view - biblioteca - universidad de mlaga
TRANSCRIPT

Calbindin-D28k in Cortical Regionsof the Lizard Psammodromus algirus
JOSE C. DAVILA, JESUS PADIAL, MANUEL J. ANDREU, AND SALVADOR GUIRADO*Departamento de Biologıa Celular, Facultad de Ciencias,
Universidad de Malaga, 29071 Malaga, Spain
ABSTRACTThe morphology, distribution, and ultrastructural features of calbindin-D28k–immunore-
active neurons and fibers in the cortical regions of the lizard Psammodromus algirus,considered homologues to the mammalian hippocampal formation, were analyzed by usingthe peroxidase anti-peroxidase technique at the light and electron microscopic level. On thebasis of staining properties and localization, two distinct populations of calbindin-D28k–immunoreactive neurons were observed in both the medial and dorsal cortices. Those locatedin the cell layer, namely principal neurons, were weakly immunostained, whereas a number ofGolgi-like stained neurons were observed in plexiform layers. Double immunocytochemistryshowed that all calbindin immunoreactive neurons in the deep plexiform layers were alsog-aminobutyric acid immunoreactive. We consider them as a population of nonprincipalneurons different from those containing the calcium-binding proteins parvalbumin andcalretinin. Two types of immunoreactive Boutons were revealed by electron microscopy on thebasis of the synaptic specialization: Boutons making asymmetrical synapses were generallysmaller in size and contacted on small dendritic profiles or cell bodies, whereas larger boutonsestablished symmetrical synapses mainly on dendritic shafts. We propose that the first type ofboutons arises from principal neurons and that the second type arises from nonprincipal ones.Finally, the staining pattern, localization, and the circuit in which nonprincipal calbindin-immunoreactive neurons and other neurochemically defined neurons could be involved incortical regions of Psammodromus are compared with those of mammalian hippocampus. J.Comp. Neurol. 405:61–74, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: calcium-binding protein; comparative neuroanatomy; intracortical circuits;
ultrastructure; reptile
The cerebral cortex of lizards consists of four regionsarranged medial to lateral, and referred to as medial,dorsomedial, dorsal, and lateral cortices. The lateral cor-tex in reptiles has been mainly compared with mammalianpiriform cortex on the basis of similar olfactory connec-tions (Martınez-Garcıa et al., 1986, 1991; Hoogland andVermeulen-VanderZee, 1988), whereas the other corticalregions have been generally compared with mammalianhippocampal formation. Thus, reptilian medial cortex couldbe considered the homologue of the dentate gyrus inmammals on the basis of a number of characteristicsincluding position and cytoarchitecture (Ramon y Cajal,1911; Curwen, 1937; Northcutt, 1967; Lacey, 1978; Molownyand Lopez-Garcıa, 1978), connections (Lohman and vanWoerden-Verkley, 1976; Lopez-Garcıa and Martınez Gui-jarro, 1988; Martınez-Garcıa and Olucha, 1988), develop-ment (Goffinet et al., 1986; Nacher et al., 1996), andpostnatal neurogenesis (Lopez-Garcıa et al., 1988). Simi-larly, the dorsomedial cortex could be compared with partof the Ammon’s horn (particularly the CA3 region) and the
dorsal cortex with other CA regions and the subiculum (fora review of similarities between these cortical regions oflizards and the mammalian hippocampus, see Nacher etal., 1996). The comparison between specific regions of thecerebral cortex in reptiles and specific regions of thehippocampus in mammals is, however, controversial be-cause more differences than similarities exist, especiallywith regard to intratelencephalic circuits that may havebeen independently evolved in reptiles and mammals. Inthis respect, the neurochemical characterization of neu-rons is a valuable tool for comparing presumptive homolo-gous regions in different species, and, in addition, it
Grant sponsor: Spanish DGICYT; Grant number: PB93–1001; Grantsponsor: Spanish DGES; Grant number: PB96–0715.
*Correspondence to: Dr. S. Guirado, Depto. Biologıa Celular, Facultad deCiencias, Campus de Teatinos, 29071 Malaga, Spain. E-mail: [email protected]
Received 5 August 1998; Revised 11 September 1998; Accepted 24September 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 405:61–74 (1999)
r 1999 WILEY-LISS, INC.

provides insights about the role of a neuronal populationin the microcircuitry of a given central nervous system(CNS) region.
Neuropeptides and calcium-binding proteins have beenrevealed as valuable markers for different subpopulationsof g-aminobutyric acid (GABA)ergic neurons either in themammalian hippocampus (Somogyi et al., 1984; Sloviterand Nilaver, 1987; Kosaka et al., 1988; Nitsch et al., 1990;Braak et al., 1991; Gulyas et al., 1991; Seress et al., 1991,1993a; Acsady et al., 1993; Nitsch and Leranth, 1993;Pitkanen and Amaral, 1993; Ribak et al., 1993; Deller etal., 1994) (for review, see Freund and Buzsaki, 1996) or inthe reptilian cerebral cortex (Davila et al., 1991, 1993;Martınez-Guijarro et al., 1991b, 1993; Martınez-Guijarroand Freund, 1992). In this regard, previous studies havedemonstrated strong similarities in the distribution of theneuropeptides somatostatin and neuropeptide Y (NPY),the calcium-binding protein parvalbumin, and the colocal-ization of these markers with GABA in both the reptiliancortical areas and mammalian hippocampus (Davila et al.,1991, 1993, 1995; Martınez-Guijarro et al., 1991b, 1993;Martınez-Guijarro and Freund, 1992).
The calcium-binding protein calbindin-D28k (CB) is alow molecular weight protein that is widely distributed inthe CNS (Celio, 1990; Baimbridge et al., 1992; Rogers andResibois, 1992; Andressen et al., 1993). In the hippocam-pus in mammals, CB is present in granule and pyramidalcells of the dentate gyrus and CA regions, respectively, aswell as in interneurons (Celio, 1990; Gulyas et al., 1991;Toth and Freund, 1992; Seress et al., 1993b) and a subsetof GABAergic neurons that innervate the middle anddistal segments of principal cell dendrites (Gulyas et al.,1996).
In this study, we analyzed the distribution and morpho-logic features of the CB-containing neurons in the cerebralcortex of the lizard Psammodromus algirus to provide newinsights on the chemically defined cellular organization ofthe presumable reptilian hippocampus and to compare itwith the basic circuitry of mammalian hippocampus.
MATERIALS AND METHODS
Adult lizards of the species P. algirus (Lacertidae), bothsexes, were used in the present work (n 5 17; body length,65–80 mm). Throughout the experimental work, animalswere treated following the European Union guidelines ontreatment of experimental animals. Animals were kept interraria for a period no longer than 2 weeks, with food(Tenebrio larvae) and water ad libitum. Lizards weredeeply anesthetized with urethane, transcardially per-fused with 0.1 M phosphate buffered saline (PBS) pH 7.4,and subsequently with 4% paraformaldehyde, 0.1% glutar-aldehyde, and 0.2% picric acid in PBS at room temperaturefor 30 minutes. The perfused brains were removed fromthe skulls and stored at 4°C overnight in 4% paraformalde-hyde and 0.2% picric acid in PBS; then they were embed-ded in 4% agar, and 50-µm-thick frontal or parasagittalsections were obtained with a Vibratome. Before theimmunocytochemical staining, sections were washed exten-sively with PBS.
Light microscopy immunocytochemistry
Free-floating sections were first incubated in 2% normalgoat serum and 0.3% Triton X-100 in PBS at room tempera-ture for 60–90 minutes to block nonspecific binding of the
secondary antibody (the same solution was used as asolvent for the primary and link antibodies) and thentransferred to the primary antibody diluted 1:2,000 at 4°Cfor 72 hours. The anti–calbindin-D28k antiserum wasraised in rabbits by immunization with recombinant ratprotein (Swant, Basel, Switzerland; Schwaller et al., 1993).After three washes in PBS for 45 minutes, the sectionswere incubated in goat anti-rabbit IgG diluted 1:35 for 1hour, washed again in PBS for 45 minutes, and incubatedin rabbit peroxidase-antiperoxidase complex diluted 1:100for 1 hour. All steps, except for the incubation in primaryantibody, were done at room temperature with constantagitation. The immunolabeling was revealed with 0.05%diaminobenzidine and 0.03% hydrogen peroxide (H2O2) inPBS. After all cytochemical procedures, sections werewashed with PBS, mounted on gelatinized slides, air dried,dehydrated in ethanol, cleared in xylene, and cover-slippedwith Eukitt.
Double immunocytochemistry
To study the putative GABAergic nature of CB-contain-ing neurons, we carried out colocalization studies by usingpostembedding immunocytochemistry on semithin sec-tions as has been described elsewhere (Davila et al., 1993,1997). Briefly, blocks from two brains fixed with 4%paraformaldehyde, 0.2% glutaraldehyde, and 0.2% picricacid in PBS, were embedded in Araldite and sectioned at 1µm. Series of alternate 1-µm sections obtained at differentrostrocaudal levels of the cerebral cortex were immuno-stained for CB (rabbit anti-CB, Swant; diluted 1:250) orGABA (rabbit anti-GABA, Sigma, St. Louis, MO; diluted1:2,000) by using the same incubation sequence describedin the preceding section. Before incubation in the primaryantibody, resin was etched from the sections with sodiummethoxide, according to Mayor et al. (1961). Neuronsimmunoreactive for CB were outlined at a magnification of3500 with the aid of a drawing tube. The matching surfacefrom the adjacent section treated for GABA immunohisto-chemistry was aligned by using blood vessels, immunoposi-tive and immunonegative neurons, and other landmarks.GABA-immunoreactive (ir) neurons were then traced onthe same drawing and studied for colocalization.
Electron microscopy immunocytochemistry
The method was essentially identical to that of lightmicroscopy except that Triton X-100 was eliminated fromall solutions. After the diaminobenzidine reaction, the50-µm-thick sections were washed in PBS, treated with 1%OsO4 in PBS for 1 hour, dehydrated in acetone, and flatembedded in Araldite between two aluminum sheets. Toincrease contrast, sections were stained with 1% uranylacetate in 70% acetone during dehydration. After a lightmicroscopic examination, selected flat embedded sectionswere glued onto pre-polymerized resin blocks and cut at70–80 nm on a Reichert Ultra-Cut E ultramicrotome.Ultrathin sections were collected on copper grids andexamined with a Philips CM100 electron microscope.
Controls
In control experiments, the anti-CB antibody was re-placed by normal rabbit serum (1:500) or by the antibodypreadsorbed at 4°C overnight with the recombinant CBprotein (1 µg per 1 ml of the diluted antibody; Swant). Noimmunostaining could be detected under these conditionsexcept for a faint background on the surface of the sections.
62 J.C. DAVILA ET AL.

Additionally, because CB is highly homologous to calreti-nin (60% of coincidence in the primary amino acid se-quence; Rogers, 1987) and the anti-CB antibody could alsorecognize calretinin, we carried out control experiments inwhich sections were incubated in the anti-CB serumpreadsorbed at 4°C overnight with the calretinin protein (1µg per 1 ml of the diluted antibody; Swant). Controlsections were then processed as those incubated with theunadsorbed antisera. As a result, no differences in thestaining pattern of the antibody was observed, except forthat the overall staining was weaker than in normalconditions.
RESULTS
Light microscopy
General. The cerebral cortex of Psammodromus con-sists of medial, dorsomedial, dorsal, and lateral regions(Fig. 1). As stated above, the medial, dorsomedial, anddorsal cortices have been considered homologous to partsof the hippocampal formation. All these lizard corticalregions display a basic tri-laminar organization. A centrallayer where most of the cell bodies are located is referred toas the cell layer. A low density of cells, mainly intrinsicneurons, is present in two plexiform layers, superficial anddeep to the cell layer, respectively (Fig. 1). For the sake ofclarity, in the following sections we will use the terminol-ogy used in mammalian hippocampus, and thus neuronslocated in the cell layer will be referred to as principalneurons (mainly projection neurons), and those present inplexiform layers as nonprincipal or local circuit neurons.
Similar results were obtained in all brains of Psammo-dromus algirus immunostained with anti-CB. On occa-sion, however, quantitative differences were observed,especially with respect to weakly immunostained neurons
in the cell layer of medial and dorsal cortices. The immuno-reactivity of these neurons varied from one animal toanother as did the number of stained cells in thoselocations.
Neurons. A moderate number of CB-immunoreactive(CB-ir) neurons was observed throughout the rostrocaudalextent of the medial and dorsal cortices (Fig. 2). Thedorsomedial cortex was almost devoid of immunoreactivecells.
Two types of staining patterns of CB-ir neurons wereobserved. Those located in the cell layers were weaklyimmunostained although clearly distinguished from theimmunonegative neighboring cells. The immunoreactivitywas mainly restricted to the cell body and in some cases tothe proximal dendrites. In contrast, CB-ir neurons locatedin the plexiform layers (as well as a very few cells in thecell layer) were strongly immunostained in a Golgi-likemanner (Fig. 3A).
Medial cortex. CB-ir neurons in the cell layer ofmedial cortex were unevenly distributed. They were al-ways located in the dorsal aspect of this cortical area,whereas its vertical medial-most part was devoid of reac-tive neurons (Fig. 3A). They displayed a small, roundedcell body (granule cells) and fine proximal dendrites (Fig.3B). A few neurons in the inner portion of the layer wereGolgi-like immunostained (Fig. 3D). Their cell bodies,located in the border with the deep plexiform layer, werepyramidal in shape and emitted poorly branched dendritesthat were sparsely decorated with long spines.
Most Golgi-like immunostained neurons were found inthe plexiform layers. CB-ir neurons in the superficialplexiform layer possessed fusiform-shaped somata withmost dendrites oriented parallel to the pial surface (Fig.3C,F). On occasion, these dendrites spread over a greatpart of the rostrocaudal extension of the cortex. Dendritic
Fig. 1. Photomicrograph of a transverse section of the brain ofPsammodromus algirus at a mid-telencephalic level stained withhematoxylin-eosin. MC, DMC, DC, and LC: medial, dorsomedial,
dorsal, and lateral cortices, respectively; spl, cl, and dpl: superficialplexiform, cell, and deep plexiform layers, respectively. ADVR, ante-rior dorsal ventricular ridge; S, septum. Scale bar 5 150 µm.
CALBINDIN IN CORTICAL REGIONS OF PSAMMODROMUS 63

Fig. 2. A–F: Rostrocaudal distribution of calbindin-immunoreac-tive neurons in the cerebral cortex of Psammodromus. Immunoreac-tive cell bodies (black circles) were plotted on camera lucida drawingsfrom six representative rostrocaudal telencephalic levels (A, rostral; F,
caudal). The cell layer of the cortical regions is outlined. MC, DMC,DC, and LC: medial, dorsomedial, dorsal, and lateral cortices, respec-tively.
64 J.C. DAVILA ET AL.

Fig. 3. Calbindin immunoreactivity in the medial cortex. A: Photo-micrograph of the medial wall of the cerebral hemispheres showingthe medial cortex. A group of weakly immunoreactive neurons (openarrow) located in the dorsal most portion of the cell layer, near of thedorsomedial cortex, contrast with a Golgi-like stained neuron (arrow)in the deep plexiform layer. Note the superficial bundle of immunore-active fibers (arrowheads). B: Detail of a principal neuron in the celllayer. Two dendrites (arrows) arise from the apical and basal poles,and the axon (arrowhead) arises laterally from the soma. C: Photomi-crograph of the vertical portion of the medial cortex in a parasagittalsection of the telencephalon (rostral pole is to the left). A bipolarneuron in the superficial plexiform layer extends their dendrites overa great distance from the soma. The primary dendrites course parallelto the surface and branch into a few secondary dendrites at right angle
(arrows). D: A Golgi-like immunoreactive neuron in the border be-tween the cell layer and the deep plexiform layer extends its dendritesinto both plexiform layers. A very few varicose axons (arrows) areobserved in the dorsal most region of the cell layer. E: Bipolarhorizontal nonprincipal neuron in the deep plexiform layer, near theependyma. F: A sparsely spiny dendrite courses parallel to the surfacein the superficial plexiform layer of the medial cortex. The area in thesquare is detailed in G. G: Detail of the area in the square in F. Anumber of dendritic spines are observed (arrowheads). Dotted lines inthis and subsequent figures mark the limits of the cell layer. spl, cl,and dpl: superficial plexiform layer, cell layer, and deep plexiformlayer, respectively. Scale bars 5 75 µm in A,D, 20 µm in B, 50 µm inC,E,F, 5 µm in G.

spines were seen to arise from the immunoreactive den-drites (Fig. 3F,G).
Immunolabeled neurons located in the deep plexiformlayer displayed rounded or ovoid cell bodies and weremainly multipolar in morphology. A distinct preferentialorientation of dendrites was observed. Dendrites of CB-irneurons located at rostral levels were oriented parallel tothe ependyma (Fig. 3E). Some of these dendrites projectedlaterally toward the deep plexiform layer of the dorsome-dial and dorsal cortices, whereas others projected mediallytoward the ventral tip of the medial cortex. In contrast,CB-ir neurons located at more caudal levels of the cortexemitted radially oriented dendrites, some of which as-cended vertically and traversed the cell layer. Immunoreac-tive dendrites were commonly long and sparsely branched,and they possessed no dendritic spines or only a few longspines.
Dorsal cortex. Many faintly immunostained neuronswere observed in the cell layer of dorsal cortex (Fig. 4A–D).The number of CB-ir neurons at this location varied fromone animal to another. They were always found in theintermediate part of the dorsal cortex (Fig. 4A). In addi-tion, a small group of neurons containing very low levels ofimmunoreactivity was located more laterally within thecell layer of dorsal cortex. When dendrites were visualized,CB-ir neurons displayed a variety of morphologies rangingfrom pyramidal to multipolar with dendrites spreading inboth plexiform layers (Fig. 4B,C). The dendritic treeextended preferentially in a rostrocaudal direction, ascould be observed in parasagittal sections (Fig. 4C).
Besides these faintly immunoreactive neurons, a fewGolgi-like CB-ir neurons were also found in the intermedi-ate zone of the cell layer of dorsal cortex. They werevertically oriented bipolar neurons with sparsely spined,unbranched dendrites (Fig. 4E). CB-ir neurons with thesame morphology were observed as well in both plexiformlayers of dorsal cortex (Fig. 4D).
A number of CB-ir neurons were located throughout thedeep plexiform layer of dorsal cortex (Fig. 4D). The smallcell body (6.84 6 1.27 µm) was strongly immunoreactivebut dendrites were much less stained. Somata wererounded in shape and the morphology appeared to be thatof multipolar neurons.
Fibers. Distinct types of CB-ir fibers were observed.Some were extremely fine, without varicosities, otherswere fine and varicose.
The first type of fibers participated in the formation ofan immunoreactive bundle in the superficial plexiformlayer of the lizard cortical areas. This bundle extendedthrough the medial, dorsomedial, and dorsal cortices asseen in frontal sections (Figs. 3A, 4A). The bundle wascontinuous and prominent at rostral levels (Fig. 4C); atintermediate levels, it seemed to disappear from thedorsomedial cortex, whereas at caudal levels, it disap-peared in the remaining cortical areas. This bundle occu-pied the middle third of the superficial plexiform layer inthe medial cortex (Fig. 3A) and the external third of thislayer in the dorsomedial and dorsal cortices. At the lateral-most portion of the dorsal cortex, the bundle becamebroader and more immunoreactive. These fibers curvedventrally and left the cortex by crossing the so-calledsuperpositio lateralis. This immunoreactive bundle occu-pies the same position and displays the same features as abundle of fibers immunoreactive to calretinin in Psammo-dromus that was described in a previous work (Davila et
al., 1997). Because it was previously described and dis-cussed, it will not be specifically discussed in the presentstudy.
On the other hand, varicose fibers were sparsely foundin all cortical regions. At rostral telencephalic levels,varicose CB-ir fibers were predominantly observed in thecell layer of all cortical regions, whereas at intermediateand caudal levels fibers were more prominent in the deepplexiform layer of the medial and dorsal cortices. A numberof immunoreactive fibers were consistently found in thezones of the cell layer of the dorsal and medial cortices inwhich CB-ir neurons were also observed (Figs. 3D, 4E).Sometimes these fibers appeared in close contact with thecell body and proximal dendrites of immunoreactive neu-rons (Fig. 4B).
Colocalization of CB and GABA
The study of colocalization of CB and GABA revealedthat all CB-ir neurons located in the deep plexiform layerof both the medial and dorsal cortices were also GABA-ir(Fig. 5A,B). On the other hand, weakly CB-ir neuronslocated either in the cell layer of the medial cortex or in thecell layer of the dorsal cortex were GABA immunonegative.In this study, double immunocytochemistry was focused onthe putative GABAergic nature of CB-ir neurons, and wedid not try to get an estimate about the proportion ofGABAergic neurons that contain CB immunoreactivity inthe lizard cerebral cortex.
Electron microscopy
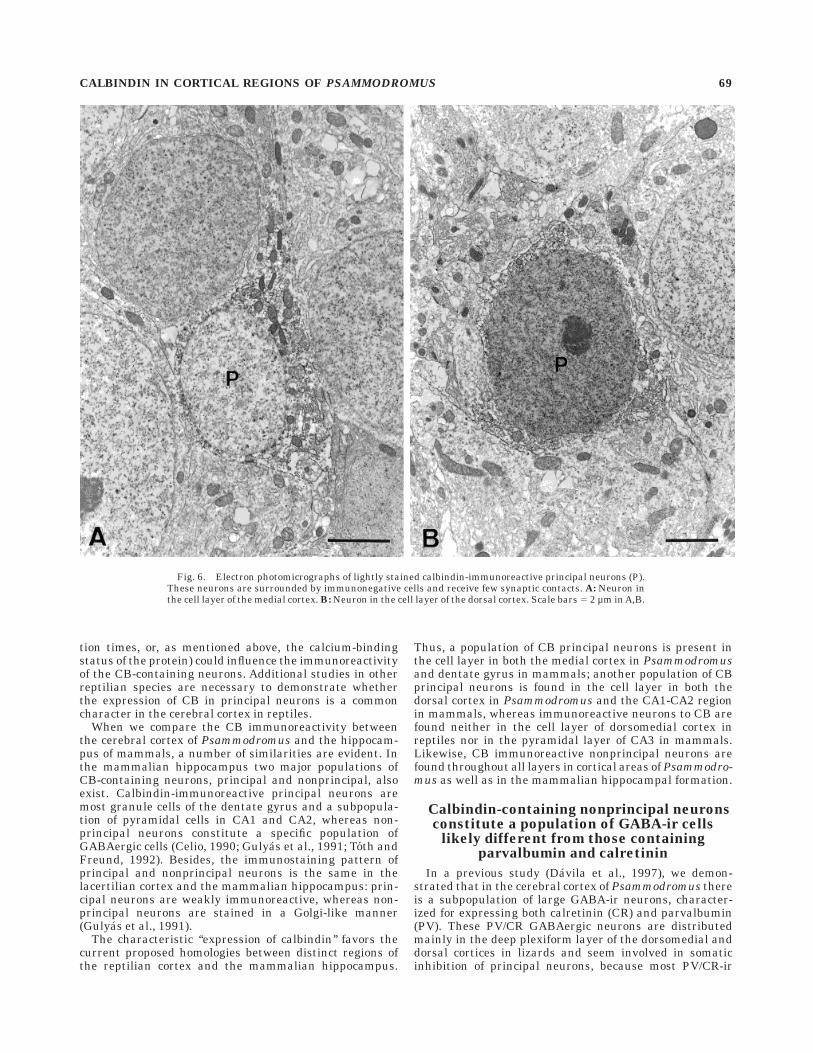
Neurons. In agreement with the light microscopicobservations, CB-ir neurons located in the cell layer showeda weak degree of immunoreactivity. They displayed simi-lar ultrastructural features as the immunonegative cellsin the layer. In the medial cortex, immunoreactive princi-pal cells displayed a small soma profile with the nucleuscommonly neither immunolabeled nor indented. The scarceperinuclear cytoplasm was filled with mitochondria, someendoplasmic reticulum and Golgi profiles, and a few densebodies (Fig. 6A). Cell bodies located within the cell layerwere tightly packed and, accordingly, CB-ir principalneurons received very few perisomatic synapses, mostlysymmetric and always immunonegative.
Immunoreactive principal neurons located in the dorsalcortex displayed a large, noninvaginated, rounded nucleus,and a rather scarce cytoplasm with the most commonorganelles (Fig. 6B). These cells received a number ofperisomatic synapses made by large immunonegative bou-tons that formed symmetric junctions.
Immunoreactivity in nonprincipal neurons was alwaysstronger than in principal ones. It is noteworthy that everyCB-ir nonprincipal neuron we observed was found to forma pair with an immunonegative cell (Fig. 7A–C). Thecontact zone between the two cells commonly included agreat extension of cell surface in which plasma membraneswere in close apposition although no membrane specializa-tion was seen (Fig. 7A). On occasion, a glial process wasfound partially intervening between the two cells (Fig.7C,E). The soma profile of immunopositive neurons wassmall. It contained a frequently immunoreactive indentednucleus eccentrically located (Fig. 7B–E) and a somewhatscarce perinuclear cytoplasm in which Golgi cisterns,abundant endoplasmic reticulum profiles, and mitochon-dria were observed. Some membrane-bounded profilesfilled with a flocculent material moderately electron dense
66 J.C. DAVILA ET AL.
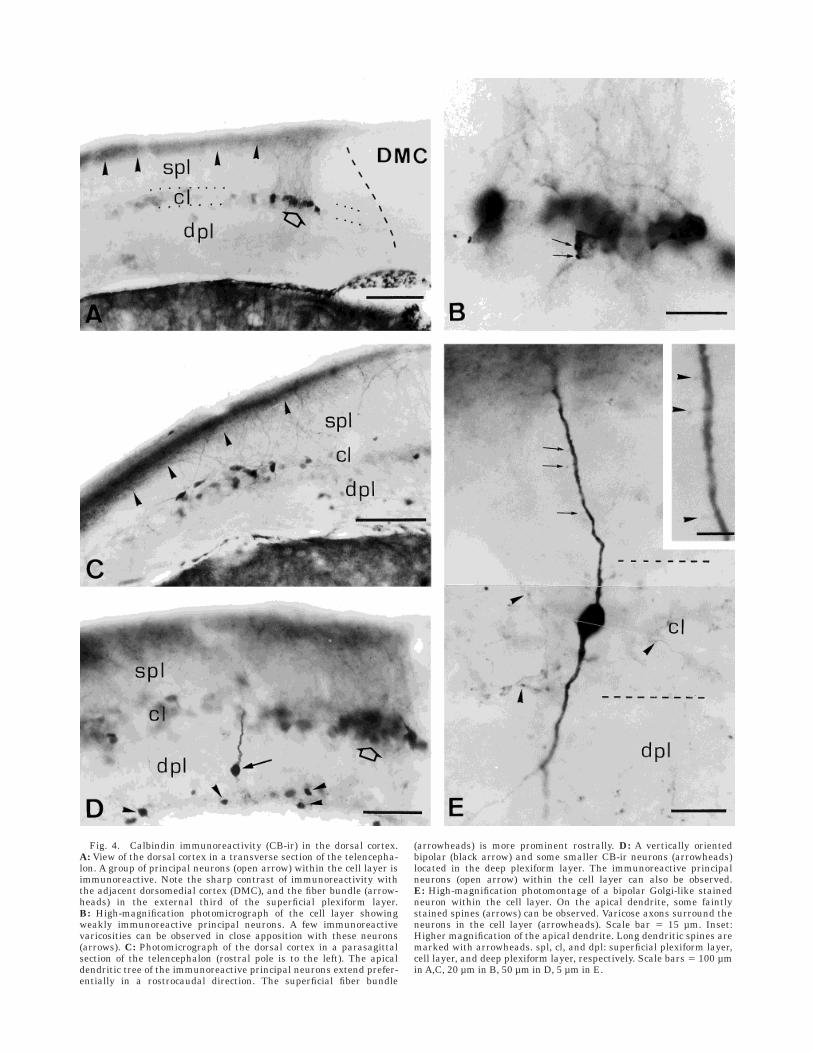
Fig. 4. Calbindin immunoreactivity (CB-ir) in the dorsal cortex.A: View of the dorsal cortex in a transverse section of the telencepha-lon. A group of principal neurons (open arrow) within the cell layer isimmunoreactive. Note the sharp contrast of immunoreactivity withthe adjacent dorsomedial cortex (DMC), and the fiber bundle (arrow-heads) in the external third of the superficial plexiform layer.B: High-magnification photomicrograph of the cell layer showingweakly immunoreactive principal neurons. A few immunoreactivevaricosities can be observed in close apposition with these neurons(arrows). C: Photomicrograph of the dorsal cortex in a parasagittalsection of the telencephalon (rostral pole is to the left). The apicaldendritic tree of the immunoreactive principal neurons extend prefer-entially in a rostrocaudal direction. The superficial fiber bundle
(arrowheads) is more prominent rostrally. D: A vertically orientedbipolar (black arrow) and some smaller CB-ir neurons (arrowheads)located in the deep plexiform layer. The immunoreactive principalneurons (open arrow) within the cell layer can also be observed.E: High-magnification photomontage of a bipolar Golgi-like stainedneuron within the cell layer. On the apical dendrite, some faintlystained spines (arrows) can be observed. Varicose axons surround theneurons in the cell layer (arrowheads). Scale bar 5 15 µm. Inset:Higher magnification of the apical dendrite. Long dendritic spines aremarked with arrowheads. spl, cl, and dpl: superficial plexiform layer,cell layer, and deep plexiform layer, respectively. Scale bars 5 100 µmin A,C, 20 µm in B, 50 µm in D, 5 µm in E.

(Fig. 7A), and multivesicular bodies were also seen. Aprimary cilium was observed arising from the basal pole ofa CB-ir nonprincipal neuron (Fig. 7D). Besides the apposi-tion of the cell membrane of the paired neuron, the cellbody of CB-ir nonprincipal neurons was partially coveredby glial processes. Few perisomatic contacts were observedon these cells. They were invariably immunonegative andboth symmetric and asymmetric synapses were observed(Fig. 8A,B). The proximal dendrites received mainly asym-metric synapses (Fig. 8C).
Axons and terminals. No myelinated immunoreac-tive axons were observed. Immunolabeled synaptic bou-tons were found in all cortical layers, but they were veryscarce in the cell layer. In the deep plexiform layer, somelarge labeled boutons were filled with clear rounded vesicles(Fig. 9B). On occasion, dense-cored vesicles were also seeninside positive boutons (Fig. 9C).
Immunoreactive terminals formed symmetric as well asasymmetric synapses. The postsynaptic element wasmainly a dendritic shaft (Fig. 9A) or a putative dendriticspine (Fig. 9C), although the cell body of both principal andnonprincipal neurons was also contacted by immunoreac-tive terminals. Despite light microscopy observations sug-
gesting that CB-ir varicose axons contact immunoreactiveneurons in the cell layer of the dorsal cortex, EM observa-tions failed to reveal these contacts. Presynaptic boutonsmaking symmetric synapses were, in general, larger thanthose making asymmetric ones, and contained more pro-files of mitochondria. Usually, symmetric contacts wereestablished on dendritic shafts (Fig. 9A), whereas asymmet-ric synapses were observed on small dendritic profiles orputative spines (Fig. 9B,C). Most immunoreactive periso-matic boutons were small in size and displayed an asym-metric synaptic specialization (Fig. 9D,E). These immuno-reactive boutons were clearly different from the typicallarge immunonegative boutons containing a number ofmitochondria and flattened vesicles and making periso-matic symmetric synapses (Fig. 9F).
DISCUSSION
Two major findings can be extracted from the presentstudy: (1) the calcium-binding protein CB is expressed inprincipal as well as nonprincipal neurons in the medialand dorsal cortices of Psammodromus; and (2) in thesecortical regions of Psammodromus, the CB-containingnonprincipal neurons constitute a population of GABA-irintrinsic cells, likely different from those containing parv-albumin and calretinin.
Before discussing the main points of this work, somecautions are to be taken into account concerning theimmunohistochemical detection of calcium-binding pro-teins. As it has been recently shown by Winsky andKuznicki (1996), the antibody recognition of calcium-binding proteins depends on their calcium-binding status.This finding might account for the quantitative differencesobserved in the number of immunostained neurons amongdifferent brains that were processed in a similar way andalso for the different degree of immunoreactivity betweenprincipal and nonprincipal neurons. In this study, how-ever, the CB immunoreactivity in the cerebral cortex ofPsammodromus was consistent from one animal to an-other, i.e., all analyzed brains presented the same patternof immunostaining, neurons weakly immunostained in thecell layers of the medial and dorsal cortices, and Golgi-likeCB-immunopositive neurons in the plexiform layers of thesame cortices.
Calbindin is present in principaland nonprincipal neurons
Two distinct populations of CB immunoreactive neuronswere observed in the cerebral cortex of Psammodromus: anumber of weakly immunoreactive principal neuronswithin the cell layer of the medial and dorsal cortices, andsome strongly immunoreactive (Golgi-like) nonprincipalneurons that can be found in all layers of these samecortices.
So far, there is only another study on the distribution ofCB in the telencephalon in a reptile, the lizard Podarcishispanica, a species closely related to Psammodromus.However, in the cerebral cortex of Podarcis, the anti-CBantibody (different from the one used in this study) labeleda population of GABAergic interneurons, but principalneurons were not immunostained (Martınez-Guijarro andFreund, 1992). This difference in the immunostainingpattern between Psammodromus and Podarcis could beattributable to an interspecific variation, although othercircumstances (specificity of the primary antibody, incuba-
Fig. 5. Alternate semithin sections immunostained for calbindin(CB) (A) or g-aminobutyric acid (GABA) (B). A neuron immunoreactivefor calbindin, located in the medial cortex near the ventricle (v), is alsoimmunoreactive for GABA (open arrows). Scale bar 5 10 µm.
68 J.C. DAVILA ET AL.
tion times, or, as mentioned above, the calcium-bindingstatus of the protein) could influence the immunoreactivityof the CB-containing neurons. Additional studies in otherreptilian species are necessary to demonstrate whetherthe expression of CB in principal neurons is a commoncharacter in the cerebral cortex in reptiles.
When we compare the CB immunoreactivity betweenthe cerebral cortex of Psammodromus and the hippocam-pus of mammals, a number of similarities are evident. Inthe mammalian hippocampus two major populations ofCB-containing neurons, principal and nonprincipal, alsoexist. Calbindin-immunoreactive principal neurons aremost granule cells of the dentate gyrus and a subpopula-tion of pyramidal cells in CA1 and CA2, whereas non-principal neurons constitute a specific population ofGABAergic cells (Celio, 1990; Gulyas et al., 1991; Toth andFreund, 1992). Besides, the immunostaining pattern ofprincipal and nonprincipal neurons is the same in thelacertilian cortex and the mammalian hippocampus: prin-cipal neurons are weakly immunoreactive, whereas non-principal neurons are stained in a Golgi-like manner(Gulyas et al., 1991).
The characteristic ‘‘expression of calbindin’’ favors thecurrent proposed homologies between distinct regions ofthe reptilian cortex and the mammalian hippocampus.
Thus, a population of CB principal neurons is present inthe cell layer in both the medial cortex in Psammodromusand dentate gyrus in mammals; another population of CBprincipal neurons is found in the cell layer in both thedorsal cortex in Psammodromus and the CA1-CA2 regionin mammals, whereas immunoreactive neurons to CB arefound neither in the cell layer of dorsomedial cortex inreptiles nor in the pyramidal layer of CA3 in mammals.Likewise, CB immunoreactive nonprincipal neurons arefound throughout all layers in cortical areas of Psammodro-mus as well as in the mammalian hippocampal formation.
Calbindin-containing nonprincipal neuronsconstitute a population of GABA-ir cells
likely different from those containingparvalbumin and calretinin
In a previous study (Davila et al., 1997), we demon-strated that in the cerebral cortex of Psammodromus thereis a subpopulation of large GABA-ir neurons, character-ized for expressing both calretinin (CR) and parvalbumin(PV). These PV/CR GABAergic neurons are distributedmainly in the deep plexiform layer of the dorsomedial anddorsal cortices in lizards and seem involved in somaticinhibition of principal neurons, because most PV/CR-ir
Fig. 6. Electron photomicrographs of lightly stained calbindin-immunoreactive principal neurons (P).These neurons are surrounded by immunonegative cells and receive few synaptic contacts. A: Neuron inthe cell layer of the medial cortex. B: Neuron in the cell layer of the dorsal cortex. Scale bars 5 2 µm in A,B.
CALBINDIN IN CORTICAL REGIONS OF PSAMMODROMUS 69

Fig. 7. Electron photomicrographs of immunoreactive nonprinci-pal neurons. A: A typical pair of an immunopositive and an immu-nonegative neuron in the deep plexiform layer of the medial cortex.Both neurons display a similar elongated profile and have a largesurface of cell membrane in close apposition. The immunoreactiveneuron possesses a nucleus with invaginations (arrows) and a cyto-plasm with cisterns of rough endoplasmic reticulum (RER), mitochon-dria, and two moderately dense bodies (arrowheads). B: A fusiformhorizontal immunoreactive neuron in the deep plexiform layer of themedial cortex forming a pair with an immunonegative cell (asterisk).
In this case, the apposition surface between both neurons is small(arrow). C: Pair of neurons in the deep plexiform layer of the dorsalcortex, near the ependyma. The area in the square is enlarged in E.D: Detail of a cilium (arrowheads) arising from an immunoreactiveneuron. An invaginated nucleus (large arrow), abundant RER, mito-chondria, and a moderately dense body (small arrow) can be observed.E: Detail of the area in the square in C. A glial process (asterisks) isobserved between the two paired neurons. The arrow labels thenuclear invagination. Scale bars 5 2 µm in A,D, 5 µm in B,C, 1 µm in E.

terminals establish mainly juxtasomatic synapses withinthe cell layer (Martınez-Guijarro et al., 1991b; Davila etal., 1993, 1997). A variety of data make it very unlikelythat CB is colocalized in the same population of PV- andCR-containing neurons in the cerebral cortex of Psammo-dromus: (1) CB-containing neurons are located in themedial and dorsal cortices but not in the dorsomedialcortex, whereas most PV/CR-containing neurons in Psam-modromus are located in the dorsomedial and dorsalcortex but not in the medial cortex; (2) CB-ir neurons aresmall or medium in size, whereas PV/CR-ir neurons dis-play a large cell body; and (3) the synaptic targets of bothgroups of neurons are quite different. This contrasts withthe results obtained in another lizard of the same family,Podarcis hispanica, by Martınez-Guijarro and Freund(1992), according to which all three calcium-binding pro-teins are expressed by the same population of GABAergicneurons in both the dorsomedial and dorsal cortices. Dueto the high degree of homology between CB and calretinin,the possibility exists that the anti-CB antibody used bythese authors cross-reacted with the calretinin-containingneurons, although interspecific variation cannot be dis-carded.
On the other hand, GABAergic neurons containing theneuropeptides somatostatin and NPY have been describedin both the cerebral cortex in reptiles (Davila et al., 1991,
1993; Martınez-Guijarro et al., 1993) and the hippocampusin mammals (Somogyi et al., 1984; Kohler et al., 1986,1987; Sloviter and Nilaver, 1987; Kosaka et al., 1988;Leranth et al., 1990). In lizards, GABA/neuropeptide-containing neurons are mainly found in the dorsomedialand dorsal cortices, and their postsynaptic targets are thedistal dendrites of principal neurons. The distribution ofneuropeptide-containing neurons partially overlaps withthe distribution of CB-ir neurons (both are found in thedorsal cortex), but the postsynaptic target of the formerpopulation is restricted to the distal dendrites; therefore, itis different from that of CB-ir neurons. Thus, most prob-ably both groups of neurons are nonoverlapped, as occurswith neuropeptide-containing neurons and the PV/CRneurons in Psammodromus.
In mammalian hippocampus, CB-ir nonprincipal neu-rons constitute a distinct population of GABAergic localcircuit neurons, which is mostly nonoverlapped with PV-,CR-, or with somatostatin- and NPY-containing neurons(Kohler et al., 1986; Sloviter and Nilaver, 1987; Sloviter,1989; Nitsch et al., 1990; Braak et al., 1991; Gulyas et al.,1991; Seress et al., 1991, 1993a; Sloviter et al., 1991;Miettinen et al., 1992; Rogers and Resibois, 1992; Acsadyet al., 1993; Nitsch and Leranth, 1993; Pitkanen andAmaral, 1993; Ribak et al., 1993).
Fig. 8. Synaptic contacts on nonprincipal neurons. A: Two negativeboutons (b) contacts on an immunoreactive soma (Som). One of themestablishes a symmetrical synapse (arrowheads). B: Perisomatic
asymmetrical synapse (arrows). C: A fine immunoreactive dendriticprofile (Den) receives an asymmetrical synapse (arrow). Scale bars 50.5 µm in A, 0.25 µm in B,; 0.2 µm in C.
CALBINDIN IN CORTICAL REGIONS OF PSAMMODROMUS 71

Fig. 9. Immunoreactive boutons. A: Two large boutons, one immu-noreactive (b) and the other immunonegative (b’), make symmetricalsynapses (arrowheads) on the same dendritic shaft (Den). Opposite,two other asymmetrical synapses (arrows). B: A large immunoreactivebouton plenty of synaptic vesicles contacting a small dendritic profile.The synaptic specialization is asymmetrical (arrows). C: An immuno-reactive bouton (b) with dense-cored vesicles makes an asymmetricalsynapse (arrows) on a putative dendritic spine. D: Low-magnification
electron photomicrograph of the cell layer of the dorsal cortex. Aprincipal neuron (P) receives a perisomatic contact with an immunore-active bouton. The area in the square is detailed in E. E: Detail of thesquared area in D. A scarcely prominent asymmetrical postsynapticdensification can be observed (arrows). F: A typical (immunonegative)perisomatic symmetrical synapse is shown for comparison. Scalebar 5 0.5 µm in A,F, 0.4 µm in B, 0.25 µm in C,E, 2 µm in D.

As a conclusion, our data on the distribution of CB in thecerebral cortex of Psammodromus reveal some additionalsimilarities between the hippocampus in mammals andthe cerebral cortex of lizards. In both structures CB isexpressed in neurons of the cell layer (putative projectingcells) and in cells outside the cell layer (probably intrinsiccells). Whereas most CB-expressing cells in the cellularlayer are weakly reactive, CB-ir neurons in the plexiformlayers of both the mammalian hippocampal formation andthe reptilian cerebral cortex display strong immunoreactiv-ity that reveals their whole dendritic morphology. Thesecells constitute a subpopulation of GABAergic neuronsthat probably does not overlap with the ones expressingneuropeptides, and PV/CR.
Functional considerations
Electron microscopy reveals two main types of synapsesmade by immunoreactive boutons throughout the lizardcortex, one of asymmetric type (presumably excitatory)and another symmetric (presumably inhibitory). Althoughwe were unable to trace the cells of origin of these synapticboutons, data from light microscopy suggest that each typeoriginates from a different population of cortical CB immu-noreactive neurons. Principal neurons in the medial cortexhave been shown to be glutamatergic (Martınez-Guijarroet al., 1991a) and give rise to the Timm-positive projectionto the dorsomedial and dorsal cortices (Lopez-Garcıa andMartınez-Guijarro, 1988; Martınez-Garcıa and Olucha,1988; Olucha et al., 1988). In addition, the dorsal-mostregion of the medial cortex projects specifically to theintermediate dorsal cortex (Martınez-Garcıa and Olucha,1988; Olucha et al., 1988; unpublished data in Psammodro-mus). Thus, CB-immunoreactive principal cells in themedial cortex could be the main source of the boutonsmaking both axosomatic and axodendritic asymmetricalsynapses on the neurons of the dorsal cortex.
On the other hand, the CB immunoreactive principalcells of the dorsal cortex, probably excitatory, could be theorigin of the boutons making asymmetrical synapses onthe neurons of the medial cortex, because it has beenshown that dorsal cortex projects to the medial cortex(Martınez-Garcıa and Olucha, 1988). Finally, we cannotdiscard a possible extrinsic origin for part of CB immunore-active boutons making asymmetrical synapses.
The most likely origin for the symmetric junctionsobserved in the medial and dorsal cortices of Psammodro-mus are the nonprincipal CB/GABA-ir neurons located inthese same regions. Symmetrical synapses were observedmainly on dendritic shafts; therefore, one may speculatethat inhibition could be performed on the dendritic do-mains of principal neurons.
The data available led us to suggest a possible role forthe different populations of GABAergic neurons in thecircuitry of the putative hippocampus in lizards. So far,three nonoverlapped populations of nonprincipal neuronshave been reported in reptilian cerebral cortex, accordingto the presence of neuropeptides or calcium-binding pro-teins: (1) GABAergic neurons with somatostatin and NPY(Davila et al., 1991, 1993; Martınez-Guijarro et al., 1993);(2) GABAergic neurons with parvalbumin and calretinin(Martınez-Guijarro and Freund, 1992; Davila et al., 1997);and (3) GABAergic neurons with CB (this study). GABA/neuropeptide-containing neurons are distributed mainlyin the dorsomedial and dorsal regions and their synaptictarget are the distal dendritic domains of principal cells
(Davila et al., 1988, 1991, 1993; Martınez-Guijarro et al.,1993). On the other hand, GABA/PV/CR-containing neu-rons are located in the dorsomedial and dorsal cortices andmediate the perisomatic inhibition of principal neurons inboth cortical areas (Davila et al., 1997). Finally, GABA/CB-containing neurons could inhibit the dendrites of principalneurons in the medial and dorsal cortices.
If we compare the populations of nonprincipal neuronsand their possible role in the cerebral cortex in lizards withthe hippocampal circuitry in mammals, strong similaritiesemerge. In the mammalian hippocampus, various types ofnonprincipal neurons have been identified on the basis oftheir neurochemical marker content, which correlateswith a selective innervation of different membrane do-mains of principal neurons (for a review, see Freund andBuzsaki, 1996). GABA/PV neurons innervate the cellbodies and axon initial segments of principal cells (Kat-sumaru et al., 1988; Nitsch et al., 1990), whereas GABA/somatostatin (Leranth et al., 1990) and GABA/CB (Sik etal., 1995; Gulyas et al., 1996) neurons are specialized toinnervate different dendritic domains of principal cells.
As a conclusion, if we look at specific cell types character-ized by the expression of neurotransmitters, neurotransmit-ter-related markers, and calcium-binding proteins, theirmorphologic and neurochemical features, and to the orga-nization of the intrinsic GABAergic system, the number ofsimilarities between the hippocampus in mammals andthe medial, dorsomedial, and dorsal cortices in reptilesallows us to suggest with a high degree of confidence somecommon basic organizational patterns in this region.
ACKNOWLEDGMENTS
The authors thank Fernando Martınez-Garcıa for provid-ing many valuable comments and suggestions on how toimprove the manuscript.
LITERATURE CITED
Acsady L, Halasy K, Freund TF. 1993. Calretinin is present in non-pyramidal cells of the rat hippocampus: III. Their inputs from themedian raphe and medial septal nuclei. Neuroscience 52:829–841.
Andressen C, Blumcke I, Celio MR. 1993. Calcium-binding proteins:selective markers of nerve cells. Cell Tissue Res 271:181–208.
Baimbridge KG, Celio MR, Rogers JH. 1992. Calcium-binding proteins inthe nervous system. Trends Neurosci 15:303–308.
Braak E, Strotkamp B, Braak H. 1991. Parvalbumin-immunoreactivestructures in the hippocampus of the human adult. Cell Tissue Res264:33–48.
Celio MR. 1990. Calbindin D-28k and parvalbumin in the rat nervoussystem. Neuroscience 35:375–475.
Curwen AO. 1937. The telencephalon of Tupinambis nigropunctatus: I.Medial and cortical areas. J Comp Neurol 66:375–404.
Davila JC, Guirado S, de la Calle A. 1988. Immunocytochemical localizationof somatostatin in cerebral cortex of lizards. Brain Res 447:52–59.
Davila JC, de la Calle A, Gutierrez A, Megıas M, Andreu MJ, Guirado S.1991. Distribution of neuropeptide Y (NPY) in the cerebral cortex of thelizards Psammodromus algirus and Podarcis hispanica: Co-localizationof NPY, somatostatin and GABA. J Comp Neurol 308:387–408.
Davila JC, Megıas M, de la Calle A, Guirado S. 1993. Subpopulations ofGABA neurons containing somatostatin, neuropeptide Y, and parvalbu-min in the dorsomedial cortex of the lizard Psammodromus algirus. JComp Neurol 336:161–173.
Davila JC, Megıas M, Andreu MJ, Real MA, Guirado S. 1995. NADPHDiaphorase-positive neurons in the lizard hippocampus: a distinctsubpopulation of GABAergic interneurons. Hippocampus 5:60–70.
Davila JC, Padial J, Andreu MJ, Real MA, Guirado S. 1997. Calretininimmunoreactivity in the cerebral cortex of the lizard Psammodromus
CALBINDIN IN CORTICAL REGIONS OF PSAMMODROMUS 73

algirus: a light and electron microscopic study. J Comp Neurol 382:382–393.
Deller T, Nitsch R, Frotscher M. 1994. Associational and commissuralafferents of parvalbumin-immunoreactive neurons in the rat hippocam-pus: a combined immunocytochemical and PHA-L study. J Comp Neurol350:612–622.
Freund TF, Buzsaki G. 1996. Interneurons of the hippocampus. Hippocam-pus 6:347–470.
Goffinet AM, Daumerie C, Langerwerf B, Pieau C. 1986. Neurogenesis inreptilian cortical structures: 3H-thymidine autoradiographic analysis. JComp Neurol 243:106–116.
Gulyas AI, Toth K, Danos P, Freund TF. 1991. Subpopulations ofGABAergic neurons containing parvalbumin, calbindin D28K, andcholecystokinin in the rat hippocampus. J Comp Neurol 312:371–378.
Gulyas AI, Hajos N, Freund TF. 1996. Interneurons containing calretininare specialized to control other interneurons in the rat hippocampus. JNeurosci 16:3397–3411.
Hoogland PV, Vermeulen-VanderZee E. 1988. Intrinsic and extrinsic connec-tions of the cerebral cortex of lizards. In: Schwerdtfeger WK, SmeetsWJAJ, editors. The forebrain of reptiles. Current concepts of structureand function. Basel: Karger. p 20–29.
Katsumaru H, Kosaka T, Heizmann CW, Hama K. 1988. Immunocytochemi-cal study of GABAergic neurons containing the calcium-binding proteinparvalbumin in the rat hippocampus. Exp Brain Res 72:347–362.
Kohler C, Eriksson L, Davies S, Chan-Palay V. 1986. Neuropeptide Yinnervation of the hippocampal region in the rat and monkey brain. JComp Neurol 244:384–400.
Kohler C, Eriksson LG, Davis S, Chan-Palay V. 1987. Co-localization ofneuropeptide tyrosin and somatostatin immunoreactivity in neurons orindividual subfields of the rat hippocampal region. Neurosci Lett78:1–6.
Kosaka T, Wu JY, Benoit R. 1988. GABAergic neurons containing somato-statin-like immunoreactivity in the rat hippocampus and dentategyrus. Exp Brain Res 71:388–398.
Lacey DJ. 1978. The organization of the hippocampus of the fence lizard: alight microscopic study. J Comp Neurol 182:247–264.
Leranth C, Malcolm AJ, Frotscher M. 1990. Afferent and efferent synapticconnections of somatostatin-immunoreactive neurons in the rat fasciadentata. J Comp Neurol 295:111–191.
Lohman AHM, van Woerden-Verkley I. 1976. Further studies on thecortical connections of the tegu lizard. Brain Res 103:9–28.
Lopez-Garcıa C, Martınez-Guijarro FJ. 1988. Neurons in the medial cortexgive rise to Timm-positive boutons in the cerebral cortex of lizards.Brain Res 463:205–217.
Lopez-Garcıa C, Molowny A, Garcıa-Verdugo JM, Ferrer I. 1988. Delayedpostnatal neurogenesis in the cerebral cortex of lizards. Dev Brain Res43:167–174.
Martınez-Garcıa F, Olucha FE. 1988. Afferent projections to the Timm-positive cortical areas of the telencephalon of lizards. In: SchwerdtfegerWK, Smeets WJAJ, editors. The forebrain of reptiles. Current conceptsof structure and function. Frankfurt, Basel: Karger. p 30–40.
Martınez-Garcıa F, Amiguet M, Olucha FE, Lopez-Garcıa C. 1986. Connec-tions of the dorsal cortex in the lizard Podarcis hispanica. Neurosci Lett63:39–44.
Martınez-Garcıa F, Olucha FE, Teruel V, Lorente MJ, Schwerdtfeger WK.1991. Afferent and efferent connections of the olfactory bulbs in thelizard Podarcis hispanica. J Comp Neurol 305:337–347.
Martınez-Guijarro FJ, Freund TF. 1992. Distribution of GABAergic inter-neurons immunoreactive for calretinin, calbindin 28K, and parvalbu-min in the cerebral cortex of the lizard Podarcis hispanica. J CompNeurol 322:449–460.
Martınez-Guijarro FJ, Soriano E, Del Rıo JA, Lopez-Garcia C. 1991a.Zinc-positive boutons in the cerebral cortex of lizards show glutamateimmunoreactivity. J Neurocytol 20:834–843.
Martınez-Guijarro FJ, Soriano E, del Rıo JA, Lopez-Garcıa C. 1991b.Parvalbumin-immunoreactive neurons in the cerebral cortex of thelizard Podarcis hispanica. Brain Res 547:339–343.
Martınez-Guijarro FJ, Soriano E, del Rıo JA, Blasco-Ibanez JM, Lopez-Garcıa C. 1993. Parvalbumin-containing neurons in the cerebral cortexof the lizard Podarcis hispanica: morphology, ultrastructure, andcoexistence with GABA, somatostatin, and neuropeptide Y. J CompNeurol 336:447–467.
Mayor HD, Hampton JC, Rosario B. 1961. A simple method for removingthe resin from epoxi embedded tissue. J Cell Biol 9:909–910.
Miettinen R, Gulyas AI, Baimbridge KG, Jacobowitz DM, Freund TF. 1992.Calretinin is present in non-pyramidal cells of the rat hippocampus: II.
Co-existence with other calcium-binding proteins and GABA. Neurosci-ence 48:29–43.
Molowny A, Lopez-Garcıa C. 1978 Estudio citoarquitectonico de la cortezacerebral de reptiles: III. Localizacion histoquımica de metales pesados ydefinicion de subregiones Timm-positivas en la corteza de Lacerta,Chalcides, Tarentola y Malpolon. Trab Inst Cajal Inv Biol 70:55–74.
Nacher J, Ramirez C, Molowny A, Lopez-Garcia C. 1996. Ontogeny ofsomatostatin immunoreactive neurons in the medial cerebral cortexand other cortical areas of the lizard Podarcis hispanica. J Comp Neurol374:118–135.
Nitsch R, Leranth C. 1993. Calretinin immunoreactivity in the monkeyhippocampal formation. II. Intrinsic GABAergic and hypothalamicnon-GABAergic systems: an experimental tracing and co-existencestudy. Neuroscience 55:797–812.
Nitsch R, Soriano E, Frotscher M. 1990. The parvalbumin-containingnonpyramidal neurons in the rat hippocampus. Anat Embryol 181:413–425.
Northcutt RG. 1967. Architectonic studies of the telencephalon of Iguanaiguana. J Comp Neurol 130:109–148.
Olucha F, Martınez-Garcıa F, Poch L, Schwerdtfeger WK, Lopez-Garcıa C.1988. Projections from the medial cortex in the brain of lizards:correlation of anterograde and retrograde transport of horseradishperoxidase with Timm staining. J Comp Neurol 276:469–480.
Pitkanen A, Amaral DG. 1993. Distribution of parvalbumin-immunoreac-tive cells and fibers in the monkey temporal lobe: the hippocampalformation. J Comp Neurol 331:37–74.
Ramon y Cajal S. 1911. Histologie du Systeme Nerveux de l’Homme et desVertebres. Paris: A. Maloine.
Ribak, CE, Seress L, Leranth C. 1993. Electron microscopic immunocyto-chemical study of the distribution of parvalbumin-containing neuronsand axon terminals in the primate dentate gyrus and Ammon’s horn. JComp Neurol 327:298–321.
Rogers JH. 1987. Calretinin: a gene for a novel calcium binding proteinexpressed principally in neurons. J Cell Biol 105:1343–1353.
Rogers JH, Resibois A. 1992. Calretinin and calbindin-D28k in rat brain:patterns of partial co-localization. Neuroscience 51:843–865.
Schwaller B, Buchwald P, Blumcke I, Celio MR, Hunziker W. 1993.Characterization of a polyclonal antiserum against the purified humanrecombinant calcium binding protein calretinin. Cell Calcium 14:639–648.
Seress L, Gulyas AI, Freund TF. 1991. Parvalbumin- and calbindin D28K-immunoreactive neurons in the hippocampal formation of the macaque.J Comp Neurol 313:162–177.
Seress L, Nitsch R, Leranth C. 1993a. Calretinin immunoreactivity in themonkey hippocampal formation: 1. Light and electron microscopiccharacteristics and colocalization with other calcium-binding proteins.Neuroscience 55:775–796.
Seress L, Gulyas AI, Ferrer I, Tunon T, Soriano E, Freund TF. 1993b.Distribution, morphological features, and synaptic connections of parv-albumin- and calbindin D28k-immunoreactive neurons in the humanhippocampal formation. J Comp Neurol 337:208–230.
Sik A, Pentonnen M, Ylinen A, Buzsaki G. 1995. Hippocampal CA1interneurons: an in vivo intracellular labeling study. J Neurosci 15:6651–6665.
Sloviter RS. 1989. Calcium-binding protein (calbindin-D28K) and parvalbu-min immunocytochemistry: localization in the rat hippocampus withspecific reference to the selective vulnerability of hippocampal neuronsto seizure activity. J Comp Neurol 280:183–196.
Sloviter RS, Nilaver G. 1987. Immunocytochemical localization of GABA-,cholecystokinin-, vasoactive intestinal polypeptide-, and somatostatin-like immunoreactivity in the area dentada and hippocampus of the rat.J Comp Neurol 256:42–60.
Sloviter RS, Sollas AN, Barbaro NM, Laxer KD. 1991. Calcium-bindingprotein (calbindin-D28K) and parvalbumin immunocytochemistry inthe normal and epileptic human hippocampus. J Comp Neurol 308:381–396.
Somogyi P, Hodgson AJ, Smith AD, Nunzi MG, Gorio A, Wu J-Y. 1984.Different populations of GABAergic neurons in the visual cortex andhippocampus of cat contain somatostatin- or cholecystokinin-immunore-active material. J Neurosci 4:2590–2603.
Toth K, Freund TF. 1992. Calbindin D 28K-containing nonpyramidal cellsin the rat hippocampus: their immunoreactivity for GABA and projec-tion to the medial septum. Neuroscience 49:793–805.
Winsky L, Kuznicki J. 1996. Antibody recognition of calcium-bindingproteins depends on their calcium-binding status. J Neurochem 66:764–771.
74 J.C. DAVILA ET AL.