validation of a single tube fluorescent multiplex assay
TRANSCRIPT

Indian Journal of Biotechnology
Vol 7, January 2008, pp 41-49
Validation of a single tube fluorescent multiplex assay for simultaneous typing of
20 Y-STR loci
Sanghamitra Sahoo1, 2
and V K Kashyap1, 2*
1National DNA Analysis Centre, Central Forensic Science Laboratory, 30 Gorachand Road, Kolkata, India
Received 12 June 2006; revised 27 April 2007; accepted 23 July 2007
This paper describes a single tube assay for 20 fluorescent labelled Y-STR loci, which is rapid and robust state-of-art
multiplex system. Reaction conditions, including annealing temperature, concentration of primers, magnesium and template
DNA, DNA polymerase and reaction volume, were optimised to yield robust amplification. The assay could withstand
moderate fluctuations in reaction parameters with no affect on haplotype analysis. Sensitivity and robustness of the
multiplex system is revealed from the examination of different matrices and environmental conditions wherein complete
haplotypes could be obtained in all the samples with no failures except those exposed to severe environmental stresses.
Genotyping of the 20 Y-STR loci showed that all loci included in the multiplex are highly polymorphic in the Indian
populations and its inclusion increases the discriminatory power of the marker system. It could be summarized that the
developed Y-STR multiplex assay is a simple, sensitive and robust system highly suitable for concurrent analysis of 20
polymorphic Y-STR loci.
Keywords: multiplex PCR, Y-STR, DNA profiling, validation
Introduction Multiplex polymerase chain reaction (PCR) is a
system in which two or more loci are simultaneously
amplified in the same reaction. This method has been
successfully applied in many areas of DNA typing,
including analyses of deletions, mutations1,2
and
polymorphisms at short tandem repeat (STR) loci3-8
since its first report9. STRs or microsatellites contain
2-6 bp repeat units which are tandemly arranged and
widely dispersed throughout the eukaryotic genome.
Analysis of these tandemly repeated DNA sequences
is of fundamental importance not only in population
biology, such as human identification, understanding
origin of man, migratory histories of populations,
phylogenetic distribution of various plant and animal
species, and other evolutionary processes, but also in
linkage analysis of genetic traits. Multiplex STR
analysis, therefore, has emerged as an indispensable
assay as it requires only sub-nanogram amounts of
DNA, is less labour intensive, provides easy
interpretation of data, high discriminatory power and
reliable results. Although a large number of
multiplexed commercial kits are available for human
autosomal STR markers [Powerplex® 2.1, Powerplex
®
16 system (Promega Corp. Madison, WI), ABI
AmpFl STR® Profiler
™, COfiler
™ and Identifiler
™)],
the same is not the state for the microsatellites of Y-
chromosome. The reason for this lacuna is because
until recent past, the number of STRs known on the Y
chromosome was limited10-14
. In addition, lack of
suitable Y-STR mutiplex system is also another
reason why the Y-chromosome has not been as
extensively used as the autosomal STRs in human
identification and population genetic studies.
Recently, however, three commercial Y-STR
multiplex systems: Y-PLEXTM
6 kit (ReliaGene
Technologies Inc.), Powerplex Y System (Promega
Corp.) and AmpFLSTR Y filer kit (Applied
Biosytems) are available; these include only 6, 12 and
17 Y-STR loci, respectively15,16
. Although the Y
chromosome harbors diversity which is only 20% as
that of the autosomes17
, a large number of Y-STRs
have been identified in recent years, and a few of
them have also been used for human identification,
particularly paternity testing, and evolutionary,
genealogical and anthropological studies18,19
.
_________________
*Author for correspondence:
Tel: 91-120-2400027; Fax: 91-120-2403014
E-mail address: [email protected]
Present Address: 2National Institute of Biologicals, A-32, Sector 62,
Institutional Area, Noida 201 307, India

INDIAN J BIOTECHNOL, JANUARY 2008
42
Majority of the STRs on the Y-chromosome are
located in the non-recombining region and thus are
passed down the generations without recombination.
In addition, because the Y-chromosome is present in a
haploid state, it is exceptional and unparalleled
compared with any of the known markers, for tracing
male movements either of the present-day or events of
the past20
. However, lack of recombination also
suggests that individual markers cannot be combined
by the product rule and to achieve power of
discrimination similar to those of autosomal STR
markers21,22
, a large number of Y-STR loci should be
included in the analysis. The Y-STR markers find
application in a wide range of forensic investigations,
especially in cases where father of the male child is
deceased17,23
, identification of human remains in mass
disasters24
and offenders, particularly in sexual assault
cases25
, where the male component needs to be
identified from the mixed male and female fractions
of the body fluids26
and where methods such as
differential lysis is unsuccessful, as in azoospermic
semen samples. In addition, these STRs are equally
important in evolutionary biology, archeological and
anthropological studies pertaining to male
population17,18,27-29
. Few studies, using Y-STRs, were
primarily based either on monoplexes or multiplexes
carried out as 2-3 tube reactions. Since these are
laborious and time-consuming protocols, single tube
multiplex methods are much in demand30
. The
development of a large multiplex will lead to newer
possibilities for studying which Y-STRs have the
most significant impact on a particular population by
enabling more rapid collection of the data from
population samples. Understanding the genetic
structure of individual populations will help in
maintaining national DNA databases, which is
increasingly essential31-33
for assessing the strength of
match evidence in DNA profiling cases. Therefore,
development of a Y-STR multiplex system is not only
of immense value in justice delivery system34
but also
in understanding the Indian scenario of male
evolutionary and population genetics. We have,
therefore, attempted to develop, standardize and
validate a set of 20 Y-STR loci for a single tube
multiplex reaction that will provide a sensitive,
reproducible, robust and rapid PCR assay.
Materials and Methods Reagents for PCR
10X PCR gold buffer containing 500 mM KCl, 150
mM Tris-HCl, pH 8.3, 25 mM MgCl2 and AmpliTaq
Gold®
DNA polymerase were procured from Applied
Biosystems, Foster City, USA. The nucleotides
(dNTP- 100 mM each) and glycerol was purchased
from Promega Corp. (Madison, WI, USA) and SRL
(Mumbai, India), respectively. Customised
fluorescent labelled and non-fluorescent primers were
procured from Isogen Bioscience BV, The
Netherlands and reconstituted in TE buffer to a final
concentration of 1 nmol/µL each. The working stocks
of all primers were prepared in milli-Q water for a
concentration of 20 pmol/µL.
Selection and Description of Y-STR loci
Our previous standardization experiments included
eight Y-STR loci35
, which constituted the minimal
haplotype required for human identification
recommended by the ISFG guidelines (protocol
unpublished). The “minimal haplotype” includes
tetranucleotide Y-STR markers DYS19, DYS385a/b,
DYS389I/II, DYS390, DYS391, DYS393 and a
trinucleotide, DYS392. However, with the availability
of information on polymorphism for new Y-STR
loci10, 36-40
, we considered their inclusion as an asset in
enhancing the power of discrimination manifold. The
loci incorporated in the new multiplex include one
dinucleotide locus YCAIIa/b, two other trinucleotide
loci DYS388 and DYS426, and four tetranucleotide
repeat loci DYS460, H4, DYS437 and DYS439. In
addition to these, there are two pentanucleotide repeat
loci DYS438 and DYS447 and one hexa repeat
nucleotide locus DYS448. Three primer pairs,
DYS389, DYS385 and YCAII generate two
amplicons each, resulting in a total of 20 Y-STRs.
The detailed information on loci used in the 20-Y-
STR multiplex is given in Table 1. We carried out
standardization and in-house validation of 20 Y-STR
loci in a single tube multiplex reaction by using these
primers described previously41
. The multiplex system
would also help in examining whether the Indian
populations harbour similar level of polymorphism in
all the selected Y-STR markers as those described by
the European populations so that a uniform set of
markers could be easily used in routine forensic
investigations to provide a high power of
discrimination.
Description of Primers
The amplification of the selected loci was
carried out using NIST primer sequences
obtained from the Y-STR database site
(http://www.cstl.nist.gov/biotech/strbase/). Although

SAHOO & KASHYAP: SINGLE TUBE FLUORESCENT MULTIPLEX ASSAY FOR SIMULTANEOUS TYPING OF 20
43
the genetic recombination between X and Y
chromosome is low, certain degree of homology exists
between the two chromosomes because of their common
evolutionary history42
. Therefore, the objective was to
incorporate those primers which have enough specificity
to ensure preferential amplification of Y-chromosome
sequences and minimize or eliminate any interference
from X chromosome. The primers were aligned with
each other and also with the sequence database at the
National Center for Biotechnology Information (NCBI)
using the Basic Local Alignment Search Tool (BLAST)
program to rule out any possibility of dimerization and
to determine the extent of homology with the X
chromosome or to any other chromosome. The primers
are labelled with four different fluorescent dyes, 6′-FAM
(blue), VIC (green), NED (yellow) and PET (red), where
different flurochrome would detect overlapping
fragment sizes.
Test Samples
Genomic DNA was obtained from blood or tissue
samples from both human and non-human sources
following organic extraction protocol43
.
Basic PCR Protocol
The basic PCR (20 µL reaction volume) included:
autoclaved milli-Q water, 1X PCR gold buffer, 1.5 mM
MgCl2, 200 µM each dNTP, 1U AmpliTaq GoldTM
, 5%
v/v glycerol (if used), primer (0.2-1.2 µM each) and
genomic DNA. The amplicons were checked by
electrophoresis on Nusieve®
Agarose gels (FMC
Bioproducts, Rockland, ME, USA) containing
Ethidium bromide in 1X TBE [0.09 M Tris-Borate;
0.002 M EDTA (pH 8.0)] buffer and detected under the
UV transilluminator. All the amplification reactions
included a known DNA sample as male positive
control, a female DNA as negative control and an
amplification negative control.
The basic PCR protocol included (1) Amplification
of each loci as monoplexes and (2) Multiplexing of the
Y-STR loci through a series of standardization steps:
i) Standardization of annealing temperature
ii) Optimization of primer concentrations
iii) Optimization of DNA polymerase and magnes-
ium concentrations
iv) Validation of multiplex assay
Results and Discussion Single Locus PCR
Each of the primer pairs was amplified individually
at the same annealing temperature (55ºC) in 1x
reaction condition with 0.5 µM primer concentration
and without glycerol (Table 2). A high primer
concentration was maintained for checking the
efficacy of primers under similar amplification
conditions. Fig. 1 shows the agarose gel image for
successful amplification of all the single Y-STR loci.
Multiplex PCR
Combining the primers together for amplifying
many loci simultaneously requires
alteration/optimization of certain parameters of the
reaction44
. Initially, all the primers were added in
equimolar amounts in various combinations under
single locus PCR conditions in addition to 5%v/v
glycerol, to test the individual primer concentrations
and other parameters for a successful multiplex
reaction. For the initial standardization, all primers
were mixed in 0.5 µM concentrations, of which only
8.0 µL of Y-STR primer mix was added to the 20 µL
reaction mix. Fig. 2 shows the 4% agarose gel
photograph for positive amplification with Y-STR
multiplex. The final primer concentrations for each of
the primer pairs are given in Table 3. Performance of
the PCR-based STR assays can be influenced by
numerous factors, including the difference in
Table 1—General information on loci used in the 20 Y-STR
multiplex system
Marker Name Repeat motif Gen Bank
accession
Reference
allele
DYS19 TAGA AC017019 (r&c) 15
DYS385 a/b GAAA AC022486 (r&c) 11
DYS389 I (TCTG) (TCTA) AC004617 (r&c) 12
DYS389 II (TCTG) (TCTA) 29
DYS390 (TCTA) (TCTG) AC011289 24
DYS391 TCTA AC011302 11
DYS392 TAT AC011745 (r&c) 13
DYS393 AGAT AC006152 12
YCAII A/B CA AC015978 23
DYS388 ATT AC004810 12
DYS426 GTT AC007034 12
DYS437 TCTA AC002992 16
DYS438 TTTTC AC002531 10
DYS439 AGAT AC002992 13
DYS447
(TAATA)
(TAAAA) AC005820 23
DYS448 AGAGAT AC025227 22
DYS460
(A7.1) ATAG AC009235 (r&c) 10
Y-GATA-H4 TAGA AC011751 (r&c) 12
Reference allele refers to the number of repeats found in the
GenBank sequence, which must sometimes be made reverse and
complement (r&c) in order to maintain consistency with previously
used repeat motifs (42,46).

INDIAN J BIOTECHNOL, JANUARY 2008
44
instrumentation, variability in quality and quantity of
DNA template being amplified, variations in reaction
and analysis conditions. Therefore, further
standardization of the multiplex was carried out after a
few preliminary rounds of detection of the amplicons
in a genetic analyzer for a high throughput format to
obtain accurate and precise size calling of the alleles.
Detection and Analysis of PCR Products
Detection of the amplicons was achieved with ABI
Prism® 3100 Genetic Analyzer 16-capillary array
system (Applied Biosystems) using the G5 matrix
filter for the 5-dye chemistry. GS500 Liz™ (Applied
Biosystems), which produced orange colour
fluorescence, was used as size standard for sizing
length of the amplicons in basepairs. Samples were
prepared in Hi-Di™ formamide (Applied Biosystems)
with GS500 Liz™ size standard (mixed in 25:1 ratio)
and 1 µL of PCR product. Electrophoresis were
performed at 15 kV for 44 min with a run temperature
of 60ºC using 3100 POP™-4 (Applied Biosystems),
1X Genetic Analyzer Buffer with EDTA and a 36 cm
array. Following data collection, samples were
analysed with Genescan® 3.7 (Windows NT, Applied
Biosystems) for sizing of the products. Allele
designations were made based on sizing bin windows
of up to ±0.15 bp.
Optimization of Reaction Conditions
Depending on the difference in peak heights of the
amplicons, appropriate adjustments were made in the
concentration of the critical reagents and
thermocycling conditions including reaction volume,
template and primer concentrations, annealing
temperature to obtain more balanced signal intensity.
Table 2—Cycling conditions for single locus and multiplex PCR
Single locus PCR Multiplex PCR
Pre-denaturation 95ºC, 11 min 95ºC, 11 min
Denaturation 94ºC, 1 min 94ºC, 1 min
Annealing 55ºC, 1 min 57ºC, 1 min
Extension 72ºC, 1 min 72ºC, 1 min
30 cycles 28 cycles
Final extension 60ºC, 45 min 60ºC, 45 min
Hold 4ºC 4ºC
Fig. 1—2% Agarose gel image of single locus PCR amplification
of 20 Y-STR using 1X buffer conditions at an annealing
temperature of 55ºC.
Fig. 2—Multiplex PCR amplification of 20Y-STRs at 57ºC using
the PE 9700 thermal cycler in a 4% agarose gel.
Table 3—Locus information, size range42 and final primer conc of
20 Y-STR loci
STR locus Size range
(bp)
Fluorescence
label
Allele
range
Primer conc
(µM)
DYS19 233-269 NED 10-19 0.76
DYS385 242-306 VIC 7-23 0.46
DYS388 151-175 NED 10-18 1.5
DYS389I 143-175 6-FAM 9-17 1.5
DYS389II 263-295 6-FAM 26-34 -
DYS390 189-233 VIC 17-28 1.5
DYS391 93-121 6-FAM 7-14 0.46
DYS392 290-318 NED 6-16 1.5
DYS393 109-133 VIC 9-16 0.92
DYS426 92-98 VIC 10-12 0.46
DYS437 186-198 6-FAM 14-17 0.46
DYS438 300-335 6-FAM 6-13 0.76
DYS439 210-230 6-FAM 16-21 0.46
DYS447 187-242 PET 22-29 0.46
DYS448 299-335 PET 18-26 0.30
DYS460 101-121 NED 7-12 0.46
H4 122-142 NED 8-13 0.46
YCAII 135-161 VIC 11-24 0.46

SAHOO & KASHYAP: SINGLE TUBE FLUORESCENT MULTIPLEX ASSAY FOR SIMULTANEOUS TYPING OF 20
45
Reaction Volume and Template Concentration
Change in the reaction volume can alter the
concentration of PCR reaction components. Varied
reaction volumes (5, 10, 12.5, 15, 20 and 25 µL) were
evaluated to determine the effects on amplification.
When the concentrations were kept uniform,
amplification was seen at >10 µL volumes. To test the
sensitivity of the multiplex system, a serial dilution of
the test samples was performed to quantify the optimal
concentration of DNA template required for a robust
multiplex amplification. Template concentration of 10
ng per reaction volume gave more than 100 RFU and
was found to be sufficient for robust amplification.
Larger loci drop-outs were observed at concentrations
less than 5 ng template DNA.
Primer Titrations
Concentrations of the primers play a crucial role in
the amplification efficacy of loci in a multiplex
reaction. Therefore, adjustments in the primer
concentrations are critical to produce equivalent signals
for each locus. Primer concentrations between 0.1 µM
and 1.5 µM with 28-30 cycles and template between 1
and 25 ng were tested for determining relative signal
balance between loci. When high primer concentrations
were used, smaller loci were preferentially amplified.
With the final primer concentrations as given in Table
3, peak heights decreased in smaller loci and increased
in larger loci, giving a relative balance of signal
intensity. Locus drop-out was seen at primer
concentrations less than 0.2 µM and 5 ng of DNA.
Final reaction volumes were adjusted to 10 µL to
which primer volume was further reduced to 2.0 µL of
primer mix.
Variation of Annealing Temperature
Changes in the annealing temperature can affect the
specificity and balance of the amplified loci. To
examine these effects, amplification reactions were
performed at various annealing temperatures (55ºC-
60ºC) with a minimum of 25 ng DNA template
concentration. Variations in annealing temperatures
can also cause cross-reactivity and allele drop-outs.
The most commonly observed loci to drop-out at
higher annealing temperature include DYS19,
DYS389II, DYS447 and DYS388. Locus drop-out and
additional artifacts were observed at temperatures
above 58ºC, whereas at lower annealing temperature an
increased yield of the smaller loci caused a locus-to-
locus imbalance. The final annealing temperature was
maintained at 57ºC (Table 2).
Optimization of DNA polymerase and Magnesium Concentrations
The DNA polymerase plays a key role in
determining the proportion of amplification that could
be achieved. Therefore, variations in the
concentrations of AmpliTaq® Gold DNA polymerase
were evaluated. Between 1 and 5U/20 µL DNA
polymerase concentrations were examined with
template concentration of 1-25 ng and 28-30 cycling
reactions. Optimal balance of signal between loci
were seen at as low as 1U per reaction volume. No
locus drop-out was observed at this enzyme
concentration but the yield of larger loci was low.
Magnesium has significant effect in polymerase
activity and specificity, but EDTA in the samples can
inadvertently alter effective concentrations of
magnesium. Various titrations between 1-2.5 mM
were examined, where the optimal balance between
loci was seen at 1.5 mM magnesium concentration.
Locus-to-locus imbalance increased on decreasing the
concentration of magnesium to less than 1.2 mM.
Lower concentration resulted in locus drop-outs while
higher concentration led to the formation of too many
non-specific products.
Comparison of Thermal Cyclers
Different models of thermal cyclers have slightly
different heating and cooling properties. To evaluate
the consistency of yield across loci on different thermal
cyclers, two different instruments— Perkin-Elmer
GeneAmp® PCR System 2400 and 9700 were
examined. This exercise was carried out to rule out
biased haplotypes due to the usage of different PCR
machines. The signal intensity of each locus (RFU)
was compared to the total signal across all the loci for a
sample. No consistent performance differences were
observed between the thermal cyclers (Table 4).
Validation of the Multiplex
STR analysis of forensic samples in particular,
relies on two important parameters (i) specificity of
primers for humans and (ii) sensitivity for detection45
.
Thus, validation studies of the multiplex system were
carried out on different samples and matrices to verify
the species specificity, sensitivity, mixture analysis
and effect of environmental stresses. The specificity
of multiplexes for amplification was examined with
five different animal species, including rat, frog, fish,
chicken and bovine sp. None of the tested samples
yielded positive amplification. This assay with high
degree of human specificity is essential for accurate
result interpretation in case of contamination from
non-human sources. The sensitivity of the multiplex

INDIAN J BIOTECHNOL, JANUARY 2008
46
system was tested by quantifying the optimal amount
of template DNA required for reliable amplification in
serially diluted DNA samples. An advantage of Y-
STR analysis even with low template concentration is
that while in the case of autosomal markers, allele
drop-out creates potential for misinterpretation at
heterozygous locus, drop-out of a Y-STR loci will
produce no information except in cases of male/male
mixtures or bi-allelic loci such as DYS385a/b and
YCAII a/b. Further, to demonstrate that the sampling
method does not produce biased haplotype profile for
the Y-STRs, reproducibility with liquid blood, dried
bloodstains, buccal swabs, semen sample and autopsy
tissue was carried out on various male samples. In all
instances, different samples produced complete,
correct and identical profiles. Exposure to different
environmental conditions, such as temperature and
sunlight exposure is known to affect the quality of
DNA resulting in degradation of larger templates and
decreased yield of larger amplicons. To evaluate this,
DNA was extracted from bloodstains exposed to
various natural and laboratory-controlled conditions
for different time periods. All the samples exposed to
50ºC or lower temperatures and a range of light
exposures produced complete haplotypes (Fig. 3). To
confirm the effect of PCR inhibitors on substrate
materials which can adversely affect the performance
of amplification, we carried out amplification and
generated complete profiles from various matrices
such as, gauze, wood, glass, and soil, etc. One of the
salient features of Y-STRs is its utility in analysis of
mixed samples. Male and female fractions in various
concentrations and combinations, from a ratio of 10:1
to 1:10 were analyzed. It was carried out to exclude
the probability of any female sample getting amplified
and secondly, to detect inhibitory concentrations of
female DNA for Y-specific amplifications. Large
concentrations of female fractions affect the
amplification of higher loci. Even with varying ratios
of the male samples in the male/male mixtures,
complete and correct haplotypes could be generated.
These data suggest that this multiplex has the level of
specificity and sensitivity to reliably produce results
for proportions to male/female mixtures generally
obtained in forensic samples. Determining the number
of donors in the male/male mixtures only becomes
difficult when bi-allelic loci are taken during
interpretation.
Population Studies
As a part of the validation study and to determine
the extent of polymorphism of the selected Y-STR
markers such that they are able to differentiate
between unrelated males with a high degree of
significance, we tested the multiplex system on
various population samples46-50
. We observed that a
Fig. 3—Validation of the 20-Y-STRs multiplex PCR in liquid blood and stain samples in guaze and Whatman paper.
SAHOO & KASHYAP: SINGLE TUBE FLUORESCENT MULTIPLEX ASSAY FOR SIMULTANEOUS TYPING OF 20
47
total of 1240 distinct Y-STR halotypes in 1297
unrelated, healthy Indian males analysed with the 20
Y-STR multiplex, with complete profiles were
generated in more than 90% of the samples analysed.
The power of discrimination between individuals with
the 20 Y-STR was estimated to be 0.999. The number
of alleles found for each locus ranged from 7-18, with
the exception of DYS385 and YCAII loci, which gave
81 and 55 genotypes, respectively. All the Y-STR loci
depicted gene diversity values greater than 0.5, even
for those loci that harbored the least number of alleles
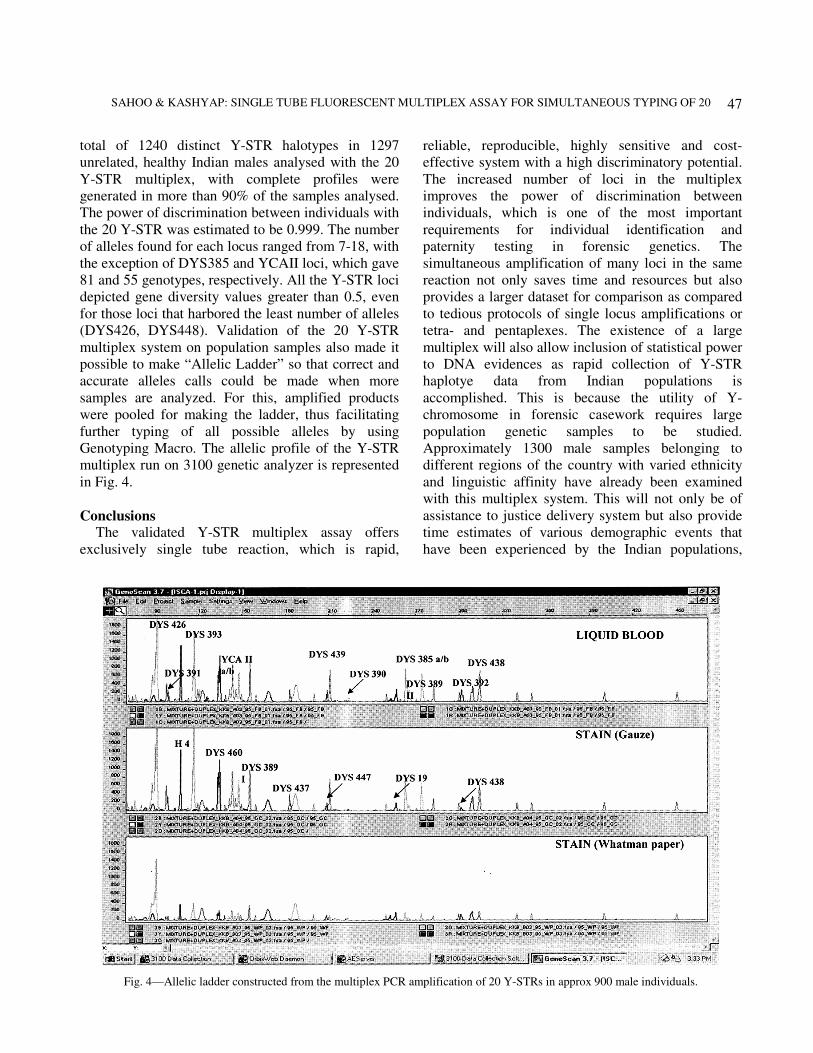
(DYS426, DYS448). Validation of the 20 Y-STR
multiplex system on population samples also made it
possible to make “Allelic Ladder” so that correct and
accurate alleles calls could be made when more
samples are analyzed. For this, amplified products
were pooled for making the ladder, thus facilitating
further typing of all possible alleles by using
Genotyping Macro. The allelic profile of the Y-STR
multiplex run on 3100 genetic analyzer is represented
in Fig. 4.
Conclusions The validated Y-STR multiplex assay offers
exclusively single tube reaction, which is rapid,
reliable, reproducible, highly sensitive and cost-
effective system with a high discriminatory potential.
The increased number of loci in the multiplex
improves the power of discrimination between
individuals, which is one of the most important
requirements for individual identification and
paternity testing in forensic genetics. The
simultaneous amplification of many loci in the same
reaction not only saves time and resources but also
provides a larger dataset for comparison as compared
to tedious protocols of single locus amplifications or
tetra- and pentaplexes. The existence of a large
multiplex will also allow inclusion of statistical power
to DNA evidences as rapid collection of Y-STR
haplotye data from Indian populations is
accomplished. This is because the utility of Y-
chromosome in forensic casework requires large
population genetic samples to be studied.
Approximately 1300 male samples belonging to
different regions of the country with varied ethnicity
and linguistic affinity have already been examined
with this multiplex system. This will not only be of
assistance to justice delivery system but also provide
time estimates of various demographic events that
have been experienced by the Indian populations,
Fig. 4—Allelic ladder constructed from the multiplex PCR amplification of 20 Y-STRs in approx 900 male individuals.

INDIAN J BIOTECHNOL, JANUARY 2008
48
thereby increasing our understanding of the Indian
scenario of male evolutionary and population
genetics.
Acknowledgement
We acknowledge the facility provided at Central
Forensic Science Laboratory, Kolkata, and would
specially like to thank Dr R Trivedi for providing
technical support required in the study, and to the
research scholars of the institute for carrying out
validation studies on population samples. This study
was funded by a research grant under the X Five Year
Plan to CFSL, Kolkata. SS was supported by a
fellowship from Ministry of Home Affairs, Govt of
India.
References 1 Henegariu O, Hirschmann P, Kilian K, Kirsch S, Lengauer C
et al, Rapid screening of Y-chromosome in idiopathic sterile
men, diagnostic for deletions in AZF, a genetic Y factor
expressed during spermatogenesis, Andrologia, 26 (1994)
97-106.
2 Shuber A P, Skoletsky J, Stern R & Handelin B L, Efficient
12-mutation testing in the CFTR gene: A general model for
complex mutation analysis, Hum Mol Genet, 2 (1993) 153-
158.
3 Edwards A, Civitello A, Hammond H A & Caskey C T,
DNA typing and genetic mapping with trimeric and
tetrameric tandem repeats, Am J Hum Genet, 49 (1991) 746-
756.
4 Fregeau C J & Fourney R M, DNA typing with fluorescently
tagged short tandem repeats: A sensitive and accurate
approach to human identification, BioTechniques, 15 (1993)
100-119.
5 Hammond H A, Jin L, Zhong Y, Caskey C T & Chakraborty
R, Evaluation of 13 short tandem repeat loci for use in
personal identification applications, Am J Hum Genet, 55
(1994) 175-189.
6 International Human Genome Sequencing Consortium,
Initial sequencing and analysis of the human genome, Nature
(Lond), 409 (2001) 860-921.
7 Kimpton C P, Gill P, Walton A, Urquhart A, Millican E S &
Adams M, Automated DNA profiling employing multiplex
amplification of short tandem repeat loci, PCR Meth Appl, 3
(1988) 13-22.
8 Urquhart A, Kimpton C P, Downes T J & Gill P, Variation in
short tandem repeat sequences-A survey of twelve
microsatellite loci for use as forensic identification markers,
Int J Legal Med, 107 (1994) 13-18.
9 Chamberlain J S, Gibbs R A, Ranier J E, Nguyen P N &
Caskey C T, Deletion screening of the duchenne muscular
dystrophy locus via multiplex DNA amplification, Nucleic
Acids Res, 16 (1988) 11141-11156.
10 Ayub Q, Mohyuddin A, Qamar R, Mazhar K, Zerjal T et al,
Identification and characterisation of novel human Y-
chromosomal microsatellites from sequence database
information, Nucleic Acids Res, 28 (2000) e8.
11 White P S, Owathan L T, Deaven L L & Longmire J L, New
male-specific microsatellite markers from the human Y-
chromosome, Genomics, 57 (1999) 433-437.
12 Grignani P, Peloso G, Fattorini P & Previdere C, Highly
informative Y-chromosomal haplotypes by the addition of
three new STRs DYS437, DYS438 and DYS439, Int J Legal
Med, 114 (2000) 125-129.
13 Gonzalez-Neira A, Elmoznino M, Lareu M V, Sanchez-Diz
P, Gusmao L et al, Sequence structure of 12 novel Y-
chromosome microsatellites and PCR amplification
strategies, Forensic Sci Int, 122 (2001)19-26.
14 Redd A J, Agellon A B, Kearney V A, Contreras V A,
Karafet T et al, Forensic value of 14 novel STRs on the
human Y-chromosome, Forensic Sci Int, 130 (2002) 97-111.
15 Sinha S K, Budowle B, Arcot S S, Richey S L, Chakraborty
R et al, Development and validation of a multiplexed Y-
chromosome STR genotyping system, Y-PLEX 6, for
forensic casework, J Forensic Sci, 48 (2003) 93-103.
16 Krenke B E, Viculis L, Richard M L, Prinz M, Milne S C et
al, Validation of a male-specific, 12-locus fluorescent short
tandem repeat (STR) multiplex, Forensic Sci Int, 151 (2005)
111-124
17 Hellborg L & Ellegren H, Low levels of nucleotide diversity
in mammalian Y-chromosomes, Mol Biol Evol, 21 (2004)
158-163.
18 Kayser M, Caglià A, Corach D, Fretwell N, Gehrig C et al,
Evaluation of Y-chromosomal STRs: A multicenter study,
Int J Legal Med, 110 (1997) 125-133; appendix: 141-149.
19 de Knijff P, Kayser M, Caglia A, Corach D, Fretwell N et al,
Chromosome Y microsatellites: Population genetic and
evolutionary aspects, Int J Legal Med, 110 (1997) 134-149.
20 Jobling M A & Tyler-Smith C, Fathers and sons: The
Y-chromosome and human evolution, Trends Genet, 11
(1995) 449-456.
21 Sahoo S & Kashyap V K, Genetic variation at 15 autosomal
microsatellite loci in the three highly endogamous tribal
populations of Orissa, India, Forensic Sci Int, 130 (2002)
189-193.
22 Sahoo S & Kashyap V K, Allele frequency data for
powerplex 16 loci in four major populations of Orissa, India,
J Forensic Sci, 47 (2002) 912-915.
23 Jobling M A, Pandaya A & Tyler-Smith C, The
Y-chromosome in forensic analysis and paternity testing, Int
J Legal Med, 110 (1997)118-124.
24 Corach D, Sala A, Penacino G, Ianucci N, Bernardi P et al,
Additional approaches to DNA typing of skeletal remains:
The search for “missing” persons killed during the last
dictatorship in Argentina, Electrophoresis, 18 (1997) 1608-
1612.
25 Honda K, Roewer L & de Knijff P, Male DNA typing from
25-year-old vaginal swabs using Y-chromosomal STR
polymorphisms in retrial request case, J Forensic Sci, 44
(1999) 868-872.
26 Prinz M, Boll K, Baum H & Shaler B, Multiplexing of
Y-chromosome specific STRs and performance for mixed
samples, Forensic Sci Int, 85 (1997) 189-218.
27 Ruiz-Linares A, Ortíz-Barrientos D, Figreon M, Mesa N,
Múnera J G et al, Microsatellites provide evidence for
Y-chromosome diversity among the founders of the New
World, Proc Natl Acad Sci (USA), 96 (1999) 6312-6317.
28 Ramana G V, Su B, Jin L, Singh L, Wang N et al,
Y-Chromosome SNP haplotypes suggest evidence of gene

SAHOO & KASHYAP: SINGLE TUBE FLUORESCENT MULTIPLEX ASSAY FOR SIMULTANEOUS TYPING OF 20
49
flow among caste, tribe and migrant Siddi populations of
Andhra Pradesh, South India, Eur J Hum Genet, 9 (2001)
695-700.
29 Su B, Xiao J, Underhill P, Deka R, Zhang W et al,
Y-Chromosome evidence for a Northward migration of
modern humans into Eastern Asia during the last ice age, Am
J Hum Genet, 65 (1999) 1718-1724.
30 Redd A J, Clifford S L & Stoneking M, Multiplex DNA
typing of short tandem repeat loci on the Y-chromosome,
Biol Chem, 378 (1997) 923-927.
31 Roewer L, Krawczak M, Willuweit S, Nagy M, Alves C et
al, Online reference database of European Y-chromosomal
short tandem repeat (STR) haplotypes, Forensic Sci Int, 118
(2-3) (2001)106-113.
32 Shin D J, Jin H J, Kwak K D, Choi J W, Han M S et al, Y-
Chromosome multiplexes and their potential for the DNA
profiling of the Koreans, Int J Legal Med, 115 (2001) 109-
117.
33 Redd A J, Chamberlain V F, Kearney V F, Stover D, Karafet
T et al, Genetic structure among 38 populations from the
United States based on 11 US core Y-chromosome STRs, J
Forensic Sci, 51 (2006) 580-585.
34 Pascali V L, Dobosz M & Brinkmann B, Coordinating
Y-chromosomal STR research for the courts, Int J Legal
Med, 112 (1998) 1.
35 Sahoo S, Chainy G B N & Kashyap V K, Allele frequency of
eight Y-chromosome STR loci in Oriya population of India,
J Forensic Sci, 48 (2003) 245-252.
36 Bär W, Brinkmann B, Budowle B, Carracedo A, Gill P et al,
DNA recommendations: Further report of the DNA
Commission of the ISFH regarding the use of short tandem
repeat systems, Int J Legal Med, 110 (1997) 175-176.
37 Caglià A, Dobosz M, Boschi I, d’ Aloja E & Pascali V L,
Increased forensic efficiency of a STR based Y-specific
haplotypes by addition of the highly polymorphic DYS385
locus, Int J Legal Med, 111 (1998) 142-146.
38 Schneider P M, Meuser S, Waiyawuth W & Rittner C,
Tandem repeat structure of the duplicated Y-chromosome
STR locus 385 and frequency studies in the German and
three Asian population, Forensic Sci Int, 97 (1998) 61-70.
39 Thomas M G, Bradman N & Flinn H M, High throughput
analysis of 10 microsatellite and 11 diallelic polymorphisms
on the human Y-chromosome, Hum Genet, 105 (1999) 577-
581.
40 White P S, Owathan L T, Deaven L L & Longmire J L, New
male-specific microsatellite markers from the human
Y-chromosome, Genomics, 57 (1999) 433-437.
41 Butler J M, Schoske R, Vallone P M, Kline M C, Redd A J et
al, A novel multiplex for simultaneous amplification of 20 Y
chromosome STR markers, Forensic Sci Int, 129 (2002) 10-
24.
42 Lahn B, Pearson N & Jegalian K, The human Y-chromosome
in the light of evolution. Nat Rev Genet, 2 (2001) 207-216.
43 Sambrook J, Fritsch E F & Maniatis T, Molecular Cloning. A
laboratory manual. 2nd edn (Cold Spring Harbor Laboratory
Press, NY) 1989.
44 Henegariu O, Heerema N A, Dlouhy S R, Vance G H &
Vogt P H, Multiplex PCR: Critical parameters and step-by-
step protocol, BioTechniques, 23 (1997) 504-511.
45 Gill P, Brenner C, Brinkmann B, Budwole B, Carracedo A et
al, DNA Commission of the International Society of
Forensic Genetics: Recommendations on the forensic
analysis using Y-chromosome STRs, Forensic Sci Int, 124
(2001) 5-10.
46 Banerjee J, Trivedi R & Kashyap V K, Y-STR haplotypes in
autochthonous tribal population of Chotanagpur Plateau,
India, J Forensic Sci, 50 (2005) 739-744.
47 Bindu G H, Trivedi R & Kashyap V K, Allele frequencies of
20 Y-chromosomal Short Tandem Repeat Loci in Tribal
population of Deccan Plateau, India, J Forensic Sci, 52
(2007) 507-510.
48 Singh A, Trivedi R & Kashyap V K, Haplotype diversity at
20 Y-chromosome microsatellites in an Indo-Caucasian
population of Bengal, India, J Forensic Sci, 51 (2006) 1434-
1437.
49 Trivedi R, Sitalaximi T, Banerjee J, Singh A, Sircar P K et
al, Molecular insights into the origins of the Shompen, a
declining population of the Nicobar archipelago, J Hum
Genet, 51 (2006) 217-226.
50 Sahoo S, Singh A, G Hima Bindu, Banerjee J, Sitalaximi T et
al, A prehistory of Indian Y-chromosomes: Evaluating demic
diffusion scenarios, Proc Natl Acad Sci USA, 103 (2006)
843-848.