utilization of developmental basic science principles in the evaluation of reproductive risks from...
TRANSCRIPT

Utilization of Developmental Basic SciencePrinciples in the Evaluation of ReproductiveRisks from Pre- and PostconceptionEnvironmental Radiation ExposuresROBERT L. BRENT*Division of Developmental Biology, Thomas Jefferson University, Philadelphia, Pennsylvania 19107; duPont Hospitalfor Children, Wilmington, Delaware 19899
ABSTRACT The subject of the reproductivetoxicity of various forms of radiation can be anxietyprovoking to the public on two accounts, since reproduc-tive failure engenders an unusual level of guilt and angerin the affected families, and radiation effects aremisunderstood and feared by the public. Reproductiveproblems include an array of genetic and acquireddiseases affecting parents and their offspring. Many ofthese problems are associated with the risk of beinginduced by preconception and/or postconception expo-sures to environmental agents. For the various forms ofradiation, namely, ionizing radiation, ultrasound, low-frequency electromagnetic fields (EMF), and micro-waves, the potential for producing reproductive effectsvaries considerably with the form of ‘‘radiation’’ and, ofcourse, the dose. Whether the exposure occurs precon-ceptionally or postconceptionally is another major con-sideration. In evaluating the actual reproductive risks,we rely on accurate dosimetry and information obtainedin epidemiological studies and animal studies. Epidemio-logical studies must demonstrate consistency of thereproductive finding, and animal studies should bedesigned to add to the findings of the epidemiologicalstudies. Most importantly, the conclusions must notcontradict the basic principles of teratology, genetics,and reproductive biology, and they should be biologi-cally plausible. But frequently important basic scienceprinciples are ignored in the evaluation process. Yetdevelopmental basic science principles can be instrumen-tal in refuting or supporting the concern about possiblerisks. Although there is some overlap with regard to thepreconception and intrauterine effects of ionizing radia-tion, there are significant differences. Preconceptioneffects are mainly stochastic effects, while intrauterineeffects are mainly deterministic effects. The stochasticgenetic risks are lower than the deterministic risks atequivalent exposures. Thus, it is frequently difficult todemonstrate the occurrence of stochastic effects inpopulations that have received low preconception expo-sures to ionizing radiation. The reproductive effectsfrom preconception and intrauterine exposures to elec-tromagnetic fields (low-frequency EMF, video displayterminals, microwaves) and ultrasound represent much
different problems, since the main effects of micro-waves and ultrasound occur because of their hyperther-mic effects at high exposures. Low-frequency EMFdoes not have the capacity to produce hyperthermia,and none of these forms of nonionizing radiation hasthe specificity to damage the DNA comparable to thespecificity of ionizing radiation. Not only do they nothave targeted mutagenic effects at the usual exposuresthat populations receive, they are not cytotoxic at theseexposure levels as well. From the viewpoint of biologi-cal plausibility, these other forms of radiation are muchless likely to have the potential for producing reproduc-tive toxicity at the usual population exposures. Teratol-ogy 59:182–204, 1999. r 1999 Wiley-Liss, Inc.
Reproductive problems include an array of geneticand acquired diseases affecting parents and theiroffspring. Many of these problems are at risk of beinginduced by preconception and/or postconception expo-sures to environmental agents. For the variousforms of radiation discussed in this National Council onRadiation Protection and Measurements (NCRP) sym-posium, namely, ionizing radiation, ultrasound, low-frequency electromagnetic fields (EMF), and micro-waves, the potential for producing reproductive effectsvaries considerably with the form of ‘‘radiation’’ and, ofcourse, the dose. Whether the exposure occurs precon-ceptionally or postconceptionally is another major deter-minant of the magnitude of the risk.
The effects of high-energy radiation (x-rays, g-rays,particulate radiation) have been studied more exten-sively than has any other environmental hazard. Ascompared with ionizing radiation the various forms ofnonionizing radiation have quite different biologic ef-fects (Table 1). In spite of the vast amount of knowledgeavailable about ionizing radiation, many physicians areunfamiliar with the quantitative and qualitative effectsof ionizing radiation (Brent, ’80a). In this respect, our
*Correspondence to: Robert L. Brent, Division of DevelopmentalBiology, Room 308 R/A, Box 269, duPont Hospital for Children,Wilmington, DE 19899.
TERATOLOGY 59:182–204 (1999)
r 1999 WILEY-LISS, INC.

laboratory is frequently consulted when women ofreproductive age are exposed to diagnostic x-rays. Moreoften than not, the exposed women have receivederroneous information because of the failure of thephysician to obtain the relevant clinical information. Inone instance, abortion was advised after exposure todiathermy because of confusion about the differencebetween microwave and ultrasound energy. In anotherinstance, the woman was advised that a diagnosticcomputed tomography (CT) scan of her abdomen re-sulted in a much higher exposure to her embryo thandid other abdominal radiographic studies and, there-fore, the embryo was at increased risk. In this particu-lar case the embryo had received a very low dosebecause it was not in the region of the exposure.
The most recent example of the hysteria that canoccur after low-dose ionizing radiation exposure topregnant women is represented by the results of theChernobyl nuclear power plant explosion that occurredin 1986. While there were reports of an increase in thefrequency of therapeutic abortions in Russia after thedisaster, this was not the case in northern and centralEurope. The exposures to the population were ex-tremely low, and there was no increase in any reproduc-tive effect studied, including congenital malformations,stillbirths, and spontaneous abortions in Norway, Swe-den, Finland, and Austria (Bengtsson, ’91; Haeusler etal., ’92; Harjulehto et al., ’91; Irgens et al., ’91; Knud-sen, ’91; Odlind and Ericson, ’91).
It would be preferable, from an educational viewpoint, tolimit the use of the term radiation, to high-energyionizing radiation. Radar, microwaves, short-wave dia-thermy, FM broadcast range, and radiofrequencies arevarious forms of long-wavelength electromagnetic wavesthat have little in common with x-rays and g-rays,at least from a biologic standpoint. The use of radiationto describe ultrasound may even be more confusingand erroneous. In England, physicians refer to diagnos-tic ultrasound as sonography and consciously refrainfrom using the term ‘‘radiation’’ in a clinical setting,
because of the anxiety-provoking connotations it maysuggest.
The term radiation provokes anxiety because of it’sassociation with the effects of ionizing radiation. Theassociations and effects include:
1. The term radiation is related to the effects of theatomic bomb. In the minds of many, it is impossibleto separate the effects of low-dose ionizing radiationfrom the psychological, physical, and radiation after-math of the atomic bomb. The horrendous outcomesassociated with the atomic and hydrogen bombs areat one end of a continuum of radiation effects. To agreater or lesser extent, this distorts the risks of allforms of energy labeled radiation.
2. Populations exposed to large doses of ionizing radia-tion have increased incidences of cancer. Thesepopulations include the radium dial workers, ura-nium miners, patients receiving radiation therapyor isotope therapy for various diseases, and thepersons who received the higher exposures in Hiro-shima and Nagasaki following the atomic bombdetonation (Polhemus and Koch, ’69). Cancer itself isalso an anxiety-provoking term. In our culture it is adisease with which people are ‘‘afflicted’’ (Fig. 1), andit is a disease that is dreaded by a large segment ofthe population. Few persons are aware of the ex-tremely small maximum risk of the occurrence ofcancer in populations exposed to much lower dosesof radiation.
3. The immense psychological consequences of high-energy radiation exposure are extremely importantand cannot be ignored, when one considers thedeleterious effects of radiation. A review of the30-year study of the survivors of the Hiroshima andNagasaki bombings showed some interesting find-ings (Brent, ’77).
More than 90 percent of survivors received much lessthan 10 rads from the A-bombs. With this knowledge,
TABLE 1. Effects and risks of different forms of radiation
X-raysg-irradiation Ultrasound
Microwavediathermy EMW* (60 Hz)
Type EMW, shortwavelength
Physical EMW, long wave-length
EMW, very longwavelength
Mechanismand effect
Tissue ionization,mutagenesis,carcinogenesis,teratogenesis
Compression,rarefaction,cavitation,tissue disrup-tion, streaminghyperthermia
Hyperthermia(cataract)
Nonthermaleffects?
Electrical andmagnetic fieldsin tissue
Cytotoxicity Yes Not at low expo-sures; yes, athigh exposures
Not at low expo-sures; yes, athigh exposures
No
Relative DNAspecificity
Yes No No No
Phenomena Stochastic andthreshold
Threshold Threshold Threshold
*EMW, electromagnetic wave.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 183

such survivors can realize that their fears of diseasefrom A-bomb exposure may be exaggerated and thatthe possibility of their developing any such disease isno greater than the nonexposed individuals. Thosewho received higher doses have greater risks. It isworth noting that, thus far, the life expectancy of theexposed population under study and of their off-spring is at least equal to that for the population inthe rest of Japan. Thirty years of study have noteradicated radiation damage but has yielded reason-able estimates of the risks involved (Okada et al.,’75).
Evaluating reproductive risks fromenvironmental agents
In the evaluation of the actual reproductive risks werely on accurate dosimetry and information obtained inepidemiological studies and animal studies (Table 2).The epidemiological studies must demonstrate consis-tency of the reproductive finding and the animal stud-ies should be designed to add to the findings of theepidemiological studies. Most importantly, the findingsmust not contradict the basic principles of teratology,genetics, and reproductive biology and should be biologi-cally plausible. But frequently important basic scienceprinciples are ignored in the evaluation process. Yetdevelopmental basic science principles can be instru-mental in refuting or supporting the concern aboutpossible risks.
Reproductive problems encompass a multiplicity ofdiseases, including sterility, infertility, abortion (miscar-riage), stillbirth, congenital malformations (due to envi-ronmental or hereditary etiologies), fetal growth retar-dation, and prematurity (Table 3). These diseases occurcommonly in the general population and thereforeenvironmental causes are not always easy to corrobo-rate. Severe congenital malformations occur in 3% ofbirths. That means that each year in the United States,120,000 newborns are born with severe birth defects.
Genetic diseases occur in approximately 11% of births.Spontaneous mutations account for less than 2–3% ofgenetic disease. Therefore, mutations induced frompreconception exposures to environmental mutagensare difficult endpoints to document (NAS/NRC, ’90).
Reproductive problems alarm the public, the press,and some scientists to a greater extent than do mostother diseases (Fig. 1). Along with cancer, psychiatricillness, and hereditary diseases, reproductive problemshave been viewed throughout history as diseases ofaffliction. Inherent in the reactions of most cultures isthat these diseases have been viewed as punishments
Fig. 1. Through the ages, these diseases have been interpreted orconsidered by multiple cultures to be stigmatizing: punishments formisdeeds or sins. In modern times, these are the diseases whosecausation is frequently adjudicated in the courts and that are mostassociated with anger and guilt.
TABLE 2. Evaluation of potential for developmentaltoxicity in the human: Evidence of a possible
causal relationship
Epidemiological studies: Controlled epidemiological studiesconsistently demonstrate an increased incidence of a par-ticular spectrum of embryonic and/or fetal effects inexposed human populations.
Secular trend data: Secular trends demonstrate a positiverelationship between the changing exposures to a commonenvironmental agent in human populations and the inci-dence of a particular embryonic and/or fetal effect.
Animal developmental toxicity studies: An animal model canbe developed that mimics the human developmental effectat clinically comparable exposures. Since mimicry may notoccur in all animal species, animal models are more likelyto be developed once there is good evidence for the embryo-toxic effects reported in the human. Developmental toxicitystudies in animals are indicative of a potential hazard ingeneral, rather than the potential for a specific adverseeffect on the fetus when there are no human data on whichto base the animal experiments.
Dose-response relationship: Developmental toxicity in thehuman increases with dose (exposure) and the develop-mental toxicity in animal occurs at a dose that is pharma-cokinetically (quantitatively) equivalent to the humanexposure.
Biological plausibility: The mechanisms of developmentaltoxicity are understood, and the effects are biologicallyplausible.
Modified from Brent (’76, ’86, ’91, ’95).
TABLE 3. Reproductive risks per million pregnancies
Reproductive risk Frequency
Immunologically and clinically diagnosedabortions per million conceptions 350,000
Clinically recognized spontaneous abortionsper million clinically recognized pregnan-cies 150,000
Genetic diseases per million births 110,000Multifactorial or polygenic genetic environ-
mental interactions 90,000Dominantly inherited disease 10,000Autosomal and sex-linked genetic disease 1,200Cytogenetic (chromosomal abnormalities) 5,000New mutations 3,000
Major malformations (genetic, unknown,environmental) 30,000
Prematurity 40,000Fetal growth retardation 30,000Stillbirths (.20 wk) 4,000–20,900Infertility 7% of couples
184 R.L. BRENT

for misdeeds (Brent, ’67) (Fig. 1). Regardless of theirrationality of this viewpoint, these feelings do exist.Ancient Babylonian writings recount tales of mothersbeing put to death because they delivered malformedinfants (Warkany, ’71). One George Spencer was slainby the Puritans in New Haven, having been convicted offathering a cyclopean pig, since the Puritans wereunable to differentiate between George Spencer’s cata-ract and the malformed pigs cloudy cornea (Brent, ’67).In modern times, some individuals with reproductiveproblems reverse the historical perspective, blamingothers for their congenital malformations, infertility,abortions, and hereditary diseases, by placing theresponsibility of their illness on environmental agentsdispensed by their health care provider or used by theiremployer (Brent, ’77). The objective evaluation of envi-ronmental causes of reproductive diseases is clouded bythe emotional climate surrounding these diseases, re-sulting in the expression of partisan positions thateither diminish or magnify the environmental risks.These nonobjective opinions can be expressed by scien-tists, the laity, or the press.
Comparison of the physical characteristics andeffects of ionizing radiation, low-frequency EMF,
microwaves, and ultrasound
The problem of emotionality is frequently magnifiedin the evaluation of radiation risks, as the unschooledare unaware of the marked differences in the biologicaleffects of different forms of radiation (Table 1).
Only the very high-frequency photons of ionizingradiation (g-rays, x-rays, and the ionizing potential ofa-rays, b-rays, neutrons, and other high-energy particu-late radiation) can remove orbital electrons and pro-duce ionization in tissues, resulting in cytotoxicity,chromosome damage and point mutations. Althoughmicrowaves (RF) can traverse tissues (depending on thewavelength), their photons do not have the energy toresult in ionization, although microwaves can producehyperthermia at high exposures. Low-frequency EMF(60 Hz) can readily traverse tissues but has insignifi-cant capacity for cytotoxicity or hyperthermia. Ultra-sound is not even a form of electromagnetic waves,being a physical or mechanical waveform that results inthe rarefaction and condensation of matter. It has thecapacity for producing hyperthermia and cytotoxicity athigh exposures, but not ionization.
Thus, one would expect biological effects from allforms of radiation. In some instances, these effects aredeleterious, but in other instances the effects are revers-ible or barely perceptible. The actual risk can only bedetermined by analysis of a large amount of physicaland biological data. As a generality, one can concludethat there are far more differences in the effects of thesevarious forms of ‘‘radiation’’ than there are similarities.Furthermore the risks from exposures to reproductivetoxicants vary both quantitatively and qualitativelybased on whether the exposures occurred preconceptu-ally or postconceptually (Tables 4, 5). It is important
that the significance of dose is not ignored whencharacterizing the manifestations of exposures to eachform of radiation. One cannot evaluate the biologicaleffects and the risks without knowing the type ofradiation exposure, its dose rate and the actual expo-sure. Too often, the principle advocated by Paracelsusin the fourteenth century is ignored: ‘‘What is therethat is not poison? All things are poison and nothing iswithout poison. Solely the dose determines that a thingis not a poison.’’
Let us first examine the risks of preconception radia-tion exposures (Table 4). Because ionizing radiation hasbeen demonstrated to produce cancer in humans andmutations in many experimental situations, it is mostappropriate that we examine the genetic risks of ioniz-ing radiation, since it is very likely that ionizingradiation represents the greatest genetic risk of all theforms of ‘‘radiation’’ listed in Table 1.
REPRODUCTIVE EFFECTS FROMPRECONCEPTION EXPOSURES
TO REPRODUCTIVE TOXICANTS
The real and hypothetical reproductive risks of pre-conception and postconception environmental expo-sures are described in Tables 4 and 5. Although there issome overlap in these effects, the qualitative andquantitative risks from environmental toxicants is sig-nificantly different. High exposures to cytotoxic agentsduring various phases of oogenesis and spermatogen-esis can result in germ cell depletion manifested aseither sterility or reduced fertility, usually considered a
TABLE 4. Risks of preconception exposureto reproductive toxicants
1. Sterility and infertility (threshold phenomenon)2. Chromosomal abnormalities that can be transmitted or
result in the death of the cell. (These abnormalities can bedeletions, duplications, or other microscopically or submi-croscopically observable configurations (stochastic phe-nomenon). These effects are an important contribution tothe dominant lethal effect of preconception ionizing radia-tion.)
3. Abnormalities of the DNA at the molecular level due todeletions, substitutions, or shifting in the sequences of thenucleotides that provide the genetic messages to the cell(stochastic phenomenon)
4. Genetically determined congenital malformations andcancer (stochastic phenomenon)
TABLE 5. Risks of postconception (in utero) exposureto reproductive toxicants
1. Pregnancy loss (abortion, stillbirths) (threshold phenom-enon)
2. Congenital malformations (anatomical defects; thresholdphenomenon)
3. Neurobehavioral abnormalities (i.e., mental retardation)(threshold phenomenon)
4. Fetal growth retardation (reversible and irreversible)(threshold phenomenon)
5. Cancer (stochastic phenomenon)
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 185

threshold or deterministic phenomenon. The occur-rence of sterility has been documented with high dosesof ionization radiation to the gonads, but the risks ofthese effects from the usual exposures of other forms of‘‘radiation’’ (low-frequency EMF, ultrasound, and micro-waves) have been equivocal or nonexistent. Preconcep-tion exposures to ionizing radiation can result in pointmutations and chromosomal abnormalities that can betransmitted or results in germ cell death. These abnor-malities can be deletions, duplications, or other micro-scopically or submicroscopically observable configura-tions and abnormalities of the DNA at the molecularlevel due to deletions, substitutions or shifting in thesequences of the nucleotides that provide the geneticmessages to the cell (NAS/NRC, ’90; Brent, ’92a,b)(Table 4). The other forms of ‘‘radiation’’ (low-frequencyEMF, microwaves, and ultrasound) being discussed inthis symposium have been reported to have equivocalor nonexistent genetic effects, which is understandable,since nonionizing radiation lacks the selective andspecific effects on the DNA manifested by ionizingradiation (Table 1).
Since ionizing radiation can result in both chromo-some damage and point mutations, it is logical topredict that radiation exposure should increase the riskof pregnancy loss (dominant lethals, genetic malforma-tions, hereditary diseases, and cancer). The risk, ofcourse, is related to the dose (Fig. 2).
The genetic and carcinogenic effects of ionizing radia-tion are considered stochastic phenomena, except forthe production of sterility (Table 6). The characteristics
of a stochastic phenomena are that although the risk isdose related, theoretically, there is no dose that does notpresent a risk. Whether the dose-response curve islinear or quadrilinear is a moot point because, at verylow doses, the risks from low-LET radiation are farbelow the spontaneous incidence of mutations. Anothercharacteristic of stochastic phenomena is that while therisk increases with dose, the severity of the disease doesnot. Thus, one cannot distinguish between a patientwith leukemia that has occurred spontaneously or afteran exposure to a high dose of ionizing radiation. This isbecause stochastic phenomena are diseases that theo-retically arise from the alteration of DNA in a singlecell.
Although numerous studies have investigated thegenetic risks of environmental toxicants, the largestand most rigorous was the study of the Japanesepopulations in Hiroshima and Nagasaki who wereexposed to the Atomic bomb. Needless to say, theperception of the public and the press is that the effectsof the atomic bomb in the F1 generation was a geneticdisaster. In fact the scientists assigned to study thesepopulations had little doubt that they would be able todemonstrate an increase in the incidence of geneticdisease in the F1 generation.
The results were somewhat surprising (Table 7).Based on the results of the extensive genetic studiesperformed in Japan, a genetic doubling dose was calcu-lated to be approximately 220 rem (2.2 Sv) (Neel et al.,’90). The implications of this finding on the incidence ofgenetic disease in the F1 generation are described inTable 7. Table 7 describes the change in the incidence ofgenetic disease in the offspring of a population of1,000,000 people who received an additional 1 rem ofradiation. It is obvious from this table that there are notenough people in the study to demonstrate a statisticaldifference between the unirradiated population and theexposed population. The reason for this is that theincidence of inherited genetic disease and spontaneousmutations are so prevalent, that a small increase in theincidence of induced genetic disease cannot be identi-fied.
What did NAS/NRC (’90) conclude about the geneticfindings in Japan?
Past BEIR committee reports have concluded thatthe increase in disease due to recessive mutationsfollowing an increase in the mutation rate fromchronic radiation will be too slight or too remote inthe future to justify quantitative estimation (BIERV, p 80).
there is no reason to suppose that humans areexempt from radiation’s mutagenic effects.
In regard to the induction of mutations, the greatercurrent risk seems to result from exposure to chemi-cal mutagens in the environment rather than from
Fig. 2. The dose-response curve of environmental toxicants (drugs,chemicals, and physical agents) can have deterministic (threshold)and/or stochastic effects. Mutagenic and carcinogenic events arestochastic phenomena and theoretically do not have a thresholdexposure below which no risk exists. At low exposures, the risk stillexists, but is usually below the spontaneous risk of cancer andmutations. Whether the curve is linear or curvilinear for stochasticphenomena can be debated but, from a theoretical point, it traverseszero. Toxicological phenomenon, such as teratogenesis, that do notinvolve mutagenic and carcinogenic effects usually follow on S-shapedcurve, with a threshold, below which no effects are expected.
186 R.L. BRENT

the exposure to populations to radiation (BEIR V,p 66).
Doubling dose in the mouse for various effects atdifferent stages of sperm and ova development anddifferent endpoints range from 40 rad to 2,500 rad.
Similarly, radiation-induced mutation rates varyfrom 1024 to 1028 depending on the mutation andstage of germ cell development when exposed.
Thus, the BEIR V committee and the geneticists whostudied the radiated populations in Japan are con-vinced that there were radiation-induced mutations.However, the calculated and demonstrated risks are sosmall that these investigators were unable to demon-strate statistically significant genetic effects, althoughthe population studied was quite large.
Is this a reasonable conclusion? Is the conclusionbiologically plausible? We can test this conclusion byestimating radiation-induced genetic risks, using theHardy-Weinberg law. The Hardy-Weinberg law wasindependently proposed at the turn of this century bytwo scientists. It is the application of a quadraticequation for the purpose of describing the distributionof genes and genotypes in the population. Two equa-tions are used, which permit the geneticist to determinethe proportion of the population that are homozygous
normal (predominant), as well as carriers and homozy-gous abnormal (rare).
The Hardy-Weinberg law consists of two formulae:
p2 1 2pq 1 q2 (genotypes)
p 1 q 5 1 (genes)
p2 is the proportion of the population that appearnormal and are homozygous
2pq is the proportion of the population that appearnormal and are carriers
q2 is the proportion of the population that are homozy-gous for the abnormal gene.
p is the proportion of the normal gene in the populationand
q is the proportion of the abnormal gene in the popula-tion.
Obviously, these two gene variants must account for allthe positions at this locus for this genetic characteristic.
The Hardy-Weinberg law allows the geneticist tocalculate the following genetic occurrences when givenone unknown from these two equations.
1. The frequency of the genetic disease2. The carrier rate3. The risk of marrying a carrier4. The combined risk of marrying a carrier and having
an affected offspring5. Determining the phenotypes from the carrier rate6. The risk of an affected individual marrying a carrier7. The increase risk of genetic disease following an
increase in the mutation rate
The Hardy-Weinberg law can be used to calculate theincrease risk of genetic disease due to an increase in themutation rate. The following calculation estimates theincreased genetic risks of an average exposure of 1 Sv(100 rem) to the world population (5 billion people) onthe incidence of cystic fibrosis (CF). Although thecurrent estimate of the doubling dose for ionizingradiation is approximately 220 rem (2.2 Sv), a doublingdose of 100 rem (1 Sv) is used for the calculation shown
TABLE 6. Stochastic and threshold dose-response relationships of diseases produced by environmental agents
Phenomenon Pathology Site Diseases Risk Definition
Stochastic Damage to a singlecell may result indisease
DNA Cancer, germ cellmutation
Some risk exists atall dosages; at lowdoses, risk may beless than sponta-neous risk
The incidence of thedisease increasesbut the severityand nature of thedisease remain thesame
Threshold Multicellular injury Multiple, variableetiology, affectingmany cell andorgan processes
Malformation,growth retarda-tion, death, tox-icity, etc.
No increase riskbelow thethreshold dose
Both the severityand incidence ofthe diseaseincrease with dose
Modified from Brent (’87a,b, ’90a,b).
TABLE 7. Genetic effects of 1 rem per generation lowLET ionizing radiation (BEIR-V)
DisorderIncidence per106 liveborn
Additional cases per 106
liveborn per remper generation
1stgeneration
2ndgeneration
Dominant 10,000 6–35 100X-linked 400 ,1 ,5Recessive 2,500 ,1
Translocation 600 ,5Trisomies 3800 ,1 ,1Malfor-
mations 20,000–30,000 10 10–100
Heart disease 600,000 Not estimated Not estimatedCancer 300,000 Not estimated Not estimated
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 187

in the following tabulation. Using this figure for thedoubling dose will overestimate the genetic risk.
The result of this calculation is that after a nucleardisaster in which the average dose to the world popula-tion was 100 rem (1 Sv), the impact on the incidence ofCF in the F1 generation would be imperceptible. Forevery case of CF that resulted from a radiation-inducedmutation, there would be 2,500 patients with CF frominherited genes or genes caused by spontaneous muta-tions. Thus, the results of the BEIR Committee’s delib-erations are consistent with those obtained by thegenetic risks of radiation, as determined by the Hardy-Weinberg law. The genetic studies completed and ongo-ing on the populations in Hiroshima and Nagasaki havebeen summarized by Neel and colleagues (Neel, ’99;Neel et al., ’90).
Epidemiological studies dealing withpreconception exposures
The background incidence of reproductive pathologyis high, affecting a large segment of the population(Table 3). Small increases in the incidence of reproduc-tive pathology may be difficult to document. Whereasmost human epidemiological studies have not found anincrease in translocations or mutations after drug,chemical, or physical agent exposures, various degreesof infertility or embryonic loss have been reported insome of these studies.
Reproduction in survivors after chemotherapyand radiation therapy. It is of interest that epidemi-ologists interested in the reproductive toxicity of precon-ception exposures to drugs, chemicals, and physicalagents have studied potentially hazardous agents withand without mutagenic and cytotoxic potential. Thisreview focuses on studies evaluating agents that arecytotoxic and/or mutagenic, since they have the great-
est potential for affecting fertility and the genetics offuture generations.
The largest group of patients that have been studiedare groups that have been treated for cancer and thathave received chemotherapeutic agents, immunosup-pressants, or ionizing radiation. Survivors of eithercancer in childhood or young adulthood have beenstudied in order to determine whether the exposure toreproductive toxicants has diminished the survivors’reproductive performance.
Mulvihill and Byrne (’85) reviewed the literaturepertaining to the normalcy of the offspring of cancersurvivors. The results of their study is summarized inTable 8. The incidence of malformations in the offspringof more than 1,000 cancer survivors was 3.85%, andtherefore within the range that one would expect in auntreated or unexposed population. It is of interest tonote that women survivors were predominant in thesestudy populations. Dr. Byrne and other members of theNational Cancer Institute (NCI) have continued thesestudies. An update of these studies is one of thepresentations included in this symposium (Byrne, ’99).
Nygaard et al. (’91) studied the reproductive perfor-mance of childhood leukemia survivors. There were 981male and female leukemia survivors. In this series ofpatients, 299 had reached the age of 18 without arelapse. These adult survivors produced 48 offspring,one of whom was malformed. There were no myeloidleukemia survivors who became parents. Of 131 malesurvivors with acute lymphatic leukemia, only 4 be-came parents. Of 149 female survivors with acutelymphatic leukemia, 23 became the parents of 41children. This study also reflected a suggestion thatmale reproductive capacity was more adversely af-fected, possibly because of a difference in therapybetween the sexes or a difference in the response to thetherapy. The study reported that 18–24 Gy to thecentral nervous system (CNS) may lower the rate offirst births. Reduced testicular size and impaired spermproduction have been associated with cranial and tes-ticular radiation. A dose of 4.0 Gy to the ovaries mayresult in permanent sterility in young females. 1.4–3.0Gy to the testes may cause permanent aspermia inmales. Although the population size of this study wassmall, the authors reported no childhood malignanciesor genetic diseases diagnosed in the offspring and noincrease in congenital malformations in the offspring.Although these same investigators (Nygaard et al., ’91)did not find an increase in the frequency of malignan-
Spontaneous mutation rate 1025/locusRadiation-induced mutation rate 1027/rem/locusRadiation-induced doubling dose 1 SvRadiation-induced mutation rate/Sv 1025/Sv/locusNew mutations per germ cell
(spontaneous) 1New mutations per germ cell/Sv 2Frequency of CF (q2) 1/1,600Frequency of CF gene (q) 1/40Frequency of carriers (2pq) 1/20Risk of mutation (new q/Sv) 1025
Radiation-induced carriers; 2pq 2 3 1025 (1/50,000)Incidence of CF in 5 3 109
individuals, the world population; q2 3,125,000Incidence of CF from radiation induced
genes (q 3 q 5 10210) ,1Incidence of CF from carriers
and radiation-induced carriers(1/20 3 1/50,000 3 1/4 3 5 3 109 1,250
Ratio of spontaneous riskto radiation-induced risk 3,125,000/1,250
Ratio of spontaneous riskto radiation-induced carrier risk 2,500/1
TABLE 8. Epidemiological studies of the offspring ofcancer survivors
Exposedparents
Fetalloss
Electiveabortion
Normalnewborn
Total majordefects
825 336 127 1,194 46
Percentage malformations in 1,194 offspring: 3.85%
Modified from Mulvihill and Byrne (’85).
188 R.L. BRENT

cies in the offspring of the cancer survivors, this was notthe case among the survivors themselves. Among acohort of 981 children who were followed 4.3–26.5 yearsafter cessation of antileukemic therapy, eight patientsin remission from acute lymphoblastic leukemia (ALL)developed a distinctively new malignant disease. Thesecond malignant neoplasms (SMN) included braintumors, basal cell carcinomas, thyroid cancer, leiomyo-sarcoma, and finally rhabdomyosarcoma in a patientwho also had suffered from Hodgkin’s disease while stillon antileukemic treatment. Individuals treated forchildhood ALL are at increased risk of a new malig-nancy, and this seems mainly to be associated withprevious irradiation. This study indicates that thesame exposure to mutagenic or cytotoxic agents presentsa much greater risk of inducing clinically recognizedmalignant disease in the exposed rather than geneticdisease in their offspring.
Byrne et al. (’88) studied the reproductive problemsand birth defects in survivors of Wilms’ Tumor and theirrelatives. In this retrospective cohort study of 47 Wilms’tumor survivors and their 77 sibling controls, femalesurvivors had a fourfold excess risk (risk ratio[RR] 5 4.1; 95% confidence interval [95% CI] 5 1.7–10.1) for any adverse live birth outcome, including birthdefects, compared with their sibling controls. Wives ofmale survivors had no apparent excess of risk problempregnancies. The implication suggested by this study isthat radiation to the female child’s abdomen and geni-tal organs resulted in deleterious reproductive out-comes (prematurity, increased neonatal mortality, andintrauterine growth retardation [IUGR[), but that theseeffects were unrelated to any alteration of the genomeof the ova.
Green and colleagues studied the reproductive histo-ries of 93 patients who had been treated for Hodgkin’sdisease, who were 18 or older, and who were at least 5years post-treatment. Forty-eight pregnancies werereported by 22 patients or their spouses. Fifteen femalepatients had 33 pregnancies and 7 male patients re-ported 14 pregnancies in their spouses. While theauthors reported no increase in birth defects or child-hood malignancies in the offspring, the number of caseswas quite small. There was no attempt to determinewhether infertility was increased or whether there wasa greater effect in the male or female (Green et al., ’82,’89). The medical records of 1,239 consecutive patientswho were younger than 20 years of age in 1960–1984and who had received chemotherapy for cancer werereviewed (Green et al., ’91a). A subpopulation of 383former patients who were currently 18 years of age orolder was chosen for further study. Of the 383 patients,100 responded to a questionnaire by mail. Of the 202pregnancies reported, there were 17 spontaneous abor-tions, 4 therapeutic abortions, 1 ectopic pregnancy, 4stillbirths, 13 premature deliveries, and 163 full-termlive births. Among the male patients, 25 reported thebirth of 39 offspring (23 girls, 16 boys), which included 1stillbirth. Of the live-born children, 2 had birthmarks
and 1 had a skin tag, demonstrating a 7.9% minoranomaly rate in infants born after chemotherapy. Norelationship was found between the congenital anoma-lies in infants of fathers who had chemotherapy and thedrug, father’s age at diagnosis, or length of time aftertreatment. For female patients, 63 children (26 girls, 37boys) included 1 stillbirth, 6 premature births, and 57live births. Of the 57 live-born children, 5 (8.1%) hadcongenital anomalies. Patients who undergo cancerchemotherapy during childhood or adolescence do nothave an increased frequency of congenital defects intheir offspring (Green et al., ’91b).
Ionizing radiation has been demonstrated to be muta-genic in both in vivo animal studies and in vitrosystems. Furthermore, radiation can readily producesterility in both males and females if the dose is highenough. The largest study ever undertaken to examinethe genetic effects of ionizing radiation occurred afterthe atomic bomb detonation in Hiroshima and Na-gasaki. The offspring of the atomic bomb survivors whowere exposed to a single dose of radiation had nomeasurable increase in induced mutations after expo-sure to an agent that has proven mutagenic potential(Neel et al., ’90). Statistical analysis of the incidence ofchromosome abnormalities demonstrated no signifi-cant increase in the frequency of chromosomal abnor-malities in children, neonates and abortuses born to theirradiated parents (Okada et al., ’75). By contrast,Martin et al. (’85) reported induced chromosomal aber-rations were present in human sperm years aftertherapeutic radiation. But the authors did not performa controlled study to determine whether these findingswere associated with a decrease in reproductive perfor-mance. The most recent summary of the mutageniceffects of ionizing radiation is provided by Neel (’99) inthis symposium.
Animal genetic studies
The extensive investigation conducted by Russell andcolleagues, using the specific locus test, unquestionablydemonstrated that point mutations could be induced byionizing radiation (Russell, ’77). While there is somecontroversy as to whether the mouse and human have adifferent or similar doubling dose, there is no questionthat ionizing radiation is mutagenic in humans andmice. Neel (’99) discusses the doubling dose controversyin humans and mice in another presentation in thissymposium. It would appear that the newest datawould indicate that the mouse and human data indi-cate doubling doses of .200 rem. The important findingis that the risk of clinical genetic disease is less thangeneticists anticipated and that if one wished to demon-strate radiation induced genetic disease in exposedhuman populations, investigators would need to studyextremely large populations, probably in the millions ortens of millions. Yet it is evident that experiments inanimals have demonstrated mutagenic effects whenanimals are exposed preconceptionally to ionizing radia-tion.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 189
Nomura (’88) examined the offspring of irradiatedanimals and animals treated with other mutagenicagents. These investigators observed a large and signifi-cant increase of phenotypic anomalies in the progeny ofICR parent mice treated before mating with x-rays(Table 9), urethane, 7,12-dimethylbenz[a]anthracene,ethylnitrosourea (ENU), and 4-nitroquinoline 1-oxide,but the increase was not significant with furylfuramide.Major types of induced anomalies were cleft palate,dwarfs, open eyelid, tail anomalies, and exencephalus.Dwarfs, open eyelid, and tail anomalies were predomi-nant types of viable anomalies and were inherited as ifthey were dominant mutations with varying expressiv-ity or penetrance. These investigators also observedthat the incidence of prenatal anomalies increased withthe dose. Spermatogonia were less sensitive to x-raysand urethane than were spermatozoa, while ENUinduced a very high incidence of prenatal anomaliesafter spermatogonial treatment. In contrast to theprevious publications, in which x-rays were used, therewas a clear, almost linear, increase of anomalies in thedose range of 0–216 rad after spermatogonial exposure.There is a significant increase in the frequency ofmalformations after exposure to x-rays during spermato-genesis (Table 9), but the increase is numerically smallas compared with the frequency of induced malforma-tions after the exposure of developing embryos duringorganogenesis to similar doses of radiation. This in-crease in the frequency of malformations was observedin the fetuses but was not observed in the live-bornmouse fetuses (Table 9).
A number of laboratories have attempted to performa risk assessment of the mutagenic effect of variousenvironmental agents. This is not an easy task becausethe actual impact of the mutagenic effects is related todose, time of gonadal development, and time afterexposure at which conception occurs. Furthermore, thebest methodology for determining human risks is toutilize human epidemiological data. The problem ofusing human data is that you have to accept theexposures to which the population has been exposed,even when the exposures are very low. When dealingwith low-risk phenomena, one needs large populationsto demonstrate an effect. In many of the human epide-miological studies, the populations are so small thateven if there were a small reproductive effect, it wouldnot be discerned. Animal investigators have capitalizedon this deficiency because they can use multiple doses
that are high and they can permit conception at timesafter exposure that will maximize any genetic or cytoge-netic effects.
Van Buul (’84) studied the enhancement of radiosen-sitivity for the induction of translocations in mousestem cell spermatogonia after treatment with chemo-therapeutic agents. The effects of pretreatment of mousespermatogonial stem cells with cyclophosphamide (100and 200 mg/kg) and Adriamycin (2.5, 5, and 7 mg/kg) onthe induction of chromosomal translocations by highdoses (800 or 900 rad) of x-rays applied 24 h later aresummarized in Table 10. Both compounds were able toalter the chromosomal radiosensitivity of survivingstem cells. The authors concluded that depletion ofdifferentiated spermatogonia is sufficient for triggeringstem cells into a more radiosensitive phase.
The mutagenic potential of 125I was studied by Lavuet al. (’84) using the dominant lethal (DL) test (Table11). Dominant lethality represents embryonic deathresulting from the chromosomal damage in the gametesof parents. Significant preimplantation losses wereobserved. Dead implantations per pregnant female inthe isotope-treated groups showed a significant in-crease from controls indicating induced levels of postim-plantation losses. All the stages of spermatogenesis(i.e., spermatozoa, spermatid, spermatocyte, and sper-matogonia) were found to be sensitive to the inductionof post-implantation losses, the spermatid stage beingthe most sensitive.
Estimating human genetic risks from in vivoanimal experiments
The following investigators have designed experi-ments in an effort to convert the results of exposure ofanimals to mutagenic agents into actual human clinicalgenetic risks. Benova et al. (’85) (Table 12) used ionizingradiation as the mutagen in mice, with g-rays, deuter-ons, and neutrons. Using conditions that would maxi-mize the genetic manifestations in the F1 generation,the investigators estimated the genetic results of expos-
TABLE 9. Comparison of anomaly rate in the F1offspring after spermatogonial treatment with x-rays
Age attreatment F1 fetuses
Live-bornmice
Day 8 embryo 0/60 (0.0) 0/255 (0.0)Day 14 fetus 1/72 (1.4) 0/175 (0.0)21 days 5/297 (1.7) 0/62 (0.0)63–65 days 9/496 (1.8) 0/239 (0.0)
Modified from Nomura (’88).
TABLE 10. Induction of chromosomal aberrations instem cell spermatogonia and testis weights after
combined treatments with Adriamycin orcyclophosphamide and X-rays with a 24-hr intervalbetween exposure to the chemical and irradiation
Treatment*No. ofmice
No. ofcells
analyzed
%translo-cations6SEM
Testisweights
in mg 6 SEM
0 11 2,200 0.1 6 0.1 133 6 5CP 200 mg/kg 5 1,000 0.2 6 0.1 135 6 9A 5 mg/kg 5 1,000 0.7 6 0.2 107 6 8
800 rad 5 930 10.5 6 2.6 79 6 10CP 200 mg/kg 1
800 rad 5 950 14.9 6 1.2 74 6 8A 7 mg/kg 1
800 rad 5 900 21.5 6 5.8 57 6 7
*CP, cyclophosphamide; A, Adriamycin.Modified from Van Buul (’84).
190 R.L. BRENT
ing a population to 0.01 Gy (1 rad) (Table 12). Even ifthese results were transformed into genetic diseaseswith clinical manifestations, it would be impossible toidentify the increase in genetic disease unless therewas a very large number of individuals in the exposedpopulation. For example, Benova et al. (’85) predictedthat 0.01 Gy would result in 39 additional transloca-tions per million births (Table 12). Since there areapproximately 5,000 chromosomal abnormalities permillion live births, it is obvious that even if the exposedand control populations were each, one million, statisti-cal analysis would not differentiate between 5,000 and5,039 karyotpe abnormalities. Similarly, 25 inducedclinical abortions would be unidentifiable in one millionbirths containing 150,000 spontaneous abortions (Tables3, 14).
The International Commission for Protection AgainstEnvironmental Mutagens and Carcinogens (ICPEMC)has issued a number of reports dealing with the basicscience of mutagenesis and the impact of environmen-tal mutagenesis on human disease. The Committee 4Final Report dealt with the estimation of genetic risksfrom environmental mutagens (Lyons, ’85) (Tables 13,14). The committee concluded that in estimating thegenetic hazards of environmental mutagens there aremajor problems in extrapolating from experimentaldata to human situations. While the committee sug-
gested methods of improving our ability to extrapolatefrom animal data, they also believed that we were farfrom achieving that goal. The Committee stated, ‘‘Atpresent extrapolation can rarely be justified exceptwhere there are data on mutagenicity in the germ cellsof animals.’’ Thus the committee had little confidence inutilizing mutation data from in vitro cellular systemsfor determining mutagenic risks in humans. They alsosuggested that the genetic dose of a mutagen may besignificantly different from the exposure dose. Utilizingin vivo animal data the committee estimated the Rela-tive Mutagenic Effect, which is the ratio of the sponta-neous plus induced mutations divided by the spontane-ous mutation rate (Tables 3, 14). The estimates indicatethat the risks for most of the mutagenic drugs that werestudied were greater during the first three months aftertreatment. The difficulty with these data is that we donot have any human epidemiological data that agreewith the estimates obtained from animal research.First-generation genetic disease predicted from animaldata indicates increases in genetic disease from 2% to
TABLE 11. Number and percentage distribution of dead implantation pregnantfemale mice mated with males treated with different doses of iodine125
Group(mCi)
Pregnantfemales
No. of deadimplantations
No. of dead implantations per female
No. of pregnant females with dead implantations0 1 2 3 4 5
Control 393 139 213 101 31 7 0 054.2% 25.7% 7.9% — — —
5 357 183 174 115 38 18 6 648.7% 32.2% 10.6% 5.0% 1.7% 1.7%
10 354 205 150 119 56 15 8 742.3% 33.6% 15.8% 4.2% 2.3% 2.0%
15 355 208 147 101 70 20 4 1341.4% 28.5% 19.7% 5.6% 1.1% 3.7%
Modified from Lavu et al. (’84).
TABLE 12. Genetic radiation risk assessment based onexperimental mutagenesis in laboratory animal (mice)
1. Estimated risk of 0.01 Gy g-irradiation39 translocations/million conceptions5 cases of multiple congenital malformations25 additional clinical abortions/million conceptions49 abortions in undiagnosed pregnancies
2. Estimated risk of chronic g-irradiation of 1.3 mrad/minute, 10 mrad/min and 17 mrad/min was 3–10 timesless effective.
3. 4.2-GeV deuterons proved inferior in effectiveness to g-ir-radiation.
4. Chronic exposure to 4.1-MeV neutrons, 80 mrad/min, was7 times as effective as chronic g-irradiation.
Modified from Benova et al. (’85).
TABLE 13. Cases of genetic disease expected to beinduced per 106 births with 3 months treatment with
some cytotoxic drugs, and the relative risk to beexpected in the exposed population*
Drug
First three months (postmeiotic stages)
Generation1
Generation2
Total allgenerations
Cases RR Cases RR Cases RR
Adria-mycin 9,000 1.09 7,600 1.07 60,000 1.6
CP 180,000 2.7 150,000 2.4 1,200,000 12MMC 2,100 1.02 1,800 1.02 14,000 1.1Myleran — — — — — —Natulan 31,000 1.3 26,000 1.25 210,000 3.0TEM 1,000,000 10 840,000 9.0 6,600,000 64Thio TEPA 76,000 1.7 63,000 1.6 510,000 5.9
CP, cyclophosphamide; MMC, mitomycin C. RR, relative risk,assuming normal incidence of 105,000 cases of genetic diseaseper 106 live births.*Results have been rounded to two significant figures.Modified from Lyon et al. (’85).
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 191
70% during the first three postmeiotic months, depend-ing on the drug (Table 13). The total number of patientswith induced genetic disease over many generationswould be substantial, provided continuous or pro-tracted exposure to the mutagenic agent. If conceptionoccurs after the first three postmeiotic months from thediscontinuation of treatment, the incidence of inducedgenetic disease is diminished (Table 14). There are nohuman epidemiological data to support these esti-mates. In fact, the data available would indicate thatthe risks are lower.
Explanation of the marked difference in theclinical manifestation of the carcinogenic and
genetic risks of ionizing radiation
It is quite apparent from the animal studies thatinfertility, decreased fertility, chromosomal aberra-tions, and point mutations can be produced by exposingthe gonads to radiation before conception. The monu-mental investigations of Russell (’77), using hundredsof thousands of mice, indicated that large populationsof mice had to be used in order to quantitate the geneticeffects of preconception radiation. Larger populationswill be necessary to demonstrate an increase in themutation rate and the occurrence of genetic disease inhumans because the clinical manifestations of themutagenic effects are difficult to demonstrate in humanpopulations. Yet the irradiated populations in Hiro-shima and Nagasaki readily demonstrated an increasein the incidence of cancer, but the investigators hadgreat difficulty in demonstrating a statistically in-creased incidence of clinical genetic disease. There arenumerous instances in which epidemiological studieshave demonstrated a causal relationship between expo-sures to ionizing radiation, chemicals, and drugs andan increase in cancer in the exposed population. Popula-tions exposed to the same ‘‘mutagenic’’ agents rarely
demonstrate an increase in clinical genetic disease.Why?
Somatic cells that acquire new mutations either‘‘spontaneously’’ or from exposure to environmentalmutagens need only to survive and grow. While it islikely that many somatic cells that mutate may die,some survive and develop into malignant or benigntumors. The cause of the malignancy may be due tosmall or large losses of chromosome material or pointmutations in the DNA.
Gonadal cells have a much more complicated andtortuous path to traverse before mutagenic effects aremanifested in clinical genetic disease in the F1 genera-tion (Brent, ’94). During the process of spermatogen-esis, oogenesis, and embryonic development, there isvery likely a preferential loss of cells with new muta-tions (chromatin loss, point mutations) that have oc-curred spontaneously or have been induced by environ-mental agents. After ionizing radiation exposure toproliferating gonadocytes, two important effects occursimultaneously: (1) the induction of mutations in surviv-ing cells, a stochastic phenomenon, and (2) cell death, athreshold phenomenon, with the incidence of cell deathbeing directly related to the radiation dose. Figure 3demonstrates the relationship between the risks of celldeath in proliferating cells and the risk of mutation.Using a conservative figure for the radiation risk ofmutation (1027/rem/locus), it can be observed that celldeath becomes a significant effect in proliferating cellswhen the radiation dose induces mutation equal to thespontaneous mutation rate. The competition betweenthese two effects is theoretically another reason thatmutagenic effects are difficult to demonstrate whencompared to carcinogenic effects in human populations.I have used the term biologic filtration to describe the
TABLE 14. Cases of genetic disease expected to beinduced per 106 births with 3 months treatment with
some cytotoxic drugs, and the relative risk to beexpected in the exposed population*
Drug
After 3-month interval (spermatogonial stages)
Generation1
Generation2
Total allgenerations
Cases RR Cases RR Cases RR
Adriamycin 36,000 1.3 30,000 1.3 240,000 3.3CP 4,800 1.05 4,200 1.04 42,000 1.4MMC 6,400 1.06 5,600 1.05 56,000 1.5Myleran 2,500 1.02 2,200 1.02 22,000 1.2Natulan 18,000 1.2 16,000 1.2 160,000 2.5TEM 25,000 1.2 22,000 1.2 220,000 3.1Thio TEPA 20,000 1.2 17,000 1.2 130,000 2.2
*CP, cyclophosphamide; MMC, mitomycin C; RR, relativerisk, assuming normal incidence of 105,000 cases of geneticdisease per 106 live births. Results have been rounded to twosignificant figures.Modified from Lyon et al. (’85).
Fig. 3. Relationship between the risks of cell death in proliferatingcells and the risk of mutation. Using a conservative figure for theradiation risk of mutation (1027/rem/locus), it can be observed that celldeath becomes a significant effect in proliferating cells when theradiation dose induces mutation equal to the spontaneous mutationrate. The competition between these two effects is theoreticallyanother reason that mutagenic effects are difficult to demonstrate ascompared with carcinogenic effects in human populations.
192 R.L. BRENT

ameliorating effects of the developmental process inreducing the manifestations of genetic effects, becauseof the propensity for the genetically damaged cells to belost during gonadogenesis, fertilization, and organogen-esis. These preferential losses would explain the quan-titative differences between the clinical manifestationsof malignancy and clinical genetic disease after expo-sure to mutagenic agents. For example:
1. These affected germ cells may be lost during gameto-genesis.
2. The mature egg or sperm may have a decreasedcapacity to be fertilized or to fertilize.
3. The embryos produced from these affected eggs andsperm may result in embryonic loss during preim-plantation or early organogenesis. In most cases, thewomen may not know that they were pregnant.
4. If there is a lapse in time between the exposure toreproductive toxicants and insemination and fertil-ization, some of the affected gonadocytes may be lost(bridge formation, lethal mutations).
5. In many instances the mutagenic exposure to thehuman populations may be so low that the calcu-lated risks would not result in a change in theincidence of genetic disease.
Thus, it is not surprising that clinical manifestations ofpreconception exposures to mutagenic agents havebeen difficult to demonstrate in the F1 generation inhumans.
But it is not surprising that some scientists haveignored the animal and human data and have indicatedthat genetic risks from environmental agents are amajor health problem. These discussions occurred dur-ing the 1960s and 1970s and are summarized in Table15. Certainly, one cannot ignore the potential risk ofgenetic disease from inherited or environmentally in-duced agents, but it is important to use the bestestimates of these genetic risks for the purposes ofcounseling patients and establishing priorities for ex-pending financial and personnel resources on healthcare matters. The best advice on these issues comesfrom Charlotte Auerbach (Auerbach, ’71) (Table 15):
I think it is absolutely essential that we do notdelude ourselves about the magnitude and complex-ity of our task. The general public is easily scaredand when they are scared, they may form pressuregroups to push governmental agencies into action.These agencies are scientifically naive and have torely on our advice. We should be very careful not togive advice that is itself naive; that is advice basedon oversimplified tests and facile interpretations.
Genetic risks: Preconception exposure of EMF,microwave, and ultrasound
The previous discussion dealing with the reproduc-tive risks of preconception radiation dealt exclusivelywith the risks of ionizing radiation, a physical agentthat has been demonstrated to have mutagenic effects
in animal studies and in vitro systems. Since Mullerdiscovered the mutagenic properties of ionizing radia-tion, genetic studies and carcinogenic studies havedemonstrated its mutagenic biologic properties. Yet theprevious discussion pointed out that in order to demon-strate the mutagenic risks of ionizing radiation that aremanifested in genetic disease, large populations areneeded, because the risks are so small.
In vitro studies, animal studies, and some epidemio-logical studies have examined the genetic effects ofnonionizing radiation, and the results have been incon-sistent. The main theme of this presentation relates tothe principles of biologic plausibility. Furthermore, theepidemiological and animal studies pertaining to thereproductive effects of low-frequency EMF, microwaves,and ultrasound are discussed in other presentations inthis symposium (Brent, ’99; Jensh and Brent, ’99;O’Connor, ’99; Robert, ’99; Ziskin, ’99). This presenta-tion deals only with the biologic plausibility that nonion-izing radiation can result in genetic effects.
The occupational, medical, and population exposuresof the above-mentioned forms of nonionizing radiationdo not have the ability to alter DNA without affecting
TABLE 15. Controversy over the magnitude of thegenetic risk from environmental agents
‘‘The threat of genetic damage is our number one health prob-lem.’’ (Legator, ’68)
‘‘There is no realistic way to predict a safe human level of asubstance shown to be teratogenic, carcinogenic or muta-genic in animals.’’ (Epstein, ’71)
‘‘Chemical mutagenesis is certainly a very small problem aswe see it at present. We view it as 1% of the very broadproblem of human genetics. We do not propose setting upwhatever Dr. Crow means by a monitoring system.’’ (Shan-non, ’68)
‘‘If the public flatly refuses to take decisive action on thebasis of the massive volumes of data linking cigarettesmoking to lung cancer, how can you expect people to actvigorously on the more hazy and abstruse things like achemical that may or may not be mutagenic in man.’’(Crow, ’69)
‘‘I think it is absolutely essential that we do not delude our-selves about the magnitude and complexity of our task.The general public is easily scared and when they arescared, they may form pressure groups to push govern-mental agencies into action. These agencies are scientifi-cally naive and have to rely on our advice. We should bevery careful not to give advice that is itself naive; that isadvice based on oversimplified tests and facile interpreta-tions.’’ (Auerbach, ’71)
‘‘The world is full of mutagens, carcinogens and reproductivetoxins, and it always has been. The important issue is thehuman exposure dose. Fortunately the exposure is usuallyminuscule’’ (Ames, ’89)
‘‘Nature’s toxic chemicals are the major carcinogens andmutagens ingested by humans. These natural chemicals inplants are present at a level 10,000 times the prevalence ofman-made pesticide residues. There is a tendency forlaymen to think of chemicals as being only man-made andto characterize them as toxic.’’ (Ames, ’89)
‘‘It would be foolish to advise anything other than extremecaution over the exposure of fathers to chemical mutagenswhen our understanding of the quantitative risks to futuregenerations is so rudimentary.’’ (Brown, ’85)
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 193

other cellular functions and there toxic effects aredeterministic (threshold effects) rather than stochasticeffects (Table 1). Whereas some in vitro studies havesuggested that these forms of radiation have mutagenicpotential, the cytotoxic effects that accompany themutagenic effect are not present at the usual exposuresof all these forms of nonionizing radiation. Therefore, ifgenetic effects are to be manifested from preconceptionexposures to ultrasound, microwaves, and low-fre-quency EMF, the risks have to be considerably smallerthan the risks from ionizing radiation. Epidemiologicalstudies and animal studies support this conclusion.
Summary of preconception radiation effects
There has been continued interest in the impact ofenvironmental reproductive toxicants including radia-tion, on the male and female reproductive capacity.Although the interest has been long-standing, society’sinterest in environmental toxicity, radiation effects,improved work safety, and the proliferation of newchemicals has more recently focused on this area. Therehas been a long-standing polarization of feelings amongscientists with regard to the risk of preconceptionexposure to reproductive toxicants (Brent, ’72a) (Table15).
What can we say with certainty about the impact ofpreconception exposures on the reproductive capacityof exposed human populations?
First, there are substantial data from animal experi-ments and epidemiological studies that indicate thationizing radiation, drugs, and chemicals can decreasemale and female fertility by decreasing the number ofsperm and or eggs. These results have been observed inoccupational epidemiological studies and in patientpopulations who have survived cancer chemotherapy orradiation therapy. In the population of male cancersurvivors, there is a suggestion that the males are atgreater risk for infertility than the females. Thus,subfertility or infertility in men and women are provenrisks from certain exposures to ionizing radiation, somechemicals, and chemotherapeutic agents.
Second, there are extensive mammalian animal inves-tigations to indicate that males and females exposed tohigh exposures of ionizing radiation and some muta-genic chemicals have offspring with an increase indominant lethals, point mutations, and chromosomalabnormalities. It is true that the maximum manifesta-tion of some of these effects varies with the stage ofsperm and egg development and the dose of theseagents. Yet, in animal experiments using very lowexposures to mutagenic agents, experimental offspringfrequently cannot be differentiated from the controls,even when the experimental conditions have beenestablished to maximize the mutagenic effect.
Third, patients who have survived therapeutic dosesof radiation and chemotherapy as children or youngadults may have problems with infertility, but thefertile population does not appear to have a measurablyincreased risk of spontaneous abortion or offspring with
congenital malformations. It is true that, even in thelarger studies, the populations are small. It is thereforepossible that if there were a small risk of induction ofchromosomal aberrations and genetically caused con-genital malformations, these studies would not be ableto recognize them. Furthermore, patients frequentlywait years after cancer treatment before having chil-dren and, of course, there may be two decades betweenchildhood cancer treatment and procreation. The verylow risk of transmitting induced mutations to theoffspring of treated cancer survivors is a reflection ofthe very low risk of mutagenesis from environmentalmutagens in general.
Fourth, the concept of ‘‘biologic filtration’’ is proposedto explain why mutagenic effects of ionizing radiationare readily demonstrated to be the cause of cancer inexposed animals and humans and why clinical geneticdisease in the F1 generation is so difficult to demon-strate in human populations.
Fifth, a great deal of attention was not directedtoward the mutagenic risks of preconception exposuresto low-frequency EMF, microwaves, and ultrasound,because these forms of radiation will be discussed byother authors in this symposium. Furthermore, thispresentation has concentrated primarily on applicationof basic science principles and biological plausibility.These forms of nonionizing radiation do not have theability to alter DNA without affecting other cellularfunctions, they are not cytotoxic at exposures whichconcern us clinically and their toxic effects are determin-istic (threshold effects), rather than stochastic (Table1). While some in vitro studies have suggested thatthese forms of radiation have mutagenic potential, thecytotoxic effects that accompanies the mutagenic effectis not present at the usual exposures of all these formsof nonionizing radiation. Therefore, if genetic effectsare to be manifested from preconception exposures toultrasound, microwaves, and low-frequency EMF, therisks have to be considerably smaller than those fromionizing radiation, and actually there may not be anyincreased risks. The epidemiological studies and ani-mal studies support this conclusion.
POSTCONCEPTION (INTRAUTERINE)EFFECTS OF RADIATION: IONIZINGRADIATION, LOW-FREQUENCY EMF,MICROWAVES, AND ULTRASOUND
Postconception effects of reproductive toxicants
While there is significant overlap between the repro-ductive effects of preconception and postconceptionreproductive toxicants, there are important qualitativeand quantitative differences (Tables 4, 5). The reproduc-tive effects of intrauterine exposure to reproductivetoxicants includes: (1) pregnancy loss (abortion, still-births), (2) congenital malformations (anatomical de-fects), (3) neurobehavioral abnormalities (i.e., mentalretardation), (4) fetal growth retardation (reversibleand irreversible), and (5) cancer (stochastic phenom-enon). All these effects are threshold phenomena with
194 R.L. BRENT
the exception of the risk of cancer (Table 5), whichmeans that regardless of the reproductive toxicant,there will be a no-effect dose for the first four reproduc-tive toxic effects.
Of all the forms of radiation that are being discussedin this symposium, the reproductive effects of ionizingradiation exposure during pregnancy has been the mostextensively studied, and for which we have the mostreliable information. Furthermore, the threshold dose,period of sensitivity and teratogenic syndrome havebeen described for ionizing radiation. (Brent, ’76, 80a,b,’89, ’94)
Effects of in utero ionizing radiation
There have been many experiments and observationsabout the effects of radiation on the developing embryo.
The subject is summarized briefly here. For furtherdetails the reader is referred to comprehensive reviewsconcerning the effects of various forms of radiation onthe developing embryo and fetus. (Brent, ’70, ’72b, ’76,’77, ’80a,b, ’89; Brent and Gorson, ’72; Brent et al., ’91;Miller, ’90; Russell and Russell, ’54; Sikov, ’92).
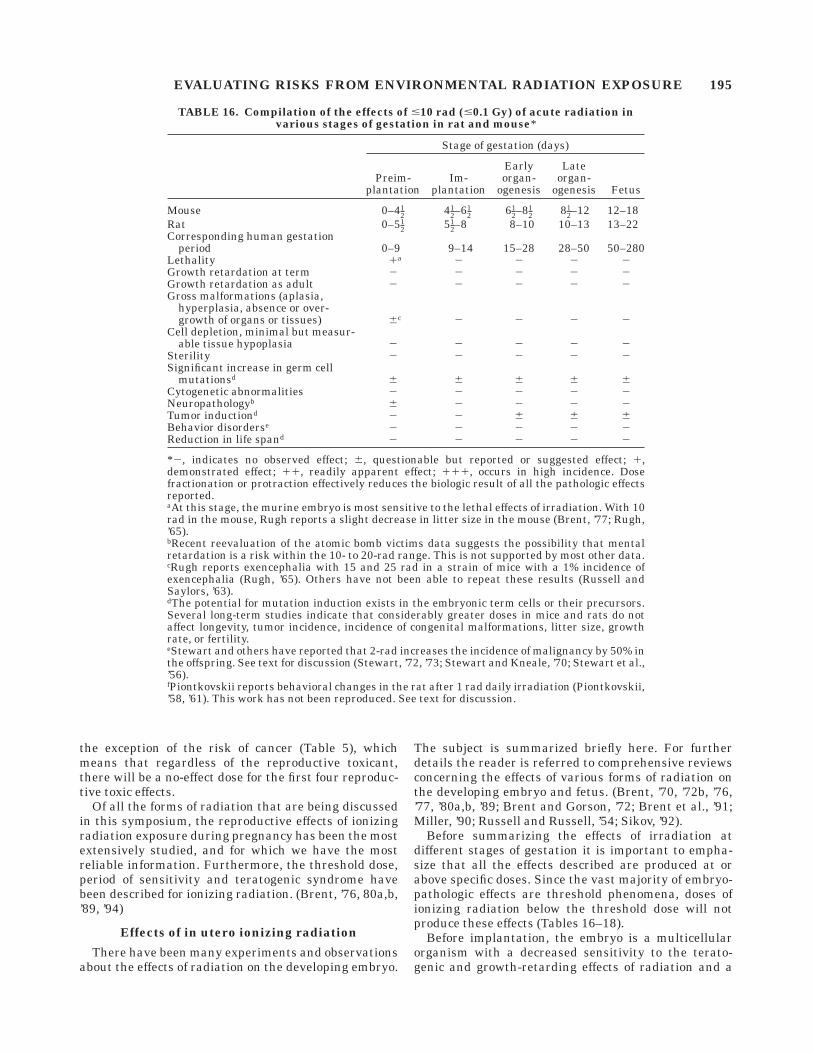
Before summarizing the effects of irradiation atdifferent stages of gestation it is important to empha-size that all the effects described are produced at orabove specific doses. Since the vast majority of embryo-pathologic effects are threshold phenomena, doses ofionizing radiation below the threshold dose will notproduce these effects (Tables 16–18).
Before implantation, the embryo is a multicellularorganism with a decreased sensitivity to the terato-genic and growth-retarding effects of radiation and a
TABLE 16. Compilation of the effects of #10 rad (#0.1 Gy) of acute radiation invarious stages of gestation in rat and mouse*
Stage of gestation (days)
Preim-plantation
Im-plantation
Earlyorgan-
ogenesis
Lateorgan-
ogenesis Fetus
Mouse 0–412 41
2–612 61
2–812 81
2–12 12–18Rat 0–51
2 512–8 8–10 10–13 13–22
Corresponding human gestationperiod 0–9 9–14 15–28 28–50 50–280
Lethality 1a 2 2 2 2Growth retardation at term 2 2 2 2 2Growth retardation as adult 2 2 2 2 2Gross malformations (aplasia,
hyperplasia, absence or over-growth of organs or tissues) 6c 2 2 2 2
Cell depletion, minimal but measur-able tissue hypoplasia 2 2 2 2 2
Sterility 2 2 2 2 2Significant increase in germ cell
mutationsd 6 6 6 6 6Cytogenetic abnormalities 2 2 2 2 2Neuropathologyb 6 2 2 2 2Tumor inductiond 2 2 6 6 6Behavior disorderse 2 2 2 2 2Reduction in life spand 2 2 2 2 2
*2, indicates no observed effect; 6, questionable but reported or suggested effect; 1,demonstrated effect; 11, readily apparent effect; 111, occurs in high incidence. Dosefractionation or protraction effectively reduces the biologic result of all the pathologic effectsreported.aAt this stage, the murine embryo is most sensitive to the lethal effects of irradiation. With 10rad in the mouse, Rugh reports a slight decrease in litter size in the mouse (Brent, ’77; Rugh,’65).bRecent reevaluation of the atomic bomb victims data suggests the possibility that mentalretardation is a risk within the 10- to 20-rad range. This is not supported by most other data.cRugh reports exencephalia with 15 and 25 rad in a strain of mice with a 1% incidence ofexencephalia (Rugh, ’65). Others have not been able to repeat these results (Russell andSaylors, ’63).dThe potential for mutation induction exists in the embryonic term cells or their precursors.Several long-term studies indicate that considerably greater doses in mice and rats do notaffect longevity, tumor incidence, incidence of congenital malformations, litter size, growthrate, or fertility.eStewart and others have reported that 2-rad increases the incidence of malignancy by 50% inthe offspring. See text for discussion (Stewart, ’72, ’73; Stewart and Kneale, ’70; Stewart et al.,’56).fPiontkovskii reports behavioral changes in the rat after 1 rad daily irradiation (Piontkovskii,’58, ’61). This work has not been reproduced. See text for discussion.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 195
greater degree of sensitivity to the lethal effects ofirradiation at this early stage (Brent and Bolden, ’67;Russell, ’50; Russell and Russell, ’50) (Tables 16–18).This stage, the preimplantation period, has been re-ferred to as ‘‘the all-or-none period,’’ since radiation ismore likely to kill the embryo than result in a livemalformed newborn. This does not indicate that malfor-
mations cannot be produced, but the cytogenetic abnor-malities and malformations, if produced, result in ahigh incidence of mortality. There are reports in theliterature indicating that radiation can produce malfor-mations at this early stage of development that surviveto term in the mouse or rat. Usually the malformationsare confined to a small group of abnormalities that
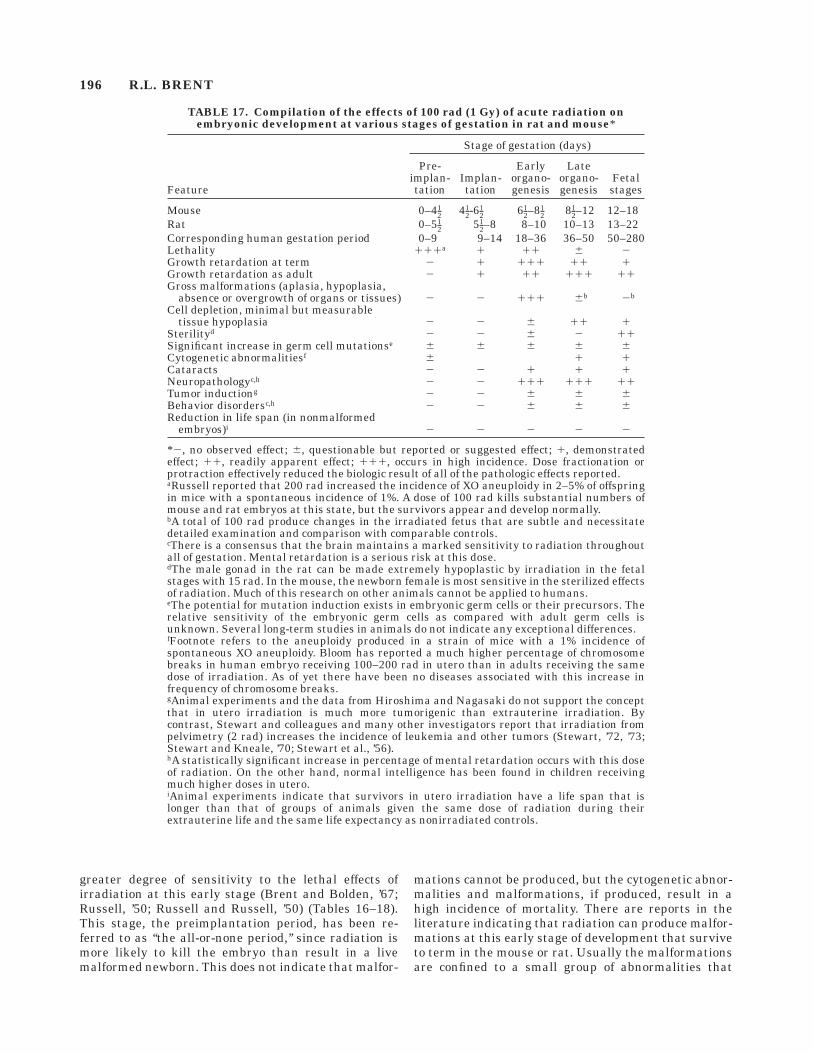
TABLE 17. Compilation of the effects of 100 rad (1 Gy) of acute radiation onembryonic development at various stages of gestation in rat and mouse*
Feature
Stage of gestation (days)
Pre-implan-tation
Implan-tation
Earlyorgano-genesis
Lateorgano-genesis
Fetalstages
Mouse 0–412 41
2-612 61
2–812 81
2–12 12–18Rat 0–51
2 512–8 8–10 10–13 13–22
Corresponding human gestation period 0–9 9–14 18–36 36–50 50–280Lethality 111a 1 11 6 2Growth retardation at term 2 1 111 11 1Growth retardation as adult 2 1 11 111 11Gross malformations (aplasia, hypoplasia,
absence or overgrowth of organs or tissues) 2 2 111 6b 2b
Cell depletion, minimal but measurabletissue hypoplasia 2 2 6 11 1
Sterilityd 2 2 6 2 11Significant increase in germ cell mutationse 6 6 6 6 6Cytogenetic abnormalitiesf 6 1 1Cataracts 2 2 1 1 1Neuropathologyc,h 2 2 111 111 11Tumor inductiong 2 2 6 6 6Behavior disordersc,h 2 2 6 6 6Reduction in life span (in nonmalformed
embryos)i 2 2 2 2 2
*2, no observed effect; 6, questionable but reported or suggested effect; 1, demonstratedeffect; 11, readily apparent effect; 111, occurs in high incidence. Dose fractionation orprotraction effectively reduced the biologic result of all of the pathologic effects reported.aRussell reported that 200 rad increased the incidence of XO aneuploidy in 2–5% of offspringin mice with a spontaneous incidence of 1%. A dose of 100 rad kills substantial numbers ofmouse and rat embryos at this state, but the survivors appear and develop normally.bA total of 100 rad produce changes in the irradiated fetus that are subtle and necessitatedetailed examination and comparison with comparable controls.cThere is a consensus that the brain maintains a marked sensitivity to radiation throughoutall of gestation. Mental retardation is a serious risk at this dose.dThe male gonad in the rat can be made extremely hypoplastic by irradiation in the fetalstages with 15 rad. In the mouse, the newborn female is most sensitive in the sterilized effectsof radiation. Much of this research on other animals cannot be applied to humans.eThe potential for mutation induction exists in embryonic germ cells or their precursors. Therelative sensitivity of the embryonic germ cells as compared with adult germ cells isunknown. Several long-term studies in animals do not indicate any exceptional differences.fFootnote refers to the aneuploidy produced in a strain of mice with a 1% incidence ofspontaneous XO aneuploidy. Bloom has reported a much higher percentage of chromosomebreaks in human embryo receiving 100–200 rad in utero than in adults receiving the samedose of irradiation. As of yet there have been no diseases associated with this increase infrequency of chromosome breaks.gAnimal experiments and the data from Hiroshima and Nagasaki do not support the conceptthat in utero irradiation is much more tumorigenic than extrauterine irradiation. Bycontrast, Stewart and colleagues and many other investigators report that irradiation frompelvimetry (2 rad) increases the incidence of leukemia and other tumors (Stewart, ’72, ’73;Stewart and Kneale, ’70; Stewart et al., ’56).hA statistically significant increase in percentage of mental retardation occurs with this doseof radiation. On the other hand, normal intelligence has been found in children receivingmuch higher doses in utero.iAnimal experiments indicate that survivors in utero irradiation have a life span that islonger than that of groups of animals given the same dose of radiation during theirextrauterine life and the same life expectancy as nonirradiated controls.
196 R.L. BRENT

occur spontaneously in that species. This has beenreferred to as an epigenetic effect, rather than a classi-cal teratogenic effect. During early organogenesis theembryo is very sensitive to the growth-retarding, terato-genic, and lethal effects of irradiation, but these em-bryos have the ability to recover somewhat from thegrowth-retarding effects in the postpartum period(Rugh, ’62; Russell, ’56; Russell and Russell, ’54).During the early fetal period, the fetus has diminishedsensitivity to multiple organ teratogenesis but retainsCNS sensitivity; it is growth retarded at term andrecovers poorly from the growth retardation in thepostpartum period. During later stages, the fetus is notgrossly deformed by radiation but can sustain perma-nent cell depletion of various organs and tissues if theradiation exposure is high enough (Brent, ’77; Brentand Gorson, ’72) (Tables 16–18).
One can postulate many mechanisms for the effects ofirradiation, including cell death, mitotic delay, distur-bances of cell migration, and others (Table 19). Of thosemechanisms listed in Table 19, the most likely mecha-nisms pertaining to ionizing radiation-induced embry-opathy are: (1) cell death or mitotic delay beyond therecuperative capacity of the embryo or fetus (ionizingradiation, chemotherapeutic agents, alcohol); (2) inhibi-tion of cell migration, differentiation and cell communi-cation; and (3) interference with histogenesis by pro-
cesses such as cell depletion, necrosis, calcification, orscarring. Until we know more about mechanisms in-volved in embryopathology in general, it is difficult todetermine those mechanisms most important in theprocess of radiation-induced embryopathologic disor-ders. Furthermore, the same pathogenic mechanismsmay not be primarily operative at all stages of gesta-tion. Radiation-induced cell death may be minimallyimportant at one stage because of the embryo’s abilityto replace the killed cells. At another stage of gestation,cell death may be a primary factor because the embryohas lost the ability to replace dead cells, and the fetusmay be permanently cell depleted. This is probably oneof the important mechanisms in the production ofmental retardation during the second trimester ofhuman development.
Other radiation-induced effects that have been in-voked to explain embryopathologic conditions are cyto-genetic abnormalities and somatic mutations (Russellet al., ’63; Russell and Major, ’57). Cytogenetic abnor-malities may be responsible for preimplantation deathin the irradiated mammalian zygote (Brent and Bolden,’67; Rugh, ’65; Russell et al., ’63), but point mutationsare less likely to be a contributing factor to abnormalmorphogenesis. If one uses the known radiation-induced mutation rate in mammalian cells, as deter-mined by Russell and colleagues in the mouse (Russell,’50, ’56; Russell et al., ’63; Russell and Major, ’57;Russell and Russell, ’50; Russell and Saylors, ’63) forestimating potential teratogenicity, it becomes per-fectly obvious that radiation-induced point mutationcould not account for even a small proportion of radia-tion-induced teratogenicity unless it simply involvedcell death. Neither phenomenon has been studied ad-equately at every stage of mammalian gestation.
Radiation effects may be manifested acutely andresult in cell death, embryonic death, or teratogenesis.Other effects may not be immediately obvious and canbe measured or ascertained only in the postpartum oradult period. For instance, neuronal depletion, infertil-ity, tissue hypoplasia, neoplasia, or shortening of lifespan are phenomena that can be evaluated only in the
TABLE 18. Estimation of the risks of radiation in the human embryo based on human epidemiological studiesand mouse and rat radiation embryological studies*
Embryonicage(days)
Minimumlethal dose
(rad)Approximate
LD50 (rad)
Minimum dosefor permanent
growth retardationin the adult
(rad)
Minimum dosefor grossanatomic
malformations(rad)
Increasedincidenceof mental
retardation
Minimum dosefor the induction
of genetic carcinogenic,and minimal cell
deletion phenomena
1–5 10 ,100 No effect in survivors — Unlikely Unknown18–36 25–50 140 20–50 20 Severe CNS
malformationsUnknown
36–50 .50 200 25–50 50 Unknown Unknown50–110 .100 .200 25–50 ** .20 rad Unknown
To term .150 Same as mother .50 — :50 rad Unknown
*Recent information to be published by Otake and Schull (’84) suggest an increased risk at lower exposures.**Anatomic malformations of a severe type cannot be produced this late in gestation, except in the genitourinary system andtissue hypoplasia in specific organ systems, such as the brain and testes.
TABLE 19. Mechanisms of action of environmentalteratogens
1. Cell death or mitotic delay beyond the recuperativecapacity of the embryo or fetus (ionizing radiation, chemo-therapeutic agents, alcohol)
2. Inhibition of cell migration, differentiation, and cell com-munication
3. Interference with histogenesis by processes such as celldepletion, necrosis, calcification, or scarring
4. Biologic and pharmacological receptor-mediated develop-mental effects (i.e., etretinate, isotretinoin, retinol, sexsteroids, streptomycin, thalidomide)
5. Metabolic inhibition (i.e., warfarin, anticonvulsants,nutritional deficiencies)
6. Physical constraint, vascular disruption, inflammatorylesions, amniotic band syndrome
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 197

postpartum or adult organism (Brent, ’70, ’72; Brentand Bolden, ’61; Brent and Gorson, ’72; Cowen andGeller, ’60; Furchgott, ’63; Hicks and D’Amato, ’66;Murphree and Pace, ’60; Rugh and Wohlfromm, ’64a,b).
Human radiation teratogenesis, includingcentral nervous system effects from ionizing
radiation
The classic effects of ionizing radiation on the develop-ing mammal are gross congenital malformations, IUGR,and embryonic death. Each of these effects has adose-response relationship and a threshold exposurebelow which there is no difference between the irradi-ated and the control populations (Tables 16–18). Be-cause human pregnancies consist predominantly of asingle embryo, it would be unusual to see the classictriad of effects, although it is conceivable, that if thedose and timing were appropriate, a stillborn, growth-retarded, malformed embryo could result.
Growth retardation and CNS effects, such as micro-cephaly or eye malformations, are the cardinal manifes-tations of intrauterine radiation effects in humans. Theoccurrence of microcephaly is such a significant compo-nent of the effects of intrauterine radiation that a‘‘teratogen update’’ has been written about this subject(Miller and Mulvihill, ’76). A large number of reportsindicate that microcephaly is the most common malfor-mation observed in human beings randomly exposed tohigh doses of radiation during pregnancy (Dekaban,’68; Gentry et al., ’59; Goldstein and Murphy, ’29a,b;Miller and Mulvihill, ’76; Murphy and Goldstein, ’30;Plummer, ’52; Wood et al., ’67a,b,c; Yamazaki et al., ’54;Zappert, ’26). In Goldstein and Murphy’s reports 19 of75 irradiated embryos became microcephalic or hydro-cephalic children, having received an estimated dose of.100 rad as a minimum (Goldstein and Murphy, ’29a,b;Murphy and Goldstein, ’30). Almost all the microce-phalic children were mentally retarded. The followingmalformations were also reported: two infants withhypoplastic genitalia, one with cleft palate and one withhypospadias, an abnormality of the large toe, and anabnormality of the ear. There were many instances ofIUGR, and three neonates had various abnormalities ofthe eyes, including microphthalmia, cataracts, strabis-mus, retinal degeneration, and optic atrophy. The uniquefinding was absence of visceral, limb, or other malforma-tions unless there was IUGR, microcephaly (congenitalmalformations of the brain), or readily apparent eyemalformations. Dekaban (’68) reported that 22 infantsexposed between the 3rd to 20th weeks of humangestation were microcephalic or mentally retarded, orboth. The estimated protracted exposure was 250 rad(2.5 Gy), and all the malformed infants exhibitedgrowth retardation. The most frequent abnormalitieswere (1) small size at birth and stunted growth, (2)microcephaly, (3) mental retardation, (4) microphthal-mia, (5) pigmentary changes in the retina, (6) genitaland skeletal malformations, and (7) cataracts. All pa-tients had received hundreds of rads, having been
treated with radiation for various reasons, includingdysmenorrhea, menorrhagia, myomas, arthritis or tu-berculosis of the sacroiliac joint, and malignant tumorof the uterus or cervix.
Irradiation of the human fetus from diagnostic expo-sures of ,5 rad (0.05 Gy) have not been observed tocause congenital malformations or growth retardation(Kinlen and Acheson, ’68; Nokkentred, ’68; Tabuchi, ’64;Tabuchi et al., ’67; Vilumsen, ’70), but not all suchepidemiological studies are negative (Jacobsen andMellemgaard, ’88). These are extremely difficult studiesto perform, and it appears that the animal data supportthe contention that gross congenital malformations willnot be increased in a human pregnant populationexposed to #20 rad (0.2 Gy). This contention is strength-ened further by the fact that most human exposures toextensive diagnostic radiation studies are fractionatedor protracted. Such exposures are less likely to producemalformations than an acute exposure of low LETradiation (Brent, ’71; Brizzee and Brannon, ’72; Cop-penger and Brown, ’67; Gentry et al., ’59; Grahn andKratchman, ’63; Konerman, ’69; Kriegal and Langen-dorff, ’64; Laskey et al., ’73; Ronnback, ’65; Segallet al., ’64; Stadler and Gowen, ’64; Vorisek, ’65; Wesley,’60).
The animal and human data support the contentionthat exposures within the 5-rad (0.05-Gy) range wouldnot be expected to increase the incidence of anatomicmalformations, growth retardation, mental retarda-tion, or abortion. We have investigated a number offunctional and biochemical parameters in the rat, suchas thyroid function, liver function, and fertility, andhave not found them to be more sensitive to radiationthan the above-mentioned parameters. That does notmean, however, that some other parameter that has notyet been studied would not result in an effect fromlow-level radiation.
General principles of teratologyand developmental biology
When evaluating studies dealing with the reproduc-tive effects of any environmental agent, importantprinciples should guide the analysis of reproductivehuman and animal studies. Paramount to this evalua-tion is the application of the basic science principles ofteratology and developmental biology. These principlesare as follows:
1. Exposure to teratogens follows a toxicological dose-response curve. There is a threshold below which noeffect will be observed and, as the dose of theteratogen is increased, both the severity and fre-quency of reproductive effects will increase (Tables6, 16–18, Fig. 2).
2. The period of exposure is critical in determiningwhat effects will be produced and whether anyeffects can be produced by a known teratogen. The
198 R.L. BRENT

period of embryonic development to which an em-bryo is sensitive to teratogenic effects can varyconsiderably, depending on the teratogen. Someteratogenic effects have a broad, and others, a verynarrow period of sensitivity.
3. Even the most potent teratogenic agent cannotproduce every malformation.
4. Most teratogens have a confined group of congenitalmalformations that result after exposure during acritical period of embryonic development. This con-fined group of malformations is referred to as thesyndrome that describes the agent’s teratogeniceffect.
5. While a group of malformations may suggest thepossibility of certain teratogens, they cannot defini-tively confirm the causal agent. On the other hand,the presence of certain malformations can eliminatethe possibility that a particular teratogenic agentwas responsible.
Based on the information already provided regardingthe intrauterine effects of ionizing radiation, we canapply these principles to the four forms of radiationbeing discussed: ionizing radiation, low-frequency EMF,microwaves, and ultrasound.
Of the four forms of radiation, ionizing radiation isthe only agent that has been completely characterizedutilizing all the above-mentioned teratology principles.The period of sensitivity for the production of micro-cephaly, severe mental retardation, growth retarda-tion, and the reported anatomical malformations hasbeen described. The threshold dose for anatomicalmalformations is approximately 0.20 Gy during theperiod of maximum sensitivity. Some malformationsand other reproductive effects have higher no-effectdoses. There is a confined group of malformationsproduced by ionizing radiation above the thresholddose. The severity and frequency of developmentaleffects increase as the exposure increases and the LD50
varies with the stage of development (Tables 16–18).But a most important aspect of the reproductive
effects concerning ionizing radiation is that all thereported deleterious effects are biologically plausible,and there are basic science mechanisms to explain theresults (Table 19). If one examines the mechanismsresponsible for teratogenesis listed in Table 19, it isobvious that three of the mechanisms are not involvedin radiation-induced teratogenesis and three of themechanisms could contribute to the radiation inducedpathology. Several mechanisms are unlikely to play arole in radiation induced teratogenesis: (1) biologic andpharmacological receptor-mediated developmental ef-fects; (2) metabolic inhibition; and (3) physical con-straint, vascular disruption, inflammatory lesions, andthe amniotic band syndrome.
Ionizing radiation has no cellular specificity, exceptfor the fact that it preferentially affects rapidly prolifer-
ating cells. Metabolic inhibition involving particularpathways or mechanical interference are not propertiesof radiation, especially at exposures that we deal withclinically. On the other hand, (1) cell death or mitoticdelay beyond the recuperative capacity of the embryo orfetus; (2) inhibition of cell migration, differentiation,and cell communication; and (3) interference withhistogenesis by processes such as cell depletion, necro-sis, calcification or scarring have been demonstrated toaccount for the reported effects of ionizing radiation onthe developing embryo.
Therefore, when we use the criteria described inTable 2 to evaluate the conclusion that ionizing radia-tion has significant reproductive toxic effects, concor-dance can be demonstrated with most of five categoriesused in the evaluation process:
1. The epidemiological studies that have been per-formed have consistently reported that high expo-sures to radiation deleteriously affect the developingembryo and fetus. These studies have also demon-strated that the severity and frequency of the terato-genic effects increase with dose and that there arethreshold doses, below which no effects are pro-duced.
2. High-dose ionizing radiation produces a definablesyndrome which consists of mental retardation,growth retardation, and microcephaly, if the dose ishigh enough and the exposure occurs during theperiod of development sensitive to these developmen-tal defects. This syndrome is produced after theperiod of major organogenesis (8th postconceptionweek to the end of pregnancy, with the most sensi-tive period the 8th to 15th weeks postconception).Animal studies indicate that hydrocephalus, eyemalformation, heart and visceral malformationscould be produced with high doses during organogen-esis along with growth retardation. But we do nothave enough data to be certain of the radiationteratogenic syndrome produced from exposure dur-ing early organogenesis.
3. Many congenital malformations and syndromes couldnot be produced by ionizing radiation, regardless ofthe exposure or the stage of gestation.
4. Mammalian animal studies mimic the qualitativeand quantitative aspects of radiation effects ob-served in the human, allowing investigators to re-fine the dose response curve and qualitative effectsfrom radiation exposure of pregnant women. Thereason for this concordance among mammalian spe-cies is that radiation exerts its reproductive effectsby directly affecting the embryo (Brent, ’60; Brentand Bolden, ’67, ’68; Brent and McLaughlin, ’60).Thus, species differences that result in discordanteffects among mammals from chemical exposureoccur much less frequently with ionizing radiation,because differences in placental transport, absorp-tion, and metabolism do not pertain to ionizingradiation.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 199

In utero effects of ionizing radiation on the riskof mental retardation and cancer
Two developmental effects of ionizing radiation have notbeen completely resolved: the risk of mental retardationand cancer after in utero radiation. These two subjects arediscussed by other presenters in these proceedings (Boice,’99; Miller, ’99; Schull and Otake, ’99). The main issue withregard to the risk of mental retardation after in uteroexposure to ionizing radiation pertains to whether theeffect is stochastic or deterministic. The possibility that0.01 Sv might double the risk of mental retardation wassuggested by Otake and Schull (’84).
From the perspective of biological plausibility andthe results of animal studies, it would appear that thedata favor the viewpoint that mental retardation is adeterministic effect with a threshold of .0.2 Sv (Tables18, 20). Histological examination of the irradiated brainexposed to 0.01 Sv shows no pathological consequencesthat could account for severe mental retardation. Thatwould mean that the pattern of effects produced byionizing radiation, which accounts for mental retarda-tion when the fetus is exposed to 0.5–2 Sv does notoccur at very low exposures. Furthermore, furtherstudies by Otake and Schull showed that these authorswere able to quantitate the risk of reduced intellectafter in utero ionizing radiation exposures. They esti-mated that there was a reduction in intellect of approxi-mately 30 IQ points per Sv in their studies. Even ifthere were a linear relationship between the dose andIQ reduction, one could predict that 0.01 Gy could notaccount for a doubling of the incidence of mentalretardation, since a linear extrapolation of Otake andSchull’s data would only represent a maximum reduc-tion of 0.3 of an IQ point at 0.01 Gy. Behavioral studiesin animals were unable to demonstrate neurobehav-ioral effects at ,0.02 Gy (Jensh and Brent, ’86, ’87;Jensh et al., ’86, ’87). Although one has to be careful inextrapolating animal data to humans, the lack ofneurobehavioral effects from in utero irradiation sup-ports the other findings that indicate that mentalretardation is a deterministic effect (Table 20).
The second question pertains to the risk of cancerafter in utero radiation. There is every likelihood of an
increased risk, but the magnitude of the risk variesconsiderably, depending on which studies one findsmore reliable or plausible. These issues are discussed indetail by other presenters in this symposium (Boice,’99; Miller, ’99).
Difficulties of performing epidemiologicalstudies dealing with the risk of abortion after
exposures to environmental toxicants
There is no more treacherous area of epidemiologyresearch than that of studies involving solely theevaluation of abortion risks of environmental agents,without the simultaneous evaluation of other reproduc-tive parameters. Spontaneous abortion can result frominherited or acquired chromosomal abnormalities, inher-ited diseases, medically or environmentally producedblighted (malformed) embryos, maternal illness, lupusanticoagulant factor (Abenhaim and Lert, ’91; Beck-man and Brent, ’86; Kline and Stein, ’85; World HealthOrganization, ’70) (Table 21). Because each week ofpregnancy has a different spontaneous abortion rate(Table 22), which can vary considerably among popula-tions of pregnant women, it is extremely difficult tomatch patients in case control studies. Epidemiologistsneed very large populations to perform cohort studiesthat evaluate the risks of abortion.
In many instances, studies involving the reproduc-tive risks of low-frequency EMF of electrical currentsand video display units (60 Hz and 17–20 Hz, respec-tively) only examined the risk of abortion. One of theadvantages of reproductive biological effects is thatthere is frequently, but not always, concordance ofeffects involving more than one parameter (e.g., growth,malformations, abortion, stillbirth, prematurity). Isolatedabortion studies that do not study the totality of reproduc-tive effects are at a serious disadvantage, since spurious ornonetiological results may be misinterpreted as beingcausally related to an environmental toxicant.
It has been estimated that up to 50% of all fertilizedova in the human are lost within the first 3 weeks ofdevelopment (Hertig, ’67) (Tables 3, 22). The WorldHealth Organization (’70) estimated that 15% of all
TABLE 20. Basic science plausibility of why 1 rad(0.01 Gy) does not have the capability of doubling
the incidence of mental retardation
1. Teratogenesis is a threshold phenomenon.2. In utero exposure to ionizing radiation indicates that
there is an approximately 30-point IQ loss per Gy duringthe most sensitive period of human brain development.This finding indicates that severe mental retardationwould not occur, even if there were not a threshold, since alinear relationship to exposure would predict a 0.3 IQ lossat 0.01 Gy.
3. At 0.01 Gy, there are no observable histologic effects in thedeveloping brain that could account for severe CNSeffects.
4. Neurobehavioral evaluations of animals exposed in uterodemonstrate a threshold for behavioral effects at the samedose as for other teratologic effects (0.2 Gy).
TABLE 21. Etiology of abortion
1. Chromosomal abnormalities: inherited or environmen-tally induced preconceptionally or periconceptionally(accounts for 50–70% of abortions)
2. Luteal-phase hormonal deficiency3. Endometriosis4. Lupus anticoagulant5. Endometritis, cervicitis, amnionitis due to bacterial or
viral infections6. Uterine abnormalities: subserosal myoma, infantile
uterus, bifid uterus, IUD, etc.7. Some teratogens, especially those with cytotoxic charac-
teristics8. Maternal diabetes, alcoholism, illicit drug abuse, phenyl-
ketonuria, hemorrhagic diatheses and many other chronicand acute maternal diseases
9. Embryos and fetuses with severe congenital malforma-tions or growth retardation
200 R.L. BRENT

clinically recognizable pregnancies end in spontaneousabortion, while 50–60% of the spontaneously abortedfetuses have chromosomal abnormalities (Boue et al.,’75; Simpson, ’80). This means that, as a conservativeestimate, 1,173 clinically recognized pregnancies willresult in approximately 173 miscarriages and 30–60 ofthe infants will have congenital anomalies in theremaining 1,000 live births. The true incidence ofpregnancy loss is much higher, but undocumentedpregnancies are not included in this risk estimate.
Epidemiological investigations dealing with thecauses of spontaneous abortions must deal with formi-dable problems:
1. Most spontaneous abortions are due to chromosomalabnormalities that are unrelated to environmentalexposures during pregnancy.
2. The risk of abortion changes with each day ofpregnancy (Table 22), so that it is essential to matchcontrols, in order to eliminate the selection of twopopulations with different spontaneous abortionrates.
3. Attempts to control for the hidden incidence oftherapeutic abortions have only limited success (Ol-sen, ’84; Susser, ’83): ‘‘The existence of high rates ofinduced abortion in the population may distortcurrently employed measures of the rate of spontane-ous abortion’’ (Susser, ’83).
Basic science aspects of the reproductive effectsof ultrasound, low-frequency EMF, and
microwaves
The human and animal studies dealing with thereproductive effects of these forms of ultrasound, low-frequency EMF and microwaves are discussed by othercontributors to this symposium (Brent, ’99; Jensh and
Brent, ’99; O’Connor, ’99; Robert, ’99; Ziskin, ’99). Butthere are certain biological principles that have beenuseful in evaluating the potential reproductive risks ofthese nonionizing forms of radiation.
For example, it is well known that persistent hyper-thermia greater than 2° above basal body temperatureincreases the risk of embryopathology (Brent et al., ’91;Edwards, ’68; Edwards and Wanner, ’77). Both ultra-sound and microwaves have this potential, but expo-sures to diagnostic ultrasound examinations or levels ofradiation from cellular phones or microwave antennaedo not significantly increase the temperature of theembryo (Klug et al., ’97).
Exposures to low-frequency EMF (power lines andvideo display terminals) do not even have the potentialfor significantly raising the temperature of the embryo.Furthermore, if one examines the mechanisms involvedin the teratogenic process, there is little evidence thatthese mechanisms can be initiated with exposures of,500 mG (Table 23). In fact, a number of findingsdiminished the probability that mG exposures to low-frequency EMF have significant reproductive effects(Table 23).
Summary of the effects of intrauterine exposureto various forms of radiation
Ionizing radiation is one of the most extensivelystudied environmental agents, which accounts for theimmense amount of quantitative and qualitative infor-mation available pertaining to the reproductive risks ofthis form of radiation. Almost all the effects of intrauter-ine radiation are deterministic, which is why counsel-ors advise pregnant women that exposures of ,0.05 Gyrepresent nonmeasurable reproductive risks, except formutagenic and carcinogenic risks, which are substan-tially below the spontaneous occurrence of cancer andspontaneous mutations. The 0.05 Gy exposure is wellbelow the 0.2 Gy threshold for congenital malforma-tions, growth retardation, neurodevelopmental abnor-malities, and other reproductive effects. The majoreffects of ionizing radiation above the threshold dose
TABLE 22. Estimated outcome of 100 pregnanciesversus time from conception*
Time fromconception
Survival to term(%)
Death duringinterval
(%)
Preimplantation0–6 days 25 54.55
Postimplantation7–13 days 55 24.66
14–20 days 73 8.183–5 wk 79.5 7.566–9 wk 90 6.52
10–13 wk 92 4.4214–17 wk 96.26 1.3318–21 wk 97.56 0.8522–25 wk 98.39 0.3126–29 wk 98.69 0.3030–33 wk 98.98 0.3034–37 wk 99.26 0.34381 wk 99.32 0.68
*An estimated 50–70% of all human conceptions are lost inthe first 30 weeks of gestation (Hertig, ’67), and 78% are lostbefore term (Robert and Lowe, ’75).Data from Kline and Stein (’85).
TABLE 23. Biological plausibility of why lowexposures to 60-Hz ELF or EMF do not have significant
reproductive toxic effects
1. Electrical fields are used to fuse zygotes for embryo fusionexperiments that result in viable newborns. The expo-sures are very high; therefore, the results do not supportthe suggestion that very low exposures of electrical ormagnetic fields are embryotoxic.
2. EMF and ELF do not have specific capabilities of alteringthe genomic or chromosome structure.
3. Even when tissues are exposed to very high exposures,cell killing does not occur.
4. There is no suggestion in the medical literature of an EMFor ELF teratogenic syndrome.
5. Natural and unavoidable thermal electric fields at the cel-lular level have current densities equivalent to thoseinduced by a 60-Hz magnetic field of 6.5 G.
ELF, electrical fields; EMF, electromagnetic field.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 201

are lethality during the preimplantation–preorganoge-netic period, malformations and growth retardationduring organogenesis, and cell depletion, as well ashistogenetic abnormalities and growth retardation dur-ing fetal development. The nature of the types ofmalformations that have been described after intrauter-ine radiation have been well characterized. It should benoted that in spite of the deleterious effects of highexposures to ionizing radiation, the effects are limitedto a spectrum of identifiable malformations. Manymalformations or groups of malformations cannot beproduced by exposures to radiation.
The other forms of radiation—electromagnetic fieldsfrom power lines, household currents and video displayterminals, microwaves, and ultrasound—have also beenstudied with regard to their reproductive risks. Biologi-cal plausibility plays an even more important role intheir evaluation than with ionizing radiation. Micro-waves and ultrasound have the capacity to producehyperthermia, which is a proven reproductive toxin.Numerous animal experiments have demonstrated thatintrauterine exposures to hyperthermia from micro-waves and ultrasound can produce malformations,growth retardation, and embryonic loss. But the usualpopulation exposures to ultrasound and microwavesare below the exposures that result in hyperthermia.Furthermore, those mechanisms that are involved inreproductive toxicity, such as cytotoxicity and abnormaldifferentiation and cell migration do not occur at thepopulation exposures to these agents.
Evaluations of pregnancy loss from intrauterine expo-sures to environmental toxicants presents special prob-lems, especially if it is the only reproductive effect beingevaluated. Many studies have ignored the basic con-cepts of reproductive toxicology and the biological plau-sibility of their findings. Investigators should be cau-tious about initiating epidemiological studies dealingwith pregnancy loss without concurrently collectingother reproductive endpoints. Studies evaluating mul-tiple reproductive endpoints have markers to assistthem in determining the validity of the pregnancy lossdata.
LITERATURE CITEDAbenhaim L, Lert F. 1991. Methodological issues for the assessment of
clusters of adverse pregnancy outcomes in the work-place: The caseof video display terminal users. J Occup Med 33:1091–1096.
Auerbach C. 1971. Newsletter of Environmental Mutagen Society, p42.
Beckman DA, Brent RL. editors. 1986. Mechanism of known environ-mental teratogens: Drugs and chemicals. Philadelphia: WB Saun-ders.
Bengtsson G. 1991. Present knowledge on the effects of radiativecontamination on pregnancy outcome. Biomed Pharmacother 45:221–223.
Benova DK, Bayrakova AK, Vyglenov AK, Kusheva RP, Rupova IM,Yagova AK, Baev IA. 1985. Genetic radiation risk assessment basedon experimental mutagenesis in laboratory mammals. Genetics21:574–581.
Boice JD Jr, Miller RW. 1999. Childhood and adult cancer followingintrauterine exposure to ionizing radiation. Teratology 59:227–233.
Boue J, BoueA, Lazar P. 1975. Retrospective and prospective epidemio-logical studies of 1,500 karyotyped spontaneous abortions. Teratol-ogy 12:11–26.
Brent RL. 1960. The indirect effect of irradiation on embryonicdevelopment. II. Irradiation of the placenta. Am J Dis Child100:103–108.
Brent RL. 1967. Medicolegal aspects of teratology. J Pediatr 71:288–289.
Brent RL. 1970. Effects of radiation on the foetus, newborn and child.In: Fry RJM, Grahn D, Griem ML, editors. Late effects of radiation.London: Taylor & Francis. p 23–64.
Brent RL. 1971. The response of the 91⁄2 day-old rat embryo to vari-ations in dose rate of 150 R X-irradiation. Radiat Res 45:127–136.
Brent RL. 1972. Irradiation in pregnancy. In: Sciarra JJ, editor. Davis’gynecology and obstetrics. New York: Harper & Row. p 1–32.
Brent RL. 1976. Environmental factors: Radiation. In: Brent RL,Harris MI, editors. Prevention of embryonic, fetal, and perinataldisease. DHEW Publication No. (NIH) 76-853. Bethesda, MD: USDepartment of Health, Education, and Welfare. p 179–197.
Brent RL. 1977. Radiations and other physical agents. In: Wilson JG,Fraser FC, editors. Handbook of teratology. New York: PlenumPress. p 153–223.
Brent RL. 1980a. Radiation teratogenesis. Teratology 21:281–298.Brent RL. 1980b. X-ray, microwave, and ultrasound: The real and
unreal hazards. Pediatr Ann 9:43–47.Brent RL. 1989. The effect of embryonic and fetal exposure to x-ray,
microwaves, and ultrasound: Counseling the pregnant and non-pregnant patient about these risks. Semin Oncol 16:347–369.
Brent RL. 1992a. Reproductive and teratologic effects of electromag-netic fields (Chap. 6). In: Davis JG, Bennett WR, Brady JV, BrentRL, Gordis L, Gordon WE, Greenhouse SW, Reiter RJ, Stein GS,Suskin C, Trichopoulos D, editors. Health effects of low-frequencyelectric and magnetic fields. Washington, DC: Oak Ridge AssociatedUniversities. p 1–55.
Brent RL. 1992b. Reproductive effects. In: Davis JG, Bennett WR,Brady JV, Brent RL, Cordis L, Gordon WE, Greenhouse SW, ReiterRJ, Stein GS, Susskind C, Trichopoulos D, editors. Health effects oflow-frequency electric and magnetic fields, Executive summary.Washington, DC: Oak Ridge Associated Universities. p 10–13.
Brent RL. 1994. Biological factors related to male mediated reproduc-tive and developmental toxicity. New York: Plenum Press.
Brent RL. 1999. Reproductive and teratologic effects of low frequencyelectromagnetic fields: A review of in vivo and in vitro studies usinganimal models. Teratology 59:261–286.
Brent RL, Bolden BT. 1961. The long-term effects of low-dosageembryonic irradiation. Radiat Res 14:453–454.
Brent RL, Bolden BT. 1967. The indirect effect of irradiation onembryonic development. III. The contribution of ovarian irradiation,uterine irradiation, oviduct irradiation, and zygote irradiation offetal mortality and growth retardation in the rat. Radiat Res30:759–773.
Brent RL, Bolden BT. 1968. Indirect effect of x-irradiation on embry-onic development. V. Utilization of high doses of maternal irradia-tion on the first day of gestation. Radiat Res 36:563–570.
Brent RL, Gorson RO. 1972. Radiation exposure in pregnancy. CurrProbl Diagn Radiol 2:1–48.
Brent RL. 1972a. Protecting the public from teratogenic and muta-genic hazards. J Clin Pharmacol 12:61–70.
Brent RL, McLaughlin MM. 1960. The indirect effect of irradiation onembryonic development. I. Irradiation of the mother while shieldingthe embryonic site. Am J Dis Child 100:94–102.
Brent RL, Jensh RP, Beckman DA. 1991. Medical sonography: Repro-ductive effects and risks. Teratology 44:123–146.
Brizzee KR, Brannon RB. 1972. Cell recovery in foetal brain afterionizing radiation. Int J Radiat Biol 21:375–378.
Byrne J. 1999. Long-term genetic and reproductive effects of ionizingradiation and chemotherapeutic agents on cancer patients and theiroffspring. Teratology 59:210–215.
Byrne J, Mulvihill JJ, Connelly RC, Myers MH, Austin DF, HolmesGF, Holmes FF, Latourette HB, Meigs JW, Strong LC. 1988.Reproductive problems and birth defects in survivors of Wilm’stumor and their relatives. Med Pediatr Oncol 16:233–240.
202 R.L. BRENT

Coppenger CJ, Brown SO. 1967. The gross manifestations of continu-ous gamma irradiation on the prenatal rat. Radiat Res 31:230–242.
Cowen D, Geller LM. 1960. Long-term pathological effects of prenatalx-irradiation on the central nervous system of the rat. J Neuro-pathol Exp Neurol 19:488–527.
Dekaban AS. 1968. Abnormalities in children exposed to x-irradiationduring various stages of gestation. Tenative timetable of radiationinjury to the human fetus. J Nucl Med 9:471–477.
Edwards MJ. 1968. Congenital malformations in the rat followinginduced hyperthermia during gestation. Teratology 1:173–178.
Edwards MJ, Wanner RA. 1977. Extremes of temperature. In: WilsonJG, Fraser FC, editors. Handbook of teratology. New York: PlenumPress. p 421–444.
Furchgott E. 1963. Behavioral effects of ionizing radiations, 1955–1961. Psychol Bull 60:157–200.
Gentry J, Parkhurst E, Bulin G. 1959. An epidemiological study ofcongenital malformations in New York State. Am J Public Health49:497.
Goldstein L, Murphy DP. 1929a. Microcephalic idiocy following ra-dium therapy for uterine cancer during pregnancy. Am J ObstetGynecol 18:189–195, 281–283.
Goldstein L, Murphy DPL. 1929b. Etiology of ill health in childrenborn after maternal pelvic irradiation. II. Defective children bornafter postconceptional maternal irradiation. AJR 22:322–331.
Grahn D, Kratchman J. 1963. Variation in neonatal death rate andbirth rate in United States and possible relations to environmentalradiation, geology and attitude. Ann J Hum Genet 15:329–352.
Green DM, Fine WE, Li FP. 1982. Offspring of patients treated forunilateral Wilm’s Tumor in childhood. Cancer 49:2285–2288.
Green DM, Hall B, Zevon MA. 1989. Pregnancy outcome aftertreatment for acute lymphoblastic leukemia during childhood oradolescence. Cancer 64:2335–2339.
Green DM, Zevon MA, Hall B. 1991a. Achievement of life goals byadult survivors of modern treatment for childhood cancer. Cancer67:206–213.
Green DM, Zevon MA, Lowrie G, Seigelstein N, Hall B. 1991b.Congenital anomalies in children of patients who received chemother-apy for cancer in childhood and adolescence. N Engl J Med 325:141–146.
Haeusler MCH, Berghold A, Schoell W, Hofer P, Schoffer M. 1992. Theinfluence of the post-Chernobyl fallout on birth defects and abortionrates in Austria. Am J Obstet Gynecol 167(4, part 1):1025–1031.
Harjulehto T, Rahola T, Suomela M. 1991. Pregnancy outcome inFinland after the Chernobyl accident. Biomed Pharmacother 45:263–266.
Hertig AT. 1967. The overall problem in man. In: Benirschke K, editor.Comparative aspects of reproductive failure. Berlin: Springer-Verlag. p 11–41.
Hicks SP, D’Amato CJ. 1966. Effects of ionizing radiation on mamma-lian development. In: Wollam DHM editor. Advances in teratology.London: Logo Press. p 196–243.
Irgens LM, Lie RT, Ulstein M. 1991. Pregnancy outcome in Norwayafter Chernobyl. Biomed Pharmacother 45:233–241.
Jacobsen L, Mellemgaard L. 1988. Anomalies of the eyes in descen-dants of women irradiated with small x-ray doses during age offertility. Acta Ophthalmol (Copenh) 46:352.
Jensh RP, Brent RL. 1986. Effects of 0.6-Gy postnatal neurophysi-ologic development in the Wistar rat. Proc Soc Exp Biol Med181:611–619.
Jensh RP, Brent RL. 1987. The effect of low-level prenatal x-irradiationon postnatal development in the Wistar rat. Proc Soc Exp Med184:256–263.
Jensh RP, Brent RL. 1999. Intrauterine effects of ultrasound: Animalstudies. Teratology 59:240–251.
Jensh RP, Brent RL, Vogel WH. 1986. Studies concerning the effects oflow level prenatal x-irradiation on postnatal growth and adultbehavior in the Wistar rat. Int J Radiat Biol 50:1069–1081.
Jensh RP, Brent RL, Vogel WH. 1987. Studies of the effect of 0.4 Gyand 0.6 Gy prenatal x-irradiation on postnatal adult behavior in theWistar rat. Teratology 35:53–61.
Kinlen LJ, Acheson FD. 1968. Diagnostic irradiation, congenitalmalformations, and spontaneous abortion. Br J Radiol 41:648–654.
Kline J, Stein Z, editors. 1985. Very early pregnancy. New York: RavenPress.
Klug S, Hetscher M, Giles S, Kohlsmann S, Kramer K. 1997. The lackof effects of nonthermal RF electromagnetic fields on the develop-ment of rat embryos grown in culture. Life Sci 61:1789–1802.
Knudsen LB. 1991. Legally-induced abortions in Denmark afterChernobyl. Biomed Pharmacother 45:229–231.
Konerman G. 1969. Die Keimesentwicklung der Maus nach Ein-wirkung wontinuierlicher 60Co Gammabestrahlung wahrend derBlastogenese, der Organogenese und fetalen period. Strahlenthera-pie 137:451–466.
Kriegal H, Langendorff H. 1964. Wirkung einer fraktionierten Roen-tegenbestrahlung auf die Embryonalentwicklung der Maus. Strahl-entherapie 123:429–437.
Laskey JW, Parrish JB, Cahill DF. 1973. Some effects of lifetimeparental exposure to low levels of tritium on the F2 generation.Radiat Res 56:171–179.
Lavu S, Reddy PP, Reddi OS. 1984. Dominant lethal induction andtesticular uptake of iodine-125 in mice. Int J Radiat Biol 45:331–343.
Lyons MF, et al. 1985. Committee 4 final report—estimation of geneticrisks and increased incidence of genetic disease due to environmen-tal mutagens. International Commission for Protection againstEnvironmental Mutagens and Carcinogens: ICPEMC document114-1982-5.3. Biol Zentralbl 104:57–87.
Martin-DeLeon PA, Boice ML. 1985. Sperm aging in the male aftersexual rest: contribution to chromosome anomalies. Gamete Res12:151–163.
Miller RW. 1990. Effects of prenatal exposure to ionizing radiation.Health Phys 59:57–61.
Miller RW. 1999. Discussion: Severe mental retardation and canceramong atomic bomb survivors exposed in utero. Teratology 59:234–235.
Miller RW, Mulvihill JJ. 1976. Small head size after atomic irradia-tion. Teratology 14:355–358.
Mulvihill JJ, Byrne J. 1985. Offspring of childhood cancer survivors.Clin Oncol 4:333–343.
Murphree R, Pace H. 1960. The effects of prenatal radiation onpostnatal development in the rat. Radiat Res 12:495–504.
Murphy DP, Goldstein L. 1930. Micromelia in a child irradiated inutero. Surg Gynecol Obstet 40:79–80.
NAS/NRC. 1990. National Academy of Sciences/National ResearchCouncil. Health effects of exposure to low levels of ionizing radia-tion. Committee on the Biological Effects of Ionizing Radiations.BEIR V. Washington, DC: National Academy Press.
Neel JV. 1999. Changing perspectives on the genetic doubling dose ofionizing radiation for humans, mice and Drosophila. Teratology59:216–221.
Neel JV, Schull WJ, Awa AA, Satoh C, Kato H, Otake M, Yoshimoto Y.1990. The children of parents exposed to atomic bombs: Estimates ofthe genetic doubling dose of radiation for humans. Am J Hum Genet46:1053–1072.
Nokkentred K. 1968. Effect of radiation upon the human fetus.Copenhagen: Munksgaard.
Nomura T. 1988. X-ray and chemically induced germ-line mutationcausing phenotypical anomalies in mice. Mutat Res 198:309–320.
Nygaard R, Clausen N, Siimes MA, Marky I, Skjeldestqad FE,Kristinsson JR, Vuoristo A, Wegelius R, Moe PJ. 1991. Reproductionfollowing treatment for childhood leukemia: A population-basedprospective cohort study of fertility and offspring. Med PediatrOncol 19:459–466.
O’Connor ME. 1999. Intrauterine effects in animals exposed to radio-frequency and microwave fields. Teratology 59:287–291.
Odlind V, Ericson A. 1991. Incidence of legal abortion in Sweden afterthe Chernobyl accident. Biomed Pharmacother 45:225–228.
Okada S, Hamilton JB, Egami N. 1975. A review of thirty-year study ofHiroshima and Nagasaki atomic bomb survivors. J Radiat Res(Tokyo) II (suppl):164.
Olsen J. 1984. Calculating risk ratios for spontaneous abortions: Theproblem of induced abortions. Int J Epidemiol 13:347–350.
Otake M, Schull WJ. 1984. In utero exposure to A-bomb radiation andmental retardation. A reassessment. Br J Radiol 57:409–414.
EVALUATING RISKS FROM ENVIRONMENTAL RADIATION EXPOSURE 203

Piontkovskii IA. 1958. Certain properties of the higher nervousactivity in adult animals irradiated prenatally by ionizing radia-tions: The problem of the effect of ionizing irradiation on offspring.Bull Exp Biol Med 46:77–80.
Piontkovskii IA. 1961. Some peculiarities of higher nervous activity inadult animals subjected to radiation in utero. Bull Exp Biol Med51(pt 3):24–31.
Plummer G. 1952. Anomalies occurring in children exposed in utero tothe atomic bomb in Hiroshima. Pediatrics 10:687–692.
Polhemus D, Koch R. 1969. Leukemia and medical irradiation.Pediatrics 23:453–461.
Robert CJ, Lowe CR. 1975. Where have all the conceptuses gone?Lancet 1:488–499.
Robert E. 1999. Intrauterine effects of electromagnetic fields (lowfrequency, mid-frequency RF and microwaves): Review of epidemio-logic studies. Teratology 59:292–298.
Ronnback C. 1965. Effects of continuous irradiation during gestationand suckling period of mice. Acta Radiol Ther 3:169–176.
Rugh R. 1962. Major radiobiological concepts and ionizing radiation onthe embryo and fetus. In: Haley, Snider, editors. Response of thenervous system to ionizing radiation. New York: Academic Press.
Rugh R. 1965. Effect of ionizing radiations, including radioisotopes, onthe placenta and embryo. Birth Defects 1:64–73.
Rugh R, Wohlfromm M. 1964a. Can x-irradiation prior to sexualmaturity affect the fertility of the male mammal (mouse)? Atom-praxis 10:33–42.
Rugh W, Wohlfromm M. 1964b. X-irradiation sterilization of thepremature female mouse. Atompraxis 10:511–518.
Russell LB. 1950. X-ray induced developmental abnormalities in themouse and their use in the analysis of embryological patterns. I.External and gross visceral changes. J Exp Zool 114:545–602.
Russell LB. 1956. X-ray induced developmental abnormalities in themouse and their use in the analysis of embryonical patterns. II.Abnormalities of the vertebral column and thorax. J Exp Zool131:329–395.
Russell LB, Badgett SK, Saylors CL. 1963. Comparison of the effects ofacute, continuous and fractionated irradiation during embryonicdevelopment. In: Sobel FH, editor. Repair from genetic radiation.New York: Pergamon Press. p 333–342.
Russell LB, Major MH. 1957. Radiation-induced presumed somaticmutations in the house mouse. Genetics 42:161–1757.
Russell LB, Russell WL. 1950. The effects of radiation on the preimplan-tation stages of the mouse embryo. Anat Res 108:521.
Russell LB, Russell WL. 1954. An analysis of the changing radiationresponse of the developing mouse embryo. J Cell Comp Physiol43:103–149.
Russell LB, Saylors CL. 1963. The relative sensitivity of variousgerm-cell stages of the mouse to radiation-induced nondysfunction,chromosome losses and deficiencies. In: Sobel FH, editor. Repairfrom genetic radiation. New York: Pergamon Press. p 313–342.
Russell WL. 1977. Mutation frequencies in female mice and theestimation of genetic hazards of radiation in women. Proc Natl AcadSci USA 74:3523–3527.
Schull WJ, Otake M. 1999. Cognitive function and prenatal exposureto ionizing radiation. Teratology 59:222–226.
Segall A, MacMahon B, Hannigan M. 1964. Congenital malformationsand background radiation in northern New England. J Chron Dis17:915–932.
Sikov MR. 1992. Hazards and risks from prenatal irradiation: Empha-sis on internal radionuclide exposures. Radiat Prot Dosimetry41:265–272.
Simpson JL. 1980. Genes, chromosomes and reproductive failure.Fertil Steril 33:116–778.
Stadler J, Gowen JW. 1964. Observations on the effects of continuousirradiation over 10 generations on reproductivities of differentstrains of mice. In: Carlson WD, Gassner FX, editors. Effects ofionizing radiation on reproductive systems. New York: PergamonPress. p 111–122.
Stewart AM. 1972. Myeloid leukaemia and cot deaths. BMJ 4:423.Stewart A. 1973. The carcinogenic effects of low-level radiation: A
reappraisal of epidemiologists’ methods and observations. HealthPhys 24:223–240.
Stewart A, Kneale GW. 1970. Radiation dose effects in relation toobstetric x-rays and childhood cancers. Lancet 1:1185–1188.
Stewart A, Webb D, Giles D, et al. 1956. Malignant disease inchildhood and diagnostic irradiation in utero. Lancet 2:447.
Susser E. 1983. Spontaneous abortion and induced abortion: Anadjustment for the presence of induced abortion when estimatingthe rate of spontaneous abortion from cross-sectional studies. Am JEpidemiol 117:305–308.
Tabuchi A. 1964. Fetal disorders due to ionizing radiation. HiroshimaJ Med Sci 13:125–173.
Tabuchi A, Nakagawa S, Hirai T. 1967. Fetal hazards due to x-raydiagnosis during pregnancy. Hiroshima J Med Sci 16:49–66.
Van Buul PPW. 1984. Enhanced radiosensitivity for the induction oftranslocation in mouse stem cell spermatogonia following treatmentwith cyclophosphamide or adriamycin. Mutat Res 128:207–211.
Vilumsen A. 1970. Environmental factors in congenital malforma-tions. Copenhagen: FDAL’s Forlag.
Vorisek P. 1965. Einfluss der kontinuierlichen intrauterinen Bestrahl-ung auf die perinatale Mortalitat der Frucht. Strahlentherapie127:112–120.
Warkany J, editor. 1971. Congenital malformation notes and com-ments. Chicago: Year Book Medical Publishers.
Wesley JP. 1960. Background radiation as a cause of congenitalmalformations. Int J Radiat Biol 2:97–118.
Wood J, Keehn R, Kawamoto S. 1967a. The growth and development ofchildren exposed in utero to the atomic bombs in Hiroshima andNagasaki. Am J Public Health 57:1374–1380.
Wood JW, Johnson KG, Omori Y. 1967b. In utero exposure to theHiroshima atomic bomb. An evaluation of head size and mentalretardation: Twenty years later. Pediatrics 39:385–392.
Wood JW, Johnson KG, Omori Y, Kawamoto S, Keehn RJ. 1967c.Mental retardation in children exposed in utero to the atomic bombsin Hiroshima and Nagasaki. Am J Public Health 57:1381.
World Health Organization. 1970. Spontaneous and induced abortion.Technical Report Series Report No. 461. Technical Report Series No.461. Geneva: World Health Organization.
Yamazaki J, Wright S, Wright P. 1954. Outcome of pregnancy inwomen exposed to the atomic bomb in Nagasaki. Am J Dis Child87:448–463.
Zappert J. 1926. Uber roentgenogene fetale microcephalie. Monatss-chr Kinderheilkd 34:490–493.
Ziskin MC. 1999. Intrauterine effects of ultrasound: Human epidemi-ology. Teratology 59:252–260.
204 R.L. BRENT