vascular endothelial growth factor-a regulation of … › images › article › pdf ›...
TRANSCRIPT

Review
Page 1 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f Int
eres
ts: n
one
decl
ared
. A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t. A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease
GW Fearnley1, GA Smith1, MA Harrison2, SB Wheatcroft3,DC Tomlinson4, S Ponnambalam1*
AbstractIntroductionVascular endothelial growth factors (VEGFs) comprise of a multi-gene-encoded family of cytokines that display remarkable effects in different tissues including the vascular, epithelial and neuronal tissues. Such factors bind to both membrane-bound and soluble receptor isoforms that regulate the ability of these cytokines to trigger profound biochemical changes within cells, resulting in changes to metab-olism, gene expression, cell prolif-eration, apoptosis and homeostasis. The cornerstone of this biochemical control is based on the interaction between soluble factors such as VEGFs and membrane-bound recep-tors such as VEGF receptor tyrosine kinases (VEGFRs). The endothelial monolayer that lines all blood vessels responds to VEGFs to regulate many aspects of vascular physiology. Dysfunction in the VEGF-A signal-ling pathways is implicated in a wide variety of diseases ranging from solid tumour metastasis to atheroscle-rosis. VEGF binding to membrane VEGFRs triggers intracellular signal-ling, post-translational protein modi-
fications, trafficking and proteolysis. Temporal and spatial co-ordination of such events is critical for program-ming downstream responses by the endothelium. In this review, we have discussed the biochemical mecha-nisms that underlie new blood vessel sprouting mediated by VEGF-like cytokines. ConclusionThe development of primary cell systems, multicellular models, organ culture and animal models, will dramatically increase our ability to understand the biochemical basis for how this important class of cytokines regulate animal physiology. Targeting such pathways directly or indirectly using humanised antibodies or small molecule inhibitors, is important for treating a wide variety of patho-logical states including solid tumour growth and metastasis, age-related macular degeneration and condi-tions involving blood vessel repair and regeneration.
IntroductionThe discovery of vascular endothe-lial growth factor A (VEGF-A) more than three decades ago trig-gered an intense interest in the biochemical mechanisms under-lying the sprouting of new blood vessels i.e. ‘angiogenesis’. This is a complex process whereby new blood vessels sprout from a pre-existing vascular network1,2, which occurs when extrinsic levels of pro-angiogenic factors exceed those of the anti-angiogenic factors. This is a biochemical switch that regulates the quiescent vasculature to proliferate and sprout new blood vessels1. Physi-ological regulation of angiogenesis is crucial as dysfunction in angiogenic
responses can cause serious patho-logical conditions such as diabetic retinopathy, atherosclerotic plaque rupture, solid tumour growth and metastasis2. This review discusses VEGF-A regulation of blood vessels sprouting in health and disease.
DiscussionThe authors have referenced som e of their own studies in this review. The protocols of these studies have been approved by the relevant ethics committees related to the institutions in which they were performed.
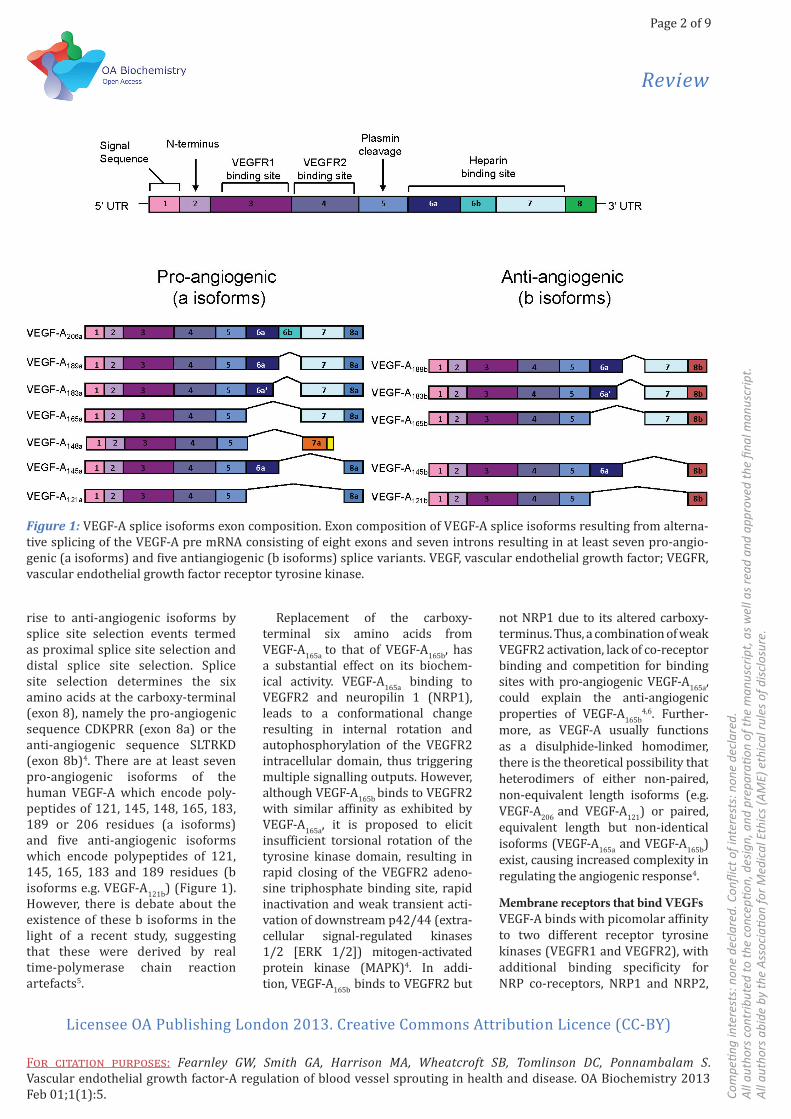
VEGFsThe human VEGF family consists of five members: VEGF-A, VEGF-B, VEGF-C, VEGF-D and placental growth factor (PlGF), which regulate angiogenesis, vasculogenesis and/or lymphangiogenesis1. The VEGF-A gene is located on the human chro-mosome 6p21.33; the encoded pre-mRNA is approximately 14 kB in length, containing eight exons and seven introns2. Alternative splicing of this pre-mRNA transcript produces multiple VEGFA isoforms (Figure 1). Each isoform contains exons 1–5 which encode the signal sequence (exon 1), N-terminus (exon 2), dimerisation domain, VEGF receptor tyrosine kinase 1 (VEGFR1)binding and N-glycosylation site (exon 3), and VEGFR2-binding site (exon 4) and a plasmin cleavage site (exon 5). The identity of each VEGF-A isoform is largely determined by variable inclu-sion of exons 6a, 6b, 7a and 7b, which encode the heparin-binding domain (Figure 1). In addition to generating VEGF-A isoforms that differ in length and domain composition, the pre-mRNA splicing machinery also gives
* Corresponding authorEmail: [email protected] Endothelial Cell Biology Unit, School of Mo-
lecular & Cellular Biology, LIGHT Laborato-ries, University of Leeds, Leeds LS2 9JT, UK
2 School of Biomedical Sciences, LIGHT Labora-tories, University of Leeds, Leeds LS2 9JT, UK
3 Division of Cardiovascular and Diabetes Re-search, LIGHT Laboratories, University of Leeds, Leeds LS2 9JT, UK
4 School of Molecular & Cellular Biology & Biomedical Health Research Centre, Astbury Building, University of Leeds, Leeds LS2 9JT, UK
Sig
nal Tr
ansd
uct
ion
Review
Page 2 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f int
eres
ts: n
one
decl
ared
.A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t.A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
rise to anti-angiogenic isoforms by splice site selection events termed as proximal splice site selection and distal splice site selection. Splice site selection determines the six amino acids at the carboxy-terminal (exon 8), namely the pro-angiogenic sequence CDKPRR (exon 8a) or the anti-angiogenic sequence SLTRKD (exon 8b)4. There are at least seven pro-angiogenic isoforms of the human VEGF-A which encode poly-peptides of 121, 145, 148, 165, 183, 189 or 206 residues (a isoforms) and five anti-angiogenic isoforms which encode polypeptides of 121, 145, 165, 183 and 189 residues (b isoforms e.g. VEGF-A121b) (Figure 1). However, there is debate about the existence of these b isoforms in the light of a recent study, suggesting that these were derived by real time-polymerase chain reaction artefacts5.
Replacement of the carboxy-terminal six amino acids from VEGF-A165a to that of VEGF-A165b, has a substantial effect on its biochem-ical activity. VEGF-A165a binding to VEGFR2 and neuropilin 1 (NRP1), leads to a conformational change resulting in internal rotation and autophosphorylation of the VEGFR2 intracellular domain, thus triggering multiple signalling outputs. However, although VEGF-A165b binds to VEGFR2 with similar affinity as exhibited by VEGF-A165a, it is proposed to elicit insufficient torsional rotation of the tyrosine kinase domain, resulting in rapid closing of the VEGFR2 adeno-sine triphosphate binding site, rapid inactivation and weak transient acti-vation of downstream p42/44 (extra-cellular signal-regulated kinases 1/2 [ERK 1/2]) mitogen-activated protein kinase (MAPK)4. In addi-tion, VEGF-A165b binds to VEGFR2 but
not NRP1 due to its altered carboxy-terminus. Thus, a combination of weak VEGFR2 activation, lack of co-receptor binding and competition for binding sites with pro-angiogenic VEGF-A165a, could explain the anti-angiogenic properties of VEGF-A165b
4,6. Further-more, as VEGF-A usually functions as a disulphide-linked homodimer, there is the theoretical possibility that heterodimers of either non-paired, non-equivalent length isoforms (e.g. VEGF-A206 and VEGF-A121) or paired, equivalent length but non-identical isoforms (VEGF-A165a and VEGF-A165b) exist, causing increased complexity in regulating the angiogenic response4.
Membrane receptors that bind VEGFsVEGF-A binds with picomolar affinity to two different receptor tyrosine kinases (VEGFR1 and VEGFR2), with additional binding specificity for NRP co-receptors, NRP1 and NRP2,
Figure 1: VEGF-A splice isoforms exon composition. Exon composition of VEGF-A splice isoforms resulting from alterna-tive splicing of the VEGF-A pre mRNA consisting of eight exons and seven introns resulting in at least seven pro-angio-genic (a isoforms) and five antiangiogenic (b isoforms) splice variants. VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor tyrosine kinase.

Review
Page 3 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f Int
eres
ts: n
one
decl
ared
. A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t. A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
which lack tyrosine kinase activity7. VEGFR1 and VEGFR2 belong to the Class III receptor tyrosine kinase family and comprise of seven immu-noglobulin-like repeats within their extracellular domain, a single transmembrane region and a split cytoplasmic tyrosine kinase domain. Both VEGFR1 and VEGFR2 play important roles in vasculogenesis and angiogenesis as gene knockout mice die in utero at E8.5 and E9.5, respectively8,9.
VEGFR1 (Flt-1) has an estimated molecular mass of 151 kDa, but the mature protein undergoes processing and glycosylation to produce a species of approximately 180 kDa, as detected by sodium dodecyl sulphate
polyacrylamide gel electropho-resis (SDS-PAGE). VEGFR1 binds to VEGF-A, VEGF-B and PlGF1. VEGFR1 has highest affinity for VEGF-A (Kd≈10–30 pM)10 but displays rela-tively weak tyrosine kinase activity following VEGF-A binding2,11. These biochemical properties have led to the suggestion that the primary physiological role of VEGFR1 is to act as a negative regulator of angiogen-esis, by ‘trapping’ VEGF-A to reduce its bioavailability for VEGFR212. The VEGFR1 gene is located on chro-mosome 13q12 and contains 30 exons; alternative splicing of this pre-mRNA gives rise to both soluble and membrane-bound protein isoforms13. VEGFR1 is expressed by
both quiescent and actively prolif-erating endothelial cells suggesting that VEGFR1 has an important role in endothelial homeostasis2.
VEGFR2 (KDR) has a molecular mass of 152 kDa, but the mature protein undergoes processing and glycosylation to produce a species of approximately 200–230 kDa, as detected by SDS-PAGE. Only this mature, glycosylated form of VEGFR2 can be efficiently autophosphoryl-ated14. The binding affinity of VEGFR2 for VEGF-A is 5–10 fold lower than that of VEGFR1 (Kd≈75–125 pM)11; however, VEGFR2 has a higher tyrosine kinase activity1,15. VEGFR2 can also bind to VEGF-C, VEGF-D and the virally encoded VEGF-E variant1,16,17. In contrast to VEGFR1, VEGFR2 expression is downregulated in quiescent adult vasculature18; this is likely due to the fact that most of the pro-angiogenic gene expression is regulated by VEGFR27. VEGFR2 activation is primarily responsible for the majority of the biological effects linked to VEGF-A bioactivity, including vascular permeability, cell migration, cell survival, cell prolifera-tion and smooth muscle relaxation (Figure 2). Thus, VEGFR2-blocking therapies which are tailored to atten-uate specific intracellular signalling events or pathways, could reduce clinical side effects of current VEGF therapies19, including excessive vascular permeability, inflammation, tumour growth and metastasis.
NRPs are approximately 140 kDa transmembrane, non-tyrosine kinase, glycoprotein co-receptors for both the semaphorin and the VEGF family1. They comprise a large extra-cellular domain of approximately 850 residues, a transmembrane region and a relatively short cytoplasmic domain (approximately 40 residues). This cytoplasmic domain is likely to function as a docking site for various downstream signalling molecules1. NRP1 binds VEGF-A165a, PIGF-152 (the heparin sulphatebinding isoform) and both VEGF-B isoforms, whereas
Figure 2: VEGFR1 and VEGFR2 and their associated ligands. Black boxes on the receptors indicate important tyrosine residues. Yellow boxes show tyrosine residues 1054 and 1059 which are important for kinase activity.c-Akt, protein kinase B; EC, endothelial cell; eNOS, endothelial nitric oxide synthase; ERK1/2, extracellular signal-regulated kinase 1/2; FAK, focal adhe-sion kinase; HPC, hematopoietic progenitor cells; HSP27, heat shock protein-27; MAPK, mitogen-activated protein kinase; MEK, MAPK and ERK kinase; PI3K, phosphatidylinositol 3-kinase; PKC, protein kinase C; PLCγ1, phospholi-pase Cγ1; PlGF, placental growth factor; Shb, SH2 domain-containing adaptor protein b; TSAd, T-cell–specific adaptor, VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor tyrosine kinase.

Review
Page 4 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f int
eres
ts: n
one
decl
ared
.A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t.A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
NRP2 binds VEGF-A145, VEGFA165a, PIGF-152 and VEGF-C. It was origi-nally thought that neither NRP1 nor NRP2 could bind VEGF-A121
1,20, however, Pan et al. (2007)21 reported that the VEGF-A121 splice variant can bind to NRP1 in vitro but cannot promote the formation of a VEGFR2/NRP1 complex21,22. Co-expression of VEGFR2 and NRP1 in stably trans-fected porcine aortic endothelial cells leads to a stronger VEGF-A–stimu-lated response compared to cells expressing VEGFR2 alone6.
VEGFR tyrosine phosphorylation and intracellular signallingA key aspect of receptor tyrosine kinase function is the capacity to become activated upon binding ligands such as growth factors. In the case of VEGFRs, binding to VEGF dimers triggers receptor dimerisa-tion followed by trans-autophos-phorylation on specific cytoplasmic tyrosine residues. VEGFRs can also undergo heterodimerisation, but the functional significance remains unclear23-25. Ligand-stimulation of VEGFR1 elevates levels of urokinase plasminogen activator (uPA) and plasminogen activator inhibitor-1 (PAI-1), which in turn regulate extra-cellular matrix degradation and cell migration. Both uPA and PAI-1 modulate actin dynamics via p38 MAPK, thus regulating endothelial cell migration2,7,26,27. VEGFR1 is also capable of recruiting phospholipase Cγ1 (PLCγ1) after phosphorylation of residue Y1169, creating a binding site for the Src homology (SH2) domains in PLCγ11 (Figure 2). However, acti-vated VEGFR1 does not seem to directly impact cellular responses such as cell proliferation.
Activated VEGFR2 is phosphoryl-ated on 6–7 tyrosine residues, notably residues Y1175, Y951 and Y12141,19. Such post-translational modifications generate binding sites for a variety of regulatory proteins containing SH2 or phosphotyrosine binding domains1 (Figure 2). PLCγ1 binds to the pY1175 site and to the phos-
photyrosine-based epitope within VEGFR21,19, and its recruitment to the plasma membrane triggers phos-phatidylinositol-4,5-bisphosphate hydrolysis to diacylglycerol (DAG) and inositol 1,4,5-triphosphate (IP3). Generation of IP3 triggers activation of the IP3 receptor, which acts as a calcium channel on the endoplasmic reticulum, by mediating IP3-activated translocation of calcium ions from the endoplasmic reticulum to the cytosol. In contrast, DAG activates the protein kinase C (PKC) enzyme, which triggers MAPK pathway acti-vation and signalling. Additionally, VEGFR2 phosphorylation stimulates the activation of the phosphatidylino-sitol 3-kinase (PI3K) pathway, leading to increased c-Akt (protein kinase B) activity, subsequent endothelial nitric oxide synthase (eNOS) phosphoryla-tion and activation, resulting in nitric oxide (NO) production1,19. Increased levels of NO in response to VEGF stimulation, promote a positive feed-back loop in angiogenesis: NO stimu-lates expression and transcriptional activity of the hypoxia inducible factor complex which stimulates VEGF-A mRNA transcription28. Activated PKC enzymes are postulated to phospho-rylate and activate MEK (MAPK and ERK kinases), which subsequently phosphorylates and activates p42/44 MAPK, leading to changes in gene expression and cell proliferation. However, studies implicate the PKCε isoform as a key regulator of VEGF-A–stimulated phosphorylation and acti-vation of c-Akt and eNOS29.
A small adaptor molecule called Shb (SH2 domain-containing adaptor protein b) also binds to the VEGFR2-pY1175 phospho-epitope and acti-vates PI3K, which sequentially stim-ulates activation of c-Akt and eNOS, thus promoting cell survival and NO production, respectively1,19. Binding of a T-cell–specific adaptor protein (TSAd) to the same phospho-epitope is also implicated in c-Src–regu-lated endothelial cell migration and vascular permeability. Generation
of the VEGFR2-pY1214 phospho-epitope is linked to increased actin remodelling and cell migration through a pathway involving CDC42, p38 MAPK and heat shock protein 27 kDa. Furthermore, the VEGFR2-pY1214 site is linked to focal adhe-sion turnover and cell migration through focal adhesion kinase and its substrate paxillin.
VEGFR ubiquitination and proteolysisA key feature of receptor-ligand complex formation is temporal and spatial regulation linked to subsequent cellular responses. For example, conventional models of receptor tyrosine kinase activation postulate that binding to ligands at the plasma membrane is followed by trafficking through the endosome-lysosome system, which is linked to receptor-ligand proteolysis. Interest-ingly, VEGFR1 and VEGFR2 display markedly different profiles in this respect. VEGFR1 levels appear to be relatively insensitive to VEGF-A stimulation, unlike VEGFR230,31. Work from our group suggests that only 20% of VEGFR1 is available at the plasma membrane, whereas the remaining pool is located within an intracellular Golgi-like compart-ment31. Other studies suggest that VEGFR1 displays both cytoplasmic and nuclear pools in both endothelial and non-endothelial cells32,33. None-theless, calcium-regulated VEGFR1 trafficking from an intracellular membrane-bound compartment to the plasma membrane may be one way of generating a negative-feed-back loop to modulate VEGF-A–stimu-lated VEGFR2 response in endothelial cells31,34.
VEGFR2 displays a complex pattern of distribution to the secretory pathway, plasma membrane and endosome-lysosome system, with marked proteolytic sensitivity30,31,35. VEGF-A binding stimulates VEGFR2 trafficking through endosome-lyso-

Review
Page 5 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f Int
eres
ts: n
one
decl
ared
. A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t. A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
some compartments30,36-38 and the nucleus33. A key aspect of VEGFR2 activation by VEGF-A binding is ubiquitination and proteolysis30,38 (Figure 3). VEGFR2 proteolysis is also tightly regulated with at least two different proteolytic activities postu-lated for its cleavage within the endo-some-lysosome system: a 26S protea-some-regulated step associated with early endosomes for processing of the cytoplasmic domain and lyso-somal activity for processing of the extracellular/luminal domain30,38 (Figure 3).
The process of ubiquitination involves a reversible post-translational modification of protein substrates39 which programmes proteolysis and/or intracellular re-distribution and thus regulates altered protein func-
tion. Intriguingly, inactive VEGFR2 may also be ubiquitinated as it under-goes slow proteolysis in the absence of VEGF-A31,40. One view is that stimu-lation of VEGF-A accentuates endog-enous VEGFR2 ubiquitination, thus promoting intracellular re-distri-bution and clearance. Sequential complex assembly, based on multiple ubiquitin–ubiquitin-binding domain interactions, enables the propagation of signalling events, which control the dynamics of receptor trafficking through the endosome-lysosome system41,42. Ubiquitin conjugation to target proteins involves E1, E2 and E3 enzymes or ligases. Thus a specific E1/E2/E3 cascade must regulate VEGFR2 ubiquitination in response to VEGF-A stimulation. Notably, de-ubiq-uitinating enzymes (DUBs) have been
implicated in regulating receptor tyrosine kinase function43-45.
VEGFR2 regulation by ESCRTsFollowing VEGF-A binding and acti-vation at the plasma membrane, the VEGFR2–VEGF-A complex is endo-cytosed and transported to early endosomes where an ubiquitin-linked receptor complex, endosomal sorting complex required for trans-port (ESCRT-0), mediates sorting and delivery to late endosomes41,46 (Figure 3). An array of endocytic regulators working with the endo-somal ubiquitin receptors, namely Hrs, STAM and Eps15, initially engage activated receptor tyrosine kinase–ligand complexes in early endosomes for subsequent delivery to multivesicular body endosomes47; this is followed by trafficking towards the lysosome for terminal degradation48. The ESCRT-0 heter-odimer can bind several ubiquitin moieties simultaneously via the ubiquitin-interacting motif of the signal transducing adaptor molecule (STAM) subunit and the di-ubiquitin motif of the Hrs subunit41. The Hrs/STAM complex shows increased co-distribution with VEGFR2 after VEGF-A stimulation30, suggesting active sorting of the receptor–ligand complex further into the endosome-lysosome system. After sorting, ubiquitin attached to cargo proteins can be removed by endo-somal DUBs, such as the associated molecule with the SH3 domain of STAM (AMSH) and ubiquitin-specific protease Y (UBPY)49. The process of de-ubiquitination enables the ESCRT machinery to dissociate from its bound cargo and recycle for subsequent use in the trafficking of other ubiquitinated molecules. Due to their functional role in epidermal growth factor receptor (ErbB1) traf-ficking, UBPY and AMSH49 can thus mediate VEGFR2 recycling from early endosomes back to the plasma membrane.
Figure 3: VEGFR2 trafficking and degradation. Non-stimulated VEGFR2 under-goes constitutive internalisation and recycling back to the plasma membrane. Following internalisation, VEGF-A–stimulated VEGFR2 is either recycled or degraded. VEGFR2 can recycle independently (Rab4a pathway) or via a NRP1 mediated pathway (Rab11 pathway). Interaction with the ESCRT machinery in early endosomes regulates transport to late endosomes where the VEGFR2 C-terminal domain can be proteolytically cleaved prior to final degradation in the lysosome. EE, early endosome; ESCRT, endosomal sorting complex required for transport; LE, late endosome; Lys, lysosome; MVB, multivesicular body; NRP, neuropilin; RE, recycling endosome; VEGFR, vascular endothelial growth factor receptor tyrosine kinase.

Review
Page 6 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f int
eres
ts: n
one
decl
ared
.A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t.A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
Traffi cking of the VEGFR2-NRP1 complexInteraction between the NRP1 binding adaptor synectin and myosin VI, mediates the internalisation of VEGFR2 into Rab5a–positive early endosomes50,51. Internalised, activated VEGFR2–VEGF-A complexes can be recycled to the plasma membrane primarily from Rab4a-positive recy-cling endosomes36. NRP1 itself recy-cles via Rab11a-positive vesicles, following transport from Rab4a-posi-tive vesicles. The NRP1 C-terminal domain contains a carboxy-terminal tripeptide motif (-SEA*) that binds a PDZ-binding motif within synectin/GIPC17,52-54. VEGF165a-stimulated inter-action between VEGFR2 and NRP1, is linked to actin-dependent recy-cling through a Rab5a-Rab4a-Rab11a endosomal pathway17.
VEGFR2 regulation by VE-cadherin and c-SrcVEGFR2 can be recycled through a non-conventional pathway in response to ligand stimulation36. Phos-phorylated VEGFR2 is delivered into endosomes via a vascular endothe-lial (VE)-cadherin–linked, clathrin-regulated endocytic pathway; here, receptor stability is retained, leading to prolonged signalling outputs which ultimately result in increased cell migration and proliferation37. Activated VEGFR2 recruits TSAd, which subsequently enables recruit-ment of the tyrosine kinase c-Src which is a potent effector of cell proliferation55. Phosphorylation of c-Src by VEGFR2 promotes activation and downstream phosphorylation of p21-activated kinase 2 (PAK2)56. Activated PAK2 phosphorylates key serine residues on VE-cadherin within the binding region for p120catenin57. Dissociation of p120-catenin exposes a short endocytic motif (DEE) within VE-cadherin58, promoting endocytosis, disruption of endothelial adherens junctions and increased vascular permeability. However, VEGFR2 and VE-cadherin
appear to follow different endocytic routes from the plasma membrane59. The c-Src enzyme shows increased co-distribution with intracellular stores of VEGFR2 upon VEGF-A stim-ulation36; however, there is uncer-tainty as to whether this is a late secretory (e.g. trans-Golgi network) or endosomal pool of VEGFR2. None-theless, both VEGFR2 and c-Src are recycled through the same endo-cytic recycling pathway. In quiescent endothelial cells, VEGFR2 is stored in intracellular Rab4/Rab11-nega-tive vesicles and delivered to the plasma membrane in a c-Src activa-tion-dependent manner in response to VEGF-A36, highlighting unique differences in comparison to other receptor tyrosine kinases.
Endosome-associated VEGFR2 cytoplasmic domain proteolysisVEGF-A stimulation results in the degradation of the mature, glyco-sylated approximately 230 kDa VEGFR2 species and an increase in the levels of a novel 160 kDa VEGFR2 cytoplasmic domain-related proteo-lytic polypeptide30,38. Production of this proteolytic 160 kDa fragment requires VEGF-A–stimulated VEGFR2 activity and subsequent endosomal trafficking38. Mono-ubiquitination of VEGFR2 precedes the production of this proteolytic fragment; this modification could act to mediate the 26S proteasome recognition of early endosomal VEGFR2, before subsequent terminal lysosomal degradation38. Carboxy-terminal post-translational modifications and recruitment of signalling effectors bestow the VEGFR2 cytoplasmic domain with important roles in endothelial function60. Inhibition of this limited cytoplasmic domain proteolysis alters endothelial cell migration and VEGF-A–regulated downstream signalling, prolonging the phosphorylation and activation of p42/44 MAPK, c-Akt and eNOS38. Thus cleavage of the VEGFR2 cyto-plasmic domain may be a key event in
controlling the duration and intensity of endothelial signalling in response to VEGF-A61.
PKC regulation of VEGFR2 function VEGF-A may promote degradation of VEGFR2 by downstream activation of non-classical PKC isozymes. Direct or indirect PKC-mediated phosphoryla-tion of VEGFR2 cytoplasmic domain residues S1188 and/or S1191, marks the receptor for PKC-regulated inter-nalisation and proteasomal degra-dation61. Thus, VEGF-A–stimulated PKC activation may promote the 26S proteasome-regulated removal of the VEGFR2 cytoplasmic domain30,38. Recent findings by Nakayama et al. (2013)62 report that atypical protein kinase C (aPKC) activity negatively regulates the endocytosis and turn-over of VEGFR2 within mature estab-lished blood vessels (where levels of aPKC are high) but not so much in sprouting vessel tips (were levels of aPKC are low)62.
ConclusionEmerging concepts in different areas of classical biochemistry and cell bi-ology highlight the importance of in-tegration between signalling path-w-ays with membrane trafficking, prot-ein biosynthesis and proteolysis, to regulate cell and tissue function. Tis is exquisitely illustrated in the VEGF regulation of animal physiology as the interaction between such ligands and their cognate receptors fine tun-es animal biology in both physiologi-cal and pathological states. A major challenge in this area is to understa-nd the complexity of VEGF isoforms in programming differential recept-or signalling, turnover and function. Many lines of evidence suggest that VEGFs have subtle effects on cell ho-meostasis in the immune, epithelial and nervous systems, but these un-derlying mechanisms remain elusive. The development of primary cell systems, multicellular models, organ culture and animal models, will dramatically increase our ability to

Review
Page 7 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f Int
eres
ts: n
one
decl
ared
. A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t. A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
understand the biochemical basis for how this important class of cytokines regulate animal physiology. Targeting such pathways directly or indirectly using humanised antibodies or small molecule inhibitors is important for treating a wide variety of patho-logical states including solid tumour growth and metastasis, age-related macular degeneration and condi-tions involving blood vessel repair and regeneration.
Abbreviations listAMSH, associated molecule with the SH3 domain of STAM; aPKC, atyp-ical protein kinase C; DAG, diacyl-glycerol; DUB, de-ubiquitinating enzyme; eNOS, endothelial nitric oxide synthase; ERK, extracel-lular signal-regulated kinase; IP3, inositol-1,4,5-triphosphate; MAPK, mitogen-activated protein kinase; NO, nitric oxide; NRP, neuropilin; PAI-1, plasminogen activator inhib-itor-1; PAK2, p21-activated kinase 2; PlGF, placental growth factor; PI3K, phosphatidylinositol 3 kinase; PKC, protein kinase C; PLCγ1, phospholi-pase Cγ1; SDS-PAGE, sodium dodecyl sulphate polyacrylamide gel electro-phoresis; SH, Src homology; STAM, signal transducing adaptor molecule; TSAd, T-cell–specific adaptor protein; UBPY, ubiquitin-specific protease Y; uPA, urokinase plasminogen activator; VE-cadherin, vascular endothelial-cadherin; VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor tyrosine kinase.
Referen ces1. Roskoski R Jr. Vascular endothelial growth factor (VEGF) signaling in tumor progression. Crit Rev Oncol. Hemat. 2007 Jun;62(3):179–213.2. Robinson CJ, Stringer SE. The splice variants of vascular endothelial growth factor (VEGF) and their receptors. J Cell Sci. 2001 Mar;114(5):853–65.3. Vincenti V, Cassano C, Rocchi M, Persico G. Assignment of the vascular endothelial growth factor gene to human chromosome 6p21.3. Circulation. 1996 Apr;93(8):1493–5.
4. Harper SJ, Bates DO. VEGF-a splicing: The key to anti-angiogenic therapeu-tics? Nat Rev Cancer. 2008 Apr;93(8):1493–5.5. Harris S, Craze M, Newton J, Fisher M, Shima DT, Tozer GM, et al. Do anti-angiogenic VEGF (VEGFxxxb) isoforms exist? A cautionary tale. Plos One. 2012 May;7(5):e35231.6. Kawamura H, Li X, Harper SJ, Bates DO, Claesson-Welsh L. Vascular endothelial growth factor (VEGF)-a165b is a weak in vitro agonist for VEGF receptor-2 due to lack of coreceptor binding and deficient regulation of kinase activity. Cancer Res. 2008 Jun;68(12):4683–92.7. Kanno S, Oda N, Abe M, Terai Y, Ito M, Shitara K, et al. Roles of two VEGF receptors, Flt-1 and KDR, in the signal transduction of VEGF effects in human vascular endothelial cells. Oncogene. 2000 Apr;19(17):2138–46.8. Shalaby F, Rossant J, Yamaguchi TP, Gertsenstein M, Wu XF, Breitman ML, et al. Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature. 1995 Jul;376(6535):62–6.9. Fong GH, Rossant J, Gertsenstein M, Breitman ML. Role of the Flt-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature. 1995 Jul;376(6535):66–70.10. Devries C, Escobedo JA, Ueno H, Houck K, Ferrara N, Williams LT. The Fms-like tyrosine kinase, a receptor for vascular endothelial growth-factor. Science. 1992 Feb;255(5047):989–91.11. Waltenberger J, Claessonwelsh L, Siegbahn A, Shibuya M, Heldin CH. Different signal-transduction properties of KDR and Flt1, 2 receptors for vascular endothelial growth-factor. J Biol Chem. 1994 Oct;269(43):26988–95.12. Rahimi N. VEGFR-1 and VEGFR-2: two non-identical twins with a unique physiog-nomy. Front Biosci. 2006 Jan;11:818–29.13. Kendall RL, Thomas KA. Inhibition of vascular endothelial-cell growth-factor activity by an endogenously encoded soluble receptor. Proc Natl Acad Sci USA. 1993 Nov;90(22):10705–9.14. Takahashi T, Shibuya M. The 230 kDa mature form of KDR/Flk-1 (VEGF receptor-2) activates the PLC-gamma pathway and partially induces mitotic signals in NIH3T3 fibroblasts. Oncogene. 1997 May;14(17):2079–89.15. Quinn TP, Peters KG, Devries C, Ferrara N, Williams LT. Fetal liver kinase-1 is a
receptor for vascular endothelial growth-factor and is selectively expressed in vascular endothelium. Proc Natl Acad Sci USA. 1993 Aug;90(16):7533–7.16. Zheng Y, Murakami M, Takahashi H, Yamauchi M, Kiba A, Yamaguchi S, et al. Chimeric VEGF-e-nz7/pigf promotes angiogenesis via VEGFR-2 without signif-icant enhancement of vascular permea-bility and inflammation. Arterioscl Throm Vasc Biol. 2006 Sep;26(9):2019–26.17. Ballmer-Hofer K, Andersson AE, Ratcliffe LE, Berger P. Neuropilin-1 promotes VEGFR-2 trafficking through rab11 vesicles thereby specifying signal output. Blood. 2011 Jul;118(3):816–26.18. Eichmann A, Corbel C, Nataf V, Vaigot P, Breant C, LeDouarin NM. Ligand-dependent development of the endothe-lial and hemopoietic lineages from embryonic mesodermal cells expressing vascular endothelial growth factor receptor 2. Proc Natl Acad Sci USA. 1997 May;94(10):5141–6.19. Olsson AK, Dimberg A, Kreuger J, Claesson-Welsh L. VEGF receptor signal-ling–in control of vascular function. Nat Rev Mol Cell Biol. 2006 May;7(5):359–71.20. Kawamura H, Li X, Goishi K, van Meeteren LA, Jakobsson L, Cebe-Suarez S, et al. Neuropilin-1 in regulation of VEGF-induced activation of p38MAPK and endothelial cell organization. Blood. 2008 Nov;112(9):3638–49.21. Pan Q, Chathery Y, Wu Y, Rathore N, Tong RK, Peale F, et al. Neuropilin-1 binds to VEGF121 and regulates endothelial cell migration and sprouting. J Biol Chem. 2007 Aug;282(33):24049–56.22. Zachary IC. How Neuropilin-1 regulates receptor tyrosine kinase signalling: the knowns and known unknowns. Biochem Soc Trans. 2011 Dec;39(6):1583–91.23. Kendall RL, Wang G, Thomas KA. Identification of a natural soluble form of the vascular endothelial growth factor receptor, Flt-1, and its heterodimeriza-tion with KDR. Biochem Biophys Res Commun. 1996 Sep;226(2):324–8.24. Neagoe PE, Lemieux C, Sirois MG. Vascular endothelial growth factor (VEGF)-A165-induced prostacyclin synthesis requires the activation of VEGF receptor-1 and -2 heterodimer. J Biol Chem. 2005 Mar;280(11):9904–12.25. Cudmore MJ, Hewett PW, Ahmad S, Wang KQ, Cai M, Al-Ani B, et al. The role of heterodimerization between VEGFR-1 and VEGFR-2 in the regulation of endothe-

Review
Page 8 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f int
eres
ts: n
one
decl
ared
.A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t.A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
lial cell homeostasis. Nat Commun. 2012 Jul;3:972.26. Landgren E, Schiller P, Cao YH, Claesson-Welsh L. Placenta growth factor stimulates map kinase and mitogenicity but not phospholipase Cg and migration of endothelial cells expressing Flt1. Onco-gene. 1998 Jan;16(3):359–67.27. Olofsson B, Korpelainen E, Pepper MS, Mandriota SJ, Aase K, Kumar V, et al. Vascular endothelial growth factor b (VEGF-B) binds to VEGF receptor-1 and regulates plasminogen activator activity in endothelial cells. Proc Natl Acad Sci USA. 1998 Sep;95(20):11709–14.28. Karar J, Maity A. PI3k/Akt/mTOR pathway in angiogenesis. Front Mol Neurosci. 2011;4:51.29. Rask-Madsen C, King GL. Differential regulation of VEGF signaling by PKC-alpha and PKC-epsilon in endothelial cells. Arterioscl Throm Vasc Biol. 2008 May;28(4):608–10.30. Ewan LC, Jopling HM, Jia H, Mittar S, Bagherzadeh A, Howell GJ, et al. Intrinsic tyrosine kinase activity is required for vascular endothelial growth factor receptor 2 ubiquitination, sorting and degradation in endothelial cells. Traffic. 2006 Sep;7(9):1270–82.31. Mittar S, Ulyatt C, Howell GJ, Bruns AF, Zachary I, Walker JH, et al. VEGFR1 receptor tyrosine kinase localization to the golgi apparatus is calcium-dependent. Exp Cell Res. 2009 Mar;315(5):877–89.32. Lee TH, Seng S, Sekine M, Hinton C, Fu Y, Avraham HK, et al. Vascular endothe-lial growth factor mediates intracrine survival in human breast carcinoma cells through internally expressed vegfr1/flt1. PLoS Med. 2007 Jun;4(6):1101–16.33. Zhang Z, Neiva KG, Lingen MW, Ellis LM, Nor JE. VEGF-dependent tumor angi-ogenesis requires inverse and reciprocal regulation of VEGFR1 and VEGFR2. Cell Death Differ. 2010 Mar;17(3):499–512.34. Bruns AF, Bao L, Walker JH, Ponnam-balam S. VEGF-A–stimulated signalling in endothelial cells via a dual receptor tyrosine kinase system is dependent on co-ordinated trafficking and proteolysis. Biochem Soc Trans. 2009 Dec;37:1193–7.35. Manickam V, Tiwari A, Jung J-J, Bhat-tacharya R, Goel A, Mukhopadhyay D, et al. Regulation of vascular endothelial growth factor receptor 2 trafficking and angiogenesis by Golgi localized t-SNARE syntaxin 6. Blood. 2011 Jan;117(4):1425–35.
36. Gampel A, Moss L, Jones MC, Brunton V, Norman JC, Mellor H. VEGF regulates the mobilization of VEGFR2/KDR from an intracellular endothelial storage compart-ment. Blood. 2006 Oct;108(8):2624–31.37. Lampugnani MG, Orsenigo F, Gagliani MC, Tacchetti C, Dejana E. Vascular endothelial cadherin controls VEGFR-2 internalization and signaling from intra-cellular compartments. J Cell Biol. 2006 Aug;174(4):593–604.38. Bruns AF, Herbert SP, Odell AF, Jopling HM, Hooper NM, Zachary IC, et al. Ligand-stimulated VEGFR2 signaling is regulated by co-ordinated trafficking and proteol-ysis. Traffic. 2010 Jan;11(1):161–74.39. Herrmann J, Lerman LO, Lerman A. Ubiquitin and ubiquitin-like proteins in protein regulation. Circ Res. 2007 May;100(9):1276–91.40. Ulyatt C, Walker J, Ponnambalam S. Hypoxia differentially regulates VEGFR1 and VEGFR2 levels and alters intracel-lular signaling and cell migration in endothelial cells. Biochem Biophys Res Commun. 2011 Jan;404(3):774–9.41. Grabbe C, Husnjak K, Dikic I. The spatial and temporal organization of ubiquitin networks. Nat Rev Mol Cell Biol. 2011 May;12(5):295–307.42. Clague MJ, Coulson JM, Urbe S. Cellular functions of the DUBs. J Cell Sci. 2012 Jan;125(2):277–86.43. McCullough J, Clague MJ, Urbe S. AMSH is an endosome-associated ubiq-uitin isopeptidase. J Cell Biol. 2004 Aug;166(4):487–92.44. Liu H, Buus R, Clague MJ, Urbe S. Regulation of erbb2 receptor status by the proteasomal DUB POH1. PLoS One. 2009;4(5):e5544.45. Carter S, Urbe S, Clague MJ. The Met receptor degradation pathway–require-ment for Lys(48)-linked polyubiquitin independent of proteasome activity. J Biol Chem. 2004 Dec;279(51):52835–9.46. Hasseine LK, Murdaca J, Suavet F, Longnus S, Giorgetti-Peraldi S, Van Obberghen E. Hrs is a positive regulator of VEGF and insulin signaling. Exp Cell Res. 2007 May;313(9):1927–42.47. de Melker AA, van der Horst G, Calafat J, Jansen H, Borst J. C-cbl ubiqui-tinates the EGF receptor at the plasma membrane and remains receptor associ-ated throughout the endocytic route. J Cell Sci. 2001 Jun;114(11):2167–78.48. Bilodeau PS, Urbanowski JL, Winis-torfer SC, Piper RC. The Vps27p-Hse1p
complex binds ubiquitin and mediates endosomal protein sorting. Nat Cell Biol. 2002 Jul;4(7):534–9.49. Clague MJ, Urbe S. Endocytosis: the DUB version. Trends Cell Biol. 2006 Nov;16(11):551–9.50. Nakamura Y, Patrushev N, Inomata H, Mehta D, Urao N, Kim HW, et al. Role of protein tyrosine phosphatase 1b in vascular endothelial growth factor sign-aling and cell-cell adhesions in endothe-lial cells. Circ Res. 2008 May;102(10):1182–91.51. Lanahan AA, Hermans K, Claes F, Kerley-Hamilton JS, Zhuang ZW, Giordano FJ, et al. VEGF receptor 2 endocytic traf-ficking regulates arterial morphogenesis. Dev Cell. 2010 May;18(5):713–24.52. Cai HB, Reed RR. Cloning and char-acterization of neuropilin-1-interacting protein: A Psd-95/Dlg/ZO-1 domain-containing protein that interacts with the cytoplasmic domain of Neuropilin-1. J Neurosci. 1999 Aug;19(15):6519–27.53. Wang L, Zeng HY, Wang P, Soker S, Mukhopadhyay D. Neuropilin-1-mediated vascular permeability factor/vascular endothelial growth factor-dependent endothelial cell migration. J Biol Chem. 2003 Dec;278(49):48848–60.54. Chittenden TW, Claes F, Lanahan AA, Autiero M, Palac RT, Tkachenko EV, et al. Selective regulation of arterial branching morphogenesis by synectin. Dev Cell. 2006 Jun;10(6):783–95.55. Sun Z, Li X, Massena S, Kutschera S, Padhan N, Gualandi L, et al. VEGFR2 induces c-Src signaling and vascular permeability in vivo via the adaptor protein TSAd. J Exp Med. 2012 Jul;209(7):1363–77.56. Weis S, Cui JH, Barnes L, Cheresh D. Endothelial barrier disruption by VEGF-mediated Src activity potentiates tumor cell extravasation and metastasis. J Cell Biol. 2004 Oct;167(2):223–9.57. Adam AP, Sharenko AL, Pumiglia K, Vincent PA. Src-induced tyrosine phos-phorylation of VE-cadherin is not suffi-cient to decrease barrier function of endothelial monolayers. J Biol Chem. 2010 Mar;285(10):7045–55.58. Nanes BA, Chiasson-MacKenzie C, Lowery AM, Ishiyama N, Faundez V, Ikura M, et al. p120-catenin binding masks an endocytic signal conserved in classical cadherins. J Cell Biol. 2012 Oct;199(2):365–80.

Review
Page 9 of 9
Com
pe n
g in
tere
sts:
non
e de
clar
ed. C
onfl i
ct o
f Int
eres
ts: n
one
decl
ared
. A
ll au
thor
s co
ntrib
uted
to th
e co
ncep
on,
des
ign,
and
pre
para
on
of th
e m
anus
crip
t, a
s w
ell a
s re
ad a
nd a
ppro
ved
the fi n
al m
anus
crip
t. A
ll au
thor
s ab
ide
by th
e A
ssoc
ia o
n fo
r Med
ical
Eth
ics
(AM
E) e
thic
al ru
les
of d
iscl
osur
e.
Licensee OA Publishing London 2013. Creative Commons Attribution Licence (CC-BY)
F : Fearnley GW, Smith GA, Harrison MA, Wheatcroft SB, Tomlinson DC, Ponnambalam S. Vascular endothelial growth factor-A regulation of blood vessel sprouting in health and disease. OA Biochemistry 2013 Feb 01;1(1):5.
59. Sandilands E, Cans C, Fincham VJ, Brunton VG, Mellor H, Prendergast GC, et al. Rhob and actin polymerization coordi-nate Src activation with endosome-medi-ated delivery to the membrane. Dev Cell. 2004 Dec;7(6):855–69.60. Meyer RD, Srinivasan S, Singh AJ, Mahoney JE, Gharahassanlou KR, Rahimi N. Pest motif serine and tyrosine phos-phorylation controls vascular endothe-
61. Singh AJ, Meyer RD, Band H, Rahimi N. The carboxyl terminus of VEGFR-2 is required for PKC-mediated down-regu-lation. Mol Biol Cell. 2005 Apr;16(4):2106–18.
62. Nakayama M, Nakayama A, van Lessen M, Yamamoto H, Hoffmann S, Drexler HC, et al. Spatial regulation of VEGF receptor endocytosis in angiogenesis. Nat Cell Biol. 2013 Jan;15(3):249–60.
lial growth factor receptor 2 stability and downregulation. Mol Cell Biol. 2011 May;31(10):2010–25.