university of eneva galime teigti, kad dvl-3 gali bti geras vaist taikinys gydant ligas, kurioms...
TRANSCRIPT

1
UNIVERSITY OF GENEVA FACULTY OF MEDICINE
DEPARTMENT OF NEUROSCIENCES
KAUNAS UNIVERSITY OF MEDICINE FACULTY OF PHARMACY
DEPARTMENT OF DRUG TECHNOLOGY AND SOCIAL PHARMACY
NEW POTENTIAL PHARMACEUTICAL TARGETS IN EPENDYMAL CELLS:
RESEARCH AND EVALUATION MASTER THESIS
MINDAUGAS JONIKAS FACULTY OF PHARMACY
KAUNAS UNIVERSITY OF MEDICINE
SUPERVISORS:
PROF. JOZSEF ZOLTAN KISS PROF. VITALIS BRIEDIS DEPARTMENT OF NEUROSCIENCES, DEPARTMENT OF DRUG TECHNOLOGY
FACULTY OF MEDICINE, AND SOCIAL PHARMACY,
UNIVERSITY OF GENEVA. FACULTY OF PHARMACY,
KAUNAS UNIVERSITY OF MEDICINE.
GENEVA, KAUNAS
2010

2
Table of Contents
1. The goal of study……..……………………………………………….14
2. Introduction…………………………………..…..…………………..15
2.1 Ependymal cells…………………………………………….………….……15
2.1.1 Cilia ………………………………………………………………………......…..16
2.1.2 Microvilli…………………………………………………………………...…….20
2.1.3 Adhesion…………………………………………………………...…………….20
2.1.4 Junctions…………………………………………………………………..……..21
2.1.5 Development…………………………………………………….…….…………22
2.1.6 The support of the neurogenic niche………………………….……….…………23
2.1.7 Other cells………………………………………………………..….……………25
2.2 Wnt Signaling system…………………………………………….………...26
2.2.1 Wnt biogenesis……………………………………………….……….………….28
2.2.2 Receptors in Wnt signaling………………………………………………………29
2.2.3 Wnt signaling and adult neurogenesis……………………………..……………..31
2.2.4 Wnt signalling and Alzheimer disease (AD)…………………………………….32
2.2.4 The structure of Dvl………………………………………….…………………..35
2.2.5 Dishevelled controls apical docking of basal bodies and planar cell
polarization………………………………………………………………….…….…... 37
3. Experimental part……………………………………………………40
3.1 Materials and methods……………………………………………………. 40
3.1.1 Cell culture………………………………………………………………………. 40
3.1.2 Brain slices……………………………………………….……………………….42

3
3.1.3 Analysis……………………………………….………………………………..45
3.2 Results……………………………………………………………………. 46
3.2.1 Analysis of Wnt ligands………………………………………………………...46
3.2.2 Analysis of Wnt signal transducer – Dishevelled……………………………….49
3.3 Discussion…………………………………………………………………..53
3.3.1 Ependymal cells can secrete Wnt 8b.....................................................................54
3.3.2 The role of Dishevelled as Wnt signal transducer in the cilia………………….. 56
3.3.3 Dishevelled nuclear shuttling…………………………………………………... 57
4. Conclusions..........................................................................................59
5. Bibliography:…………………………………….…………..............62

4
SANTRAUKA
NAUJI POTENCIAL�S VAIST� TAIKINIAI
EPENDIMIN�SE L�STEL�SE: TYRIMAI IR VERTINIMAS
Darbo tikslas:
Atlikti tyrimus, siekiant surasti galimus naujus vaist� taikinius ependimin�se l�stel�se.
Tyrimo objektas:
1. Suagusi� žiurki� smegen� skilvelin�s zonos l�stel�s bei ependemini� l�steli�
kult�ra.
2. Wnt signalinio kelio molekul�s.
Tyrimo metodai:
1. Specifiniai fluorescenciniai imunohistocheminiai tyrimai atliekami su smegen�
skilvelin�s zonos l�stel�mis ir ependemini� l�steli� kult�ra.
2. Rezultat� analizei naudojamas konfokalinis ir fluorescencinis mikroskopai.
Darbo uždaviniai:
1. Suformuoti model� tinkam� tirti suaugusi� žiurki� smegen� skilvelin�s zonos
l�steles.
2. Sukurti ependimini� l�steli� kult�ros model�.
3. Nustatyti Wnt signalinio kelio ligand� (Wnt 3A, Wnt 5A, Wnt 7A, Wnt 7B ir Wnt
8b) lokalizacij� suaugusi� žiurki� smegen� skilvelin�s zonos l�stel�se ir
ependimini� l�steli� kult�rose. Tam panaudoti specifinius imunohistocheminius
metodus.
4. Siekiant išsamesni� rezultat�, taip pat lokalizuoti Dishevelled baltymo izoformas
(Dvl-1, Dvl-2 ir Dvl-3) suaugusi� žiurki� smegen� skilvelin�s zonos l�stel�se ir
ependimini� l�steli� kult�rose. Tam pasiekti, bus pasitelkti specifiniai
imunohistocheminiai metodai.

5
Rezultatai:
1. Wnt 3A, Wnt 5A, Wnt 7A ir Wnt 7B antik�ni� specifin�s imunin�s reakcijos
ependimin�se l�stel�se neaptikome taikant konfokalin� ir fluorescencin�
mikroskopus.
2. Wnt 8b antik�nio specifin� imunin� reakcija aptikta apatin�je blakstien�li� dalyje.
3. Dvl-1 epedimin�se l�stel�se yra akumuliuojamas apatin�je blakstien�l�s dalyje ir
l�stel�s branduolyje. Tai nustat�me pagal specifin� imunin� antik�nio reakcij�.
4. Dvl-2 antik�nis nepasižym�jo specifine imunine reakcija ependimin�se l�stel�se.
5. Dvl-3 ependimin�se l�stel�se nustat�me membranoje ir nedidelis kiekis galimas
citoplazmoje.
1. Lentel�. Specifin� lokalizacija Wnt signalin�s sistemos ligand� ir signalo pardav�j�.
Wnt3A
Wnt5A
Wnt7A
Wnt7B
Wnt8B
Dvl-1
Dvl-2
Dvl-3
Ependemini� l�steli� kult�ra: • Blakstien�les
• Citoplazma
• Branduolys
• Membrana
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+
+++
-
+/-
+/-
+/-
+/-
-
++
+/-
++
Skilvelin�s zonos l�stel�s: • Blakstien�les
• Citoplazma
• Branduolys
• Membrana
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+/-
+++
-
-
-
-
-
-
++
-
+++
- - n�ra signalo; +/- - labai silpnas ar nespecifiškas signalas; + - silpnas signalas; ++ - geras signalas; +++ - stiprus signalas.

6
Išvados:
1. Suaugusi� žiurki� smegen� skilvelin�s zonos l�steles galima tirti smegen�
segmentuose, kurie yra gaunami „Vibratomo“ ar „Kryostato“ pagalba. Taikant šiuos
metodus, l�steli� pozicija yra išsaugojama ir strukt�ra nepažeidžiama; tokios
l�stel�s yra tinkamos imunohistocheminiams tyrimams. Optimaliausias segment�
storis yra 60-20µm.
2. Ependimini� l�steli� kult�ra gali b�ti sukuriama iš k� tik gimusi� žiurki� jaunikli�
smegen�. Ependimin�s l�stel�s yra išpjaunamos iš galvos smegen� tre�iojo
skilvelio ir kultivuojamos Lamininu padengtose l�kštel�se. Kultivavimas vykdomas
7-10 dien�. Atlikus l�steli� fiksacij�, galimi imunohistocheminiai tyrimai.
3. Wnt 3A, Wnt 5A, Wnt 7A ir Wnt 7B neaptikome ependimin�se l�stel�se. Pagal
šiuos rezultatus galime teigti, kad šie ligandai negali b�ti vaist� taikiniai, nes
tikriausiai n�ra ekspresuojami ependimin�se l�stel�se. Yra tikslinga šiuos
duomenis patvirtinti in situ hibridizacijos metodu.
4. Wnt 8b yra susikaup�s apatin�je blakstien�li� dalyje, ši lokalizacija yra visiškai
nauja. Ši blakstien�li� dalis pasižymi sekrecin�mis funkcijomis, tod�l manome, kad
Wnt 8b gali b�ti sekretuojamas � galvos smegen� skyst�. Tai gali b�ti labai svarbu
prenataliniam smegen� žiev�s vystimuisi ir regeneracijai po smegen� pažeidimo.
Wnt 8b galimas labai svarbus naujas vaist� taikinys.
5. Dvl -1 lokalizuota ependimini� l�steli� apatin�je blakstien�li� dalyje ir branduolyje.
Ši informacija literat�roje dar nemin�ta. Pagal Dvl-1 viet�, galima spr�sti, kad jis
dalyvauja tipiniame Wnt signaliniame kelyje. Dvl-1 gali b�ti svarbus vaist� taikinys
gydant ependimomas, nes yra žinoma, kad kitose v�žio formose tipinis Wnt
signalinis kelias yra pernelyg suaktyvintas. Ependimomose Dvl-1 gali b�ti
inhibuojamas vaistais, siekiant subalansuoti tipin� Wnt signalin� keli�.
6. Dvl-2 neaptikome ependimin�se l�stel�se, taip pat literat�roje n�ra duomen�, kad
šis baltymas yra ekspresuojamas šiose l�stel�se. Pagal šiuos duomenis galime teigti,
kad Dvl-2 negali b�ti vaist� taikinys. Yra tikslinga šiuos duomenis patvirtinti in situ
hibridizacijos metodu.

7
7. Ependimin�se l�stel�se Dvl-3 yra akumuliuojamas membranoje. Ši akumuliacijos
vieta yra visiškai nauja. Dvl-3 lokalizacija membranoje rodo jo dalyvavim� ne-
tipiniame Wnt signaliniame kelyje, kuris paveikia l�stel�s poliarizacija. Pagal šiuos
duomenis galime teigti, kad Dvl-3 gali b�ti geras vaist� taikinys gydant ligas,
kurioms pasireiškia sutrikusi blakstien�li� veikla. Kaip pavyzdys gal�t� b�ti,
sutrikusi neuron� migracija lygiagre�iai šoninio skilvelio. Tod�l, Dvl-3 gali b�ti
geras taikinys koreguojantis degeneracinius procesus, nes didesn� ekspresija Dvl-3
pagreitint� neuroblast� ir astrocit� migracij� link pažeistos vietos, taip b�t�
sustabdomi tolimesni pažeidimai.

8
SUMMARY
NEW POTENTIAL PHARMACEUTICAL TARGETS IN
EPENDYMAL CELLS: RESEARCH AND EVALUATION
The aim:
Conduct an investigation study in order to find potential new drugs targets in ependymal
cells.
Objects:
1. Ventricular zone cells of adult rat animal and ependymal cell culture.
2. WNT signaling pathway molecules.
Methods:
1. Specific fluorescent immunohistochemistry studies performed with brain
ventricular zone cells and ependymal cell culture.
2. Analysis of the results was done with confocal and fluorescence microscopes.
The objectives:
1. Develop an effective model to investigate the ventricular zone cells of adult rat
brain.
2. Establish the model of ependymal cell culture.
3. To determine the localization of the WNT signaling pathway ligands (WNT 3A,
WNT 5A, WNT 7B, WNT 7A, and WNT 8b) in ventricular zone cells of adult rat
brain and in ependymal cell culture. In order to accomplish this, use specific
immunohistochemical methods.
4. With a purpose to achieve more comprehensive result, localize Dishevelled protein
isoforms (Dvl-1, Dvl-2 and Dvl-3) in ventricular zone cells of adult rat brain and in
ependymal cell culture. In order to do this, use specific immunohistochemical
methods.
9
Results:
1. Appling confocal and fluorescence microscope specific immune response was not
observed of WNT 3A, WNT 5A, WNT 7A and WNT 7B antibodies in ependymal
cells.
2. Specific imunoreactivity of Wnt 8b was detected in the basal part of the cilia.
3. Dvl-1 is accumulated in the basal part of cilia and in nucleus of the ependymal
cells.
4. Dvl-2 antibody did not exhibit specific immune response in the ependymal cells.
5. Dvl-3 was detected on the membrane and low amount in the cytoplasm of
ependymal cells.
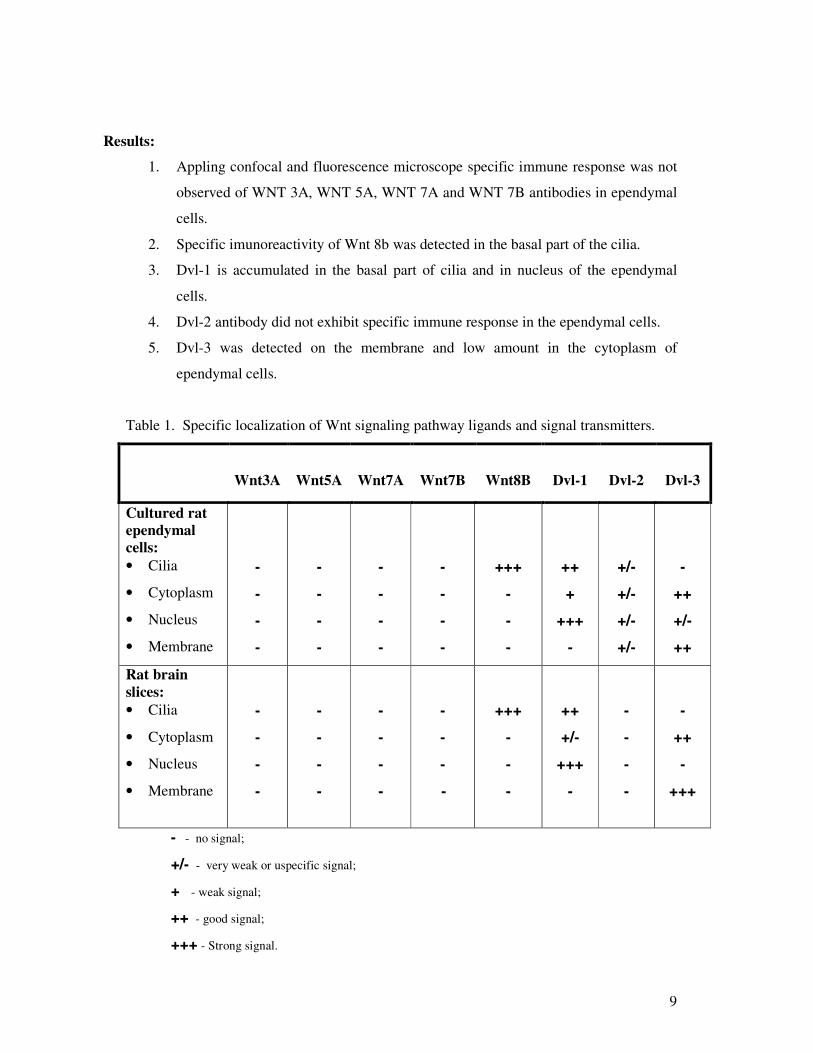
Table 1. Specific localization of Wnt signaling pathway ligands and signal transmitters.
Wnt3A
Wnt5A
Wnt7A
Wnt7B
Wnt8B
Dvl-1
Dvl-2
Dvl-3
Cultured rat ependymal cells: • Cilia
• Cytoplasm
• Nucleus
• Membrane
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+
+++
-
+/-
+/-
+/-
+/-
-
++
+/-
++
Rat brain slices: • Cilia
• Cytoplasm
• Nucleus
• Membrane
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+/-
+++
-
-
-
-
-
-
++
-
+++
- - no signal;
+/- - very weak or uspecific signal;
+ - weak signal;
++ - good signal;
+++ - Strong signal.

56
Conclusion:
1. Ventricular zone brain cells of adult rat brain can be studied in a sections,
obtained by "Vibratome" and "Cryostat”. Using these techniques, the position
and structure of the cells is maintained; these cells are suitable for
immunohistochemical experiments. Optimal thickness of sections is 60-20�m.
2. The culture of ependymal cells can be created from new-born baby rat brain.
Ependymal cells are dissected out of the third ventricle of the brain and cultured
in Laminin coated dishes. Cultivation is carried out for 7-10 days.
Immunohistochemical experiments can be done after cell fixation.
3. We did not detect WNT 3A, WNT 5A, WNT 7A and WNT 7B in the ependymal
cells. According to these results, we suggest that these ligands can not be targets
for the drugs, because it might be that there is no expression of these proteins in
ependymal cells. It is advisable to confirm these results with situ hybridization
method.
4. WNT 8b is accumulated in the basal part of the cilia, this is brand new location.
It is known, that this part of the cilia possesses secretion features that is why we
believe that WNT 8b may be secreted from ependymal cells to the cerebro spinal
fluid. This may be very important to prenatal cortical development and
regeneration after brain damage. WNT 8b may be an important new drug target.
5. Dvl-1 localization in the nucleus and basal part of cilia of ependymal cells is not
reported in the literature. According to Dvl-1 accumulation place, we can
suggest that it participates in canonical WNT signaling. Dvl-1 may be an
important drug target for treating ependymomas, because in other forms of
cancer, it is known that canonical WNT signaling pathway is over activated. In
ependymomas Dvl-1 may be drug inhibited in order to restore balance of
canonical Wnt signaling.
6. Dvl-2 was not detected in the ependymal cells, as well there is no evidence in
literature that this protein is expressed these cells. According to this data, we
strongly suggest that Dvl-2 can act as a drug target. It is advisable to confirm
these results with situ hybridization method.
7. Dvl-3 is accumulated on the membrane of the ependymal cells. This localization
is a completely new. Dvl-3 membrane localization suggests its involvement in

11
non-canonical WNT signaling, which affects planar cell polarization. According
to these data, Dvl-3 may be a good drug target for treating the diseases, which
display the impaired cell migration or ciliar activity. An example could be
impaired neuronal migration in parallel with the lateral ventricle. Thus, Dvl-3
may be a good target for adjusting the degenerative processes, because increased
expression of Dvl-3 might accelerate migration of neuroblasts and astrocyte
toward damaged areas, thereby stopping further damage.

12
LIST OF ABBREVATION:
• AChR - acetylcholine receptor.
• AD - Alzheimer disease.
• AMPA - -amino-3-hydroxyl-5-methyl-4-isoxazole-propionate.
• Ang-1 – angiopoietin.
• APC - adenomatous polyposis coli
• APP - amyloid precursor protein.
• AQP – Aquaporins.
• BBS4 - Bardet-Biedl syndrome 4 protein.
• BSA – bovine serum albumin
• CamK2 - calcium-calmodulin-dependent kinase 2
• CSF - cerebro spinal fluid
• CVO - circumventricular organs • CRD - cysteine-rich domain
• COMT - catechol O-methyltransferase.
• Dvl – Dishevelled • Evi - Evenness interrupted
• FGF - Fibroblast growth factors.
• Fz – Frizzleds
• GEF - guanine nucleotide exchange factor • GLUT - Glucose transporters.
• GSK3 - glycogen synthase kinase 3
• GTPase - family of hydrolase enzymes,which bind and hydrolyze guanosine
triphosphate.
• IGF - insulin-like growth factor
• IDA - inner dynein arms
• JNK - Jun kinase
• LEF/TCF - lymphoid enhancer factor/T cell factor

13
• LRP - lipoprotein receptor related protein
• MIF - migration inhibitory factor
• NCAM - neural cell adhesion molecule.
• NES - nuclear export sequence
• NET - norepinephrine transporters.
• NEuROD1 - pro-neurogenic transcription factor
• Nkd - Naked Cuticle.
• NMDA - N-methyl-D-aspartic acid.
• NSC - Neural stem cells.
• ODA - outer dynein arms • PAF – paraformaldehyde
• PBS - Phosphate buffered saline
• PCP – planar cell polarity.
• PKC - protein kinase C
• RPGR - retinitis pigmentosa GTPase regulator
• sFRP - Secreted frizzled-related protein
• SGZ - subgranular zone
• Srt – Sprinter
• SVZ - Subventricular zone.
• TRPV4 - Transient receptor potential cation channel subfamily V member 4.
• T2R - Taste Receptor 2.
• VEGF - Vascular endothelial growth factor.
• WG – Wingless
8.

14
1. The goal of study
The goal of my study is to investigate Wnt signaling in the ventricular zone and to
localize Wnt signaling elements (ligands and transducers) using immonohistochemistry.
WNT ligands are secreted proteins that regulate cell fate decisions, cell polarity, cell
migration, axonal morphology, and synaptic differentiation. In mammals, complexity
and specificity of Wnt signaling are in part achieved through 19 Wnt ligands. Several
studies already reported that Wnt genes continue to be expressed in the adult brain.
However, there is no immunoreactivy studies done concerning ependymal cells and Wnt
signaling elements. Therefore, I investigated the localization of Wnt elements
immunoreactivity with respect to the cilia and other structures of ependymal cells, in
order to find new drugs targets.
1. I had chosen to explore these ligands: Wnt 3A, Wnt 5A, Wnt 7A, Wnt 7B
and Wnt 8b.
2. In order to achieve comprehensive results, I had also investigated wnt
signal transmitter – Dishevelled.
As investigation object I used in vitro model of subventricular zone-derived
ependymal cells and brain section in vivo.

15
2. Introduction 2.1 Ependymal cells The ependyma is an
uninterrupted, single-cell
epithelium layer, composed of
neuroglial cells, mainly ciliated
ependymal cells. The character of
the epithelial lining was first
documented by Purkinje (1836),
and investigators subsequently
recognized that the ependyma is
heterogeneously composed, in
particular that some cells had basal
processes that extended into the
subjacent neuropil (Agduhr,
1932; Wislocki, 1932). In 1954,
Horstmann first applied the
descriptive term tanycyte to such
elongated ependymal cells. The
ependyma layer covers the entire
ventricular system of brains and
the central canal of the spinal
cord. The ependymal layer in the
adult is relatively uniform, but there are some specialized places: the area postrema at
the caudal end of the floor of the fourth ventricle and subcommissural organ at the
transitional zone between the roof of the third ventricle and the cerebral aqueduct. Those
places distinguish by a lack of the cilia and of blood-brain barrier. The ependyma at the
floor of the third ventricle becomes modified in early fetal life and also lacks blood-
brain barrier.
Figure 1. Features of normal ependymal. A. Transmission electron micrograph showing mature ependymal cells of the lateral ventricle. The top right inset shows a supraependymal axon. Bar 0.25 µm. B. Scanning electron micrograph of the surface of the caudate nucleus of an adult human showing the dense packing of cilia clusters. (Bigio et al., 2009)

16
The ependyma principally consist of ependymal cells, which develop from
neuroectoderm. Ependymal cells are neuroglial cells, morphology can be described as
cuboidal to columnar shape and a fairly round nucleus with fine stippled chromatin
pattern and inconspicuous nucleolus. The surface is covered by microvilli and most of
the cells have a central cluster of long motile cilia.
Mature mammalian ependymal cells possess the structural and enzymatic
characteristics necessary for scavenging and detoxifying a wide variety of substances in
the cerebro spinal fluid (CSF) and it is forming a metabolic barrier at the brain–CSF
interface. The presence of motile cilia, microvilli, and zonula adherens junctions at the
apical surfaces is important for these roles. The gap junctions might be used to
coordinating the cells activity.
2.1.1 Cilia
History. The field of ciliary biology is an active area of study with a rich history.
It might be, that cilia were observed first time and their motile function assessed by
Antoni Va Leeuwenhoek in 1674-75 (Dobell, 1932), but these organelles were named
by Otto Friedrich Müller in 1786 (Muller, 1786). In the second half of the nineteenth
century the non-motile cilia were observed (Kowalevsky, 1867; Langerhans, 1876;
Zimmermann, 1898). Zimmermann was the first scientist to observe these organelles in
mammalian cells, including those of humans. He named those organelles – central
flagella (“centralgeissel”) and hypothesized that they have a sensory function. However,
both Zimmermann’s name for these organelles and his proposed function for them were
soon forgotten. Sorokin in 1968 renamed these organelles “primary cilia” (Sorokin,
1968). This special class of non-motile cilia, because of their evident sensory functions,
was investigated deeper than motile cilia. Currently, however, a new idea in the field of
cilia is emerging – that all cilia have sensory functions (Christensen et al., 2007).
Evidence supporting a sensory role for motile cilia has been accumulating in the
literature for a very long time.
Structure. A striking feature of the ependymal cells is the apical cluster of the
motile cilia, which projects into the CSF. Cilia are microtubule-based cell organelles

17
extending from a basal body, a
centriole, at the apical cell
surface, containing 9+2
axonemes surrounded by a
specialized ciliary membrane.
This microtubule scaffold,
(also known axoneme), is
enveloped by an extension of
the plasma membrane. The
structure is retained in place by
a number of other associated
proteins, like the radial spoke
proteins, which help to
connect the peripheral
doublet microtubules with the
central pair. Major structures that attach to the microtubules, the outer and inner dynein
arms (ODAs and IDAs), the radial spokes, the central-pair projections, and so forth are
defined protein complexes. The ODAs and IDAs are force-producing molecular motors
that cause the doublet microtubules to slide with respect to one another. The doublet
sliding is asynchronous with the progression of activity around the axoneme, yielding a
helical beat. Maximum beat frequencies range up to approximately 100 Hz, although
most reports of mammalian ciliary beat frequency are much lower, perhaps normally
10–20 Hz.
Below plasma membrane, the axoneme remains anchored to the basal body, which
is derived from the mother centriole. The basal body is a template on which the
axoneme is built by intraflagellar transport, an intracellular cargo delivery system. The
building starts from bringing preassembled axonemal components from cytoplasm to the
tip of the axoneme. Ependymal cells have an additional kind of cilia called primary cilia.
These cilia are relatively short in size and lack the central pair of singlet microtubules as
well as dynein arms (hence they are immotile). Primary cilia appear to be associated
almost exclusively with sensory functions.
Figure 2. Basic ciliary structure. Schematic representation of a cilium and cross-section of a basal body composed of microtubule triplets and a ‘‘9+2’’ and a ‘‘9+0’’ axoneme showing the position of dynein arms and radial spokes needed for force generation and coordination. Along the outer microtubule doublets of the axoneme, molecular motors transport IFT particles. (Rodriguez et al., 2009)

18
Dynamic analysis of living ependyma shows that the cilia are beating in a
coordinated manner, trending to sweep foreign particles in the same direction as bulk
CSF flow (Yamadori and Nara, 1979). The coordination of cilia beating might be
accomplishment by communication through gap junctions or through innervations. The
CSF is also stirred at the ependymal cells surface to facilitate metabolic interactions
between ependyma and CSF content (Roth et al., 1985).
The cillary necklace. Specific proteins are localized to or concentrated in the
ciliary membrane, as opposed to the rest of the cell membrane. There are some
hypotheses that there is a selective barrier at the cilium entrance. Mode of operation of
the barrier is still uncertain. This specialized barrier region is found on all 9+2 and 9+0
mammalian and invertebrate cilia that have been studied by freeze fracture electron
microscopy (Gilula and Satir, 1972). Transport proteins have been localized to the
necklace region near the repeating intersection of particles and the membrane (Gilula
and Satir, 1972), which may imply that the particle rich regions are assembly sites for
transport of membrane and axonemal cargos.
The membrane in the necklace region of airway cilia has a different composition in
terms of lectin binding, anionic charge and free-cholesterol distribution compared to the
rest of the ciliary and cell membrane (Tuomanen, 1990). A putative guanine nucleotide
exchange factor (GEF), retinitis pigmentosa GTPase regulator (RPGR) and its
interacting protein are localized to a necklace of the connecting cilium of the
photoreceptor (Hong, 2001). RPGR isoforms are also found in the necklace region of
motile cilia of the trachea (Hong, 2003).
The importance of the necklace in tracheal cilia is reemphasizing by its disruption
and disappearance upon infection after attachment of these bacteria to the cilium and
prior to cell death. Moreover, when cilia are shed, the point of breakage and membrane
resealing occurs just above the ciliary necklace and the necklace persists.
Membrane. 9+2 and 9+0 cilia axonemes are surrounded by a ciliary membrane,
which extends from the cell membrane but is selectively different from the cell
membrane in overall composition. Surprisingly, was known very little about the ciliary
membrane until quite recently. Now, through cilia fractionation and proteomics, the
composition of the membrane proteins of the cilium is emerging.

19
Cilia response pathways for unicellular organisms are necessary for survival,
motion, control of the 9+2 motile cilium. The activity of this pathway depends on
specific receptors and channel proteins, like: cyclic nucleotide receptor, Ca2 + channels
and receptors involved in growth control pathways. All those above motioned structures
are localized to the ciliary membrane (Satir and Guerra, 2003). This suggests that all
cilia have sensory function and this hypothesis will be supported below, by discussing
studies in mammals.
Sensory reception. Motile cilia of the mammalian respiratory epithelium have
been reported to show mechanosensitivity and chemosensitivity. As mucus viscosity
around the cilia increases, ciliary mechanics are adjusted so as to maintain a ciliary beat
frequency, although it is reduced, but still sufficient to maintain transport of mucus to
the larynx, where it will be swallowed or expectorated (Johnson et al., 1991). Cytosolic
Ca2+ levels play an important role in this process of autoregulation. Increased Ca2+
concentration in cytoplasm are associated with changes in ciliary beat frequency
(Salathe, 2006).
Sanderson and Dirksen observed that mechanical stimulation of cilia, of cultured
rabbit tracheal cells with fluid movement, induced a transient increase in ciliary beat
frequency, it was dependent on the presence of Ca2+ in the extracellular medium and was
inhibited by a Ca2+-channel blocker (Sanderson and Dirksen, 1986). Lorenzo and
colleagues localized the TRPV4 cation channel in the cilia of respiratory epithelial cells
and showed that it was lost from the respiratory cilia of TRPV4 knockout mice (Lorenzo
et al., 2008). A TRPV4 agonist induced Ca2+ influx and an increase in ciliary beat
frequency in the tracheal epithelial cells from Trpv4+/+ mice but not Trpv4–/– mice. I
have to mention, that wild-type and Trpv4–/– cells were able to autoregulate ciliary beat
frequency in response to a viscous load, but just wild type cells were able to show
normal intracellular Ca2+ oscillations.
Shah and colleagues recently reported a chemoreception when they localized
different members of the bitter taste receptor family to motile cilia of airway epithelial
cells (Shah et al., 2009). There are bitter compounds, which are known to be acting
through T2R receptor and activate a signaling pathway, which induces the G protein -
gustducin (localized to the cilia), phospholipase C2 and a rise in cytosolic Ca2+. Shah
and colleagues applied those compounds to the ciliated cells. The results were induced a

20
transient, dose-dependent rise in intracellular Ca2+ in ciliated cells as well as an increase
in ciliary beat frequency.
There is evidence that than pathogens bind to respiratory cilia, the signaling
pathways can be activated: binding of Mycoplasma hyopneumoniae to the ciliary
membrane was shown to activate a G-protein-coupled receptor, which activates a
phospholipase C pathway that results in a rise in intracellular Ca2+ (Park et al., 2002).
Polycystins are now commonly known to be present in ciliary membranes of 9+0
primary cilia (Geng, 2006).It might be that similar types of channels and receptors are
present in the membranes of all mammalian motile cilia and that they play a role in
epithelial homeostasis.
The resemblance of sensory function among nearly all eukaryotic cilia (both
primary cilia and motile cilia) suggests that the original protocilium from which both
types of organelles evolved was a sensory organelle. Cilia might have evolved from a
simple „sensory membrane patch“(Jekely and Arendt, 2006; Satir et al., 2008) which
during evolutionary time extended to antenna-like structure, arising from the cell surface
and only later acquired motile function. Obviously, both sensory reception and motility
provided selective advantages for the early eukaryotic cell.
2.1.2 Microvilli
The apical surface of ependymal cells is covered by microvilli. Microvilli are a
microstructure covered with a plasma membrane, which encloses cytoplasm and
microfilaments. Although microvilli are cellular extensions, there are no or little cellular
organelles in them. Microvilli of ependymal cells are covered with glycocalyx coating.
The studies revealed presence of sialic acid, poly-N-lactosamine and D-galactose on the
ependymal microvilli (Acarin et al. 1994; Adam et al. 1993). Those enzymes along
glycoproteins are important components of chemical recognition and information
transfer mechanisms on the cell surface (Paulson, 1989). Thus the surface of ependymal
cells is adapted to interact chemically with CSF.
2.1.3 Adhesion

21
Cells in the ependyma layer are bounded to their neighbour at the apical surface by
zonula adherens type junctions (Brightman and Reese, 1969). There are tight junctions
only in between of ependymal cells covering specialized circumventricular organs
(CVO), including the choroid plexus, because there capillaries lack tight junctions.
Adherens junctions consist of cadherins, calcium-dependent transmembrane
adhesion molecules. The cadherin substances family includes cadherins,
protocadherins, desmogleins and desmocollins. On the extracellular surface cadherins
bind each other homotypically, while the intracellular domains bind p120-catenin and -
catenin (Redies et al. 1996). Numb and Numbl are required for maintenance of cadherin
based adhesion and polarity of radial glia and ependyma. In addition to apical adhesive
molecules human and mice ependymal cells reported to be expressing neural cell
adhesion molecule (NCAM, also called CD56), homophilic binding glycoprotein
(Figarella-Branger D. 1995)
Ependymal cells have a basal lamina between them (Bruni JE. 1998). The basal
lamina is a layer of extracellular matrix approximately, 40 -50 nanometers, on which
ependymal cells sticks. This layer is secreted by the ependymal cells. The basal lamina
consists of a complex of substances: laminin, utrophin, alpha-dystrobrevin and beta-
dystroglycan. This layer is anchoring ependymal cells to the ventricle wall.
2.1.4 Junctions
The membrane of ependymal cells exhibit several important classes of molecular
channels: gap junctions and aquaporins. Gap junctions forming proteins contribute to:
ion homeostasis, volume control, transferring electric current, intracellular, mechanical
sense and supporting adherent connections between neighbouring cells. Occasionally
gap junctions can be formed between ependymal cells and adjacent astrocytes. Gap
junctions are formed by hemichannels (connexons), which consist of an oligomer of six
proteins (connenxins). A complete gap junction channel is formed by two hemichannels
in mirror symmetry. (For review see Prochnow and Dermietzel, 2008). The studies of
gap junction’s proteins revealed conexins 26 and 43 in the ependymal cells. There is
data showing that ependymal cells express connexin 32 (for review see Bigio, 1995).
Colocalization of connexin 43 and basic fibroblast growth factor has been postulated to

22
play an important role in the regulation in the regulation of gap junction
communications (Yamamoto et al., 1991).
Aquaporins (AQP) are molecular channels. They are formed by two tandem
repeats of three membrane-spanning -helices, with cytoplasmic carboxyl- and
aminoterminals. Two connecting loops, each containing an Asn-Pro-Ala motif, are
believed to determine water selectivity (Murata et al., 2000). The expression and
distribution of aquaporins are organized to control water movements in the brain
(Badaut, 2002). Aquaporin 4 is expressed in the ependymal cells at the basolateral
aspects of ependymal cells, as well as in the end feet of most astrocytes (Li, 2009).
AQP4 might have structural functions beyond its role as a water channel, because AQP4
knockout mice develop hydrocephalus secondary decrease in the expression of Cx43
and loss of lateral junctions leading to disruption of the ependymal lining (for review see
Bigio, 1995). AQP1 and AQP2 immunoreactivities have been documented in human
ependymal cells (Mobasheri et al., 2004, Mobasheri et al. 2005).
2.1.5 Development
During early neurodevelopment, the embryonic ventricles are lined by a
germinal epithelium. This embryonic neuroepithelium has planar polarity that drives
morphogenetic movements essential for neural tube closure (Colas and Schoenwolf,
2001; Wallingford, 2006). Radial glial cells, in this epithelium, contain both spatial and
temporal patterning that determines cell fate and cell position in the developing brain
(Hebert and Fishell, 2008). A subpopulation of radial glia transform into ependymal
cells (Spassky et al., 2005). The majority of the ependymal cells undergo their final
nuclear DNA synthesis between E14 and E16 (Spassky et al, 2005). The ciliogenesis of
ependymal cells begins around birth and progresses in a direction from caudal to rostral
and from ventral to dorsal along the lateral wall of the lateral ventricles of the rodent
brain (Sarnat, 1992; Bruni, 1998). Spassky with others showed that ependymal cilia
appeared between P0 and P4 (Spassky et al, 2005).
The ependymal do not divide in the adult. The postmitotic characteristic of
mature ependymal cells is confirmed by studies showing that multiciliated epithelial
cells with numerous basal bodies in the apical cytoplasm are postmitotic (Lange et al.,

23
2000) and the ependymal layer in the mammalian brain does not regenerate when it is
injured (Sarnat, 1995).
2.1.6 The support of the neurogenic niche
The close contact between
ciliated ependymal cells and
pluripotent cells has led to
investigation of ependymal cells as a
modulator of stem cell populations.
The ependymal cells maintain basal
processes in regions of the frontal
horns where SVZ cells are most
abundant (Rodriguez et al., 2003).
Recently, Mirzadeh with other
observed that ependymal cells form a
remarkable pinwheel organization
specific to regions of adult
neurogenesis. The pinwheel’s center
contains the apical endings of the
neural stem cells (NSCs, B1 cells) with
a direct contact to the ventricle and a
long basal process ending on blood
vessels. The peripheral part of the pinwheel consists of two types of ependymal cells:
multiciliated (E1) and a type (E2) characterized by only two cilia and extraordinarily
complex basal bodies (Mirzadeh et al. 2008). These results suggest that adult NSCs
retain fundamental epithelial properties, including apical and basal subdivision.
There are some hypotheses that ependymal cells maintain the SVZ through
production of specific extracellular matrix and adhesion molecules (Hauwel et al., 2005)
or through release of other modulators (Sarnat et al., 1992). Ependymal cells are
producing growth factors. This might contribute to the trophic function of ependymal
cells. Fibroblast growth factors (FGF) are one the most important growth factors in brain
development (Iwata et al., 2009). It has been several times described that mature
Figure 3. Three-dimensional model of the adult SVZ
neurogenic niche illustrating B1 cells (blue; stem cells),
C cells (green; supporting cells), and A cells (red). B1
cells have a long basal process that terminates on blood
vessels (orange) and an apical ending at the ventricle
surface. Note the pinwheel organization composed of
ependymal cells (light and dark brown) encircling B1
apical surfaces. (Mirzadeh et al., 2008).

24
ependymal cells of rodents possess of FGF2 (also known as FGF) (Cuevas et al., 2000;
Fuxe et al., 1996). Some studies reported presence of mRNA for FGF2 in ependyma
(Frautschy et al., 1991), while others observed that most ependymal cells synthesize
FGFreceptor1, which allows resorption and concentration of FGF2 (Gonzalez et al.,
1995). Ependymal cells are also reported to produce FGF1 (acidic FGF) in normal and
pathological conditions (Li et al., 1998; Oomura et al., 1992; Ye et al., 2002).
Hayamizu with colleagues observed an increment of FGF2 following ischemia, this
suggest it has a role in trophic support of adjacent cells (Hayamizu et al., 2001). The
calcium binding protein S100B, which has gliotrophic and neurotrophic properties, is
well known to be present in ependymal cells, particularly at certain times in
development (Sarnat, 1992; Sarnat, 1998; Steiner et al., 2007; Vives et al., 2003)
Vascular endothelial growth factor (VEGF) is expressed by human ependyma
between 22- and 40-weeks gestations (Arai et al., 1998). There are some suggestions
that it has autocrine and paracrine functions. VEGF appears to help for the ependymal
cells to maintain their stuture, because inhibition of VEGF in mice leads to
disappearance of microvilli (Maharaj et al. 2008). VEGF is upregulated in the rat
ependyma following ischemia (Wang et al., 2008). The proteins: Ang-1, Ang-1, Tie-2,
and Flt-1, which are typically associated with endothelial growth, are present in rat
ependymal cells. Several studies have proposed that these proteins might have an
autocrine effect (Horton et al., 2009; Nourhaghighi et al., 2003; Tonchev et al., 2007).
The calcium binding protein S100B, which funtions as gliotrophic and neurotrophic
substance, is already known to be in ependymal cells, especially at particular time points
in development (Sarnat et al., 1992; Steiner et al., 2007). A variety of other growth
factors have been demonstrated in ependymal cells, but the precise functions remain to
be determined.
There are emerging evidences suggesting that ependymal cells have an impact to
the adjacent SVZ cells populations’ through metabolic regulation. Glucose uptake by
ependymal cells from CSF can occur via glucose carriers. The ependymal cells are
reported to possess GLUT1, GLUT2, GLUT3, and GLUT4 (Kobayashi et al.,1996;
Silva-Alvarez et al., 2005; Yu et al 1995). Glucokinase which is responsive to insulin
and insulin-like growth factor (IGF-1) is present in ependymal cells. Normally,
glucokinase acts like a glucose sensor, in other cells type. Glucose might be converted to

25
glycogen, later it can be mobilized by noradrenalin and serotonin, and this means that
ependymal cells might maintain glycogen as a regulated energy store (Prothmann et al.,
2001; Verleysdonk et al., 2005).
2.1.7 Other cells
As described above, ependymal cells are very close to SVZ cells and astrocytes,
but there are several additional populations worth to be mentioned. There are two
neuron systems associated with the ependymal layer. Directly applied to the ventricular
surface of ependymal cells is the supraependymal plexus of serotonergic axons (Brusco,
1998), which arise from the raphe nucleus. Immunohistochemical studies revealed
axons, positive for tyrosine hydroxylase and they are running along basal surface of
ciliated ependyma of lateral ventricles. Presumably these axons contain dopamine and
norepinephrine (Michaloudi, 1996). There are known that ependymal cells express D1
and D2 receptor subtypes, norepinephrine transporters (NETs). Monaamine oxidase B
(MAOB) and catechol O-methyltransferase (COMT) were detected in the ependymal
cells (for review see Bigio, 1995). These innervations together might be helping to
coordinate beating of cilia (Nguyen et al. 2001) or regulate metabolism (Verleysdonk et
al., 2005). In addition, there are widespread system of CSF-contacting neurons that have
extended dendritic processes between ependymal cells to contact the CSF. This
primitive system serves a non-synaptic diffuse signal transmission function (Vigh et al.
2004).
The migratory phagocytic cells (supraependymal macrophages) are well defined.
They reside on the ventricular surface of ependymal cells. Migratory phagocytic cells
are descried as scavenger cells, but they might participate in immunological response
and iron regulation in the ventricular system or the brain as a whole (Ling et al. 1998).
Interactions with ependymal cells are not well understood, but it is known that
ependymal cells store factors (migration inhibitory factor (MIF), macrophage inhibitory
cytokine-1), produced by other cells (astrocytes, choroid plexus), which are capable of
regulating macrophage function.
26
2.2 Wnt Signaling system
In 1982, Nusse and Varmus identified a proto-oncogene Wnt1 (originally called
Int-1) as a signalling molecule affects the development of mammary tumors (Nusse et
al,. 1982). After several years, WNT1 and WG (Wingless), its Drosophila melanogaster
orthologue, emerged as key morphogens, which functions as regulators of the
embryonic body plan (Barker et al., 1988; McMahon et al., 1989). Wnts ligands are
secreted glycoproteins that play essential roles in embryogenesis and cortical
development. Wnts, as ligands, interact with 7-transmembrane, G-protein-coupled
receptors called Frizzleds (Fz) to initiate several different signaling pathways.
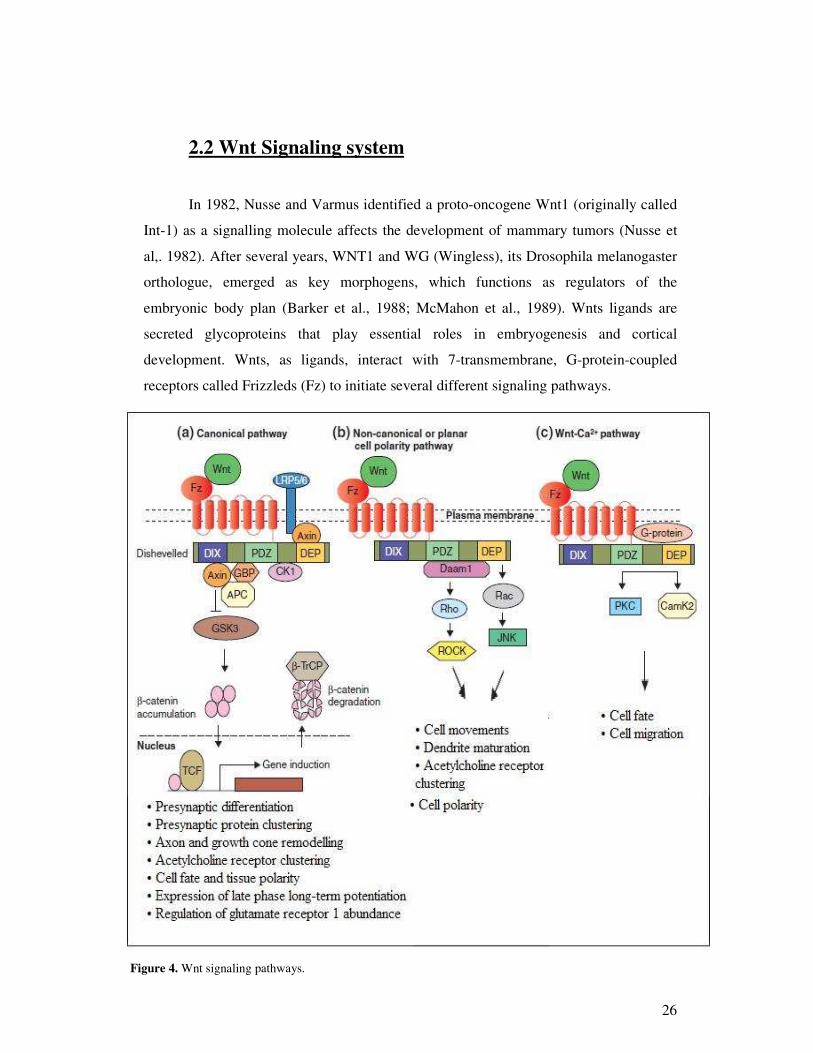
Figure 4. Wnt signaling pathways.

27
Canonical pathway (fig.4A) mediates gene induction events. This is the best-
characterized WNT signaling pathway, in which WNT binding to Frizzled receptors
activates the scaffolding protein Dishevelled (DVL), which via the DIX and PDZ
domains disassembles a so-called ‘destruction complex’ formed by glycogen synthase
kinase 3 (GSK3), Axin and adenomatous polyposis coli (APC) — a complex that
normally leads to the degradation of -catenin. WNT binding to Frizzled disrupts the
destruction complex, and this result in cytoplasmic stabilization of -catenin (He et al.,
2004; Liu et al., 1999) and its import into the nucleus, where it regulates gene
expression through association with lymphoid enhancer factor/T cell factor (LEF/TCF)
transcription factors. In this pathway, Frizzled collaborates with a co-receptor, LRP5/6
of the low-density lipoprotein receptor related protein (LRP) family.
The non-canonical or PCP pathway (fig.4B) mediates cell polarity, cell
movements during gastrulation and other processes, by signal transduction through the
PDZ and DEP domains of Dsh, leading to a modification of the actin cytoskeleton
(Weeman et al., 2003; Wallingford et al., 2002). At the level of Dsh, two independent
and parallel pathways lead to the activation of the small GTPases Rho and Rac.
Activation of Rho requires the formin-homology protein Daam1 that binds to the PDZ
domain of Dsh, leads to the activation of the Rho-associated kinase ROCK, and
mediates cytoskeletal re-organization (Habas et al. 2001; Marlow et al., 2002). Rac
activation is independent of Daam1, requires the DEP domain of Dsh, and stimulates
Jun kinase (JNK) activity (Habas et al., 2003; Boutros et al., 1998). Other Dsh-binding
molecules that influence the PCP pathway include Strabismus and Prickle, but their
mechanisms of action remain incompletely understood (Keller et al., 2002).
The Wnt-Ca2+ pathway (fig. 4C) is thought to influence both the canonical and
PCP pathways (Weeman et al., 2003). Wnt signaling through Frizzled receptors leads to
the release of intra-cellular Ca2+ in a process mediated through heterotrimeric G-
proteins and involving numerous other molecules, including phospholipase C (PLC),
calcium-calmodulin-dependent kinase 2 (CamK2) and protein kinase C (PKC) (Weeman

28
et al., 2003; Kuhl, 2002). The Wnt-Ca2+ pathway is important for cell adhesion and cell
movements during gastrulation.
2.2.1 Wnt biogenesis
Wnts are conserved in all metazoan animals. In mammals, complexity and
specificity in wnt signaling are achieved through Wnt ligands, which are cystein-rich
proteins of approximately 350-400 amino acids that contain an N-terminal signal
peptide for secretion. Murine Wnt 3a was the first purified and biochemically described
Wnt proteins (Willert et al., 2003). In addition to N-linked glycosylation, which is
reported to be required for Wnt 3a secretion (Komekado et al., 2007), Wnt 3a undergoes
two types of lipid modifications that likely account for hydrophobicity and poor
solubility of Wnt proteins (Hausmann et al. 2007). Willert and colleagues reported
lipidation for wnt ligands, it was addition of palmitate to cystein 77 (Willert et al. 2003).
Its mutation had minimal effect on Wnt 3a secretion, but diminished the ability of Wnt
3a to activate beta-catenin signaling pathaway (Galli et al., 2007; Komekado et al.,
2007; Willert et al. 2003). The second identified lipidation was a palmitoleoyl attached
to serine 209 and its mutation resulted in Wnt 3a accumulation in the endoplasmic
reticulum (ER) and a failure in secretion (Takada et al., 2006)
Two additional structures were identified for Wnt secretion: Wntless (Wls), also
known as Evenness interrupted (Evi) or Sprinter (Srt), in Drosophila, and the retromer
complex in nematodes (Hausmann et al., 2007). Wls is a multipass transmembrane
protein localized in the Golgi, edoplasmic compartments and in the plasma membrane.
Wls is essential for Wg/Wnt secretion. The retromer complex, which is composed form
five subunits was reported and defined first in the yeast. It mediates protein trafficking
between Golgi apparatus and endosomes (Hausmann et al., 2007). Loss of retromer
functions causes degradation of Wls in the lysosomes and results in reduction of Wls
and thus Wnt secretion
To conclude, Wnt is glycosylated and lipid modified by Porcupine in the ER and
is escorted by Wls from the Golgi to the plasma membrane for secretion. Wls is recycled
be endocytosis and trafficked back to Golgi by the retromer.

29
2.2.3 Wnt signalling and adult neurogenesis.
Adult neurogenesis in the mammalian brain is usually considered an active
process including the proliferation and cell fate specification of adult neural progenitors
(Duan et al., 2008). In the intact adult mammalian CNS, active neurogenesis occurs in
two ‘neurogenic’ regions: the subgranular zone (SGZ) of the dentate gyrus in the
hippocampus and the subventricular zone (SvZ) of the lateral ventricles in the forebrain
(Lie et al., 2004). New neurons are thought to originate from multipotent adult neural
stem cells, but their exact identity is still subject to debate and their multipotency at the
clonal level in vivo has not been universally demonstrated.
Recently, NEuROD1, a pro-neurogenic transcription factor in the adult brain that
is selectively expressed in dividing neural progenitors and in immature granule neurons
in the adult dentate gyrus, was identified as a downstream effector of Wnts in adult
neurogenesis (Kuwabara et al., 2009; Gao et al., 2009). WNT3A treatment induced the
expression of NEuROD1 in adult neural progenitors in vitro, and -catenin was directly
associated with the NEuROD1 gene promoter during the course of neurogenesis
(Kuwabara et al., 2009), Deletion of NEuROD1 in stem cells stopped neurogenesis in
vivo (Gao et al., 2009), and Wnt treatment of these cells did not stimulate neurogenesis
(Kuwabara et al., 2009). Wnt signalling has also been shown to modulate neurogenesis
in the SVZ. WNT3A and WNT5A increase the proliferation of cultured progenitor cells
dissected from postnatal and adult mouse SVZ and promote their neuronal
differentiation (Yu et al., 2006). In addition, retrovirus-mediated expression of stabilized
-catenin or treatment with an inhibitor of GSK3 promoted the proliferation of
progenitor cells in the SVZ. Conversely, expression of the Wnt antagonist DKK1
reduced the proliferation of progenitor cells (Adachi et al., 2007). These studies indicate
that activation of Wnt signaling regulates adult neurogenesis in the SVZ by regulating
progenitor cell proliferation.
Stem cell differentiation and proliferation are controlled by both intrinsic and
extrinsic regulators. Wnt ligands are among the extracellular factors that affect this
process (Ming et al., 2005; Nusse, 2008). During development Wnts act on CNS
progenitor cells, and the activation of -catenin leads to the proliferation of the neural

30
progenitors, resulting in the expansion of the entire neural tube (Chenn et al., 2002). In
addition, a GSK3 inhibitor was reported to induce the selective differentiation of stem
cells into neurons, and WNT7A promoted the maturation of neural precursor cells into
mature neurons (Hirabayashi et al., 2004. Wnts secreted by hippocampal progenitors
self-stimulate canonical Wnt signalling, and inhibition of this autocrine Wnt pathway
increases the number of neurons and leads to a loss of the multipotency of the
progenitors (Wexler et al., 2009). Inhibition of Wnt signalling by lentiviral expression of
a dominant negative Wnt in the dentate gyrus reduces neurogenesis in the hippocampus
(Zhou et al., 2002) and decreases long-term retention of object recognition in adult rats
(Jessberger et al., 2009)
2.2.4 Wnt signalling and Alzheimer disease (AD)
Studies in humans indicate that Wnt signalling is related to neurogenesis and is
altered or involved in the pathophysiology of AD. An example is the reduced renewal
capacity of glial-like progenitor cells dissected from the temporal cortex of patients with
AD — which correlated with elevated levels of GSK3 activity and increased
phosphorylation of -catenin — compared with that of cells from healthy controls (He et
al., 2009). Moreover, treating glial precursor cells from healthy brains with amyloid-
peptide (A) also led to increased -catenin phosphorylation level and reduced
neurogenesis. Conversely, -catenin transfection led to restoration of neurogenesis (De
Ferrari et al., 2000). These studies suggest that Wnt signaling is required for human
cortical neurogenesis, impaired Wnt signalling reduce the capacity of progenitors to
undergo neurogenesis and contribute to repair.
Almost a decade ago a relationship between loss of Wnt signalling and A
induced neurotoxicity was proposed (De Ferrari et al., 2000, De Ferrari et al., 2003).
The alteration in Wnt signalling was suggested to be the triggering factor for A
production and tau hyperphosphorylation, which induce synapse and neuron loss (De
Ferrari et al., 2003; Mudher et al.,2002). Since then, other studies have shown that
several Wnt signalling elements are altered in AD (Mudher et al., 2002; Caricasole et
al., 2003; Smal et al., 2008; Boonen et at., 2009). A directly binds to the Fz5 receptor
cysteine-rich domain at or in close proximity to the Wnt-binding place, inhibiting the

31
canonical Wnt signalling pathway (Magdesian et al., 2008). In addition, genetic studies
demonstrated a link between Wnt signaling and AD. The apolipoprotein E �4 allele,
which is supposedly associated with an increased risk of developing AD, inhibits
canonical Wnt signalling on stimulation with WNT7A (Caruso et al., 2006).
To find out whether activation of Wnt signaling could protect hippocampal
neurons from the A toxicity, the effects of activating other signalling pathways that
crosstalk with the Wnt signaling pathway was examined (Inestrosa et al., 2008). As
cholinergic dysfunction has been observed in patients with AD, treatment rodents with
an M1 muscarinic AChR receptor agonist (Farias et al., 2004) or nicotine (Inestrosa et
al., 2008) was examined. Activation of Wnt signalling through cholinergic activation
seems to be a neuroprotective mechanism against A. In fact, it is well known that M1
agonists increase the non-amyloidogenic processing of the amyloid precursor protein
(APP), reducing A production and toxicity. In addition, cholinergic activation by the
specific M1 agonist induces the phosphorylation of GSK3 in neuronal cultures from
transgenic mice that overexpress GSK3102. Ser9 phosphorylation of GSK3 by
cholinergic stimulation is probably mediated by a mechanism involving protein kinase C
(PKC), as it was blocked by a PKC inhibitor (Farias et al., 2004). The protection
observed in vitro has been confirmed in vivo. Treatment with the specific M1 agonist
improved the spatial memory and reduced the A load in the hippocampus of transgenic
mouse model of AD. These results show that cholinergic activation interacts with the
Wnt signalling pathway, leading to potential neuroprotection against A toxicity.
Recent evidence suggests that lithium is neuroprotective agent in various
neurodegenerative diseases and it it should be noted that lithium reduces the prevalence
of AD in elderly patients with bipolar disorders (Nunes et al., 2007). Studies in a mouse
model of AD indicated that lithium reduces the size of the amyloid burden, including the
A oligomers, and prevents the behavioural alterations of the animals (Toledo et al.,
2009). Under these conditions lithium activates Wnt signalling, by the inhibition of
GSK3 and the increase in -catenin (Toledo et al., 2009). These studies are consistent
with the hypothesis that loss of Wnt signalling is involved in A-dependent
neurodegeneration and activation of the canonical Wnt pathway by lithium protects
against the synaptic changes triggered in AD.

32
The known functions of Wnt signalling in the adult brain suggest that disruptions
or impairments of the Wnt pathways could have dramatic consequences. Indeed,
growing evidence implicates Wnt signaling in the pathogenesis of several
neurodegenerative and neurological diseases. In the future it will be important further
explore the molecular mechanisms that link these pathways to disease, in particular to
the pathogenesis of Alzheimer disease and Parkison disease.
The emerging roles of Wnts in adult neurogenesis, neuronal differentiation,
synaptogenesis and survival suggest that targeting Wnt signalling pathways could offer
therapeutic benefits. Drugs capable of modulating Wnt signalling may become tools for
regenerative or neuroprotective medicine, for example against diseases associated with
neuron loss.
2.2.5 The structure of Dvl
Dvl proteins have three conserved domains: an aminoterminal DIX domain of 80
amino acids, a central PDZ domain of about 90 amino acids, and a carboxyl-terminal
DEP domain of 80 amino acids (Boutros et al., 1999). In addition, another two
conserved regions, the basic region and the proline-rich region, are also implicated to
mediate protein–protein interaction and/or phosphorylation.
The DIX domain. Both overexpressed and endogenous Dvl proteins form
cytoplasmic puncta (Yanagawa et al., 1995; Yang-Snyder et al. 1996). Recent studies
Figure 5. Dvl domain structure. Dvl protein is about 700 amino acids, harboring conserved DIX, basic and serine/threonine-rich region, PDZ, praline-rich region and DEP domains.

33
revealed that Dvl puncta undergo dynamic polymerization, which to some extent
correlate with the ability of Dvl to activate Wnt/-catenin signaling (Schwarz-Romond
et al., 2005; Smalley et al., 2005). Dvl polymerization is mediated by its DIX domain
(Wharton et al., 2003). The structure of Dvl's DIX domain has not been defined, but the
crystal structure of DIX domain of rat Axin, which is closely related and shares similar
properties of Dvl's DIX domain (Schwarz-Romond et al., 2007; Fagotto, et al. 1999),
has been solved (Schwarz-Romond et al., 2007). The single DIX domain has a compact
fold with five -strands and one -helix. The structure reveals that multiple -strands
engage in head-to-tail interaction between two different surfaces of DIX domains. Some
of the residues in the core structure or those involved in the 2–4 interactions of the
Axin DIX domain are highly conserved in the DIX domains of Dvl. Mutations at the
core structure or the interaction surface strongly diminish the Wnt signaling (Schwarz-
Romond et al., 2007).
The PDZ domain. PDZ domain is a modular protein interaction domain that has
tow helices and six sheets, which together with the preceding loop form a peptide-
binding cleft (Cheyette et al.. 2002). Dvl exploits its PDZ domain to transducer signals
from the membrane receptor Fz to downstream components by direct interaction with Fz
(Wong et al., 2003; Barker and Clevers, 2006). The special role of the Dvl PDZ domain
in the Wnt pathway makes it an ideal pharmaceutical target (Barker and Clevers, 2006).
The therapeutic usefulness of PDZ protein–protein interaction interference has been
clearly shown using small peptide (Chen et al., 2009; Lee et al., 2009; Zhang et al.,
2009) or non-peptide antagonists which block the PDZ-mediated interactions (Fujii et
al., 2007; Grandy et al., 2009). For example, a virtual screen of small molecule
compounds in the three-dimensional databases has identified molecules predicted to
bind to the Dvl PDZ domain and therefore inhibit its interaction with Fz (Shan et al.,
2005). At least two promising candidates, NSC668036 (Shan et al., 2005) and
compound 3289–8625 (Grandy et al., 2009), have been subsequently synthesized and
tested for their capacity to interact with Dvl using NMR spectroscopy.
The DEP domain. The DEP domain in the C-terminal region of Dvl consists of
a bundle of three -helices, a -hairpin “arm” composed of two -strands, and two short

34
-strands. In light of the structure, the protein surface that includes K434, D445, and
D448, is predicted to be crucial for the interaction between Dvl and other proteins and to
play an important role in signal transduction (Wong et al., 2000) It was recently
suggested that the basic residues in the DEP domain are essential for the binding.
The basic region and Pro-rich region Preceding the amino terminal of the PDZ
domain is a cluster of positive charged (basic) residues. This region contains the
phosphorylation sites targeted by CK2 and PAR-1 (Willert et al. 1997), and together
with the PDZ domain, this region is involved in the direct binding with the EFX domain
of Naked Cuticle (Nkd) (Rousset et al. 2001).
2.2.6 Dishevelled controls apical docking of basal bodies and planar cell
polarization.
The motile cilia of multi-ciliated cells are very significant to the physiology of a
variety of organs. There are accumulating findings demonstrating Dvl involvement in
the formation of basal bodies. First, Dvl is required for actin erection and basal body
docking to the apical cell surface of ciliated cells. Dvl might be required for
maintenance of basal bodies at the apical surface. Dvl with Inturned, the effector
protein, are required for local activation of the Rho GTPase, specifically at the apical
surface of ciliated epithelial cells. It is already reported that Dvl links basal body
docking to vesicle traffic and Sec8 localization (Park et al., 2008).
Transmission Electron Microscopy (TEM) studies of both multi-ciliated cells
and cells with only a primary cilium have suggested that basal body docking is achieved
by association of basal bodies with cytoplasmic vesicles, which then fuse to the apical
cell surface by a mechanism similar to exocytosis (Sorokin et al., 1968; Cohen et al.,
1998). It is possible then to predict a model whereby Dvl associates closely with the
basal body complex and the recruitment of Sec8-positive exocytic vesicles. Inturned
may serve as a scaffold that places Rho closely with Dvl, thereby activating the GTPase
(Habas et al., 2001; Park et al., 2005). Because actin is crucial for fusion of vesicles to
the plasma membrane (Lanzetti, et al. 2007), it is noteworthy that apical actin assembly
is also altered in multi-ciliated cells lacking Dvl or Inturned. Dvl probably controls actin

35
assembly in multi-ciliated cells via CapZIP, which is phosphorylated downstream of
Dishevelled (Oishi et al., 2006).
Although the detailed mechanisms of Dvl function in basal body docking remain
to be covered, this link to vesicle traffic has broad implications. In addition to basal
body docking, Dvl controls canonical Wnt signaling and PCP signaling, but a unifying
mechanism for Dvl function has yet to be find out (Wallingford et al., 2006). An
association between Dvl and membranous vesicles is a matter of persistant debate
(Schwarz-Romond et al., 2005); however, several lines of evidence support this
hypothesis. First, association of Dvl with vesicles has been shown to be important for
canonical Wnt signaling (Capelluto et al., 2002), and it administers the aggregation of
the Wnt co-receptor Lrp6 into membrane-associated signalsomes (Bilic et al., 2007).
Second, intracellular PCP signaling components, including Dvl, interact functionally
and physically with elements of the membrane trafficking machinery (Yu et al., 2007;
Classen et al., 2005; Kishida et al., 2007; Chen et al., 2003). Finally, Dvl has been
included in endo- and exocytosis at synapses (Ahmad-Annuar 2006; Kishida et al.,
2007).
A connection between Dvl, membrane traffic and the basal body is especially
intriguing in light of recent results linking cilia and Wnt signaling. The cilium and basal
body are necessary organelles governing the activity of Wnt signaling pathways
(Simons et al. 2005; Ross et al., 2005; Gerdes et al., 2007). The interconnections
between Wnt signaling, vesicle traffic and ciliogenesis are best explained by the BBS4
protein. BBS4 cooperates genetically with both canonical Wnt and PCP signaling (Ross
et al., 2005; Gerdes et al., 2007), but it also mediates membrane delivery to the primary
cilium and controls microtubule organization and ciliogenesis (Kim et al., 2004;
Nachury et al., 2007). Moreover, ciliary and basal body proteins can directly regulate by
Dvl phosphorylation, localization and stability (Gerdes et al., 2007; Oishi et al., 2006).
Connecting these disparate findings will be a challenge for future investigation.
It is also important that both Dvl and Rho play roles in apical docking that are
experimentally separable from those involved in planar polarization of basal bodies. The
direction of ciliary beating is formed subsequent to basal body docking and cilia
outgrowth (Boisvieux-Ulrich et al., 1985; Frisch et al., 1968; Mitchell et al., 2007), it
seems reasonable to conclude that Dvl and Rho act first in apical docking and only later

36
affects planar polarization. Nonetheless, it will be important to find out how Dvl at the
basal body discriminates between its apical docking function and its planar polarity
function. One possibility is that Rho activity for apical docking and for planar
polarization regulated by distinct guaninenucleotide exchange molecules. This idea is
supported by the evidence that knockdown of ARHGEF11 causes a spectrum of
embryonic defects, resulting defective cilia-mediated fluid flow (Panizzi et al., 2007;
Kramer-Zucker et al., 2005). In ARHGEF11 morphants, only apical actin is disrupted in
multi-ciliated cells, but cilia are grossly normal and motile (Panizzi et al., 2007), so this
exchange factor may function exclusively in polarization of ciliary beat.

37
3. Experimental part 3.1 Materials and methods 3.1.1 Cell culture
The preparation of culture mediums and all manipulations of cells including
cell isolation and treatments were carried out under a laminar flux under sterile
conditions. For immunocytochemistry cells were culturedonto coverslips in 35-mm Petri
dishes.
Cells isolation from rat SVZ. SVZ cell cultures were prepared form newborn
rats, which were scarified at PO or P1 by decapitation. Brains were removed from the
skull and put in one drop cold Hank’s solution in a 100 mm Petri dish. By visualizing
with microscope first coronal cut was performed in order to remove the rostral part of
the hemispheres. A second coronal cut was performed at the level of anterior horn of
lateral ventricles. The ependymal cells were quickly removed with forceps at the third
ventricle and put in a 100 mm Petri dish with cold Hanks solution to preserve the tissue.
Once finished all animals SVZ cells were add in 3 ml Hanks into using and very gently
homogenized using 1 ml pipette. Then the tube was filled up to 10ml and trypsin (10
µm/brain) was added. Cells were incubated in tripsin for 15 minutes at 37ºC in order to
���������������� ����� �������������
�������������� ������������������ ��
Figure 6. Isolation of Ependymal cells. Ependymal cells were dissected from third ventricles of PO-P1 rat puppies. It later was cultured on the laminin coated dish.

38
digest intracellular contacts and separate cells. Then, tripsin was stopped with 1 ml FCS
and cells recovered be centrifugation at 1200rpm/min for 10 minutes. Cells were
resuspended in 1 ml complemented Neurobasal medium (Gibco Brl, Paisley, Uk) and
washd three times in Neurobasal medium. Cells were counted, resuspended in medium
an homogenously mixed by using a vortexing machine before plating them onto laminin
coated cell culture supports. Cells were allowed to grow in Neurobasal medium with 2
% B27 supplement, 2mM glutamaine and 1 mM sodium pyruvate at 37 ºC and 5% CO2.
Cells were cultured for 7-10 days and medium was replaced 2 times during that period.
This ependymal cells culture consisted of atrocities, neuroblasts, neurons and
ependymal cells. Ependymal cells were at the different level of maturation.
Immunostaining on ependymal cell culture. In the begging cells were with 2
ml of PAF 4% for 30 minutes. In order to remove remaining PAF 4% it was perform a
3 times washing with PBS for 10 minutes. PBS solution was removed and added 2ml
PBS-BSA-Tryton solution (for 1h) to cover unspecific antigens. Next step was to add
100µm primary antibody (Wnt 3a, 5a, 7a, 7b, 8b and Dvl 1, 3 were diluted 1:50 in PBS -
0.5%; Dvl-2 was diluted 1:100 in PBS -0.5% ) and laeve overnight at 4ºC temperature.
After ~12h all volume of primary antibody was removed and there were three series of
washing with PBS. Later secondary fluorescence antibody (diluted 1:1000 into PBS-
BSA) was added on the cells and kept it for 1h at room temperature (without light).
Solution with antibody must be removed and again washed three times with PBS. In
order to stain nucleus, I used Hoechst fluorescent DNA-binding compound. Prepared
solution (1:5000 in PBS) was deposited on the slices for 30 minutes at room
temperature. In the end slices were mounted: one drop of PBS- glycerine on the cells
and cover with slice. Slides were dried at room temperature.
During each immunolabeling experiment a negative-control was done, where
only the secondary antibody was added to the cells; in this way the background intensity
provoked by unspecific bound could be assessed.

39
Fixation. The step of fixation plays an essential role to preserve the cellular
structure and to avoid the risqué of morphological modifications. In general our cells
were fixed with paraformaldehyde solution at 4 % diluted in phosphate buffer. This
chemical fixator is the polymerized form of formaldehyde, reacting especially with
amino groups of the proteins thus creating bridges between between different
polypeptide chains. It ensures good preservation of nucleoproteins and proteins.
Sometimes this fixative preserves the enzymatic activity but never the lipids
constituents.
Specimens were carefully transferred into the dish filled with 4% PAF for an
overnight fixation at 4°C (cell culture were fixed just for 30 minutes)
3.1.2 Brain slices
Vibratome section’s preparation. Rats were anesthetized with sodium
pentobarbital (50mg/kg, intraperitoneally) and perfused through the ascending aorta
with saline rinse, followed by 500ml of PAF 4% fixative. After removal, brains were
placed in the fixation solution overnight and block for Vibratome sectioning the
following day. Tissue sections were cut into PBS buffer on Vibratome at 50 µm. Tissue
specimens are ready for immunostaining
Immunostaining on vibratome sections. The most suitable for experiment
tissue specimens were selected. Slices were separated 2-6 per well. In order to remove
remaining PAF 4% it was perform a 3 times washing with PBS for 10 minutes. PBS
solution was removed and added PBS-BSA-Tryton solution (for 1h) to cover unspecific
antigens. After removing previous solutions, primary antibody (Wnt 3a, 5a, 7a, 7b, 8b
and Dvl 1, 3 were diluted 1:50 in PBS -0.5%; Dvl-2 was diluted 1:100 in PBS -0.5% )
was added and left it for 48 hour at 4ºC temperature. After 48h all previous solution
were removed and there were three series of washing with PBS. Later secondary
fluorescence antibody (diluted 1:1000 into PBS-BSA) was added on the slices and kept
it for 2h at room temperature (without light). Solution with antibody must be removed
and again washed three times with PBS. In order to stain nucleus, I used Hoechst

40
fluorescent DNA-binding compound. Prepared solution (1:5000 in PBS) was deposited
on the slices for 30 minutes at room temperature. In the end slices were mounted: one
drop of PBS- glycerine on the sections and cover with slice. Slides were dried at room
temperature.
During each immunolabeling experiment a negative-control was done, where
only the secondary antibody was added to the cells; in this way the background intensity
provoked by unspecific bound could be assessed.
Wholemount Dissection. The animal was sacrificed by cervical dislocation
and the head is cut off. A midline incision is made, posterior to anterior, along the scalp
to reveal the skull. A series of 4 cuts in the skull are made to open the cranium: one cut
spans the two orbits anterior to the olfactory bulbs, the next two cuts are inferior to the
cerebellum and separate the cranium from the skull base and the final cut runs posterior
to anterior along the mid-sagittal suture. The cranial flaps are gently retracted and the
brain is extracted and placed into the dissecting dish. The dissection is performed under
the stereomicroscope. First, the olfactory bulbs are dissected away from the brain.
Divide the brain along the interhemispheric fissure. A coronally-oriented cut is then
made at the posterior-most aspect of the interhemispheric fissure, allowing the caudal
hippocampus to be visualized in cross-section. The hippocampus, which forms the
medial wall of the lateral ventricle at this position, must then be released from the
overlying cortex, which forms the dorsal-lateral wall of the ventricle. First, the knife is
inserted into the small ventricular space between the cortex and hippocampus dorsally,
and a cut is made in the cortex where it reflects ventrally, away from the midline, to join
the hippocampus. After this cut is made, the cortex can be slowly peeled away from the
hippocampus to reveal the lateral ventricle moving from dorsal to ventral. This
maneuver is expedited by cutting off a wedge of cortex at the corner where the
hippocampus was released. After reaching the ventral-most extent of the lateral ventricle
at this position, you may either visualize or feel where the cortex again wraps around,
this time reflecting back medially, to join the hippocampus. Another cut must be made
in this position to completely release the hippocampus or medial wall of the lateral
ventricle from the cortex or lateral wall of the lateral ventricle. The hippocampus is
pulled away from the cortex, medially and anteriorly, to open the lateral ventricle

41
widely. The hippocampus is pulled anteriorly using small strokes of the forceps and
knife to retract the medial and lateral walls apart. Once the resistance to this retraction
begins to increase, additional cuts are needed. First, to increase your exposure to the
lateral ventricle and in particular, the lateral wall and SVZ, dissect away the cortex. The
cortex is cleanly dissected away by visualizing the interface between the corpus
callosum and the VZ/SVZ. Simply cut along this interface staying on the callosal side to
avoid damaging the SVZ. In order to continue retracting the medial wall away from the
lateral wall, two more cuts are needed: one cut dorsally where the lateral wall, medial
wall, and cortex all converge, and one cut ventrally where the lateral wall, medial wall,
and thalamus converge. With these cuts made, further gentle retraction on the medial
wall allows the anterior-most extent of the lateral ventricle to be opened. Next step is to
separate the medial and lateral walls anteriorly. Cut exactly in this valley to separate
these two walls. The specimen is ready for further investigations.
Cryostat section’s preparation. Wholemount was obtain by following steps
as described above and overnight fixed with PAF 4%. Fixed wholemount specimen was
saturated in cryoprotectant (30% sucrose in 0.1M PBS for 48h).
Evenly place embedding solution on base of chilled platform. As the solution
begins to freeze, carefully align and place tissue specimen (brain or wholemount,
ventricle side down) on platform. The wholemount should adhere to the platform. Cover
entire specimen with embedding solution. This may take several application/freezing
steps to accomplish. Set the cutting temperature of cryostat to -20ºC. Section brain in
cryostat at a thickness of 20 µm. Each section were transferred to a subbed slide. Slides
were placed on warming plate at 40ºC to dry and put in holder, then freezed at -20ºC
overnight. Tissue specimens are ready for immunostaining
Immunostaining on cryostat sections. Slides with sections were defreezed at
room temperature for 1.5h. Later, they were kept in PAF 4% for 1h. In order to remove
tissue-teck and PAF 4% slides were rinsed 3 times with PBS for 10 minutes. Next step
is to paint with special pen around the sections and add the solution PBS-BSA-TX (to
cover unspecific sites) for 30 minutes. After removing previous solutions, primary anti-
Wnt 8b antibody (1:50 diluted in PBS -0.5%) was added and left it for 48 hour at 4ºC

42
temperature. After 48h all previous solution were removed and there were three series of
washing with PBS. Later secondary fluorescence antibody (diluted 1:1000 into PBS-
BSA) was added on the slices and kept it for 2h at room temperature (without light).
Solution with antibody must be removed and again washed three times with PBS. In
order to stain nucleus, I used Hoechst fluorescent DNA-binding compound. Prepared
solution (1:5000 in PBS) was deposited on the slices for 30 minutes at room
temperature. The last step is mounting the slice: several drops of the PBS- glycerine on
the sections and cover with slice. Let the slide dry at room temperature (without light)
and it is ready for analysis.
3.1.3 Analysis
Fluorescent images were obtained with a confocal microscope (Zeiss LSM 510 Meta,
Carl Zeiss Inc., Thornwood, NY), and digital images were captured and imported with
the LSM 5 image browser (Carl Zeiss Inc.).

43
Figure 7. Immunohistochemical analysis of Wnt 8b. A – Intensive signal in the ependymal cells (scale bar 100 µm). B – Clusters of cilia are labeled, no staining in ependymal cells bodies (scale bar 20µm). C – confocal analysis reveals Wnt 8b immunoreactivity just in basal part of the cilia (scale bar 5 µm). Red –Wnt8b, green – tubulin
3.2 Results
3.2.1 Analysis of Wnt ligands.
Wnt signaling proteins affects many aspects of animal development and
regeneration after injury. Wnt8b has been reported to be expressed in the dorsomedial
telencephalon, hippocampus and other brain regions (Tole et al., 2000; Theil et al.,
2002; Kimura et al., 2005; Shimogori et al., 2004), but immunohistochemical analysis
in the ventricular zone cells (ependymal) has not been done before.
Immunohistochemical experiments with WNT 8b antibody were carried out on
coronal vibratome sections from perfused and fixed adult rat brains. Low power analysis
with a fluorescence microscope of all brain structure revealed strong immunoreactivity
in the subventricular zone of third (Figure 7. A) and lateral ventricles. Also, we detected
strong signal of Wnt 8b in the hippocampus, these results corresponds to Lako and
colleagues findings in the mouse brains (Lako et al., 1998). Analysis with a high power
confocal microscope helped to detect immunoreactivity on the apical surface of
ependymal cells (Figure 7. B)
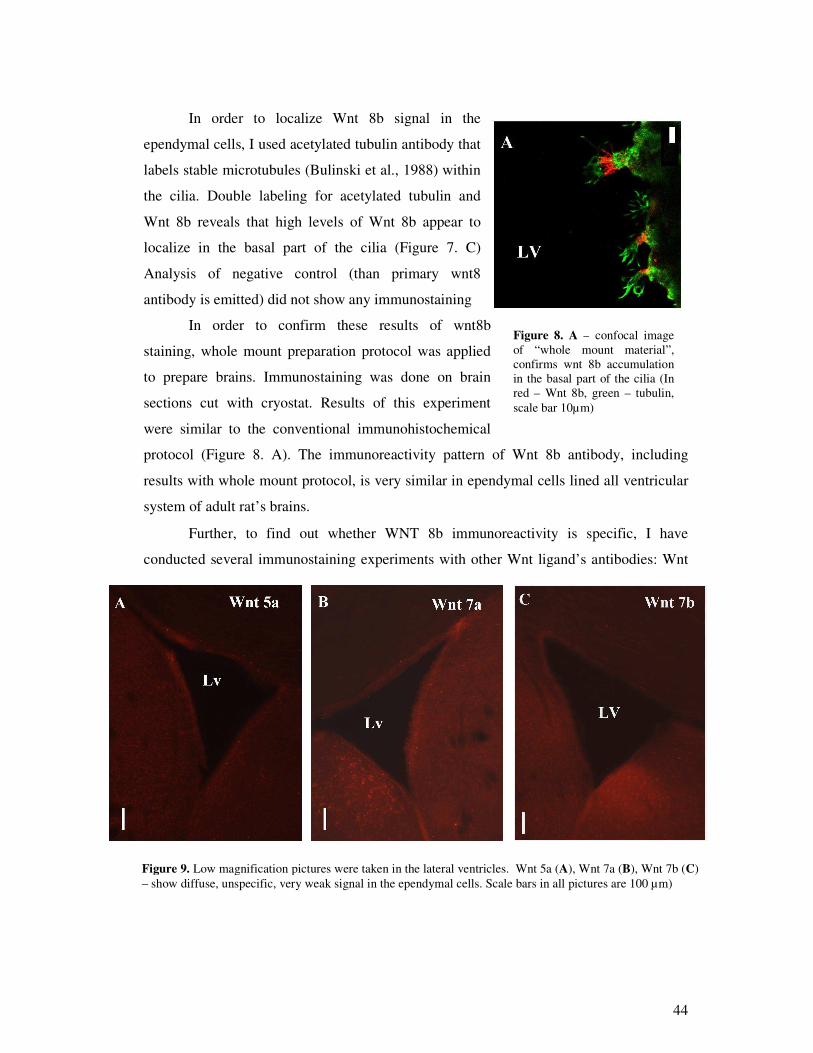
44
Figure 8. A – confocal image of “ whole mount material” , confirms wnt 8b accumulation in the basal part of the cilia (In red – Wnt 8b, green – tubulin, scale bar 10µm)
In order to localize Wnt 8b signal in the
ependymal cells, I used acetylated tubulin antibody that
labels stable microtubules (Bulinski et al., 1988) within
the cilia. Double labeling for acetylated tubulin and
Wnt 8b reveals that high levels of Wnt 8b appear to
localize in the basal part of the cilia (Figure 7. C)
Analysis of negative control (than primary wnt8
antibody is emitted) did not show any immunostaining
In order to confirm these results of wnt8b
staining, whole mount preparation protocol was applied
to prepare brains. Immunostaining was done on brain
sections cut with cryostat. Results of this experiment
were similar to the conventional immunohistochemical
protocol (Figure 8. A). The immunoreactivity pattern of Wnt 8b antibody, including
results with whole mount protocol, is very similar in ependymal cells lined all ventricular
system of adult rat’ s brains.
Further, to find out whether WNT 8b immunoreactivity is specific, I have
conducted several immunostaining experiments with other Wnt ligand’ s antibodies: Wnt
Figure 9. Low magnification pictures were taken in the lateral ventricles. Wnt 5a (A), Wnt 7a (B), Wnt 7b (C) – show diffuse, unspecific, very weak signal in the ependymal cells. Scale bars in all pictures are 100 µm)
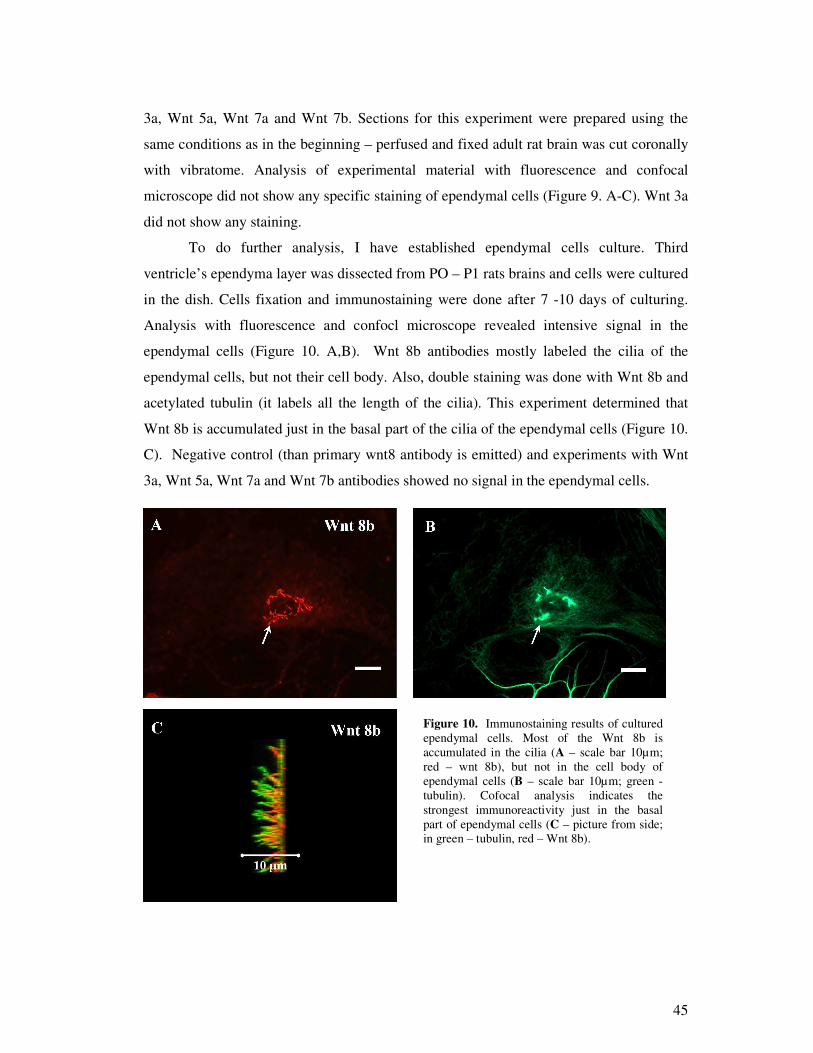
45
3a, Wnt 5a, Wnt 7a and Wnt 7b. Sections for this experiment were prepared using the
same conditions as in the beginning – perfused and fixed adult rat brain was cut coronally
with vibratome. Analysis of experimental material with fluorescence and confocal
microscope did not show any specific staining of ependymal cells (Figure 9. A-C). Wnt 3a
did not show any staining.
To do further analysis, I have established ependymal cells culture. Third
ventricle’ s ependyma layer was dissected from PO – P1 rats brains and cells were cultured
in the dish. Cells fixation and immunostaining were done after 7 -10 days of culturing.
Analysis with fluorescence and confocl microscope revealed intensive signal in the
ependymal cells (Figure 10. A,B). Wnt 8b antibodies mostly labeled the cilia of the
ependymal cells, but not their cell body. Also, double staining was done with Wnt 8b and
acetylated tubulin (it labels all the length of the cilia). This experiment determined that
Wnt 8b is accumulated just in the basal part of the cilia of the ependymal cells (Figure 10.
C). Negative control (than primary wnt8 antibody is emitted) and experiments with Wnt
3a, Wnt 5a, Wnt 7a and Wnt 7b antibodies showed no signal in the ependymal cells.
Figure 10. Immunostaining results of cultured ependymal cells. Most of the Wnt 8b is accumulated in the cilia (A – scale bar 10µm; red – wnt 8b), but not in the cell body of ependymal cells (B – scale bar 10µm; green - tubulin). Cofocal analysis indicates the strongest immunoreactivity just in the basal part of ependymal cells (C – picture from side; in green – tubulin, red – Wnt 8b).
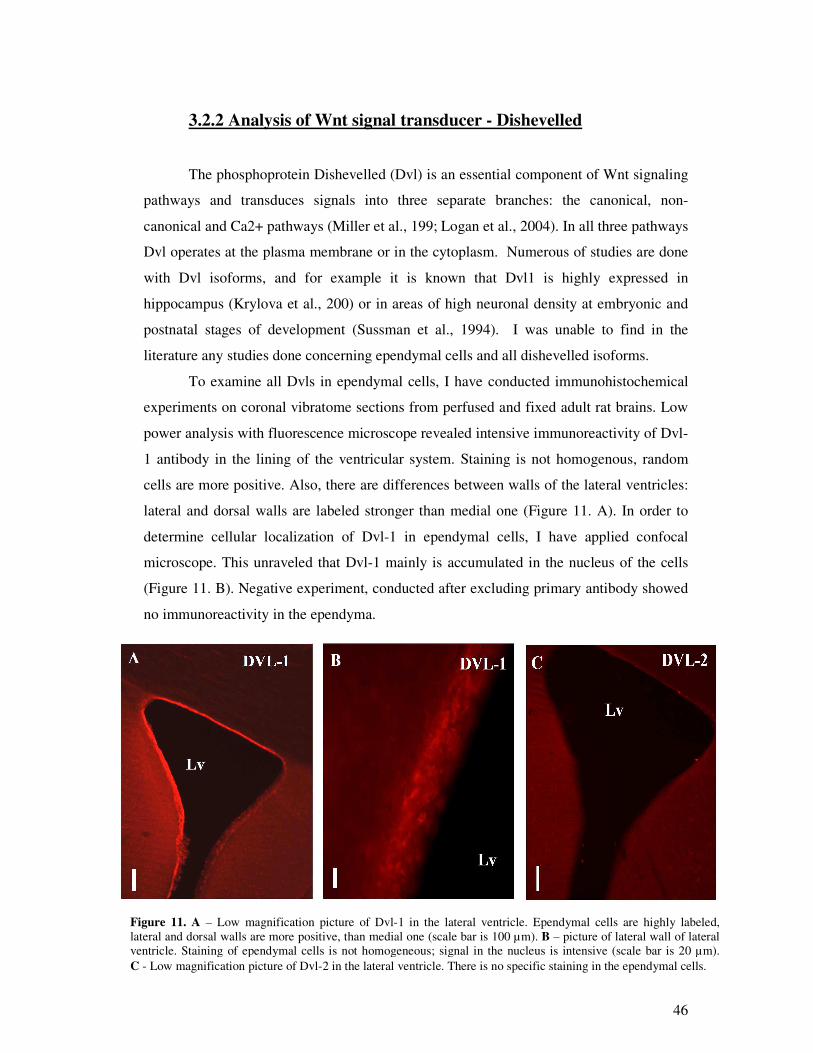
46
Figure 11. A – Low magnification picture of Dvl-1 in the lateral ventricle. Ependymal cells are highly labeled, lateral and dorsal walls are more positive, than medial one (scale bar is 100 µm). B – picture of lateral wall of lateral ventricle. Staining of ependymal cells is not homogeneous; signal in the nucleus is intensive (scale bar is 20 µm). C - Low magnification picture of Dvl-2 in the lateral ventricle. There is no specific staining in the ependymal cells.
3.2.2 Analysis of Wnt signal transducer - Dishevelled
The phosphoprotein Dishevelled (Dvl) is an essential component of Wnt signaling
pathways and transduces signals into three separate branches: the canonical, non-
canonical and Ca2+ pathways (Miller et al., 199; Logan et al., 2004). In all three pathways
Dvl operates at the plasma membrane or in the cytoplasm. Numerous of studies are done
with Dvl isoforms, and for example it is known that Dvl1 is highly expressed in
hippocampus (Krylova et al., 200) or in areas of high neuronal density at embryonic and
postnatal stages of development (Sussman et al., 1994). I was unable to find in the
literature any studies done concerning ependymal cells and all dishevelled isoforms.
To examine all Dvls in ependymal cells, I have conducted immunohistochemical
experiments on coronal vibratome sections from perfused and fixed adult rat brains. Low
power analysis with fluorescence microscope revealed intensive immunoreactivity of Dvl-
1 antibody in the lining of the ventricular system. Staining is not homogenous, random
cells are more positive. Also, there are differences between walls of the lateral ventricles:
lateral and dorsal walls are labeled stronger than medial one (Figure 11. A). In order to
determine cellular localization of Dvl-1 in ependymal cells, I have applied confocal
microscope. This unraveled that Dvl-1 mainly is accumulated in the nucleus of the cells
(Figure 11. B). Negative experiment, conducted after excluding primary antibody showed
no immunoreactivity in the ependyma.

47
Figure 12. A - Low magnification picture of Dvl-3 in the lateral ventricle. Intensive signal is in the ependymal cells; dorsal and lateral walls are labeled more positive than medial one (scale bar 100 µm). B – high magnification picture of lateral wall of lateral ventricle. Staining in ependymal cells is much stronger than in surrounding layers. Dvl-3 antibody labeled membrane and cytoplasm of ependymal cells (Scale bar 20 µm). C - Confocal image, clearly confirms that Dvl-3 in ependymal cells is accumulated close to the membrane, the concentration in the cytoplasma and nucleus is low.
The same experimental procedures, as mentioned above, were applied to
investigate Dvl-2. Analysis with fluorescence and confocal microscope detected no
immunoreactivity in the ependymal cells lined all ventricular system of the brains (Figure
10. C).
The same procedure as for Dvl-1 was used for the exploration of Dvl-3 in the
ependymal cells. Firstly, low power analysis show strong labeling of ependymal cells.
Immunoreactivity appears to be more intensive in the ependymal cells lined lateral and
dorsal walls of the lateral ventricles compare to medial wall (Figure 12. A, B). Confocal
analysis exposed intensive membranous staining of the ependymal cells (Figure 12. C).
Negative control (with omitted primary antibody) did not possess any labeling.
To do further analysis with Dvls, I have used already establish ependymal cell
culture. Confocal analysis of immunohistochemical experiments with Dvl-1 antibody
confirmed, that this prorein is mostly accumulated in nucleus (Figure 13. A). Stronger
nuclear labeling can be observed in ciliated ependymal cells comapared to not ciliated.
Double labeling for Dvl-1 and acetylated tubulin reveals that some of the Dvl-1 is
accumulated in the basal part of the cilia (Figure 13. B,C). Dvl-1 immunoreactivity in the
cilia compared to Wnt 8b is less intensive.
DVL-3 C
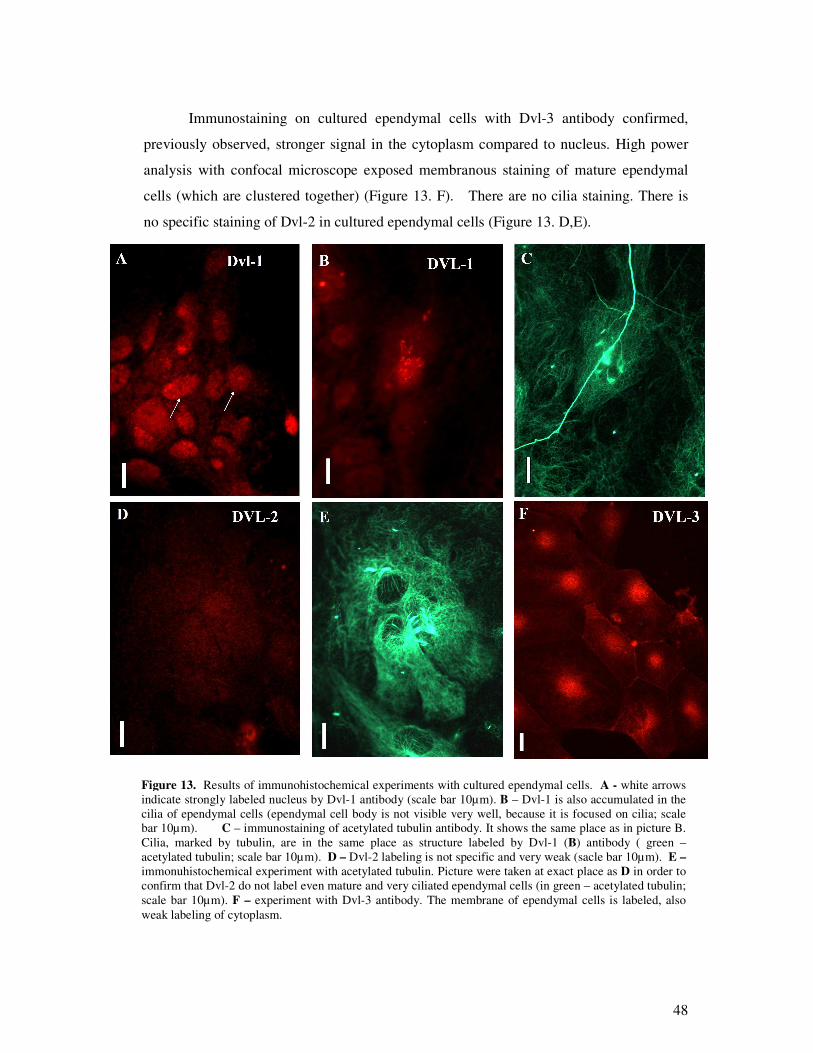
48
Immunostaining on cultured ependymal cells with Dvl-3 antibody confirmed,
previously observed, stronger signal in the cytoplasm compared to nucleus. High power
analysis with confocal microscope exposed membranous staining of mature ependymal
cells (which are clustered together) (Figure 13. F). There are no cilia staining. There is
no specific staining of Dvl-2 in cultured ependymal cells (Figure 13. D,E).
All Immunohistochemical experiments are controlled by negative control (with
omitted primary antibody), to ensure, that primary antibody is specific. In order to find
Figure 13. Results of immunohistochemical experiments with cultured ependymal cells. A - white arrows indicate strongly labeled nucleus by Dvl-1 antibody (scale bar 10µm). B – Dvl-1 is also accumulated in the cilia of ependymal cells (ependymal cell body is not visible very well, because it is focused on cilia; scale bar 10µm). C – immunostaining of acetylated tubulin antibody. It shows the same place as in picture B. Cilia, marked by tubulin, are in the same place as structure labeled by Dvl-1 (B) antibody ( green –acetylated tubulin; scale bar 10µm). D – Dvl-2 labeling is not specific and very weak (sacle bar 10µm). E – immonuhistochemical experiment with acetylated tubulin. Picture were taken at exact place as D in order to confirm that Dvl-2 do not label even mature and very ciliated ependymal cells (in green – acetylated tubulin; scale bar 10µm). F – experiment with Dvl-3 antibody. The membrane of ependymal cells is labeled, also weak labeling of cytoplasm.

49
the best immunohistochemical protocol to work with brain’ s slices, antigen retrieval
protocol was performed to all antibodies. There was no significant difference observed
between conventional and antigen retrieval protocols. Experiments were repeated several
times and different animal’ s brains were used.
Table 1. Summary of the results.
Wnt3A
Wnt5A
Wnt7A
Wnt7B
Wnt8B
Dvl-1
Dvl-2
Dvl-3
Cultured rat ependymal cells: • Cilia
• Cytoplasm
• Nucleus
• Membrane
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+
+++
-
+/-
+/-
+/-
+/-
-
++
+/-
++
Rat brain slices: • Cilia
• Cytoplasm
• Nucleus
• Membrane
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+++
-
-
-
++
+/-
+++
-
-
-
-
-
-
++
-
+++
- - no signal;
+/- - very weak or uspecific signal;
+ - weak signal;
++ - good signal;
+++ - Strong signal.

50
3.3 Discussion
WNT ligands are secreted proteins that regulate cell fate decisions, cell polarity, cell
migration, axonal morphology, and synaptic differentiation (Lucas and Salinas, 1997;
Wodarz and Nusse, 1998; Hall et al., 2000; Patapoutian and Reichardt, 2000). In
mammals, complexity and specificity of Wnt signaling are in part achieved through 19
Wnt ligands (http://www.stanford.edu/~rnusse/wntwindow.html). Several studies reported
that Wnt genes continue to be expressed in the adult brain (Parr et al. 1993) and possess
significant functions, like Wnt 5a through the non-canonical pathway regulate the
development of midbrain dopaminergic neurons (Castelo-Branco et al., 2003) or Wnt7a
affects axon guidance and remodeling (Hall et al., 2000). With that in mind, I have done
immunostaining with antibodies for Wnt 3A, Wnt 5A, Wnt 7A, Wnt 7B and Wnt 8b,
previously reported in several brain regions. Analysis of Wnt 8b immunohistochemical
experiment revealed very intensive immunoreactivity in the basal part of ependymal cell’ s
cilia. This is a novel localization of Wnt 8b in ependymal cells. There was no staining
with with Wnt 3a, Wnt 5a, Wnt 7a and Wnt 7b antibodies in vivo and in vitro. These
results suggest that staining with Wnt8b antibody in ependymal cells is specific.
The phosphoprotein Dishevelled (Dvl) is an essential component of Wnt signaling
pathways and transduces signals into three separate branches: the canonical, non-
canonical and Ca2+ pathways (Miller et al., 199; Logan et al., 2004). In all three pathways
Dvl operates at the plasma membrane or in the cytoplasm. Numerous of studies are done
with Dvls isoforms, and for example it is known that Dvl1 is highly expressed in
hippocampus (Krylova et al., 200) or in areas of high neuronal density at embryonic and
postnatal stages of development (Sussman et al., 1994). During review of the literature, I
was unable to find any immunohistochemical studies done concerning ependymal cells
and all dishevelled isoforms. Therefore, these are new results: Dvl-1 can be detected in the
nucleus and in the cilia; Dvl-3 is mostly accumulated in the cytoplasm and on the
membrane; Dvl-2 showed no specific staining.

51
3.3.1 Ependymal cells can secrete Wnt 8b
How are wnt proteins secreted? In Wnt-producing cells, the Wnt protein becomes
palmitoylated at two sites in the endoplasmic reticulum by the porcupine acyl-transferase.
The exact role of lipid modification remains unclear. A possible function is the targeting
of Wnts to particular domains of the membrane. Further transport and secretion of the
Wnt protein in secretory vesicles is controlled by the multi-pass transmembrane protein
Wntless/Evi (Ching et al., 2006), which is present in the Golgi and/or on the plasma
membrane. The mammalian ortholog of Drosophila Wntless/Evi/Sprinter is GPR177. The
retromer complex, including VPS35, may act within Wnt-producing cells to generate Wnt
forms that can be transported outside cells, possibly in the form of a lipoprotein particle.
It is already known that ependymal cells express Wnt 8b during development and in
the adult animal brain (Shimogori et al., 2004). Also, our results shows, that ependymal
cells accumulates Wnt 8b in the basal part of the cilia.
The morphology of the ependymal cells during development indicates that they
could have secretory functions. Floor plate ependymal cells in the neural tube possess
vesicles and multivesicular bodies (Yoshioka and Tanaka, 1989). Secretory droplets are
present at the apical surface of rat third ventricles ependyma prior acquisitions of cilia
(Booz, 1975). In the adult rabbit, aqueductal ependymal cells possess surface blebs and
vesicles of various sizes (Meller and Denis, 1993). Ependymal cells in adult humans show
immunoreactivity for granulophysin, a member protein associated with exocytosis
(Hatskelzon et al., 1993) and rat ependymal cells expresses mRNA for the secretory
proteins chromogranin B and secretogranin (Gee at al., 1993). Also possibly involved in
secretion is sulfated glycoprotein – 2 (clusterin) which is expressed at high levels in rat
ependymal (Danik et al., 1993; Senut et al., 1992).
The hypothesis, that ependymal cells secrete Wnt 8b, is consistent with a large body
of evidence indicating that mature ependymal cells express and synthesize Wnt 8b, also
possess proteins and organelle structure required for secretion.

52
Fig. 14. A - confocal image of
cultured ependymal cells from the
side, confirms wnt 8b accumulation in
the basal part of the cilia (green -
acetylated tubulin; red – Wnt 8b; scale
bar 2 µm). B - Maturation of Wnt.
Wnts enter into ER, where initial
glycosylation and palmitoylation
occur, and then are transported into the
Golgi apparatus, where further
glycosylation processes occur and are
finally secreted from the cell surface as
a mature and active
form. After secretion,
Wnts are associated
with proteoglycans or
other extracellular
matrix components,
and transported to the
target cells.
.

53
3.3.2 The role of Dishevelled as Wnt signal transducer in the cilia
The Wnts are a family of secreted factors that bind Frizzled receptors to activate
distinct signaling cascades depending on the specific Wnt activator, the receptor and also
the activity of Disheveled (Dvl), a molecular switch between signaling cascades. Upon
activation of the pathway, the subcellular localization of Dvl appears to determine the
final signaling outcome whereby nuclear localization of Dvl, which is required for
canonical Wnt signaling, while membrane bound Dvl favors the PCP pathway (Veeman et
al., 2003; Gerdes et al., 2008).
Fig. 15. Cilia Wnt signaling. Representation of a cilium/basal body, denoting their role in modulating
the balance between canonical and non-canonical PCP signaling. In conditions of normal ciliary/basal body

54
signaling, molecules such as Inversin (Inv) are required to favor PCP over canonical signaling by inhibiting
Dvl and activating the b-catenin destruction complex composed of Axin, GSK3-b and APC. If Dvl-1 (which
is accumulated in the cilia) is not inhibited, it will transmit further canonical Wnt signal. In contrast, if Dvl-
1 is inhibited, membranous DVL-3 should transmit non-canonical Wnt signaling. Perturbation of the basal
body (i.e., mutations in the BBS proteins) or cilia-mediated signal transduction results in decreased PCP and
the concomitant up-regulation of canonical signaling. (Cardenas et al., 2009)
Having this in mind and based on our results, we can suggest that Dvl-1 is likely
acting as canonical Wnt signal transducer; Dvl-3 – nocanonical. We observed intensive
immunoreactivity of Dvl-1 in the nucleus and in the cilia (also weak cytoplasm staining)
in the ependymal cells. Dvl-1 localization in those place shows that it is significant for
canonical Wnt signaling in this type of cells. Dvl-3 showed weak cytoplasm staining and
good signal next to the membrane. These results are not conclusive, but we can suggest
that Dvl-3 is closely relate to the membrane and possesses importance to non-canonical
Wnt signaling in the ependymal cells.
Although, studies making use of gene interruption to knock-out the expression of
Dvl1, Dvl2, or both (Hamblet et al., 2002) in mice (Lijam et al., 1997; Wang et al., 2006)
reveal some level of redundancy, also we think this is apparent in the current study.
Little is known about the regulation of Dvl expression at the levels of translation and
protein stability, but certainly interesting differences must exist among how these three
highly-homologous mammalian Dvls are processed to perform such different functions.
3.3.3 Dishevelled nuclear shuttling
Dvl has been shown to shuttle between the cytoplasm and the nuclear (Fanto et al.,
2000; Habas et al., 2005; Weitzman 2005). A nuclear export sequence (NES) and an
atypical nuclear localization sequence (NLS) were identified right after the DEP domain
and the PDZ domain (Itoh et al., 2005). Disruption of the nuclear localization of Dvl by
NLS mutation specifically impairs the -catenin pathway, but not the non-canonical
pathway. The importance in the canonical pathway of nuclear localization of mammalian
Dvl was also demonstrated (Itoh et al., 2005). In the nucleus, Dvl has been found to

55
interact with c-Jun and -catenin, followed by formation of the stable -catenin/TCF
complex and transcriptional activation of Wnt target genes (Gan et al., 2008).
We showed that portion of Dvl-1 is localized in the cilia (discussed above) and other
portion in the nucleus. Presence of Dvl 1 in the nucleus indicates, that it is “ working” as a
transducer in the canonical Wnt signaling. Dvl-3 and Dvl-2 demonstrated weak nucleus
staining, this indicates that Dvl functions are overlapping at some level.
Therefore, it is likely that there are two pools of Dvl: one translocates to the nucleus
to mediate the canonical signaling, and the other stays in the cytoplasm or moves to the
plasma membrane to mediate both canonical and non-canonical signaling, upon Wnt
stimulation.
Figure 16. Results of immunohistochemical experiments with cultured ependymal cells. Dvl-1
mainly is accumulated in the nucleus of cultured ependymal cells, white arrows indicate nucleus of the cell.
(In red – Dvl-1; scale bar 20 µm)

56
4. Conclusions
Investigation of ependymal cells in vivo and in vitro revealed significant
result to Wnt signaling and potential pharmaceutical targets in ependymal cells. I have
demonstrated that Wnt lingands and signal transducers are localized in the ependymal
cells.
1. Wnt 8b immunoreactivity, in the basal part of the cilia of ependymal cells, is the
most important finding, this is brand new location. It is know, that this part of the
cilia possesses secretion features that is why we believe that WNT 8b may be
secreted from ependymal cells to the cerebro spinal fluid. This could play
important role in prenatal cortical development and repair after injury. These
results are consistent in vivo and in vitro. To conclude, Wnt 8b is very interesting
potential target in ependymal cells.
2. We have demonstrated that Wnt 3a, Wnt 5a, Wnt 7a and Wnt 7b are not
localized in the ependymal cells. Those antibodies did not label cultured
ependymal cells or brain sections (in vivo), but there were some staining in other
brain regions, already reported in the literature. It means these antibodies work.
According to these results, we suggest that these ligands can not be targets for the
drugs, because it might be that there is no expression of these proteins in
ependymal cells. It is advisable to confirm these results with situ hybridization
method.
3. Our study possesses important results for Dishevelled molecules. We reported that
Dvl-1 is accumulated in the nucleus and in the basal part of cilia of ependymal
cells. This localization is not described in the literature. According to Dvl-1
accumulation place we can suggest that it participates in canonical WNT signaling.
It might be that drug mediated canonical Wnt signaling modification can be
achieved through Dvl-1 and it will be important drug target. Dvl-1 may be an
important drug target for treating ependymomas, because in other forms of cancer,
it is known that canonical WNT signaling pathway is over activated. In
ependymomas Dvl-1 may be drug inhibited in order to restore balance of canonical
Wnt signaling.

57
4. Analysis of Dvl-2 antibody staining with fluorescence and confocal microscope
detected no immunoreactivity in the ependymal cells lined all ventricular system
of the brains and in the cells culture. As well, there is no evidence in literature that
this protein is expressed these cells. This might be indicating that Dvl-2 does not
participate in signaling transduction processes during normal conditions in the
ependymal cells. According to this data, we strongly suggest that Dvl-2 can act as
a drug target. It is advisable to confirm these results with situ hybridization
method.
5. Dvl-3 is accumulated on the membrane of the ependymal cells. This localization is
a completely new place. Dvl-3 membrane localization suggests its involvement in
non-canonical WNT signaling, which affects planar cell polarization. According to
these data, Dvl-3 may be a good drug target for treating the diseases, which
display the impaired ciliar activity. An example could be impaired neuronal
migration in parallel with the lateral ventricle. Thus, Dvl-3 may be a good target
for adjusting the degenerative processes, because increased expression of Dvl-3
might accelerate migration of neuroblasts and astrocyte toward damaged areas,
thereby stopping further damage.

58
5. Bibliography:
1. Adachi, K. et al. -catenin signaling promotes proliferation of
progenitor cells in the adult mouse subventricular zone. Stem Cells 25, 2827–2836
(2007).
2. Adam E, Dziegielewska KM, Saunders NR, Schumacher U. (1993);
Neuraminic acid specific lectins as markers of early cortical plate neurons. Int J Dev
Neurosci. 1993 Aug;11(4):451-60.
3. Acarin L, Vela JM, González B, Castellano B (1994); Demonstration of
poly-N-acetyl lactosamine residues in ameboid and ramified microglial cells in rat
brain by tomato lectin binding. J Histochem Cytochem. 1994 Aug;42(8):1033-41.
4. Ahmad-Annuar, A. et al. Signaling across the synapse: a role for Wnt
and Dishevelled in presynaptic assembly and neurotransmitter release. J. Cell Biol.
174, 127–139 (2006).
5. Arai Y, Deguchi K, Takashima S (1998) Vascular endothelial growth
factor in brains with periventricular leukomalacia. Pediatr Neurol 19:45–49
6. Badaut J, Lasbennes F, Magistretti PJ, Regli L.,2002. Aquaporins in
brain: distribution, physiology, and pathophysiology. J Cereb Blood Flow Metab.
2002 Apr;22(4):367-78.
7. Barker N, Clevers H. Mining the Wnt pathway for cancer therapeutics.
Nat Rev Drug Discov. 2006 Dec;5(12):997-1014.
8. Baker, N. E. Localization of transcripts from the wingless gene in
whole Drosophila embryos. Development 103, 289–298 (1988).
9. Bilic, J. et al. Wnt induces LRP6 signalosomes and promotes
dishevelled-dependent LRP6 phosphorylation. Science 316, 1619–1622 (2007).
10. Bystron I, Blakemore C, Rakic P (2008) Development of the human
cerebral cortex: Boulder Committee revisited. Nat Rev Neurosci 9:110–122
11. Brightman MW, Reese TS (1969); Junctions between intimately
apposed cell membranes in the vertebrate brain. J Cell Biol. 1969 Mar;40(3):648-77.
12. Bruni JE. 1998. Ependymal development, proliferation, and functions.
Microsc Res Tech. 1998 Apr 1;41(1):2-13.

59
13. Boisvieux-Ulrich, E., Laine, M.C. & Sandoz, D. The orientation of
ciliary basal bodies in quail oviduct is related to the ciliary beating cycle
commencement. Biol. Cell 55, 147–150 (1985).
14. Boonen, R. A., van Tijn, P. & Zivkovic, D. Wnt signaling in
Alzheimer’ s disease: up or down, that is the question. Ageing Res. Rev. 8, 71–82
(2009).
15. Booz KH. 1975. Secretory phenomena at the ependyma of the IIIrd
ventricle of the embryonic rat. Anat Embryol (Berl). 1975 Aug 9;147(2):143-59.
16. Boutros M, Paricio N, Strutt DI, Mlodzik M. Dishevelled activates JNK
and discriminates between JNK pathways in planar polarity and wingless signaling.
Cell. 1998 Jul 10;94(1):109-18.
17. Boutros M, Mlodzik M. Dishevelled: at the crossroads of divergent
intracellular signaling pathways. Mech Dev. 1999 May;83(1-2):27-37.
18. Bulinski, J.C., J.E. Richards, and G. Piperno. 1988. Posttranslational
modifications of alpha-tubulin: detyrosination and acetylation differentiate
populations of interphase microtubules in cultured-cells. J. Cell Biol. 106:1213–
1220.
19. Brusco A, López-Costa JJ, Tagliaferro P, Pecci Saavedra J.
Serotonergic ependymal fibres in rat and monkey: light and electron microscopic
immunocytochemical study. Biocell. 1998 Aug;22(2):115-22.
20. G. Castelo-Branco, J.Wagner, F.J. Rodriguez, J. Kele, K. Sousa, N.
Rawal, H.A. Pasolli, E. Fuchs, J. Kitajewski, E. Arenas, Differential regulation of
midbrain dopaminergic neuron development by Wnt-1, Wnt-3a, and Wnt-5a, Proc.
Natl. Acad. Sci. USA 100 (2003) 12747–12752.
21. Caruso, A. et al. Inhibition of the canonical Wnt signaling pathway by
apolipoprotein E4 in PC12 cells. J. Neurochem. 98, 364–371 (2006).
22. Capelluto, D.G. et al. The DIX domain targets dishevelled to actin
stress fibres and vesicular membranes. Nature 419, 726–729 (2002).
23. Caricasole, A. et al. The Wnt pathway, cell-cycle activation and -
amyloid: novel therapeutic strategies in Alzheimer’ s disease? Trends Pharmacol.
Sci. 24, 233–238 (2003).

60
24. Cardenas-Rodriguez M, Badano JL. Ciliary biology: understanding the
cellular and genetic basis of human ciliopathies. Am J Med Genet C Semin Med
Genet. 2009 Nov 15;151C(4):263-80.
25. Cerpa, W., Farías, G. G., Fuenzalida, M., Bonansco, C. & Inestrosa, N.
C. Wnt-5a occludes A oligomerinduced depression of glutamatergic transmision in
CA1 pyramidal neurons from hippocampal slices. Mol. Neurodegener. 2010.
26. Cheyette BN, Waxman JS, Miller JR, Takemaru K, Sheldahl LC,
Khlebtsova N, Fox EP, Earnest T, Moon RT. Dapper, a Dishevelled-associated
antagonist of beta-catenin and JNK signaling, is required for notochord formation.
Dev Cell. 2002 Apr;2(4):449-61.
27. Chen D, Zhang J, Li M, Rayburn ER, Wang H, Zhang R. RYBP
stabilizes p53 by modulating MDM2. EMBO Rep. 2009 Feb;10(2):166-72. Epub
2008 Dec 19.
28. Chen, W. et al. Dishevelled 2 recruits b-arrestin 2 to mediate Wnt5A-
stimulated endocytosis of Frizzled 4. Science 301, 1391–1394 (2003).
29. Chenn, A. & Walsh, C. A. Regulation of cerebral cortical size by
control of cell cycle exit in neural precursors. Science 297, 365–369 (2002).
30. Ching W, Nusse R. A dedicated Wnt secretion factor. Cell 2006;
125:432-433
31. Christensen, S. T., Pedersen, L. B., Schneider, L. and Satir, P. (2007).
Sensory cilia and integration of signal transduction in human health and disease.
Traffic 8, 97-109.
32. Cohen, E., Binet, S. & Meininger, V. Ciliogenesis and centriole
formation in the mouse embryonic nervous system. An ultrastructural analysis. Biol.
Cell 62, 165–169 (1988).
33. Classen, A.K., Anderson, K.I., Marois, E. & Eaton, S. Hexagonal
packing of Drosophila wing epithelial cells by the planar cell polarity pathway. Dev.
Cell 9, 805–817 (2005).
34. Colas JF, Schoenwolf GC (2001) Towards a cellular and molecular
understanding of neurulation. Dev Dyn 221:117–145.
35. Cuevas P, Gimenez-Gallego G (2000) Fibroblast growth factor and
hydrocephalus. Neurol Res 22:102–104.

61
36. Dobell, C. (1932). Antony van Leeuwenhoek and his ‘Little Animals’ ,
p. 435. New York: Harcourt, Brace and Co.
37. Dann CE, Hsieh JC, Rattner A, Sharma D, Nathans J, Leahy DJ.
Insights into Wnt binding and signalling from the structures of two Frizzled
cysteine-rich domains. Nature. 2001 Jul 5;412(6842):86-90.
38. Danik M, Chabot JG, Hassan-Gonzalez D, Suh M, Quirion R..
Localization of sulfated glycoprotein-2/clusterin mRNA in the rat brain by in situ
hybridization. J Comp Neurol. 1993 Aug 8;334(2):209-27.
39. Duan, X., Kang, E., Liu, C. Y., Ming, G. L. & Song, H. Development
of neural stem cell in the adult brain. Curr. Opin. Neurobiol. 18, 108–115 (2008).
40. De Ferrari, G. V. & Inestrosa, N. C. Wnt signaling function in
Alzheimer’ s disease. Brain Res. Brain Res. Rev. 33, 1–12 (2000).
41. De Ferrari, G. V. et al. Activation of Wnt signaling rescues
neurodegeneration and behavioral impairments induced by -amyloid fibrils. Mol.
Psychiatry 8, 195–208 (2003).
42. Del Bigio MR., 1995. The ependyma: a protective barrier between
brain and cerebrospinal fluid. Glia. 1995 May;14(1):1-
43. Frautschy SA, Walicke PA, Baird A (1991) Localization of basic
fibroblast growth factor and its mRNA after CNS injury. Brain Res 553:291–299.
44. Fagotto F, Jho E, Zeng L, Kurth T, Joos T, Kaufmann C, Costantini F.
Domains of axin involved in protein-protein interactions, Wnt pathway inhibition,
and intracellular localization. J Cell Biol. 1999 May 17;145(4):741-56.
45. Farias, G. G. et al. M1 muscarinic receptor activation protects neurons
from -amyloid toxicity. A role for Wnt signaling pathway. Neurobiol. Dis. 17, 337–
348 (2004).
46. Fanto M, Weber U, Strutt DI, Mlodzik M. Nuclear signaling by Rac
and Rho GTPases is required in the establishment of epithelial planar polarity in the
Drosophila eye. Curr Biol. 2000 Aug 24;10(16):979-88.
47. Figarella-Branger D, Lepidi H, Poncet C, Gambarelli D, Bianco N,
Rougon G, Pellissier JF. Differential expression of cell adhesion molecules (CAM),
neural CAM and epithelial cadherin in ependymomas and choroid plexus tumors.
Acta Neuropathol. 1995;89(3):248-57.

62
48. Frisch, D. & Farbman, A.I. Development of order during ciliogenesis.
Anat. Rec. 162, 221–232 (1968).
49. Fujii N, You L, Xu Z, Uematsu K, Shan J, He B, Mikami I, Edmondson
LR, Neale G, Zheng J, Guy RK, Jablons DM. An antagonist of dishevelled protein-
protein interaction suppresses beta-catenin-dependent tumor cell growth. Cancer
Res. 2007 Jan 15;67(2):573-9.
50. Fuxe K, Tinner B, Zoli M, Pettersson RF, Baird A, Biagini G, Chadi G,
Agnati LF (1996) Computer-assisted mapping of basic fibroblast growth factor
immunoreactive nerve cell populations in the rat brain. J Chem Neuroanat 11:13–35
51. Galli LM, Barnes TL, Secrest SS, Kadowaki T, Burrus LW. Porcupine-
mediated lipid-modification regulates the activity and distribution of Wnt proteins in
the chick neural tube. Development. 2007 Sep;134(18):3339-48.
52. Gao, Z. et al. Neurod1 is essential for the survival and maturation of
adult-born neurons. Nature Neurosci. 12, 1090–1092 (2009).
53. Gee P, Rhodes CH, Fricker LD, Angeletti RH. Expression of
neuropeptide processing enzymes and neurosecretory proteins in ependyma and
choroid plexus epithelium. Brain Res. 1993 Jul 23;617(2):238-48.
54. Gerdes JM, Katsanis N. 2008. Ciliary function and Wnt signal
modulation. Curr Top Dev Biol 85:175–195.
55. Gerdes, J.M. et al. Disruption of the basal body compromises
proteasomal function and perturbs intracellular Wnt response. Nat. Genet. 39, 1350–
1360 (2007).
56. Geng L, Okuhara D, Yu Z, Tian X, Cai Y, et al. 2006. Polycystin-2
traffics to cilia independently of polycystin-1 by using an N-terminal RVxP motif. J.
Cell Sci. 119:1383–95
57. Gilula NB, Satir P. 1972. The ciliary necklace. A ciliary membrane
specialization. J. Cell Biol. 53:494–509
58. Grandy D, Shan J, Zhang X, Rao S, Akunuru S, Li H, Zhang Y,
Alpatov I, Zhang XA, Lang RA, Shi DL, Zheng JJ. Discovery and characterization
of a small molecule inhibitor of the PDZ domain of dishevelled. J Biol Chem. 2009
Jun 12;284(24):16256-63. Epub 2009 Apr 21.

63
59. Gonzalez AM, Berry M, Maher PA, Logan A, Baird A (1995) A
comprehensive analysis of the distribution of FGF-2 and FGFR1 in the rat brain.
Brain Res 701:201–226.
60. Habas R, Dawid IB, He X: Coactivation of Rac and Rho by
Wnt/Frizzled signaling is required for vertebrate gastrulation. Genes Dev 2003,
17:295-309.
61. Habas R, Dawid IB. Dishevelled and Wnt signaling: is the nucleus the
final frontier? J Biol. 2005;4(1):2. Epub 2005 Feb 17.
62. Habas R, Kato Y, He X: Wnt/Frizzled activation of Rho regulates
vertebrate gastrulation and requires a novel Formin homology protein Daam1. Cell
2001, 107:843-854.
63. Hayamizu TF, Chan PT, Johanson CE (2001) FGF-2 immunoreactivity
in adult rat ependyma and choroid plexus: responses to global forebrain ischemia
and intraventricular FGF-2. Neurol Res 23:353–358
64. Hall, A. C., Lucas, F. R. & Salinas, P. C. Axonal remodeling and
synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell
100, 525–535 (2000).
65. Hamblet NS, Lijam N, Ruiz-Lozano P, Wang J, Yang Y, Luo Z, Mei L,
Chien KR, Sussman DJ, Wynshaw-Boris A. Dishevelled 2 is essential for cardiac
outflow tract development, somite segmentation and neural tube closure.
Development 2002;129:5827–5838. [PubMed: 12421720]
66. Hatskelzon L, Dalal BI, Shalev A, Robertson C, Gerrard JM. 1993.
Wide distribution of granulophysin epitopes in granules of human tissues. Lab
Invest. 1993 May;68(5):509-19.
67. Hausmann G, Bänziger C, Basler K. Helping Wingless take flight: how
WNT proteins are secreted. Nat Rev Mol Cell Biol. 2007 Apr;8(4):331-6. Epub
2007 Mar 7.
68. Hauwel M, Furon E, Canova C, Griffiths M, Neal J, Gasque P (2005)
Innate (inherent) control of brain infection, brain inflammation and brain repair: the
role of microglia, astrocytes, ‘‘protective’ ’ glial stem cells and stromal ependymal
cells. Brain Res Brain Res Rev 48:220–233

64
69. Hebert JM, Fishell G (2008) The genetics of early telencephalon
patterning: some assembly required. Nat Rev Neurosci 9:678–685.
70. He, P. & Shen, Y. Interruption of -catenin signaling reduces
neurogenesis in Alzheimer’ s disease. J. Neurosci. 29, 6545– 6557 (2009).
71. He X, Saint-Jeannet JP, Wang Y, Nathans J, Dawid I, Varmus H. A
member of the Frizzled protein family mediating axis induction by Wnt-5A.
Science. 1997 Mar 14;275(5306):1652-4.
72. He X, Semenov M, Tamai K, Zeng X: LDL receptor-related proteins 5
and 6 in Wnt/beta-catenin signaling: arrows point the way. Development 2004,
131:1663-1677.
73. Herz J, Bock HH. Lipoprotein receptors in the nervous system. Annu
Rev Biochem. 2002;71:405-34. Epub 2001 Nov 9.
74. Hirabayashi, Y. et al. The Wnt/-catenin pathway directs neuronal
differentiation of cortical neural precursor cells. Development 131, 2791–2801
(2004).
75. Hong DH, Yue G, Adamian M, Li T. 2001. Retinitis pigmentosa
GTPase regulator (RPGRr)-interacting protein is stably associated with the
photoreceptor ciliary axoneme and anchors RPGR to the connecting cilium. J. Biol.
Chem. 276:12091–99.
76. Hong DH, Pawlyk B, Sokolov M, Strissel KJ, Yang J, et al. 2003.
RPGR isoforms in photoreceptor connecting cilia and the transitional zone of motile
cilia. Invest. Ophthalmol.Vis. Sci. 44:2413–21.
77. Horton BN, Solanki RB, Kulesza P, Ardelt AA (2009) Localization of
angiopoietin-1 and Tie2 immunoreactivity in rodent ependyma and adjacent blood
vessels suggests functional relationships. J Histochem Cytochem.
doi:10.1369/jhc.2009.954610.
78. Iwata T, Hevner RF (2009) Fibroblast growth factor signaling in
development of the cerebral cortex. Dev Growth Differ 51:299–323
79. Yang-Snyder J, Miller JR, Brown JD, Lai CJ, Moon RT. A frizzled
homolog functions in a vertebrate Wnt signaling pathway. Curr Biol. 1996 Oct
1;6(10):1302-6.

65
80. Inestrosa, N. C. & Toledo, E. M. The role of Wnt signaling in neuronal
dysfunction in Alzheimer’ s disease. Mol. Neurodegener. 3, 9 (2008).
81. Yamadori T, Nara K. (1979); The directions of ciliary beat on the wall
of the lateral ventricle and the currents of the cerebrospinal fluid in the brain
ventricles. Scan Electron Microsc. 1979;(3):335-40.
82. Yamamoto T, Kardami E, Nagy JI., 1995. Basic fibroblast growth
factor in rat brain: localization to glial gap junctions correlates with connexin43
distribution. Brain Res. 1991 Jul 19;554(1-2):336-43.
83. Yanagawa S, van Leeuwen F, Wodarz A, Klingensmith J, Nusse. The
dishevelled protein is modified by wingless signaling in Drosophila. Genes Dev.
1995 May 1;9(9):1087-97.
84. Ye X, Carp RI (2002) Increase of acidic fibroblast growth factor in the
brains of hamsters infected with either 263K or 139H strains of scrapie. J Mol
Neurosci 18:179–188.
85. Yoshioka T, Tanaka O. 1989. Ultrastructural and cytochemical
characterisation of the floor plate ependyma of the developing rat spinal cord. J
Anat. 1989 Aug;165:87-100.
86. Yu, A. et al. Association of Dishevelled with the clathrin AP-2 adaptor
is required for Frizzled endocytosis and planar cell polarity signaling. Dev. Cell 12,
129–141 (2007).
87. Yu S, Tooyama I, Ding WG, Kitasato H, Kimura H (1995)
Immunohistochemical localization of glucose transporters (GLUT1 and GLUT3) in
the rat hypothalamus. Obes Res 3(Suppl 5):753S–776S.
88. Yu, J. M., Kim, J. H., Song, G. S. & Jung, J. S. Increase in proliferation
and differentiation of neural progenitor cells isolated from postnatal and adult mice
brain by Wnt-3a and Wnt-5a. Mol. Cell Biochem. 288, 17–28 (2006).
89. Jekely, G. and Arendt, D. (2006). Evolution of intraflagellar transport
from coated vesicles and autogenous origin of the eukaryotic cilium. BioEssays 28,
191-198.
90. Jessberger, S. et al. Dentate gyrus-specific knockdown of adult
neurogenesis impairs spatial and object recognition memory in adult rats. Learn.
Mem. 16, 147–154 (2009).

66
91. Johnson, N. T., Villalon, M., Royce, F. H., Hard, R. and Verdugo, P.
(1991). Autoregulation of beat frequency in respiratory ciliated cells. Demonstration
by viscous loading. Am. Rev. Respir. Dis. 144, 1091-1094.
92. Keiji Itoh, Barbara K Brott, Gyu-Un Bae, Marianne J Ratcliffe, and
Sergei Y Sokol. Nuclear localization is required for Dishevelled function in Wnt/-
catenin signaling. J Biol. 2005; 4(1): 3. Published online 2005 February 15. doi:
10.1186/jbiol20.
93. Keller R: Shaping the vertebrate body plan by polarized embryonic cell
movements. Science 2002, 298:1950-1954.
94. Kennerdell JR, Carthew RW. Use of dsRNA-mediated genetic
interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway.
Cell. 1998 Dec 23;95(7):1017-26.
95. Kim, J.C. et al. The Bardet-Biedl protein BBS4 targets cargo to the
pericentriolar region and is required for microtubule anchoring and cell cycle
progression. Nat. Genet. 36, 462–470 (2004).
96. Kimura J, Suda Y, Kurokawa D, Hossain ZM, Nakamura M, Takahashi
M, Hara A, Aizawa S. 2005. Emx2 and Pax6 function in cooperation with Otx2 and
Otx1 to develop caudal forebrain primordium that includes future archipallium. J
Neurosci 25:5097–5108.
97. Kishida, S. et al. Dvl regulates endo- and exocytotic processes through
binding to synaptotagmin. Genes Cells 12, 49–61 (2007).
98. Kobayashi M, Nikami H, Morimatsu M, Saito M (1996) Expression
and localization of insulin-regulatable glucose transporter (GLUT4) in rat brain.
Neurosci Lett 213:103–106.
99. Komekado H, Yamamoto H, Chiba T, Kikuchi Glycosylation and
palmitoylation of Wnt-3a are coupled to produce an active form of Wnt-3a. A.
Genes Cells. 2007 Apr;12(4):521-34.
100. Korkut C, Budnik V. WNTs tune up the neuromuscular junction. Nat
Rev Neurosci. 2009 Sep;10(9):627-34.
101. Kowalevsky, A. (1867). Entwickelungsgeschichte des Amphioxus
lanceolatus. Memoires de l’ Academie Imperiale des Sciences de St.-Petersbourg
VII. 11, 1-17.

67
102. Kramer-Zucker, A.G. et al. Cilia-driven fluid flow in the zebrafish
pronephros, brain and Kupffer’ s vesicle is required for normal organogenesis.
Development 132, 1907–1921 (2005).
103. Kuhl M: Non-canonical Wnt signaling in Xenopus: regulation of axis
formation and gastrulation. Semin Cell Dev Biol 2002, 13:243-249.
104. Kuwabara, T. et al. Wnt-mediated activation of NeuroD1 and retro-
elements during adult neurogenesis. Nature Neurosci. 12, 1097–1105 (2009).
105. Lacor, P. N. et al. A oligomer-induced aberrations in synapse
composition, shape, and density provide a molecular basis for loss of connectivity in
Alzheimer’ s disease. J. Neurosci. 27, 796–807 (2007).
106. Lako MS, Lindsay S, Bullen P, Wilson DI, Robson SC, Strachan T.
1998. A novel mammalian wnt gene, WNT8B, shows brain-restricted expression in
early development, with sharply delimited expression boundaries in the developing
forebrain. Hum Mol Genet 7:813–822.
107. Langerhans, P. (1876). Zur Anatomie des Amphioxus. Archiv fur
Mikrokopische Anatomie 12, 290-348.
108. Lange BM, Faragher AJ, March P, Gull K (2000) Centriole duplication
and maturation in animal cells. Curr Top Dev Biol 49:235–249.
109. Lanzetti, L. Actin in membrane trafficking. Curr. Opin. Cell Biol. 19,
453–458 (2007)
110. Lee HJ, Wang NX, Shao Y, Zheng JJ. Identification of tripeptides
recognized by the PDZ domain of Dishevelled. Bioorg Med Chem. 2009 Feb
15;17(4):1701-8. Epub 2009 Jan 3.
111. Li X, Kong H, Wu W, Xiao M, Sun X, Hu G. Aquaporin-4 maintains
ependymal integrity in adult mice. Neuroscience. 2009 Aug 4;162(1):67-77. Epub
2009 Apr 22.
112. Lie, D. C., Song, H., Colamarino, S. A., Ming, G. L. & Gage, F. H.
Neurogenesis in the adult brain: new strategies for central nervous system diseases.
Annu. Rev. Pharmacol. Toxicol. 44, 399–421 (2004).
113. Li AJ,OomuraY,SasakiK, SuzukiK,Tooyama I,HanaiK,Kimura H, Hori
T (1998) A single pre-training glucose injection induces memory facilitation in

68
rodents performing various tasks: contribution of acidic fibroblast growth factor.
Neuroscience 85:785–794
114. Lijam N, Paylor R, McDonald MP, Crawley JN, Deng CX, Herrup K,
Stevens KE, Maccaferri G, McBain CJ, Sussman DJ, Wynshaw-Boris A. Social
interaction and sensorimotor gating abnormalities in mice lacking Dvl1. Cell
1997;90:895–905. [PubMed: 9298901]
115. Ling EA, Kaur C, Lu J (1998) Origin, nature, and some functional
considerations of intraventricular macrophages, with special reference to the
epiplexus cells. Microsc Res Tech 41:43–56.
116. Liu C, Kato Y, Zhang Z, Do VM, Yankner BA, He X: beta-Trcp
couples beta-catenin phosphorylation-degradation and regulates Xenopus axis
formation. Proc Natl Acad Sci USA 1999, 96:6273-6278.
117. Liu G, Bafico A, Harris VK, Aaronson SA. A novel mechanism for
Wnt activation of canonical signaling through the LRP6 receptor. Mol Cell Biol.
2003 Aug;23(16):5825-35.
118. Liu T, DeCostanzo AJ, Liu X, Wang Hy, Hallagan S, Moon RT,
Malbon CC. G protein signaling from activated rat frizzled-1 to the beta-catenin-
Lef-Tcf pathway. Science. 2001 Jun 1;292(5522):1718-22.
119. Logan CY, Nusse R: The Wnt signaling pathway in development and
disease. Annu Rev Cell Dev Biol 2004, 20:781-810.
120. Lorenzo, I. M., Liedtke, W., Sanderson, M. J. and Valverde, M. A.
(2008). TRPV4 channel participates in receptor-operated calcium entry and ciliary
beat frequency regulation in mouse airway epithelial cells. Proc. Natl. Acad. Sci.
USA 105, 12611-12616.
121. Lucas, F.R., and P.C. Salinas. 1997. WNT-7a induces axonal
remodeling and increases synapsin I levels in cerebellar neurons. Dev. Biol. 192:31–
44.
122. Maharaj AS, Walshe TE, Saint-Geniez M, Venkatesha S, Maldonado
AE, Himes NC, Matharu KS, Karumanchi SA, D’ Amore PA (2008) VEGF and
TGF-beta are required for the maintenance of the choroid plexus and ependyma. J
Exp Med 205:491–501.

69
123. Magdesian, M. H. et al. Amyloid- binds to the extracellular cysteine-
rich domain of Frizzled and inhibits Wnt/-catenin signaling. J. Biol. Chem. 283,
9359–9368 (2008).
124. Marlow F, Topczewski J, Sepich D, Solnica-Krezel L: Zebrafish Rho
kinase 2 acts downstream of Wnt11 to mediate cell polarity and effective
convergence and extension movements. Curr Biol 2002, 12:876-884.
125. Mao J, Wang J, Liu B, Pan W, Farr GH 3rd, Flynn C, Yuan H, Takada
S, Kimelman D, Li L, Wu D. Low-density lipoprotein receptor-related protein-5
binds to Axin and regulates the canonical Wnt signaling pathway. Mol Cell. 2001
Apr;7(4):801-9.
126. McMahon, A. P. & Moon, R. T. int-1 — a proto-oncogene involved in
cell signalling. Development 107 (Suppl.), 161–167 (1989).
127. Meller ST, Dennis BJ. 1993. A scanning and transmission electron
microscopic analysis of the cerebral aqueduct in the rabbit. Anat Rec. 1993
Sep;237(1):124-40.
128. Michaloudi HC, Papadopoulos GC (1996) Catecholaminergic and
serotoninergic fibres innervate the ventricular system of the hedgehog CNS. J Anat
189(Pt 2):273–283.
129. Ming, G. L. & Song, H. Adult neurogenesis in the mammalian central
nervous system. Annu. Rev. Neurosci. 28, 223–250 (2005).
130. Mitchell, B., Jacobs, R., Li, J., Chien, S. & Kintner, C. A positive
feedback mechanism governs the polarity and motion of motile cilia. Nature 447,
97–101 (2007).
131. Miller JR, Hocking AM, Brown JD, Moon RT: Mechanism and
function of signal transduction by the Wnt/_-catenin and Wnt/Ca2+ pathways.
Oncogene 1999, 18:7860-7872.
132. Mirzadeh Z, Merkle FT, Soriano-Navarro M, Garcia-Verdugo JM,
Alvarez-Buylla A., 2008. Neural stem cells confer unique pinwheel architecture to
the ventricular surface in neurogenic regions of the adult brain. Cell Stem Cell. 2008
Sep 11;3(3):265-78
133. Mobasheri A, Wray S, Marples D. Distribution of AQP2 and AQP3
water channels in human tissue microarrays. J Mol Histol. 2005 Feb;36(1-2):1-14.

70
134. Mobasheri A, Marples D. Expression of the AQP-1 water channel in
normal human tissues: a semiquantitative study using tissue microarray technology.
Am J Physiol Cell Physiol. 2004 Mar;286(3):C529-37. Epub 2003 Oct 30.
135. Mudher, A. & Lovestone, S. Alzheimer’ s disease-do tauists and baptists
finally shake hands? Trends Neurosci. 25, 22–26 (2002).
136. Muller, O. F. (1786). Animalcula infusoria; fluvia tilia et marina, que
detexit, systematice descripsit et ad vivum delineari curavit. Havniae: Typis N.
Molleri.
137. Murata K, Mitsuoka K, Hirai T, Walz T, Agre P, Heymann JB, Engel
A, Fujiyoshi Y. 2000. Structural determinants of water permeation through
aquaporin-1. Nature. 2000 Oct 5;407(6804):599-605.
138. Nachury, M.V. et al. A core complex of BBS proteins cooperates with
the GTPase Rab8 to promote ciliary membrane biogenesis. Cell 129, 1201–1213
(2007).
139. Nusse, R. Wnt signaling and stem cell control. Cell Res. 18, 523–527
(2008).
140. Nunes, P. V., Forlenza, O. V. & Gattaz, W. F. Lithium and risk for
Alzheimer’ s disease in elderly patients with bipolar disorder. Br. J. Psychiatry 190,
359–360 (2007).
141. Nusse, R. & Varmus, H. E. Many tumors induced by the mouse
mammary tumor virus contain a provirus integrated in the same region of the host
genome. Cell 31, 99–109 (1982).
142. Nguyen T, Chin WC, O’ Brien JA, Verdugo P, Berger AJ (2001)
Intracellular pathways regulating ciliary beating of rat brain ependymal cells. J
Physiol 531:131–140.
143. Nourhaghighi N, Teichert-Kuliszewska K,Davis J, Stewart DJ,Nag S
(2003) Altered expression of angiopoietins during blood-brain barrier breakdown
and angiogenesis. Lab Invest 83:1211–1222.
144. Oishi, I., Kawakami, Y., Raya, A., Callol-Massot, C. & Izpisua
Belmonte, J.C. Regulation of primary cilia formation and left-right patterning in
zebrafish by a noncanonical Wnt signaling mediator, duboraya. Nat. Genet. 38,
1316–1322 (2006).

71
145. Oomura Y, Sasaki K, Suzuki K, Muto T, Li AJ, Ogita Z, Hanai K,
Tooyama I, Kimura H, Yanaihara N (1992) A new brain glucosensor and its
physiological significance. Am J Clin Nutr 55:278S–282S
146. Pinson KI, Brennan J, Monkley S, Avery BJ, Skarnes WC. An LDL-
receptor-related protein mediates Wnt signalling in mice. Nature. 2000 Sep
28;407(6803):535-8.
147. Panizzi, J.R., Jessen, J.R., Drummond, I.A. & Solnica-Krezel, L. New
functions for a vertebrate Rho guanine nucleotide exchange factor in ciliated
epithelia. Development 134, 921–931 (2007).
148. Patapoutian, A., and L.F. Reichardt. 2000. Roles of Wnt proteins in
neural development and maintenance. Curr. Opin. Neurobiol. 10:392–399.
149. Park, S. C., Yibchok-Anun, S., Cheng, H., Young, T. F., Thacker, E. L.,
Minion, F. C., Ross, R. F. and Hsu, W. H. (2002). Mycoplasma hyopneumoniae
increases intracellular calcium release in porcine ciliated tracheal cells. Infect.
Immun. 70, 2502-2506.
150. Park TJ, Mitchell BJ, Abitua PB, Kintner C, Wallingford JB.
Dishevelled controls apical docking and planar polarization of basal bodies in
ciliated epithelial cells. Nat Genet. 2008 Jul;40(7):871-9. Epub 2008 Jun 15.
151. Park, T.J., Gray, R.S., Sato, A., Habas, R. & Wallingford, J.B.
Subcellular localization and signaling properties of dishevelled in developing
vertebrate embryos. Curr. Biol. 15, 1039–1044 (2005).
152. Parr BA, Shea MJ, Vassileva G, McMahon AP. 1993. Mouse Wnt
genes exhibit discrete domains of expression in the early embryonic CNS and limb
buds. Development. 1993 Sep;119(1):247-61.
153. Paulson JC. (1989); Glycoproteins: what are the sugar chains for?;
Trends Biochem Sci. 1989 Jul;14(7):272-6.
154. Rodriguez-Perez LM, Perez-Martin M, Jimenez AJ, Fernandez- Llebrez
P (2003) Immunocytochemical characterisation of the wall of the bovine lateral
ventricle. Cell Tissue Res 314:325–335
155. Prochnow N, Dermietzel R., 2008. Connexons and cell adhesion: a
romantic phase. Histochem Cell Biol. 2008 Jul;130(1):71-7. Epub 2008 May 15.

72
156. Prothmann C, Wellard J, Berger J, Hamprecht B, Verleysdonk S (2001)
Primary cultures as a model for studying ependymal functions: glycogen metabolism
in ependymal cells. Brain Res 920:74–83.
157. Redies C, Takeichi M. (1996); Cadherins in the developing central
nervous system: an adhesive code for segmental and functional subdivisions. Dev
Biol. 1996 Dec 15;180(2):413-23
158. Roth Y, Kimhi Y, Edery H, Aharonson E, Priel Z.(1985); Ciliary
motility in brain ventricular system and trachea of hamsters. Brain Res. 1985 Mar
25;330(2):291-7.
159. Ross, A.J. et al. Disruption of Bardet-Biedl syndrome ciliary proteins
perturbs planar cell polarity in vertebrates. Nat. Genet. 37, 1135–1140 (2005).
160. Rulifson EJ, Wu CH, Nusse R. Pathway specificity by the bifunctional
receptor frizzled is determined by affinity for wingless. Mol Cell. 2000 Jul;6(1):117-
26. Genes Dev. 1996 Sep 1;10(17):2189-97.
161. Sanderson, M. J. and Dirksen, E. R. (1986). Mechanosensitivity of
cultured ciliated cells from the mammalian respiratory tract: implications for the
regulation of mucociliary transport. Proc. Natl. Acad. Sci. USA 83, 7302-7306.
162. Salathe, M. (2006). Regulation of mammalian ciliary beating. Annu.
Rev. Physiol. 69, 401-422.
163. Sarnat HB (1998) Histochemistry and immunocytochemistry of the
developing ependyma and choroid plexus. Microsc Res Tech 41:14–28.
164. Sarnat HB (1992) Regional differentiation of the human fetal
ependyma: immunocytochemical markers. J Neuropathol Exp Neurol 51:58–75.
165. Sarnat HB (1992) Role of human fetal ependyma. Pediatr Neurol
8:163–178
166. Sarnat HB (1995) Ependymal reactions to injury. A review. J
Neuropathol Exp Neurol 54:1–15.
167. Satir P, Guerra C. 2003. Control of ciliary motility: a unifying
hypothesis. Europ. J. Protistol. 39:410–15
168. Satir, P., Mitchell, D. R. and Jékely, G. (2008). How did the cilium
evolve? Curr. Top. Dev. Biol. 85, 63-82.

73
169. Sawa H, Lobel L, Horvitz HR. The Caenorhabditis elegans gene lin-17,
which is required for certain asymmetric cell divisions, encodes a putative seven-
transmembrane protein similar to the Drosophila frizzled protein.
170. Schwarz-Romond, T., Merrifield, C., Nichols, B.J. & Bienz, M. The
Wnt signaling effector Dishevelled forms dynamic protein assemblies rather than
stable associations with cytoplasmic vesicles. J. Cell Sci. 118, 5269–5277 (2005).
171. Schwarz-Romond T, Fiedler M, Shibata N, Butler PJ, Kikuchi A,
Higuchi Y, Bienz M. The DIX domain of Dishevelled confers Wnt signaling by
dynamic polymerization. Nat Struct Mol Biol. 2007 Jun;14(6):484-92. Epub 2007
May 27.
172. Shankar, G. M. et al. Amyloid-�
protein dimers isolated directly from
Alzheimer’ s brains impair synaptic plasticity and memory. Nature Med. 14, 837–
842 (2008).
173. Schwarz-Romond T, Metcalfe C, Bienz M Dynamic recruitment of axin
by Dishevelled protein assemblies. J Cell Sci. 2007 Jul 15;120(Pt 14):2402-12.
174. Senut MC, Jazat F, Choi NH, Lamour Y. Protein SP40,40-like
Immunoreactivity in the Rat Brain: Progressive Increase With Age. Eur J Neurosci.
1992;4(10):917-928.
175. Shimogori T, VanSant J, Paik E, Grove EA. 2004. Members of the
Wnt, Fz, and Frp gene families expressed in postnatal mouse cerebral cortex. J
Comp Neurol. 2004 Jun 7;473(4):496-510.
176. Shah, A. S., Ben-Shahar, Y., Moninger, T. O., Kline, J. N. and Welsh,
M. J. (2009). Motile cilia of human airway epithelia are chemosensory. Science 325,
1131-1134.
177. Shan J, Shi DL, Wang J, Zheng J. Identification of a specific inhibitor
of the dishevelled PDZ domain. Biochemistry. 2005 Nov 29;44(47):15495-503.
178. Sheldahl LC, Slusarski DC, Pandur P, Miller JR, Kühl M, Moon RT.
Dishevelled activates Ca2+ flux, PKC, and CamKII in vertebrate embryos. J Cell
Biol. 2003 May 26;161(4):769-77.
179. Semënov MV, Tamai K, Brott BK, Kühl M, Sokol S, He X. Head
inducer Dickkopf-1 is a ligand for Wnt coreceptor LRP6. Curr Biol. 2001 Jun
26;11(12):951-61.

74
180. Simons, M. et al. Inversin, the gene product mutated in
nephronophthisis type II, functions as a molecular switch between Wnt signaling
pathways. Nat. Genet. 37, 537–543 (2005).
181. Silva-Alvarez C, Carrasco M, Balmaceda-Aguilera C, Pastor P, Garcia
Mde L, Reinicke K, Aguayo L, Molina B, Cifuentes M, Medina R, Nualart F (2005)
Ependymal cell differentiation and GLUT1 expression is a synchronous process in
the ventricular wall. Neurochem Res 30:1227–1236.
182. Small, S. A. & Duff, K. Linking A and tau in late-onset Alzheimer’ s
disease: a dual pathway hypothesis. Neuron 60, 534–542 (2008).
183. Smalley MJ, Signoret N, Robertson D, Tilley A, Hann A, Ewan K,
Ding Y, Paterson H, Dale TC. Dishevelled (Dvl-2) activates canonical Wnt
signalling in the absence of cytoplasmic puncta. J Cell Sci. 2005 Nov 15;118(Pt
22):5279-89. Epub 2005 Nov 1.
184. Spassky N, Merkle FT, Flames N, Tramontin AD, García-Verdugo JM,
Alvarez-Buylla A., 2005, Adult ependymal cells are postmitotic and are derived
from radial glial cells during embryogenesis. J Neurosci. 2005 Jan 5;25(1):10-8.
185. Steiner J, Bernstein HG, Bielau H, Berndt A, Brisch R, Mawrin C,
Keilhoff G, Bogerts B (2007) Evidence for a wide extraastrocytic distribution of
S100B in human brain. BMC Neurosci 8:2
186. Sorokin, S.P. Reconstructions of centriole formation and ciliogenesis in
mammalian lungs. J. Cell Sci. 3, 207–230 (1968).
187. Sussman, D.J., J. Klingensmith, P. Salinas, P.S. Adams, R. Nusse, and
N. Perrimon. 1994. Isolation and characterization of a mouse homolog of the
Drosophila segment polarity gene dishevelled. Dev. Biol. 166:73–86.
188. Tamai K, Zeng X, Liu C, Zhang X, Harada Y, Chang Z, He X. A
mechanism for Wnt coreceptor activation. Mol Cell. 2004 Jan 16;13(1):149-56.
189. Takada R, Satomi Y, Kurata T, Ueno N, Norioka S, Kondoh H, Takao
T, Takada S. Monounsaturated fatty acid modification of Wnt protein: its role in
Wnt secretion. Dev Cell. 2006 Dec;11(6):791-801.
190. Theil T, Aydin S, Koch S, Grotewold L, Ru¨ ther U. 2002. Wnt and
Bmp signaling cooperatively regulate graded Emx2 expression in the dorsal
telencephalon. Development 129:3045–3054.

75
191. Tonchev AB, Yamashima T, Guo J, Chaldakov GN, Takakura N (2007)
Expression of angiogenic and neurotrophic factors in the progenitor cell niche of
adult monkey subventricular zone. Neuroscience 144:1425–1435
192. Tole S, Ragsdale CW, Grove EA. 2000. Dorsoventral patterning of the
telencephalon is disrupted in the mouse mutant extra-toes (J). Dev Biol 217:254–
265.
193. Toledo, E. M. & Inestrosa, N. C. Activation of Wnt signaling by
lithium and rosiglitazone reduced spatial memory impairment and
neurodegeneration in brains of APPswe/PSEN1�E9 mouse model of Alzheimer’ s
disease. Mol. Psychiatry 21 Jul 2009 (doi:10.1038/ mp.2009.72).
194. Tuomanen E. 1990. The surface of mammalian respiratory cilia. In
Ciliary and Flagellar Membranes, ed. RA Bloodgood, pp. 363–88. New York:
Plenum.
195. Umbhauer M, Djiane A, Goisset C, Penzo-Méndez A, Riou JF, Boucaut
JC, Shi DL. The C-terminal cytoplasmic Lys-thr-X-X-X-Trp motif in frizzled
receptors mediates Wnt/beta-catenin signalling. EMBO J. 2000 Sep 15;19(18):4944-
54.
196. Veeman MT, Axelrod JD, Moon RT: A second canon. Functions and
mechanisms of beta-catenin-independent Wnt signaling. Dev Cell 2003, 5:367-377.
197. Verleysdonk S, Kistner S, Pfeiffer-Guglielmi B, Wellard J, Lupescu A,
Laske J, Lang F, Rapp M, Hamprecht B (2005) Glycogen metabolism in rat
ependymal primary cultures: regulation by serotonin. Brain Res 1060:89–99.
198. Vigh B, Manzano e Silva MJ, Frank CL, Vincze C, Czirok SJ, Szabo A,
Lukats A, Szel A (2004) The system of cerebrospinal fluid-contacting neurons. Its
supposed role in the nonsynaptic signal transmission of the brain. Histol Histopathol
19:607–628.
199. Vives V, Alonso G, Solal AC, Joubert D, Legraverend C (2003)
Visualization of S100B-positive neurons and glia in the central nervous system of
EGFP transgenic mice. J Comp Neurol 457:404–419
200. Wang J, Hamblet NS, Mark S, Dickinson ME, Brinkman BC, Segil N,
Fraser SE, Chen P, Wallingford JB, Wynshaw-Boris A. Dishevelled genes mediate a

76
conserved mammalian PCP pathway to regulate convergent extension during
neurulation. Development 2006;133:1767 1778. [PubMed: 16571627].
201. Wang LX, Yin RX, Sun JB (2008) Effect of Tongxinluo on nestin and
vascular endothehal growth factor mRNA expression in rat brain tissue after cerebral
ischemia-reperfusion injury. Nan Fang Yi Ke Da Xue Xue Bao 28:2131–2135.
202. Wallingford JB, Fraser SE, Harland RM: Convergent extension: the
molecular control of polarized cell movement during embryonic development. Dev
Cell 2002, 2:695-706.
203. Wallingford JB (2006) Planar cell polarity, ciliogenesis and neural tube
defects. Hum Mol Genet 15 Spec No 2:R227–R234.
204. Wallingford, J.B. & Habas, R. The developmental biology of
Dishevelled: an enigmatic protein governing cell fate and cell polarity. Development
132, 4421–4436 (2005).
205. Wexler, E. M., Paucer, A., Kornblum, H. I., Plamer, T. D. &
Geschwind, D. H. Endogenous Wnt signaling maintains neural progenitor cell
potency. Stem Cells 27, 1130–1141 (2009).
206. Wehrli M, Dougan ST, Caldwell K, O'Keefe L, Schwartz S, Vaizel-
Ohayon D, Schejter E, Tomlinson A, DiNardo S. Arrow encodes an LDL-receptor-
related protein essential for Wingless signalling. Nature. 2000 Sep
28;407(6803):527-30. Wharton KA Jr. Runnin' with the Dvl: proteins that associate
with Dsh/Dvl and their significance to Wnt signal transduction. Dev Biol. 2003 Jan
1;253(1):1-17.
207. Weitzman JB. Dishevelled nuclear shuttling. J Biol. 2005;4(1):1. Epub
2005 Feb 16.
208. Wharton KA Jr: Runnin’ with the Dvl: proteins that associate with
Dsh/Dvl and their significance to Wnt signal transduction. Dev Biol 2003, 253:1-17.
209. Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya
T, Yates JR 3rd, Nusse R. Wnt proteins are lipid-modified and can act as stem cell
growth factors. Nature. 2003 May 22;423(6938):448-52. Epub 2003 Apr 27.
210. Willert K, Brink M, Wodarz A, Varmus H, Nusse R. Casein kinase 2
associates with and phosphorylates dishevelled. EMBO J. 1997 Jun 2;16(11):3089-
96.

77
211. Wodarz, A., and R. Nusse. 1998. Mechanisms of Wnt signaling in
development. Annu. Rev. Cell Dev. Biol. 14:59–88.
212. Wong HC, Mao J, Nguyen JT, Srinivas S, Zhang W, Liu B, Li L, Wu
D, Zheng J. Structural basis of the recognition of the dishevelled DEP domain in the
Wnt signaling pathway. Nat Struct Biol. 2000 Dec;7(12):1178-84.
213. Wong HC, Bourdelas A, Krauss A, Lee HJ, Shao Y, Wu D, Mlodzik M,
Shi DL, Zheng J. Direct binding of the PDZ domain of Dishevelled to a conserved
internal sequence in the C-terminal region of Frizzled. Mol Cell. 2003
Nov;12(5):1251-60.
214. Xiao-qing Gan, Ji-yong Wang, Ying Xi, Zhi-li Wu, Yi-ping Li, and Lin
Li1. Nuclear Dvl, c-Jun, -catenin, and TCF form a complex leading to stabiLization
of -catenin–TCF interaction. J Cell Biol. 2008 March 24; 180(6): 1087–1100. doi:
10.1083/jcb.200710050.
215. Zeng, L., F. Fagotto, T. Zhang, W. Hsu, T.J. Vasicek, W.L. Perry, J.J.
Lee, S.M. Tilghman, B.M. Gumbiner, and F. Costantini. 1997. The mouse fused
locus encodes Axin, an inhibitor of the Wnt signaling pathway that regulates
embryonic axis formation. Cell. 90:181–192.
216. Zimmermann, K. W. (1898). Beitrage zur Kenntniss einiger Drusen und
Epithelien. Archiv fur Mikroskopische Anatomie 52, 552-706.
217. Zhang Y, Appleton BA, Wiesmann C, Lau T, Costa M, Hannoush RN,
Sidhu SS. Inhibition of Wnt signaling by Dishevelled PDZ peptides. Nat Chem Biol.
2009 Apr;5(4):217-9. Epub 2009 Mar 1.
218. Zhou, C.-J., Zhao, C. & Pleasure, S. J. Wnt signaling mutants have
decreased dentate granule cell production and radial glial scaffolding abnormalities.
J. Neurosci. 24, 121–126 (2004).