vigilance levels & health in hawaiian spinner
TRANSCRIPT

VIGILANCE LEVELS & HEALTH IN HAWAIIAN SPINNER DOLPHINS
By
Megan O’Toole
Dr. Dave Johnston, Advisor
April 2012
Masters project submitted in partial fulfillment of the
requirements for the Master of Environmental Management degree in
the Nicholas School of the Environment of Duke University
2012

MeganO’Toole
2
Abstract
Overthepastseveraldecades,Hawaiianspinnerdolphinshavebecomea
speciesofinterestforbothscientistsandtouristsalike.Spinnerdolphinscanbe
foundrestinginthebaysofcoastalHawaiialmostdaily,andtheirhabitshavebeen
notedbytheburgeoningecotourismindustryandincreasedhumanpresence.While
noindepthstudieshavebeenperformedonthedolphinpopulations,these
increasedinteractionshaveresearchersconcernedaboutthevulnerabilityofthe
dolphinpopulation.
Dukas&Clark(1995)hypothesizethatakeyunexploredfactorinanimal
healthisvigilancelevel,ortheabilityoftheanimaltoprocesscomplexpatterns
suchasforagingordetectingpredatorsbasedontheamountofresttheyareableto
obtain.Asthelifestyleofspinnerdolphinsputsthematextremeriskforlowered
vigilancelevels,theequationsputforwardinthearticlewereusedtocreateabio‐
energeticsmodelthatwouldillustrateitspotentialeffectsontheirabilitytocapture
preyandevadepredators.
Whileshort‐termvigilancelossdoesnotappeartobesignificantlydamaging
tothedolphin’sabilitytofeedandprotectitself,along‐termlossmaygreatlyaffect
aspinnerdolphin’soverallhealth.Thesefindingsindicatethatthecurrenthuman
presenceinthebaysduringthespinnerdolphins’restinghourshasthepotentialto
detrimentallyaffecttheirhealthandconsequentlytheirpopulationnumbersand
shouldserveasastartingpointforfurthervigilancebasedresearch.
Introduction

MeganO’Toole
3
Spinnerdolphinshavebeenstudiedforoverfortyyears,butstillmuchabout
theirbiologyisunknown.Thefederalgovernmentprovidesfortheirwellbeing
undertheMarineMammalProtectionAct(1972)butnostatelawsorprograms
providethesamelegalcoverage.InthebaysnearKona,Hawaii,thepresenceof
spinnerdolphinsisaneverydayoccurrence.Inadditiontothepodsofdolphins,
hundredsofpeople,bothtouristsandlocals,swimmingandboatingalsofillthebays
eachday(Timmeletal.2008).Theincreasingpresenceofhumansinbaysiscausing
morefrequentinteractionsbetweenthehumansandthedolphins,buttheeffectsof
theseinteractionsarestilllargelyunknown.ThewesternsideofHawaii’sbigisland
isoneofthemostnotableplaceswherehumaninteractionwithspinnerdolphins
occursdaily(Delfour2007).
Tobetterunderstandtheuniquenatureofthespinnerdolphinandtheir
susceptibilitytohumanactivities,theirdailyhabitsshouldalsobeaddressed.
Spinnerdolphinsaresmallcetaceanseasilyidentifiedbyathreetone‐greypattern
andgettheircommonnamefromtheaxialspinsandleapstheyperformduring
socialencounters(“SpinnerDolphin”2011).Theyareuniqueinthatunlikemost
othercetaceanstheyforageatnightintheopenoceanfeedingonmesopelagicfish,
shrimp,andsquid(Benoit‐Bird&Au2009)andspendthedayrestingand
socializinginbays(Norris&Dohl1980).Preyofthespinnerdolphinliveatabout
400to700mbeneaththesurfaceofthewaterduringthedaybutbecausespinners
arethoughttoonlybeabletodivetodepthsof400mmaximumtheymustwaituntil
nightfall,whenthemesopelagiclayerrisestodepthseasilyreachedbythedolphins
forabout11hoursperday(Benoit‐Bird2003).Theleadinghypothesisforrestingin

MeganO’Toole
4
thebaysisthatbyrestinginthebaysduringtheday,thepredationriskismuch
lowerthanintheopenoceanbecauseoftheenclosedareaaswellastheincreased
abilitytoseetheshadowofapredatoronthesandybottom(Heenehan2010).Just
aswithpeople,restingallowsthedolphinstorecoverfromtheirdaily,ornightly
efforts.Thedolphinsrestwithuni‐hemisphericsleepmeaningonlyonehalfofthe
brainisshutdownatatimewhiletheotherenablestheanimaltoperformfunctions
necessaryforsurvival(Cirelli&Tononi2008).Theabilitytosleepwithonlyonehalf
oftheirbrainatatimeallowsthedolphinstocontinueswimmingandreturntothe
surfacetobreathewhilealsowatchingforpredatorssotheotherhalfoftheircan
brainrest(Cirelli&Tononi2008).Currently,theirbehavioristocomeintothese
baystorestwheretheywillbelesslikelytobeattackedbypredatorsandcansleep
withlessinterruption.However,withthegrowingpresenceofhumans,this
behaviorisbeingdisruptedastheywakeupduetotheunexpectedcommotion.
MoststudiesfocusonthewesternsideofthebigislandofHawaii.Thefour
baysthatarecurrentlybeingstudiedindepthareKeauhako,Kealeakekua,
Honaunau,andMakakobays.Eachofthesebayshistoricallyhavehadfrequent
spinnerdolphinpresenceduringthedayandsohavebecomefavoredtostudy
spinnerdolphinandhumaninteractions(Norrisetal.1994).Thesebaysare
characterizedbyshallowwater,sandybottoms,areprotectedfromwindandstrong
oceancurrents,andarealllessthan1.5kmacross.Byhavingthissetof
characteristics,thebaysareideallocationsforspinnerdolphinrestingareas(Norris
etal.1994).

MeganO’Toole
5
Additionally,Konabaysarealsohometoanisolatedsub‐stockofspinner
dolphins.AccordingtotheNationalMarineFisheriesService(NMFS),theentire
chainofHawaiianislandsishosttoasinglestockofspinnerdolphins,butrecent
dataindicatesthatsixindividualstocksaremostlikelypresentalongtheislands
(Andrewsetal.,2010).TheMMPAdefinitionofastockis“agroupofmarine
mammalsofthesamespeciesorsmallertaxainacommonspatialarrangement,that
interbreedwhenmature,”andAndrewsetal.foundthatlittlegeneticmixingof
individualsinbetweenthesub‐stocksoccurs.Consequently,shouldoneofthe
individualpopulationsaroundHawaiibecomeunsustainablysmall,itisunlikelylike
thatthedolphinstockselsewhereinHawaiiwouldbeabletobolsterthedwindling
populationthroughemigrationtothesmallerstock.Additionally,spinnerdolphins
havebeenobservedtohave“long‐termsitefidelity,”meaningthatevenifconditions
ofthebaysdeterioratetothepointwherethedolphinsaredetrimentallyaffected,
theanimalswillnotseekbetterrestingbayswhichcouldexacerbatetheissueof
isolatedstocks(Marten&Psarakos1999).Bynotmovingtomorerestful,less
human‐influencedareasthedolphinsnotonlywillbecontinuingtheirexposureto
poorconditionsbutwillalsoeliminatethepossibilityofmixingwithahealthier
population.
Withtheincreasedinteractionsbetweenhumansanddolphinscomes
increasedconcernforthedolphins’wellbeingandwhetherpeople’sactivitiessuch
askayakingorscubahaveaneffectontheirbehaviorandconsequentlytheirhealth.
Thehumanpresenceinthebaysisunliketoabateanytimeintheforeseeablefuture

MeganO’Toole
6
asmillionsofdollarsayeararebroughtintoHawaiifromtouristswhowishtobe
abletoseeandorswimwiththespinnerdolphins
NoofficialpopulationestimateisavailablefortheHawaiianspinnerdolphin
(DepartmentofCommerce2011).However,arecentstudyputsthebestavailable
populationestimateforthebigislandbetween600and700dolphinsinthearea
(Hilletal.tobepublished).Theestimateisconcerningwhencomparedtothe
initialabundanceestimatedonebyNorrisof1000to2000dolphinsaboutthirty
yearsago(Norrisetal.1994).Thedecrease,however,cannotbeattributedtoany
specificfactorforseveralreasons.Littleisknownontrendsofthedolphin
populationchangingovertimeduetoalackofconsistentresearchanddifficulty
findingconclusivecorrelatingfactors.
Formanypeople,swimmingwiththedolphinshasnowbecomeviewedasa
right,whetheritisbecausetheyareontheirdreamvacationorthey’vedoneitall
theirlives.Somefeelaspiritualneedtoswimwiththedolphinswhileothersview
themasaliensfromanotherworld.Boatingtoursthatadvertiseswimmingwith
dolphinsbeganinHawaiiaround1991andhavenowbecomeamajorportionofthe
tourismindustry.Ecotourismasawholehasbeenincreasingannuallyatarateof
about10%(Delfour,2007).Whateverthereason,eachdaydolphinsandhumans
sharethesebaysandwithincreasingfrequencywillinteractwitheachother.This
includestourisminHawaii,muchofwhichoccursonthewaterandhighlightsall
varietiesofmarinemammalsincludingpilotwhales,beakedwhales,andspinner
dolphins.FabienneDelfourconductedastudyinwhichtheeffectsofkayaking,

MeganO’Toole
7
boatingandswimminginthebaysonspinnerdolphinbehavior(2007).Thestudy
foundthatasthenumberofhumanactivitiesinthebayincreased,thenumberof
timesachangeindolphinswimmingdirectionoccurredfollowinganencounterwith
humans(Delfour2007).Otherstudieshavefoundalterationsinanimaldistribution
(Lammers2004),speed(Ribeiroetal.2005),swimmingpatterns(Acevedo1991),
andbehavioratthesurface(Hastieetal.2006).Additionally,Samuelsetal(2000)
reportedthatthenumberofdolphinsreturningtothebayswithhighesthuman
presencedecreasedovertime,indicatingpoorconditionsfordolphinresting.
Humanactivityisclearlyaffectingthebehaviorofthedolphins,buttheextentto
whichthismayaffecttheirhealthisstillunknown.
Inmanycases,lawsonconservationorenvironmentalissueshaveboth
federalandstatecounterparts.However,withmarinemammalsthisisnotthecase.
Onthestatelevel,Hawaiihaslawsforbiddinginteractionwithanimalswithin
sanctuaries,butasnoneofthebaysareprotectedthestatedoesnothaveany
enforcementpower.OfthefourbaysstudiedoffoftheBigIslandofHawaii,atleast
oneisconsideredaMarineLifeConservationDistrictbythestateofHawaii,but
theirlawsmakenomentionofdolphinswithinthebay,theyonlyapplytonon‐
marinemammalspecies.In2006,thestateofHawaiiwasauthorizedtoassistinthe
enforcementofdolphinprotectionbutsofarthishasbeenmostlylimitedto
spreadingtheawarenessofMMPAguidelines(“DepartmentofLandandNatural
Resources”2006).

MeganO’Toole
8
TheonlylegislationthatcurrentlyappliestospinnerdolphinsistheMarine
MammalProtectionAct(MMPA).ThisisbecausetheMMPAprohibitsthecreationof
anystatepoliciesdealingwiththemanagementofmarinemammals,andsoHawaii
cannotcreatetheirownregulationstodealwithanypotentialissuesarisingfrom
humaninteractionswiththedolphins.Theenforcementofdolphinsunderthe
MMPAfallstoNMFSProtectedResources,andthenisdividedupintosixregions,
withthePacificIslandRegionalOffice(PIRO)dealingwiththeHawaiianSpinner
dolphins.UnderTitleI,whichaddressestheconservationandprotectionofmarine
mammals,theMMPAmakesitillegaltoharassorattempttoharassmarine
mammals.Thisincludesthealteringoftheirbehaviors,whichmayoccurinthese
bayswhenpeoplearecomeintoclosecontactwiththedolphins(MMPA1994).As
theratesofhumanactivitiesintheHawaiianbaysareincreasing,NMFSisreceiving
highernumberofcomplaintsthattheswimmersandboatersareharassingthe
dolphins(Timmeletal.2008).Whilethe1994amendmentstotheMMPAessentially
makesswimmingwithdolphinsillegalunderLevelBharassment,or“havingthe
potentialtodisturbamarinemammalstockinthewildbycausingadisruptionof
behavioralpatterns”(MMPA1994),lackofconclusivefindingsonthepartof
researchersaswellasthedifficultyofprovingsuchoccurrencesincourthavekept
theregulationdifficulttoenforce.ManyofthestudiesperformedintheHawaiian
baysarethereforeattemptingtoprovethisportionoftheMMPAisbeingviolatedin
ordertoproveacauseforstricterpolicyormoreenforcementinthesebays.

MeganO’Toole
9
AnotheroneofthegoalsoftheMarineMammalProtectionActistomaintain
thestocksofmarinemammalsabovetheoptimumsustainablepopulation(MMPA
1994).“Optimumsustainablepopulation”isthenumberofanimalsthatwillresult
inthemaximumproductivityofthepopulationkeepinginmindthecarrying
capacityofthehabitatandthehealthoftheecosystemofwhichtheyforma
constituentelement.Spinnerdolphinsarecurrentlyconsideredas“depleted”by
NMFS,whichmeansthatthespecieshavefallenbelowtheiroptimalsustainable
level(“SpinnerDolphin”2011).ThisdegradationoftheirstatuspromptedNMFS
PIROtolookforwaystomitigatehumaninteractionswiththedolphinsonaregular
basis.
Lackofenforcementinthebaysaswellasthedifficultyofprovingwhether
aninteractionhasalteredthebehaviorofthedolphinspromptedNMFStopublish
voluntarySpinnerdolphinviewingguidelines.Theseguidelines,whicharenomore
thanthenormalsuggestedbehaviorwhenencounteringanymarinemammal,are
distributedtotouroperatorsaspartofanoutreachprogramtoincreaseawareness
oftheMMPAandthepossibleeffectsofhumaninteractionsinanattempttodeter
humanpursuitofdolphinswithoutnewregulations.Theguidelinesinstructboatsto
stayover50metersawayfromthedolphinsatalltimes,tolimitobservationtimeto
halfanhour,andtomakesurethedolphinsarenotencircledortrappedbetween
boatsortheshoreatanytime(“SpinnerDolphin”2011).Additionally,outreach
programstotouristsexploringtheareasalertthemtothenocturnalnatureofthe

MeganO’Toole
10
spinnerdolphinsaswellastherequirementsoftheMMPA.Inthesummerof2011,
NOAAbeganimplementationoftheDolphinSMARTprograminHawaii,where
touristsareabletofindalistofdolphintouroperatorswhohaveagreedtoabideby
theNOAAdolphinviewingguidelines(“SwimmingwithSpinnerDolphins,”2011).
In2006,NMFSissuedanoticeofintenttocreatenewpoliciestodealwith
humaninteractionswiththespinnerdolphinsinthesebays(Departmentof
Commerce,2006).Inearly2011,NMFSannouncedtheirplanofclosingKealakekua,
Honaunau,MakakoandKauhakobaysduringthemorninghourswhenspinner
dolphinshavemostfrequentlybeenobservedtoberesting(Johnston,2011).These
fourbayshavebeenstudiedintensivelyforthepasttwoyearsinordertocollect
dataforanalysisonthebehaviorofdolphinsinthepresenceofhumans.Afterthe
closurethestudywillcontinueinanattempttodeterminewhethertheabsenceof
humanactivitybringsaboutanychangeinbehavioralpatterns.Alreadybacklash
hasbeenseenagainstthisnewpolicy,andnotwithoutwarrant.A2009reportby
theInternationalFundforAnimalWelfareestimatestheannualrevenuefortourist
relatedboatingactivitiesinHawaiiisover$40million.Anordertoshutdownfour
ofthemajortouristbaysonthebigislandduringprimebusinesshourscouldcause
millionsofdollarsinlossesforthetouroperatorsfrequentingthoseareas(“Whale
Watching”2009).WhileprotectingmarinemammalsisapriorityforNMFS,the
alienationofalargerevenuestreaminHawaiimaynotbethebestlongtermplanif
protectionofspinnerdolphinhabitatistocoexistwiththetourboatindustry.This

MeganO’Toole
11
isespeciallycontentiousasNMFSisoftenunabletoprovetheMMPAhasbeen
violated.
Oneconceptwhichhasnotbeenexploredindepthasawaytomeasure
spinnerdolphinhealthandcouldgreatlyaidinthediscussionofpotential
ramificationsofhumanactivitiesistheeffecttheamountofsleep,orlackthereof,
hasonthespinnerdolphin.InDukas&Clark’s1995articleSustainedvigilanceand
animalperformance,theydiscusstheconceptthatalackofrestleadstotheinability
ofthecentralnervoussystemto“sustainahighqualityofinformationprocessing
foranextendedperiodoftime.”A2008studyconcludedthatdolphinssubjectto
sleepdeprivationweremorelikelytohaveslowerresponsetimestostimuli,
indicatingthatacontinuousinterruptionofsleeppatternscouldpotentiallyincrease
themortalityriskofspinnerdolphinsduetoinabilitytosuccessfullyforageor
predation(Cirelli&Tononi2008).However,theseverityorproportionalresponse
tolackofsleephasnotyetbeenmeasured.Dukas&Clarkcreatedamodelwhich
measuresthevigilance,inthisinstancedefinedasa“stateofbehavioralalertnessto
predators…ageneralconditionofenhancedabilitytoprocessinformation”anda
seriesofequationswhichcorrelatemeasuresofhealthsuchasabilitytoforagefor
food,mortalityrisk,andnetenergygain.Usingatechniquesuchasmodelingwould
allowforthestudyofpossibleeffectsofhumanpresenceinthebayswithout
interferingbybecomingapresencethemselves.Fromtheseequations,amodelwas
createdspecificallytargetedtowardsthespinnerdolphininanattempttoassessthe
relationshipbetweenrest,vigilancelevel,andhealth.
MaterialsandMethods

MeganO’Toole
12
Inordertodisplayspinnerdolphinbehavioraldatawithinamodeling
framework,theenvironmentalmodelingprogramSTELLAwasused.Equations
createdbyDukasandClark(1995)wereemployedwithsomeslightmodifications
duetotheneedtotakeintoaccountcharacteristicsspecifictothespinnerdolphins
inadditiontothefeasibilityofcreatingaworkingmodelfromequationsthathave
previouslyonlybeenappliedtheoretically.Becausetheequationsinthepaperare
designedforanimalsingeneral,whencreatingaStellamodelvaluessuchas
metaboliccostoflivingandpreydensitywhileforaginghavetobealteredinorder
totailorthemodeltospinnerdolphins.Valuesfortheseparametersweretaken
frommultiplepapersdiscussingtheforaging,socializing,andsleepinghabitsofthe
dolphins(e.g.referencesandseebelow).
TheBasicModel
Thefirstsimulationrunfromthemodeldemonstratedanidealsituationin
whichthevigilanceofadolphinbeginsatthemaximumvigilancelevelbefore
foragingandendsatmaximumvigilancelevelafterarestingbout(seeTable1).
Table 1. Symbols and values used in the initial model
Symbol Description Value
t Time 0 < t < 24 υ Vigilance level 0 < υ < 100 γ Resting multiplier τ Time delay between foraging and resting 8 hours η Time Delay multiplier α Rate of vigilance decrement during foraging activity .07/hr β Rate of vigilance recovery during rest .19/hr θ Foraging multiplier
λ Number prey encountered per hour 215.7 animals
f Long-term average rate of food intake

MeganO’Toole
13
€
µ Predation risk µ Chance of encountering a predator
€
ρ Percent chance of encountering a predator while resting 0.002
€
ρ Percent chance of encountering a predator in the open ocean 0.02 M Overall mortality risk
c Metabolic cost per hour foraging 173.75 kcal/hr
€
Eprey Calories of prey item E Net rate of energy gain F Total expected lifetime energy gain L Length of prey
Thefirststepindemonstratingthevariationsinvigilanceinspinnerdolphins
isdeterminingtherateofchangeofvigilanceoverthecourseoftheiractivities.
ConstantproportionalratesofchangeareassumedintheequationsbyDukas&
Clarkof:
‐αυwhileforaging β(1‐υ)whileresting
Vigilancelevelinthismodelisdemonstratedbyν,where0<υ>100,with0
representingthedolphinatadepletedvigilanceleveland100representing
maximumvigilance(Dukas&Clark1994).Themodelwasrunforasimulation
period24hourswiththestartingpointusingavigilancelevelof100,assuming
maximumvigilanceafteraperiodofrestasthedolphinsswitchtoforaging
behaviors.Thevalueforrateofvigilancedecrement,orα, issetataproportionof
vigilanceof.07perhourandβissetatarateof0.19vigilancerecoveryperhour.
Whilethevaluesofα andβhavenotbeenrecordedinanystudy,theassumptionis
madethattherateofrecoverywouldbemuchhigherthantherateofdecrementin
animalsasamuchlargerproportionoftimeisspentawakethanasleep.For

MeganO’Toole
14
simplicitypurposes,vigilancelevelonlyincreasesduringrestingperiodsandonly
decreasesduringforagingbouts.Giventhatthedolphinsforageforan
approximately11hourseachnightwhilethemesopelagiclayerisavailabletothem
(Benoit‐Bird),itwouldmeanthatthedolphin’sdecrementtime,forthepurposeof
thismodel,wouldbelimitedtothose11hours.Additionally,KennethNorris
observedrestingspinnerdolphinstoremaininthebaysforaminimumof5hours
(1994).Therefore,thewindowoftimeinwhichvigilancerecoveryoccursissetto5
hoursper24hourcycle.Tocreatetheidealsimulation,anydecrementthatoccurs
duringtheforagingperiodwouldneedtobefullyrecoveredduringtherestingbout.
Therefore,withaninitialplaceholderrateofdecrementof.07percentageof
vigilancelostperhouroveran11hourperiod,therateofrecoverywouldhavetobe
.19perhourinordertocreatethebaseline.Inadditiontohavingboutsofforaging
andresting,atimedelayisalsotakenintoaccountforthetransitionsbetween
foragingandresting.Asdolphinsdonotforageinthesamelocationtheyrest,travel
istakenintoaccountaswellassocializationtime.Forthepurposesofthismodel,
movingfromtherestinglocationtotheforaginglocationorviceversaandanytime
spentsocializingiscategorizedasatimedelayinwhichthevigilancelevelneither
diminishesnorincreases.Forthismodel,thetimedelayissetat4hoursbetween
eachtransition,totaling8hoursperday.Consequently,themodelwouldrunwith
theforagingperiodforthefirstelevenhours,afourhourtimedelayfortraveland
socializing,arestingperiodforthenextfivehoursandthenswitchingfromrestto
foragingfor21:00‐24:00hrs.

MeganO’Toole
15
Theequationusedtomodelthevigilancedynamicsovera24hourperiodis
then
υ=‐αυθ + γβυ+ηυ
where
θ =1 t<110t>11
γ=0 t<16,t>20 1 16>t<20
η =0 t<11,16<t<20111>t<16,t>20
Assumingtheaveragerateoffoodintakeduringaforagingboutis
proportionaltothevigilancelevelatthattimeandthatanypreyencounteredbythe
dolphiniscaughtandconsumed
Averagerateoffoodintake=λυ
wecanthencalculatethetheoreticalamountoffoodadolphinwouldbeableto
consumeduringanygivenforagingperiod
€
f (υ) = λυ0
24
∑
AccordingtoDukas&Clark(1995),increasingtheamountoftimespent
foragingshouldincreasetherateatwhichtheyareabletoconsumepreydueto
increasedtimetoencounterfood.Foodintakeisalsousedtoestimatetheoverall
healthofthedolphinassumingthattheamountofpreycaughtcanindicatefuture
successintermsofgainingsizeandthusbecominglessatriskfrompredationas

MeganO’Toole
16
wellasbeinganindicatoroffuturereproductivehealth(Read2011).Therefore,the
higherthefoodintakeoverthedolphin’sdailyforagingperiod,thehigherthe
assessmentofthedolphin’shealth.
MortalityRiskandMetabolicCosts
Upuntilthispoint,predationandmetaboliccostarenotaccountedforinthe
equations,butbyutilizinganotherequationbyDukas&Clark(1995)predationrisk,
orthedolphin’ssusceptibilitytoapredationattemptcanbecalculatedby
=
Thisequationsimplyillustratesthatasforagingtimeprogressesandcauses
vigilancetodecreaseadolphin’sriskofpredationwillrise.Foragingtimeinthis
instanceincludestheactivitiesof“bothfeedingandscanningtheenvironmentfor
predators”(Dukas&Clark,1995).Withoutrestthisactivityreducesthecapacityof
thedolphintobothfeedandlookoutforpredatorsinthefuture.Assumingthereare
noothercausesofmortalityandatmaximumvigilancethedolphinwillbeableto
escapeanypredationattempts,DukasandClarktranslatethispredationriskintoan
overallmortalityrisk(M)shownasanincreasingfunctionofforagingeffortby
Ametaboliccost(c)isthentakenintoaccount.Byincludingthemetaboliccostinto
theequation,overallenergygaincanbecalculatedandanestimatedoverallcaloric
intakecanbeproduced,leadingmostlikelytoamoreaccuratepredictionoffuture
dolphinhealth(Dukas&Clark1995).Netdailyenergygainismodeledbythe
equation

MeganO’Toole
17
Themetaboliccostoflivingalsovariesbetweenrestingandforagingstates.
Atalltimes,thedolphinismovingaswellasperforminginvoluntarybodily
processeswhichrequireenergyofmaintenance.Becausenostudieshavebeen
publishedonthemetabolicratesofspinnerdolphins,thecalculatedvaluescame
fromstudiesonbottlenosedolphins.Yazdi,etal(1999)discussthevariousenergy
costsassociatedwithbaselineswimmingaswellasthecostofincreasedspeedsand
diving.Forthesimulation,themodelisdividedintotwometabolicrates,therates
whileforagingandtherateswhilenotforaging(duringthetimedelayandresting
bouts).Therestingmetabolicrateofbottlenosedolphinswasfoundtobeanenergy
consumptionrateof2.15W/kg(Yazdietal.,1999).However,duringperiodsof
efficientswimmingYazdietal.foundthemetabolicratetobeabout10%higher.
Assumingthatthedolphinsareswimmingefficientlyfortheperiodswhilenot
foraging,themetaboliccostmustfactorinthecostoftransportwhilethedolphins
travelandrest(astheyarestillswimmingduringrestingbouts).Themetaboliccost
oftransportduringtheseperiodsisthensettobeat1.16 ,whichconverts
to0.00027 (Yazdietal.,1999).Iftheaveragespinnerdolphinweighs65kg
andisswimmingatthemostefficientrateof2.5m/s(Yazdietal.1999),thenthe
dolphinwillbeusing157.95kilocaloriesperhourwhiletravellingandresting.
Williamsetal.(1999)foundthatthemetabolicrateofdolphinsincreasesupto20%
fromtherestingmetabolicrateduringdivingcausingthemetaboliccostofforaging
tobesetat173.75kilocaloriesperhour.Betweenthesetwometabolicrates,the
averagedailymaintenancerequirementforaspinnerdolphinisabout61kcal/kg.

MeganO’Toole
18
UsingtheresultsofBenoit‐Bird(2003)onpreycaloricvaluesandassuminga
61kcal/kgmaintenancerequirement,thedailyenergyneedsarecalculatedtobe
about3,965kilocaloriesperday.Themainpreyitemsofspinnerdolphinsare
knowntobefish,shrimpandsquid,withanaverageequationmeasuringthecaloric
contentofthedolphin’sdietbeing
Amajorityofthepreyfoundinspinnerdolphins’stomachsweremyctophidfish
withanaveragelengthof7.8cm(Benoit‐Bird2003)andsothiswasusedasthe
lengthfortheaveragepreyitemobtained.Whenpluggedintotheequationabove,
thecaloriccontentoftheaveragepreyofaspinnerdolphinis1,975caloriesor
1.975kilocaloriesandsothisvaluewasusedastheparameterforthecaloriesof
eachfooditemeaten.Usingthe11hourperiodasthelimitforhowlongthedolphin
willhavetoobtain3,965kilocalorieswecandeterminethatinorderforthisenergy
requirementtobemet,atleast2,007.59preyitemsmustbeeateneveryday.With
theassumptionstatedfortheaveragerateoffoodintake,thespinnerdolphinwill
eatallpreyitcomesintocontactwithshoulditbeatitshighestvigilancelevel.As
thevigilanceleveldiminishes,thesmallerpercentageofpreythedolphinwillbe
abletocapture.Therefore,thenumberofpreyencounteredperhourusedinearlier
equationswouldneedtobeatleasttheminimumamountoffoodadolphinneeds
perday.Usingthisdata,thenumberofpreyencounteredperhourissetat215.7
animalssothatwithitsvaryingvigilanceunderidealcircumstancesthedolphincan
meetitsminimummaintenanceenergyneeds.Asfoodavailabilityisnotalimiting
factorfordolphinenergymaintenance(Benoit‐Bird2003),thisnumberwould

MeganO’Toole
19
actuallybealowestimateofpreyencounters.Additionally,spinnerdolphins
activelyseekhighaggregationsofprey(Benoit‐Bird&Au2008)aswellas
cooperativelyforagesotheirforagingareaswouldhavehigherchancesof
encounteringpreythantheopenoceanorthemesopelagiclayerasawhole.
Byusingaconservativeestimateofhowmuchaspinnerdolphinwouldeatina
singleforagingperiodandaddinginthemetaboliccost,wecanthenseewhetherthe
dolphinisabletofulfillitscaloricneedsoverthecourseofthedayandareableto
haveanothercriteriabywhichtomeasurespinnerdolphinhealth.
Creatingsuboptimalconditions
Thefirstsimulationofthemodelwasrununderoptimalconditionsinwhich
thedolphinwasabletorestuntilitsvigilancewasfullyrestoredinordertocreate
baselinenumbersforoutputssuchasfoodintakeperforagingboutaswellasnet
energygainoverthelifetimeoftheanimal.Subsequentsimulationswhenthe
dolphinisonlyabletoobtain80,60,and40percentoftheminimumamountof
sleepobservedbyNorris(1994).Thesesimulationswilldepictpossible
consequencesofprevalenttouristdisruptionofdolphinsrestinginthebays.
Whilemimickingoptimalconditions,themodelissetuptorestorevigilance
toitsoriginallevelattheendofeachdaywhichprovidesthedolphinwiththe
optimalamountofvigilancetoperformthenextday’sactivities.Whilethese
conditionscontinue,thedolphinwillbeabletorestoreitsvigilanceindefinitely.
Shouldsuboptimalconditionsoccur,vigilancelevelswouldthenbealteredwithin
themodel.Takingintoaccountthepossibilitythatlongtermeffectsofsleep
deprivationmaybedifferentthanshortterm,foragingabilityaswellasmortality

MeganO’Toole
20
riskmaybemoreevidentwhenlookedatfromalongtermperspectiveandsothe
simulationwillbeextendedfromitsoriginal24hourperiodtoathirtydayperiodin
ordertoviewpotentiallongtermeffects.
Results
OptimalScenario
Vigilancelevelintheoptimalscenarioisshowntobeginat100%withthe
dolphinbeingfullyabletoprocessitssurroundings,decreaseoverthecourseofthe
foragingboutandnotriseagainuntilthedolphinisabletorestagaininthebays
withthevigilancelevelreturningtomaximumcapacityattheendoftheresting
period(Figure1).
Thevigilancedynamicwasthenusedtomodeltherateoffoodintakeadolphin
wouldbeabletoperform,asfispositivelycorrelatedwithvigilancelevel.Therate
offoodconsumptionishighestatthebeginningoftheforagingboutanddecreases
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
VigilanceLevel(Percentage)
Time(Hours)
Figure1.Vigilancedynamicovera24hourperiod
Vigilance
MeganO’Toole
21
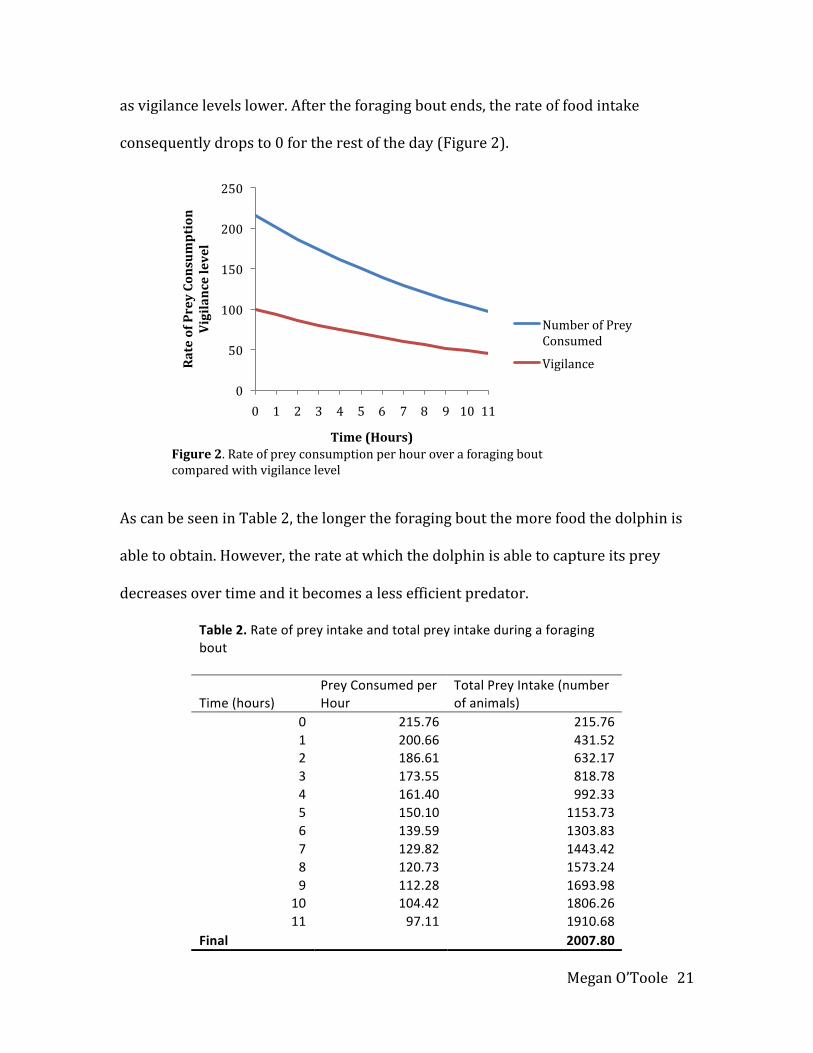
asvigilancelevelslower.Aftertheforagingboutends,therateoffoodintake
consequentlydropsto0fortherestoftheday(Figure2).
AscanbeseeninTable2,thelongertheforagingboutthemorefoodthedolphinis
abletoobtain.However,therateatwhichthedolphinisabletocaptureitsprey
decreasesovertimeanditbecomesalessefficientpredator.
Table2.Rateofpreyintakeandtotalpreyintakeduringaforagingbout
Time(hours)PreyConsumedperHour
TotalPreyIntake(numberofanimals)
0 215.76 215.761 200.66 431.522 186.61 632.173 173.55 818.784 161.40 992.335 150.10 1153.736 139.59 1303.837 129.82 1443.428 120.73 1573.249 112.28 1693.98
10 104.42 1806.2611 97.11 1910.68
Final 2007.80
0
50
100
150
200
250
0 1 2 3 4 5 6 7 8 9 10 11
RateofPreyConsumption
Vigilancelevel
Time(Hours)Figure2.Rateofpreyconsumptionperhouroveraforagingboutcomparedwithvigilancelevel
NumberofPreyConsumed
Vigilance

MeganO’Toole
22
MortalityRiskandNetEnergyGain
Predationriskwasfoundtobenegativelycorrelatedwithvigilancelevels
andsoastheforagingboutprogressed,thedolphin’sriskofpredationrisesasitis
nolongerbeabletoprocessitssurroundingsasthoroughlyasitcouldatthe
beginningoftheforagingbout.Additionally,asthedolphinrestsanditsvigilance
wouldrise,theriskofpredationlowers(Figure3).
Nextanalyzedwastheoverallmortalityrisk,whichdiffersfromthe
predationriskinthatittakesintoaccountthelikelihoodofthedolphin
encounteringapredatorratherthanjustitsriskofbeingpredatedshouldan
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
Percentage
Time(Hours)Figure3.Predationriskandvigilancelevelovera24‐hourperiodwherethepredationriskrepresentsthepercentchance,shouldapredatorattackthatthedolphinfailstonoticethepredator.
Vigilancelevel
MeganO’Toole
23
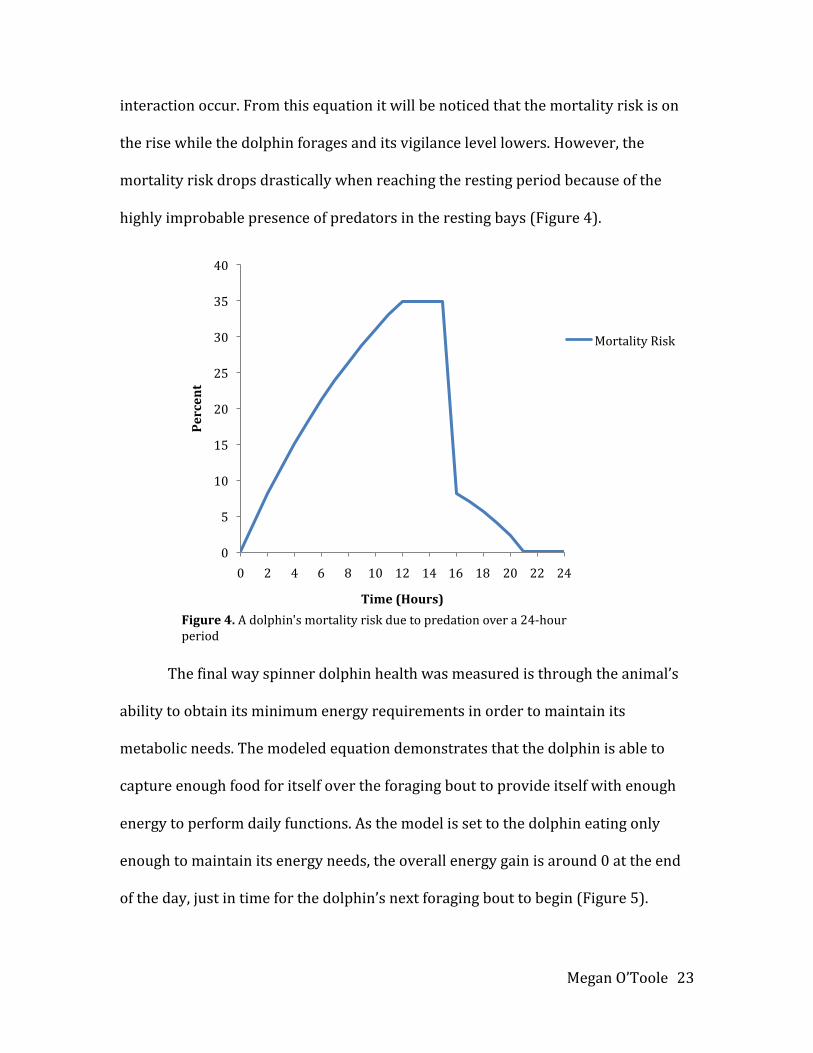
interactionoccur.Fromthisequationitwillbenoticedthatthemortalityriskison
therisewhilethedolphinforagesanditsvigilancelevellowers.However,the
mortalityriskdropsdrasticallywhenreachingtherestingperiodbecauseofthe
highlyimprobablepresenceofpredatorsintherestingbays(Figure4).
Thefinalwayspinnerdolphinhealthwasmeasuredisthroughtheanimal’s
abilitytoobtainitsminimumenergyrequirementsinordertomaintainits
metabolicneeds.Themodeledequationdemonstratesthatthedolphinisableto
captureenoughfoodforitselfovertheforagingbouttoprovideitselfwithenough
energytoperformdailyfunctions.Asthemodelissettothedolphineatingonly
enoughtomaintainitsenergyneeds,theoverallenergygainisaround0attheend
oftheday,justintimeforthedolphin’snextforagingbouttobegin(Figure5).
0
5
10
15
20
25
30
35
40
0 2 4 6 8 10 12 14 16 18 20 22 24
Percent
Time(Hours)
Figure4.Adolphin'smortalityriskduetopredationovera24‐hourperiod
MortalityRisk

MeganO’Toole
24
However,thisisaconservativeapproach,astheanimalswouldlikelyexceedtheir
dailyminimumrequirements.
Usingthisoptimalsituationinwhichthedolphinisablenotonlytoreplenish
itsvigilancelevelsattheendofeachdayaswellascaptureenoughpreytomeetits
metabolicneedsnowallowsforacomparisontosimulationsinwhichthedolphinis
notabletoobtainasmuchrestasisneeded.
SuboptimalSimulations
Inordertostudythepotentialeffectsofdisruptedsleepingtimesimulations
werethenrunat80%,60%,and40%ofthedolphin’soriginal5hoursofsleep.As
showninFigure6,theshortertherestingperiodmeansthedolphinsleavethebays
earlierinthedayandtoalongerforagingbout.Lessrestandmoretimesearching
forpreymeansthedolphin’svigilanceislowerattheendofeachforagingboutwith
lesstimetorecover,andwhenthedolphin’snextforagingboutbeginsitsvigilance
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0 2 4 6 8 10 12 14 16 18 20 22 24
NetEnergyGain(Kilocalories)
Time(Hours)Figure5.Thenetenergygainandlossduetothemetaboliccostofaspinnerdolphin
NetEnergyGain

MeganO’Toole
25
isnotrestoredtoitsoptimallevel.Theamountofrestlostisproportionaltothe
amountofvigilancenotrestoredattheendofeachday(seeFigure6).
Withloweredvigilancelevels,thedolphincontinuestocapturefoodduring
itsextrahoursofforagingbutatlowerratesastimewearson(Table3).
Table3.Rateofpreyintakeandtotalpreyintakeduringaforagingbout
Time(hours) PreyConsumedperHour TotalPreyIntake0 215.76 215.761 200.66 431.522 186.61 632.173 173.55 818.784 161.40 992.335 150.10 1153.736 139.59 1303.837 129.82 1443.428 120.73 1573.249 112.28 1693.98
10 104.42 1806.2611 97.11 1910.6812 90.32 13 83.99 14 78.11
Final(optimal) 2007.80Final(80%rest) 2098.11
0102030405060708090100
0 2 4 6 8 10 12 14 16 18 20 22 24
VigilanceLevel(Percent)
Time(Hours)
Figure6.Comparativevigilancelevelsatvaryinglengthsofrest
OptimumRest
80%rest
60%rest
40%rest

MeganO’Toole
26
Final(60%rest) 2182.11Final(40%rest) 2260.22
Whileintheshortterm,foragingforlongerperiodsoftimemaybebeneficialtothe
dolphinasincreasedcaloricintakecanbeusedasameasurementofoverallfitness
(Dukas&Clark1994)butifthelongerforagingboutsandshorterrestperiods
continuelongtermitmayaffecttheirhealthindifferentways,apossibilitythatwill
beanalyzedlaterinthepaper.
MortalityRiskandNetEnergyGain
Asvigilancelevelsfailtoincreasebacktotheirmaximumlevelattheendofa
restingperiod,theamountoffoodgainedisnottheonlyaspectofthedolphin’sday
thatisaffected.Mortalityrisk,afunctionofvigilancelevel,alsodemonstrates
markedlydifferentresultswithlesssleepthanfromthatfoundintheoptimal
scenario.Figure2,whichshowsthemortalityriskofthedolphinfallingbacktozero
attheendoftheday,iscomparedwiththedifferingmortalityrisksasaresultofa
lossofsleeptime.Figure7illustratesthedrasticdifferenceonthelevelofmortality
riskbasedontheloweredvigilancelevels.Previously,evenwhenthedolphinsleft
therestingbaysathour20theirriskofmortalityduetoincreasedchanceof
encounteringapredatordidnotrisesignificantlybecausetheirvigilancelevelwas
sohighitkepttheirpredationriskextremelylow.Duetoashorterrestingperiod,
thevigilancelevelisloweredandasaresulttheirmortalityriskjumpsexponentially
astheyexitthebayscausingwhatwaspreviouslyalmostnonexistentmortalityrisk
tobeashighas31%(Figure7).

MeganO’Toole
27
Intermsofenergygain,whileusingitasameasureofthedolphin’soverall
healthitwouldseemthatintheshorttermlongerforagingboutsandshorter
restingperiodswouldsuitthedolphin’sneedsofgrowthandreproductive
behaviorsasitproducesagreaternetenergygainattheendoftheday(seeFigure8)
asthemetaboliccostremainsthesameregardlessofvigilancelevel.
0.005.0010.0015.0020.0025.0030.0035.0040.0045.00
0 2 4 6 8 10 12 14 16 18 20 22 24
MortalityRisk(Percent)
Time(Hours)Figure7.Comparisonofvaryingmortalityriskbasedonrestinglengths
Optimalrest
80%rest
60%rest
40%rest
0200400600800100012001400160018002000
0 2 4 6 8 1012141618202224
NetEnergyGain(kilocalories)
Time(Hours)Figure8.Comparisonofnetenergygainwithvaryingrestinglengths
EnergyGainwithOptimalrest
EnergyGainwith80%rest
EnergyGainwith60%rest
EnergyGainwith40%rest

MeganO’Toole
28
However,whilethenetenergygainmaybehigherattheendofasingledaythe
overallhealthofthedolphinmaysufferevenwithregardstoenergygaininthelong
term.
ExtendedSimulation
Forthepurposesofthismodel,anextendedsimulationofthedatawasrun
for30dayswiththedolphinbeingabletoobtain80%ofitsidealrestingperiod.
Whenrunforonlyoneday,vigilancelevelwasrestoredto78%ofitsmaximum
level,enoughforthedolphintostillmaintainitsenergyneedsaswellaskeepits
mortalityriskstoareasonablelevel.However,afterbeingrunfor30daysthe
dolphin’svigilancelevelendsat0.74%ofitsmaximumlevelandquitefrequently
hitszeroduringforagingbouts(seeFigure8).Vigilancelevelsdroppingtozerodoes
notmeandeathandinrealityaspinnerdolphinwouldmostlikelyneverreacha
vigilancedecrementthatsevere.Thescalefromzerotoonehundredpercentis
merelyanarbitraryscaleusedtodemonstratethedecrementandrecoveryof
vigilanceratherthanaprecisemeasurement.

MeganO’Toole
29
Therateoffoodintakewasalsodrasticallyaffectedoverthelongterm.
Previously,whenrunattheoptimalrestingtime,thedolphinisabletocatch71.68
preyitemsperhour,butattheendof30dayslosingonly20%ofitsrestingtimethe
dolphinonlycatchesanaverageof15.79preyitemsperhourduetothedecreased
rateatwhichitisabletocatchpreybecauseofloweredvigilancelevels.Inthelatter
halfofthemonththedataoftenindicatesthedolphin’sabilitytocaptureonlyoneto
twopreyitemsperhourofforaging.Onthethirtiethdayofthesimulation,thetotal
preyintakeisjustunder19preyitems,afractionofthe2008preyitemscaptured
undertheoptimalscenarioorevenafterasingledayof80%rest(SeeTable4).
Table4.Rateofpreyintakeandtotalpreyintakeduringaforagingboutafter30daysat80%optimalsleep
Time(hours)PreyConsumedperHour TotalPreyIntake
0 2.02 2.011 1.88 3.892 1.74 5.633 1.62 7.26
0102030405060708090100
0 48 96144192240288336384432480528576624672720
VigilanceLevel(Percent)
Time(hours)Figure9.Simulationrunfor30days(or720hours)withthedolphinrestingat80%ofitsideallength
VigilanceLevel

MeganO’Toole
30
4 1.51 8.765 1.40 10.176 1.30 11.477 1.21 12.698 1.13 13.829 1.05 14.87
10 0.98 15.8411 0.91 16.7512 1.02 17.77
Final 18.78
MortalityRiskandNetEnergyGain
Asshowntheoptimalscenario(Figure4),thedolphinbeginseachdaywith
itslowestmortalityriskitwillhaveforthenext24‐hoursuntilitisabletorecover
vigilanceduringtherestingperiod.However,whensufficientrestisnotpossiblethe
dolphinfailstoregainmaximumvigilancebytheendofthedayandconsequentlyits
mortalityriskdoesnotfalltoitsideallevelsaroundzero.Overtime,thelowestlevel
ofmortalityriskrisesasthedolphinisnotabletoregainallofitslostvigilanceand
theamountunrecoveredgrowslargerasthemonthwearson.Figure10showsthis
gradualincreaseinoverallmortalityriskovera30dayperiodasthedolphin’s
vigilancelevelscontinuetodecrease.

MeganO’Toole
31
Intheshort‐termsimulations,itwouldseemthatthelongertheforaging
bout,thebetteroffthedolphinisintermsoffoodcaptureandenergygained.
However,ifthebehavioroflongerforagingboutsisexpandedtolookatthe
dolphin’snetenergygainovertheperiodofamonththebenefitquicklydisappears
andwhatintheshorttermwasbeneficialbecomesdetrimentaltothedolphin’s
overallhealth(Figure11).
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
1 3 5 7 9 11131517192123252729
MortalityRisk(Percent)
Time(Days)Figure10.Mortalityriskoveramonthperiodwith80%optimalsleepingtime
MortalityRisk
‐5000‐4000‐3000‐2000‐1000
010002000300040005000
1 4 7 10 13 16 19 22 25 28
Kilocalories
Time(Days)
Figure11.Totalkilocaloriesconsumedperdayandnetenergy
KilocaloriesConsumedperday
NetEnergy

MeganO’Toole
32
Ofcourse,themodeldoesnottakeintoaccountamultitudeoffactorsincludingthe
knowledgespinnerdolphinsoftenparticipateincooperativeforaging(Benoit‐Bird
etal2009)whichwouldreducetheamountofeffortexpendedperpreyitem
captured.Whilethisisanextremeprojectionandishighlyunlikelytobeaccurateit
showsageneraltrendthatlongtermlossofsleepandconsequentlyvigilancewill
severelyaffecttheabilityofthedolphintoprovidefoodforitself.
Discussion
Asdolphinsperformvariousactivitiesthroughouttheday,theirdecisionsto
participateincertainbehaviorsareduetoavarietyoffactors.Someofthebehavior
maybeattributedtofoodavailabilityorenergyconservationorevenpredationrisk,
butDukas&Clark(1995)arguethatvigilancedecrementisanotheroneofthese
importantdeterminingfactors.Theirequationssimulateascenarioinwhichmuch
ofthelifeofaspinnerdolphinisaffectedbyvigilancelevels.Whilecertainlynotthe
onlydeterminingfactor,Dukas&Clarkmayhavetouchedonanaspectofan
animal’slifethathasnotbeenexploredandispossiblyquitepertinenttothe
spinnerdolphincontroversies.
Vigilancemaybedefinedconciselyas“complexpatternrecognition”(Dukas
2002),butthescopeofitsinfluenceisclearlymuchgreaterthanitinitiallymay
appear.Fromthesesimulationsitwouldseemthatthevigilancelevelofananimal
affectsmorethanjustitsawarenessofitssurroundings;itaffectshowtheanimalis
abletocareforandprotectitselfeveryday.Theoptimalscenarioprovidesunder
somegeneralassumptionsthefactorswhichareacceptedtobeaffectedbyvigilance
levelandhowtheyrespondtoitsriseandfall.Intheidealscenario,thedolphinis

MeganO’Toole
33
abletoregainallofthevigilanceithaslostoverthecourseoftheforagingbout,keep
mortalityrisktoaminimumandconsumeenoughpreyitemstomeetitsmetabolic
needs.
Assoonastheamountofrestisdiminished,thedolphin’svigilancelevelfails
toreturntoitsmaximumlevelattheendofitsdailyrestingperiod.Initially,the
consequenceoffailuretoobtain5hoursofrestdoesnotseemofconsequence,as
evengetting40%ofitsoptimumsleepingperiodthevigilancelevelonlydropsto
aroundfiftypercent.However,itisnotthelevelofvigilancethatindicatesthe
dolphin’soverallhealthbutrathertheotherfactorswhichareafunctionofthe
vigilancelevelthatindicatethetrueeffectsofalackofrest.Therateofpreycapture
ispositivelycorrelatedwithvigilancelevel(Figure2)andsothelongertheforaging
boutthelowerthevigilancelevelmeaningthedolphinisslowertodetectand
captureprey.Onthefirstday,thelongerforagingboutwiththeshorterresting
periodisdemonstratedtobebeneficialforthedolphinintermsofenergygain.
However,themortalityriskindicatesthatevenwithminimallossofsleep,the
dolphinsaresignificantlymoreatrisktopredationthanwithafullrestingbout.As
showninFigure7,withafullrestingperiodof5hours,thevigilancelevelis
restoredtothepointthatevenafterthedolphinleavesthebaysfortheopenocean,
wherethechanceofencounteringapredatorjumpsexponentially,themortalityrisk
isstillveryclosetozero.Withlessrestvigilanceisnotrestoredto100%and
consequentlythepredationriskremainshighersothatthemortalityriskafter
losingonly20%restingtimeleadstoajumpinmortalityriskjustbyleavingthe
restingbays(seeFigure7).

MeganO’Toole
34
Whiletheshort‐termscenariosprovideinsightintohowsensitivethevarious
factorsaretothedynamicsofvigilancelevels,along‐termviewdemonstratesinto
thepotentialeffectsofhumaninterruptionofadolphin’srestingtime.Thethirty
daysimulationwasrunwiththedolphinsleeping4hoursperdayinordertogeta
betterideaoflongtermramificationsofsleepdeprivationwhileusingalow
estimateoftheamountofsleeplost.However,theresultsarestillquitedrastic.As
canbeseeninFigure9,vigilancelevelsdecreaseoverthethirtydayperioduntil
maximumvigilanceachievedattheendofarestingboutisonlyaquarterofa
percentofthemaximumpotential.Additionally,themortalityriskgrowsasthe
monthprogressesthedolphinisunabletohaveenoughrestingtimetorecoverits
vigilancelevelsandisthereforelesslikelytobeabletodetectapredatorintimeto
fendofftheattackorescapeasitsresponsetimewillbeslowed.
Inthemostextremeoutcomeoftheextendedsimulation,themodelpredicts
thataspinnerdolphinhavingsleptforfourhoursadayfor30daysinarowwill
onlybeabletocaptureapproximately19preyitemsoveratwelvehourforaging
boutonaccountofdiminishedvigilance.Whilethisisunrealistictoexpectthisina
realworldscenario,theindicationisclearthatalong‐termlossofsleepand
consequentlyvigilancewillseverelyaffecttheabilityofthedolphintoprovidefood
foritself.Additionally,Figure11showsthenetgainofenergytofallbelowzero
afteronlytheseconddayofgetting80%ofoptimalrestingtime.Resultssuchas
thismayindicatelongtermeffectsonthedolphin’shealthduetopoornutritionor
reproductivehealth.Oncenetenergygainedfallsbelowzero,itindicatesthatthe
dolphinmustutilizeenergyfromitsstoresinitsblubberinordertomeetits

MeganO’Toole
35
metabolicneeds.Dolphinblubberprovidestheanimalwithinsulationandpositive
buoyancy(Struntz,etal2004),importantcharacteristicsforalifelivedunderthe
water.Ifthedolphinisajuvenile,thiswouldalsodiminishgrowthcapacityasall
energywouldbeusedmerelyformaintenancepurposes.
Therearemultipleweaknessesinthismodelandmuchroomfor
improvement.Firstly,aminorprobleminthismodelisitsfailuretoaddressthe
vigilancedecrementwhichsurelyoccursduringthetravelandsocializationtime
whichoccursbetweenforagingandrestingbouts.Additionally,atthecurrentrateof
vigilancedecrement,eventheslightestlossofsleepleadstohugejumpsinthe
mortalityriskofthedolphinsaswellaslongtermenergygain.Theparametersmay
havetobechangedtoreflectthefactthatthedolphinsarenotstarvingtodeath
eventhoughtheyappeartobegettinglessthan4hoursofsleepoveraperiodlonger
thanonemonth.However,failuretoaddressthisinthemodeldoesnotnegatethe
potentialramificationsoflackofrest,asthegeneraltrendsarestillpertinentand
valuableinformationtothosestudyingthespinnerdolphins.Withlesssleepwill
comelessabilitytodetectpreyandoverthelongtermthedolphin’snutritionwill
sufferbutperhapsnottotheextentindicatedbythemodel.
Furthermore,thenetenergygainequationsandparameterscouldbemade
moreaccurate.Byonlycalculatingtheminimumamountoffoodcapturedandeaten
inasingledaybyaspinnerdolphin,themodelismostlikelytoosensitivetoa
loweredvigilancelevelinthatitalmostimmediatelyindicatesthatthedolphinwill
sufferfrommalnutritionfromtwoboutsofpoorsleep.However,thiswould
introduceanotherprobleminthatinsteadofhavingacalculatednumberbasedon

MeganO’Toole
36
studiesandestimatesformaintenanceneedsofthespinnerdolphinamore
arbitrarynumberwouldhavetotakeitsplace,makingthemodellessbasedonfacts
andmorebasedonassumptionsofhowmuchoveritsmaintenanceneedsadolphin
mayeat.
Ofcourse,aswithallmodels,theseresultsmustbetakenasmoreofa
guidelinethanfactthatwithvigilancedecrementcomesanexactproportional
responseinallotheraspectsofadolphin’slife.Eveniftheamountofvigilancelost
leadstoadecreaseinforagingabilitylessseverethanthedatafromthemodel
indicates,thetrendisclear:longtermvigilancedecrement,nomattertheamount,
caninfactcauseasizeabledecreaseintheabilitytodetectbothpreyandpredators
andleadtohigherinstancesofmortalityinthespinnerdolphin.Vigilancelevelis
nottheonlyfactorthataffectsdolphinhealthandmortality,butasoneofmanythis
modelcanprovideaninsightintothepotentialramificationsofnearconstant
humanpresenceintheirrestingbays.Ifotherfactorsweretakenintoaccount,the
resultsfromalackofsleepwouldnotbeasdrastic.However,themodelscreated
giveanideaofhowvigilancemayfactorinandbeapartofthedolphin’sdeclining
healthinthewild.
Thismodelcouldbetakenevenonestepfartherandperhapsattemptsome
sortofoptimalforagingsothatwhenthemortalityriskbecomestoohighforthe
dolphinanditisnotabletocaptureenoughpreyitwouldceaseforagingandstart
anotheractivity.Byaddingthistothemodel,perhapsitwouldcreateamore
realisticscenarioinwhichthedolphinwouldnotforageuntilitsvigilancelevelisso
lowthatitwouldbealmostimpossibletorecovertoitsmaximumcapacity.

MeganO’Toole
37
Conclusion
Vigilanceinthisinstancedoesnotprovideasolutiontotheseissuesnoran
additionalproblem.Itmerelyservesasanothersteppingstonetobetter
understandingpotentialfactorsinthedecreaseinspinnerdolphinhealthinthese
bays.Additionally,thedrasticdecreaseinallofthespinnerdolphinhealthmeasures
(mortalityrisk,totalpreycapture,netenergygain)mayprovideadditionalleverage
onthepartofthefederalgovernmenttoillustratethattheconstanthumanpresence
inthebaysisinfactaviolationoftheMMPA.Thedataproducedbythemodel
supportstheactiononthepartofthefederalgovernmenttohavepartialclosuresin
baysknowntohaverestingspinnerdolphins.
Additionalresearchmustbedoneinordertofullyunderstandthebiology
andlifehistoryofthespinnerdolphinnotonlytohelpmakethismodelmore
accuratebutalsobecausewithoutfurtherknowledgeitispossiblethatallactionsto
protectthedolphinscouldprovefutileandsubsequentregulationoftheareawould
bepremature.Greatercontrols,possiblythroughregulationandenforcementneed
tobeenactedinthebays.Spinnerdolphinhealthisdecliningandthenumbersof
peopleinthebaysareincreasingwithoutadequatemanagementofactivitiesor
thoughttoconsequencesofhumanpresence.Thenumberofcomplaintsrelatedto
spinnerdolphinharassmentincreaseseachyearandyetenforcementhasnot
reactedaccordingly.Formanytouroperatorsafiledcomplaintsimplyleadstoa
slaponthewrist,andthebusinessistoolucrativeforsuchasmallgovernmental
responsetoceasetheirbehavior.Vigilancemodelingprovidesyetanotherinsight
intoacomplexproblemwithagrowingneedforasolution.Dolphinconservationis

MeganO’Toole
38
oneoftheuniqueenvironmentaltopicsthatintermsoftheissue‐attentioncycle
(Downs1972),itisabletomaintainlong‐termpublicenthusiasm.Thisstatus
providestremendousopportunitytopreventthegradualdeclineofaspeciesintoan
endangermentofextinctionandshouldnotbewasted.

MeganO’Toole
39
WorksCited
Acevedo,A."InteractionsBetweenBoatsandBottlenoseDolphins."AquaticMammals17.3(1991):120‐24.Print.
Andrews,K.R.,Karczmarski,L.,Au,W.W.L.,Rickards,S.H.,Vanderlip,C.A.,Bowen,B.W.,Grau,E.G.,andToonen,R.J.Rollingstonesandstablehomes:socialstructure,habitatdiversityandpopulationgeneticsoftheHawaiianspinnerdolphin(Stenellalongirostris).MolecularEcology19:732‐748.(2010).
Print.Bejder,L.,A.Samuels,H.Whitehead,N.Gales,J.Mann,andR.Connor."Declinein
RelativeAbundanceofBottlenoseDolphinsExposedtoLong‐termDisturbance."ConservationBiology20.6(2006):1791‐798.Print.
Benoit‐Bird,KellyJ.,andWhitlowW.L.Au."CooperativePreyHerdingbythePelagicDolphin,StenellaLongirostris."TheJournaloftheAcousticalSocietyofAmerica125.1(2009):125.Print.
Benoit‐Bird,K.J."PreyCaloricValueandPredatorEnergyNeeds:ForagingPredictionsforWildSpinnerDolphins."MarineBiology145.3(2004).Print.
Benoit‐Bird,KellyJ."PreyDynamicsAffectForagingbyaPelagicPredatoroveraRangeofSpatialandTemporalScales."BehavioralEcologicalSociobiological53(2003):364‐73.Print.
Cirelli,Chiara,andGiulioTononi."IsSleepEssential?"PLoSBiology6.8(2008):1605‐1611.Print.
Delfour,Fabienne."HawaiianSpinnerDolphinsandtheGrowingDolphinWatchingActivityinOahu."JournaloftheMarineBiologicalAssociationoftheUK87.01(2007):109‐12.Print.
DepartmentofLandandNaturalResources.DepartmentofLandandNaturalResourcesSupportsSpinnerDolphinProtectionRules.WashingtonD.C.:USFedNewsService,2006.Print.
Dukas,Reuven."BehaviouralandEcologicalConsequencesofLimitedAttention."PhilosophicalTransactionsoftheRoyalSocietyB:BiologicalSciences357.1427(2002):1539‐547.Print.
Dukas,Reuven,andColinW.Clark."SustainedVigilanceandAnimalPerformance."AnimalBehavior45(1995):1259‐267.Print.
Heenehan,Heather."TheBigPicture:TheTotalEcologyApproachRecap."SAPPHIRE.DukeUniversity.Web.28Oct.2011.
Hill,MarieC.,AmandaL.Bradford,KimR.Andrews,RobinW.Baird,MarkH.Deakos,SabreD.Mahaffy,AlizaJ.Milette,JanÖstman‐Lind,ErinM.Oleson,AdamA.Pack,SusanH.Rickards,SuzanneYin,andDavidW.Johnston."AbundanceandMovementsofSpinnerDolphinsofftheMainHawaiianIslands."Tobe
published.Johnston,Dave."HawaiianSpinnerDolphins."Beaufort,NC.20Oct.2011.Lecture.Lammers,MarcO."OccurrenceandBehaviorofHawaiianSpinnerDolphins(Stenella
Longirostris)AlongOahu'sLeewardandSouthShores."AquaticMammals30.2(2004):237‐50.Print.
Marten,Ken,andSuchiPsarakos."Long‐TermSiteFidelityAndPossibleLong‐TermAssociationsOfWildSpinnerDolphins(StenellaLongirostris)SeenOffOahu,

MeganO’Toole
40
Hawaii."MarineMammalScience15.4(1999):1329‐336.Print.Norris,KennethS.,Wursig,Bernd,Wells,RandallS.,Wursig,Melany.TheHawaiianSpinnerDolphin.BerkeleyUniv.ofCalif.,1994.
Print.Norris,KennethS.,Dohl,T.P.“BehavioroftheHawaiianSpinnerDolphinStenella
longirostris”Fish.Bull.77(1980):821‐849.Print.Ocean,Joan.DOLPHINSWHALES.SwimwithDolphinsinHawaii,andwithWhales
Internationally:aMultidimensionalExperiencetoSharewithNewFriends.Web.6Dec.2011.<http://joanocean.com/>.
Orbach,MichaelJ."TheHumanEcology."DukeUniversityBeaufort,NC.5Sept.2011.Lecture.Read,Andy."SocialBehaviorofMarineMammals."DukeUniversity,Beaufort,NC.25
Oct.2011.Lecture.Ribiero,S.,F.A.Viddi,andT.R.O.Freitas."BehavioralResponsesofChilean
Dolphins."AquaticMammals31.2(2005):234‐42.Print."SpinnerDolphin."NationalMarineFisheriesServicePacificIslandsRegionalOffice.Web.04Dec.2011.<http://www.fpir.noaa.gov/PRD/prd_spinner.html>.
Samuels,A.,Bejder,L.,&Heinrich,S.“AReviewoftheLiteraturepertainingtoswimmingwithwilddolphins.”MarineMammalCommission.Bethesda,MD.(2000).
Struntz,D.J.,W.A.McLellan,R.M.Dillaman,J.E.Blum,J.R.Kucklick,andD.A.Pabst."BlubberDevelopmentinBottlenoseDolphins(TursiopsTruncatus)."Journal
ofMorphology259.1(2004):7‐20."SwimmingwithWildSpinnerDolphins."NationalMarineFisheriesServicePacific
IslandsRegionalOffice.Web.4Dec.2011.<http://www.fpir.noaa.gov/PRD/prd_swim_with_wild_dolphins.html>.
Timmel,Gregory,SarahCourbis,HollySargeant‐Green,andHalMarkowitz."EffectsofHumanTrafficontheMovementPatternsofHawaiianSpinnerDolphins(StenellaLongirostris)inKealakekuaBay,Hawaii."AquaticMammals34.4(2008):402‐11.Print.
"WhaleWatchingWorldwideTourismNumbers,ExpendituresandExpandingEconomicBenefits."InternationalFundforAnimalWelfare.Print.(2009).
Yazdi,P.,A.Kilian,andB.M.Culik."EnergyExpenditureofSwimmingBottlenoseDolphins(TursiopsTruncatus)."MarineBiology134.4(1999):601‐07.Print.