variability in movement patterns and habitat use of two
TRANSCRIPT

Clemson UniversityTigerPrints
All Theses Theses
5-2015
Variability in Movement Patterns and Habitat useof Two Species of PelecaniformesCaroline PoliClemson University
Follow this and additional works at: https://tigerprints.clemson.edu/all_theses
Part of the Aquaculture and Fisheries Commons, and the Biology Commons
This Thesis is brought to you for free and open access by the Theses at TigerPrints. It has been accepted for inclusion in All Theses by an authorizedadministrator of TigerPrints. For more information, please contact [email protected].
Recommended CitationPoli, Caroline, "Variability in Movement Patterns and Habitat use of Two Species of Pelecaniformes" (2015). All Theses. 2161.https://tigerprints.clemson.edu/all_theses/2161

VARIABILITY IN MOVEMENT PATTERNS AND HABITAT USE OF TWO
SPECIES OF PELECANIFORMES
___________________________________________________________________
A Thesis
Presented to
the Graduate School of
Clemson University
___________________________________________________________________
In Partial Fulfillment
of the Requirements for the Degree
Master of Science
Wildlife and Fisheries Biology
___________________________________________________________________
by
Caroline Poli
May 2015
___________________________________________________________________
Accepted by:
Patrick Jodice, Committee Chair
Patrick Gerard
Autumn-Lynn Harrison
William Mackin

ii
ABSTRACT
Information regarding movement patterns of seabirds is useful for understanding
prey abundance and distribution, predicting overlap between bird activity and
anthropogenic stressors, and assessing the effectiveness of reserves at protecting nesting
or foraging habitat(Cairns 1992, Weimerskirch 2007, Weimerskirch et al. 2010). I
examined movement patterns of 40 brown pelicans (Pelecanus occidentalis) satellite
tagged in December 2010 in South Carolina, USA, and tracked until July 2013. Birds
exhibited individual variability in movement patterns and 25% of individuals remained
near the breeding colony during nonbreeding. Birds captured in South Carolina that did
migrate wintered as far south as Guatemala, prospected and used other breeding colonies
both south and north of the capture location, and exhibited prebreeding and postbreeding
dispersal. Overlap between brown pelican home ranges and protected areas in South
Carolina was low compared to availability of protected habitat; brown pelicans frequently
used unprotected nearshore pelagic areas. The percentage of offshore locations within 10
km of the coast was high (98%), suggesting that brown pelican movements may overlap
with future offshore wind development and should be accounted for during spatial
planning.
I also examined foraging behavior and habitat use of 77 masked boobies (Sula
dactylatra) tracked with GPS units during May and November 2013 at a regionally
important breeding colony in the southern Gulf of Mexico. Birds exhibited individual
variability in foraging behavior and searched for prey at nested hierarchical scales

iii
ranging from 60 m - 30 km. Characteristics of foraging trips and foraging behavior did
not differ between sexes but did differ between stages; behavior in incubating birds was
more variable compared to chick-rearing birds. The oceanographic variables that were
related to foraging behavior at small scales (at the prey patch level) varied widely
between individuals, suggesting that individual masked boobies use different foraging
strategies. Sea surface height and velocity of water corresponded to foraging most
frequently (44% and 38% of birds respectively), a finding that is consistent with the
characterization of the Gulf of Mexico as a highly dynamic system strongly influenced by
currents and eddies.
The two data chapters included in this thesis represent a substantial enhancement
to our understanding of movement patterns and spatial ecology of Pelecaniformes in the
north Atlantic system. Data sets from both chapters can provide valuable information for
marine spatial planning efforts and provide a baseline for anthropogenic based threats
such as development, pollution, and commercial fisheries.

iv
ACKNOWLEDGEMENTS
Thank you foremost to my advisor Dr. Patrick Jodice for the opportunity to
conduct research that I am passionate about, and for spending valuable time and energy
to provide insightful guidance in all aspects of building a career as a scientist. I also thank
my committee members, Dr. Autumn-Lynn Harrison, Dr. Patrick Gerard, and Dr. Will
Mackin for scientific comments and suggestions, engaging discussions, and
encouragement throughout my graduate study. I am grateful to Dr. Adriana Vallarino
Moncada for the wonderful opportunity to work at Arrecife Alacranes as part of her team.
Each of you has helped me become a more versatile scientist.
I acknowledge the USGS South Carolina Cooperative Fish and Wildlife Research
Unit, particularly Carolyn Wakefield who offered essential support, Dr. Kate McFadden
who provided strong mentorship early in my graduate study, and lab-mates and office-
mates who helped me focus my ideas by listening through hours of caffeine-induced
chatter. This project would not have been possible without major funding from South
Carolina Department of Natural Resources, and small grants from Sacramento Zoological
Society and Clemson University Graduate Student Government.
I thank my family and friends from near and far who encouraged me to pursue
graduate study, championed me during my research, and will undoubtedly continue to
support me in the future. Three resources in particular helped me build a stronger thesis,
All In Coffee Shop, Roget's 21st Century Thesaurus, and Google.

v
TABLE OF CONTENTS
Page
TITLE PAGE .................................................................................................................... I
ABSTRACT .................................................................................................................... II
ACKNOWLEDGEMENTS ........................................................................................... IV
LIST OF TABLES ....................................................................................................... VII
LIST OF FIGURES ....................................................................................................... IX
LIST OF APPENDICES ............................................................................................. XIV
CHAPTER ONE .............................................................................................................. 1
INTRODUCTION ..................................................................................................... 1
CHAPTER TWO ............................................................................................................. 5
VARIABILITY IN HOME RANGE SIZE, MIGRATION STRATEGY, AND
PROTECTED AREA USE OF EASTERN BROWN PELICANS
(PELECANUS OCCIDENTALIS) ALONG THE US ATLANTIC COAST ...... 5
Introduction .................................................................................................. 5
Methods ......................................................................................................... 8
Deployment of satellite tags ..................................................................... 8
Data processing ....................................................................................... 9
Home range estimation .......................................................................... 10
Migration strategy ................................................................................. 12
Habitat use ............................................................................................. 12
Reserve use............................................................................................. 13
Results ......................................................................................................... 14
Tag functionality .................................................................................... 14
Distribution and home ranges ............................................................... 17
Migration strategy ................................................................................. 21
Habitat use ............................................................................................. 26
Reserve use............................................................................................. 31
Discussion.................................................................................................... 34
Distribution and home ranges ............................................................... 34
Migration ............................................................................................... 36
Habitat use ............................................................................................. 39

vi
Reserve use............................................................................................. 41
CHAPTER THREE ....................................................................................................... 43
VARIABILITY IN FORAGING BEHAVIOR OF MASKED BOOBIES
(SULA DACTYLATRA) BREEDING AT ISLA MUERTOS, MEXICO ........ 43
Introduction ................................................................................................ 43
Methods ....................................................................................................... 46
Study area .............................................................................................. 46
Data collection ....................................................................................... 48
Habitat use ............................................................................................. 49
Statistical analysis ................................................................................. 54
Results ......................................................................................................... 57
Scales of area restricted search ............................................................. 59
At-sea movements and foraging behavior.............................................. 61
Oceanographic habitat use .................................................................... 67
Discussion.................................................................................................... 71
At-sea movements and foraging behavior.............................................. 71
Sex-specific foraging behavior .............................................................. 73
Oceanographic habitat use .................................................................... 75
Scales of ARS ......................................................................................... 78
Reserve use............................................................................................. 79
CONCLUSIONS............................................................................................................ 81
APPENDICES ............................................................................................................... 83
Appendix A ................................................................................................. 84
Appendix B ................................................................................................. 86
Appendix C ................................................................................................. 88
REFERENCES .............................................................................................................. 90

vii
LIST OF TABLES
Table Page
2.1 Areal extent (mean ± sd km2) of focal region (50% utilization distribution,
UD), home range (95% UD), and percent overlap in successive years (an
estimate of site fidelity) during breeding and nonbreeding seasons for
brown pelicans satellite tagged December 2010 in South Carolina, USA,
and tracked until July 2013. Home range and focal region were log-
transformed. n = number of birds, W = Wilcoxon rank-sum test,
Significant results appear in bold; alpha = 0.1. .................................................20
2.2 Discrete migration strategies of brown pelicans satellite tagged in South
Carolina, December 2010. Satellite tags transmitted locations for 3 months
- 3.5 years. Four groupings were identified for birds with at least four
months of location data and strategies were classified as highly migratory,
migratory, semi-migratory, and stationary based on Ward hierarchical
clustering analysis of the latitudinal range for each individual. At the 0.05
significance level, stationary individuals were significantly different from
all other groups. ................................................................................................23
2.3 Proportional overlap of undesignated habitat and six protected areas in
South Carolina with individual home ranges of 20 brown pelicans satellite
tagged December 2010 in South Carolina, USA. Satellite tags transmitted
locations for 3 months - 3.5 years. Available area is defined as the total
area of coastal, wetland, estuarine, and riverine habitat in South Carolina
where pelicans are commonly seen. Estimated parameters include the
percentage of the total available area that is located within each area of
interest and the mean percentage of each individual pelican’s 95% utility
distribution that overlaps with each area of interest. Compositional
analysis was used to test whether the percent of overlap of pelican home
ranges with each area of interest was proportional to the area’s
availability; each area of interest was ranked from greatest use (1) to least
use (7)................................................................................................................30

viii
List of Tables (Continued)
Table Page
3.1 Foraging parameters of masked boobies nesting at Isla Muertos, Mexico,
May and November 2013. Wilcoxon rank-sum tests were used to compare
characteristics of foraging trips between seasons (May and November; n =
77 birds), sexes (n = 77 birds), and stages (incubating and chick-rearing in
November; n = 41 birds). Watson's Two-Sample Test of Homogeneity was
used to compare circular statistics. Values are presented as mean ± 1
standard deviation. Ranges are in parenthesis. Significant results appear in
bold; alpha = 0.05 .............................................................................................63
3.2 Foraging behavior of masked boobies nesting at Isla Muertos, Mexico
during May and November 2013. Wilcoxon rank-sum tests were used to
compare characteristics of foraging behavior between seasons (May and
November; n = 72 birds), sexes (n = 72 birds), and stages (incubating and
chick-rearing in November; n = 41 birds). Watson's Two-Sample Test of
Homogeneity was used for circular statistics. Values are presented as
mean ± 1 standard deviation. Ranges are in parenthesis. Significant results
appear in bold; alpha = 0.05. ARS: area restricted search ................................66
3.3 Physical oceanographic characteristics considered in random forest
analysis and the hypothesized link with foraging behavior of masked
boobies nesting at Isla Muertos, Mexico, May and November 2013.
Frequency and percent of models represent the number and percent of
individual models (n = 72 birds) in which a particular variable was
considered important for predicting foraging behavior. Temporal scale and
spatial resolution refer to the scale and resolution at which the
environmental variables were measured (see Methods) ...................................69

ix
LIST OF FIGURES
Figure Page
2.1 Number of satellite tags transmitting each month from 4 December 2010
(initial deployment) - 1 July 2013 (final tag ceased transmitting) for 40
brown pelicans satellite tagged in South Carolina, USA ..................................16
2.2 Distribution of 40 brown pelicans captured during the nonbreeding season
near Charleston, SC, USA, satellite-tagged December 2010 and tracked
until July 2013. Kernel density estimates of breeding (orange) and
nonbreeding (blue) distribution. Within each stage, home range (95%
utilization distribution) is light-colored and focal region (50% utilization
distribution) is dark-colored. .............................................................................19
2.3 Movements of four individual brown pelicans satellite tagged December
2010 in South Carolina, USA, illustrating examples of different migration
patterns and non-breeding destinations. For each individual, the location
of the breeding colony used during tracking is marked with an arrow; the
capture location for all birds is marked with a star in the upper left panel.
The breeding colony for bird 291 is unknown because the satellite tag
stopped transmitting before onset of the breeding season. Bird 291 =
highly migratory, destination = Guatemala. Bird 292 = migratory,
destination = southern Florida. Bird 279 = semi-migratory, destination =
Georgia. Bird 298 = stationary, destination = South Carolina ..........................33
2.4 Pre-breeding movements of five brown pelicans satellite tagged December
2010 in South Carolina, USA. Satellite tags transmitted locations for 3
months - 3.5 years. The start of each track is marked by individual bird id
number. Bird 292 arrived at the breeding colony but then dispersed south
before returning to the colony for the breeding season. All other
individuals dispersed north beyond the colony then moved south for the
breeding season .................................................................................................25

x
List of Figures (Continued)
Figure Page
2.5 Distribution of brown pelicans satellite tagged December 2010 in South
Carolina, USA, in relation to biogeography (marine provinces which are
subdivided into ecoregions) as described by Spalding et al. (2007).
Satellite tags transmitted locations for 3 months - 3.5 years. Identifiers of
ecoregions within the range of all satellite locations are as follows: 41
Virginian, 42 Carolinian, 70 Floridian, 63 Bahamian, 65 Greater Antilles,
67 Southwestern Caribbean ..............................................................................27
2.6 Generalized additive mixed models were used to examine trends in annual
occupancy of marine ecoregions described by Spalding et al. (2007) for
brown pelicans satellite tagged December 2010 in South Carolina, USA.
Satellite tags transmitted locations for 3 months - 3.5 years. Dark lines
represent model predictions and translucent areas represent 95%
confidence intervals. Probability of occupancy was estimated separately
for three frequently-used ecoregions described by Spalding et al. (2007) as
follows: a) Virginian, b) Floridian ....................................................................28
2.7 Generalized additive mixed models were used to examine trends in annual
occupancy of the Chesapeake Bay/Delmarva Peninsula region for brown
pelicans satellite tagged December 2010 in South Carolina, USA. Satellite
tags transmitted locations for 3 months - 3.5 years. Dark grey line
represents model predictions and light-gray areas represent 95%
confidence intervals. Probability of occupancy was highest from July -
October, corresponding to post-breeding and molt periods ..............................30
2.8 Coastal parks and protected areas (dark gray) in South Carolina, USA, that
encompass marine, estuarine, swamp, or riverine habitat potentially
occupied by brown pelicans 2010 - 2013. Protected areas that overlapped
with a relatively high proportion of individual pelican home ranges are
labeled and highlighted. NERR = National Estuarine Research Reserve,
NWR = National Wildlife Refuge ....................................................................32

xi
List of Figures (Continued)
Figure Page
3.1 Location of Isla Muertos, the largest breeding colony for masked boobies
in the north Atlantic region, in relation to the boundary of Arrecife
Alacranes National Park and the prominent bathymetry of the Campeche
Bank, Mexico. At-sea foraging movements of 77 breeding masked
boobies were recorded with GPS loggers during May and November 2013 ....47
3.2 Sample analysis of a GPS track illustrating delineation of locations that
correspond to area restricted search (ARS) behavior. Scale bar in (a) also
applies to panels (b) and (d). (a) A portion of a GPS track (line) with
individual locations (points). (b) For each location along the track, the
amount of time required to cross a circle of a given radius (in this case
radius = 2 km) is calculated (the first passage time; FPT). Red arrows
indicate direction of travel. The circle on the left contains an example of
direct movement at a moderate speed, resulting in short FPT. In contrast,
the circle on the right contains an example of tortuous movement at slow
speed, resulting in long FPT and potentially corresponding to ARS
behavior. (c) FPT analysis is repeated multiple times, using circles with
incrementally increasing radii (x-axis). For each radius, the variance
log(FPT) of the calculated FPT values is plotted and those points are
connected via an interpolated line for optimal visualization. The radius
that corresponds to the peak log(variance) is the dominant scale of ARS.
In this example, maximum variance log(FPT) corresponds to a search
radius of 1980 meters. (d) Once the scale of ARS is identified, the
penalized contrast method of Lavielle is used to cut the track into
segments consisting of sets of locations with similar FPT values (range 0 -
23 h). Segments with overall high FPT values correspond to ARS behavior
(blue points); segments with low FPT values correspond to non-ARS
behavior (gray points) .......................................................................................51

xii
List of Figures (Continued)
Figure Page
3.3 Arrecife Alacranes National Park boundary (black oval) in relation to
locations of small-scale area restricted search behavior (colored points)
identified from first passage time analysis. Foraging trips (gray lines; n =
129; multiple trips per bird are pictured) were recorded from masked
boobies nesting at Isla Muertos (black star), Mexico, May and November
2013...................................................................................................................58
3.4 An example of multiple scales of area restricted search (ARS) behavior
during a single foraging trip recorded for a masked booby nesting at Isla
Muertos (star), Mexico, May 2013. Multiple scales of ARS were identified
from the variance log(first passage time) plot (inset box) which has four
prominent local maxima, each indicated by a dotted line. Within the track,
the locations which correspond to foraging at each scale (large map;
delineated by colors that correspond with the dotted lines on the plot) are
nested from largest to smallest. Points indicate interpolated GPS locations,
circles indicate search radii, arrows indicate direction of travel .......................60
3.5 Distribution of azimuth from the colony during foraging trips of masked
boobies nesting at Isla Muertos, Mexico, May and November 2013.
Departure and arrival: angle between the colony and the location of the
bird when it was two km from the colony. ARS: angle between the colony
and locations of area restricted search. Numbers and dashed circles inside
each plot correspond to the number of individuals included in each bar. n
= 77 birds ..........................................................................................................62

xiii
List of Figures (Continued)
Figure Page
3.6 Generalized additive mixed models (GAMM) were used to examine the
relationship between area restricted search (ARS) behavior and physical
oceanography in foraging masked boobies nesting at Isla Muertos,
Mexico, May and November 2013. Different groups of individuals that
showed similar trends for a particular environmental variable of interest
were used to build each working model. N for each curve is listed near the
y axis of each plot, and the total number of birds that showed a strong
relationship for a particular environmental variable (as determined by
random forest analysis) and could have potentially contributed to GAMM
formation was sea surface height anomaly = 32 and velocity of water = 27.
The colored line represents model predictions and light-gray areas
represent 95% confidence intervals ..................................................................70

xiv
LIST OF APPENDICES
Appendix Page
A Individual satellite-tag functionality and latitudinal movements for brown
pelicans satellite tagged in South Carolina, USA, 2010 - 2013. Raw data
were cleaned by retaining only locations with accuracies ≤ 1.5 km and
applying a speed filter of 25 m/s .......................................................................84
B Individual breeding locations, focal region, and home ranges (in km2) for
brown pelicans satellite tagged in South Carolina, USA, 2010 - 2013.
Mean percent overlap between successive breeding or successive
nonbreeding seasons was calculated for each individual. OHCI = Old
House Channel Island. A hyphen (-) indicates that the satellite tag stopped
transmitting before enough locations could be collected to estimate the
parameter of interest .........................................................................................86
C Description of individual migration strategies and dispersal locations for
brown pelicans satellite tagged in South Carolina, USA, 2010 - 2013.
Birds used six distinct ecoregions. Proportions were calculated using the
range-wide 95% UD for each individual. Only parks and protected areas
up to 50 km inland containing marine, estuarine, swamp, or riverine
habitat were considered. A hyphen (-) indicates that the satellite tag
stopped transmitting before enough locations could be collected to
estimate the parameter of interest. NA indicates that a dispersal location
was not identified ..............................................................................................88

1
CHAPTER ONE
INTRODUCTION
Habitat use, the process in which an animal occupies an area (Johnson 1980),
influences individual behavior, body condition, mortality risk, and reproductive success
in the short-term (Cody 1985, Stephens 1986, Aubry et al. 2013). In the long term, habitat
use may impact survival, fitness, and population dynamics (Cody 1985, Kristan Iii 2003,
Pärt et al. 2011, DeCesare et al. 2013). Selective decisions that determine habitat use
occur dynamically as an animal attempts to achieve an optimal balance between
numerous interacting factors (Stephens 1986). During foraging for instance, mechanisms
that drive habitat use may include, but are not limited to, food availability (Suryan et al.
2006), physiology (Sommerfeld 2013), reproductive constraints (Preatoni et al. 2011),
risk (Freitas et al. 2008, DeCesare et al. 2013), competition (Wakefield et al. 2013), and
social facilitation (Weimerskirch et al. 2010). Adding complexity, memory and learning
may contribute to selective decisions at multiple timescales (Hutto 1985, Fronhofer et al.
2013, Janmaat et al. 2013), and an animal may repeat trips to desirable habitat in
subsequent hours, days, or years.
In marine habitat, prey are often patchily distributed in space and time within a
heterogeneous matrix (Clarke 1977, Ricklefs 1990), and the mechanisms that influence

2
habitat use by individuals in this environment may be complex and difficult to discern or
quantify. Furthermore, directly observing the behavior of marine vertebrates is costly in
terms of time and resources. The study of marine habitat use, long possible only through
airplane or vessel-based observational surveys, has advanced rapidly due to the advent of
tracking technology (Ainley et al. 2012), allowing scientists to expand the scope of their
research to include both terrestrial and marine habitat use by individuals from known
colonies of origin. Therefore, tracking information linking foraging locations of known
individuals to physiological and environmental factors may provide an enhanced
perspective on the mechanisms that underlie habitat use.
For many seabird species, behaviors that influence reproductive success, adult
survival, and population dynamics occur at sea (Furness and Tasker 2000, Croxall et al.
2002), but the mechanisms that drive at-sea habitat use are not well understood, thus
there is a need for research regarding seabird foraging ecology (Boersma et al. 2001,
Croxall et al. 2012, Lewison et al. 2012). For example, birds may adjust regional foraging
movements in response to dynamic prey distribution (Santora et al. 2009), and decisions
made by individual birds regarding the duration and location of foraging trips may affect
energy expenditure and impact physical condition (Chaurand and Weimerskirch 1994,
Weimerskirch et al. 1997, Pichegru et al. 2007). During breeding, flexible foraging
strategies may help seabirds to simultaneously meet the energetic demands of offspring
and maintain adult body condition (Chaurand and Weimerskirch 1994, Welcker et al.
2009). When prey are scarce, however, competing energetic demands may result in

3
reduced reproductive success as birds trade-off chick condition for self-maintenance
(Hamer et al. 2007, Chivers et al. 2012).
Understanding factors that influence seabird habitat use can help natural resource
managers predict foraging locations and therefore estimate spatial and temporal overlap
between bird activity and threats. For example, foraging seabirds may encounter a range
of threats and are susceptible to mortality or morbidity originating from exposure to
pollutants (Votier et al. 2005), collisions with wind-energy turbines, and entanglement in
abandoned fishing gear (Croxall et al. 1990), as well as altered breeding biology resulting
from fisheries interactions such as reduced availability of key forage fish species (Cury et
al. 2011, Bertrand et al. 2012), and increased availability of potentially low-energy
fisheries discards (James and Stahl 2000, Pichegru et al. 2007). Marine habitat use
models derived from seabird tracking data can describe the movements and behavior of
individual birds at multiple spatial and temporal scales, enabling comparison between
seabird behavior and biological, ecological, or anthropogenic features of interest. At-sea
locations of seabird movements have been used as an indicator of fish activity (Cairns
1992, Weimerskirch 2007, Weimerskirch et al. 2010), a predictor of bird exposure to
pollutants in the marine environment (Croxall et al. 1990, Votier et al. 2005), and a
scientific basis for decisions regarding the design and placement of marine reserves and
protected areas (Le Corre et al. 2012, Ludynia et al. 2012).
My goal was to investigate habitat use as it relates to the at-sea movements of
seabirds. I conducted research on two species of Pelecaniformes and examined 1)
interannual variation in movements of brown pelicans (Pelecanus occidentalis) and 2)

4
short-term habitat use by individual masked boobies (Sula dactylatra) constrained to a
central place. The first data chapter is titled “Variability in Home Range Size, Migration
Strategy, and Protected Area Use of Eastern Brown Pelicans (Pelecanus occidentalis)
Along the US Atlantic Coast”, and the second data chapter is titled “Variability in
Foraging Behavior of Masked Boobies (Sula dactylatra) Breeding at Isla Muertos,
Mexico”.

5
CHAPTER TWO
VARIABILITY IN HOME RANGE SIZE, MIGRATION STRATEGY, AND
PROTECTED AREA USE OF EASTERN BROWN PELICANS
(PELECANUS OCCIDENTALIS) ALONG THE US ATLANTIC COAST
Introduction
Marine spatial planning in the southeast and mid-Atlantic region of the United
States is a complex endeavor requiring negotiation of political, administrative, and
biological factors at the local, regional, and national levels. Interest in development of
offshore wind energy within the region is high, and to date most US states along the
Atlantic coast have initiated assessments of wind availability and potential profitability
(Michel 2013). Ecological information is necessary in order to assess the extent to which
facility construction and operation may alter oceanographic conditions, increase turbidity
and levels of pollutants, and impact wildlife (Fox et al. 2006, Furness et al. 2013). For
example, pelagic and nearshore seabirds may be affected directly from collisions with
offshore turbines or indirectly from changes in the quality of foraging habitat and altered
availability of prey species (Fox et al. 2006, Perrow et al. 2011). Predicting the impact of
construction and operation of an offshore wind energy facility on populations of interest
is facilitated by detailed knowledge of the spatial ecology of individual birds (Fox et al.

6
2006), however, movement patterns of seabirds within the southeast and mid-Atlantic
region are largely unknown and understudied (Jodice et al. 2013), leading to a lack of
information on which to inform marine spatial planning.
Within the southeast and mid-Atlantic region, knowledge of individual
movements in nearshore seabirds has accrued mainly through banding studies (Stefan
2008), but information is often limited to one or two sightings and biased towards areas
with regular human use and hence increased opportunities for band resightings. Band
recoveries do not provide comprehensive information on individual migratory routes or
wintering destinations (Guilford et al. 2009) which represent important habitat as threats,
interactions, and conditions that occur during nonbreeding may affect individual survival,
adult condition, or reproduction (Gill et al. 2001, Sorensen et al. 2009). Modern tracking
studies have the power to provide detailed information on individual movements in
species that range widely, readily cross ecological and political boundaries, and frequent
inaccessible habitat. Tracking data can be combined with other spatial data such as
oceanography, political boundaries, and ship traffic to examine the context within which
individual birds operate at relatively fine temporal scales.
The coastal region of the southeastern United States and Florida supports a
significant proportion of the breeding population of the eastern subspecies of brown
pelican (Pelecanus occidentalis). Brown pelicans declined in the late 1970s and the
species remains vulnerable to oil pollution (Shields 2002). Although delisted federally,
the species maintains various levels of state listing throughout its range (Jodice et al.
2013). The breeding range has expanded during the previous 40 years along the coast of

7
the eastern United States northward to New Jersey and southward to southern Florida
(Shields 2002). Detailed data on annual movements in the region are limited to band
returns from relatively few studies (Mason 1945, Schreiber 1976, Stefan 2008), and the
species is regularly found in nearshore and estuarine habitats, both protected and
unprotected, throughout its range (Shields 2002, Wickliffe and Jodice 2010, King et al.
2013). Large gaps in basic scientific understanding of brown pelican behavior include
lack of information regarding at-sea distribution, and movements within and between
seasons and years (Jodice et al. 2013). Application of data from other regions (e.g. Gulf
of Mexico, Pacific coast) is not ideal because brown pelican behavior may vary widely
between geographic locations as birds respond to fundamental regional differences in
landscape type, prey availability, human disturbance, and oceanographic conditions.
Current management actions that benefit the species occur primarily on colonies within
parks and protected areas, with loafing sites and wintering sites often receiving less
protection.
I examined movements of adult brown pelicans tracked with satellite tags for up
to three years. My objectives were to 1) determine home ranges during breeding and
nonbreeding, 2) examine individual site fidelity and timing of migration between years,
and 3) investigate the extent to which pelicans use parks and protected areas. I predicted
that, at an individual level, brown pelicans would exhibit behavioral differentiation,
resulting in diverse movement strategies among individuals and overall high site fidelity
across seasons within individuals. These data represent the broadest and longest term
assessment of movements to date for the species and as such can be used to inform

8
marine spatial planning and management decision-making along the east coast of the
Atlantic US.
Methods
Deployment of satellite tags
The study was initiated as a component of the Natural Resources Damage
Assessment (NRDA) to the Deepwater Horizon oil spill (Evers et al. 2011). Brown
pelicans along the coast of South Carolina were targeted for deployment of satellite tags
to act as a comparison and reference for birds tagged in the northern Gulf of Mexico. All
birds were captured at coastal sites in South Carolina, USA, within 20 km of Charleston
Harbor. The area is comprised primarily of tidal creeks, estuaries, salt marshes, and the
Atlantic Intracoastal Waterway, which is a navigable channel ca. 100 -300 m wide.
Between December 2010 and August 2013, battery powered satellite platform
terminal transmitters (PTTs; North Star Science and Technology) tracked via Argos
satellites at 401.65 MHz were used to monitor the movements of 40 individual brown
pelicans. From 4 - 19 December, 2010, nonbreeding adult pelicans were captured
opportunistically from the water with a hand-held net gun using bait fish as an attractant,
or alternatively, from loafing areas on wooden pilings using a padded leghold trap (Evers
et al. 2011). Twenty-five PTT assemblies weighing 110 grams and 15 PTT assemblies
weighing 68 grams (corresponding respectively to 3.7% and 2.3% of the mass of the
lightest adults) were attached to individual birds (n.b., permission was given by the

9
USGS Bird Banding lab for attachments to weigh ≤ 5% of body mass). All PTTs were
programmed to transmit for eight hours on and 16 hours off for the first 90 days post-
capture and then eight hours on and 137 hours (approximately six days) off until the
batteries were depleted, for a predicted longevity of 40 months for 110 g units and 22
months for 68 g units (Evers et al. 2011). PTTs were attached to the back using a harness
constructed from Teflon tubing and secured for fit with copper crimps (Evers et al. 2011).
During initial capture, adults were weighed, measured for culmen, and examined visually
for external oil; no birds showed signs of external or visible oiling. Birds were released at
the capture site after approximately 45 minutes.
Data processing
Analyses of data were performed using R (R Development Core Team 2014)
unless otherwise specified. Locations were archived daily by Argos and downloaded
remotely as individual files. Data were subsequently extracted from files (package trip;
Sumner 2013), resulting in 28088 locations from all deployed tags. Positional accuracy of
each individual location depends on the number of messages transmitted from the PTT to
the passing satellite and is provided by Argos based on seven levels. Location classes 3,
2, 1, and 0 were collected using at least four messages and have an estimated radius-of-
error of < 250, < 500, <1500, and > 1500 meters respectively. Location classes A (three
messages), B (two messages), and Z (location process failed) have no error estimates. I
discarded duplicate locations and locations of low or indeterminate accuracy, retaining
17835 locations of class 3, 2, and 1 (sensu Davis et al. 2001, Pütz et al. 2007). To remove

10
locations that represented unrealistic movements, I applied a speed filter of 25 m/s,
corresponding to moderate flight speed for brown pelicans (Schnell and Hellack 1978;
17050 locations retained). Tracks for individual birds were cut into distinct segments if
the time between transmission of two locations exceeded 19 days, an event that occurred
infrequently but resulted in large data gaps. Segments with less than 5 locations could not
be used in analysis and were discarded (17042 locations retained). For each individual
bird, I classified locations into annual stages (e.g. breeding, non-breeding, migration)
based on a combination of movement patterns and time of year. Breeding activity was not
confirmed through direct observation, therefore, to determine breeding activity and hence
breeding locations I used a combination of timing (April 1-Sept 30 was considered
breeding; Schreiber 1980, Shields 2002) and range restriction of less than 200 km from a
central place (many species of seabirds have a contracted range during the breeding
season as they are constrained to the breeding colony or nest site). All averages are
specified as mean ± standard deviation.
Home range estimation
For use in individual-based analysis, I calculated 2-dimensional Gaussian kernels
for each bird with location data that spanned > 300 days. I extracted 50% and 95% UD
contours separately for each breeding and nonbreeding stage using package
adehabitatHR (Calenge 2006) with an ad-hoc bandwidth selection (Wand and Jones
1994, Calenge 2011b). To assess site fidelity between years within individuals, I
calculated mean percent overlap of 95% UD contours between successive breeding

11
seasons and successive nonbreeding seasons. To test for seasonal differences in the
distribution of the areal extent of the focal region, home range, and home range overlap I
used Wilcoxon rank-sum tests and Kruskal Wallis tests.
To determine a combined distribution for all study birds, I calculated 2-
dimensional Gaussian kernels using the plug-in bandwidth matrix on a 0.005° grid
(package ks; Duong 2007). Individual PTTs recorded variable numbers of locations over
different time-spans within and between individuals. To evenly weight locations
collected from 40 individuals, during two stages (breeding/nonbreeding), and six seasons
(breeding 2011, 2012, 2013; nonbreeding 2010/2011, 2011/2012, 2012/2013), I divided
the data into groups of individuals and seasons with similar numbers of locations. I tallied
the number of locations collected for each season by individual (six seasons per
individual were considered). I examined the distribution of the number of locations by
individual*season combination separately for breeding and nonbreeding stages. Within
each stage, I delineated groups of individuals and seasons with similar numbers of
locations (breeding season, two groups; nonbreeding season, three groups), and
calculated separate kernel densities for each group. Each group included > 1000
locations, ensuring robust kernel estimation. I extracted the utilization distribution (UD)
contours of the 95% (home range) and 50% (focal region; as defined in Hyrenbach et al.
2002) areas for each group and merged contours together by stage to produce separate
breeding and nonbreeding distributions.

12
Migration strategy
To coarsely quantify the extent of movements that occurred during the
nonbreeding season, I calculated the span of latitude covered for each individual by
subtracting the lowest recorded latitude from the highest recorded latitude. To group
individuals into distinct migration strategies I used Ward’s method of hierarchical
clustering (Ward 1963) with Euclidean distance and a k-value = 4 (indicating the number
of means or clusters to extract). Ward’s method merges similar values, one at a time, and
minimizes the total variance within a cluster. I tested for differences between clusters
using multi-scale bootstrap resampling (package pvclust; Suzuki and Shimodaira 2006),
which assigns a p-value to clusters supported by data that are tightly grouped.
Habitat use
I used generalized additive mixed models (GAMM) to investigate timing of
occupancy for three specific ecoregions that were most frequently used by brown
pelicans during this study (Virginian, Carolinian, and Floridian; Spalding et al. 2007),
and the Chesapeake Bay/Delmarva Peninsula (CBDP) area of the mid-Atlantic coast.
Shapefiles for marine ecoregions of the world (MEOW) were downloaded from
www.marineregions.org (Claus et al. 2014), imported into ArcGIS (ESRI 2014), and
overlaid on brown pelican locations. The ecoregion corresponding to each location was
extracted using the tool near. Locations within the CBDP were identified using the tool
select. A binomial GAMM (package gamm4; Wood and Scheipl 2013) was applied
separately for each of the four areas. The binomial dependent variable indicated whether

13
or not pelicans were located within an area for each location. Day of year was the
explanatory variable, and individual was included as a random effect. Cyclic penalized
cubic regression splines were specified as the smooth and are appropriate for periodic
data, including daily or annual patterns (Wood 2006).
Reserve use
A high percentage of all locations were recorded in South Carolina (58%),
compared to other locations (Florida, 20%; North Carolina, 8%; Georgia, 6%; Virginia,
5%, Maryland, 2%, Outside US, 1%), thus I used compositional analysis to examine
whether brown pelican use of protected areas in South Carolina was proportional to area
size, adopting methods described in Adams et al. (2012). Compositional analysis
compares used to available habitat and is based on assumptions that individual animal
ranges are independent and that all animals have equal access to all areas of interest
(Aebischer et al. 1993). “Available area,” defined as coastal, wetland, estuarine, and
riverine habitat in South Carolina located <50 km inland and <20 km offshore (the
maximum inland and offshore distances recorded for brown pelicans in this study) was
delineated from satellite imagery and corroborated by assessing locations where pelicans
were commonly seen from 2004-2014 (eBird 2012). Protected area shapefiles were
downloaded from www.protectedplanet.net (IUCN and UNEP-WCMC 2014), imported
into ArcGIS, and overlaid on 95% UD contours for individual birds. All data were
transformed to two-dimensional Cartesian coordinates using a Robinson projection. I
calculated the proportion of each 95% UD area that overlapped with undesignated areas

14
and each protected area located within the available area. I used a subset of six protected
areas that had the highest mean proportion of overlap with 95% UDs in subsequent
analyses. To determine whether brown pelicans were distributed disproportionately
within any given protected area, I compared the relative proportion of each individual’s
95% UD area that overlapped each area of interest using a randomization test (R package
AdehabitatHS, function compana; Calenge 2006). The test was conducted with 500
replicates and an alpha level of 0.05; values of zero were replaced with a default value of
0.01.
Results
Tag functionality
Tags reported locations from 4 December, 2010 - 1 July, 2013. Duration of PTT
activity was 357 ± 303 days per tag from deployment to cease-function resulting in 426 ±
280 locations per tag (Fig. 2.1; Appendix A). Of the 40 tags deployed, 88% were still
transmitting one month post-deployment, 66% three months post-deployment, 50% one
year post-deployment, and 20% two years post deployment. Tags weighing 110 grams
collected significantly more locations (525 ± 292) than 68-gram tags (261 ± 163;
Wilcoxon rank-sum test, W = 283, P = 0.007), but did not necessarily transmit over a
longer time period (pooled mean 358 ± 303 days; Wilcoxon rank-sum test, W = 238, P =
0.16) despite being programmed to follow the same duty cycle. Duty cycling was daily

15
through the 2010/2011 nonbreeding season and switched to weekly coincident with the
onset of breeding in 2011.

16
Figure 2.1 Number of satellite tags
transmitting each month from 4
December 2010 (initial deployment) -
1 July 2013 (final tag ceased
transmitting) for 40 brown pelicans
satellite tagged in South Carolina,
USA.

17
Distribution and home ranges
Throughout the annual cycle, brown pelicans primarily used coastal areas along
the Atlantic coast from Maryland to south Florida and along the Gulf coast in southwest
Florida (Fig. 2.2). Birds used coastal habitat almost exclusively throughout the annual
cycle. The few locations recorded inland (n < 613) occurred near large rivers or lakes
(e.g. St. Johns River in northeast Florida; Lake Okeechobee in south Florida). Marine
locations typically occurred within 10 km of land (98%), though offshore distances of up
to 80 km (n = 60) were recorded (e.g. west of Charleston, South Carolina; between
western Cuba and the northeast Yucatán peninsula). The distribution of brown pelicans
during nonbreeding was diffuse compared to breeding and covered a wider range of
latitudes with a greater number of use areas (Fig. 2.2).
Satellite tags were operational during at least one breeding season for 26 of the 40
individuals. Breeding locations included two colonies in South Carolina (Crab Bank =
22%, Marsh Island = 19%), four colonies in North Carolina (Beacon Island, Pelican
Island, and Old House Channel Island = 11% each, New Dump Island = 7%), two
colonies in Virginia (Wreck Island = 7%, Fisherman Island = 4%), and one colony in
Maryland (Smith Island = 4%). Based on movement data as defined above, 4% of birds
with operational tags during the breeding season did not appear to breed. The areal extent
of individual home ranges was similar for breeding and nonbreeding (Table 2.1),
however, the focal region was significantly smaller during nonbreeding (Table 2.1).
For birds with tags that transmitted for multiple years (n = 21) mean home range
size was consistent between years within seasons (Kruskal Wallis test, χ2 = 3.7, df = 5,

18
P= 0.6). Individual site fidelity was high between years, especially considering the small
home range size yet large latitudinal distribution of birds, and only one individual
switched breeding colonies in successive years from Old House Channel Island in North
Carolina to Wreck Island in Virginia, a distance of 165 km. During breeding, home range
overlap within individuals between years was > 0.3 (1 = complete overlap, 0 = no
overlap) for 90% of individuals and > 0.6 for 40% of individuals (Appendix B).
Nonbreeding home range overlap was also high: > 0.3 for 81% of individuals and > 0.6
for 38% of individuals.

19
Figure 2.2 Distribution of 40 brown pelicans captured during the nonbreeding season
near Charleston, SC, USA, satellite-tagged December 2010 and tracked until July 2013.
Kernel density estimates of breeding (orange) and nonbreeding (blue) distribution.
Within each stage, home range (95% utilization distribution) is light-colored and focal
region (50% utilization distribution) is dark-colored.

20
Table 2.1 Areal extent (mean ± sd km2) of focal region
(50% utilization distribution, UD), home range (95%
UD), and percent overlap in successive years (an
estimate of site fidelity) during breeding and
nonbreeding seasons for brown pelicans satellite tagged
December 2010 in South Carolina, USA, and tracked
until July 2013. Home range and focal region were log-
transformed. n = number of birds, W = Wilcoxon rank-
sum test, Significant results appear in bold; alpha = 0.1.
n
Breed
ing
mean
± sd
n
No
n-b
reedin
g
mean
± sd
W v
alue
P-v
alue
Fo
cal regio
n (5
0U
D; k
m2)
25
62
6.4
± 4
33
.0
40
45
3.2
± 2
29
.9
63
5
0.0
7
Ho
me ran
ge (9
5U
D; k
m2)
25
34
92
.0 ±
22
53
.2
40
28
47
.9 ±
17
18
.8
57
9
0.2
9
Overlap
in su
ccessive y
ears (%)
10
52
.8 ±
23.3
2
1
55
.0 ±
21.3
9
8
0.7
9

21
Migration strategy
Brown pelicans demonstrated substantial individual variability in migration
movement patterns (Appendix C). Patterns were classified into one of four broad
strategies based on the difference in latitude between the northernmost and southernmost
locations for each individual (Fig. 2.3; Table 2.2). Of the 28 birds tracked for at least 4
months, stationary individuals (25% of birds) dispersed little from the capture location
over subsequent seasons or years, and latitudinal extent ranged from 15 - 362 km. In the
remaining classifications, latitudinal extent ranged from 525 - 792 km for semi-migratory
individuals (29% of birds), 894 - 1103 km for migratory individuals (35% of birds), and
1443 - 1831 km for highly migratory individuals (6% of birds). Individuals generally
adopted a single movement strategy and used it consistently over multiple years. Birds
that switched strategies between years (33%, n = 21 birds with multiple years of data)
made small strategic shifts, i.e. from stationary to semi-migratory or from migratory to
semi-migratory. Breeding home range was significantly smaller for stationary individuals
(1983 ± 1189 km2) compared to two other categories of migratory individuals (sample
size for highly migratory birds was not large enough for statistical testing; pooled mean
for semi-migratory and migratory birds, 4619 ± 2373 km2; Kruskal Wallis test, χ
2 = 11.1,
df = 2, P = 0.004).

22
Figure 2.3 Movements of four individual brown pelicans satellite tagged December 2010
in South Carolina, USA, illustrating examples of different migration patterns and non-
breeding destinations. For each individual, the location of the breeding colony used
during tracking is marked with an arrow; the capture location for all birds is marked with
a star in the upper left panel. The breeding colony for bird 291 is unknown because the
satellite tag stopped transmitting before onset of the breeding season. Bird 291 = highly
migratory, destination = Guatemala. Bird 292 = migratory, destination = southern
Florida. Bird 279 = semi-migratory, destination = Georgia. Bird 298 = stationary,
destination = South Carolina.

23
Table 2.2 Discrete migration strategies of brown pelicans satellite tagged in South
Carolina, December 2010. Satellite tags transmitted locations for 3 months - 3.5 years.
Four groupings were identified for birds with at least four months of location data and
strategies were classified as highly migratory, migratory, semi-migratory, and stationary
based on Ward hierarchical clustering analysis of the latitudinal range for each
individual. At the 0.05 significance level, stationary individuals were significantly
different from all other groups.
Migration strategy n
Latitudinal extent (km)
mean ± sd
Range of latitude (deg) Cluster
Highly migratory 2 1637.2 ± 274.3 15.6 - 38.1 a
Migratory 11 1008.1 ± 80.8 25.9 - 38.3 a
Semi-migratory 9 669.5 ± 25.8 25.8 - 38.4 a
Stationary 7 193.1 ± 128.1 30.3 - 35.9 b

24
Timing of dispersal from the breeding colony and migratory movements varied
among individuals (Fig. 2.3). Five areas were regularly used by multiple birds during
migratory movements including Cape Lookout, North Carolina, Charleston harbor, South
Carolina, Doboy Sound, Georgia, St. John’s River, Florida, and Cape Canaveral, Florida.
Birds moved gradually southward along the coast postbreeding, spending time in multiple
locations before arriving at a winter destination. For example, one individual migrated
from the capture location and spent 12 days in Cape Canaveral, Florida, but wintered
south of the US, traveling for ten days from Florida to coastal Guatemala (Fig. 2.3).
During migration, some individuals appeared to cross large areas of land including
continental Cuba (moving from NNE to WSW, 170 km over land) and Florida (moving
from west to east, > 200 km over land). Birds initiated movement northward towards the
breeding grounds between mid-February and mid-April (median date = 24 March, n =
41), arriving between mid-March and late May (median date = 7 April, n = 36). Five
individuals initiated a pre-breeding dispersal in late February to early April; dispersing
birds reached the breeding colony, continued to a different location and remained there
for 18-31 days, then returned to the colony (Fig. 2.4). Birds departed the breeding colony
between mid-June and late October (median date = 25 Sept, n = 27). Seven individuals
dispersed north post-breeding.

25
Figure 2.4 Pre-breeding movements of five brown pelicans satellite tagged December
2010 in South Carolina, USA. Satellite tags transmitted locations for 3 months - 3.5
years. The start of each track is marked by individual bird id number. Bird 292 arrived at
the breeding colony but then dispersed south before returning to the colony for the
breeding season. All other individuals dispersed north beyond the colony then moved
south for the breeding season.

26
Habitat use
Brown pelicans occupied six ecoregions which occur within three marine
provinces (Cold Temperate Northwest Atlantic, Warm Temperate Northwest Atlantic,
and Tropical Northwestern Atlantic; Fig. 2.5). The majority of locations occurred within
three of the six ecoregions (Fig. 2.5): Virginian (41; 12% of locations), Carolinian (42;
71% of locations), and Floridian (70; 15% of locations). One individual migrated through
two additional ecoregions, Bahamian (63; 0.02% of locations) and Greater Antilles (65;
0.05% of locations), and spent the rest of the nonbreeding season in the Southwest
Caribbean ecoregion (67; 1.3% of locations). The probability of brown pelicans occurring
in the three most commonly used ecoregions was nonlinear with respect to time of year
(Fig. 2.6). Birds were likely to use cold temperate waters from April - December, warm
temperate waters year-round, and tropical waters December - April.
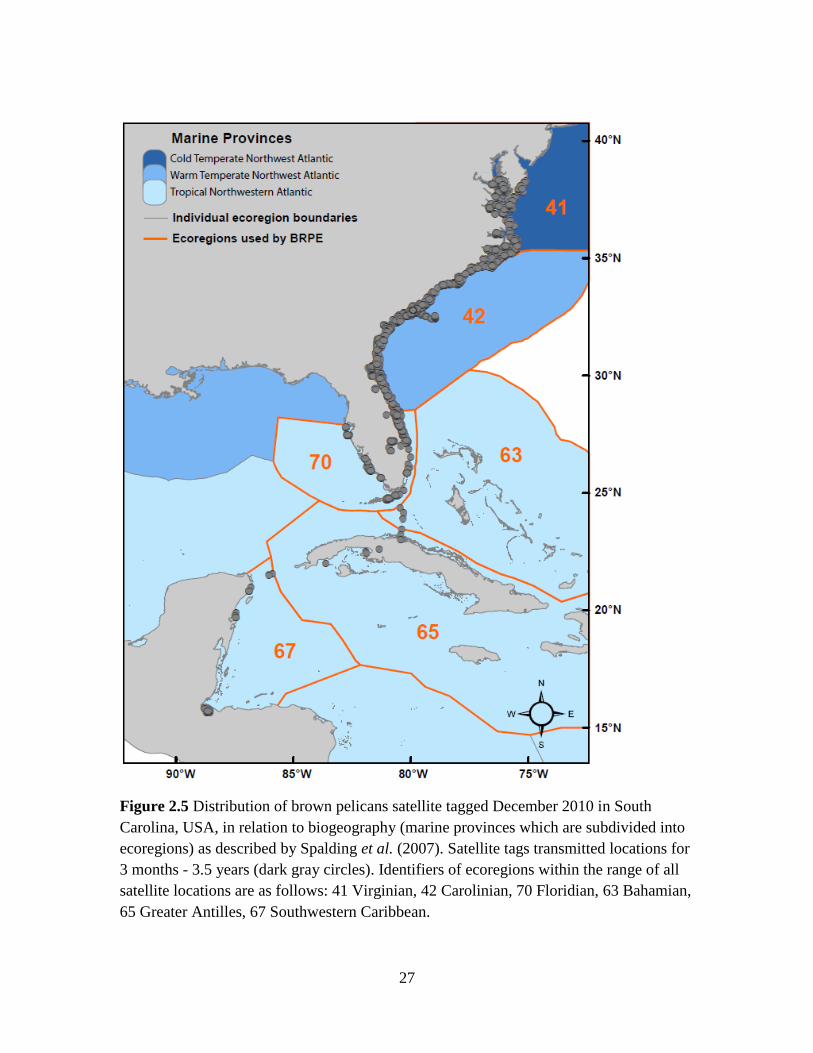
27
Figure 2.5 Distribution of brown pelicans satellite tagged December 2010 in South
Carolina, USA, in relation to biogeography (marine provinces which are subdivided into
ecoregions) as described by Spalding et al. (2007). Satellite tags transmitted locations for
3 months - 3.5 years (dark gray circles). Identifiers of ecoregions within the range of all
satellite locations are as follows: 41 Virginian, 42 Carolinian, 70 Floridian, 63 Bahamian,
65 Greater Antilles, 67 Southwestern Caribbean.

28
Figure 2.6 Generalized additive mixed models were used to examine trends in annual
occupancy of marine ecoregions described by Spalding et al. (2007) for brown pelicans
satellite tagged December 2010 in South Carolina, USA. Satellite tags transmitted
locations for 3 months - 3.5 years. Dark lines represent model predictions and translucent
areas represent 95% confidence intervals. Probability of occupancy was estimated
separately for two frequently-used ecoregions described by Spalding et al. (2007) as
follows: a) Virginian, b) Floridian.

29
The probability of brown pelicans occurring in the CBDP region of Virginia and
Maryland was nonlinear with respect to time of year (Fig. 2.7). Brown pelicans that used
the CBDP region were more likely to do so during post-breeding from late June -
November than other times of the year. Probability of occupancy of the CBDP region
during breeding in May - late June, was highly variable.

30
Figure 2.7 Generalized additive mixed models were used to examine trends in annual
occupancy of the Chesapeake Bay/Delmarva Peninsula region for brown pelicans satellite
tagged December 2010 in South Carolina, USA. Satellite tags transmitted locations for 3
months - 3.5 years. Dark grey line represents model predictions and light-gray areas
represent 95% confidence intervals. Probability of occupancy was highest from July -
October, corresponding to post-breeding and molt periods.

31
Reserve use
Within South Carolina, use of protected areas by brown pelicans varied among
individuals (Appendix C). The mean percent area of individual home ranges within South
Carolina that overlapped spatially with protected areas in the state was 15.0 ± 9.5%. Four
of 20 brown pelicans had >20% overlap with protected areas. Two birds visited all five of
the protected areas within the state considered in compositional analysis (birds had 21%
and 24% overlap). The proportion of an individual’s home range that overlapped
protected areas was not influenced by migration strategy (Kruskal Wallis test, χ2 = 2.6, df
= 2, P = 0.3).
Brown pelicans did not visit all reserves within the available area; 19 of the 61
protected areas considered (31%) did not overlap brown pelican home ranges, 36
protected areas (59%) had a mean overlap < 0.5%; six protected areas had a mean overlap
≥ 0.5% (Fig. 2.8). The two protected areas that were largest in size, Ashepoo-Combahee-
Edisto Basin National Estuarine Research Reserve (ACE) Basin (525 km2) and Cape
Romain National Wildlife Refuge (CRNWR, 228 km2; Table 2.3), appeared to greatly
influence the observed relationships. Home range overlap > 10% was observed for seven
individuals that used the ACE Basin and for six individuals that used CRNWR.

32
Figure 2.8 Coastal parks and protected areas (dark gray) in South Carolina, USA, that
encompass marine, estuarine, swamp, or riverine habitat potentially occupied by brown
pelicans 2010 - 2013. Protected areas that overlapped with a relatively high proportion of
individual pelican home ranges are labeled and highlighted. NERR = National Estuarine
Research Reserve, NWR = National Wildlife Refuge.

33
Table 2.3 Proportional overlap of
undesignated habitat and six protected
areas in South Carolina with individual
home ranges of 20 brown pelicans satellite
tagged December 2010 in South Carolina,
USA. Satellite tags transmitted locations
for 3 months - 3.5 years. Available area is
defined as the total area of coastal,
wetland, estuarine, and riverine habitat in
South Carolina where pelicans are
commonly seen. Estimated parameters
include the percentage of the total
available area that is located within each
area of interest and the mean percentage of
each individual pelican’s 95% utility
distribution that overlaps with each area of
interest. Compositional analysis was used
to test whether the percent of overlap of
pelican home ranges with each area of
interest was proportional to the area’s
availability; each area of interest was
ranked from greatest use (1) to least use
(7).

34
Among the five protected areas used in compositional analysis, brown pelican
home ranges overlapped most with the ACE Basin (ca. 6% mean overlap) and least with
Capers Island Heritage Preserve (ca. 0.5% mean overlap). However, birds utilized the
undesignated area (all area that is not protected) and Capers Island Heritage Preserve in
greater proportions relative to the availability of these areas within brown pelican habitat
in coastal South Carolina, (compositional analysis, Wilkes Lambda = 0.22, P = 0.002;
Table 2.3). Undesignated areas ranked highest according to utilization vs. available area,
followed by Capers Island Heritage Preserve. Differences among the remaining
sanctuaries were small, and rankings are likely to be less resilient.
Discussion
Distribution and home ranges
This is the first study to quantify brown pelican home ranges along the Atlantic
coast of the US. The results of this study suggest that although some brown pelicans
demonstrate high fidelity to a specific site year-round, many birds range widely along the
Atlantic coast and Florida Gulf coast throughout the year, and some move beyond US
national waters to tropical ecoregions. The overall distribution of satellite-tagged brown
pelicans captured along the central coast of South Carolina revealed several regions of
concentrated use including estuarine and marine habitat near Charleston, South Carolina,
Cape Canaveral, Florida, and three areas north of the capture location near Cape Fear,
North Carolina, Pamlico Sound, North Carolina, and the CBDP in Virginia and

35
Maryland. Stefan (2008) also documented a high incidence of band recoveries near
Charleston and Cape Canaveral from chicks banded in South Carolina between 1931 and
2005, suggesting both sites have been consistently important for pelicans for several
decades. Satellite tag data revealed that birds captured in South Carolina also wintered at
distant locations including Guatemala and along the east coast of the Yucatán peninsula.
Satellite-tag locations in coastal Georgia occurred more commonly during nonbreeding
than band encounters reported by Stefan (2008).
Home range sizes reported during this study suggest that the foraging range of
brown pelicans during the breeding season may be greater than 20 - 30 km, the distance
originally estimated for the species (Briggs et al. 1983, Wickliffe and Jodice 2010). For
example, individuals during this study regularly were located 20 - 150 km from the
colony. Other species of Pelecaniformes including boobies (Weimerskirch et al. 2009b,
Kappes et al. 2011) and spoonbills (El-Hacen et al. 2013) forage at similar distances from
colonies. Home range size within individual brown pelicans was similar between
breeding and nonbreeding, which suggests that birds may have adjusted to site-based or
seasonal differences in prey availability by spending more time searching a given area
rather than expanding their radius of travel. The extent of the focal region was larger
during breeding compared to nonbreeding, which could have been due to intraspecific
competition with other breeders for prey or increased demand for food from chicks.

36
Migration
Timing and distance of migration varied among individuals, and some birds
remained near the breeding colony year round. In other bird species, competition for
early arrival on breeding territories, combined with individual quality leads to diverse
migration strategies such as those I observed (Myers 1981, Kokko 1999). Stationary birds
that remain near the breeding colony year-round may gain a competitive advantage by
conserving energy that might otherwise be spent traveling to a nonbreeding location, and
they may procure higher quality breeding territories before migratory birds arrive at the
colony. For example, in Cory’s shearwaters (Calonectris diomedea), resident birds
occupied nesting burrows on the breeding grounds during the nonbreeding season and
were more likely attempt breeding compared to long-distance migrants (Pérez et al.
2013). Theoretically, stationary individuals that remain near the breeding colony during
nonbreeding may be more efficient at locating resources during breeding and therefore
also may have smaller home ranges compared to migratory individuals (Van Moorter et
al. 2009). In seabird species with partial residency, competition for nesting habitat is sex-
based and occurs most often among males (Pérez et al. 2013). Partial residency on the
breeding grounds has been observed in brown pelicans (King et al. 2013), and male
brown pelicans invest more effort in nest site selection and territorial defense compared
to females (Shields 2002). In this study, breeding home range was significantly smaller
for stationary birds compared to migratory birds, although information on the sex of
tagged individuals was not available. Whether reproductive success of stationary birds
differs from migratory birds is unknown.

37
Breeding site fidelity appeared high during this study and also appears to be high
in other species of seabirds and coastal waterbirds (Aebischer 1995, Cézilly et al. 2000).
High fidelity to breeding sites within individuals may conserve energy that birds would
otherwise invest searching for new colonies (Switzer 1993), competing for space or
resources (Kokko et al. 2004), and locating prey (Irons 1998). Individual fidelity to
wintering sites has also been observed in some seabird species (Croxall et al. 2005,
Phillips et al. 2005, Hatch et al. 2010), but a high degree of plasticity has been observed
in others (Croxall et al. 2005, Dias et al. 2010). In this study, within individuals, most
birds returned to the same nonbreeding sites in successive years. However, the Atlantic
coast is a dynamic system and islands or other coastal habitat used by brown pelicans
may be unstable over long periods of time, requiring birds to shift among sites
occasionally (Jodice et al. 2007). Development of coastal habitat may further affect site
availability over the long term. Therefore, management of breeding and wintering habitat
for brown pelicans may require more than protection of currently used sites.
Interactions between fisheries and seabirds may broadly impact movement
patterns and home range size as birds adjust the timing and location of foraging
movements to fishing activity (Tew Kai et al. 2013). The commercial shrimp fishery is of
major economic importance in the southeastern US, and ports are regularly located along
the southeast Atlantic coast. Studies from within the region indicate that during the
breeding season, brown pelicans and other seabirds commonly scavenge bycatch
discarded from commercial shrimp trawlers (Wickliffe and Jodice 2010, Jodice et al.
2011). In South Carolina, the energetic value of discarded fish was comparable to natural

38
prey (Jodice et al. 2011), thus birds may benefit by scavenging discards if availability is
predictable and consistent. The commercial shrimp fishery in the southeastern US
Atlantic operates June - January and peaks in intensity September - October, therefore
availability of discards to brown pelicans increases during the postbreeding and early
winter seasons. It is unclear if or to what extent the availability of discards may influence
movement strategies of individual birds, but the timing of southward movements of
satellite tagged individuals does appear to increase coincident with the decline of the
commercial shrimping season in South Carolina (Fig. 2.6).
Approximately 20% of birds in this study undertook a brief exodus from the
colony after arriving there at the end of migration but before initiating breeding. These
trips tended to last 8 - 32 days and reach a maximum distance of 350 - 420 km from the
breeding colony. Similar pre-breeding, post-arrival trips have been documented in
shearwaters (Catry et al. 2009, Guilford et al. 2009), but pre-breeding movements of
species in the family Pelecaniformes are not well understood. It is possible that female
brown pelicans disperse temporarily to locations where they can efficiently build up
reserves and thereby recover or buffer the energetic cost of egg-laying, as observed in
Manx shearwaters (Guilford et al. 2009). Alternatively, brown pelicans may prospect
other colonies and search for territories or mates before the onset of the breeding season,
as has been observed in gannets (Votier et al. 2011), kittiwakes (Cadiou et al. 1994), and
cormorants (Schjorring et al. 1999).
Prospecting in colonial breeding birds is common and may occur throughout the
year, allowing birds to gather information on nest site and resource availability (Harris

39
and Wanless 1989, Péron and Grémillet 2013). In this study, brown pelicans that
migrated were regularly observed near other breeding colonies during nonbreeding and
may have prospected other colonies and foraging areas. Variation in individual migration
strategies, heterogeneity in migratory stopover sites between individuals, and prospecting
behavior could drive range expansion for brown pelicans, including the current
distribution. Schreiber and Schrieber (1982) suggest that colonization of breeding habitat
occurs progressively over seasons and years as loafing areas used during nonbreeding
transition to roosting areas used year-round, and finally to fully realized breeding sites.
Such a process may have occurred for brown pelicans in the CBDP region, leading to a
breeding range expansion from 1978 - present (Shields 2002). Satellite-tagged birds in
this study used the CBDP during prebreeding and postbreeding, and observations of
juveniles in the region during nonbreeding are common (eBird 2012), though a relatively
cold winter climate may restrict brown pelicans from occupying the CBDP region year-
round.
Habitat use
Probability of occupancy of the CBDP for brown pelicans was highest from July -
October, corresponding to postbreeding and molt periods, and 25% of tagged birds from
this study used that area. Brown pelicans undergo an energetically demanding, complete,
postbreeding molt (Shields 2002) in which they replace all of their flight feathers. It is
therefore important that prey quality and availability at molting sites be relatively high.
Extended use of the CBDP region has been observed for diving ducks during winter

40
(Perry et al. 2007), and satellite-tagged common loons (Gavia immer) during
postbreeding migration (Paruk et al. 2014), also suggesting this region may provide
suitable conditions for migratory waterbirds. During this study, pelicans transitioned to
the Floridian ecoregion in December, suggesting that they move south after molt is
mostly complete. The CBDP was used by dispersing birds during prebreeding and
postbreeding, and is also the site of recent breeding range expansion (Brinker et al. 2007).
Active use of the area by top predators such as brown pelicans during important
phenological stages suggests that the quality of the CBDP in terms of nesting habitat or
forage fish availability is high. However, high levels of pesticides, industrial
contaminants, and lead have been recorded in waterbirds collected recently from within
the region (Rattner and McGowan 2007), and brown pelicans that spend time in the
CBDP may also be exposed to pollutants.
Throughout the annual cycle, brown pelicans crossed large marine provinces and
smaller ecoregions, encountering habitat types ranging from cold temperate to tropical
waters. A generalist foraging strategy in which birds opportunistically dive into schools
of fish may enable individuals to access resources in a variety of fundamentally different
habitats such as these ecoregions. Brown pelicans are susceptible to mortality and
morbidity from cold weather events (Shields 2002), and climate could therefore impact
the timing of movements and proportional ecoregion use. Importantly, individual
movement data indicated that although birds were captured in South Carolina, many of
them spent time in other ecoregions where they may have encountered threats not found

41
in the breeding locations including, but not limited to hunting, fisheries, energy
development, or pollution (Jodice and Suryan 2010).
Reserve use
Use of non-protected areas by brown pelicans was relatively high. Compared to
protected areas, unprotected areas are more susceptible to development, and may have
higher risk of predation and levels of human disturbance. Brown pelicans in this study
spent a large amount of time in nearshore areas and therefore outside the boundaries of
existing protected areas, which were primarily terrestrial. Percent of overlap of pelican
home ranges with five of the protected areas of interest was low compared to each area’s
availability. In contrast, Capers Island Heritage Preserve, along the central South
Carolina coast, was used frequently during both breeding and nonbreeding by satellite-
tagged pelicans. Estimates of overlap between individual home ranges and reserve area
could become inflated as a result of nesting activity within a protected area, but breeding
colonies were located at least 16 km outside the boundaries of Capers Island Heritage
Preserve, suggesting that the observed relationship was influenced by habitat features
including undeveloped beach, salt marsh, and brackish water that likely supported
foraging, loafing, or roosting within the protected area. Given the abundance of brown
pelicans as apex predators, and their association with productive nearshore and estuarine
habitats, future marine spatial planning or design of important bird areas could benefit
from inclusion of information regarding brown pelican habitat use.

42
Satellite telemetry greatly improved understanding of the movements of brown
pelicans in nearshore pelagic areas. Results of this study indicate that the offshore
distribution of pelicans, which ranged from 1 - 80 km but was most dense 0 - 10 km
offshore, overlaps generally with areas most likely to support wind or other offshore
energy development in the southeastern US (Jodice et al. 2013). However, in this study,
locations were collected at low temporal resolution and 36% of locations were collected
at night when brown pelicans are roosting on land (Shields 2002). To better assess
potential overlap with wind energy structures, location data could be collected more
frequently and during the day when birds are most likely to be foraging. Doing so would
result in kernel estimates with increased accuracy. Spatial data collected at a finer
resolution (e.g. via GPS PTTs) would provide better insight into foraging locations and
therefore improved understanding of potential overlap between brown pelican home
ranges and planned sites for offshore wind and other energy development.
Finally, interactions that occur away from the breeding colony, (i.e. on the
wintering grounds, on the foraging grounds) may influence breeding success, individual
survival, and population dynamics in brown pelicans. Population recovery range-wide is
an issue since the species is still listed as one of concern in several states. Therefore, a
holistic approach to management of areas that are important for brown pelicans year-
round, rather than an approach based solely at an individual breeding colony, may more
comprehensively address the ecological needs of the species.

43
CHAPTER THREE
VARIABILITY IN FORAGING BEHAVIOR OF MASKED BOOBIES (SULA
DACTYLATRA) BREEDING AT ISLA MUERTOS, MEXICO
Introduction
Within populations, patterns in resource use may vary widely among individuals
such that the summarized behavior of the population does not accurately represent
interactions that are ecologically important at an individual level (Van Valen 1965,
Bolnick et al. 2003). For example, species that are typically characterized as generalists
may consist of individuals with specialized behaviors (Bolnick et al. 2003) leading to a
wide array of foraging strategies that are challenging to describe at the population level.
These diverse foraging strategies may, however, serve to reduce intraspecific
competition, perhaps via mechanisms such as the expansion of diet breadth in a portion
of the population (Goss-Custard et al. 1984, Golet et al. 2000). Individual variability in
foraging behavior also may mitigate the effects of intraspecific competition when
resources are patchy and ephemeral (Goss-Custard et al. 1984), and may therefore
increase the resiliency of a population to changes in prey availability in space and time
(Durell 2000).

44
In marine environments, variable foraging strategies within populations of apex
predators may be a reflection of the patchy distribution of prey fish (Golet et al. 2000,
Staniland and Boyd 2003). In pelagic systems, features of the environment drive prey
distribution (Cushing 1990, Anderson and Rodhouse 2001, Stevens et al. 2003); for
instance, sea surface temperature and productivity influence geochemistry that
subsequently can affect the abundance of prey (Ryther 1969), and water circulation and
mixing regimes transport and aggregate prey (Lobel and Robinson 1986). Prey can
respond to these features across a wide range of spatial and temporal scales (e.g. hours to
decades, single meters to hundreds of km) and these responses can differ depending on
species, density, and behavioral patterns of prey, resulting in prey that are patchy and
vary in their associations with features of the environment (Haury et al. 1978,
Sanvicente-Añorve et al. 2000, Pattrick and Strydom 2014). Apex predators must
therefore efficiently process environmental cues derived from physical habitat features to
locate prey, and variability and patchiness of the environment may be reflected in
foraging behavior among individuals.
Habitat associations in seabirds have been commonly studied, and foraging
behavior of groups of individuals within species has been linked to a variety of
oceanographic features including but not limited to sea surface temperature (Kappes et al.
2010), mesoscale currents (Kai et al. 2009), and depth of the thermocline (Spear et al.
2001). The masked booby (Sula dactylatra) is a predatory seabird with a pantropical
distribution that seizes prey by plunge-diving into schools of fish (Anderson 2009), and
foraging behavior in the species therefore may be linked to oceanography that influences

45
the distribution of prey. Masked boobies forage on a variety of prey species (Anderson
2009), and it is possible that the average or typical behavior that characterizes
populations does not fully describe the variability in prey-sensing behavior that may
occur among individuals. Approximately 60% of the north Atlantic population of masked
boobies breeds at Isla Muertos, which is located in the southern Gulf of Mexico. Little is
known about the birds’ habitat use in relation to the marine areas in and adjacent to Isla
Muertos or the Gulf of Mexico in general, particularly as it may relate to environmental
features that correspond to foraging. Studies of masked boobies in other systems suggest
that foraging behavior during breeding varies between populations and locations (Asseid
et al. 2006, Weimerskirch et al. 2008, Weimerskirch et al. 2009a, Young et al. 2010,
Kappes et al. 2011, Sommerfeld 2013). Foraging behavior also varies among individuals
within a population in similar species including northern gannets (Morus bassanus)
during migration (Kubetzki et al. 2009), and cape gannets (Morus capensis) during chick
provisioning (Mullers and Tinbergen 2009). Information on individual variability in prey
searching behavior of masked boobies in relation to oceanography of the Gulf of Mexico
may provide a new perspective on foraging behavior in the species.
I examined at-sea movements of breeding masked boobies tracked with GPS
tags. My objectives were to 1) describe locations of foraging trips and determine foraging
behavior, 2) examine prey-searching behavior among individuals, and 3) investigate
oceanography of foraging locations and therefore prey patches. I predicted that, at an
individual level, masked boobies would exhibit behavioral differentiation, resulting in
diverse search behaviors, and that variability in the environmental characteristics

46
associated with foraging habitat would be high between individuals. These data represent
the first assessment of movements to date for the species in the Atlantic region and as
such can be used to understand how top predators respond to oceanography in the Gulf of
Mexico and to provide a template for the study of the other seabirds nesting in the region.
Methods
Study area
Data were collected 17-29 May 2013 and 1-15 November 2013 on Isla Muertos
(22.4 N 89.7 W) at Arrecife Alacranes National Park (AANP), which is located on
Campeche Bank in the southern Gulf of Mexico (Fig. 3.1). Isla Muertos is approximately
0.5 km2 and consists of coral sand platform reef (Chávez and Hidalgo 1988), with bare
ground surrounding a core of low-lying succulent vegetation, grasses, and shrubs
(Protegidas 2007). All seabird species that breed on the island also share nesting habitat
with masked boobies, including ca. 350 pairs of magnificent frigatebirds (Fregata
magnificens), ca. 200 pairs of sooty terns (Sterna fuscata), ca. 13 pairs of red-footed
boobies (Sula sula), and ca. 9 pairs of laughing gulls (Leucophaeus atricilla). There are
2500-3000 breeding pairs of masked boobies and ca. 100 additional pairs breed on other
islands in AANP (A.V. Moncada unpublished data). Masked boobies at AANP breed
year-round with peak nesting activity in May-June and November-December (A.V.
Moncada unpublished data).

47
Figure 3.1 Location of Isla Muertos, the largest breeding colony for masked boobies in
the north Atlantic region, in relation to the boundary of Arrecife Alacranes National Park
and the prominent bathymetry of the Campeche Bank, Mexico. At-sea foraging
movements of 77 breeding masked boobies were recorded with GPS loggers during May
and November 2013.

48
Data collection
I used archival GPS loggers (Mobile Action I-gotU g120) to record the foraging
tracks of individual masked boobies. Tags were removed from the commercial casing and
repackaged in heat-shrink tubing. The frequency with which tags recorded locations was
adjusted several times during the study (one location every 10, 33, 50, or 100 sec) to
optimize battery life. Birds were captured from opportunistically selected nests with a
handheld net, and tags were attached to the underside of three central retrices with 4-5
strips of Tesa tape. The total attachment (GPS tag and tape) weighed 20.6 grams,
corresponding to 1.7% of the mass of the lightest adult measured. During initial capture,
adults were sexed by vocalization (Nelson 1978), weighed (± 10 g), measured for culmen
and tarsus (± 0.1 mm), and colormarked on the chest with non-toxic paint to facilitate
detection of tagged individuals on nests; chicks were weighed (± 1 g) and measured for
relaxed wing chord (± 1 mm), head-bill, and tarsus (± 0.1 mm). Birds were released at the
nest and in all cases quickly resumed attendance. In May, all tagged birds were attending
chicks ranging in development from ca. 1 - 10 weeks post-hatch based on feather
development (e.g. feathering ranged from unfeathered to body feathers and partially
grown wing feathers). In November, few chicks were present, and tagged birds were
incubating or brooding young chicks that ranged in age from ca. 1 - 4 weeks post hatch
(e.g. unfeathered to body down and no wing feathers).
GPS-tagged individuals were recaptured 37 - 101 hours after deployment. Upon
recapture, GPS tags were removed and birds were examined for condition. Tag data were
downloaded and data were grouped into individual trips beginning with the location

49
immediately prior to colony departure and ending with the location immediately
following colony return. Only complete trips with regular locations from colony
departure to colony return were included in analyses. Each foraging trip, or track, was
processed using the R package adehabitatLT (Calenge 2011a). Locations that represented
unrealistic flight speeds over 90 km/h were removed (< 20 locations), and tracks were
interpolated at 100-second intervals. Overnight trips were subdivided to exclude
nocturnal locations where birds appeared to be loafing on the water (Sommerfeld et al.
2013a).
For each individual trip, I calculated trip duration (hours between colony
departure and return), total distance traveled (sum of the displacements between adjacent
locations), and maximum distance from the colony (largest Euclidean distance from a
location off of the colony to the colony). Outbound and inbound azimuths were
calculated by measuring the angle between the colony and the location of the bird when it
was 2 km from the colony.
Habitat use
GPS data can be used to infer locations which correspond to active foraging (i.e.,
area restricted search, ARS) by identifying areas in which the rate of turning is increased
and the rate of speed is reduced (Tinbergen et al. 1967). I applied a first passage time
(FPT) analysis (Fauchald and Tveraa 2003) to each trip (Fig. 3.2) to delimit locations
which correspond to ARS. One location from a trip was centered within a circle of a
fixed radius, and the time required for an animal to traverse forward and backward along

50
the track from that location until it exited the circle was measured. The measurement was
repeated for each location along the track. The entire iteration was repeated for circles of
radii ranging from 10 m (the approximate accuracy of GPS locations) to 35000 m (1/2 the
average distance from the colony for all recorded locations), at 10 m increments. The
difference between the FPT backwards and forwards through each circle from the central
point was log-transformed so that the variance was independent of the magnitude of the
mean (Fauchald and Tveraa 2003), which improved detection of maxima. The maxima in
variance(log(FPT)) correspond to radii at which search effort is concentrated, and this
measure was used to identify prominent scales of ARS. To designate locations as ARS or
non-ARS for each identified scale I used the penalized contrast method of Lavielle
(Lavielle 2005, Barraquand and Benhamou 2008) to group locations that have high FPT
values and are close to each other along the track. The method is a non-parametric
segmenting function based on a contrast matrix. The number of possible segments that
could be formed from a track (k-value) was set to a minimum of two and a maximum of
20. Analysis was completed using the R package adehabitatLT (Calenge 2011a). In some
cases, FPT analysis identified multiple ARS radii within a single trip. For fine-scale
analysis of foraging behavior and oceanographic habitat use, I selected only the smallest
scale for each trip. I also eliminated ARS scales over seven km that likely corresponded
to mesoscale search behavior rather than targeted prey-capture.

51
Figure 3.2 Sample analysis of a GPS track illustrating delineation of locations that
correspond to area restricted search (ARS) behavior. Scale bar in (a) also applies to
panels (b) and (d). (a) A portion of a GPS track (line) with individual locations (points).
(b) For each location along the track, the amount of time required to cross a circle of a
given radius (in this case radius = 2 km) is calculated (the first passage time; FPT). Red
arrows indicate direction of travel. The circle on the left contains an example of direct
movement at a moderate speed, resulting in short FPT. In contrast, the circle on the right
contains an example of tortuous movement at slow speed, resulting in long FPT and
potentially corresponding to ARS behavior. (c) FPT analysis is repeated multiple times,
using circles with incrementally increasing radii (x-axis). For each radius, the variance
log(FPT) of the calculated FPT values is plotted and those points are connected via an
interpolated line for optimal visualization. The radius that corresponds to the peak
log(variance) is the dominant scale of ARS. In this example, maximum variance
log(FPT) corresponds to a search radius of 1980 meters. (d) Once the scale of ARS is
identified, the penalized contrast method of Lavielle is used to cut the track into segments
consisting of sets of locations with similar FPT values (range 0 - 23 h). Segments with
overall high FPT values correspond to ARS behavior (blue points); segments with low
FPT values correspond to non-ARS behavior (gray points).

52
Environmental variables were extracted for each individual location at the
smallest available spatial scale and linearly interpolated in ArcGIS using Duke
University’s Marine Geospatial Ecology Tools (Roberts et al. 2010) unless otherwise
specified. A blended product of daily sea surface temperature (SST; °C) was derived at
three scales; daily SST at spatial resolution of 0.01°, 8-day composites at a spatial
resolution of 0.05°, and monthly composites at a spatial resolution of 0.05
°. Daily data
were compiled from various sensors and a Global 1 km SST (G1SST) analysis by the
Group for High Resolution Sea Surface Temperature (GHRSST). Eight-day and monthly
data were gathered through NASA’s Jet Propulsion Laboratory and published by the
Physical Oceanography Distributed Active Archive Center (PO.DAAC). Sensors that
gathered measurements included the Advanced Very High Resolution Radiometer
(AVHRR), the Advanced Along Track Scanning Radiometer (AATSR), the Spinning
Enhanced Visible and Infrared Imager (SEVIRI), the Advanced Microwave Scanning
Radiometer-EOS (AMSRE), the Tropical Rainfall Measuring Mission Microwave Imager
(TMI), the Moderate Resolution Imaging Spectroradiometer (MODIS), the Geostationary
Operational Environmental Satellite (GOES) Imager, the Multi-Functional Transport
Satellite 1R (MTSAT-1R) radiometer, and in situ units attached to buoys.
Monthly values of chlorophyll-a (mg/m3) were derived at a spatial resolution of
0.05° from standard mapped images provided by the Moderate Resolution Imaging
Spectroradiometer (MODIS) sensor carried onboard NASA’s Aqua satellite. Velocity
(m/s) of flowing water was calculated as the sum of eastward (u) and northward (v) sea
water velocity and interpolated in five-day increments at a spatial resolution of 0.33° from

53
satellite observations from altimeters, scatterometers, and SST sensors by the National
Oceanic and Atmospheric Administration’s (NOAA) Ocean Surface Current Analyses
Real Time (OSCAR) project. Bathymetric data (m) were obtained at a spatial resolution
of 0.016° from NOAA’s National Geophysical Data Center ETOPO1 Global Relief
Model (Amante and Eakins 2009) using the R package marmap (Pante and Simon-
Bouhet 2013), and extracted for each GPS location using R package raster (Hijmans and
van Etten 2012).
Fourteen-day oceanic front probability was generated at a spatial resolution of
0.05° through the NOAA CoastWatch Program by applying an edge detection algorithm
to daily SST datasets acquired onboard NASA’s Geostationary Operational
Environmental Satellites (GOES) (Breaker et al. 2005). The dataset was accessed by
querying ERDDAP (http:// coastwatch. pfel.noaa. gov/erddap/). I calculated distance
from GPS locations to nearest front.
Daily sea surface height anomaly (SSH; m above a reference sphere) and mixed
layer depth (MLD; m; the depth at which the temperature is 0.2° C cooler than the
temperature at the surface) were derived at a spatial resolution of 0.05° from data from
sensors and the Hybrid Coordinate Ocean Model (HYCOM; Cummings and Smedstad
2013) Gulf of Mexico analysis. Analysis and data collection are performed by the Navy
Coupled Ocean Data Assimilation (NCODA), which assimilates data from satellite
sensors collected by the Naval Oceanographic Office (NAVOCEANO) Altimeter Data
Fusion Center, satellite and in situ SST measurements, and in situ vertical temperature

54
and salinity profiles from ship-based expendable bathythermographs (XBTs), free-
drifting Argo floats, and moored buoys.
Statistical analysis
All statistical analyses were conducted in R 3.0.3 (R Development Core Team
2014). A variable number of trips were recorded for each bird, therefore, to achieve even
representation across individuals I randomly selected one trip per bird for all subsequent
analyses, resulting in 77 trips (32 in May and 45 in November). Wilcoxon rank-sum tests,
Student’s t-tests, Kruskal-Wallis tests, and generalized linear mixed models (GLMM)
with individual bird identity as a random effect were used to compare characteristics of
foraging trips and ARS behavior between seasons, sexes, and stages (i.e., incubation and
chick-rearing). Comparison of stages was conducted for November only as May data
included only chick-rearing birds and November data included chick-rearing and
incubating birds. Circular statistics for comparing azimuths of departure, arrival, and
ARS between seasons, sexes, and stages were calculated using package circular
(Agostinelli and Lund 2013). Watson's Two-Sample Test of Homogeneity was used to
compare azimuths between groups. The level of significance for each test was set to 0.05
although p-values are reported throughout.
I used a bootstrapped classifier algorithm, the random forest (RF) analysis
(Breiman 2001, Cutler et al. 2007), to determine the extent to which individual
environmental variables of interest predicted ARS behavior. For a single trip consisting
of a series of locations, I generated a classification tree with ARS as a binomial

55
dependent variable (ARS, non ARS) and nine environmental variables of interest as
predictor variables. At each branch, the environmental variable and corresponding value
that resulted in the most accurate split of locations into ARS and non-ARS behavior was
selected. The process continued until as many locations as possible were classified,
resulting in a final model. The tree was built using a random selection of two-thirds of the
locations from each trip; the remaining one-third of the locations was used for model
validation. To build a RF, the process was iterated 1000 times, resulting in 1000 possible
trees. At a given branch, RF analysis randomly selected and considered only three
predictors of interest from the pool of nine (the number of predictors used at each branch
was calculated as the square-root of the total number of predictors; Breiman 2001).
Correlation between environmental variables differed between trips, however, during
permutation of trees, variables were often subsetted so they were separate from one
another and thus could be evaluated for performance independently. The Gini Index, a
measure of the relative contribution of each variable towards formulation of a complete
model (a.k.a. reduction in node impurity), was computed across all trees.
To identify environmental conditions that correspond to foraging behavior on an
individual level, I conducted a separate RF analysis for each trip using package
randomForest (Liaw and Wiener 2002). I extracted the Gini index for nine environmental
variables for each of 72 trips. To consolidate each of the 72 models, I considered only
environmental variables with a Gini index that was greater than 15% of the total
reduction in node impurity for each model. This resulted in a majority of models with an

56
optimal amount of complexity (1-3 variables), and few models with no complexity (0
variables) or a high degree of complexity (4-5 variables).
I used generalized additive mixed models (GAMM) to investigate and visualize
the shape of several of the prevalent relationships (as identified by RF analysis) between
ARS behavior and environmental characteristics. I chose to focus on three environmental
characteristics that contributed frequently to the separation of ARS and non-ARS
behavior in RF models and were important for a large number of individuals: SSH,
velocity, and distance to front. A binomial GAMM (package mgcv; Wood 2011) was
applied separately for each of the three environmental variables, with ARS behavior at a
given location as the dependent variable (1, ARS; 0, no ARS), and SSH, velocity, or
distance to nearest front as the explanatory variable (adding more complexity and
variables decreased the likelihood that models would converge). Individual was included
as a random effect, and a first-order autoregressive time series structure was incorporated
to account for non-independence of locations within a trip. Cubic regression splines are
flexible for modeling a variety of nonlinear relationships (Zuur et al. 2009) and were
specified as the smooth.
Models were built using the following steps: 1) choose a variable of interest
(SSH, velocity, or distance to front), 2) select only the birds that extensively used the
environmental variable under consideration (extensive use was defined earlier as a
contribution to reduction of node impurity of over 15% in RF models), 3) plot and
examine the overall shape of the nonlinear relationship for each bird, 4) pool data for
birds with a similar-looking relationship (n.b. this occurred without accounting for

57
season, sex, or stage since the goal is to identify trends in behavior), 5) apply the GAMM
to the pooled data, 6) if the model converges and has small confidence intervals, increase
complexity by including more birds, 7) repeat steps 3 - 6 with that goal of building
several models for one variable that illustrate different types of trends (e.g. positive,
negative).
Results
Birds primarily traveled south-southeast of the colony towards the Yucatán
peninsula during foraging trips in both May and November (Fig. 3.3). Several birds
foraged west or north of the colony, and two individuals traveled northwest to the edge of
the Campeche Bank shelf (Fig. 3.3). Birds ranged beyond the park boundary in 87% of
trips, and 86.4% of foraging locations occurred outside of the park. Movements from the
colony to ARS zones and from ARS zones back to the colony were usually direct, though
a few birds meandered extensively.

58
Figure 3.3 Arrecife Alacranes National Park boundary (black oval) in relation to
locations of small-scale area restricted search behavior (colored points) identified from
first passage time analysis. Foraging trips (gray lines; n = 129; multiple trips per bird are
pictured) were recorded from masked boobies nesting at Isla Muertos (black star),
Mexico, May and November 2013.

59
Scales of area restricted search
Overall, one-hundred twenty-nine complete foraging trips were recorded from 77
individuals (Fig. 3.3). From the subset of trips which were selected for further study (i.e.,
one trip from each bird, complete trips only, n = 77), Masked Boobies displayed nested,
hierarchical scales of ARS (Fig. 3.4). Plots of variance of log(FPT) indicated ARS
occurred at a single scale in 35% of birds (i.e., single and distinct peak) but at multiple
scales (i.e., 2 - 5 localized peaks) in 65% of birds. FPT analysis identified at least one
scale of ARS in 97% of birds. In the remainder of trips (three out of 77), no ARS was
identified suggesting that birds did not concentrate search effort at any particular scale.

60
Figure 3.4 An example of multiple scales of area restricted search (ARS) behavior during
a single foraging trip recorded for a masked booby nesting at Isla Muertos (star), Mexico,
May 2013. Multiple scales of ARS were identified from the variance log(first passage
time) plot (inset box) which has four prominent local maxima, each indicated by a dotted
line. Within the track, the locations which correspond to foraging at each scale (large
map; delineated by colors that correspond with the dotted lines on the plot) are nested
from largest to smallest. Points indicate interpolated GPS locations, circles indicate
search radii, arrows indicate direction of travel.

61
At-sea movements and foraging behavior
ARS behavior regularly occurred at the furthest distance from the colony during
each trip (Spearman’s rho = 0.83, p < 0.001), which was near the temporal midpoint of
foraging trips. The variability in the direction of approach to the colony during inbound
legs of foraging trips was relatively low (CV = 0.18; Fig. 3.5; Table 3.1).

62
Figure 3.5 Distribution of azimuth from the colony during foraging trips of masked
boobies nesting at Isla Muertos, Mexico, May and November 2013. Departure and
arrival: angle between the colony and the location of the bird when it was two km from
the colony. ARS: angle between the colony and locations of area restricted search.
Numbers and dashed circles inside each plot correspond to the number of individuals
included in each bar. n = 77 birds.

63
m
ean ±
sd (ran
ge)
Seaso
n e
ffect
S
ex e
ffect
Stag
e effec
t
W o
r χ2
P
W
or χ
2 P
W
P
Trip
duratio
n (h
) 1
1.3
± 9
.9 (1
.0 - 4
9.0
) 5
65
0.1
87
8
0.1
6
1
30
0.0
1
To
tal distan
ce traveled
(km
) 2
05
.1 ±
11
5.8
(22
. 9 - 5
57
.0)
45
9
0.0
1
9
13
0.0
7
1
77
0.2
0
Max
imu
m d
istance fro
m co
lon
y (k
m)
74
.1 ±
38.4
(10
.2 - 2
11
.0)
47
3
0.0
1
8
84
0.1
4
1
84
0.2
6
Tim
e of co
lon
y d
epartu
re (hh:m
m)
9:2
5 ±
3:4
7 (3
:40
- 18
:06)
10
80
< 0
.01
4
93
0.0
1
2
92
0.1
6
Azim
uth
from
colo
ny at d
epartu
re (°) 1
90
.7 ±
57
.7 (1
7.2
- 34
9.7
) 0
.7
0.9
6
1
.4
0.4
9
-
-
Azim
uth
from
colo
ny at arriv
al (°)
20
7.0
± 3
7.1
(102
.6 - 2
86
.7)
6.2
0
.05
3
.7
0.1
6
-
-
Nu
mb
er of n
ested A
RS
scales
1.9
± 0
.9 (1
- 5)
73
1
0.9
1
7
65
0.7
7
2
61
0.4
7
Size o
f AR
S zo
nes (k
m)
5.3
± 5
.5 (0
.1 - 2
6.0
) 1
97
7
0.0
3
2
60
5
0.9
2
9
40
0.4
1
Table 3.1 Foraging parameters of
masked boobies nesting at Isla Muertos,
Mexico, May and November 2013.
Wilcoxon rank-sum tests were used to
compare characteristics of foraging trips
between seasons (May and November; n
= 77 birds), sexes (n = 77 birds), and
stages (incubating and chick-rearing in
November; n = 41 birds). Watson's Two-
Sample Test of Homogeneity was used to
compare circular statistics. Values are
presented as mean ± 1 standard
deviation. Ranges are in parenthesis.
Significant results appear in bold; alpha
= 0.05.

64
Within-season variability in trip duration, total distance traveled, maximum
distance from the colony, and size of ARS zones was greater in May compared to
November. Birds traveled longer total distances in November (228.4 ± 102.6 km)
compared to May (172.4 ± 126.7 km; Table 3.1). Additionally, a higher percentage of
birds traveled west in November (40%) compared to May (26%; Fig. 3.5), and birds that
traveled west in November also ranged farther from the colony than birds that traveled
west in May (Wilcoxon rank sum test W = 12, P < 0.001). ARS scales were larger in
November (6.1 ± 5.6 km) compared to May (4.2 ± 5.2 km), and ARS zones were located
farther from the colony in November (79.5 ± 32.3 km) compared to May (60.0 ± 35.6
km). Upon return from foraging trips, the distribution of angles at which birds
approached the colony differed slightly between May (204.5 ± 43.9°) and November
(209.4 ± 31.7°; Watson's Two-sample test of homogeneity χ2 = 6.2, P = 0.05).
Birds left the colony primarily during daylight, though a small percentage
departed prior to civil dawn, (3% in May, 22% in Nov; no birds departed after civil
twilight). Birds departed the colony later in May (11:07 ± 3:46 hours) compared to
November (08:14 ± 3:11 hours; Table 3.1). Females departed the colony earlier (08:31 ±
3.18 hours) compared to males (10:30 ± 3:54 hours). Birds that initiated trips earlier in
the day traveled significantly farther distances from the colony (GLMM t = -3.8, P =
0.0003) and spent more time in small-scale ARS (GLMM t = -3.0, P = 0.004) but did not
significantly increase the total duration of trips (GLMM t = -0.5, P = 0.62) compared to
birds that initiated trips later in the day. Birds spent the night away from the colony
during 21% of foraging trips (males 16% of trips, females 24% of trips) and appeared to

65
rest on the water (based on periods of little movement observed in GPS tracks) from after
civil twilight until just before civil dawn.
Trip duration in November was longer and more variable in incubating birds (14.1
± 10.2 h) compared to chick-rearing birds (7.7± 4.8 h; Table 3.1). Additionally, at the
smallest ARS scale measured for each bird (< 7 km), the size and number of ARS zones,
and the distance traveled and time spent in ARS was more variable in incubating birds
compared to chick-rearing birds. Birds spent significantly more time in ARS during
incubation (3.3 ± 2.0 h) compared to chick-rearing (1.8 ± 0.8 h; Table 3.2).

66
Table 3.2 Foraging behavior of masked
boobies nesting at Isla Muertos, Mexico
during May and November 2013.
Wilcoxon rank-sum tests were used to
compare characteristics of foraging
behavior between seasons (May and
November; n = 72 birds), sexes (n = 72
birds), and stages (incubating and chick-
rearing in November; n = 41 birds).
Watson's Two-Sample Test of
Homogeneity was used for circular
statistics. Values are presented as mean ± 1
standard deviation. Ranges are in
parenthesis. Significant results appear in
bold; alpha = 0.05. ARS: area restricted
search.
m
ean ±
sd (ran
ge)
Seaso
n e
ffect
S
ex e
ffect
Stag
e effec
t
W o
r χ2
P
W
or χ
2 P
W
P
Size o
f AR
S zo
nes (k
m)
1.9
± 1
.4 (0
.1 - 6
.1)
53
4 0
.25
6
47
0.9
9
24
6 0
.22
Nu
mb
er of A
RS
zones p
er trip 1
.4 ±
0.7
(1 - 4
) 6
51
0.8
3
66
2 0
.84
1
36
0.3
1 T
ime sp
ent in
AR
S (h
) 2
.6 ±
1.8
(0.2
- 8.1
) 5
11
0.1
6
77
8 0
.14
9
6 0
.01
Dista
nce trav
eled in
AR
S (k
m)
12
.6 ±
10.6
(1.0
- 60.9
) 6
18
0.8
5
70
4 0
.53
2
22
0.5
7 D
istance fro
m co
lon
y to
AR
S (k
m)
64
.6 ±
32
.6 (1
.8 - 1
40
.9)
33
5 <
0.0
1
68
3 0
.70
1
57
0.2
6 A
zimu
th fro
m co
lon
y to
AR
S (°)
17
0.5
± 6
1.8
(12
.6 - 3
28
.0)
0.4
0.8
1
0.4
0.8
2
- -

67
Oceanographic habitat use
Of the nine environmental variables that I assessed, there was a high degree of
variability in the number and identity that were identified as important predictors of
foraging behavior for each of the 74 birds (three individuals with no identifiable scale of
ARS were excluded from analysis). The relative contribution of each variable to
reduction in node impurity tended to be low to moderate, ranging from range 0 - 37.8%.
SSH and velocity independently appeared in the greatest number of individual models
(44.4% and 37.5% of models, respectively; Table 3.3) and had the highest percent
reduction in node impurity within each model (23.6 ± 6.3%, 23.4 ± 6.2%, respectively).
Distance to front contributed less frequently to individual models (26.4%; Table 3.3)
compared to other variables, but the percent reduction in node impurity within models
(22.0 ± 5.7%) was similar to levels achieved by SSH and velocity. In 42% of trips, two
environmental variables were identified as important predictors of foraging behavior; in
43% of trips, three variables were identified as important predictors of foraging behavior.
Two-variable combinations that appeared frequently were SSH + velocity (21%), SSH +
MLD (11%), and SSH + 8-day SST (8%). There was little consistency between birds in
the identity of variables appearing in the three-variable models, however, three-variable
combinations that appeared most frequently were SSH + velocity + distance to front and
SSH + velocity + chlorophyll-a (5% each).
Validation of the random forest models indicated that the percentage of locations
correctly classified for each model from the reserved data was high (99.0 ± 1.1%).
Sensitivity (the percentage of ARS locations correctly classified) and specificity (the

68
percentage of non-ARS locations correctly classified) of individual models was also high
(sensitivity: 98.9 ± 1.4%; specificity: 98.6 ± 2.1%). The probability of a bird engaging in
ARS behavior based on SSH, velocity, distance to front) was nonlinear for 92% of birds
(Fig. 3.6). I detected both negative and positive trends in the probability of ARS behavior
occurring in relation to SSH, velocity, and distance to front.

69
Table 3.3 Physical
oceanographic
characteristics
considered in random
forest analysis and the
hypothesized link with
foraging behavior of
masked boobies
nesting at Isla
Muertos, Mexico, May
and November 2013.
Frequency and percent
of models represent the
number and percent of
individual models (n =
72 birds) in which a
particular variable was
considered important
for predicting foraging
behavior. Temporal
scale and spatial
resolution refer to the
scale and resolution at
which the
environmental
variables were
measured (see
Methods).
Variab
le T
emp
oral
scale
Sp
atial
resolu
tion
Hyp
oth
esized lin
k
Freq
uen
cy
Percen
t
of m
od
els
Sea su
rface heig
ht an
om
aly
Daily
0
.05
° P
hysical fo
rcing in
fluences p
rey d
istributio
n an
d
turb
ule
nce in
fluences d
etection o
f pre
y b
y M
AB
O
32
44
.4
Velo
city o
f mo
vin
g w
ater F
ive-d
ay
0.3
3°
Sp
eed, d
irection, an
d m
ag
nitu
de o
f water
mo
vem
ent lin
ked
to p
rey d
istributio
n
27
37
.5
Chlo
rop
hyll-a co
ncen
tration
Mo
nth
ly 0
.05
° B
iogeo
chem
ical reg
ime lin
ked
to p
rey d
istributio
n
and
abund
ance
20
27
.8
Dista
nce to
nearest S
ST
front
Fo
urteen
-day
0.0
5°
Pro
xim
ity to
feature
s that a
ggreg
ate pre
y an
d
influ
ence p
rod
uctio
n; v
isual c
ue fo
r MA
BO
19
26
.4
Mix
ed la
yer d
epth
D
aily
0.0
5°
Influ
ences activ
ity o
f sub
surfa
ce pred
ators an
d
prey
and
therefo
re accessibility
of p
rey fo
r MA
BO
15
20
.8
Sea su
rface tem
peratu
re
Daily
0
.01
° Im
med
iate therm
al cond
ition
s linked
to p
rey
activity
level
14
19
.4
Sea su
rface tem
peratu
re E
igh
t-day
0.0
5°
Recen
t therm
al cond
ition
s linked
to p
rey activ
ity
level an
d ab
und
ance
8
11
.1
Bath
ym
etry N
one
0.0
16
° W
ater circulatio
n an
d m
ixin
g reg
ime in
fluen
ces
prey
com
mu
nity
com
po
sition an
d p
rey ab
und
ance
8
11
.1
Sea su
rface tem
peratu
re M
onth
ly 0
.05
° T
herm
al regim
e linked
to p
rey ab
und
ance
6
8
.3

70
Figure 3.6 Generalized additive mixed models (GAMM) were used to examine the
relationship between area restricted search (ARS) behavior and physical oceanography in
foraging masked boobies nesting at Isla Muertos, Mexico, May and November 2013.
Different groups of individuals that showed similar trends for a particular environmental
variable of interest were used to build each working model. N for each curve is listed
near the y axis of each plot, and the total number of birds that showed a strong
relationship for a particular environmental variable (as determined by random forest
analysis) and could have potentially contributed to GAMM formation was sea surface
height anomaly = 32 and velocity of water = 27. The colored line represents model
predictions and light-gray areas represent 95% confidence intervals.

71
Discussion
At-sea movements and foraging behavior
Variability in foraging behavior and foraging locations was high among
individuals during this study, both within and between seasons and breeding stages. The
timing and duration of foraging trips, the number and size of scales at which birds
searched for prey, and the environmental features that characterized small-scale search
locations were all highly variable. High levels of individual variability in movement
patterns have been observed in other species of Pelecaniformes at multiple spatial and
temporal scales, including the timing and location of migratory movements in northern
gannets (Morus bassanus; Kubetzki et al. 2009), the duration of foraging trips in breeding
Cape gannets (Morus capensis; Mullers and Tinbergen 2009), and the depth and
frequency of foraging dives in king cormorants (Phalacrocorax atriceps; Kato et al.
2000).
The breeding population at Isla Muertos is large (2500 - 3000 pairs), and birds
may compete for resources with conspecifics from a neighboring colony 170 km to the
west (Cayo Arenas; 100 - 300 pairs), which is within the foraging range for masked
boobies described in this study. Colony size and productivity of water surrounding the
colony may interact to shape foraging behavior in seabirds (Ashmole 1963). For example,
Grémillet et al. (2004) suggested that competition between two neighboring colonies of
Cape gannets lead to colony-based differences in habitat use. Similarly, Wakefield et al.
(2013) found that foraging ranges in northern gannets were greater when competition

72
within colonies was high and when colonies were large. Individual variability in foraging
behavior and foraging locations may be a mechanism by which intraspecific competition
is reduced (Van Valen 1965), particularly in colonial species (Bolnick et al. 2003),
especially when there is little differentiation in foraging behavior between sexes. The
variable foraging behavior observed among individuals in this study and the lack of
consistent differentiation in foraging strategies between sexes may therefore be a
reflection of intraspecific competition.
High individual variability in foraging behavior and foraging locations may also
increase the rate of detection of prey patches among individuals where the diet is not
homogeneous and where prey availability is ephemeral in time and space. In combination
with visual flock recruitment, a common behavior in Sulids (Nelson 1978, Haney et al.
1992, Tremblay et al. 2014), variability in foraging behavior may improve foraging
efficiency among individuals (Buckley 1997). For example, in social ants, individual
variability in foraging strategies lead to efficient search coverage of the area surrounding
the colony and fast identification of food patches (Deneubourg et al. 1983, Portha et al.
2002). Regurgitated fish collected from masked boobies at Isla Muertos comprised six
species in five families, with no single species appearing in more than 35% of samples
(A. Moncada, unpublished data), supporting the concept that different individuals from
the same colony capture a variety of prey and may therefore be employing different
foraging strategies.
Foraging behavior during incubation was more variable and trips were longer in
duration compared to chick-rearing. Trade-offs associated with meeting the energetic

73
demands of both parent and offspring during chick-rearing may result in a detectable shift
in behavioral patterns between incubation and chick-rearing stages resulting in more
consistency in foraging trip duration when chicks are being provisioned (Robertson et al.
2014). Attendance at the nest, and hence the duration of foraging trips, also can be
influenced by predation rates at the nest. In situations where predation at the nest is high,
parents may invest more time in nest or chick guarding (Rothenbach and Kelly 2012). At
Isla Muertos, predation of masked booby chicks (but not eggs) by breeding magnificent
frigatebirds is common (personal observation), and unguarded chicks could also be
attacked by masked boobies from neighboring territories (Weimerskirch et al. 2009a).
Nest defense during chick-rearing may therefore be an important component of
individual reproductive success and a strong driver of differences in foraging behavior
between incubating and chick-rearing birds, leading to foraging behavior that is less
flexible during the chick-rearing stage.
Sex-specific foraging behavior
Characteristics of foraging trips (i.e., range, duration) and foraging behavior
within trips (i.e., size and number of ARS zones, and time spent in ARS) did not differ
between sexes. Studies of breeding masked boobies in the western Indian Ocean (Kappes
et al. 2011), central Pacific (Young et al. 2010), and eastern Pacific (Weimerskirch et al.
2009a) also found little evidence for differentiation in foraging behavior between sexes.
Young et al. (2010) posit that lack of food resources may limit foraging locations within
commuting distance from the colony to one area, subsequently resulting in males and

74
females foraging in the same location. In environments where productivity is high and
broadly distributed, foraging behavior may differ between the sexes. Abundant or
widespread productive areas may allow for higher flexibility in behavior between males
and females, resulting in division of labor, as is the case for blue-footed boobies (Sula
nebouxii) and brown boobies (Sula leucogaster) in the Gulf of California (Weimerskirch
et al. 2009b).
Sex-based differences in foraging behavior may also be masked by resource
depletion near breeding colonies, resulting in both sexes increasing foraging ranges.
Hence, differentiation in foraging behavior between sexes may be more likely to occur at
small colonies in productive areas where competition is low. Studies of masked boobies
that have found little evidence of differentiation in foraging behavior between the sexes
are restricted to small colonies (10 - 300 pairs) in unproductive environments (Young et
al. 2010, Kappes et al. 2011, Sommerfeld 2013), or extremely large colonies (> 100,000
pairs) in highly productive environments (Weimerskirch et al. 2009a). In the former, low
intraspecific competition but limited places to forage results in overlapping use of a small
area. In the latter, an abundance of suitable foraging locations may allow for flexibility in
behavior between sexes, but high intraspecific competition could result in lack of
differentiation. Productivity in the southern Gulf of Mexico is moderate (Longhurst
2010), and foraging trips recorded during this study were distributed across multiple
locations (Fig. 3.3). Prey availability may therefore be adequate to support flexibility in
parental roles, however high intraspecific competition may have limited the extent of
differentiation between the sexes. Finally, differentiation during foraging at Isla Muertos

75
may occur within, rather than among foraging patches as female masked boobies rearing
chicks in the eastern and western Pacific dive deeper and more frequently than males
(Weimerskirch et al. 2009a, Sommerfeld et al. 2013b), and also deliver significantly more
food to the chick (Weimerskirch et al. 2009a). Comparative studies between colonies of
varying sizes and in different oceanographic regimes could provide further information
regarding factors that may influence partitioning of labor between the sexes during
foraging.
Oceanographic habitat use
In this study, direct measures of dynamic oceanographic topography such as SSH
and velocity were most often associated with foraging behavior, suggesting that search
and detection of prey was influenced by physical oceanographic processes to some
extent. SSH is a measure of topography of the ocean’s surface as influenced by tidal
forces, ocean circulation, and salinity, therefore conditions may vary widely based on
geographic location. Ocean currents circulate around and within peaks and troughs
created by areas of high and low SSH resulting in formation of small and large-scale
features of ocean surface topography, including eddies, currents, and gyres. The Gulf of
Mexico is a topographically dynamic ecosystem dominated by frequent mixing
(Longhurst 2010), and the connection between seabird foraging and SSH may therefore
be geographically and species specific, though little is known.
SSH was strongly linked to foraging movements of gray-headed albatross in the
dynamic south Indian ocean (Nel et al. 2001), and several possible mechanisms may link

76
SSH and velocity nonlinearly to masked booby foraging behavior. First, the physical
motion of horizontal currents creates habitat that influences the distribution of prey.
Second, boobies appear to possess color and near-ultraviolet vision (Reed 1987) which
suggests they can see currents and fronts, and they may use visual cues such as areas of
low turbulence and slow-moving currents to locate prey. Fish may colonize areas with
turbulence regimes (e.g. areas where SSH is low) to which they are adapted, or surface
currents may passively transport and collect fish eggs, larvae, and adults. In flying fish
(family Exocoetidae), a major prey item for masked boobies, turbulence and currents
such as those associated with areas of lower velocity may influence spawning and
dynamic suspension of eggs or larvae within the water column (Stevens et al. 2003).
Habitat preferences and local abundance of predatory fish (e.g. tuna) and dolphins
(family Delphinidae) that drive prey to the surface (i.e., within reach of plunge-diving
seabirds) may also influence the link between dynamic topography and masked boobies.
For instance, thermocline depth, which is dependent on dynamic processes and influences
the distribution of subsurface predators, was strongly linked to abundance of boobies,
tropicbirds, petrels, and storm-petrels in the eastern tropical Pacific (Au and Pitman 1986,
Vilchis et al. 2006). Data from fisheries and plankton tows would provide insight into the
hypothetical connections between regional currents, fish distribution, and seabird habitat
use. The relationship between masked booby foraging behavior and oceanographic
habitat characteristics may not be consistent within and between systems, and other
factors aside from SSH and velocity may affect ARS. These could include biological or
visual cues that indicate the presence of prey. Visual flock recruitment in Cape gannets,

77
for example, significantly reduces the amount of time individual birds spend searching
for prey (Thiebault et al. 2014). Individuals may also develop a framework for locating
prey by learning from experience gained during foraging trips and may therefore
remember and anticipate the location of productive foraging habitat within and between
years (Irons 1998).
Seabirds may be influenced by large-scale features and processes related to SST,
such as the North Pacific Transition Zone (Kappes et al. 2010), the Atlantic Gulf Stream
(Haney 1986), or mesoscale convergence zones in the Indian Ocean (Weimerskirch et al.
2004). SST has been linked to seabird habitat use at multiple time scales (i.e., daily,
annual, decadal), in temperate and tropical ecosystems, and is widely used to evaluate
habitat use and distribution of seabirds (Tremblay et al. 2009). Nonetheless, SST was not
a strong or consistent predictor of masked booby foraging behavior relative to other
variables during this study, even when considered at multiple temporal scales (daily, 8-
day, and monthly). Little is known about how SST affects seabirds in the Gulf of Mexico,
however, frequent mixing by the loop current and associated eddies may limit the long-
term persistence of large-scale SST features, leading to an ecosystem that is highly
dynamic and resulting in a weak or inconsistent relationship between SST and foraging
behavior. In this study, direct measures of dynamic oceanographic topography predicted
foraging behavior more often than habitat descriptors such as SST. SST is a complex
measurement which is influenced by dynamic topography and integrates factors including
solar heating, runoff, and precipitation (Longhurst 2010) that may not necessarily be
linked to prey. It is similarly possible that a time-lag, other than those considered in this

78
study, and intrinsic to the chain of production in the Gulf of Mexico, from nutrient
mixing to prey maturation, lead to a temporal mismatch between SST and foraging
locations of masked boobies which correspond to prey presence (Grémillet et al. 2008,
Suryan et al. 2012, Sommerfeld 2013).
Scales of ARS
The nested ARS scales observed within individual foraging trips are consistent
with hierarchical organization of patches within a given environment and multiscale
search behavior of foraging seabirds. For example, Kotliar and Wiens (1990) described a
framework for understanding the scales at which patches occur within a given landscape
in which individual fish are distributed within fine-scale aggregations of prey, which are
distributed within coarse-scale patches, which are in turn nested within large-scale
regions. Additionally, Fauchald and Tveraa (2006) predicted that foraging seabirds would
search for prey at multiple scales, following the structure of nested patches within the
environment. For example, large scales of 10 - 40 km may correspond to the farthest
distance at which masked boobies can search. Haney et al. (1992) provide evidence that
such a range would correspond closely to the radius of visibility for a seabird from a
given altitude. Scales between 1 and 10 km may correspond to search effort within an
area of suitable habitat where prey encounter is likely. Small search scales < 1 km may
correspond to the approximate size of the moving prey patch as fish are chased by
predators and try to escape capture. Tremblay et al. (2014) suggest relatively similar
scales of movement based on empirical observations of foraging behavior in Cape

79
gannets. During foraging, each nested scale may correspond to a different set of sensory
cues which are used stepwise with changing search scales to locate prey.
FPT analysis did not identify a consistent scale of search for all birds, suggesting
that birds with no determinable scale either did not forage or adjusted the amount of time
spent at each scale of search evenly and continuously during foraging trips. Conversely,
birds that searched at multiple nested scales may have done so concurrently, integrating
multiple environmental and sensory cues. Foraging seabirds may, for example, scan the
proximate eddy for prey and at the same time search the distant horizon for conspecifics.
Large gaps exist in scientific understanding of seabird perception and sensing of the
environment and prey (but see Nevitt et al. 2008, Machovsky-Capuska et al. 2012).
Reserve use
Masked boobies nesting at Isla Muertos did not forage regularly within the park
buffer zone, suggesting that critical habitat for this species may not be protected to the
extent intended during configuration of the park. The objectives for managing AANP
include protecting locally important species and the community-level ecological
interactions with which they are associated (Protegidas 2007). Protection of land and
management of waters close to the breeding colony may not entirely address the breadth
of ecological interactions related to masked booby reproduction and maintenance of the
population, especially since reproductive performance and population dynamics are
strongly linked to resource availability in boobies (Ancona et al. 2012, Bertrand et al.
2012, Weimerskirch et al. 2012, Danckwerts et al. 2014) and in many other species of

80
seabirds. Furthermore, the extent to which foraging habitat of masked boobies within and
around the park may be influenced by climate change, fisheries interactions, or energy
development remains unclear. Finally, understanding variation in behavior among
individuals may enhance understanding of biological systems, leading to the creation of
models that explain population activity by integrating individual components, and
improving management plans by considering the range of behavior within a population
(Bolnick et al. 2003).

81
CONCLUSIONS
This thesis examined movement patterns of seabirds at a range of spatial and
temporal scales. Habitat use in seabirds may include fine-scale features of the
environment as birds search for and capture prey, as well as coarse-scale occupancy of
distinct environmental regions. Understanding spatial ecology at multiple scales is
important because movement patterns may impact animal life histories in both the short-
term and long-term, influencing, for example, reproduction and survival of individuals,
and population dynamics.
Results from chapter two demonstrated that brown pelicans exhibit substantial
variability in movement patterns and are capable of long-distance migration.
Approximately 25% of individuals remain near the breeding colony year-round, which
may indicate that birds are territorial. At coarse scales, brown pelicans occupied and
moved between tropical and temperate ecoregions, and multiple individuals used the
Chesapeake Bay/Delmarva Peninsula region during prebreeding and postbreeding
dispersal. At fine scales, brown pelicans primarily used nearshore pelagic habitat outside
of protected areas, and offshore habitat within 10 km of the coast; further and more
detailed study would improve understanding of potential overlap between brown pelican
movements and future offshore wind energy development.
Results from chapter three demonstrated that breeding masked boobies initiate
foraging trips that range widely in distance, duration, and direction from the colony.
During foraging trips, birds used multiple search radii ranging in size from 30 km to 60

82
m, which suggests that birds perceive large-scale features of the environment as well as
small-scale features that may be directly related to individual schools of prey.
Accordingly, fine-scale foraging behavior corresponded with a variety of oceanographic
conditions, suggesting that individual masked boobies use a range of environmental cues
to locate prey; further study may improve understanding of whether foraging strategies
that appear to vary among individuals are consistent within individuals.
Taken together, the two data chapters included in this thesis represent a
substantial enhancement to our understanding of movement patterns of Pelecaniformes in
the north Atlantic system. The results presented in this thesis can provide valuable
information for marine spatial planning efforts and serve as a baseline for anthropogenic
based threats such as development, pollution, and commercial fisheries.

83
APPENDICES

84
Appendix A. Individual satellite-tag functionality and latitudinal movements for brown
pelicans satellite tagged in South Carolina, USA, 2010 - 2013. Raw data were cleaned by
retaining only locations with accuracies ≤ 1.5 km and applying a speed filter of 25 m/s.
Bird ID PTT mass (g) Stop Date Duration (days) # Filtered Locations Latitudinal Extent (km)
268 110 May-11 178 409 15.1
269 110 Jun-11 191 429 58.7
270 110 Apr-12 486 493 250.7
271 110 Feb-13 794 728 137.5
272 110 Dec-12 754 814 607.8
273 110 May-11 175 394 25.1
274 110 Mar-12 475 594 297.9
275 110 Feb-11 61 151 123.5
276 110 Jul-11 223 426 1082.0
277 110 Dec-10 17 65 4.3
278 110 Mar-12 454 768 193.7
279 110 Oct-11 306 604 663.7
280 110 Jan-11 42 205 62.9
281 110 Oct-12 668 793 992.6
282 110 Feb-13 807 978 525.5
283 110 Jan-11 39 106 30.9
284 110 Feb-13 783 680 1034.4
285 110 Feb-11 59 254 27.9
286 110 Jan-11 16 92 64.1
287 110 Mar-13 819 872 333.3
288 110 Feb-13 795 907 631.8
289 110 Mar-11 86 328 267.4
290 110 Jun-13 916 981 1443.3
291 110 Apr-11 106 308 1831.2
292 110 Jun-13 915 746 894.2
293 68 Mar-12 457 440 942.5
294 68 Apr-11 124 314 362.1
295 68 Dec-10 15 39 560.4
296 68 Jan-11 28 67 10.0
297 68 Dec-10 12 34 171.9
298 68 Jan-12 408 462 62.3
299 68 Feb-11 61 121 393.2
300 68 Feb-12 438 409 759.6
301 68 Jun-12 547 435 1102.9

85
302 68 Apr-12 481 401 791.9
303 68 Aug-12 612 220 312.9
304 68 Mar-12 439 414 773.5
305 68 Feb-11 71 158 16.7
306 68 Jan-11 35 97 25.3
307 68 Feb-12 420 306 602.6

86
Appendix B. Individual breeding locations, focal region, and home ranges (in km2) for
brown pelicans satellite tagged in South Carolina, USA, 2010 - 2013. Mean percent
overlap between successive breeding or successive nonbreeding seasons was calculated
for each individual. OHCI = Old House Channel Island. A hyphen (-) indicates that the
satellite tag stopped transmitting before enough locations could be collected to estimate
the parameter of interest.
ID Breeding Colony
Focal Region (50UD) Home Range (95UD) Mean Home Range Overlap
BR NB BR NB Between BR
Seasons
Between NB
Seasons
268 Crab Bank, SC 260.9 276.8 996.2 1071.2 - -
269 Marsh Is, SC 419.0 238.0 1964.7 1060.1 - -
270 Crab Bank, SC 201.6 325.3 1112.8 1606.8 - 0.72
271 Crab Bank, SC 303.4 192.8 1986.5 998.3 0.57 0.86
272 Beacon Is, NC 994.3 318.9 5540.4 2003.1 0.71 0.63
273 Crab Bank, SC 185.8 196.5 798.5 847.0 - -
274 Marsh Is, SC 759.0 812.4 4421.6 3588.6 - 0.79
275 - - 689.2 - 3603.8 - -
276 Pelican Is, NC 515.9 307.2 4686.5 6186.3 - -
277 - - 160.5 - 700.3 - -
278 Crab Bank, SC 211.5 501.0 1006.1 3711.8 - 0.64
279 Beacon Is, NC - 624.2 - 4863.2 - 0.45
280 - - 808.2 - 2934.0 - -
281 Beacon Is, NC 2308.7 701.2 11587.8 5087.4 0.42 0.23
282 OHCI, NC 530.8 379.2 2738.6 2063.0 0.43 0.72
283 - - 398.6 - 1793.6 - -
284 OHCI, NC;
Wreck Is, VA
482.3 674.6 3338.5 5011.2 0.10 0.49
285 - - 370.9 - 1608.1 - -
286 - - 907.9 - 3772.6 - -
287 Crab Bank, SC 539.6 220.5 3195.2 990.0 0.73 0.93
288 New Dump Is, NC 1343.1 598.9 7130.6 4651.0 0.45 0.59
289 - - 412.4 - 2363.8 - -
290 Fisherman's Is, VA 428.9 463.5 2805.8 5818.9 0.83 0.31
291 - - 416.0 - 2199.9 - -
292 Pelican Is, NC 581.8 332.7 3084.8 2767.2 0.56 0.44
293 Pelican Is, NC 810.4 698.7 4103.4 4680.8 - 0.28
294 Did not breed 600.9 329.4 4627.2 1818.5 - -
295 - - 243.6 - 1339.7 - -
296 - - 179.9 - 769.5 - -
297 - - 689.2 - 3360.2 - -

87
298 Marsh Is, SC 340.0 484.4 1794.8 1864.2 - 0.92
299 - - 202.1 - 1395.0 - -
300 New Dump Is, NC 950.7 393.1 4888.1 3143.0 - 0.31
301 Wreck Is, VA 713.6 557.6 3495.6 6391.4 0.70 0.19
302 Marsh Is, SC 648.3 746.9 2755.4 5465.5 - 0.49
303 Marsh Is, SC 511.9 331.2 2348.2 2019.7 - 0.26
304 Smith Is, MD 478.3 1128.9 2429.2 5981.8 - 0.37
305 - - 227.0 - 1010.3 - -
306 - - 348.8 - 1608.1 - -
307 OHCI, NC 539.6 241.0 4463.2 1769.1 - 0.46

88
Appendix C. Description of individual migration strategies and dispersal locations for
brown pelicans satellite tagged in South Carolina, USA, 2010 - 2013. Birds used six
distinct ecoregions. Proportions were calculated using the range-wide 95% UD for each
individual. Only parks and protected areas up to 50 km inland containing marine,
estuarine, swamp, or riverine habitat were considered. A hyphen (-) indicates that the
satellite tag stopped transmitting before enough locations could be collected to estimate
the parameter of interest. NA indicates that a dispersal location was not identified. Chspk
Bay = Chesapeake Bay
ID Migration
strategy
Dispersal location
#
Ecoregions
utilized
Prop of HR
in SC
Prop of SC
HR in SC
parks
Pre-breeding Post-breeding
268 Stationary NA NA 1 - -
269 Stationary NA NA 1 - -
270 Stationary NA NA 2 1.00 0.01
271 Stationary NA NA 1 1.00 0.11
272 Semi-migratory Chspk Bay, MD/VA Chspk Bay, MD/VA 2 0.24 0.00
273 Stationary NA NA 1 - -
274 Semi-migratory Onslow Bay, NC NA 2 0.87 0.25
275 - NA NA 1 - -
276 Migratory Chspk Bay, MD/VA NA 3 - -
277 - NA NA 1 - -
278 Stationary NA NA 1 0.66 0.15
279 Semi-migratory NA NA 2 - -
280 - NA NA 1 - -
281 Migratory NA NA 2 0.13 0.17
282 Semi-migratory NA Chspk Bay, MD 2 0.32 0.01
283 - NA NA 1 - -
284 Migratory NA NA 3 0.06 0.14
285 - NA NA 1 - -
286 - NA NA 1 - -
287 Stationary NA NA 1 1.00 0.16
288 Semi-migratory NA Chspk Bay, MD/VA 2 0.30 0.14
289 Stationary NA NA 1 - -
290 Highly Migratory NA NA 3 0.10 0.13
291 Highly Migratory NA NA 5 - -
292 Migratory Savannah, GA Pamlico Sound, NC 3 0.15 0.23
293 Migratory NA NA 3 0.20 0.14
294 Stationary Pamlico Sound, NC NA 2 - -

89
295 - NA NA 2 - -
296 - NA NA 1 - -
297 - NA NA 1 - -
298 Stationary NA NA 1 1.00 0.38
299 - NA NA 1 - -
300 Semi-migratory NA Chspk Bay, MD/VA 2 0.33 0.14
301 Migratory NA NA 3 0.12 0.33
302 Semi-migratory NA NA 2 0.46 0.19
303 Stationary NA NA 1 - -
304 Semi-migratory NA NA 2 0.48 0.13
305 - NA NA 1 - -
306 - NA NA 1 - -
307 Semi-migratory NA Chspk Bay, MD/VA 2 0.27 0.02

90
REFERENCES
Adams, J., C. MacLeod, R. M. Suryan, K. D. Hyrenbach, and J. T. Harvey. 2012.
Summer-time use of west coast US National Marine Sanctuaries by migrating
sooty shearwaters (Puffinus griseus). Biological Conservation 156:105-116.
Aebischer, N. 1995. Philopatry and colony fidelity of Shags Phalacrocorax aristotelis on
the east coast of Britain. Ibis 137:11-18.
Aebischer, N. J., P. A. Robertson, and R. E. Kenward. 1993. Compositional analysis of
habitat use from animal radio-tracking data. Ecology 74:1313-1325.
Agostinelli, C., and U. Lund. 2013. R package 'circular': Circular Statistics (version 0.4-
7).
Ainley, D. G., C. A. Ribic, and E. Woehler. 2012. Adding the ocean to the study of
seabirds: A brief history of at-sea seabird research. Marine Ecology Progress
Series 451:231-243.
Amante, C., and B. W. Eakins. 2009. ETOPO1 1 arc-minute global relief model:
procedures, data sources and analysis. US Department of Commerce, National
Oceanic and Atmospheric Administration, National Environmental Satellite, Data,
and Information Service, National Geophysical Data Center, Marine Geology and
Geophysics Division.
Ancona, S., I. Calixto-Albarrán, and H. Drummond. 2012. Effect of El Niño on the diet
of a specialist seabird, Sula nebouxii, in the warm eastern tropical Pacific. Marine
Ecology Progress Series 462:261-271.
Anderson, C. I., and P. G. Rodhouse. 2001. Life cycles, oceanography and variability:
ommastrephid squid in variable oceanographic environments. Fisheries Research
54:133-143.
Anderson, D. J. 2009. The Masked Booby (Sula dactylatra). Page Retrieved from the
Birds of North America Online in A. Poole, editor. The Birds of North America
Online. Cornell Lab of Ornithology, Ithaca.
Asseid, B., L. Drapeau, R. Crawford, B. Dyer, A. Hija, A. Mwinyi, P. Shinula, and L.
Upfold. 2006. The food of three seabirds at Latham Island, Tanzania, with
observations on foraging by masked boobies Sula dactylatra. African Journal of
Marine Science 28:109-114.
Au, D. W., and R. L. Pitman. 1986. Seabird interactions with dolphins and tuna in the
eastern tropical Pacific. Condor 88:304-317.

91
Aubry, L. M., R. F. Rockwell, E. G. Cooch, R. W. Brook, C. P. Mulder, and D. N.
Koons. 2013. Climate change, phenology, and habitat degradation: Drivers of
gosling body condition and juvenile survival in lesser snow geese. Global change
biology 19:149-160.
Barraquand, F., and S. Benhamou. 2008. Animal movements in heterogeneous
landscapes: identifying profitable places and homogeneous movement bouts.
Ecology 89:3336-3348.
Bertrand, S., R. Joo, C. Arbulu Smet, Y. Tremblay, C. Barbraud, and H. Weimerskirch.
2012. Local depletion by a fishery can affect seabird foraging. Journal of Applied
Ecology 49:1168-1177.
Boersma, P., J. Clark, and N. Hillgarth. 2001. Seabird conservation. Pages 559-580 in E.
Schreiber and J. Burger, editors. Biology of Marine Birds. CRC Press LLC, Boca
Raton, FL.
Bolnick, D. I., R. Svanbäck, J. A. Fordyce, L. H. Yang, J. M. Davis, C. D. Hulsey, and
M. L. Forister. 2003. The ecology of individuals: incidence and implications of
individual specialization. The American Naturalist 161:1-28.
Breaker, L. C., T. P. Mavor, and W. W. Broenkow. 2005. Mapping and monitoring large-
scale ocean fronts off the California Coast using imagery from the GOES-10
geostationary satellite. California Sea Grant College Program.
Breiman, L. 2001. Random forests. Machine learning 45:5-32.
Briggs, K. T., W. B. Tyler, D. B. Lewis, P. R. Kelly, and D. A. Croll. 1983. Brown
Pelicans in central and northern California. Journal of Field Ornithology 54:353-
373.
Brinker, D. F., J. M. McCann, B. Williams, and B. D. Watts. 2007. Colonial-nesting
seabirds in the Chesapeake Bay region: Where have we been and where are we
going? Waterbirds 30:93-104.
Buckley, N. J. 1997. Spatial-concentration effects and the importance of local
enhancement in the evolution of colonial breeding in seabirds. The American
Naturalist 149:1091-1112.
Cadiou, B., J. Y. Monnat, and E. Danchin. 1994. Prospecting in the kittiwake, Rissa
tridactyla: different behavioural patterns and the role of squatting in recruitment.
Animal Behaviour 47:847-856.
Cairns, D. K. 1992. Bridging the gap between ornithology and fisheries science: Use of
seabird data in stock assessment models. Condor 94:811-824.

92
Calenge, C. 2006. The package adehabitat for the R software: a tool for the analysis of
space and habitat use by animals. Ecological Modelling 197:516-519.
Calenge, C. 2011a. Analysis of Animal Movements in R: the adehabitatLT Package. R
Foundation for Statistical Computing, Vienna.
Calenge, C. 2011b. Home range estimation in R: the adehabitatHR package. The
Comprehensive R Archive Network.
Catry, T., J. A. Ramos, M. Le Corre, and R. A. Phillips. 2009. Movements, at-sea
distribution and behaviour of a tropical pelagic seabird: the wedge-tailed
shearwater in the western Indian Ocean. Marine Ecology-Progress Series
391:231-242.
Chaurand, T., and H. Weimerskirch. 1994. The regular alternation of short and long
foraging trips in the Blue Petrel Halobaena caerulea - a previously undescribed
strategy of food provisioning in a pelagic seabird. Journal of Animal Ecology
63:275-282.
Chivers, L. S., M. G. Lundy, K. Colhoun, S. F. Newton, J. D. Houghton, and N. Reid.
2012. Foraging trip time-activity budgets and reproductive success in the black-
legged kittiwake. Marine Ecology Progress Series 456:269-277.
Chávez, E. A., and E. Hidalgo. 1988. Los arrecifes coralinos del Caribe noroccidental y
Golfo de México en el contexto socioeconómico. Anales del Instituto de
Ciencias del Mar y Limnología 15:167-176.
Clarke, M. 1977. Beaks, nets and numbers. Symposia of the Zoological Society of
London 38:89-126.
Claus, S., N. De Hauwere, B. Vanhoorne, F. Souza Dias, F. Hernandez, and J. Mees.
2014. Marineregions.org. Flanders Marine Institute.
Cody, M. L. 1985. Habitat Selection in Birds. Academic Press Inc, Orlando, FL.
Croxall, J., P. Rothery, S. Pickering, and P. Prince. 1990. Reproductive performance,
recruitment and survival of wandering albatrosses Diomedea exulans at Bird
Island, South Georgia. The Journal of Animal Ecology 59:775-796.
Croxall, J., P. Trathan, and E. Murphy. 2002. Environmental change and Antarctic
seabird populations. Science 297:1510-1514.
Croxall, J. P., S. H. Butchart, B. Lascelles, A. J. Stattersfield, B. Sullivan, A. Symes, and
P. Taylor. 2012. Seabird conservation status, threats and priority actions: A global
assessment. Bird Conservation International 22:1-34.

93
Croxall, J. P., J. R. Silk, R. A. Phillips, V. Afanasyev, and D. R. Briggs. 2005. Global
circumnavigations: tracking year-round ranges of nonbreeding albatrosses.
Science 307:249-250.
Cummings, J. A., and O. M. Smedstad. 2013. Variational data assimilation for the global
ocean. Pages 303-343 Data Assimilation for Atmospheric, Oceanic and
Hydrologic Applications (Vol. II). Springer.
Cury, P. M., I. L. Boyd, S. Bonhommeau, T. Anker-Nilssen, R. J. Crawford, R. W.
Furness, J. A. Mills, E. J. Murphy, H. Österblom, and M. Paleczny. 2011. Global
seabird response to forage fish depletion—one-third for the birds. Science
334:1703-1706.
Cushing, D. 1990. Plankton production and year-class strength in fish populations: an
update of the match/mismatch hypothesis. Advances in marine biology 26:249-
293.
Cutler, D. R., T. C. Edwards Jr, K. H. Beard, A. Cutler, K. T. Hess, J. Gibson, and J. J.
Lawler. 2007. Random forests for classification in ecology. Ecology 88:2783-
2792.
Cézilly, F., F. Dubois, and M. Pagel. 2000. Is mate fidelity related to site fidelity? A
comparative analysis in Ciconiiforms. Animal Behaviour 59:1143-1152.
Danckwerts, D., C. McQuaid, A. Jaeger, G. McGregor, R. Dwight, M. Le Corre, and S.
Jaquemet. 2014. Biomass consumption by breeding seabirds in the western Indian
Ocean: indirect interactions with fisheries and implications for management.
ICES Journal of Marine Science: Journal du Conseil:fsu093.
Davis, L. S., R. G. Harcourt, and C. J. Bradshaw. 2001. The winter migration of Adelie
penguins breeding in the Ross Sea sector of Antarctica. Polar Biology 24:593-
597.
DeCesare, N. J., M. Hebblewhite, M. Bradley, D. Hervieux, L. Neufeld, and M. Musiani.
2013. Linking habitat selection and predation risk to spatial variation in survival.
Journal of Animal Ecology.
Deneubourg, J.-L., J. M. Pasteels, and J.-C. Verhaeghe. 1983. Probabilistic behaviour in
ants: a strategy of errors? Journal of Theoretical Biology 105:259-271.
Dias, M. P., J. P. Granadeiro, R. A. Phillips, H. Alonso, and P. Catry. 2010. Breaking the
routine: individual Cory's shearwaters shift winter destinations between
hemispheres and across ocean basins. Proceedings of the Royal Society B:
Biological Sciences:rspb20102114.
Duong, T. 2007. ks: Kernel density estimation and kernel discriminant analysis for
multivariate data in R. Journal of Statistical Software 21:1-16.

94
Durell, S. E. A. L. V. d. 2000. Individual feeding specialisation in shorebirds: population
consequences and conservation implications. Biological Reviews of the
Cambridge Philosophical Society 75:503-518.
eBird. 2012. eBird: An online database of bird distribution and abundance. eBird, Ithaca,
New York.
El-Hacen, E.-H. M., O. Overdijk, T. Lok, H. Olff, and T. Piersma. 2013. Home range,
habitat selection, and foraging rhythm in Mauritanian Spoonbills (Platalea
leucorodia balsaci): a satellite tracking study. Waterbirds 36:277-286.
ESRI. 2014. ArcMap 10.2. Environmental Systems Resource Institute, Redlands,
California.
Evers, D., P. G. R. Jodice, P. C. Frederick, L. Eggert, K. D. Meyer, M. Yates, C. S.
Flegel, J. D. Meattey, M. Duron, J. Goyette, and J. McKay. 2011. Final pre-
assessment data report: Estimating oiling and mortality of breeding colonial
waterbirds from the Deepwater Horizon oil spill, NRDA Deepwater Horizon (MC
252) Oil Spill (Bird Study #4). Page 42. Deepwater Horizon Trustee Council,
Fairhope, AL.
Fauchald, P., and T. Tveraa. 2003. Using first-passage time in the analysis of area-
restricted search and habitat selection. Ecology 84:282-288.
Fauchald, P., and T. Tveraa. 2006. Hierarchical patch dynamics and animal movement
pattern. Oecologia 149:383-395.
Fox, A., M. Desholm, J. Kahlert, T. K. Christensen, and I. Krag Petersen. 2006.
Information needs to support environmental impact assessment of the effects of
European marine offshore wind farms on birds. Ibis 148:129-144.
Freitas, C., K. M. Kovacs, R. A. Ims, M. A. Fedak, and C. Lydersen. 2008. Ringed seal
post-moulting movement tactics and habitat selection. Oecologia 155:193-204.
Fronhofer, E. A., T. Hovestadt, and H. J. Poethke. 2013. From random walks to informed
movement. Oikos 122:857-866.
Furness, R. W., and M. L. Tasker. 2000. Seabird-fishery interactions: Quantifying the
sensitivity of seabirds to reductions in sandeel abundance, and identification of
key areas for sensitive seabirds in the North Sea. Marine Ecology Progress Series
202:253-264.
Furness, R. W., H. M. Wade, and E. A. Masden. 2013. Assessing vulnerability of marine
bird populations to offshore wind farms. Journal of environmental management
119:56-66.

95
Gill, J. A., K. Norris, P. M. Potts, T. G. Gunnarsson, P. W. Atkinson, and W. J.
Sutherland. 2001. The buffer effect and large-scale population regulation in
migratory birds. Nature 412:436-438.
Golet, G. H., K. J. Kuletz, D. D. Roby, and D. B. Irons. 2000. Adult prey choice affects
chick growth and reproductive success in pigeon guillemots. The Auk 117:82-91.
Goss-Custard, J., R. Clarke, and S. l. V. Dit Durell. 1984. Rates of food intake and
aggression of oystercatchers Haematopus ostralegus on the most and least
preferred mussel Mytilus edulis beds of the Exe estuary. The Journal of Animal
Ecology 53:233-245.
Grémillet, D., G. Dell'Omo, P. G. Ryan, G. Peters, Y. Ropert-Coudert, and S. J. Weeks.
2004. Offshore diplomacy, or how seabirds mitigate intra-specific competition: a
case study based on GPS tracking of Cape gannets from neighbouring colonies.
Marine Ecology Progress Series 268:265-279.
Grémillet, D., S. Lewis, L. Drapeau, C. D. van Der Lingen, J. A. Huggett, J. C. Coetzee,
H. M. Verheye, F. Daunt, S. Wanless, and P. G. Ryan. 2008. Spatial match–
mismatch in the Benguela upwelling zone: Should we expect chlorophyll and sea‐surface temperature to predict marine predator distributions? Journal of Applied
Ecology 45:610-621.
Guilford, T., J. Meade, J. Willis, R. A. Phillips, D. Boyle, S. Roberts, M. Collett, R.
Freeman, and C. Perrins. 2009. Migration and stopover in a small pelagic seabird,
the Manx shearwater Puffinus puffinus: Insights from machine learning.
Proceedings of the Royal Society B: Biological Sciences 276:1215-1223.
Hamer, K., E. Humphreys, S. Garthe, J. Hennicke, G. Peters, D. Grémillet, R. Phillips,
M. Harris, and S. Wanless. 2007. Annual variation in diets, feeding locations and
foraging behaviour of gannets in the North Sea: flexibility, consistency and
constraint. Marine Ecology Progress Series 338:295-305.
Haney, J. C. 1986. Seabird affinities for Gulf Stream frontal eddies: responses of mobile
marine consumers to episodic upwelling. Journal of Marine Research 44:361-384.
Haney, J. C., K. M. Fristrup, and D. S. Lee. 1992. Geometry of visual recruitment by
seabirds to ephemeral foraging flocks. Ornis Scandinavica 23:49-62.
Harris, M. P., and S. Wanless. 1989. Fall colony attendance and breeding success in the
Common Murre. Condor 91:139-146.
Hatch, S. A., V. A. Gill, and D. M. Mulcahy. 2010. Individual and colony-specific
wintering areas of Pacific northern fulmars (Fulmarus glacialis). Canadian Journal
of Fisheries and Aquatic Sciences 67:386-400.

96
Haury, L., J. McGowan, and P. Wiebe. 1978. Patterns and processes in the time-space
scales of plankton distributions. Pages 277-327 Spatial pattern in plankton
communities. Springer.
Hijmans, R. J., and J. van Etten. 2012. raster: Geographic analysis and modeling with
raster data. R package version 2.0-12.
Hutto, R. L. 1985. Habitat selection by nonbreeding, migratory land birds. Pages 455-476
in M. L. Cody, editor. Habitat Selection in Birds. Academic Press Inc., Orlando,
FL.
Hyrenbach, K. D., P. Fernández, and D. J. Anderson. 2002. Oceanographic habitats of
two sympatric North Pacific albatrosses during the breeding season. Marine
Ecology Progress Series 233:283-301.
Irons, D. B. 1998. Foraging area fidelity of individual seabirds in relation to tidal cycles
and flock feeding. Ecology 79:647-655.
IUCN, and UNEP-WCMC. 2014. The World Database on Protected Areas (WDPA).
UNEP - WCMC, Cambridge, UK.
James, G. D., and J. C. Stahl. 2000. Diet of southern Buller's albatross (Diomedea bulleri
bulleri) and the importance of fishery discards during chick rearing. New Zealand
Journal of Marine and Freshwater Research 34:435-454.
Janmaat, K. R., S. D. Ban, and C. Boesch. 2013. Chimpanzees use long-term spatial
memory to monitor large fruit trees and remember feeding experiences across
seasons. Animal Behaviour 86:1183-1205.
Jodice, P., J. Tavano, and W. Mackin. 2013. Chapter 8: Marine and coastal birds and
bats. Pages Study BOEM 2013-01157 in J. Michel, editor. South Atlantic
information resources: data search and literature synthesis. U.S. Department of
the Interior, Bureau of Ocean Energy Management, Regulation, and
Enforcement, Gulf of Mexico OCS Region, New Orleans, LA.
Jodice, P. G., T. M. Murphy, F. J. Sanders, and L. M. Ferguson. 2007. Longterm trends in
nest counts of colonial seabirds in South Carolina, USA. Waterbirds 30:40-51.
Jodice, P. G., and R. M. Suryan. 2010. The transboundary nature of seabird ecology.
Pages 139-165 Landscape-scale Conservation Planning. Springer Verlag.
Jodice, P. G., L. C. Wickliffe, and E. B. Sachs. 2011. Seabird use of discards from a
nearshore shrimp fishery in the South Atlantic Bight, USA. Marine biology
158:2289-2298.
Johnson, D. H. 1980. The comparison of usage and availability measurements for
evaluating resource preference. Ecology 61:65-71.

97
Kai, E. T., V. Rossi, J. Sudre, H. Weimerskirch, C. Lopez, E. Hernandez-Garcia, F.
Marsac, and V. Garçon. 2009. Top marine predators track Lagrangian coherent
structures. Proceedings of the National Academy of Sciences 106:8245-8250.
Kappes, M. A., S. A. Shaffer, Y. Tremblay, D. G. Foley, D. M. Palacios, P. W. Robinson,
S. J. Bograd, and D. P. Costa. 2010. Hawaiian albatrosses track interannual
variability of marine habitats in the North Pacific. Progress in Oceanography
86:246-260.
Kappes, M. A., H. Weimerskirch, D. Pinaud, and M. Le Corre. 2011. Variability of
resource partitioning in sympatric tropical boobies. Marine Ecology Progress
Series 441:281-294.
Kato, A., Y. Watanuki, I. Nishiumi, M. Kuroki, P. Shaughnessy, and Y. Naito. 2000.
Variation in foraging and parental behavior of king cormorants. The Auk
117:718-730.
King, D. T., B. L. Goatcher, J. W. Fischer, J. Stanton, J. M. Lacour, S. C. Lemmons, and
G. Wang. 2013. Home Ranges and Habitat Use of Brown Pelicans (Pelecanus
occidentalis) in the Northern Gulf of Mexico. Waterbirds 36:494-500.
Kokko, H. 1999. Competition for early arrival in migratory birds. Journal of Animal
Ecology 68:940-950.
Kokko, H., M. P. Harris, and S. Wanless. 2004. Competition for breeding sites and site‐dependent population regulation in a highly colonial seabird, the common
guillemot Uria aalge. Journal of Animal Ecology 73:367-376.
Kotliar, N. B., and J. A. Wiens. 1990. Multiple scales of patchiness and patch structure: a
hierarchical framework for the study of heterogeneity. Oikos 59:253-260.
Kristan Iii, W. B. 2003. The role of habitat selection behavior in population dynamics:
Source–sink systems and ecological traps. Oikos 103:457-468.
Kubetzki, U., S. Garthe, D. Fifield, B. Mendel, and R. W. Furness. 2009. Individual
migratory schedules and wintering areas of northern gannets. Mar Ecol Prog Ser
391:257-265.
Lavielle, M. 2005. Using penalized contrasts for the change-point problem. Signal
Processing 85:1501-1510.
Le Corre, M., A. Jaeger, P. Pinet, M. A. Kappes, H. Weimerskirch, T. Catry, J. A.
Ramos, J. C. Russell, N. Shah, and S. Jaquemet. 2012. Tracking seabirds to
identify potential marine protected areas in the tropical western Indian Ocean.
Biological Conservation 156:83-93.

98
Lewison, R., D. Oro, and P. Yorio. 2012. Research priorities for seabirds: Improving
conservation and management in the 21st century. 17:93-121.
Liaw, A., and M. Wiener. 2002. Classification and Regression by randomForest. R News
2:18-22.
Lobel, P., and A. Robinson. 1986. Transport and entrapment of fish larvae by ocean
mesoscale eddies and currents in Hawaiian waters. Deep Sea Research Part A.
Oceanographic Research Papers 33:483-500.
Longhurst, A. R. 2010. Ecological geography of the sea. 2 edition. Academic Press,
Burlington, MA.
Ludynia, K., J. Kemper, and J.-P. Roux. 2012. The Namibian Islands’ marine protected
area: Using seabird tracking data to define boundaries and assess their adequacy.
Biological Conservation 156:136-145.
Machovsky-Capuska, G. E., H. C. Howland, D. Raubenheimer, R. Vaughn-Hirshorn, B.
Würsig, M. E. Hauber, and G. Katzir. 2012. Visual accommodation and active pursuit of
prey underwater in a plunge-diving bird: the Australasian gannet. Proceedings of the
Royal Society B: Biological Sciences:rspb20121519.
Mason, C. R. 1945. Pelican travels. Bird Banding 16:134-143.
Michel, J. 2013. Chapter 1: Introduction. Pages Study BOEM 2013-01157 in J. Michel,
editor. South Atlantic information resources: data search and literature synthesis.
U.S. Department of the Interior, Bureau of Ocean Energy Management,
Regulation, and Enforcement, Gulf of Mexico OCS Region, New Orleans, LA.
Mullers, R. H., and J. M. Tinbergen. 2009. Individual variation in parental provisioning
behaviour predicts survival of Cape gannet chicks under poor conditions. Ardea
97:89-98.
Myers, J. 1981. A test of three hypotheses for latitudinal segregation of the sexes in
wintering birds. Canadian Journal of Zoology 59:1527-1534.
Nel, D., J. Lutjeharms, E. Pakhomov, I. Ansorge, P. Ryan, and N. Klages. 2001.
Exploitation of mesoscale oceanographic features by grey-headed albatross
Thalassarche chrysostoma in the southern Indian Ocean. Marine Ecology Progress
Series 217:15-26.
Nelson, J. B. 1978. The Sulidae: Gannets and boobies. Oxford University Press, Oxford.
Nevitt, G. A., M. Losekoot, and H. Weimerskirch. 2008. Evidence for olfactory search in
wandering albatross, Diomedea exulans. Proceedings of the National Academy of
Sciences 105:4576-4581.

99
Pante, E., and B. Simon-Bouhet. 2013. marmap: A Package for Importing, Plotting and
Analyzing Bathymetric and Topographic Data in R PLoS ONE 8:e73051.
Paruk, J. D., D. Long IV, S. L. Ford, and D. C. Evers. 2014. Common Loons (Gavia
immer) wintering off the Louisiana coast tracked to Saskatchewan during the
breeding season. Waterbirds 37:47-52.
Pattrick, P., and N. Strydom. 2014. Larval fish variability in response to oceanographic
features in a nearshore nursery area. Journal of fish biology 85:857-881.
Perrow, M. R., J. J. Gilroy, E. R. Skeate, and M. L. Tomlinson. 2011. Effects of the
construction of Scroby Sands offshore wind farm on the prey base of Little tern
Sternula albifrons at its most important UK colony. Marine pollution bulletin
62:1661-1670.
Perry, M. C., A. M. Wells-Berlin, D. M. Kidwell, and P. C. Osenton. 2007. Temporal
changes of populations and trophic relationships of wintering diving ducks in
Chesapeake Bay. Waterbirds 30:4-16.
Phillips, R. A., J. R. Silk, J. P. Croxall, V. Afanasyev, and V. J. Bennett. 2005. Summer
distribution and migration of nonbreeding albatrosses: individual consistencies
and implications for conservation. Ecology 86:2386-2396.
Pichegru, L., P. G. Ryan, C. D. van der Lingen, J. Coetzee, Y. Ropert-Coudert, and D.
Grémillet. 2007. Foraging behaviour and energetics of Cape gannets Morus
capensis feeding on live prey and fishery discards in the Benguela upwelling
system. Marine Ecology Progress Series 350:127-136.
Portha, S., J.-L. Deneubourg, and C. Detrain. 2002. Self-organized asymmetries in ant
foraging: a functional response to food type and colony needs. Behavioral
Ecology 13:776-781.
Preatoni, D. G., M. Spada, L. A. Wauters, G. Tosi, and A. Martinoli. 2011. Habitat use in
the female Alpine long-eared bat (Plecotus macrobullaris): does breeding make
the difference? Acta Chiropterologica 13:355-364.
Protegidas, C. N. d. Á. N. 2007. Programa de conservación y manejo parque nacional
Arrecife Alacranes. Page 169 in C. N. d. Á. N. Protegidas, editor. SEPRIM,
Camino al Ajusco No. 200, Col. Jardines en la Montaña, Tlalpan C.P. 14210,
México, D. F.
Pärt, T., D. Arlt, B. Doligez, M. Low, and A. Qvarnström. 2011. Prospectors combine
social and environmental information to improve habitat selection and breeding
success in the subsequent year. Journal of Animal Ecology 80:1227-1235.

100
Pérez, C., J. P. Granadeiro, M. P. Dias, H. Alonso, and P. Catry. 2013. When males are
more inclined to stay at home: insights into the partial migration of a pelagic
seabird provided by geolocators and isotopes. Behavioral Ecology:art119.
Péron, C., and D. Grémillet. 2013. Tracking through life stages: Adult, immature and
juvenile autumn migration in a long-lived seabird. PloS one 8:e72713.
Pütz, K., A. Schiavini, A. R. Rey, and B. H. Lüthi. 2007. Winter migration of magellanic
penguins (Spheniscus magellanicus) from the southernmost distributional range.
Marine Biology 152:1227-1235.
Rattner, B. A., and P. C. McGowan. 2007. Potential hazards of environmental
contaminants to avifauna residing in the Chesapeake Bay estuary. Waterbirds
30:63-81.
Reed, J. R. 1987. Scotopic and photopic spectral sensitivities of boobies. Ethology 76:33-
55.
Ricklefs, R. E. 1990. Seabird life histories and the marine environment: Some
speculations. Colonial Waterbirds 13:1-6.
Roberts, J. J., B. D. Best, D. C. Dunn, E. A. Treml, and P. N. Halpin. 2010. Marine
Geospatial Ecology Tools: An integrated framework for ecological geoprocessing
with ArcGIS, Python, R, MATLAB, and C++. Environmental Modelling &
Software 25:1197-1207.
Robertson, G., M. Bolton, W. Grecian, and P. Monaghan. 2014. Inter-and intra-year
variation in foraging areas of breeding kittiwakes (Rissa tridactyla). Marine
biology 161:1973-1986.
Rothenbach, C. A., and J. P. Kelly. 2012. The parental dilemma under variable predation
pressure: adaptive variation in nest attendance by Great Egrets. The Condor
114:90-99.
Ryther, J. H. 1969. Photosynthesis and fish production in the sea. Science 166:72-76.
Santora, J. A., C. S. Reiss, A. M. Cossio, and R. R. Veit. 2009. Interannual spatial
variability of krill (Euphausia superba) influences seabird foraging behavior near
Elephant Island, Antarctica. Fisheries Oceanography 18:20-35.
Sanvicente-Añorve, L., C. Flores-Coto, and X. Chiappa-Carrara. 2000. Temporal and
spatial scales of ichthyoplankton distribution in the Southern Gulf of Mexico.
Estuarine, Coastal and Shelf Science 51:463-475.
Schjorring, S., J. Gregersen, and T. Bregnballe. 1999. Prospecting enhances breeding
success of first-time breeders in the great cormorant, Phalacrocorax carbo
sinensis. Animal Behaviour 57:647-654.

101
Schnell, G. D., and J. J. Hellack. 1978. Flight speeds of Brown Pelicans, Chimney Swifts,
and other birds. Bird-banding 49:108-112.
Schreiber, R. W. 1976. Movements of color marked brown pelicans. Bird Banding
47:101-111.
Schreiber, R. W. 1980. Nesting chronology of the eastern brown pelican. The Auk
97:491-508.
Schreiber, R. W., and E. A. Schreiber. 1982. Essential habitat of the Brown Pelican in
Florida. Florida Field Naturalist 10:9-17.
Shields, M. 2002. Brown Pelican (Pelecanus occidentalis). Page Retrieved from the Birds
of North America Online in A. Poole, editor. The Birds of North America Online.
Cornell Lab of Ornithology, Ithaca.
Sommerfeld, J. 2013. Foraging ecology of Masked Boobies: The importance of intrinsic
and extrinsic factors. University of Tasmania.
Sommerfeld, J., A. Kato, Y. Ropert-Coudert, S. Garthe, and M. A. Hindell. 2013a.
Foraging parameters influencing the detection and interpretation of area-restricted
search behaviour in marine predators: A case study with the masked booby. PloS
one 8:e63742.
Sommerfeld, J., A. Kato, Y. Ropert‐Coudert, S. Garthe, and M. A. Hindell. 2013b. The
individual counts: within sex differences in foraging strategies are as important as
sex‐specific differences in masked boobies Sula dactylatra. Journal of Avian
Biology 44:531-540.
Sorensen, M. C., J. M. Hipfner, T. K. Kyser, and D. R. Norris. 2009. Carry‐over effects
in a Pacific seabird: Stable isotope evidence that pre‐breeding diet quality
influences reproductive success. Journal of Animal Ecology 78:460-467.
Spalding, M. D., H. E. Fox, G. R. Allen, N. Davidson, Z. A. Ferdaña, M. Finlayson, B. S.
Halpern, M. A. Jorge, A. Lombana, and S. A. Lourie. 2007. Marine ecoregions of
the world: A bioregionalization of coastal and shelf areas. BioScience 57:573-
583.
Spear, L. B., L. T. Ballance, and D. G. Ainley. 2001. Response of seabirds to thermal
boundaries in the tropical Pacific: the thermocline versus the Equatorial Front.
Marine Ecology Progress Series 219:275-289.
Staniland, I., and I. Boyd. 2003. Variation in the foraging location of Antarctic fur seals
(Arctocephalus gazella) and the effects on diving behavior. Marine mammal
science 19:331-343.

102
Stefan, S. J. 2008. An analysis of recovery data of brown pelicans (Pelicanus
occidentalis) and royal terns (Sterna maxima) banded in South Carolina. The
College of Charleston.
Stephens, D. W. 1986. Foraging theory. Princeton University Press, Princeton, NJ.
Stevens, P. W., C. K. Bennett, and J. J. Berg. 2003. Flyingfish spawning (Parexocoetus
brachypterus) in the northeastern Gulf of Mexico. Environmental biology of
fishes 67:71-76.
Sumner, M. D. 2013. Trip: Spatial analysis of animal track data. R package version 1.1-
17.
Suryan, R. M., J. A. Santora, and W. J. Sydeman. 2012. New approach for using remotely
sensed chlorophyll a to identify seabird hotspots. Marine Ecology Progress Series
451:213-225.
Suryan, R. M., F. Sato, G. R. Balogh, K. David Hyrenbach, P. R. Sievert, and K. Ozaki.
2006. Foraging destinations and marine habitat use of short-tailed albatrosses: A
multi-scale approach using first-passage time analysis. Deep Sea Research Part II:
Topical Studies in Oceanography 53:370-386.
Suzuki, R., and H. Shimodaira. 2006. Pvclust: an R package for assessing the uncertainty
in hierarchical clustering. Bioinformatics 22:1540-1542.
Switzer, P. V. 1993. Site fidelity in predictable and unpredictable habitats. Evolutionary
Ecology 7:533-555.
Team, R. D. C. 2014. R: a language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria.
Tew Kai, E., S. Benhamou, C. D. Lingen, J. C. Coetzee, L. Pichegru, P. G. Ryan, and D.
Grémillet. 2013. Are Cape gannets dependent upon fishery waste? A multi‐scale
analysis using seabird GPS‐tracking, hydro‐acoustic surveys of pelagic fish and
vessel monitoring systems. Journal of Applied Ecology 50:659-670.
Thiebault, A., R. Mullers, P. Pistorius, M. A. Meza-Torres, L. Dubroca, D. Green, and Y.
Tremblay. 2014. From colony to first patch: Processes of prey searching and
social information in Cape Gannets. The Auk 131:595-609.
Tinbergen, N., M. Impekoven, and D. Franck. 1967. An experiment on spacing-out as a
defence against predation. Behaviour 28:307-321.
Tremblay, Y., S. Bertrand, R. W. Henry, M. A. Kappes, D. P. Costa, and S. A. Shaffer.
2009. Analytical approaches to investigating seabird-environment interactions: a
review. Marine Ecology-Progress Series 391:153-163.

103
Tremblay, Y., A. Thiebault, R. Mullers, and P. Pistorius. 2014. Bird-borne video-cameras
show that seabird movement patterns relate to previously unrevealed proximate
environment, not prey. PloS one 9:e88424.
Van Moorter, B., D. Visscher, S. Benhamou, L. Börger, M. S. Boyce, and J. M. Gaillard.
2009. Memory keeps you at home: a mechanistic model for home range
emergence. Oikos 118:641-652.
Van Valen, L. 1965. Morphological variation and width of ecological niche. American
Naturalist 908:377-390.
Vilchis, L., L. T. Ballance, and P. C. Fiedler. 2006. Pelagic habitat of seabirds in the
eastern tropical Pacific: effects of foraging ecology on habitat selection. Marine
Ecology Progress Series 315:279-292.
Votier, S. C., W. J. Grecian, S. Patrick, and J. Newton. 2011. Inter-colony movements,
at-sea behaviour and foraging in an immature seabird: results from GPS-PPT
tracking, radio-tracking and stable isotope analysis. Marine biology 158:355-362.
Votier, S. C., B. J. Hatchwell, A. Beckerman, R. H. McCleery, F. M. Hunter, J. Pellatt,
M. Trinder, and T. R. Birkhead. 2005. Oil pollution and climate have wide‐scale
impacts on seabird demographics. Ecology Letters 8:1157-1164.
Wakefield, E. D., T. W. Bodey, S. Bearhop, J. Blackburn, K. Colhoun, R. Davies, R. G.
Dwyer, J. Green, D. Grémillet, and A. L. Jackson. 2013. Space Partitioning
Without Territoriality in Gannets. Science (New York, NY) 341:68-70.
Wand, M. P., and M. C. Jones. 1994. Kernel smoothing. Chapman and Hall/CRC Press,
Boca Raton, FL.
Ward, J. H. 1963. Hierarchical grouping to optimize an objective function. Journal of the
American statistical association 58:236-244.
Weimerskirch, H. 2007. Are seabirds foraging for unpredictable resources? Deep Sea
Research Part II: Topical Studies in Oceanography 54:211-223.
Weimerskirch, H., S. Bertrand, J. Silva, C. Bost, and S. Peraltilla. 2012. Foraging in
Guanay cormorant and Peruvian booby, the major guano-producing seabirds in
the Humboldt Current System. Marine Ecology Progress Series 458:231-245.
Weimerskirch, H., S. Bertrand, J. Silva, J. C. Marques, and E. Goya. 2010. Use of social
information in seabirds: compass rafts indicate the heading of food patches. PloS
one 5:e9928.
Weimerskirch, H., Y. Cherel, F. Cuenot-Chaillet, and V. Ridoux. 1997. Alternative
foraging strategies and resource allocation by male and female wandering
albatrosses. Ecology 78:2051-2063.

104
Weimerskirch, H., M. Le Corre, and C. A. Bost. 2008. Foraging strategy of masked
boobies from the largest colony in the world: relationship to environmental
conditions and fisheries. Marine Ecology-Progress Series 362:291-302.
Weimerskirch, H., M. Le Corre, H. Gadenne, D. Pinaud, A. Kato, Y. Ropert-Coudert, and
C.-A. Bost. 2009a. Relationship between reversed sexual dimorphism, breeding
investment and foraging ecology in a pelagic seabird, the masked booby.
Oecologia 161:637-649.
Weimerskirch, H., M. Le Corre, S. Jaquemet, M. Potier, and F. Marsac. 2004. Foraging
strategy of a top predator in tropical waters: great frigatebirds in the Mozambique
Channel. Marine Ecology Progress Series 275:297-308.
Weimerskirch, H., S. A. Shaffer, Y. Tremblay, D. P. Costa, H. Gadenne, A. Kato, Y.
Ropert-Coudert, K. Sato, and D. Aurioles. 2009b. Species-and sex-specific
differences in foraging behaviour and foraging zones in blue-footed and brown
boobies in the Gulf of California. Marine Ecology Progress Series 391:267-278.
Welcker, J., H. Steen, A. M. A. Harding, and G. W. Gabrielsen. 2009. Sex-specific
provisioning behaviour in a monomorphic seabird with a bimodal foraging
strategy. Ibis 151:502-513.
Wickliffe, L. C., and P. G. Jodice. 2010. Seabird attendance at shrimp trawlers in
nearshore waters of South Carolina. Marine Ornithology 38:31-39.
Wood, S. 2006. Generalized additive models: an introduction with R. Chapman and
Hall/CRC press, Boca Raton, FL.
Wood, S., and F. Scheipl. 2013. gamm4: Generalized additive mixed models using mgcv
and lme4. R package version 0.2-2.
Wood, S. N. 2011. Fast stable restricted maximum likelihood and marginal likelihood
estimation of semiparametric generalized linear models Journal of the Royal
Statistical Society B 73:3-36.
Young, H. S., S. A. Shaffer, D. J. McCauley, D. G. Foley, R. Dirzo, and B. A. Block.
2010. Resource partitioning by species but not sex in sympatric boobies in the
central Pacific Ocean. Marine Ecology Progress Series 403:291-301.
Zuur, A., E. N. Ieno, N. Walker, A. A. Saveliev, and G. M. Smith. 2009. Mixed effects
models and extensions in ecology with R. Springer.