werner syndrome helicase has a critical role in dna damage...
TRANSCRIPT

Therapeutics, Targets, and Chemical Biology
Werner Syndrome Helicase Has a Critical Role in DNADamage Responses in the Absence of a FunctionalFanconi Anemia Pathway
Monika Aggarwal1, Taraswi Banerjee1, Joshua A. Sommers1, Chiara Iannascoli3, Pietro Pichierri3,Robert H. Shoemaker2, and Robert M. Brosh, Jr.1
AbstractWerner syndrome is genetically linked to mutations inWRN that encodes a DNA helicase-nuclease believed to
operate at stalled replication forks. Using a newly identified small-molecule inhibitor of WRN helicase (NSC617145), we investigated the role ofWRN in the interstrand cross-link (ICL) response in cells derived frompatientswith Fanconi anemia, a hereditary disorder characterized by bone marrow failure and cancer. In FA-D2�/�
cells, NSC 617145 acted synergistically with very low concentrations of mitomycin C to inhibit proliferation ina WRN-dependent manner and induce double-strand breaks (DSB) and chromosomal abnormalities. Underthese conditions, ataxia–telangiectasia mutated activation and accumulation of DNA-dependent protein kinase,catalytic subunit pS2056 foci suggested an increased number of DSBs processed by nonhomologous end-joining(NHEJ). Rad51 foci were also elevated in FA-D2�/� cells exposed to NSC 617145 andmitomycin C, suggesting thatWRN helicase inhibition interferes with later steps of homologous recombination at ICL-induced DSBs. Thus,when the Fanconi anemia pathway is defective, WRN helicase inhibition perturbs the normal ICL response,leading to NHEJ activation. Potential implication for treatment of Fanconi anemia–deficient tumors by theirsensitization to DNA cross-linking agents is discussed. Cancer Res; 73(17); 5497–507. �2013 AACR.
IntroductionWerner syndrome, characterized by premature aging and
cancer predisposition (1), is caused by autosomal recessivemutations inWRN (2), encoding a RecQDNAhelicase-nucleaseimplicated in genomic stability (3, 4). WRN helicase canunwind key homologous recombination DNA intermediatesand interacts with proteins implicated in DNA replication,recombinational repair, and telomere maintenance (5). Wer-ner syndrome cells are sensitive to certain DNA-damagingagents, and display defects in resolution of recombinationintermediates and a prolonged S-phase (6). WRN is requiredfor normal replication fork progression after DNA damage orfork arrest (7).
Werner syndrome cells are sensitive to interstrand cross-link (ICL)–inducing agents (8) and undergo apoptosis in S-phase (9). Cellular studies suggest WRN helicase, in conjunc-tion with BRCA1, is required to process DNA ICLs (10). WRN isrequired for ataxia–telangiectasia mutated (ATM) activationand the intra-S-phase checkpoint in response to ICL-induceddouble-strand breaks (DSB; ref. 11), suggesting WRN and ATMcollaborate in response to collapsed forks at ICL-inducedDSBs.Biochemical studies using a reconstituted system with a DNAsubstrate harboring a psoralen ICL suggested that WRN heli-case is required for ICL processing (12).
Patients with Fanconi anemia suffer from progressive bonemarrow failure and cancer predisposition, and their cells (suchasWerner syndrome cells) are sensitive to ICL agents, only to agreater extent (13). In fact, a chromosome breakage test usingFanconi anemia patient cells exposed to an ICL-inducing agentis used as a primary diagnostic tool for Fanconi anemia (14).The Fanconi anemia pathway is involved in initial recognitionand unhooking of an ICL and subsequent repair of the ICL-induced DSB in conjunction with homologous recombinationor translesion synthesis pathways, but the detailed molecularmechanism is an active area of investigation (15). One functionof the Fanconi anemia pathway is to channel DSBs throughthe homologous recombination pathway, thereby preventinginappropriate engagement of breaks by error-prone nonho-mologous end-joining (NHEJ; refs. 16, 17). However, the rela-tionship between DNA repair pathways is complex, as evi-denced by a recentmouse study showing that deletion of 53BP1or Ku exacerbates genomic instability in FANCD2-deficientcells (18).
Authors' Affiliations: 1Laboratory of Molecular Gerontology, NationalInstitute on Aging, NIH Biomedical Research Center, NIH, Baltimore;2Screening Technologies Branch, Developmental Therapeutics Program,Division of Cancer Treatment and Diagnosis, National Cancer Institute atFrederick, NIH, Frederick, Maryland; and 3Section of Experimental andComputational Carcinogenesis, Istituto Superiore di Sanit�a, Rome, Italy
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Current address for M. Aggarwal: Department of Oncology, LombardiComprehensive Cancer Center, Georgetown University Medical Center,GeorgetownUniversity, 3800Reservoir Road,NW,Washington,DC20007.
Corresponding Author: Robert M. Brosh Jr, National Institute on Aging,NIH, 251 Bayview Blvd, Baltimore, MD 21224. Phone: 410-558-8578; Fax:410-558-8162; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-12-2975
�2013 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 5497
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

To understand the role of WRN in the ICL response, weexamined effects of a small-molecule WRN helicase inhibitorin a Fanconi anemia–mutant genetic background. A morepotent structural analog of the previously identified parentcompound NSC 19630 (19) was discovered that is blockedfrom potential thiol reactivity but retains the ability to mod-ulate WRN function in vivo. The WRN helicase inhibitor NSC617145 acted synergistically with a very limited concentrationof mitomycin C to induce DSB accumulation and chromosom-al abnormalities, and activate the DNA damage response inFanconi anemia–mutant cells. NSC 617145 exposure resultedin enhanced accumulation of DNA-dependent protein kinase,catalytic subunit (DNA-PKcs) pS2056 foci and Rad51 foci inmitomycin C–treated Fanconi anemia–deficient cells, suggest-ing thatWRNhelicase inhibition prevents processing of Rad51-mediated recombination products and activates NHEJ. WRNhelicase may be a suitable target for chemotherapy strategiesin Fanconi anemia–deficient tumors.
Materials and MethodsCell lines and culture
HeLa (CCL-2), U2OS (HTB96), and HCT116 (CCL-247) celllines were obtained from American Type Culture Collectionwhere they were tested and authenticated. HCT116 p53 doubleknockout cells were obtained from the laboratory of Dr. B.Vogelstein (Johns Hopkins University, Baltimore, MD) wherethey were tested and authenticated (20). All these cell lineswere grown in Dulbecco's Modified EagleMedium supplemen-ted with 10% FBS, 1% penicillin–streptomycin, and 1% L-glutamine at 37�C in 5% CO2. PSNF5 [Bloom syndrome(BLM)–corrected] and PSNG13 (BLM-mutant) are isogeniccell lines that were obtained from the laboratory of Dr. I.Hickson (University of Copenhagen, Copenhagen, Denmark)where they were tested and authenticated (21, 22). PSNF5 is astable BLM cell line expressing a FLAG epitope-tagged, wild-type BLM protein from the cytomegalovirus (CMV) promoterin a pcDNA3-based construct. PSNG13 is a control BLM celltransfectant containing the pcDNA3 vector only. The BLM-mutant and -corrected cell lines were grown in the samemediaas HeLa cells but supplemented with 350 mg/mL G418. Thesimian virus 40–transformed Fanconi anemia–mutant fibro-blasts, PD20 (FA-D2), GM6914 (FA-A), and their respectivecorrected counterparts were provided by Fanconi Anemia CellRepository at Oregon Health & Science University (Portland,OR) where they were tested and authenticated. Fanconi ane-mia–mutant and -corrected cells were grown in the samemedium as HeLa cells but supplemented with 0.2 mg/mLpuromycin.
Cell proliferation assaysProliferation was measured using WST-1 assay (Roche) as
described previously (19).
siRNA transfection and Western blot analysisWRN siRNA (WH: 50-UAGAGGGAAACUUGGCAAAUU-30
and/or CG: 50-GUGUAUAGUUACGAUGCUAGUGAUU-30) andcontrol siRNA (19) were transfected using Lipofectamine 2000as per the manufacturer's protocol (Invitrogen). Cells were
plated to 50% to 60% confluence in 10-cm dishes 24 hoursbefore transfection. siRNA (0.6 nmol) was mixed with 30 mL ofLipofectamine 2000 in 3 mL of Opti-MEM (Invitrogen). Themixture was added to cells that were subsequently incubatedfor 6 hours. After 24 hours, a second transfection was carriedout similarly. Seventy-two hours after the initial transfection,cells were harvested for preparing lysate or treated with small-molecule compounds or dimethyl sulfoxide (DMSO) at theindicated concentrations and cell proliferation was measuredusing WST-1 reagent (Roche) as described for HeLa cells. ForFanconi anemia–mutant cells, siRNA transfection and treat-ment with NSC 617145 was carried out as described previouslyexcept that OD450 was measured after 48 hours of treatmentwith mitomycin C and/or NSC 617145.
For lysate preparation, cells werewashed twicewith 1� PBS.Radioimmunoprecipitation assay (RIPA) buffer [10 mmol/Lsodium phosphate (pH 7.2), 300 mmol/L NaCl, 0.1% SDS, 1%NP-40, 1% sodium deoxycholate, and 2 mmol/L EDTA] wasadded to the cells and the cells were incubated at 4�C for 30minutes. Cells were scraped and the suspension was furtherincubated on ice for 30 minutes. Cell suspension was centri-fuged at 18,500� g for 10 minutes at 4�C and supernatant wascollected. Twenty microgram of the lysate was loaded on 8% to16% SDS-PAGE. Protein was transferred onto a polyvinylidenedifluoride (PVDF) membrane and blot was probed with anti-WRN mouse monoclonal antibody (1:1,000; Spring ValleyLaboratories). For secondary antibody, peroxidase-conjugatedanti-mouse immunoglobulin G (IgG; 1:1,000; Vector Labora-tories) was used. Blot was developed using ECL Plus WesternBlot Detection Kit as per the manufacturer's protocol (Amer-sham). As a loading control, blot was stripped and thenreprobed with anti-actin antibody (1:5,000; Sigma).
Analysis of metaphase chromosomesCell harvest and metaphase slide preparation was carried
out for metaphase analysis as described previously (23).See Supplementary Data for additional information.
ResultsInhibition of WRN helicase activity by small-moleculeNSC 617145
To identify small molecules that inhibit WRN helicaseactivity in amore potentmanner than the previously identifiedcompound NSC 19630 (19), we selected four close structuralanalogs and three compounds whose pattern of activity in theNCI60 screen matched NSC 19630 for analysis (SupplementaryFig. S1). One of the latter compounds, NSC 617145, is also astructural analog, but blockedwith respect to thiol reactivity inthe five-membered rings by Cl atoms. NSC 617145 inhibitedWRNhelicase activity in a concentration-dependentmanner invitro (Supplementary Fig. S2A), yielding an IC50 value of 230nmol/L. NSC 617145 inhibited WRN ATPase in a dose-depen-dentmanner (Supplementary Fig. S2B). No detectable effect onWRN exonuclease was observed (Supplementary Fig. S2C).
To examine specificity ofWRN helicase inhibition, we tested5 mmol/L NSC 617145 (�20-fold >IC50 value) on DNA unwind-ing catalyzed by various helicases. No significant inhibition ofunwinding was observed with BLM, FANCJ, ChlR1, RecQ, and
Aggarwal et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5498
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

UvrD, and only a very modest (7%) inhibition of RECQ1(Supplementary Fig. S2D) was observed.To determine their potency in vivo, HeLa cells were exposed
to increasing concentrations of selected analogs or DMSO for 0to 3 days. Of the selected analogs, NSC 617145 showedmaximalinhibition of proliferation (98%) at the lowest concentration ofNSC 617145 (1.5 mmol/L; Fig. 1A; Supplementary Fig. S3). Toaddress whether the effect was cytostatic or cytotoxic, HeLacells were exposed to NSC 617145 for 3 days, replenishedwith media lacking NSC 617145, and allowed to recover for0 to 3 days (Fig. 1B). The lack of recovery from NSC 617145suggested a cytotoxic effect. To determine whether the effectof NSC 617145 was WRN-dependent, we compared its effecton HeLa cells depleted of WRN (�90%) with NS-siRNA–transfected cells. WRN-depleted cells grown in the presenceof 1.5 mmol/L NSC 617145 were resistant to its antiproli-ferative effects, whereas NS-siRNA HeLa cells were highlysensitive to NSC 617145 (Fig. 1C). These results were con-firmed by 2 different WRN-siRNA molecules that indepen-dently conferred resistance to NSC 617145 (SupplementaryFig. S4). Western blot analyses showed that WRN depletionby either WRN-siRNA or combination was more than 90%(Supplementary Fig. S4).NSC 617145 may exert its effect through a mechanism in
which theNSC 617145–inhibitedWRNhelicase induces greaterdamage by interfering with a compensatory mechanism(s). Todetermine whether the effect of NSC 617145 is dependent on ahelicase-active version of WRN, HeLa cells were depleted of
endogenous WRN, and siRNA-resistant WRN [wild-type orhelicase inactive K577M (24)] was expressed. The resultingcells were tested for sensitivity to NSC 617145. It was observedthat theWRN-depleted cells rescued with wild-typeWRNweresensitive to the WRN helicase inhibitor compared with emptyvector–transfected cells, supporting the finding that the inhi-bition of cell proliferation by NSC 617145 is WRN-dependent(Supplementary Fig. S5). However, WRN-depleted cells expres-sing the helicase-inactive WRN-K577M behaved similar to theWRN-depleted cells transfected with empty vector, suggestingthat they were resistant to the negative effect of WRN helicaseinhibitor NSC 617145 on cell viability.
Because NSC 617145 inhibited proliferation of p53-inacti-vated HeLa cells, we examined its effect on proliferation ofU2OS cells with wild-type p53. NSC 617145 (1.5 mmol/L)inhibited U2OS proliferation by 80% after day 2 (Supple-mentary Fig. S6A), suggesting that the effect of NSC 617145was not dependent on p53 status. We also tested a pair ofisogenic p53-negative and -positive HCT116 cells for sensi-tivity to NSC 617145 (1.5 mmol/L). Both cell lines weresensitive to the compound; however, p53-deficient cells were2-fold more resistant than p53-proficient cells (Supplemen-tary Fig. S6B). Because NSC 617145 exerted a WRN-depen-dent effect on proliferation, we evaluated whether cellsmutated for the sequence-related BLM helicase were sensi-tive. BLM-null and -corrected cells displayed similar sensi-tivity to NSC 617145 (Supplementary Fig. S6C), suggestingBLM does not play a role.
0
20
40
60
80
100
120
321.510.750
Pro
life
rati
on
(%
)
Day 0
Day 1
Day 2
Day 3
NSC 617145 (μμmol/L)
A B
C
0
20
40
60
80
100
120
3210
Pro
life
rati
on
(%
)
Days
DMSO
1.5 µmol/L NSC 617145
0
20
40
60
80
100
120
3210
Pro
life
rati
on
(%
)
Days
HeLa
WRN siRNA
NS siRNA
Figure 1. NSC 617145 inhibits cell proliferation in a WRN-specific manner. A, HeLa cells were treated with DMSO or indicated NSC 617145 concentration for0 to 3 days. B, HeLa cells were exposed to 1.5 mmol/L NSC 617145 for 3 days, replenished with media lacking NSC 617145, and allowed to recoverfor 0 to 3 days. C, HeLa cells either untransfected (HeLa) or transfectedwith nonspecific siRNA (NS siRNA) orWRN-specific siRNA (WRN siRNA) were treatedwith DMSOor 1.5mmol/L NSC617145 for indicated number of days. Cell proliferationwas then determinedwithWST-1 reagent. Percentage proliferationwascalculated. Experiments were repeated three times, and error bars indicate SD. This applies to all figures.
Role of WRN Helicase in Fanconi Anemia–Deficient Cells
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5499
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

Cellular exposure to NSC 617145 causes accumulation ofDSBs, blocked replication forks, and apoptosis
Because WRN-deficient cells are defective in resolution ofrecombination intermediates (6), NSC 617145 inhibition ofWRN helicase activity might result in DSBs at blocked repli-cation forks. Therefore, we analyzed the effect of NSC 617145on g-H2AX foci, a marker of DSBs. Exposure of HeLa cells to0.25 mmol/L NSC 617145 elevated g-H2AX foci approximately18-fold compared with DMSO-treated cells (SupplementaryFig. S7A and S7B). WRN-siRNA–treated HeLa cells showed asimilar low number of g-H2AX foci in NSC 617145- and DMSO-treated cells, suggesting that inhibition of WRN activity in vivoby NSC 617145 led to DSB accumulation in a WRN-dependentmanner. H2AX phosphorylation can be induced by a widerange of phenomenon including DSBs; therefore, we examinedthe effect of NSC 617145 on 53BP1 foci, an independentmarkerof DNA damage (25). Cellular exposure to 0.25 mmol/L NSC617145 elevated 53BP1 foci 3.5-fold compared with DMSO-treated cells, confirming NSC 617145 induced DNA damage(Supplementary Fig. S7C and S7D).
HeLa cells exposed to 1.5 mmol/L NSC 617145 showed a 4-fold increase in apoptosis as compared with DMSO-treatedcells (Supplementary Fig. S8). WRN-depleted HeLa cellsshowed no difference in level of apoptosis in NSC 617145-and DMSO-treated cells, showing WRN-dependent induc-tion of apoptosis by NSC 617145. Failure to repair DNAdamage would affect replication fork progression; therefore,we determined the effect of NSC 617145 on proliferating cellnuclear antigen (PCNA) foci formation. HeLa cells exposedto 0.25 mmol/L NSC 617145 showed an elevated number ofPCNA foci (�22-fold) compared with DMSO-treated cells(Supplementary Fig. S9A and S9B). In contrast, WRN-deplet-ed cells showed similar levels of PCNA staining for NSC617145- or DMSO-treated cells, suggesting WRN-dependentaccumulation of stalled replication foci. Consistent with thePCNA induction, exposure of HeLa cells to NSC 617145(1 mmol/L) resulted in an increased population in S-phasecompared with DMSO-treated cells (Supplementary Fig.S10). We also observed from fluorescence-activated cellsorting (FACS) analysis a sub-G1 fraction of NSC 617145–treated HeLa cells (data not shown), consistent with theinduction of apoptosis by the WRN helicase inhibitor. WRN-depleted cells exposed to 1 mmol/L NSC 617145 showed asimilar percentage of cells in S-phase compared with DMSO-treated cells, suggesting that NSC 617145 induced prolongedS-phase in a WRN-dependent manner.
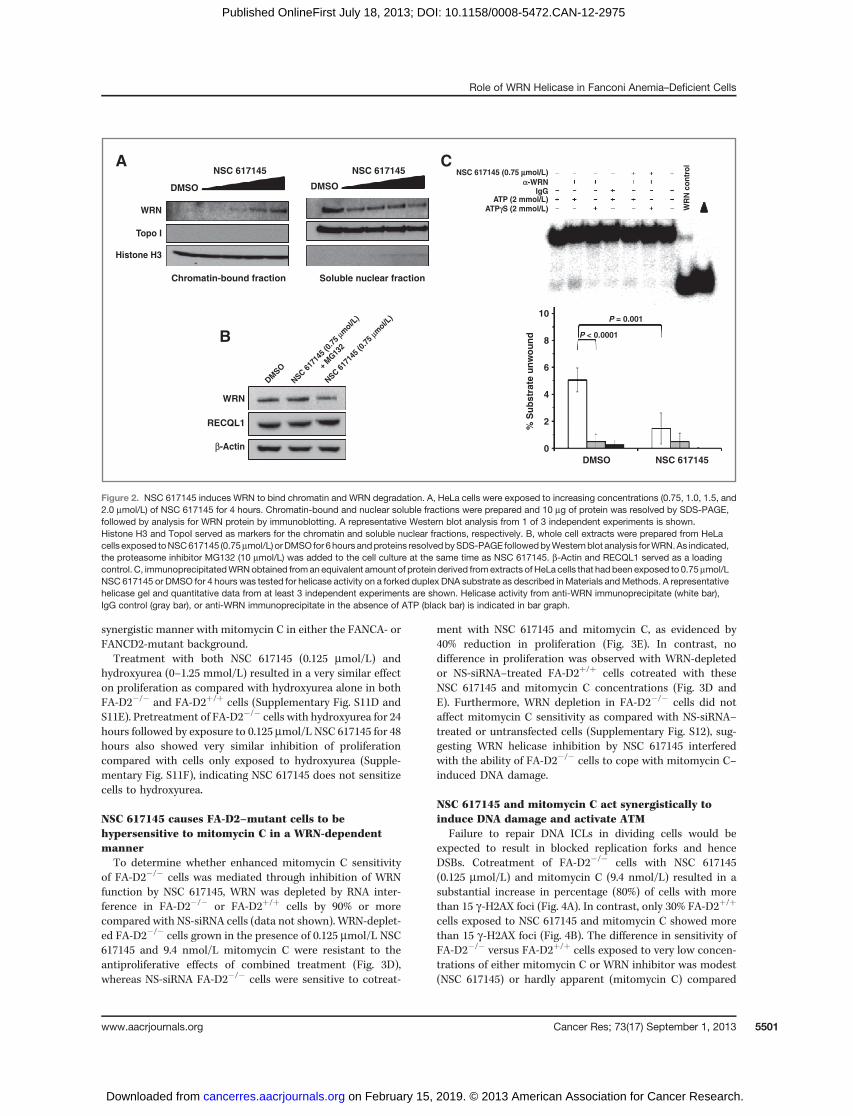
NSC 617145 induces WRN binding to chromatin andproteasomal degradation
We reasoned that NSC 617145might target cellularWRN forhelicase inhibition and induce toxic DNA lesions mediated byWRN's interaction with genomic DNA, prompting us to askwhether poisoned WRN became enriched in the chromatinfraction. Western blot analysis of nuclear soluble and chro-matin-bound fractions prepared from HeLa cells exposed toNSC 617145 showed a dose-dependent increase inWRN boundto chromatin (Fig. 2A). We also observed that the amount ofWRN from extract of HeLa cells exposed to NSC 617145 (0.75
mmol/L) for 6 hours was reduced compared with DMSO-treated cells (Fig. 2B). Inclusion of proteasome inhibitorMG132 restored WRN level to that of DMSO-treated cells (Fig.2B). Thus, NSC 617145 causes WRN to become degraded by aproteasome-mediated pathway. Consistent with reduction ofWRN protein in NSC 617145–treated cells, helicase activitycatalyzed by immunoprecipitated WRN from an equivalentamount of total extract protein from HeLa cells treated withNSC 617145 (0.75 mmol/L; 4 hours) was reduced 4-fold com-paredwith helicase activity by immunoprecipitatedWRN fromDMSO-treated cells (Fig. 2C).
Synergistic effect of NSC 617145 and DNA cross-linkingagent mitomycin C on cell proliferation
We sought to use the newly identified WRN helicase inhib-itor as a tool to explore whether WRN helicase activity helpscells cope with stress imposed by the DNA cross-linking agentmitomycin C. Treatment with either NSC 617145 (0.5 mmol/L)or mitomycin C (9.4 nmol/L) exerted no significant effect onproliferation. However, cotreatment with both NSC 617145 (0.5mmol/L) and mitomycin C (9.4 nmol/L) resulted in a 45%reduction in proliferation (Fig. 3A). In contrast, treatment withNSC 617145 (0.5 mmol/L) and hydroxyurea (0–5mmol/L) had asimilar effect on proliferation compared with hydroxyureaalone (Supplementary Fig. S11A), suggesting that the apparentsynergism between NSC 617145 and mitomycin C is not ageneral effect imposed by other forms of replicative stress suchas hydroxyurea that depletes the nucleotide pool.
NSC 617145–treated Fanconi anemia–mutant cells arehypersensitive to mitomycin C
On the basis of the synergistic effect of NSC 617145 andmitomycin C, we hypothesized that WRN might participatein a pathway that is partially redundant with Fanconianemia–mediated ICL repair. For this, we examined sensi-tivity of FA-D2–mutant and -corrected cells to mitomycin Cand NSC 617145. Dose–response curves for mitomycin C andNSC 617145 sensitivity of FA-D2 cells indicated drug con-centrations at which only modest effects on proliferationwere observed (Supplementary Fig. S11B and S11C). NSC617145 (0.125 mmol/L) or mitomycin C (9.4 nmol/L) exertedonly a mild effect on cell proliferation; however, a significant45% reduction was observed for cells treated with both NSC617145 and mitomycin C (Fig. 3B). In contrast, FA-D2þ/þ
cells showed only a 10% reduction of proliferation by com-bined treatment of NSC 617145 and mitomycin C (Fig. 3B).Sensitization of FA-D2�/� cells to mitomycin C by WRNhelicase inhibition is distinct from the effect imposed by adeficiency or inhibition of proteins in the NHEJ pathway(LIG-4 and Ku-70), which suppress ICL-sensitivity of Fanconianemia–deficient cells (16, 17).
Combinatorial treatment of FA-A�/� cells with NSC 617145(0.125 mmol/L) and mitomycin C (9.4 nmol/L) resulted in 60%reduction in proliferation compared with very modest effectsexerted by either agent alone (Fig. 3C).Moreover, the effect wasdependent on FANCA status as only 10% reduction in prolif-eration was observed for FA-Aþ/þ cells exposed to bothmitomycin C and NSC 617145. Thus, NSC 617145 acts in a
Aggarwal et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5500
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975
synergistic manner with mitomycin C in either the FANCA- orFANCD2-mutant background.Treatment with both NSC 617145 (0.125 mmol/L) and
hydroxyurea (0–1.25 mmol/L) resulted in a very similar effecton proliferation as compared with hydroxyurea alone in bothFA-D2�/� and FA-D2þ/þ cells (Supplementary Fig. S11D andS11E). Pretreatment of FA-D2�/� cells with hydroxyurea for 24hours followed by exposure to 0.125 mmol/L NSC 617145 for 48hours also showed very similar inhibition of proliferationcompared with cells only exposed to hydroxyurea (Supple-mentary Fig. S11F), indicating NSC 617145 does not sensitizecells to hydroxyurea.
NSC 617145 causes FA-D2–mutant cells to behypersensitive to mitomycin C in a WRN-dependentmannerTo determine whether enhanced mitomycin C sensitivity
of FA-D2�/� cells was mediated through inhibition of WRNfunction by NSC 617145, WRN was depleted by RNA inter-ference in FA-D2�/� or FA-D2þ/þ cells by 90% or morecompared with NS-siRNA cells (data not shown). WRN-deplet-ed FA-D2�/� cells grown in the presence of 0.125 mmol/L NSC617145 and 9.4 nmol/L mitomycin C were resistant to theantiproliferative effects of combined treatment (Fig. 3D),whereas NS-siRNA FA-D2�/� cells were sensitive to cotreat-
ment with NSC 617145 and mitomycin C, as evidenced by40% reduction in proliferation (Fig. 3E). In contrast, nodifference in proliferation was observed with WRN-depletedor NS-siRNA–treated FA-D2þ/þ cells cotreated with theseNSC 617145 and mitomycin C concentrations (Fig. 3D andE). Furthermore, WRN depletion in FA-D2�/� cells did notaffect mitomycin C sensitivity as compared with NS-siRNA–treated or untransfected cells (Supplementary Fig. S12), sug-gesting WRN helicase inhibition by NSC 617145 interferedwith the ability of FA-D2�/� cells to cope with mitomycin C–induced DNA damage.
NSC 617145 and mitomycin C act synergistically toinduce DNA damage and activate ATM
Failure to repair DNA ICLs in dividing cells would beexpected to result in blocked replication forks and henceDSBs. Cotreatment of FA-D2�/� cells with NSC 617145(0.125 mmol/L) and mitomycin C (9.4 nmol/L) resulted in asubstantial increase in percentage (80%) of cells with morethan 15 g-H2AX foci (Fig. 4A). In contrast, only 30% FA-D2þ/þ
cells exposed to NSC 617145 and mitomycin C showed morethan 15 g-H2AX foci (Fig. 4B). The difference in sensitivity ofFA-D2�/� versus FA-D2þ/þ cells exposed to very low concen-trations of either mitomycin C or WRN inhibitor was modest(NSC 617145) or hardly apparent (mitomycin C) compared
NSC 617145A
B
C
DMSO
WRN
WRN
% S
ub
str
ate
un
wo
un
d
10
8
6
4
2
0
RECQL1
Topo I
Histone H3
Chromatin-bound fraction Soluble nuclear fraction
DMSO
DMSO
DM
SO
NSC 617145
NSC 617145
P = 0.001
P < 0.0001
NSC 617145 (0.75 μμmol/L)
NSC
617
145
(0.7
5 μm
ol/L)
+ M
G13
2
NSC
617
145
(0.7
5 μm
ol/L)
α-WRN
β-Actin
WR
N c
on
tro
l
ATP (2 mmol/L)ATPγS (2 mmol/L)
IgG
Figure 2. NSC 617145 induces WRN to bind chromatin and WRN degradation. A, HeLa cells were exposed to increasing concentrations (0.75, 1.0, 1.5, and2.0 mmol/L) of NSC 617145 for 4 hours. Chromatin-bound and nuclear soluble fractions were prepared and 10 mg of protein was resolved by SDS-PAGE,followed by analysis for WRN protein by immunoblotting. A representative Western blot analysis from 1 of 3 independent experiments is shown.Histone H3 and TopoI served as markers for the chromatin and soluble nuclear fractions, respectively. B, whole cell extracts were prepared from HeLacells exposed toNSC617145 (0.75mmol/L) orDMSO for 6hours andproteins resolvedbySDS-PAGE followedbyWesternblot analysis forWRN.As indicated,the proteasome inhibitor MG132 (10 mmol/L) was added to the cell culture at the same time as NSC 617145. b-Actin and RECQL1 served as a loadingcontrol. C, immunoprecipitatedWRNobtained froman equivalent amount of protein derived fromextracts of HeLa cells that had been exposed to 0.75mmol/LNSC 617145 or DMSO for 4 hours was tested for helicase activity on a forked duplex DNA substrate as described inMaterials andMethods. A representativehelicase gel and quantitative data from at least 3 independent experiments are shown. Helicase activity from anti-WRN immunoprecipitate (white bar),IgG control (gray bar), or anti-WRN immunoprecipitate in the absence of ATP (black bar) is indicated in bar graph.
Role of WRN Helicase in Fanconi Anemia–Deficient Cells
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5501
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

with the cotreatment. Thus, accumulation of mitomycin C–induced DNA damage in FA-D2�/� cells is increased by NSC617145.
Exposure of FA-D2�/� cells with a dose range (0.125–1.5mmol/L) of NSC 617145 led to activation of ATM as detectedby pATM-Ser1981 at 0.5 mmol/L NSC 617145 (Supplemen-tary Fig. S13); however, no significant activation of ATMwas observed at lower doses. In FA-D2þ/þ cells, no appreci-able ATM activation was detected except at the highest NSC617145 concentration tested, 1.5 mmol/L. Upon coexposureof FA-D2�/� cells with a very low dose of NSC 617145 (0.125mmol/L) and mitomycin C (9.4 nmol/L), there was a sig-nificant accumulation of pATM-Ser1981, whereas ATMactivation was not detected in FA-D2þ/þ cells (Fig. 5A).FA-D2�/� cells exposed to very low dose of NSC 617145(0.125 mmol/L) and mitomycin C (9.4 nmol/L) retained highlevels of pATM-Ser1981 even after 24-hour exposure (datanot shown), suggesting a significant delay in damage repairwhen WRN helicase was inhibited. Thus, NSC 617145 andmitomycin C act synergistically to induce DNA damage andactivate ATM.
NSC 617145 elevated mitomycin C–inducedchromosomal instability and induced accumulation ofDNA-PKcs pS2056 foci in FA-D2–mutant cells
Recent studies suggest the Fanconi anemia pathway plays animportant role in preventing aberrant DNA repair (16, 17).Because NHEJ factors have high affinity for DNA ends (26), the
accumulated DSBs in FA-D2�/� cells, when WRN helicaseactivity is pharmacologically inhibited, might promote geno-mic instability when captured by error-prone pathways. Toaddress this, we examined metaphase spreads from FA-D2�/�
and FA-D2þ/þ cells for chromosomal aberrations (Fig. 5B; ref.27). FA-D2�/� cells exposed to NSC 617145 (0.125 mmol/L) andmitomycin C (9.4 nmol/L) showed a 4-fold increase in abnor-mal chromosome structures compared with either agent alone(Fig. 5C). Enhanced chromosomal instability was dependenton FANCD2 status as evidenced by the relatively low level ofchromosomal breaks in FA-D2þ/þ cells exposed to NSC 617145and/or mitomycin C.
DNA-dependent protein kinase complex is required forNHEJ in conjunction with Ku70/80 and XRCC4/Ligase IV;moreover, DNA-PKcs pS2056 is detected at DSBs (28). Tosubstantiate the role of NHEJ in DSB processing when WRNhelicase is inhibited, FA-D2�/� cells were examined for DNA-PKcs pS2056 foci formation. Cotreatment of FA-D2�/� cellswith limited concentrations of NSC 617145 (0.125 mmol/L) andmitomycin C (9.4 nmol/L) resulted in 70% of cells with morethan 15 DNA-PKcs pS2056 foci (Fig. 6A and B). In contrast, only15%of FA-D2þ/þ cells exposed toNSC617145 andmitomycinCshowed more than 15 DNA-PKcs pS2056 foci (Fig. 6A and B).The difference in sensitivity between FA-D2�/� and FA-D2þ/þ
cells exposed to very low concentrations of either mitomycin Cor NSC 617145 was modest (mitomycin C) or hardly apparent(NSC 617145) compared with cotreatment, suggesting thatwhen WRN helicase activity is inhibited, FA-D2�/� cells show
0
20
40
60
80
100
120
0 3
Pro
life
rati
on
(%
)
Days
FA-D2+/+ FA-D2–/– FA-A+/+ FA-A–/–
FA-D2+/+
NS siRNA
FA-D2–/–
NS siRNA
ED
FA-D2+/+
WRN siRNA
FA-D2–/–
WRN siRNA
MMC
NSC 617145
MMC + NSC 617145
0
20
40
60
80
100
120
Pro
life
rati
on
(%
)
MMC
NSC 617145
MMC + NSC 617145
0
20
40
60
80
100
120
140
Pro
life
rati
on
(%
)
MMC
NSC 617145
MMC + NSC 617145
0
20
40
60
80
100
120
140
Pro
life
rati
on
(%
)
MMC
NSC 617145
MMC + NSC 617145
0
20
40
60
80
100
120
140
Pro
life
rati
on
(%
)
MMC
NSC 617145
MMC + NSC 617145
CBA
Figure 3. NSC617145 exposure enhances sensitivity of Fanconi anemia–mutant cells tomitomycin C in aWRN-dependentmanner. A, HeLa cells were treatedwith NSC 617145 (0.5 mmol/L), mitomycin C (9.4 nmol/L), or both compounds for 3 days. B, FA-D2�/� and FA-D2þ/þ cells were treated with NSC 617145(0.125 mmol/L), mitomycin C (9.4 nmol/L), or both compounds for 2 days. C, FA-A�/� and FA-Aþ/þ cells were treated with NSC 617145 (0.75 mmol/L),mitomycin C (9.4 nmol/L), or both compounds for 2 days. D, FA-D2�/� and FA-D2þ/þ cell lines transfected with WRN siRNA were treated with NSC 617145(0.125 mmol/L), mitomycin C (9.4 nmol/L), or both compounds for 2 days. E, FA-D2�/� and FA-D2þ/þ cell lines transfected with NS siRNA were treated withNSC 617145 (0.125 mmol/L), mitomycin C (9.4 nmol/L), or both compounds for 2 days. Cell proliferation was determined. Percentage proliferation wascalculated.
Aggarwal et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5502
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

increased chromosomal instability due to more DSBs repairedby the error-prone pathway. Strikingly, FA-D2þ/þ cells cot-reated with mitomycin C (9.4 nmol/L) and NSC 617145 (0.125mmol/L) showed similar levels of DNA-PKcs pS2056 foci asindividually treated or DMSO control cells (Fig. 6A and B),suggesting an important role forWRNhelicase in processing ofmitomycin C–induced cross-links converted to DSBs when theFanconi anemia pathway is defective.
NSC 617145 exposure results in accumulation of Rad51foci in FA-D2–mutant cellsWRN plays a physiologic role in resolution of Rad51-depen-
dent homologous recombination products (6). Expression ofdominant-negative Rad51 or bacterial resolvase proteinRusA could suppress homologous recombination or lead tothe generation of recombinants, respectively, resulting inimproved survival of WRN�/� cells. Because Rad51 foci areformed at sites of ICL lesions independently of FA-D2 status(29), we reasoned that WRN helicase inhibition in mitomycinC–treated FA-D2�/� cells might result in accumulation ofhomologous recombination intermediates. Therefore, weinvestigated the effect of NSC 617145 and mitomycin C treat-ment on Rad51 foci formation in Fanconi anemia–mutant and
-corrected cells. We did not observe any significant differencein percentage of cells that displayed Rad51 foci betweenFA-D2�/� and FA-D2þ/þ upon cellular exposure with mito-mycin C (9.4 nmol/L), as reported previously at higher doses ofmitomycin C (29) or NSC 617145 (0.125 mmol/L; Fig. 7A and B).In both cell lines, approximately 50% showed Rad51 foci,similar to that observed for DMSO-treated cells. Cotreatmentof FA-D2�/� cells with NSC 617145 and mitomycin C resultedin 90% of cells staining positive for Rad51 foci. In contrast,FA-D2þ/þ cells exposed to both agents showed a percentage ofcells with Rad51 foci very similar toDMSO-treated cells (Fig. 7Aand B). Collectively, these results suggest that mitomycinC–induced DNA cross-links in FA-D2�/� cells exposed toWRNhelicase inhibitor are converted to DSBs and the homologousrecombination pathway is activated but homologous recom-bination intermediates fail to be subsequently resolved.
DiscussionIn this study, we discovered a new inhibitor ofWRN helicase
activity (NSC 617145) that negatively affects cell proliferationand induces DNA damage in a WRN-dependent manner.WRN depletion negates biologic activity of NSC 617145,suggesting targeted WRN helicase inhibition interferes with
A
B
DMSO
DMSO
MMC
MMC
DAPI Merge
% C
ell
s w
ith
γγ-H
2A
X f
oc
i 100
80
60
40
20
0
NSC 617145
NSC 617145
NSC 617145
+
MMC
NSC 617145
+
MMC
DMSO MMC NSC 617145 NSC 617145
+
MMC
FA-D2+/+ FA-D2–/–
FA-D2+/+ FA-D2–/–
≤ 15 γ-H2AX foci
> 15 γ-H2AX foci
γ-H2AX DAPI Mergeγ-H2AX
Figure 4. Increased accumulationof g-H2AX foci in FA-D2–mutantcells cotreated with NSC 617145andmitomycin C. A, FA-D2�/� andFA-D2þ/þ cells were treated withNSC 617145 (0.125 mmol/L),mitomycin C (9.4 nmol/L), or bothcompounds for 2 days. Cells werestained with anti-g-H2AX antibodyor 40,6-diamidino-2-phenylindole(DAPI) and imaged. B, percentageof cells with g-H2AX foci (�15 or>15, as indicated).
Role of WRN Helicase in Fanconi Anemia–Deficient Cells
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5503
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

normal cellular DNA metabolism. The synergistic effect ofNSC 617145 and mitomycin C was not observed for hydroxy-urea, suggesting that impairment of WRN helicase activityexacerbates the effect of mitomycin C–induced DNA damageduring replication but not an effect exerted by an agent(hydroxyurea) that primarily induces replication stress. Weused the WRN helicase inhibitor to sensitize a Fanconianemia–mutant because mounting evidence points towarda pivotal role of the Fanconi anemia pathway to coordinate arobust ICL response when WRN is also likely to act. WRNhelicase inhibition by NSC 617145 makes FA-A or FA-D2 cellshighly sensitive to very low mitomycin C concentrations thatwould otherwise be only marginally active in normal cells.Furthermore, the effect of combined mitomycin C/NSC617145 treatment is a consequence of a Fanconi anemiapathway deficiency rather than a defect in a specific Fanconianemia gene.
The synergistic effect of NSC 617145 andmitomycin C in theFA-D2–mutant was apparent by robust g-H2AX staining, ATMactivation, and increased chromosomal abnormalities. More-over, accumulation of DNA-PKcs pS2056 foci suggests thatFA-D2 cells attempted to deal with mitomycin C–inducedDSBs by error-prone NHEJ when WRN helicase was pharma-cologically inhibited. This finding builds on previous evidencethat the Fanconi anemia pathway plays an important role inpreventing aberrant NHEJ-mediated repair (16, 17), and impli-cates a role of WRN helicase in repair pathway choice. Inaddition to its helicase-dependent role in recombinationalrepair, WRN may have a structural role to enable repair ofmitomycin C–induced DNA damage. This would be akin to theobservation that a WRN helicase/exonuclease double-mutantcomplemented the NHEJ defect ofWRN�/� cells (30). Althoughexperimental data suggest that WRN participates in an alter-native NHEJ repair pathway of DSBs induced by increased
0
1
2
3
4
5
MMCDMSO NSC 617145 + MMC
FA-D2+/+
FA-D2–/–
NSC 617145B
NSC 617145
MMC
DMSO
MMC + NSC 617145
FA-D2+/+ FA-D2–/–
Bre
ak
s/M
eta
ph
as
e
DM
SO
MM
C
Cm
pd
MM
C +
Cm
pd
FA-D2+/+FA-D2–/–
pATM S1981
ATM
CA
DM
SO
MM
C
Cm
pd
MM
C +
Cm
pd
Figure 5. FA-D2–mutant cells cotreated with NSC 617145 and mitomycin C display ATM activation and increased chromosomal instability. A, FA-D2�/� andFA-D2þ/þ cells were cotreated with 0.125 mmol/L NSC 617145 and 9.4 nmol/L mitomycin C for 3 hours. Cell lysates were prepared and analyzed byimmunoblotting by using anti-pATM Ser1981 antibody. As a control, blot was reprobed with anti-ATM antibody. Arrow indicates phosphorylated ATM. B,FA-D2�/� and FA-D2þ/þ cells were treated with NSC 617145 (0.125 mmol/L), mitomycin C (9.4 nmol/L), or both compounds for 2 days. Chromosome spreadswere analyzed. Arrow indicates chromatid break, chromatid loss, or radial. Each radial is counted as equivalent to two chromatid breaks. C, chromatid breaksper metaphase.
Aggarwal et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5504
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

reactive oxygen species in chronic myeloid leukemia cells (31),it is unclear whether this pathway is relevant to a role of WRNhelicase to confer mitomycin C resistance in Fanconi anemia–deficient fibroblasts. Further studies will be necessary toevaluate whether an alternate end-joining pathway or homol-ogous recombination is important for repair of mitomycinC–induced lesions in the WRN helicase-inhibited condition.We also observed Rad51 foci accumulation in FA-D2�/�
cells cotreated with mitomycin C and NSC 617145, suggestingthat homologous recombination was activated and at leastsome DSBs were channeled into homology-mediated repair.However, as WRN plays a role in resolution of homologousrecombination intermediates (6), NSC 617145 inhibition ofWRN helicase activity might lead to accumulation of homol-ogous recombination structures. We observed phosphoryla-tion of DNA-PKcs only under conditions of WRN helicaseinhibition in FA-D2�/� cells, suggesting that NHEJwas elicited;however, the mechanism of DNA-PKcs recruitment is stillunclear.Development of helicase inhibitors such as NSC 617145 that
possess chemical stability and lack of nonspecific reactivity tothiol and other functional groups may prove useful for study ofcompensatory DNA repair pathways dependent on DNAunwinding by WRN or related helicases. In addition to theiruse as research tools, small-molecular inhibitors of WRN and
other helicases may be useful for development of anticancerstrategies that rely on synthetic lethality for targeting tumorswith preexisting DNA repair deficiencies (32).
Demonstration that very low concentrations of the che-motherapy drug mitomycin C are cytotoxic for Fanconianemia–mutant cells exposed to the helicase inhibitor mayhave implications for strategies that target Fanconi anemiaDNA repair pathway–deficient tumors. Certain sporadichead and neck, lung, ovarian, cervical, and hematologiccancers are characterized by epigenetic silencing of wild-type Fanconi anemia gene expression (33). It is estimatedthat 15% of all cancers harbor defects in the Fanconi anemiapathway (34). A significant number of these tumors maybecome reliant on WRN or other helicases to deal with DNAdamage such as strand breaks. WRN may be a suitable targetfor pharmacologic inhibition to sensitize Fanconi anemia–deficient tumors to chemotherapy drugs such as DNA cross-linkers.
In addition to epigenetic silencing, LOH from an additionalmutation in an Fanconi anemia gene in heterozygous carriersmay lead to increased cancer risk later in life (33, 35). Fanconianemia pathway–deficient fibroblasts were found to be highlysensitive to silencing of ATM kinase (33). FANCG- and FANCC-deficient pancreatic tumor lines were sensitive to a pharma-cologic inhibitor of ATM, raising the possibility for an
A
B
DMSO
DMSO
MMC
MMC
DAPIDNA-PKcs Merge DAPIDNA-PKcs Merge
% C
ell
s w
ith
DN
A-P
Kc
s
pS
20
56
fo
ci
100
80
60
40
20
0
NSC 617145
NSC 617145
NSC 617145
+
MMC
NSC 617145
+
MMC
DMSO MMC NSC 617145 NSC 617145
+
MMC
FA-D2+/+ FA-D2–/–
FA-D2+/+ FA-D2–/–
≤≤ 15 DNA-PKcs pS2056 foci
> 15 DNA-PKcs pS2056 foci
Figure 6. NSC 617145 exposureelicits NHEJ pathway in Fanconianemia–deficient cells uponcotreatment with mitomycin C. A,FA-D2�/� and FA-D2þ/þ cells weretreated with NSC 617145 (0.125mmol/L), mitomycin C (9.4 nmol/L),or both compounds for 2 days.Cells were stained with anti-DNA-PKcs-pS2056 antibody or 40,6-diamidino-2-phenylindole (DAPI).B, percentage of cells with DNA-PKcs-pS2056 foci (�15 or >15, asindicated) is shown.
Role of WRN Helicase in Fanconi Anemia–Deficient Cells
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5505
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

anticancer treatment. More recently, Fanconi anemia–defi-cient cell lines were shown to be hypersensitive to inhibition ofCHK1 kinase either by siRNA or a pharmacologic inhibitor ofCHK1 kinase activity (36). Unlike the hypersensitivity of Fan-coni anemia–deficient cells to CHK1 silencing or kinase inhi-bition, WRN depletion did not increase sensitivity of FA-D2�/�
cells tomitomycin C. This suggests that NSC 617145 exerted itseffect through a dominant-negative mechanism in which NSC617145–inhibited WRN helicase induced greater damage byblocking compensatory mechanism(s). Inhibition of WRN or arelated helicase by a small moleculemay provide an alternativestrategy for targeting Fanconi anemia–deficient tumors that isunique from approaches such as targeting the CHK1 or ATMkinase response.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design:M. Aggarwal, P. Pichierri, R.H. Shoemaker, R.M. Brosh,Jr.Development of methodology: M. Aggarwal, J.A. Sommers, R.M. Brosh, Jr.Acquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): M. Aggarwal, C. Iannascoli
Analysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): M. Aggarwal, T. Banerjee, J.A. Sommers, C. Iannas-coli, P. Pichierri, R.H. Shoemaker, R.M. Brosh, Jr.Writing, review, and/or revision of the manuscript:M. Aggarwal, T. Bane-rjee, J.A. Sommers, P. Pichierri, R.H. Shoemaker, R.M. Brosh, Jr.Administrative, technical, or material support (i.e., reporting or organiz-ing data, constructing databases):M. Aggarwal, J.A. Sommers, R.M. Brosh, Jr.Study supervision: R.M. Brosh, Jr.
AcknowledgmentsThe authors thank Fanconi Anemia Research Fund for FA-A and FA-D2 cell
lines, and Dr. Ian Hickson (University of Copenhagen) for recombinant BLMprotein and BLM�/� (PSNG13) and BLMþ/þ (PSNF5) cells. The authors alsothank the Flow Cytometry and Cell Sorting Shared Resource at GeorgetownUniversity, Lombardi Comprehensive Cancer Center, which is partially sup-ported by NIH/NCI grant P30-CA051008.
Grant SupportThis work was supported by Intramural Research Program of NIH, National
Institute on Aging, and National Cancer Institute, and Fanconi Anemia ResearchFund (R.M. Brosh, Jr.).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
Received July 27, 2012; revised May 10, 2013; accepted June 7, 2013;published OnlineFirst July 18, 2013.
A
B
DMSO
DMSO
MMC
MMC
DAPIRad51 Merge DAPIRad51 Merge
% N
ucle
i w
ith
Rad
51
fo
ci
100
80
60
40
20
0
NSC 617145
NSC 617145
NSC 617145
+
MMC
NSC 617145
+
MMC
FA-D2+/+ FA-D2–/–
FA-D2+/+
FA-D2–/–
Figure 7. Increased accumulationof Rad51 foci in FA-D2–mutantcells cotreated with NSC 617145and mitomycin C. FA-D2�/� andFA-D2þ/þ cells were treated withNSC 617145 (0.125 mmol/L),mitomycin C (9.4 nmol/L), or bothcompounds for 2 days. A, cellswere stained with anti-Rad51antibody or 40,6-diamidino-2-phenylindole (DAPI). B, percentageof cells with Rad51 foci is shown.
Aggarwal et al.
Cancer Res; 73(17) September 1, 2013 Cancer Research5506
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

References1. Martin GM. Genetic syndromes in man with potential relevance to the
pathobiology of aging. Birth Defects Orig Artic Ser 1978;14:5–39.2. Yu CE, Oshima J, Fu YH, Wijsman EM, Hisama F, Alisch R, et al.
Positional cloning of the Werner's syndrome gene. Science 1996;272:258–62.
3. Monnat RJ Jr. Human RECQ helicases: roles in DNA metabolism,mutagenesis and cancer biology. Semin Cancer Biol 2010;20:329–39.
4. Rossi ML, Ghosh AK, Bohr VA. Roles of Werner syndrome protein inprotection of genome integrity. DNA Repair 2010;9:331–44.
5. Sharma S, Doherty KM, Brosh RM Jr. Mechanisms of RecQ helicasesin pathways ofDNAmetabolism andmaintenance of genomic stability.Biochem J 2006;398:319–37.
6. Saintigny Y, Makienko K, Swanson C, Emond MJ, Monnat RJ Jr.Homologous recombination resolution defect in Werner syndrome.Mol Cell Biol 2002;22:6971–8.
7. Sidorova JM, Li N, Folch A, Monnat RJ Jr. The RecQ helicase WRN isrequired for normal replication fork progression after DNA damage orreplication fork arrest. Cell Cycle 2008;7:796–807.
8. Poot M, Yom JS, Whang SH, Kato JT, Gollahon KA, Rabinovitch PS.Werner syndrome cells are sensitive to DNA cross-linking drugs.FASEB J 2001;15:1224–6.
9. Poot M, Gollahon KA, Emond MJ, Silber JR, Rabinovitch PS. Wernersyndrome diploid fibroblasts are sensitive to 4-nitroquinoline-N-oxideand 8-methoxypsoralen: implications for the disease phenotype.FASEB J 2002;16:757–8.
10. Cheng WH, Kusumoto R, Opresko PL, Sui X, Huang S, Nicolette ML,et al. Collaboration of Werner syndrome protein and BRCA1 in cellularresponses to DNA interstrand cross-links. Nucleic Acids Res 2006;34:2751–60.
11. ChengWH,MufticD,MuftuogluM,Dawut L,Morris C,Helleday T, et al.WRN is required for ATM activation and the S-phase checkpoint inresponse to interstrand crosslink-induced DNA double strand breaks.Mol Biol Cell 2008;19:3923–33.
12. Zhang N, Kaur R, Lu X, Shen X, Li L, Legerski RJ. The Pso4 mRNAsplicing and DNA repair complex interacts with WRN for processing ofDNA interstrand cross-links. J Biol Chem 2005;280:40559–67.
13. D'Andrea AD, Grompe M. The Fanconi anaemia/BRCA pathway. NatRev Cancer 2003;3:23–34.
14. Auerbach AD. Fanconi anemia and its diagnosis. Mutat Res 2009;668:4–10.
15. KeeY,D'AndreaAD. Expanded roles of theFanconi anemia pathway inpreserving genomic stability. Genes Dev 2010;24:1680–94.
16. Adamo A, Collis SJ, Adelman CA, Silva N, Horejsi Z, Ward JD, et al.Preventing nonhomologous end joining suppresses DNA repairdefects of Fanconi anemia. Mol Cell 2010;39:25–35.
17. Pace P, Mosedale G, Hodskinson MR, Rosado IV, SivasubramaniamM, Patel KJ. Ku70 corrupts DNA repair in the absence of the Fanconianemia pathway. Science 2010;329:219–23.
18. Bunting SF, Callen E, Kozak ML, Kim JM, Wong N, Lopez-ContrerasAJ, et al. BRCA1 functions independently of homologous recombina-tion in DNA interstrand crosslink repair. Mol Cell 2012;46:125–35.
19. Aggarwal M, Sommers JA, Shoemaker RH, Brosh RM Jr. Inhibition ofhelicase activity by a small molecule impairs Werner syndrome heli-
case (WRN) function in the cellular response to DNA damage orreplication stress. Proc Natl Acad Sci U S A 2011;108:1525–30.
20. Bunz F, Dutriaux A, Lengauer C, Waldman T, Zhou S, Brown JP, et al.Requirement for p53 and p21 to sustain G2 arrest after DNA damage.Science 1998;282:1497–501.
21. Davies SL, North PS, Dart A, Lakin ND, Hickson ID. Phosphorylation ofthe Bloom's syndrome helicase and its role in recovery from S-phasearrest. Mol Cell Biol 2004;24:1279–91.
22. Gaymes TJ, North PS, Brady N, Hickson ID, Mufti GJ, Rassool FV.Increased error-prone non homologous DNA end-joining–a pro-posed mechanism of chromosomal instability in Bloom's syndrome.Oncogene 2002;21:2525–33.
23. Schlacher K, Wu H, Jasin M. A distinct replication fork protectionpathway connects fanconi anemia tumor suppressors to RAD51-BRCA1/2. Cancer Cell 2012;22:106–16.
24. GrayMD, Shen JC, Kamath-Loeb AS, Blank A, Sopher BL, Martin GM,et al. The Werner syndrome protein is a DNA helicase. Nat Genet1997;17:100–3.
25. Kastan MB, Bartek J. Cell-cycle checkpoints and cancer. Nature2004;432:316–23.
26. Bunting SF, Nussenzweig A. Dangerous liaisons: Fanconi anemia andtoxic nonhomologous end joining in DNA crosslink repair. Mol Cell2010;39:164–6.
27. Deans AJ, West SC. DNA interstrand crosslink repair and cancer. NatRev Cancer 2011;11:467–80.
28. Chen BP, Chan DW, Kobayashi J, Burma S, Asaithamby A, Morotomi-YanoK, et al. Cell cycle dependence of DNA-dependent protein kinasephosphorylation in response to DNA double strand breaks. J BiolChem 2005;280:14709–15.
29. Ohashi A, Zdzienicka MZ, Chen J, Couch FJ. Fanconi anemia com-plementation group D2 (FANCD2) functions independently of BRCA2and RAD51-associated homologous recombination in response toDNA damage. J Biol Chem 2005;280:14877–83.
30. Chen L,HuangS, Lee L, Davalos A, Schiestl RH,Campisi J, et al.WRN,the protein deficient inWerner syndrome, plays a critical structural rolein optimizing DNA repair. Aging Cell 2003;2:191–9.
31. Sallmyr A, TomkinsonAE, Rassool FV. Up-regulation ofWRNandDNAligase IIIalpha in chronic myeloid leukemia: consequences for therepair of DNA double-strand breaks. Blood 2008;112:1413–23.
32. Aggarwal M, Brosh RM Jr. Hitting the bull's eye: novel directed cancertherapy through helicase-targeted synthetic lethality. J Cell Biochem2009;106:758–63.
33. Kennedy RD,ChenCC, Stuckert P, Archila EM, de la VegaMA,MoreauLA, et al. Fanconi anemia pathway-deficient tumor cells are hyper-sensitive to inhibition of ataxia telangiectasia mutated. J Clin Invest2007;117:1440–9.
34. Taniguchi T,D'AndreaAD.Molecular pathogenesis of Fanconi anemia:recent progress. Blood 2006;107:4223–33.
35. Hucl T, Gallmeier E. DNA repair: exploiting the Fanconi anemia path-way as a potential therapeutic target. Physiol Res 2011;60:453–65.
36. Chen CC, Kennedy RD, Sidi S, Look AT, D'Andrea A. CHK1 inhibitionas a strategy for targeting Fanconi Anemia (FA) DNA repair pathwaydeficient tumors. Mol Cancer 2009;8:24.
Role of WRN Helicase in Fanconi Anemia–Deficient Cells
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5507
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975

2013;73:5497-5507. Published OnlineFirst July 18, 2013.Cancer Res Monika Aggarwal, Taraswi Banerjee, Joshua A. Sommers, et al. PathwayResponses in the Absence of a Functional Fanconi Anemia Werner Syndrome Helicase Has a Critical Role in DNA Damage
Updated version
10.1158/0008-5472.CAN-12-2975doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/07/18/0008-5472.CAN-12-2975.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/73/17/5497.full#ref-list-1
This article cites 36 articles, 14 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/73/17/5497.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/73/17/5497To request permission to re-use all or part of this article, use this link
on February 15, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-2975