virut thực vật phytoplasma và viroid - hà viết cường
DESCRIPTION
virut tv phytoplasma và viroidTRANSCRIPT

VIRUS THỰC
VẬT, PHYTOPLASMA
VÀ VIROID

1
TRƢỜNG ĐẠI HỌC NÔNG NGHIỆP HÀ NỘI
KHOA NÔNG HỌC
VIRUS THỰC VẬT, PHYTOPLASMA VÀ VIROID
(Bài giảng)
Biên soạn
TS. Hà Viết Cƣờng
Hà Nội, 2010

2
MỤC LỤC
MỤC LỤC .................................................................................................................................................................... 2
LỜI NÓI ĐẦU .............................................................................................................................................................. 6
DANH MỤC CHỮ VIẾT TẮT ..................................................................................................................................... 7
DANH SÁCH BẢNG .................................................................................................................................................... 8
DANH SÁCH HÌNH ..................................................................................................................................................... 9
PHẦN I: ĐẠI CƢƠNG ................................................................................................................................................12
CHUƠNG 1. GIỚI THIỆU ......................................................................................................................................13
1.1 TÓM TẮT NỘI DUNG ..................................................................................................................................13 1.2 LỊCH SỬ VIRUS ...........................................................................................................................................13 1.3 CÁC CỘT MỐC LỊCH SỬ QUAN TRỌNG KHÁC .......................................................................................14 1.4 THUÂT NGỮ VIRUS ...................................................................................................................................15 1.5 ĐỊNH NGHĨA VIRUS ...................................................................................................................................15 1.6 NGUỒN GỐC VIRUS ...................................................................................................................................16 1.7 TẦM QUAN TRỌNG CỦA VIRUS THỰC VẬT...........................................................................................17 1.8 CÂU HỎI ÔN TẬP CHƢƠNG 1 ....................................................................................................................18 1.9 TÀI LIỆU THAM KHẢO CHÍNH .................................................................................................................18
CHƢƠNG 2. PHÂN LOẠI VÀ DANH PHÁP ........................................................................................................19
2.1 TÓM TẮT NỘI DUNG ..................................................................................................................................19 2.2 PHÂN LOẠI VÀ DANH PHÁP .....................................................................................................................19 2.3 LịCH SỬ DANH PHÁP VÀ PHÂN LOẠI VIRUS .........................................................................................19 2.4 ỦY BAN PHÂN LOẠI VIRUS QUỐC TẾ (ICTV) .........................................................................................20 2.5 DANH PHÁP VIRUS HIỆN TẠI THEO ICTV ..............................................................................................22 2.6 PHÂN BIỆT VIRUS VÀ LOÀI VIRUS .........................................................................................................23 2.7 KHÁI NIỆM LOÀI VIRUS............................................................................................................................23 2.8 CÁC CHỈ TIÊU PHÂN LOẠI VIRUS ............................................................................................................24 2.9 HỆ THỐNG PHÂN LOẠI VIRUS HIỆN TẠI THEO ICTV............................................................................26 2.10 HỆ THỐNG PHÂN LOẠI VIRUS THEO BALTIMORE...........................................................................28 2.11 CÂU HỎI ÔN TẬP CHƢƠNG 2 ...............................................................................................................29 2.12 TÀI LIỆU THAM KHẢO CHÍNH ............................................................................................................29
CHƢƠNG 3. HÌNH THÁI VÀ CẤU TRÚC VIRUS ..............................................................................................31
3.1 TÓM TẮT NỘI DUNG ..................................................................................................................................31 3.2 MỘT SỐ THUẬT NGỮ.................................................................................................................................31 3.3 HÌNH THÁI VIRUS ......................................................................................................................................31 3.4 VIRUS TRẦN VÀ VIRUS CÓ VỎ BỌC .......................................................................................................31 3.5 CẤU TRÖC PHÂN TỬ VIRUS .....................................................................................................................33 3.6 CẤU TRÖC DẠNG KHỐI ĐA DIỆN ĐỐI XỨNG (ICOSAHEDRON) ..........................................................33 3.7 CÁC PHÂN TỬ VỚI KHỐI ĐA DIỆN ĐỐI XỨNG DÀI ...............................................................................36 3.8 CẤU TRÖC ĐỐI XỨNG XOẮN ...................................................................................................................36 3.9 CẤU TRÖC KHÔNG ĐỐI XỨNG (PHỨC TẠP) ...........................................................................................37 3.10 THÀNH PHẦN CẤU TẠO CỦA PHÂN TỬ VIRUS ................................................................................38 3.11 CÂU HỎI ÔN TẬP CHƢƠNG 3 ...............................................................................................................39 3.12 TÀI LIỆU THAM KHẢO CHÍNH ...........................................................................................................39
CHƢƠNG 4. TÁI SINH VIRUS (REPLICATION) .................................................................................................40
4.1 TÓM TẮT NỘI DUNG ..................................................................................................................................40 4.2 KHÁI NIỆM ..................................................................................................................................................40 4.3 ĐẶC ĐIỂM TÁI SINH VIRUS ......................................................................................................................40 4.4 TÁI SINH CỦA CÁC VIRUS CÓ BỘ GEN RNA SỢI (+) ..............................................................................40 4.5 TÁI SINH CỦA CÁC VIRUS CÓ BỘ GEN RNA SỢI (-) ..............................................................................44 4.6 TÁI SINH CỦA VIRUS RNA SỢI KÉP .........................................................................................................45 4.7 TÁI SINH CỦA VIRUS RNA QUA PHIÊN MÃ NGƢỢC (RETROVIRUS) ..................................................47 4.8 TÁI SINH CỦA VIRUS DNA SỢI VÕNG ĐƠN ............................................................................................50 4.9 TÁI SINH CỦA VIRUS DNA SỢI VÕNG KÉP PHIÊN MÃ NGƢỢC (PARARETROVIRUS) ......................51 4.10 CÁC CHIẾN LƢỢC DỊCH MÃ CỦA VIRUS THỰC VÂT .......................................................................53 4.11 CÂU HỎI ÔN TẬP CHƢƠNG 4 ...............................................................................................................58

3
4.12 TÀI LIỆU THAM KHẢO CHÍNH ............................................................................................................58
CHƢƠNG 5. SỰ DI CHUYỂN CỦA VIRUS TRONG CÂY ..................................................................................60
5.1 TÓM TẮT NỘI DUNG ..................................................................................................................................60 5.2 DI CHUYỂN GIỮA CÁC TẾ BÀO................................................................................................................60 5.3 DI CHUYỂN HỆ THỐNG QUA KHOẢNG CÁCH XA .................................................................................62 5.4 CÂU HỎI ÔN TẬP CHƢƠNG 5 ....................................................................................................................63 5.5 TÀI LIỆU THAM KHẢO CHÍNH .................................................................................................................63
CHƢƠNG 6. CƠ CHẾ GÂY BỆNH CỦA VIRUS THỰC VẬT .............................................................................64
6.1 TÓM TẮT NỘI DUNG ..................................................................................................................................64 6.2 TRIỆU CHỨNG BỆNH VIRUS .....................................................................................................................64 6.3 HAI MÔ HÌNH GIẢI THÍCH CƠ CHẾ GÂY BỆNH CỦA VIRUS.................................................................65 6.4 CÁC CƠ CHẾ GÂY BỆNH CỦA VIRUS ......................................................................................................66 6.5 CÂU HỎI ÔN TẬP CHƢƠNG 6 ....................................................................................................................68 6.6 TÀI LIỆU THAM KHẢO CHÍNH .................................................................................................................68
CHƢƠNG 7. LAN TRUYỀN CỦA VIRUS THỰC VẬT ........................................................................................69
7.1 TÓM TẮT NỘI DUNG ..................................................................................................................................69 7.2 GIỚI THIỆU .................................................................................................................................................69 7.3 LAN TRUYỀN QUA TIẾP XÖC CƠ HỌC ....................................................................................................69 7.4 LAN TRUYỀN QUA NHÂN GIỐNG VÔ TÍNH ............................................................................................69 7.5 TRUYỀN QUA HẠT GIỐNG (SEED) ...........................................................................................................70 7.6 LAN TRUYỀN QUA MÔI GIỚI (CỰC KỲ QUAN TRỌNG) ........................................................................71 7.7 CƠ CHẾ TRUYỀN VIRUS NHỜ VECTOR CÔN TRÙNG ............................................................................71 7.8 HÀNH VI CHÍCH HÖT CỦA CÔN TRÙNG BỘ HEMIPTERA ....................................................................73 7.9 LAN TRUYỀN VIRUS THEO KIỂU KHÔNG BỀN VỮNG ..........................................................................73 7.10 LAN TRUYỀN VIRUS THEO KIỂU BÁN BỀN VỮNG ..........................................................................77 7.11 LAN TRUYỀN VIRUS THEO KIỂU BỀN VỮNG TUẦN HOÀN ............................................................78 7.12 TRUYỀN THEO KIỂU BỀN VỮNG TÁI SINH .......................................................................................79 7.13 CÂU HỎI ÔN TẬP CHƢƠNG 7 ...............................................................................................................81 7.14 TÀI LIỆU THAM KHẢO CHÍNH ............................................................................................................82
CHƢƠNG 8. PHÒNG CHỐNG BỆNH VIRUS THỰC VẬT .................................................................................83
8.1 TÓM TẮT NỘI DUNG ..................................................................................................................................83 8.2 CÁC CHIẾN LƢỢC PHÕNG CHỐNG BỆNH VIRUS ...................................................................................83 8.3 SỬ DỤNG VẬT LIỆU GIỐNG SÁCH BỆNH................................................................................................83 8.4 SỬ DỤNG GIỐNG KHÁNG BỆNH MANG GEN KHÁNG CỦA CÂY .........................................................83 8.5 SỬ DỤNG GIỐNG KHÁNG BỆNH DÙNG GEN VIRUS..............................................................................86 8.6 ỨNG DỤNG TÍNH KHÁNG TẠO ĐƢỢC (INDUCED RESISTANCE) .........................................................92 8.7 CÂU HỎI ÔN TẬP CHƢƠNG 8 ....................................................................................................................96 8.8 TÀI LIỆU THAM KHẢO CHÍNH .................................................................................................................97
CHƢƠNG 9. CHẨN ĐOÁN BỆNH VIRUS THỰC VẬT .......................................................................................99
9.1 TÓM TẮT NỘI DUNG ..................................................................................................................................99 9.2 GIỚI THIỆU .................................................................................................................................................99 9.3 CHẨN ĐOÁN DỰA VÀO TRIỆU CHỨNG ..................................................................................................99 9.4 CHẨN ĐOÁN DỰA VÀO CÂY CHỈ THỊ .....................................................................................................99 9.5 KĨ THUẬT HIỂN VI ĐIỆN TỬ ................................................................................................................... 100 9.6 KĨ THUẬT CHẨN ĐOÁN DỰA VÀO THỂ VÙI ........................................................................................ 101 9.7 KĨ THUẬT PCR VÀ RT-PCR...................................................................................................................... 102 9.8 CHẨN ĐOÁN DỰA VÀO RNA SỢI KÉP ................................................................................................... 106 9.9 CHẨN ĐOÁN DỰA VÀO HUYẾT THANH HỌC ...................................................................................... 106 9.10 CÁC KĨ THUẬT DOT BLOT ................................................................................................................. 112 9.11 CÂU HỎI ÔN TẬP CHƢƠNG 9 ............................................................................................................. 113 9.12 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 113
PHẦN II: CHUYÊN KHOA ...................................................................................................................................... 114
CHƢƠNG 10. CÁC VIRUS THỰC VẬT Ở VIỆT NAM ...................................................................................... 115
10.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 115 10.2 CÁC VIRUS ĐƢỢC PHÁT HIỆN Ở VIỆT NAM ................................................................................... 115 10.3 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 119

4
CHƢƠNG 11. VIRUS DNA SỢI KÉP (PHIÊN MÃ NGƯỢC): HỌ CAULIMOVIRIDAE .................................. 121
11.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 121 11.2 HỌ CAULIMOVIRIDAE ....................................................................................................................... 121 11.3 RICE TUNGRO BACILLIFORM VIRUS ............................................................................................... 122 11.4 CÂU HỎI ÔN TẬP CHƢƠNG 11 ........................................................................................................... 124 11.5 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 124
CHƢƠNG 12. VIRUS DNA SỢI ĐƠN: CHI BEGOMOVIRUS (HỌ GEMINIVIRIDAE) ................................... 125
12.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 125 12.2 HỌ GEMINIVIRIDAE ........................................................................................................................... 125 12.3 CHI BEGOMOVIRUS ............................................................................................................................ 126 12.4 BỆNH DO BEGOMOVIRUS.................................................................................................................. 130 12.5 CÂU HỎI ÔN TẬP CHƢƠNG 12 ........................................................................................................... 133 12.6 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 134
CHƢƠNG 13. VIRUS DNA SỢI ĐƠN (TIẾP): CHI BABUVIRUS (HỌ NANOVIRIDAE) ................................ 135
13.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 135 13.2 HỌ NANOVIRIDAE .............................................................................................................................. 135 13.3 BANANA BUNCHY TOP VIRUS (BBTV) ............................................................................................ 137 13.4 CÂU HỎI ÔN TẬP CHƢƠNG 13 ........................................................................................................... 138 13.5 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 138
CHƢƠNG 14. VIRUS RNA SỢI ĐƠN CỰC DƯƠNG: CHI TOBAMOVIRUS (HỌ VIRGAVIRIDAE) ............ 139
14.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 139 14.2 HỌ VIRGAVIRIDAE ............................................................................................................................. 139 14.3 CHI TOBAMOVIRUS ............................................................................................................................ 139 14.4 TOBACCO MOSAIC VIRUS (TMV) ..................................................................................................... 140 14.5 CÂU HỎI ÔN TẬP CHƢƠNG 14 ........................................................................................................... 144 14.6 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 144
CHƢƠNG 15. VIRUS RNA SỢI ĐƠN CỰC DƯƠNG (TIẾP): CHI POTYVIRUS (HỌ POTYVIRIDAE) ........ 145
15.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 145 15.2 HỌ POTYVIRIDAE ............................................................................................................................... 145 15.3 CHI POTYVIRUS .................................................................................................................................. 146 15.4 PAPAYA RING SPOT VIRUS (PRSV) .................................................................................................. 150 15.5 POTATO VIRUS Y (PVY) ..................................................................................................................... 152 15.6 BEAN COMMON MOSAIC VIRUS (BCMV) ........................................................................................ 155 15.7 SUGARCANE MOSAIC VIRUS (SCMV) VÀ SORGHUM MOSAIC VIRUS (SRMV) .......................... 160 15.8 ONION YELLOW DWARF VIRUS (OYDV), LEEK YELLOW STRIPE VIRUS (LYSV) VÀ SHALLOT
YELLOW STRIPE VIRUS (SYSV) ................................................................................................................................. 163 15.9 TURNIP MOSAIC VIRUS (TUMV) ....................................................................................................... 164 15.10 DASHEEN MOSAIC VIRUS (DSMV) ................................................................................................... 166 15.11 SWEET POTATO FEATHERY MOTTLE VIRUS (SPFMV) .................................................................. 167 15.12 ZUCCHINI YELLOW MOSAIC VIRUS (ZYMV) .................................................................................. 168 15.13 CHILLI VEINAL MOTLE VIRUS (CHIVMV) VÀ CHILLI RINGSPOT VIRUS (CHIRSV)................... 170 15.14 CÂU HỎI ÔN TẬP CHƢƠNG 15 ........................................................................................................... 171 15.15 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 171
CHƢƠNG 16. VIRUS RNA SỢI ĐƠN CỰC ÂM: RHABDOVIRUS .................................................................. 173
16.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 173 16.2 HỌ RHABDOVIRIDAE ......................................................................................................................... 173 16.3 RICE YELLOW STUNT VIRUS (RYSV) ............................................................................................... 175 16.4 CÂU HỎI ÔN TẬP CHƢƠNG 16 ........................................................................................................... 178 16.5 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 178
CHƢƠNG 17. VIRUS RNA SỢI ĐƠN CỰC ÂM (TIẾP): TOSPOVIRUS ......................................................... 179
17.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 179 17.2 HỌ BUNYAVIRIDAE ........................................................................................................................... 179 17.3 CHI TOSPOVIRUS ................................................................................................................................ 179 17.4 TOMATO SPOTTED WILT VIRUS (TSWV)......................................................................................... 180 17.5 CÂU HỎI ÔN TẬP CHƢƠNG 17 ........................................................................................................... 183 17.6 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 183

5
CHƢƠNG 18. CÁC VIRUS RNA SỢI KÉP: HỌ REOVIRIDAE ........................................................................ 184
18.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 184 18.2 HỌ REOVIRIDAE ................................................................................................................................. 184 18.3 RICE RAGGED STUNT VIRUS (RRSV) ............................................................................................... 186 18.4 SOUTHERN RICE BLACK-STREAKED DWARF VIRUS (SRBSDV) .................................................. 189 18.5 CÂU HỎI ÔN TẬP CHƢƠNG 18 ........................................................................................................... 192 18.6 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 192
CHƢƠNG 19. VIRUS RNA SỢI KÉP (TIẾP): CHI TENUIVIRUS ..................................................................... 194
19.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 194 19.2 CHI TENUIVIRUS ................................................................................................................................. 194 19.3 RICE GRASSY STUNT VIRUS (RGSV) ................................................................................................ 194 19.4 CÂU HỎI ÔN TẬP CHƢƠNG 19 ........................................................................................................... 197 19.5 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 197
CHƢƠNG 20. PHYTOPLASMA .......................................................................................................................... 198
20.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 198 20.2 GIỚI THIỆU .......................................................................................................................................... 198 20.3 ĐẶC ĐIỂM ............................................................................................................................................ 198 20.4 PHÂN LOẠI .......................................................................................................................................... 199 20.5 TRIỆU CHỨNG ..................................................................................................................................... 199 20.6 LAN TRUYỀN ....................................................................................................................................... 200 20.7 PHÕNG CHỐNG ................................................................................................................................... 200 20.8 CA. PHYTOPLASMA ASTERIS ............................................................................................................ 200 20.9 CÂU HỎI ÔN TẬP CHƢƠNG 20 ........................................................................................................... 201 20.10 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 201
CHƢƠNG 21. VIROID .......................................................................................................................................... 202
21.1 TÓM TẮT NỘI DUNG ........................................................................................................................... 202 21.2 GIỚI THIỆU .......................................................................................................................................... 202 21.3 ĐỊNH NGHĨA VIROID .......................................................................................................................... 202 21.4 CẤU TRÖC VIROID .............................................................................................................................. 202 21.5 DANH PHÁP VÀ PHÂN LOẠI .............................................................................................................. 203 21.6 TRIỆU CHỨNG, CƠ CHẾ GÂY BỆNH VÀ LAN TRUYỀN .................................................................. 204 21.7 TÁI SINH CỦA VIROID ........................................................................................................................ 205 21.8 POTATO SPINDLE TUBER VIROID (PSTVD) ..................................................................................... 205 21.9 COCONUT CADANG-CADANG VIROID (CCCVD) ............................................................................ 206 21.10 AVOCADO SUNBLOTCH VIROID (ASBVD) ...................................................................................... 206 21.11 CÂU HỎI ÔN TẠP CHƢƠNG 21 ........................................................................................................... 206 21.12 TÀI LIỆU THAM KHẢO CHÍNH .......................................................................................................... 206
PHẦN 3: THỰC HÀNH ............................................................................................................................................ 207
BÀI 1. ĐIỀU TRA BỆNH VIRUS ........................................................................................................................ 208
BÀI 2. LÂY NHIỄM NHÂN TẠO VIRUS BẰNG TIẾP XÖC CƠ HỌC ............................................................ 210
BÀI 3. PHÁT HIỆN VIRUS BẰNG ELISA ......................................................................................................... 211
BÀI 4. PHÁT HIỆN VIRUS BẰNG PCR/RT-PCR ............................................................................................. 213

6
LỜI NÓI ĐẦU
Tài liệu này đƣợc biên soạn để giảng dạy môn virus thực vật, phytoplasma và viroid cho
sinh viên và học viên cao học khối ngành nông học, công nghệ sinh học thực vật. Tài liệu giúp
ngƣời học hiểu và có khả năng vận dụng kiến thức của môn học nhằm định hƣớng nghiên cứu
bệnh virus thực vật.
Tài liệu giới thiệu các vấn đề quan trọng của virus thực vật bao gồm: Lịch sử nghiên cứu
virus; bản chất và phân phân loại virus; các đặc điểm hình thái, sinh học, đặc biệt là cơ chế gây
bệnh, sinh sản và sự lan truyền virus; chẩn đoán và phòng chống virus. Các ví dụ cụ thể là các
virus thực vật có ý nghĩa lịch sử, kinh tế trên thế giới. Phần lớn các ví dụ đã đƣợc xác định có
mặt ở Việt Nam. Ngoài ra, 2 nhóm tác nhân gây bệnh có đặc điểm gây bênh giống với virus là
phytoplasma và viroid cũng đƣợc đề cập. Một số nội dung, đặc biệt phần đại cƣơng, cũng đƣợc
so sánh với virus động vật, thực khuẩn thể để ngƣời đọc có cái nhìn tổng thể đối với virus học
nói chung. Kiến thức cơ bản về virus trình bày trong tài liệu này đều đƣợc cập nhật và phổ quát
trên thế giới.
Tài liệu đƣợc biên soạn gồm 3 phần:
Phần đại cƣơng. Phần đại cƣơng giới thiệu các khái niệm cơ bản của virus thực vật học
liên quan đến bản chất của virus. Phần này cũng giới thiệu các kỹ thuật chẩn đoán hiện đang
đƣợc sử dụng phổ biến trên thế giới và Việt Nam cũng nhƣ các chiến lƣợc phòng chống bệnh
chính.
Phần chuyên khoa. Phần chuyên khoa giới thiệu các ví dụ cụ thể về các virus có ý nghĩa
lịch sử, có ý nghĩa kinh tế. Phần lớn các virus này đã đƣợc phát hiện thấy ở Việt Nam. Các virus
đƣợc trình bày theo đơn vị phân loại chứ không theo đối tƣợng gây hại.
Phần thực hành. Phần thực hành trình bày các bài thực hành giúp sinh viên củng cố khả
năng đánh giá bệnh virus ngoài thực tiễn cũng nhƣ nắm đƣợc kỹ thuật chẩn đoán bệnh virus thực
vật hiện đang đƣợc sử dụng rộng rãi trên thế giới và Việt Nam
Đối với chƣơng trình đại học, các nội dung học đƣợc sắp xếp tƣơng đƣơng 2 tín chỉ và đƣợc
tổ chức thành 8 bài lý thuyết (3 tiết / bài) và 3 bài thực hành. Mức độ tự học của sinh viên tập
trung ở phần chuyên khoa. Sinh viên sẽ đƣợc học lý thuyết, đƣợc thực hành và phải thực hiện
một bài tiểu luận liên quan đến nội dung môn học.
Do trình độ có hạn, việc biên soạn bài giảng này không tránh khỏi sai sót. Tác giả rất mong
nhận đƣợc ý kiến đóng góp của bạn đọc để tài liệu đƣợc hoàn chỉnh.
Tác giả
Hà Viết Cƣờng

7
DANH MỤC CHỮ VIẾT TẮT
aa Amino acid
AP Alkaline phosphatase
ATPase Adenosine triphosphatase
Avr Avirulence
bp Base pair
CP Coat protein
CTAB Cetyl trimethylammonium bromide
DAS-ELISA Double antibody sandwich - ELISA
DdRp DNA-dependent DNA polymerase
dsDNA Double-stranded DNA
dsRNA Double-stranded RNA
eIF4E Eukaryotic translation initiation factor 4E
ELISA Enzyme linked immunosorbent assay
EM Electron microscope
g Gravity
ICTV International Committee on Taxonomy of Viruses
IgG Immunoglobulin gamma
IR Induced resistance
IRES Internal Ribosome Entry Site
ISR Induced systemic resistance
kb Kilobase
LIV Longevity in vitro
Mab Monoclonal antibody
MP Movement protein
NPP Nitrophenyl phosphate
nt nucleotide
NTP Nucleotide triphosphate
NTPase Nucleotide triphosphatase
ORF Open reading frame
Pab Polyclonal antibody
PCR Polymerase Chain Reaction
PDR Pathogen derived resistance
PR Pathogenesis related (protein)
pRBR Plant retinoblastoma-related protein
PTA-ELISA Plate trapped antigen - ELISA
PTGS Post-transcriptional gene silencing
R Resistance
RdRp RNA-dependent RNA polymerase
Rep Replication
RF Replicative form
RI Replicative intermediate
RISC RNA induced silencing complex
RNAi RNA interference
RT Reverse Transcriptase
RT-PCR Reverse Transcription - Polymerase Chain Reaction
SA Salycilic acid
SAR Systemic acquired resistance
SEL Size exclusion limit
ssDNA Single-strannded DNA
ssRNA Single-strannded RNA
TDP Thermal death point
UTR Untralslated region
VRC Viral replication complex

8
DANH SÁCH BẢNG
Bảng 2-1 Các bộ, họ virus theo phân loại của ICTV (dựa trên số liệu 2009) .......................................................... 26 Bảng 2-2 Hệ thống phân loại virus theo Baltimore ............................................................................................... 28 Bảng 3-1. So sánh một số đặc điểm hình thái – cấu trúc của các nhóm virus .......................................................... 39 Bảng 7-1 Thành phần vector của virus thực vật tính đến 2007 (Hogenhout et al., 2008) ......................................... 71 Bảng 7-2. Bốn phƣơng thức truyền virus thực vật của côn trùng môi giới .............................................................. 72 Bảng 7-3. Các họ, chi virus truyền qua aphid và phƣơng thức truyền (James & Perry, 2004) ................................. 74 Bảng 8-1. Một số gen kháng R của cây và Avr tƣơng ứng của virus (Soosaar et al., 2005; Kang et al., 2005) ......... 86 Bảng 9-1 Mã suy biến trong thiết kế mồi chung .................................................................................................. 105 Bảng 11-1 Phân loại các virus thuộc họ Caulimoviridae ...................................................................................... 121 Bảng 12-1. Phân loại các virus họ Geminiviridae theo ICTV năm 2009................................................................ 125 Bảng 12-2 Các begomovirus hại cây trồng đƣợc phát hiện cho tới nay tại Việt Nam ............................................. 133 Bảng 13-1 Phân loại họ Nanoviridae ................................................................................................................... 135 Bảng 14-1 Phân loại họ Virgaviridae ................................................................................................................. 139 Bảng 15-1. Phân loại họ Potyviridae (Berger et al. 2005) ..................................................................................... 145 Bảng 15-2. Một số isolate BCMV ở Việt Nam đƣợc giải trình tự gen CP ............................................................. 157 Bảng.15-3 Một số isolate SCMV và SrMV đƣợc giải trình tự gen CP................................................................... 161 Bảng 15-4 Một số isolate TuMV tại Việt Nam đƣợc giải trình tự gen CP............................................................ 165 Bảng 15-5 Một số mẫu ZYMV tại Việt Nam đƣợc giải trình tự gen CP ................................................................ 169 Bảng 15-6 Các mẫu potyvirus trên ớt tại Việt Nam đƣợc giải trình tự ................................................................... 170 Bảng 16-1. Phân loại họ Rhabdoviridae (bộ Monogavirales) theo ICTV (2009) .................................................. 173 Bảng 16-2 So sánh khả năng truyền RYSV của 3 loài rầy xanh trên lúa................................................................ 177 Bảng 17-1. Phân loại họ Bunyaviridae theo ICTV (2009).................................................................................... 179 Bảng 18-1 Phân loại họ Reoviridae (các reovirus đƣợc ghi nhận đến 2009) .......................................................... 184 Bảng 18-2 Đặc điểm bộ gen và chức năng protein của RRSV .............................................................................. 187 Bảng 18-3 Đặc điểm bộ gen và dự đoán chức năng các protei của virus SRBSDV (Wang et al., 2010) ................. 190 Bảng 19-1 Phân loại chi Tenuivirus (loài chuẩn: Rice stripe virus) ...................................................................... 194 Bảng 20-1 Phân loại phytoplasma dựa trên đặc điểm RNA ribosome 16S............................................................. 199 Bảng 21-1 Phân loại viroid (ICTV, 2009) ............................................................................................................ 204

9
DANH SÁCH HÌNH
Hình 1-1 Triệu chứng bệnh virus trên hoa tulip. Ảnh trái, một bức tranh thế kỷ 17. Ảnh phải, hoa tulip bị nhiễm
TBV (Lesnaw & Ghabrial, 2000). 13 Hình 1-2 Ba nhà khoa học có đóng góp lớn về virus học. Từ trái sang phải: Adolf Mayer (ngƣời Hà Lan, 1843-
1942), Dimitrij Ivanovskij (ngƣời Nga, 1864-1920) và Martinus Beijerinck (ngƣời Hà Lan, 1851-1931). 14 Hình 2-1 Sơ đồ minh họa 5 thành viên của 1 lớp đa hình có 5 đặc điểm (Van Regenmortel, 2007) 24 Hình 2-2 Minh họa phân loại các chi, họ virus thực vật theo báo cáo lần thứ 8 (2005) của ICTV 27 Hình 2-3 Sơ đồ phân loại virus theo Baltimore (http://expasy.org/viralzone/all_by_protein/230.html) 28 Hình 3-1. Minh họa virus trần (hàng trên) và virus có vỏ bọc (hàng dƣới) 32 Hình 3-2 Sơ đồ cấu trúc phân tử TSWV (và các bunyavirus khác) với cấu trúc có vỏ bọc 32 Hình 3-3 Cấu trúc đối xứng đa diện icosahedron cơ bản của virus (T=1, số tiểu phần = 60) 33 Hình 3-4. Hai hình tam giác đều với số số tam giác phụ T = 4 (trái) và T = 9 (phải) 33 Hình 3-5 Minh họa cách vẽ các trục h, k và tọa độ các hình lục giác, cách tạo hình icosahedron tƣơng ứng với 1 T
xác định và cách xắp xếp các tiểu phần protein trên 1 mặt của các virus với 4 mức T khác nhau 34 Hình 3-6 Minh họa virus với T khác nhau sẽ có các đơn vị hình thái (capsomer) khác nhau 35 Hình 3-7 Hình thái Banana bunchy top virus (BBTV) 35 Hình 3-8 Hình thái Tomato bushy stunt virus (TBSV) 35 Hình 3-9 Cách cấu tạo của khối đa diện đối xứng kéo dài (hình trên) và cấu trúc phân tử với T=13 nhƣng có các Q
khác nhau (hình dƣới) 36 Hình 3-10 Cấu trúc đối xứng xoắn của TMV 37 Hình 3-11 Cấu trúc dạng nòng nọc của thực khuẩn thể T4 (Viralzone, 2009) 37 Hình 3-12 Cấu trúc phức tạp của virus đậu mùa 38 Hình 4-1 Sơ đồ giải thích cơ chế tái bản sợi (+) trên khuôn sợi (-). (b) và (c) là cơ chế tổng hợp của virus thực vật
TMV, còn (a) là dạng trung gian RI của thực khuẩn thể Qβ (Buck et al., 1999) 42 Hình 4-2 Sơ đồ tái sinh virus RNA sợi đơn, cực dƣơng 43 Hình 4-3 Sơ đồ tái sinh của rhabdovirus 44 Hình 4-4 Quá trình tái sinh của các rhabdovirus hại thực vật (cytorhabdovirus và nucleorhabdovirus) (Hull, 2002) 45 Hình 4-5 Sơ đồ tái sinh virus RNA sợi kép 46 Hình 4-6 Tổ chức bộ gen của 1 retrovirus động vật (HIV) và của 1 retrovirus thực vật (GmaSIREV) 47 Hình 4-7 Sơ đồ tái sinh của retrovirus động vật 49 Hình 4-8 Tái sinh của retrovirus thực vật 49 Hình 4-9 Sơ đồ tái sinh của begomovirus 51 Hình 4-10 Sơ đồ tái sinh của các caulimovirus 52 Hình 4-11 tổ chức bộ gen và chiến lƣợc dịch mã kiểu polyprotein của potyvirus 53 Hình 4-12 Tổ chức bộ gen và chiến lƣợc dịch mã kiểu gen phụ, bỏ qua mã kết thúc và dịch mã nội phân tử của
TMV 54 Hình 4-13 Bộ gen phân đoạn của BBTV (trái) và RRSV (phải) (Viralzone, 2009) 55 Hình 4-14 Chiến lƣợc dịch mã kiểu ―dò‖. Dấu * là mã ATG. Mũi tên chỉ hƣớng dịch mã. 55 Hình 4-15 Minh họa bộ gen của CaMV và 2 phân tử mRNA 35S và 19S (Haas et al., 2002) 56 Hình 4-16 Mô hình dịch mã của CaMV, RTBV theo cơ chế ―nhảy‖ (Thiébeauld et al., 2007) 57 Hình 4-17 Dịch mã theo kiểu chuyển khung 57 Hình 4-18 Tổ chức bộ gen của TSWV và dịch mã 2 chiều trên phân đoạn M 58 Hình 5-1 Cấu tạo sợi liên bào. A và B là mô hình sợi liên bào, trong đó A là tiết diện dọc và B là tiết diện ngang. C
là ảnh hiện vi điện tử của sợi liên bào (Lucas, 2006). 60 Hình 5-2 Chiến lƣợc tạo ống của virus thực vật để di chuyển giữa các tế bào (Hull, 2002) 61 Hình 5-3 Mô hình di chuyển giữa các tế bào của TMV 62 Hình 5-4 Hƣớng và tốc độ di chuyển hệ thống của virus trong cây (Agrios, 2005) 63 Hình 6-1 Thí nghiệm chứng minh protein P2 của RDP đã can thiệp vào đƣờng hƣớng tổng hợp gibberelline của cây
lúa (Wang et al., 2005) 67 Hình 6-2 Tƣơng tác giữa protein Rep của begomovirus với pRBR của tế bào cho phép khởi động lại chu kỳ tế bào 68 Hình 7-1. Giải phẫu chung côn trùng chích hút và phân bố virus 72 Hình 7-2 Bộ gen của potyvirus (trái) và CMV (phải) kèm theo các gen và motif cần cho lan truyền virus (Ng &
Falk, 2006) 76 Hình 7-3Mô hình tƣơng tác virus và vector theo phƣơng thức không bền vững và bền vững (Ng & Falk, 2006) 76 Hình 7-4 Sơ đồ phân tử TSWV (Hogenhout et al., 2008) 80

10
Hình 7-5 Các cơ quan bên trong của bọ trĩ liên quan đến lan truyền tospovirus và các rào cản dạng màng mà virus
phải vƣợt qua. Các số tƣơng ứng với các loại màng và tế bào liên quan đến đƣờng đi của virus từ khi đƣợc
chích nạp đến khi đƣợc chích truyền. L, lumen (tuyến dẫn); PSg, primary salivary gland (tuyến nƣớc bọt sơ
cấp); s, cross sections of muscle (tiết diện cắt ngang cơ); VP, viroplasm (vị trí tái sinh và lắp ráp virus); DM,
dense mass (vị trí có màu đậm khi chụp dƣới kính hiển vi) (Hogenhout et al., 2008). 81 Hình 8-1. Các lớp gen kháng R và một số ví dụ mỗi lớp 85 Hình 8-2. Minh họa tính kháng rất cao và bền vững đối với PRSV của giống đu đủ Rainbow chuyển gen CP so với
giống không chuyển gen Sunrise tại Hawai (Golsalves et al., 2004) 87 Hình 8-3. Cơ chế câm gen của thực vật chống virus và các các acid nucleic lạ 88 Hình 8-4. Hiệu quả PTGS với các cấu trúc chuyển gen potato virus Y (PVY) khác nhau. Hiệu quả đƣợc tính là %
cây chuyển gen miễn nhiễm với PVY; gen virus (PVY-pro) là NIa; chuỗi loop là chuỗi unidA (GUS) dài ~800
nucleotides; intron là của gen Pdk từ cây Flaveria (Smith et al., 2000). 90 Hình 8-5 Tạo cây cà chua chuyển gen kháng TYLCV bằng công nghệ RNAi (Fuentes, 2006) 91 Hình 8-6 Hai loại tính kháng tạo đƣợc: tính kháng tập nhiễm hệ thống (SAR) và tính kháng hệ thống tạo đƣợc (ISR)
(Vallad & Goodman, 2004). 93 Hình 8-7 Cấu trúc hóa học của SA, INA và BTH 93 Hình 8-8. Cây thuốc lá xử lý BTH (Bion) (trái) ức chế biểu hiện triêu chứng bệnh trên thuốc lá do TSWV gây ra.
Đối chứng không xử lý (phải) có triệu chứng dữ dội (Mandal et al., 2007) 94 Hình 8-9 Sự hình thành chitosan từ chitin 95 Hình 8-10 Hiệu quả ức chế của oligochitosan đối với TMV trên thuốc lá. (A) đối chứng, (B)–(F): xử lý chitosan lần
lƣợt ở các nồng độ 1 ppm, 10 ppm, 25 ppm, 50 ppm và 100ppm (Yin et al., 2010). 96 Hình 9-1 Lây nhiễm nhân tạo trên cây chỉ thị thuốc lá và rau muối (ảnh trái, giữa). Triệu chứng đốm chết hoại do
Citrus psorosis virus (CPsV) trên cây rau muối C. amaranticolor (http://www.DPWeb.net) 100 Hình: 9-2 Sơ đồ (trái) và ảnh chụp hiển vi điện tử (phải) của thể vùi tế bào chất của potyvirus 101 Hình 9-3 Dụng cụ và vật liệu dùng để kiểm tra thể vùi virus thực vật bằng kính hiển vi quang học
(http://plantpath.ifas.ufl.edu/pdc/Inclusionpage/Florvirus.html) 102 Hình 9-4 Phát hiện bộ gen dsRNA của virus lùn sọc đen phƣơng Nam (Zhou et al., 2008) 106 Hình 9-5 Cấu trúc kháng thể (Hull, 2002) 108 Hình 9-6 Sơ đồ sản xuất kháng thể đơn dòng 109 Hình 9-7 Kỹ thuật Outerlony chẩn đoán virus. Các mẫu ABCDE là các mẫu dƣơng 110 Hình 9-8 Các kỹ thuật ELISA khác nhau 111 Hình 9-9 Phát hiện virus bằng kỹ thuạt dot blot miễn dịch (western blot, trái, Hull, 2002) và dot blot lai DNA
(southern blot, phải, Hsu et al., 2005) 112 Hình 11-1 Hình thái phân tử và tổ chức bộ gen của RTBV (Hull, 1996) 123 Hình 11-2 Cây lúa nhiễm các tổ hợp virus khác nhau và khả năng truyền virus qua rầy xanh đuôi đen (Azzam et al.,
2000) 123 Hình 12-1 Hình thái phân tử geminiviruses. A, Phân tử Tomato yellow leaf curl virus (TYLCV), bar = 100 nm
(Gafni, 2003). B, Mô hình phân tử Maize streak virus (MSV) (Zhang et al., 2001). 126 Hình 12-2 Tổ chức bộ gen của begomovirus và DNA vệ tinh (Hà Viết Cƣờng, 2008) 129 Hình 12-3 Bệnh khảm lá sắn tại châu Phi. B và E là triệu chứng bệnh; C là vector bọ phấn và F là cây chuyển gen
kháng bệnh (Legg & Fauquet, 2004). 130 Hình 12-4 Triệu chứng bệnh cuốn lá bông tại Pakistan. Chú ý sự hình thành lá phụ ở mặt sau của lá bông ở ảnh
dƣới bên phải (Bridon & Markham, 2000) 131 Hình 13-1 Tổ chức bộ gen của các virus thuộc họ Nanoviridae (Gronenborn, 2004) 136 Hình 13-2 Bụi chuối bị bệnh chùn ngon (tại Gia Lâm) và triệu chứng trên lá 137 Hình 14-1 Sơ đồ cấu trúc phân tử của các tobamovirus (Viralzone, 2009) 139 Hình 14-2 Hình thái virion và cấu trúc của TMV 140 Hình 14-3 Tinh thể virus TMV hình thành trong tế bào thuốc lá 141 Hình 14-4 Tổ chức bộ gen chính và 2 phân tử phụ genome của TMV 142 Hình 14-5 Triệu chứng do TMV gây ra trên thuốc lá (trái), ớt (giữa) và cà chua (phải) 143 Hình 15-1 Hình thái virion và thể vùi của potyvirus. Ảnh trái: phân tử PVY, bar = 100 nm (Shukla et al., 1998).
Hình phải: thể vùi tế bào chất của PVY trong tế bào thuốc lá (Arbatova et al. 1998). 146 Hình 15-2 Tổ chức bộ gen của các potyvirus (Shukla et al., 1998). 147 Hình 15-3 Triệu chứng bệnh trên đu đủ (CABI, 2006) và bầu bí (cây bí xanh tại Việt Nam) 151 Hình 15-4 Ba nhóm phả hệ PVY trên thế giới và Việt Nam (in đậm) 153 Hình 15-5 Các triệu chứng khảm, khảm nhăn, lùn, chết hoại lá và đốm vòng chết hoại trên khoai tây do PVY
(Kerlan, 2006; Hà Viết Cƣờng, 2005) 154 Hình 15-6 Mức đa dạng của BCMV ở Việt Nam (đƣợc in đậm). 157

11
Hình 15-7 Bệnh khảm lá do BCMV trên cây họ đậu Việt Nam (ảnh trái:đậu đen, ảnh giữa: muồng 3 lá, Hà Viết
Cƣờng et al., 2008) và triệu chứng chết hoại gân của bệnh ―thối đen‖ (CABI, 2006). 158 Hình 15-8 Bệnh khảm lá do SCMV trên mía (trái), ngô (giữa) và hoàng tinh (phải) tại Việt Nam 161 Hình 15-9 Phân tích phả hệ dựa trên gen CP của các mẫu SCMV và SrMV thu tại Việt Nam (đƣợc in đậm) với các
mẫu thế giới. 162 Hình 15-10 Các potyvirus trên hành ta và tỏi tây ở Việt Nam 163 Hình 15-11 TuMV trên cây cải củ (hàng trên) và cải bẹ (hàng dƣới) ở Việt Nam 164 Hình 15-12 Phân tích phả hệ các mẫu TuMV từ Việt Nam (in đậm) và thế giới dựa trên gen CP 165 Hình 15-13 Bệnh do DsMV trên cây họ Ráy 166 Hình 15-14 Triệu chứng bệnh do SPFMV trên khoai lang Việt Nam 167 Hình 15-15 Cây phả hệ dựa trên gen CP của các mẫu SPFMV Việt Nam và các mẫu thế giới 167 Hình 15-16 Triệu chứng do ZYMV trên cây bí ngồi (Desbiez & Lecoq, 1997) 168 Hình 15-17 Cây phả hệ của các mẫu ZYMV Việt Nam và thế giới dựa trên gen CP 169 Hình 15-18 Bệnh do ChiVMV trên ớt và thuốc lá 170 Hình 16-1 Cấu tạo và tổ chức bộ gen của rhabdovirus hại thực vật 174 Hình 16-2 Sơ đồ tổ chức bộ gen của RYSV 176 Hinh 16-3 Bệnh vàng lụi lúa tại miền Bắc năm 2010 do RYSV 176 Hình 17-1 Hình thái và cấu trúc phân tử TSWV 180 Hình 17-2 Sơ đồ quá trình tái sinh của TSWV 181 Hình 17-3 Một số bệnh do TSWV gây ra trên cà chua, thuốc lá và lạc 182 Hình 18-1 Cấu trúc của một số reovirus đại diện 185 Hình 18-2 Sơ đồ virion của RRSV (hàng trên) và ảnh EM (hàng dƣới) cho thấy virion hình cầu với mấu gai (trái)
và 2 lớp vỏ (phải), thanh bar 50 nm (Shikita, 1979; Milne, 1982) 186 Hình 18-3 Tổ chức bộ gen của RRSV 187 Hình 18-4 Một số triệu chứng điển hình của bệnh lúa lùn xoắn lá do RRSV (3 hình đầu, Hà Viết Cƣờng, 2009).
Hình ngoài cùng bên phải là một phần tế bào chất của tế bào nhu mô mạch phloem tại vị trí nốt phồng với
―viroplasm‖ màu đậm chứa đầy các phân tử virus (Hibino, 1979) 188 Hình 18-5. Thể vùi virus và phân tử virus SRBSDV trong tế bào cây bị bệnh lùn sọc đen tại Việt Nam năm 2009 190 Hình 18-6. Triệu chứng bệnh LSĐ trên lúa ở miền Bắc vụ mùa năm 2009. Lá lúa bị xoắn vặn (a, b, c), trỗ không
thoát với hạt bị bị biến màu nâu (a). Có các nốt phồng màu trắng chạy dọc gân của lá (d), phiến lá (d), thân
(e). Các nốt phồng lúc đầu trắng sau chuyển màu nâu đến đen (e, f). 191 Hình 18-7. Triệu chứng bệnh LSĐ trên ngô ở miền Bắc năm 2010. 191 Hình 19-1 Phân tử virus RGSV có dạng tràng hạt khép vòng (Hibino, 1986) 195 Hình 19-2 Tổ chức bộ gen của RGSV 195 Hình 19-3 Triệu chứng bệnh lúa cỏ do RGSV tại Cần Thơ 196 Hình 20-1 Tế bào phytoplasma trong tế bào mạch rây ký chủ (Lee et al., 2000) 198 Hình 20-2 Triệu chứng chổi phù thủy và rễ tóc do Ca. Phytoplasma asteris gây ra trên cà rốt, tre 200 Hình 21-1 Cấu trúc viroid. A) hình dạng chung của viroid. B) phân tử viroid dƣới kính hiển vi điện tử. C) cấu trúc
dạng hình gậy của các pospiviroid. D) cấu trúc dạng gần hình gậy của các avsunviroid. 203 Hình 21-2 Sơ đồ tái sinh của viroid (Daros et al., 2006) 205 Hình 21-3 Bệnh củ khoai tây hình thoi 205

12
PHẦN I: ĐẠI CƢƠNG

13
Chuơng 1. GIỚI THIỆU
1.1 TÓM TẮT NỘI DUNG
Nội dung chính của chƣơng là giới thiệu lịch sử nghiên cứu virus, các cột mốc khoa học chủ
chốt liên quan đến virus và bản chất cũng nhƣ nguồn gốc của virus.
1.2 LỊCH SỬ VIRUS
Cho tới cuối thế kỷ 19, khoa học vẫn chƣa biết tới virus. Tuy nhiên, văn học đã mô tả nhiều
triệu chứng giống bệnh virus trên thực vật. Tài liệu cổ nhất là một bài thơ tiếng Nhật thời hoàng
đế Koken (năm 752 sau Công nguyên) mô tả hiện tƣợng vàng lá trên một cây cỏ dại, sau này
đƣợc xác định là cây bội lan (Eupatorium lindleyanum). Ngày nay, các nhà khoa học biết rằng
cây này mẫn cảm với nhiều virus thực vật thuộc chi Begomovirus (chẳng hạn Eupatorium yellow
vein virus, EYVV). Các begomovirus gây hiện tƣợng vàng gân và cuối cùng vàng lá trên cây bội
lan.
Ở Tây Âu, vào những năm 1600 -1660, nhiều tác phẩm nghệ thuật đã thể hiện bông hoa tulip
với triệu chứng khảm sọc rất đẹp. Một củ hoa tulip với triệu chứng khảm sọc rất có giá thời đó;
chẳng hạn củ của một giống hiếm nhƣ Semper Augustus có thể bán đƣợc 3000 guine (tiền Hà
Lan). Để so sánh, giá một con bò là 120 guine và một con tàu là 500 guine. Hiện tƣợng khảm
sọc hoa tuilip ngày nay đã đƣợc chứng minh là do một số virus thực vật, điển hình là Tulip
breaking virus (TBV) gây ra.
Hình 1-1 Triệu chứng bệnh virus trên hoa tulip. Ảnh trái, một bức tranh thế kỷ 17. Ảnh phải, hoa
tulip bị nhiễm TBV (Lesnaw & Ghabrial, 2000).
Vào cuối thế kỷ 19, kiến thức về bệnh vi khuẩn đã tích lũy khá nhiều và một thiết bị lọc vi
khuẩn đã đƣợc Chamberland (một trợ lý của Louis Pasteur) chế tạo. Thiết bị lọc này – cấu tạo
bằng sứ, có hình nến – đƣợc gọi là nến lọc Chamberland không cho phép các vi sinh vật có kích
thƣớc vi khuẩn đi qua. Năm 1882, Adolph Mayer đã mô tả một bệnh bí ẩn trên cây thuốc lá mà
ông gọi là bệnh khảm. Năm 1886, Mayer đã chứng tỏ rằng bệnh khảm thuốc lá có thể lan truyền
qua dịch cây và ông cho rằng tác nhân gây bệnh có lẽ là vi khuẩn. Năm 1892, Iwanowski đã
chứng minh rằng tác nhân gây bệnh khảm thuốc lá có thể truyền qua nến lọc vi khuẩn và ông giả
thiết rằng tác nhân gây bệnh có thể là chất độc do vi khuẩn tiết ra hoặc vi khuẩn có kích thƣớc

14
rất nhỏ để có thể qua lọc. Độc lập với Iwanowski, năm 1898, Beijerinck đã công bố kết quả
nghiên cứu thí nghiệm qua lọc với dịch cây thuốc lá bị khảm lá. Khác với Iwanowski và do
không phát hiện thấy bất cứ vi sinh vật nào, ông đã gọi tác nhân gây bệnh là contagium vivum
fluidum (tiếng Latin nghĩa là dịch sống truyền nhiễm‖ để phân biệt với các thực thể gây bệnh
dạng hạt khác. Đóng góp quan trọng của Beijerinck ở chỗ ông là ngƣời đầu tiên chứng minh sự
tồn tại của một loại tác nhân gây bệnh không phải vi khuẩn, có khả năng qua lọc, sống (nhân lên
trong cây bệnh) và hòa tan. Công trình của ông đƣợc xem là khai sinh nghành virus học và ông
đƣợc cộng đồng các nhà virus học công nhận là ―cha đẻ‖ của virus học.
Ngày nay, chúng ta biết rõ rằng tác nhân gây bệnh khảm thuốc lá là Tobacco mosaic virus
(TMV). Đây là một virus có ý nghĩa lịch sử lớn, cả trong virus học cũng nhƣ sinh học. Nhiều
phát minh quan trọng trong sinh học đều dựa trên virus này.
Hình 1-2 Ba nhà khoa học có đóng góp lớn về virus học. Từ trái sang phải: Adolf Mayer (ngƣời Hà
Lan, 1843-1942), Dimitrij Ivanovskij (ngƣời Nga, 1864-1920) và Martinus Beijerinck (ngƣời Hà
Lan, 1851-1931).
1.3 CÁC CỘT MỐC LỊCH SỬ QUAN TRỌNG KHÁC
Năm 1927, Dvorak cho thấy dịch cây nhiễm TMV và cây khỏe tạo ra kháng huyết thanh có
tính đặc hiệu khác nhau.
Năm 1935, Stanley đã kết tinh đƣợc tinh thể TMV nhƣng ông cho rằng đây là tinh thể chỉ có
bản chất protein (đoạt giải Nobel).
Năm 1936, Bawden & Pirie chứng minh đƣợc tinh thể TMV có bản chất nucleoprotein và
phân tử virus có hình gậy.
Năm 1931, Ruska & Knoll phát minh kính hiển vi điện tử.
Năm 1955-1956, Brakke phát triển kỹ thuật ly tâm gradient dùng siêu li tâm để tinh chiết
virus.
Năm 1956, Crick & Watson chứng minh vỏ protein của TMV gồm nhiều tiểu phần protein
xắp xếp theo vòng xoắn.
Năm 1977, Clack & Adam áp dụng kỹ thuật ELISA để chẩn đoán virus thực vật.
Năm 1980, Franck et al. lần đầu tiên giải mã toàn bộ bộ gen virus (Cauliflower mosaic
virus, CaMV).

15
Năm 1983, Kary Mullis phát minh kỹ thuật PCR (đoạt giải Nobel).
1.4 THUÂT NGỮ VIRUS
Về mặt lịch sử, theo từ điển Oxford English Dictionary, từ ―virus‖ đƣợc sử dụng vào thế kỷ
16 với nghĩa nọc độc đƣợc tiết ra bởi động vật độc, vào thế kỷ 18 với nghĩa bệnh lý là ―nguồn
gốc của sự ốm yếu hoặc là chất độc hình thành trong cơ thể do hậu quả của một số bệnh, đặc biệt
các bệnh có thể truyền sang ngƣời khác hoặc động vật khác‖. Cũng theo từ điển này, vào thế kỷ
19, từ ―virus‖ lại đƣợc sử dụng bởi các nhà vi khuẩn học với nghĩa vi khuẩn. Năm 1881, Paster
đã viết ―virus là ký sinh vi sinh vật, có thể nhân lên bằng nuôi cấy bên ngoài cơ thể động vật‖.
Theo nghĩa này, virus là vi khuẩn. Để phân biệt virus theo nghĩa vi khuẩn, các tác nhân gây bệnh
khảm thuốc lá và một số bệnh tƣơng tự trên động vật (ví dụ nhƣ bệnh dại) đƣợc gọi là các ―virus
qua lọc‖.
Từ 1900 – 1935, nhiều bệnh cây đƣợc cho do các virus qua lọc gây ra. Tuy nhiên, đã có nhiều
nhầm lẫn về bệnh do thiếu phƣơng pháp phân biệt virus. Chỉ tiêu ban đầu dựa vào khả năng qua
lọc. Ở một số bệnh có triệu chứng virus, mặc dù tác nhân gây bệnh không thể quan sát đƣợc
dƣới kính hiển vi (có khả năng qua lọc) nhƣng bệnh lại không truyền qua lây nhiễm cơ học. Các
bệnh này, ví dụ bệnh biến vàng cây cúc tây và củ hình thoi khoai tây, hiện đƣợc xác định là do
phytoplasma và viroid gây ra. Trong khoảng thời gian này, các nghiên cứu chủ yếu tập trung
vào mô tả bệnh, gồm triệu chứng, các biến đổi tế bào ký chủ, pham vi ký chủ và phƣơng thức
lan truyền. Cũng có một số cố gắng cải tiến lỹ thuật lọc nhằm xác định kích thƣớc virus. Các ảnh
hƣởng của nhiều tác nhân vật lý và hóa học đến sự xâm nhiễm virus cũng đƣợc nghiên cứu
nhƣng kỹ thuật khá nguyên thủy. Cho tới năm 1930, đã có rất nhiều nhầm lẫn nghiêm trọng về
bệnh virus và bản thân virus
1.5 ĐỊNH NGHĨA VIRUS
Nhờ các nghiên cứu cơ bản trong lĩnh vực sinh hóa, sinh học phân tử, di truyền, ngày nay,
bản chất virus đã đƣợc hiểu rất rõ. Có vô số khái niệm, định nghĩa nhằm giải thích thế nào là
một virus. Một trong các định nghĩa phản ánh đầy đủ bản chất virus đƣợc trình bày trong cuốn
Matthew‘s Plant Virology (Hull, 2002) là:
“Virus là các tác nhân gây bệnh không có cấu tạo tế bào, có bộ gen là acid nucleic
thƣờng đƣợc bao bọc bởi các protein vỏ, chỉ có thể tái sinh bên trong tế bào ký chủ. Quá
trình tái sinh của virus (i) phụ thuộc hoàn toàn vào bộ máy tổng hợp protein của ký chủ,
(ii) tổng hợp riêng rẽ các thành phần của virus để lắp ráp nên phân tử virus (virion) mới”.
Mặc dù các nhà virus học đồng ý rằng virus là một thực thể sinh học (và do đó thuộc đối
tƣợng nghiên cứu của sinh học) vì chúng có bộ gen, có thể tái sinh, có thể đột biến và tiến hóa
để thích nghi với điều kiện sống khác nhau thì vẫn chƣa có sự thống nhất xem liệu virus có thể
đƣợc coi là một sinh vật (organism) thực sự hay không. Hiện có 2 quan điểm khác nhau khi xét
bản chất sống của virus.
Quan điểm thứ nhất, cũng là quan điểm của Ủy ban Phân loại Virus Quốc tế (ICTV), cho
rằng virus, mặc dù có một số thuộc tính của sự sống, nhƣng không phải là một sinh vật thực sự
vì chúng vẫn thiếu một số thuộc tính cơ bản của một sinh vật sống nhƣ:
1. Virus không có khả năng thu nhận và lƣu trữ năng lƣợng tự do.
2. Virus không có chức năng sống khi ở bên ngoài tế bào ký chủ. Virus chỉ thể hiện đặc tính
của sự sống khi bộ gen của nó ở bên trong tế bào ký chủ thích hợp.

16
3. Virus không tăng trƣởng.
4. Virus không có bộ máy tổng hợp protein.
Quan điểm thứ nhất này dựa trên cơ sở cho rằng hệ thống sống đơn giản nhất là tế bào, do
vậy chỉ các sinh vật/visinh vật đơn bào hoặc đa bào mới có các đặc tính của sự sống còn các cơ
quan tử của nó thì không.
Quan điểm thứ hai coi virus là một sinh vật (chính xác là sinh vật không có cấu tạo tế bào)
dựa trên một số lập luận sau:
1. Virus có thể tái sinh.
2. Bộ gen của virus có thể bị đột biến.
3. Virus tiến hóa độc lập đối với ký chủ của chúng và có khả năng thích ứng với các ổ sinh thái
khác nhau.
4. Mặc dù virus phụ thuộc tế bào ký chủ để tái sinh (ký sinh chuyên tính) nhƣng nhiều loại tác
nhân gây bệnh khác cũng chỉ có thể sống đƣợc trong tế bào ký chủ.
5. Mặc dù virus thiếu ty thể nhƣng một số protozoa cũng thiếu.
6. Mặc dù kích thƣớc phân tử virus nhỏ nhƣng một số virus, chẳng hạn các virus thuộc nhóm
―nucleocytoplasmic large DNA viruses‖ nhƣ virus đậu mùa (pox virus) có kích thƣớc lớn
hơn vi khuẩn Chlamydia (gây bệnh viêm đƣờng tiết niệu).
7. Mặc dù virus có kích thƣớc bộ gen nhỏ nhƣng các virus thuộc nhóm ―nucleocytoplasmic
large DNA viruses‖ có kích thƣớc rất lớn. Ví dụ Acanthamoeba polyphaga mimivirus
(APMV), một virus mới đƣợc khám phá gần đây, có bộ gen lớn tới 1.2 Mb, lớn hơn bộ gen
của nhiều loài phytoplasma, riketsia và chlamydia.
8. Mặc dù virus không có hệ thống tái tạo năng lƣợng nhƣng vi khuẩn chlamydia cũng thiếu.
Lập luận chủ chốt nhất của quan điểm thứ hai là dựa vào đặc tính tái sinh của virus.
1.6 NGUỒN GỐC VIRUS
Vì đặc điểm bộ gen của virus rất đa dạng nên virus đƣợc xem là đa nguồn gốc (polythetic) và
virus DNA thì tiến hóa độc lập với virus RNA. Tuy nhiên vì chúng có một đặc điểm rất chung là
bộ gen lại đƣợc bao bọc bởi các protein vỏ nên hiện vẫn không có một quan điểm thống nhất về
nguồn gốc virus. Cho tới nay, đã có 3 giả thuyết về nguồn gốc virus.
1. Thuyết virus có trƣớc. Theo thuyết này, virus chính là dạng tồn tại hay là hóa thạch sống
của các dạng sống tiền tế bào.
Thuyết này đã bị loại bỏ từ lâu vì tất cả các virus đều là các ký sinh chuyên tính cao độ và chỉ
có thể tái sinh trong một tế bào ký chủ đang sống.
2. Thuyết suy thoái. Theo thuyết này, virus là dạng suy thoái của các sinh vật đơn bào.
Thuyết này cũng thƣờng bị phản bác vì 2 lý do:
(i) Khoa học chƣa từng khám phá ra bất kỳ một dạng trung gian nào giữa tế bào và virus.
(ii) Đối với một số dạng suy thoái từ tế bào nhƣ Mycoplasma/Phytoplasma là dạng suy thoái
của vi khuẩn, hay Microsporidia là dạng suy thoái của tế bào Eukaryote, hay Nanoarchaea là
dạng suy thoái của Archaea, thì tất cả chúng đều vẫn giữ đƣợc một số đặc trƣng của tế bào nhƣ
có ribosome cũng nhƣ bộ máy tổng hợp protein và năng lƣợng.

17
3. Thuyết trốn thoát (escape). Virus là các mảnh vật liệu di truyền của tế bào và bằng cách
nào đó thoát khỏi sự kiểm soát của tế bào và tiến hóa để trở thành các thực thể ký sinh độc lập
với bộ gen tế bào.
Thuyết này đã và đang trở nên phổ biến vì có nhiều bằng chứng ủng hộ nhƣ:
(i) Bộ gen virus có cả dạng DNA và RNA.
(ii) Một số virus có thể tổng hợp gen ký chủ vào bộ gen của nó (ví dụ begomovirus/thuốc lá).
(iii) Một số virus có thể tổng hợp vào bộ gen ký chủ (ví dụ các retrovirus, pararetrovirus).
(iv) Một số virus có đặc điểm bộ gen giống ký chủ (ví dụ geminivirus/plasmid,
potyvirus/mRNA…).
Tuy nhiên một số ý kiến khác không đồng ý khi cho rằng:
(i) Cơ chế nucleic acid của virus tiếp nhận và lắp ráp với protein vỏ vẫn không rõ.
(ii) Nếu theo thuyết này, thực khuẩn thể phải có nguồn gốc từ vi khuẩn và virus của
eukaryote phải có nguồn gốc từ eukaryote. Tuy nhiên ngƣời ta đã chứng minh một số gen thực
khuẩn thể T4 lại giống với gen của eukaryote hơn so với của vi khuẩn.
(iii) Cho tới nay, rất nhiều bộ gen của cả prokaryote lẫn eukaryote đã đƣợc giải trình tự
nhƣng phần lớn các gen virus lại chẳng giống chút nào với các gen ký chủ tƣơng ứng.
1.7 TẦM QUAN TRỌNG CỦA VIRUS THỰC VẬT
Cho tới nay (2010), hơn 2000 virus đã đƣợc phát hiện và công nhận, trong đó khoảng 1000 là
các virus gây hại thực vật. Các virus thực vật nhìn chung không làm chết cây nhƣng chúng ảnh
hƣởng nghiêm trọng đến sinh trƣởng, phát triển của cây, năng suất và chất lƣợng nông phẩm.
Nhiều trƣờng hợp, bệnh do virus gây có thể là một trong các nguyên nhân chính cản trở sản xuất
của một cây trồng nào đó. Một số ví dụ bệnh virus hại cây trồng quan trọng trên thế giới và đã
đƣợc xác định có ở Việt Nam bao gồm:
1. Bệnh xoăn vàng lá cà chua do nhiều begomovirus.
2. Bệnh chùn ngọn chuối do banana bunchytop virus (BBTV).
3. Bệnh đốm hình nhẫn đu đủ, bầu bí do papaya ring spot virus (PRSV).
4. Bệnh tàn lụi cây có múi do citrus tristeza virus (CTV).
5. Bệnh khảm lá khoai tây trên khoai tây do potato virus Y (PVY).
6. Bệnh khảm lá cây họ đậu do bean common mosaic virus virus (BCMV).
7. Bệnh tungro hại lúa nhƣ bệnh tungro do phức hợp 2 virus là rice tungro bacilliform
virus (RTBV) và rice tungro spherical virus (RTSV).
8. Bệnh vàng lùn (lúa cỏ) hại lúa do rice grasy stunt virus (RGSV).
9. Bệnh lùn xoắn lá (táp lá) hại lúa do rice ragged stunt virus (RGSV).
10. Bệnh lùn sọc đen hại lúa, ngô do southern rice black streaked dwarf virus (SRBSDV).
11. Bệnh vàng lụi (vàng tạm thời, vàng lá di động) trên lúa do rice yellow stunt virus
(RYSV).

18
1.8 CÂU HỎI ÔN TẬP CHƢƠNG 1
1. Virus đƣợc thực sự nghiên cứu từ bao giờ, gắn với bệnh gì?
2. Virus có ý nghĩa lịch sử đối với ngành virus học là gì?
3. Các nhà khoa học đã thực hiện những nghiên cứu đầu tiên về virus học.
4. Ví dụ 3 sự kiện khoa học ảnh hƣởng lớn đến nghiên cứu virus.
5. Vi dụ 3 dụng cụ có ảnh hƣởng lớn đến nghiên cứu virus.
6. Nguồn gốc của thuật ngữ ―virus‖.
7. Virus là gì?
8. Sự khác biệt cơ bản của virus đối với các tác nhân gây bệnh có bản chất tế bào (các
prokaryote và eukaryote) và phi tế bào (prion, viroid) là gì?
9. Sự khác biệt trong tranh luận về bản chất sống của virus là gì? Đâu là sự khác biệt cơ bản
nhất?
10. Các giả thuyết về nguồn gốc virus. Với bằng chứng khoa học hiện có thì khả năng nhất
virus có nguồn gốc từ đâu?
11. Ví dụ 3 virus có tầm quan trọng lớn trong nông nghiệp thế giới.
12. Ví dụ 3 virus có tầm quan trọng lớn đối với nông nghiệp Việt Nam.
1.9 TÀI LIỆU THAM KHẢO CHÍNH
1. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
2. Lesnaw, J. A. & Ghabrial, S. A. (2006). Tulip Breaking: Past, Present, and Future. Plant
Disease 84, 1052-1060.
3. Patrick, F. (2006). The origin of viruses and their possible roles in major evolutionary
transitions. Virus Research 117, 5-16.
4. Saunders, K., Bedford, I. D., Yahara, T. & Stanley, J. (2003). The earliest recorded
plant virus disease: Pathogenic DNA paints summer foliage gold, and inspired a poet
over a millennium ago. NATURE 422, 831.
5. van der Want, J. P. H. & Dijkstra, J. (2006). A history of plant virology. Archives of
Virology 151, 1467–1498.

19
Chƣơng 2. PHÂN LOẠI VÀ DANH PHÁP
2.1 TÓM TẮT NỘI DUNG
Nội dung chính của chƣơng là lịch sử và cơ sở khoa học của việc phân loại và đặt tên virus;
hệ thống tổ chức của cơ quan có thẩm quyền về mặt quốc tế đối với phân loại virus; cách phân
biệt khái niệm ―loài virus‖ và ―virus‖; cách viết tên virus và tên loài virus đúng; hệ thống phân
loại virus hiện nay.
2.2 PHÂN LOẠI VÀ DANH PHÁP
Phân loại học (Taxonomy =systemics): là khoa học phân loại gồm 2 nhánh là phân loại
―classification‖ và danh pháp ―nomenclature‖.
Phân loại là xắp xếp các các đơn vị phân loại ―taxon‖ vào các nhóm khác nhau theo mối
quan hệ của chúng dựa theo các tiêu chí xác định.
Danh pháp là đặt tên các đơn vị phân loại theo 1 qui tắc xác định.
Cả phân loại và danh pháp gắn liền với nhau và nhằm mục tiêu:
1. Tạo ra một sự sắp xếp có trật tự các virus sao cho con ngƣời có thể hiểu thấu đáo đƣợc
chúng.
2. Giúp con ngƣời có thể giao tiếp đƣợc với nhau về đối tƣợng virus nghiên cứu.
3. Giúp tiên đoán các đặc tính của một virus mới.
4. Bộc lộ mối quan hệ tiến hóa của các virus.
2.3 LịCH SỬ DANH PHÁP VÀ PHÂN LOẠI VIRUS
Khi virus đƣợc nghiên cứu lần đầu khoảng 1 thế kỷ trƣớc thì vào thời gian đó bản chất của
virus vẫn chƣa đƣợc biết: chúng không nuôi cấy đƣợc trên môi trƣờng nhân tạo, không quan sát
đƣợc dƣới kính hiển vi và chúng cũng không di chuyển qua màng lọc vi khuẩn. Do vậy, các nhà
virus học đầu tiên đã đặt tên loại tác nhân gây bệnh bí ẩn này bằng bệnh mà chúng gây ra gồm
tên cây ký chủ và triệu chứng đặc trƣng nhất, chẳng hạn virus khảm lá thuốc lá (tobacco mosaic
virus, TMV). Lúc này, virus đƣợc xem là một tác nhân gây bệnh ổn định có nghĩa trên một cây
ký chỉ cụ thể thì các bệnh khác nhau (dựa vào triệu chứng) sẽ do các virus khác nhau gây ra.
Tuy nhiên vào đầu những năm 1930, các nhà khoa học đã nhận thấy rằng: (1) các virus có thể
tồn tại dƣới dạng các chủng khác nhau và các chủng này có thể gây ra triệu chứng rất khác nhau
trên cùng một cây ký chủ; (2) các virus khác nhau có thể gây triệu chứng giống hệt nhau trên
cùng một cây ký chủ và (3) một bệnh trên một cây có thể do hai virus khác nhau gây ra.
Năm 1927, Johnson đề xuất rằng việc định tên một virus ngoài triệu chứng còn cần phải tính
đến các đặc điểm khác và đề nghị đặt tên virus bằng tên thông thƣờng (common name) của ký
chủ + từ virus + một số chỉ virus, ví dụ TMV đổi thành tobacco virus 1.
Năm 1935, Johnson và Hoggan đã sử dụng 5 đặc điểm sau để phân loại virus thực vật:
1. Phƣơng thức lan truyền.
2. Ký chủ tự nhiên và cây chỉ thị.
3. Thời gian tồn tại trong dịch chiết cây (longevity in vitro, LIV)

20
4. Ngƣỡng nhiệt độ mất hoạt tính (thermal death point, TDP).
5. Triệu chứng.
Dựa trên cách phân nhóm trên, khoảng 50 virus đã đƣợc xác định trong thời gian này.
Năm 1937, Smith đề xuất cách đặt tên và phân nhóm khác. Các virus đƣợc đặt tên và phân
nhóm theo tên khoa học của ký chủ mà trên đó virus đƣợc phát hiện đầu tiên. Nhƣ vậy TMV
đƣợc đổi thành Nicotiana virus 1. Theo hệ thống của Smith, có 15 virus thuộc nhóm Nicotiana
trong tổng số 51 virus. Với cách phân nhóm này, các virus rất khác nhau về nhiều đặc điểm cơ
bản lại đƣợc xếp vào cùng nhóm và do đó đây không phải là phân loại.
Năm 1939, Holmes đƣa ra một cách phân loại trong đó dựa chủ yếu vào phƣơng thức lan
truyền và phản ứng của ký chủ. Điểm mới là ông đã đƣa danh pháp ―tên kép La tinh‖ (Latin
binomial) để đặt tên virus giống nhƣ của các đối tƣợng khác trong phân loại sinh học. Theo cách
của ông, TMV trở thành Marmor tabaci (Marmor nghĩa là đá cẩm thạch theo tiếng la tinh, ám
chỉ triệu chứng khảm lá). Với hệ thống này, 53 trong tổng sô 89 virus đã đƣợc xếp vào chi
Marmor. Tƣơng tự nhƣ cách phân nhóm của Smith, nhiều virus thuộc chi Marmor có đặc điểm
cơ bản rất khác nhau.
Từ 1940 – 1970, một số hệ thống danh pháp virus dùng tên kép La tinh đã đƣợc đề xuất. Ví
dụ TMV đã từng đƣợc đặt là Musivum tabaci (1940), Phytovirus nicomosaicum (1941),
Nicotianavir communae (1942), Minchorda nicotianae (1957), Protovirus tabaci (1966),
Vironicotum maculans (1968), Virothrix iwanowskii (1970). Tuy nhiên việc sử dụng tên la tinh
cho virus đã bị phản đối trong thời gian dài, đặc biệt là từ các nhà virus học thực vật, vốn đi
trƣớc các nhà virus học động vật trong vấn đề phân loại virus. Lý do là họ không chấp nhận khái
niệm ―loài, species‖ đối với virus. Họ cho rằng khái niệm ―loài‖ đƣợc chấp nhận duy nhất là
―loài sinh học‖ đƣợc áp dụng cho các đối tƣợng có chung bể gen (gene pool) và biệt lập về sinh
sản. Ngoài ra theo họ, nếu chấp nhận một khái niệm loài đối với virus thì chắc chắn sẽ dẫn tới
việc la tinh hóa tên virus.
Nhƣ vậy, có thể thấy các cách phân loại và đặt tên virus ở trên chủ yếu dựa vào bệnh
chứ không phải dựa vào virus.
2.4 ỦY BAN PHÂN LOẠI VIRUS QUỐC TẾ (ICTV)
Năm 1966 tại Hội nghị Vi sinh vật học Quốc tế tổ chức tại Moscow, một Ủy ban Danh pháp
Virus Quốc tế (International Committee for the Nomenclature of Viruses) đã đƣợc thành lập với
nhiệm vụ phát triển một hệ thống phân loại và danh pháp đƣợc chấp nhận về mặt quốc tế và
thống nhất cho tất cả các loại virus.
Vào năm 1973, Ủy ban này đổi tên thành Ủy ban Phân loại Virus Quốc tế (International
Committee on Taxonomy of Viruses, ICTV).
Về mặt tổ chức, hiện nay, ICTV gồm khoảng 500 nhà virus hàng đầu trên thế giới, chia thành
5 ―tiểu ban‖ (subcommittee) chuyên môn (hình 2-1) là:
1. Virus hại động vật có xƣơng sống.
2. Virus hại động vật không xƣơng sống.
3. Virus hại thực vật.
4. Virus nhiễm vi khuẩn (thực khuẩn thể).
5. Virus hại nấm.

21
6. Một tiểu ban cơ sở dữ liệu.
Các tiểu ban chuyên môn lại bao gồm các ―nhóm nghiên cứu‖ (study-group), mỗi nhóm chịu
trách nhiệm đối với 1 họ virus hoặc 1 nhóm virus chƣa phân loại.
Cho tới nay, ICTV đã công bố 7 báo cáo trình bày hệ thống phân loại và danh pháp của tất cả
các đối tƣợng virus trên thế giới vào các năm 1971, 1976, 1979, 1982, 1991, 1995, 2000 và
2005.
Báo cáo lần thứ 7 (năm 2000) đã chính thức đƣa ra khái niệm ―loài‖ virus và qui định cách
viết tên loài. Báo cáo cũng liệt kê 1550 loài, 233 chi và 56 họ.
Báo cáo lần thứ 8 (năm 2005) đã phê chuẩn 3 bộ, 73 họ, 287 chi và hơn 5450 virus thuộc >
1950 loài.
Việc bổ sung và sửa đổi các vị trí phân loại đƣợc thực hiện thƣờng xuyên tại ICTV. ICTV chỉ
chịu trách nhiệm phân loại ở 4 mức là: bộ (order), họ (family), họ phụ (subfamily), chi (genus)
và loài (species). Đơn vị phân loại cơ bản là loài và không nhất thiết phải sử dụng cả 4 mức phân
loại. Phần lớn các loài đã đƣợc phân loại tới mức chi và phần lớn các chi đã đƣợc phân loại tới
mức họ. Chỉ một số họ đã đƣợc phân loại tới mức bộ. Theo báo cáo lần thứ 8 của ICTV, chỉ có 3
bộ đƣợc ghi nhận trong đó có một bộ bao gồm cả virus thực vật là bộ Monovirales. Ngoài ra
nhiều loài hiện mới chỉ đƣợc phân loại tới mức chi.
Ví dụ loài đã đƣợc phân loại tới mức bộ là:
Bộ Monovirales
Họ Rhabdoviridae
Chi Cyorhabdovirus
Loài Lettuce necrotic yellows virus
Ví dụ loài đã đƣợc phân loại tới mức họ là:
Họ Caulimoviridae
Chi Tungrovirus
Loài Rice tungro bacilliform virus
Ví dụ loài đã đƣợc phân loại tới mức chi là:
Chi Tenuivirus
Loài Rice grassy stunt virus
ICTV không có trách nhiệm trong phân loại và định tên các mức phân loại dƣới loài nhƣ
serotype, genotype, strain, variant và isolate. Việc phân loại ở mức dƣới loài là do các tác giả
nghiên cứu tự quyết định. ICTV cũng không có trách nhiệm phân loại và định tên các virus đƣợc
tạo ra bằng con đƣờng nhân tạo. Cần phải hiểu rõ rằng trong sinh học, đơn vị phân loại cơ bản
chính thức thấp nhất là loài (species) còn các mức phân loại dƣới loài nhƣ serotype, genotype,
strain, variant và isolate là các đơn vi phân loại không chính thức và không tƣơng đƣơng nhau
giữa các đối tƣợng nghiên cứu.

22
2.5 DANH PHÁP VIRUS HIỆN TẠI THEO ICTV
Tên virus. Tên virus thƣờng do các tác giả đặt theo trật tự sau: tên ký chủ đầu tiên virus đƣợc
phát hiện + triệu chứng điển hình + từ virus.
Cần chú ý là tên virus chính là tên thông thƣờng (common name) nên hiển nhiên là khác nhau
giữa các nƣớc. Tuy nhiên chúng ta thƣờng quen với tên thông thƣờng tiếng Anh, chẳng hạn nhƣ
tobacco mosaic virus. Do vậy, các tác giả phát hiện virus thƣờng đặt tên tiếng Anh cho virus.
Mặc dù ICTV không chính thức can thiệp vào việc đặt tên nhƣng trong một số trƣờng hợp
đặc biệt khi có rất nhiều virus thuộc các loài khác nhau cùng đƣợc phát hiện đầu tiên trên một
cây ký chủ với triệu chứng giống nhau thì các ―nhóm nghiên cứu‖ có thể hƣớng dẫn cách đặt
tên, chẳng hạn thêm tên địa phƣơng vào tên virus.
Ví dụ: Tomato yellow leaf curl Vietnam virus (TYLCVNV), tomato leaf curl Vietnam virus
(ToLCVV). Đây là 2 virus thuộc chi Begomovirus và gây bệnh xoăn vàng lá cà chua ở Việt
Nam.
Tên loài virus. Tên loài virus là tên virus tiếng Anh đƣợc viết (in) nghiêng. Ký tự đầu tiên
của tên loài phải viết hoa (không kể chữ hoa chỉ danh từ riêng nếu có trong tên loài). Tên loài
không chứa tên tác giả hoặc tên ngƣời. Danh pháp tên loài virus là danh pháp tên đơn
―monomial‖.
Ví dụ cách đặt tên loài virus:
Tên virus tiếng Anh là tobacco mosaic virus => tên loài là Tobacco mosaic virus
Tên virus tiếng Anh là tomato leaf curl Vietnam virus => tên loài là Tomato leaf curl
Vietnam virus
Một số điểm chú ý về tên loài:
Mặc dù tên loài là tên đơn nhƣng thƣờng gồm nhiều từ, ví dụ Tobacco mosaic virus.
Vì tên loài virus là tên virus tiếng Anh đƣợc viết (in) nghiêng nên dễ gây nhầm lẫn và đặc
biệt là không có sự khác nhau trong cách đọc. Để tạo ra sự khác nhau giữa tên virus và tên
loài virus, danh pháp ―tên kép‖ (binomial) tiếng Anh đã đƣợc đề xuất, chẳng hạn Tobacco
mosaic tobamovirus (trong đó từ tobamovirus là tên chi). Một ƣu điểm nữa của danh pháp
này là thông tin về chi đã đƣợc thêm vào tên loài (giống nhƣ tên loài của các đối tƣợng sinh
vật khác). Danh pháp tên kép đã đƣợc sự ủng hộ và sử dụng rộng rãi trong cộng đồng các
nhà virus thực vật. Một đề xuất chính thức đã đƣợc gửi lên ICTV vào năm 1998. Tuy nhiên
ICTV đã không chính thức công nhận danh pháp này vì 3 lý do: (1) nhiều nhà virus động vật
không ủng hộ do khi áp dụng danh pháp tên kép sang virus động vật đã tạo ra nhiều tên loài
rất dài và chứa nhiều từ trùng lặp chẳng hạn Human varicella-zoster varicellovirus hay
Influenza A influenzavirus A, (2) một số nhà virus học, những ngƣời cho rằng virus không
phải là sinh vật thực sự, phản đối vì theo họ danh pháp tên kép phải dùng tên La tinh, và (3)
tại thời điểm đó, việc thay đổi vị trí phân loại ở mức chi đã xảy ra khá phổ biến đối với nhiều
virus. Mới đây, vào năm 2002, ICTV đã tổ chức thăm dò ý kiến 250 nhà virus học hàng đầu
thế giới. Kết quả cho thấy 85 % đã ủng hộ cách sử dụng danh pháp tên kép tiếng Anh. Hiện
nay, ICTV đang xây dựng các qui định để chuyển sang sử dụng hệ thống danh pháp này.
Trong văn bản khoa học (luận văn, bài báo), tên loài chỉ xuất hiện một lần. Các thứ bậc phân
loại chính thức phải đƣợc viết (in) nghiêng và ký tự đầu tiên phải viết hoa, ví dụ: loài
Tobacco mosaic virus, chi Tobamovirus.

23
Về viết tắt. Vì tên virus thƣờng dài nên trong quá trình viết và nói, ngƣời ta hay dùng tên viết
tắt của virus, chẳng hạn TMV (tobacco mosaic virus), CMV (cucumber mosaic virus). Tên
loài virus thƣờng không cần viết tắt vì tên loài đầy đủ thƣờng chỉ xuất hiện 1 lần trong văn
bản khoa học (luận văn, bài báo).
2.6 PHÂN BIỆT VIRUS VÀ LOÀI VIRUS
Virus là thực thể cụ thể (tồn tại trong không gian và thời gian) => có thể thao tác với virus,
chẳng hạn tinh chiết, lây nhiễm…một virus.
Loài virus là khái niệm trừu tƣợng (chỉ tồn tại trong trí óc con ngƣời) => không thể thao tác
với ―loài virus‖.
Nhƣ vậy có thể thấy chỉ virus mới gây ra bệnh còn loài virus thì không gây bệnh.
2.7 KHÁI NIỆM LOÀI VIRUS
Trong sinh học, loài là đơn vị phân loại cơ bản thấp nhất. Hiện có 22 định nghĩa loài khác
nhau đƣợc đề xuất (Mayden, 1997) nhƣng chẳng có định nghĩa nào có thể áp dụng cho tất cả các
đối tƣợng. Một trong những khó khăn là thuật ngữ đƣợc sử dụng theo nhiều cách khác nhau và
không phải luôn đƣợc phân biệt rõ ràng. Một số định nghĩa chính là:
Loài kiểu hình. Định nghĩa loài kiểu hình, về măt lịch sử, có từ thời Hy Lạp cổ đại. Aristot
đã chia sinh giới tự nhiên thành các nhóm sinh vật rời rạc. Về sau, Linneaus đã củng cố khái
niệm này và xây dựng danh pháp tên kép (binomial). Một ví dụ của khái niệm loài kiều hình là:
―một loài là một nhóm các sinh vật tƣơng tự nhau và khác các nhóm khác‖ (Ridley, 1993). Định
nghĩa này hiện vẫn có ích trong phân loại, đặc biệt đối với thực vật.
Loài sinh học. Khái niệm loài nổi tiếng nhất do Mayr xây dựng lần đầu vào năm 1963: ―Loài
là các nhóm quần thể giao phối tự nhiên biệt lập về sinh sản với các loài khác‖. Định nghĩa này
đã bị chỉ trích vì nó chỉ có thể áp dụng cho các sinh vật sinh sản hữu tính. Năm 1982, Mayr đã
biến đổi định nghĩa này nhƣ sau ―Một loài là một cộng đồng các quần thể sinh sản (biệt lập) và
chiếm một ổ sinh thái tự nhiên‖. Khái niệm của Mayr còn đƣợc gọi là loài biệt lập, thƣờng đƣợc
áp dụng cho động vật nhƣng khó áp dụng cho các đối tƣợng khác.
Loài tiến hóa. Dựa trên lý thuyết tiến hóa, nhiều tác giả đƣa ra khái niệm loài tiến hóa: ―Một
loài tiến hóa là một dòng các quần thể có quan hệ tổ tiên – con cháu, khác với các dòng khác và
có xu hƣớng tiến hóa cũng nhƣ số phận lịch sử riêng‖ (Wiley, 1981). Bằng cách thêm ý tƣởng tổ
tiên – con cháu, khái niệm này đã bổ sung sự gắn kết nội tại giữa các thành viên trong cùng loài
mà sự gắn kết này không có ở các khái niệm loài dựa trên kiểu hình thuần túy.
Nhƣ vậy, không có một khái niệm loài nào có thể áp dụng cho tất cả các đối tƣợng sinh học.
Đối với một đối tƣợng, cần phải biến đổi các khái niệm hiện có. Ví dụ, một khái niệm loài tiến
hóa mới đƣợc đề xuất gần đây đối với vi sinh vật tiền nhân là ― Một nhóm các sinh vật đơn hình,
gắn kết về di truyền, có mức độ tƣơng đồng cao về về nhiều đặc điểm độc lập và có thể chẩn
đoán đƣợc nhờ một đặc tính kiểu hình rõ ràng‖ (Rosselló-Mora & Amann, 2001).
Định nghĩa loài virus. ICTV, nhƣ trình bày trong Báo cáo lần thứ 7 (2000), đã chấp nhận
định nghĩa loài áp dụng cho virus do Van Regenmorten đề xuất năm 1991: ―Một loài virus là
một lớp đa hình các virus tạo thành một dòng tái sinh và chiếm một ổ sinh thái đặc biệt‖ - ―A
virus species is a polythetic class of viruses that constitute a replicating lineage and occupy a
particular ecological niche‖

24
Một số thuật ngữ cần đƣợc giải thích:
1. Lớp (class). ―Lớp‖ trong định nghĩa này không phải là ―lớp‖ theo nghĩa thứ bậc phân loại
kiểu ―bộ-họ-lớp‖. Lớp ở đây cần đƣợc hiểu là một khái niệm trừu tƣợng chỉ tập hợp các
thực thể cùng chia sẻ một thuộc tính chung. Nếu một virus có bộ gen RNA sợi dƣơng, nó tự
động là thành viên của lớp các virus RNA sợi dƣơng. Đây đƣợc gọi là lớp chung vì nó đƣợc
xác định bởi một đặc tính chung hoặc một tập hợp các đặc tính có mặt ở tất cả các thành viên
của lớp.
2. Lớp đa hình. Lớp đa hình là một lớp mà các thành viên của nó có nhiều đặc điểm chung mặc
dù không có đặc điểm riêng biệt nào có mặt trong tất cả các thành viên của nó. Hậu quả là
không một đặc điểm duy nhất nào có thể đƣợc sử dụng làm chỉ tiêu phân biệt loài. Ví dụ,
một phản ứng ký chủ đặc biệt hoặc một mức độ tƣơng đồng chuỗi gen nào đó không thể
đƣợc sử dụng nhƣ là một chỉ tiêu tuyệt đối để phân biệt 2 loài virus của cùng một chi. Loài
không phải là một lớp chung có thế đƣợc xác định bởi một đặc tính riêng lẻ, do vậy chúng
khác các thứ bậc phân loại cao hơn, chẳng hạn chi và họ – là 2 khái niệm phân loại thuộc lớp
chung – bao gồm các thành viên cùng chia sẻ một hoặc vài các đặc tính chung cần và đủ để
phân biệt mối quan hệ.
Hình 2-1 Sơ đồ minh họa 5 thành viên của 1 lớp đa hình có 5 đặc điểm (Van Regenmortel, 2007)
3. Dòng tái sinh. Khái niệm này chỉ ra rằng các thành viên của cùng loài có cùng chung nguồn
gốc. Cũng cần chú ý rằng, chia sẻ chung nguồn gốc cũng là đặc điểm liên kết các thành viên
của cùng thứ bậc phân loại cao hơn, chẳng hạn chẳng hạn chi và họ. Các thành viên của loài
trải qua biến dị liên tục theo thời gian thông qua tái sinh bộ gen. Các biến dị tích luỹ và đạt
tới điểm mà các sai khác về kiểu hình và kiểu gen đủ để phán xét một thành viên của loài trở
thành một loài khác.
4. Ổ sinh thái. Khái niệm này chỉ ra các đặc trƣng sinh học của virus bao gồm phạm vi ký chủ,
quan hệ vector, tính gây bệnh, tính độc, tính hƣớng mô, môi trƣờng sống.
Khái niệm loài virus có một khác biệt cơ bản so với nhiều khái niêm loài khác là ở chỗ: các
khái niệm loài khác, điển hình là loài biệt lập, đƣợc xây dựng trên cơ sở cho rằng loài là một
khái niệm cụ thể, chỉ một thực thể khách quan, tồn tại độc lập với ý muốn con ngƣời; trái lại,
loài virus là một khái niêm trừu tƣợng, do con ngƣời tạo ra. Khác biệt này đã và đang là chủ đề
tranh luận nóng bỏng trong phân loại học.
2.8 CÁC CHỈ TIÊU PHÂN LOẠI VIRUS
Việc chấp nhận khái niệm loài virus bởi các nhà virus học quốc tế là một bƣớc quan trọng
nhằm thiết lập một hệ thống phân loại virus thống nhất nhƣng khái niệm này ít có giá trị trong
việc phân biệt liệu một mẫu virus cụ thể nào đó có là thành viên của một loài virus đƣợc xác
định trƣớc hay không. Lý do là ngƣời ta có thể định nghĩa các khái niệm trừu nhƣ loài nhƣng
không thể định nghĩa một mẫu virus cụ thể. Một mẫu virus chỉ có thể đƣợc đặt tên và xác định
bằng các đặc điểm chẩn đoán. Vì loài là một lớp đa hình, việc xác định này đƣợc dựa trên một

25
số các tiêu chí khác nhau qui định mức độ quan hệ của một mẫu virus với một loài đã đƣợc xác
định trƣớc (các đặc điểm chẩn đoán), thay vì chỉ vì dựa vào một chỉ tiêu đơn lẻ.
Để phân biệt ở mức loài, không cần thiết dùng các đặc điểm có mặt ở tất cả các thành viên
của chi hoặc họ, chẳng hạn nhƣ hình thái virion, tổ chức bộ gen, phƣơng thức tái sinh, số lƣợng
và kích thƣớc các protein cấu trúc và phi cấu trúc. Các chỉ tiêu dƣới đây (mà tầm quan trọng
thay đổi theo chi) thƣờng đƣợc áp dụng để phân biệt các mẫu virus ở mức loài.
1. Đặc điểm virion
1. Đặc điểm hình thái (hình dạng; kích thƣớc; có hoặc không có vỏ bọc; cấu trúc các đơn vị
hình thái capsomer)
2. Đặc tính vật lý (khối lƣợng phân tử; tỷ trọng; hệ số sa lắng; tính ổn định đối với pH,
nhiệt độ, cation, dung môi, bức xạ)
3. Đặc điểm bộ gen (loại acid nucleic (DNA hay RNA); loại sợi (đơn hay kép, mạch thẳng
hay mạch vòng); loại cực (âm, dƣơng hay lƣỡng cực); số phân đoạn (phân tử) của bộ
gen; đặc điểm đầu 5‘ và đầu 3‘; so sánh trình tự.
4. Đặc điểm protein (số lƣợng; kích thƣớc; chức năng; so sánh chuỗi)
5. Đặc điểm lipid (có hay không có; bản chất)
6. Đặc điểm carbonhydrate (có hay không có; bản chất)
2. Tổ chức bộ gen và phƣơng thức tái sinh
1. Tổ chức bộ gen.
2. Phƣơng thức tái sinh.
3. Đặc điểm phiên mã.
4. Đặc điểm dịch mã và xử lý hậu dịch mã.
5. Vị trí tích lũy protein virus, lắp ráp virion, thành thục và giải phóng virion.
6. Tế bào học, hình thành thể vùi.
3. Đặc điểm kháng nguyên
1. Quan hệ huyết thanh.
2. Bản đồ epitope.
4. Đặc điểm sinh học
1. Phạm vi ký chủ (tự nhiên và thực nghiệm).
2. Tính gây bệnh.
3. Tính hƣớng mô, bệnh học, mô học.
4. Phƣơng thức lan truyền ngoài tự nhiên.
5. Quan hệ vector.
6. Phân bố địa lý.
Ngay khi một loài đã đƣợc thiết lập trên cơ sở một tổ hợp các đặc điểm, ngƣời ta có thể xác
định một mẫu virus có là thành viên của loài đó không bằng cách đánh giá chỉ một vài đặc điểm
chẩn đoán. Ví dụ, nếu chuỗi gen của một mẫu là 99,9 % đồng nhất với một thành viên đã biết
của một loài thì mẫu đó là thành viên của loài đó.

26
2.9 HỆ THỐNG PHÂN LOẠI VIRUS HIỆN TẠI THEO ICTV
Hệ thống phân loại virus hiện nay là một hệ thống phân loại sinh học, đƣợc tổ chức thành
các thứ bậc phân loại khác nhau với các hậu tố qui định nhƣ sau:
Bộ (- virales)
Họ (- viridae)
Phân họ (- virinae)
Chi (- virus)
Loài (- virus)
Hiện nay (2009), ICTV đã phê chuẩn 2.288 loài virus đƣợc xếp vào 348 chi. Tuy nhiên,
không phải tất cả các loài đã đƣợc phân loại tới bộ. Trong tổng số loài, chỉ 549 loài (24%) đã
đƣợc phân loại đến bộ (6 bộ gồm 22 họ), còn lại 1739 loài (76%) mới chỉ đƣợc phân loại đến họ
(65 họ).
Đối với virus thực vật, hiện có khoảng ~ 1000 loài, trong đó khoảng 80 % là các virus RNA,
gồm đủ các nhóm đƣợc minh họa tóm tắt ở hình 2.2.
Bảng 2-1 Các bộ, họ virus theo phân loại của ICTV (dựa trên số liệu 2009)
Bộ Số họ Đặc điểm chính Ví dụ loài
Caudovirales 3 DNA sợi kép (thực khuẩn thể)
Enterobacteria phage T4
Herpesvirales 3 DNA, sợi kép Human herpesvirus 1
Mononegavirales 4 RNA, sợi đơn, cực âm, không phân mảnh, hại cả
động vật, thực vật
Measles virus
Rice yellow stunt virus
Nidovirales 3 RNA, sợi đơn, cực +, hại
động vật
Porcine reproductive and respiratory
syndrome virus
Picornavirales 5 RNA, sợi đơn, cực +, hại động vật, thực vật, côn
trùng
Hepatitis A virus
Sacbrood virus
Rice tungro spherical virus
Tymovirales 4 RNA, sợi đơn, cực +, hại thực vật
Potato virus X
Các virus không đƣợc xếp thành bộ
65 Rất đa dạng về đặc điểm bộ gen và ký chủ
Human immunodeficiency virus 1
Hepatitis B virus
White spot syndrome virus
Influenza A virus
Papaya ring spot virus
Tomato leaf curl Vietnam virus
Tham khảo website của ICTV (http://www.ictvonline.org) và Viralzone
(http://expasy.org/viralzone) để cập nhật hệ thống phân loại virus.

27
Hình 2-2 Minh họa phân loại các chi, họ virus thực vật theo báo cáo lần thứ 8 (2005) của ICTV

28
2.10 HỆ THỐNG PHÂN LOẠI VIRUS THEO BALTIMORE
Để đơn giản hóa phân loại virus, một hệ thống phân loại virus khác cũng thƣờng đƣợc sử
dụng gọi là hệ thống phân loại Baltimore. Hệ thống này do David Baltimore (giải Nobel 1975)
đề xuất năm 1974. Baltimore phân loại các virus thành 7 nhóm dựa trên đặc điểm bộ gen và cơ
chế tái sinh (Bảng 2.2, Hình 2.3).
Bảng 2-2 Hệ thống phân loại virus theo Baltimore
Nhóm Đặc điểm bộ gen Ví dụ virus thực vật
I DNA, sợi kép Các begomovirus
II DNA, sợi đơn Banana bunchy top virus
III RNA, sợi kép Rice ragged stunt virus
IV RNA, sợi đơn, cực (+) Papaya ring spot virus
V RNA, sợi đơn, cực (-) Rice yellow stunt virus
VI RNA, sợi đơn, cực (+), tái sinh qua trung gian DNA (Retrovirus)
Glycine max SIRE1 virus
VII DNA, sợi kép, tái sinh qua trung gian RNA
(Pararetrovirus) Rice tungro bacilliform virus
Hình 2-3 Sơ đồ phân loại virus theo Baltimore (http://expasy.org/viralzone/all_by_protein/230.html)

29
2.11 CÂU HỎI ÔN TẬP CHƢƠNG 2
1. Bản chất phân loại virus trƣớc những năm 1970. Lấy 2-3 ví dụ minh chứng.
2. Lịch sử hình thành, tổ chức và vai trò của ICTV.
3. Phân biệt 2 khái niệm ―virus‖ và ―loài virus‖.
4. Danh pháp virus hiện tại theo ICTV (tên virus, tên loài virus, các điểm chú ý; phân biệt
virus và loài virus). Lấy ví dụ 3 tên virus và tên loài virus tƣơng ứng.
5. Các cơ sở phân loại virus hiện tại. Đặc điểm nào quan trọng nhất?
6. Hệ thống phân loại virus hiện tại theo ICTV?
7. Số lƣợng loài virus (toàn bộ) đƣợc ghi nhận hiện tại. Mỗi nhóm virus động vật, virus
thực vật và thực khuẩn thể chiểm khoảng bao nhiêu phần trăm tổng số lƣợng loài?
8. Hệ thống phân loại virus thực vật hiện tại.
9. Đặc điểm chung nhất của các nhóm virus thực vật chính (DNA sợi đơn, DNA sợi kép
(phien mã ngƣợc), RNA sợi đơn cực (+), RNA sợi đơn cực (-), RNA phiên mã ngƣợc,
RNA sợi kép). Các ví dụ dại diện cho các nhóm virus chính (mức họ, mức chi, mức
loài).
10. Hệ thống phân loại Baltimor (số nhóm, đặc điểm nhóm, ví dụ). Ƣu nhƣợc điểm của hệ
thống này.
2.12 TÀI LIỆU THAM KHẢO CHÍNH
1. Hull, R. 2002. Matthews‘ Plant Virology. Fourth edition
2. Mayden, R.L. (1997). A hierarchy of species concepts: the denouement in the saga of
the species problem. In: M.F. Claridge, H.A. Dawah and M.R. Wilson (eds) ―Species:
The Units of Biodiversity‖. Chapman and Hall, London, 381–424.
3. Mayr, E. (ed) (1963). Animal Species and Evolution. Harvard University Press,
Cambridge, Mass.
4. Mayr, E. (ed) (1982). The Growth of Biological Thought. ―Diversity, Evolution, and
Inheritance‖. Harvard University Press, Cambridge, Mass.
5. Rosselló-Mora, R. and Amann, R. (2001). The species concept for prokaryotes. FEMS
Microbiology Reviews. 25, 39-67
6. Ridley, M. (1993). Evolution. Blackwell Scientific Publications. Boston, MA.
7. Van Regenmortel, M.H.V. (1990).Virus species, a much overlooked but essential
concept in virus classification. Intervirology. 31, 241–254
8. Wiley, E.O. (ed.) (1981). ―Phylogenetics: The Theory and Practice of Phylogenetic
Systematics‖. Willey, New York.
9. Van Regenmortel, M.H.V., 2001. Perspectives on binomial names of virus species.
Arch. Virol. 146, 1637–1640.
10. Van Regenmortel, M.H.V., 2003. Viruses are real, virus species are man-made
taxonomic constructions. Arch. Virol. 148, 2481–2488.

30
11. Van Regenmortel, M.H.V., 2006. Virologists, taxonomy and the demands of logic.
Arch. Virol. 151, in press.
12. Van Regenmortel, M.H.V., 2007. Virus species and virus identification: Past and
current controversies. Infection, Genetics and Evolution .7: 133–144
13. Van Regenmortel, M.H.V., Fauquet, C.M., 2002. Only italicized species names of
viruses have a taxonomic meaning. Arch. Virol. 147, 2247–2250.
14. Van Regenmortel, M.H.V., Mahy, B.W.J., 2004. Emerging issues in virus taxonomy.
Emerg. Infect. Dis. 10, 8–13.
15. Van Regenmortel, M.H.V., Maniloff, J., Calisher, C.H., 1991. The concept of virus
species. Arch. Virol. 120, 313–314.
16. Van Regenmortel, M.H.V., Bishop, D.H.L., Fauquet, C.M., Mayo, M.A., Maniloff, J.,
Calisher, C.H., 1997. Guidelines to the demarcation of virus species. Arch. Virol. 142,
1505–1518

31
Chƣơng 3. HÌNH THÁI VÀ CẤU TRÚC VIRUS
3.1 TÓM TẮT NỘI DUNG
Chƣơng này chủ yếu cung cấp các khái niệm cơ bản về đặc điểm hình thái và cấu trúc của
phân tử virus.
3.2 MỘT SỐ THUẬT NGỮ
1. Tiểu phần (subunit): Là một protein đơn lẻ, đƣợc mã hóa bởi bộ gen virus, ở trạng thái cấu
trúc không gian, cấu tạo nên vỏ protein của virus
2. Đơn vị hình thái (capsomer): Là đơn vị cấu trúc bề mặt thấy đƣợc dƣới kính hiển vi điện
tử. Một đơn vị hình thái có thể đƣợc cấu tạo bởi 1 hoặc nhiều tiểu phần và đƣợc gọi là
monomer, dimmer, trimer, tetramer, pentamer, hexamer … tùy theo số lƣợng tiểu phần hợp
thành.
3. Vỏ protein (capsid): Vỏ protein bao quanh bộ gen virus.
4. Vỏ bọc (envelope): Gồm 2 phần: (i) một lớp màng kép lipid, thƣờng có nguồn gốc từ màng
tế bào hoặc màng cơ quan tử của tế bào ký chủ; (ii) các phân tử glycoprotein (thƣờng có
nguồn gốc từ virus) nhô lên trên bề mặt.
5. Phân tử virus (virion): Là cấu trúc hình thái đầy đủ của virus. Phân tử virus cũng đƣợc gọi
là hạt virus.
3.3 HÌNH THÁI VIRUS
Phân tử virus thực vật có kích thƣớc cực kỳ nhỏ, phần lớn trong phạm vi hàng chục tới hàng
trăm nm (10-9
m), nên chỉ có thể quan sát đƣợc bằng kính hiển vi điện tử (KHVĐT). Khi đƣợc
quan sát dƣới KHVĐT, phân tử virus thực vật thƣờng có hình thái ở các dạng sau:
1. Hình cầu (thực chất là hình cầu đa diện đối xứng): rất nhiều, ví dụ nanovirus, cucumovirus,
reovirus.
2. Hình cầu kép: chỉ có các geminivirus có hình dạng này.
3. Hình giống vi khuẩn: chỉ badnavirus, tugrovirus, iflavirus, alfamovirus.
4. Hình viên đạn: chỉ rhabdovirus.
5. Hình gậy: rất nhiều, ví dụ potexvirus, tobamovirus.
6. Hình sợi mềm: chỉ potyvirus, flexivirus, closterovirus.
7. Hình tràng hạt: chỉ tenuivirus.
3.4 VIRUS TRẦN VÀ VIRUS CÓ VỎ BỌC
Virus nói chung đƣợc chia làm 2 nhóm: virus trần (naked capsid virus) và virus có vỏ bọc
(enveloped virus).
Đa số virus thực vật và thực khuẩn thể thuộc nhóm virus trần. Phân tử virus chỉ đƣợc cấu tạo
bởi lõi acid nucleic và vỏ protein. Cấu trúc acid nuclecic + protein vỏ còn đƣợc gọi là
nucleocapsid.

32
Một số virus thực vật thuộc nhóm nhóm bền vững tái sinh (nhân lên trong tế bào vector; ví dụ
nhƣ tospovirus, reovirus và nucleo/cytorhabdovirus) và nhiều virus động vật có một vỏ bọc bên
ngoài. Bên trong vỏ bọc là nucleocapsid gồm bộ gen virus đƣợc bao bọc bởi lớp vỏ preotein. Vỏ
bọc đƣợc cấu tạo bởi (i) một lớp màng kép lipid, thƣờng có nguồn gốc từ màng tế bào hoặc
màng cơ quan tử của tế bào ký chủ; (ii) các phân tử glycoprotein (thƣờng có nguồn gốc từ virus)
nhô lên trên bề mặt.
Hình 3-1. Minh họa virus trần (hàng trên) và virus có vỏ bọc (hàng dƣới)
3.4.1 Ví dụ virus thực vật có vỏ bọc
Trong khi phần lớn virus động vật là các virus có vỏ bọc thì chỉ có một số nhóm virus thực
vật có vỏ bọc. Đây là các virus thƣờng truyền qua vector côn trùng theo kiểu bền vững tái sinh
(tức là virus nhân lên trong tế bào vector).
Một ví dụ là các virus thuộc họ Bunyaviridae. Phần lớn bunyavirus gây bệnh trên động vật
(ví dụ hantavirus, nairovirus và phlebovirus) nhƣng một số hại thực vật nhƣ các virus thuộc chi
Tospovirus, điển hình là Tomato spotted wilt virus (TSWV).
Các tospovirus (cũng nhƣ các bunyavirus khác) có cấu trúc phân tử (virion) giống nhau.
Phân tử virus có hình cầu, kích thƣớc 80 – 120 nm và có lớp vỏ bọc cấu tạo bởi:
- Một lớp màng kép lipid dày ~ 5 nm, có nguồn gốc từ màng tế bào ký chủ.
- Hai loại gai (mấu) nhỏ cấu tạo bằng glycoprotein nhô trên bề mặt với phần cuống gai
nằm chìm trong lớp màng kép lipid. Độ dày của lớp gai ~ 5-10 nm. Hai loại gai này là 2
loại phân tử glycoprotein, ký hiệu là Gn và Gc (còn đƣợc ký hiệu là G1 và G2).
Bên trong là 3 phân tử nucleoprotein cấu tạo bởi RNA genome liên kết với các phân tử
protein (các phân tử protein này đƣợc gọi là các nucleoprotein) và một số enzyme phiên mã
transcriptase.
Hình 3-2 Sơ đồ cấu trúc phân tử TSWV (và các bunyavirus khác) với cấu trúc có vỏ bọc

33
3.5 CẤU TRÚC PHÂN TỬ VIRUS
Về cấu trúc, các phân tử virus thực vât (và cả các virus ký sinh động vật, nấm, vi khuẩn) gồm
2 nhóm chính: (i) cấu trúc khối đa diện đối xứng dạng icosahedral và (ii) đối xứng xoắn. Ngoài
ra, một số virus động vật và thực khuẩn thể có cấu trúc phân tử phức tạp và đƣợc xếp vào nhóm
(iii) gọi là nhóm không đối xứng
3.6 CẤU TRÚC DẠNG KHỐI ĐA DIỆN ĐỐI XỨNG (ICOSAHEDRON)
3.6.1 Cấu trúc dạng icosahedron
Các virus thực vật có cấu trúc phân tử loại icosahedron rất phổ biến. Icosahedron là một hình
đa diện cấu tạo bởi 20 tam giác đều, và do đó có 12 đỉnh và 30 cạnh. Cấu trúc này dẫn tới
icosahedron có 3 trục đối xứng. Trục đối xứng 3 lần là trục xuyên tâm tam giác; trục đối xứng 5
lần là trục xuyên đỉnh và trục đối xứng 2 lần là trục xuyên trung điểm của cạnh. Cấu trúc khối đa
diện đối xứng bắt gặp ở các virus có hình thái phân tử dạng hình cầu, hình cầu kép, hình giống
vi khuẩn và hình viên đạn.
Vì phải cần tối thiểu 3 tiểu phần để tạo thành một tam giác nên một hình icosahedron sẽ phải
cần tối thiểu 3 x 20 = 60 tiểu phần. Chỉ một vài loại virus có số lƣợng tiểu phần = 60 (ví dụ nhƣ
các nepovirus hại thực vật). Ngoài ra, tại mỗi đỉnh, các tiểu phần thƣờng lắp ráp với nhau thành
các pentamer (5 tiểu phần) nhô lên rõ rệt (Hình 3.3).
Hình 3-3 Cấu trúc đối xứng đa diện icosahedron cơ bản của virus (T=1, số tiểu phần = 60)
3.6.2 Số T
Để biểu diễn số lƣợng và cách sắp xếp của các tiểu phần protein, ngƣời ta sử dụng số T dựa
theo lý thuyết ―gần đồng đều – quasi-equivalence‖ do Caspar và Klug đề xuất năm 1962. Lý
thuyết cho rằng khi số tiểu phần = 60 thì các tiểu phần sẽ sắp xếp và chiếm các vị trí đồng đều
(dạng cơ bản). Tuy nhiên, khi số tiểu phần >60 thì các tiểu phần sẽ xắp xếp và chiếm các vị trí
gần đồng đều. Số T là số tam giác phụ trên tam giác chính của hình icosahedron (Hình 3.4).
Hình 3-4. Hai hình tam giác đều với số số tam giác phụ T = 4 (trái) và T = 9 (phải)
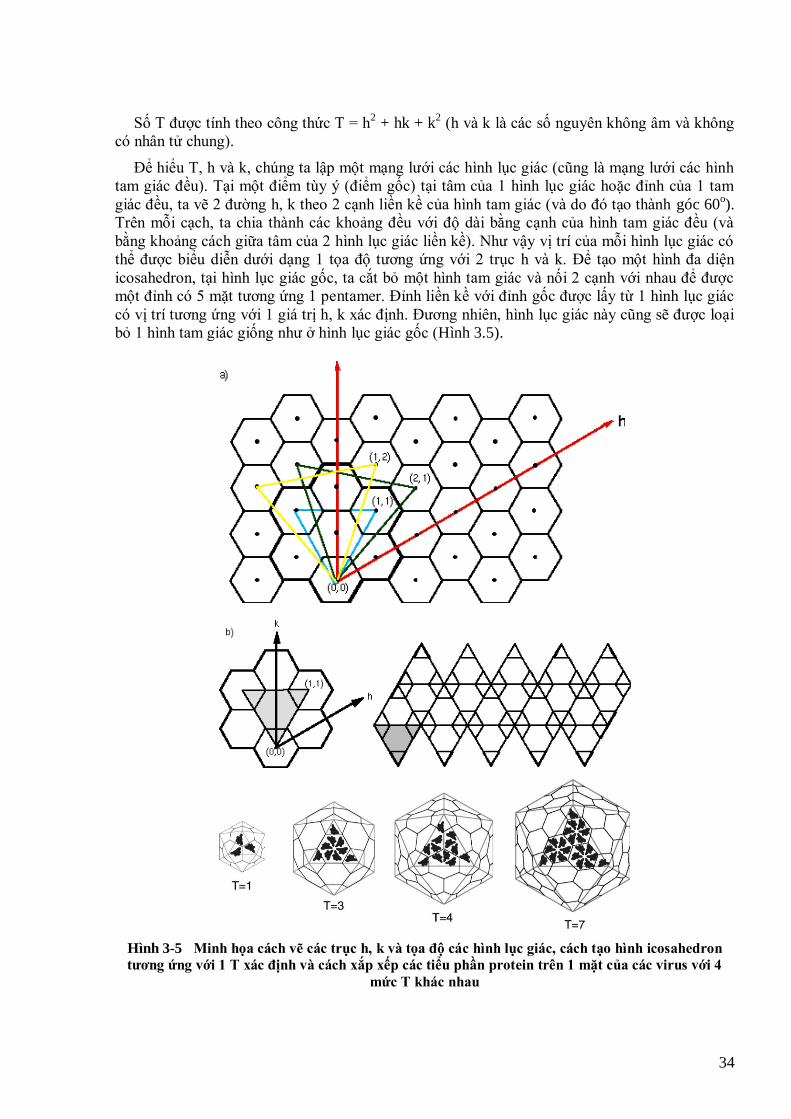
34
Số T đƣợc tính theo công thức T = h2 + hk + k
2 (h và k là các số nguyên không âm và không
có nhân tử chung).
Để hiểu T, h và k, chúng ta lập một mạng lƣới các hình lục giác (cũng là mạng lƣới các hình
tam giác đều). Tại một điểm tùy ý (điểm gốc) tại tâm của 1 hình lục giác hoặc đỉnh của 1 tam
giác đều, ta vẽ 2 đƣờng h, k theo 2 cạnh liền kề của hình tam giác (và do đó tạo thành góc 60o).
Trên mỗi cạch, ta chia thành các khoảng đều với độ dài bằng cạnh của hình tam giác đều (và
bằng khoảng cách giữa tâm của 2 hình lục giác liền kề). Nhƣ vậy vị trí của mỗi hình lục giác có
thể đƣợc biểu diễn dƣới dạng 1 tọa độ tƣơng ứng với 2 trục h và k. Để tạo một hình đa diện
icosahedron, tại hình lục giác gốc, ta cắt bỏ một hình tam giác và nối 2 cạnh với nhau để đƣợc
một đỉnh có 5 mặt tƣơng ứng 1 pentamer. Đỉnh liền kề với đỉnh gốc đƣợc lấy từ 1 hình lục giác
có vị trí tƣơng ứng với 1 giá trị h, k xác định. Đƣơng nhiên, hình lục giác này cũng sẽ đƣợc loại
bỏ 1 hình tam giác giống nhƣ ở hình lục giác gốc (Hình 3.5).
Hình 3-5 Minh họa cách vẽ các trục h, k và tọa độ các hình lục giác, cách tạo hình icosahedron
tƣơng ứng với 1 T xác định và cách xắp xếp các tiểu phần protein trên 1 mặt của các virus với 4
mức T khác nhau

35
Vì h, k có thể nhận giá trị 0 hoặc nguyên dƣơng (nhƣng không có nhân tử chung) nên T sẽ chỉ
nhận các giá trị nhƣ 1, 3, 4, 7, 9, 12, 13…
Đối với các virus icosahedral, ngƣời ta giả thiết có 12 pentamer (ở đỉnh) và các hexamer với
số lƣợng thay đổi tùy theo T. Số lƣợng và loại các đơn vị hình thái này đƣợc tính theo công thức
sau:
M = [(60T – 60)/] hexamer + 60/5 pentamer = 10 (T-1) hexamer + 12 pentamer
Ví dụ hình 3-6 ở dƣới cho thấy 2 phân tử virus với T khác nhau sẽ có có các đơn vị hình thái
khác nhau. Virus với T = 1 (hình trái ) => có 12 pentamer (tổng = 60 tiểu phần protein); với T =
4 (hình phải) => có 30 hexamer + 12 pentamer (tổng = 240 tiểu phần protein).
Hình 3-6 Minh họa virus với T khác nhau sẽ có các đơn vị hình thái (capsomer) khác nhau
3.6.3 Ví dụ virus với T khác nhau
Ví dụ 1. Banana bunchy top virus (BBTV), T = 1. Virus có bộ gen phân đoạn, ít nhất sáu
phân tử DNA dạng vòng, sợi đơn. Mỗi phân tử DNA genome đƣợc lắp ráp trong 1 virion. Virion
có T=1, 12 pentamer, với số lƣợng tiểu phần = 60.
Hình 3-7 Hình thái Banana bunchy top virus (BBTV)
Ví dụ 2: Tomato bushy stunt virus (TBSV) với T = 3. Mỗi phân tử virus chỉ có một phân tử
RNA (~ 4.8 kb). Về cấu trúc, virus có T = 3, 20 hexamer, 12 pentamer, với số lƣợng tiểu phần
=180 (387 aa; 41 kD).
Hình 3-8 Hình thái Tomato bushy stunt virus (TBSV)

36
3.7 CÁC PHÂN TỬ VỚI KHỐI ĐA DIỆN ĐỐI XỨNG DÀI
Một số phân tử virus chẳng hạn nhƣ các virus thuộc nhóm badnavirus (ví dụ RTBV), một số
thực khuẩn thể lại có cấu trúc dạng đa diện đối xứng kéo dài. Trong cấu trúc này, 2 nửa đầu và
đáy của khối đa diện đƣợc giữ nguyên nhƣng phần thân lại kéo dài (hình 3.9).
Đối với các cấu trúc kéo dài, ngƣời ta sử dụng số Q cùng với số T để xác định số tiểu phần
(N) theo công thức: N = 30 (T + Q), trong đó Q có thể là bất kỳ số nguyên >0.
Nếu Q = T => phân tử virus hoàn toàn đối xứng kiểu isohedron
Nếu Q > T => phân tử virus dạng khối đối xứng dài
Nếu Q < T => phân tử virus dạng khối đối xứng bẹt. Tuy nhiên trƣờng hợp này hiếm.
Hình 3-9 Cách cấu tạo của khối đa diện đối xứng kéo dài (hình trên) và cấu trúc phân tử với T=13
nhƣng có các Q khác nhau (hình dƣới)
3.8 CẤU TRÚC ĐỐI XỨNG XOẮN
Cấu trúc đối xứng xoắn là cấu trúc của các virus có dạng hình gậy hoặc hình sợi (phần đuôi
của nhiều thực khuẩn thể cũng có cấu trúc tƣơng tự). Trong cấu trúc đối xứng xoắn, các tiểu
phần protein xắp xếp theo đƣờng xoắn xung quanh 1 trục. Nhìn chung, cấu trúc này giúp phân tử
virus có năng lƣợng tự do phù hợp dẫn tới phân tử có thể mềm dẻo chẳng hạn ở dạng sợi mềm
hoặc gấp lại nhƣng không bị đứt gẫy.
Một virus điển hình đƣợc nghiên cứu đầy đủ nhất về cấu trúc đối xứng xoắn là TMV. Một
phân tử TMV chỉ chứa một phân tử RNA (~ 6.4 kb) đƣợc bao bọc bởi một lớp vỏ gồm 2123 tiểu
phần protein (mỗi tiểu phần là một phân tử protein có kích thƣớc 158 amino acid (17.3 kD). Các
tiểu phần protein lắp ráp với RNA virus theo cấu trúc xoắn (Hình 3.10) với chi tiết nhƣ sau:
1. Ba nucleotide trên phân tử RNA liên kết với 1 tiểu phần protein.
2. Một đơn vị lặp hoàn hảo gồm 49 tiểu phần / 3 vòng xoắn và tạo ra độ dài 6.9 nm (tổng độ dài
phân tử ~ 300 nm).
3. Đƣờng kính phân tử là 18 nm còn đƣờng kính lõi rỗng là 4 nm.
Cần chú ý là TMV có thể tự lắp ráp thành phân tử hoàn chỉnh từ 1 hỗn hợp các tiểu phần
protein + acid nucleic trong ống nghiệm.
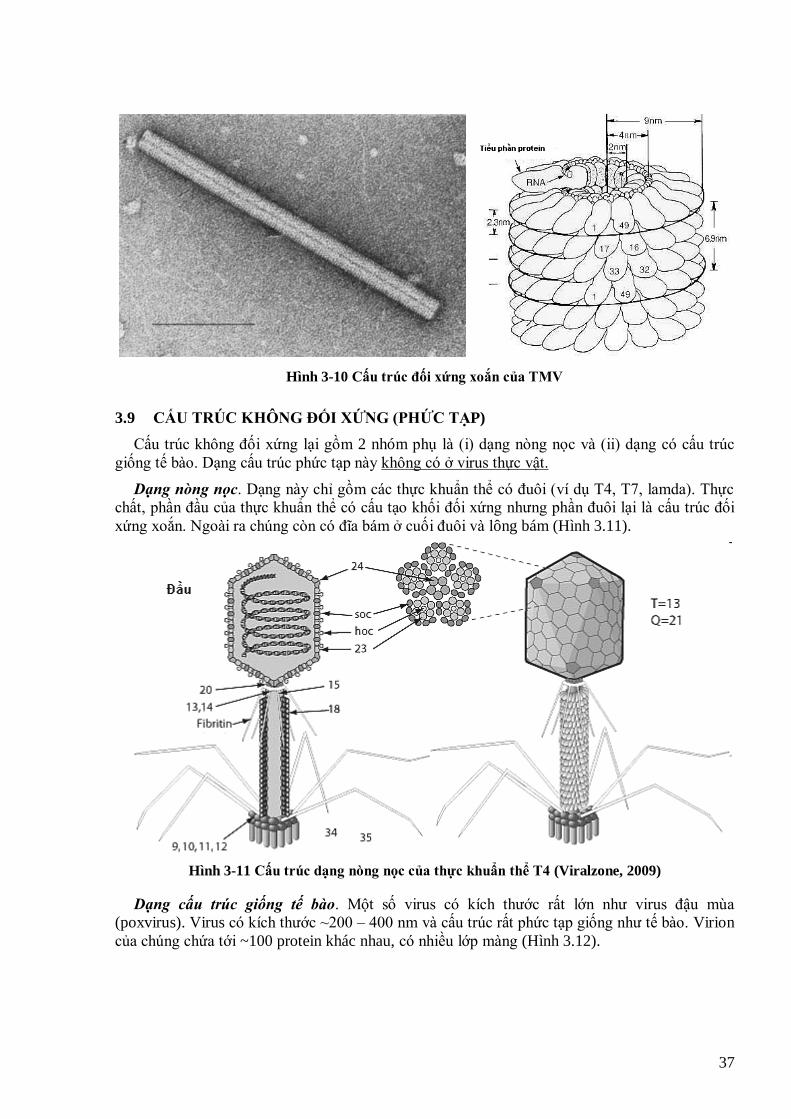
37
Hình 3-10 Cấu trúc đối xứng xoắn của TMV
3.9 CẤU TRÖC KHÔNG ĐỐI XỨNG (PHỨC TẠP)
Cấu trúc không đối xứng lại gồm 2 nhóm phụ là (i) dạng nòng nọc và (ii) dạng có cấu trúc
giống tế bào. Dạng cấu trúc phức tạp này không có ở virus thực vật.
Dạng nòng nọc. Dạng này chỉ gồm các thực khuẩn thể có đuôi (ví dụ T4, T7, lamda). Thực
chất, phần đầu của thực khuẩn thể có cấu tạo khối đối xứng nhƣng phần đuôi lại là cấu trúc đối
xứng xoắn. Ngoài ra chúng còn có đĩa bám ở cuối đuôi và lông bám (Hình 3.11).
Hình 3-11 Cấu trúc dạng nòng nọc của thực khuẩn thể T4 (Viralzone, 2009)
Dạng cấu trúc giống tế bào. Một số virus có kích thƣớc rất lớn nhƣ virus đậu mùa
(poxvirus). Virus có kích thƣớc ~200 – 400 nm và cấu trúc rất phức tạp giống nhƣ tế bào. Virion
của chúng chứa tới ~100 protein khác nhau, có nhiều lớp màng (Hình 3.12).

38
Hình 3-12 Cấu trúc phức tạp của virus đậu mùa
3.10 THÀNH PHẦN CẤU TẠO CỦA PHÂN TỬ VIRUS
3.10.1 Acid nucleic
Bộ gen có trong phân tử virus gồm DNA hoặc RNA, nhƣng nhƣng không bao giờ có cả 2
loại. Đa số virus thực vật (khoảng 80%) có bộ gen RNA. Bộ gen của virus có các đặc điểm sau:
Sợi đơn hoặc sợi kép
Mạch thẳng hoặc mạch vòng
Cực âm hoặc cực dƣơng hoặc lƣỡng cực
Phân đoạn hoặc không phân đoạn. Cần chú ý rằng tùy loại virus mà các phân đoạn đƣợc lắp
trong cùng một virion hay lắp ráp riêng biệt.
Phần lớn các virus thực vật có bộ gen RNA sợi đơn, cực dƣơng
3.10.2 Protein
Protein của virus nhìn chung đƣợc chia làm 2 loại: Protein cấu trúc (có mặt trong phân tử
virus) và protein phi cấu trúc (hình thành trong tế bào ký chủ trong quá trình gây bệnh). Các
protein cấu trúc chính của virus gồm các loại sau:
Protein vỏ (coat protein, CP): Có thể gồm 1 hay nhiều loại
Enzymes: Đối với các virus RNA thực vật, enzyme chủ yếu có mặt trên phân tử virus là
enzyme sao mã RNA (RdRp). Đối với các virus động vật, còn có thêm các enzyme cần cho
quá trình xâm nhập và nhận biết ký chủ (ví dụ neuramidase của virus cúm gia cầm). Đối với
thực khuẩn thể, virion của chúng còn có lysozim để giúp xâm nhập tế bào vi khuẩn.
Glycoproteins: Đây là các protein liên kết đƣờng, dạng protein xuyên màng, có vai trò quan
trọng đối với virus động vật và virus thực vật truyền qua vector theo kiểu bền vững tái sinh.
39
3.10.3 Lipid
Đối với các virus có vỏ bọc (một số virus thực vật, chẳng hạn các virus truyền qua vector
theo kiểu bền vững tái sinh và nhiều virus động vật) thì vỏ bọc của chúng đƣợc cấu tạo bởi lớp
màng kép phospholipid do lớp màng này có nguồn gốc từ màng tế bào hoặc màng cơ quan tử
của tế bào ký chủ.
3.10.4 So sánh cấu trúc các nhóm virus
Bảng 3-1. So sánh một số đặc điểm hình thái – cấu trúc của các nhóm virus
Đặc điểm Thực vật Động vật Vi khuẩn Nấm Protozoa
Kích thƣớc bộ gen RNA (kb) 0.3-28 5 – 30 5 – 8 2.5 – 28 5 – 10
Kích thƣớc bộ gen DNA (kb) 3 – 10 3 – 350 10 - 200 Không 180 – 1200
Số virus có vỏ bọc Ít Nhiều Ít Ít Tất cả
Mức độ phức tạp của bộ gen Ít Thay đổi Thay đổi Ít Thay đổi
Mức độ phức tạp về hình thái Ít Thay đổi Thay đổi Ít Thay đổi
Số virus có bộ gen phân mảnh Nhiều Một số Ít Một số Không
3.11 CÂU HỎI ÔN TẬP CHƢƠNG 3
1. Các thuật ngữ hình thái.
2. Các hình dạng của virus thực vật, ví dụ
3. Phân biệt virus trần và virus có vỏ bọc (đặc điểm, ví dụ)
4. Cấu trúc khối đa diện đối xứng (đặc điểm, ví dụ)
5. Ý nghĩa số T, ví dụ virus có T=1, T=3.
6. Cấu trúc đối xứng xoắn (đặc điểm, ví dụ).
7. Đặc điểm bộ gien (loại acid nucleic; loại sợi: sợi đơn, sợi kép, dạng thẳng, dạng vòng; loại
cực: cực dƣơng, cực âm, lƣỡng cực; có phân mảnh hay không phân mảnh, đƣợc lắp ráp
riêng rẽ hay cùng đƣợc lắp ráp trong virion; ví dụ mỗi loại).
8. Phân biệt protein cấu trúc và phi cấu trúc; các dạng protein cấu trúc chính. Đặc điểm lipid
của virus có vỏ bọc. Các nhóm virus nào thƣờng có protein cấu trúc và nhóm nào không có,
lấy ví dụ.
3.12 TÀI LIỆU THAM KHẢO CHÍNH
1. Ariska, F. (2008). Virus Capsid Model. Tokyo Institute of Technology.
2. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
3. Zlotnick (2005). Theoretical aspects of virus capsid assembly. Journal of Molecular
Recognition.18 (6): 479-490
4. http://www.tulane.edu/~dmsander/WWW/335/335Structure.html
5. Viralzone (http://expasy.org/viralzone)

40
Chƣơng 4. TÁI SINH VIRUS (REPLICATION)
4.1 TÓM TẮT NỘI DUNG
Chƣơng này trình bày quá trình tái sinh của tát cả các nhóm virus. Quá trình tái sinh (sinh
sản) của virus gắn liền với tế bào ký chủ và phụ thuộc hoàn toàn vào bộ máy dịch mã của tế bào.
4.2 KHÁI NIỆM
Sự tái sinh (replication) hay sinh sản là sự hình thành phân tử virus mới từ phân tử virus ban
đầu. Sau khi xâm nhập vào tế bào ký chủ, sự tái sinh virus trải qua 4 giai đoạn chính:
1. Tháo vỏ để giải phóng bộ gen virus
2. Tổng hợp protein virus
3. Tổng hợp bộ gen virus mới
4. Lắp ráp phân tử virus
4.3 ĐẶC ĐIỂM TÁI SINH VIRUS
Sự tái sinh virus, mặc dù khác nhau tùy nhóm, nhƣng đều có đặc điểm chung sau:
1. Virus sử dụng vật liệu của tế bào ký chủ (amino acid, nucleotide) để tổng hợp protein và
acid nucleic của chính virus.
2. Virus sử dụng năng lƣợng của tế bào ký chủ (chủ yếu dƣới dạng các hợp chất cao năng
nhƣ ATP) để tổng hợp protein và acid nucleic của chính virus.
3. Virus sử dụng bộ máy tổng hợp protein của tế bào ký chủ (ribosome, tRNA và các
enzyme liên quan) để tổng hợp protein của virus. Quá trình tổng hợp sẽ dựa trên khuôn
mRNA của virus. Trong nhiều trƣờng hợp (đối với các virus RNA cực dƣơng) thì bản
thân bộ gen virus đóng vai trò nhƣ mRNA. Tất cả virus thực vật sử dụng ribosome 80S
của tế bào ký chủ.
4. Hầu hết các virus thực vật tổng hợp 1 hoặc 1 số enzyme cần thiết cho quá trình tổng hợp
bộ gen virus. Ví dụ:
Tất cả các virus RNA mã hóa RdRp (RNA-dependent RNA polymerase). RdRp là
một enzyme polymer hóa và có chức năng tổng hợp RNA trên khuôn RNA.
Các geminivirus (có bộ gen DNA sợi vòng đơn) mã hóa Rep (replication) protein.
Rep không phải là một enzyme có chức năng polyme hóa nhƣng có chức năng cắt và
nối các phân tử DNA virus trong quá trình tổng hợp sợi DNA virus.
Tóm lại, sự tái sinh virus phụ thuộc hoàn toàn vào bộ máy tổng hợp protein và acid nucleic
của tế bào ký chủ. Sở dĩ nhƣ vậy là do virus nói chung và virus thực vật nói riêng chỉ mã hóa
một số ít gen; ví dụ các begomovirus chỉ mã hóa 5 – 8 gen, các potyvirus chỉ mã hóa 10 gen.
4.4 TÁI SINH CỦA CÁC VIRUS CÓ BỘ GEN RNA SỢI (+)
Phần lớn virus thực vật có bộ gen RNA sợi dƣơng (xem phần phân loại). Các bƣớc chủ chốt
của sự tái sinh của tất cả các virus thuộc nhóm này (hình 4.2) bao gốm:

41
4.4.1 Tổng hợp protein virus.
Bộ gen RNA sợi (+) đóng vai trò nhƣ là mRNA. Vì virus không có các gen mã hóa các yếu tố
cần cho quá trình dịch mã và ribosome, chúng phải sử dụng bộ máy dịch mã của ký chủ để hoàn
thành bƣớc này. Trong số các protein virus đƣợc tổng hợp, một số có vai trò quan trọng trong
quá trình tái sinh bao gồm:
RNA-dependent RNA polymerase (RdRp). RdRp xúc tác sự tổng hợp sợi RNA mới trên
khuôn RNA.
Helicase. Enzyme này có chức năng tháo xoắn (tách sợi RNA kép thành sợi RNA đơn) trong
quá trình tái sinh bằng cách phá vỡ liên kết hydro. Tháo xoắn là quá trình cần năng lƣợng nên
các helicase cũng có chức năng thủy phân adenosine triphosphate (ATP).
Methyl transferase (MT). Enzym này có chức năng tạo mũ cho đầu 5‘ của bộ gene RNA của
một số virus giống nhƣ ở mRNA của ký chủ. Chỉ các virus yêu cầu một mũ Cap đầu 5‘
(m7G5‘) nhƣ furovirus và pecluvirus mới cần chức này.
4.4.2 Hình thành phức hợp tái sinh VRC (viral replication complex)
Phức hợp tái sinh bao gồm RNA virus, các protein virus hình thành ở bƣớc 1 và các protein
ký chủ cần thiết cho quá trình tái sinh. Vì RNA virus đóng vai trò nhƣ mRNA cho quá trình dịch
mã và là khuôn cho quá trình tái sinh nên cả 2 quá trình này thƣờng liên quan chặt chẽ với nhau.
Do vậy, cần phải có một cơ chế điều hòa để cho phép chuyển từ chế độ dịch mã sang chế độ tái
sinh và để tránh sự xung đột của các ribosome, di chuyển từ đầu 5‘ sang đầu 3‘, và RdRp, di
chuyển theo chiều ngƣợc lại.
Phức hợp tái sinh luôn liên quan tới các cấu trúc màng của tế bào ký chủ bao gồm: màng lƣới
nội chất, màng ngoài lục lạp, màng không bào, màng proxisome, màng ty thể và màng nhân.
Các màng này biến đổi, hình thành các cấu trúc dạng cầu rỗng, bên trong chứa phức hợp tái
sinh. Sự gắn kết giữa phức hợp tái sinh và màng đƣợc thực hiện nhờ một hoặc nhiều protein có
nguồn gốc virus hoặc ký chủ, đƣợc gọi là các protein mỏ neo. Các protein mỏ neo thƣờng tổng
hợp vào màng, có khả năng biến đổi màng và liên kết với phức hợp tái sinh thông qua các
protein cần cho sự tái sinh nhờ tƣơng tác protein – protein. Loại màng cấu tử tế bào tham gia lắp
ráp phức hợp tái tái sinh khác nhau tùy virus, chẳng hạn đối với TMV là màng lƣới nội chất, đối
với CMV là màng không bào…
4.4.3 Tái sinh bộ gen virus
Đầu tiên, RNA (+) tức sợi virus sẽ đƣợc dùng làm khuôn để tổng hợp sợi (-). Sự tổng hợp sợi
(-) đƣợc bắt đầu từ đầu 3‘ của sợi (+). Các virus RNA có cấu trúc rất đa dạng về đầu 3‘ gồm 3
loại:
Chứa 1 vùng không dịch mã 3‘UTR + 1 đuôi polyA (ví dụ các potyvirus, potexvirus)
Chứa 1 vùng không dịch mã 3‘UTR có cấu trúc giống tRNA (ví dụ tymovirus,
cucumovirus, tobamovirus)
Chứa 1 vùng không dịch mã 3‘UTR + 1 đuôi polyA (ví dụ closterovirus, luteovirus)
Tất cả các vùng không dịch mã 3‘UTR đều có cấu trúc thứ cấp và các dấu hiệu cho phép
RdRp và các protein cần thiết tiếp cận để tổng hợp sợi (-).
Trên khuôn sợi (-), quá trình tái bản sợi (+) tức bộ gen virus bắt đầu đƣợc thực hiện. Cần chú
ý rằng một khuôn sợi (-) đƣợc sử dụng để tổng hợp nhiều sợi (+).

42
Quá trình tái bản sẽ hình thành 2 loại phân tử RNA:
Loại tái bản RF (replicative form): là 1 phân tử RNA sợi kép hoàn hảo hình thành khi
tổng hợp sợi (-) trên khuôn sợi (+).
Loại trung gian tái bản RI (replicative intermediate): là các phân tử RNA sợi kép trung
gian hình thành trong quá trình tái bản sợi (+). Ở dạng RI, nhiều phân tử RNA sợi (+) sẽ
gắn trên 1 phân tử RNA sợi (-).
Hiện có 2 giả thuyết giải thích cơ chế tái bản sợi (+) của virus thực vật:
Cơ chế bán bảo thủ. Trên khuôn sợi (-), các phân tử RNA sợi (+) đƣợc tổng hợp sau sẽ thay
thế dần sợi RNA (+) đƣợc tổng hợp trƣớc đó (hình b phía dƣới).
Cơ chế bảo thủ. Trên phân tử RF sợi kép, quá trình tháo xoắn chỉ diễn ra ngắn ngủi tại vị trí
kéo dài đầu 3‘ của các sợi (+) mới đang đƣợc tổng hơp (hình 4.1 c).
Hình 4-1 Sơ đồ giải thích cơ chế tái bản sợi (+) trên khuôn sợi (-). (b) và (c) là cơ chế tổng hợp của
virus thực vật TMV, còn (a) là dạng trung gian RI của thực khuẩn thể Qβ (Buck et al., 1999)
4.4.4 Lắp ráp thành phân tử virion mới
Khi có đủ RNA sợi (+) và protein cấu trúc, chủ yếu là CP, các phân tử này tiến hành lắp ráp
tạo viron mới. Quá trình lắp ráp này cũng đƣợc thực hiện trong phức hợp tái sinh.
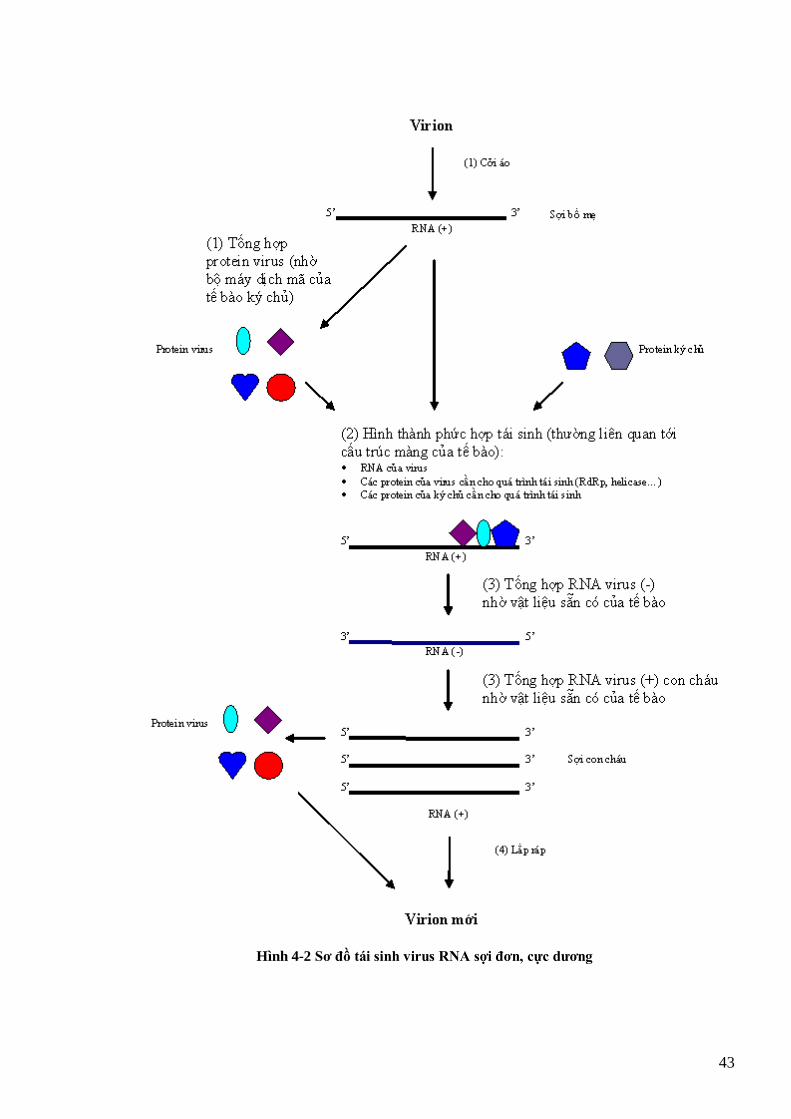
43
Hình 4-2 Sơ đồ tái sinh virus RNA sợi đơn, cực dƣơng

44
4.5 TÁI SINH CỦA CÁC VIRUS CÓ BỘ GEN RNA SỢI (-)
Điểm khác biệt cơ bản là sợi RNA genome của virus không đƣợc sử dụng để dịch mã. Về cơ
bản quá trình tái sinh của virus RNA sợi đơn cực (-) khá giống với các virus RNA sợi đơn cực
(+). Tuy nhiên có một số điểm khác biệt về trình tự, tóm tắt nhƣ sau:
1. Xâm nhập => tháo vỏ để giải phóng bộ gen virus sợi (-).
2. Sợi (-) đƣợc dùng làm khuôn để ―phiên mã‖ thành các mRNA của các gen virus và tiếp theo
tổng hợp các protein virus.
3. Sợi (-) đƣợc dùng làm khuôn để tổng hợp sợi (+) có kích thƣớc bằng sợi (-). Sợi (+) này sẽ
giữ vai trò làm khuôn để tổng hợp (-) sợi virus. Trên khuôn sợi (-) mới, các mRNA lại tiếp
tục đƣợc tổng hợp để tạo protein virus
4. Lắp ráp virion mới.
Cần chú ý 2 điểm khác biệt liên quan đến tái sinh các virus RNA sợi (-)
Mặc dù đƣợc ký hiệu là sợi (+) nhƣng sợi này không đƣợc sử dung trực tiếp để dịch mã
Đối với tất cả các virus RNA sợi âm thì trong thành phần virion của chúng bao giờ cũng có
RdRp để đảm bảo khi xâm nhập vào tế bào ký chủ, bộ gen phải đƣợc ―phiên mã‖ ngay sang
mRNA.
Hình 4-3 Sơ đồ tái sinh của rhabdovirus
Một số các họ virus quan trọng có bộ gen RNA sợi đơn, cực âm bao gồm:
a) Họ Rhabdoviridae. Đây là một họ lớn. Các rhabdovirus phần lớn gây hại động vật (ví dụ
Rabies virus, gây bệnh dại). Các rhabdovirus hại thực vật chỉ thuộc 2 chi là Nucleorhabdovirus
(ví dụ Rice yellow stunt virus (RYSV) gây bệnh vàng lụi lúa) và Cytorhabdovirus. Ngoại trừ các
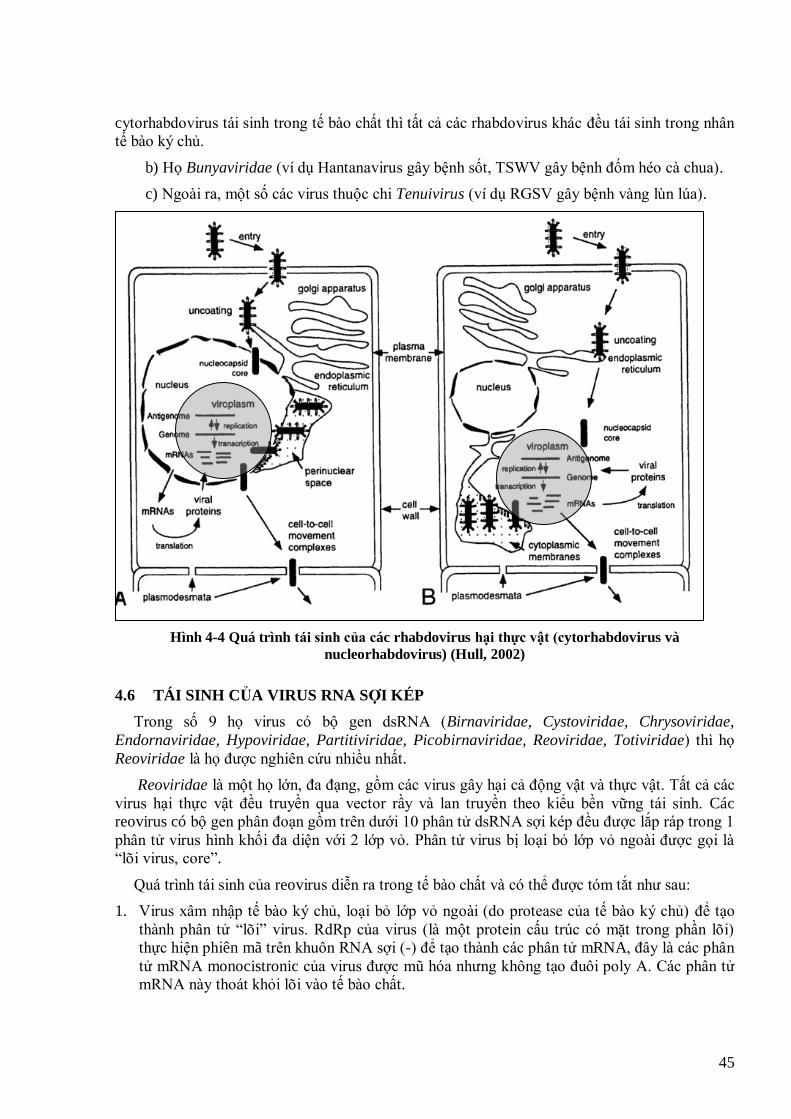
45
cytorhabdovirus tái sinh trong tế bào chất thì tất cả các rhabdovirus khác đều tái sinh trong nhân
tế bào ký chủ.
b) Họ Bunyaviridae (ví dụ Hantanavirus gây bệnh sốt, TSWV gây bệnh đốm héo cà chua).
c) Ngoài ra, một số các virus thuộc chi Tenuivirus (ví dụ RGSV gây bệnh vàng lùn lúa).
Hình 4-4 Quá trình tái sinh của các rhabdovirus hại thực vật (cytorhabdovirus và
nucleorhabdovirus) (Hull, 2002)
4.6 TÁI SINH CỦA VIRUS RNA SỢI KÉP
Trong số 9 họ virus có bộ gen dsRNA (Birnaviridae, Cystoviridae, Chrysoviridae,
Endornaviridae, Hypoviridae, Partitiviridae, Picobirnaviridae, Reoviridae, Totiviridae) thì họ
Reoviridae là họ đƣợc nghiên cứu nhiều nhất.
Reoviridae là một họ lớn, đa đạng, gồm các virus gây hại cả động vật và thực vật. Tất cả các
virus hại thực vật đều truyền qua vector rầy và lan truyền theo kiểu bền vững tái sinh. Các
reovirus có bộ gen phân đoạn gồm trên dƣới 10 phân tử dsRNA sợi kép đều đƣợc lắp ráp trong 1
phân tử virus hình khối đa diện với 2 lớp vỏ. Phân tử virus bị loại bỏ lớp vỏ ngoài đƣợc gọi là
―lõi virus, core‖.
Quá trình tái sinh của reovirus diễn ra trong tế bào chất và có thể đƣợc tóm tắt nhƣ sau:
1. Virus xâm nhập tế bào ký chủ, loại bỏ lớp vỏ ngoài (do protease của tế bào ký chủ) để tạo
thành phân tử ―lõi‖ virus. RdRp của virus (là một protein cấu trúc có mặt trong phần lõi)
thực hiện phiên mã trên khuôn RNA sợi (-) để tạo thành các phân tử mRNA, đây là các phân
tử mRNA monocistronic của virus đƣợc mũ hóa nhƣng không tạo đuôi poly A. Các phân tử
mRNA này thoát khỏi lõi vào tế bào chất.

46
2. Trong tế bào chất, các protein virus đƣợc dịch mã trên khuôn mRNA của virus.
3. Lắp ráp các các mRNA tức là các phân tử RNA sợi (+) của virus và các protein virus tạo
thành phân tử virus ―lõi‖ mới.
4. Trong phân tử ―lõi‖ mới, sợi RNA (+) của virus đƣợc phiên mã thành phân tử RNA sợi (-)
để tạo thành phân tử RNA sợi kép.
5. Trong phân tử ―lõi‖ mới, quá trình phiên mã mới trên khuôn sợi RNA (-) lại tiếp tục để tạo
các phân tử tạo mRNA (sợi +) mới.
6. Các phân tử ―lõi‖ tiếp tục lắp ráp với các protein cấu trúc khác để tạo phân tử virion hoàn
chỉnh
Nhƣ vậy có thể thấy 2 điểm khác biệt của quá trình tái sinh phân tử virus RNA sợi kép:
Quá trình tạo mRNA không phải bên ngoài tế bào chất mà đƣợc thực hiện trong phân tử
―lõi‖ của virus.
Khi bắt đầu lắp ráp, virus không phải lắp ráp các phân tử RNA sợi kép mà là các phân tử
RNA sợi đơn cực (-). Trong quá trình ―chín‖, các phân tử RNA sợi kép mới tiếp tục đƣợc
hoàn thiện.
Hình 4-5 Sơ đồ tái sinh virus RNA sợi kép

47
4.7 TÁI SINH CỦA VIRUS RNA QUA PHIÊN MÃ NGƢỢC (RETROVIRUS)
4.7.1 Các khái niệm
Transposons: là các chuỗi DNA trong bộ gen có thể tự di chuyển (transpose) từ vị trí này
sang vị trí khác trên bộ gen. Transposons là một trong nhiều loại yếu tố di động của bộ gen.
Ngƣời phát hiện ra trasposons đầu tiên là Barbara McClintock (giải Noben năm 1948 cho công
trình phát hiện transposon trên cây ngô). Tranposons đƣợc chia thành 2 nhóm:
DNA transposons: quá trình thay đổi vị trí đƣợc thực hiện thao hình thức ―cut/pate‖.
Retrotransposons: quá trình thay đổi vị trí theohình thức ―copy/pate‖ và đƣợc thực hiện qua 2
giai đoạn: (i) DNA phiên mã thành RNA => (ii) phiên mã ngƣợc lại thành DNA. Các phiên bản
copy DNA mới này đƣợc chèn vào các vị trí khác trên bộ gen.
Retrovirus: là các virus RNA có đặc điểm tái sinh bộ gen thông qua phiên mã ngƣợc nhờ
enzyme phiên mã ngƣợc Reverse transcriptase. Đối với các retrovirus, luôn có pha tổng hợp bộ
gen virus vào bộ gen ký chủ.
Các retrovirus động vật, ví dụ nhƣ các virus thuộc chi Lentivirus, họ Retrovirus nhƣ Human
immunodeficien virus (HIV) có bộ gen RNA sợi đơn, cực +, kích thƣớc khoảng 8-9 kb, đƣợc
bao quanh bởi 2 chuỗi LTR 0,2-1 Kb, có các gen nhƣ gag, pol, env. Ngoài ra các lentivirus cũng
chứa nhiều gen bổ trợ.
Các retrovirus thực vật (ví dụ các virus thuộc chi Sirevirus, họ Pseudoviridae nhƣ Glycine
max SIRE1 virus (GmaSIREV) cũng có đặc điểm giống các retrovirus động vật nhƣ: có bộ gen
RNA sợi đơn, cực +, kích thƣớc 9-10 kb, đƣợc bao quanh bởi 2 chuỗi lặp dài khoảng 0.5 - 1.2
kb ở đầu (LTR, Long Terminal Repeats), có 3 ORF mã hóa cho gen gag, pol và env. Các
sirevirus phát hiện thấy ở nhiều loài cây 1 lá mầm và 2 lá mầm.
Một số điểm chú ý về retrovirus thực vật: (i) cơ chế gây bệnh vẫn chƣa đƣợc hiểu rõ; (ii)
chúng đƣợc xem là các endovirus, có nghĩa bộ gen của chúng đƣợc tổng hợp trong bộ gen của
cây ký chủ và đƣợc lan truyền theo chiều dọc cùng với bộ gen của cây ký chủ qua các thế hệ
sau; (ii) virion chỉ hình thành trong quá trình tái sinh virus (phần lớn vẫn chƣa đƣợc nghiên cứu).
Hình 4-6 Tổ chức bộ gen của 1 retrovirus động vật (HIV) và của 1 retrovirus thực vật
(GmaSIREV)
LTR (Long Terminal Repeats) là các chuỗi đóng vai trò quan trọng trong quá trình dịch
chuyển vị trí của các retrovirus (và các yếu tố di động retroelements khác). Điển hình, một chuỗi
LTR thƣờng gồm gồm 3 phần:

48
Một vùng U3 khoảng 200-1.200 nucleotide chứa promoter.
Một vùng lặp R.
Một vùng U5 khoảng 75-250 nucleotides chứa phần đầu tiên của bộ gen.
Ngoài ra, liền kề phía hạ lƣu chuỗi LRT đầu 5‘ có một chuỗi PBS (Primer Binding Site)
khoảng 18 nucleotide là vị trí gắn mồi tRNA trong quá trình phiên mã ngƣợc; và liền kề phía
thƣợng lƣu chuỗi LRT đầu 3‘ có một chuỗi nhỏ ~10 A/G gọi là PPT (Polypurine Tract) là vị trí
bắt đầu tổng hợp sợi DNA (-).
Các retrovirus có một số các ORF mã hóa cho các protein đặc trƣng sau:
Gen gag mã hóa một polyprotein và tiếp theo đƣợc xử lý thành một số protein cấu trúc
của virus
Gen pol mã hóa một polyprotein và đƣợc xử lý sau dịch mã thành các protein chức năng
là PR (protease) chịu trách nhiệm phân cắt các protein khỏi polyprotein; RT (Reverse
Transcriptase) là enzyme phiên mã ngƣợc, RH (RNAse H) phân hủy nửa RNA trên từ
heteroduplex RNA:DNA, INT (Intergrase) tổng hợp bộ gen DNA virus vào bộ gen ký
chủ.
Gen env (envelope): tất cả các retrovirus động vật và một số retrovirus thực vật có gen
này. Gen này mã hóa cho các protein liên quan đến vỏ bọc của virus và do đó đặc biệt
quan trọng đối với các virus động vật.
Đối với các retrovirus động vật còn có các ORF phụ mã hóa cho các protein điều hòa phiên
mã nhƣ tat, rev, vif và nef.
4.7.2 Quá trình tái sinh của retrovirus
Bộ gen của các retrovirus đƣợc xem là cực (+) nhƣng không đƣợc dịch mã trực tiếp ngay.
Trong tế bào, bộ gen RNA của virus đƣợc phiên mã ngƣợc nhờ Reverse transcriptase thành
DNA sợi đơn cực (-), tiếp theo, đƣợc đƣợc tổng hợp thành phân tử DNA sợi kép. Phân tử DNA
sợi kép của virus này đƣợc tổng hợp vào bộ gen của ký chủ . Bộ gen DNA virus đƣợc phiên mã
thành RNA sợi (+). Các phân tử RNA sợi (+) này đƣợc dịch mã giống nhƣ gen ký chủ để tạo ra
các protein. Protein virus và các phân tử RNA (sợi +) này đƣợc lắp ráp thành virion.
Cần chú ý đối với retrovirus thực vật thì bộ gen DNA virus luôn nằm trong bộ gen ký chủ.
Virion chỉ hình thành trong quá trình tái sinh.
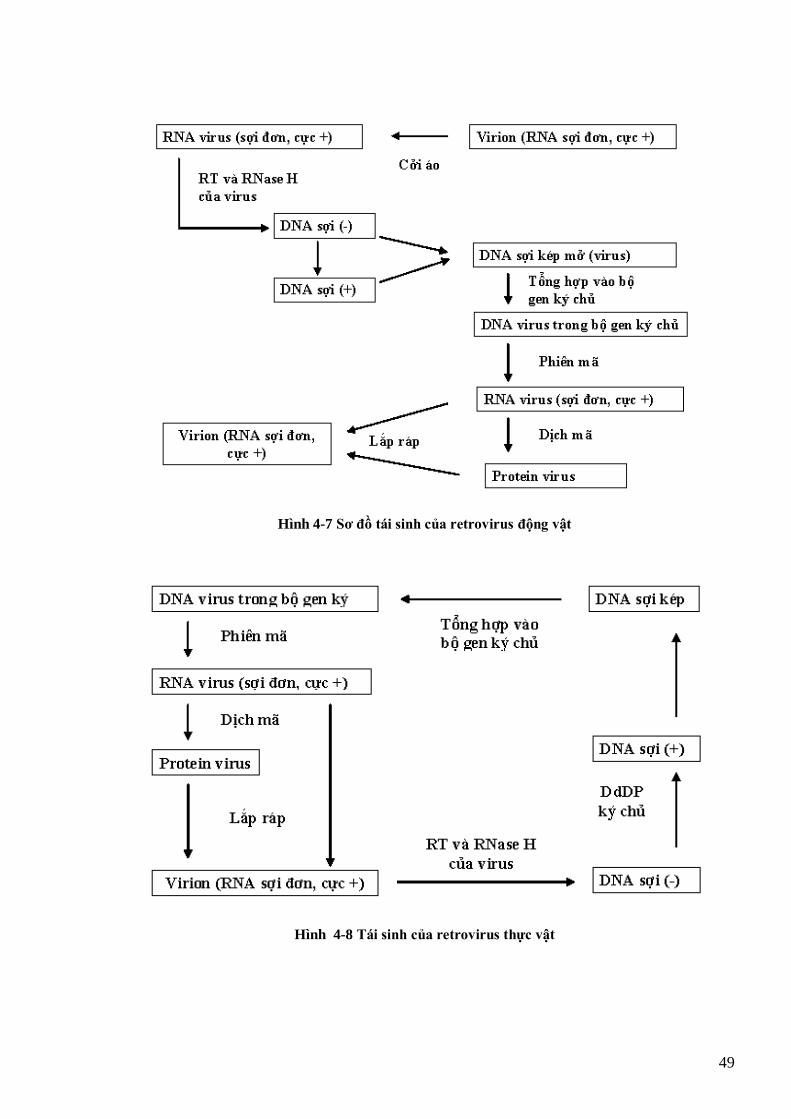
49
Hình 4-7 Sơ đồ tái sinh của retrovirus động vật
Hình 4-8 Tái sinh của retrovirus thực vật

50
4.8 TÁI SINH CỦA VIRUS DNA SỢI VÕNG ĐƠN
Đối với virus thực vật, chỉ có các virus thuộc hai họ Geminiviridae và Nanoviridae là có bộ
gen DNA sợi vòng đơn. Một số các virus động vật thuộc họ Circoviridae (ví dụ Porcine
circoviral virus type 2 (PCV2) gây bệnh còi cọc lợn sữa), họ Anelloviriade (một nhóm virus mới
phát hiện trên ngƣời) cũng có bộ gen DNA sợi vòng đơn. Các virus này đều tái sinh theo cơ chế
vòng lăn (rolling circular mechanism). Cơ chế vòng lăn có thể đƣợc chia làm 2 pha và đƣợc thực
hiện trong nhân tế bào ký chủ.
Các geminivirus và nanovirus đều mã hóa một protein gọi là Rep (đối với geminivirus) hoặc
Master Rep (đối với nanovirus). Các protein này không có chức năng polymerase nhƣng có chức
năng cắt và nối phân tử DNA ở một vị trí đặc biệt trên bộ gen virus. Vị trí này là một chuỗi bảo
thủ cao (TAATATTAC đối với geminivirus và TA(T/G)TATTAC đối với nanovirus). Vị trí TA
(gạch chân) chính là vị trí mà protein Rep/MasterRep cắt và nối phân tử DNA của virus. Chuỗi
bảo thủ này nằm trên vùng gọi là ―nguồn gốc tái bản‖ (ori, origin of replication). Ori bao gồm
một cấu trúc thân-thòng lọng (Stem-loop), chuỗi bảo thủ ở trên và các trình tự lặp đảo ngắn
(khoảng 4-5 nucleotid) gọi là interon cần thiết cho Rep nhận biết và bám vào sợi DNA của virus
để bắt đầu tái bản.
Quá trình tái sinh của geminivirus và nanovirus (Hình 4.9) diễn ra nhƣ sau:
1. Virus xâm nhập vào tế bào và cởi áo giải phóng bộ gen DNA virus.
2. Bộ gen đƣợc vận chuyển vào nhân. Trong nhân, quá trình tái sinh sảy ra theo 2 pha:
3. Pha 1 (pha tổng hợp sợi kép). Trong nhân, phân tử DNA sợi vòng đơn (bộ gen có mặt trong
phân tử virus) đƣợc tổng hợp thành thành phân tử DNA vòng kép. Quá trình khởi đầu tổng
hợp sợi kép vẫn chƣa đƣợc hiểu rõ. Nhƣ vậy sợi kép sẽ gồm một sợi virus và một sợi đối
virus. Trên sợi kép này, các gen đƣợc phiên mã (trên cả 2 sợi đối với geminivirus) để tạo các
mRNA. Các mRNA đƣợc vận chuyển ra tế bào chất và dịch mã để tạo ra các protein virus,
trong đó có Rep/Master Rep. Nhƣ vậy, quá trình phiên mã và dịch mã của virus hoàn toàn
dựa vào bộ máy phiên mã và dịch mã của tế bào ký chủ. Rep/Master Rep lại quay trở lại
nhân để tham gia tổng hợp bộ gen virus.
4. Pha 2 (pha vòng lăn). Tiếp theo là pha tái sinh theo cơ chế vòng lăn: Protein Rep/Master
Rep sẽ cắt sợi virus tại chuỗi bảo toàn. Nhờ vật liệu cũng nhƣ enzyme DNA polymearase
của tế bào, sợi virus đƣợc tổng hợp liên tục trên sợi tƣơng đồng virus. Protein Rep/Master
Rep lại tiếp tục cắt sợi virus mới đƣợc tổng hợp tại chuỗi bảo toàn (cũng vừa mới đƣợc tổng
hơp) thành một sợi virus hoàn chỉnh dƣới dạng sợi đơn mạch thẳng. Protein Rep/Master Rep
sau đó sẽ nối 2 đầu của mạch thẳng để tạo ra bộ gen virus sợi đơn mạch vòng hoàn chỉnh.
Bộ gen virus tiếp theo đƣợc vận chuyển ra tế bào chất và lắp ráp với CP để tạo các phân tử
virus mới.
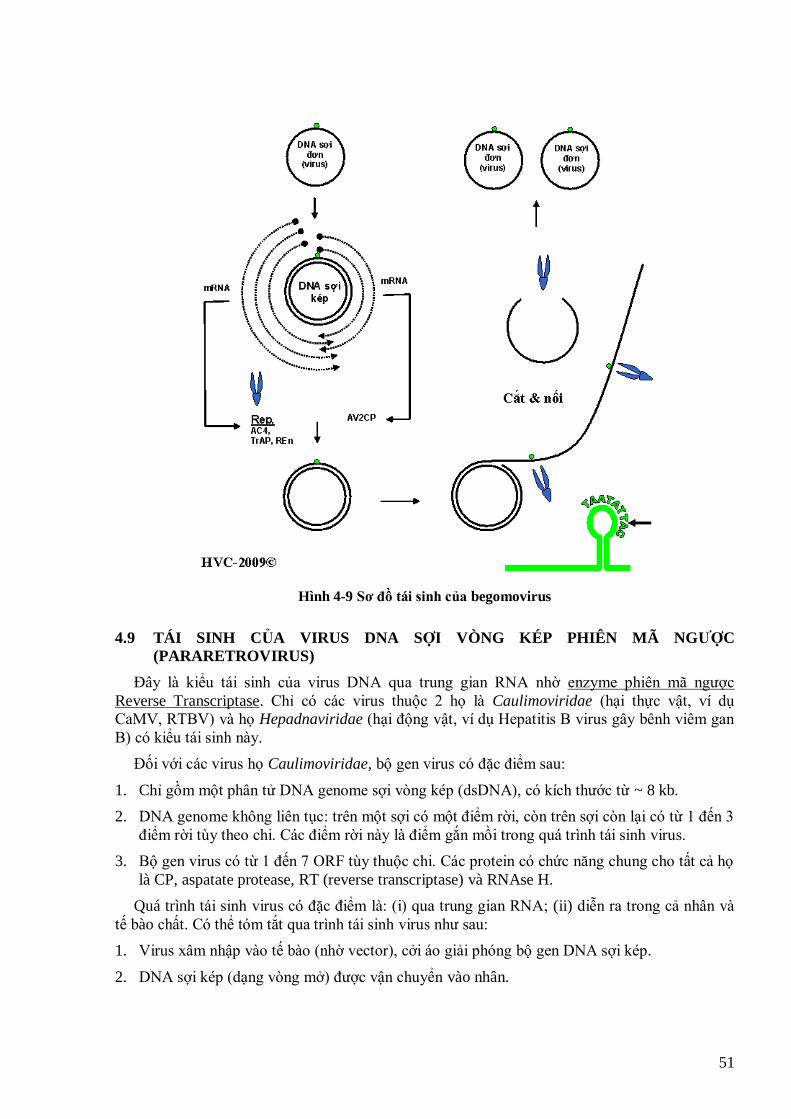
51
Hình 4-9 Sơ đồ tái sinh của begomovirus
4.9 TÁI SINH CỦA VIRUS DNA SỢI VÒNG KÉP PHIÊN MÃ NGƢỢC
(PARARETROVIRUS)
Đây là kiểu tái sinh của virus DNA qua trung gian RNA nhờ enzyme phiên mã ngƣợc
Reverse Transcriptase. Chỉ có các virus thuộc 2 họ là Caulimoviridae (hại thực vật, ví dụ
CaMV, RTBV) và họ Hepadnaviridae (hại động vật, ví dụ Hepatitis B virus gây bênh viêm gan
B) có kiểu tái sinh này.
Đối với các virus họ Caulimoviridae, bộ gen virus có đặc điểm sau:
1. Chỉ gồm một phân tử DNA genome sợi vòng kép (dsDNA), có kích thƣớc từ ~ 8 kb.
2. DNA genome không liên tục: trên một sợi có một điểm rời, còn trên sợi còn lại có từ 1 đến 3
điểm rời tùy theo chi. Các điểm rời này là điểm gắn mồi trong quá trình tái sinh virus.
3. Bộ gen virus có từ 1 đến 7 ORF tùy thuộc chi. Các protein có chức năng chung cho tất cả họ
là CP, aspatate protease, RT (reverse transcriptase) và RNAse H.
Quá trình tái sinh virus có đặc điểm là: (i) qua trung gian RNA; (ii) diễn ra trong cả nhân và
tế bào chất. Có thể tóm tắt qua trình tái sinh virus nhƣ sau:
1. Virus xâm nhập vào tế bào (nhờ vector), cởi áo giải phóng bộ gen DNA sợi kép.
2. DNA sợi kép (dạng vòng mở) đƣợc vận chuyển vào nhân.

52
3. Trong nhân, các điểm rời đƣợc bịt kín và phân tử trở thành dạng vòng kín.
4. Trong nhân, RNA pol II của ký chủ (DdRp) phiên mã thành 1 phân tử RNA có kích thƣớc >
bộ gen virus gọi là RNA 35S (hoặc 34S). Ở một số virus nhƣ CaMV và RTBV, một số phân
tử RNA này đƣợc cắt thành RNA nhỏ hơn (ví dụ RNA 19S).
5. RNA 35S (và cả các phân tử RNA nhỏ) đƣợc vận chuyển ra tế bào chất
6. Tại tế bào chất, các ORF đƣơch dịch mã trên khuôn RNA lớn (và RNA nhỏ) thành các
protein chức năng (trong đó có Reverse transcriptase và RNAse H).
7. Tại tế bào chất, phân tử RNA lớn sẽ đƣợc dùng làm khuôn để tổng hợp DNA sợi (-). RNAse
H sẽ loại bỏ phần RNA của phân tử heteroduplex RNA:DNA để giải phóng phân tử DNA
sợi đơn cực (-). Phân tử DNA này sẽ đƣợc dùng làm khuôn để tổng hợp thành phân tử DNA
genome sợi vòng kép mở.
8. Tại tế bào chất các phân tử DNA sợi kép lắp ráp với các protein CP để tạo thành virion.
Chú ý: Các virus đều thuộc nhóm pararetrovirus nên có khả năng tổng hợp gen của nó vào bộ
gen ký chủ. Cơ chế chi tiết của quá trình tổng hợp của các virus chƣa đƣợc hiểu rõ nhƣng nhiều
ký chủ của các virus thuộc chi Caulimoviridae có chứa các đoạn DNA giống của virus (ví dụ
chuối / banana streak virus; RTBV/lúa).
Hình 4-10 Sơ đồ tái sinh của các caulimovirus

53
4.10 CÁC CHIẾN LƢỢC DỊCH MÃ CỦA VIRUS THỰC VÂT
Bộ gen virus nhìn chung nhỏ nên thông tin di truyền của virus phải nén. Virus sử dụng 11
chiến lƣợc khác nhau để đảm bảo các gen của chúng có thể tiếp cận và sử dụng hiệu quả bộ máy
dịch mã của tế bào ký chủ.
4.10.1 Polyprotein
Ví dụ: Các potyvirus (PRSV, PVY…) có bộ gen RNA sợi đơn cực dƣơng, ~10 kb, 1 đầu
5‘UTR và 1 đầu 3‘UTR. Đầu 3‘UTR tận cùng bằng 1 chuỗi polyadenine. Các potyvirus có 1
ORF lớn mã hóa 1 polyprotein => đƣợc xử lý sau dịch mã thành 10 protein chức năng.
Hình 4-11 tổ chức bộ gen và chiến lƣợc dịch mã kiểu polyprotein của potyvirus
4.10.2 Gen phụ (subgenomic)
Gen phụ là các phân tử RNA đƣợc hình thành từ bộ gen chính trong quá trình tái sinh virus.
Chúng có chung đầu 3‘, một số có thể đƣợc lắp ráp trong virion, có thể ảnh hƣởng đến triệu
chứng.
Ví dụ: TMV tạo 2 phân tử gen phụ I1 và I2. Phân tử gen phụ I1 chứa ORF4 và dịch mã tạo
CP. Phân tử gen phụ I2 chứa 2 ORF và dịch mã cả CP và MP (hình 4.12).

54
Hình 4-12 Tổ chức bộ gen và chiến lƣợc dịch mã kiểu gen phụ, bỏ qua mã kết thúc và dịch mã nội
phân tử của TMV
4.10.3 Dịch mã nội phân tử
Trên bộ gen của virus có các vị trí để ribosome của tế bào ký chủ có thể tiếp cận gọi là ―vị trí
tiếp cận ribosome nội phân tử‖ (IRES, Internal Ribosome Entry Site). Đối với mRNA thông
thƣờng, quá trình dịch mã đƣợc thực hiện ở đầu 5‘ của phân tử mRNA (gọi là dịch mã phụ thuộc
mũ CAP). Tuy nhiên do có các vị trí IRES nên virus có thể dịch mã độc lập với kiểu dịch mã
phụ thuộc CAP. Thống kê tới 2009 có khoảng 60 virus động vật và 8 virus thực vật có cách dịch
mã này.
Ví dụ: TMV. Trên phân tử gen phụ I2 của TMV có 1 vị trí IRES giữa ORF3 (MP) và ORF
M4 (CP) => CP có thể đƣợc dịch mã (Hình 4.12).
4.10.4 Bỏ qua mã kết thúc (read – through)
Trên 1 ORF có nhiều mã kết thúc, tuy nhiên mã kết thúc của ORF thứ nhất bị ―dò‖ (leaky) =>
cho phép một số ribosome tiếp tục dịch mã tới mã kết thúc tiếp theo. Hậu quả là virus sẽ tạo ra 2
protein có phần đầu N giống nhau nhƣng có kích thƣớc khác nhau.
Ví dụ hai tiểu phần protein replicase của TMV có kích thƣớc khác nhau đƣợc tạo ra bằng
cách này (Hình 4.12).
4.10.5 Bộ gen phân đoạn
Khoảng 1/3 virus thực vật có bộ gen phân đoạn. Đối với phần lớn virus có bộ gen phân đoạn
thì các phân đoạn gen này đƣợc lắp ráp riêng rẽ (Ví dụ là các nanovirus nhƣ BBTV, các
begomovirus). Một số virus lại có các phân đoạn gen cùng đƣợc lắp láp trong 1 phân tử virion
(ví dụ các reovirus, tospovirus)

55
Hình 4-13 Bộ gen phân đoạn của BBTV (trái) và RRSV (phải) (Viralzone, 2009)
4.10.6 Dò (leaky scanning)
Ở các virus có cơ chế dịch mã kiểu ―dò‖ thì tiểu phần ribosome 40S sẽ trƣợt (scanning) bắt
đầu từ đầu 5‘ của mRNA nhƣng không dịch mã ở mã khởi đầu ATG (mã bị ―dò‖) mà lại dịch
mã từ mã ATG phía hạ lƣu. Có 3 dạng ―dò‖: (i) 2 mã ATG trên cùng 1 ORF, (ii) các ORF
chồng lớp (overlapping) và (iii) 2 ORF liền kề. Dịch mã kiểu dò có thể tao ra hai hay nhiều
protein giống nhau ở phần đầu C nếu quá trình dịch mã diễn ra trên cùng 1 khung đọc hoặc tạo
ra các protein hoàn toàn khác nhau nếu khác khung dịch mã (Hình 4.14).
Hình 4-14 Chiến lƣợc dịch mã kiểu “dò”. Dấu * là mã ATG. Mũi tên chỉ hƣớng dịch mã.

56
4.10.7 Cắt mRNA (Splicing)
Ở một số virus DNA nhƣ caulimovirus, tungrovirus, geminivirus, một phân tử mRNA lớn sau
khi hình thành sẽ cắt để tạo ra các mRNA nhỏ hơn, cho phép chúng có thể đƣợc dịch mã đồng
thời.
Ví dụ: mRNA 35S của CaMV, RTBV sau khi phiên mã sẽ cắt để tạo 1 mRNA 19S nhỏ hơn
(Hình 4.15).
Hình 4-15 Minh họa bộ gen của CaMV và 2 phân tử mRNA 35S và 19S (Haas et al., 2002)
4.10.8 Bổ trợ dịch mã (transactivator)
Ở một số virus DNA nhƣ caulimovirus, tungrovirus, geminivirus, một protein đƣợc dịch mã
từ 1 mRNA sẽ điều khiển (hỗ trợ sự dịch mã của các protein trên mRNA khác (transacting). Ví
dụ: protein P48 của CaMV đƣợc dịch mã từ mRNA 19S sẽ tham gia điểu khiển sự dịch mã của
các ORF trên mRNA 35S.
4.10.9 Nhảy ribosome (Ribosome shunting)
Trên bộ gen của virus, ở vùng không mã hóa có nhiều ORF nhỏ (<50 aa) không có chức
năng => chúng không thể đƣợc dịch mã => Cơ chế ―nhảy‖. Các ORF nhỏ thƣờng tạo ra các cấu
trúc thứ cấp (stem-loop) => không cho phép ribosome trƣợt thẳng qua => buộc phải ―nhảy‖. Cơ
chế dịch mã đặc biệt này đƣợc phát hiện thấy đầu tiên ở virus CaMV và RTBV, tiếp theo cũng
đƣợc phát hiện ở nhiều virus động vật nhƣ hepatitis B.
Ở 2 virus CaMV và RTBV, phân tử mRNA 35 S có một chuỗi ―leader‖ dài chứa tới 19
ORF nhỏ và có cấu trúc phức tạp để tạo ra cấu trúc thứa cấp dạng kẹp tóc. Để có thể dịch mã
theo cơ chế ―nhảy‖, một ORF nhỏ đầu tiên phải đƣợc dịch mã theo cơ chế phụ thuộc CAP. Cơ
chế shunting có thể mô tả tóm tắt ở hình 4-16 nhƣ sau:
Bƣớc 1, 2 và 3: dịch mã ORF nhỏ thứ nhất (quá trình dịch mã sẽ phụ thuộc CAP nhƣ đối với
dịch mã mRNA thông thƣờng)
Bƣớc 4 và 5: ribosome ―nhảy‖ qua cấu trúc thứ cấp (chứa các ORF nhỏ khác).
Bƣớc 6: ribosome bắt đầu dịch mã ORF chính

57
Hình 4-16 Mô hình dịch mã của CaMV, RTBV theo cơ chế “nhảy” (Thiébeauld et al., 2007)
4.10.10 Chuyển khung (Frameshift)
Cách dịch mã này có thể bắt gặp ở các virus có mRNA chứa các ORF chồng lớp
(overlapping). Trên ORF thứ nhất (khung 1), ribosome sẽ tiến hành dịch mã; tuy nhiên trƣớc khi
tới mã kết thúc, ribosome sẽ chuyển khung dịch để dịch mã tiếp trên khung 2 chứa ORF thứ 2
(Hình 4.17). Có 2 chế độ chuyển khung là lùi (-) và tăng (+) 1 nucleotide. Nhiều virus thực vật
có kiểu dịch mã này nhƣ closterovirus, tombustvirus, crinivirus, luteovirus…
Hình 4-17 Dịch mã theo kiểu chuyển khung
4.10.11 Dịch mã 2 chiều (Ambisense)
Đây là một chiến lƣợc rất phổ biến của virus. Ví dụ ở TSWV, bộ gen gồm 3 phân tử RNA
sợi đơn cực (-) hoặc lƣỡng cực ký hiệu là L (large), M (medium) và S (small) có tổng kích thƣớc
từ 11 – 19 kb.
Trên sợi M có 2 ORF đƣợc dịch mã theo 2 chiều ngƣợc nhau. Trên sợi (-) tức là sợi virus,
có 1 ORF mã hóa protein Gn và Gc - là các glycoprotein (liên kết với đƣờng). Ký hiệu n hoặc c
có nghĩa glycoprotein nằm phía đầu amin hay đầu carboxyl của chuỗi polyprotein đƣợc dịch mã

58
từ sợi M. Trên sợi (+) tức là sợi đối virus, có 1 ORF mã hóa một protein phi cấu trúc (không có
mặt trong phân tử virion) ký hiệu là NSm. Quá trình dịch mã của các ORF trên sợi M đƣợc trình
bày ở hình dƣới. Chú ý: mặc dù sợi đối virus đƣợc ký hiệu là sợi (+) nhƣng không đƣợc dịch mã
ngay mà vẫn cần phải đƣợc phiên mã sang mRNA (Hình 4.18).
Hình 4-18 Tổ chức bộ gen của TSWV và dịch mã 2 chiều trên phân đoạn M
4.11 CÂU HỎI ÔN TẬP CHƢƠNG 4
1. Đặc điểm chung của quá trình tái sinh (sinh sản) virus.
2. Tái sinh của nhóm virus RNA sợi đơn, cực dƣơng (đặc điểm, các bƣớc, hiểu sơ đồ, ví dụ).
3. Tái sinh của nhóm virus RNA sợi âm (đặc điểm, các bƣớc, hiểu sơ đồ, ví dụ).
4. Tái sinh của nhóm virus RNA sợi kép (đặc điểm, các bƣớc, hiểu sơ đồ, ví dụ).
5. Tái sinh của nhóm virus DNA sợi vòng đơn (đặc điểm, các bƣớc, hiểu sơ đồ, ví dụ).
6. Tái sinh của nhóm virus DNA sợi kép, phiên mã ngƣợc (đặc điểm, các bƣớc, hiểu sơ đồ, ví
dụ).
7. Tái sinh của nhóm retrovirus (đặc điểm, các bƣớc, hiểu sơ đồ, ví dụ).
8. Các chiến lƣợc dịch mã của virus (tại sao virus có các chiến lƣợc dịch mã khác nhau, đặc
điểm và ví dụ của mỗi chiến lƣợc).
4.12 TÀI LIỆU THAM KHẢO CHÍNH
1. Buck, K. W. (1999). Replication of tobacco mosaic virus RNA. PhilTrans R Soc Lond B
354, 613-627.
2. Dorokhov, Y. L., Ivanov, P. A., Komarova, T. V., Skulachev, M. V. & Atabekov, J. G.
(2006). An internal ribosome entry site located upstream of the crucifer-infecting

59
tobamovirus coat protein (CP) gene can be used for CP synthesis in vivo. Journal of General
Virology 87, 2693–2697.
3. Gutierrez, C., Ramirez-Parra, E., Castellano, M. M., Sanz-Burgos, A. P., Luque, A. &
Missich, R. (2004). Geminivirus DNA replication and cell cycle interactions. Veterinary
Microbiology 98, 111-119.
4. Haas, M., Bureau, M., Geldreich, A., Yot, P. & Keller, M. (2002). Cauliflower mosaic virus:
still in the news. MOLECULAR PLANT PATHOLOGY 3, 419–429.
5. Hanley-Bowdoin, L., Settlage, S. B., Orozco, B. M., Nagar, S. & Robertson, D. (2000).
Geminiviruses: models for plant DNA replication, transcription, and cell cycle regulation.
Crit Rev Biochem Mol Biol 35, 105-140.
6. Havecker, E. R., Gao, X. & Voytas, D. F. (2004). The diversity of LTR retrotransposons.
Genome Biology 5:225.
7. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
8. Mackenzie, J. (2005). Wrapping Things up about Virus RNA Replication. Traffic 6, 967–
977.
9. Sanfaçon, H. (2005). Replication of positive-strand RNA viruses in plants: contact points
between plant and virus components. Canadian Journal of Botany 83, 1529-1549.
10. Thiébeauld, O., Pooggin, M. M. & Ryabova, L. A. (2007). Alternative Translation Strategies
in Plant Viruses. Plant Viruses 1, 1-20

60
Chƣơng 5. SỰ DI CHUYỂN CỦA VIRUS TRONG CÂY
5.1 TÓM TẮT NỘI DUNG
Sau khi tái sinh, virus thực vật cần phải di chuyển sang các tế bào mới, mô mới, cơ quan mới
để gây bệnh hệ thống cho cây. Chƣơng này giới thiệu một cách đơn giản cơ chế di chuyển virus
trong cây. Sự di chuyển của virus trong cây đƣợc chia làm 2 nhóm:
1. Di chuyển giữa các tế bào (Cell-to-cell movement).
2. Di chuyển hệ thống trong cây (Long distance movement).
5.2 DI CHUYỂN GIỮA CÁC TẾ BÀO
5.2.1 Cấu tạo sợi liên bào
Sợi liên bào (plasmodesmata) là kênh giao tiếp giữa các tế bào và có cấu trúc phức tạp gắn
liền với màng tế bào và lƣới nội chất. Cấu tạo cơ bản của sợi liên bào gồm 2 ống trụ cấu tạo bởi
màng và lồng vào nhau (Hình 5.1).
Ống trụ ngoài là màng plasma (PM, plasma membrance) bao gồm màng plasma ngoài (OPM,
outer palsma membrance) hƣớng về vách tế bào (CW, cell wall) và màng plasma trong (IPM,
inner plasma membrance) hƣớng vào ống trụ trong.
Ống trụ trong cấu tạo bởi màng lƣới nội chất (ER, endoplasmic reticulum). Ống trụ trong
chính là màng lƣới nội chất bị ép chặt lại nên thƣờng đƣợc ký hiệu là AER (appresed
endoplasmic reticulum). Phần ống trụ trong cũng còn đƣợc gọi là desmotubule.
Trên bề mặt trong của ống trụ ngoài và bề mặt ngoài của ống trụ trong có các protein của ký
chủ gọi là các protein sợi liên bào (PDP, plasmodesmal proteins) gắn vào. Khoảng không giữa
ống trụ ngoài và ống trụ trong là kênh hay lỗ sợi liên bào (kênh plasmodesmata). Dịch tế bào
chất giữa các tế bào liên tục với nhau qua kênh này. Nhìn chung kích thƣớc kênh sợi liên bào
chỉ khoảng 2.5 nm. Do kích thƣớc kênh rất nhỏ, lại có rào cản là các protein sợi liên bào nên
không phải tất cả các loại phân tử đều có thể tự do di chuyển qua kênh. Khả năng sợi liên bào
không cho phép các loại phân tử sinh học đạt kích thƣớc nào đó di chuyển qua gọi là giới hạn
ngăn chặn kích thƣớc (SEL, size exclusion limit) của sợi liên bào và thƣờng đƣợc biểu diễn
dƣới dạng khối lƣợng phân tử kDa. Nhìn chung SEL chỉ cho phép các phân tử có khối lƣợng tối
đa khoảng 1 – 10 kDa đi qua, tùy loại mô và cây.
Hình 5-1 Cấu tạo sợi liên bào. A và B là mô hình sợi liên bào, trong đó A là tiết diện dọc và B là tiết
diện ngang. C là ảnh hiện vi điện tử của sợi liên bào (Lucas, 2006).

61
5.2.2 Sự di chuyển của virus thực vật giữa các tế bào
Do đặc điểm cấu tạo của tế bào và mô thực vật, nhìn chung quá trình di chuyển từ tế bào này
sang tế bào khác của virus không dễ dàng. Các tế bào thực vật liên hệ với nhau bởi sợi liên bào
(plasmodesmata) có cấu tạo phức tạp. Kích thƣớc của kênh (lỗ) sợi liên bào cũng nhƣ SEL quá
nhỏ không cho phép virion của virus nhỏ nhất, thậm chí chỉ bộ gen virus có thể qua lại tự do
đƣợc.
Để khắc phục trở ngại này, các virus thực vật đều mã hóa một hoặc một vài protein đặc biệt
gọi là protein vận chuyển (MP, movement protein). Sử dụng protein MP, kết hợp với sự tham
gia của các protein khác (của cả virus và ký chủ), phân tử virus (virion) hoặc bộ gen virus mới
có thể di chuyển từ tế bào nhiễm sang tế bào khỏe bên cạnh. Virus sử dụng 2 chiến lƣợc để di
chuyển từ tế bào này sang tế bào khác thông qua sợi liên bào.
Ƣớc lƣợng thời gian cho thấy virus nhìn chung di chuyển từ tế bào nhiễm sang tế bào khỏe
trong khoảng 2 – 20 giờ, phụ thuộc nhiệt độ, tổ hợp virus và cây.
5.2.2.1 Chiến lƣợc tạo ống (tubular)
Ít nhất các virus thuộc các họ Bromoviridae, Comoviridae, Caulimoviridae, Nepoviridae và
các chi Tospovirus, Trichovirus sử dụng chiến lƣợc tạo ống để di chuyển giữa các tế bào. Quá
trình di chuyển bao gồm các bƣớc chính sau:
Protein MP của virus tiếp cận sợi liên bào và loại bỏ ống trụ trong (desmotubule) của sợi liên
bào).
Tiếp theo các protein MP của virus lắp ráp lại thành một ống ngang qua sợi liên bào
Cuối cùng, phân tử virus (virion), thông qua tƣơng tác với protein MP của virus đƣợc vận
chuyển qua ống sang tế bào bên cạnh.
Hình 5-2 Chiến lƣợc tạo ống của virus thực vật để di chuyển giữa các tế bào (Hull, 2002)
5.2.2.2 Chiến lƣợc nucleoprotein
Đây là chiến lƣợc phổ biến nhất đối với virus thực vật. Protein MP của virus liên kết với
nucleic acid của virus tạo thành phức hợp nucleoprotein. Protein MP của virus cùng với protein

62
ký chủ sẽ biến đổi sợi liên bào và làm tăng giới hạn ngăn chặn kích thƣớc SEL của sợi liên bào
lên nhiều lần. Tiếp theo, phức hợp nucleoprotein có thể di chuyển từ tế bào nhiễm ban đầu qua
sợi liên bào đã đƣợc biến đổi để sang tế bào khỏe bên cạnh. Tại tế bào khỏe, virus lại thực hiện
tiếp quá trình tái bản bộ gen và dịch mã các protein chức năng.
Chiến lƣợc này đã đƣợc chứng minh chi tiết đối với TMV (hình 5.4). Protein MP của TMV
có thể tăng kích thƣớc SEL từ 7 kDa tới 94 kDa.
Hình 5-3 Mô hình di chuyển giữa các tế bào của TMV
5.3 DI CHUYỂN HỆ THỐNG QUA KHOẢNG CÁCH XA
Về cơ bản, kiểu di chuyển này tƣơng đối tự do đối với virus. Ngoại trừ một số ít virus nhƣ
các sobemovirus di chuyển theo mạch xylem thì phần lớn virus di chuyển theo mạch phloem.
Sau khi đạt tới mạch phloem, virus có thể di chuyển hệ thống khắp cây.
Thời gian virus phát tán từ vị trí xâm nhiễm ban đầu tới khắp cả cây phụ thuộc rất nhiều yếu
tố nhƣ tổ hợp cây và virus, loại cây, loại và tuổi cây, loại virus, kiểu xâm nhiễm và nhiệt độ.
Nhìn chung virus di chuyển trong cây rất chậm (Hình 5.5).
Hƣớng di chuyển của virus trong cây theo mạch phloem nhìn chung là từ mô nguồn ―source‖
tới mô tiêu thụ ―sink‖. Mô nguồn là nơi carbohydrates đƣợc tạo ra (trên lá thành thục) hoặc đƣợc
giải phóng từ mô dự trữ (rễ, thân). Mô tiêu thụ là nơi carbohydrates đƣợc sử dụng để tăng trƣởng
và dự trữ (Hình 5.5).

63
Hình 5-4 Hƣớng và tốc độ di chuyển hệ thống của virus trong cây (Agrios, 2005)
5.4 CÂU HỎI ÔN TẬP CHƢƠNG 5
1. Cấu tạo và chức năng của sợi liên bào.
2. Đâu là sự khác biệt giữa sự di chuyển của virus thực vật trong ký chủ nhiễm bệnh so với các
nhóm virus khác.
3. Đặc điểm sự di chuyển giữa các tế bào của virus thực vật.
4. Đặc điểm di chuyển hệ thống trong cây của virus thực vật.
5.5 TÀI LIỆU THAM KHẢO CHÍNH
1. Agrios, G. (2005). Plant Pathology. Firth Edition: Academic Press.
2. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
3. Lough, T. J. & Lucas, W. J. (2006). Integrative Plant Biology: Role of Phloem Long-
Distance Macromolecular Trafficking. Annual Review of Plant Biology 57, 203–232.
4. Lucas, W., J. (2006). Plant viral movement proteins: Agents for cell-to-cell trafficking of
viral genomes. Virology 344 169 – 184.

64
Chƣơng 6. CƠ CHẾ GÂY BỆNH CỦA VIRUS THỰC VẬT
6.1 TÓM TẮT NỘI DUNG
Nhìn chung, so với các lĩnh vực nghiên cứu khác trong virus thực vật học thì cơ chế gây bệnh
của virus ít đƣợc chú ý hơn. Chẳng hạn, thậm chí cho tới tận bây giờ, ngƣời ta vẫn không hiểu rõ
tại sao virus lại tạo ra triệu chứng khảm lá. Hai trở ngại quan trọng dẫn tới hạn chế việc nghiên
cứu cơ chế gây bệnh của virus là (i) chúng tái sinh bên trong tế bào và (ii) không tạo ra độc tố
và phytohormon giống nhƣ đối với nấm và vi khuẩn mặc dù chúng có thể can thiệp vào đƣờng
hƣớng sinh tổng hợp phytohormone.
Cho tới nay, nhờ vào các nghiên cứu ở mức phân tử, nhiều tƣơng tác giữa virus và tế bào cây
ký chủ dẫn tới biểu hiện triệu chứng bệnh đã đƣợc sáng tỏ.
Chƣơng này giới thiệu một số nội dung liên quan đến cơ chế gây bệnh của virus và biểu hiện
triệu chứng bệnh.
6.2 TRIỆU CHỨNG BỆNH VIRUS
Triệu chứng là biểu hiện phản ứng của cây đối với sự tấn công của virus. Một cây bị bệnh và
biểu hiện triệu chứng khi các chức năng sinh lý của nó bị ảnh hƣởng. Một virus ―thông minh‖
khi nó tấn công tế bào, sinh sản mạnh mẽ qua các thế hệ mà không gây bệnh cho cây.
Triệu chứng bệnh virus trên cây trồng có thể chia là 3 nhóm: (i) biến màu, (ii) biến dạng và
(iii) vết đốm. Ngoài ra, bệnh do virus gây ra cũng có thể đƣợc xếp vào 2 nhóm là (i) bệnh hệ
thống và (ii) bệnh cục bộ. Nhìn chung, phần lớn bệnh virus thuộc nhóm bệnh hệ thống. Các
nhóm triệu chứng và triệu chứng bệnh virus có thể tóm tắt nhƣ sau:
6.2.1 Biến màu
Khảm lá (mosaic). Đây có thể đƣợc xem là triệu chứng phổ biến nhất của bệnh virus
Đốm biến vàng (mottling)
Biến vàng (yellowing)
Sáng gân (vein clearing)
Vàng gân (vein yellowing)
Dải gân (vein banding)
6.2.2 Biến dạng
Xoăn lá (curling)
Cuốn lá (rolling)
Lùn (dwarfing)
Còi cọc (stunting)
Phồng gân (enation)
6.2.3 Vết đốm
Đốm chết hoại (necrotic spot)
Đốm vòng (ringspot)

65
6.3 HAI MÔ HÌNH GIẢI THÍCH CƠ CHẾ GÂY BỆNH CỦA VIRUS
6.3.1 Mô hình cạnh tranh tài nguyên
Theo mô hình này, khi virus xâm nhiễm vào tế bào, quá trình tái sinh của virus sẽ sử dụng
các tài nguyên của tế bào nhƣ amino acid, nucleotide để cấu tạo nên phân tử virus mới. Việc
chiếm đoạt quá mức tài nguyên cũng nhƣ trƣng dụng bộ máy phiên mã và dịch mã của tế bào ký
chủ sẽ dẫn tới biểu hiện triệu chứng bệnh.
Ngƣời ta đã chứng minh rằng hàm lƣợng protein và acid nucleic của TMV có thể chiếm tới 1
% tổng số khối lƣợng vật chất của cây thuốc lá nhiễm bệnh nhƣng khi virus dịch mã, nó có thể
huy động tới hơn 50% tổng số protein của cây.
Ngƣời ta cũng quan sát thấy tại vị trí xâm nhiễm, nhiều gen của cây bị suy giảm mức độ
phiên mã và dịch mã. Sự suy giảm này giúp virus dễ dàng tiếp cận với tài nguyên của tế bào
hơn. Ngoài ra trên bộ gen của nhiều virus có chứa các cấu trúc đặc biệt nhƣ các vị trí lắp ráp
ribosome, các chuỗi tăng cƣờng dịch mã ―translation enhancer‖ làm cho mRNA của virus có lợi
thế cạnh tranh hơn so với mRNA của tế bào ký chủ trong quá trình tiếp cận bộ máy dịch mã của
tế bào.
Tuy nhiên, cơ chế gây bệnh của virus theo mô hình cạnh tranh không phải là chủ yếu vì:
Ở nhiều bênh virus, mức độ dữ dội của triệu chứng không tỷ lệ thuận với mức độ tích lũy
virus.
Trong tế bào, vòng đời của virus rất ngắn, quá trình tái sinh của virus xảy ra rất nhanh,
khoảng vài giờ, sau đó suy giảm do đó quá trình cạnh tranh tài nguyên tế bào của virus cũng
diễn ra rất nhanh. Ngoài ra, ngay sau khi virus ngừng tái sinh thì tế bào thƣờng phục hồi tài
nguyên của nó dễ dàng
6.3.2 Mô hình tƣơng tác
Theo mô hình này, virus gây bệnh cho cây thông qua sự tƣơng tác giữa các thành phần của
virus, thƣờng là các protein, với các thành phần của tế bào ký chủ, qua đó ảnh hƣởng đến các
đƣờng hƣớng sinh hóa của tế bào.
Mô hình này phức tạp hơn nhƣng hiện thực và đã đƣợc chứng minh trong nhiều trƣờng hợp
(xem các phần sau). Một trong các đặc điểm quan trọng nhất của mối tƣơng tác này là tính đặc
hiệu. Điều này giải thích tại sao triệu chứng bệnh có thể rất khác nhau khi các virus có quan hệ
gần gũi tấn công cùng cùng loại cây hoặc cùng một virus tấn công các cây khác nhau
Dựa vào hậu quả của sự tƣơng tác đến các chức năng sinh lý của cây, ngƣời ta chia các tƣơng
tác giữa virus và cây thành 2 nhóm:
6.3.2.1 Tƣơng tác gây hậu quả đến ký chủ
Các tƣơng tác nhóm này cần thiết cho các chức năng sống của virus và cũng ảnh hƣởng trực
tiếp đến các chức năng sinh lý của cây dẫn tới triệu chứng bệnh. Có rất nhiều ví dụ về các tƣơng
tác nhóm này (xem các phần sau).
6.3.2.2 Tƣơng tác không gây hậu quả đến ký chủ
Các tƣơng tác này cần thiết cho các chức năng sống của virus nhƣng không hoặc rất ít ảnh
hƣởng đến các chức năng sinh lý của cây và hậu quả là cây không bị bệnh. Ví dụ điển hình cho
các tƣơng tác này là các virus ẩn (cryptic virus). Ví dụ tƣơng tự là tƣơng tác của protein

66
replicase của TMV với 2 protein xuyên màng (TOM1 và TOM3) của tế bào thuốc lá. Tƣơng tác
này cần thiết cho sự tái sinh của TMV và không ảnh hƣởng đến cây. Khi gây bất hoạt 2 gen này
(silencing) thì mức độ tái sinh của virus giảm nhƣng không ảnh hƣởng đến sinh trƣởng của cây.
Cần chú ý là việc phân nhóm này không bất biến vì hậu quả của các tƣơng tác bị ảnh hƣởng
bởi nhiều yếu tố nhƣ môi trƣờng, trạng thái sinh lý của cây.
6.4 CÁC CƠ CHẾ GÂY BỆNH CỦA VIRUS
6.4.1 Virus gây mất cân bằng phytohormone
Nhiều triệu chứng của bệnh virus nhóm biến dạng chẳng hạn nhƣ cây lùn, còi cọc, xoăn cuốn
lá thƣờng liên quan đến mất cân bằng phytohormone trong cây.
6.4.1.1 Auxin
Auxin là một phytohormone chủ yếu của cây, điều khiển nhiều đƣờng hƣớng sinh hóa liên
quan đến sinh trƣởng của cây.
Ví dụ. Thí nghiệm trên cây Arabidopsis với TMV đã xác định đƣợc tƣơng tác của protein
replicase của virus với 1 nhóm các protein của ký chủ gọi là các protein Aux/IAA. Các protein
Aux/IAA là các protein hoạt động trong nhân với chức năng kìm hãm các protein là yếu tố phiên
mã điều khiển sự phiên mã các gen ký chủ đƣợc kích hoạt bằng auxin (ký hiệu là ARFs = auxin-
responsive transcription factors).
Khi không có mặt auxin, protein Aux/IAA liên kết với protein ARF => các gen ký chủ liên
qua đến đƣờng hƣớng auxin không đƣợc hoạt hóa.
Khi có mặt auxin, protein Aux/IAA bị phân hủy và giải phóng protein ARF => hoạt hóa các
gen ký chủ liên qua đến đƣờng hƣớng auxin.
Khi tế bào nhiễm TMV, protein replicase của virus tƣơng tác với protein Aux/IAA và không
cho nó di chuyển vào trong nhân => protein ARF hoạt động một cách không kiểm soát => tạo
triệu chứng bệnh.
Khi nhiễm TMV mang đột biến trên gen replicase làm mất chức năng tƣơng tác với protein
Aux/IAA của nó => cây biểu hiện triệu chứng nhẹ hơn rất nhiều mặc dù mức độ tái sinh và di
chuyển hệ thống của virus vân tƣơng tự nhƣ của virus TMV dạng hoang dại.
Khi gây bất hoạt gen mã hóa protein Aux/IAA của cây Arabidopsis thì cây biểu hiện kiểu
hình giống nhƣ bị nhiễm TMV.
6.4.1.2 Gibberellins
Ví dụ. Cây lúa nhiễm một số virus họ Reoviridae nhƣ RRSV, SRBSDV, RGDV thƣờng biểu
hiện triệu chứng lùn, bộ lá xanh đậm. Triệu chứng này giống với hiện tƣợng cây lúa bị đột biến
mất khả năng tổng hợp gibebrellin. Ảnh hƣởng của sự nhiễm bệnh virus đến cân bằng
gibberellin đã đƣợc chứng minh đối với 1 virus tƣơng tự thuộc họ này là RDV (Rice dwarf
virus).
RDV mã hóa 1 protein gọi là P2. Đây là protein tạo vỏ của virus, qui định tính đặc hiêu
vector và qui định triệu chứng lùn + bộ lá xanh đậm trên cây lúa.
P2 tƣơng tác với 1 protein ký chủ là ent-kaurene oxidase. Ent-Kaurene oxidases là một
protein chủ chốt trong đƣờng hƣớng sinh tổng hợp GA trong cây. Cây lúa nhiễm RDV bị lùn.

67
Khi xử lý cây bệnh với GA3 dẫn tới biến mất triệu chứng trên cây bệnh trong khi xử lý IAA thì
vẫn giống với đối chứng nhiễm (Hình 6.1).
Hình 6-1 Thí nghiệm chứng minh protein P2 của RDP đã can thiệp vào đƣờng hƣớng tổng hợp
gibberelline của cây lúa (Wang et al., 2005)
6.4.1.3 Ethylene
Ethylen là một phytohormone tham gia điều khiển quá trình già hóa và phòng thủ của cây.
Khi cây bị nhiễm virus, hàm lƣợng ethylen thƣờng tăng. Ethylen đã đƣợc chứng minh có liên
quan tới triệu chứng biến vàng và chết hoại của cây bị bệnh virus.
Ví dụ. Cây chuyển gen P6 của virus CaMV biểu hiện triệu chứng giống hệt bị nhiễm virus
nhƣ biến vàng, còi cọc và dải gân.
6.4.2 Protein của virus khởi động lại chu kỳ tế bào
Một ví dụ điển hình cho cơ chế này là các begomovirus. Quá trình tái bản của begomovirus
xảy ra trong nhân tế bào, kể cả ở các tế bào không phân chia. Do vậy, sau khi nhiễm vào tế bào,
begomovirus phải khởi động bộ máy tái sinh của tế bào. Một trong các protein của tế bào ký chủ
thực vật điều khiển chu kỳ tế bào là pRBR (plant retinoblastoma-related protein) - chịu trách
nhiệm chuyển chu kỳ tế bào từ pha G1 sang pha S (pha tổng hợp). Protein Rep của TYLCV đã
đƣợc chứng minh tƣơng tác với pRBR của tế bào ký chủ. Hậu quả là chu kỳ tế bào đƣợc khởi
động lại, tạo các enzym cần thiết cho sự tái bản DNA của virus (Hình 6.2).

68
Hình 6-2 Tƣơng tác giữa protein Rep của begomovirus với pRBR của tế bào cho phép khởi động
lại chu kỳ tế bào
6.5 CÂU HỎI ÔN TẬP CHƢƠNG 6
1. Các nhóm triệu chứng chính trên cây do virus gây ra. Ví dụ
2. Các triệu chứng chính trên cây do virus gây ra. Ví dụ.
3. Khái niệm 2 mô hình giải thích cơ chế gây bệnh của cây. Mô hình nào hiện thực hơn ? Tại
sao ?
4. Ví dụ cơ chế gây bệnh hƣởng đến mất cân bằng phytohormone.
5. Ví dụ cơ chế gây bệnh do virus khởi động lại chu kỳ tế bào.
6.6 TÀI LIỆU THAM KHẢO CHÍNH
1. Culver, J. N. & Padmanabhan, M. S. (2007). Virus-Induced Disease: Altering Host
Physiology One Interaction at a Time. Annual Review of Phytopathology 45, 221–243.
2. Gutierrez, C. (2002). Strategies for geminivirus DNA replication and cell cycle interference.
Physiological and Molecular Plant Pathology 60, 219-230.
3. Gutierrez, C., Ramirez-Parra, E., Castellano, M. M., Sanz-Burgos, A. P., Luque, A. &
Missich, R. (2004). Geminivirus DNA replication and cell cycle interactions. Veterinary
Microbiology 98, 111-119.
4. Hanley-Bowdoin, L., Settlage, S. B., Orozco, B. M., Nagar, S. & Robertson, D. (2000).
Geminiviruses: models for plant DNA replication, transcription, and cell cycle regulation.
Crit Rev Biochem Mol Biol 35, 105-140.
5. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
6. Zhu, S., Gao, F., Cao, X., Chen, M., Ye, G. & al., e. (2005). The rice dwarf virus P2 protein
interacts with ent-kaurene oxidases in vivo, leading to reduced biosynthesis of gibberellins
and rice dwarf symptoms. Plant Physiology 139, 1935–1945.

69
Chƣơng 7. LAN TRUYỀN CỦA VIRUS THỰC VẬT
7.1 TÓM TẮT NỘI DUNG
Chƣơng này đề cập đến các cách lan truyền ngoài tự nhiên của virus thực vật, đặc biệt nhấn
mạnh đến phƣơng thức lan truyền qua vector sinh học là côn trùng chích hút.
7.2 GIỚI THIỆU
Virus thực vật lan truyền ngoài tự nhiên theo không gian (từ cây này sang cây khác, từ ruộng
này sang ruộng khác, từ nƣớc này sang nƣớc khác…) và theo thời gian (trong một vụ trồng hoặc
từ vụ này sang vụ khác… ) nhờ các phƣơng thức sau:
1. Tiếp xúc cơ học
2. Nhân giống vô tính
3. Hạt giống/phấn hoa
4. Môi giới
Cần chú ý là một virus có thể có nhiều phƣơng thức lan truyền khác nhau.
7.3 LAN TRUYỀN QUA TIẾP XÖC CƠ HỌC
Lan truyền qua tiếp xúc cơ học là lan truyền nhờ các tổn thƣơng cơ học nhƣ tiếp xúc, cọ xát,
va chạm, chăm sóc, vun xới, tỉa cành lá.
Đây là phƣơng thức lan truyền không phổ biến ngoài tự nhiên mặc dù trong điều kiện thí
nghiệm, nhiều virus vẫn có khả năng này. Các virus có khả năng lan truyền qua tiếp xúc cơ học
phải là các virus có thể tái sinh đƣợc ở tế bào biểu bì, tế bào nhu mô.
Ngoài tự nhiên, chỉ một vài nhóm có khả năng truyền dễ dàng qua tiếp xúc cơ học là các
virus thuộc chi Tobamovirus (ví dụ TMV).
7.4 LAN TRUYỀN QUA NHÂN GIỐNG VÔ TÍNH
Đây là con đƣờng lan truyền rất quan trọng của nhiều bệnh virus vì nhìn chung các bệnh virus
phần lớn là bệnh hệ thống. Một khi cây đã nhiễm virus thì các bộ phận dùng làm giống nhƣ củ
giống, hom giống, thân ngầm cũng mang virus.
Các ví dụ về virus truyền qua nhân giống vô tính đã phát hiện thấy ở Việt Nam gồm:
BBTV (banana bunchy top virus). Virus gây bệnh chùn ngọn chuối, có thể truyền dễ dàng từ
cây mẹ sang cây con qua thân ngầm.
Các potyvirus gây hại hành tỏi nhƣ OYDV, SYSV và LYSV.
Các potyvirus gây hại trên mía nhƣ SCMV, SrMV (hai virus này cùng gây bệnh trên ngô
nhƣng không truyền qua hạt ngô).
Các virus hại khoai tây nhƣ PVY, PVX

70
7.5 TRUYỀN QUA HẠT GIỐNG (SEED)
7.5.1 Phân biệt 2 khái niệm
―Seed-borne‖ (virus tồn tại trong/trên hạt giống): tồn tại trên/trong hạt nhƣng chƣa chắc
truyền bệnh sang cây con.
―Seed transmitable‖ (virus truyền qua hạt giống): tồn tại trên/trong hạt và truyền bệnh
sang cây con.
Phát hiện thấy virus trên hạt nhƣng chƣa chắc cây đã bị bệnh.
7.5.2 Tầm quan trọng
Truyền qua hạt giống là một trong các cách lan truyền virus thực vật quan trọng. Virus có
thể tồn tại lâu dài trong hạt dẫn tới tăng khả năng phát tán cao cả theo không gian và thời gian.
Khoảng 20 % virus thực vật truyền qua hạt giống. Một số các chi virus có virus truyền qua
hạt giống quan trọng là:
Alfamovirus (1/1)
Comovirus (6/15)
Cucumovirus (3/3)
Ilarvirus (8/17)
Nepovirus (17/40)
Potyvirus (16/179)
Sobemovirus (4/14)
Tobamovirus (7/17)
Tobravirus (3/3)
7.5.3 Đặc điểm của virus truyền qua hạt giống
1. Khả năng truyền virus qua hạt giống khác nhau theo loại cây. Khoảng 2/3 virus truyền qua
hạt giống nhiễm trên cây họ đậu nhƣng rất ít virus nhiễm trên cây hòa thảo truyền qua hạt
giống.
2. Các virus truyền qua hạt giống chủ yếu tồn tại trong phôi hạt (embryo). Nhìn chung phần lớn
các virus thực vật có thể nhiễm vào các bộ phận khác nhau của hạt (ví dụ vỏ hạt, nội nhũ).
Tuy nhiên trong quá trình chín của hạt, các virus tồn tại bên ngoài phôi hạt thƣờng bị bất
hoạt. Chỉ một số ít các virus có thể tồn tại bên ngoài phôi hạt là các tobamovirus nhƣ TMV,
ToMV, Cucumber green mottle mosaic virus (CGMMV) và một sobemovirus là Southern
bean mosaic virus (SBMV).
3. Không phải tất cả các virus tồn tại trong hạt đều nhiễm sang cây con. Nhiều virus nhiễm vào
phôi nhƣng cũng tồn tại dƣới dạng bất hoạt ở các bộ phận khác của hạt. Ví dụ, trong một thí
nghiệm, 50% hạt đậu có phản ứng ELISA dƣơng với Pea seed borne mosaic virus (PSbMV)
nhƣng chỉ có 2—3 % có mang virus có sức sống (có khả năng truyền virus sang cây con).
4. Khả năng truyền qua hạt nhờ noãn phụ thuộc khả năng nhiễm sớm vào hoa của virus.
5. Hai yếu tố chính qui đinh tỷ lệ nhiễm qua hạt giống của một virus là (i) tƣơng tác ký sinh ký
chủ và (ii) thời điểm nhiễm.

71
6. Virus truyền qua hạt giống có khả năng truyền qua tiếp xúc cơ học và thƣờng nhiễm tế bào
nhu mô. Các virus giới hạn phloem nhìn chung không truyền qua hạt giống.
7. Hạt nhiễm virus có thể hình thành trên cây khỏe do hạt đƣợc thụ phấn từ hạt phấn mang
virus. Tỷ lệ truyền virus sang cây con qua hạt phấn thấp hơn nhiều so với truyền qua noãn.
7.5.4 Các ví dụ về virus truyền qua hạt giống ở Việt Nam
TMV/ToMV tồn tại trên vỏ hạt thuốc lá, cà chua
ZYMV tồn tại trong phôi hạt cây bầu bí, BCMV tồn tại trong phôi hạt cây họ đậu.
7.6 LAN TRUYỀN QUA MÔI GIỚI (CỰC KỲ QUAN TRỌNG)
Truyền qua môi giới là con đƣờng lan truyền quan trọng nhất của virus ngoài tự nhiên. Phần
lớn các bệnh virus thực vật lan truyền ngoài tự nhiên nhờ môi giới. Các môi giới của virus là côn
trùng, nhện hại cây, tuyến trùng, nấm, tơ hồng. Môi giới còn đƣợc gọi là vectơ truyền bệnh.
Nhóm môi giới quan trọng nhất của virus thực vật là côn trùng, trong đó, nhóm côn trùng
chích hút bộ Hemiptera nhƣ rệp muội, bọ rầy, bọ phấn là các nhóm chịu trách nhiệm truyền
nhiều virus có ý nghĩa kinh tế. Khoảng 75 % virus thực vật lan truyền nhờ vector côn trùng.
Bảng 7-1 Thành phần vector của virus thực vật tính đến 2007 (Hogenhout et al., 2008)
Bộ Nhóm vector
Nhóm virus
Tổng % RNA,
Cầu đa
diện
RNA,
Gậy, sợi DNA
RNA,
vỏ bọc
Hemiptera
Rệp muội (aphids) 26 153 13 5 197 28
Bọ phấn (whiteflies) - 13 115 - 128 18
Rầy lá (Leafhoppers) 8 - 15 3 26 4
Rầy thân (planthoppers) 10 4 - 4 18 3
Khác - 8 5 - 13 2
Thysanoptera Bọ trĩ (thrips) 2 - - 14 16 2
Coleoptera Bọ cánh cứng (beetles) 50 1 - - 51 7
Acari Nhện (mites) 10 9 - - 10 1
Nematoda Tuyến trùng 45 3 - - 48 7
Mycota Nấm 8 16 - - 24 3
Vector chƣa xác định 84 60 19 3 166 24
Tổng 133 268 167 30 697
% 33 39 24
7.7 CƠ CHẾ TRUYỀN VIRUS NHỜ VECTOR CÔN TRÙNG
7.7.1 Các kiểu truyền
Gồm 4 kiểu truyền, đặc điểm mỗi kiểu truyền đƣợc trình bày tóm tắt ở Bảng 7.2.
1. Không bền vững (non-persistence)
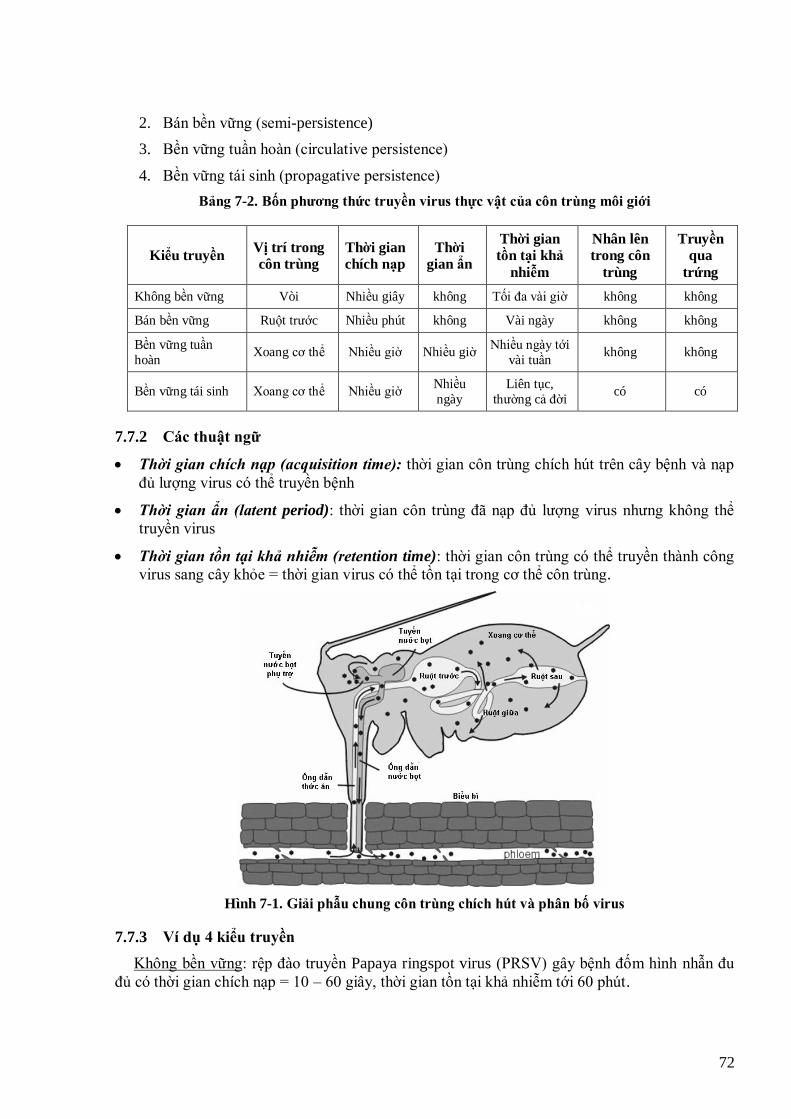
72
2. Bán bền vững (semi-persistence)
3. Bền vững tuần hoàn (circulative persistence)
4. Bền vững tái sinh (propagative persistence)
Bảng 7-2. Bốn phƣơng thức truyền virus thực vật của côn trùng môi giới
Kiểu truyền Vị trí trong
côn trùng
Thời gian
chích nạp
Thời
gian ẩn
Thời gian
tồn tại khả
nhiễm
Nhân lên
trong côn
trùng
Truyền
qua
trứng
Không bền vững Vòi Nhiều giây không Tối đa vài giờ không không
Bán bền vững Ruột trƣớc Nhiều phút không Vài ngày không không
Bền vững tuần
hoàn Xoang cơ thể Nhiều giờ Nhiều giờ
Nhiều ngày tới
vài tuần không không
Bền vững tái sinh Xoang cơ thể Nhiều giờ Nhiều
ngày
Liên tục,
thƣờng cả đời có có
7.7.2 Các thuật ngữ
Thời gian chích nạp (acquisition time): thời gian côn trùng chích hút trên cây bệnh và nạp
đủ lƣợng virus có thể truyền bệnh
Thời gian ẩn (latent period): thời gian côn trùng đã nạp đủ lƣợng virus nhƣng không thể
truyền virus
Thời gian tồn tại khả nhiễm (retention time): thời gian côn trùng có thể truyền thành công
virus sang cây khỏe = thời gian virus có thể tồn tại trong cơ thể côn trùng.
Hình 7-1. Giải phẫu chung côn trùng chích hút và phân bố virus
7.7.3 Ví dụ 4 kiểu truyền
Không bền vững: rệp đào truyền Papaya ringspot virus (PRSV) gây bệnh đốm hình nhẫn đu
đủ có thời gian chích nạp = 10 – 60 giây, thời gian tồn tại khả nhiễm tới 60 phút.

73
Bán bền vững: rầy xanh đuôi đen truyền Rice tungo bacilliform virus (RTBV) gây bệnh
tungro trên lúa có thời gian chích nạp = 30 phút, thời gian tồn tại khả nhiễm tới 4 ngày.
Bền vững tuần hoàn: bọ phấn truyền African cassava mosaic virus (ACMV) gây bệnh khảm
lá sắn có thời gian chích nạp tối thiểu 3.5 giờ, thời kỳ ẩn = 8 giờ, thời gian tồn tại khả nhiễm
khoảng 9 giờ.
Bền vững tái sinh: rầy nâu truyền Rice grassy stunt virus (RGSV) gây bệnh lúa cỏ có thời
gian chích nạp = 1 giờ, thời kỳ ẩn = 3-28 ngày, thời gian tồn tại khả nhiễm cả đời.
7.8 HÀNH VI CHÍCH HÚT CỦA CÔN TRÙNG BỘ HEMIPTERA
Các côn trùng chích hút bộ Hemiptera (gọi chung là hemipterans) có cấu trúc phần miệng
thích hợp cho quá trình chích hút dịch cây và đƣợc gọi vòi chích (stylets). Đối với nhóm truyền
virus theo kiểu không bền vững và bán bền vững thì phần miệng cũng chính lá nơi bám dính của
virus. Vòi chích của chúng có cấu trúc đủ để xuyên qua vách tế bào mà không làm tổn thƣơng tế
bào. Hành vi chích hút của các hemipterans tƣơng tự nhau, nhƣng chỉ có aphid có khả năng
truyền virus theo kiểu không bền vững (xem phần cơ chế và tính đặc hiệu)
Khi bay, hemipterans không thể phân biệt đƣợc đâu là ký chủ thích hợp, nơi chúng có thể hút
dịch cây, sinh sản, phát triển thành quần thể. Ngay khi hạ cánh trên một cây (chú ý hemipteran
có xu tính với màu vàng nên cây có lá biến vàng dễ hấp dẫn chúng), việc đầu tiên là hemipteran
thực hiện các cú chích hút thăm dò (probing). Khi chích hút, chúng tiết 1 giọt nƣớc bọt dạng keo
dính và nhanh chóng chọc vòi vào tế bào biểu bì để thử thức ăn. Các cú chích hút thăm dò này
thƣờng nông (có nghĩa trên lớp tế bào biểu bì) và thƣờng kéo dài ≤ 1 phút. Sau khi thăm dò trên
tế bào biểu bì, và thấy thức ăn phù hợp, chúng tiếp tục chọc vòi sâu hơn, thƣờng len lỏi giữa các
tế bào, cho tới tận ống rây của mạch phloem. Quá trình này có thể mất nhiều phút tới nhiều giờ.
Giọt nƣớc bọt dạng keo dính này sẽ hình thành 1 lớp bao vòi chích (style sheath) của vector Tuy
nhiên, khi chọc với vào tới mạch phloem, chúng lại bơm một loại nƣớc bọt thứ 2, dạng nƣớc,
chứa nhiều enzyme vào ống rây. Khi rút vòi ra, loại nƣớc bọt thứ 2 này sẽ bịt kín lớp bao vòi
chích.
Hành vi chích hút của vetor cho thấy quá trình chích hút thăm dò sẽ thích hợp để truyền các
virus thuốc nhóm không bền vững hoặc bán bền vững vì các virus này, có mặt ở khắp mọi nơi,
kể cả lớp tế bào biểu bì. Trái lại chỉ khi vòi chích của vector đạt tới mạch phloem, chúng mới có
khả năng truyền các virus nhóm bền vững vì các virus này thƣờng giới hạn ở mạch phloem.
7.9 LAN TRUYỀN VIRUS THEO KIỂU KHÔNG BỀN VỮNG
7.9.1 Lan truyền virus thực vật của aphid họ Aphididae
Trong số côn trùng, aphid là nhóm tiến hóa hóa nhất về khả năng sử dụng thức ăn là thực vật.
Do vậy có thể xem aphid là nhóm vector quan trọng nhất của virus thực vật. Aphid truyền
khoảng 50 % số virus thực vật truyền qua vector côn trùng.
Aphid có một số lợi thế để trở thành một vector hiệu quả của virus thực vật:
Nhiều loài aphid là đa thực (polyphagous). Ví dụ rệp đào (Myzus pesicae) có phổ ký chủ
rất rộng tới 50 họ thực vật.
Chúng có thể sinh sản theo kiểu trinh sinh (parthenogenesis), do đó mật độ quần thể có
thể gia tăng nhanh chóng.

74
Có cấu tạo vòi chích và hành vi chích hút thích hợp với lan truyền virus, đặc biệt đối với
các virus tồn tại ở biểu bì (phần lớn các virus).
Bảng 7-3. Các họ, chi virus truyền qua aphid và phƣơng thức truyền (James & Perry, 2004)
Phƣơng thức Họ Chi Số loài Ví dụ virus
Không bền
vững
Bromoviridae Alfamovirus 1 Alfalfa mosaic virus
Bromoviridae Cucumovirus 3 Cucumber mosaic virus
Comoviridae Fabavirus 4 Broad bean wilt virus-1
Potyviridae Macluravirus 2 Maclura mosaic virus
Potyviridae Potyvirus 91 Potato virus Y
Unassigned Carlavirus 31 Carnation latent virus
132
Bán bền
vững
Caulimoviridae Caulimovirus 9 Cauliflower mosaic virus
Closteroviridae Closterovirus 8 Beet yellows virus
Sequiviridae Sequivirus 2 Parsnip yellow fleck virus§
Sequiviridae Waikavirus 3 Anthriscus yellows virus
Unassigned Unassigned 1 Black raspberry necrosis virus
Unassigned Unassigned 1 Strawberry mottle virus
24
Bền vững
tuần hoàn
Luteoviridae Enamovirus 1 Pea enation mosaic virus-1
Luteoviridae Luteovirus 2 Barley yellow dwarf virus
Luteoviridae Polerovirus 5 Potato leaf roll virus
Luteoviridae Umbravirus 7 Carrot mottle virus‡
Unassigned Nanovirus 4 Banana bunchy top virus
Unassigned Sobemovirus† 11 Blueberry shoestring virus
30
Bền vững tái
sinh
Rhabdoviridae Cytorhabdovirus$ 8 Lettuce necrotic yellows virus
Rhabdoviridae Nucleorhabdovirus‡ 7 Sonchus yellow net virus
15
Tổng 201
7.9.2 Truyền virus bằng aphid theo kiểu không bền vững
Trong số các virus truyền qua aphid thì phần lớn truyền theo kiểu không bền vững
(132/201, 66 %, năm 2000). Cần chú ý rằng: Chỉ aphid mới có thể truyền đƣợc virus theo kiểu
không bền vững.
7.9.3 Đặc điểm truyền theo kiểu không bền vững
1. Thời gian chích nạp ngắn, thƣờng chỉ cần vài giây. Các nghiên cứu cho thấy hiệu quả truyền
virus nhóm không bền vững của aphid tỷ lệ thuận với số cú chích hút thăm dò ở lớp tế bào
biểu bì và ngay khi chúng chọc vòi xuống phía dƣới lớp tế bào biểu bì tới các tế bào nhu mô
hoặc bó mạch thì hiệu quả truyền bệnh sẽ suy giảm nhanh chóng. Các nghiên cứu cũng cho

75
thấy, nếu bỏ đói rệp thì hiệu quả truyền virus cũng tăng vì làm tăng các cú chích hút thăm
dò.
2. Quá trình chích nạp và chích truyền diễn ra tại tế bào biểu bì. Các virus truyền theo kiểu này
thƣờng không đặc hiệu mô và có mặt ở tế bào biểu bì của cây.
3. Virus chỉ tồn tại ở tầng cutile ở phần vòi mà tầng này sẽ bị tách bỏ sau mỗi lần lột xác =>
virus sẽ mất sau mỗi lần lột xác => vector trở nên sạch virus => virus không thể truyền qua
giao phối cũng nhƣ qua thế hệ sau.
4. Virus có thời gian tồn tại khả nhiễm ngắn, tối đa vài giờ. Nhìn chung, khả năng truyền virus
của rệp giảm ngay lập tức sau khi chích nạp. Tốc độ mất khả năng truyền virus phụ thuộc
nhiều yếu tố nhƣng có lẽ chủ yếu do bản chất tồn tại chỉ ở phần vòi của của virus.
5. Virus không có thời gian ẩn có nghĩa sau khi chích nạp, vector có thể truyền ngay virus.
6. Việc giải phóng virus từ vector trong quá trình chích truyền chƣa đƣợc hiểu rõ nhƣng vì
virus tồn tại ở phần vòi nên 2 cơ chế đã đƣợc đề xuất: ―chỉ do hành vi bơm nƣớc bọt‖, (ii)
kết hợp giữa ―bơm nƣớc bọt và chớ (regurgitation) thức ăn‖.
7.9.4 Cơ chế tƣơng tác virus - vector
Nhìn qua kiểu truyền thấy có vẻ không có liên hệ sinh học giữa virus và vector. Tuy nhiên
các bằng chứng khoa học cho thấy có sự tƣơng tác giữa protein của virus và phần vòi của vector.
Tƣơng tác này qui định khả năng, hiệu quả truyền và tính đặc hiệu virus – vector. Hiện có 2 mô
hình tƣơng tác giữa virus và vector:
Mô hình vỏ protein (tƣơng tác trực tiếp)
Theo mô hình này, vỏ protein của phân tử virus sẽ liên kết với receptor nằm phía trong vòi
chích của aphid. Tƣơng tác giữa vỏ protein virus với vòi của rệp qui định cả khả năng truyền lẫn
tính đặc hiệu. Không có tƣơng tác tác này, virus sẽ không thể truyền đƣợc bằng aphid. Mô hình
này đã đƣợc chứng minh ở nhiều virus, đặc biệt là CMV (Cucumber mosaic virus).
CMV là một virus hình cầu đa diện, với bộ gen gồm 3 phân tử RNA (RNA-1, RNA-1 và
RNA-3), sợi đơn, cực (+). Virus có phổ ký chủ rộng nhất trong số các virus thực vật và lan
truyền bằng nhiều loài rệp muôi theo kiểu không bền vững. Vỏ protein của virus có kích thƣớc
218 amino acid. Ngƣời ta đã chứng minh rằng thay đổi các aa ở 2 vị trí 129 và 162 có thể làm
một chủng CMV mất hoặc phục hồi khả năng truyền nhờ rệp bông Aphis gosypii; và thay đổi ở
các vị trí 24, 168 và 214 có thể làm một chủng CMV vốn chỉ truyền đƣợc bằng rệp bông có thể
truuyền đƣợc bằng rệp đào M. persicae.
Mô hình bắc cầu (tƣơng tác gián tiếp)
Theo mô hình này, vòi của aphid sẽ liên kết với một protein vỏ của phân tử virus thông qua
một cầu nối là một protein cũng của virus gọi là protein hỗ trợ. Mô hình này đã đƣợc nghiên
cứu rất kỹ đối với các potyvirus.
Potyvirus là các virus có phân tử dạng sợi mềm, có bộ gen RNA sợi đơn, cực (+). Số lƣợng
các potyvirus lớn nhất trong số các virus thực vật (khoảng 200 virus). Các potyvirus lan truyền
theo kiểu không bền vững bằng nhiều loài aphid và gây nhiều bệnh có ý nghĩa kinh tế (Ví dụ
PRSV gây bệnh đốm hình nhẫn đu đủ, PVY gây bệnh khảm lá khoai tây…).
Tất cả các potyvirus mã hóa cho 1 protein hỗ trợ ký hiệu là Hc-Pro (helper component
protein). Hc-Pro có một motif gồm 4 aa bảo thủ (KITC) nằm ở đầu amin chịu trách nhiệm tƣơng
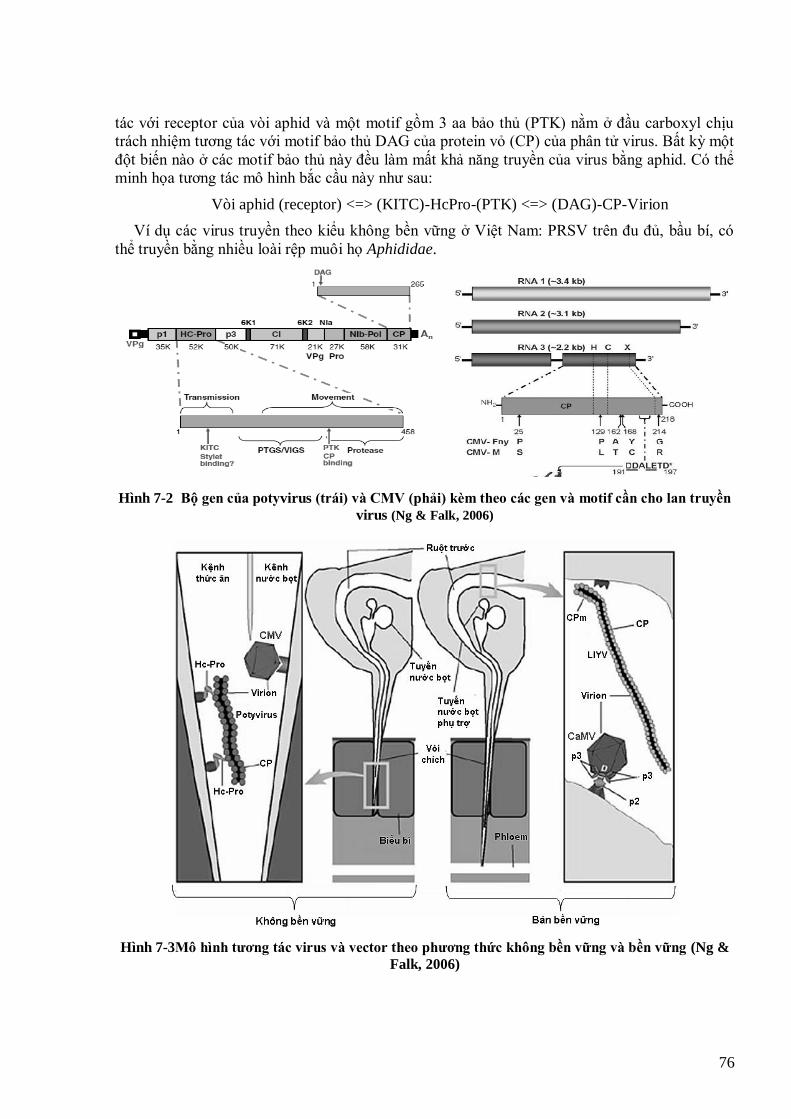
76
tác với receptor của vòi aphid và một motif gồm 3 aa bảo thủ (PTK) nằm ở đầu carboxyl chịu
trách nhiệm tƣơng tác với motif bảo thủ DAG của protein vỏ (CP) của phân tử virus. Bất kỳ một
đột biến nào ở các motif bảo thủ này đều làm mất khả năng truyền của virus bằng aphid. Có thể
minh họa tƣơng tác mô hình bắc cầu này nhƣ sau:
Vòi aphid (receptor) <=> (KITC)-HcPro-(PTK) <=> (DAG)-CP-Virion
Ví dụ các virus truyền theo kiểu không bền vững ở Việt Nam: PRSV trên đu đủ, bầu bí, có
thể truyền bằng nhiều loài rệp muôi họ Aphididae.
Hình 7-2 Bộ gen của potyvirus (trái) và CMV (phải) kèm theo các gen và motif cần cho lan truyền
virus (Ng & Falk, 2006)
Hình 7-3Mô hình tƣơng tác virus và vector theo phƣơng thức không bền vững và bền vững (Ng &
Falk, 2006)

77
7.10 LAN TRUYỀN VIRUS THEO KIỂU BÁN BỀN VỮNG
7.10.1 Đặc điểm
1. Nhiều nhóm côn trùng khác nhau có kiểu truyền bán bền vững; trong khi chỉ aphid mới có
kiểu truyền không bền vững
2. Trái với kiểu truyền không bền vững, ở kiểu truyền bán bền vững, quá trình chích nạp cũng
nhƣ chích truyền phần lớn xảy ra ở mạch phloem. Điều này giải thích ở kiểu truyền này, thời
gian chích nạp dài hơn, tới nhiều phút và nhiều virus truyền theo kiểu bán bền vững thƣờng
cũng khó truyền bằng tiếp xúc cơ học.
3. Virus chỉ tồn tại ở tầng cutile phần ruột trƣớc (foregut) => tầng này sẽ bị tách bỏ sau mỗi
lần lột xác => giống nhƣ ở nhóm không bền vững virus sẽ mất sau mỗi lần lột xác => vector
trở nên sạch virus => virus không thể truyền qua giao phối cũng nhƣ qua thế hệ sau.
4. Virus có thời gian tồn tại khả nhiễm dài hơn so với nhóm không bền vững, có thể tới vài
ngày.
5. Virus không có thời gian ẩn có nghĩa sau khi chích nạp, vector có thể truyền ngay virus.
6. Việc giải phóng virus từ vector trong quá trình chích truyền chƣa đƣợc hiểu rõ nhƣng có lẽ
do hành vi chớ thức ăn ―regurgitation‖ vì virus tồn tại ở ruột trƣớc.
7.10.2 Tƣơng tác virus - vector
Tƣơng tác vector – virus ở nhóm này dựa trên cả mô hình ―vỏ protein‖ và mô hình ― bắc cầu‖
với sự tham gia của một hoặc nhiều protein hỗ trợ.
Tƣơng tác trực tiếp theo mô hình ―vỏ protein‖ đã đƣợc chứng minh với các virus họ
Closteroviridae nhƣ CTV (Citrus tristeza virus) gây bệnh tàn lụi cây có múi và đặc biệt LIYV
(Lettuce infectious yellows virus) gây bệnh vàng lá rau diếp. Các virus thuộc họ Closteroviridae
là các virus RNA, sợi đơn cực (+) và có hình thái dạng sợi dài mềm. Trong khi CTV truyền
bằng nhiều loài aphid thì LIYV lại truyền bằng bọ phấn theo kiểu bán bền vững. Cả 2 virus đều
có vỏ protein đƣợc cấu tạo từ 4 -5 phân tử protein vỏ khác nhau, trong đó có một protein đƣợc
dịch mã ít trong tế bào gọi là CPm (minor coat protein) đóng vai trò tƣơng tác trực tiếp với vòi
của vector.
Tƣơng tác gián tiếp theo cơ chế ―bắc cầu‖ đã đƣợc chứng minh đối với CaMV, một DNA
virus có phân tử hình cầu với bộ gen DNA dạng vòng kép, lan truyền bằng nhiều loài rệp theo
kiểu bán bền vững. CaMV có tới 3 protein (p2, p3 và p4 – là protein vỏ) tham gia vào quá trình
lan truyền qua vector. Protein p2 sẽ liên kết với vòi của rệp (và do đó nó đóng vai trò là protein
bổ trợ). Protein p3 lại liên kết với p4. Để lan truyền đƣợc, p2 sẽ liên kết với p3 và tạo cầu nối
hoàn chỉnh để virus có thể truyền đƣợc qua vector.
Tƣơng tác gián tiếp còn đƣợc chứng minh với virus tungro. Bệnh tungro là bệnh nguy hiểm
nhất trên lúa. Bệnh gây ra do 2 virus thuộc 2 đơn vị phân loại khác hẳn nhau cùng xâm nhiễm là
RTBV (một DNA virus, chi Badnavirus, có bộ gen DNA dạng vòng kép) và RTSV (một RNA
virus, chi Waikavirus, có bộ gen RNA mạch thẳng, sợi đơn, cực (+)). Bệnh do rầy xanh đuôi đen
(Nephotettix virescens) truyền theo kiểu bán bền vững. RTSV truyền dễ dàng bằng rầy xanh
đuôi đen nhƣng không tạo triệu chứng bệnh. RTBV không thể truyền đƣợc bằng rầy xanh đuôi
đen (nếu bằng cách nào đó, chẳng hạn bằng agroinoculation, chỉ nhiễm một mình trên cây lúa thì
cũng tạo triệu chứng nhẹ). RTBV chỉ lan truyền nhờ rầy khi rầy chích trên cây nhiễm hỗn hợp 2
virus hoặc chích trên cây nhiễm RTSV trƣớc. Khi rầy chích hút trên cây bị nhiễm RTSV, một

78
protein phi cấu trúc của virus này sẽ liên kết với vòi của rầy và đóng vai trò là một protein hỗ trợ
để liên kết với RTBV và do đó tạo điều kiện truyền virus này. Trong trƣờng hợp này RTSV
đƣợc gọi là virus hỗ trợ cho lan truyền RTBV bằng rầy.
7.11 LAN TRUYỀN VIRUS THEO KIỂU BỀN VỮNG TUẦN HOÀN
Các virus truyền theo kiểu bền vững tuần hoàn gồm các virus thuộc họ Luteoviridae,
Geminiviridae và Nanoviridae. Tất cả các virus này đều có phân tử dạng hình cầu đa diện đối
xứng. Các virus họ Luteoviridae và Nanoviridae chỉ truyền bằng aphid còn các virus họ
Geminiviridae thì phần lớn đƣợc truyền nhờ bọ phấn (chi Begomovirus) hoặc bằng rầy (chỉ các
topocuvirus và mastrevirus).
7.11.1 Đặc điểm
1. Quá trình chích nạp cũng nhƣ chích truyền phần lớn xảy ra ở mạch phloem => thời gian
chích nạp và chích truyền khá lâu và phần lớn virus có kiểu truyền này thì rất khó hoặc
không thể truyền đƣợc bằng tiếp xúc cơ học vì virus thƣờng chỉ giới hạn ở mạch phloem.
2. Virus tồn tại trong xoang cơ thể nhƣng nhìn chung không tồn tại ở cơ quan sinh sản. Điều
này dẫn tới:
(i) Virus sẽ không bị mất sau mỗi lần lột xác.
(ii) Virus nhìn chung không truyền qua giao phối cũng nhƣ qua thế hệ sau.
3. Tuyến luân chuyển của virus: cây bệnh => vòi chích => ruột trƣớc => ruột giữa, ruột sau =>
xoang cơ thể => tuyến nƣớc bọt => cây. Nhƣ vậy trong cơ thể côn trùng, virus nhóm bền
vững tuần hoàn phải vƣợt qua ít nhất 2 rào cản:
(i) Từ ruột vào xoang cơ thể
(ii) Từ xoang cơ thể vào tuyến nƣớc bọt
4. Virus có thời gian tồn tại khả nhiễm dài, thƣờng kéo dài nhiều ngày tới nhiều tuần.
5. Virus có thời gian ẩn khá dài vì phải vƣợt qua nhiều rào cản tế bào.
6. Việc giải phóng virus từ vector trong quá trình chích truyền do hành vi bơm nƣớc bọt mang
virus vào phloem.
7.11.2 Quan hệ virus vector
7.11.2.1 Họ Luteoviridae.
Các virus họ Luteoviridae gồm 3 chi là Enamovirus, Luteovirus, và Polerovirus. Chúng đều
là các virus RNA, sợi đơn, cực (+). Hai virus có thể có mặt ở Việt Nam là PLRV (Potato leaf
roll virus) trên khoai tây và một bệnh khá nghiêm trọng trên bông ở Việt Nam là Cotton leafroll
dwarf virus (CLRDV) gây bệnh xanh lùn.
Vỏ protein của các luteovirus đƣợc cấu tạo bởi 2 protein là CP (coat protein), đƣợc dịch mã
nhiều trong tế bào cây và một protein là RT đƣợc dịch mã ít (RT là read through, tức khi dịch
mã ribosome sẽ bỏ qua mã ngừng của protein trƣớc). Cả 2 protein CP và RT đều cần cho tƣơng
tác đặc hiệu virus – aphid. Đặc biệt, RT là protein qui định tính đặc hiệu vị trí xâm nhập của
virus ở ruột (vào chỉ ruột giữa hay cả ruột giữa và ruột sau).

79
7.11.2.2 Họ Geminiviridae
Các geminivirus là các virus DNA sợi vòng đơn. Trong số các geminivirus, các begomovirus
chiếm số lƣợng lớn nhất (~ 20% tổng số virus thực vật). Các begomovirus chỉ mã hóa một
protein vỏ (CP) và tính đặc hiệu vector do CP qui định.
Ở cả luteovirus và begomovirus, ngƣời ta đã chứng minh có sự tƣơng tác giữa phân tử virus
với 1 protein của vi khuẩn nội sinh phổ biến trong vector (vi khuẩn Buchnera). Protein này
(GroEL) thuộc nhóm symbionin và có chức năng bảo vệ phân tử virus khỏi bị phân hủy trong
xoang cơ thể côn trùng. Ngƣời ta đã lợi dụng đặc điểm này để chuyển gen mã hóa GroEL vào
cây cà chua để chống các begomovirus gây bệnh xoăn vàng lá.
7.12 TRUYỀN THEO KIỂU BỀN VỮNG TÁI SINH
Tất cả các virus thực vật có vỏ bọc đều truyền theo kiểu bền vững tái sinh. Các virus thực vật
có vỏ bọc chỉ thuộc 2 họ là Bunyaviridae (đều truyền bằng bọ trĩ) và Rhabdoviridae (phần lớn
truyền bằng rầy). Một số virus có vỏ bọc có ý nghĩa kinh tế là:
Virus héo đốm cà chua (TSWV, Tomato spotted wilt virus, họ Buniaviridae) gây bệnh
trên nhiều cây trồng và phân bố khắp thế giới (truyền bằng bọ trĩ).
Virus vàng lụi (vàng lá Bắc Giang) (RYSV, Rice yellow stunt virus, họ Rhabdoviridae)
đang gây bệnh vàng lùn (vàng lá) ở miền Bắc (truyền bằng rầy xanh đuôi đen).
Các virus thực vật không có vỏ bọc mà truyền theo phƣơng thức bền vững tái sinh đều truyền
qua rầy. Các virus này thuộc 2 họ là Reoviridae, Tymoviridae và các virus thuộc chi Tenuivirus.
Một số virus thuộc nhóm này là các virus có ý nghĩa ở Việt Nam
Virus lùn sọc đen phƣơng Nam (SRBSDV, họ Reoviridae), truyền bằng rầy lƣng trắng,
rầy nâu nhỏ
Virus lùn xoắn lá (RRSV, họ Reoviridae), truyền bằng rầy nâu.
Virus lúa cỏ (RGSV, chi Tenuivirus), truyền bằng rầy nâu.
7.12.1 Đặc điểm
1. Nhìn chung nhóm này có nhiều đặc điểm chung với virus nhóm bền vững tái sinh.
2. Quá trình chích nạp cũng nhƣ chích truyền phần lớn xảy ra ở mạch phloem => thời gian
chích nạp và chích truyền rất lâu và các virus có kiểu truyền này thì rất khó hoặc không thể
truyền đƣợc bằng tiếp xúc cơ học vì virus thƣờng chỉ giới hạn ở mạch phloem.
3. Virus tồn tại trong xoang cơ thể và nhiều trƣờng hợp ở cả cơ quan sinh sản. Điều này dẫn
tới:
(i) Virus sẽ không bị mất sau mỗi lần lột xác
(ii) Virus nhìn chung có truyền qua giao phối cũng nhƣ qua thế hệ sau
4. Tuyến luân chuyển của virus: cây bệnh => vòi chích => ruột trƣớc => ruột giữa, ruột sau =>
xoang cơ thể => tuyến nƣớc bọt/cơ quan sinh sản => cây. Nhƣ vậy trong cơ thể côn trùng,
virus nhóm bền vững tái sinh phải vƣợt qua ít nhất 3 rào cản:
(i) Từ ruột vào xoang cơ thể
(ii) Từ xoang cơ thể vào tuyến nƣớc bọt

80
(iii) Từ xoang cơ thể vào cơ quan sinh sản nhƣ buồng trứng.
5. Virus nhân lên trong cơ thể côn trùng. Điều này dẫn tới virus có nhiều đặc điểm chung với
các virus động vật về đặc tính xâm nhập tế bào, sinh sản và lắp ráp phân tử.
6. Virus có thời gian tồn tại khả nhiễm dài, thƣờng kéo dài cả đời.
7. Virus có thời gian ẩn khá dài vì cần thời gian cho virus nhân lên.
8. Việc giải phóng virus từ vector trong quá trình chích truyền do hành vi bơm nƣớc bọt đƣa
virus vào phloem.
7.12.2 Ví dụ lan truyền nhờ bọ trĩ theo kiểu bền vững tái sinh
Các virus lan truyền nhờ bọ trĩ theo kiểu bền vững tái sinh đều thuộc chi Tospovirus với
virus điển hình là TSWV. Bọ trĩ là một vector hiệu quả vì chúng là các loài đa thực
Các tospovirus có virion hình cầu, kích thƣớc từ 80 – 120 nm (của tospovirus là từ 80 – 110
nm). Virion có vỏ bọc (envelope), trên có gai nhỏ cấu tạo bằng glycoprotein (Gn, Gc), phần
cuống gai nằm chìm trong một lớp màng kép lipid. Bên trong là 3 phân tử nucleoprotein và một
số transcriptase (RdRp).
Hình 7-4 Sơ đồ phân tử TSWV (Hogenhout et al., 2008)
Mối quan hệ bọ trĩ – tospovirus khá không bình thƣờng vì bọ trĩ trƣởng thành không thể
chích nạp virus và chỉ có thể truyền đƣợc nếu sâu non chích nạp virus. .
Bọ trĩ Frankliniella occidentalis là một vector rất hiệu quả vì truyền 5/14 tospovirus.
Tospovirus phải đƣơng đầu với khoảng 6 loại tế bào và màng khác nhau kể từ khi chúng
đƣợc chích nạp qua vòi và quay trở lại cây khi bọ trĩ chích truyền. Cơ chế lan truyền của
tospovirus có thế đƣợc mô tả nhƣ sau.
Ngay khi đƣợc chích nạp, phân tử virus di chuyển theo ruột trƣớc tới ruột giữa là vị trí chủ
yếu để virus xâm nhập vào tế bào vector. Ngoài cùng thành ruột giữa có một lớp tế bào biểu mô
(epithelial cells) dạng cột đƣợc ngăn cách với ống ruột bởi 1 lớp màng đỉnh (apical membrane)
[1]. Do cấu tạo của lớp tế bào biểu mô nên thành ruột giống nhƣ có một lớp lông (microvilii).
Virus sẽ xâm nhập qua lớp màng đỉnh để nhiễm vào tế bào biểu mô theo lối ―nhập bào‖, tức là
sau khi gai vỏ của virus nhận biết các thụ thể trên bề mặt tế bào biểu mô thì lớp vỏ bọc của virus
sẽ dung hợp với màng tế bào biểu mô và sau đó giải phóng nucleoprotein và enzyme RdRp của
virus vào bên trong tế bào chất của tế bào biểu mô. Tại tế bào biểu mô, virus sẽ sinh sản và tiếp

81
theo thoát khỏi tế bào biểu mô, vƣợt qua lớp màng thứ 2 là lớp màng đáy (basement membrane)
[2] để xâm nhập vào tế bào cơ bao quanh ruột. Tại tế bào cơ, virus tiếp tục sinh sản và hình
thành nhiều viroplasm [hình nhỏ, 3]. Sau khi virus mới đƣợc hình thành, chúng thoát khỏi các tế
bào cơ ruột và di chuyển qua màng đáy [hình nhỏ, 4] của tuyến nƣớc bọt [5], tiếp tục di chuyển
qua lớp màng đỉnh của tuyến nƣớc bọt [6] để vào ống dẫn của tuyến nƣớc bọt và theo dòng nƣớc
bọt để xâm nhập vào cây khi bọ trĩ chích hút cây.
Hình 7-5 Các cơ quan bên trong của bọ trĩ liên quan đến lan truyền tospovirus và các rào cản dạng
màng mà virus phải vƣợt qua. Các số tƣơng ứng với các loại màng và tế bào liên quan đến đƣờng
đi của virus từ khi đƣợc chích nạp đến khi đƣợc chích truyền. L, lumen (tuyến dẫn); PSg, primary
salivary gland (tuyến nƣớc bọt sơ cấp); s, cross sections of muscle (tiết diện cắt ngang cơ); VP,
viroplasm (vị trí tái sinh và lắp ráp virus); DM, dense mass (vị trí có màu đậm khi chụp dƣới kính
hiển vi) (Hogenhout et al., 2008).
7.13 CÂU HỎI ÔN TẬP CHƢƠNG 7
1. Các phƣơng thức lan truyền của virus thực vật, phƣơng thức lan truyền quan trọng nhất
ngoài tự nhiên.
2. Đặc điểm truyền qua hạt giống (vidụ TMV, ZYMV).
3. Các nhóm vector của virus thực vật.
4. Đặc điểm các kiểu truyền virus nhờ vector côn trùng. So sánh thuật ngữ ―bền vững‖ và
―không bền vững‖.
5. Kiểu truyền không bền vững (đặc điểm, tƣơng tác phân tử của cơ chế trực tiếp và bắc cầu, ví
dụ).
6. Kiểu truyền bán bền vững (đặc điểm, tƣơng tác phân tử của cơ chế trực tiếp và bắc cầu, ví
dụ)
7. Kiểu truyền bền vững tuần hoàn (đặc điểm, vai trò của vi khuẩn nội sinh, ví dụ)
8. Kiểu bền vững tái sinh (đặc điểm, ví dụ).

82
9. Đặc điểm truyền tospovirus nhờ bọ trĩ, so sánh bản chất virus, cơ chế nhân lên trong bọ trĩ
với virus động vật.
7.14 TÀI LIỆU THAM KHẢO CHÍNH
1. Andret-Link, P. & Fuchs, M. (2005). Transmission specificity of plant viruses by vectors.
Journal of Plant Pathology 87, 153-165.
2. Ghanim, M., Morin, S., Zeidan, M. & Czosnek, H. (1998). Evidence for transovarial
transmission of tomato yellow leaf curl virus by its vector, the whitefly Bemisia tabaci.
Virology 240, 295-303.
3. Hogenhout, S. A., Ammar, E., Whitfield, A. E. & Redinbaugh, M. G. (2008). Insect Vector
Interactions with Persistently Transmitted Viruses. Annual Review of Phytopathology 46,
327–359.
4. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
5. Jones, D. R. (2003). Plant viruses transmitted by whiteflies. European Journal of Plant
Pathology 109, 195–219.
6. Morin, S., Ghanim, M., Sobol, I. & Czosnek, H. (2000). The GroEL protein of the whitefly
Bemisia tabaci interacts with the coat protein of transmissible and nontransmissible
begomoviruses in the yeast two-hybrid system. Virology 276, 404-416.
7. Moritz, G., Kumma, S. & Moundb, L. (2004). Tospovirus transmission depends on thrips
ontogeny. Virus Research 100, 143–149.
8. Ng, J. C. & Perry, K. L. (2004). Transmission of plant viruses by aphid vectors. Molecular
Plant Pathology 5, 505–511.
9. Ng, J. C. K. & Falk, B. W. (2006). Virus-Vector Interactions Mediating Nonpersistent and
Semipersistent Transmission of Plant Viruses. Annual Review of Phytopathology
10. 44, 183–212.
11. Power, A. G. (2000). Insect transmission of plant viruses: a constraint on virus variability.
Current Opinion in Plant Biology 3, 336–340.
12. Syller, J. (2006). The roles and mechanisms of helper component proteins encoded by
potyviruses and caulimoviruses. Physiological and Molecular Plant Pathology 67, 119–130.
13. Whitfield, A. E., Ullman, D. E. & German, T. L. (2005). Tospovirus-Thrips Interactions.
Annual Review of Phytopathology 43, 459–489.

83
Chƣơng 8. PHÒNG CHỐNG BỆNH VIRUS THỰC VẬT
8.1 TÓM TẮT NỘI DUNG
Nội dung chƣơng này giới thiệu các nguyên lý phòng chống bệnh virus, các phƣơng pháp
hiện đang đƣợc sử dụng để phòng chống virus thực vật dựa vào tính kháng tự nhiên và tính
kháng chuyển gen. Bản chất của gen kháng virus cũng nhƣ tính kháng hình thành thông qua cơ
chế câm gen (gene silencing) đƣợc thảo luận kỹ.
8.2 CÁC CHIẾN LƢỢC PHÒNG CHỐNG BỆNH VIRUS
Không giống nhƣ đối với nấm, vi khuẩn và tuyến trùng, hiện nay có thể thấy không có thuốc
hóa học tiêu diệt trực tiếp virus trong cây. Tuy nhiên, ngƣời ta vẫn có thể phòng chống virus
thực vật bằng nhiều chiến lƣợc khác nhau. Các chiến lƣợc chính bao gồm:
1. Sử dụng giống sạch bệnh.
2. Sử dụng giống kháng bệnh dựa vào:
Tính kháng tự nhiên thông qua gen kháng của cây.
Tính kháng từ tác nhân gây bệnh.
3. Tạo tính kháng tập nhiễm hệ thống (SAR, systemic acquired resistance).
4. Tiêu diệt nguồn virus (trên cây vụ trƣớc, trên ký chủ phụ, nhổ bỏ cây bệnh vụ hiện tại)
5. Phòng chống vector (hóa học, sinh học, nhà lƣới, bẫy…)
Tùy thuộc đặc điểm sinh học và dịch tễ học của mỗi loại virus mà ngƣời ta có thể sử dụng
một hay nhiều chiến lƣợc khác nhau.
8.3 SỬ DỤNG VẬT LIỆU GIỐNG SÁCH BỆNH
Phƣơng pháp này áp dụng đối với các virus truyền qua vật liệu giống (hạt giống, các cây nhân
giống vô tính). Vật liệu giống sạch bệnh có thể đƣợc tạo ra bằng một số cách nhƣ:
1. Chọn giống từ nguồn sạch (ruộng không bị bệnh, khu vực không bị bệnh)
2. Xử lý nhiệt.
3. Xử lý hóa chất để tiêu diệt virus tồn tại trên vỏ hạt.
4. Nuôi cấy đỉnh sinh trƣởng.
5. Tạo giống invitro.
6. Sử dụng hóa chất (ribavirin) để tiêu diệt virus trong nuôi cấy mô.
8.4 SỬ DỤNG GIỐNG KHÁNG BỆNH MANG GEN KHÁNG CỦA CÂY
8.4.1 Khái niệm gen –đối - gen
Quan hệ gen-đối-gen (gene-for-gene) do Harol Henry Flor (1900-1991), một nhà bệnh
cây học ngƣời Mỹ xây dựng. Mặc dù không phải là ngƣời đầu tiên sử dụng thuật ngữ gen-đối-
gen, nhƣng ông là ngƣời đầu tiên chứng minh rằng biểu hiện bệnh là hậu quả mối tƣơng tác của
cả tác nhân gây bệnh và ký chủ và di truyền đƣợc.

84
Định nghĩa ―đối với mỗi gen qui định tính kháng trong cây ký chủ có một gen tƣơng ứng qui
định tính không độc trong ký sinh và 2 gen này tƣơng tác đặc hiệu với nhau‖
Quan hệ gen-đối-gen cho tới nay đã đƣợc chứng minh là tồn tại trong rất nhiều loại bệnh do
hầu hết các nhóm tác nhân gây bệnh (nấm, vi khuẩn, virus, tuyến trùng, mollicus...) gây ra.
Các gen cây ký chủ qui định tính kháng đƣợc gọi chung là gen R (Resistance). Các gen ký
sinh qui định tính không độc của ký sinh đƣợc gọi chung là gen Avr (Avirulence).
Tính kháng của cây trồng tuân theo quan hệ gen-đối-gen đƣợc gọi là tính kháng gen-đối-gen.
Tính kháng gen-đối-gen thƣờng là tính kháng đặc hiệu ký chủ, tính kháng đơn gen, tính kháng
gen chủ, tính kháng không bền vững. Hậu quả của một phản ứng kháng gen-đối-gen thƣờng là
phản ứng siêu nhạy/apoptosis.
8.4.2 Gen kháng R và protein R của cây chống virus
8.4.2.1 Gen kháng trội
Cho tới nay, rất nhiều gen kháng R của cây đã đƣợc xác định. Sản phẩm của các gen này gọi
là các protein kháng R.
Hiện nay, các protein kháng R đƣợc chia thành 5 lớp dựa trên đặc điểm cấu trúc và vị trí hoạt
động của chúng trong tế bào ký chủ (Hình 8-1). Các protein trong cùng lớp nhìn chung khá bảo
thủ. Các protein kháng R của cây chống virus phần lớn thuộc 2 lớp CNL và TNL. Đây là 2
lớp chiếm số lƣợng lớn nhất trong tất cả các loại protein kháng R của thực vật.
CNL (CC-NB-LRR). Là các protein có cấu trúc gồm (1) một vùng khóa kéo leucine
(leucine-zipper – LZ) hoặc một chuỗi xoắn kép (coiled-coil – CC) ở đầu amin; (2) một vùng
có khả năng liên kết nucleotide (nucleotide binding – NB) ở vùng trung tâm; và (3) một
vùng lặp giàu leucine (leucine-rich repeats - LRR) phía đầu carboxyl. Các protein của lớp
này hoạt động trong tế bào chất.
TNL (TIR-NB-LRR). Là các protein kháng chỉ có ở cây 2 lá mầm. Về cấu trúc, lớp protein
TNL tƣơng tự lớp CNL nhƣng thay vì chuỗi CC là một chuỗi TIR. Chuỗi TIR là một vùng
protein tƣơng đồng với receptor Toll (của ruồi dấm) và receptor Interleukin-1 (của ngƣời)
(TIR = Toll/Interleukin-1 Receptor). Tƣơng tự protein kháng lớp CNL, các protein của lớp
TNL cũng hoạt động trong tế bào chất.
Cả 2 lớp CNL và TNL còn đƣợc xếp vào chung một họ protein kháng gọi là họ NB-LRR.
Số lƣợng gen kháng của họ này đƣơc xem là chiếm số lƣợng lớn nhất trong các họ protein.

85
Hình 8-1. Các lớp gen kháng R và một số ví dụ mỗi lớp
8.4.2.2 Gen kháng lặn
Các lớp gen kháng R chống virus ở trên là các gen kháng trội. Tuy nhiên, có một nhóm khá
lớn các gen kháng lặn chống virus đã đƣợc xác định trên nhiều loại cây. Phần lớn các gen kháng
lặn này đƣợc nghiên cứu trên cây chống potyvirus. Các nghiên cứu cho thấy nhiều gen cây ký
chủ khi bị đột biến lại tạo ra tính kháng đối với virus. Một trong các gen kháng lặn quan trọng
nhất, phổ biến nhất và đƣợc ký hiệu khác nhau trên các cây khác nhau là gen mã hóa yếu tố khởi
đầu dịch mã eIF4E. Môt số ví dụ về các các gen này là pvr1 (trên ớt), mo1 (trên rau diếp), sbm1
(trên đậu), rym4/5 (trên lúa miến). Dạng isoform của eIF4E là eIF(iso)4E cũng có vai trò tƣơng
tự. Quá trình dịch mã của potyvirus là quá trình độc lập Cap. Tuy nhiên, các yếu tố ký chủ, trong
đó có eIF4E hoặc eIF(iso)4 đều cần thiết cho quá trình này. Nếu các yếu tố phiên mã của ký chủ
mất chức năng thì quá trình dịch mã của potyvirus không thể thực hiện đƣợc và hậu quả là cây
kháng bệnh.
8.4.3 Các gen Avr của virus
Không giống nhƣ gen/protein kháng R của cây, nhìn chung, gen/protein Avr của tác nhân
gây bệnh chia sẻ ít đặc điểm chung với nhau.
Đối với virus thực vật, các gen Avr thƣờng liên quan đến 1 protein chức năng của virus.
Nhiều gen Avr của virus thực vật là protein vỏ CP.
Đối với gen mã hóa yếu tố khởi đầu dịch qui định tính kháng lặn chống potyvirus thì Avr
tƣơng ứng của virus chính là VPg.
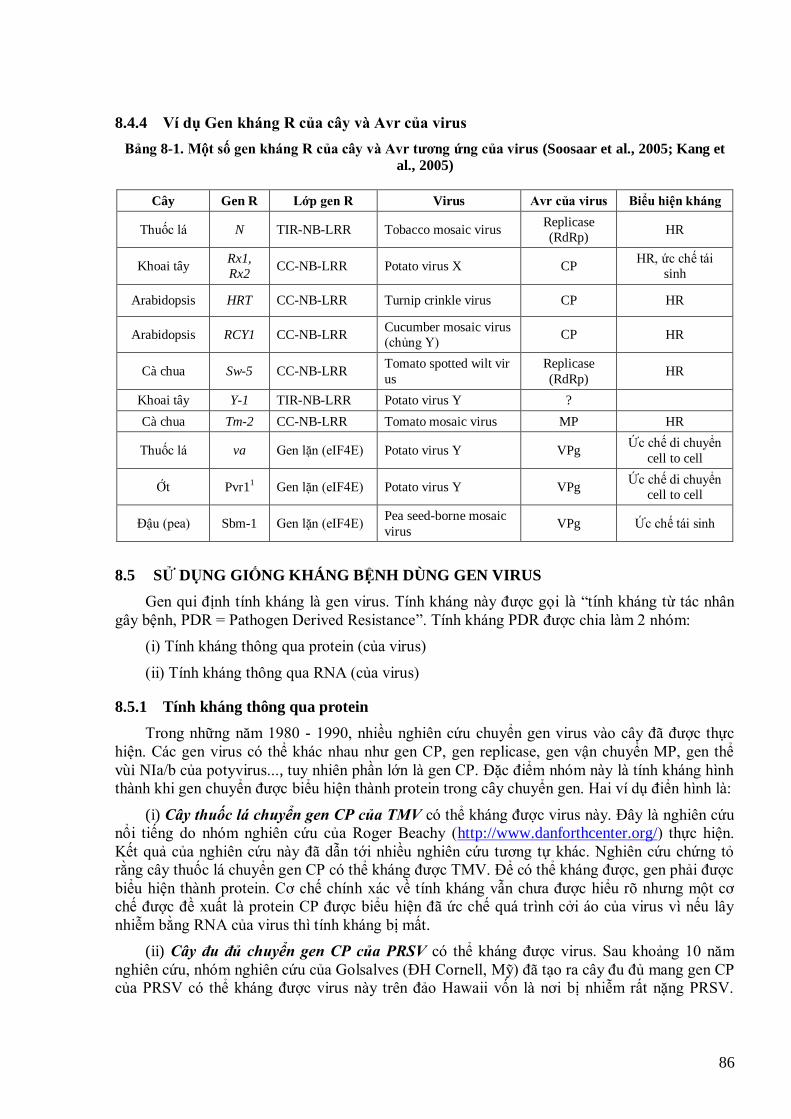
86
8.4.4 Ví dụ Gen kháng R của cây và Avr của virus
Bảng 8-1. Một số gen kháng R của cây và Avr tƣơng ứng của virus (Soosaar et al., 2005; Kang et
al., 2005)
Cây Gen R Lớp gen R Virus Avr của virus Biểu hiện kháng
Thuốc lá N TIR-NB-LRR Tobacco mosaic virus Replicase
(RdRp) HR
Khoai tây Rx1,
Rx2 CC-NB-LRR Potato virus X CP
HR, ức chế tái
sinh
Arabidopsis HRT CC-NB-LRR Turnip crinkle virus CP HR
Arabidopsis RCY1 CC-NB-LRR Cucumber mosaic virus
(chủng Y) CP HR
Cà chua Sw-5 CC-NB-LRR Tomato spotted wilt vir
us
Replicase
(RdRp) HR
Khoai tây Y-1 TIR-NB-LRR Potato virus Y ?
Cà chua Tm-2 CC-NB-LRR Tomato mosaic virus MP HR
Thuốc lá va Gen lặn (eIF4E) Potato virus Y VPg Ức chế di chuyển
cell to cell
Ớt Pvr11 Gen lặn (eIF4E) Potato virus Y VPg Ức chế di chuyển
cell to cell
Đậu (pea) Sbm-1 Gen lặn (eIF4E) Pea seed-borne mosaic
virus VPg Ức chế tái sinh
8.5 SỬ DỤNG GIỐNG KHÁNG BỆNH DÙNG GEN VIRUS
Gen qui định tính kháng là gen virus. Tính kháng này đƣợc gọi là ―tính kháng từ tác nhân
gây bệnh, PDR = Pathogen Derived Resistance‖. Tính kháng PDR đƣợc chia làm 2 nhóm:
(i) Tính kháng thông qua protein (của virus)
(ii) Tính kháng thông qua RNA (của virus)
8.5.1 Tính kháng thông qua protein
Trong những năm 1980 - 1990, nhiều nghiên cứu chuyển gen virus vào cây đã đƣợc thực
hiện. Các gen virus có thể khác nhau nhƣ gen CP, gen replicase, gen vận chuyển MP, gen thể
vùi NIa/b của potyvirus..., tuy nhiên phần lớn là gen CP. Đặc điểm nhóm này là tính kháng hình
thành khi gen chuyển đƣợc biểu hiện thành protein trong cây chuyển gen. Hai ví dụ điển hình là:
(i) Cây thuốc lá chuyển gen CP của TMV có thể kháng đƣợc virus này. Đây là nghiên cứu
nổi tiếng do nhóm nghiên cứu của Roger Beachy (http://www.danforthcenter.org/) thực hiện.
Kết quả của nghiên cứu này đã dẫn tới nhiều nghiên cứu tƣơng tự khác. Nghiên cứu chứng tỏ
rằng cây thuốc lá chuyển gen CP có thể kháng đƣợc TMV. Để có thể kháng đƣợc, gen phải đƣợc
biểu hiện thành protein. Cơ chế chính xác về tính kháng vẫn chƣa đƣợc hiểu rõ nhƣng một cơ
chế đƣợc đề xuất là protein CP đƣợc biểu hiện đã ức chế quá trình cởi áo của virus vì nếu lây
nhiễm bằng RNA của virus thì tính kháng bị mất.
(ii) Cây đu đủ chuyển gen CP của PRSV có thể kháng đƣợc virus. Sau khoảng 10 năm
nghiên cứu, nhóm nghiên cứu của Golsalves (ĐH Cornell, Mỹ) đã tạo ra cây đu đủ mang gen CP
của PRSV có thể kháng đƣợc virus này trên đảo Hawaii vốn là nơi bị nhiễm rất nặng PRSV.

87
Hiện nay, cây đu đủ chuyển gen đang chiếm một tỷ lệ lớn tại Hawaii và kỹ thuật này đã đƣợc
chuyển gia cho một số nƣớc nhƣ Brazil, Thái Lan, Đài Loan, Jamaica, Venezuela, Bangladesh,
Tanzania, Uganda, và Kenya. Khi áp dụng công nghệ này cần chú ý là cây đu đủ chuyển gen chỉ
kháng đƣợc các chủng virus tƣơng ứng với gen chuyển. Ngoài ra, các vấn đề liên quan tới sinh
vật biến đổi gen GMO (genetically modified organism) cũng gây trở ngại cho chuyển giao công
nghệ này rộng rãi hơn.
Hình 8-2. Minh họa tính kháng rất cao và bền vững đối với PRSV của giống đu đủ Rainbow
chuyển gen CP so với giống không chuyển gen Sunrise tại Hawai (Golsalves et al., 2004)
Do thành công của chƣơng trình này tại Mỹ nên một số cây trồng chuyển gen dùng kỹ thuật
tƣơng tự cũng đƣợc tạo ra và đƣợc thƣơng mại hóa, chủ yếu tại Mỹ nhƣ:
Các cây dƣa chuyển gen CP chống water melon virus 2 (WMV2), zuchini yellow mosaic
virus (ZYMV), cucumber mosaic virus (CMV).
Khoai tây chuyển gen CP chống PVY, chuyển gen replicase chống Potato leaf roll virus
(PLRV).
8.5.2 Tính kháng thông qua RNA
Cũng trong những năm 1980, một số nghiên cứu chuyển gen kháng bệnh virus đã chứng
minh rằng gen chuyển không nhất thiết phải biểu hiện thành protein. Chẳng hạn các gen vệ tinh
của Cucumber mosaic virus (CMV) hay Tobacco ringspot virus (TRSV) cũng tạo tính kháng
trên cây chuyển gen. Tiếp theo các nhà khoa học thấy rằng, gen chuyển đƣợc lấy từ vùng không
mã hóa cũng tạo ra tính kháng.
Một nghiên cứu quan trọng trong thời gian này, do nhóm Dougherty (Mỹ) thực hiện, là
chuyển gen mã hóa CP của Tobacco eatch virus (TEV) vào thuốc lá để chống virus. Các quan
sát cho thấy các dòng chuyển gen có tính kháng mạnh nhất là các dòng không hoặc có rất ít
mRNA của gen chuyển và cây chuyển gen lúc đầu bị nhiễm thì hàm lƣợng mRNA cao, dần dần
khi cây trở nên kháng thì hàm lƣợng mRNA của gen chuyển cũng dần biến mất. Các tác giả cho
rằng tính kháng này liên quan đến 1 cơ chế gọi là ―câm gen hậu phiên mã‖ (posttranscriptional
gene silencing, PTGS).
Hiện nay, tính kháng của virus thông qua RNA đã đƣợc chứng minh là do cơ chế câm gen
(RNA silencing) hay là cơ chế kiểm soát RNA (RNA interference) của các sinh vật nhân chuẩn.

88
Tính kháng virus của thực vật thông qua cơ chế câm gen có thể tóm tắt qua các bƣớc sau
(Hình 8-3):
1. Hình thành các phân RNA sợi kép (dsRNA) của virus trong tế bào chất
2. Enzyme Dicer của tế bào ký chủ cắt các phân tử dsRNA thành các phân tử RNA nhỏ
(siRNA) có kích thƣớc 21-22 nucleotides
3. Lắp ráp siRNA (sợi đơn) thành phức hợp RISC (RNA induced silencing complex)
4. Phức hợp RISC dƣới sự hƣớng dẫn của siRNA sẽ cắt đặc hiệu các phân tử RNA tƣơng
ứng (RNA virus hoặc các phân tử transcript của nó)
Hình 8-3. Cơ chế câm gen của thực vật chống virus và các các acid nucleic lạ
8.5.3 Hiện tƣợng câm gen
8.5.3.1 Công trình của Craig Mello và Andrew Fire
Năm 1998, Craig Mello và Andrew Fire (và 4 tác giả khác) đã công bố trên tạp chí Nature
một công trình nghiên cứu nhằm tìm hiểu vai trò của RNA đến sự biểu hiện gen. Các tác giả đã
sử dụng một số gen của tuyến trùng Caenorhabditis elegens nhƣ unc-22 (mã hóa protein
myofilament, liên quan đến co cơ), unc-54 (mã hóa protein myosin liên quan đến co cơ), fem-1
(mã hóa protein chứa ankyrin liên quan đến sinh sản), hlh-1 (mã hóa protein họ myoD cần cho
di chuyển và biệt hóa hình dạng), và mex (mã hóa cho 1 protein có nhiều ở phôi). Ba phát hiện
quan trọng từ công trình này là:
Các đoạn dsRNA tƣơng ứng với vùng intron hoặc promoter không tạo ra hiện tƣợng câm
(silencing) của gen tƣơng ứng. Nhƣ vây sự can thiệp (interfering) của RNA là ở mức hậu
phiên mã (post-transcriptional level).
Khi tiêm các RNA ở dạng cùng chiều (sense) hay ngƣợc chiều (antisense) vào tuyến trùng đã
không ảnh hƣởng tới lƣợng mRNA của gen tƣơng ứng tức không làm ảnh hƣởng tới sự biểu

89
hiện của gen. Tuy nhiên khi tiêm các RNA ở dạng sợi kép dsRNA (hỗn hợp của RNA cùng
chiều và ngƣợc chiều) đã làm câm (silence) gen tƣơng ứng. Nhƣ vậy hiện tƣợng câm gen
(gene silencing) đƣợc thực hiện thông qua trung gian là RNA sợi kép (dsRNA).
Khi tiêm dsRNA vào một vị trí thì hiện tƣợng câm gen xuất hiện ở các vị trí khác trong cơ
thể tuyến trùng. Nhƣ vậy dấu hiệu câm gen có thể di truyển hệ thống.
Hiện tƣợng câm gen do dsRNA đã đƣợc các tác giả đặt tên là RNA interfering (sự can thiệp
của RNA). Vì nghiên cứu này, Craig Mello và Andrew Fire đã đƣợc trao giải Nobel y học năm
2006.
8.5.3.2 Lịch sử khám phá siRNA
siRNA đã đƣợc khám phá bởi Hamilton và Baulcombe năm 1999. Các tác giả đã chuyển
gen aco, gus vào cây cà chua và thuốc lá. Trên các cây biểu hiện hiện tƣợng PTGS, các tác giả
đã phát hiện đƣợc các phân tử RNA nhỏ, kích thƣớc khoảng 25 nucleotides, đặc hiệu, nhƣng
ngƣợc chiều với gen chuyển (chứng tỏ không phải là sản phẩm phân hủy của mRNA của các gen
trên). Ngoài ra các tác giả cũng phát phát hiện thấy sau khi lây nhiễm potato virus X (PVX) 4
ngày, trên cây cũng hình thành các phân tử kích thƣớc khoảng 25 nucleotides, đặc hiệu nhƣng
ngƣợc nghĩa với PVX.
8.5.3.3 Định nghĩa siRNAs
siRNAs là các phân tử RNA sợi kép nhỏ, kích thƣớc khoảng 21-25 nucleotides, đƣợc tạo ra
bởi Dicer, một RNA endonuclease nhóm III, là thành phần quan trong của phức hợp RISC gọi là
siRISC có chức năng phân hủy mRNA đồng dạng của nó.
8.5.3.4 Nguồn gốc siRNAs
Phần lớn siRNA có nguồn gốc từ mRNAs, các yếu tố di động (transposons), virus hoặc
DNA dị nhiễm sắc thể (heterochromatic DNA). Nhƣ vậy siRNA có cùng nguồn gốc với gen mục
tiêu.
8.5.3.5 Sinh tổng hợp siRNAs
Quá trình tạo ra các phân tử siRNA có thể đƣợc tóm tắt qua các bƣớc sau:
Hình thành phân tử phiên mã sợi kép dài (dsRNA). Đầu tiên là hình thành các sản phẩm
RNA sợi kép. Các sản phẩm sợi kép này có thể là các gen chuyển đƣợc thiết kế ngƣợc chiều
nhau, hoặc là các sản phẩm trung gian trong quá trình tái sinh của virus.
Hình thành siRNA thành thục. Các phân tử dsRNA đƣợc Dicer cắt thành các phân tử RNA
nhỏ sợi kép có kích thƣớc khoảng 22 nucleotides. Do bị cắt bởi Dicer, các phân tử này có
đầu so le với gốc phosphat ở đầu 5‘ và 2-3 nucleotides đầu 3‘ (overhang). Tiếp theo, phân tử
RNA nhỏ sợi kép này đƣợc đƣợc tách bởi helicase thành 2 phân tử sợi đơn. Các phân tử
siRNA thành thục sợi đơn này sẽ tham gia hình thành phức hợp RISC.
8.5.3.6 Vai trò của siRNA
RNA silencing thông qua siRNA là (i) một cơ chế phòng thủ chống virus, (ii) ngăn chặn sự
biểu hiện quá mức hoặc không cần thiết của các mRNA, và (iii) bảo vệ bộ gen khỏi bị gián đoạn
bởi transposon.

90
8.5.4 Công nghệ RNAi trong tạo giống kháng bệnh virus
8.5.4.1 Thiết kế cấu trúc chuyển gen
Đây là bƣớc quan trọng nhất, quyết định thành công của một chƣơng trình tạo giống kháng
thông qua RNAi (RNA interference).
Có 3 cách để thiết kế cấu trúc chuyển gen virus
- Tạo cấu trúc chứa gen đồng nghĩa (sene).
- Tạo cấu trúc chứa gen ngƣợc nghĩa (antisense).
- Tạo cấu trúc chứa gen theo cả 2 chiều đồng nghĩa và ngƣợc nghĩa (cấu trúc tự bắt cặp).
Tất cả các loại cấu trúc trên muốn tạo tính kháng thì đều phải hình thành cấu trúc dsRNA
trong cây chuyển gen. Có nhiều cách để có thể hình thành cấu trúc dsRNA của gen chuyển.
Chẳng hạn, các cấu trúc đồng nghĩa và ngƣợc nghĩa có thể (i) đƣợc chuyển đồng thời vào cây,
hoặc (ii) đƣợc chuyển riêng biệt để tạo các dòng chứa gen đồng nghĩa hoặc ngƣợc nghĩa. Các
dòng này sẽ đƣợc lai để tạo dòng mang cả gen đồng nghĩa và ngƣợc nghĩa, hoặc (iii) trong
trƣờng hợp cây chỉ đƣợc chuyển với cấu trúc đồng nghĩa hoặc ngƣợc nghĩa thì RdRp của cây sẽ
tạo ra các phân tử dsRNA từ các transcripts của gen đƣợc chuyển.
Tuy nhiên, chỉ loại cấu trúc tự bắt cặp đã đƣợc chứng tỏ là hiệu quả hơn cả trong việc tạo ra
tính kháng. Hiện nay, hầu hết các nghiên cứu tạo tính kháng thông qua RNAi trên cây đều sử
dụng các cấu trúc tự bắt cặp (hình).
Đối với cấu trúc tự bắt cặp, nếu 2 chuỗi gen virus bao quanh một chuỗi intron thì hiệu quả
PTGS sẽ tăng lên rất nhiều.
Hình 8-4. Hiệu quả PTGS với các cấu trúc chuyển gen potato virus Y (PVY) khác nhau. Hiệu quả
đƣợc tính là % cây chuyển gen miễn nhiễm với PVY; gen virus (PVY-pro) là NIa; chuỗi loop là
chuỗi unidA (GUS) dài ~800 nucleotides; intron là của gen Pdk từ cây Flaveria (Smith et al., 2000).

91
8.5.4.2 Ví dụ tạo cây chuyển gen kháng TYLCV thông qua RNAi
TYLCV là một trong nhiều begomovirus gây bệnh xoăn vàng lá cà chua. Các begomovirus
lan truyền ngoài tự nhiên nhờ bọ phấn (B. tabaci) theo kiểu bền vững tuần hoàn.
TYLCV có bộ gen DNA vòng đơn có kích thƣớc khoảng 2.6 – 2.8 kb, chứa 6 gen đƣợc tổ
chức theo 2 chiều ngƣợc nhau.
Trên chiều kim đồng hồ (chiều virus) có 2 gen là gen mã hóa vỏ protein CP và gen AV2.
Trên chiều ngƣợc kim đồng hồ (chiều sợi tƣơng đồng virus) có 4 gen là (i) C1 mã hóa protein tái
sinh hay còn gọi là protein Rep có chức năng chính là cắt - nối bộ gen virus tại một vị trí đặc
biệt ở vùng nguồn gốc tái sinh và tƣơng tác với protein ký chủ để tạo điều kiện thuận lợi cho tái
sinh virus, (ii) C2 mã hóa 1 protein hoạt hóa phiên mã, (iii) C3 mã hóa 1 protein tăng cƣờng tái
sinh và (iv) C4 mã hóa protein C4 với chức năng liên quan đến phổ ký chủ và phát triển triệu
chứng.
Năm 2006, một nhóm nghiên cứu tại đại học Havana (University of Havana), Cuba đã
chuyển gen Rep của TYLCV và cây cà chua và chứng minh rằng cây kháng virus rất tốt.
Hình 8-5 Tạo cây cà chua chuyển gen kháng TYLCV bằng công nghệ RNAi (Fuentes, 2006)
Cấu trúc chuyển gen kháng TYLCV: (a) Tổ chức bộ gen của TYLCV; (b) Bản đồ đoạn
chứa cấu trúc chuyển gien; (c) Sơ đồ hairpin RNA sau khi intron bị cắt khỏi transcript
CaMV 35S: promoter
Intron: chuỗi intron của Castor bean catalase.
LB, LR: bờ trái và bờ phải.
tNos: chuỗi terminator của nopaline synthase.
nptII: neomycin phosphotransferase II (kháng kanamycine).
pNos: chuỗi promoter của nopaline synthase.

92
8.6 ỨNG DỤNG TÍNH KHÁNG TẠO ĐƢỢC (INDUCED RESISTANCE)
8.6.1 Giới thiệu
Cây trồng trong quá trình tiến hóa đã phát triển một số cơ chế phòng thủ chống lại các ức chế
hữu sinh nhƣ tác nhân gây bệnh truyền nhiễm, côn trùng và cả các ức chế vô sinh nhƣ hạn, nóng
và mặn. Trong những năm 1960, Ross đã quan sát thấy rằng khi lây nhiễm nhân cây thuốc lá với
virus TMV thì cây sẽ hình thành tính kháng hệ thống vì khi lây nhiễm tiếp TMV lần thứ 2 tại vị
trí cách xa điểm lây nhiễm lần đầu thì vết đốm chết hoại hình thành nhỏ hơn. Tính kháng kiểu
này đã đƣợc gọi là Tính kháng tập nhiễm hệ thống (SAR, systemic acquired resistance).
Tiếp theo, ngƣời ta thấy rằng các vi khuẩn vùng rễ vốn thúc đẩy tăng trƣởng rễ cây trồng, khi
phát triển trên rễ cũng tạo ra tính kháng hệ thống của cây. Tính kháng kiểu này đã đƣợc gọi là
Tính kháng hệ thống tạo đƣợc (ISR, induced systemic resistance).
Cả 2 loại tính kháng trên đều là biểu hiện khác nhau của một loại tính kháng đƣợc gọi là Tính
kháng tạo đƣợc (IR, induced resistance).
8.6.2 Định nghĩa
8.6.2.1 Tính kháng tạo đƣợc
Tính kháng tạo đƣợc là ―một trạng thái sinh lý của cây với khả năng phòng thủ đƣợc tăng
cƣờng nhờ đƣợc kích hoạt bởi các kích thích môi trƣờng đặc biệt‖. Nhờ khả năng phòng thủ
đƣợc tăng cƣờng này mà cây có thể chống lại đƣợc sự tấn công tiếp theo của các tác nhân gây
bệnh. Tính kháng tạo đƣợc mang tính phổ rộng, chống lại đƣợc nấm, vi khuẩn, virus, tuyến
trùng, côn trùng. Nhƣ vậy có thể thấy tính kháng tạo đƣợc chỉ hình thành sau khi đƣợc kích thích
bởi nguồn kích hoạt (elicitor).
Dựa vào bản chất của nguồn kích hoạt và đƣờng hƣớng dẫn truyền tín hiệu, tính kháng tạo
đƣợc có thể đƣợc chia làm 2 loại chính là là tính kháng tập nhiễm hệ thống SAR và tính kháng
hệ thống tạo đƣợc ISR (Hình 8-6).
8.6.2.2 Tính kháng tập nhiễm hệ thống SAR
Tính kháng tập nhiễm hệ thống SAR là loại tính kháng tạo đƣợc có tính lƣu dẫn (hệ
thống), phổ rộng (chống lại nhiều tác nhân gây bệnh và thậm chí các ức chế vô sinh), thƣờng
dẫn tới biểu hiện protein PR (pathogenesis-related protein) và thông qua đƣờng hƣớng dẫn
truyền tín hiệu SA (salycilic acid).
Nguồn tạo SAR là vi sinh vật gồm nấm, vi khuẩn, virus…Các vi sinh vật này có thể gây bệnh
hoặc không gây bệnh cho cây. Ngoài ra, SAR cũng có thể là các hóa chất nhƣ SA, INA. BTH….
Dấu hiệu phân tử tín hiệu đặc trƣng của SAR là SA có thể đƣợc tạo ra rất nhanh trong vòng
vài giờ (4-6 giờ) sau lây nhiễm và nhân lên nhanh chóng; sau khoảng 24 giờ SAR đã biểu hiện
toàn cây. Sự di chuyển của SAR thƣờng theo hƣớng từ dƣới gốc lên trên ngọn.
Cây có thể duy trì SAR trong thời gian rất lâu, trong nhiều trƣờng hợp kéo dài cả đời của cây.
Cần chú ý nếu tính kháng tạo đƣợc nhƣng không lƣu dẫn tức tính kháng chỉ hình thành xung
quanh vị trí xâm nhiễm thì đƣợc gọi là tính kháng tập nhiễm cục bộ (local acquired resistance).

93
8.6.2.3 Tính kháng hệ thống tạo ISR
Tính kháng hệ thống tạo ISR là loại tính kháng tạo đƣợc có tính lƣu dẫn (hệ thống), phổ
rộng (chống lại nhiều tác nhân gây bệnh), không dẫn tới biểu hiện proteinPR và thông qua
đƣờng hƣớng dẫn truyền tín hiệu JA/ET (jasmonic acid và ethylen).
Nguồn tạo ISR là vi sinh vật vùng rễ không gây bệnh cho cây, điển hình là các vi khuẩn vùng
rễ nhƣ Pseudomonas, Bacillus hoặc Bradyrhizobium.
Hình 8-6 Hai loại tính kháng tạo đƣợc: tính kháng tập nhiễm hệ thống (SAR) và tính kháng hệ
thống tạo đƣợc (ISR) (Vallad & Goodman, 2004).
8.6.3 Các hợp chất có khả năng cảm ứng SAR
SAR hình thành khi có sự tấn công của vi sinh vật. Tuy nhiên nhiều thí nghiệm đã chứng tỏ
rằng SAR có thể đƣợc tạo ra khi xử lý các hợp chất tự nhiên hoặc tổng hợp. Tất cả các hóa chất
có khả năng cảm ứng SAR đƣợc gọi là các chất kích hoạt SAR (chất kích kháng).
Để có thể đƣợc xem là chất kích hoạt SAR thực sự, một hóa chất hoặc sản phẩm chuyển hóa
của nó phải không có hoạt tính kháng sinh. Một số thuốc hóa học trừ bệnh ngoài hoạt tính kháng
nấm còn có khả năng cảm ứng SAR nhƣ Fosetyl-Al (Aliet), metalaxyl, Cu(OH)2.
Dựa vào nguồn gốc hình thành, các hợp chất có thể cảm ứng tạo SAR bao gồm:
8.6.3.1 Các hóa chất tổng hợp có khả năng cảm ứng SAR
Một số hóa chất tổng hợp có khả năng kích thích cây hình thành SAR đƣợc ứng dụng nhiều
trong nghiên cứu phòng chống virus (và các tác nhân gây bệnh cây khác) là SA, INA và BTH
(Hình 8-7).
Hình 8-7 Cấu trúc hóa học của SA, INA và BTH

94
SA (Salicyclic acid) đƣợc biết là một phytohormone nội bào tham gia dẫn truyền tín hiệu
nhằm điều khiển nhiều quá trình sinh lý của cây nhƣ nảy mầm, tăng trƣởng tế bào, hô hấp, đóng
mở khí khổng, già hóa. SA cũng khởi động các phản ứng kháng chống lại các ức chế (stress) vô
sinh nhƣ hạn, mặn và hữu sinh nhƣ tác nhân gây bệnh. Trong cây SA đƣợc tổng hợp bằng 2
đƣờng hƣớng enzyme:
Từ chorismate nhờ enzyme Isochrosimate synthase (ICS) và isochrosimate pyruvate lyase
(IPL).
Từ phenylalanine (một amino acid), nhờ enzym Phenyl alanine ammonia lyase (PAL).
Vì SA tự do ở nồng độ cao có thể ảnh hƣởng bất lợi đến các đƣờng hƣớng sinh hóa của cây
nên trong cây nên để tránh nồng độ gây độc, SA đƣợc cây tổng hợp ra phần lớn đƣợc chuyển
hóa thành dạng SA O-β-glucoside (SAG) và đƣợc dự trữ trong không bào, ngoài ra SA cũng
đƣợc chuyển hóa thành các dạng methyl salicylate (MeSA), methyl salicylate O-β-glucoside
(MeSAG), và salicyloyl glucose ester (SGE). Tất cả các hợp chất này là dạng bất hoạt của SA và
không gây độc tế bào. Khi cần thiết, các hợp chất trên lại đƣợc chuyển hóa lại thành SA.
Sử dụng SA ngoại bào (tức là phun dung dịch SA lên cây) đã đƣợc chứng minh cảm ứng tạo
tính kháng SAR đối với nhiều tác nhân gây bệnh nhóm sinh dƣỡng gồm cả nấm, vi khuẩn và
virus. Tuy nhiên vì nồng độ hiệu quả để tạo tính kháng của SA (> 100 µM) có thể gây độc tế bào
nên SA tự do không đƣợc ứng dụng ngoài thực tiễn để tạo tính kháng.
INA (dichloroisonicotinic acid). INA có cơ chế cảm ứng SAR giống nhƣ SA chống lại nhiều
tác nhân gây bệnh. INA có thể cảm ứng SAR trƣớc hoặc sau khi lây nhiễm. Điểm khác biệt so
với SA là INA cảm ứng SAR độc lập với SA và hoạt động ở phía hạ lƣu đƣờng hƣớng dẫn
truyền tín hiệu so với SA. Tƣơng tự nhƣ SA, INA có thể gây độc cho cây nên cũng không đƣợc
ứng dụng ngoài thực tiễn.
BTH (benzo(1,2,3)-thiadiazole-7-carbothiolic acid (BTH, acibenzolar-S-methyl). BTH có cơ
chế tạo tính kháng SAR giống nhƣ SA và có thể chống đƣợc nhiều nhóm tác nhân gây hại kể cả
virus. BTH có hiệu quả chống nấm Cercospora nicotianae, Peronospora tabacina,
Phytophthora parasitica, nhiều nấm phấn trắng, gỉ sắt và sƣơng mai khác, vi khuẩn
Pseudomonas syringae, virus TMV, CMV và TSWV (Hình 8-8). Điểm quan trọng của BTH so
với các hợp chất tạo tính kháng SAR khác là BTH không gây độc cho cây. Chính vì vậy, BTH
đã đƣợc ứng dụng trong thực tiễn (thƣơng mại hóa) làm thuốc kích kháng. Sản phẩm thƣơng mại
của BTH gồm BION (hãng Syngenta) hoặc Actiguard (hãng Novartis).
Hình 8-8. Cây thuốc lá xử lý BTH (Bion) (trái) ức chế biểu hiện triêu chứng bệnh trên thuốc lá do
TSWV gây ra. Đối chứng không xử lý (phải) có triệu chứng dữ dội (Mandal et al., 2007)

95
8.6.3.2 Các thuốc BVTV có hoạt tính tạo SAR
Một số thuốc bảo vệ thực vật, bao gồm cả thuốc trừ bệnh và thuốc trừ sâu, ngoài khả năng ức
chế trực tiếp dịch hại cũng có hoạt tính tạo tính kháng SAR. Các ví dụ của nhóm này bao gồm:
Thuốc trừ nấm chứa gốc phosphate nhƣ fosetyl-Al (Al(PO3)3) (ví dụ thuốc thƣơng mại là
Aliette®). Đây là thuốc đặc hiệu với nấm Phytophthora. Ngoài tác động trực tiếp của fosetyl-Al
do giải phóng H3PO3 thì thuốc này (và cả các chứa gốc phosphate khác nhƣ phosphate kali) có
khả năng cảm ứng tạo SAR trong cây. Muối phosphate có thể cô lập ion Ca+2
dẫn tới phá hủy
vách tế bào và tạo thành các oligomer có khả năng cảm ứng SAR, tăng cƣờng hoạt tính enzyme
PAL và tích lũy SA.
Thuốc trừ nấm Probenazole (tên thƣơng mại Oryzemate, đặc trị nấm đạo ôn lúa, không đăng
ký tại Việt Nam) tăng cƣờng biểu hiện protein PR, tăng cƣờng hoạt tính enzyme PAL và tích lũy
SA trên lúa và thuốc lá.
Thuốc trừ nấm nhóm Strobilurin. Đây là các thuốc có có nguồn gốc từ nấm mũ Strobilurus
tenacellus. Nhóm thuốc này có nhiều hoạt chất khác nhau. Một trong số các hoạt chất là
Pyraclostrobin (một sản phẩm thƣơng mại chứa hoạt chất này là Cabrio Top, có đăng ký tại Việt
Nam). Xử lý thuốc này trên thuốc lá làm tăng tính kháng TMV do thuốc kích thích biểu hiện các
enzyme oxidase ở ty thể, làm tăng biểu hiện protein PR. Thuốc cũng làm tăng tính chịu hạn,
giảm sự già hóa và kéo dài thời gian chín của quả, hạt trên nhiều cây trồng (thuốc có ―hiệu ứng
xanh‖).
Thuốc trừ sâu Imidacloprid (ví dụ tên thƣơng mại Confidor, Admire). Đây là thuốc trừ côn
trùng chích hút nhƣng cũng tạo tính kháng SAR nhƣ thuốc trừ nấm nhóm strobilurin.
8.6.4 Hóa chất có khả năng cảm ứng tính kháng tạo đƣợc khác
8.6.4.1 Chitosan
Chitosan là polymer mạch thẳng đƣợc cấu tạo bởi liên kết β-(1-4) của 2 tiểu phần D-
glucosamine (tiểu phần bị loại bỏ nhóm acetyl) và tiểu phần N-acetyl-D-glucosamine (tiểu phần
chứa nhóm acetyl). Chitosan đƣợc hình thành từ sự loại bỏ nhóm acetyl của chitin (dạng
polymer mach thẳng của N-acetyl-D-glucosamine) (Hình 8-9). Chitosan có nhiều ứng dụng
trong nông nghiệp, công nghiệp thực phẩm và y học. Chitosan không đƣợc xem là chất kích
kháng thực sự vì nó có hoạt tính kháng nấm. Tính kháng tạo đƣợc của chitosan rẩt rộng, chống
lại nhiều tác nhân gây bệnh nhƣ nấm, vi khuẩn và virus.
.
Hình 8-9 Sự hình thành chitosan từ chitin

96
Tuy nhiên chitosan có nhƣợc điểm lớn là không hòa tan trong nƣớc cũng nhƣ dung môi hữu
cơ. Để khắc phục nhƣợc điểm này, chitosan đƣợc chuyển thành dạng oligochitosan bằng cách
thủy phân chitosan. Oligochitosan có hoạt tính sinh học mạnh hơn nhiều so chitosan thô. Khả
năng kích kháng của oligochitosan phức tạp, liên quan đến cả đƣờng hƣớng SA và JA/ET và do
đó tạo tính kháng với các tác nhân gây bệnh thuộc cả 2 nhóm biotroph và necrotroph. Mức độ
kháng và tính đặc hiệu của oligochitosan phụ thuộc vào trọng lƣợng phân tử, mức độ acetyl hóa
và pH của nó. Vi dụ hình 8-10 cho thấy oligochitosan có thể ức chế sự xâm nhiễm của TMV
trên thuốc lá.
Hình 8-10 Hiệu quả ức chế của oligochitosan đối với TMV trên thuốc lá. (A) đối chứng, (B)–(F): xử
lý chitosan lần lƣợt ở các nồng độ 1 ppm, 10 ppm, 25 ppm, 50 ppm và 100ppm (Yin et al., 2010).
8.6.4.2 BABA
BABA (Beta-aminobutyric acid). Hợp chất chất này có khả năng tạo tính kháng chống cả các
ức chế vô sinh và hữu sinh. Đƣờng hƣớng hình thành tính kháng tạo đƣợc của BABA khá phức
tạp. Trong một số trƣờng hợp, tính kháng cảm ứng bởi BABA hình thành thông qua tích lũy SA
nhƣng trong nhiều trƣờng hợp khác lại liên quan đến một phytohormone nội sinh khác là ABA
(abssicic acid).
8.6.4.3 Các hợp chất khác
Nhiều loại hợp chất khác, có nguồn gốc tự nhiên hoặc tổng hợp cũng đƣợc biết có khả năng
kích thích cây hình thành tạo tính kháng. Một số ví dụ là vitamin (B1, B2, K), 3-acetonyl-3-
hydroxyindole (AHO), polyamines, catechin, polyacrylic acid, các oligosaccharide có nguồn gốc
từ cây (oligogalacturonide, cellodextrin), một số hợp chất có nguồn gốc từ tác nhân gây bệnh
(glucan, peptide, ergosterol, syringolin).
8.7 CÂU HỎI ÔN TẬP CHƢƠNG 8
1. Các chiến lƣợc phòng chống bệnh virus.
2. Các phƣơng pháp tạo giống sạch bệnh.
3. Sử dụng giống kháng bệnh mang gen kháng có nguồn gốc từ cây (đặc điểm gen kháng R
của cây chống virus và đặc điểm ge Avr của virus; ví dụ).

97
4. Sử dụng giống kháng dựa cơ chế PDR (khái niệm chung; ví dụ tạo cây chuyển gen chống
PRSV)
5. Khái niệm tính kháng thông qua RNA và cơ chế câm gen; ví du tạo cây chuyển gen kháng
TYLCV).
6. Cơ chế và đặc điểm của tính kháng tạo đƣợc và tính kháng tập nhiễm hệ thống SAR
7. Các hóa chất có thể ứng dụng tạo tính kháng SAR chống virus.
8.8 TÀI LIỆU THAM KHẢO CHÍNH
1. Beachy, R. N. (1999). Coat-protein-mediated resistance to tobacco mosaic virus: discovery
mechanisms and exploitation. PhilTrans R Soc Lond B 354, 659-664.
2. Beckers, G. J. & Conrath, U. (2007). Priming for stress resistance: from the lab to the field.
Current Opinion in Plant Biology 10, 425-431.
3. Bent, A. F. & Mackey, D. (2007). Elicitors, effectors, and R genes: The new paradigm and a
lifetime supply of questions. Annu Rev Phytopathol 45, 399-436.
4. Chisholm, S. T., Coaker, G., Day, B. & Staskawic, B. J. (2006). Host-Microbe Interactions:
Shaping the Evolution of the Plant Immune Response. Cell 124, 803–814,.
5. Cieslinska, M. (2007). Application of thermo-and chemotherapy in vitro for eliminating
some viruses infecting Prunus sp. fruit trees. Journal of Fruit and Ornamental Plant Research
15, 117-124.
6. Durrant, W. E. & Dong, X. (2004). Systemic acquired resistance. Annu Rev Phytopathol 42,
185-209.
7. Gonsalves, D., Gonsalves, C., Ferreira, S., Pitz, K., Fitch, M., Manshardt, R., and Slightom, J.
(2004). Transgenic virus resistant papaya: From Hope to Reality for Controlling of Papaya ringspot virus in Hawaii. Online. APSnet feature, American Phytopathological Society.
8. Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E. & Mello, C. C. (1998).
Potent and specific genetic interference by double-strandedRNA in Caenorhabditis elegans.
Nature 391, 806-811.
9. Fuentes, A., Ramos, P. L., Fiallo, E., Callard, D., Sanchez, Y., Peral, R., Rodrıguez, R. &
Pujol, M. (2006). Intron–hairpin RNA derived from replication associated protein C1 gene
confers immunity to Tomato Yellow Leaf Curl Virus infection in transgenic tomato plants.
Transgenic Research 15, 291–304.
10. Hamilton, A. J. & Baulcombe, D. C. (1999). A species of small antisense RNA in
postranscriptional gene silencing in plants. Science 286, 950-952.
11. Hammerschmidt, R. (2009). Systemic Acquired Resistance. In Plant Innate Immunity, pp.
173-222.
12. Hayat, Q., Hayat, S., Irfan, M. & Ahmad, A. (2010). Effect of exogenous salicylic acid
under changing environment: A review. Environmental and Experimental Botany 68, 14-25.
13. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
14. Jones, J. D. G. & Dangl, J. L. (2006). The plant immune system. NATURE 444, 323-329.

98
15. Kang, B.-C., Yeam, I. & Jahn, M. M. (2005). GENETICS OF PLANT VIRUS
RESISTANCE. Annu Rev Phytopathol 43, 581–621.
16. Kiraly, L., Barnaz, B. & Kiralyz, Z. (2007). Plant resistance to pathogen infection: Forms
and mechanisms of innate and acquired resistance. J Phytopathol 155, 385-396.
17. Mandal, B., Mandal, S., Csinos, A. S., Martinez, N., Culbreath, A. K., & Pappu, H. R.
(2008). Biological and molecular analyses of the acibenzolar- S-methyl-induced systemic
acquired resistance in flue-cured tobacco against Tomato spotted wilt virus. Phytopathology
98,196-204.
18. Mysore, K. S. & Ryu, C. M. (2004). Nonhost resistance: how much do we know? Trends
Plant Sci 9, 97-104.
19. Ryals, J. A., Neuenschwander, U. H., Willits, M. G., Molina, A., Steiner, H. Y. & Hunt, M.
D. (1996). Systemic acquired resistance. Plant Cell 8, 1809-1819.
20. Smith, N. A., Singh, S. P., Wang, M.-B., Stoutjesdijk, P. A., Green, A. G. & Waterhouse, P.
M. (2000). Total silencing by intronspliced hairpin RNAs. Nature 407, 319-320.
21. Soosaar, J. L. M., Burch-Smith, T. M. & Dinesh-Kumar, S. P. (2005). MECHANISMS OF
PLANT RESISTANCE TO VIRUSES. NATURE REVIEWS 3, 789-798.
22. Takken, F. L. W. & Joosten, M. (2000). Plant resistance genes: their structure, function and
evolution. Eur J Plant Pathol 106, 699-713.
23. Vallad, G.E. & Goodman, R.M. (2004). Systemic Acquired Resistance and Induced
Systemic Resistance in Conventional Agriculture Crop Sci. 44,1920–1934
24. Vanderschuren, H., Stupak, M., Fütterer, J., Gruissem, W. & Zhang, P. (2007). Engineering
resistance to geminiviruses – review and perspectives. Plant Biotechnology Journal 5, 207–
220.
25. Vlot, A. C., Dempsey, D. A. & Klessig, D. F. (2009). Salicylic Acid, a Multifaceted
Hormone to Combat Disease. Annu Rev Phytopathol 47, 177-206.
26. Yin, H., Zhao, X. & Du., Y. (2010). Oligochitosan: A plant diseases vaccine—A review.
Carbohydrate Polymers 82, 1–8
27. Zhang, Y.-Y., Li, H.-X., Ouyang, B. & Ye, Z.-B. (2006). Regulation of Eukaryotic Initiation
Factor 4E and Its Isoform: Implications for Antiviral Strategy in Plants. Journal of
Integrative Plant Biology 48, 1129−1139.

99
Chƣơng 9. CHẨN ĐOÁN BỆNH VIRUS THỰC VẬT
9.1 TÓM TẮT NỘI DUNG
Nội dung chƣơng này tập trung vào kỹ thuật chẩn đoán virus dựa trên đặc trƣng phân tử, cả
mức protein và acid nucleic vủa virus.
9.2 GIỚI THIỆU
Chẩn đoán là xác định chính xác tác nhân gây bệnh. Nhiều phƣơng pháp đã đƣợc sử dụng để
chẩn đoán virus thực vật dựa vào:
1. Đặc trƣng sinh học: Triệu chứng, cây chỉ thị
2. Đặc trƣng hình thái: Kính hiển vi điện tử, thể vùi
3. Đặc trƣng phân tử (mức protein): Khuyếch tán trong gel, ELISA, dot blot miễn dịch
4. Đặc trƣng phân tử (mức acid nucleic): PCR, RT-PCR, dot blot (lai DNA), dsRNA, giải
trình tự
Hiện nay, kỹ thuật chẩn đoán bệnh virus phổ biến nhất, đơn giản và tƣơng đối rẻ là kỹ thuật
ELISA. Ngoài ra, cùng với sự phát triển của công nghệ sinh học thì các kỹ thuật liên quan đến
PCR, RT-PCR và giải trình tự cũng đang trở nên thông dụng.
9.3 CHẨN ĐOÁN DỰA VÀO TRIỆU CHỨNG
Hiển nhiên, chẩn đoán bằng triệu chứng phụ thuộc lớn vào kinh nghiệm của ngƣời nghiên
cứu.
Ƣu điểm: là kỹ thuật chẩn đoán nhanh chóng, rẻ nhất và trong nhiều trƣờng hợp khá chính
xác.
Nhƣợc điểm: nhìn chung, chẩn đoán dựa vào triệu chứng bệnh virus rất khó và trong nhiều
trƣờng hợp là không thể vì một số lý do sau:
Mức độ biến động triệu chứng quá lớn, phụ thuộc nhiều vào điều kiện ngoại cảnh và
trạng thái sinh lý của cây.
Một virus có thể tạo rất nhiều triệu chứng khác nhau trên cùng một cây, trên các giống
cây khác nhau.
Nhiều virus khác nhau, thậm chí thuộc các đơn vị phân loại khác nhau có thể tạo triệu
chứng giống nhau.
Trên một cây có thể nhiễm 2 hay nhiều virus khác nhau. Tƣơng tác của các virus có
thể rất phức tạp.
9.4 CHẨN ĐOÁN DỰA VÀO CÂY CHỈ THỊ
Cây chỉ thị là các cây tạo ra các triệu chứng khá đặc trƣng cho 1 virus. Đây là kỹ thuật chẩn
đoán đƣợc thực hiện nhiều trƣớc giai đoạn phát triển của công nghệ sinh học.

100
Nhiều cây chỉ thị thông dụng cho virus thực vật thuộc các chi Chenopodium, Nicotiana,
Cucumis, Phaseolus, Solanum, Vicia, Vigna và Brassica. Các cây chỉ thị thị có thể tạo vết bệnh
cục bộ hay nhiễm bệnh hệ thống
Một số cây chỉ thị nhƣ rau muối (Chenopodium amaranticolor, C. quinoa), thuốc lá cảnh (N.
benthamiana) phản ứng với rất nhiều virus khác nhau.
Ƣu điểm. Vì là một thử nghiệm sinh học nên phản ứng trên cây chỉ thị cho phép xác định
chính xác bản chất virus của tác nhân gây bệnh. Trong nhiều trƣờng hợp, sử dụng cây chỉ thị
thích hợp cho phép phân biệt đƣợc các chủng của virus.
Nhƣợc điểm.
Đắt (tốn công lao động, đòi hỏi phƣơng tiện chăm sóc nhƣ nhà lƣới, nhà kính chuẩn)
Nhiều trƣờng hợp không thể phân biệt đƣợc mức loài. Một số cây chỉ thị nhƣ thuốc lá phản
ứng với rất nhiều virus.
Tốn thời gian: để phản ứng có thể đánh giá đƣợc thƣờng mất nhiều ngày, nhiều tuần, thậm
chí nhiều tháng.
Hình 9-1 Lây nhiễm nhân tạo trên cây chỉ thị thuốc lá và rau muối (ảnh trái, giữa). Triệu chứng
đốm chết hoại do Citrus psorosis virus (CPsV) trên cây rau muối C. amaranticolor
(http://www.DPWeb.net)
9.5 KĨ THUẬT HIỂN VI ĐIỆN TỬ
Quan sát mẫu virus dƣới kính hiển vi điện tử EM (Electron microscope) là một kỹ thuật quan
trọng trong phát hiện và chẩn đoán virus. Có 2 loại kính hiển vi điện tử chính đƣợc sử dụng là
kính hiển vi xuyên TEM (Transmission EM) và kính hiển vi quét SEM (Scanning EM). Kính
hiển vi điện tử sử dụng sóng điện tử (có bƣớc sóng > sóng ánh sáng 100.000x) để khuyếch đại
hình ảnh nên có thể phóng đại hình ảnh tối đa 2.000.000x
Mẫu virus quan sát có thể từ phân tử virus đã tinh chiết hoặc trực tiếp trong mô (dùng kỹ
thuật lát cắt siêu mỏng)
Ƣu: + Phát hiện trực tiếp virus trong mô
+ Cho phép dự đoán tới chi
Nhƣợc: đắt, phức tạp

101
9.6 KĨ THUẬT CHẨN ĐOÁN DỰA VÀO THỂ VÙI
9.6.1 Loại thể vùi
Thể vùi (inclusion body) là sự kết tụ của:
Các phân tử virus (tất cả các nhóm virus),
Các protein vỏ virus (ví dụ tymovirus),
Các protein không phải protein vỏ virus (ví dụ tenuivirus, potyvirus),
Các cấu tử tế bào bị biến đổi do sự nhiễm bệnh (ví dụ chloroplasts của tế bào cây bị
nhiễm tymovirus).
9.6.2 Vị trí hình thành thể vùi trong tế bào
Trong tế bào chất (ví dụ thể vùi hình trụ của potyvirus),
Trong nhân (ví dụ thể vùi trong nhân của geminivirus),
Trong cả tế bào chất và nhân (ví dụ thể vùi tinh thể của potyvirus),
Trong không bào và tế bào chất (ví dụ thể vùi tinh thể của cucumovirus).
9.6.3 Ví dụ một số loại thể vùi
1. Thể vùi trong tế bào chất của potyvirus. Đƣợc cấu tạo từ các phân tử protein CI
(cytoplasm inclusion). phân tử protein xắp xếp thành các phiến protein. Các phiến này lại lắp ráp
thành các dạng nhƣ dạng vòng bánh xe, dạng cuốn, dạng phiến. Hình thái thể vùi đặc trƣng cho
các potyvirus và có giá trị chẩn đoán.
Hình: 9-2 Sơ đồ (trái) và ảnh chụp hiển vi điện tử (phải) của thể vùi tế bào chất của potyvirus
2. Thể vùi của TMV. Cấu tạo từ phân tử virus. Các phân tử virus hình gậy xếp thành hàng.
Các hàng lại chồng lớp lên nhau thành phiến tinh thể hình lục giác. Thể vùi của TMV hình thành
trong tế báo chất và không bào.
Ƣu điểm của chẩn đoán bằng thể vùi. Một trong các ƣu điểm của thể vùi là rất đơn giản và
có thể chẩn đoán bằng kính hiển vi quang học. Qui trình kiểm tra thể vùi virus có thể tóm tắt nhƣ
sau:
Tƣớc 1 mảnh biểu bì hoặc cắt một mảnh mô mỏng.

102
Làm trong mô bằng dung dịch Triton X-100 5 % trong khoảng 10-15 phút.
Nhuộm thể vùi bằng thuốc nhuộm (10-15 phút). Một số thuốc nhuộm phổ biến là (i)
Calcomine orange / 2-methoxyethanol, (ii) Brilliant green / 2- methoxyethanol và (iii)
Azure A / 2-methoxyethanol + 0.2 M Na2HPO4.
Xử lý mô bằng dung dịch rửa (70 % 2-methoxyethyl acetate + 30% ethanol).
Kiểm tra dƣới kính hiển vi quang học
Hình 9-3 Dụng cụ và vật liệu dùng để kiểm tra thể vùi virus thực vật bằng kính hiển vi quang học
(http://plantpath.ifas.ufl.edu/pdc/Inclusionpage/Florvirus.html)
9.7 KĨ THUẬT PCR VÀ RT-PCR
9.7.1 Giới thiệu và nguyên lý
PCR là một trong các phát minh quan trọng nhất của thế kỷ 20 trong sinh học phân tử. PCR là
một kỹ thuật đơn giản nhƣng đƣợc sử dụng trong hầu hết các nghiên cứu CNSH. Có vô số tài
liệu tiếng Việt và tiếng Anh về kỹ thuật PCR. Dƣới đây là tóm tắt kỹ thuật
Tùy thuộc khuôn là DNA hay RNA mà có tên phản ứng là PCR hay RT-PCR
Khuôn là DNA => PCR (polymerase chain reaction)
Khuôn là RNA => RT-PCR (Reverse transcription – PCR)
Các bƣớc cơ bản trong 1 phản ứng PCR gồm 3 bƣớc :
Biến tính. Hỗn hợp phản ứng đƣợc đặt ở nhiêt độ cao (92 – 94 OC) trong thời gian ngắn
(khoảng 20 – 60 giây). Ỏ nhiệt độ này, khuôn DNA dạng sợi kép tách thành sợi đơn.
Gắn mồi. Hỗn hợp phản ứng đƣợc đặt ở nhiêt độ gắn mồi (Ta) trong thời gian ngắn. Ta
đƣợc tính toán tùy thuôch đặc điểm mồi, thƣờng trong khoảng 40 – 60 OC). Ở nhiệt độ này, mồi
gắn vào khuôn sợi đơn ở vị trí đặc hiệu.

103
Tổng hợp sản phẩm PCR. Hỗn hợp phản ứng đƣợc tăng tới nhiệt độ tối ƣu cho phản ứng
tổng hợp chuỗi (72 hoặc 68 OC tùy DNA polymerase chịu nhiệt).
Trong trƣờng hợp RT-PCR, trƣớc khi thực hiện PCR, cần phải có thêm một bƣớc chuyển
khuôn RNA thành cDNA bằng enzyme phiên mã ngƣợc (reverse transcriptase). Hai bƣớc RT và
PCR có thể đƣợc thực hiện riêng rẽ hay trong cùng một tube phản ứng.
Các bƣớc trong chẩn đoán bệnh cây bằng PCR/RT-PCR :
Thiết kế mồi (primer)
Chiết acid nucleic tổng số (DNA hoặc RNA) từ mô cây; dùng các kỹ thuật chiết thông
thƣờng (rẻ) hoặc kit chiết thƣơng mại (đắt)
Chạy PCR hoặc RT-PCR
Kiểm tra và đánh giá kết quả bằng điện di
9.7.2 Thiết kế và lựa chọn mồi (primer)
Mồi là một chuỗi oligonucleotide sợi đơn ngắn (kích thƣớc thƣờng từ 18 – 24 nucleotides).
Có hai loại mồi đƣợc sử dụng cho chẩn đoán bệnh cây
Mồi chung (universal/degenerate primers): đƣợc thiết kế trên các vùng bảo thủ cao, có khả
năng phát hiện các đối tƣợng khác hẳn nhau (thƣờng các đối tƣợng của một chi, một họ).
Mồi đặc hiệu (specific primers): đƣợc thiết kế trên các vùng có thể phân biệt đƣợc loài, thậm
chí dƣới loài nhƣ chủng/nòi/type sinh học…
Có đƣợc các mồi tốt là bƣớc quan trọng nhất trong chẩn đoán bệnh cây. Có 2 cách để có thể
nhận đƣợc các mồi tốt.
Tham khảo và lựa chọn từ các nghiên cứu đã đƣợc công bố.
Tự thiết kế mồi.
9.7.3 Tự thiết kế mồi
Để tự thiết kế mồi dựa trên các chuỗi gen sẵn có trên Genbank, ngƣời ta cần sử dụng một
phần mềm có khả năng căn trình tự đa chuỗi (multiple sequence alignment), chẳng hạn
ClustalX, Bioedit. Ngay sau khi các chuỗi DNA đã đƣợc căn trình tự, chúng ta có thể thiết kế
các mồi chung hay mồi đặc hiệu tùy theo yêu cầu.
Các chú ý khi thiết kế mồi:
Độ dài mồi. Nên nằm trong khoảng 18-22 nucleotides. Độ dài này đủ dài để đảm bảo tính đặc
hiệu và đủ ngắn để mồi gắn đễ dàng vào khuôn.
Nhiệt độ tách chuỗi (Tm = Melting Temperature). Tm là nhiệt độ mà tại đó 1/2 số sợi DNA
kép tách thành sợi đơn. Tm của mồi nên nằm trong khoảng 52-58 OC.
Nhiệt độ gắn mồi (Ta = annealing temperature). Ta đƣợc tính dựa trên Tm. Ta quá cao làm
mồi khó gắn vào khuôn dẫn tới năng xuất PCR thấp. Ta quá thấp làm mồi dễ gắn không đặc hiệu
vào khuôn dẫn tới tạo các sản phẩm không đặc hiệu. Ta của 2 mồi xuôi và ngƣợc dòng nên
tƣơng đƣơng. Có nhiều cách tính Ta, dƣới đây là một công thức:
Ta = 0.3 x Tm(mồi) + 0.7 Tm (sản phẩm) – 14.9
Hàm lƣợng GC. Số các gốc G và C nên nămg trong khoảng 40-60%.

104
Chuỗi GC đầu 3’. Đầu tận cùng 3 ‗ nên là G hoặc C hoặc GC hoặc CG. Đầu 3‘ không nên
có nhiều hơn 3 gốc liên tục G/C.
Cấu trúc thứ cấp. Cấu trúc thứ cấp có thể hình thành trong cùng một mồi hoặc giữa 2 mồi.
Mồi chứa cấu trúc thứ cấp xấu thƣờng dẫn tới năng suất PCR bị giảm, thậm chí không có sản
phẩm. Tính ổn định của cấu trúc thứ cấp đƣợc đo bằng ΔG (là năng lƣợng cần để phá vỡ cấu
trúc thứ cấp, đƣợc tính từ các phần mềm thiết kế mồi). Các loại cấu trúc thứ cấp là:
Hairpins. Là cấu trúc thứ cấp hình thành trong nội bộ mồi. Giá trị ΔG của các hairpin đầu 3‘
không nên <-2 kcal/mol và của các hairpin ở giữa không nên <-3 kcal/mol.
Tự mồi (Self Dimer). Là cấu trúc hình thành giƣã các phân tử của cùng loại mồi. Giá trị ΔG
của các hairpin đầu 3‘ không nên <-5 kcal/mol và của các hairpin ở giữa không nên <-6
kcal/mol.
Tự mồi chéo (Cross Dimer). Là cấu trúc hình thành giƣã các phân tử của 2 mồi khác nhau
(cặp mồi trong phản ứng PCR). Giá trị ΔG của các hairpin đầu 3‘ không nên <-5 kcal/mol và
của các hairpin ở giữa không nên <-6 kcal/mol.
Các chuỗi lặp. Mồi không nên chứa nhiều chuỗi lặp kép (ví dụ TATATATA) hoặc chuỗi lặp
đơn (ví dụ TTTT) vì có thể gây ra hiện tƣợng bắt cặp nhầm (misprime). Trong một mồi, chỉ nên
có tối đa 4 chuỗi lặp. Các chuỗi lặp cũng không nên xuất hiện nhiều lần trong 1 mồi
Độ ổn định đầu 3 ‗. Tận cùng đầu 3‘ (khoảng 5 nucleotides) không nên chứa toàn gốc G
hoặc C vì có giá trị ΔG cao dễ dẫn tới bắt cặp không đặc hiệu vào khuôn. Trái lại nếu đầu 3‘
chứa quá nhiều gốc T hoặc A sẽ khó bắt cặp.
Vùng thiết kế mồi của khuôn. vùng thiết kế mồi của khuôn không nên chứa quá nhiều cấu
trúc thứ cấp. Nếu các cấu trúc thứ cấp này ổn định ở nhiệt độ gắn mồi thì mồi sẽ không thể bắt
cặp đƣợc.
9.7.4 Thiết kế mồi chung (degenerate primer)
Mồi chung là một mồi đƣợc thiết kế trên vùng bảo thủ. Nếu vùng bảo thủ là đồng nhất thì
mồi chung sẽ chỉ là một chuỗi mồi duy nhất. Nếu vùng bảo thủ chứa các vị trí sai khác thì mồi
chung thực chất là một hỗn hợp các mồi, mỗi mồi tƣơng ứng với một sai khác nucleotide.
Trong thiết kế mồi chung, ngƣời ta sử dụng các mã suy biến tại các vị trí sai khác, ví dụ R là
A hoặc G (số suy biến = 2); Y là C hoặc T (số suy biến bằng 2); N là A hoặc T hoặc G hoặc C
(số suy biến bằng 4). Số sai khác nucleotide tại mỗi vị trí của mồi đƣợc gọi là các số suy biến.
Tổng số mồi trong hỗn hợp sẽ bằng tích của tất cả các số suy biến tại mỗi vị trí. Để làm giảm số
lƣợng mồi trong hỗn hợp, ngƣời ta thƣờng sử dụng một nucleotide đặc biệt là inosine. Inosine có
thể ghép cặp với bất kỳ nucleotide nào trong số 4 nucleotide A, T, G, C.
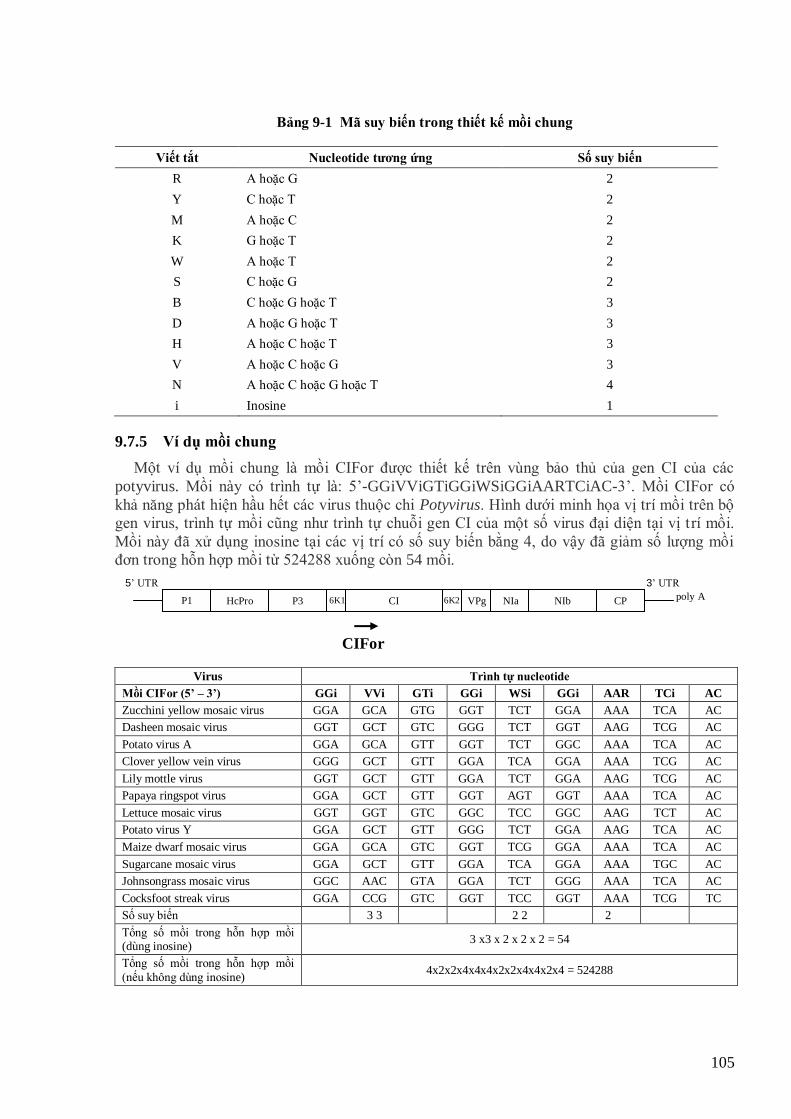
105
Bảng 9-1 Mã suy biến trong thiết kế mồi chung
Viết tắt Nucleotide tƣơng ứng Số suy biến
R A hoặc G 2
Y C hoặc T 2
M A hoặc C 2
K G hoặc T 2
W A hoặc T 2
S C hoặc G 2
B C hoặc G hoặc T 3
D A hoặc G hoặc T 3
H A hoặc C hoặc T 3
V A hoặc C hoặc G 3
N A hoặc C hoặc G hoặc T 4
i Inosine 1
9.7.5 Ví dụ mồi chung
Một ví dụ mồi chung là mồi CIFor đƣợc thiết kế trên vùng bảo thủ của gen CI của các
potyvirus. Mồi này có trình tự là: 5‘-GGiVViGTiGGiWSiGGiAARTCiAC-3‘. Mồi CIFor có
khả năng phát hiện hầu hết các virus thuộc chi Potyvirus. Hình dƣới minh họa vị trí mồi trên bộ
gen virus, trình tự mồi cũng nhƣ trình tự chuỗi gen CI của một số virus đại diện tại vị trí mồi.
Mồi này đã xử dụng inosine tại các vị trí có số suy biến bằng 4, do vậy đã giảm số lƣợng mồi
đơn trong hỗn hợp mồi từ 524288 xuống còn 54 mồi.
Virus Trình tự nucleotide
Mồi CIFor (5’ – 3’) GGi VVi GTi GGi WSi GGi AAR TCi AC
Zucchini yellow mosaic virus GGA GCA GTG GGT TCT GGA AAA TCA AC
Dasheen mosaic virus GGT GCT GTC GGG TCT GGT AAG TCG AC
Potato virus A GGA GCA GTT GGT TCT GGC AAA TCA AC
Clover yellow vein virus GGG GCT GTT GGA TCA GGA AAA TCG AC
Lily mottle virus GGT GCT GTT GGA TCT GGA AAG TCG AC
Papaya ringspot virus GGA GCT GTT GGT AGT GGT AAA TCA AC
Lettuce mosaic virus GGT GGT GTC GGC TCC GGC AAG TCT AC
Potato virus Y GGA GCT GTT GGG TCT GGA AAG TCA AC
Maize dwarf mosaic virus GGA GCA GTC GGT TCG GGA AAA TCA AC
Sugarcane mosaic virus GGA GCT GTT GGA TCA GGA AAA TGC AC
Johnsongrass mosaic virus GGC AAC GTA GGA TCT GGG AAA TCA AC
Cocksfoot streak virus GGA CCG GTC GGT TCC GGT AAA TCG TC
Số suy biến 3 3 2 2 2
Tổng số mồi trong hỗn hợp mồi (dùng inosine)
3 x3 x 2 x 2 x 2 = 54
Tổng số mồi trong hỗn hợp mồi
(nếu không dùng inosine) 4x2x2x4x4x4x2x2x4x4x2x4 = 524288
HcPro VPg 6K1 P1 P3 CI NIb NIa CP 6K2
3‘ UTR 5‘ UTR
poly A
CIFor

106
9.8 CHẨN ĐOÁN DỰA VÀO RNA SỢI KÉP
Nhiều virus thực vật có bộ gen RNA. Các virus RNA sợi kép thƣờng có bộ gen phân đoạn,
trong đó số lƣợng và kích thƣớc các phân đoạn có thể có giá trị chẩn đoán vì đặc trƣng cho chi
(ví dụ nhƣ các reovirus). Các virus có bộ gen RNA sợi đơn cũng có thể chẩn đoán bằng kỹ thuật
này vì chúng hình thành các phân tử dsRNA là dạng trung gian trong quá trình tái bản. Ngoài ra,
nhiều virus hình thành các phân tử phụ gen nome (subgenome) và các phân tử này cũng tạo các
phân tử dsRNA có kích thƣớc đặc trƣng.
Về mặt kỹ thuật, chẩn đoán dsRNA của mẫu virus đƣợcthực hiện qua các bƣớc sau:
Chiết RNA hoặc acid nucleic tổng số từ mô cây
Tinh chiết dsRNA bằng cách cho dịch RNA hoặc acid nucleic tổng số vào cột chứa bột
cellulose (loại CF11, Whatman) với điều kiện nồng độ ethanol = 16.5%. Ở nồng độ
ethanol này, chỉ các phân tử dsRNA liên kết vào hạt cellulose. Tiếp theo, các loại acid
nucleic khác (ssRNA, DNA) bị loại bỏ khỏ cột bằng rửa với đệm chứa ethanol 16.5 %.
Cuối cùng, dsRNA đƣợc giải hấp thụ bằng đệm không chứa ethanol.
Phân tích kết quả bằng điện di polyacrylamid hoặc agarose
Kỹ thuật này phổ biến trong những năm 1990 nhằm chẩn đoán nhiều virus, đặc biệt các
virus trên cây có múi nhƣ CTV. Hiện nay, do kỹ thuật quá tốn công sức nên hầu nhƣ không
đƣợc sử dụng, ngoại trừ để phát hiện và xác định các virus mới.
Hình 9-4 Phát hiện bộ gen dsRNA của virus lùn sọc đen phƣơng Nam (Zhou et al., 2008)
9.9 CHẨN ĐOÁN DỰA VÀO HUYẾT THANH HỌC
9.9.1 Các khái niệm
Cơ sở của phƣơng pháp huyết thanh học là dựa vào sự tƣơng tác đặc hiệu giữa kháng
nguyên và kháng thể đặc hiệu với kháng nguyên đó
a. Kháng nguyên (antigen)
Kháng nguyên thƣờng là các đại phân tử chứa protein hoặc polysaccharide. Kháng nguyên
phải có hai đặc tính quan trọng: (1) hoạt tính sinh miễn dịch tức khả năng kích thích động vật
máu nóng hình thành kháng thể đặc hiệu và (2) tính đặc hiệu tức khả năng liên kết với kháng
thể đặc hiệu tƣơng ứng. Một số chất có trọng lƣợng phân tử thấp nhƣ các amino acid, một số loại
polysaccharide... không có hoạt tính sinh miễn dich nhƣng lại có khả năng liên kết với kháng thể

107
đặc hiệu với kháng nguyên có chứa các chất này. Các chất đó đƣợc gọi là bán kháng nguyên
(hapten).
b. Nhóm quyết định kháng nguyên (epitope)
Trên bề mặt kháng nguyên có các vùng đặc biệt có chức năng cảm ứng hình thành kháng
thể và là vị trí liên kết kháng thể. Các vùng này đƣợc gọi là các nhóm quyết định kháng nguyên
(epitope).
Một nhóm quyết định kháng nguyên chỉ kích thích hệ miễn dịch của động vật máu nóng
hình thành một dòng phân tử kháng thể. Kháng nguyên chứa một nhóm quyết định kháng
nguyên gọi là kháng nguyên đơn giá, chứa nhiều nhóm quyết định kháng nguyên gọi là kháng
nguyên đa giá. Các epitope có thể đƣợc chia thành:
Epitope trình tự: chỉ phụ thuộc trình tự amino acid
Epitope hình thái: phụ thuộc vào cả cấu trúc không gian
Epitope liên tục: là một chuỗi amino acid liên tục
Epitope không liên tục: là các amino acid không liền nhau (trên chuỗi cấp 1) nhƣng do sự
gấp của protein nên xắp xếp gần nhau dẫn tới cùng nhau tạo thành 1 epitope
c. Kháng nguyên của virus
Đối với virus, nhìn chung bất kỳ một protein nào của virus cũng có thể đƣợc sử dụng làm
kháng nguyên. Tuy nhiên, loại kháng nguyên phổ biến nhất là protein vỏ virus. Kháng nguyên
vỏ protein virus có thể đƣợc tạo ra bằng: (i) tinh chiết phân tử virus từ cây nhiễm bệnh hoặc (ii)
biểu hiện vỏ protein trong tế bào vi khuẩn dùng công nghệ sinh học.
d. Kháng thể (antibody)
Sự hình thành kháng thể: Hệ thống miễn dịch của cơ thể động vật máu nóng bao gồm các
cơ quan có thẩm quyền miễn dịch và các tế bào có thẩm quyền miễn dịch. Các cơ quan có thẩm
quyền miễn dịch (tuyến ức, lách, tủy...) là nơi sản sinh, duy trì, biệt hoá và điều khiển sự hoạt
động của các tế bào có thẩm quyền miễn dịch. Các tế bào có thẩm quyền miễn dịch (chủ yếu
gồm 2 loại là lympho bào B và lympho bào C) tham gia vào quá trình đáp ứng miễn dịch của cơ
thể nhằm chống lại các tác nhân lạ xâm nhập. Có 2 kiểu đáp ứng miễn dịch:
Kiểu đáp ứng miễn dịch dịch thể: Khi kháng nguyên xâm nhập vào cơ thể, bị đại thực bào
vây bắt, xử lý và trình diện cho lympho bào B. Lympho bào B đƣợc biệt hoá thành tế bào
plasma sản xuất ra kháng thể dịch thể. Kháng thể dịch thể tồn tại trong dịch cơ thể (huyết
tƣơng) và sẽ kết hợp đặc hiệu với kháng nguyên.
Kiểu đáp ứng miễn dịch tế bào: Khi kháng nguyên xâm nhập vào cơ thể, bị đại thực bào vây
bắt, xử lý và trình diện cho lympho bào T. Lympho bào T đƣợc biệt hoá thành lympho bào T
mẫn cảm kháng nguyên và chính bản thân chúng là kháng thể tế bào. Kháng thể tế bào tồn
tại trên bề mặt tế bào và cũng kết hợp đặc hiệu với kháng nguyên.
Nhƣ vậy, theo nghĩa rộng, kháng thể là bất kỳ phân tử nào có khả năng liên kết đặc
hiệu với kháng nguyên. Tuy nhiên, kháng thể sử dụng trong chẩn đoán bằng kỹ thuật huyết
thanh học chính là các kháng thể dịch thể
Kháng thể dịch thể là các globulin miễn dịch (Immunoglobulin, Ig). Có 5 lớp Ig là
IgG, IgM, IgA, IgD và IgE nhƣng trong chẩn đoán huyết thanh học, thƣờng chỉ sử dụng IgG.

108
Cấu tạo của IgG (xem hình vẽ): gồm 4 phân tử polypeptid (2 chuỗi nặng, 2 chuỗi nhẹ)
liên kết với nhau bằng các cầu nối disulfide. Phân tử IgG có 3 vùng chức năng chính (có thể
tách đƣợc bằng một protease là papain)
Hai mảnh Fab (antigen binding fragment) giống nhau về cấu trúc và mang tính chất
kháng thể do chứa vị trí liên kết với kháng nguyên. Các mảnh Fab, khi đƣợc tách ra bằng
papain, sẽ vẫn giữ nguyên hoạt tính kháng thể nhƣng không thể tạo ra phản ứng kết tủa
hoặc phản ứng ngƣng kết.
Một mảnh Fc (cristallizable fragment) không có hoạt tính kháng thể nhƣng chứa nhóm
quyết định kháng nguyên của phân tử IgG do vậy khi tiêm phân tử IgG nguyên vẹn hoặc
chỉ cần mảnh Fc từ 1 loài (chẳng hạn thỏ) sang 1 loài khác (chẳng hạn dê) thì loài thứ 2
sẽ vẫn tạo ra kháng thể đặc hiệu kháng thể của loài thứ nhất (anti-antibody). Đây là cơ sở
cho các kiểu ELISA gián tiếp.
Hình 9-5 Cấu trúc kháng thể (Hull, 2002)
e. Kháng thể đơn dòng và kháng thể đa dòng
Kháng thể đơn dòng (Monoclonal antibody, MAb)
Là kháng thể đƣợc hình thành chỉ từ một dòng lympho bào B
Nhận biết và liên kết với chỉ 1 epitope
Sản xuất phức tạp
Kháng thể đa dòng (Polyclonal antibody, PAb)
Là một hỗn hợp của các kháng thể đơn dòng
Sản xuất đơn giản. Việc sản xuất bao gồm tiêm kháng nguyên (phân tử virus, protein
virus, protein vi khuẩn, nấm…) vào động vật máu nóng (nhƣ thỏ, dê, chuột…) và thu
kháng huyết thanh. Kháng huyết thanh sẽ chứa một hỗn hợp nhiều dòng kháng thể, mỗi
dòng đặc hiệu cho 1 epitope.

109
9.9.2 Sản xuất kháng thể đơn dòng
Các bƣớc chính để sản xuất kháng thể đơn dòng gồm
1. Gây miễn dịch trên chuột. Bƣớc này giống nhƣ sản xuất kháng thể đa dòng.
2. Dung hợp (lai) tế bào myeloma chuột và tế bào lá lách chuột đã gây miễn dịch.
Nuôi cấy một dòng tế bào myeloma chuột. Tế bào myeloma là một loại tế bào bạch cầu
bị ung thƣ nên có 2 đặc điểm: (1) không thể sản xuất kháng thể và (2) có thể sinh trƣởng
vô hạn định. Ngoài ra, vì chúng bị mất chức năng của gen HGPRT (hypoxantine guanine
phosphoribosyl transferase) nên không thể sống đƣợc trên môi trƣờng HTA
(Hypoxanthine Aminopterin Thymidine) vì aminopterin trong môi trƣờng ức chế một
đƣờng hƣớng sinh hóa liên quan đến biểu hiện gen HGPRT.
Tinh chiết tế bào lá lách của chuột đã gây miễn dịch. Dịch tế bào lá lách sẽ chứa lympho
bào B sản xuất kháng thể. Loại tế bào miễn dịch này có thời gian sống ngắn và không thể
sống đƣợc trên môi trƣờng nuôi cấy nhân tạo.
Dung hợp các tế bào lá lách với các tế bào myeloma nhờ PEG (polyethylene glycol) để
tạo ra các tế bào lai (hybridoma). Tế bào lai kết hợp đặc điểm của lympho bào B (tạo
kháng thể) và đặc điểm của tế bào myeloma (sinh trƣởng vô hạn định trên môi trƣờng
nhân tạo).
Nuôi cấy tế bào lai trong môi trƣờng chọn lọc. Môi trƣờng chọn lọc chứa HTA
(Hypoxanthine Aminopterin Thymidine) không cho phép sự sinh trƣởng của các tế bào
không lai.
3. Kiểm tra sự có mặt của Mabs trong dịch nuôi cấy của từng dòng tế bào lai bằng ELISA
4. Nuôi cấy phục hồi từng dòng tế bào lai có kháng thể mong muốn (ELISA +)
5. Tạo kháng thể từ các dòng tế bào lai có kháng thể mong muốn bằng invitro (trên môi
trƣờng nhân tạo hoặc in vivo (trong cơ thể chuột).
Hình 9-6 Sơ đồ sản xuất kháng thể đơn dòng

110
9.9.3 Kỹ thuật chẩn đoán Ouchterlony
Kỹ thuật Ouchterlony đƣợc còn đƣợc gọi là kỹ thuật ―Khuếch tán miễn dịch kép trong gel‖.
Đây là một trong các kỹ thuật huyết thanh học đƣợc sử dụng phổ biến trƣớc khi áp dụng kỹ thuật
ELISA. Nguyên lý của kỹ thuật rất đơn giản: ngƣời ta chế tạo các đĩa gel với các giếng gel. Ở
giếng giữa là kháng huyết thanh chứa kháng thể virus. Các giếng xung quanh sẽ nhỏ các mẫu
cần kiểm tra. Virus và IgG sẽ khuếch tán ra xung quanh giếng. Nếu kháng thể và virus là đồng
dạng của nhau thì chúng sẽ kết hợp tạo thành một vạch mờ (Hình)
Hình 9-7 Kỹ thuật Outerlony chẩn đoán virus. Các mẫu ABCDE là các mẫu dƣơng
9.9.4 Kỹ thuật chẩn đoán bằng ELISA
ELISA (Enzyme Linked Immunosorbent Assay) là một kỹ thuật chẩn đoán quan trọng và
phổ biến nhất hiện nay so với các kỹ thuật huyết thanh học khác. Trong bệnh cây, ELISA đƣợc
sử dụng chủ yếu để chẩn đoán virus và một số vi khuẩn. Một trong số các hãng nổi tiếng có bán
các kít ELISA chẩn đoán bệnh cây là Agdia (http://www.agdia.com/).
9.9.4.1 Vật liệu chính cho ELISA
Bản ELISA: bằng nhựa (polystyrene), 96 giếng, liên kết không đặc hiệu các loại protein
Các loại dung dich đệm: Đệm nghiền mẫu, đệm pha kháng thể, đệm rửa, đệm pha chất nền
(cơ chất).
Kháng thể các loại
Chất nền: nhiều loại, tùy thuộc enzyme, phổ biến nhất là NPP (nitrophenyl phosphate)
Enzyme: nhiều loại, cần cơ chất tƣơng ứng, phổ biến nhất là alkaline phosphatase (cơ chất
tƣơng ứng là NPP)
Máy đọc ELISA: là một loại thiết bị so màu vi phiến nhằm lƣợng hóa màu phản ứng.
Dụng cụ nghiền, chiết mẫu cây: chày, cối sứ…
9.9.4.2 Các kỹ thuật ELISA
Có rất nhiều kỹ thuật ELISA khác nhau. Dựa vào quan hệ của kháng thể liên kết enzyme
với kháng nguyên, ELISA có thể đƣợc chia làm 2 nhóm (xem hình) là:
Nhóm trực tiếp (direct ELISA): Kháng thể liên kết enzyme là kháng thể phát hiện kháng
nguyên (ví dụ các kỹ thuật I, VIII và IX trong sơ đồ).

111
Nhóm gián tiếp (indirect ELISA): Kháng thể liên kết enzyme không phải là kháng thể phát
hiện kháng nguyên (ví dụ các kỹ thuật II, III, IV, V, VI, VI trong sơ đồ).
ELISA cũng gồm rất nhiều kiểu khác nhau. Hai ví dụ đang đƣợc sử dụng là:
Kẹp kép kháng thể (double antibody sandwich = DAS) (tất cả các kỹ thuật ngoài trừ kỹ
thuật II trong sơ đồ)
Bẫy kháng nguyên trƣớc (plate-trapped antigen = PTA) nhƣ kỹ thuật II trong sơ đồ.
Hình 9-8 Các kỹ thuật ELISA khác nhau
9.9.4.3 Qui trình thử 2 kiểu phản ứng ELISA đại diện cho mỗi nhóm
a. Kỹ thuật ELISA trực tiếp kiểu kẹp kép kháng thể (DAS-ELISA)
Cố định IgG đặc hiệu virus vào bản ELISA: IgG đƣợc hoà trong dung dịch đệm phủ
(thƣờng là đệm carbonate) và đƣợc cho vào giếng với lƣợng 100 l/giếng. Bản ELISA đƣợc
ủ trong hộp ẩm ở 37 OC khoảng 2-4 giờ. Bản ELISA, thƣờng cấu tạo bằng polystirene có khả
năng liên kết không đặc hiệu với protein do vậy sẽ liên kết với các phân tử IgG (là protein
miễn dịch). Sau khi ủ, giếng đƣợc rửa bằng đêm rửa và nƣớc.
Cố định dịch cây vào bản ELISA: Nghiền mẫu cây đệm chiêt mẫu thành dịch cây. Dịch cây
đƣợc cho vào giếng với lƣợng100l/giếng. Bản ELISA đƣợc ủ ở 37 OC khoảng 2-4 giờ hoặc
qua đêm ở 4-6 OC. Nếu trong dịch cây có chứa virus thì virus sẽ liên kết đặc hiệu với IgG có
sẵn trong giếng còn protein của cây hoặc các virus khác sẽ không liên kết. Sau khi ủ, bản
ELISA đƣợc rửa nhƣ bƣớc 1.
Cố định IgG liên kết enzyme: Hoà IgG liên kết enzyme (IgG-E) trong đệm liên kết và cho
vào giếng với lƣợng 100 l/giếng. IgG này giống nhƣ IgG của bƣớc 1 nhƣng chỉ khác là nó
đƣợc liên kết từ trƣớc với 1 enzyme là Alkaline Phosphatase (AP). Bản ELISA đƣợc ủ và
rửa nhƣ bƣớc 1.
Cố định chất nền và đánh giá kết quả: Hoà chất nền trong đệm chất nền theo tỷ lệ 0.6
mg/ml) và cho giếng với lƣợng100 l/giếng. Chất nền ở đây là Nitrophenyl phosphate
(NPP). Bản ELISA đƣợc ủ ở nhiệt độ phòng trong hộp ẩm trong tối 60 phút. Phản ứng hoá
học ở đây là NPP sẽ bị thuỷ phân thành Nitrophenol phosphate dƣới sự xúc tác của enzyme
AP. Nitrophenol phosphate là chất có màu vàng, do vậy có thể nhận biết đƣợc bằng mắt hoặc
bằng máy đọc ELISA (máy so màu). Hiển nhiên, do nồng độ NPP đều giống nhau ở các
giếng nên giếng nào có màu vàng càng đậm chứng tỏ nồng độ enzyme càng cao tức nồng độ
virus trong mẫu thử càng cao. Có thể ngừng phản ứng thuỷ phân bằng cách bổ sung dung
dịch NaOH 3M với lƣợng 25-50 l/giếng.

112
b. Phản ứng ELISA gián tiếp kiểu bẫy kháng nguyên trước (PTA-ELISA)
1. Cố định dịch cây vào bản ELISA: Nghiền mẫu cây trong đệm phủ và cho 100l/giếng. ủ
bản ELISA qua đêm ở 4-6OC trong hộp ẩm. Sáng hôm sau, rửa 3 lần bằng đệm rửa. Protein
của cây và virus trong dịch cây (nếu có) sẽ liên kết không đặc hiệu vào thành giếng.
2. Cố định IgG thỏ đặc hiệu virus vào bản ELISA: Hoà IgG đặc hiệu virus trong đệm huyết
thanh và cho 100l/giếng. Ủ bản ELISA ở 37 OC 1-2 giờ trong hộp ẩm. Rửa bản ELISA nhƣ
bƣớc 1.
3. Cố định IgG đặc hiệu IgG thỏ liên kết AP vào bản ELISA: Hoà IgG đặc hiệu IgG thỏ liên
kết AP trong đệm liên kết và cho 100l/giếng. Ủ bản ở 37 OC 1-2 giờ. IgG liên kết AP thứ 2
này là IgG đƣợc tạo ra khi tiêm IgG của thỏ vào 1 loài động vật khác thỏ (chẳng hạn dê) nên
không liên kết với virus mà liên kết với IgG thỏ đặc hiệu virus ở bƣớc 2. Sau khi ủ, rửa bản
nhƣ bƣớc 1.
4. Cố định chất nền vào bản ELISA: Thực hiện tƣơng tự nhƣ ở DAS-ELISA.
9.10 CÁC KĨ THUẬT DOT BLOT
Nhiều kiểu dot blot đã đƣợc sử dụng để phát hiện virus. Các kỹ thuật này thực chất là các
dạng đơn giản hóa của western blot (lai protein) hoặc southern blot (lai DNA). Nhƣ vậy dựa vào
bản chất lai hóa, dot blot có thể gồm 2 nhóm:
Dựa trên western blot: Dịch cây chứa virus hoặc protein đƣợc cố định luôn lên màng mà
không cần tách bằng điện di polyacrylamid. Các bƣớc phát hiện tiếp theo về cơ bản giống
ELISA hoặc western blot.
Dựa trên southern blot: Dịch chiết DNA, sản phẩm cDNA, PCR/ RT-PCR đƣợc cố định lên
màng mà không cần tách trƣớc bằng điện di agarose. Các bƣớc phát hiện tiếp theo về cơ bản
giống nhƣ southern blot.
Hình 9-9 Phát hiện virus bằng kỹ thuạt dot blot miễn dịch (western blot, trái, Hull, 2002) và dot
blot lai DNA (southern blot, phải, Hsu et al., 2005)

113
9.11 CÂU HỎI ÔN TẬP CHƢƠNG 9
1. Các cơ sở chẩn đoán bệnh virus
2. Chẩn đoán dựa vào triệu chứng (ƣu, nhƣợc điểm.
3. Chẩn đoán dựa vào huyết thanh học (đặc điểm kháng nguyên, kháng thể, epitope, phân biệt
kháng thể đơn dòng, đa dòng, đặc điểm bản ELISA, enzyme sử dụng trong ELISA, phân biệt
ELISA trực tiếp và gián tiếp, trình tự DAS-ELISA và PTA-ELLSA).
4. Chẩn đoán bằng PCR/RT-PCR (đặc điểm mồi chung, so sánh mồi chung mồi đặc hiệu, đặc
điểm một mồi tốt).
9.12 TÀI LIỆU THAM KHẢO CHÍNH
1. Clark, M. F. & Adams, A. N. (1977). Characteristics of the microplate method of enzyme-
linked immunosorbent assay for the detection of plant viruses. Journal of General Virology
34, 475–483.
2. Ha, C. V., Revill, P. A., Harding, R. M., Vu, M. T. & Dale, J. L. (2008). Design and
application of two novel degenerate primer pairs for the detection and complete genomic
characterization of potyviruses. Archives of Virology 153, 25–36.
3. Hsu, Y. C., Yeh, T. J. & Chang, Y. C. (2005). A new combination of RT-PCR and reverse
dot blot hybridization for rapid detection and identification of potyviruses. Journal of
Virological Methods 128, 54–60.
4. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
5. Koenig, R. & Paul, H. L. (1982). Variants of ELISA in Plant Virus Diagnosis. Journal of
Virological Methods 5, 113-125.
6. Martin, R. R., James, D. & Levesque, C. A. (2000). Impacts of molecular diagnostic
technologies on plant disease management. Annual Review of Phytopathology. 38, 207–239
38, 207–239.
7. Mathews, R. E. F. (1993). Diagnosis of Plant Virus Diseases: CRC Press.
8. Mowat, W. P. & Dawson, S. (1987). Detection and identification of plant viruses by ELISA
using crude sap extracts and unfractionated antisera. Journal of Virological Methods 15 233-
247.

114
PHẦN II: CHUYÊN KHOA

115
Chƣơng 10. CÁC VIRUS THỰC VẬT Ở VIỆT NAM
10.1 TÓM TẮT NỘI DUNG
Chƣơng này tóm tắt lịch sử phát hiện virus thực vật ở Việt Nam và cung cấp danh sách các
virus đƣợc phát hiện cho tới nay (2010). Chƣơng này chỉ cung cấp thông tin mang tính chất
tham khảo cho ngƣời đọc.
10.2 CÁC VIRUS ĐƢỢC PHÁT HIỆN Ở VIỆT NAM
Bệnh virus đƣợc ghi nhận sớm nhất tại Việt Nam là bệnh chùn ngọn chuối, đƣợc phát hiện ở
Tây Nguyên trong những năm 60 (Vakili, 1969). Hiện nay, bệnh này đã đƣợc biết là do Banana
bunchy top virus (BBTV) gây ra.
Trong những năm 70, ở miền Bắc, đặc biệt ở các tỉnh trung du và miền núi, đã xuất hiện một
bệnh trên lúa gọi là bệnh vàng lụi với triệu chứng điển hình là cây lúa bị biến vàng, lùn, có thể
chết. Mặc dù tác nhân gây bệnh không thể đƣợc xác định tại thời điểm đó nhƣng bệnh đã đƣợc
cho là do virus gây ra vì bệnh có thể truyền qua rầy xanh đuôi đen (Nephotettix virescens) (Hà
Minh Trung, 1984).
Trong những năm 80, hai virus là Potato virus X (PVX) và Potato virus Y (PVY) nhiễm trên
khoai tây đã đƣợc xác đinh ở miền Bắc bằng kính hiển vi điện tử và ELISA (Vũ Triệu Mân,
1984). Trong cùng thời gian, các bệnh giống nhƣ bị nhiễm geminivirus (xoăn vàng lá/ngọn)
cũng đƣợc qua sát thấy trên một số cây trồng nhƣ khoai tây, thuốc lá và đặc biệt là cà chua. Các
thí nghiệm lan truyền cho thấy tác nhân gây bệnh truyền qua bọ phấn (Bemisia tabaci) và qua
ghép (Nguyễn Thơ, 1984).
Từ khoảng nửa cuối những năm 90 trở lại đây, do tăng cƣờng hợp tác nghiên cứu với các
phòng thí nghiệm virus trên thế giới, sự phát triển của công nghệ sinh học cũng nhƣ điều kiện
nghiên cứu đƣợc tăng cƣờng, nhiều virus thực vật đã đƣợc phát hiện và xác định ở Việt Nam.
Cho tới nay (2010), 52 virus thực vật thuộc 52 loài đã đƣợc xác định ở Việt Nam (bảng 10-1),
phần lớn đã đƣợc giải trình tự. Trong số này, đa số là các virus thuộc 2 chi Begomovirus (22
virus) và Potyvirus (18 virus). Đây cũng là 2 chi virus thực vật lớn nhất với mỗi chi chiếm
khoảng 20 % tổng số virus thực vật toàn thế giới. Ngoài ra, nhiều virus có ý nghĩa kinh tế, chẳng
hạn các virus gây bệnh trên lúa nhƣ bệnh lùn xoắn lá (Rice ragged stunt virus, RRSV), bệnh lúa
cỏ hay còn gọi là bệnh vàng lùn (Rice grassy stunt virus), bệnh lúa lùn sọc đen (southern rice
black streaked dwarf virus, SRBSDV) cũng mới đƣợc xác định chính xác.
Cần phải thấy rằng, số virus đƣợc phát hiện này chiếm một tỷ lệ nhỏ so với khoảng hơn 1000
virus thực vật đƣợc ghi nhận trên thế giới hiện nay. Trong khi đó, Việt Nam đã đƣợc chứng
minh là thuộc một trong các trung tâm đa dạng của nhiều virus, nhóm virus thực vật, chẳng hạn
nhƣ các begomovirus, một số potyvirus.
Do chẩn đoán bệnh virus dựa vào triệu chứng thƣờng khó, nhiều khi không thể nên trong
bảng 1, chỉ 52 virus đƣợc liệt kê do danh tính của chúng đã đƣợc xác định rõ với nguồn tham
khảo tin cậy. Chắc chắn rằng còn nhiều virus thực vật nữa hiện đang có mặt ở Việt Nam và chờ
đƣợc khám phá.

116
Bảng 10.1. Thành phần các virus thực vật đƣợc phát hiện tại Việt Nam tới 2010
TT Virus Viết tắt Chi Họ Ký chủ Tham khảo/mức
xác định*
1 Banana bunchy top virus BBTV Babuvirus Nanoviridae Chuối Bell et al., 2002;
Karan et al., 1994
2 Piper yellow mottle virus PYMoV Badnavirus Caulimoviridae Hồ tiêu, lá
nốt Giải trình tƣ?
3 Banana streak virus strain acuminata
Vietnam BSAcVNV Badnavirus Caulimoviridae Chuối
Lheureux et al.,
2007
4 Luffa yellow mosaic virus LYMV Begomovirus Geminiviridae Mƣớp Revil et al., 2003
5 Squash leaf curl China virus SLCCNV Begomovirus Geminiviridae Bí ngô, bầu
bí Revil et al., 2004
6 Corchorus yellow vein virus CoYVV Begomovirus Geminiviridae Rau đay Ha et al., 2006
7 Corchorus golden mosaic virus CoGMV Begomovirus Geminiviridae Rau đay Ha et al., 2008a
8 Kudzu mosaic virus KuMV Begomovirus Geminiviridae Sắn dây, đậu
tƣơng? Ha et al., 2008a
9 Clerodendrum golden mosaic virus ClGMV Begomovirus Geminiviridae Mò Ha et al., 2008a
10 Spilanthes yellow vein virus SpYVV Begomovirus Geminiviridae Cúc nút áo Ha et al., 2008a
11 Mimosa yellow leaf curl virus MiYLCV Begomovirus Geminiviridae Mimosa Ha et al., 2008a
12 Sida yellow vein Vietnam virus SiYVVNV Begomovirus Geminiviridae Ké hoa vàng Ha et al., 2008a
13 Erectites yellow mosaic virus ErYMV Begomovirus Geminiviridae Rau tàu bay Ha et al., 2008a
14 Ludwigia yellow vein Vietnam virus LuYVVNV Begomovirus Geminiviridae Rau mƣơng Ha et al., 2008a
15 Ludwigia yellow vein virus LuYVV Begomovirus Geminiviridae Rau mƣơng Ha et al., 2008a
16 Papaya leaf curl China virus PaLCuCNV Begomovirus Geminiviridae Thuốc lá, cà
chua Ha et al., 2008a
17 Lindernia anagallis yellow vein virus LaYVV Begomovirus Geminiviridae Lữ đằng Ha et al., 2008a
18 Alternanthera yellow vein virus AlYVV Begomovirus Geminiviridae Cúc (Zinha),
nhọ nồi Ha et al., 2008a
19 Tomato yellow leaf curl Kanchanaburi TYLCKaV Begomovirus Geminiviridae Cà chua, cà Ha et al., 2008a;
117
virus pháo DQ169054
20 Tomato leaf curl Vietnam virus ToLCVV Begomovirus Geminiviridae Cà chua Ha et al., 2008a;
Green et al., 2001
21 Tomato yellow leaf curl Vietnam virus TYLCVNV Begomovirus Geminiviridae Cà chua Ha et al., 2008a
22 Tomato leaf curl Dan Xa virus ToLCDXV Begomovirus Geminiviridae Cà chua Badwil et al., 2008
23 Tomato leaf curl Hainan virus ToLCHV Begomovirus Geminiviridae Cà chua, đu
đủ Giải trình tƣ
24 Tomato leaf curl Hanoi virus ToLCHanV Begomovirus Geminiviridae Cà chua Giải trình tƣ
25 Sida leaf curl virus SiLCV Begomovirus Geminiviridae Cối xay Ha et al., 2008a
26 Citrus tristeza virus CTV Closterovirus Closteroviridae Cây có múi Kiritani & Su, 1999
27 Cucumber mosaic virus CMV Cucumovirus Bromoviridae Bầu bí, cà
chua Revil et al., 2004
28 Southern rice black streaked dwarf
virus SRBSDV Fijivirus Reoviridae Lúa, ngô Giải trình tƣ
29 Rice yellow stunt virus (Rice transitory
yellowing virus) RYSV (RTYV) Nucleorhabdovirus Rhabdoviridae Lúa Giải trình tƣ
30 Rice ragged stunt virus RRSV Oryzavirus Reoviridae Lúa Giải trình tƣ
31 Banana bract mosaic virus BBrMV Potyvirus Potyviridae Chuối Rodoni et al., 1999
32 Bean common mosaic virus BCMV Potyvirus Potyviridae Đậu đỗ Ha et al., 2008c;
Hao et al., 2003
33 Chilli veinal mottle virus ChiVMoV Potyvirus Potyviridae Ớt Ha et al., 2008c
34 Chilli ringspot virus ChiRSV Potyvirus Potyviridae Ớt Ha et al., 2008c
35 Dasheen mosaic virus DsMV Potyvirus Potyviridae Khoai sọ, bán
hạ 3 thùy Ha et al., 2008c
36 Shallot yellow stripe virus SYSV Potyvirus Potyviridae Hành ta, tỏi
tây Ha et al., 2008c
37 Leek yellow stripe virus LYSV Potyvirus Potyviridae Hành ta, tỏi
tây Ha et al., 2008c
38 Onion yellow dwarf virus OYDV Potyvirus Potyviridae Tỏi tây Ha et al., 2008c

118
39 Papaya ringspot virus PRSV Potyvirus Potyviridae Đu đủ, bầu bí Bateson et al., 2002
40 Potato virus Y PVY Potyvirus Potyviridae Khoai tây, ớt Ha et al., 2008c
41 Sorghum mosaic virus SrMV Potyvirus Potyviridae Mía Ha et al., 2008c
42 Sugarcane mosaic virus SCMV Potyvirus Potyviridae Ngô, mía Ha et al., 2008c
43 Sweet potato feathery mottle virus SPFMV Potyvirus Potyviridae Khoai lang Ha et al., 2008c
44 Turnip mosaic virus TuMV Potyvirus Potyviridae Cải củ, cải bẹ Ha et al., 2008c
45 Zucchini yellow mosaic virus ZYMV Potyvirus Potyviridae Bầu bí Ha et al., 2008c
46 Telosma mosaic virus TelMV Potyvirus Potyviridae Hoa thiên lý Ha et al., 2008b
47 Basella rugose mosaic virus (Peace lily
mosaic virus)
BaRMV
(PeLMV) Potyvirus Potyviridae
Hoa lan ý,
mồng tơi Ha et al., 2008b
48 Wild tomato mosaic virus WTMV Potyvirus Potyviridae Cà độc dƣợc Ha et al., 2008b
49 Rice grassy stunt virus RGSV Tenuivirus Lúa Giải trình tƣ
50 Tobacco mosaic virus TMV Tobamovirus Virgaviridae Cà chua,
thuốc lá, ớt ELISA
51 Rice tungro bacilliform virus RTBV Tungrovirus Caulimoviridae Lúa Azzam et al., 2000
52 Rice tungro spherical virus RTSV Waikavirus Lúa ELISA

119
10.3 TÀI LIỆU THAM KHẢO CHÍNH
1. Azzam, O., Arboleda, M., Umadhay, K.M., de los Reyes, J.B., Cruz, F.S., Mackenzie, A. and
McNally, K.L. (2000). Genetic composition and complexity of virus populations at tungro-
endemic and outbreak rice sites. Archives of Virology. 145(12):2643-57.
2. Bateson, M. F., Lines, R. E., Revill, P., Chaleeprom, W., Ha, C. V., Gibbs, A. J. & Dale, J. L.
(2002). On the evolution and molecular epidemiology of the potyvirus Papaya ringspot virus.
Journal of General Virology 83, 2575-2585.
3. Bell, K. E., Dale, J. L., Ha, C. V., Vu, M. T. & Revill, P. A. (2002). Characterisation of Rep-
encoding components associated with banana bunchy top nanovirus in Vietnam. Archives of
Virology 147, 695-707.
4. Blawida, R., Van, D.T. and Maiss, E. (2008). Transreplication of a Tomato yellow leaf curl
Thailand virus DNA-B and replication of a DNAß component by Tomato leaf curl Vietnam
virus and Tomato yellow leaf curl Vietnam virus. Virus Research. Volume 136, 1-2: 107-117
5. Green, S. K., Tsai, W. S., Shih, S. L., Black, L. L., Rezaian, A., Rashid, M. H., Roff, M. M.
N., Myint, Y. Y. & Hong, L. T. A. (2001). Molecular characterisation of begomoviruses
associated with leafcurl diseases of tomato in Bangladesh, Laos, Malaysia, Myanmar, and
Vietnam. Plant Disease 85.
6. Ha Viet Cuong, Coombs, S., Revill, P., Harding, R., Vu, M., and Dale, J. (2006). Corchorus
yellow vein virus, a New World geminivirus from the Old World. Journal of General
Virology 87: 997-1003.
7. Ha Viet Cuong, Steven Coombs, Peter Revill, Rob Harding, Man Vu and James Dale.
(2008a). Molecular characterization of begomoviruses and DNA satellites from Vietnam:
additional evidence that the New World geminiviruses were present in the Old World prior to
continental separation. Journal of General Virology. 89: 312-326.
8. Ha Viet Cuong, S. Coombs, P. A. Revill, R. M. Harding, M. Vu, J. L. Dale. (2008b). Design
and application of two novel degenerate primer pairs for the detection and complete genomic
characterization of potyviruses. Archives of virology. 153:25-36
9. Ha Viet Cuong, S. Coombs, P. A. Revill, R. M. Harding, M. Vu, J. L. Dale. (2008c).
Identification and sequence analysis of potyviruses infecting crops in Vietnam. Archives of
virology. 153:45-60
10. Hà Minh Trung. (1984). Bệnh hại lúa ở Việt Nam. Khoa học và kỹ thuật Nông nghiệp. Số
12: 540 - 546.
11. Hao, N. B., Albrechtsen, S. E. & Nicolaisen, M. (2003). Detection and identification of the
blackeye cowpea mosaic strain of Bean common mosaic virus in seeds of Vigna unguiculata
sspp. from North Vietnam. Australasian Plant Pathology 32, 505-509.
12. Karan, M., Harding, R. M. & Dale, J. L. (1994). Evidence for 2 Groups of Banana Bunchy
Top Virus Isolates. Journal of General Virology 75, 3541-3546.
13. Kiritani, K. & Su, H. J. (1999). Papaya ring spot, banana bunchy top, and citrus greening in
the Asia and Pacific region: Occurrence and control strategy. Jarq-Japan Agricultural
Research Quarterly 33, 23-30.

120
14. Lheureux, F., Laboureau, N., Muller, E., Lockhart, B.E.L. and Iskra-Caruana, M.L. 2007.
Molecular characterization of banana streak acuminata Vietnam virus isolated from Musa
acuminata siamea (banana cultivar. Archives of Virology. Volume 152, 7:1409-1416.
15. Nguyễn Thơ. (1984). Điều tra nghiên cứu bệnh virus trên cà chua, thuốc lá và khoai tây. Tạp
chí Khoa học Nông nghiệp của Đại học Nông nghiệp I.
16. Revill, P. A., Ha, C. V., Porchun, S. C., Vu, M. T. & Dale, J. L. (2003). The complete
nucleotide sequence of two distinct geminiviruses infecting cucurbits in Vietnam. Archives
of Virology 148, 1523-1541.
17. Revill, P. A., Ha, C. V., Lines, R. E., Bell, K. E., Vu, M. T. & Dale, J. L. (2004). PCR and
ELISA-based virus surveys of banana, papaya and cucurbit crops in Vietnam. Asia Pacific
Journal of Molecular Biology and Biotechnology 12, 27-32.
18. Rodoni, B. C., Dale, J. L. & Harding, R. M. (1999). Characterisation and expression of the
coat protein-coding region of banana bract mosaic potyvirus, development of diagnostic
assays and detection of the virus in banana plants from five countries in southeast Asia.
Archives of Virology 144, 1725-1737.
19. Vakili, N. G. (1969). Bunchy top disease of bananas in the central Highlands of South
Vietnam. Plant Disease Reporter 53, 634-638.
20. Vũ Triệu Mân (1984). Nghiên cứu bệnh virus khoai tây ở miền Bắc Việt Nam. Tạp chí Khoa
học Nông nghiệp của Đại học Nông nghiệp I.

121
Chƣơng 11. VIRUS DNA SỢI KÉP (PHIÊN MÃ NGƯỢC): HỌ CAULIMOVIRIDAE
11.1 TÓM TẮT NỘI DUNG
Chƣơng 11 tóm tắt đặc điểm phân loại, phân tử, sinh học của nhóm virus DNA phiên mã
ngƣợc (pararetrovirus) hại thực vật là họ Caulimoviridae. Chƣơng này cũng mô tả đặc điểm của
một virus quan trọng bậc nhất trên lúa và có có mặt ở Việt Nam là RTBV (rice tungro
bacilliiorm virus).
11.2 HỌ CAULIMOVIRIDAE
11.2.1 Phân loại
Họ gồm 6 chi phân biệt bởi tổ chức bộ gen và hình thái phân tử. Trong số các caulimovirus
có 2 virus quan trọng là CaMV (có ý nghĩa trong CNSH thực vật) và RTBV (là một trong số các
virus nguy hiểm nhất trên lúa).
Bảng 11-1 Phân loại các virus thuộc họ Caulimoviridae
Chi Loài điển hình Số loài Hình thái
Caulimovirus Cauliflower mosaic virus 8 Cầu
Cavemovirus Cassava vein mosaic virus 2 Cầu
Petuvirus Petunia vein clearing virus 1 Cầu
Soymovirus Soybean chlorotic mottle virus 3 Cầu
Badnavirus Commelina yellow mottle virus 18 Nhộng
Tungrovirus Rice tungro bacilliform virus 1 Nhộng
11.2.2 Đặc điểm phân tử virus (virion)
Hình thái. Phân tử virus không có vỏ bọc, có hình cầu hoặc hình nhộng tùy theo chi; chẳng
hạn các virus thuộc chi Caulimovirus có hình cầu còn virus thuộc chi Tungrovirus có hình
nhộng. Các virus có hình cầu gọi là các caulimovirus còn các virus có hình nhộng còn đƣợc gọi
là các badnavirus.
Acid nucleic. Bộ gen virus chỉ gồm một phân tử DNA genome sợi vòng kép (dsDNA), có kích
thƣớc từ 7,8 – 8,2 kbp. DNA genome không liên tục: trên một sợi có một điểm rời, còn trên sợi
còn lại có từ 1 đến 3 điểm rời tùy theo chi. Các điểm rời này là điểm gắn mồi trong quá trình tái
sinh virus.
Protein. Bộ gen virus có từ 1 đến 7 ORF tùy thuộc chi. Các protein có chức năng chung cho
tất cả họ là CP, aspatate protease, RT và RNAse H.
11.2.3 Tổ chức bộ gen và phƣơng thức tái sinh
Tổ chức bộ gen là đặc điểm chính để phân biệt các chi. Các gen virus đƣợc mã hóa trên một
sợi DNA (sợi +, theo chiều kim đồng hồ). Sau khi xâm nhập vào nhân tế bào ký chủ, các điểm
rời đƣợc gắn kín và bộ gen virus trở thành sợi vòng kép hoàn chỉnh (còn gọi là

122
minichromosome). Bộ gen virus tiếp theo đƣợc phiên mã theo kiểu bất đối xứng, tức là từ khuôn
sợi (-), nhờ DNA-dependent RNA polymerase của ký chủ để tạo ra phiên bản RNA (transcript)
có kích thƣớc lớn hơn genome. Phiên bản RNA có 2 chức năng: (1) làm khuôn cho quá trình
phiên mã ngƣợc để tạo sợi DNA của virus (sợi -, ngƣợc chiều kim đồng hồ) và (2) làm khuôn để
dịch mã gen virus.
11.2.4 Đặc điểm sinh học
Phần lớn các virus có phổ ký chủ hẹp. Các virus thuộc chi Petuvirus, Soymovirus và
Cavemovirus chỉ nhiễm cây hai lá mầm. Virus thuộc chi Tungrovirus (hiện có 1 đại diện duy
nhất là RTBV) chỉ nhiễm cây một lá mầm. Các virus thuộc chi Badnavirus nhiễm cả cây một và
hai lá mầm.
11.2.5 Các virus thuộc họ Caulimoviridae có ở Việt Nam
Hiện nay, mới có 2 virus thuộc họ này chắc chắn có ở Việt Nam là:
1. RTBV (cùng với RTSV) gây bệnh tungrovirus. Bênh mặc dù đã xuất hiện từ lâu tại Việt
Nam nhƣng chƣa có ghi nhận bệnh gây thành dịch ở Việt Nam.
2. BSAcVNV (Banana streak acuminata Vietnam virus). Đây là một virus mới đƣợc công bố
năm 2007. Virus đƣợc phân lập từ mẫu cây chuối Musa acuminata siamea ở Việt Nam.
Lúc đầu, virus đã đƣợc cho là do BSV (Banana streak virus) gây ra nhƣng sau khi giải
trình tự toàn bộ bộ gen (7801 bp), các tác giả đã phát hiện đây là một loài mới và đặt tên
là BSAcVNV (Lheureux et al., 2007)
11.3 RICE TUNGRO BACILLIFORM VIRUS
11.3.1 Tầm quan trọng
RTBV là một virus quan trọng bậc nhất trên lúa khắp thế giới. Bệnh có ở Việt Nam nhƣng
chƣa có công bố chính thức rằng bệnh đã gây thành dịch nghiêm trọng.
11.3.2 Virus
RTBV thuộc chi Tungrovirus, họ Caulimoviridae. Tên ―bacilliform‖ đƣợc đặt dựa trên hình
dạng của virus là có hình vi khuẩn hay hình nhộng, kích thƣớc phân tử ~ 130 x 30 nm.
RTBV, giống nhƣ các virus trong họ, là một pararetrovirus (có nghĩa là có bản chất bộ gen là
DNA nhƣng khi tái sinh phải qua một bƣớc trung gian là phiên mã ngƣợc từ DNA sang RNA).
Bộ gen virus là một phân tử DNA sợi vòng kép, kích thƣớc ~ 8 kb, chứa 2 vị trí không liên
tục là D1 và D2.
Trên bộ gen virus có 4 ORF đƣợc tổ chức theo chiều kim đồng hồ, mã hóa 4 protein có kích
thƣớc 24, 12, 194, và 46 kDa (đƣợc gọi là các protein P24, P12, P194 và P46). Các ORF 1, 2, 3
xắp xếp gần nhau và gối lên nhau.
P24 có chức năng chƣa rõ nhƣng có thể liên quan đến quá trình lắp ráp phân tử. Đáng chú ý là
ORF của protein này không có mã khởi đầu ATG (ở cả 3 khung) nhƣng lại đƣợc dịch từ mã thứ
2 ATT. ORF này cũng không có mã kết thúc.
P194 là protei quan trong nhất, chứa 4 vùng chức năng: CP (coat protein), aspartate protease,
RT (reverse transcriptase) và RH (RNase H).

123
P12 có chức năng tăng cƣờng dịch mã, đƣợc biểu hiện từ phân tử RNA nhỏ riêng (phân tử
này đƣợc cắt ra từ phân tử RNA lớn)
Hình 11-1 Hình thái phân tử và tổ chức bộ gen của RTBV (Hull, 1996)
11.3.3 Triệu chứng
RTBV khi nhiễm cùng với một virus RNA là RTSV (Rice tungro spherical virus) sẽ gây ra
một bệnh rất nghiêm trọng trên lúa gọi là bệnh tungro. Triệu chứng điển hình là cây lùn mạnh, lá
biến màu vàng, để nhánh giảm. Lá biến vàng bắt đầu từ chót lá. Lá bệnh có thể nhỏ, không cân
đối, có thể có các vệt màu nâu tối. Cây bị bệnh tungro có thể không trỗ bông, hoặc nếu trỗ thì hạt
cũng bị lép và biến màu
Cây bị nhiễm với chỉ RTBV triệu chứng nhẹ hơn còn cây bị nhiễm với chỉ RTSV có thể hơi
còi cọc nhƣng không biến vàng.
Hình 11-2 Cây lúa nhiễm các tổ hợp virus khác nhau và khả năng truyền virus qua rầy xanh đuôi
đen (Azzam et al., 2000)

124
11.3.4 Lan truyền
Ngoài tự nhiên virus truyền qua nhiều loài rầy xanh đuôi đen (Nephortettix spp.), trong đó
hiệu quả nhất là loài N. virescens. Tuy nhiên RTBV không thể tự lan truyền nhờ rầy đƣợc mà
phải nhờ sự hỗ trợ của virus RTSV. Điều này có nghĩa rầy chích hút trên cây chỉ nhiễm RTBV sẽ
không truyền virus. Rầy chích trên cây nhiễm hỗn hợp RTBV + RTSV sẽ truyền đƣợc virus.
Phƣơng thức truyền qua rầy là kiểu bán bền vững. Thời gian ẩn trong vector trung bình khoảng 3
ngày (thay đổi theo loài vector, điều kiện nhiệt độ). Thời gian tồn tại khả nhiễm chỉ vài ngày.
Virus không truyền qua tiếp xúc cơ học cũng nhƣ hạt giống và cây lúa là ký chủ tự nhiên duy
nhất của virus.
11.3.5 Phòng chống
Hiện không có biện pháp nào có thể trực tiếp chống virus. Các biện pháp có thể áp dụng đều
liên quan tới quản lý vector rầy xanh đuôi đen (dùng giống kháng rầy, thuốc hóa học…).
11.4 CÂU HỎI ÔN TẬP CHƢƠNG 11
1. Đặc điểm họ Caulimoviridae (hình thái, tổ chức bộ gen và phƣơng thức tái sinh)
2. Virus RTBV (tầm quan trọng, tổ chức bộ gen, triệu chứng, lan truyền)..
3. Virus CaMV (tầm quan trọng, tổ chức bộ gen, triệu chứng, lan truyền).
4. Tại sao promoter của CaMV đƣợc sử dụng nhiều trong CNSH và các nguy cơ có thể khi sử
dụng promoter này trong công nghệ chuyển gen thực vật.
11.5 TÀI LIỆU THAM KHẢO CHÍNH
1. Azzam, O., Arboleda, M., Umadhay, K. M., de los Reyes, J. B., Cruz, F. S., Mackenzie, A. &
McNally, K. L. (2000). Genetic composition and complexity of virus populations at tungro-
endemic and outbreak rice sites. Archives of Virology 12, 2643-2657.
2. Hass, M., Bureau, M., Geldreich, A., Yot, P. & Keller, M. (2002). Cauliflower mosaic virus:
still in the news. MOLECULAR PLANT PATHOLOGY 3, 419–429.
3. Hull, R. (1996). Molecular Biology of Rice tungro viruses. Annual Review of
Phytopathology 34, 275–297.
4. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
5. Hull, R., Geering, A., Harper, G., Lockhart, B. E. & Schoelz, J. E. (2005). Caulimoviridae.
In: Virus Taxonomy. VIIIth report of the International Committee on Taxonomy of Viruses.
(C.M. Fauquet, M.A Mayo, J. Maniloff, U. Desselberger and L.A. Ball, eds).
Elsevier/Academic Press. London., 385 - 396.
6. Lheureux, F., Laboureau, N., Muller, E., Lockhart, B. E. L. & Iskra-Caruana, M. L. (2007).
Molecular characterization of banana streak acuminata Vietnam virus isolated from Musa
acuminata siamea (banana cultivar). Archives of Virology 152, 1409-1416

125
Chƣơng 12. VIRUS DNA SỢI ĐƠN: CHI BEGOMOVIRUS (HỌ GEMINIVIRIDAE)
12.1 TÓM TẮT NỘI DUNG
Chƣơng 12 mô tả tóm tắt đặc điểm phân loại, phân tử, sinh học của họ Geminiviridae, một
trong hai họ virus thực vật lớn nhất. Chƣơng này cũng mô tả chi tiết đặc điểm của các virus
thuộc chi Begomovirus - là chi virus quan trọng nhất của họ. Một số bệnh do begomovirus có ý
nghĩa kinh tế trên thế giới và Việt Nam cũng đƣợc mô tả, đặc biệt chi tiết đối với bệnh xoăn vàng
lá cà chua.
12.2 HỌ GEMINIVIRIDAE
Họ Geminiviridae là một trong 2 họ virus thực vật lớn nhất (cùng với họ Potyviridae). Số loài
đƣợc công nhận chính thức bởi ICTV năm 2009 là 218 loài (Bảng 1), chiếm khoảng 20% tổng số
loài virus thực vật. Từ ―Gemini‖ có nghĩa là ―sinh đôi hay cặp đôi‖, ám chỉ hình thái dạng cầu
kép của phân tử virus.
Các geminivirus có hình thái phân tử dạng hình cầu kép, có bộ gen DNA dạng vòng, sợi đơn,
kích thƣớc ~ 2.7 kb. Các geminivirus có thể có bộ gen đơn (monopartite) gồm 1 phân tử DNA
hoặc bộ gen kép (bipartite) nếu có 2 phân tử DNA genome.
Các geminivirus đƣợc phân loại trên cơ sở tổ chức bộ gen và vector truyền bệnh. Cho tới nay
(số liệu năm 2009 của ICTV), 4 chi trong họ đƣợc ghi nhận bao gồm: Mastrevirus, Curtovirus,
Topocuvirus và Begomovirus.
Bảng 12-1. Phân loại các virus họ Geminiviridae theo ICTV năm 2009
Nhiều geminivirus là các virus gây bệnh cây có ý nghĩa kinh tế. Một số ví dụ điển hình là:
MSV (Maize streak virus, chi Mastrevirus), gây bệnh khảm sọc ngô, rất phổ biến ở châu
Phi.
Các begomovirus (chi Begomovirus) gây bệnh khảm lá sắn ở châu Phi, tiểu lục địa Ấn
Độ, bệnh cuốn lá bông, và đặc biệt bệnh xoăn vàng lá cà chua khắp thế giới.
Chi Loài điển hình Bộ gen Vector Số loài
Mastrevirus Maize streak virus (MSV) Đơn Rầy lá 14
Curtovirus Beet curly top virus (BCTV) Đơn Rầy lá 7
Topocuvirus Tomato pseudo-curly top virus (TPCTV) Đơn Rầy than 1
Begomovirus Bean golden mosaic virus (BGMV) Đơn hoặc kép Bọ phấn 196
Tổng số 218

126
12.3 CHI BEGOMOVIRUS
12.3.1 Giới thiệu
Begomovirus là 1 trong 4 chi của họ Geminiviridae và là chi lớn nhất trong họ với tổng số
loài đƣợc công nhận bởi ICTV năm 2009 là 196 loài (trong tổng số 218 loài của họ).
Có thể thấy chi Begomovirus cũng là 1 trong 2 chi (cùng với chi Potyvirus, họ Potyviridae)
lớn nhất trong số các chi virus thực vật (chiếm ~ 20 tổng số virus).
Do số lƣợng virus rất nhiều, lại thƣờng gây bệnh với triệu chứng giống nhau trên một cây ký
chủ nên tên virus gây bệnh thƣờng thêm tên địa danh (nƣớc, tỉnh), ví dụ nhƣ Tomato leaf curl
Vietnam virus.
12.3.2 Hình thái phân tử
Hình thái phân tử (virion) của begomovirus giống nhƣ của các geminivirus khác, đều có
hình cầu kép. Phân tử virus thực chất là 2 hình cầu 20 mặt (icosahedron), mỗi mặt là 1 tam giác
đều với số đơn vị tam giác (T) trên mỗi mặt = 1, nối với nhau để tạo ra phân tử hình cầu đa diện
kép (gemini). Do nối với nhau nên 2 hình cầu này không hoàn thiện dẫn tới trên mỗi hình cầu chỉ
có 55 tiểu phần protein (protein vỏ) đƣợc xắp xếp thành 11 đơn vị hình thái, mỗi đơn vị gồm 5
tiểu phần protein (pentameric capsomer). Kết quả là toàn bộ phân tử có 110 tiểu phần và 22 đơn
vị hình thái.
Cần chú ý là đối với các begomovirus có bộ gen kép thì mỗi phân tử chỉ chứa 1 phân tử
DNA genome.
Hình 12-1 Hình thái phân tử geminiviruses. A, Phân tử Tomato yellow leaf curl virus (TYLCV),
bar = 100 nm (Gafni, 2003). B, Mô hình phân tử Maize streak virus (MSV) (Zhang et al., 2001).
12.3.3 Đặc điểm bộ gen
Các begomovirus là các virus thực vật có bộ gen DNA sợi vòng đơn có kích thƣớc khoảng 2.6
– 2.8 kb. Chúng hoặc có bộ gen kép (bipartite) gồm 2 phân tử DNA gọi là DNA-A và DNA-B
hoặc có bộ gen đơn (monopartite) tƣơng đƣơng DNA-A.
Đối với các virus có bộ gen đơn thì DNA của chúng chứa 6 gen đƣợc tổ chức theo 2 chiều
ngƣợc nhau.
A B

127
Trên chiều kim đồng hồ (chiều virus) có 2 ORF là (i) V1 mã hóa vỏ protein (CP) có chức
năng chính là tạo vỏ phân tử virus, lan truyên qua vector, vận chuyển bộ gen virus vào và ra khỏi
nhân tế bào ký chủ và vận chuyển bộ gen virus giữa các tế bào; và (ii) V2 mã hóa protein V2 có
chức năng liên quan đến vận chuyển virus giữa các tế bào.
Trên chiều ngƣợc kim đồng hồ (chiều sợi tƣơng đồng virus) có 4 ORF là (i) C1 mã hóa
protein tái sinh hay còn gọi là protein Rep có chức năng chính là cắt - nối bộ gen virus tại một vị
trí đặc biệt ở vùng nguồn gốc tái sinh và tƣơng tác với protein ký chủ để tạo điều kiện thuận lợi
cho tái sinh virus, (ii) C2 mã hóa 1 protein hoạt hóa phiên mã (TrAP, transcriptional activator
protein), (iii) C3 mã hóa 1 protein tăng cƣờng tái sinh (REn, replication enhancer) và (iv) C4 mã
hóa protein C4 với chức năng liên quan đến phổ ký chủ và phát triển triệu chứng, ức chế hoạt
động câm gen của tế bào ký chủ.
Giữa 2 vùng gen mã hóa ngƣợc chiều nhau ở trên là một vùng liên gen không mã hóa IR
(intergenic region). Vùng IR chứa nguồn gốc tái sinh (ori = origin of replication) gồm (i) các
chuỗi lặp đảo (iteron) cần thiết cho sự nhận biết và gắn kết của protein Rep và (ii) một cấu trúc
kẹp tóc (hairpin) hay còn gọi là thân – thòng lọng (stem – loop), trong đó chuỗi TAATATTAC
trên vùng thòng lọng là giống nhau ở tất cả các begomovirus. Vị trí T7-C
8 của chuỗi này là nơi
protein Rep cắt và nối bộ gen begomovirus trong quá trình tái sinh.
Đối với các begomovirus có bộ gen kép thì DNA-A có tổ chức bộ gen và chức năng các gen
tƣơng tự nhƣ DNA-A của các begomovirus có bộ gen đơn (tên các ORF đƣợc thêm ký tự A vào
đằng trƣớc thành AV1, AV2, AC1, AC2, AC3 và AC4). Cần chú ý là tất cả các begomovirus ở
Tân thế giới (châu Mỹ) đều thiếu ORF AV2.
Phân tử DNA-B của chúng chỉ chứa 2 ORF đƣợc sắp xếp theo 2 chiều ngƣợc nhau. Trên
chiều kim đồng hồ có ORF BV1 mã hóa protein con thoi (NSP, nuclear shuttle protein) có chức
năng chính là vận chuyển bộ gen virus vào và ra khỏi nhân tế bào. Trên chiều ngƣợc kim đồng
hồ có ORF BC1 mã hóa protein vận chuyển (MP, movement protein) có chức năng vận chuyển
bộ gen virus giữa các tế bào ký chủ.
Ở vùng IR của DNA-A và DNA-B của các begomovirus có bộ gen kép có một chuỗi bảo thủ
cao (khoảng 150 nucleotide) giữa 2 phân tử gọi là vùng chung CR (common region). Vùng CR
chứa chuỗi ori. Sở dĩ có vùng chung này vì DNA-B không thể tự tái sinh mà phải cần sự nhận
biết và cắt – nối của protein Rep mã hóa trên DNA-A để tái sinh đƣợc trong tế bào ký chủ.
12.3.4 DNA vệ tinh của begomovirus
Có 2 loại phân tử DNA vệ tinh thƣờng đƣợc phát hiện có liên quan đến các begomovirus có
bộ gen đơn tại Cựu thế giới (Châu Á, Châu Phi, Châu Âu và Châu Uc). Các phân tử này đều có
bộ gen DNA sợi vòng đơn với kích thƣớc xấp xỉ ½ kích thƣớc bộ gen virus chủ. Các phân tử này
gồm 2 loại là:
Alphasatellite (trƣớc đây gọi là DNA-1). Loại phân tử vệ tinh này có đặc điểm cấu trúc giống
nhƣ DNA-1 của nanovirus nhƣ DNA sợi vòng đơn, kích thƣớc khoảng 1.2 kb, chứa 1 vùng ori,
mã hóa 1 protein tái sinh (Rep) theo chiều kim đồng hồ và 1 vùng giàu adenine. Alphasatellite có
thể tự tái sinh trong tế bào cây, đƣợc lắp ráp trong phân tử virus và nhờ virus để phát tán qua bọ
phấn. Tuy nhiên vệ tinh này không có vai trò trong tính gây bệnh của virus.
Betasatellite (trƣớc đây gọi là DNA-β). Loại phân tử vệ tinh này đã thu hút sự chú ý kể từ khi
Saunders et al. (2000) và Briddon et al. (2001) chứng minh rằng triệu chứng điển hình trên cây
ageratum (bệnh vàng gân cây ageratum) và cây bông (bệnh cuốn lá bông) chỉ có thể hình thành
khi Ageratum yellow vein virus (AYVV) và Cotton leaf curl virus (CLCV) cùng đƣợc lây nhiễm

128
với 1 phân tử DNA-β tƣơng ứng. Tuy nhiên vai trò của phân tử vệ tinh này đối với sự biểu hiện
triệu chứng bệnh không thống nhất. Nhiều begomovirus bộ gen đơn đã đƣợc chứng minh là có
thể tạo ra triệu chứng điển hình mà không cần sự có mặt của phân tử betasatellite chẳng hạn nhƣ
đối với Tobacco yellow leaf curl Yunnan virus (TYLCYNV)
Betasatellite có bộ gen DNA sợi vòng đơn, có kích thƣớc bằng 1 nửa của begomovirus (phần
lớn là 1350 nucleotides) và mã hóa trên sợi tƣơng đồng 1 protein là betaC1 (βC1) có kích thƣớc
điển hình 118 amino acids. Phân tử chứa 1 vùng giàu adenin và 1 vùng bảo thủ cao giữa các
phân tử betasatellite khác nhau gọi là vùng SCR (satellite conserve region). Trên vùng SCR có
một trình tự bảo thủ TAATATTAC giống nhƣ của begomovirus. Betasatellite đƣợc lắp ráp trong
phân tử virus, chỉ có thể tái sinh nhờ virus. Betasatllite có thể ức chế phản ứng phòng thủ của cây
và do đó có vai trò quan trọng trong tính gây bệnh của nhiều begomovirus có bộ gen đơn.
12.3.5 Tái sinh của begomovirus
Begomovirus, giống nhƣ các geminivirus khác, tái sinh trong tế bào thực vật theo cơ chế vòng
lăn (rolling circular mechanism). Cơ chế vòng lăn có thể đƣợc chia làm 2 pha và đƣợc thực hiện
trong nhân tế bào ký chủ (xem hình vẽ phần đại cƣơng)
1. Pha tổng hợp sợi DNA vòng đơn (bộ gen có mặt trong phân tử virus) thành sợi DNA vòng
kép khi bộ gen virus đƣợc chuyển vào nhân tế bào. Nhƣ vậy sợi kép sẽ gồm một sợi virus và
một sợi tƣơng đồng virus. Pha này vẫn chƣa đƣợc hiểu rõ.
2. Pha tái sinh theo cơ chế vòng lăn: Protein Rep (sau khi đƣợc tổng hợp) sẽ cắt sợi virus tại
chuỗi bảo toàn TATATTAC. Nhờ vật liệu cũng nhƣ enzyme DNA polymearase của tế bào,
sợi virus đƣợc tổng hợp liên tục trên sợi tƣơng đồng virus. Protein Rep lại tiếp tục cắt sợi
virus mới đƣợc tổng hợp tại chuỗi TATATTAC (cũng vừa mới đƣợc tổng hơp) thành một sợi
virus hoàn chỉnh dƣới dạng sợi đơn mạch thẳng. Protein Rep sau đó sẽ nối 2 đầu của mạch
thẳng để tạo ra bộ gen virus sợi đơn mạch vòng hoàn chỉnh.
Ngoài cơ chế vòng lăn, gần đây ngƣời ta còn phát hiện begommovirus thƣờng tái sinh theo cơ
chế gọi là ―tái sinh phụ thuộc tái tổ hợp‖ (Recombination dependent replication (RDR)). Cách tái
sinh này cho phép virus có thể tái tổ hợp dễ dàng khi nhiễm hỗn hợp trên một cây ký chủ.
12.3.6 Cơ chế gây bệnh của begomovirus
Tương tác với các yếu tố ký chủ liên quan đến bộ máy tái bản. Quá trình tái bản của
begomovirus xảy ra trong nhân tế bào, kể cả ở các tế bào không phân chia. Do vậy, sau khi
nhiễm vào tế bào, begomovirus phải khởi động bộ máy tái sinh của tế bào. Một trong các protein
của tế bào ký chủ thực vật điều khiển chu kỳ tế bào là pRBR (plant retinoblastoma-related
protein) - chịu trách nhiệm chuyển chu kỳ tế bào từ pha G1 sang pha S (pha tổng hợp). Protein
Rep của begomovirus đã đƣợc chứng minh tƣơng tác với pRBR của tế bào ký chủ, cho phép
khởi động lại chu kỳ tế bào để tạo điều kiện cho virus tái sinh.
12.3.7 Lan truyền của begomovirus
Tất cả các begomovirus lan truyền ngoài tự nhiên nhờ bọ phấn (B. tabaci) theo kiểu bền vững
tuần hoàn. Chƣa có bằng chứng chứng minh begomovirus nhân lên trong cơ thể bọ phấn.

129
Hình 12-2 Tổ chức bộ gen của begomovirus và DNA vệ tinh (Hà Viết Cƣờng, 2008)
12.3.8 Phòng trừ begomovirus
Phòng trừ begomovirus chủ yếu dựa vào 2 chiến lƣợc chính là là (i) phòng chống vector và
(ii) tạo cây kháng bệnh.
Phòng chống vector. Có nhiều kỹ thuật phòng trừ bọ phấn nhƣng đƣợc áp dụng tùy điều kiện:
Dùng giống kháng bọ phấn
Dùng thuốc hóa học.
Trồng cây trong nhà lƣới, nhà kính
Dùng bẫy hấp dẫn màu vàng hoặc bề mặt phản xạ (chỉ áp dụng có hiệu quả trong điều
kiện nhà lƣới)
Tạo giống kháng virus. Tính kháng begomovirus có thể đƣợc tạo ra nhờ 2 cơ chế: Tính
kháng từ cây và tính kháng từ tác nhân gây bệnh (PRD).
α satellite βC1
Rep
SCR
A-rich
β satellite
A-rich
C2 (TrAP)
V2
V1 (CP)
C4
C1 (Rep)
C3 (REn)
Begomovirus
(bộ gen đơn)
DNA genome
AV1
(CP)
AC3
(REn) AC2
(TrAP)
AC1
(Rep)
AC4
CR
BV1
(NSP)
CR
BC1
(MP)
AV2
Begomovirus
(bộ gen kép)
DNA-B
Begomovirus
(bộ gen kép)
DNA-A

130
Trên cây cà chua, ngƣời ta chƣa phát hiện thấy gen kháng R chống lại begomovirus trên cây
cà chua trồng (Lycopersicon esculentum) nhƣng một số gen kháng chống lại begomovirus đã
đƣợc phát hiện thấy trên một số giống cà chua dại (gọi là các gen Ty. Ví dụ Ty1 gen đƣợc phân
lập từ cây cà chua dại (Lycopersicon chilense) và là một gen kháng trội không hoàn toàn. Ty-1
dƣờng nhƣ tƣơng tác với các protein chịu trách nhiệm di chuyển của virus để tạo tính kháng. Ty-
1 đã đƣợc chuyển vào cà chua trồng để tạo giống cà chua kháng. Hiện nay, nhiều giống cà chua
kháng begomovirus là các giống kháng chuyển gen dùng gen kháng từ cây.
Gần đây nhiều giống cây (cà chua, bông, sắn) đang đƣợc thử nghiệm tạo tính kháng thông qua
cơ chế RNAi. Các gen thƣờng đƣợc sử dụng để tạo cấu trúc chuyển gen là CP hoặc Rep.
12.4 BỆNH DO BEGOMOVIRUS
Begomovirus gây nhiều bệnh quan trọng trên cây trồng. Các triệu chứng điển hình do
begomvirus gây ra bao gồm: Vàng gân, khảm biến vàng (kèm theo biến dạng lá), biến vàng lá,
lùn cây, xoăn lá. Một đặc điểm quan trọng là nhiều bệnh có thể do các begomovirus thuộc các
loài khác hay gây ra. Dƣới đây trình bày một số bệnh quan trọng trên thế giới và Việt Nam
12.4.1 Bệnh khảm lá sắn
Triệu chứng điển hình là lá bị khảm, biến dạng dữ dội, cây lùn. Bệnh khảm lá sắn là bệnh
nguy hiểm nhất trên cây sắn tại châu Phi do một số begomovirus có bộ gen kép gây ra nhƣ
ACMV (African cassava mosaic virus), EACMV (East African cassava mosaic virus), SACMV
(South African cassava mosaic virus). Ngoài châu Phi, bệnh cũng đƣợc phát hiện tại tiểu lục địa
Ấn Độ nhƣ ICMV (Indian cassava mosaic virus); SLCAM (Srilancan cassava mosaic virus). Ở
Việt Nam, chƣa phát hiện thấy bệnh. Vì bệnh có thể truyền qua hom sắn giống nên vấn đề kiểm
dịch bệnh này rất quan trọng do cây sắn là một cây công nghiệp có ý nghĩa kinh tế của Việt
Nam.
Hình 12-3 Bệnh khảm lá sắn tại châu Phi. B và E là triệu chứng bệnh; C là vector bọ phấn và F là
cây chuyển gen kháng bệnh (Legg & Fauquet, 2004).
12.4.2 Bệnh cuốn lá bông
Bệnh cuốn lá là một trong các bệnh nguy hiểm nhất tại các nƣớc trồng bông thuộc tiểu lục địa
Ấn Độ nhƣ Pakistan, Ấn Độ. Triệu chứng điển hình là cuốn lá và lùn cây. Ngoài ra trên lá bông

131
bệnh thƣờng biểu hiện triệu chứng phồng gân và mọc lá phụ. Ở châu Á, bệnh do ít nhất 5
begomovirus gây ra bao gồm Cotton leaf curl Alabad virus, Cotton leaf curl Bangalore virus,
Cotton leaf curl Gezira virus, Cotton leaf curl Kokhran virus, Cotton leaf curl Multan virus. Tất
cả các begomovirus gây hại trên bông ở châu Á đều là các virus có bộ gen đơn và đều cần một
phân tử betasatellite nhiễm kèm để tạo triệu chứng.
Bệnh cuốn lá bông đã đƣợc quan sát thấy tại miền Bắc Việt Nam. Tuy nhiên nguyên nhân gây
bệnh chƣa đƣợc xác định.
Hình 12-4 Triệu chứng bệnh cuốn lá bông tại Pakistan. Chú ý sự hình thành lá phụ ở mặt sau của
lá bông ở ảnh dƣới bên phải (Bridon & Markham, 2000)
12.4.3 Bệnh xoăn vàng lá cà chua
12.4.3.1 Tầm quan trọng
Bệnh xoăn lá cà chua đƣợc ghi nhận đầu tiên trên thế giới từ cuối những năm 40 tại Israel.
Hiện nay, bệnh xoăn vàng lá đã trở thành bệnh virus quan trọng nhất trên cây cà chua khắp thế
giới. Tại Việt Nam, bệnh xoăn vàng lá là bệnh virus nguy hiểm nhất trên cà chua.
12.4.3.2 Triệu chứng bệnh
Bệnh xoăn vàng lá xuất hiện triệu chứng trong vòng 2-4 tuần sau khi nhiễm bệnh và phát triển
đầy đủ triệu chứng trong vòng 2 tháng. Triệu chứng có thể thay đổi theo điều kiện môi trƣờng,
giai đoạn sinh trƣởng và điều kiện sinh lý của cây tại thời điểm nhiễm bệnh
Triệu chứng sớm nhất là lá cong xuống dƣới vào phía bên trong. Về sau, lá không có hình
dạng, nhỏ hẹp, biến vàng từ mép và chót lá lan vào giữa gân; lá cuốn cong lên phía trên thành
hình thuyền; lá non biến vàng mạnh, giòn và nhỏ hẹp. Triệu chứng biến vàng đặc biệt rõ ở các lá
non. Cuống lá có thể xoắn vặn. Cây lùn còi cọc, mọc nhiều cành nhánh nhỏ, đốt thân ngắn. Cây
nhiễm sớm thƣờng không ra quả do hoa bị rụng.
Chú ý quan trọng: Triệu chứng do các begomovirus khác nhau gây ra trên cà chua thƣờng
tƣơng tự nhau.

132
12.4.3.3 Tác nhân gây bệnh là một phức hợp nhiều virus
Trƣớc những năm 90, tác nhân gây bệnh xoăn vàng lá cà chua đã đƣợc xác định do các
begomovirus. Các begomovirus gây bệnh thƣờng đƣợc đặt tên theo cây ký chủ (tomato) và triệu
chứng, chủ yếu dƣới 2 dạng xoăn lá (leaf curl) và biến vàng (yellow), nên tên virus gây bệnh là
―Tomato yellow leaf curl virus‖ và tên các isolate virus gây bệnh thƣờng đƣợc kèm theo 1 từ chỉ
địa phƣơng nơi bệnh xuất hiện; ví dụ:
Tomato yellow leaf curl virus- Is (TYLCV-Is) phân lập từ Israel,
Tomato yellow leaf curl virus – Sar (TYLCV-Sar) phân lập từ đảo Sardinia –Ý
Tomato yellow leaf curl virus – Th (TYLCV-Th) phân lập từ Thái Lan…
Do có ít lựa chọn trong việc đặt tên, các tác giả cũng có xu hƣớng lấy chỉ triệu chứng xoăn lá
―leaf curl‖ để đặt tên virus mặc dù các virus này đều tạo triệu chứng biến vàng trên cà chua;
chẳng hạn:
Tomato leaf curl virus-Au (ToLCV-Au) phân lập từ Öc
Tomato leaf curl Guzarat virus (ToLCGV) phân lập từ tỉnh Guzarat (Pakistans)
Từ những năm 90 trở lại đây, sự tiến bộ của công nghệ sinh học đã cho phép giải trình tự toàn
bộ bộ gen virus và xác định danh tính virus một cách dễ dàng. Dựa vào các phân tích phân tử,
Ủy ban Phân loại Virus Quốc tế (ICTV) đã xác định ngƣỡng phân biệt loài begomovirus là 89 %
mức đồng nhất trình tự nucleotide của phân tử DNA-A. Chỉ tiêu này đã cho phép tách đƣợc
nhiều begomovirus, vốn đƣợc xem là các isolate khác nhau của cùng một virus gây bệnh xoăn
vàng lá cà chua, thành các virus phân biệt; ví dụ:
TYLCV-Sar trở thành Tomato yellow leaf curl Sardinia virus (TYLCSV)
TYLCV-Ch trở thành Tomato yellow leaf curl China virus (TYLCCNV)
TYLCV-Th trở thành Tomato yellow leaf curl Thailand virus (TYLCSV).
TYLCV-Is phân lập từ Israel, do quyền ƣu tiên, đƣợc công nhận là Tomato yellow
leaf curl virus (TYLCV).
Nhƣ vậy, tác nhân gây bệnh xoăn vàng lá cà chua đƣợc xem là một phức hợp nhiều
begomovirus khác nhau. Hiện nay, có khoảng 60 begomovirus gây bệnh xăn vàng lá cà chua
khắp thế giới. Hậu quả là, dựa trên triệu chứng quan sát, bệnh không thể đƣợc gán cho một loài
begomovirus cụ thể trừ phi danh tính của virus đã đƣợc xác định.
Nhiều begomovirus gây bệnh xoăn vàng lá cà chua ở Cựu thế giới (Châu Á, Âu, Phi, Đại
Dƣơng) là các begogomovirus có bộ gen đơn và thƣờng đƣợc phát hiện kèm với các phân tử
DNA vệ tinh là alphasatellite và betasatellite.
12.4.3.4 Lan truyền
Tất cả các begomovirus lan truyền ngoài tự nhiên nhờ bọ phấn theo kiểu bền vững tuần hoàn.
Chƣa có bằng chứng chứng minh begomovirus nhân lên trong cơ thể bọ phấn. Bọ phấn dùng vòi
chọc vào mô mạch dẫn để hút dịch cây từ mạch phloem. Virus đƣợc hút qua vòi, tới diều, thấm
qua màng ruột vào xoang cơ thể, đạt tới tuyến nƣớc bọt và cuối cùng vào ống nƣớc bọt.
Nghiên cứu với TYLCV tại Israel cho thấy thời gian chích nạp và chích truyền tối thiểu của
bọ phấn là khoảng 15 – 20 phút. Kể từ khi bắt đầu chích nạp, virus đƣợc phát hiện có mặt ở

133
phần đầu sau khoảng 10 phút, ở ruột giữa sau khoảng 50 phút, ở xoang cơ thể sau khoảng 90
phút và ở tuyến nƣớc bọt sau khoảng 7 giờ. Thời gian từ khi virus đƣợc phát hiện thấy ở tuyến
nƣớc bọt tới khi bọ phấn có thể truyền đƣợc bênh là khoảng 1 giờ. Nhƣ vậy thời kỳ ẩn của
TYLCV trong cơ thể bọ phấn là khoảng 8 giờ.
Thí nghiệm với TYLCV cũng cho thấy, số lƣợng virus đƣợc tích lũy đạt cực đại (khoảng 600
triệu phân tử virus) nếu để bọ phấn chích nạp trên cây bệnh 12 giờ. Thời gian này đối với
TYLCSV là 24 giờ.
Sau thời gian chích nạp khoảng 1-2 ngày, virus có thể đƣợc duy trì trong cơ thể bọ phấn nhiều
tuần (3 tuần đối với TYLCSV) hoặc cả đời nhƣ đối với TYLCV.
Virus có thể truyền qua giao phối từ bọ phấn đực sang bọ phấn cái và ngƣợc lại. Đã có bằng
chứng cho thấy TYLCV có thể truyền qua trứng mặc dù tỷ lệ truyền bệnh của bọ phấn trƣởng
thành phát triển từ trứng mang virus là rất nhỏ. Trái lại, thí nghiệm với Tomato mottle virus
(ToMoV) cho thấy bọ phấn không truyền virus này qua trứng.
Một cá thể bọ phấn có thể truyền bệnh sau khi chích nạp 24 giờ mặc dù tỷ lệ truyền bệnh
không cao; tỷ lệ truyền bệnh đạt 100 % nếu sử dụng 5 – 15 bọ phấn. Bọ phấn cái truyền virus
hiệu quả hơn bọ phấn đực và hiệu quả truyền tốt nhất khi bọ phấn trƣởng thành ở 1 -2 tuần tuổi
và giảm dần theo thời gian sinh trƣởng của bọ phấn. Hiệu quả truyền virus giảm theo tuổi là do
lƣợng virus đƣợc chích nạp giảm.
12.4.3.5 Các begomovirus hại cà chua ở Việt Nam
Việt Nam là một trong các trung tâm đa dạng chính của begomovirus. Tuy nhiên, hiện nay
mới chỉ xác định đƣợc một số begomovirus sau gây bệnh trên cà chua:
Bảng 12-2 Các begomovirus hại cây trồng đƣợc phát hiện cho tới nay tại Việt Nam
TT Virus Khu vực
1 Tomato leaf curl Vietnam virus (ToLCVV) Cả nƣớc
2 Tomato yellow leaf curl Vietnam virus (TYLCVNV) Miền Bắc
3 Tomato yellow leaf curl Kanchanaburi virus (TYLCKaV) Miền Trung, miền Nam
4 Tomato leaf curl Dan Xa virus (ToLCDXV)* Miền Bắc
5 Tomato leaf curl Hainan virus (ToLCHV) Miền Bắc
6 Tomato leaf curl Hanoi virus (ToLCHanV) Miền Bắc
Ghi chú * ToLCDXV mới được ICTV phân loại lại, còn trong công bố ban đầu, virus này đã được các tác
giả phân loại không chính xác là Tomato yellow leaf curl Vietnam virus-DX1.
12.5 CÂU HỎI ÔN TẬP CHƢƠNG 12
1. Đặc điểm chung họ Geminiviridae.
2. Đặc điểm chi Bgomovirus (hình thái, đặc điểm bộ gen, tái sinh, lan truyền).
3. Bệnh xoăn vàng lá cà chua (triệu chứng, bản chất tác nhân gây bệnh, lan truyền (thời
gian chích nạp, chích truyền, ẩn), phƣơng hƣớng phòng trừ).

134
12.6 TÀI LIỆU THAM KHẢO CHÍNH
1. Fauquet, C. M. & Stanley, J. (2005). Revising the way we conceive and name viruses below
the species level: A review of geminivirus taxonomy calls for new standardized isolate
descriptors. Archives of Virology 150, 2151-2179.
2. Ghanim, M., Morin, S., Zeidan, M. & Czosnek, H. (1998). Evidence for transovarial
transmission of tomato yellow leaf curl virus by its vector, the whitefly Bemisia tabaci.
Virology 240, 295-303.
3. Gutierrez, C., Ramirez-Parra, E., Castellano, M. M., Sanz-Burgos, A. P., Luque, A. &
Missich, R. (2004). Geminivirus DNA replication and cell cycle interactions. Veterinary
Microbiology 98, 111-119.
4. Ha, C., Coombs, S., Revill, P., Harding, R., Vu, M. & Dale, J. (2006). Corchorus yellow vein
virus, a New World geminivirus from the Old World. Journal of General Virology 87, 997–
1003.
5. Ha, C., Coombs, S., Revill, P., Harding, R., Vu, M. & Dale, J. (2008). Molecular
characterization of begomoviruses and DNA satellites from Vietnam: additional evidence
that the New World geminiviruses were present in the Old World prior to continental
separation. Journal of General Virology 89, 312–326.
6. Mansoor, S., Briddon, R. W., Zafar, Y. & Stanley, J. (2003). Geminivirus disease complexes:
an emerging threat. TRENDS in Plant Science 8 128-134.
7. Moriones, E. & Navas-Castillo, J. (2000). Tomato yellow leaf curl virus, an emerging virus
complex causing epidemics worldwide. Virus Research 71, 123-134.
8. Nawaz-ul-Rehman, M. S. & Fauquet, C. M. (2009). Evolution of geminiviruses and their
satellites. FEBS Letters 583, 1825–1832.
9. Revill, P. A., Ha, C. V., Porchun, S. C., Vu, M. T. & Dale, J. L. (2003). The complete
nucleotide sequence of two distinct geminiviruses infecting cucurbits in Vietnam. Archives
of Virology 148, 1523-1541.
10. Rojas, M. R., Hagen, C., Lucas, W. J. & Gilbertson, R. L. (2005). Exploiting chinks in the
plant's armor: Evolution and emergence of geminiviruses. Annual Review of Phytopathology
43, 361-394.
11. Seal, S. E., vandenBosch, F. & Jeger, M. J. (2006). Factors influencing begomovirus
evolution and their increasing global significance: Implications for sustainable control.
Critical Reviews in Plant Sciences 25, 23-46.
12. Stanley, J., Bisaro, D. M., Briddon, R. W., Brown, J. K., Fauquet, C. M., Harrison, B. D.,
Rybicki, E. P. & Stenger, D. C. (2005). Geminiviridae. In Virus Taxonomy VIIIth report of
the International Committee on Taxonomy of Viruses, pp. 301 - 326. Edited by C. M.
Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger & L. A. Ball. London:
Elsevier/Academic Press.

135
Chƣơng 13. VIRUS DNA SỢI ĐƠN (TIẾP): CHI BABUVIRUS (HỌ NANOVIRIDAE)
13.1 TÓM TẮT NỘI DUNG
Chƣơng 13 mô tả chi tiết đặc điểm phân loại, phân tử, sinh học của các virus họ Nanoviridae,
một trong 3 họ virus thực vật có bộ gen DNA. Chƣơng này cũng mô tả chi tiết đặc điểm của
BBTV (banana bunchy top virus) gây bệnh chùn ngọn chuối ở Việt Nam.
13.2 HỌ NANOVIRIDAE
13.2.1 Phân loại
Họ Nanoviridae chỉ gồm 6 thành viên hại thực vật đƣợc phân loại vào 2 chi là Nanovirus và
Babuvirus.
Bảng 13-1 Phân loại họ Nanoviridae
Virus Chi Ký chủ
Abaca bunchy top virus Babuvirus Musa
Banana bunchy top virus Babuvirus Musa
Cardamom bushy dwarf virus Babuvirus ?
Faba bean necrotic yellows virus Nanovirus Cây họ đậu
Subterranean clover stunt virus Nanoviru Cây họ đậu
Milk vetch dwarf virus Nanovirus Cây họ đậu
13.2.2 Đặc điểm phân tử virus (virion)
Hình thái. Virion có hình cầu đa diện đối xứng 20 mặt (icosahedral, T=1, 60 tiểu phần
protein), kích thƣớc 17-20 nm, không có vỏ bọc.
Acid nucleic. Bộ gen virus gồm 6 – 8 phân tử DNA sợi vòng đơn (ssDNA) tùy theo chi. Mỗi
virion chỉ chứa một phân tử DNA. Các phân tử DNA genome có kích thƣớc từ 977 - 1111 đến
nucleotid.
Protein. Virion chỉ chứa 1 loại protein là CP.
13.2.3 Tổ chức bộ gen và đặc điểm tái sinh
Các phân tử DNA genome có cấu trúc tƣơng tự nhau gồm một vùng không dịch mã (NCR =
non coding region) và vùng dịch mã. Vùng không dịch mã có trình tự nucleic acid giống nhau
cho tất cả các phân tử DNA genome của loài. Vùng không dịch mã chứa nguồn gốc tái sinh (ori),
trên đó là một cấu trúc thân – thòng lọng (stem-loop structure). Trên vùng dịch mã, mỗi phân tử
DNA genome đều chứa 1 ORF mã hóa cho một protein chức năng. Các protein này đƣợc mã hóa
theo cực (+) tức mã hóa trên sợi virus (chiều kim đồng hồ). Các protein của mỗi virus gồm:
M-Rep (major replication protein): chịu trách nhiệm tái sinh tất cả các phân tử DNA genome.
CP (coat protein): protein vỏ

136
Clink (cell-cycle link protein): protein tham gia quá trình biến đổi chu kỳ tế bào ký chủ theo
hƣớng có lợi cho sự tái sinh của virus.
MP (movement protein): protein vân chuyển.
NSP (nuclear shuttle protein) : protein con thoi tham gia vận chuyển DNA genome của virus
vào và ra khỏi nhân tế bào ký chủ.
Ngoài ra là các protein khác ký hiệu từ U1 – U5 với chức năng chƣa đƣợc xác định.
Các virus thuộc họ Nanoviridae tái sinh trong nhân tế bào theo cơ chế vòng lăn (rolling cicle)
tƣơng tự nhƣ các geminivirus. Ở trong nhân tế bào, phân tử DNA sợi đơn của virus chuyển thành
phân tử DNA trung gian dạng sợi kép. Đồng thời các protein của virus cũng đƣợc dịch mã thành
các protein chức năng, trong đó vai trò quan trọng đối với sự tái sinh của virus là M-Rep và
Clink. Clink tƣơng tác với protein ký chủ để khởi động và tăng cƣờng bộ máy tái bản DNA của
tế bào ký chủ. Trên phân tử DNA trung gian sợi kép, M-Rep sẽ cắt tại vị trí (T-A) của chuỗi
TAT/GTAT-AC trên vùng thòng lọng của cấu trúc thân-thòng lọng và quá trình tái sinh sẽ diễn
ra liên tục theo cơ chế vòng lăn nhờ DNA polymerase của tế bào. M-Rep lại tiếp tục cắt các sợi
DNA genome mới hình thành và nối chúng lại để tạo thành phân tử DNA genome hoàn chỉnh.
Hình 13-1 Tổ chức bộ gen của các virus thuộc họ Nanoviridae (Gronenborn, 2004)
13.2.4 Đặc điểm sinh học
Phạm vi ký chủ. Các virus, nhìn chung, có phạm vi ký chủ hep. Các virus thuộc chi Nanovirus
nhiễm chủ yếu cây họ đậu còn các virus thuộc chi Babuvirus nhƣ BBTV nhiễm trên cây họ
Musaceae (chuối).
Lan truyền. Tất cả các virus không truyền qua tiếp xúc cơ học hoặc qua hạt giống. Trong điều
kiện tự nhiên, virus lan truyền nhờ rệp muội (họ Aphididae) theo kiểu bền vững tuần hoàn. Đối
với BBTV, virus có thể lan truyền dễ dàng qua thân ngầm hoặc qua nuôi cấy mô.

137
13.3 BANANA BUNCHY TOP VIRUS (BBTV)
13.3.1 Tầm quan trọng
Bệnh chùn ngọn do BBTV là bệnh virus quan trọng nhất trên chuối. Bệnh đã từng là nguyên
nhân phá hủy nhiều khu vực trồng chuối tại Öc và một số nƣớc Nam Thái Bình dƣơng nhƣ Fiji.
Tại Việt Nam, bệnh khá phổ biến trên nhóm chuối tiêu nhƣng tầm quan trọng ít đƣợc chú ý.
Bệnh còn đƣợc gọi với các tên nhƣ chuối đực, chuối xẹ, chuối hủi.
13.3.2 Virus
BBTV có hình thái phân tử dạng hình khối đa diện, kích thƣớc 18 – 20 nm. BBTV có bộ gen
phân mảnh gồm ít nhất 6 phân tử DNA sợi vòng đơn, kích thƣớc mỗi phân tử ~ 1000 – 1100
nucleotides. Các đặc điểm bộ gen và tái sinh giống nhƣ các nanovirus khác (xem phần giới thiệu
họ Nanoviridae)
13.3.3 Triệu chứng
Triệu chứng bệnh chùn ngọn chuối rất điển hình và có giá trị chẩn đoán. Các triệu chứng bao
gồm:
Cây sinh trƣởng còi cọc, lùn dữ dội (đặc biệt nếu do truyền qua thân ngầm);
Lá cây bệnh có các sọc xanh đậm dài ngắn khác nhau chạy dọc gân lá. Gần gân chính,
các sọc này thƣờng cong lại dạng móc.
Lá bệnh thƣờng nhỏ, hẹp, dựng đứng; mép lá biến vàng, lƣợn sóng, có thể bị chết hoại.
Không ra buồng. Nếu nhiễm muôn thì vẫn có thể có quả nhƣng chất lƣợng quả kém
Hình 13-2 Bụi chuối bị bệnh chùn ngon (tại Gia Lâm) và triệu chứng trên lá
13.3.4 Lan truyền
BBTV lan truyền ngoài tự nhiên theo 2 cách:
Bằng 1 loài rệp muội là rệp chuối (Pentalonia nigronervosa) theo kiểu bền vững tuần hoàn.
Rệp có thời gian chích nạp 4 – 17 giờ, thời gian chích truyền tối thiểu 30 phút – 2 giờ; thời
gian ẩn 13 – 20 ngày. Hiệu quả truyền virus trong quần thể rệp khoảng 50 %.

138
Qua nhân giống vô tính. BBTV dễ dàng truyền qua thân ngầm từ cây mẹ sang cây con. Thậm
chí nhân giống chuối bằng invitro cũng rất dễ dàng lan truyền virus.
13.3.5 Phòng chống
Các biện pháp phòng chống bệnh có thể đƣợc áp dụng bao gồm:
Nhổ bỏ cây bệnh kết hợp dùng vật liệu giống sạch bệnh. Biện pháp này đã áp dụng thành
công tại Öc.
Áp dụng nghiêm ngặt biện pháp kiểm dịch đối với vật liệu giống giữa các vùng trong và
ngoài nƣớc.
Quản lý vector bằng thuốc hóa học hoặc sinh học.
13.4 CÂU HỎI ÔN TẬP CHƢƠNG 13
1. Đặc điểm chung họ Nanoviridae (hình thái, đặc điểm bộ gen, tái sinh).
2. Bệnh chùn ngọn chuối (triệu chứng, bản chất tác nhân gây bệnh, lan truyền (thời gian chích
nạp, chích truyền, ẩn), phƣơng hƣớng phòng trừ).
13.5 TÀI LIỆU THAM KHẢO CHÍNH
1. CABI (2006). Banana bunchy top virus. Crop Protection Compendium CD ROM
2. Gronenborn, B. (2004)). Nanoviruses: genome organisation and protein function. Veterinary
Microbiology 98 103–109.
3. Timchenko, T., Katul, L., Sano, Y., de-Kouchkovsky, F., Vetten, H. J. & Gronenborn, B.
(2000). The Master Rep Concept in Nanovirus Replication: Identification of Missing
Genome Components and Potential for Natural Genetic Reassortment. Virology 274, 189-
195.
4. Vetten, H. J., Chu, P. W. G., Dale, J. L., Harding, R., Hu, J., Katul, L., Kojima, M., Randles,
J. W., Sano, Y. & Thomas, J. E. (2005). Nanoviridae. In Virus Taxonomy VIIIth report of
the International Committee on Taxonomy of Viruses, pp. 343-352. Edited by C. M. Fauquet,
M. A. Mayo, J. Maniloff, U. Desselberger & L. A. Ball. London: Elsevier/Academic Press.

139
Chƣơng 14. VIRUS RNA SỢI ĐƠN CỰC DƯƠNG: CHI TOBAMOVIRUS (HỌ VIRGAVIRIDAE)
14.1 TÓM TẮT NỘI DUNG
Chƣơng 14 mô tả tóm tắt đặc điểm phân loại, phân tử của họ Virgavirridae cũng nhƣ chi quan
trọng nhất của họ là chi Tobamovirus. Các thành viên của họ là các virus RNA sợi đơn, cực (+).
Chƣơng này cũng mô tả chi tiết đặc điểm của TMV (Tobacco mosaic virus), một virus có ý
nghĩa lịch sử lớn.
14.2 HỌ VIRGAVIRIDAE
Các virus họ Virgaviridae đều là các virus RNA sợi đơn, cực (+) hại thực vật. Hiện tại họ này
đƣợc phân loại thành 6 chi với tổng số 43 loài, trong đó chi Tobamovirus chiếm số lƣợng lớn
nhất với 25 loài.
Bảng 14-1 Phân loại họ Virgaviridae
TT Chi Số phân
đoạn gen Loài điển hình Số loài
1 Furovirus 2 Soil-borne wheat mosaic virus 5
2 Hordeivirus 3 Barley stripe mosaic virus 4
3 Pecluvirus 2 Peanut clump virus 2
4 Pomovirus 3 Potato mop-top virus 4
5 Tobamovirus 1 Tobacco mosaic virus 25
6 Tobravirus 2 Tobacco rattle virus 3
Tổng 43
Các virgavirus đều có dạng hình gậy cứng, không có vỏ bọc, có cấu trúc đối xứng xoắn. Phân
tử virus chỉ có vỏ protein bao quanh lõi RNA genome. Các virgavirus có bộ gen RNA sợi đơn,
cực (+), có mũ Cap đầu 5‘ nhƣng không có đuôi polyA.
Ngoại trừ chi Tobamovirus có bộ gen gồm 1 phân tử RNA thì các chi còn lại đều có bộ gen
phân đoạn gồm 2 – 3 phân tử RNA. Các phân đoạn RNA nếu có đƣợc lắp ráp riêng rẽ.
14.3 CHI TOBAMOVIRUS
Là chi lớn nhất với 25 thành viên. Tobacco mosaic virus (TMV), thành viên điển hình của chi,
là một virus có ý nghĩa rất lớn trong lịch sử virus học và sinh học.
Phân tử virus hình gậy, kích thƣớc 300-310 nm x 18 nm. Các tobamovirus có bộ gen RNA sợi
đơn, cực (+), không phân đoạn, kích thƣớc ~ 6.3-6.5 kb. Trong quá trình tái sinh, các
tobamovirus tạo 2 phân tử phụ geome. Phân tử RNA genome và 2 phân tử phụ genome đều có
đầu 5‘ đƣợc mũ hóa, còn đầu 3‘ có cấu trúc thức cấp dạng tRNA và không có đuôi poly A.
Hình 14-1 Sơ đồ cấu trúc phân tử của các tobamovirus (Viralzone, 2009)

140
14.4 TOBACCO MOSAIC VIRUS (TMV)
14.4.1 Tầm quan trọng
TMV (virus khảm lá thuốc lá) thuộc chi Tobamovirus.
TMV là một virus có ý nghĩa lịch sử lớn trong sinh học và trong ngành virus học.
TMV là một trong các virus thực vật phân bố khắp thế giới, có phổ ký chủ rất rộng, có thể
nhiễm trên 200 loài cây thuộc 30 họ thực vật khác nhau. Các cây ký chủ quan trọng nhất của
TMV thuộc các họ: Solanaceae (họ cà)), Scrophulariaceae, Labiatae, Leguminosae (họ đậu),
Cucurbitaceae (họ bầu bí) và Alliaceae.
Tại Việt Nam, TMV là virus quan trọng nhất trên thuốc lá.
14.4.2 Virus
14.4.2.1 Hình thái phân tử
Phân tử TMV có hình gậy, kích thƣớc ~300 x 18 nm. Phân tử virus gồm 1 phân tử RNA
genome kích thƣớc ~ 6.4 kb đƣợc bao bọc bởi một lớp vỏ gồm ~ 2300 tiểu phần protein (mỗi
tiểu phần là một phân tử protein có kích thƣớc 158 amino acid (17.3 kD).
Giống nhƣ các virus hình gậy khác, phân tử TMV có cấu trúc đối xứng xoắn nhƣ sau:
Ba nucleotide trên phân tử RNA liên kết với 1 tiểu phần protein
Một đơn vị lặp hoàn hảo gồm 49 tiểu phần protein / 3 vòng xoắn và tạo ra độ dài 6.9 nm
(tổng độ dài phân tử ~ 300 nm)
Đƣờng kính phân tử là 18 nm còn đƣờng kính lõi rỗng là 4 nm
Cần chú ý là TMV có thể tự lắp ráp thành phân tử hoàn chỉnh từ 1 hỗn hợp các tiểu phần
protein và bộ gen RNA trong ống nghiệm.
Hình 14-2 Hình thái virion và cấu trúc của TMV
14.4.2.2 Đặc điểm vật lý
TMV có ngƣỡng pha loãng giới hạn là 10-6
- 10-7
, khả năng tồn tại trong chiết dịch cây bệnh
của virus là khoảng 1 tháng, nhiệt độ mất hoạt tính Q10 là 930C. Virus có khả năng tích luỹ trong

141
dịch cây bệnh với nồng độ rất cao (2 g/l). Virus rất mẫn cảm với tia Rơnghen và dễ dàng bị phân
huỷ ở ngƣỡng pH < 1 và pH > 11.
Trong tế bào cây ký chủ, đặc biệt là tế bào lông lá, TMA có thể hình thành nên thể vùi dạng
tinh thể lục giác cấu tạo bởi các phân tử virus xếp thành hàng, thành phiến.
Là virus thực vật bền nhất: virus tinh chiết (bảo quản ở 40OC) vẫn duy trì khả năng nhiễm
bệnh sau 50 năm; virus tồn tại lâu dài trong tàn dƣ và thuốc lá chế biến; virus trong mô giữ đƣợc
hoạt tính gây bệnh sau 30 phút sấy ở nhiệt độ 120OC.
Hình 14-3 Tinh thể virus TMV hình thành trong tế bào thuốc lá
14.4.3 Tổ chức bộ gen và tái sinh
TMV và các tobamovirus khác có bộ gen RNA sợi đơn, cực +, không phân đoạn, kích thƣớc ~
6.4 kb.
Tổ chức bộ gen của TMV và các tobamovirus khác tƣơng tự nhau. Tính từ trái qua phải, bộ
gen của TMV đƣợc tố chức nhƣ sau:
1. Một chuỗi không dịch mã đầu 5‘ (untranslated leader).
2. Bốn ORF là:
ORF1 mã hóa protein MET-Hel (126 kDa) có chức năng tạo mũ (methyltransferase) đầu
5‘ phân tử RNA và tháo xoắn (helicase) trong quá trình tái sinh virus.
ORF2 là 1 ORF lồng của ORF1. ORF 1 cùng với ORF2 đƣợc dịch mã theo cơ chế
―readthrough‖ để tạo 1 protein lớn là RdRp (183 kDa)
ORF3 mã hóa protein vận chuyển MP (30 kDa)
ORF4 mã hóa CP (17.5 kDa)
3. Một chuỗi không dịch mã đầu 3‘ có cấu trúc thứ cấp giống tRNA.

142
Trong quá trình tái sinh, TMV tạo 2 phân tử RNA phụ genome có chung đầu 3‘ với phân tử
RNA genome nhƣng có kích thƣớc ngắn hơn, tƣơng ứng gen MP và CP.
Phân tử RNA genome và 2 phân tử phụ genome đều đƣợc mũ hóa m7G5'ppp5'Gp (Cap) ở đầu
5‘ nhƣng không có đuôi polyA.
Hình 14-4 Tổ chức bộ gen chính và 2 phân tử phụ genome của TMV
Quá trình tái sinh của TMV đã đƣợc nghiên cứu kỹ và trải qua các bƣớc sau:
1. Phân tử virus xâm nhập vào tế bào chất nhờ tổn thƣơng cơ học.
2. Cởi áo một phần protein vỏ ở đầu 5‘ của phân tử virus.
3. Ribosome của tế bào ký chủ tiếp cận đầu 5‘ của RNA genome và tiến hành dịch mã 2
protein tái sinh là Met-Hel và RdRp. Trong quá trình dịch mã thì quá trình cởi áo cũng
đƣợc tiếp tục. Đây là cơ chế ―cởi áo cùng dịch mã‖ (co-traslational disassembly)
4. Hai protein MET-Hel và RdRp sử dụng sợi RNA genome (sợi +) để sao mã sợi RNA (-).
5. Tiếp theo, sợi RNA (-) này lại đƣợc dùng làm khuôn để tổng hợp sợi (+) và các phân tử
RNA phụ genome (có thể xem là các mRNA của protein MP và CP). Trên các phân tử
phụ genome này, các protein MP và CP đƣợc dịch mã.
6. MP kết hợp với sợi RNA (+) để di chuyển sang tế bào khác qua sợi liên bào theo cơ chế
nucleoprotein.
7. Các phân tử RNA sợi (+) và CP khi tích lũy đủ kích hoạt quá trình lắp ráp phân tử virus
mới.
8. Các phân tử virus cũng kết tụ lại để tạo các thể vùi (tinh thể) dạng lục giác trong tế bào
chất.

143
14.4.4 Triệu chứng
Triệu chứng của TMV có thể thay đổi theo chủng virus, cây ký chủ.
Trên cây thuốc lá, cây bệnh nhiễm hệ thống với triệu chứng điển hình là lá bị khảm rõ rệt
giống nhƣ da ếch. Triệu chứng đầu tiên xuất hiện trên lá non gồm các vùng xanh đậm, xanh nhạt,
vàng xen kẽ nhau. Lá phát triển kém, phiến lá nhỏ hẹp, mặt lá gồ ghề. Cây nhỏ chỉ bằng 1/2 - 1/4
lần so với cây khoẻ.
Trên cây ớt, virus TMV nhiễm hệ thống gây hiện tƣợng khảm lá, lùn cây. Đôi khi cây bệnh
xuất hiện các vết chết hoại ở phần cuống lá.
Trên cây cà chua: virus TMV gây hiện tƣợng đốm biến vàng, khảm lá; lá ít biến dạng hơn.
Hình 14-5 Triệu chứng do TMV gây ra trên thuốc lá (trái), ớt (giữa) và cà chua (phải)
14.4.5 Lan truyền
TMV là một trong các virus có khả năng lan truyền rất cao
Virus xâm nhập và lan truyền rất dễ dàng qua tiếp xúc cơ học (cọ xát lá, cắt tỉa, tiếp xúc của
rễ trong đất). Virus có thể lan truyền qua côn trùng miệng nhai (theo kiểu cơ học).
Thời kì tiềm dục của bệnh phụ thuộc vào điều kiện nhiệt độ. Ở nhiệt độ 300C là ~5 ngày, ở
nhiệt độ thấp hơn là 2 tháng, trung bình là 8 - 10 ngày.
TMV là một loại virus truyền qua hạt của nhiều loại cây (ví dụ hạt cà chua, thuốc lá). Tuy
nhiên, virus không tồn tại bên trong hạt giống mà bám ở bề mặt vỏ hạt.
TMV (và các tobamovirus khác) là virus thực vật cực kỳ bền vững (có thể đƣợc xem là ngoại
lệ duy nhất trong số các virus thực vật). TMVcó thể tồn tại rất lâu dài (hàng tháng tới hàng năm)
trên tàn dƣ cây bệnh trong đất. Thậm chí thuốc lá đã qua chế biến vẫn có thể truyền bệnh (virus
giữ đƣợc hoạt tính gây bệnh sau 30 phút sấy ở nhiệt độ 1200C).
14.4.6 Biện pháp phòng trừ
1. Sử dụng giống chống bệnh dựa trên tính kháng gen đối gen (nhƣ gen N trên thuốc lá, gen L
trên ớt), dựa trên tính kháng từ tác nhân gây bệnh PDR (ví dụ cây thuốc lá chuyển gen CP có
thể kháng rất tốt virus).

144
2. Sử dụng giống sạch bệnh: Chọn giống từ những ruộng không bị nhiễm bệnh; thu hoạch riêng
cây bệnh, cây khoẻ.
3. Sử lý hạt giống. Vì TMV có thể tồn tại ở vỏ hạt nên có thể hạn chế virus bằng xử lí hạt giống
với hóa chất. Ví dụ ngâm hạt trong dung dịch Na3PO4 10% trong 2 giờ; hoặc ngâm hạt trong
dung dịch Na3PO4 1% trong 15 phút, sau đó ngâm trong dung dịch NaOCl 0.53% trong 30
phút. Xử lý hạt với dung dịch HCl 20% cũng cho kết quả tốt.
4. Khử trùng dụng cụ thu hái bằng Formalin 4 %, rửa tay và dụng cụ bằng xà phòng, sữa.
5. Nhổ bỏ cây bệnh trên đồng ruộng, dọn sạch tàn dƣ cây bệnh và diệt côn trùng môi giới là sâu
ăn lá (truyền kiểu cơ giới).
14.5 CÂU HỎI ÔN TẬP CHƢƠNG 14
1. Đặc điểm (hình thái, tổ chức bộ gen) của họ Tobamoviridae.
2. TMV (hình thái, tổ chức bộ gen và tái sinh, triệu chứng, lan truyền vầ phòng trừ).
14.6 TÀI LIỆU THAM KHẢO CHÍNH
1. CABI (2006). Tobacco mosaic virus. Crop Protection Compendium CD ROM
2. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
3. Lewandowski, D. J. (2005). Tobamovirus. In Virus Taxonomy VIIIth report of the
International Committee on Taxonomy of Viruses, pp. 1009-1014. Edited by C. M. Fauquet,
M. A. Mayo, J. Maniloff, U. Desselberger & L. A. Ball. London: Elsevier/Academic Press.
4. Scholthof, K.-B. G. (2004). TOBACCOMOSAIC VIRUS: AModel System for Plant
Biology. Annual Review of Phytopathology 42, 13–34.

145
Chƣơng 15. VIRUS RNA SỢI ĐƠN CỰC DƯƠNG (TIẾP): CHI POTYVIRUS (HỌ POTYVIRIDAE)
15.1 TÓM TẮT NỘI DUNG
Chƣơng 15 mô tả tóm tắt đặc điểm phân loại, phân tử, sinh học của họ Potyviridae, một trong
hai họ virus thực vật lớn nhất. Chƣơng này cũng mô tả chi tiết đặc điểm phân loại, sinh học phân
tử, đặc biệt là vai trò của các protein, của chi Potyvirus - là chi virus thực vật có ý nghĩa kinh tế
bậc nhất trên thế giới và cũng là chi có số lƣợng virus lớn nhất trong họ. Tất cả các potyvirus
đƣợc phát hiện tới nay ở Việt Nam đều đƣợc đề cập với mức độ chi tiết tùy thuộc thông tin sẵn
có.
15.2 HỌ POTYVIRIDAE
Họ Potyviridae (tên có nguồn gốc từ Potato virus Y) là họ virus thực vật lớn nhất với trên
200 loài (Bảng 15-1). Tất cả thành viên trong học đều có bộ gen RNA sợi đơn, cực dƣơng, có
phân tử hình sợi mềm đƣờng kính 11-15 nm và dài 650 – 950 nm đối với nhóm có bộ gen đơn và
200-300 + 500-600 nm đối với các virus có bộ gen kép.
Hiện nay, các potyvirus đƣợc phân loại thành 6 chi trên cơ sở (i) tổ chức bộ gen, (ii) vector
lan truyền và (iii), so sánh trình tự. Đặc điểm bộ gen, mối quan hệ vector và số loài của các chi
đƣợc trình bày ở Bảng 15-1. Trong số 6 chi, chi Potyvirus là quan trọng nhất, cả về số lƣợng loài
cũng nhƣ các bệnh có ý nghĩa kinh tế mà chúng gây ra.
Các virus của 5 chi Potyvirus, Macluravirus, Ipomovirus, Rymovirus và Tritimovirus có bộ
gen không phân đoạn tức bộ gen chỉ gồm 1 phân tử RNA duy nhất nên gọi là virus có bộ gen
đơn. Các virus của chi còn lại, chi Bymovirus, có bộ gen kép gồm 2 phân tử RNA-1 và RNA-2.
Đối với các bymovirus, phân tử RNA-1 tƣơng đƣơng với 2/3 kích thƣớc tính từ đầu 3‘ còn phân
tử RNA-2 tƣơng đƣơng phần còn lại của bộ gen của các virus có bộ gen đơn.
Bảng 15-1. Phân loại họ Potyviridae (Berger et al. 2005)
Chi Loài chuẩn Bộ gen Vector Số loài
1. Potyvirus Potato virus Y (PVY) Bộ gen đơn Rệp (không bền vững) 197
2. Ipomovirus Sweet potato mild mottle virus
(SPMMV)
Bộ gen đơn Bọ phấn (không bền vững) 4
3. Macluravirus Maclura mosaic virus (MacMV) Bộ gen đơn Rệp (không bền vững) 4
4. Rymovirus Ryegrass mosaic virus (RGMV) Bộ gen đơn Nhện (bền vững tuần hoàn) 3
5. Tritimovirus Wheat streak mosaic virus
(WSMV)
Bộ gen đơn Nhện (bền vững tuần hoàn) 4
6. Bymovirus Barley yellow mosaic virus (BaYMV)
Bộ gen kép Nấm 6
Tổng 218

146
15.3 CHI POTYVIRUS
15.3.1 Giới thiệu
Chi Potyvirus là chi lớn nhất trong họ Potyviridae với khoảng 200 loài. Nhƣ vậy, cùng với
chi Begomovirus (họ Geminivirisdae), Potyirus là chi virus thực vật lớn nhất, chiếm khoảng 20%
tổng số virus thực vật.
Nhiều potyvirus là các tác nhân gây bệnh cây có ý nghĩa kinh tế. Ví dụ:
Papaya ringspot virus (PRSV) gây bệnh virus quan trọng nhất trên đu đủ và bầu bí.
Turnip mosaic virus (TuMV) hại cây rau họ thập tự và đƣợc xem là virus quan trọng thứ 2 trên cây rau khắp thế giới.
Bean common mosaic virus (BCMV) trên cây họ đậu.
Potato virus Y (PVY) trên cây khoai tây và các cây họ cà khác.
Plum pox virus (PPV) là virus qua trọng nhất trên cây quả hạch nhƣ đào, táo, lê, mận.
Trong tế bào cây, potyvirus thƣờng tạo các loại thể vùi đặc trƣng. Tất cả các potyvirus đều
hình thành thể vùi hình trụ trong tế bào chất. Thể vùi tế bào chất hình thành từ protein CI
(cytoplasmic inclusions (CI)). Các thể vùi CI này có hình dạng đặc trƣng (hình vòng bánh xe,
hình phiến…) cho potyvirus và do đó có giá trị chẩn đoán. Một số virus lại hình thành thể vùi
trong nhân gọi là thể vùi nhân (nuclear inclusion, NI). Thể vùi nhân đƣợc cấu tạo bởi 2 protein là
NIb và NIa. Một số virus cũng hình thành thể vùi trong nhân nhƣng hình dạng thể vùi không xác
định gọi là thể vùi vô định hình (amorphor inclusions, AI) đƣợc cấu tạo bởi protein HcPro.
Phần lớn virus trong họ có phổ ký chủ hẹp hoặc rất hẹp nhƣng một số ít lại có phổ ký chủ rất
rộng, nhiễm cả trên cây 1 và 2 lá mầm
Hình 15-1 Hình thái virion và thể vùi của potyvirus. Ảnh trái: phân tử PVY, bar = 100 nm (Shukla
et al., 1998). Hình phải: thể vùi tế bào chất của PVY trong tế bào thuốc lá (Arbatova et al. 1998).
15.3.2 Đặc điểm virion
Các potyvirus có phân tử hình sợi mềm đƣờng kính 11-15 nm và dài 650 – 950 nm. Mỗi
phân tử virion đƣợc cấu tạo từ 1700-2000 tiểu phần protein xắp xếp theo kiểu xoắn đối xứng
xung quanh phân tử RNA genome. Ngoài ra, phân tử virus còn có 1 protein gắn ở đầu 5‘ là VPg
(genome-linked protein, VPg). Thành phần phân tử bao gồm 95 % protein và 5% RNA.
A B

147
15.3.3 Tổ chức bộ gen
Tất cả các potyvirus có bộ gen là một phân tử RNA sợi đơn, cực dƣơng, kích thƣớc ~ 10 kb.
Tổ chức bộ gen của các potyvirus giống nhau, tính từ trái sang phải là:
Một vùng 5‘ không dịch mã (5‘UTR)
Một ORF đơn, lớn gọi là ORF chính (major ORF).
Một đầu 3‘ không dịch mã (3‘UTR), kết thúc bằng 1 đuôi poly-A.
Hình 15-2 Tổ chức bộ gen của các potyvirus (Shukla et al., 1998).
ORF chính mã hóa 1 polyprotein, sau khi dịch mã sẽ đƣợc xử lý thành khoảng 10 protein
chức năng
1. Protein P1
2. Protein bổ trợ (Helper component protein, HC-Pro)
3. Protein P3,
4. Protein 6K1,
5. Protein thể vùi tế bào chất (CI)
6. Protein 6K2
7. Protein VPg (genome-linked protein)
8. Protein NIa-Pro (major protease of small nuclear inclusion protein -NIa)
9. Protein NIb (large nuclear inclusion protein)
10. Protei vỏ (coat protein, CP)
Đối với các virus thuộc chi Bymovirus thì RNA-1 mã hóa các protein tƣơng tự P3, 6K1, CI,
6K2, NIa, NIb và CP, còn RNA-2 mã hóa 2 protein P1 và P2, trong đó P1 tƣơng tự HC-Pr còn
P2 tƣơng tự protein ―read through― của các furovirus và cần thiết để virus có thể truyền đƣợc qua
nấm.
15.3.4 Chức năng các protein
Chức năng của các protein của potyvirus đã đƣợc nghiên cứu khá kỹ. Hầu hết các protein của
potyvirus đều là các protein đa chức năng.
a. P1
1. P1 là 1 proteinase. P1 là vùng biến động nhất của bộ gen. P1 có hoạt tính protenase và sẽ tự
cắt liên kết P1/HC-Pro sau dịch mã để giải phóng P1.

148
2. P1 tương tác với RNA. P1 có khả năng liên kết với RNA của virus, do vậy nó có vai trò trong
quá trình vận chuyển virus
3. P1 tham gia ức chế phản ứng phòng thủ của tế bào ký chủ (thông qua cơ chế câm gen).
b. HC-Pro
1. HC-Pro cần thiết cho sự lan truyền qua vector. Vai trò này của protein Hc-Pro đƣợc thể hiện
qua cơ chế bắc cầu. HcPro đóng vai trò là 1 phân tử cầu nối, trong đó phần đầu N của nó
(thông qua motif bảo thủ cao KITC) sẽ liên kết với receptor ở bề mặt phía trong của vòi
aphid còn phần trung tâm của nó (thông qua motif PTK) sẽ liên kết với protein vỏ CP của
virion. Nhờ cầu nối này mà phân tử virus đƣợc giữ lại ở phần vòi chích của vector.
2. HC-Pro cần cho virus di chuyển hệ thống và di chuyển giữa các tế bào. HC-Pro là một
protein vận chuyển (MP) của potyvirus. HC-Pro làm tăng giới han ngăn chặn kích thƣớc SEL
(size exclusion limit) của sợi liên bào và do đó cho phép RNA của virus có thể di chuyển gữa
các tế bào.
3. HC-Pro có hoạt tính proteinase. Đầu C của HC-Pro có hoạt tính proteinase, cho phép nó tự
cắt khỏi liên kết giữa HC-Pro và P3.
4. HC-Pro cần cho qua tái bản (thông qua motif IGN).
5. HC-Pro có thể ức chế bộ máy câm gen của tế bào. Đây là một chức năng quan trọng của
potyvirus chống lại phản ứng phòng thủ của cây.
c. P3
P3 cũng là vùng biến động và ít đƣợc nghiên cứu chức năng. P3 đƣợc biết là qui định tính gây
bệnh của virus thông qua tƣơng tác với các protein khác của virus.
d. CI
1. CI là thành phần chính của phức hợp tái sinh virus. CI có hoạt tính helicase, hoạt tính liên
kết RNA, hoạt tính thủy phân ATP. Tất cả các hoạt tính này đều cần thiết cho CI tháo xoắn
cấu trúc trung gian sợi kép của RNA virus trong quá trình tái sinh.
2. CI cần cho sự di chuyển của virus gữa các tế bào . Quá trình di chuyển giữa các tế bào yêu
cầu năng lƣợng. Do CI có hoạt tính ATPase nên nó đảm nhiệm vai trò cung cấp năng lƣợng
cho quá trình vận chuyển virus qua các tế bào. Ngoài ra, CI tƣơng tác trực tiếp với
plasmodesmata và phức hợp ribonucleoprotein của virus để tạo điều kiện thuận lợi cho quá
trình di chuyển.
e. 6K1 và 6K2
6K1 tƣơng tác với P3 để qui định tính gây bệnh của virus. 6K2 là một protein mỏ neo giúp
liên kết phức hợp tái sinh virus vào mạng lƣới nội chất – nơi thực hiện quá trình tái sinh.
f. VPg
1. VPg chính là phần đầu N của NIa. VPg cũng là 1 protein cấu trúc vì ngoài protein CP, nó có
mặt trong phân tử virus. VPg lien kết vào đầu 5‘ của RNA của virus và bảo vệ phân tử RNA
khỏi bị phân hủy
2. VPg cần cho quá trình tái sinh bộ gen virus, di chuyển hệ thống của virus trong cây.
3. VPg tương tác với yếu tố khởi đầu dịch mã của tế bào nhƣ eIF4E và eIF(iso)4E, do đó cho
phép ribosome có thể tiếp cận RNA virus ngay sau khi tháo vỏ để thực hiện quá trình dịch

149
mã. Cần chú ý là potyvirus có bộ gen RNA sợi đơn cực (+), phân tử RNA genome của virus
không đƣợc mũ hóa nên quá trình dịch mã ORF trên bộ gen virus là một quá trình dịch mã
độc lập mũ Cap.
4. VPg là một protein Avr của potyvirus. Nhiều gen kháng của thực vật đối với potyvirus đã
đƣợc chứng minh là các gen lặn ví dụ nhƣ pvr1 (ớt), mo1 (rau diếp), sbm1 (đâu) và rym4/5
(lúa miến). Các gen này thực chất mã hóa yếu tố khởi đầu dịch mã eIF4E. Khi các gen này (ở
trạng thái đồng hợp tử) mang các đột biến phá vỡ tƣơng tác VPg – eIF4E sẽ dẫn tới kiểu hình
kháng.
g. NIa-Pro
NIa-Pro là phần đầu C của protein NIa. Nó là một protease chính của virus. Đầu C của NIa-
Pro có hoạt tính protease cho phép nó cắt các liên kết P3/6K1, 6K1/CI, CI/6K2, 6K2/VPg,
VPg/NIa-Pro, NIa-Pro/NIb và NIb/CP để giải phóng ra các protein chức năng đơn lẻ.
h. NIb
NIb là một RNA dependent RNA polymerase (RdRp). Đây là một chức năng quan trọng cho
phép bộ gen virus đƣợc tái bản từ sợi (+) sang sợi (-) và ngƣợc lại.
i. CP
1. CP của potyvirus đã đƣợc nghiên cứu rất kỹ. CP có thể đƣợc chia làm 3 phần (đầu N, trung
tâm và đầu C). Đầu N là phần nhô lên bề mặt phân tử virus, khá biến động về trình tự và
chứa các epitope đặc hiệu cho virus ở mức loài và mức chủng. Phần trung tâm và đầu C bảo
thủ hơn và mức biến động của 2 phần này tƣơng đƣơng mức biến động chung trên toàn bộ bộ
gen. Chính vì vậy, CP, đặc biệt phần đầu C và trung tâm thƣờng hay đƣợc sử dụng trong
đánh giá đa dạng virus.
2. CP có vai trò quan trọng trong lan truyền qua vector. Đầu N của CP chứa một motif bảo thủ
cao (DAG). Thông qua motif này, phân tử virus sẽ liên kết với Hc-Pro theo lý thuyết bắc cầu
để lan truyền qua vector rệp muội.
3. CP có vai trò quan trọng trong di chuyển hệ thống và di chuyển giữa các tế bào. CP (cùng
với Hc-Pro) là 2 protein vận chuyển (MP) của potyvirus. Cả 2 protein này đều làm tăng giới
hạn ngăn chặn kích thƣớc SEL của sợi liên bào và do đó cho phép virus di chuyển giữa các tế
bào.
4. CP là một protei cấu trúc tạo virion.
5. CP điều khiển qua trình tái sinh RNA virus
15.3.5 Tái sinh của potyvirus
Quá trình tái sinh của potyvirus (và các virus thực vật có bộ gen RNA) diễn ra trong tế bào
chất. Ngay sau khi xâm nhập vào tế bào chất của tế bào ký chủ (nhờ vector hoặc tiếp xúc cơ
học), quá trình tái sinh của potyvirus diễn ra theo các bƣớc sau:
1. Trong tế bào chất, phân tử virus tháo vỏ, giải phóng bộ gen virus (RNA sợi +).
2. Bộ máy dịch mã của tế bào sẽ dịch mã tạo polyprotein ngay trên khuôn RNA genome của
virus (RNA sợi +). Tiếp theo, polyprotein này đƣợc xử lý hậu dịch mã nhờ chính các vùng có
hoạt tính protease của polyprotein để tạo thành các protein chức năng
3. Các protein của virus và của tế bào ký chủ thực hiện quá trình tái bản. Quá trình tái bản yêu
cầu 2 protein chủ chốt của virus là RdRp (NIb) và helicase (CI). Các protein tham gia tái bản

150
cùng với RNA sợi (+) lắp ráp thành ―phức hợp tái sinh virus‖ ký hiệu là VRC (viral
replication complex) gắn trên màng lƣới nội chất. Trong quá trình này còn có sự tham gia
của một số protein khác nhƣ protein mỏ neo 6K2 của virus. Tại VRC, sợi RNA (-) sẽ đƣợc
tổng hợp trên sợi RNA (+).
4. Sợi (-) mới đƣợc tổng hợp ra lại dùng làm khuôn để tổng hợp nhiều sợi (+) mới.
5. Lắp ráp thành virion mới từ protein CP và sợi (+) đƣợc tổng hợp mới.
15.4 PAPAYA RING SPOT VIRUS (PRSV)
15.4.1 Tầm quan trọng
PRSV (virus đốm hình nhẫn đu đủ), thuộc chi Potyvirus, là virus nguy hiểm nhất, gây bệnh
đốm hình nhẫn trên đu đủ khắp thế giới. Tại Việt Nam, chỉ sau trồng khoảng 3-4 tháng, tỷ lệ
bệnh trên đu đủ đã rất cao. Không hiếm gặp các vƣờn đu đủ bị nhiễm 100% sau vài tháng trồng.
PRSV cũng là virus phổ biến trên bầu bí khắp thế giới. Tại Việt Nam, các nghiên cứu dựa vào
RT-PCR và ELISA cho thấy PRSV cũng là virus phổ biến nhất, gây bệnh khảm lá, trên cây bầu
bí. Các cây bị nhiễm bao gồm: bí ngô, dƣa chuột, bầu, bí xanh, các loại dƣa…
15.4.2 Virus
PRSV có hình thái và tổ chức bộ gen điển hình của 1 potyvirus. Phân tử virus hình sợi mềm,
dài 760-800 nm, đƣờng kính 12 nm.
Virus có 2 type: type P nhiễm trên đu đủ và type W nhiễm trên cây họ bầu bí. Hiện vẫn chƣa
có số liệu chứng minh liệu ở Việt Nam 2 type này có nhiễm chéo sang nhau không. Nghiên cứu
đa dạng dựa trên trình tự gen mã hóa vỏ protein của virus từ các mẫu thu thập khắp cả nƣớc năm
2003 cho thấy (i) không có sự khác biệt về trình tự giữa 2 type P và W ở Việt Nam (ii) có sự
khác biệt theo vùng địa lý, có nghĩa quần thể PRSV ở miền Bắc tách biệt với quần thể PRSV ở
miền Nam và (iii) đã có đột biến từ type W thành type P.
15.4.3 Triệu chứng
15.4.3.1 Trên đu đủ
1. Trên quả, virus gây đốm hình nhẫn và biến dạng quả. Các đốm lúc đầu trong ƣớt sau bị chết
hoại màu trắng bạc – trắng xám. Quả bị biến dạng, chất lƣợng giảm.
2. Trên cuống lá và thân có các sọc trong ƣớt, về sau cũng trở lên chết hoại
3. Trên lá, các triệu chứng là khảm lá, đốm biến vàng, biến dạng (lá co quắp, mất thùy, chỉ còn
trơ gân). Trên lá cũng có thể có các nốt phồng xanh đậm. Triệu chứng thƣờng dữ dội trên lá
non.
4. Cây bệnh thƣờng thấp và còi cọc.
5. Mùa đông, triệu chứng biểu hiện nặng hơn mùa hè.
15.4.3.2 Trên bầu bí
Trên cây bầu bí, triệu chứng khó đánh giá chính xác vì cây có thể bị nhiễm các virus khác.
Nhìn chung, trên cây bầu bí, PRSV (và các potyvirus) tạo triệu chứng khảm và biến dạng trên lá,
cây còi cọc, quả bị biến dạng

151
Hình 15-3 Triệu chứng bệnh trên đu đủ (CABI, 2006) và bầu bí (cây bí xanh tại Việt Nam)
15.4.4 Lan truyền
PRSV không truyền qua hạt giống đu đủ cũng nhƣ bầu bí. Virus truyền đƣợc bằng tiếp xúc cơ
học nhƣng đây không phải là con đƣờng lan truyền ngoài tự nhiên của virus.
Ngoài tự nhiên, virus lan truyền qua rệp muội theo kiểu không bền vững. Trên 20 loài rệp
muội có khả năng truyền virus.
15.4.5 Chẩn đoán
Trên đu đủ, PRSV có thể đƣợc chẩn đoán khá chính xác dựa theo triệu chứng. Tuy nhiên, trên
bầu bí, không thể chẩn đoán đƣợc bệnh vì nhiều virus bầu bí, đặc biệt là các potyvirus nhƣ
ZYMV, cũng tạo triệu chứng tƣơng tự.
Ngoài ra, PRSV cũng có thể đƣợc chẩn đoán bằng ELISA (virus có hoạt tính miễn dịch tốt)
và RT-PCR.
15.4.6 Phòng chống bệnh trên đu đủ
1. Trồng cây ở khu vực biệt lập và nhổ bỏ cây bệnh. Biện pháp đã áp dụng thành công ở một số
vùng thuộc Brazil. Chƣơng trình phòng chống kéo dài hơn 10 năm ở bang Espirito Santo
(Brazil) đã làm giảm áp lực bệnh trên đồng ruộng. Do hiệu quả của biện pháp, chính quyền
bang đã thông qua luật bắt buộc các chủ vƣờn phải nhổ bổ cây bệnh.
2. Trồng cây trong nhà lưới. Để ngăn chặn triệt để vector rệp, kỹ thuật này đã đƣợc áp dụng
thành công ở Đài Loan. Tuy nhiên hiệu quả kinh tế không cho phép áp dụng rộng rãi kỹ thuật
này.
3. Dùng chế phẩm EM. Sử dụng chế phẩm EM (Effective microorganisms), một hỗn hợp các vi
sinh vật có ích gồm vi khuẩn lên men lactic, nấm men, actinomycetes và các vi khuẩn quang
dƣỡng, đã làm giảm tỷ lệ bệnh ở Đài Loan. Cơ chế phòng chống có lẽ liên quan đến tính
kháng tạo đƣợc (Tsai, 1998).

152
4. Dùng giống kháng bệnh/chịu bệnh. Cây đu đủ trồng (Carica papaya) không có gen kháng
virus. Một số loài đu đủ dại có gen kháng nhƣng rất khó để chuyển đƣợc các gen kháng này
vào đu đủ trồng thông qua lai tạo. Trong những năm 80, các nhà khoa học đã tạo ra một số
giống chịu bệnh nhƣ giống Cariflora ở Mỹ, giống lai Tainung 5 ở Đài Loan. Các giống này
có chất lƣợng tốt và chịu đƣợc bệnh.
5. Bảo vệ chéo bằng chủng yếu. Để tạo khả năng bảo vện chéo, một chủng virus yếu (tạo triệu
chứng nhẹ) sẽ đƣợc lây nhiễm trƣớc trên cây để bảo vệ cây chống các chủng mạnh tại địa
phƣơng. Trong những năm 80, 90, các nhà khoa học Mỹ và Đài Loan đã tạo ra một chủng
yếu là HA 5-1 bằng cách gây đột biến 1 chủng mạnh của Hawai. Biện pháp đã đƣợc áp dụng
tại Đài Loan trong gần 10 năm tuy nhiên hiện nay ít đƣợc sử dụng do có quá nhiều trở ngại
(chi phí lớn, khả năng xuất hiện lại chủng mạnh, chủng yếu không thể bảo vệ cây trong điều
kiện lạnh, chủng yếu chỉ bảo vệ cây chống chủng mạnh có cùng nguồn gốc). Tại Hawai, mặc
dù chủng HA 5-1 bảo vệ cây rất tốt chống các chủng mạnh địa phƣơng (vì cùng nguồn gốc)
nhƣng biện pháp vẫn không thể áp dụng do nông dân không muốn nhiễm virus trên cây khỏe.
6. Sử dụng cây chuyển gen thông qua tính kháng PDR. Trong những năm 80, 90, các nhà khoa
học Mỹ (đại học Cornell) đã tạo thành công cây đu đủ chuyển gen mang gen CP của PRSV.
Hai giống đu đủ chuyển gen (Rainbow và Sun Up) có khả năng kháng rất tốt PRSV và đã
đƣợc thƣơng mại rộng rãi tại Mỹ. Kỹ thuật này đã và đang đƣợc áp dụng tại nhiều nƣớc nhƣ
Đài Loan, Thái Lan, Brasil, Mexico…. Hiện nay, dùng đu đủ chuyển gen mang gen virus là
biện pháp hiệu quả nhất để chống PRSV.
15.5 POTATO VIRUS Y (PVY)
15.5.1 Tầm quan trọng
PVY là một virus thuộc chi Potyvirus, đƣợc ghi nhận đầu tiên trên khoai tây từ 1931 và là một
trong các virus thực vật đƣợc nghiên cứu nhiều và đầy đủ nhất. PVY có phổ ký chủ rộng, đặc
biệt nguy hiểm trên cây họ cà. PVY đƣợc xem là virus quan trọng nhất trên khoai tây và trong
nhiều trƣờng hợp trên thuốc lá, ớt và cà chua.
15.5.2 Virus
PVY là loài chuẩn của chi Potyvirus, và giống nhƣ các potyvirus khác, có phân tử hình sợi
mềm, kích thƣớc 730-740 x 11-12 nm với ~ 6 % nucleic acid.
PVY là một trong các virus rất đa dạng về di truyền và tính gây bệnh. Về mặt lịch sử, dựa trên
phản ứng đối với cây ký chủ và chỉ thị (Nicotiana tabacum cv Samsun, Solanum tuberosum và
Physalis floridana), PVY đƣợc chia thành 3 chủng chính là PVYO, PVY
N và PVY
C .
1. Chủng PVYO xuất hiện khắp thế giới, tạo phản ứng siêu nhạy trên cây khoai tây mang gen
kháng Ny.
2. Chủng PVYC phân bố ở Öc, Ấn Độ và châu Âu, tạo phản ứng siêu nhạy trên cây khoai tây
mang gen kháng Nc.
3. Chủng PVYN
phân bố ở châu Âu, châu Phi và Nam Mỹ, gây chết hoại gân trên thuốc lá.
Gần đây, một số chủng mới gần gũi với chủng PVYN cũng đƣợc mô tả. Chẳng hạn, chủng
PVYN-Wilga có nguồn gốc từ chủng PVY
N phân lập từ khoai tây Ba Lan nhƣng tạo triệu
chứng nhẹ hơn; chủng PVYNTN
tạo vết đốm vòng chết hoại trên củ khoai tây.

153
Nhìn chung việc phân chia chủng PVY khá phức tạp vì các chủng thƣờng hay tái tổ hợp với
nhau. Mới đây (1998), dựa trên phân tích phân tử, các isolate PVY trên thế giới đƣợc chia thành
3 cụm phả hệ là:
1. PVYN bao gồm các mẫu virus khoai tây thuộc chủng PVY
N và PVY
NTN,
2. PVYO bao gồm các mẫu virus khoai tây tƣơng ứng với chủng PVY
O
3. PVYNP
bao gồm các mẫu virus không phải phân lập từ khoai tây (NP là viết tắt của non-
potato). Phần lớn các mẫu thuộc chủng PVYC.
15.5.3 PVY tại Việt Nam
Môt nghiên cứu dựa vào trình tự gen CP của 3 mẫu PVY Việt Nam (thu trên khoai tây và ớt )
năm 2008 cho thấy cả 3 mẫu này phân nhóm vào 3 cụm khác nhau ở trên, trong đó mẫu virus
phân lập từ cây ớt tại Đà Lạt phân nhóm vào cụm PVYNP
. Kết quả này chứng tỏ virus PVY tại
Việt Nam khá đa dạng và có thể đã xâm nhập vào Việt Nam bằng nhiều đƣờng khác nhau.
Hình 15-4 Ba nhóm phả hệ PVY trên thế giới và Việt Nam (in đậm)
AJ390290-[NUCLEOTIDEN/v951156-2]-UK-potato
PVY-[VN/P2]-Vietnam-potato
X68223-[Europe-H]-Hungary-potato
AF321554-[NUCLEOTIDEN]-Slovenia-potato
AJ133454-[NUCLEOTIDEN]-The Netherland-petunia
AJ535662-[NUCLEOTIDEN-Ca/H]-Hungary-pepper
M95491-Hungary
AY166866-[NUCLEOTIDEN-Tu660]-Canada potato
D12570-[T]-Japan-potato
AF255660-[NBR]-Brazil-potato
M22470-[N]-New Zealand-potato
X97895-[N-605]-Switzerland
S74813-[T13]
AJ390296-[NUCLEOTIDEN-NN-UK-N]-UK-potato
AJ390308--NUCLEOTIDEN-S-RBS96]-UK-potato
AF525081-Solanum palinacanucleotidehum
X12456-[N]-France-potato-ref.
U09509-[O]-Canada-potato
X68222-[O-US]-USA-potato
Z70238-[N-Wilga]-Poland-potao
Z70239-[O-LW]-Poland-potato
AF118153-[O]-India-eggplanucleotide
DQ157179-[N:O-OR1]-USA-potato
PVY-[VN/P1]-Vietnam-potato
AJ223593-[O-768]-Switzerland
AJ390305-[O-Des]-UK-potato
AF012028-[C-30]-Germany
AF463399-[MrNs]-USA-tobacco
X68224-[NsNr]-USA-tobacco
PVY-[VN/C10]-Vietnam-chilli
AJ303096-[PN-82]-Spain-pepper
AJ439544-[Son41]-France-Solanum nigrum
AJ005639-[P21-82]-Spain-pepper
AJ390307-[C-O-Tom]-Portugal-tomato
AF012027-[C-28]-Germany
AF012029-[C-45]-Germany
AJ303093-[Si15]-Italy-pepper
AJ303094-[K16.94]-Tunisia-pepper
AF237963-[nnp]-Italy-pepper
AJ303095-[Tu12.3]-Turkey-pepper
AJ439545-[LYE84.2]-Spain-tomato
100
99
67
88
100
64
82
99
98
92
100
98
76
69
61
100
97
93
92
80
80
77
74
100
PVYN
PVYO
PVYNP
(Non-potato)

154
15.5.4 Triệu chứng
Trên khoai tây, PVY thƣờng gây bệnh khảm hoặc khảm nhăn. Triệu chứng rất biến động phụ
thuộc chủng virus, giống cây, điều kiện thời tiết và cách nhiễm bệnh (bằng rệp hay từ củ). Các
triệu chứng thƣờng bắt gặp trên khoai tây bao gồm khảm, khảm nhăn, lùn, lá giòn, đốm chết hoại
gân (thƣờng ở lá phía dƣới), đốm vòng chết hoại trên củ.
Trên thuốc lá, ớt, cà chua, PVY thƣờng gây các triệu chứng đốm biến vàng, khảm, hoặc đốm
chết hoại trên gân. Nhìn chung lá cây bệnh ít bị biến dạng hơn so với khoai tây.
Hình 15-5 Các triệu chứng khảm, khảm nhăn, lùn, chết hoại lá và đốm vòng chết hoại trên khoai
tây do PVY (Kerlan, 2006; Hà Viết Cƣờng, 2005)
15.5.5 Lan truyền
PVY truyền dễ dàng qua củ khoai tây nhƣng không truyền qua hạt thật (hạt khoai tây, cà
chua, thuốc lá). Củ bệnh là một trong các nguồn bệnh quan trọng nhất trên khoai tây.
Khoảng 50 loài rệp muội có thể truyền PVY theo kiểu không bền vững. Trong số các loài rệp
này, rệp đào M. persicae là vector hiệu quả nhất.
Ngoài ra, nhiều cây ký chủ dại họ cà (thuộc các chi Datura, Solanum, Physalis) là nguồn bệnh
quan trọng đối với PVY.

155
15.5.6 Phòng chống
15.5.6.1 Đối với khoai tây
1. Tạo giống sạch bệnh: trồng khoai tây từ hạt, nuôi cấy đỉnh sinh trƣởng, trồng ruộng giống tại
các khu vực biệt lập, điều tra và vệ sinh đồng ruộng (nhổ bỏ cây bệnh, ký chủ dại).
2. Phòng trừ vector. Mặc dù kém hiệu quả nhƣng có thể làm giảm mật độ vector.
3. Tạo giống kháng với gen kháng có ngồn gốc từ cây. Ba loại tính kháng trên khoai tây đã
đƣợc áp dụng là (i) kháng ngang, (ii) tạo phản ứng siêu nhạy và (iii) cực kháng. Tính kháng
ngang là tính kháng đa gen. Tính kháng tạo phản ứng siêu nhạy do một gen kháng chủ, trội là
Ny qui định. Dạng cực kháng cũng do một gen kháng chủ, trội là Ry qui định. Các gen qui
định khả năng cực kháng đều lấy từ khoai tây dại nhƣ Rysto từ S. stoloniferum và Ryadg từ S.
andigena.
4. Tạo giống kháng kiểu DPR. Tạo các giống mang gen của PVY (nhƣ CP, P1, NIb) cũng đã
đƣợc nghiên cứu. Cây chuyển gen CP thƣờng tạo ra kiểu hình dạng cực kháng. Ngoài ra, tạo
giống kháng thông qua cơ chế RNAi cũng đã và đang đƣợc nghiên cứu từ khoảng 10 năm trở
lại đây. Một số giống khoai tây chuyển gen kháng PVY đã đƣợc thƣơng mại hóa nhƣ giống
Russett Burbank và Shepody tại Mỹ.
15.5.6.2 Đối với thuốc lá
Trên thuốc lá, tạo giống kháng dùng cả gen kháng từ cây và từ virus là biện pháp chính để
chống lại PVY. Tuy nhiên giống mang gen kháng từ cây (ví dụ gen va) thƣờng bị giảm năng suất
và chất lƣợng thuốc. Vì vậy, tính kháng thông qua PDR thƣờng đƣợc chú ý hơn.
15.5.6.3 Đối với cà chua và ớt
1. Tạo giống kháng dựa vào gen kháng của cây. Nhiều gen kháng chủ (cả trội và lặn) chống
PVY đã đƣợc phát hiện trên cà chua và ớt. Ví dụ gen lặn pot-1 từ cây L. hirsutum, các gen
lặn pvr1, pvr2, pvr4 và pvr5 từ Capsicum annuum; các gen kháng trội Pvr4 từ C. annuum và
Pvr7 từ C. chinense. Các gen kháng lặn đặc biệt pvr2 đã đƣợc đƣa vào nhiều giống ớt ở châu
Âu và Mỹ. Gen kháng trội Pvr4 kháng đƣợc tất cả các chủng PVY cũng đƣợc đƣa vào giống
các giống ớt lai ở châu Âu.
2. Ngoài ra, các kỹ thuật canh tác nhƣ dùng bẫy dính màu vàng hoặc nhà lƣới đã đƣợc áp dụng
thành công trong một số trƣờng hợp đặc biệt (nơi áp lực bệnh không cao, mô hình canh tác
kiểu công nghiệp) nhƣ ở Israel để chống phát tán bệnh trên ruộng.
15.6 BEAN COMMON MOSAIC VIRUS (BCMV)
15.6.1 Tầm quan trọng
BCMV, thuộc chi Potyvirus, là một trong số các virus có ý nghĩa kinh tế nhất trên cây họ đậu.
Ngoài tự nhiên, BCMV chủ yếu nhiễm trên các cây đậu đỗ thuộc chi Phaseolus, đặc biệt là P.
vulgaris. Trên cây P. vulgaris, BCMV có thể làm giảm năng suất tới 60 %. Ngoài tự nhiên,
BCMV có thể lan truyền bằng nhiều loài rệp muôi (aphididae), và đặc biệt, có thể lan truyền qua
hạt giống. Do có khả năng truyền qua hạt nên tác động của BCMV đối với cây họ đậu có thể rất
lớn trên qui mô toàn cầu.

156
15.6.2 Virus
BCMV là một virus rất đa dạng về di truyền và phức tạp về phân loại. BCMV là một phức
hợp các virus gây bệnh trên cây đậu đỗ và có một hiện trạng phân loại rất dễ gây nhầm lẫn. Về
mặt lịch sử, các chủng BCMV nhiễm trên đậu đỗ đã đƣợc chia thành 2 nhóm phân biệt về mặt
huyết thanh học (serotype) là nhóm serotype A và nhóm serotype B. Trong những năm 90, nhiều
nghiên cứu, chủ yếu dựa trên đặc điểm phân tử đã phân loại lại BCMV. Hiện nay, BCMV chỉ
bao gồm các chủng thuộc nhóm serotype B và nhiều virus đậu đỗ (vốn trƣớc kia đƣợc xem là
các loài riêng biệt nhƣ Blackeye cowpea mosaic virus (BlCMV), Peanut stripe virus (PStV) và
Azuki bean mosaic virus (AzMV).
Các nghiên cứu phả hệ học cũng cho thấy BCMV và nhiều potyvirus nhiễm trên cây đậu đỗ
nhƣ soybean mosaic virus (SMV), bean yellow mosaic virus (BYMV) và cowpea aphid borne
mosaic virus (CABMV) cũng nhƣ nhiều potyvirus không nhiễm trên đậu đỗ nhƣ dasheen mosaic
virus (DsMV), passion fruit woodiness virus (PWV), South African passiflora virus (SAPV),
watermelon mosaic virus II (WMVII) và zucchini yellow mosaic virus (ZYMV) phân nhóm với
nhau và hình thành nên một nhóm thuộc chi Potyvirus gọi là ―BCMV subgroup‖.
15.6.3 BCMV ở Việt Nam
Ở Việt Nam, mặc dù các cây đậu đỗ thƣờng biểu hiện triệu chứng giống nhƣ bị nhiễm BCMV
nhƣng cho tới nay, chỉ có 2 công bố về virus quan trọng này. Đầu tiên là một nghiên cứu dựa
trên phân tích ELISA và giải trình tự từ các mẫu hạt giống đậu đỗ thu thập ở miền Bắc Việt
Nam. Dựa trên nghiên cứu này, các tác giả (Ngô Bích Hảo et al., 2003) đã xác nhận sự có mặt
của BCMV trên hạt đậu (Vigna unguiculata) và cho thấy tỷ lệ hạt đậu nhiễm virus là từ 0,8 to
12,4 % . Nghiên cứu thứ 2 là các phân tích trình tự gen từ 9 mẫu virus phân lập từ các cây họ đậu
biểu hiện triệu chứng bệnh nhƣ đậu đen, đậu đỏ, đậu đũa, đậu tƣơng và muồng 3 lá (một cây che
phủ cà phê) thu thập tại nhiều tỉnh Việt Nam (Hà Viết Cƣờng et al., 2008).
Các nghiên cứu gần đây cho thấy vùng Đông Nam Á và đặc biệt Việt Nam là một trong
những trung tâm phát sinh và đa dạng của BCMV vì:
(i) BCMV đã đƣợc phát hiện thấy trên khá nhiều cây họ đậu nhƣ đậụ đen, đậu đỏ (P.
vulgaris), đậu đũa (V. unguiculata), đậu tƣơng (Glycin max) và muồng 3 lá (Crotalaria
anagyroides) – một cây che phủ cà phê tại Đắc Lắc.
(ii) Phần lớn các mẫu BCMV phân lập tại Việt Nam đã phân nhóm với 2 nhóm chủng chính là
chủng black eye cowpea mosaic (BlCM) và chủng peanut stunt virus (PSt)
(iii) Đặc biệt, 2 mẫu BCMV phân lập từ đậu đen (BCMV-VN-BB2-5) và đậu đũa (BCMV-
YB2) đã biểu hiện mực độ biến động trình tự gen rất cao, chẳng hạn thiếu các chuỗi epitope nằm
ở đầu amin của protein CP và trong phân tích phả hệ, chúng nằm ở vị trí trung gian giữa cụm
chứa tất cả các chủng/isolate BCMV và cụm chứa các potyvirus khác thuộc nhóm ―BCMV
subgroup‖.
(iv) Một phân tích phả hệ học mới đây nhất dựa trên so sánh trình tự khoảng 400 chuỗi
nucleotide mã hóa phần trung tâm và phần đầu carboxyl protein vỏ virus đại diện cho 34 loài
potyvirus thuộc nhóm ―BCMV subgroup‖ (Gibb et al., 2008) đã chứng minh rõ ràng rằng các
virus của nhóm này (kể cả BCMV) có nguồn gốc từ vùng Đông Nam Á, Nam Á, và khu vực
châu Đại dƣơng.
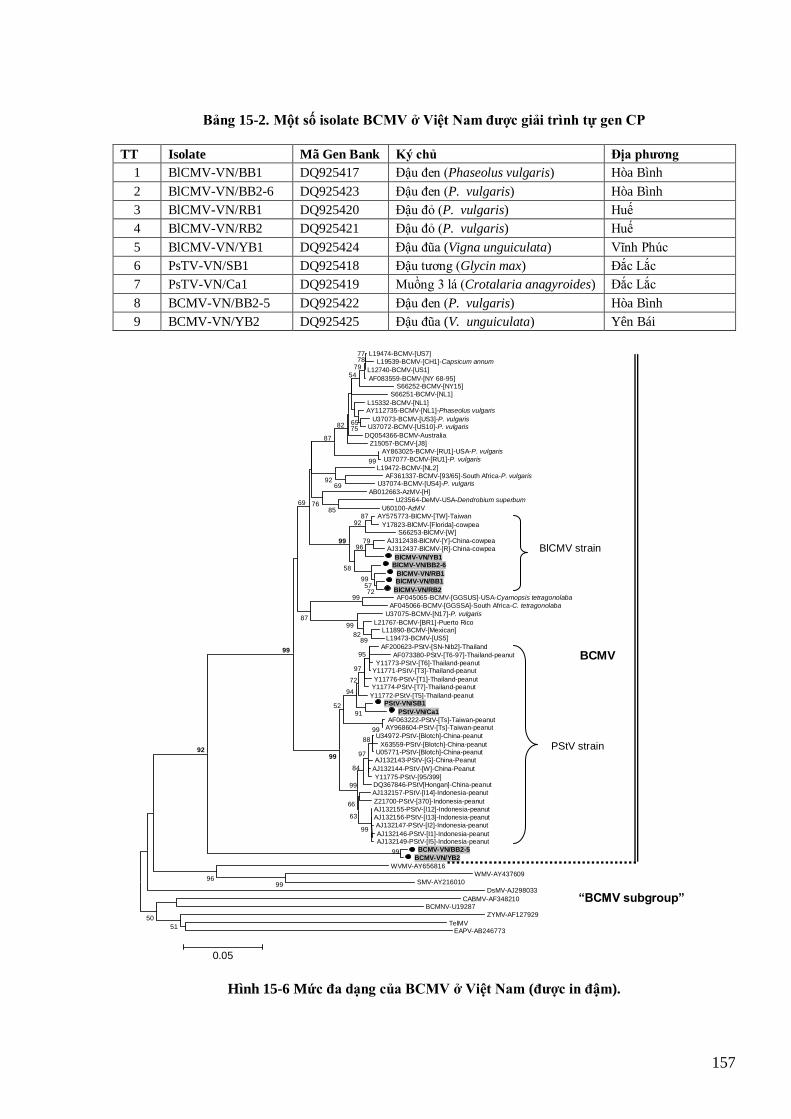
157
Bảng 15-2. Một số isolate BCMV ở Việt Nam đƣợc giải trình tự gen CP
TT Isolate Mã Gen Bank Ký chủ Địa phƣơng
1 BlCMV-VN/BB1 DQ925417 Đậu đen (Phaseolus vulgaris) Hòa Bình
2 BlCMV-VN/BB2-6 DQ925423 Đậu đen (P. vulgaris) Hòa Bình
3 BlCMV-VN/RB1 DQ925420 Đậu đỏ (P. vulgaris) Huế
4 BlCMV-VN/RB2 DQ925421 Đậu đỏ (P. vulgaris) Huế
5 BlCMV-VN/YB1 DQ925424 Đậu đũa (Vigna unguiculata) Vĩnh Phúc
6 PsTV-VN/SB1 DQ925418 Đậu tƣơng (Glycin max) Đắc Lắc
7 PsTV-VN/Ca1 DQ925419 Muồng 3 lá (Crotalaria anagyroides) Đắc Lắc
8 BCMV-VN/BB2-5 DQ925422 Đậu đen (P. vulgaris) Hòa Bình
9 BCMV-VN/YB2 DQ925425 Đậu đũa (V. unguiculata) Yên Bái
Hình 15-6 Mức đa dạng của BCMV ở Việt Nam (đƣợc in đậm).
L19474-BCMV-[US7] L19539-BCMV-[CH1]-Capsicum annum
L12740-BCMV-[US1] AF083559-BCMV-[NY 68-95]
S66252-BCMV-[NY15] S66251-BCMV-[NL1]
L15332-BCMV-[NL1] AY112735-BCMV-[NL1]-Phaseolus vulgaris
U37073-BCMV-[US3]-P. vulgaris U37072-BCMV-[US10]-P. vulgaris
DQ054366-BCMV-Australia Z15057-BCMV-[J8]
AY863025-BCMV-[RU1]-USA-P. vulgaris U37077-BCMV-[RU1]-P. vulgaris
L19472-BCMV-[NL2] AF361337-BCMV-[93/65]-South Africa-P. vulgaris
U37074-BCMV-[US4]-P. vulgaris AB012663-AzMV-[H]
U23564-DeMV-USA-Dendrobium superbum U60100-AzMV
AY575773-BlCMV-[TW]-Taiwan Y17823-BlCMV-[Florida]-cowpea
S66253-BlCMV-[W] AJ312438-BlCMV-[Y]-China-cowpea AJ312437-BlCMV-[R]-China-cowpea
BlCMV-VN/YB1 BlCMV-VN/BB2-6
BlCMV-VN/RB1 BlCMV-VN/BB1
BlCMV-VN/RB2 AF045065-BCMV-[GGSUS]-USA-Cyamopsis tetragonolaba
AF045066-BCMV-[GGSSA]-South Africa-C. tetragonolaba U37075-BCMV-[N17]-P. vulgaris
L21767-BCMV-[BR1]-Puerto Rico L11890-BCMV-[Mexican]
L19473-BCMV-[US5] AF200623-PStV-[SN-Nib2]-Thailand
AF073380-PStV-[T6-97]-Thailand-peanut Y11773-PStV-[T6]-Thailand-peanut
Y11771-PStV-[T3]-Thailand-peanut Y11776-PStV-[T1]-Thailand-peanut
Y11774-PStV-[T7]-Thailand-peanut Y11772-PStV-[T5]-Thailand-peanut
PStV-VN/SB1 PStV-VN/Ca1
AF063222-PStV-[Ts]-Taiwan-peanut AY968604-PStV-[Ts]-Taiwan-peanut
U34972-PStV-[Blotch]-China-peanut X63559-PStV-[Blotch]-China-peanut
U05771-PStV-[Blotch]-China-peanut AJ132143-PStV-[G]-China-Peanut
AJ132144-PStV-[W]-China-Peanut Y11775-PStV-[95/399] DQ367846-PStV[Hongan]-China-peanut AJ132157-PStV-[I14]-Indonesia-peanut Z21700-PStV-[370]-Indonesia-peanut AJ132155-PStV-[I12]-Indonesia-peanut AJ132156-PStV-[I13]-Indonesia-peanut AJ132147-PStV-[I2]-Indonesia-peanut AJ132146-PStV-[I1]-Indonesia-peanut AJ132149-PStV-[I5]-Indonesia-peanut
BCMV-VN/BB2-5 BCMV-VN/YB2
WVMV-AY656816 WMV-AY437609
SMV-AY216010 DsMV-AJ298033
CABMV-AF348210 BCMNV-U19287
ZYMV-AF127929 TelMV
EAPV-AB246773
99
99 88
97 84
66 63
99
95 97
91
72 94
52
99
72 57 99
79 96
87 92
58 99
99
99
89 82 99
69 92
87
77 78
79
75 65
54
82 87
85 76 69
99
99
99 96
92
51 50
0.05
“BCMV subgroup”
BCMV
BlCMV strain
PStV strain

158
15.6.4 Triệu chứng
BCMV tạo một số các triệu chứng phổ biến sau trên cây đâu: khảm thƣờng, khảm vàng, khảm
xanh (có các nốt phồng xanh đậm), lá thƣờng nhăn, có thể cuốn, cây còi cọc. Tuy nhiên triệu
chứng rất biến động phụ thuộc nhiều vào chủng virus, giống cây, giai đoạn phát triển, thời điểm
nhiễm và điều kiện ngoại cảnh.
Nếu cây nhiễm từ hạt thì các lá non đầu tiên bị đốm biến vàng và biến dạng.
Cây bệnh có ít quả, quả nhỏ, chín chậm; vỏ quả có các đốm màu xah đậm.
Một số chủng virus có thể tạo triệu chứng chết hoại màu nâu đen ở gân lá gọi là bệnh ―thối
đen‖. Triệu chứng này chỉ xuất hiệ trên các giống có gen kháng trội I. Vết chết hoại có thể phát
triển tới tới thân, rễ và đỉnh sinh trƣởng. Cây có thể chết nếu nhiễm sớm. Nếu nhiễm muộn, cây
vẫn sống nhƣng có thể chết từng phần.
Hình 15-7 Bệnh khảm lá do BCMV trên cây họ đậu Việt Nam (ảnh trái:đậu đen, ảnh giữa: muồng
3 lá, Hà Viết Cƣờng et al., 2008) và triệu chứng chết hoại gân của bệnh “thối đen” (CABI, 2006).
15.6.5 Lan truyền
15.6.5.1 Qua hạt giống/phấn hoa
BCMV là một virus nhiễm trên hạt (seed-borne) nhiều loại cây họ đậu, đặc biệt trên các giống
đậu P. vulgaris.
BCMV có khả năng truyền qua phấn hóa vì virus đã đƣợc phát hiện thấy trong hạt phấn.
BCMV có mặt ở nhiều bộ phận nhƣ lá đài (sepals), cánh hóa (petals), nhị (stamens) và nhụy
(pistils). Dịch chiết từ quả và phôi của hạt non có khả năng lây nhiễm rất cao. Cây bị nhiễm sớm
có tỷ lệ hạt nhiễm virus rất cao, có thể > 50%.
Thời điểm nhiễm bệnh ảnh hƣởng đến khả năng truyền qua hạt (seed-transmission). Cây bị
nhiễm ở giai đoạn lá thật đầu tiên thì tỷ lệ truyền qua hạt cao, có thể tới ~ 90%. Cây bị nhiễm ở
giai đoạn 3 lá thật thì tỷ hạt truyền virus giảm mạnh, chỉ còn 1%. Nếu cây bị nhiễm sau ra hoa
khoảng 1 tuần thì hạt không có khả năng truyền virus (mặc dù vẫn có thể bị nhiễm virus). Trong

159
một nghiên cứu khác, tỷ lệ hạt truyền virus đạt tối đa ở giai đoạn lá thật thứ nhất và hầu nhƣ
không có khả năng truyền qua hạt khi cây bị nhiễm bệnh khoảng 1 tháng sau trồng.
Lây nhiễm virus vào lá mầm dẫn tới biểu hiện bệnh cũng nhƣ khả năng truyền qua hạt đạt tối
đa.
Virus tồn tại chủ yếu ở phôi nhƣng cũng có thể ở các bộ phân khác của hạt. Virus có mặt ở vỏ
hạt nhƣng biến mất khi hạt khô. Virus không có mặt ở lá mầm.
Virus có khả năng tồn tại trên hạt rất lâu mà vẫn duy trì khả năng truyền bệnh. Có thí nghiệm
cho biết hạt vẫn có thể truyền bệnh sang cây con sau 30 năm Có thí nghiệm cho thấy hạt bảo
quản ở 2-4 oC sau 6 năm vẫn có khả năng truyền bệnh với tỷ lệ 3-4 %.
Hạt nhiễm virus nhìn chung không ảnh hƣởng đến sức nảy mầm nhƣng chất lƣợng hạt giảm
sút mạnh.
Truyền qua hạt là cách tạo sự nhiễm bệnh sơ cấp quan trọng nhất đối với đậu. Tuy nhiên khả
năng này cũng khác nhau tùy giống, chủng virus cũng nhƣ thời điểm cây mẹ bị nhễm bệnh.
15.6.5.2 Qua vector
BCMV lan truyền tự nhiên ngoài đồng ruộng bằng nhiều loài rệp muội theo kiểu không bền
vững. Nhiều loài rệp mặc dù không sống trên cây đậu nhƣng vẫn có khả năng truyền virus khi
chúng di chuyển tìm thức ăn.
Truyền qua rệp là cách truyền bệnh thứ cấp quan trọng trên đồng ruộng.
15.6.6 Chẩn đoán
Chẩn đoán dựa vào triệu chứng bệnh do BCMV không dễ vì triệu chứng đa dạng và cây họ
đậu bị nhiễm nhiều virus với triệu chứng tƣơng tự, đặc biệt các triệu chứng khảm lá.
Virus có thể đƣợc chẩn đoán bằng cây chỉ thị. Mƣời một giống đậu (P. vulgaris) có thể phân
biệt đƣợc 10 type gây bệnh (pathotypes) khác nhau
Virus có thể đƣợc chẩn đoán bằng ELISA dùng các kháng thể đơn dòng/đa dòng, đặc hiệu
loài thậm chí chủng. Virus cũng có thể đƣợc chẩn đoán bằng RT-PCR.
15.6.7 Phòng chống
Có thể phòng chống BCMV bằng một số các biện pháp nhƣ (i) trồng hạt khỏe, sạch virus, (ii)
trồng giống kháng, (iii) cải thiện điều kiện canh tác, và (iv) phòng chống vector. Trong các biện
pháp này, 2 biện pháp đầu (i) và (ii) là quan trọng nhất.
15.6.7.1 Trồng giống khỏe và sạch
Trồng giống khỏe và sạch virus đòi hỏi phải có một chƣơng trình sản xuất hạt giống đƣợc
kiểm soát nghiêm ngặt. Ruộng giống thƣờng đƣợc trồng tại khu vự biệt lập (có mật độ rệp thấp,
ít hoặc không có cây ký chủ là nguồn bệnh của BCMV). Ruộng giống đƣợc chăm sóc tốt. Cây
đƣợc kiểm tra sự có mặt của virus thƣờng xuyên bằng ELISA. Vệ sinh đồng ruộng để loại bỏ cây
bệnh trong vụ và giữa các vụ. Hạn chế vector bằng thuốc hóa học hoặc các kỹ thuật thích hợp.
Hạt giống phải đƣợc xác nhận chất lƣợng thông qua kiểm tra sự có mặt của virus bằng các kỹ
thuật thích hợp nhƣ ELISA.

160
15.6.7.2 Tạo giống kháng bệnh
Đối với cây họ đậu, nhiều gen kháng và giống kháng BCMV đã đƣợc xác định và ứng dụng.
Các gen kháng BCMV này tuân theo quan hệ gen đối gen và có thể chia làm 2 nhóm là các gen
kháng lặn và gen kháng trội.
Gen kháng lặn. Các yếu tố khởi đầu dịch mã eIF của cây ký chủ luôn cần cho quá trình dịch
mã và qua đó là quá trình gây bệnh của virus. Các đột biến trên gen mã hóa eIF4E đã đƣợc xác
định là nguồn các gen kháng lặn quan trọng ở nhiều loài cây chống virus. Quan hệ của các gen
kháng này với virus tuân theo qua hệ gen-đối-gen. Gen không độc tƣơng ứng của các potyvirus
chính là VPg.
Trên cây đậu (P. vulgaris), bốn gen kháng lặn, bc-1, bc-2, bc-3 và bc-u, đã đƣợc xác định là
các gen kháng lặn chống BCMV và BCMNV (Bean common mosaic necrosis virus). (Chú ý ký
hiệu alle của các gen này: ví dụ gen bc-1 và bc-12 là alle của nhau, bc-2 và bc-22 là alle của
nhau…). Trong số 4 gen kháng lặn này, bc-1, bc-2, bc-3 qui định tính kháng đặc hiệu chủng còn
bc-u là gen kháng không đặc hiệu chủng. Bản thân bc-u tự nó không kháng đƣợc BCMV nhƣng
nó cần thiết để các gen kháng lặn khác tạo ra tính kháng.
Gen kháng trội. Trên cây họ đậu, một gen kháng trội là gen I đã đƣợc xác định. Gen I thuộc
nhóm gen kháng NB. Tuy nhiên cây chỉ mang gen kháng I thì không chống đƣợc các chủng
BCMV gây chết hoại. Để có tính kháng tốt với các chủng này, cây cần kết hợp cả gen kháng I và
các gen kháng lặn.
Hiện nay, nhiều giống đậu kháng BCMV kết hợp cả gen kháng trội I và các gen kháng lặn
bc đã đƣợc tạo ra tại Trung tâm Nông nghiệp Nhiệt đới Quốc tế (CIAT:
http://www.ciat.cgiar.org/Paginas/index.aspx)
15.7 SUGARCANE MOSAIC VIRUS (SCMV) VÀ SORGHUM MOSAIC VIRUS
(SRMV)
15.7.1 Các potyvirus hại họ hòa thảo
Có 6 potyvirus nhiễm trên cây hòa thảo họ Graminae nhƣ mía, ngô, lúa miến (sorghum) và
các cây cỏ hòa thảo khác là SCMV, SrMV, MDMV (Maize dwarf mosaic virus), JGMV
(Johnsongrass mosaic virus), ZeMV (Zea mosaic virus) và PenMV (Pennisetum mosaic virus).
Các virus này tạo thành một nhóm gọi là ―SCMV sub-group‖.
1. JGMV có phân bố hẹp, chỉ ở Öc, Mỹ, Nigeria. Virus không truyền qua hạt.
2. MDMV có phân bố địa lý rộng hơn (có thể khắp thế giới) và nhiễm tự nhiên trên các cây
ngô, cỏ Johnsongrass và lúa lúa miến. Tuy nhiên danh tính của virus rất dễ gây nhầm lẫn vì
nhiều bệnh virus đƣợc cho là do MDMV gây ra thực chất là do SCMV. Virus có thể truyền
qua hạt ngô với tỷ lệ rất thấp (0.007% to 0.4%).
3. SrMV phát hiện thấy ở Mỹ và nhiều nƣớc châu Á. Virus chỉ gây bệnh trên lúa miến và mía.
4. ZeMV phát hiện thấy trên ngô ở Israel năm 2000.
5. PenMV phát hiện thấy trên cỏ trắng (Pennisetum centrasiaticum) ở phía Bắc Trung Quốc
năm 2003.
6. SCMV là virus phân bố khắp thế giới, nhiễm tự nhiên trên nhiều loài cây họ Gramineae. Hai
chủng SCMV mới cũng đƣợc báo cáo gây bệnh trên cây không phải họ Graminae là cây cọ ở
Colombia, cây chuối sợi abaca và cây hoàng tinh (Maranta arundinacea) ở Philippines.

161
15.7.2 SCMV và SrMV tại Việt Nam
Ở Việt Nam, bệnh khảm lá ngô đã đƣợc quan sát thấy từ lâu. Các nghiên cứu gần đây đã phát
hiện đƣợc 2 potyvirus hại họ hòa thảo là SCMV và SrMV.
SCMV gây bệnh khảm lá ngô, mía và hoàng tinh. Đây là bệnh virus phổ biến nhất trên ngô và
mía ở Việt Nam. Phân tích phả hệ dựa vào trình tự gen CP cho thấy virus ở Việt Nam khá đa
dạng, phân nhóm với các mẫu có nguồn gốc ký chủ khác nhau thuộc nhiều khu vực địa lý là châu
Mỹ, châu Âu và châu Á. Đặc biệt 3 mẫu SCMV phân lập từ mía ở Việt Nam tạo thành một nhóm
riêng biệt (nhóm IV) chia sẻ chung tổ tiên với tất cả các mẫu SCMV trên thế giới.
SrMV gây bệnh khảm lá mía. Trên mía, không thể phân biệt đƣợc triệu chứng khảm lá do
SrMV và SCMV gây ra. Phân tích phả hệ cũng cho thấy SrMV của Việt Nam phân nhóm với các
mẫu SrMV có nguồn gốc phía Nam Trung Quốc.
Hình 15-8 Bệnh khảm lá do SCMV trên mía (trái), ngô (giữa) và hoàng tinh (phải) tại Việt Nam
Bảng.15-3 Một số isolate SCMV và SrMV đƣợc giải trình tự gen CP
TT Isolate Mã GenBank Ký chủ Địa điểm
1 SCMV-VN/AR1 DQ925432 Hoàng tinh Hà Nội
2 SCMV-VN/M1 DQ925426 Ngô Hòa Bình
3 SCMV-VN/M2 DQ925429 Ngô Hà Tây
4 SCMV-VN/SC1 DQ925431 Mía Yên Bái
5 SCMV-VN/SC2 DQ925427 Mía Hòa Bình
6 SCMV-VN/SC3 DQ925430 Mía Bắc Giang
7 SCMV-VN/SC4 DQ925428 Mía Hà Nội
8 SrMV-VN/SC5 DQ925433 Mía Hòa Bình
9 SrMV-VN/SC6 DQ925434 Mía Hà Tây
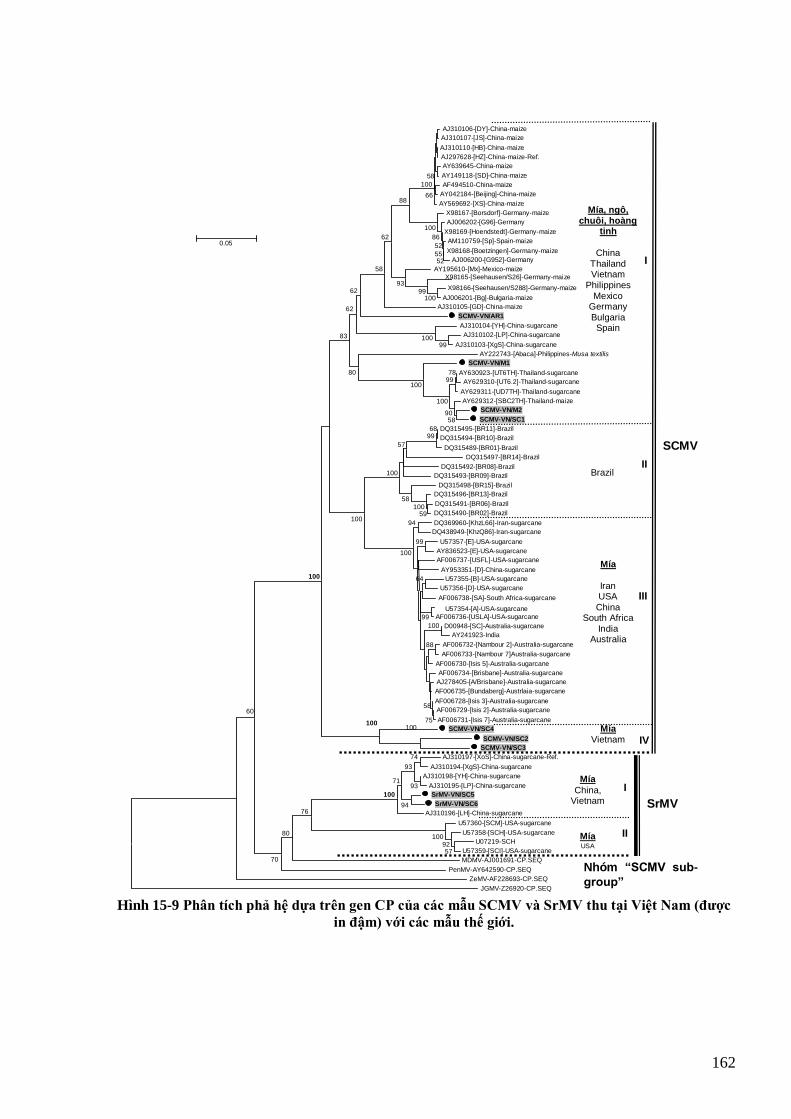
162
Hình 15-9 Phân tích phả hệ dựa trên gen CP của các mẫu SCMV và SrMV thu tại Việt Nam (đƣợc
in đậm) với các mẫu thế giới.
AJ310106-[DY]-China-maize
AJ310107-[JS]-China-maize
AJ310110-[HB]-China-maize
AJ297628-[HZ]-China-maize-Ref.
AY639645-China-maize
AY149118-[SD]-China-maize
AF494510-China-maize
AY042184-[Beijing]-China-maize
AY569692-[XS]-China-maize
X98167-[Borsdorf]-Germany-maize
AJ006202-[G96]-Germany
X98169-[Hoendstedt]-Germany-maize
AM110759-[Sp]-Spain-maize
X98168-[Boetzingen]-Germany-maize
AJ006200-[G952]-Germany
AY195610-[Mx]-Mexico-maize
X98165-[Seehausen/S26]-Germany-maize
X98166-[Seehausen/S288]-Germany-maize
AJ006201-[Bg]-Bulgaria-maize
AJ310105-[GD]-China-maize
SCMV-VN/AR1
AJ310104-[YH]-China-sugarcane
AJ310102-[LP]-China-sugarcane
AJ310103-[XgS]-China-sugarcane
AY222743-[Abaca]-Philippines-Musa textilis
SCMV-VN/M1
AY630923-[UT6TH]-Thailand-sugarcane
AY629310-[UT6.2]-Thailand-sugarcane
AY629311-[UD7TH]-Thailand-sugarcane
AY629312-[SBC2TH]-Thailand-maize
SCMV-VN/M2
SCMV-VN/SC1
DQ315495-[BR11]-Brazil
DQ315494-[BR10]-Brazil
DQ315489-[BR01]-Brazil
DQ315497-[BR14]-Brazil
DQ315492-[BR08]-Brazil
DQ315493-[BR09]-Brazil
DQ315498-[BR15]-Brazil
DQ315496-[BR13]-Brazil
DQ315491-[BR06]-Brazil
DQ315490-[BR02]-Brazil
DQ369960-[KhzL66]-Iran-sugarcane
DQ438949-[KhzQ86]-Iran-sugarcane
U57357-[E]-USA-sugarcane
AY836523-[E]-USA-sugarcane
AF006737-[USFL]-USA-sugarcane
AY953351-[D]-China-sugarcane
U57355-[B]-USA-sugarcane
U57356-[D]-USA-sugarcane
AF006738-[SA]-South Africa-sugarcane
U57354-[A]-USA-sugarcane
AF006736-[USLA]-USA-sugarcane
D00948-[SC]-Australia-sugarcane
AY241923-India
AF006732-[Nambour 2]-Australia-sugarcane
AF006733-[Nambour 7]Australia-sugarcane
AF006730-[Isis 5]-Australia-sugarcane
AF006734-[Brisbane]-Australia-sugarcane
AJ278405-[A/Brisbane]-Australia-sugarcane
AF006735-[Bundaberg]-Austrlaia-sugarcane
AF006728-[Isis 3]-Australia-sugarcane
AF006729-[Isis 2]-Australia-sugarcane
AF006731-[Isis 7]-Australia-sugarcane
SCMV-VN/SC4
SCMV-VN/SC2
SCMV-VN/SC3
AJ310197-[XoS]-China-sugarcane-Ref.
AJ310194-[XgS]-China-sugarcane
AJ310198-[YH]-China-sugarcane
AJ310195-[LP]-China-sugarcane
SrMV-VN/SC5
SrMV-VN/SC6
AJ310196-[LH]-China-sugarcane
U57360-[SCM]-USA-sugarcane
U57358-[SCH]-USA-sugarcane
U07219-SCH
U57359-[SCI]-USA-sugarcane
MDMV-AJ001691-CP.SEQ
PenMV-AY642590-CP.SEQ
ZeMV-AF228693-CP.SEQ
JGMV-Z26920-CP.SEQ
78 99
58 90
100
100
100
100 100
99 100
52 55
52
86
100
59 100
68 99
57
58
100
100
99
99
94
88
75
64
58
100
66
58
100
100
99
93
88
80
62
58
62
62
83
100
94
93
74
93
71
100
57 92
100
76
80
70
60
0.05
Mía
China, Vietnam
Mía
Iran USA
China South Africa
India Australia
Brazil
Mía Vietnam
Mía
USA
Mía, ngô, chuôi, hoàng
tinh
China
Thailand Vietnam
Philippines Mexico
Germany Bulgaria Spain
SCMV
SrMV
IV
III
I
II
I
II
Nhóm “SCMV sub-
group”

163
15.8 ONION YELLOW DWARF VIRUS (OYDV), LEEK YELLOW STRIPE VIRUS
(LYSV) VÀ SHALLOT YELLOW STRIPE VIRUS (SYSV)
15.8.1 Các virus hành tỏi
Cây họ hành tỏi (Alliace) có thể bị nhiễm nhiều loại virus RNA thuộc các chi Carlavirus,
Allexivirus và Potyviruses. Trong số các virus hành tỏi, các potyvirus có ý nghĩa kinh tế nhất.
Năm 1993, Van Dijk đã thực hiện một khảo sát dựa trên gần 6000 mẫu hành tỏi khắp thế giới
và xác định đƣợc 4 potyvirus là LYSV (Leek yellow stripe virus), OYDV (Onion yellow dwarf
virus), SYSV (Shallot yellow stripe virus) và WoYSV (Welsh onion yellow stripe virus). Các
nghiên cứu tiếp theo của Tsuneyoshi et al. (1998) và van de Vlugt et al. (1999) đã kết luận lại
WoYSV chỉ là một chủng của SYSV.
Trên cây hành tỏi, các potyvirus tạo các triệu chứng gồm còi cọc, lá có các đốm hoặc đốm sọc
biến màu từ vàng rất nhạt tới rõ. Tất cả 3 virus trên đều tạo triệu chứng tƣơng tự nhau nên triệu
chứng ít có giá trị chẩn đoán. Tất cả các virus hành tỏi đều truyền qua củ giống và rệp muội theo
kiểu không bền vững. Vì các cây hành tỏi thƣờng đƣợc trồng từ củ giống nên củ giống nhiễm
virus là cách lan truyền quan trọng nhất của các virus này.
15.8.2 OYDV, LYSV và SYSV tại Việt Nam
Dựa vào trình tự gen CP, cả 3 potyvirus là OYDV, LYSV và SYSV đều đã đƣợc phát hiện
thấy trên các cây hành ta (shallot) và tỏi tây (leek) tại Việt Nam.
OYDV đã đƣợc phát hiện trên cây tỏi tây còn SYSV và LYSV đƣợc phát hiện trên hành ta và
tỏi tây.
Các cây này biểu hiện triệu chứng còi cọc với các đốm sọc vàng trên lá. Nhiễm hỗn hợp 2
virus (SYSV và LYSV) cũng phổ biến.
Hình 15-10 Các potyvirus trên hành ta và tỏi tây ở Việt Nam

164
15.9 TURNIP MOSAIC VIRUS (TUMV)
15.9.1 Giới thiệu
Turnip mosaic virus (TuMV) đƣợc đánh giá là virus có tầm quan trọng hàng thức 2 (sau
Cucumber mosaic virus, CMV) trên cây rau khắp thế giới. TuMV là potyvirus duy nhất nhiễm
trên cây rau họ thập tự. TuMV có phổ ký chủ cực kỳ rộng, nhiễm ít nhất 318 loài thuộc 43 họ
cây 2 lá mầm. Virus cũng nhiễm cả trên một số cây 1 lá mầm.
Dựa trên tính gây bệnh trên cây cải thuộc 2 chi Brassica và Raphanus, các isolate của TuMV
đƣợc chia thành 2 nhóm ―pathotypes‖ hay ―host types‖ ký hiệu là BR và B. Các isolate TuMV
nhóm BR nhiễm cả trên cải Brassica và Raphanus còn nhóm B chỉ nhiễm cây cải chi Brassica. .
Phân tích phả hệ dựa trên toàn bộ bộ gen virus lại chia các isolate TuMV khắp thế giới thành 4
nhóm là basal-B, basal-BR, Asian-BR và world-B (basal ám chỉ virus thuộc nhánh xuất phát từ
gốc trên cây phả hệ).
15.9.2 TuMV ở Việt Nam
TuMV đã đƣợc phát hiện thấy trên cây cải củ và cải bẹ tại nhiều tỉnh. Cây nhiễm bệnh còi
cọc, biểu hiện triệu chứng khảm lá điển hình. Trên cây cải bẹ bị nhiễm nặng, lá có thể bị chết
hoại.
Phân tích phả hệ dựa trên gen CP cho thấy các mẫu TuMV Việt Nam phân nhóm với nhóm
―world-B‖ – là nhóm rất đa dạng, đƣợc phân lập từ khắp nới trên thế giới. Đặc biệt 4/5 mẫu
TuMV Việt Nam có qua hệ gần gũi với các isolate từ Đông Á (Bắc Trung Quốc, Nhật Bản và
Triều Tiên).
Hình 15-11 TuMV trên cây cải củ (hàng trên) và cải bẹ (hàng dƣới) ở Việt Nam

165
Bảng 15-4 Một số isolate TuMV tại Việt Nam đƣợc giải trình tự gen CP
TT Isolate Mã Genbạnk Cây Địa điểm
1 TuMV-VN/Rs1 DQ925459 Cải củ (Raphanus sativus) Đắk Lắc
2 TuMV-VN/Rs2 DQ925463 Cải củ (R. sativus) Lai Châu
3 TuMV-VN/Bj1 DQ925460 Cải bẹ (Brassica juncea) Đăk Lăc
4 TuMV-VN/Bj2 DQ925461 Cải bẹ (B. juncea) Hòa Bình
5 TuMV-VN/Bj3 DQ925462 Cải bẹ (B. juncea) Lai Châu
Hình 15-12 Phân tích phả hệ các mẫu TuMV từ Việt Nam (in đậm) và thế giới dựa trên gen CP
AB180015-CHZH33
AB180016-CHZH34
AB180025-HZ6
AB180008-CHsE1
AB076504-2J
AB076502-K1J
AB105135-TU-2RI
AB076551-CHN4
AB076552-CHN5
AF394602-TW
AB076549-CHN2
AB076550-CHN3
AB076487-ST19J
AB093625-C42J
AB076499-ON16J
AB180019-CHZJ25
AB180013-CHZC21
AB180014-CHZC22
AB179995-CHHH29
AB179996-CHHH30
AB179997-CHHZ3
AB076545-NPL3
AB093623-DMJ TuMV-VN/Rs1-R. sativus
TuMV-VN/Bj2-B. juncea
AB180027-RADI
AB188983-DNK4
AB105134-TU3 TuMV-VN/Bj3-B. juncea
AB180011-CHYK56
AB180009-CHYK19
AB180010-CHYK20
AB180021-CHZJ27
AB180000-CHK55
TuMV-VN/Rs2-R. sativus
AB180026-CQS1
AB180007-CHSC2
AB076553-HJIS
AY090660-CHN12
AB180012-CHZC1
AB180022-HBI2
AB188987-GBR8
AB189017-NLD2
AB189001-GRC17
AB188989-GBR30
AB093605-KEN1
AB188988-GBR27
AB188990-GBR31
AB188991-GBR32
AB188992-GBR36
AB188993-GBR38
AB194796-UK1
AB188994-GBR50
AB189003-GRC31
AB188998-GRC2
AB188999-GRC6
AB189000-GRC12
AB093604-PV376Br
AB188979-DEU5
AB093612-NZ290
AB093610-CDN1
D10927-Q-Ca
AB093609-USA1
AB188972-CZE4
AB188982-DNK3
AB076531-PV0054
AB188986-FRD1
AB189016-NLD1
AB093611-BZ1
AB188985-FRA2
AB189002-GRC18
AB189004-GRC32
AB188977-DEU1
AB189020-POL4
AB188973-CZE5
AB189018-POL1 TuMV-VN/Bj1-B. juncea
AB188995-GBR51
AB093608-CZE1
AB093606-RUS1
AB188981-DNK2
AB093607-RUS2
AB188975-CZE18
AB189021-PRT1
AB180018-CHZJ23
AB076555-HZ4
AB180017-CHZH36
AB093627-HRD
AB179990-CH6
AB179994-CHBJ2
AB179993-CHBJ1
AB179992-CH8
AB179991-CH7
AB180020-CHZJ26
AB076520-Rn98
AB093601-Cal1
AB093600-ITA7
AB188978-DEU4
AB093603-PV0104
AB189014-ITA8
AB093614-CP845J
AB093613-KYD81J
AB189006-GRC42
AB189007-GRC43
AB188984-ERU1D
AB093599-A64
AB093598-Al
AB189005-GRC41
AB189008-GRC44
AB189013-ITA6
AB189012-ITA5
AB189010-ITA3
AB189011-ITA4
ScMV-AJ316084
JYMV-AB027007
100
100
63
100
71
100
100
100
100
87
100
100
95
91
66
99
99
99
99
95
88
62
90
80
89
82
89
79
62
75
73
70
66 85
87
61
65
63
62
93
99
74
63
51
77
74
80
100
0.05
Wo
rld-B
A
sia
n-B
R B
asal-B
R B
sal-B
East Asia China Korea Japan
Vietnam

166
15.10 DASHEEN MOSAIC VIRUS (DSMV)
DsMV là một potyvirus thuộc nhóm phụ ―BCMV subgroup‖. DsMV là virus quan trọng nhất
trên cây họ ráy (Araceae) khắp thế giới, đặc biệt là vùng nhiệt đới và á nhiệt đới. Virus lan
truyền ngoài tự nhiên qua củ giống và nhiều loài rệp muội theo kiểu không bền vững. Một đặc
điểm phân tử khác biệt của DsMV so với các potyvirus khác là vùng đầu N của gen CP chứa
nhiều chuỗi lặp gồm các amino acid trung tính nhƣ G, T, P, N (ví dụ một chuỗi lặp hoàn hảo
GGGNNTNNTPPANNTTT).
Nhiều cây họ Ráy nhiễm bệnh là cây trồng nhƣ khoai môn, khoai sọ (Colocasia esculenta) và
cây cảnh. Trên cây bệnh, DsMV thƣờng tạo các triệu chứng: khảm lá, khảm dọc gân kiểu ―lông
chim‖ và biến dạng lá.
Tại Việt Nam, DsMV mới đƣợc xác định bằng giải trình tự gen CP từ 2 mẫu là khoai sọ và
bán hạ 3 thùy (Typhonium trilobatum) – một cây dƣợc liệu và là một ký chủ mới của virus (Hà
Viết Cƣờng et al., 2008).
Hình 15-13 Bệnh do DsMV trên cây họ Ráy
Hàng trên (từ trái sang phải): Khảm dạng ―lông chim‖ trên Môn hồng diệp (Philodendron
sp.), khảm sọc giữa gân trên Môn xanh (Philodendron oxycardium), khảm trên Môn trƣờng sinh
(Dieffenbachia sp.) (CABI, 2006)
Hàng dƣới (trái sang phải): Khảm dạng ―lông chim‖ trên khoai sọ và khảm thông thƣờng trên
cây bán hạ 3 thùy ở Việt Nam.

167
15.11 SWEET POTATO FEATHERY MOTTLE VIRUS (SPFMV)
Sweet potato feathery mottle virus (SPFMV) cùng với Sweet potato chlorotic stunt virus
(SPCSV; chi Crinivirus, họ Closteroviridae) và Sweet potato mild mottle virus (SPMMV; chi
Ipomovirus, họ Potyviridae) là 3 virus chính trong tổng số khoảng 20 virus nhiễm trên khoai lang
toàn thế giới.
Đầu tiên, dựa vào đặc tính huyết thanh học và triệu chứng, SPFMV đƣợc chia thành 2 chủng
là chủng RC (russet crack) gây triệu chứng nứt và hóa bần củ và chủng C (common) không gây
triệu chứng trên củ. Gần đây hơn, dựa trên phân tích gen CP, các isolate của SPFMV chia làm 4
cụm phả hệ tƣơng đƣơng 4 chủng là RC, C, O và EA (East Africa).
Tại Việt Nam, SPFMV mới đƣợc phát hiện gần đây trên 2 mẫu khoai lang ở Huế và Bắc
Giang. Cây bệnh biểu hiện triệu chứng khảm và biến dạng. Phân tích phả hệ dựa vào gen CP của
2 mẫu này thấy chúng thuộc chủng Đông Phi (EA). Đáng chú ý là cụm 2 mẫu Việt Nam này nằm
ở vị trí gốc của cả cụm chủng EA.
Hình 15-14 Triệu chứng bệnh do SPFMV trên khoai lang Việt Nam
Hình 15-15 Cây phả hệ dựa trên gen CP của các mẫu SPFMV Việt Nam và các mẫu thế giới
Chủng RE
Chủng O
Chủng RC
Chủng C
AJ781786-EA-Unj1-Tanzania
AJ010702-EA-Nam3-Uganda
AY518938-RC-Spain1RC
AJ781781-EA-BkB1-Tanzania
AY459593-EA-85
AJ010704-EA-Nam1-Uganda
AJ781783-EA-Mis1-Tanzania
AJ781784-EA-Tar1-Tanzania
AJ010700-Mad-Madagaska
AY459597-EA-Putisrabe-Madagasc
SPFMV-1091-CP.SEQ
AY523551-RC-Zambia-Zambia
AY459599-EA-Portugal-Portugal
AY459600-EA-Carnar3-Spain
SPFMV-1214-CP.SEQ
AF015541-O-K2-Korea
AJ010705-O-Nig3-Nigeria
S43450-RC-RC-USA
AF015540-RC-K1-Korea
D86371-RC-S-Japan
AJ515378-RC-Eg1-Egypt
AJ515379-RC-Eg9-Egypt
AY459591-C-51
AJ781778-C-Au4-Australia
AJ781779-C-Au5C-Australia
AJ010702-Nam3-Uganda
AY459601-C-MD1
AM076411-Ita1-Italia
U96625-C-6-Argentina
71
53
99
90
89
79
58
76
66
59
55
52
76
74
97
7797
73
78
99

168
15.12 ZUCCHINI YELLOW MOSAIC VIRUS (ZYMV)
ZYMV, đƣợc phát hiện đầu tiên ở Ý năm 1973, đang là một trong số các virus quan trong
hàng đầu trên cây họ bầu bí khắp thế giới. Virus phân bố ở ít nhất 50 quốc gia kể cả các khu vực
biệt lập về địa lý nhƣ các hải đảo. Virus khá đa dạng, gồm nhiều chủng, nhóm phả hệ khác nhau.
Virus có phổ ký chủ tƣơng đối hẹp, chủ yếu gây hại trên các cây họ bầu bí. ZYMV tạo triệu
chứng đặc trƣng là khảm lá, biến dạng lá, nốt phồng xanh đậm trên lá, quả; quả biến dạng, cây
lùn. Nhìn chung, triệu chứng do ZYMV gây ra giống với các potyvirus khác trên họ bầu bí và do
đó ít có giá trị chẩn đoán.
Virus lan truyền ngoài tự nhiên bằng nhiều loài rệp muội theo kiểu không bền vững. Virus
cũng truyền qua hạt. Tuy nhiên khả năng truyền qua hạt không cao và không phải tất cả các loài
cây họ bầu bí đều có thể truyền đƣợc (chƣa thấy có công bố cho thấy ZYMV truyền qua dƣa
(Cucurbita melo) và dƣa chuột (Cucumis stivus). Các nghiên cứu cho thấy tỷ lệ hạt truyền virus
thay đổi từ 0.05 – 5 %.
ZYMV có khả năng phát tán rất mạnh. Lý do là (i) virus có thể truyền qua hạt (mặc dù tỷ lệ
không cao) và (2) quan trọng hơn là virus có thời gian tồn tại khả nhiễm khá lâu mặc dù virus
đƣợc truyền theo kiểu không bền vững. Tỷ lệ rệp truyền virus vẫn có thể tới 1 % sau khi chích
nạp 30 giờ (đối với rệp đào), 10-20 giờ (đối với rệp bông).
Về phòng chống: Biện pháp phòng chống ZYMV hiện nay là sử dụng giống kháng mang gen
kháng Zym. Ngoài ra, dùng cây chuyển gen CP (mức protein) hoặc thông qua công nghệ RNAi
cũng đã và đang đƣợc nghiên cứu.
Hình 15-16 Triệu chứng do ZYMV trên cây bí ngồi (Desbiez & Lecoq, 1997)
Tại Việt Nam, ZYMV đã đƣợc phát hiện thấy trên một số cây nhƣ dƣa chuột, bí ngô, bầu.
Phân tích gen CP của một số mẫu ZYMV Việt Nam cho thấy các kết quả thú vị là:
(i) Tất cả các mẫu không phân nhóm với nhóm I là nhóm gồm rất nhiều các isolate có nguồn
gốc từ khắp nơi trên thế giới,
(ii) Có một isolate nằm trong nhóm II là nhóm chỉ có nguồn gốc từ Singapore và đảo
Reunion thuộc Pháp
(iii) Các isolate còn lại cùng với một số isolate có nguồn gốc từ phía Nam Trung Quốc hình
thành nhóm III. Đây là nhóm nằm ở vị trí gốc nhất của cây.
Kết quả này cho thấy Việt Nam và phía Nam Trung Quốc là trung tâm đa dạng của virus
này (Hà Viết Cƣờng et al., 2008).
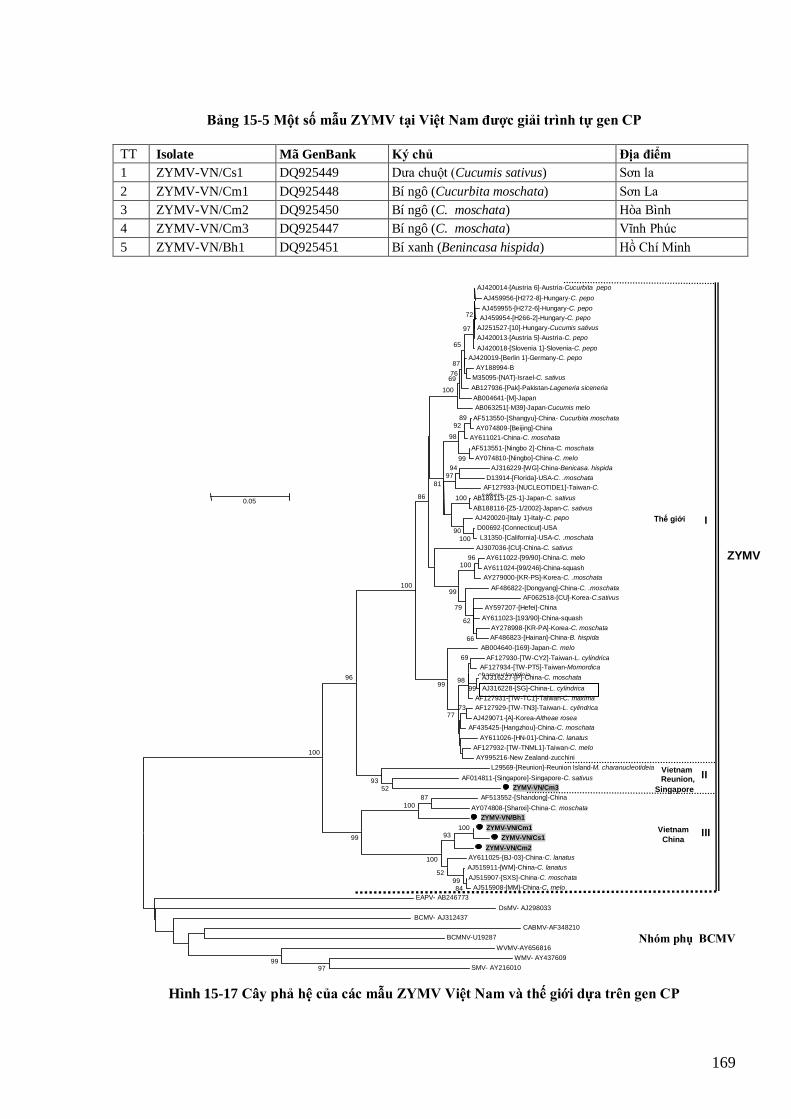
169
Bảng 15-5 Một số mẫu ZYMV tại Việt Nam đƣợc giải trình tự gen CP
TT Isolate Mã GenBank Ký chủ Địa điểm
1 ZYMV-VN/Cs1 DQ925449 Dƣa chuột (Cucumis sativus) Sơn la
2 ZYMV-VN/Cm1 DQ925448 Bí ngô (Cucurbita moschata) Sơn La
3 ZYMV-VN/Cm2 DQ925450 Bí ngô (C. moschata) Hòa Bình
4 ZYMV-VN/Cm3 DQ925447 Bí ngô (C. moschata) Vĩnh Phúc
5 ZYMV-VN/Bh1 DQ925451 Bí xanh (Benincasa hispida) Hồ Chí Minh
Hình 15-17 Cây phả hệ của các mẫu ZYMV Việt Nam và thế giới dựa trên gen CP
AJ420014-[Austria 6]-Austria-Cucurbita pepo
AJ459956-[H272-8]-Hungary-C. pepo
AJ459955-[H272-6]-Hungary-C. pepo
AJ459954-[H266-2]-Hungary-C. pepo
AJ251527-[10]-Hungary-Cucumis sativus
AJ420013-[Austria 5]-Austria-C. pepo
AJ420018-[Slovenia 1]-Slovenia-C. pepo
AJ420019-[Berlin 1]-Germany-C. pepo
AY188994-B
M35095-[NAT]-Israel-C. sativus
AB127936-[Pak]-Pakistan-Lageneria siceneria
AB004641-[M]-Japan
AB063251[-M39]-Japan-Cucumis melo
AF513550-[Shangyu]-China- Cucurbita moschata
AY074809-[Beijing]-China
AY611021-China-C. moschata
AF513551-[Ningbo 2]-China-C. moschata
AY074810-[Ningbo]-China-C. melo
AJ316229-[WG]-China-Benicasa. hispida
D13914-[Florida]-USA-C. .moschata
AF127933-[NUCLEOTIDE1]-Taiwan-C. sativus AB188115-[Z5-1]-Japan-C. sativus
AB188116-[Z5-1/2002]-Japan-C. sativus
AJ420020-[Italy 1]-Italy-C. pepo
D00692-[Connecticut]-USA
L31350-[California]-USA-C. .moschata
AJ307036-[CU]-China-C. sativus
AY611022-[99/90]-China-C. melo
AY611024-[99/246]-China-squash
AY279000-[KR-PS]-Korea-C. .moschata
AF486822-[Dongyang]-China-C. .moschata
AF062518-[CU]-Korea-C.sativus
AY597207-[Hefei]-China
AY611023-[193/90]-China-squash
AY278998-[KR-PA]-Korea-C. moschata
AF486823-[Hainan]-China-B. hispida
AB004640-[169]-Japan-C. melo
AF127930-[TW-CY2]-Taiwan-L. cylindrica
AF127934-[TW-PT5]-Taiwan-Momordica charanucleotideia AJ316227-[P]-China-C. moschata
AJ316228-[SG]-China-L. cylindrica
AF127931-[TW-TC1]-Taiwan-C. maxima
AF127929-[TW-TN3]-Taiwan-L. cylindrica
AJ429071-[A]-Korea-Altheae rosea
AF435425-[Hangzhou]-China-C. moschata
AY611026-[HN-01]-China-C. lanatus
AF127932-[TW-TNML1]-Taiwan-C. melo
AY995216-New Zealand-zucchini
L29569-[Reunion]-Reunion Island-M. charanucleotideia
AF014811-[Singapore]-Singapore-C. sativus
ZYMV-VN/Cm3
AF513552-[Shandong]-China
AY074808-[Shanxi]-China-C. moschata
ZYMV-VN/Bh1
ZYMV-VN/Cm1
ZYMV-VN/Cs1
ZYMV-VN/Cm2
AY611025-[BJ-03]-China-C. lanatus
AJ515911-[WM]-China-C. lanatus
AJ515907-[SXS]-China-C. moschata
AJ515908-[MM]-China-C. melo
EAPV- AB246773
DsMV- AJ298033
BCMV- AJ312437
CABMV-AF348210
BCMNV-U19287
WVMV-AY656816
WMV- AY437609
SMV- AY216010
96 100
100
84 99
93
52
100
87
100
72
97
76
65
87
69
100
100
100
99
66
62
79
99
99
69
98
73 77
99
89 92
98
94 97
90
81
86
100
99
52 93
96
100
97 99
0.05
I
III
II
ZYMV
Nhóm phụ BCMV
Thế giới
Vietnam
China
Vietnam Reunion,
Singapore

170
15.13 CHILLI VEINAL MOTLE VIRUS (CHIVMV) VÀ CHILLI RINGSPOT VIRUS
(CHIRSV)
ChiVMV là một potyvirus hại ớt, đƣợc phát hiện đầu tiên tại Malaysia năm 1979 trên cơ sở
triệu chứng, hình thái phân tử, lan truyền, đặc tính vật lý và huyết thanh học. Virus không truyền
qua hạt giống, truyền bằng rệp theo kiểu không bền vững.
Triệu chứng điển hình nhất trên ớt là là khảm gân xanh. Tuy nhiên, virus cũng gây một loạt
các triệu chứng khác nhƣ biến dạng lá, cây còi cọc. Nhìn chung, chẩn đoán ChiVMV trên đồng
ruộng rất khó do triệu chứng biến động mạnh theo giống, giai đoạn sinh trƣởng và thời điểm
nhiễm bệnh của cây. Ngoài ra, cây ớt có thể bị nhiễm nhiều loại virus khác nhau dẫn tới triệu
chứng biểu hiện rất phức tạp.
Các nghiên cứu thực hiện bởi Trung tâm Rau Châu Á cho thấy ChiVMV cực kỳ phổ biến trên
ớt tại nhiều nƣớc Châu Á, đặc biệt tại Đông Nam Á.
Tại Việt Nam, ChiVMV là virus phổ biến nhất trên ớt. Hầu hết các mẫu ớt biểu hiện triệu
chứng bệnh virus đều nhiễm ChiVMV.
Một potyvirus thuộc một loài mới cũng đƣợc phát hiện tại Việt Nam và đƣợc đặt tên là Chilli
ringspot virus (ChiRSV) dựa trên triệu chứng. ChiRSV mới chỉ đƣợc phát hiện thấy trên 2 mẫu
ớt tại Tháp Chàm và Điện Biên Phủ và không phải là virus phổ biến.
Nhƣ vậy, trên cây ớt tại Việt Nam hiện có 3 potyvirus đã đƣợc phát hiện là ChiVMV,
ChiRSV và PVY, trong đó ChiVMV là virus phổ biến nhất.
Bảng 15-6 Các mẫu potyvirus trên ớt tại Việt Nam đƣợc giải trình tự
TT Isolate Mã GenBank Địa điểm
1 PVY-VN/C10 DQ925436 Đà Lạt
2 ChiVMV-VN/C1 DQ925440 Hà Nội
3 ChiVMV-VN/C2 DQ925441 Hà Nội
4 ChiVMV-VN/C3 DQ925442 Hà Nội
5 ChiVMV-VN/C4 DQ925443 Hà Nội
6 ChiVMV-VN/C5 DQ925444 Hồ Chí Minh
7 ChiVMV-VN/C6 DQ925446 Vĩnh Phúc
8 ChiVMV-VN/C7 DQ925445 Huế
9 ChiRSV-VN/C8 DQ925438 Ninh Thuận
10 ChiRSV-VN/C9 DQ925439 Điện Biên Phủ
Hình 15-18 Bệnh do ChiVMV trên ớt và thuốc lá

171
15.14 CÂU HỎI ÔN TẬP CHƢƠNG 15
1. Đặc điểm họ Potyviridae (phân loại, hình thái, quan hệ vector, bản chất bộ gen)
2. Đặc điểm chi Potyvirus (hình thái, tổ chức bộ gen và tái sinh, chức năng protein HcPro, CI,
VPg, NIb, CP).
3. PRSV (tầm quan trọng, triệu chứng, lan truyền, phòng chống).
4. PVY (tầm quan trọng, triệu chứng, lan truyền, phòng chống).
5. BCMV (tầm quan trọng, triệu chứng, lan truyền, phòng chống).
6. Các giả thuyết có thể về nguồn gốc của ZYMV, BCMV, PVY, SCMV/SrMV
15.15 TÀI LIỆU THAM KHẢO CHÍNH
1. Adams, M. J., Antoniw, J. F. & Beaudoin, F. (2005a). Overview and analysis of the
polyprotein cleavage sites in the family Potyviridae. Molecular Plant Pathology 6, 471-487.
2. Adams, M. J., Antoniw, J. F. & Fauquet, C. M. (2005b). Molecular criteria for genus and
species discrimination within the family Potyviridae. Archives of Virology 150, 459-479.
3. AVRDC. (1993). Vegetable Research and Development in Southeast Asia: The AVNET
Final Report.
4. Bateson, M. F., Lines, R. E., Revill, P., Chaleeprom, W., Ha, C. V., Gibbs, A. J. & Dale, J. L.
(2002). On the evolution and molecular epidemiology of the potyvirus Papaya ringspot virus.
Journal of General Virology 83, 2575-2585.
5. Berger, P. H., Adams, M. J., Barnett, O. W., Brunt, A. A., Hammond, J., Hill, J. H., Jordan,
R. L., Kashiwazaki, S., Rybicki, E., Spence, N., Stenger, D. C., Ohki, S. T., Uyeda, I., van
Jaayen, A., Valkonen, J. & Vetten, H. J. (2005). Potyviridae. In Virus Taxonomy VIIIth
report of the International Committee on Taxonomy of Viruses. Edited by C. M. Fauquet, M.
A. Mayo, J. Maniloff, U. Desselberger & L. A. Ball. London: Elsevier/Academic Press.
6. Berger, P. H., Wyatt, S. D., Shiel, P. J., Silbernagel, M. J., Druffel, K. & Mink, G. I. (1997).
Phylogenetic analysis of the Potyviridae with emphasis on legume-infecting potyviruses.
Archives of Virology 142, 1979-1999.
7. CABI (2006a). Bean common mosaic virus. Crop Protection Compendium CD ROM
8. CABI (2006b). Papaya ring spot virus. Crop Protection Compendium CD ROM
9. CABI (2006c). Potato virus Y. Crop Protection Compendium CD ROM
10. Chen, J., Chen, J. P. & Adams, M. J. (2002). Characterisation of potyviruses from sugarcane
and maize in China. Archives of Virology 147, 1237-1246.
11. Desbiez, C., Wipf-Scheibel, C. & Lecoq, H. (2002). Biological and serological variability,
evolution and molecular epidemiology of Zucchini yellow mosaic virus (ZYMV, Potyvirus)
with special reference to Caribbean islands. Virus Res 85, 5-16.
21. Ha Viet Cuong, S. Coombs, P. A. Revill, R. M. Harding, M. Vu, J. L. Dale. (2008b). Design
and application of two novel degenerate primer pairs for the detection and complete genomic
characterization of potyviruses. Archives of virology. 153:25-36

172
22. Ha Viet Cuong, S. Coombs, P. A. Revill, R. M. Harding, M. Vu, J. L. Dale. (2008c).
Identification and sequence analysis of potyviruses infecting crops in Vietnam. Archives of
virology. 153:45-60
12. Hao, N. B., Albrechtsen, S. E. & Nicolaisen, M. (2003). Detection and identification of the
blackeye cowpea mosaic strain of Bean common mosaic virus in seeds of Vigna unguiculata
sspp. from North Vietnam. Australasian Plant Pathology 32, 505-509.
13. Kreuze, J. F., Karyeija, R. F., Gibson, R. W. & Valkonen, J. P. T. (2000). Comparisons of
coat protein gene sequences show that East African isolates of Sweet potato feathery mottle
virus form a genetically distinct group. Archives of Virology 145, 567-574.
14. Mink, G. I., Vetten, J., Ward, C. W., Berger, P. H., Morales, F., Myers, J. R., Silbernagel, M.
J. & Barnett, O. W. (1994). Taxonomy and Classification of Legume-Infecting Potyviruses -
a Proposal from the Potyviridae Study-Group of the Plant-Virus Subcommittee of ICTV.
Archives of Virology 39, 231-235.
15. Revill, P. A., Ha, C. V., Lines, R. E., Bell, K. E., Vu, M. T. & Dale, J. L. (2004). PCR and
ELISA-based virus surveys of banana, papaya and cucurbit crops in Vietnam. Asia Pacific
Journal of Molecular Biology and Biotechnology 12, 27-32.
16. Shukla, D. D., Ward, C. W., Brunt, A. A. & Berger, P. H. (1998). Potyviridae family.
Description of plant viruses No. 366. AAB (Association of Applied Biologists).
17. Tomimura, K., Spak, J., Katis, N., Jenner, C. E., Walsh, J. A., Gibbs, A. J. & Ohshima, K.
(2004). Comparisons of the genetic structure of populations of Turnip mosaic virus in West
and East Eurasia. Virology 330, 408-423.
18. Tsuneyoshi, T., Matsumi, T., Natsuaki, K. T. & Sumi, S. (1998). Nucleotide sequence
analysis of virus isolates indicates the presence of three potyvirus species in Allium plants.
Archives of Virology 143, 97-113.
19. van der Vlugt, R. A. A., Steffens, P., Cuperus, C., Barg, E., Lesemann, D. E., Bos, L. &
Vetten, H. J. (1999). Further evidence that shallot yellow stripe virus (SYSV) is a distinct
potyvirus and reidentification of Welsh onion yellow stripe virus as a SYSV strain.
Phytopathology 89, 148-155.
20. Van Dijk, P. (1993). Survey and Characterization of Potyviruses and Their Strains of Allium
Species. Neth J Plant Pathol 99, 1-48.

173
Chƣơng 16. VIRUS RNA SỢI ĐƠN CỰC ÂM: RHABDOVIRUS
16.1 TÓM TẮT NỘI DUNG
Chƣơng 16 mô tả phân loại và các đặc trƣng phân tử của họ Rhabdoviridae, đại diện cho
nhóm các virus RNA, sợi đơn, cực âm Chƣơng này cũng đề cập đến một virus gây bệnh vàng lụi
(vàng lá) lúa, một bệnh đã từng gây thành dịch ở miền Bắc trong những năm 70 và đang tái xuất
hiện trên lúa tại nhiều tỉnh miền Bắc.
16.2 HỌ RHABDOVIRIDAE
Tổng số loài rhabdovirus đƣợc công nhận bởi ICTV cho tới năm 2009 là 51. Các
rhabdovirus (họ Rhabdoviridae, bộ Mononegavirales) đƣợc phân loại chính thức vào 6 chi và
một số chƣa phân loại đến chi. ―Rhabdo‖ có nguồn gốc từ từ Hy Lạp ―rhabdos‖ có nghĩa là ―hình
gậy‖ ám chỉ phân tử virus có hình nhộng (gậy ngắn).
Các rhabdovirus gây hại thực vật chỉ thuộc 2 chi Nucleorhabdovirus và Cytorhabdovirus,
còn lại là các virus động vật.
Trong số các rhabdovirus hại động vật, Rabies virus (gây bệnh dại) là nguy hiểm nhất.
Trong số các rhabdovirus hại thực vật, Rice yellow stunt virus (RYSV) – chi Nucleorhabdovirus
gây bệnh vàng lụi lúa là virus quan trọng nhất.
Bảng 16-1. Phân loại họ Rhabdoviridae (bộ Monogavirales) theo ICTV (2009)
TT Chi Loài điển hình Ký chủ Số loài
1 Cytorhabdovirus Lettuce necrotic yellows virus Thực vật 9
2 Nucleorhabdovirus Potato yellow dwarf virus Thực vật 9
3 Vesiculovirus Vesicular stomatitis Indiana virus Động vật 10
4 Ephemerovirus Bovine ephemeral fever virus Động vật 3
5 Lyssavirus Rabies virus Động vật 11
6 Novirhabdovirus Infectious hematopoietic necrosis virus Động vật 4
7 Chưa phân loại Động vật 5
Tổng số 51
16.2.1 Hình thái, cấu trúc phân tử và tổ chức bộ gen
Các rhabdovirus là các virus có bộ gen RNA, sợi đơn, không phân mảnh, cực âm, kích
thƣớc 11-15 kb, chiếm khoảng 1-2% khối lƣợng phân tử virus.
Bộ gen virus chứa ít nhất 5 ORF không liên tục theo chiều từ đầu 3‘ – 5‘ (chú ý RNA của
virus là cực âm) theo thứ tự sau: 3‘-N-P-M-G-L-5‘. Các ORF đƣợc phiên mã thành mRNA và
dịch mã độc lập thành các protein cấu trúc sau:
N: protein nucleoprotein. Bên trong tế bào, N đóng vai trò cân bằng giữa quá trình phiên mã
các gen virus và tái sinh bộ gen virus do tác động đến sự nhận biết các dấu hiệu phiên mã.
L: protein lớn (large protein). L là một RdRp.
M : protein tạo protein nền (matrix protein). M còn đƣợc ký hiệu là M1. M đóng vai trò điều
khiển quá trình phiên mã của virus, ức chế phiên mã của ký chủ, ảnh hƣởng đến sự gây bệnh.
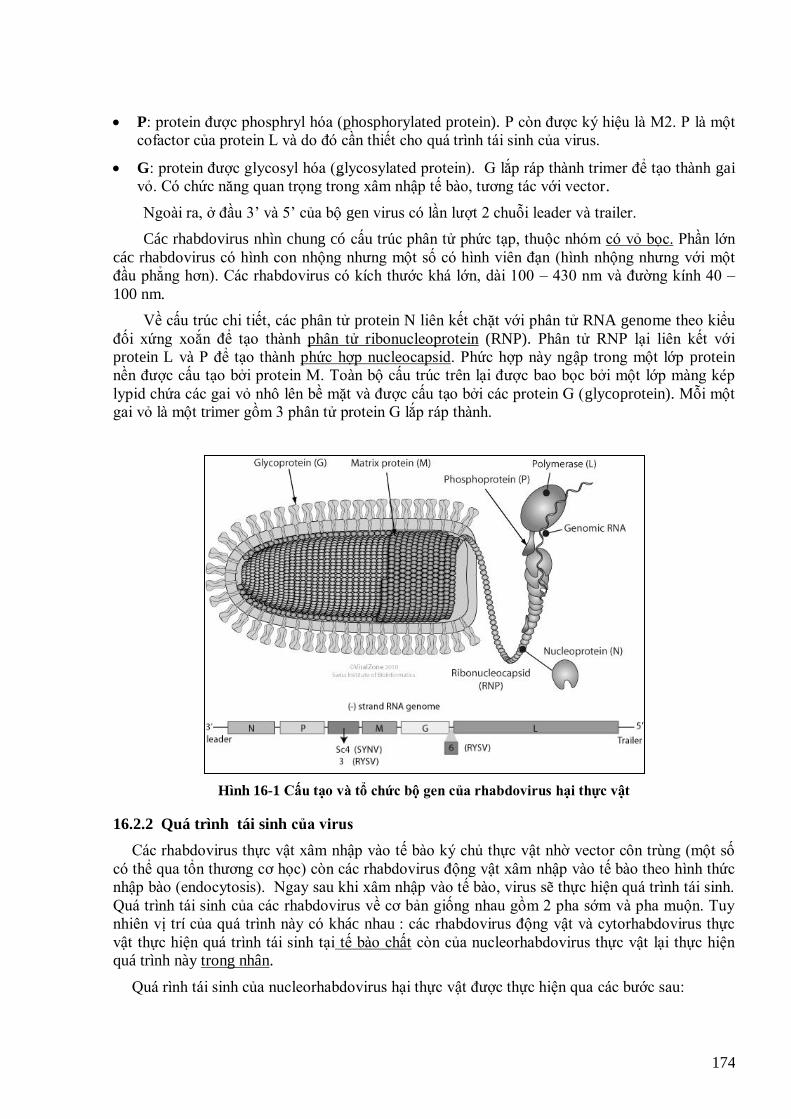
174
P: protein đƣợc phosphryl hóa (phosphorylated protein). P còn đƣợc ký hiệu là M2. P là một
cofactor của protein L và do đó cần thiết cho quá trình tái sinh của virus.
G: protein đƣợc glycosyl hóa (glycosylated protein). G lắp ráp thành trimer để tạo thành gai
vỏ. Có chức năng quan trọng trong xâm nhập tế bào, tƣơng tác với vector.
Ngoài ra, ở đầu 3‘ và 5‘ của bộ gen virus có lần lƣợt 2 chuỗi leader và trailer.
Các rhabdovirus nhìn chung có cấu trúc phân tử phức tạp, thuộc nhóm có vỏ bọc. Phần lớn
các rhabdovirus có hình con nhộng nhƣng một số có hình viên đạn (hình nhộng nhƣng với một
đầu phẳng hơn). Các rhabdovirus có kích thƣớc khá lớn, dài 100 – 430 nm và đƣờng kính 40 –
100 nm.
Về cấu trúc chi tiết, các phân tử protein N liên kết chặt với phân tử RNA genome theo kiểu
đối xứng xoắn để tạo thành phân tử ribonucleoprotein (RNP). Phân tử RNP lại liên kết với
protein L và P để tạo thành phức hợp nucleocapsid. Phức hợp này ngập trong một lớp protein
nền đƣợc cấu tạo bởi protein M. Toàn bộ cấu trúc trên lại đƣợc bao bọc bởi một lớp màng kép
lypid chứa các gai vỏ nhô lên bề mặt và đƣợc cấu tạo bởi các protein G (glycoprotein). Mỗi một
gai vỏ là một trimer gồm 3 phân tử protein G lắp ráp thành.
Hình 16-1 Cấu tạo và tổ chức bộ gen của rhabdovirus hại thực vật
16.2.2 Quá trình tái sinh của virus
Các rhabdovirus thực vật xâm nhập vào tế bào ký chủ thực vật nhờ vector côn trùng (một số
có thể qua tổn thƣơng cơ học) còn các rhabdovirus động vật xâm nhập vào tế bào theo hình thức
nhập bào (endocytosis). Ngay sau khi xâm nhập vào tế bào, virus sẽ thực hiện quá trình tái sinh.
Quá trình tái sinh của các rhabdovirus về cơ bản giống nhau gồm 2 pha sớm và pha muộn. Tuy
nhiên vị trí của quá trình này có khác nhau : các rhabdovirus động vật và cytorhabdovirus thực
vật thực hiện quá trình tái sinh tại tế bào chất còn của nucleorhabdovirus thực vật lại thực hiện
quá trình này trong nhân.
Quá rình tái sinh của nucleorhabdovirus hại thực vật đƣợc thực hiện qua các bƣớc sau:

175
1. Ngay sau khi xâm nhập vào tế bào, phân tử virus gắn vào mạng lƣới nội chất và giải phóng
phức hợp nucleocapsid (gồm RNA-N-L-P) vào tế bào chất.
2. Phức hợp nucleocapsid xâm nhập nhân tế bào nhờ các dấu hiệu nhập nhân NLS có mặt ở
phân tử N, L và P.
3. Tại nhân tế bào, quá trình phiên mã sơ cấp (pha sớm) xảy ra : protein L của virus (là một
RdRp) phiên mã các mRNA của các protein sớm (N, L, P) của virus trên khuôn RNA virus.
Các mRNA này đƣợc vận chuyển ra tế bào chất để dịch mã thành các protein N, L, P.
4. Tiếp theo, các protein N, L, P lại đƣợc vận chuyển trở lại nhân và bắt đầu quá trình phiên mã
thứ cấp (pha muộn) tổng hợp tất cả các mRNA của virus cũng nhƣ bắt đầu quá trình tái bản
bộ gen (tổng hợp RNA sợi (+) => làm khuôn để tổng hợp sợi (-) là sợi virus ; bản thân sợi (-)
lại làm khuôn để tổng hợp mRNA virus).
5. Tại màng trong của nhân, các protein virus cảm ứng để hình thành cấu trúc viroplasms chứa
protein N, P, L của virus. Viroplasm là nơi thực hiện quá trình tái sinh và lắp ráp các thành
phần virus.
6. Vào giai đoạn cuối của quá trình tái sinh, protein M sẽ liên kết và gấp xoắn các phân tử
nucleocapsid (RNA-N-P-L) mới đƣợc lắp ráp. Phức hợp nucleocapsid-M sẽ nảy chồi qua
màng trong của nhân để hình thành phân tử virus hoàn chỉnh và tụ tập tại vùng ngoại vi của
nhân (khoảng không giữa màng trong và màng ngoài của nhân)
16.3 RICE YELLOW STUNT VIRUS (RYSV)
16.3.1 Giới thiệu
Bệnh vàng lùn ―rice yellow stunt disease‖ hay còn đƣợc gọi là bệnh vàng tạm thời ―rice
transitory yellowing disease‖ đã đƣợc phát hiện thấy tại nhiều nƣớc trồng lúa thuộc khu vực châu
Á và Đông Nam Á từ những năm 60. Tại Việt Nam, bệnh vàng tạm thời còn đƣợc dịch là bệnh
vàng lá di động.
Hai mẫu virus đƣợc gọi là Rice yellow stunt virus (RYSV) gây bệnh vàng lùn và Rice
transitory yellowing virus (RTYV) gây bệnh vàng tạm thời đã đƣợc giải trình tự toàn bộ bộ gen.
So sánh trình tự của 2 virus này đã chứng tỏ chúng chỉ là thành viên của cùng 1 loài (Hiraguri et
al., 2009). Vì vậy chúng ta sẽ sử dụng tên RYSV để chỉ tác nhân gây bệnh.
Tại Trung Quốc, bệnh vàng lùn đã gây thành dịch tại nhiều tỉnh phía Nam trong những năm
60 và 70 nhƣ Đài Loan (1960 – 1962, 1973 – 1980), Quảng Đông (1964-1966, 1979), Phúc Kiến
(1966, 1969, 1973), Chiết Giang (1970 – 1973). Tại Việt Nam, trong cùng thời gian, 1 bệnh
biến vàng trên lúa gọi là bệnh “vàng lụi‖ do rầy xanh đuôi đen truyền đã gây thành dịch tại
nhiều tỉnh phía Bắc, đặc biệt tại các tỉnh miền núi.
Mặc dù mẫu bệnh vàng lụi của Việt Nam không đƣợc lƣu giữ nhƣng đối chiếu bệnh vàng lùn
của Trung Quốc và bệnh vàng lụi của Việt Nam thấy có nhiều điểm tƣơng đồng nhƣ: (i) triệu
chứng giống nhau, (ii) sự xuất hiện các vụ dịch cùng thời gian, tại các khu vực gần gũi về mặt
địa lý, (iii) cùng vector rầy xanh đuôi đen, (iv) sự có mặt của 1virus hình viên đạn trên mẫu bệnh
Việt Nam
Trong các năm 2009, 2010, một bệnh vàng lá lúa đã gây thành dịch nghiêm trọng tại một số
địa phƣơng miền Bắc nhƣ Hà Tây, Bắc Giang. Kiểm tra bằng RT-PCR và giải trình tự cho thấy
tác nhân gây bệnh là RYSV.

176
16.3.2 Virus
RYSV có hình viên đạn, dài 180–210 nm, đƣờng kính 94 nm. Virus có bộ gen RNA sợi đơn,
cực âm, không phân mảnh, kích thƣớc ~ 14 kb.
Virus có 1 chuỗi nucleotide khoảng 203 nucleotide gọi là leader ở đầu 3‘, một chuỗi 181
nucleotide gọi là trailer ở đầu 5‘. Virus mã hóa 7 protein, theo thứ tự từ trái sang phải là N-P-3-
M-G-6-L. Tuy nhiên chỉ có 5 protein cấu trúc có mặt ở phân tử virus là N, P, M, G, L (giống nhƣ
ở các rhabdovirus khác).
Hình 16-2 Sơ đồ tổ chức bộ gen của RYSV
16.3.3 Triệu chứng bệnh
Lá biến vàng, đẻ nhánh giảm, cây lùn.
Hai đến ba tuần sau cấy, cây bệnh điển hình có 1 – 2 lá phía dƣới bị biến vàng sau
chuyển màu vàng sáng hoặc vàng cam. Cuối cùng, các lá này trở nên nhăn, héo.
Trên giống mẫn cảm, cây bị lùn và đẻ nhánh giảm mạnh, trỗ bông giảm hoặc không trỗ.
Cây nhiễm sớm biểu hiện triệu chứng rõ nhƣng cây nhiễm muộn có thể không biểu hiện
triệu chứng. Trƣờng hợp nặng, cây có thể chết trƣớc khi trỗ.
Trong điều kiện nhà kính, cây bệnh biểu hiện khả năng phục hồi triệu chứng tốt hơn
ngoài đồng ruộng. Sau khi biểu hiện triệu chứng vàng lá điển hình thì cây bệnh dần phục
hồi thậm chí không biểu hiện triệu chứng. Thông thƣờng, các dảnh ―trông bình thƣờng‖
có thể hình thành từ 1 cây bị bệnh. Vì lý do này nên bệnh mới đƣợc đặt tên là ―biến vàng
tạm thời – transitory yellowing‖.
Hinh 16-3 Bệnh vàng lụi lúa tại miền Bắc năm 2010 do RYSV
16.3.4 Lan truyền
Virus RYSV thuộc nhóm gây hại ở bó mạch phloem. Trong tế bào nhiễm bệnh, các phân tử
virus tập trung ở ngoại vi nhân (giữa màng trong và màng ngoài nhân).
Ký chủ thực vật tự nhiên duy nhất của virus là lúa. Sau khi thu hoạch, virus tồn tại trên lúa
chét hoặc trong vector rầy xanh.
RYSV -AB011257, kích thước 14042 nucleotides
N P 3 M G 6 L Trailer Leader
3’ 5’

177
RYSV lan truyền theo kiểu bền vững tái sinh (nhân lên trong vector), mặc dù không truyền
qua trứng, bằng 3 loài rầy xanh trên lúa là: Nephotettix nigropictus, N. cincticeps và N. virescens
Thí nghiệm so sánh hiệu quả truyền của 3 loài rầy (Inoue, 1978) cho thấy:
Hiệu quả truyền bệnh cao nhất là của N. nigropictus, tiếp theo là N. cincticeps và thấp nhất là
của N. virescens. Thí nghiệm của các tác giả khác cũng cho kết quả tƣơng tự.
Nhìn chung, đối với rầy N. nigropictus, hiệu quả truyền của rầy đực cao hơn rầy cái; còn đối
với 2 loài rầy còn lại thì không có sự khác biệt đáng kể.
Theo Chiu et al. (1965), Chiu & Jane (1967), khi nghiên cứu cả 3 loài:
Khoảng 41 – 62 % cá thể rầy có khả năng truyền virus
Thời gian chích nạp của đa số (>50%) rầy là 1 vài giờ. Nếu tăng thời gian chích nạp lên 1
ngày thì 90 % rầy có khả năng truyền. Thời kỳ ẩn trong vector 3 – 34 ngày nhƣng thƣờng 9 –
16 ngày.
Thời gian chích nạp thay đổi từ 5 - 10 phút, 5 – 10 giờ, 24 giờ sẽ làm tăng tƣơng ứng hiệu quả truyền từ 28 %, 45 %, 88 %.
Thời kỳ ủ bệnh trong cây 2 – 4 tuần
Nghiên cứu với rầy N. nigropictus, Hsieh (1969) cho biết:
Nhiệt độ < 16 oC hoặc > 38 oC, rầy không có khả năng truyền virus
Trong khoảng nhiệt độ 20 – 36 oC, nhiệt độ càng cao thì thời gian ẩn càng rút ngắn.
Theo Chen (1979), Chen & Chiu (1980):
Cho rầy chích nạp trên lá biến vàng hay trên giống mẫn cảm thì hiệu quả truyền tƣơng ứng là
28.8 % và 34.4 %. Còn khi cho rầy chích nạp trên lá đã phục hồi hoặc trên giống kháng thì hiệu quả truyền chỉ tƣơng ứng 3.3 % và 9%.
Rầy đực và rầy cám có khả năng truyền virus hiệu quả hơn rầy cái.
Mỗi rầy mang virus có thể truyền bệnh cho 3 cây lúa/ ngày
Khi nhiễm virus, rầy có khả năng truyền cả đời.
Virus ảnh hƣởng bất lợi đến rầy: làm rầy cám có thể chết trƣớc khi khi trƣởng thành, giảm số trứng đẻ, rút ngắn vòng đời
Bảng 16-2 So sánh khả năng truyền RYSV của 3 loài rầy xanh trên lúa
Chỉ tiêu Tham khảo N. nigropictus N. cincticeps N. virescens
Hiệu quả truyền (%) Inoue (1978) 35 - 75 27 - 59 0 - 5
Chiu et al. 1968) 41 - 65 25 - 36 47
Thời giân ẩn (ngày)
Inoue (1978) 7 - 17 7 - 22 17 -25
Chiu et al. 1968) 9 - 16 25 - 34
Hsieh (1970) 4 - 20 4 - 20
Thời gian tồn tại khả nhiễm (ngày) Inoue (1978) 30 28 10
Ghi chú: các số liệu của Chiu et al. và Hsieh được dẫn theo Ou (1985).
16.3.5 Phòng trừ
Biện pháp phòng trừ hiện tại:

178
Bảo vệ mạ, đặc biệt vụ mùa
Điều tra ruộng thƣờng xuyên, nhổ bỏ cây bệnh
Kiểm soát vector rầy xanh đuôi đen
Chọn tạo giống kháng rầy, kháng virus
16.4 CÂU HỎI ÔN TẬP CHƢƠNG 16
1. Đặc điểm họ Rhabdoviridae (phân loại, hình thái, quan hệ vector, bản chất bộ gen). So
sánh điểm khác biệt cơ bản của các rhabdovirus thực vật với các virus thực vật và động
vật khác
2. RYSV (tầm quan trọng, triệu chứng, đặc điểm bộ gen, lan truyền, phòng chống).
16.5 TÀI LIỆU THAM KHẢO CHÍNH
1. Hà Viết Cƣờng, Lê Văn Hải và Vũ Triệu Mân (2010). Xác định tác nhân gây bệnh vàng
lá lúa tại tỉnh Bắc Giang vụ mùa năm 2010. Tạp chí BVTV. Số 4: 8-12.
2. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
3. Hibino, H. (1996). Biology and epidemiology of rice viruses. Annual Review of
Phytopathology 34, 249-274.
4. Hiraguri, A., Hibino, H., Hayashi, Shimizu, T.T., Uehara-Ichiki, T., Omura, T. & Sasaya, T. (2009). Complete sequence analysis of rice transitory yellowing virus and its comparison to rice yellow stunt virus. Archives of Virology. 155:243–245
5. Inoue, H. (1978). Transmission efficiency of rice transitory yellowing virus by the green plant hoppers, Nephotettix spp. (Hemiptera: Cicadellidae). Applied Entomology and
Zoology. 14:123-126
6. Jackson, A. O., Dietzgen, R. G., Goodin, M. M., Bragg, J. N. & Deng, M. (2005).
Biology of Plant Rhabdoviruses. Annual Review of Phytopathology 43, 623–660.
7. Ou, S.H. (1985). Rice Diseases. Second Edition, p. 23-25 & 48-49. Commonwealth Mycological Institute Publication. Kew, Surrey, UK.
8. Shikata, E. & Chen, M.J. (1969) Electron microscopy of rice transitory yellowing virus. Journal of Virology. 3:261–264
9. Shikata, E. (1972). Rice transitory yellowing virus. Description of Plant Viruses. CMI/AAB No 100.

179
Chƣơng 17. VIRUS RNA SỢI ĐƠN CỰC ÂM (TIẾP): TOSPOVIRUS
17.1 TÓM TẮT NỘI DUNG
Chƣơng 17 mô tả phân loại và các đặc trƣng phân tử của họ Bunyaviridae, đại diện cho nhóm
các virus RNA, sợi đơn, cực âm và lƣỡng cực. Chƣơng này cũng đề cập đến một virus gây hại
phổ biến trên nhiều loài cây khắp thế giới là TSWV (tomato spotted wilt virus).
17.2 HỌ BUNYAVIRIDAE
Họ này gồm 95 loài đƣợc phân loại vào 5 chi trong đó 4 chi hại động vật và chỉ có 1 chi hại
thực vật là Tospovirus. Các bunyavirus hại động vật (ví dụ Hantaan virus gây bệnh sốt xuất
huyết Hataan) đều lan truyền qua vector côn trùng.
Các bunyavirus có virion hình khối đa diện, kích thƣớc 80 – 120 nm. Phân tử virus có vỏ
bọc. Vỏ có mấu nhỏ cấu tạo bằng glycoprotein. Bộ gen virus gồm 3 phân tử RNA sợi đơn cực (-)
hoặc lƣỡng cực lý hiệu là L (large), M (medium) và S (small) có tổng kích thƣớc từ 11 – 19 kb.
Tất cả 3 phân tử đều đƣợc lắp ráp trong 1 phân tử virus. Các nucleotide ở 2 đầu của mỗi phân tử
RNA bảo thủ trong mỗi chi nhƣng khác nhau giữa các chi. Trình tự ở các đầu này bổ sung nhau
nên có thể bắt cặp với nhau tạo phân tử RNA mạch vòng giả.
Bảng 17-1. Phân loại họ Bunyaviridae theo ICTV (2009)
TT Chi Loài điển hình Ký chủ Số loài
1 Hantavirus Hantaan virus Động vật 23
2 Nairovirus Dugbe virus Động vật 7
3 Orthobunyavirus Bunyamwera virus Động vật 48
4 Phlebovirus Rift Valley fever virus Động vật 9
5 Tospovirus Tomato spotted wilt virus Thực vật 8
Tổng số 95
17.3 CHI TOSPOVIRUS
Hiện có khoảng 8 tospovirus hại thực vật đã đƣợc ghi nhận, trong đó virus điển hình và có ý
nghĩa kinh tế là Tomato spotted wilt virus (TSWV).
Các tospovirus đều lan truyền ngoài từ nhiên nhờ bọ trĩ theo kiểu bền vững tái sinh.
Các tospovirus hại thực vật có hình thái phân tử và tổ chức bộ gen giống các bunyavirus khác
hại động vật. Chúng có phân tử hình cầu, kích thƣớc từ 80 – 120 nm. Phân tử có vỏ bọc, vỏ có
mấu nhỏ cấu tạo bằng glycoprotein nhô lên trên bề mặt. Tất cả các tospovirus có bộ gen phân
mảnh gồm 3 phân tử RNA sợi đơn cực (-) hoặc lƣỡng cực ký hiệu là L (large), M (medium) và S
(small) có tổng kích thƣớc từ 11 – 19 kb. Tất cả 3 phân tử đều đƣợc lắp ráp trong 1 phân tử virus.

180
Các nucleotides ở 2 đầu của mỗi phân tử RNA bảo thủ trong mỗi chi nhƣng khác nhau giữa
các chi. Trình tự ở các đầu này có thể bắt cặp với nhau tạo phân tử RNA mạch vòng giả.
17.4 TOMATO SPOTTED WILT VIRUS (TSWV)
17.4.1 Tầm quan trọng
Tomato spotted wilt virus (TSWV) (virus đốm héo cà chua) là virus quan trọng nhất của chi
Tospovirus.
TSWV phân bố khắp thế giới và là virus thực vật có phổ ký chủ lớn nhất, nhiễm hơn 940 loài
cây thuộc 8 họ cây một lá mầm và 90 họ cây hai lá mầm.
TSWV gây thiệt đáng kể trên một loạt các cây trồng quan trọng nhƣ lạc, đậu đỗ, khoai lang,
thuốc lá, đu đủ, cà chua, ớt và nhiều cây cảnh.
17.4.2 Virus
17.4.2.1 Hình thái
Virion có hình khối đa diện (hình cầu), kích thƣớc từ 80 – 120 nm. Phân tử có vỏ bọc. Vỏ có
mấu nhỏ cấu tạo bằng glycoprotein (kích thƣớc 5-10 nm), phần cuống mấu nằm chìm trong một
lớp màng kép lipid (dày khoảng 5 nm) có nguồn gốc từ màng tế bào ký chủ (vector hoặc cây).
Bên trong lớp vỏ là lõi virus gồm 3 phân tử RNA dạng mạch vòng giả do 2 đầu của mỗi phân
tử có thể tự bắt cặp. Các phân tử RNA genome này liên kết với các phân tử protein N
(nucleoprotein). Các phân tử RNA genome liên kết protein N này đƣợc gọi là các phân tử RNP
(Ribonucleocapsid). Ngoài ra phần lõi phân tử virus cũng chứa một số lƣợng nhỏ protein L (là
RdRp của virus).
Cần chú ý là các phân tử ribonucleocapsid này có khả năng xâm nhiễm nếu đƣợc đƣa vào tế
bào. Trái lại chỉ các phân tử virion nguyên vẹn (có vỏ bọc và mấu) mới có khả năng truyền qua
vector bọ trĩ.
Phân tử virus chứa khoảng 65% protein, 20% lipid, 7% carbohydrate và 5% RNA.
Hình 17-1 Hình thái và cấu trúc phân tử TSWV

181
17.4.3 Tổ chức bộ gen và tái sinh
Tổ chức bộ gen của TSWV giống nhƣ của các tospovirus khác gồm 3 phân tử RNA sợi đơn
cực (-) hoặc lƣỡng cực ký hiệu là L (large), M (medium) và S (small) có tổng kích thƣớc ~ 17
kb, cụ thể là:
Sợi L: 8897 nucleotides, có cực (-), chứa 1 ORF lớn, mã hóa protein L trên sợi đối virus
Sợi M: 4821 nucleotides, lƣỡng cực, chứa 2 ORF mã hóa 3 protein là NSm trên sợi virus
và Gc (G2) và Gn (G1) trên sợi đối virus. NSm là viết tắt của ―Non-structural protein on
M segment‖ có nghĩa là protein phi cấu trúc đƣợc mã hóa trên sợi M.
Sợi S: 2916 nucleotides, lƣỡng cực, chứa 2 ORF mã hóa 2 protein là NSs trên sợi virus
và N trên sợi đối virus. NSm là viết tắt của ―Non-structural protein on S segment‖ có
nghĩa là protein phi cấu trúc đƣợc mã hóa trên sợi S.
Tất cả 3 phân tử RNA genome đều đƣợc lắp ráp trong 1 phân tử virus.
Hai đầu của mỗi phân tử RNA genome có trình tự bổ sung khoảng 65 – 70 nucleotides dẫn tới
chúng có thể tự bắt cặp từ đầu mút 5‘ và 3‘ trông giống ―cán chảo‖. Hậu quả mặc dù các phân tử
genome là mạch thẳng nhƣng chúng tạo cấu trúc dạng ―vòng giả‖. Tám nucleotides ở đầu mút 5‘
và 3‘ của trình tự bổ sung này bảo thủ ở cả 3 phân tử. Đây là một đặc điểm giống nhau của các
virus có bộ gen RNA sợi âm, phân đoạn ở nhiều họ khác nhƣ tenuivirus, reovirus.
Hình 17-2 Sơ đồ quá trình tái sinh của TSWV
17.4.4 Chức năng protein
Phân tử virus (virion) của TSWV và các tospovirus có 4 protein cấu trúc là G1, G2, N và L.
G1 và G2 là các protein liên kết với đƣờng nên còn gọi là glycoprotein hình thành nên các gai
mấu trên bề mặt phân tử virus. G1 và G2 cũng đƣợc ký hiệu Gn hoặc Gc tùy thuộc vị trí của nó ở
đầu amin hay đầu carboxyl của chuỗi polyprotein trên sợi RNA M. Các glycoprotein này có vai
trò quan trọng trong lan truyền qua vector bọ trĩ: chúng liên kết với các receptor bề mặt của ruột
giữa bọ trĩ

182
Protein N là các nucleoprotein đƣợc mã hóa trên sợi RNA S. Protein N sẽ liên kết với các sợi
RNA của virus để tạo thành các phân tử nucleocapsid.
Protein L là một protein có kích thƣớc lớn (L= large) và đƣợc mã hóa trên sợi RNA L. Protein
L là một RdRp của virus chịu trách nhiệm tái bản RNA và phiên mã tạo mRNA. Protein L có
mặt ở phần lõi (bên trong virion), nhƣng số lƣợng ít hơn nhiều so với protein N.
Virus cũng mã hóa 2 protein phi cấu trúc là NSm đƣợc mã hóa trên sợi RNA M và NSs đƣợc
mã hóa trên sợi RNA S. NSm có vai trò trong di chuyển giữa các tế bào của phân tử virus qua
sợi liên bào (kích thƣớc các tospovirus quá lớn để có thể di chuyển tự do qua sợi liên bào). NSs
có chức năng ức chế phản ứng phòng thủ của tế bào ký chủ qua cơ chế câm gen.
17.4.5 Triệu chứng
TSWV gây ra nhiều triệu chứng khác nhau. Các triệu chứng này thay đổi theo giống cây ký
chủ, trạng thái sinh lý cây ký chủ (tuổi, điều kiện dinh dƣỡng và môi trƣờng). Các chủng virus
không khác nhau nhiều về đặc tính gây bệnh mà chủ yếu khác nhau về đặc trƣng phân tử và
huyết thanh học. Nhìn chung, triệu chứng ít có giá trị chẩn đoán bệnh do TSWV và các
tospovirus khác.
Trên cà chua. Các triêu chứng chính là: các đốm hoặc sọc chết hoại trên lá, cuống lá, thân,
đỉnh sinh trƣởng; lá nhăn, hóa màu nâu đồng thau; cây nhỏ và còi cọc. Trên quả, hàm lƣợng
carotene và sắc tố giảm; quả chín có các đốm màu vàng hoặc đỏ nhạt (đốm ma). Đôi khi, triệu
chứng chỉ thấy trên quả.
Trên ớt (ngọt). Triệu chứng chủ yếu là cây còi cọc và biến vàng. Lá có thể có đốm chết hoại,
đốm vòng, hoặc khảm. Trên thân có các sọc chết hoại kéo tới chồi đỉnh. Trên quả chín có các
đốm vòng đồng tâm hoặc các sọc chết hoại.
Hình 17-3 Một số bệnh do TSWV gây ra trên cà chua, thuốc lá và lạc

183
17.4.6 Lan truyền
TSWV lan truyền ngoài tự nhiên bằng nhiều loài bọ trĩ thuộc 2 chi Frankliniella và Thrips (họ
Thripidae, bộ Thysanoptera) theo kiểu bền vững tái sinh. Tám loài bọ trĩ đƣợc biết là vector của
TSWV bao gồm Frankliniella occidentalis, F. fusca (bọ trĩ thuốc lá), F. intonsa (bọ trĩ hoa), F.
schultzei (bọ trĩ bông), F. bispinosa, Thrips palmi (bọ trĩ dƣa), Thrips tabaci (bọ trĩ hành) và T.
setosus. Trong số các loài này, F. occidentalis có thể đƣợc coi là vector quan trọng nhất. Từ năm
1980, sự phát tán nhanh chóng của loài bọ trĩ này khắp thế giới đã tạo điều kiện để virus phân bố
rộng hơn.
TSWV truyền bằng bọ trĩ theo kiểu bền vững tái sinh khá đặc biệt. Chỉ bọ trĩ non tuổi 1 và
tuổi 2 mới có thể chích nạp virus và chỉ giai đoạn tuổi 2 và trƣởng thành mới có khả năng truyền
virus. Đối với F. occidentalis, thời gian chích nạp và chích truyền khoảng 60 phút và thời gian
ẩn trung bình 3 – 7 ngày. Virus duy trì khả năng lây nhiễm cả đời nhƣng chƣa có bằng chứng
cho thấy virus truyền qua trứng.
Cả 2 loại protein bề mặt là Gn và Gc đều cần cho sự lan truyền của virus qua vector, trong đó
Gc giúp virus bám vào thành ruột trong qua trình chích nạp.
TSWV không truyền qua hạt giống.
17.4.7 Phòng chống
Biện pháp phòng chống TSWV hiện nay là kết hợp 3 biện pháp:
1. Phòng chống vector bọ trĩ. Tuy nhiên vì bọ trĩ hiện đã kháng hầu hết các loại thuốc trừ
sâu nên việc phòng chống vector bọ trĩ cực kỳ khó.
2. Phòng chống cỏ dại. Do TSWV có phổ ký chủ cực kỳ lớn nên cỏ dại cũng là nguồn
bệnh rất quan trọng.
3. Sử dụng giống kháng. Hiện nay, chỉ có một số ít gen kháng TSWV đã đƣợc xác định, ví
dụ gen SW5. Ngoài ra, dùng giống kháng mang gen kháng từ virus nhƣ vùng mã hóa và
không mã hóa của gen N và NSm cũng đang đƣợc thử nghiệm.
17.5 CÂU HỎI ÔN TẬP CHƢƠNG 17
1. Đặc điểm virus họ Bunyaviridae (phân loại, hình thái, bản chất bộ gen). So sánh điểm khác
biệt cơ bản của các tospovirus với các virus thực vật và động vật khác
2. TSWV (tầm quan trọng, triệu chứng, đặc điểm bộ gen, lan truyền, phòng chống).
17.6 TÀI LIỆU THAM KHẢO CHÍNH
1. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.
2. Moritz, G., Kumm, S. & Mound, a. (2004a). Tospovirus transmission depends on thrips ontogeny. Virus Research 100 143–149.
3. Moritz, G., Kumma, S. & Moundb, L. (2004b). Tospovirus transmission depends on thrips
ontogeny. Virus Research 100, 143–149.
4. Rosello, S., Diez, M. J. & Nuez, F. (1996). Viral diseases causing the greatest economic losses tomato crop. I. The Tomato spotted wilt virus. Scientia Horticuhurae 67 117- 150.
5. Whitfield, A. E., Ullman, D. E. & German, T. L. (2005). Tospovirus-Thrips Interactions.
Annual Review of Phytopathology 43, 459–489.

184
Chƣơng 18. CÁC VIRUS RNA SỢI KÉP: HỌ REOVIRIDAE
18.1 TÓM TẮT NỘI DUNG
Chƣơng 18 mô tả phân loại và các đặc trƣng phân tử của họ Reoviridae, đại diện cho nhóm
các virus RNA sợi kép, có bộ gen phân đoạn, cùng đƣợc lắp ráp trong một virion. Chƣơng này
cũng đề cập đến 2 reovirus rất quan trọng đối với sản xuất lúa của Việt Nam là RRSV (Rice
ragged stunt virus) và SRBSDV (Southern rice black streaked dwarf virus).
18.2 HỌ REOVIRIDAE
18.2.1 Phân loại
Các virus thuộc họ Reoviridae (đƣợc gọi chung là các reovirus) là một nhóm các virus rất đa
dạng về ký chủ. Trong số 15 chi virus thì tới 11 chi hại động vật, 1 chi hại nấm và 3 chi hại thực
vật là Phytoreovirus, Oryzaevirus và Fijivirus. Các reovirus hại thực vật đều gây hại cây một lá
mầm, phần lớn họ hòa thảo, đặc biệt lúa. Chúng đều lan truyền ngoài tự nhiên bằng rầy theo kiểu
bền vững tái sinh. Nhiều reovirus hại thực vật là các virus quan trọng trên lúa, chẳng hạn ở Việt
Nam là virus lùn xoắn lá (RRSV) và virus lùn sọc đen phƣơng Nam (SRBSDV) gây bệnh lúa lùn
sọc đen mới đƣợc phát hiện năm 2009 ở miền Bắc.
Cho tới 2009, đã có 84 reovirus đƣợc ghi nhận trên thế giới, trong đó có 14 virus hại thực
vật thuộc các chi Fijivirus, Phytoreovirus và Oryzaevirus (Bảng 18-1)
Bảng 18-1 Phân loại họ Reoviridae (các reovirus đƣợc ghi nhận đến 2009)
TT Chi Loài chuẩn (gạch chân) Ký chủ Số loài
1 Cardoreovirus Eriocheir sinensis reovirus Động vật (biển) 1
2 Mimoreovirus Micromonas pusilla reovirus Động vật (biển) 1
3 Orbivirus Bluetongue virus Động vật 22
4 Rotavirus Rotavirus A Động vật 5
5 Seadornavirus Banna virus Động vật 3
6 Aquareovirus Aquareovirus A Động vật biển 7
7 Coltivirus Colorado tick fever virus Động vật 2
8 Cypovirus Cypovirus 1 Động vật (côn trùng) 16
9 Dinovernavirus Aedes pseudoscutellaris reovirus
10 Idnoreovirus Idnoreovirus - 1 Động vật (cô trùng) 5
11 Mycoreovirus Mycoreovirus 1 Nấm 3
12 Orthoreovirus Mammalian orthoreovirus Động vật 5
13 Phytoreovirus
Wound tumor virus,
Rice dwarf virus,
Rice gall dwarf virus Thực vật 3
14 Fijivirus
Fiji disease virus
Garlic dwarf virus
Maize rough dwarf virus
Mal de Rio Cuarto virus
Nilaparvata lugens reovirus
Oat sterile dwarf virus Pangola stunt virus
Rice black streaked dwarf virus
Southern rice black streaked virus
Thực vật 9
15 Oryzavirus Rice ragged stunt virus,
Echinochloa ragged stunt virus Thực vật 2

185
18.2.2 Hình thái
Virion của các reovirus đều có hình cầu đa diện đối xứng (icosahedral), kích thƣớc từ 60 –
80 nm. Lớp vỏ protein đƣợc cấu tạo thành 1, 2 hoặc 3 lớp đồng tâm. Dựa vào cấu trúc lớp vỏ,
các reovirus đƣợc chia làm 2 nhóm (Hình):
(i) Có gai tƣơng đối lớn (spike) trên trên bề mặt (ví dụ chi Fijivirus, Oryzavirus, Cypovirus,
Orthoreovirus, Aquareovirus, Idnoreovirus và Mycoreovirus)
(ii) Không có gai (ví dụ chi Phytoreovirus, Rotavirus, Orbivirus, Coltivirus và
Seadornavirus)
Hình 18-1 Cấu trúc của một số reovirus đại diện
18.2.3 Tổ chức bộ gen
Các reovirus là các virus RNA có bộ gen phân đoạn gồm là 10 – 12 phân tử RNA sợi kép
mạch thẳng. Tất cả các phân tử RNA này đều đƣợc lắp ráp trong một virion. Đầu 5‘ của sợi (+)
của mỗi phân tử RNA sợi kép đƣợc mũ hóa (Cap). RNA virus thiếu một chuỗi polyadenine ở đầu
3‘.
18.2.4 Protein
Virion chứa ít nhất 3 protein có hoạt tính enzyme liên quan đến tổng hợp RNA và tạo mũ:
RNA dependent RNA polymerase (RdRp) với vai trò kép: là 1 enzyme phiên mã
(transcriptase) tổng hợp sợi (+) = mRNA trên khuôn RNA sợi kép hoặc là 1 enzyme
tái sinh (replicase) tổng hợp sợi (-) trên khuôn sợi (+)
Transmethylase: tạo mũ (cap) đầu 5‘ của RNA
Helicase: tháo xoắn phân tử RNA sợi kép
Virion chứa ít nhất 3 protein chính liên quan đến cấu tạo vỏ protein (major capsid proteins)
Virion cũng chứa một số protein phụ với chức năng chƣa rõ.
Các protein có kích thƣớc 15 – 155 kDa và chiếm 80-85% khối lƣợng virion
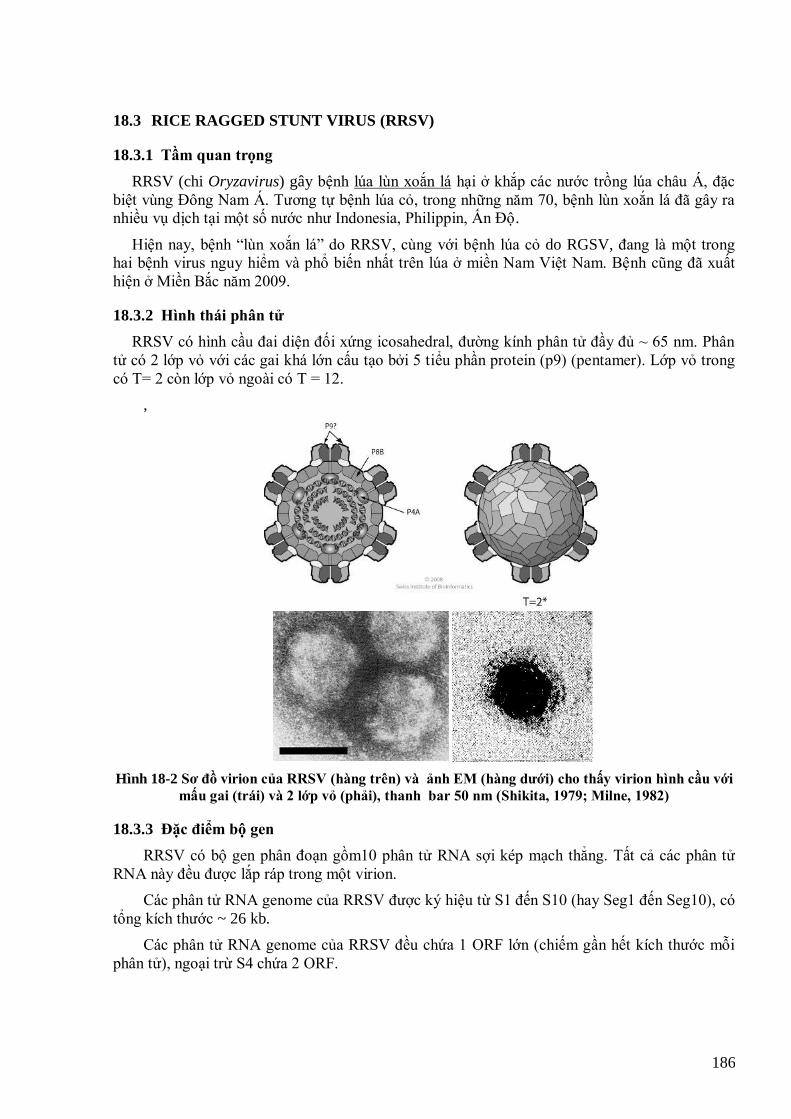
186
18.3 RICE RAGGED STUNT VIRUS (RRSV)
18.3.1 Tầm quan trọng
RRSV (chi Oryzavirus) gây bệnh lúa lùn xoắn lá hại ở khắp các nƣớc trồng lúa châu Á, đặc
biệt vùng Đông Nam Á. Tƣơng tự bệnh lúa cỏ, trong những năm 70, bệnh lùn xoắn lá đã gây ra
nhiều vụ dịch tại một số nƣớc nhƣ Indonesia, Philippin, Ấn Độ.
Hiện nay, bệnh ―lùn xoắn lá‖ do RRSV, cùng với bệnh lúa cỏ do RGSV, đang là một trong
hai bệnh virus nguy hiểm và phổ biến nhất trên lúa ở miền Nam Việt Nam. Bệnh cũng đã xuất
hiện ở Miền Bắc năm 2009.
18.3.2 Hình thái phân tử
RRSV có hình cầu đai diện đối xứng icosahedral, đƣờng kính phân tử đầy đủ ~ 65 nm. Phân
tử có 2 lớp vỏ với các gai khá lớn cấu tạo bởi 5 tiểu phần protein (p9) (pentamer). Lớp vỏ trong
có T= 2 còn lớp vỏ ngoài có T = 12.
,
Hình 18-2 Sơ đồ virion của RRSV (hàng trên) và ảnh EM (hàng dƣới) cho thấy virion hình cầu với
mấu gai (trái) và 2 lớp vỏ (phải), thanh bar 50 nm (Shikita, 1979; Milne, 1982)
18.3.3 Đặc điểm bộ gen
RRSV có bộ gen phân đoạn gồm10 phân tử RNA sợi kép mạch thẳng. Tất cả các phân tử
RNA này đều đƣợc lắp ráp trong một virion.
Các phân tử RNA genome của RRSV đƣợc ký hiệu từ S1 đến S10 (hay Seg1 đến Seg10), có
tổng kích thƣớc ~ 26 kb.
Các phân tử RNA genome của RRSV đều chứa 1 ORF lớn (chiếm gần hết kích thƣớc mỗi
phân tử), ngoại trừ S4 chứa 2 ORF.

187
Hình 18-3 Tổ chức bộ gen của RRSV
Các protein mã hóa trên 3 phân tử S3, S8 và S9 là các protein cấu trúc chính của phân tử
virus (với các protein tƣơng ứng là P3, P8B vafP9). Bộ gen của RRSV mã hóa 14 protein, trong
đó ít nhất có 6 protein cấu trúc là P1, P2, P3, P4A, P8B và P9. Vị trí và chức năng của các
protein này trình bày ở Bảng 18-2.
Bảng 18-2 Đặc điểm bộ gen và chức năng protein của RRSV
Phân
đoan
Kích thƣớc
(bp)
Protein Chức năng
S1 3849 P1 Gai vỏ (spike B)
S2 3810 P2 Protein vỏ của lớp vỏ trong
S3 3699 P3 Protein vỏ chính của lớp vỏ trong
S4 3823 P4A (Pol) và P4B RdRP (P4A) còn P4B chƣa rõ
S5 2682 P5 (Cap) Enzyme tạo mũ (cap) và guanintransferase
S6 2157 P6 Chƣa rõ
S7 1938 NS7 Protein phi cấu trúc, chƣa rõ chức năng
S8 1814 P8, P8A và P8B Protease (P8, P8A); Protein vỏ chính của lớp vỏ ngoài (P8B)
S9 1132 P9 Protein gai vỏ ngoài (spike A), đặc hiệu vector
S10 1162 NS10 Protein phi cấu trúc, chƣa rõ chức năng
18.3.4 Phạm vi kí chủ
Phạm vi kí chủ của RRSV rất hẹp, chỉ gây hại tự nhiên trên lúa Oryza spp.
18.3.5 Triệu chứng bệnh
Bệnh lùn xoắn lá (hay là bệnh lùn táp lá, ragged stunt disease) biểu hiện 5 triệu chứng đặc
trƣng là (i) cây lùn, (ii) bộ lá xanh đậm, cứng, (iii) lá xoắn vặn một chiều, đặc biệt phần chót lá,
(iv) một bên mép lá (ở các lá bị xoắn vặn hoặc đang phục hồi) bị rách táp dạng chữ V (ragged)
và có thể lan sâu vào gân chính, (v) có các sọc phồng màu trắng sáp chạy dọc gân lá, bẹ lá và đặc
biệt nhiều ở phần cuống lá (tiếp giáp giữa phiến lá và bẹ lá). Trong số các triệu chứng này, táp lá
(ragged) đƣợc xem là đặc trƣng nhất của bệnh.
Trong cây bệnh, phân tử virus hình thành rất nhiều trong tế bào chất của tế bào nhu mô và tế
bào ống rây của mạch dẫn phloem. Trong tế bào chất của các tế bào này, vị trí sinh sản và lắp ráp

188
phân tử virus hình thành một cấu trúc gọi là ‗viroplasms‘ dạng sợi thƣờng bắt màu đậm dƣới
kính hiển vi điện tử (Hình 18-4) trongg cây, vị trí có nhiều virus nhất là các sọc phồng trắng
dạng sáp. Các sọc phồng là sự sinh sản quá mức cả về kích thƣớc và số lƣợng của các tế bào nhu
mô của mạch phloem
Hình 18-4 Một số triệu chứng điển hình của bệnh lúa lùn xoắn lá do RRSV (3 hình đầu, Hà Viết
Cƣờng, 2009). Hình ngoài cùng bên phải là một phần tế bào chất của tế bào nhu mô mạch phloem
tại vị trí nốt phồng với “viroplasm” màu đậm chứa đầy các phân tử virus (Hibino, 1979)
18.3.6 Lan truyền
Virus không truyền qua hạt giống, hạt phấn và tiếp xúc cơ học.
Vector duy nhất của virus RRSV là rầy nâu Nilaparvata lugens. Rầy truyền virus theo kiểu
bền vững tái sinh nhƣng không truyền qua trứng rầy.
Rây nâu N. lugens thƣờng gây hại trên các giống lúa nhiễm rầy nâu, với mật độ cao có thể gây
cháy rầy. Cây lúa nhiễm rầy chuyển mầu vàng rồi chết. Rầy trƣởng thành cánh dài xâm nhập vào
ruộng lúa và đẻ trứng trên các bẹ lá hoặc ở các gân lá. Trứng đẻ có nắp rộng và dẹt. Thời gian
trứng là 7-9 ngày, rầy có 5 tuổi. Giai đoạn rầy non kéo dài 13-15 ngày. Rầy non tuổi một có mầu
trắng, các tuổi sau có mầu nâu. Rầy trƣởng thành có 2 loại: cánh ngắn và cánh dài.
Rầy nâu có đặc điểm truyền RRSV tƣơng tự nhƣ đối với truyền RGSV. Khả năng truyền
virus không có sự khác biệt rõ ràng giữa rầy đực và rầy cái, giữa rầy cánh dài và cánh ngắn. Tuy
nhiên, rầy non có khả năng truyền virus cao hơn với thời gian ẩn ngắn hơn rầy trƣởng thành.
Thời gian chích nạp tối thiểu là 3 giờ. Thời gian ẩn của virus trong vector từ 2 -33 ngày, trung
bình 9 ngày. Thời gian chích truyền tối thiểu là 1 giờ. Triệu chứng trên cây biểu hiện trong vòng
10-36 ngày sau lây nhiễm. Trung bình khoảng 40 % (trong phạm vi 6-76%) rầy truyền đƣợc
virus RRSV. Rầy có thể duy trì khả năng truyền virus từ 1 – 4 tuần hoặc tới khi chết.
18.3.7 Chẩn đoán
Có thể chẩn đoán bệnh bằng 3 biện pháp
Chẩn đoán dựa vào triệu chứng. Nhìn chung bệnh lùn xoắn lá tạo triệu chứng khá đặc trƣng
nên có thể chẩn đoán dễ dàng dựa trên triệu chứng với mức độ tin cậy cao. Các triệu chứng quan
trọng là (i) cây lùn, (ii) bộ lá xanh đậm và cứng, (iii) lá xoắn vặn đặc biệt phần chót lá; (iv) một

189
bên mép lá rách táp, viền vết rách biến vàng, chỗ rách táp thƣờng có dang chữ V, (iv) có nhiều
sọc phồng màu trắng sáp dọc gân lá, bẹ phiến lá và đặc biệt nhiều ở phần tiếp giáp gữa lá và bẹ
lá. Tuy nhiên bệnh có thể rất khó phân biệt với bệnh lùn sọc đen do virus lùn sọc đen phƣơng
Nam gây ra. Điểm chủ yếu để phân biệt 2 bệnh là ở bệnh lùn sọc đen có các sọc phồng ở thân.
Chẩn đoán bằng ELISA. Virus RRSV có hoạt tính miễn dịch tốt. Nhiều nguồn kháng thể virus
hiện sẵn có (IRRI, Nhật Bản, Việt Nam) giúp chẩn đoán bệnh dễ dàng. Tuy nhiên khi chẩn đoán
virus RRSV bằng ELISA cần chú rằng kháng thể đặc hiệu phân tử RRSV có thể tạo phản ứng
ELISA rất mạnh đối với rầy nâu cái mang trứng không nhiễm virus (có thể do kháng thể của
RRSV đã phản ứng với protein của trứng rầy).
Chẩn đoán bằng RT-PCR. Nhiều bộ mồi đặc hiệu virus RRSV hiện cũng sẵn có để chẩn đoán
bệnh.
18.3.8 Phòng chống
Hiện chƣa có công bố tại Việt Nam về đặc tính kháng nhiễm của các giống lúa đối với virus
RRSV. Tƣơng tự đối với bệnh lúa cỏ (do RGSV), chiến lƣợc phòng chống bệnh tại Việt Nam, do
vậy, vẫn chủ yếu dựa vào các biện pháp quản lý vector rầy nâu nhƣ:
Dùng giống kháng rầy
Sử dụng thuốc hóa học phòng trừ rầy
Né rầy di cƣ mang virus bằng điều chỉnh thời vụ trồng (biện pháp đã đƣợc áp dụng khá
hiệu quả tại đồng bằng sông Cửu Long).
Ngoài ra, quy hoạch lại mùa vụ theo hƣớng chấm dứt việc rải vụ quanh năm, giảm cơ cấu 3
vụ lúa nhằm giảm áp lực nguồn bệnh và môi giới truyền bệnh.
Khi cây lúa đã bị nhiễm bệnh thì tiêu huỷ kịp thời nguồn bệnh để tránh lây lan.
18.4 SOUTHERN RICE BLACK-STREAKED DWARF VIRUS (SRBSDV)
18.4.1 Tầm quan trọng
SRBSDV (chi Fijivirus) gây bệnh lúa lùn sọc đen (bệnh lùn lụi) là một virus mới đƣợc xác
định đầu tiên năm 2008 tại các tỉnh phía Nam Trung Quốc nhƣ Quảng Đông, Hải Nam. Tiếp
theo, tại Việt Nam, virus đã đƣợc xác định đầu tiên ở vụ mùa năm 2009 tại nhiều tỉnh miền Bắc
và đang là 1 tác nhân gây bệnh nguy hiểm trên lúa, đặc biệt trong vụ mùa ở miền Bắc và Bắc
Trung Bộ. Theo thống kê của Cục BVTV, vụ mùa năm 2010 , bệnh LSĐ đã phát sinh gây hại tại
28 tỉnh/thành miền Bắc và Bắc Trung Bộ với tổng diện tích bị nhiễm tính là 24.000 ha, trong đó
diện tích phải nhổ tỉa là 9.000 ha và phải tiêu hủy là 1.700 ha.
Chú ý phân biệt một virus tƣơng tự (nhƣng không phát hiện thấy ở Việt Nam) cũng gây ra
bệnh lùn sọc đen là Rice black streaked dwarf virus (RBSDV):
Tên virus tiếng Việt Tên virus tiếng Anh Bệnh
Virus lúa lùn sọc đen Rice black streaked dwarf virus (RBSDV)
Lúa lùn sọc đen
Virus lúa lùn sọc đen phƣơng Nam Southern rice black streaked dwarf virus (SRBSDV)

190
18.4.2 Hình thái phân tử
Virus SRBSDV là một fijivirus nên có các đặc điểm hình thái tƣơng tự nhƣ đối với các
fijivirus khác. Virus có dạng hình cầu đa diện đối xứng icosahedral, đƣờng kính phân tử đầy đủ ~
70 nm. Phân tử có 2 lớp vỏ, trên bề mặt vỏ ngoài có 12 gai loại A (A spike) ngắn gắn trên 12
đỉnh của hình đa diện icosahedral. Vỏ ngoài dễ bị phân hủy để lộ ra lớp vỏ trong cũng mang 12
gai loại B (B spike) trên 12 đỉnh của hình đa diện. Lớp vỏ trong có T= 2 còn lớp vỏ ngoài có T =
13.
Trong tế bào, các phân tử virus thƣờng kết tụ thành thể vùi xếp thành lớp (Hình 18-5).
Hình 18-5. Thể vùi virus và phân tử virus SRBSDV trong tế bào cây bị bệnh lùn sọc đen tại Việt
Nam năm 2009
18.4.3 Đặc điểm bộ gen
Cấu trúc bộ gene của virus LSĐ phƣơng Nam (SRBSDV) gồm 10 phân đoạn RNA sợi kép
(dsRNA) và mã hóa cho các protein chức năng khác nhau. Vì là một virus mới nên tất cả các gen
của virus này đều chƣa đƣợc xác định chức năng. Dựa trên một virus gần gũi là virus LSĐ
(RBSDV) và các fijivirus khác, chức năng một số protein của virus LSĐ phƣơng Nam có thể
đƣợc dự đoán và tóm tắt ở Bảng 18-3.
Bảng 18-3 Đặc điểm bộ gen và dự đoán chức năng các protei của virus SRBSDV (Wang et al., 2010)
Phân
đoan
Kích thƣớc
(bp)
Protein Chức năng
S1 4500 VP1 RdRp
S2 3815 VP2 Protein vỏ của lớp vỏ trong
S3 3618 VP3 Chƣa rõ chức năng
S4 3571 VP4 Chƣa rõ chức năng
S5 3167 VP5 Chƣa rõ chức năng
S6 2651/+2 VP6 Ức chế phản ứng phòng thủ kiểu câm gen của cây
S7 2176 VP7 (a, b) Tạo cấu trúc ống liên quan đến vận chuyển?
S8 1928 VP8 Protein vỏ của lớp vỏ trong
S9 1899/+1 VP9 (a, b) Chƣa rõ chức năng
S10 1797/+1 VP10 Protein vỏ của lớp vỏ ngoài

191
18.4.4 Triệu chứng
Trên lúa. Bệnh lùn sọc đen do virus LSĐ (RBSDV) hay virus LSĐ phƣơng Nam (SRBSDV)
gây ra đều tạo các triệu chứng đặc trƣng sau: (i) cây bị lùn đặc biệt khi nhiễm sớm, (ii) không
trỗ bông hoặc nếu trỗ thì chỉ trỗ một nửa với hạt bị biến màu nâu đen, (iii) bộ lá xanh đậm, cứng,
(iv) lá, đặc biệt phần chót lá, có thể xoắn vặn nhƣng thƣờng không bị rách táp, (v) có nhiều sọc
phồng trắng sau chuyển màu nâu đến đen chạy dọc gân lá, bẹ lá và thân cây, (vi) cây bị bệnh
nặng nhổ lên dễ dàng.
Hình 18-6. Triệu chứng bệnh LSĐ trên lúa ở miền Bắc vụ mùa năm 2009. Lá lúa bị xoắn vặn (a, b,
c), trỗ không thoát với hạt bị bị biến màu nâu (a). Có các nốt phồng màu trắng chạy dọc gân của lá
(d), phiến lá (d), thân (e). Các nốt phồng lúc đầu trắng sau chuyển màu nâu đến đen (e, f).
Trên ngô. Cây ngô nhiễm virus LSĐPN biểu hiện một số triệu chứng sau (Hình 18-7): (i)
các nốt phồng màu trắng sáp trên gân dƣới của lá và gân lá bao bắp, (ii) lá ngọn thƣờng xoắn
vặn, rách táp mép lá, (ii) cây lùn rõ rệt so với cây khỏe.
Hình 18-7. Triệu chứng bệnh LSĐ trên ngô ở miền Bắc năm 2010.
18.4.5 Lan truyền
Vì các reovirus gây hại trên lúa không truyền qua hạt giống cũng nhƣ tiếp xúc cơ học nên khả
năng truyền qua vector rầy đã đƣợc chú ý. Nghiên cứu của các nhà khoa học Trung Quốc với 3
loài rầy là rầy lƣng trắng (Sogatella furcifera), rầy nâu (Nilaparavata lugens) và rầy nâu nhỏ
(Laodelphax striatellus) cho thấy cả rầy lƣng trắng và rầy nâu nhỏ đều có khả năng truyền
a b c f e d

192
SRBSDV từ lúa sang lúa với hiệu quả truyền rất cao (100 % cây nhiễm bệnh với chỉ 3 – 4
rầy/cây). Tuy nhiên, chỉ có rầy lƣng trắng mới có khả năng truyền SRBSDV từ lúa sang ngô.
Nghiên cứu này cũng cho thấy rầy nâu không thể truyền đƣợc SRBSDV.
Tại Việt Nam, đã có một số nghiên cứu về biến động quần thể cũng nhƣ khả năng di cƣ của
rầy trên lúa. Các nghiên cứu của Viện BVTV cho thấy trong những năm gần đây, quần thể rầy
trên lúa miền Bắc đã có sự thay đổi đáng kể. Tỷ lệ rầy nâu chiếm khoảng gần 70 % vào năm
1981 đã giảm xuống còn khoảng gần 30 % vào năm 2007. Trái lại, quần thể rầy lƣng trắng đã
tăng từ khoảng 35 % (1981) lên tới trên 70 % (năm 2007). Đặc biệt, rầy nâu nhỏ đã tái xuất hiện
vào tháng 5-6 năm 2008 tại nhiều tỉnh miền Bắc nhƣ Hƣng Yên, Hải Dƣơng, Thái Bình và Bắc
Ninh.
18.4.6 Chẩn đoán
Có thể chẩn đoán bệnh bằng 2 biện pháp:
Chẩn đoán dựa vào triệu chứng. Nhìn chung bệnh LSĐ phƣơng Nam tạo triệu chứng khá đặc
trƣng nên có thể chẩn đoán dễ dàng dựa trên triệu chứng với mức độ tin cậy cao. Các triệu chứng
quan trọng là (i) cây lùn, (ii) bộ lá xanh đậm và cứng, (iii) lá xoắn vặn đặc biệt phần chót lá; (iv)
có sọc phồng màu trắng sáp dọc gân lá, bẹ phiến lá và đặc biệt ở thân, (v) cây bệnh nhổ lên dễ
dàng. Tuy nhiên bệnh có thể rất khó phân biệt với bệnh lùn xoắn lá. Điểm chủ yếu để phân biệt
2 bệnh là ở bệnh lùn sọc đen có các sọc phồng ở thân. Bệnh ở vụ xuân, triệu chứng thƣờng
không điển hình, đặc biệt đầu vụ.
Chẩn đoán bằng RT-PCR. Các mồi đặc hiệu virus đã sẵn có tại nhiều đơn vị nghiên cứu và
quản lý tại Việt Nam.
18.4.7 Phòng chống
Hiện chƣa có công bố tại Việt Nam về đặc tính kháng nhiễm của các giống lúa đối với virus
LSĐ phƣơng Nam. Tƣơng tự đối với bệnh virus hại lúa, chiến lƣợc phòng chống bệnh tại Việt
Nam, do vậy vẫn chủ yếu dựa vào các biện pháp quản lý vector rầy. Trong điều kiện miền Bắc,
các biện pháp nên đƣợc áp dụng là:
Dùng giống kháng rầy.
Dùng thuốc hóa học, sinh học phòng trừ rầy; bảo vệ mạ và cây lúa ít nhất tới làm đòng.
Tiêu hủy nguồn bệnh để làm giảm áp lực nguồn bệnh và tránh lây lan.
18.5 CÂU HỎI ÔN TẬP CHƢƠNG 18
1. Đặc điểm họ Reoviridae (phân loại, hình thái, quan hệ vector, bản chất bộ gen).
2. RRSV (tầm quan trọng, triệu chứng, đặc điểm bộ gen, lan truyền, phòng chống).
3. SRBSDV (tầm quan trọng, triệu chứng, đặc điểm bộ gen, lan truyền, phòng chống)
18.6 TÀI LIỆU THAM KHẢO CHÍNH
1. Ha, V. C., Nguyen, V.H., Vu, T.M. and Matsumoto, M. (2009). Rice dwarf disease in North
Vietnam in 2009 is caused by southern rice black-streaked dwarf virus (SRBSDV). Bulletin of the Institute of Tropical Agriculture, Kyushu University. Vol. 32, No. 1: 85-92.

193
2. Hibino, H. (1996). Biology and epidemiology of rice viruses. Annual Review of
Phytopathology 34, 249-274.
3. Hogenhout, S.A., Ammar, E.D., Whitfield, A.E. and Redinbaugh, M.G. (2008). Insect Vector
Interactions with Persistently Transmitted Viruses. Annual Review of Phytopathology.
46:327–59.
4. Milne, R.G., del Vas M., Harding, R.M., Marzachi, R. and Mertens, P.P.C. (2005). Genus
Fijivirus. In: Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U. and Ball, L.A. (eds).
Virus taxonomy. Eighth report of the international committee on taxonomy of viruses.
Elsevier Academic Press, San Diego, pp 534–542.
5. Ou, S.H. (1985). Rice Diseases. Second Edition, p. 23-25 & 48-49. Commonwealth
Mycological Institute Publication. Kew, Surrey, UK.
6. Zhang, H.M., Yang, E.J., Chen, J.P. and Adams, M.J. (2008). A black-streaked dwarf disease
on rice in China is caused by a novel fijivirus. Archives of Virology. 153:1893–1898.
7. Zhou, G.H., Wen, J.J., Cai, D.J., Li, P., Xu, D.L. and Zhang, S.G. (2008). Southern rice
black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae.
Chinese Science Bulletin. Vol. 53. No. 23: 3677-3685.
8. Upadhyaya,N.M. and Mertens, P.P.C. (2005) Genus Oryzavirus. In Virus Taxonomy, Eighth
Report of ICTV. Ed. Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U. and Ball, L.A. 550-555.

194
Chƣơng 19. VIRUS RNA SỢI KÉP (TIẾP): CHI TENUIVIRUS
19.1 TÓM TẮT NỘI DUNG
Chƣơng 19 mô tả phân loại và các đặc trƣng phân tử của virus chi Tenuivirus, đại diện cho
nhóm các virus RNA sợi kép, có bộ gen phân đoạn và có hình thái phân tử không hoàn chỉnh.
Chƣơng này cũng đề cập đến 1 tenuivirus quan trọng đối với sản xuất lúa của Việt Nam là
RGSV (Rice grassy stunt virus).
19.2 CHI TENUIVIRUS
Các tenuivirus là các virus RNA có hình thái phân tử dạng tràng hạt. Phân tử virus đƣợc gọi là
nucleoprotein gồm các phân tử RNA genome liên kết với protein vỏ (đƣợc gọi là nucleocapsid).
Các tenuivirus là các virus có bộ gen phân đoạn, gồm 4 – 6 phân tử RNA sợi kép tùy theo
loài, có kích thƣớc từ ~ 1.3 – 9 kb.
Hiện nay, có 6 tenuivirus đƣợc ghi nhận trên thế giới (Bảng 19-1). Các tenuivirus đều gây
bệnh trên cây họ hòa thảo, mỗi virus chỉ lan truyền bằng 1 loài rầy theo kiểu bền vững tuần hoàn
hoặc bền vững tái sinh. Trong số các virus này, hiện chỉ có virus lúa cỏ (RGSV) đã đƣợc phát
hiện ở Việt Nam.
Bảng 19-1 Phân loại chi Tenuivirus (loài chuẩn: Rice stripe virus)
Loài virus Viết tắt
Echinochloa hoja blanca virus EHBV
Maize stripe virus MsPV
Rice grassy stunt virus RGSV
Rice hoja blanca virus RHBV
Rice stripe virus RSV
Urocholoa hoja blanca virus UHBV
19.3 RICE GRASSY STUNT VIRUS (RGSV)
19.3.1 Tầm quan trọng
RGSV gây hại ở khắp các nƣớc trồng lúa châu Á, đặc biệt vùng Đông Nam Á. Trong những
năm 70, bệnh đã gây ra nhiều vụ dịch tại một số nƣớc nhƣ Indonesia, Philippin, Ấn Độ
Tại Việt Nam, virus gây bệnh trên lúa với một số tên gọi khác nhau nhƣ bệnh ―lại mạ‖, ―lúa
cỏ‖, ―vàng lùn‖. Hiện nay, bệnh ―vàng lùn‖ do RGSV gây ra là một trong hai bệnh virus (cùng
với bệnh lùn xoắn lá) nguy hiểm và phổ biến nhất trên lúa ở miền Nam Việt Nam.
19.3.2 Hình thái phân tử
Giống nhƣ các teinuivirus khác, RGSV có hình thái phân tử (virion) dạng sợi tràng hạt, đƣờng
kính 6 – 8 nm, không có vỏ bọc. Hầu hết các phân tử virus ở dạng khép vòng với chu vi 200-
2400nm (trung bình 950-1350 nm).

195
Hình 19-1 Phân tử virus RGSV có dạng tràng hạt khép vòng (Hibino, 1986)
19.3.3 Đặc điểm bộ gen
Bộ gen của RGSV có tổng kích thƣớc là 25 kb, gồm 6 phân đoạn RNA sợi kép, mạch
thẳng khác nhau đƣợc kí hiệu là RNA1, RNA2, RNA3, RNA4, RNA5 và RNA6 theo thứ tự
giảm dần.
Tất cả 6 phân đoạn RNA của virus đều lƣỡng cực (ambiense). Trên mỗi phân đoạn mã hóa 2
protein ngƣợc chiều nhau. Chỉ một số protein của virus đã đƣợc biết chức năng bao gồm:
pC1 mã hóa protein tái bản RdRp của virus
pC5 mã hóa protein N là protein vỏ (capsid)
p6 mã hóa protein NCP (non-capsid protein) là một protein hình thành nhiều trong tế bào
cây lúa bị bệnh.
Hình 19-2 Tổ chức bộ gen của RGSV
19.3.4 Phạm vi kí chủ
Phạm vi kí chủ của RGSV rất hẹp, chỉ gây hại trên lúa Oryza spp.

196
19.3.5 Triệu chứng bệnh
Cây lúa bị nhiễm bệnh, đặc biệt ở giai đoạn mạ, bị lùn mạnh, đẻ nhiều nhánh, lá nhỏ hẹp dựng
đứng xếp xít vào nhau giống búi cỏ nên bệnh đƣợc gọi là bệnh ―lúa cỏ‖. Lá bệnh biến màu lục
nhạt hoặc vàng nhạt, thƣờng có các đốm nâu – đen nhỏ.
Lá bệnh có thể vẫn giữ màu xanh nếu bón đủ đạm.
Cây bệnh không trỗ bông hoăc trỗ rất kém, hạt bị lép hoặc biến màu nâu – đen.
Cây nhiễm muộn thì lá biến vàng, hạt bị biến dạng.
Thí nghiệm chậu vại cho thấy: tùy theo giống, năng suất có thể giảm 70 – 100% khi nhiễm
sau trồng 1 – 1.5 tháng.
Hình 19-3 Triệu chứng bệnh lúa cỏ do RGSV tại Cần Thơ
19.3.6 Lan truyền
Virus không truyền qua hạt giống, hạt phấn và tiếp xúc cơ học.
Virus lan truyền bằng rầy nâu Nilaparvata spp., trong đó quan trọng nhất là rầy nâu N. lugens.
Rầy truyền virus theo kiểu bền vững tái sinh nhƣng không truyền qua trứng rầy.
Rây nâu N. lugens thƣờng gây hại trên các giống lúa nhiễm rầy nâu, với mật độ cao có thể gây
cháy rầy. Cây lúa nhiễm rầy chuyển mầu vàng rồi chết. Rầy trƣởng thành cánh dài xâm nhập vào
ruộng lúa và đẻ trứng trên các bẹ lá hoặc ở các gân lá. Trứng đẻ có nắp rộng và dẹt. Thời gian
trứng là 7-9 ngày, rầy có 5 tuổi. Giai đoạn rầy non kéo dài 13-15 ngày. Rầy non tuổi một có mầu
trắng, các tuổi sau có mầu nâu. Rầy trƣởng thành có 2 loại: cánh ngắn và cánh dài.
Khả năng truyền virus không có sự khác biệt rõ ràng giữa rầy đực và rầy cái, giữa rầy cánh
dài và cánh ngắn. Tuy nhiên, rầy non có khả năng truyền virus cao hơn với thời gian ẩn ngắn hơn
rầy trƣởng thành. Rầy nâu có thời gian chích nạp tối thiểu là 1 giờ, thời gian ẩn của virus trong
vector khoảng 5-28 ngày. Sau thời gian ẩn, rầy bắt đầu truyền virus với thời gian chích truyền tối
thiểu 9 phút. Triệu chứng bệnh biểu hiện rõ sau khi nhiễm virus khoảng 10-20. Rầy nâu mang
virus có thể truyền bệnh cho đến lúc chết.

197
19.3.7 Chẩn đoán
Có thể chẩn đoán bệnh bằng 3 biện pháp:
Chẩn đoán dựa vào triệu chứng. Nhìn chung bệnh lúa cỏ tạo triệu chứng khá đặc trƣng nên có
thể chẩn đoán dựa trên triệu chứng với mức độ tin cậy cao. Các triệu chứng quan trọng là (i) lùn,
(ii) lá biến màu xanh nhợt, nhỏ hẹp, dựng đứng, (iii) đẻ nhiều nhánh xếp xít vào nhau, (iv) lá có
các đốm nâu – đen nhỏ. Tuy nhiên, khi bệnh nhiễm muộn, việc chẩn đoán dựa theo triệu chứng
sẽ khó hơn.
Chẩn đoán bằng ELISA. Virus RGSV có hoạt tính miễn dịch tốt. Nhiều nguồn kháng thể
virus hiện sẵn có (IIRRI, Nhật Bản, Việt Nam) giúp chẩn đoán bệnh dễ dàng. Cần chú ý kháng
huyết thanh đặc hiệu phân tử RGSV phản ứng yếu với Rice stripe virus (RSV) nhƣng không
phản ứng với Maize stripe virus (MsPV) hoặc Rice hoja blanca virus (RHBV).
Chẩn đoán bằng RT-PCR. Nhiều bộ mồi đặc hiệu virus RGSV hiện cũng sẵn có để chẩn đoán
bệnh.
19.3.8 Phòng chống
Hiện chƣa có công bố tại Việt Nam về đặc tính kháng nhiễm của các giống lúa đối với virus
RGSV. Chiến lƣợc phòng chống bệnh tại Việt Nam, do vậy vẫn chủ yếu dựa vào các biện pháp
quản lý vector rầy nâu nhƣ:
Dùng giống kháng rầy
Sử dụng thuốc hóa học phòng trừ rầy
Né rầy di cƣ mang virus bằng điều chỉnh thời vụ trồng (biện pháp đã đƣợc áp dụng khá
hiệu quả tại ĐB sông Cửu long).
Ngoài ra, quy hoạch lại mùa vụ theo hƣớng chấm dứt việc rải vụ quanh năm, giảm cơ cấu 3
vụ lúa nhằm giảm áp lực nguồn bệnh và môi giới truyền bệnh.
Khi cây lúa đã bị nhiễm bệnh thì tiêu hủy kịp thời nguồn bệnh để tránh lây lan.
19.4 CÂU HỎI ÔN TẬP CHƢƠNG 19
4. Đặc điểm chi Tenuivirus (phân loại, hình thái, quan hệ vector, bản chất bộ gen).
5. RGSV (tầm quan trọng, triệu chứng, đặc điểm bộ gen, lan truyền, phòng chống).
19.5 TÀI LIỆU THAM KHẢO CHÍNH
1. Haenni, A.L., de Miranda, J.R., Falk, B.W., Goldbach, R., Mayo, M.A., Shirako, Y. and
Toriyama, S. (2005). Genus Tenuivirus. In: Virus Taxonomy, Eighth Report of ICTV. Ed.
Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U. and Ball, L.A. 717-723.
2. Hibino, H. (1996). Biology and epidemiology of rice viruses. Annual Review of
Phytopathology 34, 249-274.
3. Hogenhout, S.A., Ammar, E.D., Whitfield, A.E. and Redinbaugh, M.G. (2008). Insect Vector
Interactions with Persistently Transmitted Viruses. Annual Review of Phytopathology.
46:327–59.
4. Ou, S.H. (1985). Rice Diseases. Second Edition, p. 23-25 & 48-49. Commonwealth
Mycological Institute Publication. Kew, Surrey, UK

198
Chƣơng 20. PHYTOPLASMA
20.1 TÓM TẮT NỘI DUNG
Chƣơng 20 mô tả tóm tắt lịch sử nghiên cứu cũng nhƣ các đặc điểm khái quát nhất (bản chất,
phân loại, lan truyền, tính chất gây bệnh) của một nhóm tác nhân gây bệnh thực vật đặc biệt: là
vi khuẩn nhƣng lại có nhiều đặc điểm bệnh học giống virus.
20.2 GIỚI THIỆU
Vào năm 1967, các nhà khoa học Nhật Bản đã phát hiện ra một nhóm tác nhân gây bệnh có
đặc điểm cấu trúc giống nhƣ Mycoplasma (Mycoplasma - Like Organism, MLO) là nguyên nhân
gây ra nhiều bệnh biến vàng cây trồng mà trƣớc đó hầu hết các nhà khoa học đều cho rằng
nguyên nhân có lẽ do virus. Hiện nay tất cả các tác nhân MLO gây bệnh trên thực vật đƣợc gọi là
Phytoplasma.
Bản chất thực sự của Phytoplasma chỉ đƣợc khám phá vào cuối những năm 1980 và đầu
những năm 1990 do áp dụng các công cụ phân tích phân tử. Các phân tích dựa trên chuỗi gen mã
hóa tiểu phần 16S RNA ribosome và chuỗi gen mã hóa tiểu phần protein của ribosome đã cho
thấy Phytoplasma (vào thời điểm đó vẫn đƣợc gọi là MLO) thuộc nhóm dịch khuẩn bào
(mollicustes). Tiếp theo, các phân tích dựa vào phản ứng trùng hợp chuỗi (PCR) dùng các cặp
mồi đặc hiệu chuỗi gen 16S RNA cũng nhƣ phân tích đa hình các các đoạn cắt giới hạn RFLP
các sản phẩm PCR đã cho phép phát hiện và xác định nguyên nhân của hàng trăm bệnh cây là do
Phytoplasma gây ra.
20.3 ĐẶC ĐIỂM
Phytoplasma là loại vi khuẩn hại thực vật đặc biệt, có 3 đặc điểm quan trọng sau:
(i) Chúng không thể nuôi cấy đƣợc trên môi trƣờng nhân tạo (đây là lý do tại sao phần lớn
các nghiên cứu về Phytoplasma thƣờng đƣợc thực hiện bởi các nhà virus học).
(ii) Chúng thiếu lớp vách tế bào. Tế bào của chúng chỉ đƣợc bao bọc bởi một lớp màng tê bào,
bên trong là các cơ quan tử tƣơng tự nhƣ của vi khuẩn. Do không có lớp vách rắn chắc bên ngoài
nên tế bào Phytoplasma có dạng vô định hình, thay đổi từ hình cầu (với đƣờng kính trung bình từ
200µm đến 800µm), hình ovan, hình sợi, hình ống. Kích thƣớc bộ gen của Phytolasma thay đổi
từ 530 kilobases tới 1130 kilobases.
(iii) Chúng tồn tại ở tế bào mạch phloem (chủ yếu ở tế bào mạch rây).
Hình 20-1 Tế bào phytoplasma trong tế bào mạch rây ký chủ (Lee et al., 2000)

199
20.4 PHÂN LOẠI
Hiện nay, theo khóa phân loại vi khuẩn của Bergey (2004), Phytolasma đƣợc xếp vào chi
Candidatus Phytoplasma (Ca. Phytoplasma), Bộ Acholeplasmatales, Lớp Mollicutes, Ngành
Firmucutes, Giới Vi khuẩn (Bacteria). Trên cơ sở so sánh chuỗi DNA ribosome 16S (16S
rDNA) bằng kỹ thuật RFLP hoặc giải trình tự gen, các loài Ca. Phytoplasma đƣợc xếp vào ít
nhất 16 nhóm phả hệ (ký hiệu là 16SrI...16SrXIV), mỗi nhóm có tên riêng, chẳng hạn nhóm
16SrI là nhóm Aster yellowing (Biến vàng cây cúc).
Bảng 20-1 Phân loại phytoplasma dựa trên đặc điểm RNA ribosome 16S
Nhóm
16Sr Tên nhóm Loài
16SrI Aster yellows Ca. Phytoplasma asteris, Ca. Phytoplasma japonicum
16SrII Peanut witch's broom Ca. Phytoplasma aurantifolia
16SrIII X-disease Ca. Phytoplasma pruni
16SrIV Coconut lethal yellowing Ca. Phytoplasma palmae, Ca. Phytoplasma castaneae, Ca. Phytoplasma cocosnigeriae
16SrV Elm yellows Ca. Phytoplasma ziziphi, Ca. Phytoplasma vitis, Ca. Phytoplasma
ulmi
16SrVI Clover proliferation Ca. Phytoplasma trifolii
16SrVII Ash yellows Ca. Phytoplasma fraxini
16SrVIII Luffa witch's-broom Ca. Phytoplasma luffae
16SrIX Pidgeon pea witch's broom Ca. Phytoplasma phoenicium
16SrX Apple proliferation Ca. Phytopalsma Mali, Ca. Phytoplasma pyri, Ca. Phytoplasma
prunorum, Ca. Phytoplasma spartii, Ca. Phytoplasma rhamnii, Ca.
Phytoplasma allocasuarinae
16SrXI Rice Yellow Dwarf Ca. Phytopalsma oryzae
16SrXII Stolbur Ca. Phytoplasma solani, Ca. Phytoplasma australiense
16SrXIII Mexican periwinkle
virescence Chưa xác định
16SrXIV Bermuda white leaf Ca. Phytoplasma cynodontis
16SrXV Hibiscus witch's-broom Ca. Phytoplasma brasiliense
20.5 TRIỆU CHỨNG
Do làm biến đổi cân bằng phytohormon trong cây nên Phytoplasma thƣờng gây ra một số
triệu chứng đặc trƣng nhƣ:
1. Diệp hóa (phyllody): các bộ phận của hoa (cánh hoa) phát triển thành cấu trúc giống nhƣ lá.
2. Lục hóa (virescence): thay vì có màu sắc bình thƣờng, hoa phát triển sắc tố lục.
3. Chổi phù thủy (witches’-broom): sự phát triển quá mức chồi bên, chồi phụ + lóng kéo dài bất thƣờng (nhƣng mảnh).
4. Chùn ngon, chùn hoa
5. Cây còi cọc, lùn

200
6. Lá biến màu (vàng, đỏ).
20.6 LAN TRUYỀN
Phytoplasma lan truyền chủ yếu nhờ côn trùng chích hút họ Cicadellidea (rầy lá) và
Fulgoridea (rầy thân) theo kiểu bền vững tái sinh.
Phytoplasma không lan truyền qua hạt giống nhƣng có thể lan truyền dễ dàng qua vật liệu
giống đƣợc nhân giống vô tính (hom giống, củ giống, thân ngầm).
Phytoplasma không lan truyền qua lây nhiễm nhân tạo bằng tiếp xúc cơ học dùng dịch chiết
cây bệnh nhƣng truyền dễ dàng qua chiết, ghép và tơ hồng.
20.7 PHÒNG CHỐNG
Do phytoplasma truyền qua vector rầy và quan nhân giống vô tính nên các chiến lƣợc chính
để phòng trừ phytoplssma thƣờng dùng hiện nay là:
- Kiểm soát vector rầy bằng biện pháp hóa học, sinh học.
- Nhổ bỏ, tiêu hủy tàn dƣ cây bệnh càng sớm càng tốt.
- Không dùng những cây bị bệnh dù đã lớn làm hom giống. Có thể xử lý hom giống bằng
nƣớc nóng 54oC trong 40 phút (kinh nghiệm của Đài Loan đối với bệnh phytoplasma trên
mía).
- Không vận chuyển trao đổi hom giống từ các vùng bị bệnh tới các vùng trồng mới, tổ
chức sản xuất và kiểm tra các giống sạch bệnh
20.8 CA. PHYTOPLASMA ASTERIS
Đây là phytoplasma phổ biến khắp thế giới và quan trọng nhất trên thực vật. Phytoplasma hại
khoảng 300 loài cây thuộc 48 họ. Hai cây ký chủ quan trọng nhất là cà rốt và rau diếp. Năng suất
cây giảm thay đổi theo cây ký chủ (nặng nhất trên cà rốt: thƣờng giảm 10 -25 %, có thể tới 90%).
Phytoplasma lan truyền bằng nhiều loài rầy lá, quan trọng nhất là Macrosteles quadrilineatus.
Phytoplasma cũng truyền qua ghép và tơ hồng.
Hình 20-2 Triệu chứng chổi phù thủy và rễ tóc do Ca. Phytoplasma asteris gây ra trên cà rốt, tre

201
20.9 CÂU HỎI ÔN TẬP CHƢƠNG 20
1. Lịch sử nghiên cứu và phát hiện Phytoplasma
2. Các đặc điểm chính của phtoplasma (bản chất, phân loại, tính chất gây bệnh)
3. Phóng chống phytoplasma
4. Bệnh do Ca. Phytoplasma asteris.
20.10 TÀI LIỆU THAM KHẢO CHÍNH
1. Bertaccini, A. (2007). Phytoplasmas: diversity, taxonomy, and epidemiology. Front Biosci
12, 673-689.
2. Christensen, N. M., Axelsen, K. B., Nicolaisen, M. & Schulz, A. (2005). Phytoplasmas and
their interactions with hosts. Trends Plant Sci 10, 526-535.
3. Lee, I. M., Gundersen-Rindal, D. E., Davis, R. E., Bottner, K. D., Marcone, C. & Seemuller,
E. (2004). 'Candidatus Phytoplasma asteris', a novel phytoplasma taxon associated with aster
yellows and related diseases. Int J Syst Evol Microbiol 54, 1037-1048.
4. Lee, I. M., Davis, R. E. & Gundersen-Rindal, D. E. (2000). Phytoplasma: phytopathogenic
mollicutes. Annu Rev Microbiol 54, 221-255.
5. Weintraub, P. G. & Beanland, L. (2006). Insect vectors of phytoplasmas. Annu Rev Entomol
51, 91-111.
6. Weintraub, P. G. & Wilson, M. R. (2010). Control of Phytoplasma Diseases and Vector. In
Phytoplasma: Genomes, Plant Hosts and Vectors Edited by P. G. Weintraub & P. Jones:
CABI.

202
Chƣơng 21. VIROID
21.1 TÓM TẮT NỘI DUNG
Chƣơng 21 mô tả tóm tắt lịch sử nghiên cứu cũng nhƣ các đặc điểm khái quát nhất (bản chất,
cấu trúc phân tử, phân loại, tái sinh, lan truyền, tính chất gây bệnh) của một nhóm tác nhân gây
bệnh thực vật đặc biệt: chỉ là một phân tử RNA rất nhỏ nhƣng có thể gây bệnh rất giống virus.
21.2 GIỚI THIỆU
Vào năm 1922, một bệnh lạ trên khoai tây đã đƣợc quan sát thấy trên cây khoai tây. Cây bệnh
biểu hiện triệu chứng cây thấp nhỏ, lá hẹp, chót lá thót nhọn và quan trọng nhất là củ bệnh dài,
nhỏ hẹp, vỏ củ nhẵn, có nhiều mắt. Cây bệnh truyền dễ dàng qua tiếp xúc (kể cả qua dụng cụ
nhiễm bệnh), qua hạt và phấn hoa. Trong vòng 40 năm tiếp theo, tác nhân gây bệnh đã đƣợc qui
cho 1 virus thực vật. Tiếp theo, ngƣời ta chứng minh rằng tác nhân gây bệnh có hệ số sa lắng
thấp, mẫn cảm với ribonuclease nhƣng không mẫn cảm với deoxyribonuclease, phenol,
chloroform, n-butanol và ethanol. Các đặc điểm trên chứng tỏ tác nhân gây bệnh là một phân tử
RNA nhỏ. Khi bản chất sinh hóa của tác nhân gây bệnh đƣợc làm rõ thì thuật ngữ ―viroid‖ đƣợc
đề xuất vào năm 1971 để phân biệt các tác nhân gây bệnh là các phân tử RNA nhỏ không có vỏ
protein với các tác nhân gây bệnh là virus. Bệnh trên củ khoai tây đã đƣợc đặt tên là bệnh ―củ
hình thoi‖ (Potato spindle tuber viroid – PSTVd).
Các bệnh tiếp theo đƣợc phát hiện với nguyên nhân tƣơng tự là bệnh lùn cây cúc
(Chrysanthemum stunt viroid, CSVd) và bệnh vẩy vỏ cây có múi (Citrus exocortis viroid,
CEVd).
Cho tới nay, hơn 30 bệnh viroid đã đƣợc phát hiện thấy trên thực vật, phần lớn trên cây trồng
có ý nghĩa kinh tế nhƣ khoai tây, cà chua, cọ, nho, cây có múi, bơ, đào, táo, lê, hoa cúc…
21.3 ĐỊNH NGHĨA VIROID
Viroid là các phân tử RNA sợi vòng đơn, không có vỏ protein, tự tái sinh trong tế bào ký chủ
thực vật. Một số là tác nhân gây bệnh nhƣng một số không tạo triệu chứng nhiễm bệnh trên cây.
21.4 CẤU TRÚC VIROID
Cấu trúc sơ cấp. Ở dạng cấu trúc sơ cấp, viroids là 1 phân tử RNA sợi đơn, dạng vòng, kích
thƣớc từ 246 - 375 nt.
Cấu trúc thứ cấp. Nhìn tổng thể, viroid có dạng hình gậy xoắn nhƣng về cấu trúc thứ cấp chi
tiết, phân tử gồm 3 dạng khác nhau.
Dạng 1, đặc trƣng cho họ Pospoviroidae, có cấu trúc hình gậy. Phân tử viroid trở thành dạng
sợi kép, trên đó gồm các đoạn bắt cặp hoàn hảo (đoạn xoắn kép) xen kẽ với các đoạn không bắt
cặp (đoạn thòng lọng). Phân tử đƣợc chia làm 5 vùng chức năng theo thứ tự từ trái sang phải là:
Vùng đầu trái TL (terminal – left), vùng qui định tính gây bệnh P (pathogenic), vùng trung tâm C
(central), vùng biến động V (variable) và vùng đầu phải TR (terminal – right). Vùng trung tâm C
chứa một vùng bảo thủ gọi là vùng bảo thủ trung tâm CCR (central conserved region) liên quan
đến tái sinh viroid.
Dạng 2, đặc trƣng cho họ Avsunviroidae, có cấu trúc gần hình gậy. Phân tử viroid cũng có
dạng sợi kép gồm các đoạn bắt cặp hoàn hảo (đoạn xoắn kép) xen kẽ với các đoạn không bắt cặp

203
(đoạn thòng lọng). Tuy nhiên, ở đầu trái của phân tử có 2 cấu trúc kẹp tóc (hairpin). Ngoài ra, 2
thành viên của họ này là Chrysanthemum chlorotic mottle viroid (CChMVd) và Peach latent
mosaic viroid (PLMVd) có cấu trúc phân nhánh phức tạp ở gần đầu trái của phân tử. Các viroid
họ Avsunviroidae thiếu vùng bảo thủ trung tâm CCG nhƣng chứa một vùng tạo cấu trúc thứ cấp
giống nhƣ đầu búa và có hoạt tính enzyme (tự cắt) gọi là ribozyme đầu búa (hammerhead
ribozyme)
Hình 21-1 Cấu trúc viroid. A) hình dạng chung của viroid. B) phân tử viroid dƣới kính hiển vi điện
tử. C) cấu trúc dạng hình gậy của các pospiviroid. D) cấu trúc dạng gần hình gậy của các
avsunviroid.
21.5 DANH PHÁP VÀ PHÂN LOẠI
21.5.1 Danh pháp viroid
Các viroid đƣợc đặt tên giống nhƣ virus, tuy nhiên từ cuối cùng là ―viroid‖. Ví dụ:
- Potato spindle tuber viroid (PSTVd)
- Avocado sunblotch viroid (ASBVd)
A
B
C
D

204
21.5.2 Phân loại viroid
Dựa vào cấu trúc phân tử và quan hệ chuỗi nucleic acid (ngƣỡng đồng nhất trình tự 75%), các
viroid hiện nay đƣợc phân loại thành 2 họ Avsunviroidae và Pospiviroidae.
Các thành viên thuộc họ Avsunviroidae có khả năng tự cắt các multimers đƣợc tạo ra trong
quá trình tái sinh và không có vùng bảo thủ trung tâm (central conserved region (CCR)).
Các thành viên của họ Pospiviroidae không có có khả năng tự cắt các multimers đƣợc tạo ra
trong quá trình tái sinh và có một vùng bảo thủ trung tâm (central conserved region (CCR)).
Các loài trong mỗi họ đƣợc xác định chủ yếu dựa trên mức độ tƣơng đồng chuỗi RNA.
Ngƣỡng phân biệt loài là 90% mức đồng nhất chuỗi RNA.
Các chi đƣợc phân biệt trên cơ sở sự có mặt hay không có mặt và đặc điểm của vùng bảo thủ
trung tâm (CCR).
Bảng 21-1 Phân loại viroid (ICTV, 2009)
Họ Chi Loài chuẩn Số loài
Avsunviroidae
Avsunviroid Avocado sunblotch viroid 1
Elaviroid Eggplant latent viroid 1
Pelamoviroid Peach latent mosaic viroid 2
Pospiviroidae
Apscaviroid Apple scar skin viroid 10
Cocadviroid Coconut cadang-cadang viroid 4
Coleviroid Coleus blumei viroid 1 3
Hostuviroid Hop stunt viroid 1
Pospiviroid Potato spindle tuber viroid 9
31
21.6 TRIỆU CHỨNG, CƠ CHẾ GÂY BỆNH VÀ LAN TRUYỀN
21.6.1 Triệu chứng
Triệu chứng bệnh viroid nhìn chung tƣơng tự triệu chứng do virus gây ra. Triệu chứng thay
đổi theo loài/chủng viroid, loài/giống cây ký chủ, và cả điều kiện môi trƣờng. Nhìn chung, một
trong các triệu chứng phổ biến nhât của bệnh viroid là ―còi cọc‖ (stunt) có nghĩa cây thấp nhỏ.
21.6.2 Cơ chế gây bệnh
RNA của viroid có thể có các đoạn tƣơng đồng với RNA của ký chủ dẫn tới (i) viroid có thể
bắt cặp với mRNA của ký chủ và ức chế quá trình vận chuyển, dịch mã của các mRNA này hoặc
(ii) tƣơng tác với quá trình điều khiển biểu hiện gen ký chủ thông qua cơ chế câm gen. Ngoài ra,
viroid cũng đƣợc chứng minh là có thể tƣơng tác với protein ký chủ để cảm ứng gây bệnh
21.6.3 Lan truyền
Viroid lan truyền qua tiếp xúc cơ học, ghép, vật liệu giống là chủ yếu.

205
21.7 TÁI SINH CỦA VIROID
Viroid tái sinh theo cơ chế vòng lăn (rolling-circle) và gồm 2 phƣơng thức:
21.7.1 Phƣơng thức bất đối xứng (asymmetric)
Đây là cách tái sinh của các viroid họ Pospivirodae. Phân tử RNA genome mạch vòng, cực
(+), dạng monomer của viroid đƣợc phiên mã bởi RdRp của ký chủ để tạo ra sợi cực (-) dạng
oligomer. Trên khuôn sợi cực (-), sợi cực (+) cũng dạng oligomer đƣợc tổng hợp. Tiếp theo, với
sự tham gia của yếu tố ký chủ (HF, host factor), sợi cực (+) dạng oligomer sẽ đƣợc cắt để tạo
thành sợi cực (+) dạng monomer và đƣợc nối lại thành mạch vòng.
21.7.2 Phƣơng thức đối xứng (symmetric)
Đây là cách tái sinh của các viroid họ Avsunvirodae. Phân tử RNA genome mạch vòng, cực
(+), dạng monomer của viroid đƣợc phiên mã bởi RdRp của ký chủ để tạo ra sợi cực (-) nhƣng
đƣợc cắt luôn bởi ribozyme đầu búa của viroid để tạo dạng monomer và đƣợc nối lại ngay để tạo
thành sợi mạch vòng cực (-). Phân tử sợi mạch vòng, cực (-) này đƣợc dùng làm khuôn để tổng
hợp sợi (+) và cũng đƣợc cắt và nối ngay thành sợi mạch vòng, cực (+) dạng monomer.
Hình 21-2 Sơ đồ tái sinh của viroid (Daros et al., 2006)
21.8 POTATO SPINDLE TUBER VIROID (PSTVD)
Tác nhân: Potato spindle tuber viroid (PSTVd), họ Pospiviroidae
Bệnh: củ khoai tây hình thoi
Tầm quan trọng: có thể giảm 25 % năng suất
Ký chủ: Khoai tây (chính); cũng nhiễm tự nhiên trên nhiều cây họ cà khác
Triệu chứng: củ: nhỏ, dài nhiều mắt; cây: lùn, đẻ nhiều nhánh
Phân bố: khắp thế giới
Lan truyền: dễ dàng qua tiếp xúc cơ học; côn trùng miệng nhai; có thể qua phấn hoa
Hình 21-3 Bệnh củ khoai tây hình thoi

206
21.9 COCONUT CADANG-CADANG VIROID (CCCVD)
Tác nhân: Coconut cadang-cadang viroid (CCCVd), họ Pospiviroidae
Bệnh: cadang-cadang trên cây cọ
Tầm quan trọng: nguy hiểm nhất trên cọ tại Philippin (30+ triệu cây cọ đã chết vào 1982)
Ký chủ: Cọ
Triệu chứng: kéo dài hàng năm. Quả tròn, nhỏ; đốm biến vàng trên lá, không hình thành hoa,
cây còi cọc.
Phân bố: Philippin
Lan truyền: Chƣa rõ. Bằng thực nghiệm, viroid có thể truyền qua phấn hoa, hạt, dụng cụ thu
hái.
21.10 AVOCADO SUNBLOTCH VIROID (ASBVD)
Tác nhân: Avocado sunblotch viroid (ASBVd), họ Avsunviroidae
1. Tầm quan trọng: giảm năng suất từ 30 – 95% tùy giống
2. Ký chủ: Cây bơ là ký chủ tự nhiên duy nhât
3. Triệu chứng: cây thấp nhỏ; quả nhỏ, có nhiều vết biến màu nhƣ bị bỏng nắng
4. Phân bố: châu Mỹ, châu Âu, Öc, châu Á (Israel)
5. Lan truyền: chủ yếu qua ghép; cây con làm gốc ghép trồng từ hạt cây nhiễm bệnh (tỷ lệ
truyền qua hạt có thể tới 95%).
21.11 CÂU HỎI ÔN TẠP CHƢƠNG 21
1. Lịch sử nghiên cứu viroid
2. Bản chất viroid (định nghĩa, cấu trúc sơ cấp, thứ cấp, tái sinh)
3. Phân loại và danh pháp viroid.
4. Đặc điểm gây bệnh và lan truyền
5. Potato spindle tuber viroid
21.12 TÀI LIỆU THAM KHẢO CHÍNH
1. Daròs, J.-A., Elena, S. F. & Flores, R. (2006). Viroids: an Ariadne‘s thread into the RNA
labyrinth. EMBO reports 7, 593–598.
2. Diener, T. O. (2003). Discovering viroids—a personal perspective. Nature Review
Microbiology 1, 75–80.
3. Flores, R., Hernandez, C., Alba, A. E. M. ı. d., Daros, J.-A. & Serio, F. D. (2005). VIROIDS
AND VIROID-HOST INTERACTIONS. Annu Rev Phytopathol 43, 117–139.
4. Góra-Sochacka, A. (2004). Viroids: unusual small pathogenic RNAs. Acta Biochimica
Polonica 51, 587–607.
5. Hull, R. (2002). Matthews's Plant Virology. Fourth edition: Academic Press.

207
PHẦN 3: THỰC HÀNH

208
Bài 1. ĐIỀU TRA BỆNH VIRUS
Mục đích
Giúp sinh viên biết đƣợc triệu chứng bệnh virus ngoài đồng ruộng
Biết cách điều tra đồng ruộng
Biết cách thu thập và bảo quản mẫu bệnh virus
Nội dung
Điều tra một bệnh ngoài đồng ruộng theo yêu cầu của giáo viên (bệnh sẽ thay đổi theo
thực tế đồng ruộng)
Thu thập mẫu: Mỗi nhóm thu 10 mẫu bệnh virus trên 10 loại cây trồng khác nhau. Các
mẫu cần thu thập trƣớc buổi thực tập 1-2 ngày, bảo quản trong hộp ẩm, túi ẩm, hoặc
trồng lại trong cốc nhựa (đối với mẫu cây nhỏ) để đảm bảo mẫu vẫn còn tƣơi khi mang
đến phòng thực hành. Giáo viên sẽ hƣớng dẫn phân biệt các mẫu tại phòng thực tập. Xem
bảng gợi ý mẫu bệnh virus/cây trồng sẵn có tại miền BắcViệt Nam.
Xử lý khô mẫu bằng silicagel để bảo quản lâu dài
Vật liệu
Silicagel tự chỉ thị, giấy thấm, lọ có nắp hoặc ống falcon loại 50 mL.
Yêu cầu cụ thể đối với sinh viên
Lớp chia nhóm (5 sinh viên/nhóm)
Mỗi nhóm thực hiện tất cả các nội dung trên và viết báo cáo riêng
Báo cáo
o Trình bày trên word
o Tên nhóm và các thành viên
o Về điều tra bệnh:
– Tên bệnh, tên cây ký chủ (giống nếu có), giai đoạn sinh trƣởng, địa điểm,
ngày điều tra, thu mẫu. Tham khảo tài liệu dịch bệnh cây để biết phƣơng
pháp điều tra.
– Kết quả điều tra:
– Tỷ lệ bệnh (%), cần kèm dung lƣợng mẫu
– Mô tả triệu chứng điển hình (kèm ảnh minh họa)
– Nhận xét chung về tình hình bệnh
– Tham khảo 2-3 tài liệu để mô tả khoảng ½ trang về virus gây bệnh (hình
thái, phân loại, đặc điểm bộ gen, bệnh gây ra trên cây, quan hệ vector).
o Về thu thập mẫu bệnh:
– Lập bảng ghi tên mẫu bệnh thu thập gồm cây, tên bệnh và tên virus (dự
đoán). Đối chiếu với hƣớng dẫn của giáo viên

209
Bảng hƣớng dẫn tìm cây và bệnh virus (Chú ý có thể tìm cả các cây trồng và cây dại
khác nếu phát hiện thấy triệu chứng giống bị nhiễm virus)
TT Cây Virus
1 Lúa SRBSDV, RRSV, RYSV
2 Ngô SCMV (Sugar cane mosaic potyvirus), SRBSDV
3 Hoa loa kèn LMoV (Lily mottle potyvirus)
4 Hoàng tinh SCMV
5 Dâu tằm MRSV (Mulberry ring spot nepovirus)
6 Chuối tiêu BBTV (Banana bunchytop babuvirus)
7 Chuối ta CMV (Cucumber mosaic cucumovirus)
8 Đu đủ PRSV
9 Bí ngô PRSV, SLCCNV (Squash leaf curl China begomovirus)
10 Dƣa chuột, bầu… PRSV, ZYMV (Zucchini yellow mosaic potyvirus)
11 Đậu tƣơng, sắn dây KuMV (Kudzu mosaic begomovirus)
Sắn dây KuMV (Kudzu mosaic begomovirus)
12 Đậu đỗ khác BCMV (Bean common mosaic potyvirus)
13 Cà chua Begomoviruses
14 Ớt ChiVMoV (Chilli veinal mottle potyvirus)
15 Khoai sọ DsMV (Dasheen mosaic potyvirus)
16 Khoai tây PVY (Potato virus Y)
PVX (Potato virus X)
17 Hành, tỏi OYDV (Onion yellow dwarf potyvirus), LYSV (Leek yellow
stripe potyvirus), SYSV (Shallot yellow stripe poyvirus)
18 Lữ đằng Begomovirus
19 Hoa ngũ sắc Begomovirus
20 Nhọ nồi Begomovirus

210
Bài 2. LÂY NHIỄM NHÂN TẠO VIRUS BẰNG TIẾP XÖC CƠ HỌC
Mục đích
Sinh viên biết cách lây nhiễm nhân tạo bệnh virus bằng tiếp xúc cơ học
Nội dung
Tự trồng cây thí nghiệm.
Mang cây thí nghiệm đến phòng thực tập để lây nhiễm
Tự theo dõi và đánh giá kết quả lây nhiễm
Yêu cầu cụ thể đối với sinh viên
Lớp chia nhóm (5 sinh viên / nhóm)
Mỗi nhóm thực hiện các nhiệm vụ cụ thể sau
o Trồng cây thí nghiệm: Trƣớc ngày thí nghiệm khoảng 1 tuần, gieo các hạt đậu
tƣơng và bầu bí vào cốc nhựa nhỏ (loại cốc nhựa uống nƣớc dùng 1 lần) và mua
đu đủ con.
Đậu tƣơng: 10 hạt / 5 cốc
Bầu bí: 10 hạt/5 cốc
o Mua cây thí nghiệm: mỗi nhóm mua 3 cây đu đủ con. Các cây đƣợc trồng vào túi
bầu trồng cây
o Mang cây thí nghiệm tới phòng thực tập và thực hiện lây nhiễm theo hƣớng dẫn
của giáo viên.
o Lây PRSV trên đu đủ, bầu bí, cây chỉ thị (rau muối hoặc thuốc lá)
o Lây KuMV trên đậu tƣơng
o Viết báo cáo
Báo cáo
Trình bày trên word
Tên nhóm và các thành viên
Kết quả lây nhiễm sau 3 ngày, 7 ngày, 14 ngày (Ghi triệu chứng quan sát thấy + ảnh).
Trình tự lây bệnh nhân tạo bằng tiếp xúc cơ học
Thao tác
Nghiền lá bệnh bằng chày và cối sứ sạch trong đệm phosphate 0.1 M, pH 7.5 theo tỷ lệ 1
g mô lá/2 ml đệm; lọc qua vải lọc hoặc giấy lọc (chú ý giữ lạnh dịch nghiền trên đá)
Thổi bột carborandum 600mesh lên lá cây cần lây
Dùng ngón tay hoặc tăm bông tẩm dịch virus và lƣớt nhẹ trên lá từ cuống đến chót, chú ý
không quá mạnh
Rửa ngay lá lây bằng nƣớc. Đánh dấu lá đã lây bằng bút viết kính
Theo dõi kết quả sau 1 - 2 tuần

211
Bài 3. PHÁT HIỆN VIRUS BẰNG ELISA
Mục đích
Sinh viên biết sử dụng kỹ thuật ELISA để chẩn đoán virus thực vật
Nội dung
Tự thu thập mẫu bệnh ngoài đồng ruộng
Thực hiện phản ứng ELISA gián tiếp kiểu PTA-ELISA (Plate Trapped Antigen - ELISA)
để phát hiện virus PRSV
Yêu cầu cụ thể đối với sinh viên
Lớp chia nhóm (5 sinh viên / nhóm)
Mỗi nhóm thực hiện phản ứng PTA-ELISA trên 4 mẫu để phát hiện PRSV gồm:
– Mẫu đu đủ bệnh (đối chứng dƣơng): 1 mẫu
– Mẫu bầu bí (bất kỳ cây học bầu bí biểu hiện triệu chứng khảm): 2 mẫu
– Mẫu đu đủ khỏe (đối chứng cây khỏe): 1 mẫu
Báo cáo
Trình bày trên word
Tên nhóm và các thành viên
Kết quả ELISA đối với các mẫu
Qui trình chẩn đoán bệnh virus bằng kỹ thuật PTA-ELISA
Chuẩn bị
1. Dụng cụ: chày cối sứ sạch, tube 1,5ml, pipet, giấy thấm, bản ELISA, hộp nhựa
2. Các đệm ELISA: Đệm nghiền mẫu (đệm carbonate), đệm rửa (đệm PBS-T), đệm pha kháng
thể (đệm PBS-T- ovalbumin), đệm cơ chất (chất nền).
3. Hai loại kháng thể: IgG thỏ đặc hiệu PRSV, IgG chuột/dê đặc hiệu IgG thỏ liên kết enzyme
Alkaline Phosphatase (AP)
4. Chất nền Nitrophenyl phosphate (NPP), dạng viên hoặc bột.
Thực hiện phản ứng
1. Cố định dịch cây vào bản ELISA: Nghiền mẫu cây trong đệm nghiền mẫu và cho 100
l/giếng. Ủ bản ELISA qua đêm ở 4-6OC trong hộp ẩm (hoặc 2- 4 giở ở 37
oC). Rửa bản 3
lần bằng đệm rửa (PBS-T). Protein của cây và virus trong dịch cây (nếu có) sẽ liên kết không
đặc hiệu vào thành giếng.
2. Cố định IgG thỏ đặc hiệu virus vào bản ELISA: Hoà IgG đặc hiệu virus trong đệm PBS-T-
ovalbumin và cho 100 l/giếng. Ủ bản ELISA ở 37 OC 2-4 giờ trong hộp ẩm. Rửa bản
ELISA nhƣ bƣớc 1.
3. Cố định IgG đặc hiệu IgG thỏ liên kết AP vào bản ELISA: Hoà IgG đặc hiệu IgG thỏ liên
kết AP trong đệm PBS-T-ovalbumin và cho 100 l/giếng. Ủ bản ELISA ở 37 OC 2-4 giờ
trong hộp ẩm. IgG liên kết AP thứ 2 này là IgG đƣợc tạo ra khi tiêm IgG của thỏ vào 1 loài

212
động vật khác thỏ (chẳng hạn chuột hoặc dê) nên không liên kết với virus mà liên kết với IgG
thỏ đặc hiệu virus ở bƣớc 2. Sau khi ủ, rửa bản nhƣ bƣớc 1
4. Cố định chất nền và đánh giá kết quả: Hoà chất nền trong đệm chất nền theo tỷ lệ 0.6
mg/ml) và cho giếng với lƣợng 100l/giếng. Chất nền ở đây là Nitrophenyl phosphate (NPP).
Bản ELISA đƣợc ủ ở nhiệt độ phòng trong hộp ẩm trong tối 60 phút. Phản ứng hoá học ở đây
là NPP sẽ bị thuỷ phân thành Nitrophenol phosphate dƣới sự xúc tác của enzyme AP.
Nitrophenol phosphate là chất có màu vàng, do vậy có thể nhận biết đƣợc bằng mắt hoặc
bằng máy đọc ELISA (máy so màu). Hiển nhiên, do nồng độ NPP đều giống nhau ở các
giếng nên giếng nào có màu vàng càng đậm chứng tỏ nồng độ enzyme càng cao tức nồng độ
virus trong mẫu thử càng cao. Có thể ngừng phản ứng thuỷ phân bằng cách bổ sung dung
dịch NaOH 3M với lƣợng 25-50 l/giếng.

213
Bài 4. PHÁT HIỆN VIRUS BẰNG PCR/RT-PCR
(Tùy thuộc điều kiện)
Mục đích
Sinh viên biết sử dụng kỹ thuật PCR/RT-PCR để chẩn đoán virus thực vật
Nội dung
Tự thu thập mẫu bệnh ngoài đồng ruộng
Thực hiện phản ứng PCR để phát hiện begomovirus hoặc RT-PCR để phát hiện virus
RYSV. Loại virus có thể thay đổi theo thực tế đồng ruộng.
Yêu cầu
Lớp chia nhóm (5 sinh viên / nhóm)
Mỗi nhóm thực hiện phản ứng PCR hoặc RT-PCR trên một mẫu thu thập ngoài đồng
ruộng
Nội dung
Chiết DNA hoặc RNA tổng số từ mẫu cây
Thực hiện phản ứng PCR hoặc RT-PCR
Kiểm tra kết quả phản ứng bằng điện di agarose
Qui trình chẩn đoán begomovirus bằng PCR dùng mồi chung
Mồi phát hiện
Begomovirus là các virus có bộ gien DNA sợi vòng đơn, kích thƣớc khoảng 2.6 – 2.8 kb.
Sử dụng một cặp mồi chung (degenerate primers) BegoAFor1 và BegoARev1 đƣợc thiết kế ở
vùng bảo thủ của phân tử DNA-A để phát hiện begomovirus (ví dụ trên cây cà chua bị bệnh xoăn
vàng lá). Sản phẩm của phản ứng ~ 1.2 kb. Vị trí mồi trình bày ở Hình 1.
Hình 1. Sơ đồ tổ chức bộ gen của begomovirus và vị trí cặp mồi chung
Chiết DNA tổng số t mô cây
Chuẩn bị
1. Đệm chiết (đệm CTAB, Cetyl trimethylammonium bromide)
2. Chloroform: Isoamylalcolhol (24:1)
3. β Mercaptoethanol (BME)
4. Ethanol 70%
5. 2-Propanol
6. Đệm TE (Tris EDTA)

214
7. Chày, cối sứ (hoặc tube 1.5 mL và chày nhựa): rửa sạch ngâm trong dung dịch NaOH 3M
(1 ngày), rửa lại bằng nƣớc cất rồi hấp vô trùng.
8. Tube 1,5ml
9. Bút viết kính
10. Pipet (1000, 200) và tip
11. Máy li tâm để bàn (15000 g)
12. Bể nhiệt
Quy trình chiết
1. Ủ đệm chiết CTAB (cứ 1 mẫu chiết cần 1ml đệm) đã bổ sung BME (với tỉ lệ 1ml đệm
CTAB :10 μL BME) trong bể nhiệt ở nhiệt độ 60oC trong 30 phút.
2. Lấy 0,1 g mô lá bệnh nghiền với 1 ml CTAB bằng chày cối sứ (nghiền đến khi dung dịch
thu đƣợc là một thể đồng nhất). Chuyển dịch nghiền vào tube 1,5 ml. Ủ ở 65 oC trong 10
phút ở bể nhiệt.
3. Li tâm 13000 g trong 5 phút, thu dịch trên tủa (700 μL) cho vào tube mới
4. Cho Chloroform: Isoamylalcolhol (24 :1) vào tube với tỉ lệ thể tích tƣơng đƣơng. Đảo đều tube.
5. Li tâm 13000 g trong 5 phút, thu dịch trên tủa (600 μL).
6. Lặp lại các bƣớc 4, 5 thêm một lần nữa và thu dịch trên tủa (400 μL).
7. Cho thể tích tƣơng đƣơng (400 μL) 2-propanol lạnh vào tube. Đảo đều tube và để lạnh (-
20 oC) 30 phút.
8. Li tâm 13000 g trong 15 phút. Loại bỏ dịch phía trên, thu lấy cặn DNA tổng số.
9. Rửa cặn bằng Ethanol 70% hai lần (mỗi lần 0.7 – 1 mL)
10. Làm khô cặn DNA bằng cách mở nắp tube, để ở nhiệt độ phòng trong 30 phút (trong buồng cấy).
11. Hòa cặn trong 20-50 μL nƣớc cất 2 lần vô trùng hoặc đệm TE và bảo quản dich DNA ở -
20 oC
Thực hiện phản ứng PCR
Cho vào mỗi tube PCR loại 0.5 mL các thành phần sau:
Thành phần Thể tích cần lấy (μL)
Dd H2O 19.3
Đệm PCR (10X) 2.5
dNTPs (10 mM mỗi loại) 0.5
Mồi xuôi dòng: BegoAFor1 (20 μM) 1.0
Mồi ngƣợc dòng: BegoARev1 (20 μM) 1.0
DNA polymerase (Taq) (5U/ μL) 0.2
DNA tổng số 0.5
Tổng thể tích 25
Cho vào máy PCR và thực hiện phản ứng PCR với điều kiện phản ứng sau:

215
Nhiệt độ (oC) Thời gian Số chu kỳ
Khởi đầu biến tính 92 4 phút 1
Các chu kỳ tổng hợp
92 35 giây
30 50 35 giây
72 1 phút
Kết thúc 72 5 phút 1
Qui trình chẩn đoán RYSV gây bệnh vàng lụi lúa bằng RT-PCR dung mồi chung
RYSV là một virus có bộ gen RNA sợi đơn, cực âm, không phân mảnh, kích thƣớc ~ 14 kb.
Sử dụng cặp mồi đặc hiệu RY-L-F2 và RY-L-R2 thiết kế trên gen L (large) (Hình 2) để phát hiện
virus bằng RT-PCR trên mẫu cây lúa bị bệnh vàng lụi.
Mồi phản ứng
Hình 2. Tổ chức bộ gen RYSV và vị trí cặp mồi đặc hiệu
Chiết RNA tổng số t mô cây
Chuẩn bị
1. Đệm chiết (đệm CTAB)
2. LiCl 10 M
3. Chloroform: Isoamylalcolhol (24:1)
4. β Mercaptoethanol (BME)
5. Ethanol 70%
6. Chày, cối sứ (hoặc tube 1.5 mL và chày nhựa): rửa sạch ngâm trong dung dịch NaOH 3M
(1 ngày), rửa lại bằng nƣớc cất rồi hấp vô trùng.
7. Tube 1,5ml
8. Bút viết kính
9. Pipet (1000, 200) và tip
10. Máy li tâm để bàn (15000 g)
11. Bể nhiệt
Quy trình chiết
1. Ủ đệm chiết CTAB (cứ 1 mẫu chiết cần 1ml đệm) đã bổ sung BME (với tỉ lệ 1ml đệm
CTAB :10 μL BME) trong bể nhiệt ở nhiệt độ 60 oC trong 30 phút.
2. Lấy 0,1 g mô lá bệnh nghiền với 1ml CTAB bằng chày cối sứ (nghiền đến khi dung dịch
thu đƣợc là một thể đồng nhất). Chuyển dịch nghiền vào tube 1,5 ml. Ủ ở 65 oC trong 10
phút ở bể nhiệt.
3. Li tâm 13000 g trong 5 phút, thu dịch trên tủa (700 μL) cho vào tube mới

216
4. Cho Chloroform: Isoamylalcolhol (24 :1) vào tube với tỉ lệ thể tích tƣơng đƣơng. Đảo
đều tube.
5. Li tâm 13000 g trong 5 phút, thu dịch trên tủa (600 μL).
6. Lặp lại các bƣớc 4, 5 thêm một lần nữa và thu dịch trên tủa (400 μL).
7. Cho ¼ thể tích LiCl 10 M (100 μL) vào tube. Đảo đều tube và để lạnh (-20 oC) 30 phút.
8. Li tâm 13000 g trong 15 phút. Loại bỏ dịch trên tủa, giữ lại cặn RNA tổng số.
9. Rửa cặn RNA bằng Ethanol 70 % hai lần (mỗi lần 0.7 – 1 mL)
10. Làm khô cặn RNA bằng cách mở nắp tube, để ở nhiệt độ phòng trong 30 phút (trong buồng cấy).
11. Hòa cặn trong 20-50 μL nƣớc cất 2 lần vô trùng hoặc đệm TE và bảo quản dịch RNA ở -
80 oC
Thực hiện phản ứng PCR
Cho vào mỗi tube PCR loại 0.5 mL các thành phần sau:
Thành phần Thể tích cần lấy (µl)
H2O 15. 5
Dream Taq Buffer 5X 2.0
dNTPs 0.3
Mồi RY-L-F2 0.3
Mồi RY-L-R2 0.3
Reverse Aid enzyme (fermentas) 0.3
Dream Taq enzyme (Fermentas) 0.3
RNA tổng số 1
Tổng 20
Cho vào máy PCR và thực hiện phản ứng RT-PCR với điều kiện sau
Nhiệt độ ( oC) Thời gian Số chu kỳ
Tổng hợp cDNA 50 30 phút 1
Khởi đầu biến tính 92 4 phút 1
Các chu kỳ tổng hợp
92 35 giây
30 50 35 giây
72 1 phút
Kết thúc 72 5 phút 1
Kiểm tra kết quả PCR hoặc RT-PCR bằng điện di agarose
Chuẩn bị
1. Pha đệm điện di TAE 1x từ dung dịch gốc (TAE 5x): lấy 100 ml TAE 5x cho vào cốc đong,
bổ sung thêm 400 ml nƣớc cất, lắc đều.

217
2. Chuẩn bị bản gel: Cân 1g agarose cho vào trong lọ, bổ sung 100 ml TAE 1x, lắc đều và cho
vào lò vi sóng để hoà tan agarose (khoảng 3 phuts). Sau khi agarose đã tan hoàn toàn, bổ
sung 2 µl Ethidium Bromide, lắc đều, để lọ ở nhiệt độ phòng. Khi dung dịch agarose 1% nằm
trong khoảng 50oC thì đổ vào khay gel đã đặt sẵn lƣợc (dày khoảng 0.6 cm). Để khay gel ở
nhiệt độ phòng khoảng 30 phút để đông gel.
3. Chuẩn bị mẫu điện di: Lấy các tube 0,5 ml sạch, đánh số thứ tự. Cho vào mỗi tube 2 µl đệm
cài gel (Loading Dye) 6x và 10 µl sản phẩm PCR hoặc RT-PCR. Trộn đều và ly tâm nhanh
(spin) trong 5 giây.
Điện di
1. Sau 30 phút để khay gel ở nhiệt độ phòng thì tiến hành rút lƣợc (cầm chính giữa lƣợc, rút đều
tay để không vỡ giếng).
2. Đặt cả khay gel vào bể điện di, đổ dung dịch đệm TAE 1x phủ ngập gel (gel ngập sâu
khoảng 1mm).
3. Dùng pipet cho tất cả mẫu điện di (12 µl) vào giếng. Chú ý: (i) luôn cho thang DNA (DNA
marker) vào bản gel (thƣờng là giếng đầu tiên) và (ii) thao tác phải cẩn thận để tránh tràn
mẫu.
4. Cắm nguồn điện cho máy điện di ở hiệu điện thế 100 V trong 30 phút.
5. Sau khi điện di, đặt bản agarose lên máy soi gel (phát tia UV) và quan sát các băng DNA
hình thành và so sánh với thang DNA để đánh giá kết quả. Chụp ảnh nếu cần thiết.