Ecology 34: 165-174.
Martof, B. S. (1953b) Home range and movemen ts o f th e green frog, Rana clami tans. Ecology 34: 529-543.
Martof, B. S. (1956a) Facto rs influencing s ize and c ompos it ion of popula tions of Rana clami tans. Am. Midl. Nat. 56: 224-245.
Martof, B. S. (1956b) Growth and development of th e green frog, Rana clami tans, unde r natural condit ions . Am. Midl. Nat. 55: 101-117.
Martof, B. S.; Palmer , W. M.; Bailey, J. R.; et al . (1980). Amphibia ns and r ept iles of th e Carolina s and Virginia . Chapel Hill, NC: Univ ersi ty of Nor th Carolina Press.
McAlpine , D. F.; Dilw or th, T. G. (1989) Microhabitat an d prey si ze among t hree species of Rana (Anura: Ranidae) symp atr ic in easter n Canada. Can. J. Zool. 67: 2241-2252.
Mele, J. A. (1980) The role of lipids in s torage and ut iliza tion of en ergy fo r reproduc tion a nd maintenance in the green frog, Rana clami tans [Ph.D. disser tat ion] . New Br uns wick, NJ: Rutgers Univ ersi ty.
2-450 Green Frog

Pope, C. H. (1947) Amphibia ns and r ept iles of th e Chic ago area. Chic ago, IL: Chic ago Nat. Hist. Mus. Press.
Pough, F. H.; Kamel, S. (1984) Post-metamo rphic phy siologic al change in r elation t o anur an lif e his tories. Oecologia 65: 138-144.
Robins on, F. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Ryan, R. A. (1953) Growth rates o f so me ranids unde r natural condit ions . Copeia 1953:73-80.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28.
Stewar t, M. M.; Sandis on, P. (1973) Comparative food ha bit s of symp atr ic mi nk fr ogs, bullf rogs , and gr een frogs . J. Herpetol. 6: 241-244.
Wells, K. D. (1976) Mult iple egg c lut ches in t he green frog (Rana clami tans). Herpetologic a 32: 85-87.
Wells, K. D. (1977) Territoriality an d male mat ing s uccess i n the green frog (Rana clami tans). Ecology 58: 750-762.
Wells, K. D. (1978) Territoriality i n the green frog (Rana clami tans): vocaliza tions an d agonis tic behaviour . Anim . Behav. 26: 1051-1063.
Wright , A. H. (1914) North Amer ican anura: life histo ries of th e anurans of Ithaca, New York. Washingt on, DC: Carnegie Insti tute; Publ. No. 197.
2-451 Green Frog

2.3.8. Bullfrog (true frog fam ily )
Order Anur a, Family Ranidae. These are typ ical frogs wi th adult s be ing t ruly amphibious . They tend t o liv e at the edge of water bodie s and enter th e water to catch prey, fl ee danger, and s pawn (Behler and King, 1979). This profi le covers large ranids . Medium -sized ranids ar e covered in t he previous prof ile (Sect ion 2 .3.7).
Selected species
The bullf rog' s (Rana catesb eiana) natural range includes th e easter n and central United States an d southeaster n Canada; however , it has been introduced in many areas in the wester n United States an d other parts o f North Amer ica. It is co ntinuing t o expand it s range, apparently at th e expense of sever al native sp ecies in many locat ions (Bur y and Whelan, 1984). There are no subspecies fo r the bullf rog.
Body size. The bullf rog is th e largest North Amer ican ranid. Adult s us ually r ange between 9 and 15 cm i n length from snout-to -vent length (SVL) and excep tiona l indiv idua ls can reach one half k ilogr am or more in w eight (Cona nt and Collins , 1991; Durham and Bennett, 1963). Males are usually smal ler than femal es (Smi th, 1961). Frogs exhibit indeterminate growth , and bullf rogs cont inue to inc rease in size for at least 6 year s after metamo rphos is (Dur ham and Be nnett, 1963; Howard, 1981a). Hutchins on et al. (1968) develope d an allo metr ic equation r elating bullf rog s urface ar ea (SA in cm) to body weight (Wt in grams):
SA = 0.953 Wt0.725.
This equa tion a lso is presented in Chapter 3 as Equation 3-24.
Habitat . Adult bullf rogs liv e at the edges of ponds , lakes, and slow-mo ving s treams large enough t o avoid c rowding a nd w ith suffi cient vegetation t o prov ide easi ly access ible cover (B ehler and King, 1979). Smal l str eams ar e used when better habitat i s lacki ng (Conant and Collins , 1991). Bullf rogs r equir e permanent bodie s of water , because th e tadpole s generally r equir e 1 or more years to develop pr ior to metamo rphos is (How ard, 1981b). Smal l frogs favo r areas of ver y sh allow water where short grasses o r other vegetat ion or debr is o ffer cover (Durham and Bennett, 1963). Larger bullf rogs seem to avoid s uch areas (Durham and Bennett, 1963). Tadpole s tend t o congr egate around gr een plants (Jaeg er and Hailman, 1976, ci ted in Bury and Whelan, 1984).
Food ha bi ts . Adult R. catesb eiana are indis cr iminate and aggressi ve predators, feeding a t the edge of th e water and among water weeds on any avai lable smal l animals, inc luding in sects, crayfi sh, other frogs and tadpole s, minnow s, sn ails , young t urtles, and occas iona lly smal l birds, smal l mammal s, and young s nakes (B ehler and King, 1979; DeGraaf and Rudis , 1983; Korsch gen and Baskett, 1963). Bullf rogs o ften focus on local ly abunda nt foods (e. g., cicadas, meadow vo les) (Korsch gen and Baskett, 1963). Crustacean s and insects p robably make u p the bulk of th e die t in m ost areas (Carpenter and Morrison, 1973; Fulk and W hitaker , 1968; Smith, 1961; Tyler and Hoesten bach, 1979). Bullf rog t adpole s cons ume pr imarily aqua tic pla nt mater ial and some inver tebrates,
2-453 Bullf rog

�
�
�
�
�
�
� �
�
�
�
but also scaven ge dead fish and eat l ive or dead tadpole s and eggs (Bur y and Whelan, 1984; Ehrlich, 1979).
Temperature regulation and daily acti vi ties. Bullf rogs fo rage by day (Behler and King, 1979). They th ermoregulate behaviorally by posi tioning t hemsel ves r elative to the sun a nd by en ter ing or leaving t he water (Lilly whit e, 1970). In one stu dy, body temp eratures measu red in bullf rogs dur ing t heir nor mal daily acti vi ties aver aged 30�C and ranged from 26 to 33�C (Lilly whit e, 1970). At night , their body temp eratures wer e found t o range between 14.4 and 24.9�C (Lilly whit e, 1970). Tadpole s also select r elatively war m areas, 24 to 30�C (Bury and Whelan, 1984). Despite th is narrow range of temp eratures in whic h bullf rogs nor mally maint ain t hemsel ves, they are not immobilize d by mode rately lower temp eratures (Lilly whit e, 1970). The metabolic r ate of bullf rogs inc reases wi th inc reasing body temp erature. Between 15 and 25�C, the Q10 for oxygen cons umpt ion is 1.87; between 25 and 33�C, the Q10 is 2.41 (Burggren et al ., 1983).
Hibernation. Most bullf rogs hibe rnate in mud and leaves unde r water beginning in the fall, but so me bullf rogs in t he sout hern states may b e acti ve year round ( Bur y and Whelan, 1984). They emer ge someti me in the spring, usually wh en air temp eratures are about 19 to 24�C and water temp eratures are at least 13 to 14�C (Wright , 1914; Willis et al., 1956). Bullf rogs emerge from hibe rnation l ater than other ranid s pecies (Ryan, 1953).
Breeding acti vi ties and social organization. Bullf rogs sp awn at night clos e to shorelines in areas sh eltered by sh rubs (Raney, 1940, ci ted in DeGraaf and Rudis , 1983). The timing a nd dur ation of th e breeding seaso n var ies depending on t he location. In the southern states, the breeding seaso n exten ds fr om sp ring t o fall, whereas in the northern states, it is restr icted to late sp ring a nd s ummer (Behler and King, 1979). Males ten d to be ter ritorial dur ing t he breeding seaso n, defending t heir calling po sts an d ovipos it ion s ites (i.e., submerged vegetat ion ne ar shor e) (Howard, 1978b; Ryan, 1980). Female visi ts to the pond t end t o be br ief and s por adic (Emlen, 1976). Some mal es mate wi th sever al femal es whereas others, usually younge r and smal ler males, may n ot breed at al l in a given year (DeGraaf and Rudis , 1983). Females attach their eggs, contained in floating f ilm s of jelly , to submerged vegetat ion ( Behler and King, 1979). Adults ar e otherwise rather solitary occupy ing t heir own part of a str eam or pond (Smith, 1961).
Tadpol e and metamo rphos is. Eggs hatch in 3 to 5 days (Clarkso n and DeVos, 1986; Smith, 1956). Temperatures above 32�C have been shown to cause abnor malit ies in tadpole s and abov e 35.9�C to kill e mbryos (How ard, 1978a). Tadpole growth rates i ncrease wi th inc reasing ox ygen levels, food availa bilit y, and water temp erature (Bury and Whelan, 1984). Tadpole gill v entila tion a t 20�C can generate a branchial water flow of almost 0.3 ml/g-min (Burggren and West, 1982). Metamo rphos is fr om a tadpole to a frog c an occur as early as 4 to 6 months in the southern parts o f i ts r ange; however , most ta dpole s metamo rphose fr om 1 to 3 years after hatching, depending on la ti tude and temp erature (DeGraaf and Rudis , 1983; Martof et al ., 1980).
Home range and r esour ces. The species' hom e range inc lude s it s foraging a reas and r efuge s in a nd around a quatic environm ents. Home range size decreases wi th inc reasing bullf rog de nsity, and m ales tend t o use larger hom e ranges than femal es (Currie and Be llis , 1969). Bullf rogs tend t o stay i n the same pools th roughout th e summer months
2-454 Bullf rog

�
if th e water level is stab le (Raney, 1940, ci ted in DeGraaf and Rudis , 1983). During t he breeding seaso n, adult mal es establis h territories that they defend against co nspeci fic males (Eml en, 1968). During t he non- breeding seaso n, Currie and Bellis (1969) found no evidence of ter ritorial defense. Males often do not return to the same pond t he follow ing spring ( Durham and Be nnett, 1963).
Popul ation de nsity . Dur ing t he breeding seaso n, each breeding m ale may d efend a few meter s of sh oreline (Cur rie and Be llis , 1969; Emlen, 1968). The densi ties of femal es and non- breeding m ales vary wi th time of day and seaso n and are diffi cult to esti mate. Tadpole s can be present local ly in extr emely high de nsit ies (Cecil a nd J ust, 1979).
Popul ation dy namics. Sexual matu rity i s attai ned in about 1 to 3 years after metamo rphos is , depending on la ti tude (How ard, 1978a; Raney and Ingr am, 1941, ci ted in Bury and Whelan, 1984). Only femal es th at are at least 2 year s past metamo rphos is mate dur ing t he early breeding seaso n; males and femal es 1 year past metamo rphos is may breed dur ing t he later breeding pe riods (How ard, 1978a, 1981b). Also, some olde r femal es have been observed to mate an d to lay a sec ond c lut ch dur ing t he later breeding pe riod (Howard, 1978a). Willis et al. (1956) esti mated the minim um breeding le ngt h for femal es in Misso uri to be 123 to 125 mm SVL . Mortal ity o f tadpole s is high ( Cecil a nd J ust, 1979), and adult fr ogs ar e unlik ely to liv e beyond 5 to 8 years postmetamo rphos is (How ard, 1978b). In some areas, snapping t urtles may b e respons ible fo r a large compone nt of adult bullf rog mortal ity (Howard, 1981a).
Simi lar species (from general references)
� The pig f rog (Rana grylio ) is smalle r th an the bullf rog (8 to 14 cm) an d is found in s out h Sout h Carolina to sout h Flor ida and s out h Texas.
The remaining r anid s pecies are more simila r in s ize to the green (or bronze) frog. See Sect ion 2 .3.7 for a descr iption of th ese fr ogs.
General references
Behler and King (1979); Bury and Whelan (1984); Conant and Collins (1991); DeGraaf and Rudis (1983); Smith (1961).
2-455 Bullf rog
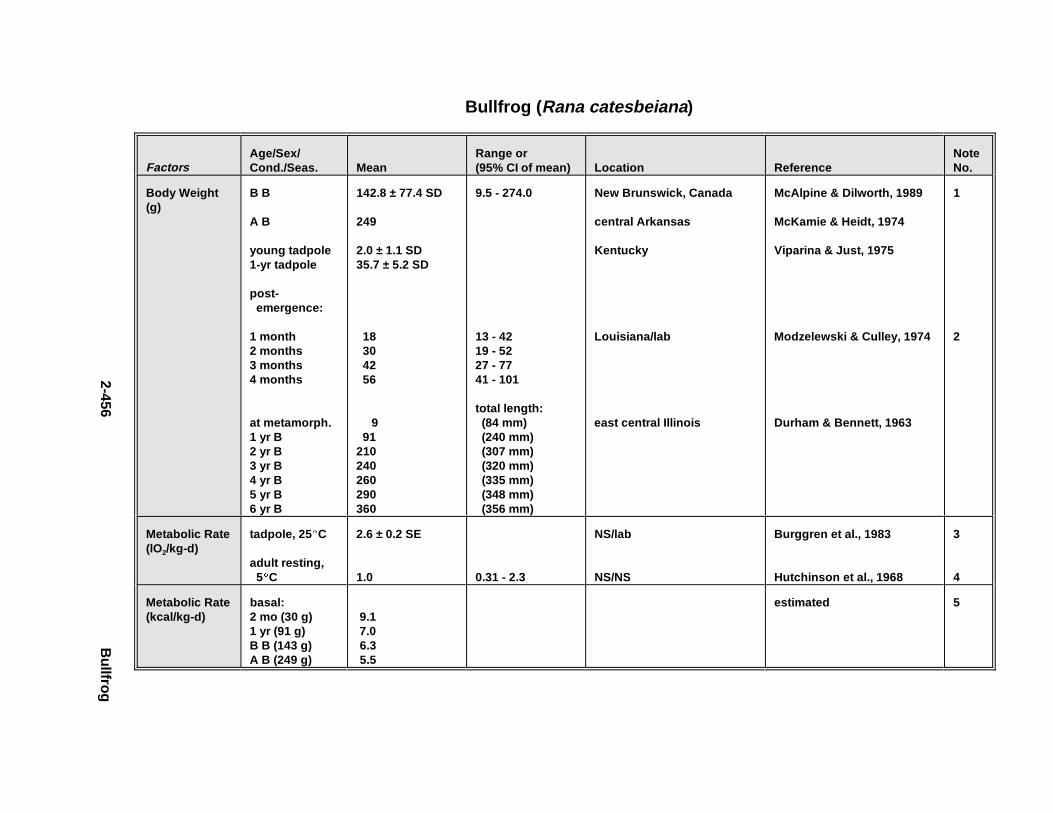
Bullfrog ( Rana catesbeiana )
2-456 B
ullfrog
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Body Weight B B 142.8 ± 77.4 SD 9.5 - 274.0 New Brunsw ick, Canada McAlpine & Di lworth, 1989 1 (g)
A B 249 cent ral Arkansas McKamie & Heidt, 1974
young t adpol e 2.0 ± 1.1 SD Kent ucky Vipar ina & Just , 1975 1-yr tadpol e 35.7 ± 5.2 SD
post -emergence:
1 mont h 18 13 - 42 Loui siana/lab Modzelewski & Cul ley, 1974 2 2 mont hs 30 19 - 52 3 mont hs 42 27 - 77 4 mont hs 56 41 - 101
at metamorph. 9 (84 mm) east cent ral Illinoi s Durham & Bennet t, 1963 1 yr B 91 (240 mm) 2 yr B 210 (307 mm) 3 yr B 240 (320 mm) 4 yr B 260 (335 mm) 5 yr B 290 (348 mm) 6 yr B 360 (356 mm)
total lengt h:
Metabol ic Rate tadpol e, 25��C 2.6 ± 0.2 SE NS/lab Burggren et al., 1983 3 (lO /kg-d)2
adul t rest ing, 5��C 1.0 0.31 - 2.3 NS/NS Hutchinson et al., 1968 4
Metabol ic Rate basal : est imated 5 (kcal /kg-d) 2 mo (30 g) 9.1
1 yr (91 g) 7.0 B B (143 g) 6.3 A B (249 g) 5.5

Bullfrog ( Rana catesbeiana )
Factors Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion Reference No.
Food Ingest ion (13 - 42 g) 0.071 Loui siana (24 - 27��C) Modzelewski & Cul ley, 1974 Rate (g/g-d) (18 - 52 g) 0.059
(28 - 77 g) 0.040 (40 - 100 g) 0.033
Surface Ar ea 2 mo (30 g) 11 est imated 6 (cm ) 1 yr (91 g) 252
B B (143 g) 35 A B (249 g) 52
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
adul ts: Kent ucky /NS Bush, 1959 Decapoda- Astacidae 47.7 Lepi dopt era 19.0 (% wet vol ume; stomach Coleopt era 16.0 cont ents) (Lampryidae) (5.8) (Chrysomelid ae) (5.8) (Carabidae) (4.1) Pulmonat a-Zoni tidae 8.3 Chi lipoda 7.7 sand, rock, gravel 1.2
adul ts: New Yor k/mount ain lake Stewart & Sandi son, 1973 plant 19.7 animal 65.2 (% vol ume; stomach (Odonat a) (8.8) cont ents) (Coleopt era) (15.8) (Hemiptera) (0.5) (Hymenopt era) (2.2) (Amphi bia) (26.4) unaccount ed 15.1
2-457 B
ullfrog

Bullfrog ( Rana catesbeiana )
Dietary Compositio n Spr ing Summer Fall Winter (measur e) Reference No.
Locat ion/Habi tat Note
adul ts: Missour i/bait minnow pond Corse & Metter, 1980 frogs 35 33 39 tadpol es 8 11 0 (num ber of items found; shi ners 305 157 25 stomach cont ents) other fish 7 2 5 Gastropoda 55 70 26 crayfish 22 162 18 other crust acea 71 42 47 Arachni da 3 23 3 Coleopt era (adul t) 31 33 15 Diptera (larvae) 2 7 0 Hemiptera 41 43 16
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Home Range A M nonbr eed 2.9 0.76 - 11.3 Ontario, Canada/pond Currie & Bellis , 1969 Size (m radius) A F nonbr eed 2.4 0.61 - 10.2
A M territory 2.7 Michigan/pond Emlen, 1968 7
Popul ation B B (1960) 1,376 Ontario, Canada/pond Currie & Bellis , 1969 Densi ty B B (1961) 892 (N/ha)
tadpol es: Kent ucky /pond Ceci l & Just , 1979 Novem ber 130,000 March 69,000 May 16,000
Clutch Si ze 10,000 - 20,000 Kansas/ NS Smith, 1956 7,360 ± 741.7 SE New Jer sey/pond Ryan, 1980
Clutches/ Year 93% of F 1 Michi gan/pond Emlen, 1977 7% of F 2
Days to 2 - 4 Arizona, California/river Clarkson & DeVos, 1986 Hatching 4 - 5 Kansas/ NS Smith, 1956
2-458 B
ullfrog

Bullfrog ( Rana catesbeiana )
Population Dynamics
Age/Sex/ Range or Note Cond. /Seas. Mean (95% CI of mean) Locat ion/Habi tat Reference No.
Age at B 1 yr Caro lin as, Virg in ia/NS Marto f et al., 1980 Metamor- B 1 - 2 yr Michi gan/pond Collin s, 1979 phosi s B 2 - 3 yr New Yor k/NS Ryan, 1953
B 3 yr Nova Scot ia, Canada/NS Bleakney , 1952 8
Age at Sexual M 1 yr after metam. Michi gan/pond Howar d, 1978a Maturity F 1 - 2 yr afte r
B New York/NS Ryan, 1953 metam.
1 - 2 yr after metam.
Annual A M 1 - 2 yr 58 Michi gan/pond Howar d, 1984 Mortality Rates A M 2 - 3 yr 58 (%) A M 3 - 4 yr 48
Mortality Rates tadpol es (to 85.5 82.4 - 88.2 Kentucky /shal low ponds Ceci l & Just , 1979 (%) metamorph.)
A M 4 - 5 yr 77
Longevi ty A B up t o 5 - 8 Michigan/ponds Howar d, 1978b (yr)
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Mating/Laying February October sout hern range i n N America Behler & Ki ng, 1979 Apr il May late June California, Ar izona Clarkson & DeVos, 1986 May late June August Missour i Willis et al., 1956 late May July July nor thern range i n N America DeGraaf & Rudi s, 1983;
Behl er & Ki ng, 1979
Metamor- August (1st clutch) October California, Ar izona Clarkson & DeVos, 1986 phosi s March (2nd cl utch) Apr il California, Ar izona Clarkson & DeVos, 1986
June late June-Aug. early October Missouri Willis et al., 1956 Jul y Sept., October New York Ryan, 1953
2-459 B
ullfrog

�
�
Bullfrog ( Rana catesbeiana )
Seasonal Act ivi ty Begi n Peak End Locat ion Reference No.
Note
Hibernation late October late March east cent ral Illinoi s Durham & Bennet t, 1963 mid-October March Missouri Willis et al., 1956
1 Mean snout-to -vent length (SVL) of fro gs was 98 mm SVL and the range was 45 to 128 mm SVL.2 Age post metamorphosi s; maintained at a temperature of 24 to 27�C and f ed mosqui tofish, cr icket s, and ear thworms.3 Restrained, cannul ated; weight 5.7 g.4 Mean weight of frogs w as 74.8 g.5 Est imated assum ing t emperature of 20�C using Equatio n 3-50 (Robinson et al., 1983). Body weight s (1) for 2-mont h post metamorphosi s frog
from Modzelewski and Cul ley (1974); (2) for a 1-year post metamorphosi s frog f rom Durham and Bennet t (1963), Farrar and Dupr e (1983); (3) for bot h juveni les and adul ts of bot h sexes, McAlpine and Di lworth (1989); and (4) for adul ts of bot h sexes, McKamie and Hei dt (1974).
6 Estimated using Equatio n 3-24 (Hutchinson et al., 1968) and body weight s as descr ibed in not e 5. 7 Based on aver age di stance bet ween frogs. 8 Cited in Bur y and Whelan (1984). 2-460
Bullfro
g

References (including A ppendix)
Behler, J. L.; King, F. W. (1979) The Audubon S ociety fi eld guide to Nor th Amer ican rept iles and amphibia ns. New York, NY: Alfred A. Knopf , Inc .
Bleakney, J. S. (1952) The amphibia ns and r ept iles of Nova Scotia. Can. Field-Nat. 66: 125-129.
Brooks, G. R., Jr . (1964) An analysi s of th e food ha bit s of th e bullf rog by body size , sex, month, and habitat. Va. J. Sci . 15: 173-186.
Bruneau, M.; Magnin, E. (1980) Croissan ce, nutr ition, et reproduc tion de s oua ouarons Rana catesb eiana Shaw (Amphibia A nur a) des Laurentide s au nor d de Mont real. Can J . Zool. 58: 175-183.
Burggren, W. W.; West, N. H. (1982) Changing r espir atory im por tance of gills , lungs an d skin dur ing metamo rphos is in t he bullf rog Rana catesb eiana. Physiol. Zool. 56: 263-273.
Bur ggr en, W. W.; Feder, M. E.; Pinde r, A. W. (1983) Temperature and the balance between aerial and aquatic respir ation in la rvae of Rana berlandi eri and Rana catesb eiana. Physiol. Zool. 56: 263-273.
Bury, R. B.; Whelan, J. A. (1984) Ecology and m anagement of th e bullf rog. U.S. Fish Wildl. Serv. Resour . Publ. No. 155; 23 pp.
Bush, F. M. (1959) Foods of so me Kentucky h erpti les. Herpetologic a 15: 73-77.
Carpenter , H. L.; Morrison, E. O. (1973) Feeding be havior of th e bullf rog, Rana catesb eiana, in north central Texas. Bios 44: 188-193.
Ceci l, S. G.; Just, J. J. (1979) Survival rate, popula tion de nsity and d evelopment of a naturally occurring a nur an larvae (Rana catesb eiana). Copeia 1979: 447-453.
Clarkso n, R. W.; DeVos, J. C., Jr . (1986) The bullf rog, Rana catesb eiana Shaw, in the lower Color ado Riv er, Arizona -Calif ornia . J. Herpetol. 20: 42-49.
Cohen, N. W.; Howard, W. E. (1958) Bullf rog f ood a nd gr owth at the San Joaquin Experimental Range, Calif ornia . Copeia 1958: 223-225.
Collins , J. P. (1975) A comparative stu dy of l ife histo ry str ategies in a communit y of fr ogs [Ph.D. disser tat ion] . Ann A rbor , MI: Univ ersi ty of Michiga n.
Collins , J. P. (1979) Intrapopula tion v ariation in t he body size at metamo rphos is and t iming of metamo rphos is in t he bullf rog. Ecology 60: 738-749.
2-461 Bullf rog

Conant , R.; Collins , J. T. (1991) A field guide to rept iles and amphibia ns - easter n and central Nor th Amer ica. Bos ton, MA: Hought on Mif flin Co.
Corse, W. A.; Metter , D. A. (1980) Econom ics, adult feeding a nd la rval growth of Rana catesb eiana on a fish hatch ery. J. Herpetol. 14: 231-238.
Cur rie, W.; Bellis , E. D. (1969) Home range and movemen ts o f th e bullf rog, Rana catesb eiana (Shaw), in an Ontar io pond. Copeia 1969: 688-692.
DeGraaf, R. M.; Rudis , D. D. (1983) Amphibia ns and r ept iles of New Engla nd. Amherst, MA: Universi ty o f Massach usetts Pr ess.
Dowe, B. J. (1979) The effect o f ti me of ovipos it ion a nd m icroenvironm ent on gr owth of larval bullf rogs ( Rana catesb eiana) in A rizona [master 's th esis]. Tempe, AZ: Arizona State Universi ty.
Durham, L.; Bennett, G. W. (1963) Age, growth , and homing in t he bullf rog. J. Wildl. Manage. 27: 107-123.
Ehrlich, D. (1979) Predation by bullf rog t adpole s (Rana catesb eiana) on eggs and ne wly hatch ed larvae of th e plains leopard frog (Rana blairi ). Bull. Md. Herpetol. Soc. 15: 25-26.
Emlen, S. T. (1968) Territoriality i n the bullf rog, Rana catesb eiana. Copeia 1968: 240-243.
Emlen, S. T. (1976) Lek organization a nd mating s trategie s of th e bullf rog. Behav. Ecol. Sociobiol. 1: 283-313.
Emlen, S. T. (1977) "Double clut ching" a nd it s pos sible signif icance in t he bullf rog. Copeia 1977: 749-751.
Farrar, E. S.; Dupr e, R. K. (1983) The role of diet in glyco gen sto rage by juvenile bullf rogs pr ior to overwint ering. Comp. Bioc hem. Physiol. A: Comp. Physiol. 75: 255-260.
Frost, S. W. (1935) The food of Rana catesb eiana Shaw. Copeia 1935: 15-18.
Fulk , F. D.; Whitaker , J. O., Jr . (1968) The food of Rana catesb eiana in three habitats i n Owen Count y, India na. India na Acad. Sci . 78: 491-496.
George, I. D. (1940) A stu dy of th e bullf rog, Rana catesb eiana Shaw, at Ba ton Rouge , Louis iana [Ph .D. disser tat ion] . Ann A rbor , MI: Univ ersi ty of Michiga n.
Gibbons , J. W.; Semlitsch , R. D. (1991) Guide to the rept iles and amphibia ns of th e Savannah River Site. Athens, GA: The Universi ty o f Georgia Press.
Glass, M. L.; Burggren, W. W.; Johansen, K. (1981) Pulmonary diffu sing c apaci ty of th e bullf rog (Rana catesb eiana). Acta Ph ys iol. Scand. 113: 485-490.
2-462 Bullf rog

Hammer , D. A.; Linde r, R. L. (1971) Bullf rog f ood ha bit s on a water fowl produc tion a rea in South Dakota. Proc. SD Acad. Sci . 50: 216-219.
Howard, R. D. (1978a) The influence of mal e-defended ovipos it ion s ites o n early emb ryo mortalit y in bullf rogs . Ecology 59: 789-798.
Howard, R. D. (1978b) The evolut ion of mating s trategie s in bullf rogs , Rana catesb eiana. Evolut ion 3 2: 850-871.
Howard, R. D. (1981a) Sexual dimorphis m in bullf rogs . Ecology 62: 303-310.
Howard, R. D. (1981b) Male age-size distr ibut ion a nd m ale mating s uccess i n bullf rogs . In: Alexande r, R. D.; Tink le, D. W., ed. Natural selection a nd s ocial behavior : recent research and new th eory; pp. 61-77.
Howard, R. D. (1984) Alternative mat ing be havior s of young m ale bullf rogs . Am. Zool. 24: 397-406.
Hutchins on, V. H.; Whit ford, W. G.; Kohl, M. (1968) Relation of body size and s urface ar ea to gas exch ange in anurans. Physiol. Zool. 41: 65-85.
Jaeger, R. G.; Hailman, J. P. (1976) Ontogenetic sh ift o f sp ectr al phot otacti c preferences i n anur an tadpole s. J. Comp. Physiol. Psyc hol. 90: 930-945.
Korsch gen, L. J.; Baskett, T. S. (1963) Foods of impoundm ent and s tream dwelling bullf rogs in Mis sour i. Herpetologic a 19: 89-97.
Korsch gen, L. J.; Moyle, D. L. (1955) Food ha bit s of th e bullf rog in c entral Missour i farm ponds . Amer . Midl. Nat. 54: 332-341.
Lilly whit e, H. B. (1970) Behavioral temp erature regula tion in t he bullf rog, Rana catesb eiana. Copeia 1970: 158-168.
Martof, B. S.; Palmer , W. M.; Bailey, J. R.; et al . (1980) Amphibia ns and r ept iles of th e Carolina s and Virginia . Chapel Hill, NC: Univ ersi ty of Nor th Carolina Press.
McAlpine , D. F.; Dilw or th, T. G. (1989) Microhabitat an d prey si ze among t hree species of Rana (Anura: Ranidae) symp atr ic in easter n Canada. Can. J. Zool. 67: 2241-2252.
McAuliffe, J. R. (1978) Biologic al survey an d management of sp ort-hunt ed bullf rog popula tions in Nebraska. Linc oln, NE: Nebraska Game an d Parks Commi ssion; 78 pp.
McKamie, J. A.; Heidt, G. A. (1974) A comparison of sp ring f ood ha bit s of th e bullf rog, Rana catesb eiana, in three habitats o f cen tral Arkansas. Southwest. Nat. 19: 107-111.
Modzelewski , E. H., Jr .; Culle y, D. D., Jr . (1974) Growth responses o f th e bullf rog, Rana catesb eiana fed var ious liv e foods . Herpetologic a 30: 396-405.
2-463 Bullf rog

Oliver , J. A. (1955) The natural histo ry of North Amer ican amphibia ns and r ept iles. Princ eton, NJ: Van Nostrand Co.
Raney, E. C. (1940) Summer movemen ts o f th e bullf rog, Rana catesb eiana (Shaw), as determined by th e jaw-tag method. Am. Midl. Nat. 23: 733-745.
Raney, E. C.; Ingr am, W. M. (1941) Growth of tag ged frogs (Rana catesb eiana Shaw and Rana clami tans Daudin) unde r natural condit ions . Am. Midl. Nat. 26: 201-206.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Ryan, M. J. (1980) The reproduc tive behavior of th e bullf rog (Rana catesb eiana). Copeia 1980: 108-114.
Ryan, R. A. (1953) Growth rates o f so me ranids unde r natural condit ions . Copeia 1953: 73-80.
Smith, H. M. (1956) Handbook of amphibia ns and r ept iles of Kansas. Univ . Kansas Mus. Nat. His t. Misc. Publ. 9.
Smith, P. W. (1961) The amphibia ns and r ept iles of Illinois . Ill. Nat. His t. Surv. Bull. 28.
Stewar t, M. M.; Sandis on, P. (1973) Comparative food ha bit s of symp atr ic mi nk fr ogs, bullf rogs , and gr een frogs . J. Herpetol. 6: 241-244.
Storer, T. I. (1922) The easter n bullf rog in Ca lif ornia . Calif . Fish and Game 8: 219-224.
Treanor , R. R.; Nichola , S. J. (1972) A preliminary stu dy of th e commer cial and sporting ut iliza tion of th e bullf rog, R. catesb eiana Shaw in Ca lif ornia . Calif . Dept . Fish and Game, Inland Fis h. Admin. Rep. 72-4; 23 pp.
Turner, F. B. (1960) Postmetamo rphic growth in anur ans. Am. Midl. Nat. 64: 327-338.
Tyler, J. D.; Hoesten bach, R. D., Jr . (1979) Differ ences i n food of bullf rogs ( Rana catesb eiana) from pond a nd s tream habitats i n southwester n Oklahoma. Southwest. Nat. 24: 33-38.
Viparina, S.; Just, J. J. (1975) The life period, growth and di ffer entiation of Rana catesb eiana larvae occu rring in na ture. Copeia 1975: 103-109.
Weathers, W. W. (1976) Influence of temp erature accl imat ion on ox ygen cons umpt ion, haemody namics and ox ygen transpor t in bullf rogs . Aust. J. Zool. 24: 321-330.
Willis , Y. L.; Moyle, D. L.; Baskett, T. S. (1956) Emergence, breeding, hibe rnation, movemen ts an d transfo rmat ion of th e bullf rog, Rana catesb eiana, in Mis sour i. Copeia 1956: 30-41.
2-464 Bullf rog

Wright , A. H. (1914) North Amer ican Anura: life histo ries of th e Anurans of Ithaca, New York. Washingt on, DC: Carnegie Insti tute; Publ. No. 197.
Wright , A. H.; Wright , A. A. (1949) Handbook of fr ogs and t oads of th e Unit ed States an d Canada. Ithaca, New York: Comsto ck Publis hing Co.
2-465 Bullf rog

3. ALLOMETRIC EQUATIONS
Values fo r key co ntact r ate facto rs su ch as food a nd water inge stion r ates h ave
been measu red for few w ildlif e species. In this sect ion, we descr ibe allometr ic equations
that can be used to esti mate sever al exposure facto rs on the basis of animal body weight
us ing m odels derived from taxonom ically si mila r sp ecies. We empha size, however , that
measu red values fr om wel l-conducted studies on the species of co ncern are likely to be
more accu rate and to have narrower confidence l imi ts.
Allometr y is defined as th e stu dy of th e relations hips b etween the growth and size
of one body part to the growth and s ize of th e whole organis m; however , allometr ic
relations hips al so exist between body size and ot her biologic al parameter s (e.g., metabolic
rate). The relations hip b etween the phys iologic al and phy sical parameter s and body
weight fr equent ly can be exp ressed as:
Y = a Wtb ± SE of Y, or [3-1]
log Y = log a + b log W t ± SE of log Y [3-2]
where Y is th e biologic al character isti c to be predicted , Wt is th e animal 's body weight
(mass), a and b are empirical ly derived constan ts, and SE is th e stan dard error of th e mean
value of th e parameter .
Equation 3-2 is th e log t ransfo rmation of Equa tion 3-1. Equation 3-2 represents a
str aight line , wi th b equal to the slope of th e line and log a equal to the Y-intercept of th e
line . Values for a and b usually are determined empirical ly fr om measu red values using
line ar regression a nalysi s. Once value s are determined for a and b, Equation 3-1 can be
used to predic t a value of Y from th e body weight of th e anim al. The SE of Y is th e
stan dard error of th e mean Y esti mated for the mean of th e Wt val ues; the SE of log Y is
the stan dard error of th e mean log Y esti mated for the mean of th e log W t value s.
3-1

Allometr ic equations can be used to esti mate p arameter values fo r species fo r
which measu red values are not avai lable. The equations presented in th is ch apter ,
however , should not be used for taxonom ic categ ories other than the categ ory fo r which
each was d evelope d. For examp le, equations d evelope d for igua nid liza rds cannot be used
for amphibia ns and s hould not be used for other groups of r ept iles wi thout car eful
evalua tion of lik ely di ffer ences b etween the groups . It also is impor tant to rememb er that
the allometr ic equations presented in th is ch apter have been develope d us ing mean values
for a num ber of sp ecies wi thin a taxonom ic categ ory. Indiv idua l species us ually exhibit
values so mewh at differ ent fr om th ose predicted by an allometr ic mo del based on sever al
species. Furthermore, differ ent-si zed indiv idua ls wi thin a sp ecies and indiv idua ls at
varying stag es of matu ration a re lik ely to exhibit a di ffer ent allometr ic relations hip b etween
body weight and t he dependent var iable . For further dis cussion of wi thin- species
allometr ic equations r elated to growth and reproduc tion, see Reiss (1989).
In the next fi ve sect ions , we descr ibe empirical ly derived allometr ic equations th at
relate food ing estion r ates (Sect ion 3 .1), water intake r ates (Sect ion 3 .2), inha lation r ates
(Sect ion 3 .3), surface ar ea (Sect ion 3 .4), and metabolic r ate (Sect ion 3 .5) to body weight .
As discu ssed above, most o f th e allometr ic mo dels differ for birds, mammal s, repti les, and
amphibia ns, and m any also vary wi thin t hese tax onom ic groups . In Sect ion 3 .6, we pr ov ide
a summar y of operations inv olv ing loga rithms and pow ers and unit conv ersion facto rs fo r
those persons wh o may wan t to modify al lometr ic equations found in t he lit erature.
Fina lly , in Sect ion 3 .7 we descr ibe how to esti mate 95-p ercent co nfidence intervals fo r food
inge stion r ates an d free-living metabolic r ates p redicted on the basis of allometr ic
equations presented in th is ch apter . We present mo st eq uations in t he unt ransformed
form only . For equations fo r whic h an investi gator reported stan dard errors fo r the log
transfo rmation of th e relations hip, we pr esent th e equation bot h ways. For those persons
interested in esti mat ing c onf idence int ervals for other allo metr ic equations , Peters (1983)
provides a si mple review of how to esti mate r egression stati sti cs fo r equations of th e form
of Equa tion 3-2. Sect ion 3 .8 cont ains th e references fo r th is ch apter .
3-2

3.1. FOOD INGESTION RATES
Food ing estion r ates var y wi th many facto rs, including metabolic r ate, the energy
devoted to growth and r eproduc tion, and c ompos it ion of th e die t. The metabolic r ate of
free-ranging a nim als is a func tion of sever al facto rs, including a mbie nt temp erature,
acti vi ty l evels, and body weight . In bir ds and mammal s, thermoregula tion c an
considerably increase an animal 's meta bolic r equir ements d uring t he wint er, whereas
reproducti ve effo rts can replace th ermoregula tion a s the predom inant extr a metabolic
expendit ure in t he spring a nd s ummer . Many r ept iles and amphibia ns, on t he other hand,
drop their acti vi ty l evels and metabolic r ates i n the winter .
For homeotherms (i .e., animals th at mai ntain a relatively co nstan t body temp erature
such as mo st b irds and mammal s), metabolic r ate generally decreases wi th increasing
body mass (see Sect ion 3 .5). The smal lest b irds and mammal s mu st co nsume quanti ties
of food e qual to their body weight or more daily ; in c ont rast, the larger homeotherms may
cons ume only a small f raction of th eir body weight in f ood da ily . Herbiv ores tend t o
cons ume larger quant it ies of food t han carniv ores because of th e lower energy co ntent of
their food. Ingestion r ates, expressed in unit s of food e nergy nor malize d to body size (e. g.,
kcal /kg-day), are not s ignif icantly di ffer ent fo r herbivores and carnivores (Peter s, 1983).
Four -legged poik ilot herms (thos e anim als whos e usual body temp eratures are the same as
that of th eir environm ent, su ch as rept iles and amphibia ns) exhibit th e same s lope of
decreasing ing estion r ates p er unit body weight wi th inc reasing body size but show a low er
int ercept (i.e., low er inge stion r ate fo r a given body weight ) than hom eotherms (Nagy,
1987).
The rate of food c ons umpt ion t hat an anim al must achi eve to meet i ts meta bolic
needs can be calculated by dividing it s free-liv ing ( or fi eld) metabolic r ate (FMR) (see
Sect ion 3 .5) by th e metaboliza ble en ergy in it s food ( Nagy, 1987). Metaboliza ble en ergy
(ME) is th e gross en ergy (GE) in a unit of food c ons umed minus th e energy los t in feces
and ur ine. Assi mila tion effi ciency (AE) equals th e ratio ME/GE, or the fract ion of GE that is
metaboliza ble. AE is r elatively con stan t among di ffer ent groups of cons umer sp ecies of
mammal s and birds th at are all either carnivorous, insecti vorous, herbivorous, or
3-3

granivorous (Hume, 1982; Peters, 1983; Nagy, 1987; Robbins , 1983). Nagy (1987)
calculated the mean ME (i.e., kilojoule s of ME per gram of dry matter ) of var ious diets fo r
birds and mammal s fr om aver age values of AE for birds and mammal s and typ ical GE
contents o f th ose diets as r eported by Golley (1961) and Robbins (1983). These val ues are
presented in Table 3-1. (For more informat ion on ME and A E, see Sect ion 4 .1.2.) Using t he
values presented in Table 3-1, Nagy (1987) develope d allo metr ic equations fo r food
inge stion ( FI) rates as a fu nct ion of body weight (W t) for bir ds, mammal s, and lizards us ing
esti mated FMRs and ge neral di etary co mposi tion. In the remainde r of this sect ion, we
present th ese equations fo r bir ds (Sect ion 3 .1.1) and mammal s (Sect ion 3 .1.2). Secti on
3.1.3 summar izes Nagy's food ing estion a llo metr ic equations fo r igua nid liza rds. We repor t
this inf ormation even though no igua nid liza rds were among our selected species because
it is th e only inf ormation of this typ e we ide nt if ied for any amphibia n or rept ile .
Nagy's (1987) esti mates o f FMR are based on doubly labeled water measu rements
of CO2 produc tion in f ree-liv ing a nim als. When perfor med correctl y, th is met hod is mo re
accu rate fo r esti mat ing t he metabolic r ate of fr ee-living a nim als than other methods
commonly used (King, 1974). Other allometr ic equations fo r food ing estion r ates th at we
identi fied in the open literature are based largely on captive animals wi thout co rrections fo r
the addit iona l energy r equir ements o f fr ee-living a nim als. For more accu rate esti mates o f
food ingestion r ates b y typ e of diet, we recommen d follow ing t he procedur es out line d in
Sect ion 4 .1.2 instead of using t hese generic equations .
3.1.1. Birds
For birds, Nagy (1987) cal culated FI rates (i n grams d ry matter per day) fr om ME and
FMR and d evelope d the follow ing e quations :
FI (g/day) = 0.648 Wt0.651 (g), or
FI (kg/day) = 0.0582 Wt0.651 (kg)
FI (g/day) = 0.398 Wt0.850 (g)
all birds [3-3]
passer ines [3-4]
3-4

Table 3-1. Metabolizable Energy (ME) of Various Diets fo r Birds and Mammal s
Diet
insects
fish
Metabolizable Energy a a(kJ /g) (kca l/g)
18.7 = 4.47
18.0 = 4.30
18.7 = 4.47
16.2 = 3.87
Animal Group
mammal s
birds
mammals
birds
mamma ls
mamma ls
hu mmingbir ds
mamma ls and birds
vegetati on 10.3 = 2.26
seeds 18.4 = 4.92
nectar 20.6 = 4.92
14 = 3.35omnivo ry
ag = gr ams dr y weight . Sour ce: Nagy, 1987.
FI (g/day) = 0.301 Wt0.751 (g)
FI (g/day) = 0.495 Wt0.704 (g)
non-p asser ines
seabirds
[3-5]
[3-6]
where Wt equals the body weight (wet) o f th e animal in grams (g ) or kilogr ams (kg ) as
indi cated . We provide the regression stati sti cs fo r these equations (including sample size
and r egression co effi cient) and informat ion r equir ed to esti mate a 95-p ercent co nfidence
interval for an FI rate predicted for a speci fied body weight in Sect ion 3. 7. More accu rate
esti mates o f food r equir ements can be made from esti mates o f FMR (Sect ion 3. 5), dietary
composi tion, and AE for th e species of inter est, as outlined in Sect ion 4. 1.2.
3.1.2. Mammals
For placental mammal s, Nagy (1987) cal culated FI rates (i n grams d ry matter per
day) fr om ME and FMR values and d eveloped the following equations:
3-5

FI (g/day) = 0.235 Wt0.822 (g), or all mammal s [3-7]
FI (kg/day) = 0.0687 Wt0.822 (kg)
FI (g/day) = 0.621 Wt0.564 (g) rodents [3-8]
FI (g/day) = 0.577 Wt0.727 (g) herbivores [3-9]
We prov ide th e regression stati sti cs fo r these equations (inc luding samp le size and
regression c oeffi cient) and informat ion r equir ed to esti mate a 95-p ercent co nfidence
interval for an FI rate predicted for a speci fied body weight in Sect ion 3 .7. More accu rate
esti mates o f food r equir ements can be made from esti mates o f FMR (Sect ion 3 .5), dietary
compos it ion, and AE for th e species of int erest, as outline d in Sect ion 4 .1.2.
Herbiv ores tend t o cons ume more food t han carniv ores or omniv ores on a dry-
weight basis because of th e lower energy co ntent of th e herbivores' diets. On an energy
basis (e.g., ki localories), the inge stion r ates o f car nivores and herbivores are not
signif icantly di ffer ent (Farlow, 1976):
FI (kjoule /day) = 971 Wt0.73 (kg) (r2 = 0.942), or herbivores [3-10]
FI (kcal /day) = 1.518 Wt0.73 (g)
FI (kjoule /day) = 975 Wt0.70 (kg) (r2 = 0.968), or carnivores [3-11]
FI (kcal /day) = 1.894 Wt0.70 (g)
3.1.3. Reptiles and A mphibians
This sect ion s ummar izes food ing estion a llo metr ic equations fo r igua nid liza rds,
whic h is th e only inf ormation of this typ e we ide nt if ied for any amphibia n or rept ile . Nagy
(1987) calculated FI rates (i n grams d ry matter per day) fr om ME and FMR values on s pring
and summer days an d develope d the follow ing e quations :
3-6

FI (g/day) = 0.019 Wt0.841 (g) herbivores [3-12]
FI (g/day) = 0.013 Wt0.773 (g) insecti vores [3-13]
Again, on an energy basis, car niv ores and he rbiv ores are not signif icantly di ffer ent and can
be represented by a s ingle r elations hip:
FI (kjoule /day) = 0.224 Wt0.799 (g), or all igua nids [3-14]
FI (kcal /day) = 0.054 Wt0.799 (g)
We prov ide th e regression stati sti cs fo r these equations (inc luding samp le size and
regression c oeffi cient) and informat ion r equir ed to esti mate a 95-p ercent co nfidence
interval for an FI rate predicted for a speci fied body weight in Sect ion 3 .7. More accu rate
esti mates o f food r equir ements fo r these and other groups of r ept iles and amphibia ns can
be made from esti mates o f FMR (Sect ion 3 .5), dietary co mposi tion, and AE for th e species
of interest, as outline d in Sect ion 4 .1.2.
Allometr ic equations fo r FI rates fo r other groups of r ept iles and amphibia ns wer e
not found. For other groups , we recommen d esti mat ing FI r ates fr om FMR and die t, as
descr ibed in Sect ion 4 .1.2.
3.2. WATER INTAKE RATES
Daily water requir ements d epend on the rate at wh ich animals lose water to the
environm ent due to evapor ation a nd excr etion. Los s rates d epend on sever al facto rs,
inc luding body size , ambie nt temp erature, and phys iologic al adaptations fo r cons erving
water . Drinking water is only one way i n which animals may meet th eir water
requir ements. All animals produce so me water as a product o f th eir metabolis m. The
degree to which metabolic water produc tion a nd di etary water content can sati sfy an
animal 's water requir ements var ies fr om sp ecies to species and wi th environmental
condit ions . Exten sive l iterature descr ibes th e allometr y of to tal water flux fo r var ious
3-7

groups of anim als. Allo metr ic mo dels to predict d rinking water intake, on the other hand,
are limi ted.
3.2.1. Birds
Based on measu red body weight s and dr ink ing water values fr om Calder (1981) and
Skadhauge (1975), Calder and Braun (1983) develope d an equation f or dr ink ing water
inge stion ( WI) for bir ds:
WI (L/day) = 0.059 Wt0.67 (kg) all birds [3-15]
where Wt equals th e average body weight in k ilogr ams (kg ) of th e bird species. This
equation is b ased on data fr om 21 sp ecies of 11 to 3,150 g body weight . Total water
turnov er should be propor tiona l to metabolic r ate (body weight to the 3/4 pow er, see
Sect ion 3 .5.2.1). The expone nt fo r Equa tion 3-15 is not s ignif icantly di ffer ent fr om 0.75
(Calder and Braun, 1983). Addit iona l sour ces o f water not acc ount ed for in t his equa tion
(metabolic water and water contained in food) al so help t o balance the anim als' daily water
losses. For allometr ic equations fo r to tal water flux (including water obtained from food)
for birds, see Nagy and Peterson (1988).
To esti mate d aily drinking water intake as a p ropor tion of an anim al's body weight
(e.g., as g/g-day), the WI rate esti mated above is divided by th e animal 's body weight in k g:
WI (g/g-day) = WI (kg/kg-day), or [3-16]
= WI (L/day) / Wt (kg )
In general, birds drink less water than do mammal s of equiv alent body weight s.
Because of th eir relatively high metabolic r ates, the quanti ty o f meta bolic water produc ed
by birds is greater in relations hip t o body size th an that pr oduc ed by other ver tebrates
(Bartholomew an d Cade, 1963). In addit ion, bir ds ar e able to cons erve water by excr eting
nitrogen as uric aci d instead of urea (as excr eted by mammal s); uric aci d can be excr eted
3-8

�
� � �
� �
�
in a semi -solid s uspension, whereas urea must b e excr eted in aqueous solut ion. On the
other hand, bir ds exhibit a high r ate of water loss fr om th e respiratory system an d use
pant ing a nd evaporative water loss to prevent overheating a t high a mbie nt temp eratures.
For examp le, Dawso n (1954) found evaporative losses i n two species of to whees to
increase fo urfold b etween 30 and 40�C.
Although bir ds may sati sfy so me of th eir water needs by oxidative food metabolis m,
it has not been demonstr ated that any normal ly acti ve bird can sati sfy i ts water
requir ements wi th metabolic water alone (B artholo mew an d Cade, 1963). The balance
must b e obtained from water contained in foods su ch as in sects o r succu lent plant
mater ial and from drinking water .
As would be exp ected , birds drink mo re water at war mer temp eratures to make u p
for evaporative losses. Seibert (1949) found t hat junc os (weighing 1 6 to 18 g) cons umed
an average of 11 percent of th eir body weight in water daily at an ambient temp erature of
0�C, 16 percent at 23�C, and 21 percent at 37�C. The white-th roated sparrow increased
water consumption f rom 18 pe rcent of its body weight at 0�C to 27 percent at 23�C and 44
percent at 37�C.
Water consumption r ates p er unit body weight al so tend t o decrease wi th increasing
body weight wi thin a sp ecies. For examp le, in white leghor n chicken s, water intake p er
gram of body weight is high est i n the youngest ch icks (45 p ercent of th e body weight at 1
week wh en chicks aver age 62 g) and decreases wi th age thereafter (13 percent of th e body
weight at 16 weeks wh en chicks aver age 2.0 kg) unti l egg-laying, when water consumption
increases fo r the produc tion of eggs (24 percent of th e body weight fo r laying he ns)
(Medway an d Kare, 1959).
Some sp ecies obtain more of th eir daily water needs fr om th eir diet and therefore
drink less water than others; therefore, measu red water inge stion v alues from well-
condu cted studie s should be used when avai lable. In the absence of measu red values,
Equation 3-15 should pr ov ide a reasona ble cen tral value . Addit iona l inf ormation r equir ed
to esti mate a 95-p ercent co nfidence interval was n ot provided along w ith this equa tion.
3-9

3.2.2. Mammals
Based on measu red body weight s and dr ink ing water values fr om Calder (1981) and
Skadhauge (1975), Calder and Braun (1983) develope d an allo metr ic equation f or dr ink ing
water inge stion ( WI) for mammal s:
WI (L/day) = 0.099 Wt0.90 (kg) all mammal s [3-17]
where Wt equals th e average body weight in k ilogr ams (kg ). Addit iona l sour ces o f water
not account ed for in t his equa tion ( i.e., metabolic water and water contained in food) help
to balance th e animals' daily water losses. The empirical ly determined expone nt of 0.90
does not su ggest a si mple physiologic al expla nation. If to tal water turnover (metabolic
water combine d with water obtained from food) is propor tiona l to metabolic r ate (body
weight to the 3/4 pow er, see Sect ion 3 .5.2.1), then dr ink ing water inge stion w ould be
expected to scal e simi larly, as was th e case fo r birds (see Sect ion 3 .2.1). For allo metr ic
equations r elating body weight to total water flux (including water obtained from food) fo r
mammal s, see Nagy and Peterson (1988).
To nor malize dr ink ing water intake to body weight (e. g., as g/g-day; see Chapter 4,
Equation 4-4), the WI rate esti mated above is divided by th e animal 's body weight in k g:
NWI (g/g-day) = WI (kg/kg-day), or [3-18]
= WI (L/day) / Wt (kg )
We present normalized dr ink ing water intakes i n the species profi les.
3.2.3. Reptiles and A mphibians
Allometr ic equations r elating body weight to dr ink ing water inge stion r ates wer e not
ident if ied for rept iles and amphibia ns. The water balance of th ese groups is co mple x, in
part because th ey can absorb water through t heir skin a s well a s dr ink water and extr act
water from th eir food ( Duellm an and T rueb, 1986; Minnic h, 1982). The relative
3-10

cont ribut ion of th ese th ree routes o f water intake d epends on the species, habitat,
temp erature, and body su rface ar ea. In general, the ski n of repti les is less p ermeable than
that of amphibia ns. Aquatic turtles (e.g., snapping t urtle , paint ed turtle) also may inge st
large amounts o f water when feeding on a quatic pla nts and anim als; however , the
magnit ude of su ch ing estion ha s not been quant if ied (Mahmoud a nd Kli cka, 1979). For
further dis cussion of water balance fo r these groups , see Duellman and Trueb (1986),
Feder and Burggren (1992), Minnic h (1982), and Nagy and Peterson (1988).
3.3. INHALATION RATES
Inhalation r ate is one of th e respiratory parameter s needed to esti mate p otential
expos ure of wildlif e to airbor ne con tamina nts. Inhalation r ates var y wi th species, body
size, body temp erature, ambient temp erature, and acti vi ty l evels. When inha lation r ate is
increased, either because of increased acti vi ty l evels or to promote evap orative cooling,
exposure to airborne contami nants may b e increased. As discu ssed in Sect ion 4 .1.4, an
inha lation t oxicologis t should be cons ult ed when assess ing t his pathway b ecause
addit iona l respir atory parameter s also must b e considered (see U.S. EPA, 1990).
3.3.1. Birds
Lasiewski and Calder (1971) develope d an allo metr ic relations hip f or inha lation r ate
(IR) asso ciated wi th stan dard metabolis m (i.e., post-d igesti ve, at rest) fo r non- passer ine
bir ds (N = 6 species ranging in w eight fr om 43 to 88,000 grams). They exc lude d
passer ines, which have a so mewh at highe r metabolic r ate th an non- passer ines (see
Sect ion 3 .5):
IR (ml/min) = 284 Wt0.77 (kg), or all non- passer ines [3-19]
IR (m3/day) = 0.4089 Wt0.77 (kg), or
IR (m3/day) = 0.002002 Wt0.77 (g)
As noted above, these inha lation r ates wer e asso ciated wi th stan dard metabolic r ates.
Free-liv ing metabolic r ates ar e likely to be highe r by a facto r of at l east 2 o r 3 (see Secti on
3-11

3.5); therefore, IRs esti mated from th ese equations should be adju sted acco rdingly (e. g.,
mult iplie d by 2 or 3) although IRs might not be dir ectl y propor tiona l to metabolic r ate.
3.3.2. Mammals
Using measu red values fr om sever al reports o f respiration r ates i n mammal s
(cover ing 691 data points), Stahl (1967) develope d an allo metr ic relations hip f or inha lation
rate wi th body size fo r mammal s (N = 691, r = 0.98, SE Y = 45):
IR (ml/min) = 379 Wt0.80 (kg), or all mammal s [3-20]
IR (m3/day) = 0.5458 Wt0.80 (kg), or
IR (m3/day) = 0.002173 Wt0.80 (g)
As for th e equations giv en for bir ds, these IRs wer e asso ciated wi th stan dard metabolic
rates. Field metabolic r ates ar e likely to be highe r by a facto r of at l east 2 o r 3 (see Secti on
3.5); therefore, IRs determined from th ese equations should be adju sted acco rdingly (e. g.,
mult iplie d by 2 or 3, although IRs may n ot be directl y propor tiona l to metabolic r ate).
3.3.3. Reptiles and A mphibians
In contrast to the fairly regula r breathing p atter ns of mo st b irds and mammal s, most
repti les breath air in disti nct ep isodes. They may take s ingle breaths, or exhibit an epis ode
of sever al breaths, and then hold t heir breath for varying le ngt hs of ti me (Mils om and Cha n,
1986). Inhalation r ate var ies fo r repti les and amphibia ns not only wi th body size an d
acti vi ty l evel , as fo r birds and mammal s, but also wi th body temp erature. Some gas
exch ange occu rs normal ly th rough t he int egum ent of bot h rept iles and amphibia ns
(Duellman and Trueb, 1986; Lilly whit e and Maderson, 1982). Moreover , for semi aquatic
species, a signif icant pr opor tion of gas exch ange can occu r unde rwater through t he skin,
reduc ing t he need to ins pir e air (Seymo ur, 1982). For examp le, in adult bullf rogs , gas
exch ange through t he skin c an account fo r 18 percent of to tal oxygen uptake (B urggren
and West, 1982). Given the complexi ty o f th e subject, we refer those interested in
3-12

inha lation e xpos ures for rept iles or amphibia ns to more speci fic treatmen ts o f th ese topic s
(e.g., Duellman and Trueb, 1986; Feder and Burggren, 1992; Gans and Dawso n, 1976;
Jackso n, 1979; Hutchins on et al., 1968; Lilly whit e and Maderson, 1982).
3.4. SURFACE AREAS
The degree to which an animal may ab sorb contami nants th rough dir ect co ntact
wi th its ski n depends on many facto rs, including t he surface ar ea of th e ski n avai lable for
contact. Summar izing measu red surface ar eas fo r more than 100 animals reported by
Hemmi ngsen (1960), Schmidt-Nielsen (1970, 1972) determined that animals have su rface
areas th at usually are approximatel y twi ce th at of a sp here of th e same we ight (assu ming a
speci fic gr avi ty of 1 for bot h the sphe re and t he anim al). The permeabilit y of an anim al's
ski n to contami nants, however , depends on character isti cs o f th e ski n (e.g., presence of
keratinize d scal es) as wel l as th e contami nant (e.g., molecule size, lipophilic ity). This
sect ion pr esents a llometr ic equations fo r esti mat ing s kin s urface ar ea; character isti cs
affecting s kin pe rmeabilit y are not dis cussed .
3.4.1. Birds
In s tudie s of avi an thermal biology , skin s urface ar ea is co mmonly esti mated using
Meeh's (1879, ci ted in Walsberg and King, 1978) formula wi th Rubne r's (1883, ci ted in
Walsberg and King, 1978) constan t of 10:
SAski n (cm 2) = 10 Wt0.667 (g) all birds [3-21]
where SAski n is th e ski n surface ar ea beneath the feath ers and Wt is body weight (W alsberg
and King, 1978). Although Rubne r's con stan t of 10 was d erived origina lly fr om domesti c
fowl, Drent and Stonehouse (1971) have ver ified the formula for birds in a var iety o f taxa
and of weight s spanning t hree orders of magnit ude. For passer ines, beak su rface ar ea
tends to be about 1 pe rcent (range 0.7 percent to 1.6 percent of 10 passer ine species) of
ski n surface ar ea, and leg surface ar ea about 7 pe rcent (range 5.9 percent to
3-13

7.9 percent of 10 passer ine species) (Walsberg and King, 1978). These ratios would be
expected to vary fo r many non- passer ines (e.g., herons, woodc ock).
3.4.2. Mammals
Summar izing d ata fr om mo re than 100 mammal s, Stahl (1967) develope d a
relations hip b etween surface an d body weight :
SAski n (m2)= 0.11 Wt0.65 (kg), or all mammal s
SAski n (cm 2) = 12.3 Wt0.65 (g)
[3-22]
This r elations hip is ver y simila r to that develope d for bir ds (Equa tion 3-21).
3.4.3. Reptiles and A mphibians
Surface ar ea has been found t o be a di ffer ent fu nction of body weight fo r adult
amphibia ns th an for bir ds or mammal s (Hutchins on et al., 1968; Whitfo rd and Hutchins on,
1967):
SAski n (cm 2) = 1.131 Wt0.579
SAski n (cm 2) = 0.953 Wt0.725
SAski n (cm 2) = 0.997 Wt0.712
SAski n (cm 2) = 8.42 Wt0.694
(g) all frogs
(g) bullf rog
(g) green frog
(g) salamanders
[3-23]
[3-24]
[3-25]
[3-26]
Models by whic h to esti mate su rface ar eas fo r turtles (excl usive of th e shell and
plastr on) and snakes wer e not found. The general formula fo r th e surface ar ea of a
cylinde r can be used to appr ox imate th e surface ar ea of a sn ake i f th e length and girth are
known or esti mated .
3-14

3.5. ALLOMETRIC EQUATIONS FOR METABOLIC RATE
The allometr ic equations fo r esti mat ing f ood ing estion r ates p rovided in Sect ion 3 .1
were derived us ing v ery simple assu mptions about th e energetic cont ent and dig estibilit y
of th e die t for th e species inc lude d in t he regression e quations . Cons equent ly , the
equations will pr ov ide only ver y rough esti mates o f food ing estion r ates fo r any given
species. For a si te-sp eci fic exp osure assessmen t, it may b e more appropriate to evaluate
inge stion r ates fo r a diet th at is l ikely to represent th e species and stu dy area. The caloric
content and percent water , fat, and protein of wildlif e diets var y not only among s pecies,
but al so among indiv idua ls wi thin t he same sp ecies depending on facto rs su ch as locat ion,
time of year , age, and sex. If one can esti mate th e energetic requir ements o f th e animal in
the field and its d ietary co mposi tion f or a speci fied si tuation, one can esti mate fo od
inge stion r ates fo r that diet and si tuation. In the remainde r of this sect ion, we dis cuss
metabolic r ate and provide allometr ic equations to esti mate fi eld free-living metabolic r ates
(FMRs) for wildlif e species. Chapter 4 descr ibes how to use FMR esti mates an d
inf ormation a bout th e energy cont ent of sp eci fic di ets to esti mate f ood ing estion r ates.
Several facto rs influence meta bolic r ates o f fr ee-ranging a nim als, inc luding body
size, body temp erature, and typ e and level of acti vi ty. For homeotherms, metabolic en ergy
must b e expended to keep core body temp erature wi thin r elatively narrow lim its. At
moderate amb ient temp eratures, homeotherms l ose heat to the surrounding e nvironm ent
as rapidly as they ga in it and t herefore need not exp end extr a metabolic en ergy to maint ain
core body temp erature. That range of amb ient temp eratures over which an animal 's
metabolic r ate is at a m inim um and c onstan t level is cal led the thermoneutral zone. Below
the thermone utral zone , the organis m lo ses h eat to the environment and must i ncrease i ts
metabolic acti vi ty to compensate. Above th e thermoneutral zone, the organis m gains heat
from i ts en vironment and must i ncrease i ts meta bolic r ate to use evap oration t o cool it s
body .
Thermoneutral zones var y so mewh at among s pecies depending upon t he ins ulating
properties and c olor of th e fur or feath ers, surface-to -volume ratios, and other facto rs. The
degree to which metabolic r ate increases wi th changes in ambient temp erature outsi de of
3-15

�
�
�
the thermone utral zone is r efer red to as th e temp erature coeffi cient (TC). Temperature acoeffi cients al so vary wi th body size , ins ulation, and ot her facto rs.
There are sever al ways to measu re and express meta bolic r ate, including b asal
metabolic r ate (BMR), resting metabolic r ate (RMR), existen ce meta bolic r ate (EMR),
average daily meta bolic r ate (ADMR), and f ree-liv ing or field metabolic r ate (FMR). The
differ ent measu res are dist inguis hed by th e range of anim al acti vi ties include d in t he
measu re:
� Basal metabolic r ate (BMR), also someti mes l abeled stan dard metabolic r ate
(SMR), represents th e minimal value of heat produc tion f or hom eotherms.
BMR must be measu red wi thin t he thermone utral zone of ambie nt
temp eratures wh en the animal is at r est an d in a post-ab sorptive state (i .e.,
all f ood ha s been dig ested ) (Gessaman , 1973).
� Standard metabolic r ate (SMR) has been used in the literature in more than
one way. Many authors define SMR as BMR (see above). Others use SMR if
the thermone utral zone has not been define d so that some cost of
thermoregula tion may b e include d (Bennett an d Harvey, 1987).
� Resting metabolic r ate (RMR) is usually measu red at temp eratures below th e
thermone utral zone wh en the anim al is at rest, but not post-ab sorptive (i .e.,
the anim al is eating r egula rly and may b e expending e nergy to dig est i ts
food) . The RMR exceed s th e BMR by th e heat libe rated in the dige stion of
food ( i.e., the speci fic dy namic action, or SDA) and by so me cost of
thermoregula tion. RMR and BMR a re usually measu red using indir ect
calorimetr y (i .e., oxyg en consumption a nd c arbon diox ide produc tion) over a
period of 1 to 3 hour s.
aWater has a much higher heat conduc tance than air. When subm erged or s wimming, the degreeto which metabolic rate i ncreases w ith decreasi ng water tem peratu re depends on the animal's insul ation (e.g., whether t he fur t raps an ai r layer next to the ski n over part or al l of the body or whether t here is an insul ative layer of blubber) , durat ion of subm ergence, and body si ze.
3-16

�
�
�
� Existen ce meta bolic r ate (EMR) is th e metabolic r ate necessar y fo r an animal
to maintain itsel f in captivi ty wi thout a change in body weight . EMR is
greater than RMR due to the cost of loc omotor and ot her acti vi ties requir ed
for sel f-mai ntenance. Most r esearchers measu re EMR on the basis of fo od
cons umpt ion a nd energy excr etion a t a con stan t weight over th e period of
sever al days o r weeks (K endeigh, 1969).
� Average daily meta bolic r ate (ADMR) is usually measu red over 24 hour s at a
temp erature simi lar to the animal 's natural environment and wi th food a nd
water avai lable ad lib itum. ADMR is th e sum of BMR and t he metabolic co sts
of th ermoregula tion, dig estion, and da ily acti vi ties.
� Free-liv ing or field metabolic r ate (FMR) can be measu red using doubly -
labeled water , and it represents th e total daily energy requir ement fo r an
anim al in t he wild. FMR inc lude s the costs o f BMR, SDA, thermoregula tion,
loc omot ion, feeding, predator avoida nce, alertness, postu re, and other
energy exp endit ures. Various mode ls and measu res have indi cated that a
constan t val ue of approximatel y th ree times BMR is a reasonable esti mate o f
FMR for bir ds and mammal s (Lamprey, 1964; Buechner and Golley, 1967;
Koplin e t al., 1980), although m ore precise esti mates al so have been
develope d (see Sect ions 3. 5.1.3, 3.5.2.3, and 3.5.3.2).
FMR also has been used in t he lit erature to represent fast ing metabolic r ate (e.g.,
Gessaman , 1973), but we d o not discu ss fast ing metabolic r ate esti mates i n th is Handbook .
The relations hips b etween metabolic r ate and body weight fall int o two broad
categ ories: those fo r homeothermic animals (i .e., most b irds and mammal s), and those fo r
poik ilot hermic anim als (i.e., most rept iles and amphibia ns). For poik ilot herms, metabolic
rate mu st b e related to body temp erature. It also is important to rememb er that
poik ilot herms can adjus t their body temp eratures relative to ambient temp eratures
3-17

somewh at by modif ying t heir behavior (e.g., basking in t he sun, adopt ing pos tures to
minim ize or maxi mize absorption of sola r radiation) .
Allometr ic mo dels relating metabolic r ate to body size fo r bir ds and mammal s are
descr ibed in Sect ions 3. 5.1 and 3.5.2, respecti vely. Allometr ic mo dels fo r repti les and
amphibia ns ar e descr ibed in Sect ion 3 .5.3. We have attemp ted to identi fy th e most
accu rate allometr ic equations cu rrent ly avai lable for esti mat ing f ree-liv ing metabolic r ates.
We also present allometr ic equations fo r basal and existen ce meta bolis m, whic h in
combina tion w ith appr opr iate informat ion on acti vi ty budgets an d energy co sts can be
used to esti mate fi eld metabolic r ates. Furthermore, measu res of basal and existen ce
metabolis m are avai lable for considerably mo re species th an are measu res (or esti mates)
of fr ee-liv ing metabolic r ates. Consequently, more allometr ic mo dels have been develope d
that dis tinguis h the metabolic r ate-weight r elations hip a mong taxonom ic groups us ing
measu res of basal and existen ce meta bolis m than us ing measu res of fi eld metabolic r ates.
We caution us ers to pay clos e atten tion t o the unit s for th e parameter s in the allometr ic
equations . For most equations , energy is exp ressed as kcal (wi th the excep tion of so me
equations fo r rept iles and amphibia ns). Mass may b e expressed either in g or kg,
depending on how th e equation was r eported.
We emphasize that th e literature on allometr ic relations hips and metabolic r ate is
exten sive and complex. We provide a very si mplif ied ov erview that should be of
assi stan ce fo r scr eening- level exposure assessmen ts only . For addit iona l inf ormation on
methods of esti mat ing metabolic co sts o f fr ee-ranging a nim als, please co nsult exp ert
reviews o n the subject (e.g., Bennett an d Dawso n, 1976; Bennett an d Harvey, 1987; Ellis ,
1984; Gans and Dawso n, 1976; Gessaman , 1973; Kendeigh e t al., 1977; King, 1974; Peters,
1983; Robins on et al., 1983; Wiens, 1984).
3.5.1. Birds
In birds, metabolic r ate generally decreases wi th increasing body mass. Several
authors have found passer ine birds to have highe r metabolic r ates o verall for their body
size th an non- passer ines (Lasiewski and Dawso n, 1967; Nagy, 1987; Kendeigh, 1970;
3-18

Zar, 1968). In th is sect ion, we pr esent allometr ic mo dels fo r three measu res of meta bolic
rate on the basis of body size in bir ds: basal metabolic r ate (BMR), existen ce meta bolic
rate (EMR), and field metabolic r ate (FMR). All equations take th e general form of Y = aWtb,
but can also be represented in their log- transfo rmed form (the equation of a str aight line ).
We conc lude this sect ion by dis cussing t he inf luence of ambie nt temp erature on avian
metabolic r ates. Addit iona l inf ormation r equir ed to esti mate a 95-p ercent co nfidence
interval (CI) for a predicted FMR (the expression of metabolic r ate th at is generally mo st
appropriate fo r wildlif e expos ure assessmen ts) i s provided in Sect ion 3 .7.
3.5.1.1. Basal Metabolic Rate
Several investi gators have derived values fo r the constan ts a and b for the equation
relating BMR t o body weight (W t) from empir ical data on birds. Lasiewski and Dawso n
(1967) compile d body weight and BMR f or almost 100 species of birds. They found BMR
for passer ines to be highe r th an BMR for non- passer ines (i .e., the Y-intercept fo r
passer ines is highe r th an the Y-int ercept for non- passer ines):
Passer ines
log BMR (kcal /day)
BMR (kcal /day)
Non-passer ines
log BMR (kcal /day)
BMR (kcal /day)
= 2.11 + 0.724 log W t (kg) ± 0.113, or
= 128 Wt0.724 (kg)
= 1.89 + 0.723 log W t (kg) ± 0.068, or
[3-27]
[3-28]
= 77.6 Wt0.723 (kg)
Ellis (1984) found t he Y-int ercept for seabirdsb to be somewh at highe r th an the Y-
int ercept for non- passer ines determined by Lasiewski and Dawso n (1967):
bSeabirds included pengui ns , albatross, petrels, shearwaters , pelicans, skuas, gul ls, terns , noddy s, murres, cormorants, and fri gatebirds.
3-19

�
�
�
Seabir ds
log BMR (kcal /day) = 1.96 + 0.721 log W t (kg) (no SE prov ided), or [3-29]
BMR (kcal /day) = 91.2 Wt0.721 (kg)
Zar (1968) reexami ned the data co mpile d by Lasiewski and Dawso n (1967) and
develope d models fo r relating BMR t o body weight (kg ) for sever al orders and familie s of
birds (Table 3-2). These may b e used to esti mate wh ether the FMR for a species of int erest
is l ikely to fal l above or below th at predicted on the basis of th e allometr ic equations
derived for "a ll bir ds."
3.5.1.2. Existen ce Metabolic Rates
Kendeigh (1970) develope d allo metr ic equations fo r EMRs as a func tion of weight
(Wt) at 30�C separately fo r passer ines and for non- passer ines. As was th e case fo r BMRs,
passer ines sh owed highe r EMRs th an did non- passer ines:
Passer ines (N = 15 species)
log E MR (kcal /day) = 0.1965 + 0.6210 log W t (g) ± 0.0633, or [3-30]
EMR (kcal /day) = 1.572 Wt0.6210 (g), or
log E MR (kcal /day) = 2.060 + 0.6210 log W t (kg), or
EMR (kcal /day) = 114.8 Wt0.6210 (kg)
Non-passer ines (N = 9 species)
log E MR (kcal /day) = -0.2673 + 0.7545 log W t (g) ± 0.0630, or [3-31]
EMR (kcal /day) = 0.5404 Wt0.7545 (g), or
log E MR (kcal /day) = 1.996 + 0.7545 log W t (kg), or
EMR (kcal /day) = 99.03 Wt0.7545 (kg), or
The average increase of EMR at 30�C over BMR is 31 and 26 percent in p asser ine and non-
passer ine species, respecti vely (Kendeigh, 1970). At 0�C, on t he other hand, EMR of
passer ine and non- passer ine species is si mi lar, indi cating t hat non- passer ines are affected
3-20

�
Table 3-2. Allometri c Equations for Basal Metabolic Rate (BMR) in Birdsa
Number SE of SE ofb b
of data mean mean Avian group poi nts a log a b BMR log B MR
Apodi formes 9 114 2.06 0.769 0.201 0.0558
Strigiformes 7 66.4 1.82 0.69 11.1 0.0989
Columbiformes 10 92.1 1.96 0.858 2.68 0.0491
Gallif ormes 13 72.6 1.86 0.698 15.3 0.0904
Falconiformes 5 65.3 1.82 0.648 45.3 0.108
Anseri formes 9 95.8 1.98 0.634 23.4 0.0524
Ciconiiformes 7 86.9 1.94 0.737 22.0 0.0464
Passeri formes 48 129 2.11 0.724 8.71 0.0806
Corvi dae 8 126 2.10 0.709 23.3 0.147
Ploeceidae 17 164 2.21 0.794 1.40 0.0808
Fringillid ae 19 125 2.10 0.714 1.02 0.0473
All Nonpasseri nes 72 78.5 1.90 0.723 42.8 0.111
All Species 120 86.3 1.94 0.668 52.8 0.133
aValues for the equat ion r elating BM R to body weight (Wt): log BM R (kcal /day) = log a + b l og Wt (kg). bEst imated f rom the mean log Wt used t o devel op t he allometric equat ion. Sour ce: Zar, 1968.
more by cold t han passer ines. Kendeigh (1970) esti mated the equation f or all bir d species
(N = 24) at 0�C to equal:
All birds (24 sp ecies)
log E MR (kcal /day) = 0.6372 + 0.5300 log W t (g) ± 0.0613, or [3-32]
EMR (kcal /day) = 4.337 Wt0.5300 (g)
3-21

�
�
The equations al so indi cate th at smal ler species are affected more by cold t han are larger
species. The slope s of th e regression line s for EMR on body weight is l ess steep at 0�C
than at 30�C, indi cating t hat smal l birds mu st i ncrease heat produc tion m ore than large
birds to regulate body temp erature during c old weath er.
To nor malize EMR t o body weight , div ide th e daily EMR by body weight :
NEMR (kcal /kg-day) = EMR (kcal /day) / Wt (kg ) [3-33]
3.5.1.3. Free-Living Metabolic Rate
FMRs have been measu red using doubly -labeled water (DLW) to measu re CO2
produc tion in a nim als in t he field. Based on DLW measu rements wi th 25 species of birds,
Nagy (1987) develope d an equation r elating FMR f or bir ds to body weight :
FMR (kjoule s/day) = 10.89 Wt0.640 (g), or all bir ds [3-34]
FMR (kcal /day) = 2.601 Wt0.640 (g)
In bir ds, the slope of FMR (i.e., 0.640) does not differ signif icantly from th e BMR s lope of
0.668 (see Table 3-2). This indi cates th at FMR may b e a relatively co nstan t mu ltiple of BMR
in bir ds over a large range of body mass.
Using esti mates o f FMR deter mined for 42 species by a var iety o f met hods ,
Walsberg (1983) found a si mila r relations hip ( r2 = 0.98, SE Y = 0.415, SE b = 0.012):
FMR (kjoule s/day) = 13.05 Wt0.605 (g), or all bir ds [3-35]
FMR (kcal /day) = 3.12 Wt0.605 (g)
Separating t he passer ine from th e non- passer ine species, Nagy (1987) found a
highe r FMR among p asser ines th an non- passer ines of co mparable weight (i .e., the Y-
intercept fo r passer ines is highe r th an the Y-int ercept for non- passer ines), as exp ected on
the basis of basal metabolic r ate:
3-22

FMR (kjoule s/day) = 8.892 Wt0.749 (g), or passer ines [3-36]
FMR (kcal /day) = 2.123 Wt0.749 (g)
FMR (kjoule s/day) = 4.797 Wt0.749 (g), or non- passer ines [3-37]
FMR (kcal /day) = 1.146 Wt0.749 (g)
FMR (kjoule s/day) = 8.017 Wt0.704 (g), or seabirds [3-38]
FMR (kcal /day) = 1.916 Wt0.704 (g)
FMR (kjoule s/day) = 21.13 Wt0.440 (g), or non-seabirdsc [3-39]
FMR (kcal /day) = 5.051 Wt0.440 (g)
We prov ide th e regression stati sti cs fo r Nagy's (1987) equations (inc luding samp le
size and t he regression c oeffi cient) and informat ion r equir ed to esti mate a 95-p ercent
conf idence int erval for an FMR in Sect ion 3 .7.d
Nagy (1987) esti mated the accu racy o f th e doubly -labeled water method t o be ± 8
percent or better . Because of diffi culties in recapturing bir ds dur ing t he nonbr eeding
seaso n, most o f th e measu red FMRs wer e for breeding bir ds (Nagy, 1987).
King (1974) esti mated that FMR exceed s BMR by a facto r of 3.5 on average (based
on a samp le of 18 measu res fo r species ranging f rom 4 to 400 g in weight). Gessaman
(1973) summar ized data on mockingbir ds and pur ple mar tins fr om Utter (1971) that
indi cated an FMR equal to 1.6 to 2.4 times th e predicted BMR for adult s not acti vely feeding
nestlings . Feeding n estlings inc reased the ratio of FMR to BMR f rom 2.7 to 3.4 in pur ple
mar tins (Utter , 1971, ci ted in Gessaman , 1973).
cAll of the large bi rds i ncluded i n the database w ere seabi rds such as noddy, ki ttiwake, shearw ater,albatro ss, tern , and petrel (Nagy, 1987). Other large birds, such as herons, hawks, and owls, were not included. Accordi ngl y, non- passeri ne and non- seabi rd equat ions shoul d be used w ith caut ion.
dInsuffi cient information is provided in Walsberg (1983) to esti mate co nfidence interval s fo r a predicted FMR for species with body weight s abov e or be low the mean log body weight value of his data set.
3-23

� �
�
�
To nor malize FMR to body weight , div ide th e daily FMR by body weight :
NFMR (kcal /kg-day) = FMR (kcal /day) / Wt (kg ) [3-40]
Figur e 3-1 illus trates ap proximate mo nthly var iations in t he total energy budge t of
an adult hous e sparrow in Illinois th roughout th e year and the relations hip b etween BMR
and FMR (adapted from Kendeigh e t al., 1977). For th is bird, FMR varies seaso nally, wi th a
maxi mum val ue in midwinter (28 kcal /day) and a minim um in Augus t pr ior to molt ing ( 20
kcal /day). Other species, however (e.g., willow ptarmiga n), show no s ignif icant var iation in
FMR with seaso n (King, 1974). For examp les of nest ling e nergy budg ets, see Kendeigh e t
al. (1977) and Dunn (1980). For a discu ssion of mode ling e nergy budg ets fo r birds in
general and for seabirds in particular, see Wiens (1984).
3.5.1.4. Temperature and Metabolic Rate
Below an anim al's thermone utral zone , metabolis m inc reases wi th decreasing
ambient temp erature. Sect ion 3 .5.1.2 presented equations fo r EMR at 30�C and at 0�C, but
these are not particularly helpful for esti mat ing E MR at ot her temp eratures. Although f ew
researchers have attemp ted general multiple r egressions of metabolic r ate on both body
size and temp erature for birds, some relations hips h ave been investi gated in general terms
(Peters, 1983):
� Low temp eratures induc e a greater propor tiona l r ise in metabolic r ate
relative to basal metabolic r ate in smal ler birds th an in larger ones.e
� At high temp eratures, metabolic r ate increases to increase blood f low an d
evapor ative cooling ( via pant ing) .
eThis is because conduct ance and heat loss f or a gi ven t hermal gradi ent between body t emperatureand am bient temperature ri se more sl ow ly with body si ze than do basal metabol ic rates.
3-24


�
Peters (1983) develope d an equation r elating t he ratio of SMR t o BMR to thermal gradient
(i.e., the differ ence between ambient temp erature and body temp erature) for birds:
SMR/BMR = 0.029 (thermal gradient in �C) Wt-0.249 (kg) [3-41]
Thus, stan dard metabolic co sts i ncrease relative to basal metabolis m at low er
temp eratures, but less so for larger birds th an for smal ler birds. Despite th e str ong
dependence of meta bolic r ate on ambient temp erature, for scr eening- level risk
assessmen ts, it should not be necessar y to adjust esti mates o f FMR for seaso nal
temp erature changes. As Figur e 3-1 illus trates, high metabolic d emands of
thermoregula tion in t he wint er can be replaced by th ose of reproduc tion a nd m olt ing
dur ing s pring, summer , and fal l.
3.5.2. Mammals
As fo r birds, metabolic r ate in mammal s generally decreases wi th increasing body
size. The metabolic r ates o f herbivorous and carnivorous mammal s are simi lar for
simi larly si zed species. In th is sect ion, we pr esent allometr ic mo dels fo r three measu res of
metabolic r ate on the basis of body size in mammal s: basal metabolic r ate (BMR), resti ng
metabolic r ate (RMR), and f ree-liv ing metabolic r ate (FMR). All equations take th e general
form of Y = aWtb, but also can be represented in their log-transfo rmed form (th e equation
of a str aight line ). We conc lude this sect ion by dis cussing t he inf luence of ambie nt
temp erature on mammal ian metabolic r ates. Addit iona l inf ormation t hat allow s one to
esti mate a 95-p ercent co nfidence interval for a predicted FMR, the expression of metabolic
rate th at is generally mo st ap propriate fo r wildlif e expos ure assessmen ts, is provided in
Sect ion 3 .7.
3.5.2.1. Basal Metabolic Rate
On the basis of BMR measu rements fo r 26 species weighing 3 .5 to 600 kg, Kleiber
(1961) esti mated that BMR was r elated to body weight in mammal s acco rding t o the 3/4
power :
3-26

BMR (kcal /day) = 70 Wt0.75 (kg) ± 0.004
Boddingt on' s (1978) analysi s produced simi lar results:
[3-42]
[3-43]BMR (kcal /day) = 75 Wt0.73
3.5.2.2. Resti ng Metabolism
(kg) ± 0.013
Stahl (1967) used an exten sive database (349 sp ecies) to determine slight ly highe r
value s for RMR than had been determined for BMR (Sect ion 2 .5.2.1):
RMR (kcal /day) = 80 Wt0.76
3.5.2.3. Field Metabolic Rate
(kg) [3-44]
Based on doubly -labeled water measu rements wi th 23 species of placental
mammal s, Nagy (1987) develope d an equation r elating FMR t o body weight :
FMR (kjoule s/day) = 3.35 Wt0.813 (g), or placental mammal s
FMR (kcal /day) = 0.800 Wt0.813 (g)
[3-45]
The slope of 0.813 is s ignif icantly highe r th an the BMR s lope s of 0. 73 to 0.76 repor ted
abov e. Thus , the FMR does not appe ar to be a con stan t mu ltiple of BMR ov er a range of
body size s as was th e case i n birds. However , no FMR measu rements h ave yet b een made
on shrews o r other very acti ve smal l mammal s, and whales wer e include d in t he FMR data
set (Nagy, 1987).
Separating t he herbiv ores from non- herbiv ores, Nagy (1987) develope d two
addit iona l equations :
FMR (kjoule s/day) = 5.943 Wt0.727
FMR (kcal /day) = 1.419 Wt0.727
(g), or herbivores [3-46]
(g)
3-27

�
�
FMR (kjoule s/day) = 2.582 Wt0.862 (g), or non- herbiv ores [3-47]
FMR (kcal /day) = 0.6167 Wt0.862 (g)
Separating r odents from other anim als, Nagy (1987) found:
FMR (kjoule s/day) = 10.51 Wt0.507 (g), or rodents [3-48]
FMR (kcal /day) = 2.514 Wt0.507 (g)
Nagy (1987) esti mated the accu racy o f th e doubly -labeled water method t o be ± 8 percent
or better .
To nor malize FMR to body weight (e. g., kcal /kg-day), divide the daily FMR by body
weight . In Sect ion 3 .7, we pr ov ide th e regression stati sti cs fo r Nagy's (1987) equations
(inc luding sample size and t he regression c oeffi cient) and informat ion t hat allow s one to
esti mate a 95-p ercent co nfidence interval for an FMR value predicted for a speci fied body
weight .
3.5.2.4. Temperature and Metabolic Rate
Few researchers have attemp ted general multiple r egressions of metabolic r ate wi th
bot h body mass an d temp erature for mammal s. However , sever al relations hips h ave been
investi gated qualitati vely (Peter s, 1983):
� Low temp eratures induc e a greater propor tiona l r ise in metabolic r ate
relative to basal metabolic r ate in smal ler mammal s th an in larger ones.f
� At high temp eratures, metabolic r ate increases to increase blood f low an d
evapor ative cooling ( e.g., pant ing) .
fThis is because conduct ance and heat loss f or a gi ven t hermal gradi ent between body t emperature and am bient temperature ri se more sl ow ly with body si ze than do basal metabol ic rates (Peters, 1983).
3-28

�
�
�
Peters (1983) develope d an equation r elating t he ratio of SMR t o BMR to thermal gradient
for mammal s:
SMR/BMR = 0.068 (thermal gradient in �C) Wt-0.182 (kg) [3-49]
Thus, stan dard metabolic co sts i ncrease relative to basal metabolis m at low er
temp eratures, but less so for larger than for smal ler mammal s.
3.5.3. Reptiles and A mphibians
Most rept iles and amphibia ns tend t o have mu ch lower metabolic r ates th an birds or
mammal s because th ey are poik ilot hermic . For examp le, at temp eratures si mi lar to normal
body temp eratures of birds and mammal s (around 3 7 to 39�C), resting metabolic r ates o f
rept iles and amphibia ns tend t o be only 10 to 20 percent of thos e of bir ds and mammal s of
simila r body weight (B ennett an d Dawso n, 1976). In th is sect ion, we pr ov ide so me
examp les of allometr ic equations fo r metabolic r ate. Because meta bolic r ate depends on
body temp erature, which in poik ilot herms can vary sub stan tially over time, we recommen d
that th ose persons interested in esti mat ing metabolic r ates co nsult mo re complete
treatmen ts o f th e subject, including t hermoregula tion in poik ilot herms (e.g., Bennett an d
Dawso n, 1976; Congdon e t al., 1982; Duellman and Trueb, 1986; Feder and Burggren, 1992;
Harless an d Morlock, 1979; Hutchins on, 1979).
3.5.3.1. Basal and Resti ng Metabolic Rates
Robins on et al. (1983) develope d an equation f or th e relations hip b etween BMR and
body mass fo r repti les and amphibia ns at 20�C:
BMR (Watts) = 0.19 Wt0.76 (kg), or [3-50]
BMR (kcal /day) = 3.92 Wt0.76 (kg)
Thus , the BMR of hom eotherms (Sect ions 3. 5.1 and 3.5.2) is appr ox imatel y 30 ti mes th e
BMR of poik ilot herms at this ambie nt temp erature (Peters, 1983). The differ ence in
3-29

�
�
�
metabolic r ates b etween homeotherms an d poik ilot herms is l essen ed when poik ilot herms
modif y their body temp eratures by behavioral adjustmen ts (su ch as basking in t he sun) .
Andrews an d Pough (1985) used multiple r egression a nalysi s to evaluate th e
relations hip b etween metabolic r ate and three var iables—mass, temp erature, and stan dard
or resting metabolic state—fo r snakes an d lizards. From a total of 226 observat ions on 107
species (between 20 and 30�C for mo st obs ervations ), they develope d the follow ing
equation:
MR (ml O2/hr) = 0.013 Wt0.80 (g) × 100.038 tem peratu re (�C) [3-51]
× 100.14 metabolic state
where MR equals ei ther SMR or RMR and metabolic state eq uals zero (0) for stan dard
metabolis mg and equals 1 for resting metabolis m.h The Q10 value s for th e inf luence of
temp erature on metabolic r ate (i .e., quot ient of th e rate measu red at one temp erature
divided by th e rate measu red at a temp erature 10�C low er) were 2.4 for resting metabolis m
and 1.4 for stan dard metabolis m. Thus SMR de pended less o n ambient temp erature than
did RMR.
Equation 3-51 is based on adult anim als and s hould not be used to esti mate
metabolic r ates o f juvenile sn akes an d lizards. Andr ews an d Pough (1985) reviewed
allometr ic equations r elating r esting metabolic r ate to body weight wi thin s pecies and
found t hat the expone nts were signif icantly low er th an the value of 0.80 in Equation 3-51.
See Andrews an d Pough (1985) for intraspeci fic allometr ic mo dels fo r th is group.
3.5.3.2. Free-Living Metabolic Rates
Nagy (1987) develope d an equation f or th e relations hip b etween FMR and body size
in igua nid liza rds:
gMeasured for fasti ng individuals during the period of normal inacti vity (at n ight for most sp ecies).hMeasured for fasti ng individuals during the period of normal acti vity (dayti me for most sp ecies).
3-30

�
�
� �
FMR (kjoule s/day) = 0.224 Wt0.799 (g), or [3-52]
FMR (kcal /day) = 0.0535 Wt0.799 (g)
Bennett an d Nagy (1977) esti mated that th e ratio of FMR to EMR for lizards is 2. 0.
Robins on et al. (1983) esti mated the value to be 2.9, assu ming t hat liza rds rest at
maintenance levels fo r 8 hour s per day at 35�C.
Feder (1981, 1982) presented equations r elating FMR t o body size of unr estr ained
ranid ( frog) tadpole s at 25�C:
dry mass (mg ) = 0.047 (wet mass) 1.06 (mg) [3-53]
and
FMR (µlO2/hr) = 2.5 (dry mass) 0.878 (mg), or [3-54]
FMR (mlO2/day) = 0.06 (dry mass) 0.878 (mg)
Assu ming 1 millilit er of oxygen is metabolic ally equiv alent to appr ox imatel y 4.80 calories
(Dawso n, 1974):
FMR (cal /day) = 0.288 (dry mass) 0.878 (mg) [3-55]
Burggren et al . (1983) esti mated Q10 values fo r metabolic r ates fo r bullf rog la rvae of 1.87
between temp eratures of 15 and 25�C and of 2.41 between temp eratures of 25 and 33�C.
Q10 value s for a second r anid s pecies (Rana berlandi eri) were simi lar (1.97 and 1.76,
respecti vely). Thus, the metabolic r ate fo r ranid f rogs appr ox imatel y double s wi th each 10-
degree rise in temp erature over th is range of temp eratures.
The equations presented in th is sect ion s how th at poik ilot herm metabolic r ate
depends str ongly on temp erature. The avai lable literature on the subject i s exten sive and
complex, and again, interested readers are encouraged to consult su bstan tive tr eatmen ts
of th e subject (see r eferences ci ted in the introduc tion t o Sect ion 3 .5.3).
3-31

3.6. MATH PRIMER AND UNIT CONVERSIONS
To assi st r eaders in using or modif ying a llo metr ic equations presented in th is
Handbook or in us ing a llo metr ic equations presented in the open literature, we provide a
brief su mmar y of loga rithm and pow er func tions in Sect ions 3. 6.1 and 3.6.2. Sect ion 3 .6.3
cont ains fr equent ly used unit conv ersion facto rs.
3.6.1. Summary of Operations Inv olv ing Logarithm s
log 1 = 0
log ( N1 N2) = log N 1 + log N 2
log ( N1 / N2) = log N 1 - log N 2
log ( 1 / N1) = -log N 1
clog ( N1 ) = c log N 1
= log ( N11/c) = (1/c) log N 1log c r oot of N1
3.6.2. Summary of Operations Inv olv ing Pow ers
aW Wb = Wa+b
a b(W ) = Wab
a a a(W W ) = W1 W21 2
aW / Wb = Wa-b
aW / W = Wa-1
1/Wb = W-b
0W = 1a a a(W1 / W2) = W1 /W2
c root of W a = (Wa)1/c = Wa/c
3-32

3.6.3. Unit Conv ersions
3.6.3.1. Approximate Facto rs for Metabolic Equations
1 kg dry mass
1 kg dry mass
1 kg wet mass
1 kg fat
tissu e densi ty
1 kg wet mass
1 kg dry mass
1 ml O2
= 3 to 10 kg wet mass (Peters, 1983)
(Peters, 1983)= 22 × 106 joule s
= 2 to 7 × 106 joule s
= 40 × 106 joule s
= 1 kg/liter
= 1 × 1015 µm3
= 0.4 kg carbon
= 20.1 joule s
= 4.8 calories
(Peters, 1983)
(Peters, 1983)
(Peters, 1983)
(Peters, 1983)
(Peters, 1983)
(Peters, 1983)
(Dawso n, 1974)
0.4047 hectar es (ha)259 ha1 × 10-4 ha100 ha
3.6.3.2. Exact C onversions
Area 1 acre = 1 square mi le mi2) = 1 square meter (m2) = 1 square ki lometer (km2) =
Lengt h 1 inch 1 foot =
1 mi le (mi)
Volum e 1 m3
Mass 1 ounce (oz) 1 pound ( lb) 1 lb
= 2.54 centimeter s (cm) 0.3 meter s (m) = =
= =
= = =
30.48 cm 1.61 ki lometer s (km)
1 × 103 liters (L) 31 × 106 cm
28.35 grams (g ) 453.6 g 0.4536 kilogr ams (kg )
3-33

Work and energy (fo rce × distan ce) 1 joule (J) = 1 kg-m2/s2
= 0.239 calories (cal )
Power (en ergy per unit ti me) 1 watt (W ) = 1 kg-m2/s3
= 1 joule /s = 20.64 kcal /day
1 ml O2/s = 0.0446 mMol O 2/s = 1.43 mg O2/s
3.7. ESTIMATING CONFIDENCE INTERVALS
A commonly r epor ted measu re of th e precision of esti mat ing log Y fr om log W t (or Y
from Wt) fo r allometr ic equations is th e stan dard error (SE) of log Y :
log Y = log a + b log W t ± SE of log Y [3-2]
The SE of log Y is th e stan dard error of th e esti mate o f log Y fr om log W t at a value of log
Wt th at represents th e mean of th e log W t value s us ed to esti mate th e allometr ic
relations hip. This value cannot be used to esti mate a co nfidence interval (CI) for a log Y
value predicted from log W t value s ot her th an the mean log W t value . The CI of a pr edicted
log Y value is smal lest at th e mean log Y and mean log W t value s and inc reases as log W t
for the species of interest d eviates fr om mean log W t. Thus , to esti mate th e CI for a single
predicted value of Y, one also must kn ow th e samp le size and the mean of th e log W t
value s us ed in de veloping t he allo metr ic equation, whic h many in vesti gators do not report.
Nagy (1987), however , did pr ov ide su ffi cient stati sti cal informat ion t o esti mate a 95-
percent CI for a predicted value of Y given any val ue of Wt fo r his fr ee-living ( field)
metabolic r ate (FMR) and f ood ingestion ( FI) rate equations . In this sect ion, we out line
Nagy's sh ort-cu t fo r esti mat ing t his CI and pr ov ide th e stati sti cal values requir ed for each
of Nagy 's equations presented in th is Handbook .
3-34

To esti mate 95-p ercent CIs fo r the predicted FMR and FI r ate, use th e values fr om
Table 3-3 (for FI rate equations) or 3-4 (for FMR equations ) in t he follow ing f ormula :
______ 2 0.595% CIlog y = log y ± c [d + e (log W t - log W t) ]
where y is FMR in k ilojoule s/day or FI in gr ams (d ry weight )/day. Log W t is th e log of th e
body weight in gr ams o f th e species fo r which y is being esti mated . Log W t bar is th e
mean log W t of th e species us ed to develop t he allo metr ic equation. Values for c, d, e, and
log W t bar are prov ided in T ables 3-3 and 3-4. Tables 3-3 and 3-4 also provide samp le
sizes (N), regression c oeffi cients (r 2), and SE esti mates fo r b and log a in t he applic able
equations .
3-35

Table 3-3. Regressi on Statistics f or Nagy's ( 1987) Allometric Equat ions f or Food I ngest ion Rates for Free-Living Animals
Regressi on Statistics f or Allometric Equat ions f or Food I ngest ion (FI) Rates (Dry Matter Ingest ion) Rates of Free-Living Mammals, Birds, and Li zards . Equations are in the form Y = aWt where Y is Food Ingestion Rate (in gra ms dry weight /day) and Wt is body weight ofb
species s (g rams wet weight). ______
95% CI = log FI ± c [d + e (log W t - log W t) ]log FI(species s) (species s) (species s) 2 0.5
Group subgroup
Equation a log a (SE log a) b (SE b) N r2
______ log Wt c d e
Birds 3-3 0.64 -0.188 (0.060) 0.651 (0.028) 50 0.919 1.983 0.347 1.020 0.026
passeri nes 3-4 0.40 -0.400 (0.075) 0.850 (0.053) 26 0.915 1.378 0.158 1.038 0.480
non- passeri nes 3-5 0.30 -0.521 (0.132) 0.751 (0.048) 24 0.919 2.638 0.401 1.042 0.061
seabirds 3-6 0.49 -0.306 (0.187) 0.704 (0.061) 15 0.911 2.958 0.399 1.067 0.109
Eutherian Mammals (i.e., placental)
3-7 0.23 -0.629 (0.065) 0.822 (0.026) 46 0.958 2.196 0.425 1.022 0.015
rodents 3-8 0.62 -0.207 (0.194) 0.564 (0.119) 33 0.421 1.598 0.434 1.030 0.313
herb ivores 3-9 0.58 -0.239 (0.109) 0.727 (0.039) 17 0.960 2.566 0.405 1.059 0.041
Iguanids
herb ivores 3-12 0.019 -1.713 (0.123) 0.841 (0.059) 5 0.985 1.896 0.358 1.200 0.278
insecti vores 3-13 0.012 -1.890 (0.037) 0.773 (0.038) 20 0.958 0.870 0.151 1.050 0.279
3-36
Source: Nagy, 1987.

Table 3-4. Regressi on Statistics fo r Nagy's (1987) Allometri c Equations for Free-L iving (Field) Metabolic Rates
Regressi on Statistics fo r Allometri c Equations for Free-Living Metabolic Rates (FMR) of Free-Living Mammals, Birds, and Lizards. Equations are in the form Y = aWt where Y is FMR (in k ilojoul es/day) and Wt is body weight of species s (grams wet weight ). b
______ 95% CI = log FMR ± c [d + e (log W t - log W t) ]log FMR(species s) (species s) (species s)
2 0.5
Group subgroup
Equation a log a (SE log a) b (SE b) N r2
______ log Wt c d e
Birds 3-34 10.9 1.037 (0.064) 0.640 (0.030) 50 0.907 1.983 0.368 1.020 0.026
passeri nes 3-36 8.89 0.949 (0.059)
non- passeri nes 3-37 4.79 0.681 (0.102)
seabirds 3-38 8.02 0.904 (0.187)
non- seabi rds 3-39 21.1 1.325 (0.081)
0.749 (0.037) 26 0.899 1.378 2.014 0.026
24 0.899 2.638 2.014 0.026
15 0.911 2.958 0.399 1.067
35 0.709 1.565 0.297 1.029
0.0014
0.749 (0.037) 0.0014
0.704 (0.061) 0.109
0.1130.440 (0.049)
Eutherian Mammals (i.e., placental)
3-45 3.35 0.525 (0.057) 0.813 (0.023) 46 0.967 2.196 0.371 1.022 0.015
rodents 3-48 10.5 1.022 (0.141) 0.507 (0.087) 33 0.524 1.598 0.316 1.030 0.313
3-37
herb ivores 3-46 5.94 0.774 (0.109)
non- herbi vores 3-47 2.58 0.412 (0.058)
Iguanids 3-52 0.224 -0.650 (0.029)
0.727 (0.039) 17 0.959 2.566 0.406 1.059
29 0.977 1.980 0.321 1.035
25 0.981 1.075 0.161 1.040
0.041
0.862 (0.026) 0.027
0.799 (0.023)
Source: Nagy, 1987.
0.088

3.8. REFERENCES
Andrews, R. M.; Pough, F. H. (1985) Metabolis m of squ amate r epti les: allometr ic and ecologic al relations hips . Physiol. Zool. 58: 214-231.
Bartholo mew, G. A.; Cade, T. J. (1963) The water economy o f land birds. Auk 80: 504-539.
Bennett, A. F.; Dawso n, W. R. (1976) Metabolis m. In: Gans, C.; Dawso n, W. R., eds. The biology of r ept ilia : v. 5, Physiology A . New York, NY: Academic Press; pp. 127-223.
Bennett, P. M.; Harvey, P. H. (1987) Acti ve and rest ing metabolis m in bir ds: allo metr y, phy loge ny and e cology . J. Zool. Lond. 213: 327-363.
Bennett, A. F.; Nagy, K. A. (1977) Energy exp endit ure in f ree-ranging liza rds. Ecology 58: 697-700.
Boddingt on, M. J. (1978) An absolute meta bolic scope fo r acti vi ty. J. Theor. Biol. 75: 443-449.
Buechner, H. K.; Golley, F. B. (1967) Preliminary esti mat ion of en ergy flow in Uga nda ko b (Adenota ko b thomasi Neumann). In: Petrusewi cz, L., ed. Seconda ry pr oduc tivi ty of ter restr ial ecosystems. Warszawa-Krakow; pp. 243-254.
Burggren, W. W.; West, N. H. (1982) Changing r espir atory im por tance of gills , lungs an d skin dur ing metamo rphos is in t he bullf rog Rana catesb eiana. Physiol. Zool. 56: 263-273.
Bur ggr en, W. W.; Feder, M. E.; Pinde r, A. W. (1983) Temperature and the balance between aerial and aquatic respir ation in t he larvae of Rana berlandi eri and Rana catesb eiana. Physiol. Zool. 56: 263-273.
Calder, W. A. (1981) Scaling of phy siologic al processes i n homeothermic animals. Ann. Rev. Physiol. 43: 301-322.
Calder, W. A.; Braun, E. J. (1983) Scaling of osmotic regula tion in mammal s and birds. Am. J. Physiol. 244: R601-R606.
Congdon, J. D.; Dunha m, A. E.; Tink le, D. W. (1982) Energy budgets an d life histo ries of rept iles. In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia , phy siology D; phy siologic al ecology : v. 13. New York, NY: Academic Press; pp. 233-271.
Dawso n, W. R. (1954) Temperature regula tion a nd water requir ements o f th e brown and abert to whees, Pipilo fuscu s and Pipilo aberti . Univ . Calif ornia Publ. Zool. 59: 81-124.
3-38

Dawso n, W. R. (1974) Appendix : conv ersion facto rs fo r unit s us ed in t he sympos ium . In: Paynter , R. A., ed. Avian energetics. Cambridge , MA: Nuttal Ornithologic al Club; Publi cation no. 15.
Drent, R. H.; Stonehouse, B. (1971) Thermoregulatory responses o f th e Peruvian penguin Spheniscu s hum bol ti . Comp. Bioc hem. Physiol. A: Comp. Physiol. 40: 689-710.
Duellman, W. E.; Trueb, L. (1986) Biology of amphibia ns. New York, NY: McGraw-Hill Book Company.
Dunn, E. H. (1980) On the var iabilit y in e nergy allo cation of n estling bir ds. Auk 97: 19-27.
Ellis , H. I. (1984) Energetics o f fr ee-ranging seabir ds. In: Whit tow, G. C.; Rahn, H., ed. Seabird energetics. New York, NY: Plenum Press; pp. 203-234.
Farlow, J. O. (1976) A consideration of th e trophic dynamics of a late Cretaceo us large-dinos aur co mmunit y (Oldm an Formation) . Ecology 57: 841-857.
Feder, M. E. (1981) Effect o f body size , trophic state, time of day, and experimental str ess on ox ygen cons umpt ion of anur an larvae: an experimental assessmen t and evalua tion of th e lit erature. Comp. Bioc hem. Physiol. A: 70: 497-508.
Feder, M. E. (1982) Effect o f developmental stag e and body size on ox ygen cons umpt ion of anuran larvae: a reappraisal . J. Exp. Zool. 220: 33-42.
Feder, M. E.; Burggren, W. W., eds. (1992) Environmental physiology of th e amphibia . Chicago, IL: Universi ty o f Chicago Press.
Gans, C.; Dawso n, W. R., eds. (1976) Biology of th e rept ilia : v. 5, phy siology A . New York, NY: Academic Press.
Gessaman , J. A. (1973) Methods of esti mat ing t he energy co st of fr ee existen ce. In: Gessaman , J. A., ed. Ecologic al energetics of hom eotherms. Monogr . Ser. 20 ed. Loga n, UT: Utah State Universi ty Pr ess; pp. 3-31.
Golley, F. B. (1961) Energy val ues of ecologic al mater ials. Ecology 42: 581-584.
Harless, M.; Morlock, H., eds. (1979) Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.
Hemmi ngsen, A. M. (1960) Energy meta bolis m as related to body size and r espir atory surfaces, and its ev olut ion. Rept . Steno Me m. Hosp. Nord. Insulin La b., Part II 9: 1-95.
Hume, I. D. (1982) Digesti ve phys iology and nut rition of mar supia ls . Cambridge , Engla nd: Cambridge Univ ersi ty Press.
3-39

Hutchins on, V. H. (1979) Thermoregula tion. In: Harless, M.; Morlock, H., ed. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 207-227.
Hutchins on, V. H.; Whit ford, W. G.; Kohl, M. (1968) Relation of body size and s urface ar ea to gas exch ange in anurans. Physiol. Zool. 41: 65-85.
Jackso n, D. C. (1979) Respiration. In: Harless, M.; Morlock, H., ed. Turtles: perspecti ves and research. Toronto, Canada: John W iley and Sons , Inc.; pp. 165-191.
Kendeigh, S. C. (1969) Energy responses o f birds to their thermal environments. Wilson Bull. 81: 441-449.
Kendeigh, S. C. (1970) Energy requir ements fo r existen ce in relation t o size of bir d. Condor 72: 60-65.
Kendeigh, S. C.; Dol' nik , V. R.; Govr ilov , V. M. (1977) Avian energetics. In: Pinowski , J.; Kendeigh, S. C., eds. Graniv orous bir ds in ecosystems. Cambridge , MA: Cambridge Universi ty Pr ess.
King, J. R. (1974) Seasonal alloc ation of ti me and energy r esour ces i n birds. In: Paynter , R. A. Jr., ed. Avian energetics. Cambridge , MA: Nuttal Ornithologic al Club; Publi cati on no. 15; pp. 4-85.
Kleiber, M. (1961) The fi re of l ife. New York, NY: John W iley.
Koplin, J. R.; Collopy , M. W.; Bamman n, A. R.; et al . (1980) Energetics o f two winter ing raptors. Auk 97: 795-806.
Lamprey, H. F. (1964) Esti mat ion of th e large mammal densi ties, biomass an d energy exch ange in the Tarangir e Game Reserve and the Masai Steppe in Tanganyika. E. Afr . Wild. J. 2: 1-46.
Lasiewski , R. C.; Calder, W. A. (1971) A preliminary allometr ic analysi s of respiratory var iables in r esting bir ds. Resp. Phys. 11: 152-166.
Lasiewski , R. C.; Dawso n, W. R. (1967) A reexami nation of th e relation b etween stan dard metabolic r ate and body weight in bir ds. Condor 69: 12-23.
Lilly whit e, H. B.; Maderson, P. F. (1982) Skin str uctu re and permeabilit y. In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia : v. 12, phy siology C; phy siologic al ecology . New York, NY: Academic Press; pp. 397-442.
Mahmoud, I. Y.; Kli cka, J. (1979) Feeding, dr ink ing, and excr etion. In: Harless, M.; Morlock, H., ed. Turtles: perspecti ves an d research. Toronto, Canada: John W iley and Sons , Inc.; pp. 229-243.
3-40

Medway, W.; Kare, M. R. (1959) Water metabolis m of th e growing do mesti c fo wl wi th speci fic reference to water balance. Poult ry Sci . 38: 631-637.
Meeh, K. (1879) Oberflachenmess unge n des mensclic hen Kor pers. Z. Biol. 15: 426-458.
Mils om, W. K.; Chan, P. (1986) The relations hip b etween lung v olum e, respir atory dr ive and breathing p atter n in the turtle Chrysemys p icta. J. Exp . Biol. 120: 233-247.
Minnic h, J. E. (1982) The use of water . In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia : v. 12, phy siology C; phy siologic al ecology . New York, NY: Academic Press; pp. 325-395.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Nagy, K. A.; Peterson, C. C. (1988) Scaling of water flux rate in animals. Berkeley, CA: Universi ty o f California Press.
Peters, R. H. (1983) The ecologic al impli cations of body size . Cambridge , Engla nd: Cambridge Univ ersi ty Press.
Reiss, M. J. (1989) The allometr y of growth and reproduc tion. Cambridge , Unit ed Kingdom : Cambridge Univ ersi ty Press.
Robbins , C. T. (1983) Wildlif e feeding a nd nut rition. New York, NY: Academic Press.
Robins on, R. W.; Peters, R. H.; Zimmer mann, J. (1983) The effects o f body size an d temp erature on metabolic r ate of organisms. Can. J. Zool. 61: 281-288.
Rubne r, M. (1883) Uber den Einfluss d er Korpergrosse au f Sto ff- und Kr aftwesch sel . Z. Biol. 19: 535-562.
Schmidt-Nielsen, K. (1970) Energy meta bolis m, body size and pr obl ems o f sca ling. Fed. Proc. Am. Soc. Exp . Biol. 29: 1524-1532.
Schmidt-Nielsen, K. (1972) How animals wo rk. Cambridge , MA: Cambridge Univ ersi ty Press.
Seibert, H. C. (1949) Differ ences b etween migrant and non- migr ant bir ds in f ood a nd water intake at var ious temp eratures and phot operiods . Auk 66: 128-153.
Seymo ur, R. S. (1982) Physiologic al adaptations to aquatic lif e. In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia : v. 13, phy siology D; phy siologic al ecology . New York, NY: Academic Press; pp. 1-51.
Skadhauge, R. (1975) Renal and cloacal transport of sal t and water . Symp . Zool. Soc. London 3 5: 97-106.
3-41

Stahl, W. R. (1967) Scaling of r espir atory var iables in mammal s. J. Appl. Physiol. 22: 453-460.
U. S. Environmental Protect ion A gency. (1990) Inter im met hods fo r development of inha lation r eference co ncentrations , review dr aft. Washingt on, DC: Offi ce of Research and Development; EPA report no. EPA/600/8-90/066A.
Utter , J. M. (1971) Daily energy exp endit ures of fr ee-liv ing pur ple mar tins ( Progne subi s) and m ockingbir ds (Mimus polyglotto s) wi th a comparison of two northern popula tions of mo ckingbir ds [Ph .D. disser tat ion] . Rutgers, NJ: Rutgers Univ ersi ty; 173 pp.
Walsberg, G. E. (1983) Avian ecologic al energetics. In: Farner, D. S.; King, J. R.; Parkes, K. C., eds. Avian biology , v. 7. New York, NY: Academic Press; pp. 161-220.
Walsberg, G. E.; King, J. R. (1978) The relations hip of th e exter nal surface ar ea of birds to ski n surface ar ea and body mass. J. Exp. Biol. 76: 185-189.
Whit ford, W. G.; Hutchins on, V. H. (1967) Body size and metabolic r ate in salamanders. Physiol. Zool. 40: 127-133.
Wiens, J. A. (1984) Modelling t he energy r equir ements o f seab ird popula tions . In: Whit tow, G. C.; Rahn, H., eds. Seabird energetics. New York, NY: Plenum Press; pp. 284.
Zar, J. H. (1968) Standard metabolis m comparisons b etween orders of birds. Condor 10: 278.
3-42

� �
4. EXPOSURE ESTIMATES
This sect ion pr ov ides equations to esti mate o ral doses o f ch emical contami nants
for wildlif e, along w ith a dis cussion of dos e esti mates fo r other exposure routes. Secti on
4.1 prov ides general dos e equations . Equations fo r dr ink ing water exposures are
presented in Sect ion 4 .1.1, follow ed by equa tions fo r dietary exp osures in Sect ion 4 .1.2. In
the dietary exp osure sect ion, data on the caloric and water content of var ious food t ypes
and diet assi mi lation effi ciencies are also provided. An equation a nd d ata to fac ilitate
esti mat ing do ses r eceived through s oil or sedim ent ing estion a re dis cussed in Secti on
4.1.3. Sect ions 4. 1.4 and 4.1.5 prov ide a qua li tati ve discu ss ion of inha lation a nd de rmal
dose esti mates. Sect ion 4 .2 descr ibes co nsiderations fo r analyses o f uncer tainty i n
exposure assessmen ts. References ar e provided in Sect ion 4 .3.
4.1. GENERAL DOSE EQUATIONS
EPA's (1992a) Framewo rk for Ecological Risk A ssessmen t defines exp osure as th e
co-occu rrence of or contact b etween a str esso r and an ecologic al compone nt . When
assess ing r isks o f exp osure to chemical contami nants, potential dose is often the metr ic
used to quant ify expos ure. Potential dos e is define d as the amount of ch emical present in
food or water ingested , air inha led, or mater ial applie d to the skin ( U.S. EPA, 1992b).
Potential dos e is an alogous to the admini ster ed dose in a toxici ty test. Because exp osure
to chemicals in the environment is generally inadver tent, rather than administer ed, EPA's
(1992b) Guidelines fo r Exposure Assessmen t use th e term potential dose rather than
administer ed dose.
A general equation f or esti mat ing dos e for intake p rocesses i s:
t2
Dpot � � C(t) IR(t) dt [4-1]
t1
4-1

where Dpot is th e total potential dose over time (e.g., total mg contami nant intake b etween
t1 and t2), C(t) is th e contami nant co ncentration in t he con tacted medium at time t (e.g., mg
contami nant/kg medium), and IR(t) is th e intake r ate of th e contami nated medium at time t
measu red as mass ingested or inha led by an anim al per unit ti me (e.g., kg medium /day). If
C and IR are constan t over time, then the total potential dose can be esti mated as:
Dpot = C × IR × ED [4-2]
where ED is th e expos ure dur ation a nd equals t2 - t1.
Therefore, if C and IR are constan t, the potential average daily dose (ADDpot ) for the
dur ation of th e expos ure, nor malize d to the anim al's body weight (e. g., mg/kg-day), is
esti mated by dividing t otal pot ential dos e by ED and by body weight (B W):
ADDpot = (C × IR × ED) / (BW × ED), or [4-3]
ADDpot = (C × IR) / BW
If C or IR vary over time, they may b e averaged over ED. However , it is not always
appropriate to average intake o ver the enti re exposure duration: For examp le, a given
quant ity of a chemical might acu tely pois on an anim al if ing ested in a single event, but i f
that amount is averaged over a longe r period, effects m ight not be exp ected at al l.
Simi larly, developmental effects o ccu r only dur ing s peci fic periods of g estat ion or
developm ent. A toxicologis t should be cons ult ed to determine which effects may b e of
concern given the exposure patter n and chemicals of interest. For carcinoge nic
compounds , it may b e more appropriate to average exposure over the animal 's l ifeti me.
Again, address an y quest ions to a toxicologis t.
In addit ion, IR and BW can be co mbine d int o a nor malize d ing estion or inha lation
rate (NIR) (e.g., kg medium /kg body weight - day):
NIR = IR / BW [4-4]
4-2

Therefore,
ADDpot = C × NIR [4-5]
It is important to rememb er that NIR can vary wi th changes in a ge, size, and r eproduc tive
statu s of an animal.
Two other var iables often are used in calculations of average daily dose. A
frequency term (FR) is used to denot e the fraction of th e time that an anim al is expos ed to
contami nated media. In ecologic al expos ure assessmen ts, th is ter m often is used when athe foraging r ange of an anim al is larger th an the area of con tamina tion. An absorption
facto r (ABS) is used when an esti mate o f absorbed dose rather than potential dose is
desired. It is co mmonly assu med that absorption in t he species of conc ern in t he field is
the same as i n the test o rganism, so no absorption facto r is needed. However , if
absorption is exp ected to differ , a ratio of th e absorption facto rs would be used in t he
expos ure equation.
4.1.1. Drinking Water
Figur e 4-1 presents two wildlif e oral expos ure equations co rresponding t o two patter ns of
cont amina tion of water :
(1) the anim al obt ains so me of its dr ink ing water from a co ntami nated source
and t he remainde r fr om unc ontami nated sources; and
(2) the animal consumes d rinking water from sever al sources co ntami nated at
differ ent levels.
aThe frequency t erm shoul d be est imated w ith care. For exam ple, if a feature at tract ive to wildlifeis cont aminated, an animal may spend a proport iona lly longe r t ime in the cont aminated area. Similarly, if only part o f an animal's th eoreti cal foraging range has su itable habitat, the animal may spend more ti me feeding in that habitat. Finally, animals may avo id areas o r media with contamination they can detect.
4-3

Figur e 4-1. Wildlif e Dose Equations fo r Drink ing W ater Exposures
One Sour ce of Cont amina tion
ADD = C × FR × NIR [4-6]pot
Differ ent Sources With Varying L evels of Cont amina tion
n
ADD = � (C × FR ) x NIR [4-7]pot i i
i=1
ADDpot = Potential averag e daily dose (e.g., in mg/kg-day).
C = Averag e contaminant co ncentrati on in a single water so urce (e.g., in mg/L or in mg/kg, because 1 l iter o f water w eighs 1 kg).
FR = Fraction of total water ingestion f rom the cont aminated water sourc e (unitless).
NIR = Norm alized water ingestion ra te (i.e., fraction of body weight cons umed as water per uni t time; e.g., in g/g-day)
and
thCi = Averag e contaminant co ncentrati on in the i water so urce (e.g., in mg/L).
FRi = Fracti on of water co nsumed fro m the ith water so urce (unitless).
n = Number of contaminated water so urces.
In the fi rst case, the distr ibut ion a nd mean value of th e contami nant co ncentration in t he
one sour ce could be d etermined. In the second case, the differ ent water sources ar e likely
to be character ized by differ ent mean levels of co ntami nation, and c ons umpt ion f rom th ese
sources w ould be weight ed by th e fraction ( FRi) of th e anim al's total daily water inge stion
obtained from each source. FR (or FRi) in Figur e 4-1 is a fu nct ion of th e degree of overlap
of th e contami nated water source(s) an d the animal 's home range. If th e area of th e
contami nated water source is larger than the typ ical home range for the species, FR could
4-4

equal one fo r man y indiv idua ls . The num ber of indiv idua ls fo r whic h FR equals one could
be esti mated from informat ion on popula tion de nsity, dis tr ibut ion, and s ocial str ucture.
For large, mobile anim als, th e area of con tamina tion may b e smal ler than the area over
whic h a single anim al is lik ely to move. In these cases, FR for an animal wi th the
contami nated area enti rely wi thin it s hom e range can be esti mated using inf ormation on
the home range, attr ibutes o f th e contami nated area, and drinking be havior of th e anim al.
Home range esti mates s hould be used wi th care because (1) th e area in which an animal
moves var ies wi th sever al facto rs, including r eproduc tive statu s, seaso n, and habitat
quality; (2) most an imals do not drink or feed random ly wi thin t heir hom e range; (3) the
term home range has been used inconsisten tly in the literature; and (4) esti mates o f home
range can vary su bstan tially wi th the measu rement tec hnique used. In this Handbook an d
acco mpanying A ppendix , we have tr ied to identi fy cl early wh ich esti mates o f home range
correspond t o a daily acti vi ty an d foraging hom e range.
When us ing hom e range d ata, we recommen d that users co nsult th e Appendix
tables fo r the species of interest to become fam ilia r wi th how esti mates o f home range size
vary wi th geogr aphic ar ea, seaso n, typ e of habitat, animal reproducti ve statu s, and
measu rement tec hnique . The Appendix table s pr ov ide bot h the sample size and a br ief
descr iption of th e method us ed to esti mate h ome range size, which can help indi cate th e
robustn ess o f an esti mate an d whether it is l ikely to over - or unde resti mate h ome range
size. For mark-and-recapture studies, the number of recaptures per animal is provided
when pos sible to assi st th e user in determining t he degree to whic h the repor ted value s
may unde resti mate tr ue home range size. If a stu dy indi cated that th e home range
esti mate i s l ikely to include ar eas outsi de of th e animals' usual acti vi ty r ange (e.g., distan t
egg- laying s ites u sed only onc e per seaso n), th is would be not ed in t he Appendix table s,
and t he value would not be inc lude d in Cha pter 2. Some anim als us e a fixed "hom e base"
some distan ce fr om feeding gr ounds su ch as a rook ery. For these animals, we have
repor ted foraging r adius (th e distan ce th ey will t ravel to a feeding a rea). Foraging r adius
can be used to determine whether the animal might feed or drink in a given contami nated
area.
4-5

4.1.2. Diet
Wildlif e can be expos ed to con tamina nts in one or more compone nts of th eir die t,
and differ ent co mpone nts can be con tami nated at differ ent levels. In th is sect ion, we
out line methods of esti mat ing f ood ing estion r ates th at allow to tal doses to be esti mated
when differ ent co mpone nts of th e die t are con tami nated, either at si mi lar or differ ent levels
(Sect ion 4 .1.2.1). We also prov ide d ata on caloric co ntent of foods and assi mi lation
effi ciencies th at can be used in the dose equations prov ided (Sect ion 4 .1.2.2).
4.1.2.1. Dose Equations
Figur e 4-2 presents a g eneric equation f or esti mat ing or al doses o f co ntami nants i n
food f or wildlif e species. FRk is a func tion of th e degree of overlap of th e k typ e ofth
simplest case, the normalized ing estion r ate fo r each food t ype, NIRk, is know n on a wet-
Figur e 4-2. Wildlif e Dose Equations fo r Dietary Exp osures
m
ADDpot = � (Ck × FRk × NIRk) [4-8] k=1
ADDpot = Potential averag e daily dose (e.g., in mg/kg-day).
Ck = Average cont aminant conc entration i n the kth type of food ( e.g., in mg/kg wet weight).
FRk = Fract ion of intake of the kth food t ype that is cont aminated (uni tless) . For example, if the kth com ponent of an ani mal's di et were sal mon, FRk for sal mon would equal the fracti on of the salmon consumed that is contaminated at level Ck. If all of the salmon co nsumed were co ntaminated at level Ck, then FRk
woul d equal one.
NIRk = Normalized ingestion ra te of the kth food t ype on a wet-weight basis (e.g., in g/g-day).
m = Number of cont aminated food t ypes.
4-6

contami nated forage or prey and the animal 's home range (see Sect ion 4 .1.1). In the
weight basis, and E quation 4-8 can be used directl y. In many cases, however , NIRk is
unknown or has been determined for laboratory diets th at differ signif icantly from natural
diets i n terms o f cal oric val ue per unit wet we ight . Ingestion r ates b ased on relatively dry
laboratory diets m ight unde resti mate th e amount of food a fr ee-liv ing a nim al cons umes.
There are sever al ways to esti mate NIRk, depending on t he typ e of inf ormation t hat
is avai lable. If d ietary co mposi tion is exp ressed as th e number of each prey typ e captured
on a daily basis (Nk), esti mat ing t he nor malize d ing estion r ate fo r each prey typ e (NIRk)
requir es only one step :
NIRk = (Nk × Wtk) / BW [4-9]
where Wtk is th e body weight of th e kth prey typ e and BW is th e body weight of th e
predator.
Figur e 4-3 presents a fl ow ch art depict ing e quations th at can be used if th e
propor tion of th e die t for a giv en food t ype has been measu red or esti mated on a wet-
weight basis. These equations may r equir e esti mates o f th e free-living metabolic r ate
(FMR) of th e organis m and t he metaboliza ble en ergy (ME) of th e organis m's forage or prey.
Esti mated FMRs can be found in t he species pr of iles in Cha pter 2, and a llo metr ic equations
for esti mat ing FMR on t he basis of body weight ar e prov ided in Cha pter 3 (Sect ion 3 .5). ME
should be averaged over the food t ypes when ME on a wet-we ight basis (e.g., cal /g wet
weight) d iffer s su bstan tially among t he di ffer ent foods . Sect ion 4 .1.2.2 descr ibes how to
esti mate ME.
A common s ituation facing s omeone conduc ting a wildlif e expos ure assessmen t fo r
predators is th at in a key stu dy, dietary co mposi tion is exp ressed as a percentage of th e
total num ber of prey captured ov er a period of ti me in stead of as a p ercentage of th e total
wet we ight of food ingested daily. Because so me prey can be substan tially larger than
others (e.g., rabbits co mpared wi th voles), and because ME of differ ent typ es of prey may
4-7


Figur e 4-4. Esti mat ing NIR k Based on Differ ent ME Values When Dietary Composi tion Is Expressed as Percentage of Total Prey Captured
Step 1: Calculate th e metaboliza ble en ergy (ME) cont ent of each prey or food t ype on a wet-we ight basis:
ME(wet wt) k = GE(wet wt) k × AEk [4-13]
Step 2: Esti mate th e average number of prey (or other food i tems) co nsumed each day:
Navg = FMR / (weighted average prey ME) m
Navg = FMR / (� PNk × Wtk × ME(wet wt) k) k=1
Step 3: Calculate IRk:
IRk = Nto t × PNk × Wtk
Step 4: Normalize to body weight :
NIRk = IRk / BW
[4-14]
[4-15]
[4-16]
thME(wet wt)k = Metabol izable energy i n the k prey or f ood t ype (e.g., in kcal /g wet weight ). thGE(wet wt)k = Gross energy cont ent of the k food t ype (e.g., in kcal /g wet weight ).
thAEk = Assi milation ef ficiency f or the speci es for the k food t ype (uni tless) .Navg = Average num ber of prey ( or ot her f ood i tems) eaten each day.FMR = Free-liv ing metabolic rate (e.g., in kcal/day).m = Number of di fferent types of prey or ot her foods .
thPNk = Proport ion of the total num ber of prey that is compos ed of the k prey type(unitless). It often is the case th at larger numbers o f relatively sm all prey and smaller numbers o f relatively large prey are cap tured. (If th e to tal number of prey of each typ e captured each day are rep orted in the literatu re, calcu lations of IRk are very si mple [i .e., Nk × Wtk] and step s 1 and 2 are unnecessary. )
thWtk = Body weight of an indi vidua l of the k food t ype (e.g., in g). thIRk = Ingestion ra te of the k food t ype (e.g., in g/day).
4-9

and the weighted average ME of th e prey. Given Navg , the ingestion r ate fo r each prey typ e
(IRk) can be computed on a wet-we ight basis and nor malized to body weight (NIRk).
Because Navg is esti mated using pr ey weight, di ffer ent si zes of th e same p rey sp ecies (e.g.,
smaller and lar ger fi sh) should be separated into appropriate si ze intervals to reduce
uncer tainty i n the esti mate.
4.1.2.2. Energy Content and Assi mi lation Effi ciencies
The total or gross en ergy (GE) co ntent of a food typ e is a function only of
character isti cs o f th e food. On the other hand, metabolizable energy (ME) depends on
character isti cs o f both the food and th e organism eating it. To clarify the meaning of ME,
Figur e 4-5 presents a fl ow ch art of energy utilization by animal s. Digestible energy in a diet
is GE consu med minus the energy lost as feces; digestible energy effi ciency (DE) is
digestible energy divided by GE. ME is GE consu med minus the energy lost as both feces
and ur ine. Assi milation effi ciency (AE, also cal led metabolizable energy effi ciency) i s ME
divided by GE. Rearranging this relationship, ME is equal to GE of the diet multiplied by
the animal 's AE for th e diet as shown in Figur e 4-6, Equation 4-17. General ME values can
be found in T able 3-1 or more speci fic ones cal culated from GE co ntent of th e food and th e
AE of the animal eating th at food, as discu ssed below.
The GE conten t of food typically is repor ted using one (or mo re) of three measu res:
(1) energy per unit total dry weight, (2) energy per unit ash-free dry weight, or (3) energy
per unit fresh bio mass (i .e., per unit wet we ight) (Górecki , 1975). Caloric co ntent per unit
total dry weight is obtained dir ectl y fr om th e combust ion of dried mater ial in a calorimeter .
Ash-free dry weight is the dry weight after subtract ing th e ash conten t.b The ash-free dry-
weight calor ic value exceed s th e total dry-weight calor ic value by the ratio of the total dry
weight to the ash-free dry weight. Typically, animal (excl usive of th ick sh ells) and plant
mater ials are 1 to 10 percent ash on a wet-we ight basis and 5 to 30 percent ash on a dry-
weight basis (Ashwell-Er ickso n and Elsner, 1981; Cummi ns and Wuych eck, 1971;
bAsh cons tituents typically include calcium carbona te (e.g., shell), calcium phos phate (vertebra te bone ), and hy drated silic a salts.
4-10


Figure 4-6. Metabol izable Energy (ME) Equation
ME = GE × AE [4-17]
where:
ME = Metabolizable energy (e.g., in kcal/g)
GE = Gross energy (e.g., in kcal/g)
AE = Assi milation effi ciency (u nitless)
This Handbook assum es ME and GE are est imated on a w et-weight basi s. To est imate ME or GE of the k th food t ype on a w et-weight basi s f rom dry- weight measurem ents, the fol low ing equations can be used:
GE(wet wt)k = GE(dry wt)k × (1 - proport ion w aterk) or [4-18]
GE(wet wt)k = GE(dry wt)k × (dry weight
k/wet weightk) [4-19]
and ME(wet wt)k = ME(dry wt)k × (1 - proport ion w aterk) or [4-20]
ME(wet wt)k = ME(dry wt)k × (dry weight
k/wet weightk) [4-21]
Hunt , 1972). The ash content of th e diet is not meta bolize d and t hus does not prov ide
energy to the anim al. Figur e 4-6 (Equations 4-18 through 4-21) illus trates h ow th e caloric
cont ent per unit of fr esh bio mass can be obtained by adjus ting t he dry-weight value b ased
on the water content of th e biomass. A summar y of GE co ntents o f man y wildlif e food
typ es are presented in Tables 4-1 a given species on a wet-we ight basis tends to be mo re
var iable than caloric co ntent on a dry-weight basis because plants, and to a lesser degree
animals, vary in their water content depending on e nvironm ental condit ions . Ash- free dry-
weight calor ic value s are not presented because i t is not appropriate to use th em wi th the
equations and A Es in t his ch apter . Ash c ont ents are acco unted for in the AEs presented in
Table 4-3.
4-12
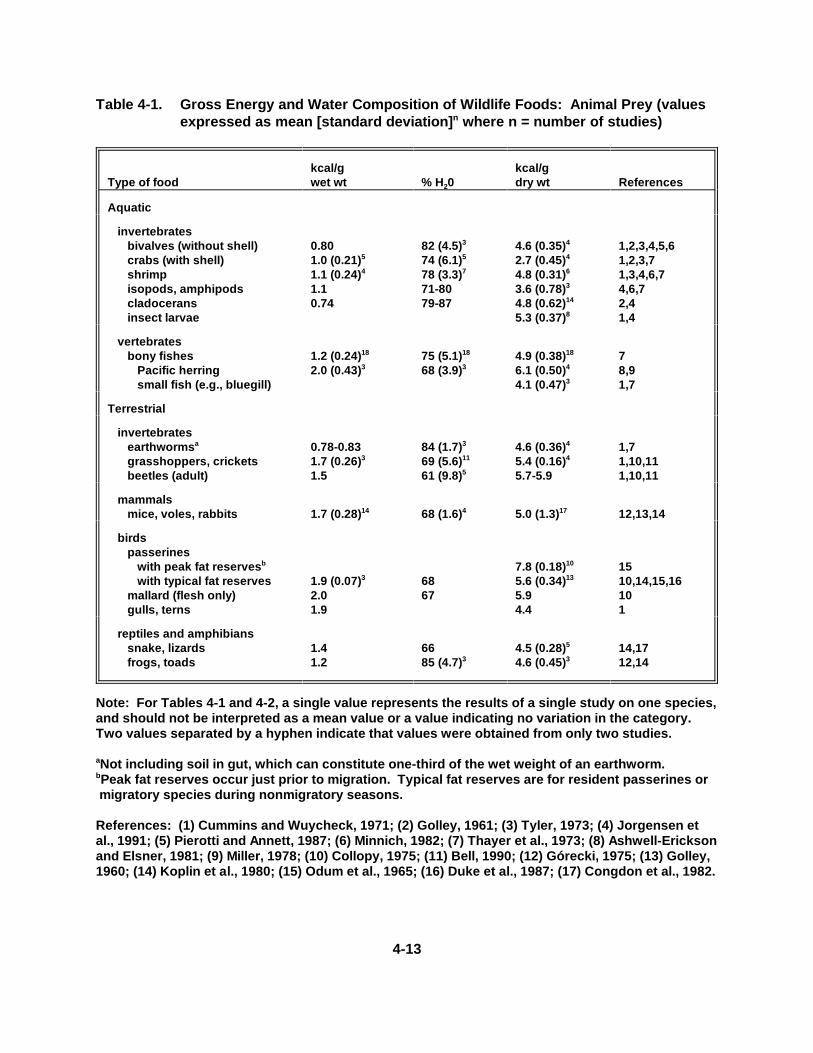
Table 4-1. Gross En ergy and Water Composi tion of W ildlif e Foods : Anim al Prey (values nexpressed as mean [stan dard deviation] where n = num ber of studie s)
Type of food
Aquat ic
invertebrates bivalves (without shel l) crabs (with shel l) shr imp isopods, amphi pods cladocer ans insect larvae
vertebrates bony fishes
Paci fic her ring small fis h (e.g., bluegill)
Terrestrial
invertebrates earthwormsa
grasshopper s, cr icket s beet les (adul t)
mammals mice, vol es, rabbi ts
bi rds passer ines
with peak f at reserves b
with typical fat reserves malla rd (flesh only) gulls , terns
rept iles and am phi bians snake, lizards fro gs, toads
kcal /g kcal /g wet wt % H20 dry wt References
0.80 82 (4.5)3 4.6 (0.35)4 1,2,3,4,5,6 1.0 (0.21)5 74 (6.1)5 2.7 (0.45)4 1,2,3,7 1.1 (0.24)4 78 (3.3)7 4.8 (0.31)6 1,3,4,6,7 1.1 71-80 3.6 (0.78)3 4,6,7 0.74 79-87 4.8 (0.62)14 2,4
5.3 (0.37)8 1,4
1.2 (0.24)18 75 (5.1)18 4.9 (0.38)18 7 2.0 (0.43)3 68 (3.9)3 6.1 (0.50)4 8,9
4.1 (0.47)3 1,7
0.78-0.83 84 (1.7)3 4.6 (0.36)4 1,7 1.7 (0.26)3 69 (5.6)11 5.4 (0.16)4 1,10,11 1.5 61 (9.8)5 5.7-5.9 1,10,11
1.7 (0.28)14 68 (1.6)4 5.0 (1.3)17 12,13,14
7.8 (0.18)10 15 1.9 (0.07)3 68 5.6 (0.34)13 10,14,15,16 2.0 67 5.9 10 1.9 4.4 1
1.4 66 4.5 (0.28)5
1.2 85 (4.7)3 4.6 (0.45)3 14,17 12,14
Note: For Tables 4-1 and 4-2, a single value represents th e resu lts o f a single stu dy on one species, and s houl d not be interpre ted as a mean value or a value indi cating no v ariation i n the category . Two values sep arated by a hyphen indicate th at val ues were obtained fro m only tw o stu dies.
aNot includi ng s oil in gut , which can cons titute one-third of the wet weight of an earthworm. bPeak fat reserves o ccu r just p rior to migration. Typical fat reserves are fo r resi dent passeri nes or migrat ory speci es duri ng nonm igratory seasons.
Referen ces: (1) Cummins and Wuych eck, 1971; (2) Golley, 1961; (3) Tyler, 1973; (4) Jo rgensen et al., 1991; (5) Pierotti and Annett, 1987; (6) Minnich, 1982; (7) Thayer et a l., 1973; (8) Ashwell-Erickso n and Elsner, 1981; (9) Mille r, 1978; (10) Collopy, 1975; (11) Bell, 1990; (12) Górecki , 1975; (13) Golley, 1960; (14) Ko plin et al., 1980; (15) Odum et al., 1965; (16) Duke et a l., 1987; (17) Congdon et al., 1982.
4-13

algae aquatic macrophy tes emergent vegetati on
Terrestri al
monoc ots young grasses mature dry grasses
dicots leaves root s bul bs , rhi zomes stem s, branches seeds
fruit pul p, skin pulp, ski n, seeds
kcal /g Type of food wet wt a % H20
Aquatic
0.41-0.61 84 (4.7)3
87 (3.1)3
[45-80] b
1.3 70-88 7-10
85 (3.5)3
9.3 (3.1)12
1.1 (0.30)3 77 (3.6)3
kcal /g dry w t
2.36 (0.64)4
4.0 (0.31)12
4.3 (0.13)3
4.2 4.3 (0.33)5
4.2 (0.49)57
4.7 (0.43)52
3.6 (0.68)3
4.3 (0.34)51
5.1 (1.1)57
2.0 (3.4)28
2.2 (1.6)10
Referen ces
1,2,31,2,41,2,4
5,61,5,7,8
992,7,1096,9,11,12
10,13 10
Note: For Tables 4-1 and 4-2, a single value represents th e resu lts o f a single stu dy on one species, and s houl d not be interpre ted as a mean value or a value indi cating no v ariation i n the category . Two values sep arated by a hyphen indicate th at val ues were obtained fro m only tw o stu dies.
a Few determ inations of the energy co ntent o f plants h ave been made on a wet-weight basisbecause pl ants fluct uate widely in water cont ent dependi ng on envi ronm ental condi tions.
b Values in brackets rep resent to tal range of field measurements, instead of values fro m only tw o studi es, as for the remainder of the table. Buchsbaum and Valiela (1987) found t he water cont ent of the emergent marsh vegetati on Spartina alternif lora, S. patens, and Junc us gerardi to decrease over a su mmer fro m 80 to 60 percen t, 70 to 45 percen t, and 78 to 61 percen t, resp ecti vely, as th e marsh dri ed. In cont rast, they found a subm erged macrophy te to maintain water cont ent within a few percent throughout the season.
Referen ces: (1) Cummins and Wuych eck, 1971; (2) Jo rgensen et al., 1991; (3) Minnich, 1982; (4) Boyd and Goodyear, 1971; (5) Davi s and Golley, 1963; (6) Dro zdz, 1968; (7) Golley, 1960; (8) Kendeigh and West, 1965; (9) Golley, 1961; (10) Karaso v, 1990; (11) Dice, 1922; (12) Robel et al ., 1979; (13) Levey an d Karaso v, 1989.
4-14
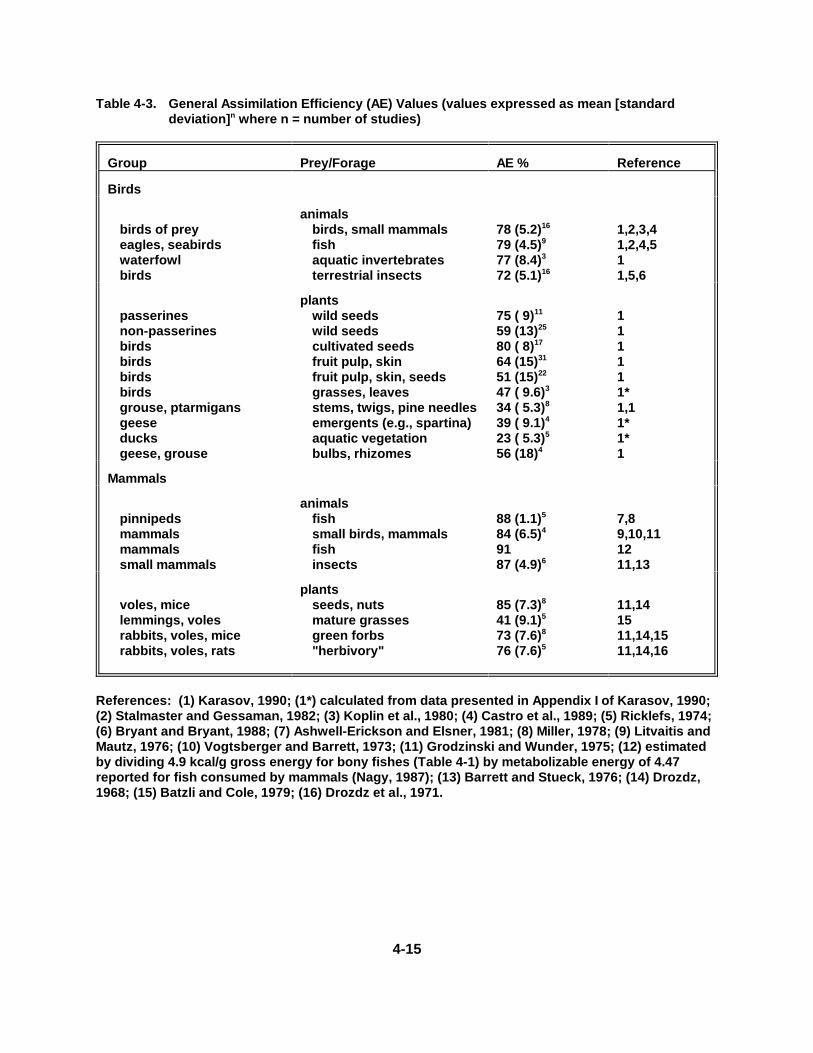
Table 4-3. General Assi milation Effi ciency (AE) Values (val ues exp ressed as mean [stan dard ndeviation] where n = num ber of studi es)
Group Prey/Forage
Birds
animals birds of prey birds, small ma mmals eagles, seabirds fish waterfowl aquatic inverteb rates birds terrestri al insects
plants passeri nes wild seeds non- passeri nes wild seeds birds cu ltivated seeds birds frui t pul p, skin birds fru it pulp, ski n, seeds birds grasses, leaves grous e, ptarmigans stem s, tw igs, pine needles geese emergents (e.g., sparti na) ducks aquatic veg etati on geese, grouse bul bs , rhi zomes
Mammals
animals pinni peds fish mammals small b irds, mammals mammals fish small ma mmals insects
vo les, mice lemmin gs, voles rabbits, vo les, mice rabbits, vo les, rats
plants seeds, nuts mature grasses green forbs "herb ivory"
AE % Referen ce
78 (5.2)16 1,2,3,4 79 (4.5)9 1,2,4,5 77 (8.4)3 1 72 (5.1)16 1,5,6
75 ( 9)11 1 59 (13)25 1 80 ( 8)17 1 64 (15)31 1 51 (15)22 1 47 ( 9.6)3 1* 34 ( 5.3)8 1,1 39 ( 9.1)4 1* 23 ( 5.3)5 1* 56 (18)4 1
88 (1.1)5 7,8 84 (6.5)4 9,10,11 91 12 87 (4.9)6 11,13
85 (7.3)8
41 (9.1)5
73 (7.6)8
76 (7.6)5
11,14 15 11,14,15 11,14,16
Referen ces: (1) Karaso v, 1990; (1*) cal cu lated fro m data presented i n Appendi x I of Karaso v, 1990; (2) Stalmaster an d Gessam an, 1982; (3) Ko plin et al., 1980; (4) Castro et al., 1989; (5) Rickl efs, 1974; (6) Bryan t and Bryan t, 1988; (7) Ashwell-Erickso n and Elsner, 1981; (8) Mille r, 1978; (9) L itvai tis and Mautz, 1976; (10) Vogt sberger and B arret t, 1973; (11) Grodz inski and Wunder, 1975; (12) est imated by dividing 4 .9 kcal/g gros s energy for bony fishes (Table 4-1) by metabol izable energy of 4.47 reported for fi sh consumed by mammals (Nagy, 1987); (13) Barrett an d Stueck, 1976; (14) Dro zdz, 1968; (15) Batz li and Cole, 1979; (16) Dro zdz et al., 1971.
4-15

Table 4-3 summar izes AEs fo r sever al differ ent typ es of foods and s pecies. Assi mila tion
effi ciency i s a fu nct ion of bot h the cons umer sp ecies' phy siology and t he typ e of die t.
Facto rs th at reduce man y sp ecies' abilit y to assi mi late th e energy co ntained in food
inc lude th e ash cont ent of th e die t and t he percentage of r elatively indig estible organic
mater ials su ch as ch itin (arthropods ) or cellulos e (plants). The highe r th e ash cont ent, th e
low er th e AE, all e lse be ing e qual.
Fat co ntent also influences GE. For examp le, carbohy drates (ap proximatel y 4.3
kcal /g) and proteins (approximatel y 5.7 kcal /g) typ ical ly provide about half as many
calories per gram as fat (ap proximatel y 9.5 kcal /g) (Peters, 1983). Thus, smal l changes in
fat co ntent of animal tissu es or plant seed s cau se s ignif icant changes in t heir calor ic value .
For examp le, just p rior to fal l migration, passer ine birds have ach ieved peak fat d eposi tion
and average 7.8 kcal /g dry weight . Non-migr ating p asser ines (i .e., permanent residents o r
migr atory species dur ing nonm igr ating seaso ns) aver age only 5. 6 kcal /g dry weight . Two
references wi th substan tial compila tion of d ata on caloric co ntent of biologic al mater ials
are Jorgensen et al . (1991) and Cummi ns and Wuych eck (1971). The latter include s
exten sive data on inver tebrates.
Figur e 4-7 provides a samp le calculation of food ing estion r ates u sing t he
methodology out line d abov e.
4.1.3. Soil and Sedim ent Ingestion
In this sect ion, we review inf ormation on t he ing estion of soil a nd s edim ent for th e
species inc lude d in t his Handbook (and s imila r sp ecies). Despite th e potential importance
of soil a nd s edim ent ing estion a s a rout e of expos ure of wildlif e to environm ental
contami nants, data to quanti fy th ese inge stion r ates ar e limi ted at th is ti me.
4-16

Figure 4-7. Example of Estimating Food I ngestion Rates for Wildlife Species From Free-Living Metabolic Rate and Dietary Composit ion: Male Mink
1. Estimate Field Metabolic FMR (kcal /day) = 0.6167 (g Wt)0.862
Rate (FMR) [Equation 3-47] = 0.6167 (1,040)0.862
= 246 (kcal /day)
2. Normalize to Body Weight NFMR (kcal /g-day) = 246 (kcal /day)/1,040 (g Wt) a
(Wt) [Equation 3-40] = 0.24 (kcal /g-day)
3. Estimate Averag e Metabolizable Energy (ME ) of Diet [Equation 4-12]avg
Dietary Item (k=5)
Proport ion of Diet (P )k
b
Gross Energy (GE )k
c
(kcal/g wet wt )
Assi milation
Effi ciency (AE )k
d
Metabolizable Energy (ME )k
(kcal/g wet wt) (ME = GE × AE )k k
(P × ME )k
Fish 0.85 1.2 0.91 1.1 0.93
Crustacea 0.04 1.1 0.87 0.96 0.038
Amphibia 0.03 1.2 0.91 1.1 0.033
Birds/ Mammals
0.06 1.8 0.84 1.5 0.090
Vegetati on 0.02 1.3 0.73 0.95 0.019
ME (kcal /g wet w t) = �(P × ME ) 1.1avg k e
4. Est imate Total NIR (g/g-day) = 0.24 (kcal /g-day)total
Normalized Ingestion Rate 1.1 (kcal/g wet wt) (i.e ., ME )avg
(NIR ) [Equatio n 4-11] = 0.22 (g/g-day)total
5. Estim ate Prey-specific NIR (g/g-day) = 0.85 (P ) × 0.22 (g/g-day)fish fish
Normalized Ingest ion Rat es = 0.19 (g/g-day) (e.g., NIR ) [Equatio n 4-10]fish
Body weight for Mont ana popul ation i n the sum mer (Mitche ll, 1961). a
Dietary com posi tion based on Al exander (1977). b
Values f rom Tables 4-1 and 4-2 (for veget ation, assum ing val ue for young gr asses) . c
Values f rom Table 4-3 (for veget ation, assum ing gr een forbs; for crust acea, assum ing equi valent AEd
for insect s; for amphi bia, assum ing equi valent to mammals consum ing f ish). In this example, ME is the same as the ME value for fish, which com prises 85 per cent of the diet. e
avg
k
k
=k
4-17

4.1.3.1. Backg round
Soil is ing ested both intentiona lly and inc identally by man y species of wildlif e and
can be a signif icant expos ure pathway fo r some co ntami nants (A rthur and Alldr edge, 1979;
Garten, 1980). Many ungulates d elibe rately eat s oil t o obt ain nut rients; some may tr avel a
considerable distan ce to reach cer tain areas (sal t l icks) th at are used by man y animals.
Some birds gather mud in their beaks fo r nest-building, and ot hers cons ume it fo r cal cium
(Kreulen and Jager, 1984). Many animals can incidental ly ingest s oil w hile groom ing,
digging, grazing c los e to the soil, or feeding on i tems th at are covered wi th soil ( such as
root s and t ubers) or cont ain s edim ent (such as mollu scs). Earthworms ingest s oil dir ectl y;
the soil in t heir gut s may b e an important exp osure medium fo r anim als that eat th ese
organisms (B eyer et al ., 1993).c
Soil ing estion r ates h ave been esti mated for only a few w ildlif e species and w ere not
availa ble in t he publis hed lit erature for mo st of th e anim als in t his Handbook . The
percentage of soil ing ested is often esti mated from th e acid-insoluble ash cont ent of
wildlif e scats o r digesti ve tr act co ntents. Scat an alysi s on smal l animals is often diffi cult
because scat ar e smal l. Soil ing estion by large mammal s also has been esti mated using
ins oluble ch emical tracers (Mayland et al ., 1977) and using stan dard x-ray diffr acti on
analysi s (Gar ten, 1980).
4.1.3.2. Methods
Garten (1980) esti mated the amount of soil in t he gastr ointesti nal (GI) tract o f a
smal l mammal (the hispid c otton r at) using t he follow ing e quation:
I = (S - F)W [4-22]
cSeed-eating birds often consume "gri t" to aid in digesti on, which makes th em vulnerable topoisoning by granular formulations of pestic ides and fertiliz ers. In this secti on, however, we restri ct o ur discu ssi on to so ils and sediments, which are co mposed of much smaller parti cle sizes.
4-18

where I equals th e amount of soil in t he GI tract, S equals th e ratio of insoluble ash to dry
contents i n the GI tract, F equals th e ratio of insoluble ash to dry cont ents in fescu e (the
dominant veg etat ion in t he rat's habitat), and W equals th e dry weight of GI-tract co ntents.
It is al so pos sible to esti mate s oil ing estion r ates fr om th e acid-insoluble ash
content of th e animal 's scat b ecause th e percentage of aci d-insoluble ash in m ineral soil is
much highe r (usually at least 90 p ercent) th an in plant or animal tissu e (usually no more
than a few p ercent). Beyer et al . (in press) u sed scat samp les to esti mate th e fract ion of
soil in t he die t for sever al species. The equation f or this esti mat ion a ppr oach is slight ly
more complicated than Equation 4-22, because i t acc ount s for dige stibilit y and t he mine ral
cont ent of th e soil. They found a signif icant correlation b etween the measu red and
predicted relations hips of th e ratio of acid- ins oluble ash to dry weight of scat an d the
percentage of soil in t he die t.
4.1.3.3. Results
Percent soil in t he die t for so me of th e selected and simi lar species include d in
Chapter 2 are inc lude d in T ables 4-4 and 4-5. Of th e species st udie d, the sandpipe r group,
which feeds on mud-dwelling inv ertebrates, was f ound t o have th e highest r ates o f
soil/ sedim ent ing estion (30, 18, 17, and 7.3 percent of d iet, respecti vely, for semi palmated ,
wester n, sti lt, and least sa ndpipe rs, al though only a single samp le was an alyzed for each
species). Wood ducks al so can ingest a high pr opor tion of sedim ent (24 percent) wi th their
food. Relatively high s oil in takes wer e esti mated for the raccoon ( 9.4 percent), an
omniv ore, and t he woodc ock (10.4 percent), which feeds exten sively on earthworms.
Other species th at eat ear thworms m ight be exp ected to exhibit si mila rly high s oil in takes.
The Canada goose, which browses o n grasses, also exhibit ed a high pe rcentage of soil in
its die t (8.2 percent). Soil ing estion was l owest fo r the white-fo oted mouse, meadow vo le,
fox, and box tu rtle (<2, 2.4, 2.8, and 4.5 percent, respecti vely). Box tu rtles, tortoises, and
other repti les, however , have been known to intentiona lly ing est s oil, perhaps for its
nutr ient co ntent (Kramer , 1973; Sokal , 1971). Beyer et al .'s (in press) d ata should be used
with caution, because er ror was i ntroduced by esti mat ing v ariables in
4-19

Table 4-4. Percent Soil or Sediment in Diet Estimated From Acid-Insolubl e Ash of Scat
Species
Birds
Canada goos e
Mallard
Wood du ck
Blue-winged tea l
Ring-necked du ck
American woodcock
Semipalmated sandpiper
Western sandpiper
Sti lt sandpiper
Least sandpiper
Mammal s
Red f ox
Raccoon
White-f ooted m ouse
Meadow vole
Repti les and Amphibians
Scat Samplesa
% Inso lub le Ash
Mean (SE) Range
Estimated %
Digestibilit y of Diet
Estimated Percen t Soi l
in Diet (dry w eight)
23 12 (1.5) 3.9 - 38 25 8.2
88 6.9 (1.1) 0.36 - 47 30 <2
7 24 (13) 0 - 75 60 11
12 2.3 (0.36) 0.72 - 5.1 60 <2
6 0.72 (5.5) 0.50 - 1.2 60 <2
7 22 (5.5) 6.3 - 40 55 10.4
1 56 70 30
1 42 70 18
1 40 70 17
1 24 70 7.3
7 14 (2.6) 4.8 - 25 70 2.8
4 28 (8.9) 13 - 50 70 9.4
9 8.5 (0.71) 5.7 - 11 65 <2
7 8.9 4.2 - 14 55 2.4 (1.2)
Eastern painted turtl e
9 21 (2.9) 11 - 41 70 5.9
Box tu rtl e 8 18 (6.5) 3.6 - 49 70 4.5
aFor the sandpi pers, the whi te-foot ed mouse, and t he meadow vol e, scat samples f rom more than oneanimal had t o be com bined into one sam ple to provi de suf ficient quant ity for chem ical analysis.
Sour ce: Adapt ed f rom Beyer et al. (in pr ess).
4-20

Table 4-5. Other Esti mates o f Percent Soil or Sedim ent in Die t
Species Estimated % soil in diet
(dry weight) Reference
Jackrab bit 6.3 Arthur and Gates 1988
Hisp id cotto n rats 2.8 Garten 1980
Shorebirds 10-60 Reeder 1951
the equation ( e.g., dig estibili ty) an d by th e smal l samp les th ey obtained from so me of th e
smal ler animals.
Other studie s of soil ing estion by sp ecies simila r to thos e presented in th is
Handbook ar e summar ized in Table 4-5. Sediment has been found in t he sto mach s of
white-fo oted mice (Gar ten, 1980) and ruddy ducks an d shovelers (Goodm an and Fis her,
1962). Sediment in the gut of tadpole s inha bit ing high way d rainages may b e respons ible
for high c onc entrations of lead detected in these organisms (B irdsal l et al ., 1986).
4.1.3.4. Dose Equations
To esti mate exp osures to contami nants i n soils or sedim ents from th e data
provided in Tables 4-4 and 4-5, Equation 4-23 (Figur e 4-8) can be used. If th e percent so il
in the diet is measu red on a dry-weight basis, as it usually is , total dietary intake s hould
also be expressed on a dry-weight basis.
4.1.4. Air
Inhalation t oxici ty value s and expos ure esti mates ar e usually exp ressed in unit s of
conc entration in a ir (e.g., mg/m3) rather th an as average daily doses. Assessmen t of th e
inha lation pa thway b ecomes co mplicated if th e toxici ty val ues mu st b e extr apolated from a
test sp ecies (e.g., rat) to a differ ent sp ecies (e.g., shrew). Inhalation t oxicologis ts
extr apolate to xici ty val ues fr om sp ecies to species on the basis of th e dose deposi ted and
retained in the respiratory tr act (th e dose th at is avai lable for absorption, dis tr ibut ion,
4-21

Figur e 4-8. Wildlif e Oral Dose Equation f or Soil or Sedim ent Ing estion E xpos ures
m ADDpot = (� (Ck × FS × IRtot al(dry wei ght) × FRk))/BW [4-23]
k=1
ADDpot = Potential averag e daily dose (e.g., in mg/kg-day).
thCk = Averag e contaminant co ncentrati on in so ils in the k foraging area (e.g., inmg/kg dry weight ).
FS = Fraction of soil in diet (as percentage of diet on a dry -weight basis divided by 100; unitless).
IRtotal = Food i ngest ion rat e on a dry- weight basi s (e.g., in kg/day). Nagy's ( 1987) equations for esti mating FI rates o n a dry-weight basis (presen ted in Secti on 3.1) can be used to esti mate a val ue for th is facto r. If the equations for esti mating FI rates o n a wet-w eight basi s presented i n Secti on 4.2 are used, conversi on to i ngesti on rates o n a dry-w eight basi s w oul d be necessary.
FRk = Fract ion of total food i ntake f rom the kth foragi ng area ( uni tless) .
BW = Body weight (e.g., in kg).
m = Total number of foraging areas.
metabolis m, and e lim ination) . Once the appr opr iate to xici ty b enchmark (in terms o f dose)
has been esti mated for the species of co ncern (e.g., shrew), the corresponding a ir
conc entration is esti mated based on the respiratory physiology of th at species. EPA uses
this approach because i t can account fo r nonline ar relations hips b etween exposure
conc entrations , inha led dos e, and dos e to the target or gan(s). Because of th e complexi ties
asso ciated wi th the extr apola tions , an inha lation t oxicologis t should be cons ult ed when
assess ing t his pathway.
The dose deposi ted, retained, and absorbed in the respiratory tr act i s a fu nct ion of
species anatomy and phy siology as well a s phy sicochemical properties of th e
contami nant. The assesso r will ne ed to cons ider facto rs su ch as th e target sp ecies' airway
4-22

size, branching p atter n, breathing r ate (volume and frequency), and clearance
mech anisms, as wel l as wh ether the contami nant is a gas or aerosol and whether its
effects ar e systemi c or confined to the respiratory tr act. Key informat ion on t he
cont amina nt inc lude s partic le size dis tr ibut ion ( for aerosols ), temp erature and vapor
pressu re (for gaseous agents), and pharmaco kinetic data (e.g., air/blood pa rtit ion
coeffi cients, metabolic parameter s). While phy siologic ally b ased pharmaco kinetic mo dels
have been usefu l for these cal culations , they are availa ble fo r only a few l aboratory
species. These issu es are discu ssed in detai l in Inter im Methods for Development of
Inhalation Reference Concentrations (U.S. EPA, 1990). Although t he doc ument speci fically
descr ibes how to calculate inha lation r eference co ncentrations fo r hum ans, th e pr inc iple s
are useful f or any air-breathing s pecies.
4.1.5. Dermal Exposure
Dermal toxici ty val ues and exposure esti mates ar e usually exp ressed as an
absorbed dos e result ing f rom skin c ontact wi th a contami nated medium . This expos ure
pathway can be of great importance to wildlif e, partic ularly when an anim al is dir ectl y
sprayed (Driver et al ., 1991). Dermal exposures may al so be a concern for wildlif e that
swi m or bur row. Dermal absorption of con tamina nts is a func tion of ch emical properties of
the contami nated medium , the permeabilit y of th e anim als' int egum ent, th e area of
integument in contact wi th the contami nated medium , and t he dur ation a nd p atter n of
contact. A full dis cussion of quant ifying a bsorbed dos e through t he skin is beyond t he
scope of this doc ument, and m any of th e requir ed parameter s have not been measu red for
wildlif e species. Readers int erested in pursuing t his expos ure pathway may f ind u sefu l
inf ormation in Dermal Exposure Assessmen t: Principles and Applicati ons (U.S. EPA,
1992c).
4.2. ANALYSIS OF UNCERTAINTY
In the risk assessmen t process, sever al sources o f uncer tainty s hould be evaluated,
inc luding t he unc ertaint ies asso ciated wi th the exposure assessmen t and the toxici ty
4-23

assessmen t. The follow ing sect ions dis cuss three sour ces o f uncer tainty r elated to the
exposure assessmen t: (1) natural var iabilit y in t he popula tion in qu estion, (2) uncer tainty
about popula tion pa rameter s as a co nsequence of l imi ts o n sampling t he popula tion ( i.e.,
sampling unc ertain ty), and (3) uncer tainty about mode ls used to esti mate val ues. There
are other categ ories of uncer tainties asso ciated wi th si te-sp eci fic r isk assessmen ts th at
also need to be cons idered (e.g., selection of sub stan ces o f co ncern, data gaps, toxici ty
assessmen ts). Addit iona l dis cussion of sour ces an d treatmen t of uncer tainty i s avai lable
in Framewo rk for Ecological Risk A ssessmen t (U.S. EPA, 1992a) and Guidelines fo r
Exposure Assessmen t (U.S. EPA, 1992b). For treatmen t of si te-sp eci fic uncer tainties in
particular, see th e Risk A ssessmen t Guidance fo r Superfund, Volume I; Human Health
Evaluation Manual (Part A) Inter im Final (U.S. EPA, 1989).
4.2.1. Natural Variation
As a review of th e data provided in th is Handbook makes cl ear, there is natural
var iation in t he value s exhibit ed by popula tions fo r all e xpos ure facto rs. Popula tion v alues
for some parameter s (e.g., body weight ) can assu me a normal distr ibut ion t hat can be
character ized by a mean and var iance. We have provided the stan dard deviation ( SD) as
the measu re of popula tion v ariance whenever poss ible . If a risk assesso r is co ncerned
with expos ures that might be exp erienced by anim als exhibit ing c haracter isti cs n ear the
extr emes o f th e popula tion' s dis tr ibut ion, the SD can be used wi th the mean value for a
nor mally dis tr ibut ed popula tion t o esti mate th e parameter value for animals wi th
character isti cs at sp eci fied point s in t he dis tr ibut ion ( e.g., 95th percenti le). We also have
provided the total range of val ues reported for each of th e exposure facto rs wh enever
pos sible . The ranges can be partic ularly he lpf ul f or parameter s th at are not normal ly
distr ibuted, such as home-range size.
Another aspect o f natural var iation, however , is th at differ ent popula tions or the
same popula tion a t di ffer ent ti mes o r locat ions can exhibit di ffer ent mean values fo r any
parameter (e.g., body weight ) and even differ ent var iances. We have tr ied to present
enough d ata to give users of th e Handbook a feel for the range of val ues th at differ ent
popula tions can assu me depending on ge ogr aphic location, seaso n, and other facto rs
4-24

�
(e.g., habitat q uality). We recommen d that r isk assesso rs review th e data presented in the
Appendix to appr eciate th e potential for var iation in t he parameter s of interest.
Dietary co mposi tion, in pa rtic ular, can vary markedly wi th seaso n, locat ion, and
availa bilit y of prey or forage. The latter facto r var ies wi th local condit ions and us ually is
not avai lable for risk assessmen ts. Thus, it can be one of th e larger sources o f uncer tainty
in w ildlif e expos ure assessmen ts. State an d local wildlif e experts might be able to help
speci fy th e local dietary habits o f a sp ecies of co ncern and should be cons ult ed if
scr eening a nalyses s uggest th at exp osure at levels of co ncern is a possibilit y.
4.2.2. Sampling Uncertainty
Another source of uncer tainty i n exposure esti mates r esults fr om l imi ted sampling
of popula tions . Esti mates o f a popula tion mean and var iance become mo re accu rate as
the number of samp les taken from th e popula tion inc reases. With only a few samp les fr om
a popula tion, our conf idence that the true popula tion mean is near the esti mated mean is
low; as th e number of samp les increases, our confidence increases. The stan dard error
(SE) of th e mean is equal to the var iance of th e popula tion ( �) div ided by th e squa re root of
the samp le size (n). SE can be esti mated from th e stan dard deviation of th e popula tion
divided by th e square root of n. SE can be used to calculate co nfidence l imi ts o n an
esti mate o f th e mean value for a popula tion. For a nor mally dis tr ibut ed popula tion, the 95-
percent co nfidence l imi t of th e mean is th e esti mated mean plus or minus appr ox imatel y 2
SEs fo r reasonable samp le sizes (e.g., n = at least 20).
Sampling unc ertaint y oc curs in m any areas of exp osure assessmen t. Cont amina nt
conc entration is one key p arameter subject to sampling e rror. For si te-sp eci fic r isk
assessmen ts, as th e number of environmental samp les increases, the uncer tainty about
the true dis tr ibut ion of value s decreases. Even wi th large samp le sizes, however , th is
uncer tainty can dominate th e total uncer tainty i n the exposure assessmen t. Other
parameter s subject to sampling e rror are the expos ure facto rs presented in th is Handbook .
One of our cr iteria fo r sel ecting v alues from th e Appendix to inc lude in Cha pter 2 was a
sample size large enough t o ensure that SE was only a few p ercent of th e mean value.
4-25

4.2.3. Model Uncertainty
Two m ain t ypes of mode ls ar e lik ely to be used in w ildlif e expos ure assessmen ts:
(1) allometr ic mo dels to predict co ntact-r ate parameter s (e.g., food ing estion r ates) an d (2)
fate an d transport mo dels to predict co ntami nant co ncentrations to whic h wildlif e are
exposed.
In this Handbook , we have tr ied to present stati sti cal confidence l imi ts asso ciated
wi th allometr ic equations wh enever poss ible . To reduc e the conf idence lim its asso ciated
wi th allometr ic mo dels, it is important to use a mo del derived from th e smal lest an d most
simila r taxonom ic/dietary group a ppr opr iate fo r the extr apola tion. For examp le, to esti mate
a metabolic r ate fo r a red-winge d bl ackb ird, it is preferable to use a meta bolic r ate mo del
derived from data on passer ines rather than a model derived from data on many differ ent
groups of bir ds (e.g., raptors, seabirds, geese), and best to use a mo del for Icter ids (th e
subf amily to whic h the red-winge d bl ackbir d be longs ) rather th an a mode l derived from
data on passer ines.
Uncertaint ies in e xpos ure mode ls can inc lude how well t he expos ure mode l or its
math emati cal expression a ppr ox imates th e true relations hips in t he field as well a s how
realisti c th e exposure model assu mptions ar e for th e si tuation a t hand. Judic ious fi eld
sampling ( e.g., of con tamina nt conc entrations in c ertain pr ey species) can help calibr ate or
confi rm esti mates i n the exposure model (e.g., food-chain exposures). Often a sensi tivi ty
analysi s can help a risk assesso r identi fy wh ich model parameter s and assu mptions ar e
most i mportant in determining r isk so that atten tion c an be fo cused on r educ ing
uncer tainty i n these elements.
4.3. REFERENCES
Alexander, G. (1977) Food of ver tebrate predators on trout water s in north central lower Michiga n. Michiga n Acad. 10: 181-195.
Arthur , W. J., III; Alldr edge, A. W. (1979) Soil ing estion by mule deer in nor th central Color ado. J. Range Manage. 32: 67-70.
4-26

Arthur W. J., III; Gates, R. J. (1988) Trace element intake vi a soil ing estion in pr onghor ns and in black-tai led jackr abbits. J. Range Manage. 41: 162-166.
Ashwel l-Erickso n, S.; Elsner, R. (1981) The energy co st o f fr ee existen ce fo r Bering S ea harbor and spotted seals. In: Hood, D. W.; Calder, J. A., eds. The Easter n Bering S ea shelf: oceanogr aphy and r esour ces: v. 2, Washingt on, DC: Departmen t of Commer ce; pp. 869-899.
Barrett, G. W.; Stueck, K. L. (1976) Caloric inge stion r ate and assi mi lation effi ciency o f th e short-tai led shrew, Blarina brevicauda. Ohio J . Sci . 76: 25-26.
Batzli, G. O.; Cole, F. R. (1979) Nutr itiona l ecology of mi crot ine r odents: dig estibilit y of forage. J. Mammal . 60: 740-750.
Bell, G. P. (1990) Birds and mammal s on an insect d iet: a primer on diet co mposi tion analysi s in r elation t o ecologic al energetics. Studie s Avian Biol. 13: 391-415.
Beyer , N.; Connor , E.; Gerould, S. (In pr ess) Esti mates o f soil ing estion by wildlif e. J. Wildl. Manage. 1993.
Beyer , W. N.; Staffo rd, C.; Best, D. (1993) Survey an d evaluation of con tami nants i n earthworms from conf ined dis pos al facilit ies for dredged mater ial in the Great Lakes. Environ. Monit . Assess. 24: 151-165.
Birdsal l, C. W.; Grue, C. E.; Anderson, A. (1986) Lead concentrations in bullf rog Rana catesb eiana and gr een frog R. clami tans inha bit ing high way d rainages. Environ. Poll. (Series A) 40: 233-247.
Boyd, C. E.; Goodyear , C. P. (1971) Nutr itive quality o f food in e cologic al systems. Arch. Hydrobiol. 69: 256-270.
Bryant, D. M.; Bryant, V. M. (1988) Assi mi lation effi ciency an d growth of nest ling insecti vores. Ibis 130: 268-274.
Buchsbaum, R.; Valiela, I. (1987) Variabilit y in t he chemistr y of estu arine plants an d its effect o n feeding by Canada geese. Oecologia (B erl.) 73: 146-153.
Buchsbaum, R.; Wilson, J.; Valiela, I. (1986) Digestibilit y of plant cons ti tuents by Canada geese and Atlantic brant. Ecology 67: 386-393.
Castr o, G.; Stoyan, N.; Myers, J. P. (1989) Assi mi lation effi ciency i n birds: a funct ion of taxon or food t ype? Comp. Bioc hem. Physiol. A. Comp. Physiol. 92: 271-278.
Collopy , M. W. (1975) Behavioral and predatory dynamics o f kestr els wi nter ing in t he Arcata B otto ms [master 's th esis]. Arcata, CA: Humboldt State Universi ty.
Congdon, J. D.; Dunha m, A. E.; Tink le, D. W. (1982) Energy budgets an d life histo ries of rept iles. In: Gans, C., ed. Biology of th e rept ilia : v. 13. New York, NY: Academic
4-27

Press; pp. 233-271.
Cummi ns, K. W.; Wuych eck, J. C. (1971) Caloric equivalents fo r investi gations in ecologic al energetics. Stuttg art, West Ger many: Internationa l Asso ciation of T heoretical and Applie d Lim nology .
Davis, D. E.; Golley, F. B. (1963) Principles in mamma logy . New York, NY: Van Nostrand Rheinhold.
Dice, L. R. (1922) Some facto rs affect ing t he dis tr ibut ion of th e prairie vole , forest d eer mous e, and pr airie deer mous e. Ecology 3: 29-47.
Driver, C. J.; Ligo tke, M. W.; Van Voris, P., et al . (1991) Routes o f uptake an d their relative cont ribut ion t o the toxicologic al respons e of nor thern bobw hit e (Colin us virg inianus) to an organophos phate pesti cide. Environ. Toxicol. Chem. 10: 21-33.
Drozdz, A. (1968) Digestibilit y and assi mi lation of natural foods in smal l rodents. Acta Theriol. 13: 367-389.
Drozdz, A.; Górecki , A.; Grodzins ki , W.; et al. (1971) Bioe nergetics of water voles (Arv icola ter restr is L.) from sout hern Moravia. Ann. Zool. Fennic i 8: 97-103.
Duke, G. E.; Mauro, L.; Bird, D. M. (1987) Physiology . In: Pendle ton, B. A.; Mills ap, B. A.; Cline , K. W.; et al., eds. Raptor management tec hnique s manua l. Washingt on, DC: Insti tute for Wildlif e Research, Nationa l Wildlif e Federation. Sci . Tech. Ser. No. 10; pp. 262-267.
Garten, C. T. (1980) Ingestion of soil by his pid c otton r ats, white-fo oted mice, and easter n chipm unk s. J. Mammal . 61: 136-137.
Golley, F. B. (1960) Energy dynamics o f a food c hain of an old- field c ommunit y. Ecol. Monogr . 30: 187-206.
Golley, F. B. (1961) Energy val ues of ecologic al mater ials. Ecology 42: 581-584.
Goodman, D. C.; Fisher, H. I. (1962) Func tiona l anatomy of th e feeding a pparatus in water fowl (Aves: Anatidae). Carbonda le, IL: Sout hern Illinois Univ ersi ty Press; 193 pp.
Górecki , A. (1975) Calorimetr y in ecologic al studie s. In: Grodzins ki , W.; Klekowski , R. Z.; Dunc an, A., eds. IPB handbook no. 24: methods fo r ecologic al energetics. Oxfo rd, London, Edinbur gh, Melbour ne: Blackwel l Scienti fic Publi cations ; pp. 275-281.
Grodzins ki , W.; Wunde r, B. A. (1975) Ecologic al energetics of smal l mammal s. In: Golle y, F. B.; Petrusewi cz, K.; Ryszkowski , L., eds. Smal l mammal s: their producti vi ty an d popula tion dy namics. Cambridge , MA: Cambridge Univ ersi ty Press; pp. 173-204.
Hunt , G. L., Jr . (1972) Influence of food dis tr ibut ion a nd hum an dis turbance on t he
4-28

reproducti ve su ccess o f herring gulls . Ecology 53: 1051-1061.
Jorgensen, S. E.; Nielsen, S. N.; Jorgensen, L. A. (1991) Handbook of ecologic al parameter s and ecotoxicology . Amster dam, The Netherlands: Elsevi er Science Publis hers.
Karasov, W. H. (1990) Digestion in bir ds: chemical and phy siologic al determinants an d ecologic al impli cations . Studie s in A vian Biology 13: 391-415.
Kendeigh, S. C.; West, G. C. (1965) Caloric val ues of plant seed s eaten by birds. Ecology 46: 553-555.
Koplin, J. R.; Collopy , M. W.; Bamman n, A. R.; et al . (1980) Energetics o f two winter ing raptors. Auk 97: 795-806.
Kramer , D. C. (1973) Geopha gy in Terrepene ornata ornata Agassi z. J. Herpetol. 7: 138-139.
Kreulen, D. A.; Jager, T. (1984) The signif icance of soil ing estion in t he ut iliza tion of ar id rangelands by large herbivores, wi th special reference to natural licks o n the Kalahari pans. In: Internationa l sympos ium on he rbiv ore nut rition in t he subt ropic s and tropics (1983: Pretoria, South Afr ica). Draigna ll, Sout h Afr ica: Science Press; pp. 204-221.
Levey, D. J.; Karasov, W. H. (1989) Digesti ve responses o f temp erate birds swi tch ed to fruit or insect d iets. Auk 106: 675-686.
Litvai tis, J. A.; Mautz, W. W. (1976) Energy utiliza tion of th ree diets fed to a captive red fox. J. Wildl. Manage. 40: 365-368.
Mayland, H. F.; Shewmaker , G. E.; Bull, R. C. (1977) Soil ing estion by cattl e grazing c rested wheatgrass. J. Range Manage. 30: 264-265.
Mille r, L. K. (1978) Energetics o f th e northern fur seal in relation t o cli mate an d food resources o f th e Bering Sea. Nat. Tech. Inf. Serv. P. B. 275-96.
Mitch ell, J. L. (1961) Mink mo vemen ts an d popula tions on a Mont ana river . J. Wildl. Manage. 25: 48-54.
Minnic h, J. E. (1982) The use of water . In: Gans, C.; Pough, F. H., eds. Biology of th e rept ilia , phy siology C; phy siologic al ecology : v. 12. New York, NY: Academic Press; pp. 325-395.
Nagy, K. A. (1987) Field metabolic r ate and food r equir ement sca ling in mammal s and bir ds. Ecol. Monogr . 57: 111-128.
Odum, E. P.; Marshall, S. G.; Marples, T. G. (1965) The caloric co ntent of mi grating bir ds. Ecology 46: 901-904.
4-29

Peters, R. H. (1983) The ecologic al impli cations of body size . Cambridge , Engla nd: Cambridge Univ ersi ty Press.
Pierotti , R.; Annett, C. (1987) Reproducti ve co nsequences o f dietary sp ecialization a nd swi tching in a n ecologic al generalis t. In: Kamil, A. C.; Krebs, J. ; H. R. Pullia m, eds. Foraging be havior . New York, NY: Plenum Pr ess; pp. 417-442.
Reeder, W. G. (1951) Stomach analysi s of a group of shor ebir ds. Condor 53: 43-45.
Rickl efs, R. E. (1974) Energetics o f reproduc tion in bir ds. In: Paynter , R. A., ed. Avian energetics. Cambridge , MA: Nuttall O rnit hologic al Club.
Robel, R. J.; Bisset, A. R.; Dayto n, A. D.; et al . (1979) Comparative energetics o f bobw hit es on six differ ent foods . J. Wildl. Manage. 43: 987-992.
Sokal , O. M. (1971) Lithopha gy and ge opha gy in r ept iles. J. Herpetol. 5: 69-71.
Stalmaster , M. V.; Gessaman , J. A. (1982) Food c ons umpt ion a nd energy r equir ements o f captive ba ld eagles. J. Wildl. Manage. 46: 646-654.
Thayer , G. W.; Schaaf, W. E.; Angelovic, J. W.; et al . (1973) Caloric measu rements o f so me estu arine organisms. Fishery Bull. 71: 289-296.
Tyler, A. V. (1973) Caloric val ues of so me North Atlantic inver tebrates. Mar. Biol. 19: 258-261.
U. S. Environmental Protect ion A gency. (1989) Risk assessmen t guida nce for Supe rfund: volum e I - hum an health evalua tion m anual, int erim fina l. Washingt on, DC: Offi ce of Solid W aste, Offi ce of Emer gency an d Remedial Respons e; EPA repor t no. EPA/540/1-89/002.
U. S. Environmental Protect ion A gency. (1990) Inter im met hods fo r development of inha lation r eference co ncentrations , review dr aft. Washingt on, DC: Offi ce of Research and Development; EPA report no. EPA/600/8-90/066A.
U. S. Environmental Protect ion A gency. (1992a) Framewo rk fo r ecologic al risk assessmen t. Washingt on, DC: Risk Assessmen t Forum; EPA report no. EPA/630/R-92/001.
U. S. Environmental Protect ion A gency. (1992b) Guide line s for expos ure assessmen t. Washingt on, DC: Science Advisory Boa rd; EPA repor t no. EPA/600/Z-92/001.
U. S. Environmental Protect ion A gency. (1992c) Dermal exposure assessmen t: principle s and appli cations , int erim r epor t. Washingt on, DC: Offi ce of Research and Development; EPA report no. EPA/600/8-91/001B.
Vogtsb erger, L. M.; Barrett, G. W. (1973) Bioe nergetics of cap tive red foxes. J. Wildl. Manage. 37: 495-500.
4-30