vliv zvýšené koncentrace co na anatomii listu...
TRANSCRIPT

Univerzita Karlova v Praze
Přírodovědecká fakulta
Katedra fyziologie rostlin
Vliv zvýšené koncentrace CO2 na anatomii listu jehličnanů
Bakalářská práce
Zuzana Kubínová
Praha 2008

1
Vedoucí bakalářské práce:
Mgr. Zuzana Lhotáková, Katedra fyziologie rostlin PřF UK
Konzultant bakalářské práce:
doc. RNDr. Jana Albrechtová, Ph.D., Katedra fyziologie rostlin PřF UK
Prohlašuji, že jsem tuto bakalářskou práci vypracovala samostatně s použitím citované
literatury pod vedením Mgr. Zuzany Lhotákové a doc. Jany Albrechtové a souhlasím s jejím
zveřejňováním.
V Praze, dne Zuzana Kubínová

2
Poděkování
Děkuji Zuzce Lhotákové a doc. Janě Albrechtové za nezměrnou vstřícnost, podporu a
trpělivost a za cenné připomínky v průběhu vytváření této práce.
Děkuji za příjemné prostředí a podporu dalším členům Katedry fyziologie rostlin PřF UK
v Praze.
Děkuji svým rodičům za povzbuzování a toleranci a své sestře za přípravu pokrmů, které mi
dodaly síly do další práce.

3
OBSAH:
Seznam použitých zkratek.......................................................................................................... 4 Abstrakt ...................................................................................................................................... 5 Abstract ...................................................................................................................................... 6 1. Úvod ....................................................................................................................................... 7
1.1 Anatomie listu jehličnanů – stavba jehlice....................................................................... 7 1.1.1 Pokožka ..................................................................................................................... 8 1.1.2 Mezofyl ..................................................................................................................... 9 1.1.3 Vodivá pletiva ........................................................................................................... 9
1.2 Koncentrace CO2 a její vliv na rostliny.......................................................................... 11 1.2.1 Změny koncentrace CO2 v atmosféře Země ........................................................... 11 1.2.2 Studium vlivu zvýšené koncentrace CO2 na rostliny .............................................. 13 1.2.3 Vliv zvýšeného obsahu CO2 v atmosféře na rostliny.............................................. 18
2. Vliv oxidu uhličitého na strukturu listu................................................................................ 20 2.1 Změny v morfogenezi listu ............................................................................................ 20
2.1.1 Změny v zakládání a senescenci listů ..................................................................... 20 2.1.2 Změny v relativní rychlosti růstu listů .................................................................... 21 2.1.3 Změny ve zvětšování a tvorbě buněčné stěny......................................................... 23 2.1.4 Změny v morfologických parametrech listu ........................................................... 25 2.1.5 Změny v tvorbě sušiny a asimilátů.......................................................................... 26
2.2 Mikroskopické změny struktury listu............................................................................. 28 2.2.1 Krycí pletiva............................................................................................................ 28
2.2.1.1 Průduchy........................................................................................................... 28 2.2.2 Mezofyl ................................................................................................................... 30
2.2.2.1 Chloroplasty ..................................................................................................... 31 2.2.2.2 Obsah nestrukturních sacharidů v listu ............................................................ 33
2.2.3 Vodivá pletiva ......................................................................................................... 33 3. Závěr..................................................................................................................................... 34 4. Citovaná literatura ................................................................................................................ 36

4
Seznam použitých zkratek
AC (ambient concentration) normální koncentrace CO2
EC (elevated concentration) zvýšená koncentrace CO2
FACE free air CO2 enrichment
OTC (open top chamber) otevřená růstová komora
ppm (parts per milion) jednotka koncentrace – miliontina objemu (1:106), počet objemových
jednotek z milionu těchto jednotek
RGR (relative growth rate) relativní rychlost růstu, tj. změna hmotnosti biomasy na jednotku
biomasy za čas
SLA (specific leaf area) specifická listová plocha, tj. plocha listu vztažená na jednotku
hmotnosti sušiny listu
SLW (specific leaf weight) specifická hmotnost listu, tj. hmotnost sušiny listu vztažená na
plochu listu; též LMA (specific leaf mass area), SLM (specific leaf mass), LSM (leaf specific
mass)
UDP-Glc dehydrogenáza uridindifosfát-glukóza dehydrogenáza, enzym, který se podílí na
dráze tvorby součástí buněčné stěny
XET (xyloglucan endotransglycosylase) xyloglukan endotransglykosyláza
[CO2] koncentrace oxidu uhličitého

5
Abstrakt
V posledních desetiletích narůstá koncentrace CO2 v atmosféře Země, což má vliv na
rostliny, neboť CO2 je substrátem pro fotosyntézu a zdrojem uhlíku, jenž je vázán do jejich
biomasy. Bylo zjištěno, že zvýšená koncentrace CO2 v atmosféře ovlivňuje fyziologické
procesy v rostlinách, zejména má vliv na rychlost fotosyntézy, tvorbu a ukládání zásobních
látek a na fyziologické procesy ovlivňované průduchy. Vzhledem k propojenosti struktur a
funkce též působí změny stavby rostlinných orgánů, především listů, které jsou místem, kde
fotosyntéza probíhá.
Změny v anatomii listu se mohou projevit na všech úrovních pozorování. Nejsou však
u všech zkoumaných druhů rostlin konzistentní, což naznačuje komplikovanost působení
změn koncentrace CO2 na rostliny. U anatomických struktur se vliv zvýšené koncentrace CO2
nejvíce projevuje při jejich vytváření, zatímco po ukončení vývoje listu nedochází
k výrazným změnám jeho stavby. Zvýšenou koncentrací CO2 jsou proto více ovlivněny
mladé, rostoucí rostliny. Při morfogenezi listu se může změnit velikost a rychlost tvorby
buněk, a tím i tvar a velikost celého listu. Změny se dále projevují ve fyziologických a
strukturálních vlastnostech pletiv, např. v parametrech mezofylu a vodivých pletiv či v počtu
průduchů, kdy se při vyšší koncentraci CO2 obvykle snižuje hustota průduchů. V neposlední
řadě může být ovlivněna ultrastruktura chloroplastů.
Ze studií působení zvýšené koncentrace CO2 na anatomii listu často i u příbuzných
druhů vyplývají protichůdné výsledky. Tato skutečnost je zřejmě způsobena kombinací
environmentálních a vnitřních faktorů, jejichž vliv je třeba brát v potaz při odhalování
mechanismů tohoto komplikovaného procesu.
Cílem předkládané práce je zhodnotit vliv zvýšené koncentrace CO2 v atmosféře na
anatomii listu jehličnanů. První část práce nabízí nejprve shrnutí základních anatomických
charakteristik listů jehličnanů, dále je věnována změnám koncentrace oxidu uhličitého
v atmosféře, vlivům CO2 na rostliny a souhrnu experimentálních metod studia působení CO2
na rostliny. Ve druhé, hlavní části práce je popsáno působení oxidu uhličitého na anatomii
listu se zaměřením na jehlice jehličnanů ve vztahu k procesům morfogeneze listu a
mikroskopickým změnám struktury jehlic.
Klíčová slova: anatomie listu, jehlice, koncentrace CO2, mezofyl, morfogeneze listu,
pokožka, průduchy, strukturální parametry listu, rychlost růstu

6
Abstract
During the last decades CO2 concentration has been increasing in the Earth
atmosphere, thus affecting plants because CO2 represents a substrate for photosynthesis and a
source of carbon, which is bound into their biomass. It was found that the elevated CO2
concentration affects physiological processes in plants, namely photosynthetic rate,
production and accumulation of storage substances and the physiological processes affected
by stomata. Regarding interconnection of structure and function, elevated CO2 concentration
also causes changes in the structure of plant organs, particularly leaves, in which
photosynthesis takes place.
Changes in the leaf anatomy can be found at all levels of observation. However, they
are not consistent, which indicates the complexity of the effect of the elevated CO2
concentration on plants. In the case of leaf anatomical structures, this effect is strongest
during the leaf development, while in the structure of mature leaves there are not so distinct
changes. Young, developing plants are thus more strongly affected by the elevated CO2
concentration. During the leaf morphogenesis the cell size and cell production rate can
change, resulting also in shape and size changes of entire leaf. Further, there are changes in
physiological and structural characteristics of tissues, e.g. in parameters of mesophyll,
vascular tissues and stomata. The elevated CO2 concentration can result in the increase of the
stomatal density and stomatal index. It can also influence the chloroplast ultrastructure.
The results of studies of the effect of the elevated CO2 concentration on leaf anatomy
are often contradictory, even in the case of related species. This fact is caused by the
combined effect of different environmental and internal factors, which has to be taken into
account in elucidation of mechanisms of this complicated process.
The aim of the present review was to evaluate the effect of elevated CO2 concentration
on leaf anatomy of coniferous needles. The first part of the review briefly describes basic
anatomical characteristics of needles, then shortly refers to changes in CO2 concentration in
atmosphere and finally it reviews currently available methods of simulation of CO2
enrichment. The second, main part then describes the effects of elevated CO2 concentration on
leaf anatomy with emphasis given to coniferous needles. This part is focused on processes of
leaf morphogenesis and on microscopic changes of needles.
Keywords: leaf anatomy, needles, CO2 concentration, mesophyl, leaf morphogenesis,
epidermis, stomata, leaf structural parameteres, growth rate

7
1. Úvod
Anatomie listu rostlin je ovlivněna podmínkami, ve kterých rostliny rostou. Jedním
z faktorů prostředí, který ovlivňuje strukturu listů, je hodnota koncentrace oxidu uhličitého
v okolním vzduchu. Cílem této práce je shrnout poznatky o vlivech zvýšené koncentrace CO2
v atmosféře na anatomii listu jehličnanů. Původním záměrem práce bylo soustředit se na
danou problematiku pouze u jehličnanů, avšak vzhledem k menšímu množství literatury
zabývající se jehličnany byly využity i výsledky prací, které zkoumaly vliv zvýšené
koncentrace CO2 na strukturu listů jiných rostlin.
První, úvodní část práce nabízí shrnutí základních anatomických charakteristik listů
jehličnanů, dále je věnována změnám koncentrace oxidu uhličitého v atmosféře, vlivům CO2
na rostliny a souhrnu experimentálních metod studia působení CO2 na rostliny. Ve druhé,
hlavní části práce – v kapitole 2 – je popsáno působení oxidu uhličitého na anatomii listu se
zaměřením na jehlice jehličnanů na různých úrovních pozorování – od změn na
makroskopické úrovni až po změny v ultrastruktuře. Tato část pojednává o změnách
v morfogenezi listu způsobených zvýšenou koncentrací CO2 (kap. 2.1.) a dále o
mikroskopických změnách struktury zralých jehlic (kap. 2.2). Následuje závěr a seznam
citované literatury.
1.1 Anatomie listu jehličnanů – stavba jehlice
Anatomická stavba listu byla studována již od objevení mikroskopie a byla popsána
v mnoha učebních textech, které se zabývají anatomií rostlin (Esau, 1977; Mauseth, 1988;
Fahn, 1990; Pazourek a Votrubová, 1997; Dickison, 2000; Votrubová, 2001; Evert, 2006),
poznatky z uvedených zdrojů byly využity v celé této kapitole. Jehlice smrku ztepilého byly
experimentálním materiálem několika diplomových a dizertačních prací týmu doc.
Albrechtové a následných publikací (Soukupová et al., 2000a; Soukupová et al., 2000b), ze
kterých byla v této kapitole využita některá jejich mikroskopická dokumentace.
List je nejrozmanitějším rostlinným orgánem. Existuje více typů listu, nejčastější jsou
asimilační listy (trofofyly), kterých se týkají dále uváděné údaje. Stavba listu je dána jeho
funkcí – list slouží hlavně k fotosyntéze a transpiraci. Je třeba, aby zachytil co nejvíce záření,
a proto bývá u většiny rostlin plochý a zároveň tenký, aby se záření dostalo skrze list do všech
jeho částí. Listy jehličnanů jsou tenké, úzké a protáhlé; pro svůj tvar se nazývají jehlice. Jsou
to jednožilné xeromorfní listy, které vyrůstají ve svazcích o jedné až pěti jehlicích. Počtem

8
jehlic ve svazku může být ovlivněn jejich tvar na příčném průřezu, příkladem je tvar jehlice
borovice, který záleží na počtu jehlic ve svazku – svazek je na průřezu kruhový – viz obr.
1.1 c. Na obrázcích 1.1a-f je znázorněna stavba jehlic několika druhů stromů. Listy jehličnanů
většinou přes zimu neopadávají, výjimkou je například modřín a tis.
1.1.1 Pokožka
List je ohraničen krycím pletivem – pokožkou (epidermis), která bývá tvořena jednou
vrstvou dlaždicovitých, těsně se dotýkajících buněk (obr. 1.1a, e). Části jejich buněčných stěn
vystavené okolnímu prostředí bývají ztlustlé a nacházejí se v nich hydrofóbní látky – kutin,
vosky a na povrchu nebuněčná vrstva kutikula. Tyto látky omezují propustnost stěn pro vodu.
Jehlice bývají často vystaveny působení faktorů prostředí i po několik let, a proto mají buňky
jejich pokožky tlusté buněčné stěny a povrch jehlic je potažen silnou kutikulou
s epikutikulárními vosky.
V pokožce se nacházejí průduchy, které umožňují výměnu plynů mezi vnitřním
prostorem listu a okolím. U jehlic, stejně jako u jiných listů, obsahují svěrací buňky průduchů
narozdíl od ostatních buněk pokožky chloroplasty. Míra dostupnosti CO2 pro biochemický a
fotochemický aparát fotosyntézy, která reguluje tok CO2 podle jeho potřebnosti uvnitř listu, je
vyjádřena parametrem vodivosti průduchů. Při uzavření průduchů je rychlost fotosyntézy
limitována dostupností CO2, jehož koncentrace je v intercelulárách nízká. Průduchy mohou
být u jehlic zanořené pod povrch listu, jako je tomu například u smrku ztepilého a některých
druhů borovic a douglasek (obr. 1.1b), aby se z nich uvolněné molekuly vody nedostaly
snadno do volného vzduchu, ale hromadily se v epistomatální dutině. Tím vytvoří vyšší
koncentraci vodní páry v okolí průduchu, čímž se sníží výdej vody z listu.
Průduchy jsou na všech stranách jehlice, obvykle uspořádané v řadách, jako je tomu
např. u smrku a borovice. Počet a hustota průduchů záleží druhu rostliny i na podmínkách
okolního prostředí Hustota průduchů, tedy počet průduchů na mm2, se liší mezi druhy rostlin i
mezi oblastmi listu, protože v rámci listu existují vývojové gradienty, a její hodnota se
pohybuje od desítek po stovky. Hustota průduchů je citlivá na změny koncentrace CO2, čehož
je využíváno při paleoklimatologických studiích (viz kapitola 2.2.1.1). Na krátkodobé nebo
náhlé změny vlastností okolního prostředí průduchy odpovídají změnami otevřenosti, která
ovlivňuje vodivost průduchů a tím i rychlost transpirace.

9
1.1.2 Mezofyl
Pod pokožkou se u jehlic obvykle nachází vrstva sklerenchymatických buněk, které
tvoří spodní pokožku, takzvanou hypodermis (Fahn, 1990). Tato vrstva není v místech, kde se
nacházejí vertikální řady průduchů (obr. 1.1e). Hypodermis se nalézá u většiny druhů
jehličnanů, například u smrku ztepilého Picea abies, u borovic, u čeledi Araucariaceae a u
dalších druhů, zatímco u tisu a toreje se nevyskytuje vůbec (Esau, 1977).
Základní listové pletivo – listový mezofyl – tvoří parenchymatické buňky, v nichž se
nachází velký počet chloroplastů. Mezofylové buňky bývají u jehlic laločnaté, s výběžky
buněčných stěn trčícími do mezibuněčných prostor, které se nazývají interceluláry, a jimiž
proudí do mezofylu plyny přijaté prostřednictvím průduchů. Vnitřní povrch listu, tj. součet
povrchů všech stěn mezofylových buněk vystavených intercelulárám, je důležitý kvantitativní
parametr listu.
Jehlice jsou listy ekvifaciální, tj. mají podobnou stavbu mezofylu na obou stranách
listu. Jejich mezofyl je tvořen buď několika vrstvami nerozlišených laločnatých
chlorenchymatických buněk – např. u borovice, smrku a cedru, nebo je rozlišen na houbovitý
a palisádový parenchym – např. u jedle, tisu, douglasky, blahočetu, ostrolistce, sekvoje a
Podocarpu (Mauseth, 1988). V mezofylu jehlic se může vyskytovat různý počet
pryskyřičných kanálků, které mají na průřezu kruhový okraj tvořený epiteliálními buňkami
(obr. 1.1a).
1.1.3 Vodivá pletiva
Vodivá pletiva se nacházejí ve středním válci jehlice, který je ohraničen endodermis,
vrstvou zpravidla tlustostěnných buněk (obr. 1.1a, e). Jehlicemi probíhá jeden (např. rod
Picea) nebo dva (např. rod Pinus) cévní svazky, které bývají nerozvětvené. Sestávají
z xylému, umístěného blíže k přivrácené (adaxiální) straně a z floému blíže k spodní,
odvrácené (abaxiální) straně jehlice. Prostor mezi cévními svazky a endodermis vyplňuje u
jehlic transfúzní pletivo, tvořené tracheidami a parenchymatickými buňkami, z nichž mnohé
obsahují velké vakuoly s polyfenolickými látkami. Ve středním válci se nevyskytují žádné
mezibuněčné prostory (Esau, 1977).

10

11
Obr. 1.1: Struktura jehlic na příčném řezu a Jehlice Pinus monophylla. A příčný řez celou jehlicí, B podélný řez mezofylem a trasfúzním pletivem, C příčný řez středem jehlice. Převzato z Esau (1977), upraveno. b Detail zanořeného průduchu smrku ztepilého Picea abies ucpaného voskem. Převzato ze Soukupová et al. (2000a). c Schéma uspořádání jehlic borovic ve svazcích a tvarů jejich příčných řezů. Převzato z < http://www.lovett-pinetum.org/drawingneedles.jpg> a upraveno. d Příčný řez jehlicí Picea pungens. Převzato ze Soukupová et al. (2000b). e Popis základních struktur na příkladu jehlice P. pungens. Převzato ze Soukupová et al. (2000b) a upraveno. f Příčné řezy jehlicemi: A borovice černé Pinus nigra, B modřínu Larix sp., C tisu Taxus canadensis, D Abies sp. Převzato z Esau (1977), upraveno.
1.2 Koncentrace CO2 a její vliv na rostliny
1.2.1 Změny koncentrace CO2 v atmosféře Země
Oxid uhličitý patří mezi tzv. skleníkové plyny, které zachycují a odrážejí infračervené
záření, jež by se bez jejich přítomnosti v atmosféře rychleji uvolnilo do vesmíru, čímž se
podílejí na zvyšování teploty zemské atmosféry. Tento jev se nazývá skleníkový efekt. Je
popsán na obr. 1.2.
Obr 1.2: Schéma skleníkového efektu. Převzato z Wikimedia <http://upload.wikimedia.org/wikipedia/commons/e/ec/Schema_sklenikovy_efekt.gif>
Z údajů mnoha měření vyplývá, že se v posledních desítkách let zvyšuje obsah oxidu
uhličitého v zemské atmosféře (viz grafy na obr. 1.3 a 1.4). V důsledku lidské činnosti, tj.
spalováním fosilních paliv, průmyslovou výrobou a změnami využití krajiny, zejména
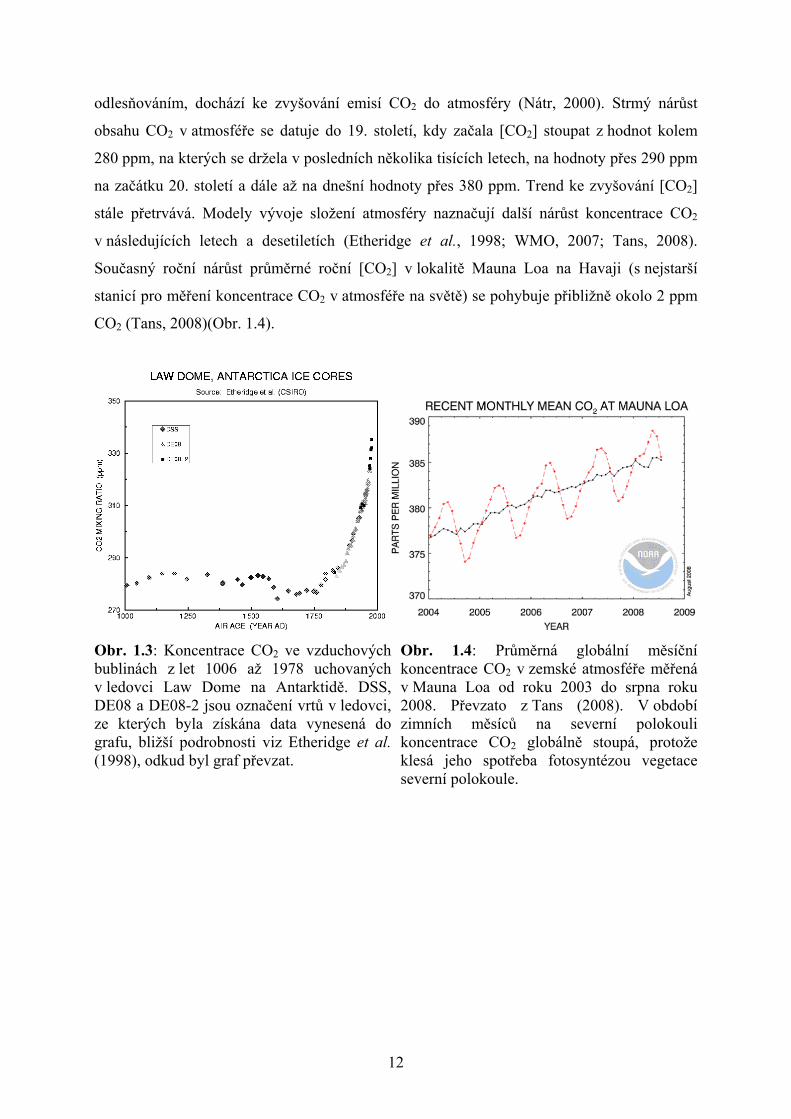
12
odlesňováním, dochází ke zvyšování emisí CO2 do atmosféry (Nátr, 2000). Strmý nárůst
obsahu CO2 v atmosféře se datuje do 19. století, kdy začala [CO2] stoupat z hodnot kolem
280 ppm, na kterých se držela v posledních několika tisících letech, na hodnoty přes 290 ppm
na začátku 20. století a dále až na dnešní hodnoty přes 380 ppm. Trend ke zvyšování [CO2]
stále přetrvává. Modely vývoje složení atmosféry naznačují další nárůst koncentrace CO2
v následujících letech a desetiletích (Etheridge et al., 1998; WMO, 2007; Tans, 2008).
Současný roční nárůst průměrné roční [CO2] v lokalitě Mauna Loa na Havaji (s nejstarší
stanicí pro měření koncentrace CO2 v atmosféře na světě) se pohybuje přibližně okolo 2 ppm
CO2 (Tans, 2008)(Obr. 1.4).
Obr. 1.3: Koncentrace CO2 ve vzduchových bublinách z let 1006 až 1978 uchovaných v ledovci Law Dome na Antarktidě. DSS, DE08 a DE08-2 jsou označení vrtů v ledovci, ze kterých byla získána data vynesená do grafu, bližší podrobnosti viz Etheridge et al. (1998), odkud byl graf převzat.
Obr. 1.4: Průměrná globální měsíční koncentrace CO2 v zemské atmosféře měřená v Mauna Loa od roku 2003 do srpna roku 2008. Převzato z Tans (2008). V období zimních měsíců na severní polokouli koncentrace CO2 globálně stoupá, protože klesá jeho spotřeba fotosyntézou vegetace severní polokoule.

13
1.2.2 Studium vlivu zvýšené koncentrace CO2 na rostliny
Při studiu vlivů zvýšené [CO2] na rostliny se používají různé metody obohacování
vzduchu oxidem uhličitým, které mají své výhody i nevýhody, přičemž v současné době se
jeví jako nejvhodnější metoda FACE, tj. Free Air CO2 Enrichment (přehled viz Nátr, 2000).
Nyní je ve světe v provozu několik desítek těchto systémů, viz např. seznam na webových
stránkách <http://public.ornl.gov/face/global_face.shtml>. Na obr. 1.5a-o jsou fotografie
něktrých lokalit. Systém FACE sestává z dutých sloupků, postavených okolo pokusné plochy,
ze kterých na pokusné rostliny tryská oxid uhličitý. Díky informacím z čidel je pomocí
počítačů regulován přívod CO2 tak, aby byl na pokusné ploše homogenní vzduch se zadaným
obsahem CO2. Mezi výhody této metody patří zejména skutečnost, že pokusné rostliny rostou
při zvýšené [CO2], ale jinak téměř ve svém přirozeném prostředí, a že jsou zvýšenou [CO2]
ovlivněna celá společenstva rostlin na ekosystémové úrovni. Hlavní nevýhodou metody
FACE je její nákladnost, zejména kvůli velké spotřebě CO2 (Ceulemans a Mousseau, 1994).
Další metodou, používanou při studiu vlivu EC na rostliny, je pěstování rostlin v
růstových komorách. Nejdříve se začaly používat uzavřené růstové komory, často umístěné
uvnitř skleníků. Jejich hlavní nevýhodou je snížená ozářenost pěstovaných rostlin. Tato
nevýhoda se téměř vyloučí použitím otevřených růstových komor (OTC, Open Top
Chamber), uvnitř kterých je množství světla srovnatelné s okolním prostředím (obr. 1.5p-r).
OTC mohou obklopovat nejen rostliny v květináčích, ale i jedince rostoucí v přirozené půdě.
Nevýhodou těchto komor je mírně odlišná teplota a relativní vlhkost vzduchu a také snížená
rychlost větru vzhledem k okolí. Tato metoda je také nákladná díky velké spotřebě oxidu
uhličitého (Ceulemans a Mousseau, 1994). V České republice se OTC nacházejí na
experimentálním pracovišti ÚSBE na Bílém Kříži v Beskydech (obr. 1.5p).
Při měření fyziologických parametrů rostlin, např. při stanovování rychlosti
fotosyntézy a transpirace při EC, se uplatňují kyvety nebo vaky připevňované na jednotlivé
větve nebo části větví. Hlavní nevýhodou zmiňované metody je to, že jsou zvýšenou [CO2]
ovlivněny pouze části rostlin (Nátr, 2000). V neposlední řadě lze ke studiu vlivu EC na
rostliny využít přirozených výronů CO2. Výhodou je skutečně dlouhodobé působení EC na
okolní rostliny, ale tato metoda má i četné nevýhody – např. vypouštění CO2 ze zdroje není
v průběhu času rovnoměrné a [CO2] kolísá působením větrů. Často se též k ovlivněným
rostlinám obtížně hledají takové protějšky, které by rostly poblíž v obdobných podmínkách,
avšak v AC (Nátr, 2000).

14

15
Obr. 1.5: Systémy FACE (Free-Air CO2 Enrichment): a Aspen FACE použitý na topoly v Harshaw Experimental Forest poblíž Rhinelanderu ve státě Wisconsin v USA. Převzato z <http://aspenface.mtu.edu> b Systém FACE o průměru 22 m v porostu topolů Populus alba, Populus nigra a Populus x euramericana poblíž města Tuscania v provincii Viterbo ve střední Itálii. Převzato z <http://www.unitus.it/euroface/gallery/aero.htm> c Systémy FACE v lese s opadavými stromy v Oak Ridge National Laboratory v Tennesee v USA. Převzato z <http://www.rmrs.nau.edu/USAMAB/images/southern-appalacian_aerial-Aug00.jpg> d, e, f: Experimentální systém v lese Duke Forest poblíž Chapel Hill v Orange County v Severní Karolíně v USA: d Pohled shora, foto Les Todd, převzato z <http://www.dukemagazine.duke.edu/dukemag/issues/111205/depgaz17.html> e Pohled zespodu na centrální věž uprostřed kruhu FACE sloužící k odběru vzorků. Převzato z <http://www.duke.edu/~jspippen/work/face.htm> f Mnoho senzorů zaznamenává nejrůznější parametry půdy, stromů i ovzduší na experimentální ploše. <http://www.duke.edu/~jspippen/work/face.htm> g Kruhy o průměru 1,5 m v projektu TasFACE v Pontville v Tasmánii, převzato z <http://www.utas.edu.au/docs/plant_science/ps/ps/face_approach.html> h Pokusné pracoviště Sky Oaks, San Diego State University, USA. Převzato z <http://www.sci.sdsu.edu/GCRG/skyoaks/overhead.htm> i Pokusná plocha projektu BioCON v Cedar Creek v Minnesotě v USA. Převzato z <http://geology.about.com/library/weekly/aa071600b.htm> j Systém FACE o průměru 8 m v německém Giessenu Převzato z <http://www.uni-giessen.de/cms/fbz/fb08/biologie/pflanzenoek/forschung/Foeinr/UKL/projekte/GiFACE/GiFACE-System/image1> k Systém SoyFace pro zkoumání sóji – University of Illinois, Urbana Champaign, USA. Převzato z <http://www.soyface.uiuc.edu/WebImages/images/Slide11_tif.jpg> l Systém FACE v Braunschweigu, Německo. Převzato z <http://www.fal.de/cln_044/nn_792590/DE/aktuell/pressemitteilungen/2007/presse__30__2007__aoe.html__nnn=true> m Experimentální rýžové pole se systémem FACE v Shizukuishi, Iwate v Japonsku. Převzato z <www.niaes.affrc.go.jp/annual/r2000/html/rp4.html> n Systém SoyFace pro zkoumání sóji – University of Illinois, Urbana Champaign, USA. Převzato z <http://www.soyface.uiuc.edu/WebImages/images/ring6.jpg> o Systém FACE v Mohavské poušti ve státě Nevada v USA. Převzato z <http://www.unlv.edu/Climate_Change_Research/Pictures/NDFF1.htm> OTC – otevřené komory: p Kultivační sféry se smrkem na pracovišti ÚSBE na Bílém Kříži v Beskydech <http://www.usbe.cas.cz/lefr/kultivacni_sfery.htm> q Tervuren, Belgie <www.var.fgov.be/chip.php> r Jokioinen, Finsko <www.var.fgov.be/chip.php>
Výsledky experimentů záleží nejen na použité metodě zvýšení [CO2], délce působení
EC a na stáří zkoumaných rostlin, ale také na dalších faktorech. Körner (2006) zdůrazňuje, že

16
větší vliv na výsledek experimentu než použitá metoda obohacení vzduchu CO2 mají další
podmínky pro růst, zejména kvalita půdy, ve které jsou rostliny pěstovány.
K rozporům ve výsledcích může docházet i při srovnávání rostlin téhož druhu
zkoumaného za velmi podobných podmínek. Aby se zamezilo nesrovnalostem, je třeba
srovnávat dostatečný počet vzorků a při jejich odběru si všímat dalších faktorů, které mohou
vlastnosti vzorku ovlivnit – např. u jehlic vzít v úvahu jejich stáří a vertikální polohu v koruně
(Lin et al., 2001). Například se mezi vzorky odebranými v různých vrstvách koruny smrku
projevily rozdíly v aktivitě enzymu ribulózy-1,5-bisfosfát karboxylázy/oxygenázy (Hrstka et
al., 2005). Při anatomických analýzách je třeba uvážit možnost gradientu sledovaných
parametrů v rámci listu (Lhotáková et al., 2007).
Nelze jednoznačně říci, zda jsou výsledky studií vlivu zvýšené koncentrace oxidu
uhličitého na strukturu rostlin vždy porovnatelné, a to nejen proto, že jsou zkoumány odlišné
druhy rostlin, ale také pro různé pojetí normální a zvýšené koncentrace oxidu uhličitého.
Vzhledem k tomu, že [CO2] v atmosféře vzrůstá (obr. 1.3 a 1.4), vzrůstají během let i hodnoty
koncentrace CO2 považované v pokusech za normální – pohybují se od 250 do 400 ppm.
Hodnoty zvýšené koncentrace bývají většinou dvojnásobkem normálních hodnot, ale zdaleka
ne vždy. Tabulka 1.1 nabízí srovnání zkoumaných rostlinných druhů a použitých koncentrací
CO2, které byly uvedeny ve studiích citovaných v této práci.
Tab. 1.1: Shrnutí zkoumaných druhů rostlin a koncentrací považovaných v jednotlivých
studiích za normální (AC) a zvýšené (EC) koncentrace oxidu uhličitého.
Vysvětlivky: d – den; n – noc; ? – autoři neuvádějí hodnotu koncentrace, kterou považovali
v experimentu za normální, uvádějí pouze o kolik se zvýšená [CO2] lišila od AC; d* -
koncentrace CO2 byla zvýšena na uvedenou hodnotu jen ve dne.
Autoři, rok vydání AC (ppm) EC (ppm) Zkoumané rostliny
Apple et al., 2000 ?350 ?350+200 Pseudotsuga menziesii
Cavender-Bares et al., 2000 380 700 Quercus rubra Conroy et al., 1986 330 660 Pinus radiata Eguchi et al., 2004 360 720 Larix kaempferi Carr.
Fendrych, 2005 370 700 Picea abies
Ferris et al., 2001 360 550 Populus x euramericana (hybrid P.
deltoides x P. nigra, klon I-214)
Populus nigra (Jean Pourtet)
Populus alba (2AS-11)

17
Gibeaut et al., 2000 360 1000 Arabidopsis thaliana (Landsberg
erecta)
Griffin et al., 2001 370d/450n 570d/650n Liquidambar styraciflua
Griffin et al., 2001 250 1000 Glycine max
Griffin et al., 2001 360 730 Abutilon theophrasti
Griffin et al., 2001 360 830±52 Piper auritum
Epipremnum pinnatum
Griffin et al., 2001 360 560 Pinus taeda
Pinus radiata
Acer rubrum
Cercis canadensis
Liquidambar styraciflua
Kinsman et al., 1997 400±1,5 699±2,6 Dactylis glomerata L.
Leadley et al., 1987 348 645 Glycine max L. Merr. ‘Bragg’
Lin et al., 2001 350 750 Pinus sylvestris L.
Luomala et al., 2005 362±43 693±30 Pinus sylvestris L.
Norby a O’Neill, 1991 371±17 493±10 787±30
Liriodendron tulipifera L.
Oksanen et al., 2001 360 560d* Populus tremuloides Michx.
Oksanen et al., 2005
1. rok 365
2. rok 371
3. rok 373
1. rok 651
2. rok 720
3. rok 729
Betula pendula Roth (klon 4 a klon 80)
Olszyk et al., 2005 ? ?+180 Pseudotsuga menziesii (Mirb.) Franco
Onoda et al., 2007 370 530 Polygonum sachalinense
Sasa kurilensis
Tiarella polyphylla
Onoda et al., 2007 370 760 Hydrangea paniculata
Phragmites australis
Plantago asiatica
Onoda et al., 2007 375 670 Hydrangea paniculata
Polygonum sachalinense
Sasa kurilensis
Pritchard et al., 1997 365 720 Pinus palustris
Ranasinghe a Taylor, 1996 330–350 620–640 Phaseolus vulgaris L. cv. Tender Green

18
Sholtis et al., 2004 364 553 Liquidambar styraciflua
Taylor et al., 2003 370 550 Populus x euramericana (hybrid P. deltoides x P. nigra, klon I-214)
Teng et al., 2006 370 700 Arabidopsis thaliana L. (Col-0)
Thomas a Bazzaz, 1996 350 700 Taraxacum officinale Weber
Plantago major
Rumex crispus
Thomas a Harvey, 1983 340 520;718;910 Zea mays L. ‘Golden Bantam’
Glycine max L. Merril ‘Ransom’
Pinus taeda L.
Liquidambar styraciflua L.
Tissue et al., 1996 366±2 516±2; 666±3
Pinus taeda L.
Při studiu působení zvýšené [CO2] na rostliny lze vyžít i jiných metod, které nejsou
založeny na simulaci atmosféry se zvýšenou koncentrací CO2, např. metod
dendrochronologie, která se zabývá studiem letokruhů dřevin (Ceulemans a Mousseau, 1994;
Nátr, 2000), nebo paleobotaniky, například pro studium hustoty průduchů na fragmentech
pokožky fosilních listů (Beerling a Chaloner, 1993).
1.2.3 Vliv zvýšeného obsahu CO2 v atmosféře na rostliny
Oxid uhličitý, jakožto substrát pro fotosyntézu, je pro rostliny nepostradatelný. Nárůst
jeho koncentrace v atmosféře je však v poslední době velmi rychlý, pravděpodobně zejména
vlivem rozvoje průmyslu a dopravy (a z toho vyplývajícího zvyšování emisí CO2) ve světě,
takže nastává otázka, zda se rostliny dokáží tomuto tempu nárůstu přizpůsobit (Nátr, 2000).
V této kapitole jsou zmíněny jen některé aspekty vlivu zvýšené koncentrace CO2 v atmosféře
– zkráceně EC – na rostliny, které jsou podrobněji popsány v dalších kapitolách spolu
s dalšími změnami, které nejsou v této kapitole uvedeny.
Zvýšená [CO2] způsobuje změny mnoha fyziologických procesů v rostlinách, které
spolu různým způsobem interagují, čímž komplikují porozumění vlivům EC na rostliny, které
jsou vzájemně složitě propletené. Například v důsledku působení zvýšené koncentrace CO2 se
snižuje vodivost průduchů, a tím se zvyšuje účinnost využití vody (Nátr, 1998). Rostliny
mohou díky přivření průduchů a následné větší účinnosti využití vody čerpat méně vody

19
z půdy, což má za následek, že v půdě zůstává více vody. Větší množství vody v půdě
umožňuje rostlinám lépe přijímat živiny (Körner, 2003).
U rostlin pěstovaných při EC se zvyšuje obsah sacharidů, čímž klesá relativní obsah
bílkovin. To vede ke změně kvality rostlinné potravy pro herbivory, čímž může být ovlivněno
jejich přežívání a množení a tím druhové složení ekosystémů. Rozdíl v reakci rychlosti čisté
fotosyntézy na zvýšenou [CO2] rostlin C3 a C4 také může mít vliv na druhové složení
ekosystémů, protože budou zvýhodněny druhy rostlin C3, které nejsou nynější koncentrací
CO2 saturovány (Nátr, 1998); viz obr. 1.6.
Obr. 1.6: Graf závislosti rychlosti čisté fotosyntézy na koncentraci CO2 v atmosféře – porovnání rostlin C3 (pšenice – wheat) a C4 (kukuřice – maize). Převzato z <http://www.ciesin.columbia.edu/docs/004-038/fig4-1.gif>.
V důsledku přímých vlivů EC na rostliny (např. změn v rychlosti fotosyntézy,
fotorespirace a dýchání, dále změn ve vodivosti průduchů a v účinnosti využití vody) dochází
k vlivům nepřímým, například ke zvýšenému vytváření asimilátů, ke změně ukládání a
složení sušiny, ke změně relativního obsahu uhlíku k dusíku, a z toho vyplývající změně
kvality biomasy pro herbivory. Zvýšená [CO2] dále ovlivňuje výnos, rychlost vývoje, vodní
provoz, růst a vývoj, migraci, biodiverzitu a interakci rostlin s parazity. Působení CO2 je
široké a může být umocněno interakcemi s dalšími faktory, například s teplotou a dostupností
živin a vody (Nátr, 2000).
Ukázalo se, že EC působí na různé druhy rostlin ve stejných experimentálních
podmínkách odlišně. Např. u Betula platyphylla nastalo vlivem EC snížení rychlosti
fotosyntézy nezávisle na tom, zda byly rostliny pěstovány v půdě s malým nebo velkým
obsahem živin, kdežto u Betula Maximowicziana došlo ke snížení rychlosti fotosyntézy v EC
pouze u rostlin pěstovaných v půdě s malým množstvím živin (Eguchi et al., 2008). Dalším
příkladem je odlišná reakce rychlosti růstu tří druhů topolů (viz kapitola 2.1.2) (Ferris et al.,
2001).

20
K nepřímým vlivům na rostliny náleží také změny v anatomii rostlin. V mnoha
studiích bylo zjištěno, že růst rostlin při zvýšené koncentraci oxidu uhličitého v okolním
vzduchu ovlivňuje stavbu rostlinných orgánů včetně listů. Změny ve stavbě listů se projevily
na úrovni makroskopické, mikroskopické, na úrovni samotných buněk i v látkovém složení
listů. Pro pochopení celkových vlivů EC na růst a vývoj rostlin, musí být změny v anatomii
listů při zvýšené [CO2] posuzovány v souvislostech se změnami fyziologických procesů
(Leadley et al., 1987).
2. Vliv oxidu uhličitého na strukturu listu
Struktura listu je utvářena a ovlivňována mnoha endogenními i exogenními faktory.
Tvorba listu je dynamický proces zahrnující v sobě celou řadu fyziologických procesů, v
jejichž důsledku se vytváří určitá struktura,, ve které se odehrávají fyziologické procesy
probíhající v listu. Tato kapitola pojednává o vlivu EC na vzájemně spjatou strukturu a funkci
listu, které jsou vzájemně spjaté. První část je věnována změnám procesů v důsledku zvýšené
koncentrace CO2, které mají vliv na morfogenezi listu. Druhá část pojednává o změnách
anatomické struktury listu v důsledku působení zvýšené koncentrace CO2.
2.1 Změny v morfogenezi listu
Morfogeneze je podmíněna buněčným dělením, tj. tvorbou buněk, a jejich růstem. Oba
tyto procesy jsou citlivé na koncentraci CO2, ale velikost jejich změn závisí na druhu rostliny
a na dalších podmínkách prostředí (Taylor et al., 2003).
2.1.1 Změny v zakládání a senescenci listů
U různých druhů rostlin bylo pozorováno, že růst ve zvýšené [CO2] ovlivňuje
fenologii listů různým, až protikladným způsobem, přičemž důvody rozdílů nejsou zatím
zcela známy.
U semenáčků Quercus rubra neměla zvýšená koncentrace CO2 na časovou
posloupnost procesů vzniku a senescence listů žádný vliv (Cavender-Bares et al., 2000).
Naopak na semenáčky Pinus sylvestris L. zvýšená [CO2] vliv měla – jejich pupeny
přezimovaly v pokročilejším vývojovém stádiu, na jaře vyrašily dříve a vyvíjely se rychleji a
na podzim jehlice předčasně opadávaly (Jach a Ceulemans, 1999). U listů Populus nigra,

21
Populus alba a Populus x euramericana došlo při EC k oddálení podzimní senescence
(Tricker et al., 2004).
Obvykle dochází ke zvýšení počtu vytvořených listů, často tedy ke zkrácení
plastochronu, což je doba, která uběhne od založení jednoho listového primordia ve vrcholu
stonku do založení dalšího, po něm bezprostředně následujícího. Šestileté působení EC na
mladé stromky smrku ztepilého Picea abies L. Karst. se projevilo mimo jiné neprůkazným
nárůstem počtu jehlic na délku výhonu (Tomaskova et al., nepublikováno). V jiné práci,
zabývající se topoly, Ferris et al. (2001) zjistili, že Populus x euramericana (I-214), Populus
nigra (Jean Pourtet) a Populus alba (2AS-11) měly v prostředí s vyšší koncentrací oxidu
uhličitého větší počet listů. K podobným zjištěním došli také Tissue et al. (1996), kteří
zaznamenali při EC větší počet rašících výhonů na semenáčcích borovice kadidlové Pinus
taeda L. a Conroy et al., (1986), kteří uvádějí, že semenáčky borovice montereyské Pinus
radiata D. Don měly v EC větší počet jehlic. Kinsman et al. (1997) se narozdíl od výše
uvedených autorů zabývali ve své práci bylinami (srhou říznačkou, Dactylis glomerata L.) a
naznačují příčiny zvýšeného počtu listů a jejich rychlejší tvorby na rostlinách ve zvýšené
[CO2]: V apikálním dómu stonkového meristému D. glomerata vzrostl počet buněk s kratším
trváním buněčného cyklu a díky tomu poklesl plastochron.
Zkrácení doby trvání buněčného cyklu může vést k vytváření většího počtu buněk a
k větší rychlosti růstu pletiv a tím i ke změnám strukturních parametrů a rychlosti tvorby
orgánů. Důvodem zkrácení doby trvání buněčného cyklu v EC je pravděpodobně zvýšená
rychlost fotosyntézy a účinnější fixace CO2 a s nimi spojená zvýšená tvorba sacharidů, které
mohou působit jako signální molekuly a ovlivňovat tak průběh buněčného cyklu. Zvýšená
[CO2] vede ke zvýšení rychlosti fotosyntézy, která způsobí zvýšenou tvorbu asimilátů, a tudíž
stoupá i koncentrace sacharózy, která funguje jako signální molekula (Kinsman et al., 1997).
Sacharóza zvyšuje expresi genu pro cyklin D (CYCD3;1) a přítomnost sacharózy je nezbytná
pro udržení hladiny tohoto proteinu a tím i pro přechod z G1 do S fáze. Pokud je při přechodu
z G1 do S fáze nízká koncentrace cukrů, k přechodu do S fáze nedojde (Smith a Stitt, 2007).
2.1.2 Změny v relativní rychlosti růstu listů
S urychlením dělení buněk souvisí také rychlost růstu, která je ve zvýšené koncentraci
CO2 zpravidla vyšší. Nepodařilo se mi nalézt práci, která by se zabývala relativní rychlostí
růstu (RGR) u listu jehličnanů. Jediné práce o jehličnanech pojednávají o relativní rychlosti
přírůstku výšky hlavního výhonu borovice lesní. U semenáčků borovice lesní P. sylvestris, na

22
začátku experimentu starých 3 roky, byl v prvním roce pěstování nárůst výšky hlavního
výhonu ve zvýšené [CO2] větší, ale ve druhém roce již byl prakticky shodný s relativní
rychlostí růstu výšky v normální [CO2]. Z toho vyplývá, že došlo ke snížení míry působení
EC na růst v čase (Jach a Ceulemans, 1999). Podobné závěry plynou i ze zkoumání relativní
rychlosti růstu (RGR), tedy rychlosti přírůstku biomasy, rostlin semenáčků borovice
kadidlové P. taeda. RGR byla v prvních 11 měsících působení zvýšené [CO2] vyšší u rostlin
ve zvýšené koncentraci CO2, ale po delší době byla srovnatelná s RGR v normální
koncentraci CO2 (Tissue et al., 2004).
Listy opadavých dřevin přetrvávají pouze jednu vegetační sezónu, během níž nastává
jejich morfogeneze i senescence, kteréžto procesy jsou citlivé k působení zvýšené
koncentrace CO2. Vlivem EC nastávají změny v RGR listů listnatých dřevin obvykle vedoucí
ke zvýšené expanzi listů. Ferris et al. (2001) se zabývali vlivem [CO2] na rychlost zvětšování
listové plochy semenáčků tří druhů topolů. Zjistili druhové rozdíly v rychlosti expanze listové
plochy i ve způsobu odpovědi na EC. Rychlost byla u P. x euramericana nezávisle na [CO2]
v daných podmínkách zhruba dvakrát vyšší než u P. nigra a P. alba. Ukázalo se, že u různých
druhů existují rozdílné mechanismy reakce na EC: V listech P. x euramericana a P. nigra
bylo zvětšení listů způsobené produkcí více a větších buněk, zatímco v listech P. alba tvorbou
více malých buněk. Taylor et al. (2003) zkoumali zhruba o rok později na stejném
experimentálním stanovišti expanzi listů přibližně rok a čtvrt starých jedinců P. x
euramericana (hybrid P. deltoides x P. nigra, klon I-214). Jejich výsledky byly ve shodě s
výsledky Ferrise et al. (2001) – zvýšená [CO2] urychlila expanzi listů. Počet buněk
vytvořených za den v obou prostředích narostl na počátku vývoje listů, v dalších fázích
vývoje klesal při AC dříve a rychleji než při EC. Ve zvýšené [CO2] se přechodně a v celém
listu rovnoměrně zvýšila expanze epidermálních buněk, v pozdějších stádiích vývoje listu se
velikost buněk mezi prostředími nelišila. Ani v jednom prostředí nebyly pozorovány zjevné
rozdíly ve velikosti epidermálních buněk na různých místech čepele, tudíž nerovnoměrná
expanze listů v obou koncentracích CO2 byla pravděpodobně způsobena různou rychlostí
dělení buněk v různých oblastech listu. To se potvrdilo při porovnání rychlosti tvorby buněk
mezi sedmi segmenty listu vymezenými sekundárními žilkami od báze listu ke špičce. Byl
pozorován zřetelný gradient rychlosti tvorby buněk od báze ke špičce listu. U báze byla
rychlost tvorby buněk nejvyšší, což vedlo k tomu, že tam byla rychlost expanze listové plochy
P. x euramericana výrazně vyšší než u zbylých částí listu (Taylor et al., 2003). Ferris et al.
(2001) došli k podobným výsledkům když u tří druhů topolů porovnávali relativní nárůst
plochy listu mezi pěti segmenty listů, taktéž vymezenými sekundárními žilkami. Kromě

23
mezidruhových rozdílů se zde projevil i vliv koncentrace CO2: Rychlost růstu při EC byla
průkazně vyšší v segmentech na bázích listů všech tří druhů, v ostatních segmentech měla
tentýž trend, ale výsledky nebyly statisticky průkazné. Taylor et al. (2003) dále uvádějí, že
toto prostorové rozložení rychlosti tvorby buněk bylo patrné při obou koncentracích CO2. Ve
zvýšené [CO2] byla rychlost tvorby buněk ve všech segmentech větší, nejvíce se mezi
koncentracemi lišil segment č. 2 u báze listu, ve kterém se při EC vytvářelo zhruba dvakrát
tolik buněk než při AC.
Obdobné důsledky působení zvýšené [CO2] na RGR jsou známy i u bylin. Relativní
rychlost růstu listů (RGR), tedy rychlost přírůstku biomasy, byla v počátečních třech týdnech
vývoje listů huseníčku rolního Arabidopsis thaliana L. vyšší u rostlin ve zvýšené [CO2],
později měla přibližně stejnou hodnotu jako RGR v AC. Autoři dávají RGR listů do
souvislosti s aktivitou enzymu UDP-Glc dehydrogenázy, která měla podobný průběh, tj. byla
nejvyšší na začátku vývoje rostlin a postupně klesala, přičemž zhruba po třech týdnech začala
klesat strměji. RGR i aktivita UDP-Glc dehydrogenázy měly v AC podobný průběh jako v
EC, ale jejich hodnoty byly nižší. Regulace aktivity UDP-Glc dehydrogenázy by mohla být
klíčová pro syntézu xyloglukanů a dalších polysacharidů z matrixu buněčné stěny se vztahem
k jejímu růstu. (Gibeaut et al., 2000). Díky růstu buněčné stěny je umožněno zvětšování
buněk. To znamená, že zvýšená aktivita UDP-Glc dehydrogenázy pravděpodobně umožňuje
větší rychlost růstu listů díky zvětšování buněčné stěny a tím objemu buněk. Domnívám se, že
by aktivita UDP-Glc dehydrogenázy mohla být zvýšena díky většímu množství sacharidů,
kterých je v EC více (viz kapitola 2.2.2.2).
2.1.3 Změny ve zvětšování a tvorbě buněčné stěny
Buněčné stěny ohraničují buňky a částečně je omezují ve zvětšování. Rychlost
zvětšování buňky je ovlivněna turgorem tlačícím na stěnu zevnitř buňky a extensibilitou
buněčné stěny. Extensibilita (roztažitelnost) buněčné stěny je důležitá, protože díky
rozvolnění stěny snižují buňky svůj vodní potenciál, a to vede k příjmu vody protoplastem a
ke zvýšení turgoru, čímž je umožněno zvětšení buňky. Elasticita buněčných stěn je vratná
extensibilita, plasticita je nevratná extensibilita. Extensibilitu ovlivňují hormony – auxiny a
gibereliny ji zvyšují a kyselina abscisová a etylén ji snižují (Evert, 2006). Podle hypotézy
kyselého růstu auxiny aktivují ATPázy, které pumpují H+ z cytosolu do buněčné stěny a tím
se v ní sníží pH, což vede k rozvolnění stěny a následnému zvýšení turgoru. Kyselý růst
zprostředkovávají proteiny expanziny, které rozpojují nekovalentní spojení mezi

24
polysacharidy, čímž je umožněno roztažení buněčné stěny, aniž by byla poškozena.
Sekundárními rozvolňujícími faktory jsou XET (xyloglukan endotransglykosylázy),
pektinázy, endoglukanázy a hydroxylové radikály. XET rozpojují a spojují xyloglukanové
řetězce, přičemž nenarušují strukturu buněčné stěny (Evert, 2006).
Ve zvýšené [CO2] byla v rostlinách A. thaliana zjištěna vyšší aktivita UDP-Glc
dehydrogenázy, která je klíčovým enzymem působícím v dráze vedoucí k biosyntéze
mnohých polysacharidů buněčné stěny, např. xyloglukanu. Zvýšená aktivita UDP-Glc
dehydrogenázy byla zaznamenána zejména v počátečním období vývoje rostlin A. thaliana
v EC, kdy byla vyšší také relativní rychlost růstu a kdy byly nižší hodnoty poměru plochy
listu k hmotnosti buněčné stěny (Gibeaut et al., 2000). Buněčné stěny buněk listů A. thaliana
byly při zvýšené koncentraci CO2 výrazně tlustší a vzrostla rovněž koncentrace celulózy
vztažená na hmotnost sušiny listu a obsah rozpustných sacharidů (Teng et al., 2006). Zároveň
byla pozorována vyšší koncentrace auxinu a kyseliny giberelové a nižší koncentrace kyseliny
abscisové (Teng et al., 2006). To mohlo zvýšit extensibilitu buněčných stěn, protože
gibereliny ji zvyšují a kyselina abscisová ji snižuje (viz výše). Zvýšená extensibilita umožňuje
rychlejší zvětšování buněk. U tří druhů topolu (P. x euramericana, P. nigra a P. alba) byla při
zvýšené [CO2] pozorována vyšší extensibilita buněčných stěn. Výsledky stanovení
prostorového rozložení extensibility buněčných stěn na různých místech listové čepele rostlin
fazolu obecného Phaseolus vulgaris L. při zvýšené [CO2] ve fázi zvětšování buněk ukázaly,
že se extensibilita buněčných stěn zvýšila zejména v oblasti spodních okrajů listu, což by
mohlo způsobit změnu tvaru listu (Ranasinghe a Taylor, 1996). Tyto výsledky jsou ve shodě
s výsledky Ferrise et al. (2001), kteří uvádějí, že se segmenty listu topolu blíže k bázi listu
zvětšovaly rychleji než listové segmenty v ostatních částech listu (viz kapitola 2.1.2).
V mladých rostoucích listech topolů byla aktivita XET při EC vyšší než při normální
koncentraci (Ferris et al., 2001), následkem čehož vzrostla plasticita buněčných stěn.
V listech P. vulgaris aktivita XET při EC vzrostla, ale jen pokud byla koncentrace CO2
zvýšena pouze ve fázi zvětšování buněk a nikoli ve fázi dělení buněk. V ostatních
kombinacích působení zvýšené [CO2] aktivita XET poklesla (Ranasinghe a Taylor, 1996), což
opět ukazuje na druhovou specificitu odpovědi na působení EC. Při vzájemném porovnání
druhů topolu byla celková plasticita největší u P. alba, dále u P. nigra a nejmenší byla u P. x
euramericana. Výsledky naznačují, že některé účinky a mechanismy působení zvýšené [CO2]
jsou druhové specifické i v rámci jednoho rodu. Aktivita XET také velmi pravděpodobně
ovlivňovala rychlost růstu – např. u P. alba odpovídala nižší aktivita XET pomalejší rychlosti

25
růstu. Elasticita buněčných stěn se narozdíl od plasticity nelišila mezi třemi druhy topolu
navzájem, ani u jednotlivých druhů mezi prostředími s různou koncentrací CO2 (Ferris et al.,
2001). Při studiu vlivu EC na vlastností buněčných stěn se ukazuje, že i zde jsou zvýšenou
[CO2] více ovlivněny ontogeneticky mladší listy a jedinci.
2.1.4 Změny v morfologických parametrech listu
Se změnami rychlosti růstu některých úseků čepele listů nebo celých listů souvisejí
změny rozměrů a tvarů listů, které jsou patrně závislé zejména na genotypu rostlin a na
uspořádání experimentů – např. na různém stáří zkoumaných rostlin a rozdílné délce trvání
působení EC. Často dochází k zvětšení některých rozměrů a biomasy listů.
Větší tloušťku jehlic přičítanou nárůstu mezofylu uvádějí Thomas a Harvey (1983).
Zjistili, že při působení vyšší [CO2] = 910 ppm po dobu 45 dní byly jehlice ročních
semenáčků P. taeda L. díky nárůstu mezofylu tlustší než jehlice v normální koncentraci CO2;
nutno poznamenat, že se tloušťka v ostatních hodnotách EC (viz Tab. 1.1) průkazně nelišila
od tloušťky v AC. Lin et al. (2001) zjistili, že se u jehlic sedmiletých stromů P. sylvestris,
pěstovaných první tři roky v květináčích a poté přesazených do OTC a čtyři roky rostoucích
v EC nebo v AC, zvětšil počet i rozměr mezofylových buněk v EC, v důsledku čehož se
zvětšila poměrná plocha, kterou na ploše příčného řezu zabíral mezofyl. Vzrostla též plocha
celého příčného řezu a jehlice měly v EC prokazatelně větší tloušťku a trend k větší šířce.
Výsledky studií ukazují, že důsledky působení zvýšené [CO2] záleží na stáří rostlin – jsou
ovlivněny zejména ontogeneticky mladší rostliny. To se potvrdilo v případě srovnání
výsledků Lin et al. (2001) a Luomala et al. (2005), protože ve druhé z těchto studií nebyly
zaznamenány průkazné rozdíly v tloušťce, délce a projekční ploše jehlic přibližně 20 let
starých stromů P. sylvestris L., pěstovaných po tři roky v komorách při zvýšené [CO2].
Účinky EC na tloušťku listu byly též zaznamenány u bylin. Při zvýšené [CO2] byly
tlustší listy A. thaliana (Teng et al., 2006), dvou kultivarů sóji Glycine max L. – (Leadley et
al., 1987; Thomas a Harvey, 1983)– a dřeviny ambroně Liquidambar styraciflua L. (Thomas
a Harvey, 1983), zatímco se tloušťka listů nezměnila u mladých stromů P. x euramericana
(Taylor et al., 2003) a u kukuřice Zea mays L. (Thomas a Harvey, 1983). S komplikovaným
objasňováním působení EC na rozměry listu souvisí i to, že výsledky některých prací ukazují
opačný trend – menší tloušťku listu při EC. Například u listů na začátku pokusu osmiletých
stromů břízy bělokoré Betula pendula L., pěstovaných po tři roky v OTC, poklesla tloušťka

26
listů (Oksanen et al., 2005) a rovněž listy smetánky lékařské Taraxacum officinale (L.) Web.
byly po čtyřech měsících růstu v EC tenčí oproti listům v AC (Thomas a Bazzaz, 1996). Listy
T. officinale ve zvýšené [CO2] navíc změnily svůj tvar – vykazovaly větší poměr obvodu k
ploše, byly tedy více vykrojené (Thomas a Bazzaz, 1996).
Ke změně tvaru došlo i u listu topolu P. x euramericana, který se v EC rozšířil a u nějž
se zvětšila jeho plocha. Výsledky ukázaly, že rychlost a časová posloupnost procesů tvorby
buněk byly pro určení tvaru listu důležitější než expanze buněk, protože ta byla v celé čepeli
stejná (viz kapitola 2.1.2) (Taylor et al., 2003). Ferris et al. (2001) zaznamenali nárůst délky a
konečné velikosti listů v prvním roce růstu mladých stromů tří druhů topolu (P. x
euramericana, P. nigra a P. alba) v EC; přisuzují ho vyšší produkci biomasy při zvýšené
[CO2].
Závěry ohledně změn rozměrů listů v EC nejsou ve všech studiích konzistentní.
Například jehlice dvouletých semenáčků modřínu japonského Larix kaempferi Carr.
pěstovaných 160 dní při zvýšené [CO2] měly přibližně stejnou tloušťku jako jehlice v AC
(Eguchi et al., 2004). Podobně se nezměnila šířka, délka ani plocha jehlic po tříletém
působení zvýšené [CO2] na původně dvouleté semenáčky douglasky tisolisté Pseudotsuga
menziesii Mirb. Franco (Apple et al., 2000), stejně jako plocha příčného řezu jehlicí a jeho
obvod u semenáčků téhož druhu pěstovaných po čtyři roky v EC (Olszyk et al., 2005).
Při studiu vlivu EC na rostliny byly zjištěny rozdíly v jeho působení v závislosti
na délce vystavení zvýšené koncentraci CO2. Například v krátkodobých pokusech u
semenáčků P. radiata, na začátku experimentu starých osm týdnů, pěstovaných po dobu 22
týdnů v EC, byla zjištěna větší délka a průměr příčného řezu jehlic, přičemž rozdíl v průměru
jehlic byl způsoben spíše větší velikostí buněk než jejich větším počtem (Conroy et al., 1986).
Dlouhodobější pokusy Pritcharda et al. (1998) ukázaly, že po 12 měsících působení EC na
semenáčky borovice bahenní Pinus palustris Mill. byly jejich jehlice tlustší, měly větší objem
cévních svazků a větší plochu příčného řezu než v AC. I proporce jednotlivých pletiv na řezu
se změnily – transfúzní pletiva zaujímala průkazně větší část plochy a stejnou tendenci měl
též mezofyl a vodivá pletiva. Avšak po 20 měsících v EC se anatomické parametry jehlic
průkazně nelišily od AC, jehlice jehlice pouze měly neprůkazně menší objem cévních svazků.
Tyto výsledky potvrzují domněnku, že velikost dospělých jehlic je nakonec obdobná v obou
koncentracích CO2 a naznačují fotosyntetickou aklimaci (Pritchard et al., 1998).
2.1.5 Změny v tvorbě sušiny a asimilátů

27
Rozmanitý vliv působení EC na vlastnosti listů se projevuje také při studiu vlivu EC
na tvorbu sušiny a asimilátů, která ovlivňuje hmotnost listů. Pro svou lepší vypovídací
hodnotu bývá často místo samotné hmotnosti sušiny používána hodnota specifické hmotnosti
listu (s anglickými zkratkami SLW, LMA a SLM), která vyjadřuje poměr hmotnosti sušiny
listu k ploše listu; v některých případech se uvádí její převrácená hodnota – specifická listová
plocha (SLA), tj. poměr plochy k hmotnosti sušiny listu.
Ani v tomto případě nejsou výsledky studií jednotné a převládají zde spíše malé
změny těchto parametrů. Například nebyla ovlivněna SLA jehlic L. kaempferi (Eguchi et al.,
2004), ani SLW a hmotnost jehlic P. menziesii nebyly průkazně větší (Apple et al., 2000), a
stejně tak ani čerstvá hmotnost a hmotnost sušiny jehlic 20 let starých stromů P. sylvestris
(Luomala et al., 2005). K průkazným změnám vlivem EC došlo pouze u jehlic semenáčků P.
menziesii, které měly po čtyřech letech pěstování v EC větší SLW (Olszyk et al., 2005).
Podobně jako jehlice jehličnanů reagují na EC i listy listnatých dřevin. SLA a/nebo
SLW se vlivem EC průkazně nezměnily u listů tří druhů topolu (Ferris et al., 2001), jitrocele
většího Plantago major L. a šťovíku kadeřavého Rumex crispus L. (Thomas a Bazzaz, 1996).
Naproti tomu se SLA snížilo vlivem EC u listů B. pendula (Oksanen et al., 2005), A. thaliana
(Gibeaut et al., 2000) a Taraxacum officinale (Thomas a Bazzaz, 1996). Pokles SLA bývá
vysvětlován nárůstem tloušťky listu, ale mohou existovat i jiná vysvětlení. Např. Oksanen et
al. (2005) přičítají nižší SLA mírnému nárůstu tloušťky buněčných stěn mezofylových buněk
a nárůstu velikosti škrobových zrn a počtu mitochondrií.
Onoda et al. (2007) srovnávali vlastnosti listů dvouděložných (D) i jednoděložných (J)
rostlin, po desetiletí rostoucích na třech stanovištích poblíž přírodních zdrojů CO2, s
rostlinami stejných druhů rostoucími nedaleko, ale v AC. Změny v SLW nebyly mezi druhy
jednotné: Polygonum sachalinense (D) a Plantago asiatica (D) měly při EC vyšší SLW,
zatímco Sasa kurilensis (J) měla nižší hodnotu SLW a na SLW listů Hydrangea paniculata
(D), Tiarella polyphylla (J), Phragmites australis (J) neměla zvýšená [CO2] vliv. Autorům se
nepodařilo potvrdit domněnky o rozdílné reakci listů jedno- a dvouděložných rostlin. Spíše
lze očekávat rozdíly způsobené zvýšenou koncentrací CO2 mezi rostlinami C3 a C4, u nichž
je rozdílná závislost rychlosti čisté fotosyntézy na koncentraci CO2 v atmosféře (obr. 1.6).
Z výše uvedených údajů je zřejmé, že interpretace důsledků působení EC je
komplikovaná, protože spektrum procesů, které zvýšená koncentrace oxidu uhličitého
ovlivňuje, je velké a záleží na mnoha podmínkách.

28
2.2 Mikroskopické změny struktury listu
2.2.1 Krycí pletiva
Z výsledků studií vyplývá, že u různých druhů rostlin reagují krycí pletiva listu na EC
rozdílně. V dostupné literatuře však nebyly u jehličnanů změny ve strukturálních parametrech
krycích pletiv studovány.
Vlivem zvýšené koncentrace CO2 se na příčném řezu nezměnila plocha epidermis
jehlic P. palustris (Pritchard et al., 1998), ani plocha epidermis a hypodermis a jejich tloušťka
měřená souhrnně (Lin et al., 2001) i odděleně (Luomala et al., 2005) v jehlicích P. sylvestris,
a též tloušťka epidermální vrstvy jehlic P. taeda nepodlehla změnám (Thomas a Harvey,
1983). Stejně tak u bylin k změně tloušťky epidermis nedochází, jako například u listů Z.
mays a G. max (Thomas a Harvey, 1983). Změny v počtu a velikosti epidermálních buněk
vlivem EC byly studovány na bylinách i listnatých dřevinách. Ve velikosti epidermálních
buněk na paradermálním řezu listem pšenice Triticum aestivum L. nebyly zaznamenány
změny (Masle, 2000), zatímco studie na topolech (Ferris et al., 2001) ukázala vliv EC i
mezidruhových rozdílů: U P. x euramericana a P. nigra se na obou stranách listu kromě počtu
zvětšila i velikost epidermálních buněk, kdežto u P. alba se vlivem EC zvýšil počet buněk
pouze na abaxiální straně listu a navíc bez výrazných změn jejich velikosti.
Při studiu epidermis v EC bylo zkoumáno i množství a struktura epikutikulárních
vosků a zase s ukázaly mezidruhové rozdíly působení EC. Struktura epikutikulárních vosků se
u jehlic P. menziesii (Apple et al., 2000) a P. sylvestris (Lin et al., 2001) výrazně nezměnila.
Ukládání epikutikulárních vosků na listech kukuřice nebylo působením EC dotčeno, narozdíl
od sóji a dřeviny L. styraciflua, u kterých se při hodnotě [CO2] = 910 ppm na abaxiální straně
listu ukládalo více epikutikulárních vosků (Thomas a Harvey, 1983).
2.2.1.1 Průduchy
Množství průduchů a jejich vlastnosti ovlivňují rychlost fotosyntetické fixace CO2,
proto lze předpokládat, že budou významně ovlivněny změnami koncentrace CO2.
Bylo zjištěno, že se hustota průduchů, tedy jejich počet na jednotku plochy, mění
v závislosti na koncentraci CO2 – při vzrůstající [CO2] se snižuje a naopak. Například hustota
průduchů, naměřená na fosilních nálezech vrby Salix herbacea, ukazuje, že se v posledních
140 tisících letech se zvyšující se [CO2] hustota průduchů snižovala (Beerling a Chaloner,

29
1993). Tohoto vztahu mezi hustotou průduchů a [CO2] v atmosféře se využívá pro
rekonstrukce historických hodnot [CO2]. Ukázalo se, že fosilie jehličnanů jsou velmi vhodné
pro rekonstrukci historických hodnot [CO2], a to díky způsobu vývoje jejich jehlic. Při
dospívání jehlic totiž dochází hlavně k zvětšování velikosti buněk namísto tvorby nových
buněk, a proto se nemění parametry hustoty ani frekvence průduchů, tj. počtu průduchů
vztažených k délce pozorované části jehlice (Kouwenberg et al., 2004).
Autoři několika studií zabývajících se ovlivněním parametrů průduchů zvýšenou
[CO2] u několika druhů jehličnanů dospěli k podobným závěrům, a sice, že v těchto
parametrech téměř nenastaly změny. Nebyla ovlivněna hustota, vodivost, ani rychlost
transpirace průduchů semenáčků P. menziesii (Apple et al., 2000), ani počet řad průduchů a
hustota průduchů na jehlicích semenáčků P. palustris (Pritchard et al., 1998), ani frekvence
průduchů u P. abies (Fendrych, 2005) a konečně ani hustota průduchů P. sylvestris (Luomala
et al., 2005). Jedině na jehlicích P. sylvestris ve studii Lin et al. (2001) při zachování počtu
řad poklesla hustota průduchů, dle autorů pravděpodobně kvůli nárůstu expanze
epidermálních buněk; což by odpovídalo výše zmíněným údajům o vlivu EC na rozvolňování
buněčné stěny a na expanzi buněk (viz kapitola 2.1.3).
Studie na bylinách přinesly pestřejší výsledky. Zatímco Ranasinghe a Taylor (1996)
nezaznamenali u P. vulgaris rozdíly v hustotě průduchů ani ve stomatálním indexu listů, tj. v
poměrném zastoupení stomatálních buněk v celkovém počtu stomatálních a epidermálních
buněk, u Glycine max se stomatální index zvýšil společně s frekvencí průduchů, s výjimkou
listů [CO2] = 718 ppm, kdy naopak hodnoty obou parametrů poklesly (Thomas a Harvey,
1983). Pokles hodnoty stomatálního indexu a hustoty průduchů u A. thaliana vyústil v pokles
vodivosti průduchů a rychlosti transpirace (Teng et al., 2006). Snížení vodivosti průduchů při
zvýšené [CO2] bylo také zjištěno u křídlatky sachalinské P. sachalinense a jitrocele asijského
P. asiatica (Onoda et al., 2007) a též u téměř všech 17 zkoumaných druhů rostlin, převážně
bylin, rostoucích po mnoho generací na stanovišti s přirozeným výronem CO2. Pokles
vodivosti u nich nebyl doprovázen téměř žádnými změnami ve stomatálním indexu a hustotě
průduchů, pouze u dvou ze 17 zkoumaných druhů došlo s poklesem vodivosti zároveň
k poklesu hustoty průduchů, takže se neprokázala hypotéza o přizpůsobení se dlouhodobému
působení EC změnou počtu průduchů (Bettarini et al., 1998). Naznačení opačného vlivu EC
na vodivost průduchů zaznamenali Norby a O’Neill (1991): U listů semenáčků liliovníku
tulipánokvětého Liriodendron tulipifera L. měla vodivost tendenci se zvyšující se koncentrací
CO2 stoupat. Autoři tuto rozdílnou změnu vodivosti průduchů přičítají tomu, že mohla být
ovlivněna nárůstem účinnosti využití vody.

30
2.2.2 Mezofyl
Mezofylové buňky zprostředkovávají výměnu plynů, které vnikly do listu z vnějšího
prostředí skrze průduchy. Příjem a výdej plynů do intercelulár závisí na struktuře mezofylu,
která je ovlivněna genotypem rostlin a vlastnostmi vnějšího prostředí.
V listech bývá při zvýšené koncentraci CO2 často naměřena větší tloušťka mezofylu.
Děje se tomu tak často i u jehlic, například u P. taeda při [CO2] = 910 ppm byla tloušťka
mezofylu vlivem EC větší (Thomas a Harvey, 1983) a u jehlic P. sylvestris byla
nezanedbatelně větší plocha mezofylu na řezu jehlice a zvětšily se také mezofylové buňky i
jejich počet (Lin et al., 2001). Výsledky studií ovlivnění tloušťky mezofylu zvýšenou [CO2]
však nejsou jednotné. V tohoročních jehlicích P. sylvestris byla vrstva mezofylu na abaxiální
straně signifikantně tenčí při EC, ale nebyla celkově průkazně ovlivněna plocha intercelulár
na příčném řezu jehlicemi (Luomala et al., 2005). Podobně zůstala beze změn i plocha
příčného řezu jehlic semenáčků P. menziesii a poměrné zastoupení mezofylu a intercelulár v
něm (Olszyk et al., 2005). Změny ve struktuře vlivem EC závisí i na délce doby jeho
působení. Například u jehlic P. palustris, po 12 měsíců pěstovaných v EC, mezofyl vykazoval
trend k zaujímání větší plochy na příčném řezu než v AC, ale po 20 měsících v EC se již
v tomto parametru nelišily od AC (Pritchard et al., 1998).
Koncentrace CO2 působí jako morfogenní faktor spolu s dalšími faktory prostředí.
Spolupůsobení vlivu EC a množství dostupných živin na stavbu mezofylu zkoumali Eguchi et
al. (2004). Při zvýšené [CO2] a zvýšeném množství živin (dusíku, fosforu a draslíku) bylo
v jehlicích L. kaempferi přítomno více mezofylových buněk, v těchto podmínkách měly
buňky významně menší výšku i průměr. Při sníženém množství živin byl naopak zjištěn
menší počet mezofylových buněk a buňky měly větší průměr a spíše větší výšku. Autoři také
uvádějí, že se u jehlic stromů ošetřovaných různým hnojením lišil poměr vnitřního povrchu
jehlic k ploše jehlic následujícím způsobem: Ve zvýšené [CO2] v kombinaci se sníženým
množstvím živin poklesl a se zvýšeným množstvím živin naopak vzrostl. Tento poměr je
pravděpodobně odpovědný za změny rychlosti fotosyntézy, která poklesla při EC za současné
nízké dostupnosti živin a vzrostla při EC a vysoké dostupnosti živin (Eguchi et al., 2004).
U listnáčů byly pozorovány rozdílné změny ve stavbě mezofylu. U břízy B. pendula
byl zjištěn pokles tloušťky palisádového i houbovitého parenchymu v listech (Oksanen et al.,
2005). Odpověď různých druhů topolů na EC se lišila: Zatímco se plocha, kterou zaujímaly
buňky palisádového parenchymu na příčném řezu listem, zvětšila v listech u tří zkoumaných

31
druhů; plocha, kterou zaujímaly buňky houbovitého parenchymu na příčném řezu listem,
narostla pouze u dvou druhů – P. x euramericana a P. nigra – a u P. alba se naopak snížila
(Ferris et al., 2001). Pouze u jednoho druhu topolu ze tří studovaných (u P. x euramericana)
byl pozorován nárůst velikosti buněk palisádového i houbovitého parenchymu vlivem EC
(Taylor et al., 2003). U A. thaliana se plocha jednotlivých mezofylových buněk na řezu listem
v rozdílných [CO2] výrazně nelišila (Teng et al., 2006).
Leadley et al. (1987) zjistili, že poměr vnitřního povrchu listu k ploše listu u G. max,
poměr objemu celého mezofylu k celkovému objemu listu a ani poměr vnitřního povrchu listu
k objemu celého mezofylu nebyl v EC rozdílný od AC. Při zkoumání vlastností samotného
palisádového parenchymu však přišli na to, že poměr povrchu palisádového parenchymu k
ploše listu, objem palisádového parenchymu vztažený k celkovému objemu listu i poměr
povrchu k objemu palisádového parenchymu samotného při zvýšené [CO2] signifikantně
klesl.
Ve slunných listech Populus tremuloides vzrostla tloušťka mezofylu a také objem
intercelulár pod vlivem zvýšené [CO2] (Oksanen et al., 2001). Je známo, že u slunných listů
bývá jako jedna ze strukturálních adaptací pozorováno zvětšení počtu vrstev palisádového
parenchymu oproti stinným listům. Podobný jev byl pozorován i v důsledku působení
zvýšené [CO2] u listů sóji G. max, u kterých byla tlustší vrstva houbovitého i palisádového
parenchymu, který byl v EC tvořen třemi vrstvami namísto dvěma, jak tomu bylo v AC
(Thomas a Harvey 1983).
2.2.2.1 Chloroplasty
Chloroplasty jsou organely, ve kterých probíhají fotosyntetické reakce. Jelikož je oxid
uhličitý substrátem pro fotosyntézu, lze při změně jeho koncentrace očekávat změny ve
struktuře chloroplastů.
Při zvýšené koncentraci oxidu uhličitého obsahovaly buňky mezofylu A. thaliana větší
počet chloroplastů na buňku. Mechanismus, jakým EC ovlivňuje počet chloroplastů zatím
není znám (Teng et al., 2006). Ve slunných listech P. tremuloides vzrostl podíl plochy
chloroplastů k ploše buněk na řezu listem (Oksanen et al., 2001). U jehlic P. palustris,
odebraných na jaře po 12 měsících vystavení zvýšené koncentraci CO2, bylo pozorováno, že
chloroplasty zaujímají na příčném řezu jehlicí větší plochu a obsahují v důsledku většího
ukládání škrobu více škrobových zrn, která jsou větší než v AC (Pritchard et al., 1997). Ve
shodě s výše uvedenými daty jsou i výsledky měření objemu chloroplastů v pochvách cévních

32
svazků kukuřice, které měly trend být při EC větší (Thomas a Harvey, 1983). Naproti tomu se
u jehlic P. palustris, vystavených EC po dobu 20 měsíců a odebraných na podzim, žádné
rozdíly mezi koncentracemi CO2 neprojevily, pouze v případě, kdy byly pěstovány při
nedostatku dusíku a vody, měly v EC větší škrobová zrna i větší plochu chloroplastů na
příčném řezu. Obdobné skutečnosti, tj. větší plocha chloroplastů na řezu a větší velikost i
počet škrobových zrn, byly zjištěny též u listů bylin, např. A. thaliana (Teng et al., 2006) a
listnatých dřevin, např. B. pendula (Oksanen et al., 2005).
Zvýšená koncentrace CO2 vede k větší rychlosti čisté fotosyntézy (Griffin et al., 2001;
Sholtis et al., 2004), z čehož vyplývá zvýšené ukládání škrobu, které způsobuje změny ve
struktuře chloroplastů (Pritchard et al., 1997, Teng et al., 2006). Zvětšená škrobová zrna
pravděpodobně tlačí na thylakoidní membrány a způsobují tím zprohýbání thylakoidních
membrán; tento jev byl pozorován v chloroplastech listů A. thaliana (Teng et al., 2006) i
v chloroplastech jehlic P. palustris odebraných na jaře po 12 měsících vystavení působení EC
(Pritchard et al., 1997). Někdy může při dlouhodobém působení zvýšené [CO2] docházet k tak
velké akumulaci škrobových zrn v chloroplastu, že se tím naruší struktura thylakoidních
membrán, až dochází k tzv. aklimační depresi, tedy k poklesu rychlosti asimilace (Pritchard et
al., 1997).
Chloroplasty jehlic P. palustris, odebraných na podzim po 20 měsících vystavení
zvýšené [CO2], měly na řezu více plastoglobulů – což jsou kulovité útvary, ve kterých se
hromadí lipidy a které obvykle vznikají v důsledku stresu – než v normální [CO2].
Chloroplasty jehlic P. palustris, odebraných na jaře po 12 měsících vystavení zvýšené [CO2],
měly na řezu méně plastoglobulů než v normální koncentraci CO2, ale tento jev nastal pouze
při pěstování rostlin v půdě s dostatkem dusíku; při nízkém obsahu dusíku v půdě se rozdíly
mezi koncentracemi CO2 neprojevily (Pritchard et al., 1997). U A. thaliana se při EC počet
plastoglobulů v chloroplastu výrazně nelišil od počtu v AC (Teng et al., 2006). Ve slunných
listech topolu P. tremuloides při EC poklesl počet plastoglobulů (Oksanen et al., 2001).
Při zvýšené [CO2] byl zjištěn větší poměr agranálních (stromatálních) thylakoidních
membrán ke granálním v chloroplastech na řezu listů bylin A. thaliana (Teng et al., 2006) a
Piper auritum a dřevin Acer rubrum, L. styraciflua a Cercis canadensis (Griffin et al., 2001) a
u jehlic P. palustris, odebraných po 20 měsících v EC (Pritchard et al., 1997). Tento jev
vysvětlují Griffin et al. (2001) následovně: Granální thylakoidy obsahují zejména fotosystém
II a kyslík uvolňující komplex, zatímco stromatální thylakoidy obsahují zejména fotosystém I,
kde je redukován NADPH potřebný pro redukci CO2 v Calvinově cyklu. Nárůstem podílu

33
stromatálních thylakoidů ke granálním se rostlina pravděpodobně přizpůsobuje EC, protože
toto uspořádání vede k účinnější fixaci CO2 a tvorbě cukerných produktů.
2.2.2.2 Obsah nestrukturních sacharidů v listu
Pod vlivem zvýšené [CO2] obvykle dochází k nárůstu tvorby a akumulace rozpustných
sacharidů a škrobu v listech. Obsah škrobu v jehlicích semenáčků L. kaempferi (Eguchi et al.,
2004) a v listech dvouděložných rostlin A. thaliana (Teng et al., 2006), P. sachalinense, P.
asiatica, H. paniculata a jednoděložné T. polyphylla (Onoda et al., 2007) byl vyšší při jejich
pěstování ve zvýšené koncentraci CO2. V listech dvouděložné rostliny L. styraciflua také
vzrostl v EC obsah nestrukturních sacharidů – tj. škrobu a rozpustných cukrů – na hmotnost
listu; obsah škrobu nejvíce vzrostl na vrcholu vegetační sezóny, kdy byla rychlost čisté
fotosyntézy nejvyšší (Sholtis et al., 2004). Též u A. thaliana byla při zvýšené koncentraci CO2
pozorována vyšší koncentrace nestrukturních sacharidů vztažená na hmotnost sušiny listu
(Teng et al., 2006). Podobná zjištění učinili i Griffin et al. (2001), kteří uvádějí jako
pravděpodobnou příčinu zvýšeného hromadění škrobu nejen zvýšení rychlosti fotosyntézy,
ale i účinnější fixaci CO2 díky změně struktury thylakoidů (viz kapitola 2.2.2.1). Účinnější
asimilace CO2 při EC nastala v listech G. max. Ukázalo se, že zvýšená [CO2] podněcuje
respirační odbourávání sacharidů, jenž poskytuje více energie a biochemických prekurzorů
pro zvětšování a růst listů při EC (Ainsworth et al., 2006). Naproti tomu listy jednoděložné
rostliny S. kurilensis a P. australis obsahovaly při zvýšené i normální koncentraci CO2
podobné množství škrobu na hmotnost sušiny listu, což bylo pravděpodobně způsobeno tím,
že některé jednoděložné rostliny zřejmě ukládají místo škrobu sacharózu (Onoda et al., 2007).
Ve slunných listech P. tremuloides vzrostlo množství škrobu v chloroplastech
(Oksanen et al., 2001). Záleží na době experimentálního působení, protože po delší době
může nastat aklimace a zprvu pozorované rozdíly vymizí. Například jehlice P. palustris
vystavené 20 měsíců EC a odebrané na podzim, obsahovaly méně škrobu než jehlice
odebrané na jaře po 12 měsících (Pritchard et al., 1997).
2.2.3 Vodivá pletiva
Zvýšená [CO2] způsobuje větší rychlost fotosyntézy a tedy i větší produkci asimilátů,
které jsou z míst jejich vytvoření odváděny vodivými pletivy (floémem) do sinků. Proto se dá
očekávat, že struktura cévních svazků bude EC ovlivněna.

34
Objem cévních svazků se pod vlivem zvýšené [CO2] spíše zvětšuje či zůstává
nezměněn. Po 12 měsících působení EC na semenáčky P. palustris měly jejich jehlice větší
objem cévních svazků, po 20 měsících stále jevily podobný trend (Pritchard et al., 1998). U P.
taeda také zaujímala vodivá a transfúzní pletiva v jehlicích větší díl plochy příčného řezu
jehlic pěstovaných v EC (Thomas a Harvey, 1983). U P. palustris po 12 měsících v EC
zaujímala transfúzní pletiva průkazně větší díl plochy na příčném řezu a stejnou tendenci měla
i vodivá pletiva, po 20 měsících v EC již jehlice tyto rozdíly mezi EC a AC nevykazovaly
(Pritchard et al., 1998). Podobně na EC reagovaly i rostliny Glycine max – objem jejich
vodivých pletiv v listech byl při zvýšené [CO2] spíše vyšší, ale rozdíl nebyl průkazný
(Leadley et al., 1987).
Naproti tomu podíl plochy středního válce k ploše příčného řezu jehlicemi semenáčků
P. menziesii nebyl zvýšenou [CO2] ovlivněn (Olszyk et al., 2005). Zatímco se tloušťka
středního válce se při EC v jehlicích 20letých stromů P. sylvestris nezměnila (Luomala et al.
2005), Lin et al. (2001) zaznamenali pokles podílu plochy středního válce v celém příčném
řezu jehlicí P. sylvestris zároveň s poklesem podílu xylému, zatímco podíl floému se zvýšil
(Lin et al., 2001). Pritchard et al. (1997) naopak zjistili, že floém na příčném řezu zaujímal v
jehlicích P. palustris vystavených zvýšené koncentraci CO2 spíše menší relativní plochu, což
bylo důsledkem menšího počtu sítkových buněk, nikoli tím, že by měly menší velikost.
Působení zvýšené [CO2] při svém vlivu na strukturální parametry vodivých pletiv
interaguje s dalšími faktory prostředí. V případě jednoho z nastavení experimentálních
podmínek – při nedostatku dusíku a dostatku vody – byly sítkové buňky v EC zploštělé ve
směru rovnoběžném k rovině buněčných dělení v kambiu oproti buňkám v normální
koncentraci CO2, jinak se žádné rozdíly v rozměrech buněk mezi koncentracemi CO2
neprojevily. Zmenšení velikosti a počtu sítkových buněk by mohlo negativně ovlivnit
schopnost transportovat asimiláty, kterých je v EC produkováno více (Pritchard et al., 1997).
3. Závěr
Zvýšená [CO2] způsobuje změny mnoha fyziologických procesů a struktur, v nichž
tyto procesy probíhají. Z literatury je zřejmé, že zvýšená koncentrace oxidu uhličitého
v atmosféře často způsobuje určité změny ve struktuře listu jehličnanů i listů jiných rostlin,
avšak tyto změny nejsou zcela konzistentní. Jsou druhově specifické a závislé na faktorech
prostředí, které spolu různým způsobem interagují, čímž komplikují porozumění vlivům EC
na rostliny, které jsou vzájemně složitě propojené.

35
Vlivem zvýšené koncentrace dochází ve velké části případů ke zrychlení fotosyntézy a
tím ke zvýšené tvorbě asimilátů, která může být spojena s hromaděním zásobních látek
v listech, s rychlejší tvorbou buněk nebo s jejich zvětšováním, což může za následek např.
zvětšování plochy listu a změnu jeho tvaru. Často dochází ke snížení hustoty průduchů a ke
změnám ve struktuře mezofylu.
Změny ve struktuře jehlic a listů jsou obecně převážně započaty již při vzniku listů a
v dospělých listech přetrvávají. Důvodem změn je ovlivnění fyziologických procesů, zejména
fotosyntézy, buněčného cyklu a růstu buněk, které dále působí na utváření stavby listů.
Při studiu vlivu zvýšené koncentrace CO2 na rostliny musí být vzat v úvahu možný
vznik artefaktů, který závisí na použitých metodách experimentu. Záleží nejen na metodě
obohacování vzduchu oxidem uhličitým (mezi nejlépe hodnocené metody se v současné době
řadí metoda FACE), ale také na obsahu vody a živin v půdě, na ozářenosti a na dalších
faktorech prostředí. Dalším z problémů při snaze o porovnání výsledků různých experimentů
je – mimo rozdílného stáří rostlin a odlišné délky trvání experimentu – rozdílné pojetí hodnot
normální a zvýšené [CO2], které je způsobeno zejména rychle se měnící [CO2] v atmosféře.
Autoři většinou volili jako hodnotu EC dvojnásobek hodnoty AC, ale ani to nebylo vždy
pravidlem. Dále musí být bráno v potaz, zda jsou vybrané vzorky reprezentativní (například
z hlediska umístění jehlic v prostoru a gradientu strukturálních charakteristik v rámci jehlice).
Dle mého názoru také velmi záleží na metodě získání a zpracování dat, včetně počtu vzorků
zahrnutých do analýz. Z výše uvedeného vyplývá, že výsledky dosažené v odlišných
experimentálních podmínkách jsou velmi obtížně porovnatelné. Pro lepší možnosti
srovnávání by bylo třeba ustanovit nějaká všeobecně přijatelná pravidla pro zkoumání vlivu
EC na strukturu listu.
Mechanismy působení zvýšené koncentrace oxidu uhličitého na anatomii listu nejsou
v současné době zdaleka objasněny a zaslouží si proto další zkoumání vedoucí k vysvětlení
působení zvýšené koncentrace CO2 na list, včetně jejích interakcí s dalšími faktory.

36
4. Citovaná literatura
Ainsworth EA, Rogers A, Vodkin LO, Walter A, Schurr U. 2006. The effects of elevated CO2 concentration on soybean gene expresion. An analysis of growing and mature leaves. Plant Physiology 142, 135–147.
Apple ME, Olszyk DM, Ormrod DP, Lewis J, Southworth D, Tingey DT. 2000. Morphology and stomatal function of douglas fir needles exposed to climate change: Elevated CO2 and temperature. International Journal of Plant Sciences 161, 127–132.
Beerling DJ, Chaloner WG. 1993. Evolutionary responses of stomatal density to global CO2 change. Biological Journal of Linnean Society 48, 343–353.
Bettarini I, Vaccari F, Miglietta F. 1998. Elevated CO2 concentrations and stomatal density: observations from 17 plant species growing in a CO2 spring in central Italy. Global Change Biology 4, 17–22.
Cavender-Bares J, Potts M, Zacharias E, Bazzaz FA. 2000. Consequences of CO2 and light interactions for leaf phenology, growth, and senescence in Quercus rubra. Global Change Biology 6, 877–887.
Ceulemans R, Mousseau M. 1994. Effects of elevated atmospheric CO2 on woody plants. New Phytologist 127, 425–446.
Conroy J, Barlow EWR, Bevege DI. 1986. Response of Pinus radiata Seedlings to Carbon Dioxide Enrichment at Different Levels of Water and Phosphorus: Growth, Morphology and Anatomy. Annals of Botany 57, 165–177.
Dickison WC. 2000. Integrative Plant Anatomy. Harcourt/Academic Press, San Diego, xvii 533p. ISBN 0-12-2151710-4.
Eguchi E, Fukatsu E, Funada R, Tobita H, Kitao M, Maruyama Y, Koike T. 2004. Changes in morphology, anatomy, and photosynthetic capacity of needles of Japanese larch (Larix kaempferi) seedlings grown in high CO2 concentrations. Photosynthetica 42, 173–178.
Eguchi N, Karatsu K, Ueda T, Funada R, Takagi K, Hiura T, Sasa K, Koike T. 2008. Photosynthetic responses of birch and alder saplings grown in a free air CO2 enrichment system in northern Japan. Trees 22, 437–447.
Esau K. 1977. Anatomy of Seed Plants. 2. ed. John Wiley & Sons, New York, xx, 550 p. ISBN 0-471-24520-8.
Etheridge DM, Steele LP, Langenfelds RL, Francey RJ, Barnola J-M, Morgan VI. 1998. Historical CO2 records from the Law Dome DE08, DE08-2, and DSS ice cores. In Trends: A Compendium of Data on Global Change [online]. CDIAC, ORNL, June 1998. WWW: <http://cdiac.ornl.gov/trends/co2/lawdome.html>.
Evert RF. 2006. Esau’s Plant Anatomy. John Wiley & Sons, Inc., Hoboken, xx, 601 p. ISBN 978-0-471-73843-5.
Fahn A. 1990. Plant Anatomy. Pergamon Press, Oxford, xi, 588 p. ISBN 0-08-037491-3.
Fendrych M. 2005. Kvantitativní analýza průduchů jehlic smrku ztepilého pěstovaného ve zvýšené koncentraci CO2. Praha, 34 s. Katedra fyziologie rostlin, Přírodovědecká fakulta Univerzity Karlovy. Vedoucí seminární práce doc. Jana Albrechtová.
Ferris R, Sabatti M, Miglietta F, Mills RF, Taylor G. 2001. Leaf area is stimulated in Populus by free air CO2 enrichment (POPFACE), through increased cell expansion and production. Plant, Cell and Environment 24, 305–315.
Gibeaut MD, Cramer GR, Seeman JR., 2000. Growth, cell walls, and UDP-Glc dehydrogenase activity of Arabidopsis thaliana grown in elevated carbon dioxide. Journal of Plant Physiology, 158, 569–576.
Griffin KL, Anderson OR, Gastrich MD, Lewis JD, Lin G, Schuster W, Seeman JR, Tissue DT, Turnbull MH, Whitehead D. 2001. Plant growth in elevated CO2 alters mitochondrial number and chloroplast fine structure. Proceedings of the National Academy of Sciences 98, 2473–2478.

37
Jach ME, Ceulemans R. 1999. Effects of elevated atmospheric CO2 on phenology, growth and crown structure of Scots pine (Pinus sylvestris) seedlings after two years of exposure in the field. Tree Physiology 19, 289–300.
Hrstka M, Urban O, Marek MV. 2005. Long-term effect of elevated CO2 on spatial differentiation of ribulose-1,5-bisphosphate carboxylase/oxygenase activity in Norway spruce canopy. Photosynthetica 43, 211–216.
Kinsman EA, Lewis C, Davies MS, Young JE, Francis D, Vilhar B, Ougham HJ. 1997. Elevated CO2 stimulates cells to divide in grass meristems: a diferential effect in two natural populations of Dactylis glomerata. Plant, Cell and Environment 20, 1309–1316.
Körner C. 2003. Atmospheric CO2 enrichment – an ecological perspective. in Larcher W. 2003. Physiological Plant Ecology. 4th edition. Springer, pages 177-184.
Körner C. 2006. Plant CO2 responses: an issue of definition, time and resource supply. New Phytologist 172, 393–411.
Leadley PW, Reynolds JA, Thomas JF, Reynolds JF. 1987. Effects of CO2 enrichment on leaf surface area in soybeans. Botanical Gazette 148, 137–140.
Lhotáková Z, Albrechtová J, Malenovský Z, Rock BN, Polák T, Cudlín, P. 2007. Does the azimuth orientation of Norway spruce (Picea abies/L./Karst.) branches within sunlit crown part influence the heterogeneity of biochemical, structural and spectral characteristics of needles? Environmental and Experimental Botany 59, 283–292.
Lin J, Jach ME, Ceulemans R. 2001. Stomatal density and needle anatomy of Scots pine (Pinus sylvestris) are affected by elevated CO2. New Phytologist 150, 665–674.
Luomala E-M, Laitinen K, Sutinen S, Kellomäki S, Vapaavuori E. 2005. Stomatal density, anatomy and nutrient concentrations of Scots pine needles are affected by elevated CO2 and temperature. Plant, cell and envinronment 28, 733–749.
Masle J. 2000. The effects of elevated CO2 concentrations on cell division rates, growth patterns, and blade anatomy in young wheat plants are modulated by factors related to leaf position, vernalization, and genotype. Plant Physiology 122, 1399–1415.
Mauseth JD. 1988. Plant Anatomy. The Benjamin/Cummings Publishing Company, Inc., Menlo Park, xiv, 560 p. ISBN 0-8053-4570-1.
Nátr L 1998. Fotosyntéza. In: Procházka S, Macháčková I, Krekule J, Šebánek J a kolektiv. Fyziologie rostlin., 1. vyd. Academia, Praha, 484 s. ISBN 80-200-0586-2.
Nátr L. 2000. Koncentrace CO2 a rostliny. ISV nakladatelství, Praha, 257 s. ISBN 80-85866-62-5.
Norby RJ, O’Neill EG. 1991. Leaf area compensation and nutrient interactions in CO2-enriched seedling of yellow poplar (Liriodendron tulipifera L.). New Phytologist 117, 515–528.
Oksanen E, Riikonen J, Kaakinen S, Holopainen T, Vapaavuori E. 2005. Structural characteristics and chemical composition of birch (Betula pendula) leaves are modifired by increasing CO2 and ozone. Global Change Biology 11, 732–748.
Oksanen E, Sober J, Karnosky DF. 2001. Impacts of elevated CO2 and/or O3 on leaf ultrastructure of aspen (Populus tremuloides) and birch (Betula papyrifera) in the Aspen FACE experiment. Envinronmental Pollution 115, 437–446.
Olszyk D, Apple A, Gartner B, Spicer R, Wise C, Buckner E, Benson-Scott A, Tingey D. 2005. Xeromorphy increases in shoots of Pseudotsuga menziesii (Mirb.) Franco seedlings with exposure to elevated temperature but not elevated CO2. Trees 19, 552–563.
Onoda Y, Hirose T, Hikosaka K. 2007. Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three natural CO2 springs in Japan. Ecological Research 22, 475–484.
Pazourek J, Votrubová O. 1997. Atlas of Plant Anatomy. PERES Publishers, Praha, 447 p. ISBN 80-901691-2-0.

38
Pritchard SG, Mosjidis C, Peterson CM, Runion B, Rogers HH. 1998. Anatomical and morphological alterations in longleaf pine needles resulting from growth in elevated CO2: Interactions with soil resource availability. International Journal of Plant Sciences 159, 1002–1009
Pritchard SG, Peterson CM, Prior SA, Rogers HH. 1997. Elevated atmospheric CO2 differentially affects needle chloroplast ultrastructure and phloem anatomy in Pinus palustris: interactions with soil resource availability. Plant, Cell and Environment 20, 461–471.
Ranasinghe S, Taylor G. 1996. Mechanism for increased leaf growth in elevated CO2. Journal of Experimental Botany 47, 349–358.
Sholtis JD, Gunderson CA, Norby RJ, Tissue DT. 2004. Persistent stimulation of photosynthesis by elevated CO2 in a sweetgum (Liquidambar styraciflua) forest stand. New Phytologist 162, 343–354.
Smith AM, Stitt M. 2007. Coordination of carbon supply and plant growth. Plant, Cell and Environment 30, 1126–1149.
Soukupova J, Cvikrova M, Albrechtova J,Rock BN, Eder J. 2000a. Histochemical and biochemical approaches to the study of phenolic compounds and peroxidases in needles of Norway spruce (Picea abies). New Phytologist 146, 403–414.
Soukupova J, Rock BN, Albrechtova J. 2000b. Comparative study of two spruce species in a polluted mountain region. New Phytologist 150, 133–145.
Tans P. Trends in Carbon Dioxide [online]. NOAA/ESRL, March 2008. WWW: <http://www.esrl.noaa.gov/gmd/ccgg/trends/>.
Taylor G, Tricker PJ, Zhang FZ, Alston VJ, Miglietta F, Kuzminsky E. 2003. Spatial and temporal effects of Free-Air CO2 Enrichment (POPFACE) on leaf growth, cell expansion, and cell production in a closed canopy of poplar. Plant Physiology 131, 177–185.
Teng N, Wang J, Chen T, Wu X, Wang Y, Lin J. 2006. Elevated CO2 induces physiological, biochemical and structural changes in leaves of Arabidopsis thaliana. New Phytologist 172, 92–103.
Thomas JF, Harvey CN. 1983. Leaf anatomy of four species grown under continuous CO2 enrichment. Botanical Gazette 144, 303–309.
Thomas SC, Bazzaz FA. 1996. Elevated CO2 and leaf shape: Are dandelions getting toothier? American Journal of Botany 83, 106–111.
Tissue TD, Thomas RB, Strain BR. 1996. Growth and photosynthesis of loblolly pine (Pinus taeda) after exposure to elevated CO2 for 12 months in the field. Tree Physiology 16, 49–59.
Tomaskova I, Pokorny R, Marek M.V. Effect of elevated atmospheric CO2 concentration on Norway spruce needle parameters. Nepublikováno.
Tricker PJ, Calfapietra C, Kuzminsky E, Puleggi R, Ferris R, Nathoo M, Pleasants LJ, Alston V, de Angelis P, Taylor G. 2004. Long-term acclimation of leaf production, development, longevity and quality following 3 yr exposure to free-air CO2 enrichment during canopy closure in Populus. New Phytologist 162, 413–426.
Votrubová O. 2001. Anatomie rostlin. 2. vyd. Nakladatelství Karolinum, Praha, 89 s. ISBN 80-246-0367-5.
WMO. 2007. Greenhouse Gas Bulletin [online]. WMO, November 2007. WWW: <http://www.wmo.int/pages/prog/arep/gaw/ghg/documents/ghg-bulletin-3.pdf>.